THE EVOLUTION OF MAN
Volume I
CHAPTER VII
CONCEPTION
The recognition of the fact that every man begins
his individual existence as a simple cell is the solid foundation
of all research into the genesis of man. From this fact we are
forced, in virtue of our biogenetic law, to draw the weighty
phylogenetic conclusion that the earliest ancestors of the human
race were also unicellular organisms; and among these protozoa we
may single out the vague form of the amœba as particularly
important (cf. Chapter VI). That these unicellular ancestral forms
did once exist follows directly from the phenomena which we
perceive every day in the fertilised ovum. The development of the
multicellular organism from the ovum, and the formation of the
germinal layers and the tissues, follow the same laws in man and
all the higher animals. It will, therefore, be our next task to
consider more closely the impregnated ovum and the process of
conception which produces it.
The process of impregnation or sexual conception is one of those
phenomena that people love to conceal behind the mystic veil of
supernatural power. We shall soon see, however, that it is a purely
mechanical process, and can be reduced to familiar physiological
functions. Moreover, this process of conception is of the same
type, and is effected by the same organs, in man as in all the
other mammals. The pairing of the male and female has in both cases
for its main purpose the introduction of the ripe matter of the
male seed or sperm into the female body, in the sexual canals of
which it encounters the ovum. Conception then ensues by the
blending of the two.
We must observe, first, that this important process is by no
means so widely distributed in the animal and plant world as is
commonly supposed. There is a very large number of lower organisms
which propagate unsexually, or by monogamy; these are especially
the sexless monera (chromacea, bacteria, etc.) but also many other
protists, such as the amœbæ, foraminifera, radiolaria,
myxomycetæ, etc. In these the multiplication of individuals
takes place by unsexual reproduction, which takes the form of
cleavage, budding, or spore-formation. The copulation of two
coalescing cells, which in these cases often precedes the
reproduction, cannot be regarded as a sexual act unless the two
copulating plastids differ in size or structure. On the other hand,
sexual reproduction is the general rule with all the higher
organisms, both animal and plant; very rarely do we find asexual
reproduction among them. There are, in particular, no cases of
parthenogenesis (virginal conception) among the vertebrates.
Sexual reproduction offers an infinite variety of interesting
forms in the different classes of animals and plants, especially as
regards the mode of conception, and the conveyance of the
spermatozoon to the ovum. These features are of great importance
not only as regards conception itself, but for the development of
the organic form, and especially for the differentiation of the
sexes. There is a particularly curious correlation of plants and
animals in this respect. The splendid studies of Charles Darwin and
Hermann Müller on the fertilisation of flowers by insects have
given us very interesting particulars of this.1 This
reciprocal service has given rise to a most intricate sexual
apparatus. Equally elaborate structures have been developed in man
and the higher animals, serving partly for the isolation of the
sexual products on each side, partly for bringing them together in
conception. But, however interesting these phenomena are in
themselves, we cannot go into them here, as they have only a minor
importance—if any at all—in the real process of
conception. We must, however, try to get a very clear idea of this
process and the meaning of sexual reproduction.
1. See Darwin’s work, On the Various
Contrivances by which Orchids are Fertilised (1862).
[ 52 ]
In every act of conception we have, as I said, to
consider two different kinds of cells—a female and a male
cell. The female cell of the animal organism is always called the
ovum (or ovulum, egg, or egg-cell); the male cells are known
as the sperm or seed-cells, or the spermatozoa (also spermium and
zoospermium). The ripe ovum is, on the whole, one of the largest
cells we know. It attains colossal dimensions when it absorbs great
quantities of nutritive yelk, as is the case with birds and
reptiles and many of the fishes. In the great majority of the
animals the ripe ovum is rich in yelk and much larger than the
other cells. On the other hand, the next cell which we have to
consider in the process of conception, the male sperm-cell or
spermatozoon, is one of the smallest cells in the animal body.
Conception usually consists in the bringing into contact with the
ovum of a slimy fluid secreted by the male, and this may take place
either inside or out of the female body. This fluid is called
sperm, or the male seed. Sperm, like saliva or blood, is not a
simple fluid, but a thick agglomeration of innumerable cells,
swimming about in a comparatively small quantity of fluid. It is
not the fluid, but the independent male cells that swim in it, that
cause conception.
