THE EVOLUTION OF MAN
Volume II
CHAPTER XXIII
OUR APE ANCESTORS
The long series of animal forms which we must regard
as the ancestors of our race has been confined within narrower and
narrower circles as our phylogenetic inquiry has progressed. The
great majority of known animals do not fall in the line of our
ancestry, and even within the vertebrate stem only a small number
are found to do so. In the most advanced class of the stem, the
mammals, there are only a few families that belong directly to our
genealogical tree. The most important of these are the apes and
their predecessors, the half-apes, and the earliest Placentals
(Prochoriata).
The Placentals (also called Choriata, Monodelphia,
Eutheria or Epitheria) are distinguished from the lower
mammals we have just considered, the Monotremes and Marsupials, by
a number of striking peculiarities. Man has all these distinctive
features; that is a very significant fact. We may, on the ground of
the most careful comparative-anatomical and ontogenetic research,
formulate the thesis: “Man is in every respect a true
Placental.” He has all the characteristics of structure and
development that distinguish the Placentals from the two lower
divisions of the mammals, and, in fact, from all other animals.
Among these characteristics we must especially notice the more
advanced development of the brain. The fore-brain or cerebrum
especially is much more developed in them than in the lower
animals. The corpus callosum, which forms a sort of wide
bridge connecting the two hemispheres of the cerebrum, is only
fully formed in the Placentals; it is very rudimentary in the
Marsupials and Monotremes. It is true that the lowest Placentals
are not far removed from the Marsupials in cerebral development;
but within the placental group we can trace an unbroken gradation
of progressive development of the brain, rising gradually from this
lowest stage up to the elaborate psychic organ of the apes and man.
The human soul—a physiological function of the brain—is
in reality only a more advanced ape-soul.
The mammary glands of the Placentals are provided with teats
like those of the Marsupials; but we never find in the Placentals
the pouch in which the latter carry and suckle their young. Nor
have they the marsupial bones in the ventral wall at the anterior
border of the pelvis, which the Marsupials have in common with the
Monotremes, and which are formed by a partial ossification of the
sinews of the inner oblique abdominal muscle. There are merely a
few insignificant remnants of them in some of the Carnivora. The
Placentals are also generally without the hook-shaped process at
the angle of the lower jaw which is found in the Marsupials.
However, the feature that characterises the Placentals above all
others, and that has given its name to the whole sub-class, is the
formation of the placenta. We have already considered the formation
and significance of this remarkable embryonic organ when we traced
the development of the chorion and the allantois in the human
embryo (pp.165–9) The
urinary sac or the allantois, the
[ 254 ]
curious vesicle that grows out of the hind part of
the gut, has essentially the same structure and function in the
human embryo as in that of all the other Amniotes (cf. Figs. 194–6). There is a quite
secondary difference, on which great stress has wrongly been laid,
in the fact that in man and the higher apes the original cavity of
the allantois quickly degenerates, and the rudiment of it sticks
out as a solid projection from the primitive gut. The thin wall of
the allantois consists of the same two layers or membranes as the
wall of the gut—the gut-gland layer within and the gut-fibre
layer without. In the gut-fibre layer of the allantois there are
large blood-vessels, which serve for the nutrition, and especially
the respiration, of the embryo—the umbilical vessels (p. 170). In the reptiles and birds the
allantois enlarges into a spacious sac, which encloses the embryo
with the amnion, and does not combine with the outer fœtal
membrane (the chorion). This is the case also with the lowest
mammals, the oviparous Monotremes and most of the Marsupials. It is
only in some of the later Marsupials (Peramelida) and all
the Placentals that the allantois develops into the distinctive and
remarkable structure that we call the placenta.
Fig. 273—Fœtal membranes of the human
embryo (diagrammatic). m the thick muscular wall of the
womb. plu placenta [the inner layer (plu′) of
which penetrates into the chorion-villi (chz) with its
processes]. chf tufted, chl smooth chorion. a
amnion, ah amniotic cavity, as amniotic sheath of the
umbilical cord (which passes under into the navel of the
embryo—not given here), dg vitelline duct, ds
yelk sac, dv, dr decidua (vera and reflexa). The uterine
cavity (uh) opens below into the vagina and above on the
right into an oviduct (t). (From Kölliker.) |
The placenta is formed by the branches of the blood-vessels in
the wall of the allantois growing into the hollow ectodermic tufts
(villi) of the chorion, which run into corresponding depressions in
the mucous membrane of the womb. The latter also is richly
permeated with blood-vessels which bring the mother’s blood
to the embryo. As the partition in the villi between the maternal
blood-vessels and those of the fœtus is extremely thin, there
is a direct exchange of fluid between the two, and this is of the
greatest importance in the nutrition of the young mammal. It is
true that the maternal vessels do not entirely pass into the
fœtal vessels, so that the two kinds of blood are simply
mixed. But the partition between them is so thin that the nutritive
fluid easily transudes through it. By means of this transudation or
diosmosis the exchange of fluids takes place without difficulty.
The larger the embryo is in the placentals, and the longer it
remains in the womb, the more necessary it is to have special
structures to meet its great consumption of food.
