*** START OF THE PROJECT GUTENBERG EBOOK IE TIER- UND PFLANZENWELT DES SüSSWASSERS. ZWEITER BAND. ***
Anmerkungen zur Transkription
Der vorliegende Text wurde anhand der Buchausgabe von
1891 so weit wie möglich originalgetreu wiedergegeben. Typographische
Fehler wurden stillschweigend korrigiert. Ungewöhnliche und heute
nicht mehr verwendete Schreibweisen bleiben gegenüber dem Original
unverändert; fremdsprachliche Ausdrücke wurden nicht korrigiert.
Fußnoten wurden direkt nach dem betreffenden Absatz
eingefügt, Literaturhinweise als Endnoten dagegen am Ende eines
jeden Kapitels. Darüberhinaus haben einige der Autoren dort weitere
nummerierte Literaturhinweise angefügt, welche durch Zahlen gefolgt von
runden Klammern gekennzeichnet werden.
Botanische und zoologische Bezeichnungen werden meist in
kursiver Schrift dargestellt, Personennamen meist
gesperrt. Dies wird
allerdings nicht durchweg konsistent gehandhabt; in der vorliegenden
Bearbeitung wurde dies auch nicht harmonisiert.
Abhängig von der im jeweiligen Lesegerät installierten
Schriftart können die im Original gesperrt
gedruckten Passagen gesperrt, in serifenloser Schrift, oder aber sowohl
serifenlos als auch gesperrt erscheinen.
Die Tier- und Pflanzenwelt des Süsswassers.
Die Tier- und Pflanzenwelt des Süsswassers.
Einführung in das Studium derselben.
Unter Mitwirkung von
Dr. C. Apstein (Kiel), Fr. Borcherding (Vegesack), S. Clessin (Ochsenfurt),
Prof. Dr. F. A. Forel (Morges, Schweiz), Prof. Dr. A. Gruber (Freiburg
i. Br.), Prof. Dr. P. Kramer (Halle a. d. S.), Prof. Dr. P. Ludwig (Greiz),
Dr. W. Migula (Karlsruhe), Dr. L. Plate (Marburg), Dr. E. Schmidt-Schwedt
(Berlin), Dr. A. Seligo (Danzig), Dr. J. Vosseler (Tübingen),
Dr. W. Weltner (Berlin) und Prof. Dr. F. Zschokke (Basel)
herausgegeben
von
Dr. Otto Zacharias,
Direktor der Biologischen Station am Grossen Plöner See in
Holstein
Zweiter Band
Mit 51 in den Text gedruckten Abbildungen.
Leipzig
Verlagsbuchhandlung von J. J. Weber
1891
Alle Rechte vorbehalten.
[S. v]
Vorwort.
Nach der Aufnahme zu urteilen, welche der erste Teil des vorliegenden
Werkes in den nächst interessierten Kreisen sowohl als auch in der
Tagespresse gefunden hat, ist mit unserer „Einführung in das Studium
der Tier- und Pflanzenwelt des Süsswassers“ eine in der biologischen
Litteratur wirklich vorhandene Lücke ausgefüllt worden.
Die Absicht des Herausgebers ist demnach vollständig erreicht; aber
mit der Befriedigung, die er hierüber empfindet, wird sogleich auch
der Wunsch rege, sämtlichen Herren, welche der Aufforderung zur
Mitarbeiterschaft an diesem Buche in so freundlicher Weise entsprochen
haben, beim Erscheinen des Schlussbandes den verbindlichsten Dank für
ihre wertvollen Beiträge abzustatten. Der Unterzeichnete nimmt an,[S. vi]
dass er damit nicht nur seinen eigenen Empfindungen Ausdruck giebt,
sondern zugleich auch im Namen aller Derjenigen spricht, welche eine
umfassende, gemeinverständliche und sichere Unterweisung in
betreff der einheimischen Wasserwelt bisher vermisst haben.
Biologische Station am Plöner See. Ende Juni 1891.
Dr. Otto Zacharias.
[S. vii]
Inhaltsverzeichnis.
Seite
I.
Die Hydrachniden (Wassermilben).
Von Prof. Dr. P. Kramer in Halle.