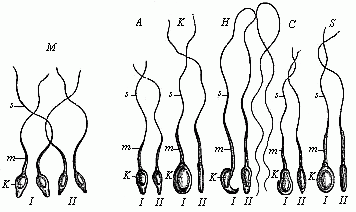
Fig. 20—Spermia or
spermatozoa of various mammals. The pear-shaped flattened
nucleus is seen from the front in I and sideways in II.
k is the nucleus, m its middle part (protoplasm),
s the mobile, serpent-like tail (or whip); M four human
spermatozoa, A four spermatozoa from the ape; K from
the rabbit; H from the mouse; C from the dog;
S from the pig. |
The spermatozoa of the great majority of animals have two
characteristic features. Firstly, they are extraordinarily small,
being usually the smallest cells in the body; and, secondly, they
have, as a rule, a peculiarly lively motion, which is known as
spermatozoic motion. The shape of the cell has a good deal to do
with this motion. In most of the animals, and also in many of the
lower plants (but not the higher) each of these spermatozoa has a
very small, naked cell-body, enclosing an elongated nucleus, and a
long thread hanging from it (Fig. 20). It was long before we could
recognise that these structures are simple cells. They were
formerly held to be special organisms, and were called “seed
animals” (spermato-zoa, or spermato-zoidia); they are now
scientifically known as spermia or spermidia, or as
spermatosomata (seed-bodies) or spermatofila (seed
threads). It took a good deal of comparative research to convince
us that each of these spermatozoa is really a simple cell. They
have the same shape as in many other vertebrates and most of the
invertebrates. However, in many of the lower animals they have
quite a different shape. Thus, for instance, in the craw fish they
are large round cells, without any movement, equipped with stiff
outgrowths like bristles (Fig. 21
f ). They have also a peculiar form in some of the
worms, such as the thread-worms (filaria); in this case they
are sometimes
[ 53 ]
amœboid and like very small ova (Fig. 21
c to e). But in most of the lower animals (such as the
sponges and polyps) they have the same pine-cone shape as in man
and the other animals (Fig. 21 a, h).
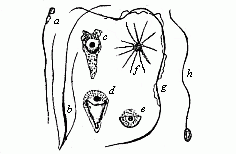 Fig. 21—Spermatozoa
or spermidia of various animals. (From Lang). a
of a fish, b of a turbellaria worm (with two side-lashes),
c to e of a nematode worm (amœboid
spermatozoa), f from a craw fish (star-shaped), g
from the salamander (with undulating membrane), h of an
annelid (a and h are the usual shape).
Fig. 21—Spermatozoa
or spermidia of various animals. (From Lang). a
of a fish, b of a turbellaria worm (with two side-lashes),
c to e of a nematode worm (amœboid
spermatozoa), f from a craw fish (star-shaped), g
from the salamander (with undulating membrane), h of an
annelid (a and h are the usual shape). |
When the Dutch naturalist Leeuwenhoek discovered these
thread-like lively particles in 1677 in the male sperm, it was
generally believed that they were special, independent, tiny
animalcules, like the infusoria, and that the whole mature organism
existed already, with all its parts, but very small and packed
together, in each spermatozoon (see
p.12). We now know that the mobile spermatozoa are nothing but
simple and real cells, of the kind that we call
“ciliated” (equipped with lashes, or cilia). In
the previous illustrations we have distinguished in the
spermatozoon a head, trunk, and tail. The “head” (Fig. 20 k) is merely the oval nucleus
of the cell; the body or middle-part (m) is an accumulation
of cell-matter; and the tail (s) is a thread-like
prolongation of the same.
Moreover, we now know that these spermatozoa are not at all a
peculiar form of cell; precisely similar cells are found in various
other parts of the body. If they have many short threads
projecting, they are called ciliated; if only one long,
whip-shaped process (or, more rarely, two or four), caudate
(tailed) cells.
