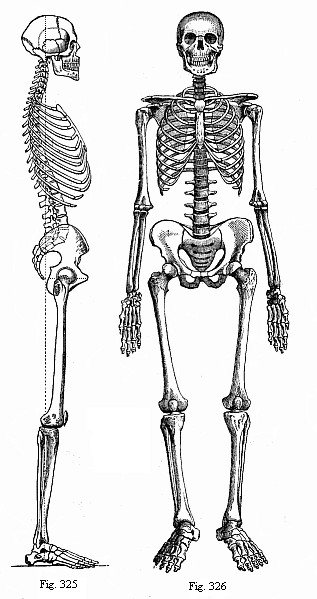
 Fig. 325—The human skeleton. From the right.
Fig. 325—The human skeleton. From the right.Fig. 326—The human skeleton. Front.
The peculiar structure of the locomotive apparatus is one of the features that are most distinctive of the vertebrate stem. The chief part of this apparatus is formed, as in all the higher animals, by the active organs of movement, the muscles; in consequence of their contractility they have the power to draw up and shorten themselves. This effects the movement of the various parts of the body, and thus the whole body is conveyed from place to place. But the arrangement of these muscles and their relation to the solid skeleton are different in the Vertebrates from the Invertebrates.
In most of the lower animals, especially the Platodes and Vermalia, we find that the muscles form a simple, thin layer of flesh immediately underneath the skin. This muscular layer is very closely connected with the skin itself; it is the same in the Mollusc stem. Even in the large division of the Articulates, the classes of crabs, spiders, myriapods, and
|
Fig. 325—The human skeleton. From the right. Fig. 326—The human skeleton. Front. |
insects, we find a similar feature, with the difference that in this case the skin forms a solid armour—a rigid cutaneous skeleton made of chitine (and often also of carbonate of lime). This external chitine coat undergoes a very elaborate articulation both on the trunk and the limbs of the Articulates, and in consequence the muscular system also, the contractile fibres of which are attached inside the chitine tubes, is highly articulated. The Vertebrates form a direct contrast to this. In these alone a solid internal skeleton is developed, of cartilage or bone, to which the muscles are attached. This bony skeleton is a complex lever apparatus, or passive apparatus of movement. Its rigid parts, the arms of the levers, or the bones, are brought together by the actively mobile muscles, as if by drawing-ropes. This admirable locomotorium, especially its solid central axis, the vertebral column, is a special feature of the Vertebrates, and has given the name to the group.
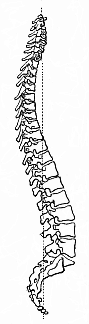 Fig. 327—The human vertebral column (standing upright, from the right side). (From H. Meyer.)
Fig. 327—The human vertebral column (standing upright, from the right side). (From H. Meyer.) |
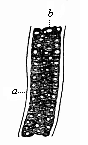 Fig. 328—A piece of the axial rod (chorda dorsalis), from a sheep embryo. a cuticular sheath, b cells. (From Kölliker.)
Fig. 328—A piece of the axial rod (chorda dorsalis), from a sheep embryo. a cuticular sheath, b cells. (From Kölliker.) |
In order to get a clear idea of the chief features of the development of the human skeleton, we must first examine its composition in the adult frame (Fig. 325, the human skeleton seen from the right; Fig. 326, front view of the whole skeleton). As in other mammals, we distinguish first between the axial or dorsal skeleton and the skeleton of the limbs. The axial skeleton consists of the vertebral column (the skeleton of the trunk) and the skull (skeleton of the head); the latter is a peculiarly modified part of the former. As appendages of the vertebral column we have the ribs, and of the skull we have the hyoid bone, the lower jaw, and the other products of the gill-arches.
The skeleton of the limbs or extremities is composed of two groups of parts—the skeleton of the extremities proper and the zone-skeleton, which connects these with the vertebral column. The zone-skeleton of the arms (or fore legs) is the shoulder-zone; the zone-skeleton of the legs (or hind legs) is the pelvic zone.
The vertebral column (Fig. 327) in man is composed of thirty-three to thirty-five ring-shaped bones in a continuous series (above each other, in man’s upright position). These vertebræ are separated from each other by elastic ligaments, and at the same time connected by joints, so that the whole column forms a firm and solid, but flexible and elastic, axial skeleton, moving freely in all directions. The vertebræ differ in shape and connection at the various parts of the trunk, and we distinguish the following groups in the series, beginning at the top: Seven cervical vertebræ, twelve dorsal vertebræ, five lumbar vertebræ, five sacral vertebræ, and four to six caudal vertebræ. The uppermost, or those next to the skull, are the cervical vertebræ (Fig. 327); they have a hole in each of the lateral processes. There are seven of these vertebræ in man and almost all the other mammals, even if the neck is as long as that of the camel or giraffe, or as short as that of the mole or hedgehog. This constant number, which has few exceptions (due to adaptation), is a strong proof of the common descent of the mammals; it can only be explained by faithful heredity from a common stem-form, a primitive mammal with seven cervical vertebræ. If each species had been created separately, it would have been better to have given the long-necked mammals more, and the short-necked animals less, cervical vertebræ. Next to these come the dorsal (or pectoral)
vertebræ, which number twelve to thirteen (usually twelve) in man and most of the other mammals. Each dorsal vertebra (Fig. 165) has at the side, connected by joints, a couple of ribs, long bony arches that lie in and protect the wall of the chest. The twelve pairs of ribs, together with the connecting intercostal muscles and the sternum, which joins the ends of the right and left ribs in front, form the chest (thorax). In this strong and elastic frame are the lungs, and between them the heart. Next to the dorsal vertebræ comes a short but stronger section of the column, formed of five large vertebræ. These are the lumbar vertebræ (Fig. 166); they have no ribs and no holes in the transverse processes. To these succeeds the sacral bone, which is fitted between the two halves of the pelvic zone. The sacrum is formed of five vertebræ, completely blended together. Finally, we have at the end a small rudimentary caudal column, the coccyx. This consists of a varying number (usually four, more rarely three, or five or six) of small degenerated vertebræ, and is a useless rudimentary organ with no actual physiological significance. Morphologically, however, it is of great interest as an irrefragable proof of the descent of man and the anthropoids from long-tailed apes. On no other theory can we explain the existence of this rudimentary tail. In the earlier stages of development the tail of the human embryo protrudes considerably. It afterwards atrophies; but the relic of the atrophied caudal vertebræ and of the rudimentary muscles that once moved it remains permanently. Sometimes, in fact, the external tail is preserved. The older anatomists say that the tail is usually one vertebra longer in the human female than in the male (or four against five); Steinbach says it is the reverse.
