THE EVOLUTION OF MAN
Volume II
CHAPTER XXIV
EVOLUTION OF THE NERVOUS SYSTEM
The previous chapters have taught us how the human body as a whole develops from the first simple rudiment, a single layer of cells. The whole human race owes its origin, like the individual man, to a simple cell. The unicellular stem-form of the race is reproduced daily in the unicellular embryonic stage of the individual. We have now to consider in detail the evolution of the various parts that make up the human frame. I must, naturally, confine myself to the most general and principal outlines; to make a special study of the evolution of each organ and tissue is both beyond the scope of this work, and probably beyond the anatomic capacity of most of my readers to appreciate. In tracing the evolution of the various organs we shall follow the method that has hitherto guided us, except that we shall now have to consider the ontogeny and phylogeny of the organs together. We have seen, in studying the evolution of the body as a whole, that phylogeny casts a light over the darker paths of ontogeny, and that we should be almost unable to find our way in it without the aid of the former. We shall have the same experience in the study of the organs in detail, and I shall be compelled to give simultaneously their ontogenetic and phylogenetic origin. The more we go into the details of organic development, and the more closely we follow the rise of the various parts, the more we see the inseparable connection of embryology and stem-history. The ontogeny of the organs can only be understood in the light of their phylogeny, just as we found of the embryology of the whole body. Each embryonic form is determined by a corresponding stem-form. This is true of details as well as of the whole.
We will consider first the animal and then the vegetal systems of organs of the body. The first group consists of the psychic and the motor apparatus. To the former belong the skin, the nervous system, and the sense-organs. The motor apparatus is composed of the passive and the active organs of movement (the skeleton and the muscles). The second or vegetal group consists of the nutritive and the reproductive apparatus. To the nutritive apparatus belong the alimentary canal with all its appendages, the vascular system, and the renal (kidney) system. The reproductive apparatus comprises the different organs of sex (embryonic glands, sexual ducts, and copulative organs).
As we know from previous chapters (XI–XIII), the animal systems of organs (the organs of sensation and presentation) develop for the most part out of the outer primary germ-layer, or the cutaneous (skin) layer. On the other hand, the vegetal systems of organs arise for the most part from the inner primary germ-layer, the visceral layer. It is true that this antithesis of the animal and vegetal spheres of the body in man and all the higher animals is by no means rigid; several parts of the animal apparatus (for instance, the greater part of the muscles) are formed from cells that come originally from the entoderm; and a great part of the vegetative apparatus (for instance, the mouth-cavity and the gonoducts) are composed of cells that come from the ectoderm.
In the more advanced animal body there is so much interlacing and displacement of the various parts that it is often very difficult to indicate the sources of them. But, broadly speaking, we may take it as a positive and important fact that in man and the higher animals the chief part of the animal organs comes from the ectoderm, and the greater part of the vegetative organs from the entoderm. It was for this reason that Carl Ernst von Baer called the one the animal and the other the vegetative layer (see p. 16).
The solid foundation of this important thesis is the gastrula, the most instructive embryonic form in the animal world, which we still find in the same shape in the most diverse classes of animals. This form points demonstrably to a
[ 267 ]
common stem-form of all the Metazoa, the Gastræa; in this long-extinct stem-form the whole body consisted throughout life of the two primary germinal layers, as is now the case temporarily in the gastrula; in the Gastræa the simple cutaneous (skin) layer actually represented all the animal organs and functions, and the simple visceral (gut) layer all the vegetal organs and functions. This is the case with the modern Gastræads (Fig. 233); and it is also the case potentially with the gastrula.
We shall easily see that the gastræa theory is thus able to throw a good deal of light, both morphologically and physiologically, on some of the chief features of embryonic development, if we take up first the consideration of the chief element in the animal sphere, the psychic apparatus or sensorium and its evolution. This apparatus consists of two very different parts, which seem at first to have very little connection with each other—the outer skin, with all its hairs, nails, sweat-glands, etc., and the nervous system. The latter comprises the central nervous system (brain and spinal cord), the peripheral, cerebral, and spinal nerves, and the sense-organs. In the fully-formed vertebrate body these two chief elements of the sensorium lie far apart, the skin being external to, and the central nervous system in the very centre of, the body. The one is only connected with the other by a section of the peripheral nervous system and the sense-organs. Nevertheless, as we know from human embryology, the medullary tube is formed from the cutaneous layer. The organs that discharge the most advanced functions of the animal body—the organs of the soul, or of psychic life—develop from the external skin. This is a perfectly natural and necessary process. If we reflect on the historical evolution of the psychic and sensory functions, we are forced to conclude that the cells which accomplish them must originally have been located on the outer surface of the body. Only elementary organs in this superficial position could directly receive the influences of the environment. Afterwards, under the influence of natural selection, the cellular group in the skin which was specifically “sensitive” withdrew into the inner and more protected part of the body, and formed there the foundation of a central nervous organ. As a result of increased differentiation, the skin and the central nervous system became further and further separated, and in the end the two were only permanently connected by the afferent peripheral sensory nerves.
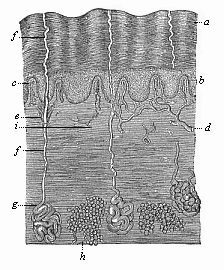 Fig. 284—The human skin in vertical section (from Ecker), highly magnified, a horny layer of the epidermis, b mucous layer of the epidermis, c papillæ of the corium, d blood-vessels of same, ef ducts of the sweat-glands (g), h fat-glands in the corium, i nerve, passing into a tactile corpuscle above.
Fig. 284—The human skin in vertical section (from Ecker), highly magnified, a horny layer of the epidermis, b mucous layer of the epidermis, c papillæ of the corium, d blood-vessels of same, ef ducts of the sweat-glands (g), h fat-glands in the corium, i nerve, passing into a tactile corpuscle above. |
The observations of the comparative anatomist are in complete accord with this view. He tells us that large numbers of the lower animals have no nervous system, though they exercise the functions of sensation and will like the higher animals. In the unicellular Protozoa, which do not form germinal layers, there is, of course, neither nervous system nor skin. But in the second division of the animal kingdom also, the Metazoa, there is at first no nervous system. Its functions are represented by the simple cell-layer of the ectoderm, which the lower Metazoa have inherited from the Gastræa (Fig. 30 e). We find this in the lowest Zoophytes—the Gastræads, Physemaria, and Sponges (Figs. 233–238). The lowest Cnidaria (the hydroid polyps) also are little superior to the Gastræads in structure. Their vegetative functions are accomplished by the simple visceral layer, and their animal functions by the simple cutaneous layer. In these
[ 268 ]
cases the simple cell-layer of the ectoderm is at once skin, locomotive apparatus, and nervous system.