Very careful recent examination of the spermia, under a very
high microscopic power (Fig. 22 a, b), has detected some further
details in the finer structure of the ciliated cell, and these are
common to man and the anthropoid ape. The head (k) encloses
the elliptic nucleus in a thin envelope of cytoplasm; it is a
little flattened on one side, and thus looks rather pear-shaped
from the front (b). In the central piece (m) we can
distinguish a short neck and a longer connective piece (with
central body). The tail consists of a long main section (h)
and a short, very fine tail (e).
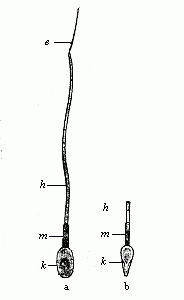 Fig. 22—A single
human spermatozoon magnified; a shows it from the broader and b
from the narrower side. k head (with nucleus), m
middle-stem, h long-stem, and e tail. (From
Retzius.)
Fig. 22—A single
human spermatozoon magnified; a shows it from the broader and b
from the narrower side. k head (with nucleus), m
middle-stem, h long-stem, and e tail. (From
Retzius.) |
The process of fertilisation by sexual conception consists,
therefore, essentially in the coalescence and fusing together of
two different cells. The lively spermatozoon travels towards the
ovum by its serpentine movements, and bores its way into the female
cell (Fig. 23). The nuclei of both sexual
cells, attracted by a certain “affinity,” approach each
other and melt into one.
The fertilised cell is quite another thing from the unfertilised
cell. For if we must regard the spermia as real cells no less than
the ova, and the process of conception as a coalescence of the two,
we must consider the resultant cell as a quite new and independent
organism. It bears in the cell and nuclear matter of the
penetrating spermatozoon a part of the father’s body, and in
the protoplasm and caryoplasm of the ovum a part of the
mother’s body. This is clear from the fact that the child
inherits many features from both parents. It inherits from the
father by means of the spermatozoon, and from the mother by means
of the ovum. The
[ 54 ]
actual blending of the two cells produces a third
cell, which is the germ of the child, or the new organism
conceived. One may also say of this sexual coalescence that the
stem-cell is a simple hermaphrodite; it unites both sexual
substances in itself.
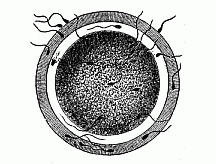 Fig. 23—The fertilisation of the ovum by the spermatozoon (of a mammal). One of the many thread-like, lively spermidia pierces through a fine pore-canal into the nuclear yelk. The nucleus of the ovum is invisible.
Fig. 23—The fertilisation of the ovum by the spermatozoon (of a mammal). One of the many thread-like, lively spermidia pierces through a fine pore-canal into the nuclear yelk. The nucleus of the ovum is invisible. |
I think it necessary to emphasise the fundamental importance of
this simple, but often unappreciated, feature in order to have a
correct and clear idea of conception. With that end, I have given a
special name to the new cell from which the child develops, and
which is generally loosely called “the fertilised
ovum,” or “the first segmentation sphere.” I call
it “the stem-cell” (cytula). The name
“stem-cell” seems to me the simplest and most suitable,
because all the other cells of the body are derived from it, and
because it is, in the strictest sense, the stem-father and
stem-mother of all the countless generations of cells of which the
multicellular organism is to be composed. That complicated
molecular movement of the protoplasm which we call
“life” is, naturally, something quite different in this
stem-cell from what we find in the two parent-cells, from the
coalescence of which it has issued. The life of the stem-cell or
cytula is the product or resultant of the paternal life-movement
that is conveyed in the spermatozoon and the maternal life-movement
that is contributed by the ovum.
The admirable work done by recent observers has shown that the
individual development, in man and the other animals, commences
with the formation of a simple “stem-cell” of this
character, and that this then passes, by repeated segmentation (or
cleavage), into a cluster of cells, known as “the
segmentation sphere” or “segmentation cells.” The
process is most clearly observed in the ova of the echinoderms
(star-fishes, sea-urchins, etc.). The investigations of Oscar and
Richard Hertwig were chiefly directed to these. The main results
may be summed up as follows:—
Conception is preceded by certain preliminary changes, which are
very necessary—in fact, usually indispensable—for its
occurrence. They are comprised under the general heading of
“Changes prior to impregnation.” In these the original
nucleus of the ovum, the germinal vesicle, is lost. Part of it is
extruded, and part dissolved in the cell contents; only a very
small part of it is left to form the basis of a fresh nucleus, the
pronucleus femininus. It is the latter alone that combines
in conception with the invading nucleus of the fertilising
spermatozoon (the pronucleus masculinus).