Geschichtliches. — Stellung der Süsswassermilben zu den übrigen
Milben. — Beschreibung der äussern Gestalt von Piona flavescens. —
Die hauptsächlichsten inneren Organe der Hydrachnidae. — Die typischen
Gruppen der Wassermilben, durch Beispiele erläutert. — Geographische
Verbreitung und Lebensweise. — Entwickelung, erläutert an Nesaeafuscata, Diplodontus filipes und Hydrachna globosa. — Anhang: Tabelle
zur Bestimmung der bis jetzt unterschiedenen Gattungen
Kerfe und Kerflarven des süssen Wassers,
besonders der stehenden Gewässer.
Von Dr. E. Schmidt-Schwedt in Berlin.
Einleitende Bemerkungen. — Vergleich mit den Wassersäugetieren. —
Besondere Wichtigkeit von Atmung und Bewegung. — 1. Käfer:
a) Taumelkäfer. — b) Schwimmkäfer. — c) Kolbenwasserkäfer. —
d) Parnus, Cyphon, Donacia. — 2. Zweiflügler: Larven und Puppen. —
Kennzeichnung derselben: a) Mücken: Culex, Anopheles, Dixa, Corethra,
Mochlonyx, Chironomus, Tanypus, Simulia. — b) Phalacrocera. —
c) Stratiomyden. — d) Eristalis. — 3. Schmetterlingslarven: Paraponyx,
Hydrocampa, Cataclysta. — 4. Netzflüglerlarven: a) Frühlingsfliegen:
Limnophilus, Polycentropus, Hydropsyche. — b) Sialis, Sisyra. —
[S. viii]
5. Geradflüglerlarven: a) Libellen: Agrion-, Libellula-, Aeschna-Gruppe,
Calopteryx, Gomphus. — b) Eintagsfliegen: Chloëon, Caenis. —
c) Afterfrühlingsfliegen: Nemura. — Gegensatz der Netzflügler und
Geradflügler hinsichtlich des Wasserlebens zu den übrigen Ordnungen. —
6. Schnabelkerfe: a) Hydrometriden. — b) Notonecta, Plea. —
c) Corisa. — d) Nepa, Ranatra, Naucoris. — Schlussbemerkungen:
Hinweis auf die Kerfe des Meeres. — Anhang: Tabelle zu annähernder
Bestimmung der Kerflarven des Süsswassers
Die Mollusken des Süsswassers.
Von S. Clessin in Ochsenfurt.
Einteilung der Mollusken. — Wohnorte und Gewohnheiten. — Entwickelung
und Alter der Mollusken. — Anpassungsfähigkeit der
Mollusken. — Die Mollusken der Tiefenfauna. — Höhlen-Mollusken. —
Die Perlenmuschel
Die deutschen Süsswasserfische und ihre
Lebensverhältnisse.
Von Dr. A. Seligo in Heiligenbrunn bei Danzig.
Das Wasser als Lebenselement der Fische. — Das Süsswasser. —
Ausbreitung der Süsswasserfische. — Umgrenzung des zu besprechenden
Gewässergebietes. — Der Ursprung der Cypriniden und Salmoniden. —
Aufzählung der im Gebiete vorkommenden Arten der Knochenfische,
Ganoidfische und Neunaugen und die Verbreitung derselben. — Die
Organe der Fische und ihre Verrichtungen: Haut, Schuppen, Glanz,
Farbe. — Wirbelsäule. — Körperform. — Ortsbewegung, Flossen,
Muskeln. — Leibeshöhle, Zwerchfell, Brusthöhle, Herz, Leber, Nieren,
Milz. — Mundhöhle und Bezahnung. — Kiemen. — Atmung und
Sauerstoffbedürfnis, Fischregionen der Gewässer. — Darm und Magen. —
Verdauung. — Körpertemperatur, Einflüsse der Temperatur des
Mediums. — Nahrung, Fütterung, Wachstum. — Schwimmblase. —
Fortpflanzungsorgane und das Laichen. — Samenfäden und Eier. —
Fortpflanzung des Aals, des Lachses und der Forellen. — Künstliche
Fischzucht. — Teichwirtschaft. — Einführung ausländischer und Ausbreitung
einheimischer Fischarten auf künstlichem Wege. — Brutpflege.
