THE EVOLUTION OF MAN
Volume II
CHAPTER XXIX
EVOLUTION OF THE SEXUAL ORGANS
If we measure the importance of the systems of organs in the animal frame according to the richness and variety of their phenomena and the physiological interest that this implies, we must regard as one of the principal and most interesting systems the one which we are now going to examine—the system of the reproductive organs. Just as nutrition is the first and most urgent condition for the self-maintenance of the individual organism, so reproduction alone secures the maintenance of the species—or, rather, the maintenance of the long series of generations which the totality of the organic stem represents in their genealogical connection. No individual organism has the prerogative of immortality. To each is allotted only a brief span of personal development, an evanescent moment in the million-year course of the history of life.
Hence, reproduction and the correlative phenomenon, heredity, have long been regarded, together with nutrition, as the most important and fundamental function of living things, and it has been attempted to distinguish them from “lifeless bodies” on this very score. As a matter of fact, this division is not so profound and thorough as it seems to be, and is generally supposed to be. If we examine carefully the nature of the reproductive process, we soon see that it can be reduced to a general property that is found in inorganic as well as organic bodies—growth. Reproduction is a nutrition and growth of the organism beyond the individual limit, which raises a part of it into the whole. This is most clearly seen when we study it in the simplest and lowest organisms, especially the Monera (Figs. 226–228) and the unicellular Amœbæ (Fig. 17). There the simple individual is a single plastid. As soon as it has reached a certain limit of size by continuous feeding and normal growth, it cannot pass it, but divides, by simple cleavage, into two equal halves. Each of these halves then continues its independent life, and grows on until it in turn reaches the limit of growth, and divides. In each of these acts of self-cleavage two new centres of attraction are formed for the particles of bodies, the foundations of the two new-formed individuals. There is no such thing as immortality even in these unicellulars.
[ 331 ]
The individual as such is annihilated in the act of cleavage (cf. p. 48).
In many other Protozoa reproduction takes place not by cleavage, but by budding (gemmation). In this case the growth that determines reproduction is not total (as in segmentation), but partial. Hence in gemmation also we may oppose the local growth-product, that becomes a new individual in the bud, as a child-organism to the parent-organism from which it is formed. The latter is older and larger than the former. In cleavage the two products are equal in age and morphological value. Next to gemmation we have, as other forms of asexual reproduction, the forming of embryonic buds and the forming of embryonic cells. But the latter leads us at once to sexual generation, the distinctive feature of which is the separation of the sexes. I have dealt fully with these various types of reproduction in my History of Creation (chap. viii) and my Wonders of Life (chap. xi).
The earliest ancestors of man and the higher animals had no faculty of sexual reproduction, but multiplied solely by asexual means—cleavage, gemmation, or the formation of embryonic buds or cells, as many Protozoa still do. The differentiation of the sexes came at a later stage. We see this most plainly in the Protists, in which the union of two individuals precedes the continuous cleavage of the unicellular organism (transitory conjugation and permanent copulation of the Infusoria). We may say that in this case the growth (the condition of reproduction) is attained by the coalescence of two full-grown cells into a single, disproportionately large individual. At the same time, the mixture of the two plastids causes a rejuvenation of the plasm. At first the copulating cells are quite homogeneous; but natural selection soon brings about a certain contrast between them—larger female cells (macrospores) and smaller male cells (microspores). It must be a great advantage in the struggle for life for the new individual to have inherited different qualities from the two cellular parents. The further advance of this contrast between the generating cells led to sexual differentiation. One cell became the female ovum (macrogonidion), and the other the male sperm-cell (microgonidion).
The simplest forms of sexual reproduction among the living Metazoa are seen in the Gastræads p. 233, the lower sponges, the common fresh-water polyp (Hydra), and other Cœlenteria of the lowest rank. Prophysema (Fig. 234), Olynthus (Fig. 238), Hydra, etc., have very simple tubular bodies, the thin wall of which consists (as in the original gastrula) only of the two primary germinal layers. As soon as the body reaches sexual maturity, a number of the cells in its wall become female ova, and others male sperm-cells: the former become very large, as they accumulate a considerable quantity of yelk-granules in their protoplasm (Fig. 235 e); the latter are very small on account of their repeated cleavage, and change into mobile cone-shaped spermatozoa (Fig. 20). Both kinds of cells detach from their source of origin, the primary germinal layers, fall either into the surrounding water or into the cavity of the gut, and unite there by fusing together. This is the momentous process of fecundation, which we have examined in Chapter VII (cf. Figs. 23–29).
From these simplest forms of sexual propagation, as we can observe them to-day in the lowest Zoophytes, the Gastræads, Sponges, and Polyps, we gather most important data. In the first place, we learn that, properly speaking, nothing is required for sexual reproduction except the fusion or coalescence of two different cells—a female ovum and male sperm-cell. All other features, and all the very complex phenomena that accompany the sexual act in the higher animals, are of a subordinate and secondary character, and are later additions to this simple, primary process of copulation and fecundation. But if we bear in mind how extremely important a part this relation of the two sexes plays in the whole of organic nature, in the life of plants, of animals, and of man; how the mutual attraction of the sexes, love, is the mainspring of the most remarkable processes—in fact, one of the chief mechanical causes of the highest development of life—we cannot too greatly emphasise this tracing of love to its source, the attractive force of two erotic cells.
Throughout the whole of living nature the greatest effects proceed from this very small cause. Consider the part that the flowers, the sexual organs of the flowering plants, play in nature; or the exuberance of wonderful phenomena that sexual selection produces in animal life; or the
[ 332 ]
momentous influence of love in the life of man. In every case the fusion of two cells is the sole original motive power; in every case this invisible process profoundly affects the development of the most varied structures. We may say, indeed, that no other organic process can be compared to it for a moment in comprehensiveness and intensity of action. Are not the Semitic myth of Adam and Eve, the old Greek legend of Paris and Helena, and so many other famous traditions, only the poetic expression of the vast influence that love and sexual selection have exercised over the course of history ever since the differentiation of the sexes? All the other passions that agitate the heart of man are far outstripped in their joint influence by this sense-inflaming and mind-benumbing Eros. On the one hand, we look to love with gratitude as the source of the greatest artistic achievements—the noblest creations of poetry, plastic art, and music; we see in it the chief factor in the moral advance of humanity, the foundation of family life, and therefore of social advance. On the other hand, we dread it as the devouring flame that brings destruction on so many, and has caused more misery, vice, and crime than all the other evils of human life put together. So wonderful is love and so momentous its influence on the life of the soul, or on the different functions of the medullary tube, that here more than anywhere else the “supernatural” result seems to mock any attempt at natural explanation. Yet comparative evolution leads us clearly and indubitably to the first source of love—the affinity of two different erotic cells, the sperm-cell and ovum.1
The lowest Metazoa throw light on this very simple origin of the intricate phenomena of reproduction, and they also teach us that the earliest sexual form was hermaphrodism, and that the separation of the sexes (by division of labour) is a secondary and later phenomenon. Hermaphrodism predominates in the most varied groups of the lower animals; each sexually-mature individual, each person, contains female and male sexual cells, and is therefore able to fertilise itself and reproduce. Thus we find ova and sperm-cells in the same individual, not only in the lowest Zoophytes (Gastræads, Sponges, and many Polyps), but also in many worms (leeches and earthworms), many of the snails (the common garden and vineyard snails), all the Tunicates, and many other invertebrate animals. All man’s earlier invertebrate ancestors, from the Gastræads up to the Prochordonia, were hermaphrodites; possibly even the earliest Acrania. We have an instructive proof of this in the remarkable circumstance that many genera of fishes are still hermaphrodites, and that it is occasionally found in the higher Vertebrates of all classes (as atavism). We may conclude from this that gonochorism (separation of the sexes) was a later stage in our development. At first, male and female individuals differ only in the possession of one or other kind of gonads; in other respects they were identical, as we still find in the Amphioxus and the Cyclostomes. Afterwards, accessory organs (ducts, etc.) are associated with the primary sexual glands; and much later again sexual selection has given rise to the secondary sexual characters—those differences between the sexes which do not affect the sexual organs themselves, but other parts of the body (such as the man’s beard or the woman’s breast).
