THE EVOLUTION OF MAN
Volume I
CHAPTER XI
THE VERTEBRATE CHARACTER OF MAN
We have now secured a number of firm standing-places in the labyrinthian course of our individual development by our study of the important embryonic forms which we have called the cytula, morula, blastula, gastrula, cœlomula, and chordula. But we have still in front of us the difficult task of deriving the complicated frame of the human body, with all its different parts, organs, members, etc., from the simple form of the chordula. We have previously considered the origin of this four-layered embryonic form from the two-layered gastrula. The two primary germinal layers, which form the entire body of the gastrula, and the two middle layers of the cœlomula that develop between them, are the four simple cell-strata, or epithelia, which alone go to the formation of the complex body of man and the higher animals. It is so difficult to understand this construction that we will first seek a companion who may help us out of many difficulties.
This helpful associate is the science of comparative anatomy. Its task is, by comparing the fully-developed bodily forms in the various groups of animals, to learn the general laws of organisation according to which the body is constructed; at the same time, it has to determine the affinities of the various groups by critical appreciation of the degrees of difference between them. Formerly, this work was conceived in a teleological sense, and it was sought to find traces of the plan of the Creator in the actual purposive organisation of animals. But comparative anatomy has gone much deeper since the establishment of the theory of descent; its philosophic aim now is to explain the variety of organic forms by adaptation, and their similarity by heredity. At the same time, it has to recognise in the shades of difference in form the degree of blood-relationship, and make an effort to construct the ancestral tree of the animal world. In this way, comparative anatomy enters into the closest relations with comparative embryology on the one hand, and with the science of classification on the other.
Now, when we ask what position man occupies among the other organisms according to the latest teaching of comparative anatomy and classification, and how man’s place in the zoological system is determined by comparison of the mature bodily forms, we get a very definite and significant reply; and this reply gives us extremely important conclusions that enable us to understand the embryonic development and its evolutionary purport. Since Cuvier and Baer, since the immense progress that was effected in the early decades of the nineteenth century by these two great zoologists, the opinion has generally prevailed that the whole animal kingdom may be distributed in a small number of great divisions or types. They are called types because a certain typical or characteristic structure is constantly preserved within each of these large sections. Since we applied the theory of descent to this doctrine of types, we have learned that this common type is an outcome of heredity; all the animals of one type are blood-relatives, or members of one stem, and can be traced to a common ancestral form. Cuvier and Baer set up four of these types: the vertebrates, articulates, molluscs, and radiates. The first three of these are still retained, and may be conceived as natural phylogenetic unities, as stems or phyla in the sense of the theory of descent. It is quite otherwise with the fourth type—the radiata. These animals, little known as yet at the beginning of the nineteenth century, were made to form a sort of lumber-room, into which were cast all the lower animals that did not belong to the other three types. As we obtained a closer acquaintance with them in the course of the last sixty years, it was found that we must distinguish among them from four to eight different types. In this way the total number of animal stems or phyla has been raised to eight or twelve (cf. Chapter XX).
[ 104 ]
These twelve stems of the animal kingdom are, however, by no means co-ordinate and independent types, but have definite relations, partly of subordination, to each other, and a very different phylogenetic meaning. Hence they must not be arranged simply in a row one after the other, as was generally done until thirty years ago, and is still done in some manuals. We must distribute them in three subordinate principal groups of very different value, and arrange the various stems phylogenetically on the principles which I laid down in my Monograph on the Sponges, and developed in the Study of the Gastræa Theory. We have first to distinguish the unicellular animals (protozoa) from the multicellular tissue-forming (metazoa). Only the latter exhibit the important processes of segmentation and gastrulation; and they alone have a primitive gut, and form germinal layers and tissues.
The metazoa, the tissue-animals or gut-animals, then sub-divide into two main sections, according as a body-cavity is or is not developed between the primary germinal layers. We may call these the cœlenteria and cœlomaria, the former are often also called zoophytes or cœlenterata, and the latter bilaterals. This division is the more important as the cœlenteria (without cœlom) have no blood and blood-vessels, nor an anus. The cœlomaria (with body-cavity) have generally an anus, and blood and blood-vessels. There are four stems belonging to the cœlenteria: the gastræads (“primitive-gut animals”), sponges, cnidaria, and platodes. Of the cœlomaria we can distinguish six stems: the vermalia at the bottom represent the common stem-group (derived from the platodes) of these, the other five typical stems of the cœlomaria—the molluscs, echinoderms, articulates, tunicates, and vertebrates—being evolved from them.
Man is, in his whole structure, a true vertebrate, and develops from an impregnated ovum in just the same characteristic way as the other vertebrates. There can no longer be the slightest doubt about this fundamental fact, nor of the fact that all the vertebrates form a natural phylogenetic unity, a single stem. The whole of the members of this stem, from the amphioxus and the cyclostoma to the apes and man, have the same characteristic disposition, connection, and development of the central organs, and arise in the same way from the common embryonic form of the chordula. Without going into the difficult question of the origin of this stem, we must emphasise the fact that the vertebrate stem has no direct affinity whatever to five of the other ten stems; these five isolated phyla are the sponges, cnidaria, molluscs, articulates, and echinoderms. On the other hand, there are important and, to an extent, close phylogenetic relations to the other five stems—the protozoa (through the amœbæ), the gastræads (through the blastula and gastrula), the platodes and vermalia (through the cœlomula), and the tunicates (through the chordula).