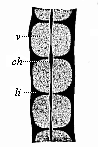 | Fig. 329—Three dorsal vertebræ, from a human embryo, eight weeks old, in lateral longitudinal section. v cartilaginous vertebral body, li inter-vertebral disks, ch chorda. (From Kölliker.) |
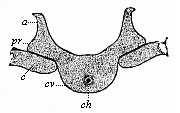
| Fig. 330—A dorsal vertebra of the same embryo, in lateral transverse section. cv cartilaginous vertebral body, ch chorda, pr transverse process, a vertebral arch (upper arch), c upper end of the rib (lower arch). (From Kölliker.) |
In the human vertebral column there are usually thirty-three vertebræ. It is interesting to find, however, that the number often changes, one or two vertebræ dropping out or an additional one appearing. Often, also, a mobile rib is formed at the last cervical or the first lumbar vertebra, so that there are then thirteen dorsal vertebræ, besides six cervical and four lumbar. In this way the contiguous vertebræ of the various sections of the column may take each other’s places.
In order to understand the embryology of the human vertebral column we must first carefully consider the shape and connection of the vertebræ. Each vertebra has, in general, the shape of a seal-ring (Figs. 164–166). The thicker portion, which is turned towards the ventral side, is called the body of the vertebra, and forms a short osseous disk; the thinner part forms a semi-circular arch, the vertebral arch, and is turned towards the back. The arches of the successive vertebræ are connected by thin intercrural ligaments in such a way that the cavity they collectively enclose represents a long canal. In this vertebral canal we find the trunk part of the central nervous system, the spinal cord. Its head part, the brain, is enclosed by the skull, and the skull itself is merely the uppermost part of the vertebral column, distinctively modified. The base or ventral side of the vesicular cranial capsule corresponds originally to a number of developed vertebral bodies; its vault or dorsal side to their combined upper vertebral arches.
While the solid, massive bodies of the vertebræ represent the real central axis of the skeleton, the dorsal arches serve to protect the central marrow they enclose. But similar arches develop on the ventral side for the protection of the viscera in the breast and belly. These lower or
ventral vertebral arches, proceeding from the ventral side of the vertebral bodies, form, in many of the lower Vertebrates, a canal in which the large blood-vessels are enclosed on the lower surface of the vertebral column (aorta and caudal vein). In the higher Vertebrates the majority of these vertebral arches are lost or become rudimentary. But at the thoracic section of the column they develop into independent strong osseous arches, the ribs (costæ). In reality the ribs are merely large and independent lower vertebral arches, which have lost their original connection with the vertebral bodies.
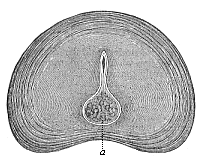 Fig. 331—Intervertebral disk of a new-born infant, transverse section. a rest of the chorda. (From Kölliker.)
Fig. 331—Intervertebral disk of a new-born infant, transverse section. a rest of the chorda. (From Kölliker.) |
If we turn from this anatomic survey of the composition of the column to the question of its development, I may refer the reader to earlier pages with regard to the first and most important points (pp. 145–148). It will be remembered that in the human embryo and that of the other vertebrates we find at first, instead of the segmented column, only a simple unarticulated cartilaginous rod. This solid but flexible and elastic rod is the axial rod (or the chorda dorsalis). In the lowest Vertebrate, the Amphioxus, it retains this simple form throughout life, and permanently represents the whole internal skeleton (Fig. 210 i). In the Tunicates, also, the nearest Invertebrate relatives of the Vertebrates, we meet the same chorda—transitorily in the passing larva tail of the Ascidia, permanently in the Copelata (Fig. 225 c). Undoubtedly both the Tunicates and Acrania have inherited the chorda from a common unsegmented stem-form; and these ancient, long-extinct ancestors of all the chordonia are our hypothetical Prochordonia.
Long before there is any trace of the skull, limbs, etc., in the embryo of man or any of the higher Vertebrates—at the early stage in which the whole body is merely a sole-shaped embryonic shield—there appears in the middle line of the shield, directly under the medullary furrow, the simple chorda. (Cf. Figs. 131–135 ch). It follows the long axis of the body in the shape of a cylindrical axial rod of elastic but firm composition, equally pointed at both ends. In every case the chorda originates from the dorsal wall of the primitive gut; the cells that compose it (Fig. 328 b) belong to the entoderm (Figs. 216–221). At an early stage the chorda develops a transparent structureless sheath, which is secreted from its cells (Fig. 328 a). This chordalemma is often called the “inner chorda-sheath,” and must not be confused with the real external sheath, the mesoblastic perichorda.