In this respect there is a very conspicuous difference between
the lower and higher mammals. In the Marsupials, in which the
embryo is only a comparatively short time in the womb and is born
in a very immature condition, the vascular arrangements in the
yelk-sac and the allantois suffice for its nutrition, as we find
them in the Monotremes, birds, and reptiles. But in the Placentals,
where gestation lasts a long time, and the embryo reaches its full
development under the protection of its enveloping membranes, there
has to be a new mechanism for the direct supply of a large quantity
of food, and this is admirably met by the formation of the
placenta.
Branches of the blood-vessels penetrate into the chorion-villi
from within, starting from the gut-fibre layer of the allantois,
and bringing the blood of the fœtus through the umbilical
vessels (Fig. 273 chz). On the other hand, a thick network
of blood-vessels develops in the mucous membrane that clothes the
inner surface of the womb, especially in the region of the
depressions into which the chorion-villi penetrate (plu).
This network of arteries contains maternal blood, brought by the
uterine vessels. As the connective tissue between the enlarged
capillaries of
[ 255 ]
the uterus disappears, wide cavities filled with
maternal blood appear, and into these the chorion-villi of the
embryo penetrate. The sum of these vessels of both kinds, that are
so intimately correlated at this point, together with the
connective and enveloping tissue, is the placenta. The
placenta consists, therefore, properly speaking, of two different
though intimately connected parts—the fœtal placenta
(Fig. 273 chz) within and the maternal or uterine placenta
(plu) without. The latter is made up of the mucous coat of
the uterus and its blood-vessels, the former of the tufted chorion
and the umbilical vessels of the embryo (cf. Fig. 196).
 Fig. 274—Skull of a fossil
lemur (Adapis parisiensis), from the Miocene at Quercy.
A lateral view from the right. B lower jaw, C
lower molar, i incisors, c canines, p
premolars, m molars.
Fig. 274—Skull of a fossil
lemur (Adapis parisiensis), from the Miocene at Quercy.
A lateral view from the right. B lower jaw, C
lower molar, i incisors, c canines, p
premolars, m molars. |
The manner in which these two kinds of vessels combine in the
placenta, and the structure, form, and size of it, differ a good
deal in the various Placentals; to some extent they give us
valuable data for the natural classification, and therefore the
phylogeny, of the whole of this sub-class. On the ground of these
differences we divide it into two principal sections; the lower
Placentals or Indecidua, and the higher Placentals or
Deciduata.
To the Indecidua belong three important groups of mammals: the
Lemurs (Prosimiæ), the Ungulates (tapirs, horses,
pigs, ruminants, etc.), and the Cetacea (dolphins and whales). In
these Indecidua the villi are distributed over the whole surface of
the chorion (or its greater part) either singly or in groups. They
are only loosely connected with the mucous coat of the uterus, so
that the whole fœtal membrane with its villi can be easily
withdrawn from the uterine depressions like a hand from a glove.
There is no real coalescence of the two placentas at any part of
the surface of contact. Hence at birth the fœtal placenta
alone comes away; the uterine placenta is not torn away with
it.
The formation of the placenta is very different in the second
and higher section of the Placentals, the Deciduata. Here
again the whole surface of the chorion is thickly covered with the
villi in the beginning. But they afterwards disappear from one part
of the surface, and grow proportionately thicker on the other part.
We thus get a differentiation between the smooth chorion
(chorion laeve, Fig. 273 chl) and the thickly-tufted
chorion (chorion frondosum, Fig. 273 chf). The former
has only a few small villi or none at all; the latter is thickly
covered with large and well-developed villi; this alone now
constitutes the placenta. In the great majority of the Deciduata
the placenta has the same shape as in man (Figs. 197 and 200)—namely a thick, circular disk
like a cake; so we find in the Insectivora, Chiroptera, Rodents,
and Apes. This discoplacenta lies on one side of the
chorion. But in the Sarcotheria (both the Carnivora and the seals,
Pinnipedia) and in the elephant and several other Deciduates
we find a zonoplacenta; in these the rich mass of villi runs
like a girdle round the middle of the ellipsoid chorion, the two
poles of it being free from them.
Still more characteristic of the Deciduates is the peculiar and
very intimate connection between the chorion frondosum and
the corresponding part of the mucous coat of the womb, which we
must regard as a real coalescence of the two. The villi of the
chorion push their branches into the blood-filled tissues of the
coat of the uterus, and the vessels of each loop together so
intimately that it is no longer possible to separate the
fœtal
[ 256 ]
from the maternal placenta; they form henceforth a
compact and apparently simple placenta. In consequence of this
coalescence, a whole piece of the lining of the womb comes away at
birth with the fœtal membrane that is interlaced with it.
This piece is called the “falling-away” membrane
(decidua). It is also called the serous (spongy) membrane,
because it is pierced like a sieve or sponge. All the higher
Placentals that have this decidua are classed together as the
“Deciduates.” The tearing away of the decidua at birth
naturally causes the mother to lose a quantity of blood, which does
not happen in the Indecidua. The last part of the uterine coat has
to be repaired by a new growth after birth in the Deciduates. (Cf.
Figs. 199, 200, pp.