How we are to explain these phylogenetic relations in the present state of our knowledge, and what place is assigned to the vertebrates in the animal ancestral tree, will be considered later (Chapter XX). For the present our task is to make plainer the vertebrate character of man, and especially to point out the chief peculiarities of organisation by which the vertebrate stem is profoundly separated from the other eleven stems of the animal kingdom. Only after these comparative-anatomical considerations shall we be in a position to attack the difficult question of our embryology. The development of even the simplest and lowest vertebrate from the simple chordula (Figs. 83–86) is so complicated and difficult to follow that it is necessary to understand the organic features of the fully-formed vertebrate in order to grasp the course of its embryonic evolution. But it is equally necessary to confine our attention, in this general anatomic description of the vertebrate-body, to the essential facts, and pass by all the unessential. Hence, in giving now an ideal anatomic description of the chief features of the vertebrate and its internal organisation, I omit all the subordinate points, and restrict myself to the most important characteristics.
Much, of course, will seem to the reader to be essential that is only of subordinate and secondary interest, or even not essential at all, in the light of comparative anatomy and embryology. For instance, the skull and vertebral column and the extremities are non-essential in this sense. It is true that these parts are very important physiologically; but for the morphological conception of the vertebrate they are not essential, because they are only found in the higher, not the lower, vertebrates. The lowest vertebrates have
[ 105 ]
neither skull nor vertebræ, and no extremities or limbs. Even the human embryo passes through a stage in which it has no skull or vertebræ; the trunk is quite simple, and there is yet no trace of arms and legs. At this stage of development man, like every other higher vertebrate, is essentially similar to the simplest vertebrate form, which we now find in only one living specimen. This one lowest vertebrate that merits the closest study—undoubtedly the most interesting of all the vertebrates after man—is the famous lancelet or amphioxus, to which we have already often referred. As we are going to study it more closely later on (Chapters XVI and XVII), I will only make one or two passing observations on it here.
The amphioxus lives buried in the sand of the sea, is about one or two inches in length, and has, when fully developed, the shape of a very simple, longish, lancet-like leaf; hence its name of the lancelet. The narrow body is compressed on both sides, almost equally pointed at the fore and hind ends, without any trace of external appendages or articulation of the body into head, neck, breast, abdomen, etc. Its whole shape is so simple that its first discoverer thought it was a naked snail. It was not until much later—half a century ago—that the tiny creature was studied more carefully, and was found to be a true vertebrate. More recent investigations have shown that it is of the greatest importance in connection with the comparative anatomy and ontogeny of the vertebrates, and therefore with human phylogeny. The amphioxus reveals the great secret of the origin of the vertebrates from the invertebrate vermalia, and in its development and structure connects directly with certain lower tunicates, the ascidia.
When we make a number of sections of the body of the amphioxus, firstly vertical longitudinal sections through the whole body from end to end, and secondly transverse sections from right to left, we get anatomic pictures of the utmost instructiveness (cf. Figs. 98–102). In the main they correspond to the ideal which we form, with the aid of comparative anatomy and ontogeny, of the primitive type or build of the vertebrate—the long-extinct form to which the whole stem owes its origin. As we take the phylogenetic unity of the vertebrate stem to be beyond dispute, and assume a common origin from a primitive stem-form for all the vertebrates, from amphioxus to man, we are justified in forming a definite morphological idea of this primitive vertebrate (Prospondylus or Vertebræa). We need only imagine a few slight and unessential changes in the real sections of the amphioxus in order to have this ideal anatomic figure or diagram of the primitive vertebrate form, as we see in Figs. 98–102. The amphioxus departs so little from this primitive form that we may, in a certain sense, describe it as a modified “primitive vertebrate.”1
The outer form of our hypothetical primitive vertebrate was at all events very simple, and probably more or less similar to that of the lancelet. The bilateral or bilateral-symmetrical body is stretched out lengthways and compressed at the sides (Figs. 98–100), oval in section (Figs. 101, 102). There are no external articulation and no external appendages, in the shape of limbs, legs, or fins. On the other hand, the division of the body into two sections, head and trunk, was probably clearer in Prospondylus than it is in its little-changed ancestor, the amphioxus. In both animals the fore or head-half of the body contains different organs from the trunk, and different on the dorsal from on the ventral side. As this important division is found even in the sea-squirt, the remarkable invertebrate stem-relative of the vertebrates, we may assume that it was also found in the prochordonia, the common ancestors of both stems. It is also very pronounced in the young larvæ of the cyclostoma; this fact is particularly interesting, as this palingenetic larva-form is in other respects also an important connecting-link between the higher vertebrates and the acrania.