— Gehirn und Sinnesorgane: Auge, Hörorgan, Seitenorgan,
Geschmack und Geruch, Tastorgane. — Fischfang
Die quantitative Bestimmung des Plankton
im Süsswasser.
Von Dr. C. Apstein in Kiel.
Einleitung. — Vertikalnetz. — Filtrator. — Konservierung. — Anwendung
der Apparate. — Volumenbestimmung. — Vorbereitung zur
Zählung. — Stempelpipetten. — Das Hensensche Zählmikroskop. —
Zählung und Protokoll derselben. — Ein Beispiel zur Methodik
Die Fauna des Süsswassers in ihren
Beziehungen zu der des Meeres.
Von Dr. Otto Zacharias in Plön (Holstein).
Das Vorkommen von marinen Gattungen im Süsswasser. — Reliktenseen.
— Eine Meduse als Bewohnerin von Strandseen auf Trinidad. —
Einwanderung von Meerestieren in den Ortoire-Fluss. — Freischwimmende
Muschellarven (Dreyssena polymorpha) im Süsswasser. —
Die Verbreitung der kleinen Wasserfauna durch „passive Wanderung“. —
Der Süsswasser-Monotus. — Die „Fauna relegata“ des Professors
Pavesi. — Der Transport kleiner Wasserorganismen durch Schwimmvögel,
Wasserkäfer und strömende Luft. — Hakenborsten und Klebzellen
der Würmer als Anheftungswerkzeuge. — Das Wandern der
Wasserschnecken und Muscheln. — Spezialisierte Haftorgane bei
Protozoen (Difflugia)
Über die wissenschaftlichen Aufgaben
biologischer Süsswasser-Stationen.
Von Dr. Otto Zacharias in Plön (Holstein).
Die Begründung der „Biologischen Station“ zu Plön. — Vorteile eines
solchen Forschungsinstituts. — Die pelagischen Organismen des Grossen
Plöner Sees. — Die besonderen Aufgaben von Süsswasserstationen. —
Die Winterfauna unserer Binnenseen. — Beobachtung der Wasserinsekten
und der im Wasser lebenden Larven von Landkerbtieren. —
Erforschung der eigentümlichen Fortpflanzungsverhältnisse mancher
Turbellarien und Oligochäten. — Faunistische Exkursionen und vergleichende
Untersuchungen. — Praktische Gesichtspunkte. — Beschreibung
der Plöner Station. — Die Erforschung der böhmischen
Gewässer durch Prof. Anton Fritsch
Das Tierleben auf Flussinseln und am Ufer
der Flüsse und Seen.
Von Fr. Borcherding in Vegesack.
Einleitende Bemerkungen. — Die Säugetiere an und in dem Süsswasser.
— Die Brutvögel. — Die Gäste auf dem Frühjahrs- und
Herbstzuge. — Die Sumpfschildkröte, Emys europaea Gray. — Die
Anuren und Urodelen des süssen Wassers. — Die Fischfauna eines
Flusses, eines Geest- und eines Moorsees. — Die Mollusken an und
in den Gewässern. — Die niedere Tierwelt
Die Hydrachniden oder Süsswassermilben gehören mit ihren auf dem Lande
lebenden Verwandten jenem unermesslichen Heere spinnenartiger Tiere
an, welche im Systeme der Zoologen den Namen Acarida tragen, und deren
Formenreichtum bei einem gewissen gemeinsamen Grundzug der Gestalt ein
ausserordentlich grosser ist.
Noch bis gegen Ende des vorigen Jahrhunderts waren es im ganzen nur
wenige Milben, auf welche sich die Aufmerksamkeit der Beobachter
gelenkt hatte. Sie gehörten zumeist den auf dem Menschen und den
Haustieren lebenden Schmarotzern an und wurden um der Krankheiten
willen, die sie hervorrufen, beachtet und beschrieben. Die erste
grössere Arbeit über andere Acarida lieferte erst der sorgfältig
beobachtende und scharfblickende dänische Naturforscher O. Fr.
Müller, indem er im Jahre 1781 eine grosse Zahl der in Dänemark
einheimischen Süsswassermilben abbildete und die vortrefflichen
Abbildungen durch kurze Erklärungen erläuterte. So war es also gerade
die uns beschäftigende Milbengruppe, welche zuerst mit einer für lange
Zeit unerreichten Vollständigkeit behandelt worden ist.