168–70.")
 Fig. 275—The Slender Lori
(Stenops gracilis) of Ceylon, a tail-less lemur."
Fig. 275—The Slender Lori
(Stenops gracilis) of Ceylon, a tail-less lemur." |
In the various orders of the Deciduates, the placenta differs
considerably both in outer form and internal structure. The
extensive investigations of the last ten years have shown that
there is more variation in these respects among the higher mammals
than was formerly supposed. The physiological work of this
important embryonic organ, the nutrition of the fœtus during
its long sojourn in the womb, is accomplished in the various groups
of the Placentals by very different and sometimes very elaborate
structures. They have lately been fully described by Hans
Strahl.
The phylogeny of the placenta has become more intelligible from
the fact that we have found a number of transitional forms of it.
Some of the Marsupials (Perameles) have the beginning of a
placenta. In some of the Lemurs (Tarsius) a discoid placenta
with decidua is developed.
While these important results of comparative embryology have
been throwing further light on the close blood-relationship of man
and the anthropoid apes in the last few years (p. 172), the great advance of
paleontology has at the same time been affording us a deeper
insight into the stem-history of the Placental group. In the
seventh chapter of my Systematic Phylogeny of the
Vertebrates I advanced the hypothesis that the Placentals form
a single stem with many branches, which has been evolved from an
older group of the Marsupials (Prodidelphia). The four great
legions of the Placentals—Rodents, Ungulates, Carnassia, and
Primates—are sharply separated to-day by important features
of organisation. But if we consider their extinct ancestors of the
Tertiary period, the differences gradually disappear, the deeper we
go in the Cenozoic deposits; in the end we find that they vanish
altogether.
[ 257 ]
The primitive stem-forms of the Rodents
(Esthonychida), the Ungulates (Chondylarthra), the
Carnassia (Ictopsida), and the Primates (Lemuravida)
are so closely related at the beginning of the Tertiary period that
we might group them together as different families of one order,
the Proplacentals (Mallotheria or Prochoriata).
Hence the great majority of the Placentals have no direct and
close relationship to man, but only the legion of the
Primates. This is now generally divided into three
orders—the half-apes (Prosimiæ), apes
(Simiæ), and man (Anthropi). The lemurs or
half-apes are the stem-group, descending from the older
Mallotheria of the Cretaceous period. From them the apes were
evolved in the Tertiary period, and man was formed from these
towards its close.
The Lemurs (Prosimiæ) have few living
representatives. But they are very interesting, and are the last
survivors of a once extensive group. We find many fossil remains of
them in the older Tertiary deposits of Europe and North America, in
the Eocene and Miocene. We distinguish two sub-orders, the fossil
Lemuravida and the modern Lemurogona. The earliest
and most primitive forms of the Lemuravida are the Pachylemurs
(Hypopsodina); they come next to the earliest Placentals
(Prochoriata), and have the typical full dentition, with
forty-four teeth (3.1.4.3. over 3.1.4.3.). The Necrolemurs
(Adapida, Fig. 274) have only forty teeth, and have lost an
incisor in each jaw (2.1.4.3. over 2.1.4.3.). The dentition is
still further reduced in the Lemurogona (Autolemures), which
usually have only thirty-six teeth (2.1.3.3. over 2.1.3.3.). These
living survivors are scattered far over the southern part of the
Old World. Most of the species live in Madagascar, some in the
Sunda Islands, others on the mainland of Asia and Africa. They are
gloomy and melancholic animals; they live a quiet life, climbing
trees, and eating fruit and insects. They are of different kinds.
Some are closely related to the Marsupials (especially the
opossum). Others (Macrotarsi) are nearer to the Insectivora,
others again (Chiromys) to the Rodents. Some of the lemurs
(Brachytarsi) approach closely to the true apes. The
numerous fossil remains of half-apes and apes that have been
recently found in the Tertiary deposits justify us in thinking that
man’s ancestors were represented by several different species
during this long period. Some of these were almost as big as men,
such as the diluvial lemurogonon Megaladapis of
Madagascar.
 Fig. 276—The white-nosed ape
(Cercopithecus petaurista).
Fig. 276—The white-nosed ape
(Cercopithecus petaurista). |
Next to the lemurs come the true apes (Simiæ), the
twenty-sixth stage in our ancestry. It has been beyond question for
some time now that the apes approach nearest to man in every
respect of all the animals. Just as the lowest apes come close to
the lemurs, so the highest come next to man. When we carefully
study the comparative anatomy of the apes and man, we can trace a
gradual and uninterrupted advance in the organisation of the ape up
to the purely human frame, and, after impartial examination of the
“ape problem” that has been discussed of late years
with such passionate interest, we come infallibly to the important
conclusion, first formulated by Huxley in 1863: “Whatever
systems of organs we take, the comparison of their modifications in
the series of apes leads to the same result: that the anatomic
differences that separate man from the gorilla and chimpanzee are
not as great as those that separate the gorilla from the lower
apes.” Translated into phylogenetic language, this
“pithecometra-law,” formulated in such masterly fashion
by Huxley, is quite equivalent to the popular saying: “Man is
descended from the apes.”