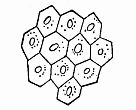 Fig. 285—Epidermic cells of a human embryo of two months. (From Kölliker.)
Fig. 285—Epidermic cells of a human embryo of two months. (From Kölliker.) |
When we come to the higher Metazoa, in which the sensory functions and their organs are more advanced, we find a division of labour among the ectodermic cells. Groups of sensitive nerve cells separate from the ordinary epidermic cells; they retire into the more protected tissue of the mesodermic under-skin, and form special neural ganglia there. Even in the Platodes, especially the Turbellaria, we find an independent nervous system, which has separated from the outer skin. This is the “upper pharyngeal ganglion,” or acroganglion, situated above the gullet (Fig. 241 g). From this rudimentary structure has been developed the elaborate central nervous system of the higher animals. In some of the higher worms, such as the earth-worm, the first rudiment of the central nervous system (Fig. 74 n) is a local thickening of the skin-sense layer (hs), which afterwards separates altogether from the horny plate. In the earliest Platodes (Cryptocœla) and Vermalia (Gastrotricha) the acroganglion remains in the epidermis. But the medullary tube of the Vertebrates originates in the same way. Our embryology has taught us that this first structure of the central nervous system also develops originally from the outer germinal layer.
Let us now examine more closely the evolution of the human skin, with its various appendages, the hairs and glands. This external covering has, physiologically, a double and important part to play. It is, in the first place, the common integument that covers the whole surface of the body, and forms a protective envelope for the other organs. As such it also effects a certain exchange of matter between the body and the surrounding atmosphere (exhalation, perspiration). In the second place, it is the earliest and original sense organ, the common organ of feeling that experiences the sensation of the temperature of the environment and the pressure or resistance of bodies that come into contact.
The human skin (like that of all the higher animals) is composed of two layers, the outer and the inner or underlying skin. The outer skin or epidermis, consists of simple ectodermic cells, and contains no blood-vessels (Fig. 284 a, b). It develops from the outer germinal layer, or skin-sense layer. The underlying skin (corium or hypodermis) consists chiefly of connective tissue, contains numerous blood-vessels and nerves, and has a totally different origin. It comes from the outermost parietal stratum of the middle germinal layer, or the skin-fibre layer. The corium is much thicker than the epidermis. In its deeper strata (the subcutis) there are clusters of fat-cells (Fig. 284 h). Its uppermost stratum (the cutis proper, or the papillary stratum) forms, over almost the whole surface of the body, a number of conical microscopic papillæ (something like warts), which push into the overlying epidermis (c). These tactile or sensory particles contain the finest sensory organs of the skin, the touch corpuscles. Others contain merely end-loops of the blood-vessels that nourish the skin (c, d). The various parts of the corium arise by division of labour from the originally homogeneous cells of the cutis-plate, the outermost lamina of the mesodermic skin-fibre layer (Fig. 145 hpr, and Figs. 161, 162 cp).
In the same way, all the parts and appendages of the epidermis develop by differentiation from the homogeneous cells of this horny plate (Fig. 285). At an early stage the simple cellular layer of this horny plate divides into two. The inner and softer stratum (Fig. 284 b) is known as the mucous stratum, the outer and harder (a) as the horny (corneous) stratum. This horny layer is being constantly used up and rubbed away at the surface; new layers of cells grow up in their place out of the underlying mucous stratum. At first the epidermis is a simple covering of the surface of the body. Afterwards various appendages develop from it, some internally, others externally. The internal appendages are the cutaneous glands—sweat, fat, etc.
[ 269 ]
The external appendages are the hairs and nails.
The cutaneous glands are originally merely solid cone-shaped growths of the epidermis, which sink into the underlying corium (Fig. 286 1). Afterwards a canal (2, 3) is formed inside them, either by the softening and dissolution of the central cells or by the secretion of fluid internally. Some of the glands, such as the sudoriferous, do not ramify (Fig. 284 efg). These glands, which secrete the perspiration, are very long, and have a spiral coil at the end, but they never ramify; so also the wax-glands of the ears. Most of the other cutaneous glands give out buds and ramify; thus, for instance, the lachrymal glands of the upper eye-lid that secrete tears (Fig. 286), and the sebaceous glands which secrete the fat in the skin and generally open into the hair-follicles. Sudoriferous and sebaceous glands are found only in mammals. But we find lachrymal glands in all the three classes of Amniotes—reptiles, birds, and mammals. They are wanting in the lower aquatic vertebrates.
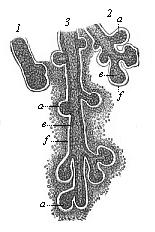 Fig. 286—Rudimentary lachrymal glands from a human embryo of four months. (From Kölliker.) 1 earliest structure, in the shape of a simple solid cone, 2 and 3 more advanced structures, ramifying and hollowing out. a solid buds, e cellular coat of the hollow buds, f structure of the fibrous envelope, which afterwards forms the corium about the glands.