The impregnation of the ovum commences with a decay of the
germinal vesicle, or the original nucleus of the ovum (Fig. 8). We have seen that this is in most
unripe ova a large, transparent, round vesicle. This germinal
vesicle contains a viscous fluid (the caryolymph). The firm
nuclear frame (caryobasis) is formed of the enveloping
membrane and a mesh-work of nuclear threads running across the
interior, which is filled with the nuclear sap. In a knot of the
network is contained the dark, stiff, opaque nuclear corpuscle or
nucleolus. When the impregnation of the ovum sets in, the greater
part of the germinal vesicle is dissolved in the cell; the nuclear
membrane and mesh-work disappear; the nuclear sap is distributed in
the protoplasm; a small portion of the nuclear base is extruded;
another small portion is left, and is converted into the secondary
nucleus, or the female pro-nucleus (Fig. 24 e
k).
The small portion of the nuclear base which is extruded from the
impregnated ovum is known as the “directive bodies” or
“polar cells”; there are many disputes as to their
origin and significance, but we are as yet imperfectly acquainted
with them. As a rule, they are two small round granules, of the
same size and appearance as the remaining pro-nucleus. They are
detached cell-buds; their separation from the large mother-cell
takes
[ 55 ]
place in the same way as in ordinary “indirect
cell-division.” Hence, the polar cells are probably to be
conceived as “abortive ova,” or “rudimentary
ova,” which proceed from a simple original ovum by cleavage
in the same way that several sperm-cells arise from one
“sperm-mother-cell,” in reproduction from sperm. The
male sperm-cells in the testicles must undergo similar changes in
view of the coming impregnation as the ova in the female ovary. In
this maturing of the sperm each of the original seed-cells divides
by double segmentation into four daughter-cells, each furnished
with a fourth of the original nuclear matter (the hereditary
chromatin); and each of these four descendant cells becomes a
spermatozoon, ready for impregnation. Thus is prevented the
doubling of the chromatin in the coalescence of the two nuclei at
conception. As the two polar cells are extruded and lost, and have
no further part in the fertilisation of the ovum, we need not
discuss them any further. But we must give more attention to the
female pro-nucleus which alone remains after the extrusion of the
polar cells and the dissolving of the germinal vesicle (Fig. 23 e k). This tiny round corpuscle of
chromatin now acts as a centre of attraction for the invading
spermatozoon in the large ripe ovum, and coalesces with its
“head,” the male pro-nucleus. The product of this
blending, which is the most important part of the act of
impregnation, is the stem-nucleus, or the first segmentation
nucleus (archicaryon)—that is to say, the nucleus of
the new-born embryonic stem-cell or “first segmentation
cell.” This stem-cell is the starting point of the subsequent
embryonic processes.
Hertwig has shown that the tiny transparent ova of the
echinoderms are the most convenient for following the details of
this important process of impregnation. We can, in this case,
easily and successfully accomplish artificial impregnation, and
follow the formation of the stem-cell step by step within the space
of ten minutes. If we put ripe ova of the star-fish or sea-urchin
in a watch glass with sea-water and add a drop of ripe sperm-fluid,
we find each ovum impregnated within five minutes. Thousands of the
fine, mobile ciliated cells, which we have described as
“sperm-threads” (Fig. 20), make
their way to the ova, owing to a sort of chemical sensitive action
which may be called “smell.” But only one of these
innumerable spermatozoa is chosen—namely, the one that first
reaches the ovum by the serpentine motions of its tail, and touches
the ovum with its head. At the spot where the point of its head
touches the surface of the ovum the protoplasm of the latter is
raised in the form of a small wart, the “impregnation
rise” (Fig. 25 A). The
spermatozoon then bores its way into this with its head, the tail
outside wriggling about all the time (Fig. 25 B, C).