The head of the acrania, or the anterior half of the body (both of the real amphioxus and the ideal prospondylus), contains the branchial (gill) gut and heart in the ventral section and the brain and sense-organs in the dorsal section. The trunk, or posterior half of the body, contains the hepatic (liver) gut and sexual-
1. The ideal figure of the vertebrate as given in Figs. 98–102 is a hypothetical scheme or diagram, that has been chiefly constructed on the lines of the amphioxus, but with a certain attention to the comparative anatomy and ontogeny of the ascidia and appendicularia on the one hand, and of the cyclostoma and selachii on the other. This diagram has no pretension whatever to be an “exact picture,” but merely an attempt to reconstruct hypothetically the unknown and long extinct vertebrate stem-form, an ideal “archetype.”
[ 106 ]
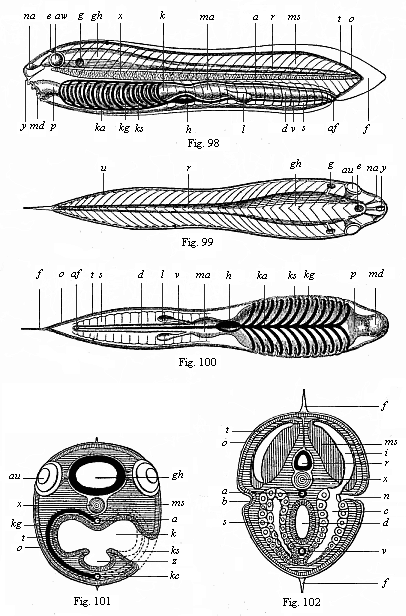
Figs. 98–102.—The ideal primitive vertebrate (prospondylus). Diagram. Fig. 98 side-view (from the left). Fig. 99 back-view. Fig. 100 front view. Fig. 101 transverse section through the head (to the left through the gill-pouches, to the right through the gill-clefts). Fig. 102 transverse section of the trunk (to the right a pro-renal canal is affected). a aorta, af anus, au eye, b lateral furrow (primitive renal process), c cœloma (body-cavity), d small intestine, e parietal eye (epiphysis), f fin border of the skin, g auditory vesicle, gh brain, h heart, i muscular cavity (dorsal cœlom-pouch), k gill-gut, ka gill-artery, kg gill-arch, ks gill-folds, l liver, ma stomach, md mouth, ms muscles, na nose (smell pit), n renal canals, u apertures of same, o outer skin, p gullet, r spinal marrow, a sexual glands (gonads), t corium, u kidney-openings (pores of the lateral furrow), v visceral vein (chief vein). x chorda, y hypophysis (urinary appendage), z gullet-groove or gill-groove (hypobranchial groove)./td> |
[ 107 ]
glands in the ventral part, and the spinal marrow and most of the muscles in the dorsal part.
In the longitudinal section of the ideal vertebrate (Fig. 98) we have in the middle of the body a thin and flexible, but stiff, cylindrical rod, pointed at both ends (ch). It goes the whole length through the middle of the body, and forms, as the central skeletal axis, the original structure of the later vertebral column. This is the axial rod, or chorda dorsalis, also called chorda vertebralis, vertebral cord, axial cord, dorsal cord, notochorda, or, briefly, chorda. This solid, but flexible and elastic, axial rod consists of a cartilaginous mass of cells, and forms the inner axial skeleton or central frame of the body; it is only found in vertebrates and tunicates, not in any other animals. As the first structure of the spinal column it has the same radical significance in all vertebrates, from the amphioxus to man. But it is only in the amphioxus and the cyclostoma that the axial rod retains its simplest form throughout life. In man and all the higher vertebrates it is found only in the earlier embryonic period, and is afterwards replaced by the articulated vertebral column.
The axial rod or chorda is the real solid chief axis of the vertebrate body, and at the same time corresponds to the ideal long-axis, and serves to direct us with some confidence in the orientation of the principal organs. We therefore take the vertebrate-body in its original, natural disposition, in which the long-axis lies horizontally, the dorsal side upward and the ventral side downward (Fig. 98). When we make a vertical section through the whole length of this long axis, the body divides into two equal and symmetrical halves, right and left. In each half we have originally the same organs in the same disposition and connection; only their disposal in relation to the vertical plane of section, or median plane, is exactly reversed: the left half is the reflection of the right. We call the two halves antimera (opposed-parts). In the vertical plane of section that divides the two halves the sagittal (“arrow”) axis, or “dorsoventral axis,” goes from the back to the belly, corresponding to the sagittal seam of the skull. But when we make a horizontal longitudinal section through the chorda, the whole body divides into a dorsal and a ventral half. The line of section that passes through the body from right to left is the transverse, frontal, or lateral axis.
The two halves of the vertebrate body that are separated by this horizontal transverse axis and by the chorda have quite different characters. The dorsal half is mainly the animal part of the body, and contains the greater part of what are called the animal organs, the nervous system, muscular system, osseous system, etc.—the instruments of movement and sensation. The ventral half is essentially the vegetative half of the body, and contains the greater part of the vertebrate’s vegetal organs, the visceral and vascular systems, sexual system, etc.—the instruments of nutrition and reproduction. Hence in the construction of the dorsal half it is chiefly the outer, and in the construction of the ventral half chiefly the inner, germinal layer that is engaged. Each of the two halves develops in the shape of a tube, and encloses a cavity in which another tube is found. The dorsal half contains the narrow spinal-column cavity or vertebral canal above the chorda, in which lies the tube-shaped central nervous system, the medullary tube. The ventral half contains the much more spacious visceral cavity or body-cavity underneath the chorda, in which we find the alimentary canal and all its appendages.