In the very first exposition of his profound natural
classification (1735) Linné
[ 258 ]

Fig. 277—The drill-baboon
(Cynocephalus leucophæus).
(From Brehm.) |
placed the anthropoid mammals at the head of the
animal kingdom, with three genera: man, the ape, and the sloth. He
afterwards called them the “Primates”—the
“lords” of the animal world; he then also separated the
lemur from the true ape, and rejected the sloth. Later zoologists
divided the order of Primates. First the Gottingen anatomist,
Blumenbach, founded a special order for man, which he called
Bimana (“two-handed”); in a second order he united
the apes and lemurs under the name of Quadrumana
(“four-handed”); and a third order was formed of the
distantly-related Chiroptera (bats, etc.). The separation of
the Bimana and Quadrumana was retained by Cuvier and most of the
subsequent zoologists. It seems to be extremely important, but, as
a matter of fact, it is totally wrong. This was first shown in 1863
by Huxley, in his famous Man’s Place in Nature. On the
strength of careful comparative anatomical research he proved that
the apes are just as truly “two-handed” as man; or, if
we prefer to reverse it, that man is as truly four-handed as the
ape. He showed convincingly that the ideas of hand and foot had
been wrongly defined, and had been improperly based on
physiological instead of morphological grounds. The circumstance
that we oppose the
[ 259 ]
thumb to the other four fingers in our hand, and so
can grasp things, seemed to be a special distinction of the hand in
contrast to the foot, in which the corresponding great toe cannot
be opposed in this way to the others. But the apes can grasp with
the hind-foot as well as the fore, and so were regarded as
quadrumanous. However, the inability to grasp that we find in the
foot of civilised man is a consequence of the habit of clothing it
with tight coverings for thousands of years. Many of the
bare-footed lower races of men, especially among the negroes, use
the foot very freely in the same way as the hand. As a result of
early habit and continued practice, they can grasp with the foot
(in climbing trees, for instance) just as well as with the hand.
Even new-born infants of our own race can grasp very strongly with
the great toe, and hold a spoon with it as firmly as with the hand.
Hence the physiological distinction between hand and foot can
neither be pressed very far, nor has it a scientific basis. We must
look to morphological characters.
As a matter of fact, it is possible to draw such a sharp
morphological distinction—a distinction based on anatomic
structure—between the fore and hind extremity. In the
formation both of the bony skeleton and of the muscles that are
connected with the hand and foot before and behind there are
material and constant differences; and these are found both in man
and the ape. For instance, the number and arrangement of the
smaller bones of the hand and foot are quite different. There are
similar constant differences in the muscles. The hind extremity
always has three muscles (a short flexor muscle, a short extensor
muscle, and a long calf-muscle) that are not found in the fore
extremity. The arrangement of the muscles also is different before
and behind. These characteristic differences between the fore and
hind extremities are found in man as well as the ape. There can be
no doubt, therefore, that the ape’s foot deserves that name
just as much as the human foot does, and that all true apes are
just as “bimanous” as man. The common distinction of
the apes as “quadrumanous” is altogether wrong
morphologically.
But it may be asked whether, quite apart from this, we can find
any other features that distinguish man more sharply from the ape
than the various species of apes are distinguished from each other.
Huxley gave so complete and demonstrative a reply to this question
that the opposition still raised on many sides is absolutely
without foundation. On the ground of careful comparative anatomical
research, Huxley proved that in all morphological respects the
differences between the highest and lowest apes are greater than
the corresponding differences between the highest apes and man. He
thus restored Linné’s order of the Primates (excluding
the bats), and divided it into three sub-orders, the first composed
of the half-apes (Lemuridæ), the second of the true
apes (Simiadæ), the third of men
(Anthropidæ).
But, as we wish to proceed quite consistently and impartially on
the laws of systematic logic, we may, on the strength of
Huxley’s own law, go a good deal farther in this division. We
are justified in going at least one important step farther, and
assigning man his natural place within one of the sections of the
order of apes. All the features that characterise this group of
apes are found in man, and not found in the other apes. We do not
seem to be justified, therefore, in founding for man a special
order distinct from the apes.
The order of the true apes (Simiæ or
Pitheca)—excluding the lemurs—has long been divided
into two principal groups, which also differ in their geographical
distribution. One group (Hesperopitheca, or western apes)
live in America. The other group, to which man belongs, are the
Eopitheca or eastern apes; they are found in Asia and Africa,
and were formerly in Europe. All the eastern apes agree with man in
the features that are chiefly used in zoological classification to
distinguish between the two simian groups, especially in the
dentition. The objection might be raised that the teeth are too
subordinate an organ physiologically for us to lay stress on them
in so important a question. But there is a good reason for it; it
is with perfect justice that zoologists have for more than a
century paid particular attention to the teeth in the systematic
division and arrangement of the orders of mammals. The number,
form, and arrangement of the teeth are much more faithfully
inherited in the various orders than most other characters.