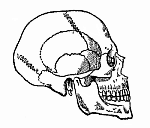 Fig. 332—Human skull.
Fig. 332—Human skull. |
But this unsegmented primary axial skeleton is soon replaced by the segmented secondary axial skeleton, which we know as the vertebral column. The provertebral plates (Fig. 124 s) differentiate from the innermost, median part of the visceral layer of the cœlom-pouches at each side of the chorda. As they grow round the chorda and enclose it they form the skeleton plate or skeletogenetic layer—that is to say, the skeleton-forming stratum of cells, which provides the mobile foundation of the permanent vertebral column and skull (scleroblast). In the head-half of the embryo the skeletal plate remains a continuous, simple, undivided layer of tissue, and presently enlarges into a thin-walled capsule enclosing the brain, the primordial skull. In the trunk-half the provertebral
plate divides into a number of homogeneous, cubical, successive pieces; these are the several primitive vertebræ. They are not numerous at first, but soon increase as the embryo grows longer (Figs. 153–155).
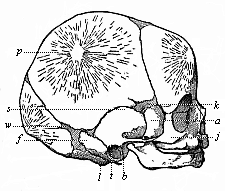 | Fig. 333—Skull of a new-born child. (From Kollmann.) Above, in the three bones of the roof of the skull, we see the lines that radiate from the central points of ossification; in front, the frontal bone; behind, the occipital bone; between the two the large parietal bone, p. s the scurf bone, w mastoid fontanelle, f petrous bone, t tympanic bone, l lateral part, b bulla, j cheek-bone, a large wing of cuneiform bone, k fontanelle of cuneiform bone. |
In all the Craniotes the soft, indifferent cells of the mesoderm, which originally compose the skeletal plate, are afterwards converted for the most part into cartilaginous cells, and these secrete a firm and elastic intercellular substance between them, and form cartilaginous tissue. Like most of the other parts of the skeleton, the membranous rudiments of the vertebræ soon pass into a cartilaginous state, and in the higher Vertebrates this is afterwards replaced by the hard osseous tissue with its characteristic stellate cells (Fig. 6). The primary axial skeleton remains a simple chorda throughout life in the Acrania, the Cyclostomes, and the lowest fishes. In most of the other Vertebrates the chorda is more or less replaced by the cartilaginous tissue of the secondary perichorda that grows round it. In the lower Craniotes (especially the fishes) a more or less considerable part of the chorda is preserved in the bodies of the vertebræ. In the mammals it disappears for the most part. By the end of the second month in the human embryo the chorda is merely a slender thread, running through the axis of the thick, cartilaginous vertebral column (Figs. 182 ch, 329 ch). In the cartilaginous vertebral bodies themselves, which afterwards ossify, the slender remnant of the chorda presently disappears (Fig. 330 ch). But in the elastic inter-vertebral disks, which develop from the skeletal plate between each pair of vertebral bodies (Fig. 329 li), a relic of the chorda remains permanently. In the new-born child there is a large pear-shaped cavity in each intervertebral disk, filled with a gelatinous mass of cells (Fig. 331 a).
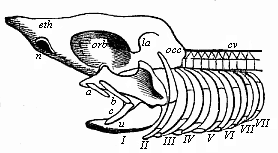 | Fig. 334—Head-skeleton of a primitive fish. n nasal pit, eth cribriform bone region, orb orbit of eye, la wall of auscultory labyrinth, occ occipital region of primitive skull, cv vertebral column, a fore, bc hind-lip cartilage, o primitive upper jaw (palato-quadratum), u primitive lower jaw, II hyaloid bone, III–VIII first to sixth branchial arches. (From Gegenbaur.) |
Though less sharply defined, this gelatinous nucleus of the elastic cartilaginous disks persists throughout life in the mammals, but in the birds and most reptiles the last trace of the chorda disappears. In the subsequent ossification of the cartilaginous vertebra the first deposit of bony matter (“first osseous nucleus”) takes place in the vertebral body immediately round the remainder of the chorda, and soon displaces it altogether. Then there is a special osseous nucleus formed in each half of the vertebral arch. The ossification does not reach the point at which the three nuclei are joined until after birth. In the first year the two osseous halves of the arches unite; but it is much later—in the second to the eighth year—
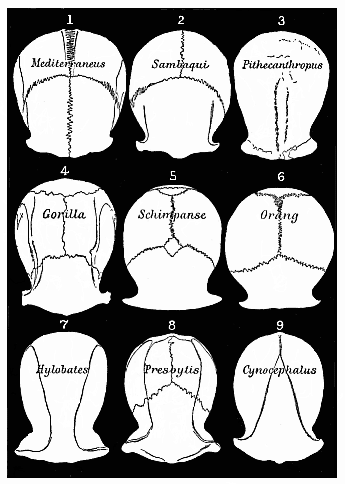 Fig. 335—Roofs of the skulls of nine Primates (Cattarrhines), seen from above and reduced to a common size. 1 European, 2 Brazilian, 3 Pithecanthropus, 4 Gorilla, 5 Chimpanzee, 6 Orang, 7 Gibbon, 8 Tailed ape, 9 Baboon.
Fig. 335—Roofs of the skulls of nine Primates (Cattarrhines), seen from above and reduced to a common size. 1 European, 2 Brazilian, 3 Pithecanthropus, 4 Gorilla, 5 Chimpanzee, 6 Orang, 7 Gibbon, 8 Tailed ape, 9 Baboon. |
that they connect with the osseous vertebral bodies.