Fig. 286—Rudimentary lachrymal glands from a human embryo of four months. (From Kölliker.) 1 earliest structure, in the shape of a simple solid cone, 2 and 3 more advanced structures, ramifying and hollowing out. a solid buds, e cellular coat of the hollow buds, f structure of the fibrous envelope, which afterwards forms the corium about the glands. |
The mammary glands (Figs. 287, 288) are very remarkable; they are found in all mammals, and in these alone. They secrete the milk for the feeding of the new-born mammal. In spite of their unusual size these structures are nothing more than large sebaceous glands in the skin. The milk is formed by the liquefaction of the fatty milk-cells inside the branching mammary-gland tubes (Fig. 287 c), in the same way as the skin-grease or hair-fat, by the solution of fatty cells inside the sebaceous glands. The outlets of the mammary glands enlarge and form sac-like mammary ducts (b); these narrow again (a), and open in the teats or nipples of the breast by sixteen to twenty-four fine apertures. The first structure of this large and elaborate gland is a very simple cone in the epidermis, which penetrates into the corium and ramifies. In the new-born infant it consists of twelve to eighteen radiating lobes (Fig. 288). These gradually ramify, their ducts become hollow and larger, and rich masses of fat accumulate between the lobes. Thus is formed the prominent female breast (mamma), on the top of which rises the teat or nipple (mammilla). The latter is only developed later on, when the mammary gland is fully-formed; and this ontogenetic phenomenon is extremely interesting, because the earlier mammals (the stem-forms of the whole class) have no teats. In them the milk comes out through a flat portion of the ventral skin that is pierced like a sieve, as we still find in the lowest living mammals, the oviparous Monotremes of Australia. The young animal licks the milk from the mother instead of sucking it. In many of the lower mammals we find a number of milk-glands at different parts of the ventral surface. In the human female there is usually only one pair of glands, at the breast; and it is the same with the apes, bats, elephants, and several other mammals. Sometimes, however, we find two successive pairs of glands (or even more) in the human female. Some women have four or five pairs of breasts, like pigs and hedgehogs (Fig. 103). This polymastism points back to an older stem-form. We often find these accessory breasts in the male also (Fig. 103 D). Sometimes, moreover, the normal mammary glands are fully developed and can suckle in the male; but as a rule they are merely rudimentary organs without functions in the male. We have already (Chapter XI) dealt with this remarkable and interesting instance of atavism.
While the cutaneous glands are inner growths of the epidermis, the appendages
[ 270 ]
which we call hairs and nails are external local growths in it. The nails (Ungues) which form important protective structures on the back of the most sensitive parts of our limbs, the tips of the fingers and toes, are horny growths of the epidermis, which we share with the apes. The lower mammals usually have claws instead of them; the ungulates, hoofs. The stem-form of the mammals certainly had claws; we find them in a rudimentary form even in the salamander. The horny claws are highly developed in most of the reptiles (Fig. 264), and the mammals have inherited them from the earliest representatives of this class, the stem-reptiles (Tocosauria). Like the hoofs (ungulæ) of the Ungulates, the nails of apes and men have been evolved from the claws of the older mammals. In the human embryo the first rudiment of the nails is found (between the horny and the mucous stratum of the epidermis) in the fourth month. But their edges do not penetrate through until the end of the sixth month.
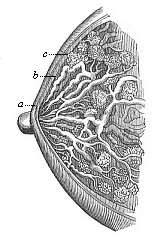 Fig. 287—The female breast (mamma) in vertical section. c racemose glandular lobes, b enlarged milk-ducts, a narrower outlets, which open into the nipple. (From H. Meyer.)
Fig. 287—The female breast (mamma) in vertical section. c racemose glandular lobes, b enlarged milk-ducts, a narrower outlets, which open into the nipple. (From H. Meyer.) |
The most interesting and important appendages of the epidermis are the hairs; on account of their peculiar composition and origin we must regard them as highly characteristic of the whole mammalian class. It is true that we also find hairs in many of the lower animals, such as insects and worms. But these hairs, like the hairs of plants, are thread-like appendages of the surface, and differ entirely from the hairs of the mammals in the details of their structure and development.
The embryology of the hairs is known in all its details, but there are two different views as to their phylogeny. On the older view the hairs of the mammals are equivalent or homologous to the feathers of the bird or the horny scales of the reptile. As we deduce all three classes of Amniotes from a common stem-group, we must assume that these Permian stem-reptiles had a complete scaly coat, inherited from their Carboniferous ancestors, the mailed amphibia (Stegocephala); the bony scales of their corium were covered with horny scales. In passing from aquatic to terrestrial life the horny scales were further developed, and the bony scales degenerated in most of the reptiles. As regards the bird’s feathers, it is certain that they are modifications of the horny scales of their reptilian ancestors. But it is otherwise with the hairs of the mammals. In their case the hypothesis has lately been advanced on the strength of very extensive research, especially by Friedrich Maurer, that they have been evolved from the cutaneous sense-organs of amphibian ancestors by modification of functions; the epidermic structure is very similar in both in its embryonic rudiments. This modern view, which had the support of the greatest expert on the vertebrates, Carl Gegenbaur, can be harmonised with the older theory to an extent, in the sense that both formations, scales and hairs, were very closely connected originally. Probably the conical budding of the skin-sense layer grew up under the protection of the horny scale, and became an organ of touch subsequently by the cornification of the hairs; many hairs are still sensory organs (tactile hairs on the muzzle and cheeks of many mammals: pubic hairs).
This middle position of the genetic connection of scales and hairs was advanced in my Systematic Phylogeny of the Vertebrates (p. 433). It is confirmed by the similar arrangement of the two cutaneous formations. As Maurer pointed out, the hairs, as well as the cutaneous sense-organs and the scales, are at first arranged in regular longitudinal series, and they afterwards break into alternate groups. In the embryo of a bear two
[ 271 ]
inches long, which I owe to the kindness of Herr von Schmertzing (of Arva Varallia, Hungary), the back is covered with sixteen to twenty alternating longitudinal rows of scaly protuberances (Fig. 289). They are at the same time arranged in regular transverse rows, which converge at an acute angle from both sides towards the middle of the back. The tip of the scale-like wart is turned inwards. Between these larger hard scales (or groups of hairs) we find numbers of rudimentary smaller hairs.
The human embryo is, as a rule, entirely clothed with a thick coat of fine wool during the last three or four weeks of gestation. This embryonic woollen coat (Lanugo) generally disappears in part during the last weeks of fœtal life but in any case, as a rule, it is lost immediately after birth, and is replaced by the thinner coat of the permanent hair. These permanent hairs grow out of hair-follicles, which are formed from the root-sheaths of the disappearing wool-fibres. The embryonic wool-coat usually, in the case of the human embryo, covers the whole body, with the exception of the palms of the hands and soles of the feet. These parts are always bare, as in the case of apes and of most other mammals. Sometimes the wool-coat of the embryo has a striking effect, by its colour, on the later permanent hair-coat. Hence it happens occasionally, for instance, among our Indo-Germanic races, that children of blond parents seem—to the dismay of the latter—to be covered at birth with a dark brown or even a black woolly coat. Not until this has disappeared do we see the permanent blond hair which the child has inherited. Sometimes the darker coat remains for weeks, and even months, after birth. This remarkable woolly coat of the human embryo is a legacy from the apes, our ancient long-haired ancestors.