Hence the form of dentition in man is very important. In the
fully developed
[ 260 ]
condition we have thirty-two teeth; of these eight
are incisors, four canine, and twenty molars. The eight incisors,
in the middle of the jaws, have certain characteristic differences
above and below. In the upper jaw the inner incisors are larger
than the outer; in the lower jaw the inner are the smaller. Next to
these, at each side of both jaws, is a canine (or “eye
tooth”), which is larger than the incisors. Sometimes it is
very prominent in man, as it is in most apes and many of the other
mammals, and forms a sort of tusk. Next to this there are five
molars above and below on each side, the first two of which
(the
 Fig. 278—Skeletons of a man and the
four anthropoid apes.
Fig. 278—Skeletons of a man and the
four anthropoid apes.
(Fig. 278, Gibbon; Fig. 279, Orang; Fig. 280, Chimpanzee; Fig. 281, Gorilla; Fig. 282, Man.
(From Huxley.) Cf. Figs. 203–209. |
[ 261 ]
“pre-molars”) are small, have only one
root, and are included in the change of teeth; the three back ones
are much larger, have two roots, and only come with the second
teeth. The apes of the Old World, or all the living or fossil apes
of Asia, Africa, and Europe, have the same dentition as man.
On the other hand, all the American apes have an additional
pre-molar in each half of the jaw. They have six molars above and
below on each side, or thirty-six teeth altogether. This
characteristic difference between the eastern and western apes has
been so faithfully inherited that it is very instructive for us. It
is true that there seems to be an exception in the case of a small
family of South American apes. The small silky apes
(Arctopitheca or Hapalidæ), which include the
tamarin (Midas) and the brush-monkey (Jacchus), have
only five molars in each half of the jaw (instead of six), and so
seem to be nearer to the eastern apes. But it is found, on closer
examination, that they have three premolars, like all the western
apes, and that only the last molar has been lost. Hence the
apparent exception really confirms the above distinction.
Of the other features in which the two groups of apes differ,
the structure of the nose is particularly instructive and
conspicuous. All the eastern apes have the same type of nose as
man—a comparatively narrow partition between the two halves,
so that the nostrils run downwards. In some of them the nose
protrudes as far as in man, and has the same characteristic
structure. We have already alluded to the curious long-nosed apes,
which have a long, finely-curved nose. Most of the eastern apes
have, it is true, rather flat noses, like, for instance, the
white-nosed monkey (Fig. 276); but the nasal partition is thin and
narrow in them all. The American apes have a different type of
nose. The partition is very broad and thick at the bottom, and the
wings of the nostrils are not developed, so that they point
outwards instead of downwards. This difference in the form of the
nose is so constantly inherited in both groups that the apes of the
New World are called “flat-nosed”
(Platyrrhinæ), and those of the Old World
“narrow-nosed” (Catarrhinæ). The bony
passage of the ear (at the bottom of which is the tympanum) is
short and wide in all the Platyrrhines, but long and narrow in all
the Catarrhines; and in man this difference also is
significant.
This division of the apes into Platyrrhines and Catarrhines, on
the ground of the above hereditary features, is now generally
admitted in zoology, and receives strong support from the
geographical distribution of the two groups in the east and west.
It follows at once, as regards the phylogeny of the apes, that two
divergent lines proceeded from the common stem-form of the
ape-order in the early Tertiary period, one of which spread over
the Old, the other over the New, World. It is certain that all the
Platyrrhines come of one stock, and also all the Catarrhines; but
the former are phylogenetically older, and must be regarded as the
stem-group of the latter.
What can we deduce from this with regard to our own genealogy?
Man has just the same characters, the same form of dentition,
auditory passage, and nose, as all the Catarrhines; in this he
radically differs from the Platyrrhines. We are thus forced to
assign him a position among the eastern apes in the order of
Primates, or at least place him alongside of them. But it follows
that man is a direct blood relative of the apes of the Old World,
and can be traced to a common stem-form together with all the
Catarrhines. In his whole organisation and in his origin man is a
true Catarrhine; he originated in the Old World from an unknown,
extinct group of the eastern apes. The apes of the New World, or
the Platyrrhines, form a divergent branch of our genealogical tree,
and this is only distantly related at its root to the human race.
We must assume, of course, that the earliest Eocene apes had the
full dentition of the Platyrrhines; hence we may regard this
stem-group as a special stage (the twenty-sixth) in our ancestry,
and deduce from it (as the twenty-seventh stage) the earliest
Catarrhines.
We have now reduced the circle of our nearest relatives to the
small and comparatively scanty group that is represented by the
sub-order of the Catarrhines; and we are in a position to answer
the question of man’s place in this sub-order, and say
whether we can deduce anything further from this position as to our
immediate ancestors. In answering this question the comprehensive
and able studies that Huxley gives of
[ 262 ]
the comparative anatomy of man and the various
Catarrhines in his Man’s Place in Nature are of great
assistance to us. It is quite clear from these that the differences
between man and the highest Catarrhines (gorilla, chimpanzee, and
orang) are in every respect slighter than the corresponding
differences between the highest and the lowest Catarrhines
(white-nosed monkey, macaco, baboon, etc.). In fact, within the
small group of the tail-less anthropoid apes the differences
between the various genera are not less than the differences
between them and man. This is seen by a glance at the skeletons
that Huxley has put together (Figs. 278–282). Whether we take
the skull or the vertebral column or the ribs or the fore or hind
limbs, or whether we extend the comparison to the muscles,
blood-vessels, brain, placenta, etc., we always reach the same
result on impartial examination—that man is not more
different from the other Catarrhines than the extreme forms of them
(for instance, the gorilla and baboon) differ from each other. We
may now, therefore, complete the Huxleian law we have already
quoted with the following thesis: “Whatever system of organs
we take, a comparison of their modifications in the series of
Catarrhines always leads to the same conclusion; the anatomic
differences that separate man from the most advanced Catarrhines
(orang, gorilla, chimpanzee) are not as great as those that
separate the latter from the lowest Catarrhines (white-nosed
monkey, macaco, baboon).”