The bony skull (cranium), the head-part of the secondary axial skeleton, develops in just the same way as the vertebral column. The skull forms a bony envelope for the brain, just as the vertebral canal does for the spinal cord; and as the brain is only a peculiarly differentiated part of the head, while the spinal cord represents the longer trunk-section of the originally homogeneous medullary tube, we shall expect to find that the osseous coat of the one is a special modification of the osseous envelope of the other. When we examine the adult human skull in itself (Fig. 332), it is difficult to conceive how it can be merely the modified fore part of the vertebral column. It is an elaborate and extensive bony structure, composed of no less than twenty bones of different shapes and sizes. Seven of them form the spacious shell that surrounds the brain, in which we distinguish the solid ventral base below and the curved dorsal vault above. The other thirteen bones form the facial skull, which is especially the bony envelope of the higher sense-organs, and at the same time encloses the entrance of the alimentary canal. The lower jaw is articulated at the base of the skull (usually regarded as the XXI cranial bone). Behind the lower jaw we find the hyoid bone at the root of the tongue, also formed from the gill-arches, and a part of the lower arches that have developed as “head-ribs” from the ventral side of the base of the cranium.
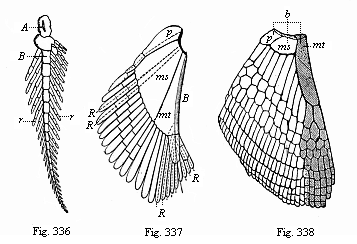 Fig. 336—Skeleton of the breast-fin of Ceratodus (biserial feathered skeleton). A, B, cartilaginous series of the fin-stem. rr cartilaginous fin-radii. (From Gunther.)
Fig. 336—Skeleton of the breast-fin of Ceratodus (biserial feathered skeleton). A, B, cartilaginous series of the fin-stem. rr cartilaginous fin-radii. (From Gunther.)Fig. 337—Skeleton of the breast-fin of an early Selachius (Acanthias). The radii of the median fin-border (B) have disappeared for the most part; a few only (R) are left. R, R, radii of the lateral fin-border, mt metapterygium, ms mesopterygium, p propterygium. (From Gegenbaur.) Fig. 338—Skeleton of the breast-fin of a young Selachius. The radii of the median fin-border have wholly disappeared. The shaded part on the right is the section that persists in the five-fingered hand of the higher Vertebrates. (b the three basal pieces of the fin: mt metapterygium, rudiment of the humerus, ms mesopterygium, p propterygium.) (From Gegenbaur.) |
Although the fully-developed skull of the higher Vertebrates, with its peculiar shape, its enormous size, and its complex composition, seems to have nothing in common with the ordinary vertebræ, nevertheless even the older comparative anatomists came to recognise at the end of the eighteenth century that it is really nothing else originally than a series of modified vertebræ. When Goethe in 1790 “picked up the skull of a slain victim from the sand of the Jewish cemetery at Venice, he noticed at once
that the bones of the face also could be traced to vertebræ (like the three hind-most cranial vertebræ).” And when Oken (without knowing anything of Goethe’s discovery) found at Ilenstein, “a fine bleached skull of a hind, the thought flashed across him like lightning: ‘It is a vertebral column.’”
This famous vertebral theory of the skull has interested the most distinguished zoologists for more than a century: the chief representatives of comparative anatomy have devoted their highest powers to the solution of the problem, and the interest has spread far beyond their circle. But it was not until 1872 that it was happily solved, after seven years’ labour, by the comparative anatomist who surpassed all other experts of this science in the second half of the nineteenth century by the richness of his empirical knowledge and the acuteness and depth of his philosophic speculations. Carl Gegenbaur has shown, in his classic Studies of the Comparative Anatomy of the Vertebrates (third section), that we find the most solid foundation for the vertebral theory of the skull in the head-skeleton of the Selachii. Earlier anatomists had wrongly started from the mammal skull, and had compared the several bones that compose it with the several parts of the vertebra (Fig. 333) they thought they could prove in this way that the fully-formed mammal skull was made of from three to six vertebræ.
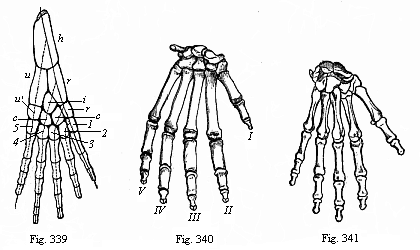 Fig. 339—Skeleton of the fore leg of an amphibian. h upper-arm (humerus), ru lower arm (r radius, u ulna), rcicu′, wrist-bones of first series (r radiale, i intermedium, c centrale, u′ ulnare). 1, 2, 3, 4, 5 wrist-bones of the second series. (From Gegenbaur.)