Manches Jahrzehnt hindurch geschah darnach für die nähere Kenntnis der
Acariden wenig Entscheidendes. Erst die Arbeiten Dugès’ 1834
und des Forstrats C. L. Koch 1837–1850 bezeichnen einen neuen
wesentlichen Fortschritt. So brachte namentlich des ersteren eingehende
Darstellung die Kenntnis der Entwickelung von Hydrachna globosa,
während letzterer durch die[S. 4] grosse Zahl der von ihm beobachteten
Milben, unter denen sich auch sehr zahlreiche Süsswassermilben
befanden, zuerst überhaupt eine Vorstellung von dem Reichtum der
Milbenklasse gab, wenn auch freilich die nicht hinreichende Genauigkeit
seiner Abbildungen und Beschreibungen dem Systematiker noch heute viel
Mühe bereitet.
In der neuesten Zeit ist den Acariden ein immer grösseres Interesse
entgegengebracht worden, wenn auch lange noch nicht in dem Masse,
als es die in mehr als einer Hinsicht merkwürdige Lebens- und
Entwickelungsgeschichte derselben verdient. Allerdings sind die
Beobachtungsobjekte meist sehr klein und schwierig zu behandeln und
daraus mag sich der im ganzen langsame Fortschritt unserer Kenntnis
über diese Tiergruppe erklären. Die Süsswassermilben bilden aber
noch die am wenigsten Schwierigkeiten bietende Gruppe und laden
durch ihre zierliche Gestalt und Munterkeit des Wesens, auch durch
verhältnismässige Grösse zur Beobachtung ein. Auch sind sie fast
überall reichlich zu finden, wo nur irgend fliessendes oder stehendes
Wasser Jahr für Jahr vorhanden ist.
Die meisten anderen Milben bleiben unserem Auge in der Regel
verborgen, obwohl es kaum einen Ort geben dürfte, wo einem genaueren
Beobachter nicht irgend ein charakteristischer Vertreter dieser
Tiergruppe begegnete. Zumeist möchte wohl eine feuchte Umgebung dem
Leben dieser der Mehrzahl nach zarten Geschöpfe günstig sein, aber
doch wird man auch an den kahlen, in trockenster Luft des Sommers
am Wege liegenden Steinen nicht umsonst nach einer mit zierlichem
Stechapparat versehenen blauroten Acaride (Bryobia speciosa)
suchen, der sich noch manche Vertreter unserer zierlichen Panzermilben
(Oribatidae) anschliessen. Milben finden sich unter Laub und
Steinen, im Moose und auf den Blättern der Bäume, auf und unter ihrer
Rinde und im anbrüchigen Holze, auf und unter der Haut zahlreicher
kalt- und warmblütiger Tiere, auf den Federn der Vögel, ja sogar in
denselben: wo sich nur überhaupt irgendwelche Nahrung darbieten mag,
sei sie natürlichen oder künstlichen Ursprungs, überall begegnen wir
Milben, ihre Leibesgestalt oft in wunderbarer Weise[S. 5] dem Aufenthaltsort
anpassend und ihre Lebensgewohnheiten einrichtend nach den Bedingungen,
die derselbe bietet.