We must, therefore, consider the descent of man from other
Catarrhines to be fully proved. Whatever further information on the
comparative anatomy and ontogeny of the living Catarrhines we may
obtain in the future, it cannot possibly disturb this conclusion.
Naturally, our Catarrhine ancestors must have passed through a long
series of different forms before the human type was produced. The
chief advances that effected this “creation of man,” or
his differentiation from the nearest related Catarrhines, were: the
adoption of the erect posture and the consequent greater
differentiation of the fore and hind limbs, the evolution of
articulate speech and its organ, the larynx, and the further
development of the brain and its function, the soul; sexual
selection had a great influence in this, as Darwin showed in his
famous work.
With an eye to these advances we can distinguish at least four
important stages in our simian ancestry, which represent prominent
points in the historical process of the making of man. We may take,
after the Lemurs, the earliest and lowest Platyrrhines of South
America, with thirty-six teeth, as the twenty-sixth stage of our
genealogy; they were developed from the Lemurs by a peculiar
modification of the brain, teeth, nose, and fingers. From these
Eocene stem-apes were formed the earliest Catarrhines or eastern
apes, with the human dentition (thirty-two teeth), by modification
of the nose, lengthening of the bony channel of the ear, and the
loss of four pre-molars. These oldest stem-forms of the whole
Catarrhine group were still thickly coated with hair, and had long
tails—baboons (Cynopitheca) or tailed apes
(Menocerca, Fig. 276). They lived during the Tertiary
period, and are found fossilised in the Miocene. Of the actual
tailed apes perhaps the nearest to them are the
Semnopitheci.
If we take these Semnopitheci as the twenty-seventh stage in our
ancestry, we may put next to them, as the twenty-eighth, the
tail-less anthropoid apes. This name is given to the most advanced
and man-like of the existing Catarrhines. They were developed from
the other Catarrhines by losing the tail and part of the hair, and
by a higher development of the brain, which found expression in the
enormous growth of the skull. Of this remarkable family there are
only a few genera to-day, and we have already dealt with them
(Chapter XV)—the gibbon (Hylobates, Fig. 203) and
orang (Satyrus, Figs. 204, 205) in South-Eastern Asia and
the Archipelago; and the chimpanzee (Anthropithecus, Figs.
206, 207) and gorilla (Gorilla, Fig. 208) in Equatorial
Africa.
The great interest that every thoughtful man takes in these
nearest relatives of ours has found expression recently in a fairly
large literature. The most distinguished of these works for
impartial treatment of the question of affinity is Robert
Hartmann’s little work on The Anthropoid Apes.
Hartmann divides the primate order into two families: (1)
Primarii (man and the anthropoid apes); and (2)
Simianæ (true apes, Catarrhines and Platyrrhines).
Professor Klaatsch, of Heidelberg, has advanced a different view in
his interesting and richly illustrated work on The Origin and
Development of the Human
[ 263 ]
Race. This is a substantial supplement to my
Anthropogeny, in so far as it gives the chief results of
modern research on the early history of man and civilisation. But
when Klaatsch declares the descent of man from the apes to be
“irrational, narrow-minded, and false,” in the belief
that we are thinking of some living species of ape, we must remind
him that no competent scientist has ever held so narrow a view. All
of us look merely—in the sense of Lamarck and Darwin—to
the original unity (admitted by Klaatsch) of the primate stem. This
common descent of all the Primates (men, apes, and lemurs) from one
primitive stem-form, from which the most far-reaching conclusions
follow for the whole of anthropology and philosophy, is admitted by
Klaatsch as well as by myself and all other competent zoologists
who accept the theory of evolution in general. He says explicitly
(p. 172): “The three anthropoid apes—gorilla,
chimpanzee, and orang—seem to be branches from a common root,
and this was not far from that of the gibbon and man.” That
is in the main the opinion that I have maintained (especially
against Virchow) in a number of works ever since 1866. The
hypothetical common ancestor of all the Primates, which must have
lived in the earliest Tertiary period (more probably in the
Cretaceous), was called by me Archiprimus; Klaatsch now
calls it Primatoid. Dubois has proposed the appropriate name
of Prothylobates for the common and much younger stem-form
of the anthropomorpha (man and the anthropoid apes). The actual
Hylobates is nearer to it than the other three existing
anthropoids. None of these can be said to be absolutely the most
man-like. The gorilla comes next to man in the structure of the
hand and foot, the chimpanzee in the chief features of the skull,
the orang in brain development, and the gibbon in the formation of
the chest. None of these existing anthropoid apes is among the
direct ancestors of our race; they are scattered survivors of an
ancient branch of the Catarrhines, from which the human race
developed in a particular direction.