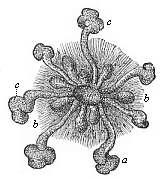 Fig. 288—Mammary gland of a new-born infant, a original central gland, b small and c large buds of same. (From Langer.)
Fig. 288—Mammary gland of a new-born infant, a original central gland, b small and c large buds of same. (From Langer.) |
It is not less noteworthy that many of the higher apes approach man in the thinness of the hair on various parts of the body. With most of the apes, especially the higher Catarrhines (or narrow-nosed apes), the face is mostly, or entirely, bare, or at least it has hair no longer or thicker than that of man. In their case, too, the back of the head is usually provided with a thicker growth of hair; this is lacking, however, in the case of the bald-headed chimpanzee (Anthropithecus calvus). The males of many species of apes have a considerable beard on the cheeks and chin; this sign of the masculine sex has been acquired by sexual selection. Many species of apes have a very thin covering of hair on the breast and the upper side of the limbs—much thinner than on the back or the under side of the limbs. On the other hand, we are often astonished to find tufts of hair on the shoulders, back, and extremities of members of our Indo-Germanic and of the Semitic races. Exceptional hair on the face, as on the whole body, is hereditary in certain families of hairy men. The quantity and the quality of the hair on head and chin are also conspicuously transmitted in families. These extraordinary variations in the total and partial hairy coat of the body, which are so noticeable, not only in comparing different races of men, but also in comparing different families of the same race, can only be explained on the assumption that in man the hairy coat is, on the whole, a rudimentary organ, a useless inheritance from the more thickly-coated apes. In this man resembles the elephant, rhinoceros, hippopotamus, whale, and other mammals of various orders, which have also, almost entirely or for the most part, lost their hairy coats by adaptation.
The particular process of adaptation by which man lost the growth of hair on most parts of his body, and retained or augmented it at some points, was most probably sexual selection. As Darwin luminously showed in his Descent of Man, sexual selection has been very active
[ 272 ]
in this respect. As the male anthropoid apes chose the females with the least hair, and the females favoured the males with the finest growths on chin and head, the general coating of the body gradually degenerated, and the hair of the beard and head was more strongly developed. The growth of hair at other parts of the body (arm-pit, pubic region) was also probably due to sexual selection. Moreover, changes of climate, or habits, and other adaptations unknown to us, may have assisted the disappearance of the hairy coat.
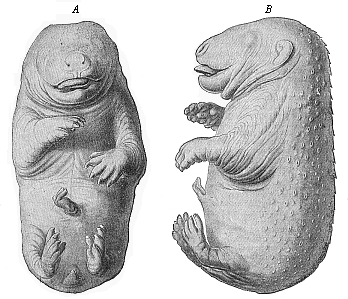 Fig. 289—Embryo of a bear (Ursus arctos). A seen from ventral side, B from the left.
Fig. 289—Embryo of a bear (Ursus arctos). A seen from ventral side, B from the left. |
The fact that our coat of hair is inherited directly from the anthropoid apes is proved in an interesting way, according to Darwin, by the direction of the rudimentary hairs on our arms, which cannot be explained in any other way. Both on the upper and the lower part of the arm they point towards the elbow. Here they meet at an obtuse angle. This curious arrangement is found only in the anthropoid apes—gorilla, chimpanzee, orang, and several species of gibbons—besides man (Figs. 203, 207). In other species of gibbon the hairs are pointed towards the hand both in the upper and lower arm, as in the rest of the mammals. We can easily explain this remarkable peculiarity of the anthropoids and man on the theory that our common ancestors were accustomed (as the anthropoid apes are to-day) to place their hands over their heads, or across a branch above their heads, during rain. In this position, the fact that the hairs point downwards helps the rain to run off. Thus the direction of the hair on the lower part of our arm reminds us to-day of that useful custom of our anthropoid ancestors.
The nervous system in man and all the other Vertebrates is, when fully formed, an extremely complex apparatus, that we may compare, in anatomic structure and physiological function, with an extensive telegraphic system. The chief station of
[ 273 ]
the system is the central marrow or central nervous system, the innumerable ganglionic cells or neurona (Fig. 9) of which are connected by branching processes with each other and with numbers of very fine conducting wires. The latter are the peripheral and ubiquitous nerve-fibres; with their terminal apparatus, the sense-organs, etc., they constitute the conducting marrow or peripheral nervous system. Some of them—the sensory nerve-fibres—conduct the impressions from the skin and other sense-organs to the central marrow; others—the motor nerve-fibres—convey the commands of the will to the muscles.
The central nervous system or central marrow (medulla centralis) is the real organ of psychic action in the narrower sense. However we conceive the intimate connection of this organ and its functions, it is certain that its characteristic actions, which we call sensation, will, and thought, are inseparably dependent on the normal development of the material organ in man and all the higher animals. We must, therefore, pay particular attention to the evolution of the latter. As it can give us most important information regarding the nature of the “soul,” it should be full of interest. If the central marrow develops in just the same way in the human embryo as in the embryo of the other mammals, the evolution of the human psychic organ from the central organ of the other mammals, and through them from the lower vertebrates, must be beyond question. No one can doubt the momentous bearing of these embryonic phenomena.
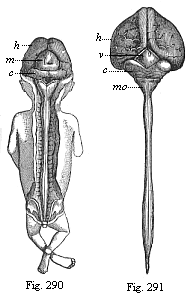 Fig. 290—Human embryo, three months old, from the dorsal side: brain and spinal cord exposed. (From Kölliker.) h cerebral hemispheres (fore brain), m corpora quadrigemina (middle brain), c cerebellum (hind brain): under the latter is the triangular medulla oblongata (after brain).
Fig. 290—Human embryo, three months old, from the dorsal side: brain and spinal cord exposed. (From Kölliker.) h cerebral hemispheres (fore brain), m corpora quadrigemina (middle brain), c cerebellum (hind brain): under the latter is the triangular medulla oblongata (after brain).