Fig. 339—Skeleton of the fore leg of an amphibian. h upper-arm (humerus), ru lower arm (r radius, u ulna), rcicu′, wrist-bones of first series (r radiale, i intermedium, c centrale, u′ ulnare). 1, 2, 3, 4, 5 wrist-bones of the second series. (From Gegenbaur.)Fig. 340—Skeleton of gorilla’s hand. (From Huxley.) Fig. 341—Skeleton of human hand, back. (From Meyer.) |
The older theory was refuted by simple and obvious facts, which were first pointed out by Huxley. Nevertheless, the fundamental idea of it—the belief that the skull is formed from the head-part of the perichordal axial skeleton, just as the brain is from the simple medullary tube, by differentiation and modification—remained. The work now was to discover the proper way of supplying this philosophic theory with an empirical foundation, and it was reserved for Gegenbaur to achieve this. He first opened out the phylogenetic path which here, as in all morphological questions, leads most confidently to the goal. He showed that the primitive fishes (Figs. 249–251), the ancestors of all the Gnathostomes, still preserve permanently in the form of their skull the structure out of which the transformed skull of the higher Vertebrates, including man, has been evolved. He further showed that
the branchial arches of the Selachii prove that their skull originally consisted of a large number of (at least nine or ten) provertebræ, and that the cerebral nerves that proceed from the base of the brain entirely confirm this. These cerebral nerves are (with the exception of the first and second pair, the olfactory and optic nerves) merely modifications of spinal nerves, and are essentially similar to them in their peripheral expansion. The comparative anatomy of these cerebral nerves, their origin and their expansion, furnishes one of the strongest arguments for the new vertebral theory of the skull.
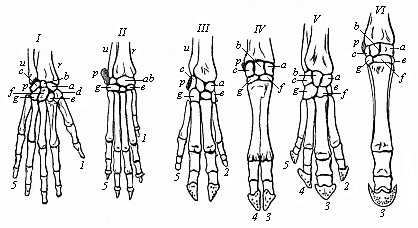 Fig. 342—Skeleton of the hand or fore foot of six mammals. I man, II dog, III pig, IV ox, V tapir, VI horse. r radius, u ulna, a scaphoideum, b lunare, a triquetrum, d trapezium, e trapezoid, f capitatum, g hamatum, p pisiforme. 1 thumb, 2 index finger, 3 middle finger, 4 ring finger, 5 little finger. (From Gegenbaur.)
Fig. 342—Skeleton of the hand or fore foot of six mammals. I man, II dog, III pig, IV ox, V tapir, VI horse. r radius, u ulna, a scaphoideum, b lunare, a triquetrum, d trapezium, e trapezoid, f capitatum, g hamatum, p pisiforme. 1 thumb, 2 index finger, 3 middle finger, 4 ring finger, 5 little finger. (From Gegenbaur.) |
We have not space here to go into the details of Gegenbaur’s theory of the skull. I must be content to refer the reader to the great work I have mentioned, in which it is thoroughly established from the empirico-philosophical point of view. He has also given a comprehensive and up-to-date treatment of the subject in his Comparative Anatomy of the Vertebrates (1898). Gegenbaur indicates as original “cranial ribs,” or “lower arches of the cranial vertebræ,” at each side of the head of the Selachii (Fig. 334), the following pairs of arches: I and II, two lip-cartilages, the anterior (a) of which is composed of an upper piece only, the posterior (bc) from an upper and lower piece; III, the maxillary arches, also consisting of two pieces on each side—the primitive upper jaw (os palato-quadratum, o) and the primitive lower jaw (u); IV, the hyaloid bone (II); finally, V–X, six branchial arches in the narrower sense (III–VIII). From the anatomic features of these nine to ten cranial ribs or “lower vertebral arches” and the cranial nerves that spread over them, it is clear that the apparently simple cartilaginous primitive skull of the Selachii was originally formed from so many (at least nine) somites or provertebræ. The blending of these primitive segments into a single capsule is, however, so ancient that, in virtue of the law of curtailed heredity, the original division seems to have disappeared; in the embryonic development it is very difficult to detect it in isolated traces, and in some respects quite impossible. It is claimed that several (three to six) traces of provertebræ have been discovered in the anterior (pre-chordal) part of the Selachii-skull; this would bring up the number of cranial somites to twelve or sixteen, or even more.
In the primitive skull of man (Fig. 323) and the higher Vertebrates, which has been evolved from that of the Selachii, five consecutive sections are discoverable at a certain early period of development, and one might be induced to trace these to five primitive vertebræ; but these sections are due entirely to adaptation to
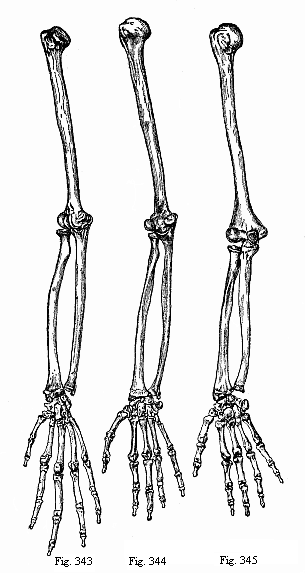 Figs. 343–345—Arm and hand of three anthropoids. Fig. 343—Chimpanzee (Anthropithecus niger). Fig. 344—Veddah of Ceylon (Homo veddalis). Fig. 345—European (Homo mediterraneus). (From Paul and Fritz Sarasin.)
Figs. 343–345—Arm and hand of three anthropoids. Fig. 343—Chimpanzee (Anthropithecus niger). Fig. 344—Veddah of Ceylon (Homo veddalis). Fig. 345—European (Homo mediterraneus). (From Paul and Fritz Sarasin.) |
the five primitive cerebral vesicles, and correspond, like these, to a large number of metamera. That we have in the primitive skull of the mammals a greatly modified and transformed organ, and not at all a primitive formation, is clear from the circumstance that its original soft membranous form only assumes the cartilaginous character for the most part at the base and the sides, and remains membranous at the roof. At this part the bones of the subsequent osseous skull develop as external coverings over the membranous structure, without an intermediate cartilaginous stage, as there is at the base of the skull. Thus a large part of the cranial bones develop originally as covering bones from the corium, and only secondarily come into close touch with the primitive skull (Fig. 333). We have previously seen how this very rudimentary beginning of the skull in man is formed ontogenetically from the “head-plates,” and thus the fore end of the chorda is enclosed in the base of the skull. (Cf. Fig. 145 and pp. 138, 144, and 149.)