The third important fact that we learn from the lower Zoophytes relates to the earliest origin of the two kinds of sexual cells. As in the Gastræads (the lowest sponges and hydroids), in which we find the first beginnings of sexual differentiation, the whole body consists merely of the two primary germinal layers, it follows that the sexual cells also must have proceeded from the cells of these primary layers, either the inner or outer, or from both. This simple fact is extremely important, because the first trace of the ova as well as the spermatozoa is found in the middle germinal layer or mesoderm in the higher animals, especially the Vertebrates. This arrangement is a later development from the preceding (in connection with the secondary formation of the mesoderm).
If we trace the phylogeny of the sexual organs in our earliest Metazoa ancestors, as the comparative anatomy and ontogeny of the lowest Cœlenteria (Cnidaria, Platodaria) exhibit it to us, we find that the first step in advance is the localisation or concentration of the two kinds of sexual
1. The sensual perception (probably related to smell) of the two copulating sex-cells, which causes their mutual attraction, is a little understood, but very interesting, chemical function of the cell-soul (cf. p. 58 and The Riddle of the Universe, chap. ix.)
[ 333 ]
cells scattered in the epithelium into definite groups. In the Sponges and lowest Hydropolyps isolated cells are detached from the cell-strata of the two primary germinal layers, and become free sexual cells; but in the Cnidaria and Platodes we find these associated in groups which we call sexual glands (gonads). We can now for the first time speak of sexual organs in the morphological sense. The female germinative glands, which in this simplest form are merely groups of homogeneous cells, are the ovaries (Fig. 241 c). The male germinative glands, which also in their first form consist of a cluster of sperm-cells, are the testicles (Fig. 241 h). In the medusæ, which descend, both ontogenetically and phylogenetically, from the more simply organised Polyps, we find these simple sexual glands sometimes as gastric pouches, sometimes as outgrowths of the radial canals that proceed from the stomach. Particularly interesting in connection with the question of the first origin of the gonads are the lowest forms of the Platodes, the Cryptocœla that have of late been separated as a special class (Platodaria) from the Turbellaria proper (Fig. 239). In these very primitive Platodes the two pairs of sexual glands are merely two pairs of rows of differentiated cells in the entodermic wall of the primitive gut—two median ovaries (o) within, and two lateral spermaries (s) without. The mature sexual cells are ejected by the posterior outlets; the female (f) lies in front of the male (m).
 Fig. 383—Embryos of Sagitta, in three earlier stages of development. (From Hertwig.) A gastrula, B cœlomula with open primitive mouth, C the same primitive mouth closed, ua primitive gut, bl primitive mouth, g progonidia (hermaphroditic primitive sexual cells), cs cœlom-pouches, pm parietal layer, vm visceral layer of same, d permanent gut (enteron), st mouth-pit (stomodæum).
Fig. 383—Embryos of Sagitta, in three earlier stages of development. (From Hertwig.) A gastrula, B cœlomula with open primitive mouth, C the same primitive mouth closed, ua primitive gut, bl primitive mouth, g progonidia (hermaphroditic primitive sexual cells), cs cœlom-pouches, pm parietal layer, vm visceral layer of same, d permanent gut (enteron), st mouth-pit (stomodæum). |
In the great majority of the Bilateria or Cœlomaria it is the mesoderm from which the gonads develop. Probably the first traces of them are the two large cells that appear at the edge of the primitive mouth (right and left), as a rule during gastrulation or immediately afterwards—the important promesoblasts, or “polar cells of the mesoderm,” or “primitive cells of the middle germinal layer” (p. 194). In the real Enterocœla, in which the mesoderm appears from the first in the shape of a couple of cœlom-pouches, these are very probably the original gonads (p. 194). This is seen very clearly in the arrow-worm (Sagitta). In the gastrula of Sagitta (Fig. 383 A) we find at an early stage a couple of entodermic cells of an unusual size (g) at the base of the primitive gut (ud). These primitive sexual cells (progonidia) are symmetrically placed to the right and left of the middle plane, like the two promesoblasts of the bilateral gastrula of the Amphioxus (Fig. 38 p). A little outwards from them the two cœlom pouches (B, cs) are developed out of the primitive gut, and each progonidion divides into a male and a female sexual cell (B, g). The two male cells (at first rather the larger) lie close together within, and are the parent-cells of the testicles (prospermaria). The two female cells lie outwards from these, and are the parent-cells of the ovary (protovaria). Afterwards, when the cœlom-pouches have detached from the permanent gut (C, d) and the primitive mouth (A, bl) is closed, the female cells advance towards the mouth (C, st), and the male towards the rear. The foremost pair of ovaries are then separated by a transverse partition from the hind pair. Thus the first structures of the sexual glands of the Sagitta are a couple of hermaphroditic entodermic cells; each of these divides
[ 334 ]
into a male and a female cell; and these four cells are the parent-cells of the four sexual glands. Probably the two promesoblasts of the Amphioxus-gastrula (Fig. 38) are also hermaphroditic primitive sexual cells in the same sense, inherited by this earliest vertebrate from its ancient bilateral gastræad ancestors.
 Fig. 384—A, Part of the kidneys of Bdellostoma. a prorenal duct (nephroductus), b segmental or primitive urinary canals (pronephridia), c renal or Malpighian capsules. B Portion of same, highly magnified. c renal capsules with the glomerulus, d afferent artery, e efferent artery. From Johannes Müller (Myxinoides).