Fig. 291—Central marrow of a human embryo, four months old, from the back. (From Kölliker.) h large hemispheres, v quadrigemina, c cerebellum, mo medulla oblongata: underneath it the spinal cord. |
In order to understand them fully we must first say a word or two of the general form and the anatomic composition of the mature human central marrow. Like the central nervous system of all the other Craniotes, it consists of two parts, the head-marrow or brain (medulla capitis or encephalon) and the spinal-marrow (medulla spinalis or notomyelon). The one is enclosed in the bony skull, the other in the bony vertebral column. Twelve pairs of cerebral nerves proceed from the brain, and thirty-one pairs of spinal nerves from the spinal cord, to the rest of the body (Fig. 171). On general anatomic investigation the spinal marrow is found to be a cylindrical cord, with a spindle-shaped bulb both in the region of the neck above (at the last cervical vertebra) and the region of the loins (at the first lumbar vertebra) below (Fig. 291). At the cervical bulb the strong nerves of the upper limbs, and at the lumbar bulb those of the lower limbs, proceed from the spinal cord. Above, the latter passes into the brain through the medulla oblongata (Fig. 291 mo). The spinal cord seems to be a thick mass of nervous matter, but it has a narrow canal at its axis, which passes into the further cerebral ventricles above, and is filled, like these, with a clear fluid.
The brain is a large nerve-mass, occupying the greater part of the skull, of most elaborate structure. On general examination it divides into two parts, the cerebrum and cerebellum. The cerebrum lies in front and above, and has the familiar characteristic convolutions and furrows on its surface (Figs. 292, 293). On the upper side it is divided by a deep longitudinal fissure into two halves, the
[ 274 ]
cerebral hemispheres; these are connected by the corpus callosum. The large cerebrum is separated from the small cerebellum by a deep transverse furrow. The latter lies behind and below, and has also numbers of furrows, but much finer and more regular, with convolutions between, at its surface. The cerebellum also is divided by a longitudinal fissure into two halves, the “small hemispheres”; these are connected by a worm-shaped piece, the vermis cerebelli, above, and by the broad pons Varolii below (Fig. 292 VI).
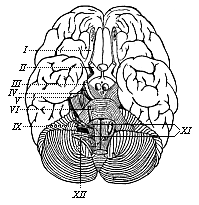 Fig. 292—The human brain, seen from below. (From H. Meyer.) Above (in front) is the cerebrum with its extensive branching furrows; below (behind) the cerebellum with its narrow parallel furrows. The Roman numbers indicate the roots of the twelve pairs of cerebral nerves in a series towards the rear.
Fig. 292—The human brain, seen from below. (From H. Meyer.) Above (in front) is the cerebrum with its extensive branching furrows; below (behind) the cerebellum with its narrow parallel furrows. The Roman numbers indicate the roots of the twelve pairs of cerebral nerves in a series towards the rear. |
But comparative anatomy and ontogeny teach us that in man and all the other Craniotes the brain is at first composed, not of these two, but of three, and afterwards five, consecutive parts. These are found in just the same form—as five consecutive vesicles—in the embryo of all the Craniotes, from the Cyclostoma and fishes to man. But, however much they agree in their rudimentary condition, they differ considerably afterwards. In man and the higher mammals the first of these ventricles, the cerebrum, grows so much that in its mature condition it is by far the largest and heaviest part of the brain. To it belong not only the large hemispheres, but also the corpus callosum that unites them, the olfactory lobes, from which the olfactory nerves start, and most of the structures that are found at the roof and bottom of the large lateral ventricles inside the two hemispheres, such as the corpora striata. On the other hand, the optic thalami, which lie between the latter, belong to the second division, which develops from the “intermediate brain ”; to the same section belong the single third cerebral ventricle and the structures that are known as the corpora geniculata, the infundibulum, and the pineal gland. Behind these parts we find, between the cerebrum and cerebellum, a small ganglion composed of two prominences, which is called the corpus quadrigeminum on account of a superficial transverse fissure cutting across (Figs. 290 m and 291 v). Although this quadrigeminum is very insignificant in man and the higher mammals, it forms a special third section, greatly developed in the lower vertebrates, the “middle brain.” The fourth section is the “hind-brain” or little brain (cerebellum) in the narrower sense, with the single median part, the vermis, and the pair of lateral parts, the “small hemispheres” (Fig. 291 c). Finally, we have the fifth and last section, the medulla oblongata (Fig. 291 mo), which contains the single fourth cerebral cavity and the contiguous parts (pyramids, olivary bodies, corpora restiformia). The medulla oblongata passes straight into the medulla spinalis (spinal cord). The narrow central canal of the spinal cord continues above into the quadrangular fourth cerebral cavity of the medulla oblongata, the floor of which is the quadrangular depression. From here a narrow duct, called “the aqueduct of Sylvius,” passes through the corpus quadrigeminum to the third cerebral ventricle, which lies between the two optic thalami; and this in turn is connected with the pairs of lateral ventricles which lie to the right and left in the large hemispheres. Thus all the cavities of the central marrow are directly interconnected. All these parts of the brain have an infinitely complex structure in detail, but we cannot go into this. Although it is much more elaborate in man and the higher Vertebrates than in the lower classes, it develops in them all from the same rudimentary structure, the five simple cerebral vesicles of the embryonic brain.
But before we consider the development of the complicated structure of the brain from this simple series of vesicles, let
[ 275 ]
us glance for a moment at the lower animals, which have no brain. Even in the skull-less vertebrate, the Amphioxus, we find no independent brain, as we have seen. The whole central marrow is merely a simple cylindrical cord which runs the length of the body, and ends equally simply at both extremities—a plain medullary tube. All that we can discover is a small vesicular bulb at the foremost part of the tube, a degenerate rudiment of a primitive brain. We meet the same simple medullary tube in the first structure of the ascidia larva, in the same characteristic position, above the chorda. On closer examination we find here also a small vesicular swelling at the fore end of the tube, the first trace of a differentiation of it into brain and spinal cord. It is probable that this differentiation was more advanced in the extinct Provertebrates, and the brain-bulb more pronounced (Figs. 98–102). The brain is phylogenetically older than the spinal cord, as the trunk was not developed until after the head. If we consider the undeniable affinity of the Ascidiæ to the Vermalia, and remember that we can trace all the Chordonia to lower Vermalia, it seems probable that the simple central marrow of the former is equivalent to the simple nervous ganglion, which lies above the gullet in the lower worms, and has long been known as the “upper pharyngeal ganglion” (ganglion pharyngeum superius); it would be better to call it the primitive or vertical brain (acroganglion).