Bei der immer noch vorhandenen sehr unvollständigen Kenntnis auch
unserer heimatlichen Milben ist es noch nicht möglich gewesen, eine
sogenannte natürliche Anordnung dieser Tiere vorzunehmen, d. h. eine
solche, bei welcher die Abstammung, die gegenseitige Verwandtschaft
ausschlaggebend ist, doch heben sich schon einige grössere
Verwandtschaftskreise aus dem Schwarm der überhaupt hierhergehörigen
Tiere ab. So bilden die soeben schon erwähnten Panzermilben, denen
jeder Sammler am häufigsten im Moose begegnet, einen in sich völlig
abgeschlossenen Stamm. Sie sind reine Landbewohner, und wenn man auch
in jüngster Zeit im Meere einige Vertreter gefunden haben will, so ist
das doch mit Vorsicht aufzunehmen. Ebenso stellen die Gamasidae,
diejenigen Milben, zu welchen die auf den Dungkäfern so häufig
scharenweise anzutreffenden braunen Acariden gehören, eine wohl
abgeschlossene Gruppe dar. So vielgestaltig aber auch die Wohnstätten
derselben sind, in das Wasser ist doch keins davon hinabgestiegen. Zwar
hat man einen ihrer Vertreter in den wohl gewiss mit Wasser stets und
reichlich bespülten Nasengängen einer Seehundsart aufgefunden, auch
haben eifrige Naturforscher unter den durch die Flutwelle regelmässig
überspülten Steinen des Seestrandes einige Gamasiden entdeckt, aber
wirkliche Wassertiere haben wir damit doch nicht vor uns. Nicht besser
steht es mit den zahllosen Geschlechtern der die Haut und die Federn
der Vögel oder die Haare der kleinen Säugetiere bevölkernden Milben
(Sarcoptidae) oder denjenigen, welche dem grossen Stamm der
durch die Mehlmilbe gekennzeichneten Acariden (Tyroglyphidae)
angehören. Wenn sie auch meist der Feuchtigkeit als einer notwendigen
Voraussetzung ihres Lebens bedürfen, so sind sie doch niemals Bewohner
unserer Teiche und Flüsse geworden. Einzig und allein diejenige Gruppe
unter den Milben, denen ich die allgemeine Bezeichnung Vorderatmer
gegeben habe, weil sie ihre beiden kleinen Luftlöcher ganz vorn an
dem kegelförmig hervorspringenden Mundabschnitt führen, bietet uns
Beispiele von auch dem Leben im Wasser angepassten[S. 6] Milben dar.
Diese Vorderatmer werden am besten durch die so häufig in unseren
Gärten am Fusse der Obstbäume anzutreffende Samtmilbe (Trombidium
fuliginosum) veranschaulicht. Trombidiumartige Milben also sind es,
welche in grosser Zahl die süssen Gewässer, nur mit wenigen Arten die
See bewohnen.
Eine Naturgeschichte dieser Hydrachniden muss vor allen Dingen ein Bild
der äusserlich wahrnehmbaren Gestalt entwerfen und so mag sich denn
zunächst darauf die Aufmerksamkeit richten.
Ein Zug mit dem Fangnetz durch das klare Wasser eines Teiches
fördert in der Regel ausser zahlreichen kleinen Krustern auch manche
undurchsichtige und durchsichtige Hydrachnide zutage. Wir entnehmen
eine Milbe der letzteren Sorte, es ist eine Piona flavescens,
und betrachten sie, nachdem sie in ein Uhrgläschen übergeführt ist,
zunächst mit der Lupe.
Fig. 1. Piona flavescens, von der Seite gesehen.
Der rundliche Rumpf ist, wie wir bald bemerken, ungeteilt, oben
hochgewölbt, unten abgeflacht, so wie Fig. 1 es zeigt. Bei den meisten
Hydrachniden hat er diese Gestalt, nur ausnahmsweise treffen wir einen
flachen Rumpf, einen stark in die Länge gezogenen, oder einen durch
besondere Anhänge am hinteren Rande ausgezeichneten. Der vorderen
Hälfte der Unterfläche entspringen die acht Füsse.
[S. 7]
Wie schlank und zierlich sind diese hellen völlig durchsichtigen
Füsschen, die das Tier oft in ganzer Länge von sich streckt und so eine
Zeit lang still und unbeweglich liegen bleibt, um mit einem plötzlichen
Ruck eine Strecke fortzueilen. Wieder liegt es still da und fängt nach
einer kurzen Ruhe an langsam auf dem Grunde fortzukriechen. Jetzt eilt
es wieder, sich wie im Wirbel überschlagend, in hastiger Bewegung eine
Strecke fort, um bald zu ruhen, bald langsam im Wasser zu wandeln.