Presently the tail also disappears within the ovum. At the same
time the ovum secretes a thin external yelk-membrane (Fig. 25
C), starting from the point of impregnation; and this prevents
any more spermatozoa from entering.
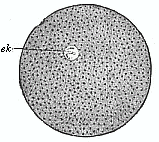 Fig. 24—An
impregnated echinoderm ovum, with small homogeneous nucleus
(e k).
Fig. 24—An
impregnated echinoderm ovum, with small homogeneous nucleus
(e k).
(From Hertwig.) |
Inside the impregnated ovum we now see a rapid series of most
important changes. The pear-shaped head of the sperm-cell, or the
“head of the spermatozoon,” grows larger and rounder,
and is converted into the male pro-nucleus (Fig.
26 s k). This has an attractive influence on the fine
granules or particles which are distributed in the protoplasm of
the ovum; they arrange themselves in lines in the figure of a star.
But the attraction or the “affinity” between the two
nuclei is even stronger. They move towards each other inside the
yelk with increasing speed, the male (Fig. 27
s k) going more quickly than the female nucleus (e
k). The tiny male nucleus takes with it the radiating mantle
which spreads like a star about it. At last the two sexual nuclei
touch (usually in the centre of the globular ovum), lie close
together, are flattened at the points of contact, and coalesce into
a common mass. The small central particle of
[ 56 ]
nuclein which is formed from this combination of the
nuclei is the stem-nucleus, or the first segmentation nucleus; the
new-formed cell, the product of the impregnation, is our stem-cell,
or “first segmentation sphere” (Fig. 2).

Fig. 25—Impregnation of the ovum
of a star-fish. (From Hertwig.) Only a small part of the
surface of the ovum is shown. One of the numerous spermatozoa
approaches the “impregnation rise” (A), touches
it (B), and then penetrates into the protoplasm of the ovum
(C). |
Hence the one essential point in the process of sexual
reproduction or impregnation is the formation of a new cell, the
stem-cell, by the combination of two originally different cells,
the female ovum and the male spermatozoon. This process is of the
highest importance, and merits our closest attention; all that
happens in the later development of this first cell and in the life
of the organism that comes of it is determined from the first by
the chemical and morphological composition of the stem-cell, its
nucleus and its body. We must, therefore, make a very careful study
of the rise and structure of the stem-cell.
The first question that arises is as to the two different active
elements, the nucleus and the protoplasm, in the actual
coalescence. It is obvious that the nucleus plays the more
important part in this. Hence Hertwig puts his theory of conception
in the principle: “Conception consists in the copulation of
two cell-nuclei, which come from a male and a female cell.”
And as the phenomenon of heredity is inseparably connected with the
reproductive process, we may further conclude that these two
copulating nuclei “convey the characteristics which are
transmitted from parents to offspring.” In this sense I had
in 1866 (in the ninth chapter of the General Morphology)
ascribed to the reproductive nucleus the function of generation and
heredity, and to the nutritive protoplasm the duties of
nutrition and adaptation. As, moreover, there is a complete
coalescence of the mutually attracted nuclear substances in
conception, and the new nucleus formed (the stem-nucleus) is the
real starting-point for the development of the fresh organism, the
further conclusion may be drawn that the male nucleus conveys to
the child the qualities of the father, and the female nucleus the
features of the mother. We must not forget, however, that the
protoplasmic bodies of the copulating cells also fuse together in
the act of impregnation; the cell-body of the invading spermatozoon
(the trunk and tail of the male ciliated cell) is dissolved in the
yelk of the female ovum. This coalescence is not so important as
that of the nuclei, but it must not be overlooked; and, though this
process is not so well known to us, we see clearly at least the
formation of the star-like figure (the radial arrangement of the
particles in the plasma) in it (Figs.
26–27).
The older theories of impregnation generally went astray in
regarding the large ovum as the sole base of the new organism, and
only ascribed to the spermatozoon the work of stimulating and
originating its development. The stimulus which it gave to the ovum
was sometimes thought to be purely chemical, at other times rather
physical (on the principle of transferred movement), or again a
mystic and transcendental process. This error was partly due to the
imperfect knowledge at that time of the facts of impregnation, and
partly to the striking
[ 57 ]
difference in the sizes of the two sexual cells.