Probably this upper pharyngeal ganglion of the lower worms is the structure from which the complex central marrow of the higher animals has been evolved. The medullary tube of the Chordonia has been formed by the lengthening of the vertical brain on the dorsal side. In all the other animals the central nervous system has been developed in a totally different way from the upper pharyngeal ganglion; in the Articulates, especially, a pharyngeal ring, with ventral marrow, has been added. The Molluscs also have a pharyngeal ring, but it is not found in the Vertebrates. In these the central marrow has been prolonged down the dorsal side; in the Articulates down the ventral side. This fact proves of itself that there is no direct relationship between the Vertebrates and the Articulates. The unfortunate attempts to derive the dorsal marrow of the former from the ventral marrow of the latter have totally failed (cf. p. 219).
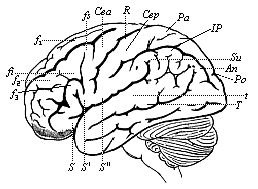 Fig. 293—The human brain, seen from the left. (From H. Meyer.) The furrows of the cerebrum are indicated by thick, and those of the cerebellum by finer lines. Under the latter we can see the medulla oblongata. f1–f2 frontal convolutions, C central convolutions, S fissure of Sylvius, T temporal furrow, Pa parietal lobes, An angular gyrus, Po parieto-occipital fissure.
Fig. 293—The human brain, seen from the left. (From H. Meyer.) The furrows of the cerebrum are indicated by thick, and those of the cerebellum by finer lines. Under the latter we can see the medulla oblongata. f1–f2 frontal convolutions, C central convolutions, S fissure of Sylvius, T temporal furrow, Pa parietal lobes, An angular gyrus, Po parieto-occipital fissure. |
When we examine the embryology of the human nervous system, we must start from the important fact, which we have already seen, that the first structure of it in man and all the higher Vertebrates is the simple medullary tube, and that this separates from the outer germinal layer in the middle line of the sole-shaped embryonic shield. As the reader will remember, the straight medullary furrow first appears in the middle of the sandal-shaped embryonic shield. At each side of it the parallel borders curve over in the form of dorsal or medullary swellings. These bend together with their free borders, and thus form the closed medullary tube (Figs. 133–137). At first this tube lies directly underneath the horny plate; but it afterwards travels inwards, the upper edges of the provertebral plates growing together between the horny plate and the tube, joining above the latter, and forming a completely closed canal. As Gegenbaur very properly observes, “this gradual imbedding in the
[ 276 ]
inner part of the body is a process acquired with the progressive differentiation and the higher potentiality that this secures; by this process the organ of greater value to the organism is buried within the frame.” (Cf. Figs. 143–146).
In the Cyclostoma—a stage above the Acrania—the fore end of the cylindrical medullary tube begins early to expand into a pear-shaped vesicle; this is the first outline of an independent brain. In this way the central marrow of the Vertebrates divides clearly into its two chief sections, brain and spinal cord. The simple vesicular form of the brain, which persists for some time in the Cyclostoma, is found also at first in all the higher Vertebrates (Fig. 153 hb). But in these it soon passes away, the one vesicle being divided into several successive parts by transverse constrictions. There are first two of these constrictions, dividing the brain into three consecutive vesicles (fore brain, middle brain, and hind brain, Fig. 154 v, m, h). Then the first and third are sub-divided by fresh constrictions, and thus we get five successive sections (Fig. 155).
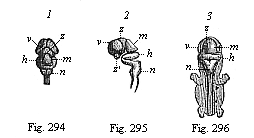 Fig. 294–296—Central marrow of the human embryo from the seventh week, 4/5 inch long. (From Kölliker.) Fig. 294. The brain from above, v fore brain, z intermediate brain, m middle brain, h hind brain, n after brain. Fig. 2955. The brain with the uppermost part of the cord, from the left. Fig. 296. Back view of the whole embryo: brain and spinal cord exposed.
Fig. 294–296—Central marrow of the human embryo from the seventh week, 4/5 inch long. (From Kölliker.) Fig. 294. The brain from above, v fore brain, z intermediate brain, m middle brain, h hind brain, n after brain. Fig. 2955. The brain with the uppermost part of the cord, from the left. Fig. 296. Back view of the whole embryo: brain and spinal cord exposed. |
In all the Craniotes, from the Cyclostoma up to man, the same parts develop from these five original cerebral vesicles, though in very different ways. The first vesicle, the fore brain (Fig. 155 v), forms by far the largest part of the cerebrum—namely, the large hemispheres, the olfactory lobes, the corpora striata, the callosum, and the fornix. From the second vesicle, the intermediate brain (z), originate especially the optic thalami, the other parts that surround the third cerebral ventricle, and the infundibulum and pineal gland. The third vesicle, the middle brain (m), produces the corpora quadrigemina and the aqueduct of Sylvius. From the fourth vesicle, the hind brain (h), develops the greater part of the cerebellum—namely, the vermis and the two small hemispheres. Finally, the fifth vesicle, the after brain (n), forms the medulla oblongata, with the quadrangular pit (the floor of the fourth ventricle), the pyramids, olivary bodies, etc.
We must certainly regard it as a comparative-anatomical and ontogenetic fact of the greatest significance that in all the Craniotes, from the lowest Cyclostomes and fishes up to the apes and man, the brain develops in just the same way in the embryo. The first rudiment of it is always a simple vesicular enlargement of the fore end of the medullary tube. In every case, first three, then five, vesicles develop from this bulb, and the permanent brain with all its complex anatomic structures, of so great a variety in the various classes of Vertebrates, is formed from the five primitive vesicles. When we compare the mature brain of a fish, an amphibian, a reptile, a bird, and a mammal, it seems incredible that we can trace the various parts of these organs, that differ so much internally and externally, to common types. Yet all these different Craniote brains have started with the same rudimentary structure. To convince ourselves of this we have only to compare the corresponding stages of development of the embryos of these different animals.