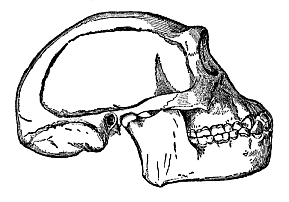 Fig. 283—Skull of the fossil
ape-man of Java (Pithecanthropus erectus), restored by
Eugen Dubois.
Fig. 283—Skull of the fossil
ape-man of Java (Pithecanthropus erectus), restored by
Eugen Dubois. |
Although man is directly connected with this anthropoid family
and originates from it, we may assign an important intermediate
form between the Prothylobates and him (the twenty-ninth
stage in our ancestry), the ape-men (Pithecanthropi). I gave
this name in the History of Creation to the
“speechless primitive men” (Alali), which were
men in the ordinary sense as far as the general structure is
concerned (especially in the differentiation of the limbs), but
lacked one of the chief human characteristics, articulate speech
and the higher intelligence that goes with it, and so had a less
developed brain. The phylogenetic hypothesis of the organisation of
this “ape-man” which I then advanced was brilliantly
confirmed twenty-four years afterwards by the famous discovery of
the fossil Pithecanthropus erectus by Eugen Dubois (then
military surgeon in Java, afterwards professor at Amsterdam). In
1892 he found at Trinil, in the residency of Madiun in Java, in
Pliocene deposits, certain remains of a large and very man-like ape
(roof of the skull, femur, and teeth), which he described as
“an erect ape-man” and a survivor of a “stem-form
of man” (Fig. 283). Naturally, the Pithecanthropus excited
the liveliest interest, as the long-sought transitional form
between man and the ape: we seemed to have found “the missing
link.” There were very interesting scientific discussions of
it at the last three International Congresses of Zoology (Leyden,
1895, Cambridge, 1898, and Berlin, 1901). I took an active part in
the discussion at
[ 264 ]
Cambridge, and may refer the reader to the paper I
read there on “The Present Position of Our Knowledge of the
Origin of Man” (translated by Dr. Gadow with the title of
The Last Link).
An extensive and valuable literature has grown up in the last
ten years on the Pithecanthropus and the pithecoid theory connected
with it. A number of distinguished anthropologists, anatomists,
paleontologists, and phylogenists have taken part in the
controversy, and made use of the important data furnished by the
new science of pre-historic research. Hermann Klaatsch has given a
good summary of them, with many fine illustrations, in the
above-mentioned work. I refer the reader to it as a valuable
supplement to the present work, especially as I cannot go any
further here into these anthropological and pre-historic questions.
I will only repeat that I think he is wrong in the attitude of
hostility that he affects to take up with regard to my own views on
the descent of man from the apes.
The most powerful opponent of the pithecoid theory—and the
theory of evolution in general—during the last thirty years
(until his death in September, 1902) was the famous Berlin
anatomist, Rudolf Virchow. In the speeches which he delivered every
year at various congresses and meetings on this question, he was
never tired of attacking the hated “ape theory.” His
constant categorical position was: “It is quite certain that
man does not descend from the ape or any other animal.” This
has been repeated incessantly by opponents of the theory,
especially theologians and philosophers. In the inaugural speech
that he delivered in 1894 at the Anthropological Congress at
Vienna, he said that “man might just as well have descended
from a sheep or an elephant as from an ape.” Absurd
expressions like this only show that the famous pathological
anatomist, who did so much for medicine in the establishment of
cellular pathology, had not the requisite attainments in
comparative anatomy and ontogeny, systematic zoology and
paleontology, for sound judgment in the province of anthropology.
The Strassburg anatomist, Gustav Schwalbe, deserved great praise
for having the moral courage to oppose this dogmatic and ungrounded
teaching of Virchow, and showing its untenability. The recent
admirable works of Schwalbe on the Pithecanthropus, the earliest
races of men, and the Neanderthal skull (1897–1901) will
supply any candid and judicious reader with the empirical material
with which he can convince himself of the baselessness of the
erroneous dogmas of Virchow and his clerical friends (J. Ranke, J.
Bumüller, etc.).
As the Pithecanthropus walked erect, and his brain (judging from
the capacity of his skull, Fig. 283) was midway between the lowest
men and the anthropoid apes, we must assume that the next great
step in the advance from the Pithecanthropus to man was the further
development of human speech and reason.
Comparative philology has recently shown that human speech is
polyphyletic in origin; that we must distinguish several (probably
many) different primitive tongues that were developed
independently. The evolution of language also teaches us (both from
its ontogeny in the child and its phylogeny in the race) that human
speech proper was only gradually developed after the rest of the
body had attained its characteristic form. It is probable that
language was not evolved until after the dispersal of the various
species and races of men, and this probably took place at the
commencement of the Quaternary or Diluvial period. The speechless
ape-men or Alali certainly existed towards the end of the
Tertiary period, during the Pliocene, possibly even the Miocene,
period.