Fig. 384—A, Part of the kidneys of Bdellostoma. a prorenal duct (nephroductus), b segmental or primitive urinary canals (pronephridia), c renal or Malpighian capsules. B Portion of same, highly magnified. c renal capsules with the glomerulus, d afferent artery, e efferent artery. From Johannes Müller (Myxinoides). |
The sexually-mature Amphioxus is not hermaphroditic, as its nearest invertebrate relatives, the Tunicates, are, and as the long-extinct pre-Silurian Primitive Vertebrate (Prospondylus, Figs. 98–102) probably was. The actual lancelet has gonochoristic structures of a very interesting kind. As we saw in the anatomy of the Amphioxus, we find the ovaries of the female and the spermaries of the male in the shape of twenty to thirty pairs of elliptical or roundish four-cornered sacs, which lie on either side of the gut on the parietal surface of the respiratory pore (Fig. 219 g). According to the important discovery of Rückert (1888), the sexual glands of the earliest fishes, the Selachii, are similarly arranged. They only unite afterwards to form a pair of simple gonads. These have been transmitted by heredity to all the rest of the Craniotes. In every case they lie originally on each side of the mesentery, underneath the chorda, at the bottom of the body-cavity. The first traces of them are found in the cœlom-epithelium, at the spot where the skin-fibre layer and gut-fibre layer meet in the middle of the mesenteric plate (Fig. 93 mp). At this point we observe at an early stage in all craniote embryos a small string-like cluster of cells, which we may call, with Waldeyer, the “germ epithelium,” or (in harmony with the other plate-shaped rudimentary organs) the sexual plate (Fig. 173 g). This germinal or sexual plate is found in the fifth week in the human embryo, in the shape of a couple of long whitish streaks, on the inner side of the primitive kidneys (Fig. 183 t). The cells of this sexual plate are distinguished by their cylindrical form and chemical composition from the rest of the cœlom-cells; they have a different purport from the flat cells which line the rest of the body-cavity. As the germ epithelium of the sexual plate becomes thicker, and supporting tissue grows into it from the mesoderm, it becomes a rudimentary sexual gland. This ventral gonad then develops into the ovary in the female Craniotes, and the testicles in the male.
In the formation of the gonidia or erotic sexual cells and their conjunction at fecundation we have the sole essential features of sexual reproduction; but in the great majority of animals we find other organs taking part in it. The chief of these secondary sexual organs are the gonoducts, which serve to convey the mature sexual cells out of the body, and the copulative organs, which bring the fecundating male sperm into touch with the ovum-bearing female. The latter organs are, as a rule, only found in the higher animals, and are much less widely distributed than the gonoducts. But these also are secondary formations, and are wanting in many animals of the lower groups.
In the lower animals the mature sexual cells are generally ejected directly from
[ 335 ]
the body. Sometimes they pass out immediately through the skin (Hydra and many hydroids); sometimes they fall into the gastric cavity, and are evacuated by the mouth (gastræads, sponges, many medusæ, and corals); sometimes they fall into the body-cavity, and are ejected by a special pore (porus genitalis) in the ventral wall. The latter procedure is found in many of the worms, and also in the lowest Vertebrates. Amphioxus has the peculiar feature that the mature sexual products fall first into the mantle-cavity; from there they are either evacuated by the respiratory pore, or else they pass through the gill-clefts into the branchial gut, and so out by the mouth (p. 185). In the Cyclostomes they fall into the body-cavity, and are ejected by a genital pore in its wall; so also in some of the fishes. From these we gather the features of our earlier ancestors in this respect. On the other hand, in all the higher and most of the lower Vertebrates (and most of the higher Invertebrates) we find in both sexes special tubular passages of the sexual gland, which are called “gonoducts.” In the female they conduct the ova from the ovary, and so are called “oviducts,” or “Fallopian tubes.” In the male they convey the spermatozoa from the testicles, and are called “spermaducts,” or vasa deferentia.
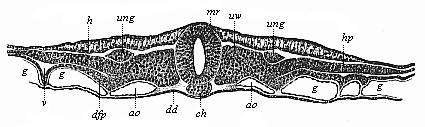 Fig. 385—Transverse section of the embryonic shield of a chick, forty-two hours old. (From Kölliker.) mr medullary tube, ch chorda, h horny plate (skin-sense layer), ung nephroduct, vw episomites (dorsal primitive segments), hp skin-fibre layer (parietal layer of the hyposomites), dfp gut-fibre layer (visceral layer of hyposomites), ao aorta, g vessels. (Cf. transverse section of duck-embryo, Fig. 152.)
Fig. 385—Transverse section of the embryonic shield of a chick, forty-two hours old. (From Kölliker.) mr medullary tube, ch chorda, h horny plate (skin-sense layer), ung nephroduct, vw episomites (dorsal primitive segments), hp skin-fibre layer (parietal layer of the hyposomites), dfp gut-fibre layer (visceral layer of hyposomites), ao aorta, g vessels. (Cf. transverse section of duck-embryo, Fig. 152.) |
The original and genetic relation of these two kinds of ducts is just the same in man as in the rest of the higher Vertebrates, and quite different from what we find in most of the Invertebrates. In the latter, as a rule, the gonoducts develop directly from the embryonic glands or from the outer skin; but in the Vertebrates an independent organic system is employed to convey the sexual products, and this had originally a totally different function—namely, the system of urinary organs. These organs have primarily the sole duty of removing unusable matter from the body in a fluid form. Their liquid excretory product, the urine, is either evacuated directly through the skin or through the last section of the gut. It is only at a later stage that the tubular urinary passages also convey the sexual products from the body. In this way they become “urogenital ducts.” This remarkable secondary conjunction of the urinary and sexual organs into a common urogenital system is very characteristic of the Gnathostomes, the six higher classes of Vertebrates. It is wanting in the lower classes. In order to appreciate it fully, we must give a comparative glance at the structure of the urinary organs.
The renal or urinary system is one of the oldest and most important systems of organs in the differentiated animal body, as I have pointed out on several previous occasions (cf. Chapter XVII). We find it not only in the higher stems, but also very generally distributed in the earlier group of the Vermalia. Here we meet it in the lowest worms, the Rotatoria (Gastrotricha, Fig. 242), and in the instructive stem of the Platodes. It consists of a pair of simple or branching canals, which are lined with one layer of cells, absorb unusable juices from the tissue, and eject them by an outlet in the outer skin (Fig. 240 nm). Not only the free-living Turbellaria, but also the parasitic Suctoria, and even the still more degenerate tapeworms, which have lost their alimentary canal in consequence of their parasitic life, are equipped with these renal canals
[ 336 ]
or nephridia. In the first embryonic structure they are merely a pair of simple cutaneous glands, or depressions in the ectoderm. They are generally described as excretory organs in the worms, but formerly often as “water vessels.” They may be conceived as largely-developed tubular cutaneous glands, formed by invagination of the cutaneous layer. According to another view, they owe their origin to a later rupture of the body-cavity outwards. In most of the Vermalia each nephridium has an inner opening (with cilia) into the body-cavity and an outer one on the epidermis.

Fig. 386—Rudimentary primitive kidneys of a dog-embryo. The hind end of the embryonic body is seen from the ventral side and covered with the visceral layer of the yelk-sac, which is torn away and folded down in front in order to show the nephroducts with the primitive urinary canals (a). b primitive vertebræ, c spinal cord, d entrance into the pelvic-gut cavity. (From Bischoff.) | |

Fig. 387—Primitive kidneys of a human embryo. u the urinary canals of the primitive kidneys, w Wolffian duct, w′ uppermost end of the same (Morgagni’s hydatid), m Mullerian duct. m′ uppermost end of same (Fallopian hydatid), g gonad (sexual gland). (From Kobelt.) |
In these lowest, unsegmented worms, and in the unsegmented Molluscs, there is only one pair of renal canals. They are more numerous in the higher Articulates. In the Annelids, the body of which is composed of a large number of joints, there is a pair of these pronephridia in each segment (hence they are called segmental canals or organs). Even here they are still simple tubes; on account of their coiled or looped form they are often called “looped canals.” In most of the Annelids, and many of the Vermalia, we can distinguish three sections in the nephridium—an outer muscular duct, a glandular middle part, and an inner part that opens by a ciliated funnel into the body-cavity. This opening is furnished with whirling cilia, and can, therefore, take up the juices to be excreted directly from the body-cavity and convey them from the body. But in these worms the sexual cells, which develop in very primitive form on the inner surface of the body-cavity, also fall into it when mature, and are sucked up by the funnel-shaped inner ciliated openings of the renal canals, and ejected with the urine. Thus the urine-forming looped canals, or pronephridia, serve as oviducts in the female Annelids and as spermaducts in the male.