This comparison is extremely instructive. If we extend it through the whole series of the Craniotes, we soon discover this interesting fact: In the Cyclostomes (the Myxinoida and Petromyzonta), which we have recognised as the lowest and earliest Craniotes, the whole brain remains throughout life at a very low stage, which is very brief and passing in the embryos of the higher Craniotes; they retain the five original sections of the brain unchanged. In the fishes we find an essential and considerable modification of the five vesicles; it is clearly the brain of the Selachii in the first place, and subsequently the brain of the Ganoids, from which the brain of the rest of the fishes on the one hand and of the Dipneusts and Amphibia, and through these of the higher Vertebrates, on the other hand, must be derived. In the fishes and Amphibia (Fig. 300) there is a preponderant development of the middle brain, and also the after brain, the first, second, and
[ 277 ]
fourth sections remaining very primitive. It is just the reverse in the higher Vertebrates, in which the first and third sections, the cerebrum and cerebellum, are exceptionally developed; while the middle brain and after brain remain small. The corpora quadrigemina are mostly covered by the cerebrum, and the oblongata by the cerebellum. But we find a number of stages of development within the higher Vertebrates themselves. From the Amphibia upwards the brain (and with it the psychic life) develops in two different directions; one of these is followed by the reptiles and birds, and the other by the mammals. The development of the first section, the fore brain, is particularly characteristic of the mammals. It is only in them that the cerebrum becomes so large as to cover all the other parts of the brain (Figs. 293, 301–304).
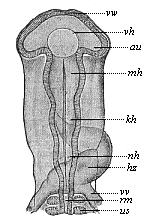 Fig. 297—Head of a chick embryo (hatched fifty-eight hours), from the back. (From Mihalkovics.) vw anterior wall of the fore brain. vh its ventricle. au optic vesicles, mh middle brain, kh hind brain, nh after brain, hz heart (seen from below), vw vitelline veins, us primitive segment, rm spinal cord.
Fig. 297—Head of a chick embryo (hatched fifty-eight hours), from the back. (From Mihalkovics.) vw anterior wall of the fore brain. vh its ventricle. au optic vesicles, mh middle brain, kh hind brain, nh after brain, hz heart (seen from below), vw vitelline veins, us primitive segment, rm spinal cord. | | 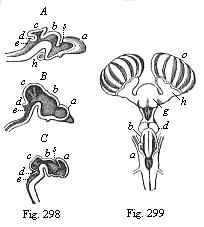 Fig. 298—Brain of three craniote embryos in vertical section. A of a shark (Heptarchus), B of a serpent (Coluber), C of a goat (Capra). a fore brain, b intermediate brain, c middle brain, d hind brain, e after brain, s primitive cleft. (From Gegenbaur.)
Fig. 298—Brain of three craniote embryos in vertical section. A of a shark (Heptarchus), B of a serpent (Coluber), C of a goat (Capra). a fore brain, b intermediate brain, c middle brain, d hind brain, e after brain, s primitive cleft. (From Gegenbaur.)
Fig. 299—Brain of a shark (Scyllium), back view. g fore-brain, h olfactory lobes, which send the large olfactory nerves to the nasal capsule (o), d intermediate brain, b middle brain; behind this the insignificant structure of the hind brain, a after brain. (From Gegenbaur.) |
There are also notable variations in the relative position of the cerebral vesicles. In the lower Craniotes they lie originally almost in the same plane. When we examine the brain laterally, we can cut through all five vesicles with a straight line. But in the Amniotes there is a considerable curve in the brain along with the bending of the head and neck; the whole of the upper dorsal surface of the brain develops much more than the under ventral surface. This causes a curve, so that the parts come to lie as follows: The fore brain is right in front and below, the intermediate brain a little higher, and the middle brain highest of all; the hind brain lies a little lower, and the after brain lower still. We find this only in the Amniotes—the reptiles, birds, and mammals.
Thus, while the brain of the mammals agrees a good deal in general growth with that of the birds and reptiles, there are some striking differences between the two. In the Sauropsids (birds and reptiles) the middle brain and the middle part of the hind brain are well developed. In the mammals these parts do not grow, and the fore-brain develops so much that it overlies the other vesicles. As it continues to grow towards the rear, it at last covers the whole of the rest of the brain, and also encloses the middle parts from the sides (Figs. 301–303). This process is of great importance, because the fore brain is the organ of the higher psychic life, and in it those functions of the nerve-cells are discharged which we sum up in
[ 278 ]
the word “soul.” The highest achievements of the animal body—the wonderful manifestations of consciousness and the complex molecular processes of thought—have their seat in the fore brain. We can remove the large hemispheres, piece by piece, from the mammal without killing it, and we then see how the higher functions of consciousness, thought, will, and sensation, are gradually destroyed, and in the end completely extinguished. If the animal is fed artificially, it may be kept alive for a long time, as the destruction of the psychic organs by no means involves the extinction of the faculties of digestion, respiration, circulation, urination—in a word, the vegetative functions. It is only conscious sensation, voluntary movement, thought, and the combination of various higher psychic functions that are affected.
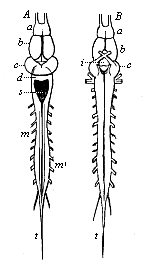 Fig. 300—Brain and spinal cord of the frog. A from the dorsal, B from the ventral side. a olfactory lobes before the (b) fore brain, i infundibulum at the base of the intermediate brain, c middle brain, d hind brain, s quadrangular pit in the after brain, m spinal cord (very short in the frog), m′ roots of the spinal nerves, t terminal fibres of the spinal cord. (From Gegenbaur.)
Fig. 300—Brain and spinal cord of the frog. A from the dorsal, B from the ventral side. a olfactory lobes before the (b) fore brain, i infundibulum at the base of the intermediate brain, c middle brain, d hind brain, s quadrangular pit in the after brain, m spinal cord (very short in the frog), m′ roots of the spinal nerves, t terminal fibres of the spinal cord. (From Gegenbaur.) |
The fore brain, the organ of these functions, only attains this high level of development in the more advanced Placentals, and thus we have the simple explanation of the intellectual superiority of the higher mammals. The soul of most of the lower Placentals is not much above that of the reptiles, but among the higher Placentals we find an uninterrupted gradation of mental power up to the apes and man. In harmony with this we find an astonishing variation in the degree of development of their fore brain, not only qualitatively, but also quantitatively. The mass and weight of the brain are much greater in modern mammals, and the differentiation of its various parts more important, than in their extinct Tertiary ancestors. This can be shown paleontologically in any particular order. The brains of the living ungulates are (relatively to the size of the body) four to six times (in the highest groups even eight times) as large as those of their earlier Tertiary ancestors, the well-preserved skulls of which enable us to determine the size and weight of the brain.