The phylogeny of the skull has made great progress during the last three decades through the joint attainments of comparative anatomy, ontogeny, and paleontology. By the judicious and comprehensive application of the phylogenetic method (in the sense of Gegenbaur) we have found the key to the great and important problems that arise from the thorough comparative study of the skull. Another school of research, the school of what is called “exact craniology” (in the sense of Virchow), has, meantime, made fruitless efforts to obtain this result. We may gratefully acknowledge all that this descriptive school has done in the way of accurately describing the various forms and measurements of the human skull, as compared with those of other mammals. But the vast empirical material that it has accumulated in its extensive literature is mere dead and sterile erudition until it is vivified and illumined by phylogenetic speculation.
Virchow confined himself to the most careful analysis of large numbers of human skulls and those of anthropoid mammals. He saw only the differences between them, and sought to express these in figures.
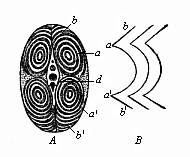 Fig. 346—Transverse section of a fish’s tail (from the tunny). (From Johannes Müller.) a upper (dorsal) lateral muscles, a′, b′ lower (ventral) lateral muscles, d vertebral bodies, b sections of incomplete conical mantle, B attachment lines of the inter-muscular ligaments (from the side).
Fig. 346—Transverse section of a fish’s tail (from the tunny). (From Johannes Müller.) a upper (dorsal) lateral muscles, a′, b′ lower (ventral) lateral muscles, d vertebral bodies, b sections of incomplete conical mantle, B attachment lines of the inter-muscular ligaments (from the side). |
Without adducing a single solid reason, or offering any alternative explanation, he rejected evolution as an unproved hypothesis. He played a most unfortunate part in the controversy as to the significance of the fossil human skulls of Spy and Neanderthal, and the comparison of them with the skull of the Pithecanthropus (Fig. 283). All the interesting features of these skulls that clearly indicated the transition from the anthropoid to the man were declared by Virchow to be chance pathological variations. He said that the roof of the skull of Pithecanthropus (Fig. 335, 3) must have belonged to an ape, because so pronounced an orbital stricture (the horizontal constriction between the outer edge of the eye-orbit and the temples) is not found in any human being. Immediately afterwards Nehring showed in the skull of a Brazilian Indian (Fig. 335, 2), found in the Sambaquis of Santos, that this stricture can be even deeper in man than in many of the apes. It is very instructive in this connection to compare the roofs of the skulls (seen from above) of different primates. I have, therefore, arranged nine such skulls in Fig. 335, and reduced them to a common size.
We turn now to the branchial arches, which were regarded even by the earlier natural philosophers as “head-ribs.” (Cf. Figs. 167–170). Of the four original gill-arches of the mammals the first lies between the primitive mouth and the first gill-cleft. From the base of this arch is formed the upper-jaw process, which joins with the inner and outer nasal processes on each side, in the manner we have previously explained, and forms the chief parts of the skeleton of the upper jaw (palate bone, pterygoid bone, etc.) (Cf. p. 284.) The remainder of the first branchial arch, which is now called, by
way of contrast, the “upper-jaw process,” forms from its base two of the ear-ossicles (hammer and anvil), and as to the rest is converted into a long strip of cartilage that is known, after its discoverer, as “Meckel’s cartilage,” or the promandibula. At the outer surface of the latter is formed from the cellular matter of the corium, as covering or accessory bone, the permanent bony lower jaw. From the first part or base of the second branchial arch we get, in the mammals, the third ossicle of the ear, the stirrup; and from the succeeding parts we get (in this order) the muscle of the stirrup, the styloid process of the temporal bone, the styloid-hyoid ligament, and the little horn of the hyoid bone. The third branchial arch is only cartilaginous at the foremost part, and here the body of the hyoid bone and its larger horn are formed at each side by the junction of its two halves. The fourth branchial arch is only found transitorily in the mammal embryo as a rudimentary organ, and does not develop special parts; and there is no trace in the embryo of the higher Vertebrates of the posterior branchial arches (fifth and sixth pair), which are permanent in the Selachii. They have been lost long ago. Moreover, the four gill-clefts of the human embryo are only interesting as rudimentary organs, and they soon close up and disappear. The first alone (between the first and second branchial arches) has any permanent significance; from it are developed the tympanic cavity and the Eustachian tube. (Cf. Figs. 169, 320.)
It was Carl Gegenbaur again who solved the difficult problem of tracing the skeleton of the limbs of the Vertebrates to a common type. Few parts of the vertebrate body have undergone such infinitely varied modifications in regard to size, shape, and adaptation of structure as the limbs or extremities; yet we are in a position to reduce them all to the same hereditary standard. We may generally distinguish three groups among the Vertebrates in relation to the formation of their limbs. The lowest and earliest Vertebrates, the Acrania and Cyclostomes, had, like their invertebrate ancestors, no pairs of limbs, as we see in the Amphioxus and the Cyclostomes to-day (Figs. 210, 247). The second group is formed of the two classes of the true fishes and the Dipneusts; here there are always two pairs of limbs at first, in the shape of many-toed fins—one pair of breast-fins or fore legs, and one pair of belly-fins or hind legs (Figs. 248–259). The third group comprises the four higher classes of Vertebrates—the amphibia, reptiles, birds, and mammals; in these quadrupeds there are at first the same two pairs of limbs, but in the shape of five-toed feet. Frequently we find less than five toes, and sometimes the feet are wholly atrophied (as in the serpents). But the original stem-form of the group had five toes or fingers before and behind (Figs. 263–265).