The renal system of the Vertebrates is similar to, yet materially different from, these segmental canals of the Annelids. The peculiar development of it and its relations to the sexual organs are among the most difficult problems in the morphology of our stem. If we examine briefly the vertebrate renal system from the phylogenetic point of view, as confirmed by recent discoveries, we may distinguish three forms of it: (1) Fore-kidneys or head-kidneys (pronephros); (2) primitive or middle kidneys (mesonephros); (3) permanent kidneys (metanephros). These three systems of kidneys are not fundamentally and completely distinct, as earlier students (such as Semper) wrongly supposed; they represent three different generations of one and the same excretory apparatus; they correspond to three phylogenetic stages,
[ 337 ]
and succeed each other in the stem-history of the Vertebrates in such wise that each younger and more advanced generation develops farther behind in the body, and replaces the older and less advanced generation that preceded it in time and space. The fore kidneys, first accurately described by Wilhelm Müller in 1875 in the Cyclostomes and Ichthyoda, form the sole excretory organ of the Acrania (Amphioxus); they continue in the Cyclostomes and some of the fishes, but are found only in slight traces and for a time in the embryos of the six other classes of Vertebrates. The primitive kidneys are first found in the Cyclostomes, behind the fore kidneys; they have been transmitted from the Selachii to all the Gnathostomes. In the Anamnia they act permanently as urinary glands; in the Amniotes their anterior part (“germinal kidneys”) changes into organs of the sexual apparatus, while the third generation develops from the end of their posterior part (“urinal kidneys”)—the characteristic after or permanent kidneys of the three higher classes of Vertebrates. The order in which the three renal systems succeed each other in the embryo of man and the higher Vertebrates corresponds to their phylogenetic succession in the history of our stem, and, consequently, in the natural classification of the Vertebrates.
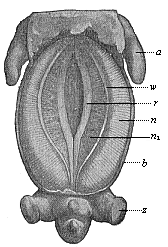
Fig. 388—Pig-embryo, three-fifths of an inch long, seen from the ventral side. a fore leg, z hind leg, b ventral wall, r sexual prominence, w nephroduct, n primitive kidneys, n1 their inner part. (From Oscar Schultze.) |
|
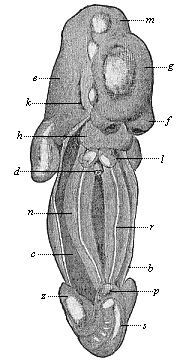
Fig. 389—Human embryo of the fifth week, two-fifths of an inch long, seen from the ventral side (the anterior ventral wall, b, is removed, the body-cavity, c, opened). d gut (cut off), f frontal process, g cerebrum, m middle brain, e after brain, h heart, k first gill-cleft, l pulmonary sac, n primitive kidneys, r sexual region, p phallus (sexual prominences), s tail. (From Kollmann.) |
As in the morphology of any other system of organs, so in the case of the urinary and sexual organs the Amphioxus is the real typical primitive Vertebrate; it affords the key to the mysteries of the structure of man and the higher Vertebrates. The kidneys of the Amphioxus—first discovered by Boveri in 1890—are typical “fore kidneys,” composed of a double row of short segmental canals (Fig. 217 x). The inner aperture of these pronephridia opens into the mesodermic body-cavity (the middle part of the cœloma, B); the external aperture into the ectodermic mantle or peribranchial cavity (C). Their position, their
[ 338 ]
structure, and their relation to the branchial vessel make it clear that these segmental pronephridia correspond to the rudimentary fore kidneys of the Craniotes. The mantle-cavity into which they open seems to correspond to the prorenal duct of the latter.

| Figs. 390, 391, 392—Primitive kidneys and rudimentary sexual organs. Figs. 390 and 391 of Amphibia (frog-larvæ); Fig. 390 earlier, 391 later stage. Fig. 392 of a mammal (ox-embryo). u primitive kidney, k sexual gland (rudiment of testicle and ovary). The primary nephroduct (ug in Fig. 390) divides (in Figs. 391 and 392) into the two secondary nephroducts—the Mullerian (m) and Wolffian (ug′) ducts, joined together behind in the genital cord (g). l ligament of the primitive kidneys. (From Gegenbaur.) |
 Figs. 393, 394—Urinary and sexual organs of an Amphibian (water salamander or Triton). Fig. 393 of a female, 394 of a male. r primitive kidney, ov ovary, od oviduct and c Rathke’s duct, both developed from the Müllerian duct, u primitive ureter (also acting as spermaduct [ve] in the male, opening below into the Wolffian duct [u apostrophe]), ms mesovarium. (From Gegenbaur.)
Figs. 393, 394—Urinary and sexual organs of an Amphibian (water salamander or Triton). Fig. 393 of a female, 394 of a male. r primitive kidney, ov ovary, od oviduct and c Rathke’s duct, both developed from the Müllerian duct, u primitive ureter (also acting as spermaduct [ve] in the male, opening below into the Wolffian duct [u apostrophe]), ms mesovarium. (From Gegenbaur.) |
The next higher Vertebrates, the Cyclostomes, yield some very interesting data. Both orders of this class, the hags and lampreys, have still the fore kidneys inherited from the Acrania—the former permanently, the latter in their earlier stages. Behind these the primitive kidneys soon develop, and in a very characteristic form. The remarkable structure of the mesonephros of the Cyclostomes, discovered by Johannes Müller, explains the intricate formation of the kidneys in the higher Vertebrates. We find in the hag-fishes (Bdellostoma) a long tube, the prorenal duct (nephroductus, Fig. 384 a). This opens with its anterior end into the cœloma by a ciliated aperture, and externally with its posterior end by an outlet in the skin. Inside it open a large number of small transverse canals (“segmental or primitive urinary canals,” b). Each of these terminates blindly in a vesicular capsule (c), and this encloses a coil of blood-vessel (glomerulus, an arterial network, Fig. 384 B, c). Afferent branches of arteries conduct arterial blood into the coiled branches of the glomerulus (d), and efferent arterial branches conduct it away from the net (c). The primitive renal canals (mesonephridia) are distinguished by this net-formation from their predecessors.
In the Selachii also we find a longitudinal row of segmental canals on each side, which open outwards into the primitive renal ducts (nephrotomes, p. 149. The segmental canals (a pair in each segment of the middle part of the body) open internally by a ciliated funnel into the body-cavity. From the posterior group of these organs a compact primitive kidney is formed, the anterior group taking part in the construction of the sexual organs.