The medullary tube, as the central nervous system or psychic organ of the vertebrate is called in its first stage, consists, in man and all the higher vertebrates, of two different parts: the large brain, contained in the skull, and the long spinal cord which stretches from there over the whole dorsal part of the trunk. Even in the primitive vertebrate this composition is plainly indicated. The fore half of the body, which corresponds to the head, encloses a knob-shaped vesicle, the brain (gh); this is prolonged backwards into the thin cylindrical tube of the spinal marrow (r). Hence we find here this very important psychic organ, which accomplishes sensation, will, and thought, in the vertebrates, in its simplest form. The thick wall of the nerve-tube, which runs through the long axis of the body immediately over the axial rod, encloses a narrow central canal filled with fluid (Figs. 98–102 r). We still find the medullary tube in this very simple form for a time in the embryo of all the vertebrates, and it retains this form in the amphioxus throughout life;
[ 108 ]
only in the latter case the cylindrical medullary tube barely indicates the separation of brain and spinal cord. The lancelet’s medullary tube runs nearly the whole length of the body, above the chorda, in the shape of a long thin tube of almost equal diameter throughout, and there is only a slight swelling of it right at the front to represent the rudiment of a cerebral lobe. It is probable that this peculiarity of the amphioxus is connected with the partial atrophy of its head, as the ascidian larvæ on the one hand and the young cyclostoma on the other clearly show a division of the vesicular brain, or head marrow, from the thinner, tubular spinal marrow.
Probably we must trace to the same phylogenetic cause the defective nature of the sense organs of the amphioxus, which we will describe later (Chapter XVI). Prospondylus, on the other hand, probably had three pairs of sense-organs, though of a simple character, a pair of, or a single olfactory depression, right in front (Figs. 98, 99, na), a pair of eyes (au) in the lateral walls of the brain, and a pair of simple auscultory vesicles (g) behind. There was also, perhaps, a single parietal or “pineal” eye at the top of the skull (epiphysis, e).
In the vertical median plane (or middle plane, dividing the bilateral body into right and left halves) we have in the acrania, underneath the chorda, the mesentery and visceral tube, and above it the medullary tube; and above the latter a membranous partition of the two halves of the body. With this partition is connected the mass of connective tissue which acts as a sheath both for the medullary tube and the underlying chorda, and is, therefore, called the chord-sheath (perichorda); it originates from the dorsal and median part of the cœlom-pouches, which we shall call the skeleton plate or “sclerotom” in the craniote embryo. In the latter the chief part of the skeleton—the vertebral column and skull—develops from this chord-sheath; in the acrania it retains its simple form as a soft connective matter, from which are formed the membranous partitions between the various muscular plates or myotomes (Figs. 98, 99 ms).
To the right and left of the cord-sheath, at each side of the medullary tube and the underlying axial rod, we find in all the vertebrates the large masses of muscle that constitute the musculature of the trunk and effect its movements. Although these are very elaborately differentiated and connected in the developed vertebrate (corresponding to the various parts of the bony skeleton), in our ideal primitive vertebrate we can distinguish only two pairs of these principal muscles, which run the whole length of the body parallel to the chorda. These are the upper (dorsal) and lower (ventral) lateral muscles of the trunk. The upper (dorsal) muscles, or the original dorsal muscles (Fig. 102 ms), form the thick mass of flesh on the back. The lower (ventral) muscles, or the original muscles of the belly, form the fleshy wall of the abdomen. Both sets are segmented, and consist of a double row of muscular plates (Figs. 98, 99 ms); the number of these myotomes determines the number of joints in the trunk, or metamera. The myotomes are also developed from the thick wall of the cœlom-pouches (Fig. 102 i).
Outside this muscular tube we have the external envelope of the vertebrate body, which is known as the corium or cutis. This strong and thick envelope consists, in its deeper strata, chiefly of fat and loose connective tissue, and in its upper layers of cutaneous muscles and firmer connective tissue. It covers the whole surface of the fleshy body, and is of considerable thickness in all the craniota. But in the acrania the corium is merely a thin plate of connective tissue, an insignificant “corium-plate” (lamella corii, Figs. 98–102 t).
Immediately above the corium is the outer skin (epidermis, o), the general covering of the whole outer surface. In the higher vertebrates the hairs, nails, feathers, claws, scales, etc., grow out of this epidermis. It consists, with all its appendages and products, of simple cells, and has no blood-vessels. Its cells are connected with the terminations of the sensory nerves. Originally, the outer skin is a perfectly simple covering of the outer surface of the body, composed only of homogeneous cells—a permanent horn-plate. In this simplest form, as a one-layered epithelium, we find it, at first, in all the vertebrates, and throughout life in the acrania. It afterwards grows thicker in the higher vertebrates, and divides into two strata—an outer, firmer corneous (horn) layer and an inner, softer mucus-layer; also a number of external and internal appendages grow out of it: outwardly, the hairs, nails, claws, etc., and
[ 109 ]
inwardly, the sweat-glands, fat-glands, etc.