The third, and last, stage of our animal ancestry is the true or
speaking man (Homo), who was gradually evolved from the
preceding stage by the advance of animal language into articulate
human speech. As to the time and place of this real “creation
of man” we can only express tentative opinions. It was
probably during the Diluvial period in the hotter zone of the Old
World, either on the mainland in tropical Africa or Asia or on an
earlier continent (Lemuria—now sunk below the waves of the
Indian Ocean), which stretched from East Africa (Madagascar,
Abyssinia) to East Asia (Sunda Islands, Further India). I have
given fully in my History of Creation, (chapter xxviii) the
weighty reasons for claiming this descent of man from the
anthropoid eastern apes, and shown how we may conceive the spread
of the various races from this “Paradise” over the
whole earth. I have also dealt fully with the relations of the
various races and species of men to each other.
[ 265 ]
SYNOPSIS OF THE CHIEF SECTIONS OF OUR STEM-HISTORY
First Stage: The Protists
Man’s ancestors are unicellular protozoa,
originally unnucleated Monera like the Chromacea, structureless
green particles of plasm; afterwards real nucleated cells (first
plasmodomous Protophyta, like the Palmella; then
plasmophagous Protozoa, like the Amœba).
Second Stage: The Blastæads
Man’s ancestors are round cœnobia or
colonies of Protozoa; they consist of a close association of many
homogeneous cells, and thus are individuals of the second order.
They resemble the round cell-communities of the Magospheræ
and Volvocina, equivalent to the ontogenetic blastula: hollow
globules, the wall of which consists of a single layer of ciliated
cells (blastoderm).
Third Stage: The Gastræads
Man’s ancestors are Gastræads, like
the simplest of the actual Metazoa (Prophysema, Olynthus, Hydra,
Pemmatodiscus). Their body consists merely of a primitive gut, the
wall of which is made up of the two primary germinal layers.
Fourth Stage: The Platodes
Man’s ancestors have substantially the
organisation of simple Platodes (at first like the
cryptocœlic Platodaria, later like the rhabdocœlic
Turbellaria). The leaf-shaped bilateral-symmetrical body has only
one gut-opening, and develops the first trace of a nervous centre
from the ectoderm in the middle line of the back (Figs. 239,
240).
Fifth Stage: The Vermalia
Man’s ancestors have substantially the
organisation of unarticulated Vermalia, at first Gastrotricha
(Ichthydina), afterwards Frontonia (Nemertina, Enteropneusta). Four
secondary germinal layers develop, two middle layers arising
between the limiting layers (cœloma). The dorsal ectoderm
forms the vertical plate, acroganglion (Fig. 243).
Sixth Stage: The Prochordonia
Man’s ancestors have substantially the
organisation of a simple unarticulated Chordonium (Copelata and
Ascidia-larvæ). The unsegmented chorda develops between the
dorsal medullary tube and the ventral gut-tube. The simple
cœlom-pouches divide by a frontal septum into two on each
side; the dorsal pouch (episomite) forms a muscle-plate; the
ventral pouch (hyposomite) forms a gonad. Head-gut with
gill-clefts.
Seventh Stage: The Acrania
Man’s ancestors are skull-less Vertebrates,
like the Amphioxus. The body is a series of metamera, as several of
the primitive segments are developed. The head contains in the
ventral half the branchial gut, the trunk the hepatic gut. The
medullary tube is still simple. No skull, jaws, or limbs.
Eighth Stage: The Cyclostoma
Man’s ancestors are jaw-less Craniotes (like
the Myxinoida and Petromyzonta). The number of metamera increases.
The fore-end of the medullary tube expands into a vesicle and forms
the brain, which soon divides into five cerebral vesicles. In the
sides of it appear the three higher sense-organs: nose, eyes, and
auditory vesicles. No jaws, limbs, or floating bladder.
Ninth Stage: The Ichthyoda
Man’s ancestors are fish-like Craniotes: (1)
Primitive fishes (Selachii); (2) plated fishes (Ganoida); (3)
amphibian fishes (Dipneusta); (4) mailed amphibia (Stegocephala).
The ancestors of this series develop two pairs of limbs: a pair of
fore (breast-fins) and of hind (belly-fins) legs. The gill-arches
are formed between the gill-clefts: the first pair form the
maxillary arches (the upper and lower jaws). The floating bladder
(lung) and pancreas grow out of the gut.
Tenth Stage: The Amniotes
Man’s ancestors are Amniotes or gill-less
Vertebrates: (1) Primitive Amniotes (Proreptilia); (2)
Sauromammals; (3) Primitive Mammals (Monotremes); (4) Marsupials;
(5) Lemurs (Prosimiæ); (6) Western apes (Platyrrhinæ);
(7) Eastern apes (Catarrhinæ): at first tailed Cynopitheca;
then tail-less anthropoids; later speechless ape-men (Alali);
finally speaking man. The ancestors of these Amniotes develop an
amnion and allantois, and gradually assume the mammal, and finally
the specifically human, form.
Title and Contents
Vol. II Title and Contents
Glossary
Chapter XXII
Chapter XXIV
Figs. 1–209
Figs. 210–408