In the same simple form that remains
[ 339 ]
throughout life in the Myxinoides and partly in the Selachii we find the primitive kidney first developing in the embryo of man and the higher Craniotes (Figs. 386, 387). Of the two parts that compose the comb-shaped primitive kidney the longitudinal channel, or nephroduct, is always the first to appear; afterwards the transverse “canals,” the excreting nephridia, are formed in the mesoderm; and after this again the Malpighian capsules with their arterial coils are associated with these as cœlous outgrowths. The primitive renal duct, which appears first, is found in all craniote embryos at the early stage in which the differentiation of the medullary tube takes place in the ectoderm, the severance of the chorda from the visceral layer in the entoderm, and the first trace of the cœlom-pouches arises between the limiting layers (Fig. 385). The nephroduct (ung) is seen on each side, directly under the horny plate, in the shape of a long, thin, thread-like string of cells. It presently hollows out and becomes a canal, running straight from front to back, and clearly showing in the transverse section of the embryo its original position in the space between horny plate (h), primitive segments (uw), and lateral plates (hpl). As the originally very short urinary canals lengthen and multiply, each of the two primitive kidneys assumes the form of a half-feathered leaf (Fig. 387). The lines of the leaf are represented by the urinary canals (u), and the rib by the outlying nephroduct (w). At the inner edge of the primitive kidneys the rudiment of the ventral sexual gland (g) can now be seen as a body of some size. The hindermost end of the nephroduct opens right behind into the last section of the rectum, thus making a cloaca of it. However, this opening of the nephroducts into the intestine must be regarded as a secondary formation. Originally they open, as the Cyclostomes clearly show, quite independently of the gut, in the external skin of the abdomen.
 Fig. 395—Primitive kidneys and germinal glands of a human embryo, three inches in length (beginning of the sixth week), magnified. k germinal gland, u primitive kidney, z diaphragmatic ligament of same, w Wolffian duct (opened on the right), g directing ligament (gubernaculum), a allantoic duct. (From Kollmann.)
Fig. 395—Primitive kidneys and germinal glands of a human embryo, three inches in length (beginning of the sixth week), magnified. k germinal gland, u primitive kidney, z diaphragmatic ligament of same, w Wolffian duct (opened on the right), g directing ligament (gubernaculum), a allantoic duct. (From Kollmann.) |
In the Myxinoides the primitive kidneys retain this simple comb-shaped structure, and a part of it is preserved in the Selachii; but in all the other Craniotes it is only found for a short time in the embryo, as an ontogenetic reproduction of the earlier phylogenetic structure. In these the primitive kidney soon assumes the form (by the rapid growth, lengthening, increase, and serpentining of the urinary canals) of a large compact gland, of a long, oval or spindle-shaped character, which passes through the greater part of the embryonic body-cavity (Figs. 183 m, 184 m, 388 n). It lies near the middle line, directly under the primitive vertebral column, and reaches from the cardiac region to the cloaca. The right and left kidneys are parallel to each other, quite close together, and only separated by the mesentery—the thin narrow layer that attaches the middle gut to the under surface of the vertebral column. The passage of each primitive kidney, the nephroduct, runs towards the back on the lower and outer side of the gland, and opens in the cloaca, close to the starting-point of the allantois; it afterwards opens into the allantois itself.
The primitive or primordial kidneys of the amniote embryo were formerly called the “Wolffian bodies,” and sometimes “Oken’s bodies.” They act for a time as
[ 340 ]
kidneys, absorbing unusable juices from the embryonic body and conducting them to the cloaca—afterwards to the allantois. There the primitive urine accumulates, and thus the allantois acts as bladder or urinary sac in the embryos of man and the other Amniotes. It has, however, no genetic connection with the primitive kidneys, but is a pouch-like growth from the anterior wall of the rectum (Fig. 147 u). Thus it is a product of the visceral layer, whereas the primitive kidneys are a product of the middle layer. Phylogenetically we must suppose that the allantois originated as a pouch-like growth from the cloaca-wall in consequence of the expansion caused by the urine accumulated in it and excreted by the kidneys. It is originally a blind sac of the rectum. The real bladder of the vertebrate certainly made its first appearance among the Dipneusts (in Lepidosiren), and has been transmitted from them to the Amphibia, and from these to the Amniotes. In the embryo of the latter it protrudes far out of the not yet closed ventral wall. It is true that many of the fishes also have a “bladder.” But this is merely a local enlargement of the lower section of the nephroducts, and so totally different in origin and composition from the real bladder. The two structures can be compared from the physiological point of view, and so are analogous, as they have the same function; but not from the morphological point of view, and are therefore not homologous. The false bladder of the fishes is a mesodermic product of the nephroducts; the true bladder of the Dipneusts, Amphibia, and Amniotes is an entodermic blind sac of the rectum.
 Figs. 396–398—Urinary and sexual organs of ox-embryos. Fig. 396, female embryo one and a half inches long; Fig. 397, male embryo, one and a half inches long. Fig. 398 female embryo two and a half inches long. w primitive kidney, wg Wolffian duct, m Müllerian duct, m′ upper end of same (opened at t), i lower and thicker part of same (rudiment of uterus), g genital cord, h testicle, (h′, lower and h″, upper testicular ligament), o ovary, o′ lower ovarian ligament, i inguinal ligament of primitive kidney, d diaphragmatic ligament of primitive kidney, nn accessory kidneys, n permanent kidneys, under them the S-shaped ureters, between these the rectum, v bladder, a umbilical artery. (From Kölliker.)
Figs. 396–398—Urinary and sexual organs of ox-embryos. Fig. 396, female embryo one and a half inches long; Fig. 397, male embryo, one and a half inches long. Fig. 398 female embryo two and a half inches long. w primitive kidney, wg Wolffian duct, m Müllerian duct, m′ upper end of same (opened at t), i lower and thicker part of same (rudiment of uterus), g genital cord, h testicle, (h′, lower and h″, upper testicular ligament), o ovary, o′ lower ovarian ligament, i inguinal ligament of primitive kidney, d diaphragmatic ligament of primitive kidney, nn accessory kidneys, n permanent kidneys, under them the S-shaped ureters, between these the rectum, v bladder, a umbilical artery. (From Kölliker.) |
In all the Anamnia (the lower amnionless Craniotes, Cyclostomes, Fishes, Dipneusts, and Amphibia) the urinary organs remain at a lower stage of development to this extent, that the primitive kidneys (protonephri) act permanently as urinary glands. This is only so as a passing phase of the early embryonic life in the three higher classes of Vertebrates, the Amniotes. In these the permanent or after or secondary (really tertiary) kidneys (renes or metanephri) that are distinctive of these three classes soon make their appearance. They represent the third and last generation of the vertebrate kidneys. The permanent kidneys do not arise (as was long supposed) as independent glands from the alimentary tube, but from the last section of the primitive kidneys and the nephroduct. Here a simple tube, the secondary renal duct, develops, near the point of its entry into the cloaca; and this tube grows considerably forward. With its blind upper or anterior end is connected a glandular renal growth, that owes its origin to a differentiation of the last part of the primitive kidneys. This rudiment of the
[ 341 ]
permanent kidneys consists of coiled urinary canals with Malpighian capsules and vascular coils (without ciliated funnels), of the same structure as the segmental mesonephridia of the primitive kidneys. The further growth of these metanephridia gives rise to the compact permanent kidneys, which have the familiar bean-shape in man and most of the higher mammals, but consist of a number of separate folds in the lower mammals, birds, and reptiles. As the permanent kidneys grow rapidly and advance forward, their passage, the ureter, detaches altogether from its birth-place, the posterior end of the nephroduct; it passes to the posterior surface of the allantois. At first in the oldest Amniotes this ureter opens into the cloaca together with the last section of the nephroduct, but afterwards separately from this, and finally into the permanent bladder apart from the rectum altogether. The bladder originates from the hindmost and lowest part of the allantoic pedicle (urachus), which enlarges in spindle shape before the entry into the cloaca. The anterior or upper part of the pedicle, which runs to the navel in the ventral wall of the embryo, atrophies subsequently, and only a useless string-like relic of it is left as a rudimentary organ; that is the single vesico-umbilical ligament. To the right and left of it in the adult male are a couple of other rudimentary organs, the lateral vesico-umbilical ligaments. These are the degenerate string-like relics of the earlier umbilical arteries.