It is probable that in our primitive vertebrate the skin was raised in the middle line of the body in the shape of a vertical fin border (f). A similar fringe, going round the greater part of the body, is found to-day in the amphioxus and the cyclostoma; we also find one in the tail of fish-larvæ and tadpoles.
Now that we have considered the external parts of the vertebrate and the animal organs, which mainly lie in the dorsal half, above the chorda, we turn to the vegetal organs, which lie for the most part in the ventral half, below the axial rod. Here we find a large body-cavity or visceral cavity in all the craniota. The spacious cavity that encloses the greater part of the viscera corresponds to only a part of the original cœloma, which we considered in Chapter X; hence it nay be called the metacœloma. As a rule, it is still briefly called the cœloma; formerly it was known in anatomy as the pleuroperitoneal cavity. In man and the other mammals (but only in these) this cœloma divides, when fully developed, into two different cavities, which are separated by a transverse partition—the muscular diaphragm. The fore or pectoral cavity (pleura-cavity) contains the œsophagus (gullet), heart, and lungs; the hind or peritoneal or abdominal cavity contains the stomach, small and large intestines, liver, pancreas, kidneys, etc. But in the vertebrate embryo, before the diaphragm is developed, the two cavities form a single continuous body-cavity, and we find it thus in all the lower vertebrates throughout life. This body-cavity is clothed with a delicate layer of cells, the cœlom-epithelium. In the acrania the cœlom is segmented both dorsally and ventrally, as their muscular pouches and primitive genital organs plainly show (Fig. 102).
The chief of the viscera in the body-cavity is the alimentary canal, the organ that represents the whole body in the gastrula. In all the vertebrates it is a long tube, enclosed in the body-cavity and more or less differentiated in length, and has two apertures—a mouth for taking in food (Figs. 98, 100 md) and an anus for the ejection of unusable matter or excrements (af). With the alimentary canal a number of glands are connected which are of great importance for the vertebrate body, and which all grow out of the canal. Glands of this kind are the salivary glands, the lungs, the liver, and many smaller glands. Nearly all these glands are wanting in the acrania; probably there were merely a couple of simple hepatic tubes (Figs. 98, 100 l) in the vertebrate stem-form. The wall of the alimentary canal and all its appendages consists of two different layers; the inner, cellular clothing is the gut-gland-layer, and the outer, fibrous envelope consists of the gut-fibre-layer; it is mainly composed of muscular fibres which accomplish the digestive movements of the canal, and of connective-tissue fibres that form a firm envelope. We have a continuation of it in the mesentery, a thin, bandage-like layer, by means of which the alimentary canal is fastened to the ventral side of the chorda, originally the dorsal partition of the two cœlom-pouches. The alimentary canal is variously modified in the vertebrates both as a whole and in its several sections, though the original structure is always the same, and is very simple. As a rule, it is longer (often several times longer) than the body, and therefore folded and winding within the body-cavity, especially at the lower end. In man and the higher vertebrates it is divided into several sections, often separated by valves—the mouth, pharynx, œsophagus, stomach, small and large intestine, and rectum. All these parts develop from a very simple structure, which originally (throughout life in the amphioxus) runs from end to end under the chorda in the shape of a straight cylindrical canal.
As the alimentary canal may be regarded morphologically as the oldest and most important organ in the body, it is interesting to understand its essential features in the vertebrate more fully, and distinguish them from unessential features. In this connection we must particularly note that the alimentary canal of every vertebrate shows a very characteristic division into two sections—a fore and a hind chamber. The fore chamber is the head-gut or branchial gut (Figs. 98–100 p, k), and is chiefly occupied with respiration. The hind section is the trunk-gut or hepatic gut, which accomplishes digestion (ma, d). In all vertebrates there are formed, at an early stage, to the right and left in the fore-part of the head-gut, certain special clefts that have an intimate connection with the original respiratory apparatus of
[ 110 ]
the vertebrate—the branchial (gill) clefts (ks). All the lower vertebrates, the lancelets, lampreys, and fishes, are constantly taking in water at the mouth, and letting it out again by the lateral clefts of the gullet. This water serves for breathing. The oxygen contained in it is inspired by the blood-canals, which spread out on the parts between the gill-clefts, the gill-arches (kg). These very characteristic branchial clefts and arches are found in the embryo of man and all the higher vertebrates at an early stage of development, just as we find them throughout life in the lower vertebrates. However, these clefts and arches never act as respiratory organs in the mammals, birds, and reptiles, but gradually develop into quite different parts. Still, the fact that they are found at first in the same form as in the fishes is one of the most interesting proofs of the descent of these three higher classes from the fishes.
Not less interesting and important is an organ that develops from the ventral wall in all vertebrates—the gill-groove or hypobranchial groove. In the acrania and the ascidiæ it consists throughout life of a glandular ciliated groove, which runs down from the mouth in the ventral middle line of the gill-gut, and takes small particles of food to the stomach (Fig. 101 z). But in the craniota the thyroid gland (thyreoidea) is developed from it, the gland that lies in front of the larynx, and which, when pathologically enlarged, forms goitre (struma).