The true primitive form of the pairs of limbs, such as they were found in the primitive fishes of the Silurian period, is preserved for us in the Australian dipneust, the remarkable Ceratodus (Fig. 257). Both the breast-fin and the belly-fin are flat oval paddles, in which we find a biserial cartilaginous skeleton (Fig. 336). This consists, firstly, of a much segmented fin-rod or “stem” (A, B), which runs through the fin from base to tip; and secondly of a double row of thin articulated fin-radii (r, r), which are attached to both sides of the fin-rod, like the feathers of a feathered leaf. This primitive fin, which Gegenbaur first recognised, is attached to the vertebral column by a simple zone in the shape of a cartilaginous arch. It has probably originated from the branchial arches.1
We find the same biserial primitive fin more or less preserved in the fossilised remains of the earliest Selachii (Fig. 248), Ganoids (Fig. 253), and Dipneusts (Fig. 256). It is also found in modified form in some of the actual sharks and pikes. But in the majority of the Selachii it has already degenerated to the extent that the radii on one side of the fin-rod have been partly or entirely lost, and are retained only on the other (Fig. 337). We thus get the uniserial fin, which has been transmitted from the Selachii to the rest of the fishes (Fig. 338).
Gegenbaur has shown how the five-toed leg of the Amphibia, that has been inherited by the three classes of Amniotes, was evolved from the uniserial fish-fin.2
1. While Gegenbaur derives the fins from two pairs of posterior separated branchial arches, Balfour holds that they have been developed from segments of a pair of originally continuous lateral fins or folds of the skin.)
2. The limb of the four higher classes of Vertebrates is now explained in the sense that the original fin-rod passes along its outer (ulnar or fibular) side, and ends in the fifth toe. It was formerly believed to go along the inner (radial or tibial) side, and end in the first toe, as Fig. 339 shows.)
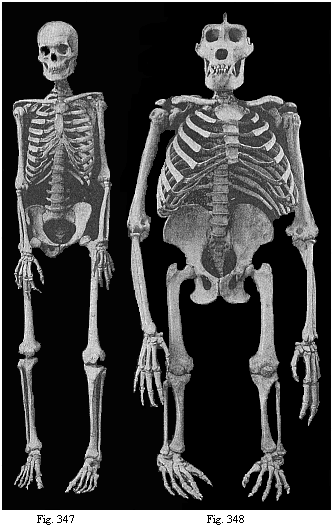 Fig. 347—Human skeleton. (Cf. Figure 326.)
Fig. 347—Human skeleton. (Cf. Figure 326.)Fig. 348—Skeleton of the giant gorilla. (Cf. Figure 209.) |
In the dipneust ancestors of the Amphibia the radii gradually atrophy, and are lost, for the most part, on the other side of the fin-rod as well (the lighter cartilages in Fig. 338). Only the four lowest radii (shaded in the illustration) are preserved; and these are the four inner toes of the foot (first to fourth). The little or fifth toe is developed from the lower end of the fin-rod. From the middle and upper part of the fin-rod was developed the long stem of the limb—the important radius and ulna (Fig. 339 r and u) and humerus (h) of the higher Vertebrates.
In this way the five-toed foot of the Amphibia, which we first meet in the Carboniferous Stegocephala (Fig. 260), and which was inherited from them by the reptiles on one side and the mammals on the other, was formed by gradual degeneration and differentiation from the many-toed fish-fin (Fig. 341). The reduction of the radii to four was accompanied by a further differentiation of the fin-rod, its transverse segmentation into upper and lower halves, and the formation of the zone of the limb, which is composed originally of three limbs before and behind in the higher Vertebrates. The simple arch of the original shoulder-zone divides on each side into an upper (dorsal) piece, the shoulder-blade (scapula), and a lower (ventral) piece; the anterior part of the latter forms the primitive clavicle (procoracoideum), and the posterior part the coracoideum. In the same way the simple arch of the pelvic zone breaks up into an upper (dorsal) piece, the iliac-bone (os ilium), and a lower (ventral) piece; the anterior part of the latter forms the pubic bone (os pubis), and the posterior the ischial bone (os ischii).
There is also a complete agreement between the fore and hind limb in the stem or shaft. The first section of the stem is supported by a single strong bone—the humerus in the fore, the femur in the hind limb. The second section contains two bones: in front the radius (r) and ulna (u), behind the tibia and fibula. (Cf. the skeletons in Figs. 260, 265, 270, 278–282, and 348.) The succeeding numerous small bones of the wrist (carpus) and ankle (tarsus) are also similarly arranged in the fore and hind extremities, and so are the five bones of the middle-hand (metacarpus) and middle-foot (metatarsus). Finally, it is the same with the toes themselves, which have a similar characteristic composition from a series of bony pieces before and behind. We find a complete parallel in all the parts of the fore leg and the hind leg.