Though in man and all the other Amniotes the primitive kidneys are thus early replaced by the permanent kidneys, and these alone then act as urinary organs, all the parts of the former are by no means lost. The nephroducts become very important physiologically by being converted into the passages of the sexual glands. In all the Gnathostomes—or all the Vertebrates from the fishes up to man—a second similar canal develops beside the nephroduct at an early stage of embryonic evolution. The latter is usually called the Müllerian duct, after its discoverer, Johannes Müller, while the former is called the Wolffian duct. The origin of the Müllerian duct is still obscure; comparative anatomy and ontogeny seem to indicate that it originates by differentiation from the Wolffian duct. Perhaps it would be best to say: “The original primary nephroduct divides by differentiation (or longitudinal cleavage) into two secondary nephroducts, the Wolffian and the Müllerian ducts.” The latter (Fig. 387 m) lies just on the inner side of the former (Fig. 387 w). Both open behind into the cloaca.
 Fig. 399—Female sexual organs of a Monotreme (Ornithorhynchus, Fig. 269). o ovaries, t oviducts, u womb, sug urogenital sinus; at u′ is the outlet of the two wombs, and between them the bladder (vu). cl cloaca. (From Gegenbaur.)
Fig. 399—Female sexual organs of a Monotreme (Ornithorhynchus, Fig. 269). o ovaries, t oviducts, u womb, sug urogenital sinus; at u′ is the outlet of the two wombs, and between them the bladder (vu). cl cloaca. (From Gegenbaur.) |
However uncertain the origin of the nephroduct and its two products, the Müllerian and the Wolffian ducts, may be, its later development is clear enough. In all the Gnathostomes the Wolffian duct is converted into the spermaduct, and the Müllerian duct into the oviduct. Only one of them is retained in each sex; the other either disappears altogether, or only leaves relics in the shape of rudimentary organs. In the male sex, in which the two Wolffian ducts become the spermaducts, we often find traces of the Müllerian ducts, which I have called “Rathke’s canals” (Fig. 394 c). In the female sex, in which the two Müllerian ducts form the oviducts, there are relics of the Wolffian ducts, which are called “the ducts of Gaertner.”
We obtain the most interesting information with regard to this remarkable evolution of the nephroducts and their association with the sexual glands from the Amphibia (Figs. 390–395). The first structure of the nephroduct and its differentiation into Müllerian and Wolffian ducts are just the same in both sexes in the Amphibia, as in the mammal embryos (Figs. 392, 396). In the female Amphibia
[ 342 ]
the Müllerian duct develops on either side into a large oviduct (Fig. 393 od), while the Wolffian duct acts permanently as ureter (u). In the male Amphibia the Müllerian duct only remains as a rudimentary organ without any functional significance, as Rathke’s canal (Fig. 394 c); the Wolffian duct serves also as ureter, but at the same time as spermaduct, the sperm-canals (ve) that proceed from the testicles (t) entering the fore part of the primitive kidneys and combining there with the urinary canals.
 |
Figs. 400, 401—Original position of the sexual glands in the ventral cavity of the human embryo (three months old). Fig. 400, male. h testicles, gh conducting ligament of the testicles, wg spermaduct, h bladder, uh inferior vena cava, nn accessory kidneys, n kidneys. Fig. 401, female. r round maternal ligament (underneath it the bladder, over it the ovaries). r′ kidneys, s accessory kidneys, c cæcum, o small reticle, om large reticle (stomach between the two), l spleen. (From Kölliker.) |
In the mammals these permanent amphibian features are only seen as brief phases of the earlier period of embryonic development (Fig. 392). Here the primitive kidneys, which act as excretory organs of urine throughout life in the amnion-less Vertebrates, are replaced in the mammals by the permanent kidneys. The real primitive kidneys disappear for the most part at an early stage of development, and only small relics of them remain. In the male mammal the epididymis develops from the uppermost part of the primitive kidney; in the female a useless rudimentary organ, the epovarium, is formed from the same part. The atrophied relic of the former is known as the paradidymis, that of the latter as the parovarium.
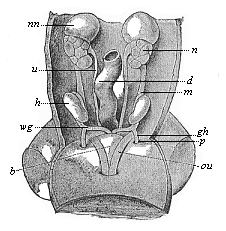 Fig. 402—Urogenital system of a human embryo of three inches in length. h testicles, wg spermaducts, gh conducting ligament, p processus vaginalis, b bladder, au umbilical arteries, m mesorchium, d intestine, u ureter, n kidney, nn accessory kidney. (From Kollman.)
Fig. 402—Urogenital system of a human embryo of three inches in length. h testicles, wg spermaducts, gh conducting ligament, p processus vaginalis, b bladder, au umbilical arteries, m mesorchium, d intestine, u ureter, n kidney, nn accessory kidney. (From Kollman.) |
The Müllerian ducts undergo very important changes in the female mammal. The oviducts proper are developed only from their upper part; the lower part dilates into a spindle-shaped tube with thick muscular wall, in which the impregnated ovum develops into the embryo. This is the womb (uterus). At first the two wombs (Fig. 399 u) are completely separate, and open into the cloaca on either side of the bladder (vu), as is still the case in the lowest living mammals, the Monotremes. But in the Marsupials a communication is opened between the two Müllerian ducts, and in the Placentals they combine below with the rudimentary Wolffian ducts to form a single “genital cord.” The original independence of the two wombs and the vaginal canals formed from their lower ends are retained in many of the lower Placentals, but in the higher they gradually blend and form a single organ. The conjunction proceeds from below (or behind) upwards (or forwards). In many of the Rodents (such as the rabbit and squirrel) two separate wombs still open into the simple and single vaginal canal; but in others, and in the Carnivora, Cetacea, and Ungulates, the lower halves of the wombs have already fused into a single piece, though the upper halves (or “horns”) are still separate (“two-horned” womb, uteris bicornis). In the bats and lemurs the “horns” are
[ 343 ]
very short, and the lower common part is longer. Finally, in the apes and in man the blending of the two halves is complete, and there is only the one simple, pear-shaped uterine pouch, into which the oviducts open on each side. This simple uterus is a late evolutionary product, and is found only in the ape and man.