From the head-gut we get not only the gills, the organs of water-breathing in the lower vertebrates, but also the lungs, the organs of atmospheric breathing in the five higher classes. In these cases a vesicular fold appears in the gullet of the embryo at an early stage, and gradually takes the shape of two spacious sacs, which are afterwards filled with air. These sacs are the two air-breathing lungs, which take the place of the water-breathing gills. But the vesicular invagination, from which the lungs arise, is merely the familiar air-filled vesicle, which we call the floating-bladder of the fish, and which alters its specific weight, acting as hydrostatic organ or floating apparatus. This structure is not found in the lowest vertebrate classes—the acrania and cyclostoma. We shall see more of it in Volume II.
The second chief section of the vertebrate-gut, the trunk or liver-gut, which accomplishes digestion, is of very simple construction in the acrania. It consists of two different chambers. The first chamber, immediately behind the gill-gut, is the expanded stomach (ma); the second, narrower and longer chamber, is the straight small intestine (d): it issues behind on the ventral side by the anus (af). Near the limit of the two chambers in the visceral cavity we find the liver, in the shape of a simple tube or blind sac (l); in the amphioxus it is single; in the prospondylus it was probably double (Figs. 98, 100 l).
Closely related morphologically and physiologically to the alimentary canal is the vascular system of the vertebrate, the chief sections of which develop from the fibrous gut-layer. It consists of two different but directly connected parts, the system of blood-vessels and that of lymph-vessels. In the passages of the one we find red blood, and in the other colourless lymph. To the lymphatic system belong, first of all, the lymphatic canals proper or absorbent veins, which are distributed among all the organs, and absorb the used-up juices from the tissues, and conduct them into the venous blood; but besides these there are the chyle-vessels, which absorb the white chyle, the milky fluid prepared by the alimentary canal from the food, and conduct this also to the blood.
The blood-vessel system of the vertebrate has a very elaborate construction, but seems to have had a very simple form in the primitive vertebrate, as we find it to-day permanently in the annelids (for instance, earth-worms) and the amphioxus. We accordingly distinguish first of all as essential, original parts of it two large single blood-canals, which lie in the fibrous wall of the gut, and run along the alimentary canal in the median plane of the body, one above and the other underneath the canal. These principal canals give out numerous branches to all parts of the body, and pass into each other by arches before and behind; we will call them the primitive artery and the primitive vein. The first corresponds to the dorsal vessel, the second to the ventral vessel, of the worms. The primitive or principal artery, usually called the aorta (Fig. 98 a), lies above the gut in the middle line of its dorsal side, and conducts oxidised or arterial blood from the gills to the body. The primitive or principal vein (Fig. 100 v) lies below the
[ 111 ]
gut, in the middle line of its ventral side, and is therefore also called the vena subintestinalis; it conducts carbonised or venous blood back from the body to the gills. At the branchial section of the gut in front the two canals are connected by a number of branches, which rise in arches between the gill-clefts. These “branchial vascular arches” (kg) run along the gill-arches, and have a direct share in the work of respiration. The anterior continuation of the principal vein which runs on the ventral wall of the gill-gut, and gives off these vascular arches upwards, is the branchial artery (ka). At the border of the two sections of the ventral vessel it enlarges into a contractile spindle-shaped tube (Figs. 98, 100 h). This is the first outline of the heart, which afterwards becomes a four-chambered pump in the higher vertebrates and man. There is no heart in the amphioxus, probably owing to degeneration. In prospondylus the ventral gill-heart probably had the simple form in which we still find it in the ascidia and the embryos of the craniota (Figs. 98, 100 h).
The kidneys, which act as organs of excretion or urinary organs in all vertebrates, have a very different and elaborate construction in the various sections of this stem; we will consider them further in Chapter 2.29. Here I need only mention that in our hypothetical primitive vertebrate they probably had the same form as in the actual amphioxus—the primitive kidneys (protonephra). These are originally made up of a double row of little canals, which directly convey the used-up juices or the urine out of the body-cavity (Fig. 102 n). The inner aperture of these pronephridial canals opens with a ciliated funnel into the body-cavity; the external aperture opens in lateral grooves of the epidermis, a couple of longitudinal grooves in the lateral surface of the outer skin (Fig. 102 b). The pronephridial duct is formed by the closing of this groove to the right and left at the sides. In all the craniota it develops at an early stage in the horny plate; in the amphioxus it seems to be converted into a wide cavity, the atrium, or peribranchial space.
Next to the kidneys we have the sexual organs of the vertebrate. In most of the members of this stem the two are united in a single urogenital system; it is only in a few groups that the urinary and sexual organs are separated (in the amphioxus, the cyclostoma, and some sections of the fish-class). In man and all the higher vertebrates the sexual apparatus is made up of various parts, which we will consider in Chapter XXIX. But in the two lowest classes of our stem, the acrania and cyclostoma, they consist merely of simple sexual glands or gonads, the ovaries of the female sex and the testicles (spermaria) of the male; the former provide the ova, the latter the sperm. In the craniota we always find only one pair of gonads; in the amphioxus several pairs, arranged in succession. They must have had the same form in our hypothetical prospondylus (Figs. 98, 100 s). These segmental pairs of gonads are the original ventral halves of the cœlom-pouches.