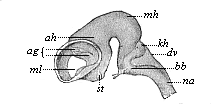 Fig. 301—Brain of an ox-embryo, two inches in length. (From Mihalkovics.) Left view; the lateral wall of the left hemisphere has been removed, st corpora striata, ml Monro-foramen, ag arterial plexus, ah Ammon’s horn, mh middle brain, kh cerebellum, dv roof of the fourth ventricle, bb pons Varolii, na medulla oblongata.
Fig. 301—Brain of an ox-embryo, two inches in length. (From Mihalkovics.) Left view; the lateral wall of the left hemisphere has been removed, st corpora striata, ml Monro-foramen, ag arterial plexus, ah Ammon’s horn, mh middle brain, kh cerebellum, dv roof of the fourth ventricle, bb pons Varolii, na medulla oblongata. | |
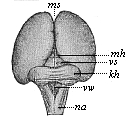 Fig. 302—Brain of a human embryo, twelve weeks old. (From Mihalkovics.) Seen from behind and above. ms mantle-furrow, mh corpora quadrigemina (middle brain), vs anterior medullary ala, kh cerebellum, vv fourth ventricle, na medulla oblongata.
Fig. 302—Brain of a human embryo, twelve weeks old. (From Mihalkovics.) Seen from behind and above. ms mantle-furrow, mh corpora quadrigemina (middle brain), vs anterior medullary ala, kh cerebellum, vv fourth ventricle, na medulla oblongata. |
In the lower mammals the surface of the cerebral hemispheres is quite smooth and level, as in the rabbit (Fig. 304). Moreover, the fore brain remains so small that it does not cover the middle brain. At a stage higher the middle
[ 279 ]
brain is covered, but the hind brain remains free. Finally, in the apes and man, the latter also is covered by the fore brain. We can trace a similar gradual development in the fissures and convolutions that are found on the surface of the cerebrum of the higher mammals (Figs. 292, 293). If we compare different groups of mammals in regard to these fissures and convolutions, we find that their development proceeds step by step with the advance of mental life.
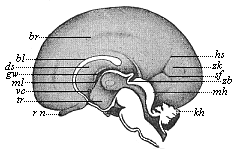 |
Fig. 303—Brain of a human embryo, twenty-four weeks old, halved in the median plane: right hemisphere seen from inside. (From Mihalkovics.) rn olfactory nerve, tr funnel of the intermediate brain, vc anterior commissure, ml Monro-foramen, gw fornix, ds transparent sheath, bl corpus callosum, br fissure at its border, hs occipital fissure, zh cuneus, sf occipital transverse fissure, zb pineal gland, mh corpora quadrigemina, kh cerebellum. |
Of late years great attention has been paid to this special branch of cerebral anatomy, and very striking individual differences have been detected within the limits of the human race. In all human beings of special gifts and high intelligence the convolutions and fissures are much more developed than in the average man; and they are more developed in the latter than in idiots and others of low mental capacity. There is a similar gradation among the mammals in the internal structure of the fore brain. In particular the corpus callosum, that unites the two cerebral hemispheres, is only developed in the Placentals. Other structures—for instance, in the lateral ventricles—that seem at first to be peculiar to man, are also found in the higher apes, and these alone. It was long thought that man had certain distinctive organs in his cerebrum which were not found in any other animal. But careful examination has discovered that this is not the case, but that the characteristic features of the human brain are found in a rudimentary form in the lower apes, and are more or less fully developed in the higher apes. Huxley has convincingly shown, in his Man’s Place in Nature (1863), that the differences in the formation of the brain within the ape-group constitute a deeper gulf between the lower and higher apes than between the higher apes and man.
|
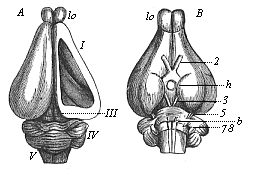
Fig. 304—Brain of the rabbit. A from the dorsal, B from the ventral side, lo olfactory lobes, I fore brain, h hypophysis at the base of the intermediate brain, III middle brain, IV hind brain, V after brain, 2 optic nerve, 3 oculo-motor nerve, 5–8 cerebral nerves. In A the roof of the right hemisphere (I) is removed, so that we can see the corpora striata in the lateral ventricle. (From Gegenbaur.) |
The comparative anatomy and physiology of the brain of the higher and lower mammals are very instructive, and give important information in connection with the chief questions of psychology.
The central marrow (brain and spinal cord) develops from the medullary tube in man just as in all the other mammals, and the same applies to the conducting marrow or “peripheral nervous system.” It consists of the sensory nerves, which conduct centripetally the impressions from the skin and the sense-organs to the central marrow, and of the motor nerves, which convey centrifugally the movements of the will from the central marrow to the muscles. All these
[ 280 ]
peripheral nerves grow out of the medullary tube (Fig. 171), and are, like it, products of the skin-sense layer.
The complete agreement in the structure and development of the psychic organs which we find between man and the highest mammals, and which can only be explained by their common origin, is of profound importance in the monistic psychology. This is only seen in its full light when we compare these morphological facts with the corresponding physiological phenomena, and remember that every psychic action requires the complete and normal condition of the correlative brain structure for its full and normal exercise. The very complex molecular movements inside the neural cells, which we describe comprehensively as “the life of the soul,” can no more exist in the vertebrate, and therefore in man, without their organs than the circulation without the heart and blood. And as the central marrow develops in man from the same medullary tube as that of the other vertebrates, and as man shares the characteristic structure of his cerebrum (the organ of thought) with the anthropoid apes, his psychic life also must have the same origin as theirs.
If we appreciate the full weight of these morphological and physiological facts, and put a proper phylogenetic interpretation on the observations of embryology, we see that the older idea of the personal immortality of the human soul is scientifically untenable. Death puts an end, in man as in any other vertebrate, to the physiological function of the cerebral neurona, the countless microscopic ganglionic cells, the collective activity of which is known as “the soul.” I have shown this fully in the eleventh chapter of my Riddle of the Universe.
Title and Contents
Vol. II Title and Contents
Glossary
Chapter XXIII
Chapter XXV
Figs. 1–209
Figs. 210–408