 Figs. 403–406—Origin of human ova in the female ovary. Fig. 403. Vertical section of the ovary of a new-born female infant, a ovarian epithelium, b rudimentary string of ova, c young ova in the epithelium, d long string of ova with follicle-formation (Pflüger’s tube), e group of young follicles, f isolated young follicle, g blood-vessels in connective tissue (stroma) of the ovary. In the strings the young ova are distinguished by their considerable size from the surrounding follicle-cells. (From Waldeyer.)
Figs. 403–406—Origin of human ova in the female ovary. Fig. 403. Vertical section of the ovary of a new-born female infant, a ovarian epithelium, b rudimentary string of ova, c young ova in the epithelium, d long string of ova with follicle-formation (Pflüger’s tube), e group of young follicles, f isolated young follicle, g blood-vessels in connective tissue (stroma) of the ovary. In the strings the young ova are distinguished by their considerable size from the surrounding follicle-cells. (From Waldeyer.)
Fig. 404—Two young Graafian follicles, isolated. In 1 the follicle-cells still form a simple, and in 2 a double, stratum round the young ovum; in 2 they are beginning to form the ovolemma or the zona pellucida (a).
Figs. 405 and 406—Two older Graafian follicles, in which fluid is beginning to accumulate inside the eccentrically thickened epithelial mass of the follicle-cells (Fig. 405 with little, 406 with much, follicle-water). ei the young ovum, with embryonic vesicle and spot, zp ovolemma or zona pellucida, dp discus proligerus, formed of an accumulation of follicle-cells, which surround the ovum, ff follicle-liquid (liquor folliculi), gathered inside the stratified follicle-epithelium (fe), fk connective-tissue fibrous capsule of the Graafian follicle (theca folliculi). |
In the male mammals there is the same fusion of the Müllerian and Wolffian ducts at their lower ends. Here again they form a single genital cord (Fig. 397 g), and this opens similarly into the
[ 344 ]
original urogenital sinus, which develops from the lowest section of the bladder (v). But while in the male mammal the Wolffian ducts develop into the permanent spermaducts, there are only rudimentary relics left of the Müllerian ducts. The most notable of these is the “male womb” (uterus masculinus), which originates from the lowest fused part of the ducts, and corresponds to the female uterus. It is a small, flask-shaped vesicle without any physiological significance, which opens into the ureter between the two spermaducts and the prostate folds (vesicula prostatica).
 |
Fig. 407—A ripe human Graafian follicle. a the mature ovum, b the surrounding follicle-cells, c the epithelial cells of the follicle, d the fibrous membrane of the follicle, e its outer surface. |
The internal sexual organs of the mammals undergo very distinctive changes of position. At first the germinal glands of both sexes lie deep inside the ventral cavity, at the inner edge of the primitive kidneys (Figs. 386 g, 392 k), attached to the vertebral column by a short mesentery (mesorchium in the male, mesovarium in the female). But this primary arrangement is retained permanently only in the Monotremes (and the lower Vertebrates). In all other mammals (both Marsupials and Placentals) they leave their original cradle and travel more or less far down (or behind), following the direction of a ligament that goes from the primitive kidneys to the inguinal region of the ventral wall. This is the inguinal ligament of the primitive kidneys, known in the male as the Hunterian ligament (Fig. 400 gh), and in the female as the “round maternal ligament” (Fig. 401 r). In woman the ovaries travel more or less towards the small pelvis, or enter into it altogether. In the male the testicles pass out of the ventral cavity, and penetrate by the inguinal canal into a sac-shaped fold of the outer skin. When the right and left folds (“sexual swellings”) join together they form the scrotum. The various mammals bring before us the successive stages of this displacement. In the elephant and the whale the testicles descend very little, and remain underneath the kidneys. In many of the rodents and carnassia they enter the inguinal canal. In most of the higher mammals they pass through this into the scrotum. As a rule, the inguinal canal closes up. When it remains open the testicles may periodically pass into the scrotum, and withdraw into the ventral cavity again in time of rut (as in many of the marsupials, rodents, bats, etc.).
The structure of the external sexual organs, the copulative organs that convey the fecundating sperm from the male to the female organism in the act of copulation, is also peculiar to the mammals. There are no organs of this character in most of the other Vertebrates. In those that live in water (such as the Acrania and Cyclostomes, and most of the fishes) the ova and sperm-cells are simply ejected into the water, where their conjunction and fertilisation are left to chance. But in many of the fishes and amphibia, which are viviparous, there is a direct conveyance of the male sperm into the female body; and this is the case with all the Amniotes (reptiles, birds, and mammals). In these the urinary and sexual organs always open originally into the last section of the rectum, which thus forms a cloaca
[ 345 ]
(p. 249). Among the mammals this arrangement is permanent only in the Monotremes, which take their name from it (Fig. 399 cl). In all the other mammals a frontal partition is developed in the cloaca (in the human embryo about the beginning of the third month), and this divides it into two cavities. The anterior cavity receives the urogenital canal, and is the sole outlet of the urine and the sexual products; the hind or anus-cavity passes the excrements only.
Even before this partition has been formed in the Marsupials and Placentals, we see the first trace of the external sexual organs. First a conical protuberance rises at the anterior border of the cloaca-outlet—the sexual prominence (phallus, Fig. 402 A, e, B, e). At the tip it is swollen in the shape of a club (“acorn” glans). On its under side there is a furrow, the sexual groove (sulcus genitalis, f), and on each side of this a fold of skin, the “sexual pad” (torus genitalis, h l). The sexual protuberance or phallus is the chief organ of the sexual sense (p. 282); the sexual nerves spread on it, and these are the principal organs of the specific sexual sensation. As erectile bodies (corpora cavernosa) are developed in the male phallus by peculiar modifications of the blood-vessels, it becomes capable of erecting periodically on a strong accession of blood, becoming stiff, so as to penetrate into the female vagina and thus effect copulation. In the male the phallus becomes the penis; in the female it becomes the much smaller clitoris; this is only found to be very large in certain apes (Ateles). A prepuce (“foreskin”) is developed in both sexes as a protecting fold on the anterior surface of the phallus.
 Fig. 408—The human ovum after issuing from the Graafian follicle, surrounded by the clinging cells of the discus proligerus (in two radiating crowns). z ovolemma (zona pellucida, with radial porous canals), p cytosoma (protoplasm of the cell-body, darker within, lighter without), k nucleus of the ovum (embryonic vesicle). (From Nagel.) (Cf. Figs. 1 and 14.)
Fig. 408—The human ovum after issuing from the Graafian follicle, surrounded by the clinging cells of the discus proligerus (in two radiating crowns). z ovolemma (zona pellucida, with radial porous canals), p cytosoma (protoplasm of the cell-body, darker within, lighter without), k nucleus of the ovum (embryonic vesicle). (From Nagel.) (Cf. Figs. 1 and 14.) |
The external sexual member (phallus) is found at various stages of development within the mammal class, both in regard to size and shape, and the differentiation and structure of its various parts; this applies especially to the terminal part of the phallus, the glans, both the larger glans penis of the male and the smaller glans clitoridis of the female. The part of the cloaca from the upper wall of which it forms belongs to the proctodæum, the ectodermic invagination of the rectum (p. 311); hence its epithelial covering can develop the same horny growths as the corneous layer of the epidermis. Thus the glans, which is quite smooth in man and the higher apes, is covered with spines in many of the lower apes and in the cat, and in many of the rodents with hairs (marmot) or scales (guinea-pig) or solid horny warts (beaver). Many of the Ungulates have a free conical projection on the glans, and in many of the Ruminants this “phallus-tentacle” grows into a long cone, bent hook-wise at the base (as in the goat, antelope, gazelle, etc.). The different forms of the phallus are connected with variations in the structure and distribution of the sensory corpuscles—i.e. the real organs of the sexual sense, which develop in certain papillæ of the corium of the phallus, and have been evolved from ordinary tactile corpuscles of the corium by erotic adaptation (p. 282).