The organs which we have now enumerated in this general survey, and of which we have noted the characteristic disposition, are those parts of the organism that are found in all vertebrates without exception in the same relation to each other, however much they may be modified. We have chiefly had in view the transverse section of the body (Figs. 101, 102), because in this we see most clearly the distinctive arrangement of them. But to complete our picture we must also consider the segmentation or metamera-formation of them, which has yet been hardly noticed, and which is seen best in the longitudinal section. In man and all the more advanced vertebrates the body is made up of a series or chain of similar members, which succeed each other in the long axis of the body—the segments or metamera of the organism. In man these homogeneous parts number thirty-three in the trunk, but they run to several hundred in many of the vertebrates (such as serpents or eels). As this internal articulation or metamerism is mainly found in the vertebral column and the surrounding muscles, the sections or metamera were formerly called pro-vertebræ. As a fact, the articulation is by no means chiefly determined and caused by the skeleton, but by the muscular system and the segmental arrangement of the kidneys and gonads. However, the composition from these pro-vertebræ or internal metamera is usually, and rightly, put forward as a prominent character of the vertebrate, and the manifold division or differentiation of them is of great importance in the various groups of the vertebrates. But as far as our present
[ 112 ]
task—the derivation of the simple body of the primitive vertebrate from the chordula—is concerned, the articulate parts or metamera are of secondary interest, and we need not go into them just now.
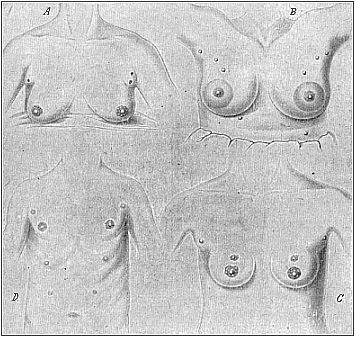
Fig. 103 A, B, C, D.—Instances of redundant mammary glands and nipples (hypermastism). A a pair of small redundant breasts (with two nipples on the left) above the large normal ones; from a 45-year-old Berlin woman, who had had children 17 times (twins twice). (From Hansemann.) B the highest number: ten nipples (all giving milk), three pairs above, one pair below, the large normal breasts; from a 22-year-old servant at Warschau. (From Neugebaur.) C three pairs of nipples: two pairs on the normal glands and one pair above; from a 19-year-old Japanese girl. D four pairs of nipples: one pair above the normal and two pairs of small accessory nipples underneath; from a 22-year-old Bavarian soldier. (From Wiedersheim.) |
The characteristic composition of the vertebrate body develops from the embryonic structure in the same way in man as in all the other vertebrates. As all competent experts now admit the monophyletic origin of the vertebrates on the strength of this significant agreement, and this “common descent of all the vertebrates from one original stem-form” is admitted as an historical fact, we have found the answer to “the question of questions.” We may, moreover, point out that this answer is just as certain and precise in the case of the origin of man from the mammals. This advanced vertebrate class is also monophyletic, or has evolved from one common stem-group of lower vertebrates (reptiles, and, earlier still, amphibia). This follows from the fact that the mammals are clearly distinguished from the other classes of the stem, not merely in one striking particular, but in a whole group of distinctive characters.
It is only in the mammals that we find the skin covered with hair, the breast-cavity separated from the abdominal cavity by a complete diaphragm, and the larynx provided with an epiglottis. The
[ 113 ]
mammals alone have three small auscultory bones in the tympanic cavity—a feature that is connected with the characteristic modification of their maxillary joint. Their red blood-cells have no nucleus, whereas this is retained in all other vertebrates. Finally, it is only in the mammals that we find the remarkable function of the breast structure which has given its name to the whole class—the feeding of the young by the mother’s milk. The mammary glands which serve this purpose are interesting in so many ways that we may devote a few lines to them here.
As is well known, the lower mammals, especially those which beget a number of young at a time, have several mammary glands at the breast. Hedgehogs and sows have five pairs, mice four or five pairs, dogs and squirrels four pairs, cats and bears three pairs, most of the ruminants and many of the rodents two pairs, each provided with a teat or nipple (mastos). In the various genera of the half-apes (lemurs) the number varies a good deal. On the other hand, the bats and apes, which only beget one young at a time as a rule, have only one pair of mammary glands, and these are found at the breast, as in man.
These variations in the number or structure of the mammary apparatus (mammarium) have become doubly interesting in the light of recent research in comparative anatomy. It has been shown that in man and the apes we often find redundant mammary glands (hyper-mastism) and corresponding teats (hyper-thelism) in both sexes. Fig. 103 shows four cases of this kind—A, B, and C of three women, and D of a man. They prove that all the above-mentioned numbers may be found occasionally in man. Fig. 103 A shows the breast of a Berlin woman who had had children seventeen times, and who has a pair of small accessory breasts (with two nipples on the left one) above the two normal breasts; this is a common occurrence, and the small soft pad above the breast is not infrequently represented in ancient statues of Venus. In Fig. 103 C we have the same phenomenon in a Japanese girl of nineteen, who has two nipples on each breast besides (three pairs altogether). Fig. 103 D is a man of twenty-two with four pairs of nipples (as in the dog), a small pair above and two small pairs beneath the large normal teats. The maximum number of five pairs (as in the sow and hedgehog) was found in a Polish servant of twenty-two who had had several children; milk was given by each nipple; there were three pairs of redundant nipples above and one pair underneath the normal and very large breasts (Fig. 103 B).