When we thus learn from comparative anatomy that the skeleton of the human limbs is composed of just the same bones, put together in the same way, as the skeleton in the four higher classes of Vertebrates, we may at once infer a common descent of them from a single stem-form. This stem-form was the earliest amphibian that had five toes on each foot. It is particularly the outer parts of the limbs that have been modified by adaptation to different conditions. We need only recall the immense variations they offer within the mammal class. We have the slender legs of the deer and the strong springing legs of the kangaroo, the climbing feet of the sloth and the digging feet of the mole, the fins of the whale and the wings of the bat. It will readily be granted that these organs of locomotion differ as much in regard to size, shape, and special function as can be conceived. Nevertheless, the bony skeleton is substantially the same in every case. In the different limbs we always find the same characteristic bones in essentially the same rigidly hereditary connection; this is as splendid a proof of the theory of evolution as comparative anatomy can discover in any organ of the body. It is true that the skeleton of the limbs of the various mammals undergoes many distortions and degenerations besides the special adaptations (Fig. 342). Thus we find the first finger or the thumb atrophied in the fore-foot (or hand) of the dog (II). It has entirely disappeared in the pig (III) and tapir (V). In the ruminants (such as the ox, IV) the second and fifth toes are also atrophied, and only the third and fourth are well developed (VI, 3). Nevertheless, all these different fore-feet, as well as the hand of the ape (Fig. 340) and of man (Fig. 341), were originally developed from a common pentadactyle stem-form. This is proved by the rudiments of the degenerated toes, and by the similarity of the arrangement of the wrist-bones in all the pentanomes (Fig. 342 a–p).
If we candidly compare the bony skeleton of the human arm and hand with that of the nearest anthropoid apes, we find an almost perfect identity. This is especially true of the chimpanzee. In regard to the proportions of the various
parts, the lowest living races of men (the Veddahs of Ceylon, Fig. 344) are midway between the chimpanzee (Fig. 343) and the European (Fig. 345). More considerable are the differences in structure and the proportions of the various parts between the different genera of anthropoid apes (Figs. 278–282); and still greater is the morphological distance between these and the lowest apes (the Cynopitheca). Here, again, impartial and thorough anatomic comparison confirms the accuracy of Huxley’s pithecometra principle p. 171.
The complete unity of structure which is thus revealed by the comparative anatomy of the limbs is fully confirmed by their embryology. However different the extremities of the four-footed Craniotes may be in their adult state, they all develop from the same rudimentary structure. In every case the first trace of the limb in the embryo is a very simple protuberance that grows out of the side of the hyposoma. These simple structures develop directly into fins in the fishes and Dipneusts by differentiation of their cells. In the higher classes of Vertebrates each of the four takes the shape in its further growth of a leaf with a stalk, the inner half becoming narrower and thicker and the outer half broader and thinner. The inner half (the stalk of the leaf) then divides into two sections—the upper and lower parts of the limb. Afterwards four shallow indentations are formed at the free edge of the leaf, and gradually deepen; these are the intervals between the five toes (Fig. 174). The toes soon make their appearance. But at first all five toes, both of fore and hind feet, are connected by a thin membrane like a swimming-web; they remind us of the original shaping of the foot as a paddling fin. The further development of the limbs from this rudimentary structure takes place in the same way in all the Vertebrates according to the laws of heredity.
The embryonic development of the muscles, or active organs of locomotion, is not less interesting than that of the skeleton, or passive organs. But the comparative anatomy and ontogeny of the muscular system are much more difficult and inaccessible, and consequently have hitherto been less studied. We can therefore only draw some general phylogenetic conclusions therefrom.
It is incontestable that the musculature of the Vertebrates has been evolved from that of lower Invertebrates; and among these we have to consider especially the unarticulated Vermalia. They have a simple cutaneous muscular layer, developing from the mesoderm. This was afterwards replaced by a pair of internal lateral muscles, that developed from the middle wall of the cœlom-pouches; we still find the first rudiments of the muscles arising from the muscle-plate of these in the embryos of all the Vertebrates (cf. Figs. 124, 158–160, 222–224 mp). In the unarticulated stem-forms of the Chordonia, which we have called the Prochordonia, the two cœlom-pouches, and therefore also the muscle-plates of their walls, were not yet segmented. A great advance was made in the articulation of them, as we have followed it step by step in the Amphioxus (Figs. 124, 158). This segmentation of the muscles was the momentous historical process with which vertebration, and the development of the vertebrate stem, began. The articulation of the skeleton came after this segmentation of the muscular system, and the two entered into very close correlation.
The episomites or dorsal cœlom-pouches of the Acrania, Cyclostomes, and Selachii (Fig. 161 h) first develop from their inner or median wall (from the cell-layer that lies directly on the skeletal plate [sk] and the medullary tube [nr]) a strong muscle-plate (mp). By dorsal growth (w) it also reaches the external wall of the cœlom-pouches, and proceeds from the dorsal to the ventral wall. From these segmental muscle-plates, which are chiefly concerned in the segmentation of the Vertebrates, proceed the lateral muscles of the stem, as we find in the simplest form in the Amphioxus (Fig. 210). By the formation of a horizontal frontal septum they divide on each side into an upper and lower series of myotomes, dorsal and ventral lateral muscles. This is seen with typical regularity in the transverse section of the tail of a fish (Fig. 346). From these earlier lateral muscles of the trunk develop the greater part of the subsequent muscles of the trunk, and also the much later “muscular buds” of the limbs.1
1. The ontogeny of the muscles is mostly cenogenetic. The greater part of the muscles of the head (or the visceral muscles) belong originally to the hyposoma of the vertebrate organism, and develop from the wall of the hyposomites or ventral cœlom-pouches. This also applies originally to the primary muscles of the limbs, as these too belong phylogenetically to the hyposoma. (Cf. Chapter XIV.)
Title and Contents
Vol. II Title and Contents
Glossary
Chapter XXV
Chapter XXVII
Figs. 1–209
Figs. 210–408