[ 346 ]
The formation of the corpora cavernosa, which cause the stiffness of the phallus and its capability of penetrating the vagina, by certain special structures of their spongy vascular spaces, also shows a good deal of variety within the vertebrate stem. This stiffness is increased in many orders of mammals (especially the carnassia and rodents) by the ossification of a part of the fibrous body (corpus fibrosum). This penis-bone (os priapi) is very large in the badger and dog, and bent like a hook in the marten; it is also very large in some of the lower apes, and protrudes far out into the glans. It is wanting in most of the anthropoid apes; it seems to have been lost in their case (and in man) by atrophy.
The sexual groove on the under side of the phallus receives in the male the mouth of the urogenital canal, and is changed into a continuation of this, becoming a closed canal by the juncture of its parallel edges, the male urethra. In the female this only takes place in a few cases (some of the lemurs, rodents, and moles); as a rule, the groove remains open, and the borders of this “vestibule of the vagina” develop into the smaller labia (nymphæ). The large labia of the female develop from the sexual pads (tori genitales), the two parallel folds of the skin that are found on each side of the genital groove. They join together in the male, and form the closed scrotum. These striking differences between the two sexes cannot yet be detected in the human embryo of the ninth week. We begin to trace them in the tenth week of development, and they are accentuated in proportion as the difference of the sexes develops.
Sometimes the normal juncture of the two sexual pads in the male fails to take place, and the sexual groove may also remain open (hypospadia). In these cases the external male genitals resemble the female, and they are often wrongly regarded as cases of hermaphrodism. Other malformations of various kinds are not infrequently found in the human external sexual organs, and some of them have a great morphological interest. The reverse of hypospadia, in which the penis is split open below, is seen in epispadia, in which the urethra is open above. In this case the urogenital canal opens above at the dorsal root of the penis; in the former case down below. These and similar obstructions interfere with a man’s generative power, and thus prejudicially affect his whole development. They clearly prove that our history is not guided by a “kind Providence,” but left to the play of blind chance.
We must carefully distinguish the rarer cases of real hermaphrodism from the preceding. This is only found when the essential organs of reproduction, the genital glands of both kinds, are united in one individual. In these cases either an ovary is developed on the right and a testicle on the left (or vice versa); or else there are testicles and ovaries on both sides, some more and others less developed. As hermaphrodism was probably the original arrangement in all the Vertebrates, and the division of the sexes only followed by later differentiation of this, these curious cases offer no theoretical difficulty. But they are rarely found in man and the higher mammals. On the other hand, we constantly find the original hermaphrodism in some of the lower Vertebrates, such as the Myxinoides, many fishes of the perch-type (serranus), and some of the Amphibia (ringed snake, toad). In these cases the male often has a rudimentary ovary at the fore end of the testicle; and the female sometimes has a rudimentary, inactive testicle. In the carp also and some other fishes this is found occasionally. We have already seen how traces of the earlier hemaphrodism can be traced in the passages of the Amphibia.
Man has faithfully preserved the main features of his stem-history in the ontogeny of his urinary and sexual organs. We can follow their development step by step in the human embryo in the same advancing gradation that is presented to us by the comparison of the urogenital organs in the Acrania, Cyclostomes; Fishes, Amphibia, Reptiles, and then (within the mammal series) in the Monotremes, Marsupials, and the various Placentals. All the peculiarities of urogenital structure that distinguish the mammals from the rest of the Vertebrates are found in man; and in all special structural features he resembles the apes, particularly the anthropoid apes. In proof of the fact that the special features of the mammals have been inherited by man, I will, in conclusion, point out the identical way in which the ova are formed in the ovary. In all the mammals the mature ova are contained in special capsules, which are known as the Graafian
[ 347 ]
follicles, after their discoverer, Roger de Graaf (1677). They were formerly supposed to be the ova themselves; but Baer discovered the ova within the follicles (p. 16). Each follicle (Fig. 407) consists of a round fibrous capsule (d), which contains fluid and is lined with several strata of cells (c). The layer is thickened like a knob at one point (b); this ovum-capsule encloses the ovum proper (a). The mammal ovary is originally a very simple oval body (Fig. 387 g), formed only of connective tissue and blood-vessels, covered with a layer of cells, the ovarian epithelium or the female germ epithelium. From this germ epithelium strings of cells grow out into the connective tissue or “stroma” of the ovary (Fig. 403 b). Some of the cells of these strings (or Pflüger’s tubes) grow larger and become ova (primitive ova, c); but the great majority remain small, and form a protective and nutritive stratum of cells round each ovum—the “follicle-epithelium” (e).
The follicle-epithelium of the mammal has at first one stratum (Fig. 404 1), but afterwards several (2). It is true that in all the other Vertebrates the ova are enclosed in a membrane, or “follicle,” that consists of smaller cells. But it is only in the mammals that fluid accumulates between the growing follicle-cells, and distends the follicle into a large round capsule, on the inside wall of which the ovum lies, at one side (Figs. 405, 406). There again, as in the whole of his morphology, man proves indubitably his descent from the mammals.
In the lower Vertebrates the formation of ova in the germ-epithelium of the ovary continues throughout life; but in the higher it is restricted to the earlier stages, or even to the period of embryonic development. In man it seems to cease in the first year; in the second year we find no new-formed ova or chains of ova (Pflüger’s tubes). However, the number of ova in the two ovaries is very large in the young girl; there are calculated to be 72,000 in the sexually-mature maiden. In the production of the ova men resemble most of the anthropoid apes.
Generally speaking, the natural history of the human sexual organs is one of those parts of anthropology that furnish the most convincing proofs of the animal origin of the human race. Any man who is acquainted with the facts and impartially weighs them will conclude from them alone that we have been evolved from the lower Vertebrates. The larger and the detailed structure, the action, and the embryological development of the sexual organs are just the same in man as in the apes. This applies equally to the male and the female, the internal and the external organs. The differences we find in this respect between man and the anthropoid apes are much slighter than the differences between the various species of apes. But all the apes have certainly a common origin, and have been evolved from a long-extinct early-Tertiary stem-form, which we must trace to a branch of the lemurs. If we had this unknown pithecoid stem-form before us, we should certainly put it in the order of the true apes in the primate system; but within this order we cannot, for the anatomic and ontogenetic reasons we have seen, separate man from the group of the anthropoid apes. Here again, therefore, on the ground of the pithecometra-principle, comparative anatomy and ontogeny teach with full confidence the descent of man from the ape.
Title and Contents
Vol. II Title and Contents
Glossary
Chapter XXVIII
Chapter XXX
Figs. 1–209
Figs. 210–408