A number of recent investigations (especially among recruits) have shown that these things are not uncommon in the male as well as the female sex. They can only be explained by evolution, which attributes them to atavism and latent heredity. The earlier ancestors of all the primates (including man) were lower placentals, which had, like the hedgehog (one of the oldest forms of the living placentals), several mammary glands (five or more pairs) in the abdominal skin. In the apes and man only a couple of them are normally developed, but from time to time we get a development of the atrophied structures. Special notice should be taken of the arrangement of these accessory mammæ; they form, as is clearly seen in Fig. 103 B and D, two long rows, which diverge forward (towards the arm-pit), and converge behind in the middle line (towards the loins). The milk-glands of the polymastic lower placentals are arranged in similar lines.
The phylogenetic explanation of polymastism, as given in comparative anatomy, has lately found considerable support in ontogeny. Hans Strahl, E. Schmitt, and others, have found that there are always in the human embryo at the sixth week (when it is three-fifths of an inch long) the microscopic traces of five pairs of mammary glands, and that they are arranged at regular distances in two lateral and divergent lines, which correspond to the mammary lines. Only one pair of them—the central pair—are normally developed, the others atrophying. Hence there is for a time in the human embryo a normal hyperthelism, and this can only be explained by the descent of man from lower primates (lemurs) with several pairs.
But the milk-gland of the mammal has a great morphological interest from another point of view. This organ for feeding the young in man and the higher mammals is, as is known, found in both sexes. However, it is usually active only in the female sex, and yields the valuable “mother’s milk”; in the male sex it is
[ 114 ]
small and inactive, a real rudimentary organ of no physiological interest. Nevertheless, in certain cases we find the breast as fully developed in man as in woman, and it may give milk for feeding the young.
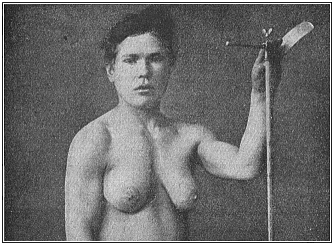
We have a striking instance of this gynecomastism (large milk-giving breasts in a male) in Fig. 104. I owe the photograph (taken from life) to the kindness of Dr. Ornstein, of Athens, a German physician, who has rendered service by a number of anthropological observations, (for instance, in several cases of tailed men). The gynecomast in question is a Greek recruit in his twentieth year, who has both normally developed male organs and very pronounced female breasts. It is noteworthy that the other features of his structure are in accord with the softer forms of the female sex. It reminds us of the marble statues of hermaphrodites which the ancient Greek and Roman sculptors often produced. But the man would only be a real hermaphrodite if he had ovaries internally besides the (externally visible) testicles.
I observed a very similar case during my stay in Ceylon (at Belligemma) in 1881. A young Cinghalese in his twenty-fifth year was brought to me as a curious hermaphrodite, half-man and half-woman. His large breasts gave plenty of milk; he was employed as “male nurse” to suckle a new-born infant whose mother had died at birth. The outline of his body was softer and more feminine than in the Greek shown in Fig. 104. As the Cinghalese are small of stature and of graceful build, and as the men often resemble the women in clothing (upper part of the body naked, female dress on the lower part) and the dressing of the hair (with a comb), I first took the beardless youth to be a woman. The illusion was greater, as in this remarkable case gynecomastism was associated with cryptorchism—that is to say, the testicles had kept to their original place in the visceral cavity, and had not travelled in the normal way down into the scrotum. (Cf. Chapter XXIX.) Hence the latter was very small, soft, and empty. Moreover, one could feel nothing of the testicles in the inguinal canal. On the other hand, the male organ was very small, but normally developed. It was
[ 115 ]
clear that this apparent hermaphrodite also was a real male.
Another case of practical gynecomastism has been described by Alexander von Humboldt. In a South American forest he found a solitary settler whose wife had died in child-birth. The man had laid the new-born child on his own breast in despair; and the continuous stimulus of the child’s sucking movements had revived the activity of the mammary glands. It is possible that nervous suggestion had some share in it. Similar cases have been often observed in recent years, even among other male mammals (such as sheep and goats).
The great scientific interest of these facts is in their bearing on the question of heredity. The stem-history of the mammarium rests partly on its embryology (Chapter XXIV.) and partly on the facts of comparative anatomy and physiology. As in the lower and higher mammals (the monotremes, and most of the marsupials) the whole lactiferous apparatus is only found in the female; and as there are traces of it in the male only in a few younger marsupials, there can be no doubt that these important organs were originally found only in the female mammal, and that they were acquired by these through a special adaptation to habits of life.
Later, these female organs were communicated to both sexes by heredity; and they have been maintained in all persons of either sex, although they are not physiologically active in the males. This normal permanence of the female lactiferous organs in both sexes of the higher mammals and man is independent of any selection, and is a fine instance of the much-disputed “inheritance of acquired characters.”
Title and Contents
Glossary
Chapter X
Chapter XII
Figs. 1–209
Figs. 210–408