Most of the earlier observers thought that the spermatozoon did not
penetrate into the ovum. And even when this had been demonstrated,
the spermatozoon was believed to disappear in the ovum without
leaving a trace. However, the splendid research made in the last
three decades with the finer technical methods of our time has
completely exposed the error of this. It has been shown that the
tiny sperm-cell is not subordinated to, but coordinated
with, the large ovum. The nuclei of the two cells, as the
vehicles of the hereditary features of the parents, are of equal
physiological importance. In some cases we have succeeded in
proving that the mass of the active nuclear substance which
combines in the copulation of the two sexual nuclei is originally
the same for both.
These morphological facts are in perfect harmony with the
familiar physiological truth that the child inherits from both
parents, and that on the average they are equally distributed. I
say “on the average,” because it is well known that a
child may have a greater likeness to the father or to the mother;
that goes without saying, as far as the primary sexual characters
(the sexual glands) are concerned. But it is also possible that the
determination of the latter—the weighty determination whether
the child is to be a boy or a girl—depends on a slight
qualitative or quantitative difference in the nuclein or the
coloured nuclear matter which comes from both parents in the act of
conception.
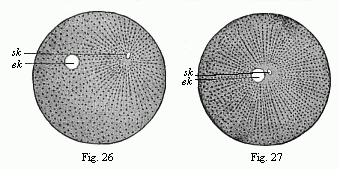
Figs. 26 and 27.—Impregnation of
the ovum of the sea-urchin. (From Hertwig.) In Fig. 26
the little sperm-nucleus (sk) moves towards the larger
nucleus of the ovum (ek). In Fig. 27 they nearly touch, and
are surrounded by the radiating mantle of protoplasm. |
The striking differences of the respective sexual cells in size
and shape, which occasioned the erroneous views of earlier
scientists, are easily explained on the principle of division of
labour. The inert, motionless ovum grows in size according to the
quantity of provision it stores up in the form of nutritive yelk
for the development of the germ. The active swimming sperm-cell is
reduced in size in proportion to its need to seek the ovum and bore
its way into its yelk. These differences are very conspicuous in
the higher animals, but they are much less in the lower animals. In
those protists (unicellular plants and animals) which have the
first rudiments of sexual reproduction the two copulating cells are
at first quite equal. In these cases the act of impregnation is
nothing more than a sudden growth, in which the originally
simple cell doubles its volume, and is thus prepared for
reproduction (cell-division). Afterwards slight differences are
seen in the size of the copulating cells; though the smaller ones
still have the same shape as the larger ones. It is only when the
difference in size is very pronounced that a notable difference in
shape is found: the sprightly sperm-cell changes more in shape and
the ovum in size.
Quite in harmony with this new conception of the equivalence
of the two gonads, or the equal physiological importance of the
male and female sex-cells and their equal share in the process of
heredity, is the important fact established by Hertwig (1875), that
in normal impregnation only one single spermatozoon
[ 58 ]
copulates with one ovum; the membrane which is
raised on the surface of the yelk immediately after one sperm-cell
has penetrated (Fig. 25 C) prevents
any others from entering. All the rivals of the fortunate
penetrator are excluded, and die without. But if the ovum passes
into a morbid state, if it is made stiff by a lowering of its
temperature or stupefied with narcotics (chloroform, morphia,
nicotine, etc.), two or more spermatozoa may penetrate into its
yelk-body. We then witness polyspermism. The more Hertwig
chloroformed the ovum, the more spermatozoa were able to bore their
way into its unconscious body.
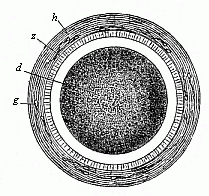 Fig. 28—Stem-cell of
a rabbit, magnified. In the centre of the granular protoplasm
of the fertilised ovum (d) is seen the little, bright
stem-nucleus, z is the ovolemma, with a mucous membrane
(h). s are dead spermatozoa.
Fig. 28—Stem-cell of
a rabbit, magnified. In the centre of the granular protoplasm
of the fertilised ovum (d) is seen the little, bright
stem-nucleus, z is the ovolemma, with a mucous membrane
(h). s are dead spermatozoa. |
These remarkable facts of impregnation are also of the greatest
interest in psychology, especially as regards the theory of the
cell-soul, which I consider to be its chief foundation. The
phenomena we have described can only be understood and explained by
ascribing a certain lower degree of psychic activity to the sexual
principles. They feel each other’s proximity, and are
drawn together by a sensitive impulse (probably related to
smell); they move towards each other, and do not rest until
they fuse together. Physiologists may say that it is only a
question of a peculiar physico-chemical phenomenon, and not a
psychic action; but the two cannot be separated. Even the psychic
functions, in the strict sense of the word, are only complex
physical processes, or “psycho-physical” phenomena,
which are determined in all cases exclusively by the chemical
composition of their material substratum.
The monistic view of the matter becomes clear enough when we
remember the radical importance of impregnation as regards
heredity. It is well known that not only the most delicate bodily
structures, but also the subtlest traits of mind, are transmitted
from the parents to the children. In this the chromatic matter of
the male nucleus is just as important a vehicle as the large
caryoplasmic substance of the female nucleus; the one transmits the
mental features of the father, and the other those of the mother.
The blending of the two parental nuclei determines the individual
psychic character of the child.
But there is another important psychological question—the
most important of all—that has been definitely answered by
the recent discoveries in connection with conception. This is the
question of the immortality of the soul. No fact throws more light
on it and refutes it more convincingly than the elementary process
of conception that we have described. For this copulation of the
two sexual nuclei (Figs. 26 and 27)
indicates the precise moment at which the individual begins to
exist. All the bodily and mental features of the new-born child are
the sum-total of the hereditary qualities which it has received in
reproduction from parents and ancestors. All that man acquires
afterwards in life by the exercise of his organs, the influence of
his environment, and education—in a word, by
adaptation—cannot obliterate that general outline of his
being which he inherited from his parents. But this hereditary
disposition, the essence of every human soul, is not
“eternal,” but “temporal”; it comes into
being only at the moment when the sperm-nucleus of the father and
the nucleus of the maternal ovum meet and fuse together. It is
clearly irrational to assume an “eternal life without
end” for an individual phenomenon, the commencement of which
we can indicate to a moment by direct visual observation.
The great importance of the process of impregnation in answering
such questions is quite clear. It is true that conception has never
been studied microscopically in all its details in the human
case—notwithstanding its occurrence at every moment—for
reasons that are
[ 59 ]
obvious enough. However, the two cells which need
consideration, the female ovum and the male spermatozoon, proceed
in the case of man in just the same way as in all the other
mammals; the human fœtus or embryo which results from
copulation has the same form as with the other animals. Hence, no
scientist who is acquainted with the facts doubts that the
processes of impregnation are just the same in man as in the other
animals.
The stem-cell which is produced, and with which every man begins
his career, cannot be distinguished in appearance from those of
other mammals, such as the rabbit (Fig. 28). In the case of man,
also, this stem-cell differs materially from the original ovum,
both in regard to form (morphologically), in regard to material
composition (chemically), and in regard to vital properties
(physiologically). It comes partly from the father and partly from
the mother. Hence it is not surprising that the child who is
developed from it inherits from both parents. The vital movements
of each of these cells form a sum of mechanical processes which in
the last analysis are due to movements of the smallest vital parts,
or the molecules, of the living substance. If we agree to call this
active substance plasson, and its molecules
plastidules, we may say that the individual physiological
character of each of these cells is due to its molecular
plastidule-movement. Hence, the plastidule-movement of the
cytula is the resultant of the combined plastidule-movements of the
female ovum and the male sperm-cell.1
1. The plasson of the stem-cell or cytula may, from
the anatomical point of view, be regarded as homogeneous and
structureless, like that of the monera. This is not inconsistent
with our hypothetical ascription to the plastidules (or molecules
of the plasson) of a complex molecular structure. The complexity of
this is the greater in proportion to the complexity of the organism
that is developed from it and the length of the chain of its
ancestry, or to the multitude of antecedent processes of heredity
and adaptation.
Title and Contents
Glossary
Chapter VI
Chapter VIII
Figs. 1–209
Figs. 210–408