Betrachten wir die einzelnen Füsse noch genauer, so fallen besonders
an den hinteren Paaren lange, seidenglänzende Haarborsten auf,
welche gedrängt stehen und leicht beweglich sind. Die zahlreichsten
bemerken wir am vorletzten und drittletzten Gliede der beiden hinteren
Fusspaare. Es sind dies die für unsere Süsswassermilben ganz besonders
charakteristischen Schwimmborsten, und von ihrer Anzahl, ihrer Breite
und Stellung hängt zum grossen Teil die Gewandtheit und Schnelligkeit
ab, mit der sich die Tierchen im Wasser bewegen. Als Regel können
wir annehmen, dass bei den erwachsenen Milben die vorderen Füsse nur
wenige, die hinteren dagegen zahlreiche Schwimmborsten führen, und dass
wiederum an jedem Fusse, der überhaupt welche besitzt, das vorletzte
Glied die meisten, die dem Körper näheren Glieder immer weniger solche
Borsten tragen. Ausnahmen von dieser Regel sind allerdings beobachtet.
So bemerkt man, dass bei einer der grössten einheimischen Arten,
einer der schnellsten und gewandtesten Schwimmerinnen, Eylaïs
extendens, welche als tiefrote Jägerin die Wasser durcheilt, das
vierte Fusspaar gar keine Schwimmborsten besitzt. Das Tier hat sich
daher gewöhnt, den letzten Fuss jeder Körperseite beim Schwimmen ruhig
nach hinten gestreckt zu tragen. Diese Haltung giebt ein untrügliches
Erkennungsmittel für die soeben namhaft gemachte, in unseren
stillstehenden Gewässern häufigere Milbe ab. Einigen Wassermilben
fehlen die Schwimmborsten sogar gänzlich. Sie sind dadurch gezwungen,
eine durchaus kriechende Lebensweise zu führen, und leben meist im
Schlamme verborgen.
Wie wenig übrigens der Besitz oder Mangel von Schwimmborsten
eine Verwandtschaft zwischen manchen in diesem einen[S. 8] Punkte
übereinstimmenden Milben mit sich bringt, beweisen zwei Gattungen,
deren Vertretern die Schwimmborsten fehlen, nämlich Bradybates
und Limnochares (die Tiefschreiter und Schlammfreunde). Diese
Milben sind in ihrer ganzen Erscheinung ausserordentlich verschieden
von einander. Erstere möchte wohl als das Urbild einer, der Lebensweise
im Wasser angepassten Samtmilbe angesehen werden können, so vollständig
wiederholt sie Zug um Zug die Gestalt dieser Landmilbe. Ganz anders
bietet sich Limnochares dem Beobachter dar. Ein unförmlicher
linsengrosser roter Klumpen, welcher mühsam von den auffallend kurzen
dünnen Füssen fortgeschleppt wird und das langgezogene schnabelförmige
Mundstück beim Zurückziehen ganz in sich aufzunehmen vermag, ist sie
ganz darauf angewiesen, dass die Tragkraft des Wassers ihre Muskeln bei
der Fortbewegung ihres Leibes unterstützt.
Hervorgehoben mag noch werden, dass das Schwimmborstensystem
am ausgebildetsten bei den Hartmilben des süssen Wassers, den
Hartschwimmern, deren hervorstechendste Gattung Arrenurus
(Hartschwanz) ist, gefunden wird. Hier zeigt jeder der beiden
Hinterfüsse eine doppelte Reihe solcher Borsten.
Steht es nun auch gewiss fest, dass die Schwimmfertigkeit durch die
Schwimmborsten wesentlich bedingt ist, so beweist doch die Beobachtung
derjenigen Milben, welche die tiefen Regionen des Genfersees
bewohnen, dass damit die Frage nach den Gründen des Schwimmens noch
nicht ganz erledigt ist. Dort im Genfersee, und vermutlich auch in
anderen sehr tiefen Seen der Schweiz, hat Prof. Forel in den
tiefsten Schichten eine Hydrachnide gefunden, welche sich trotz
aller vorhandenen Schwimmborsten nicht imstande zeigt, den Boden
zu verlassen und schwimmend die Oberfläche zu erreichen. Wird eine
solche Milbe in ein noch so flaches Gefäss gethan, so vermag sie nur
auf dem Boden zu kriechen. Wird ihr, wenn man sie an die Oberfläche
gezogen hat, der Unterstützungspunkt genommen, so sinkt sie trotz allen
Fussbewegungen, die sie ausführt, doch auf den Boden zurück. Offenbar
hat sie sich unter dem Einfluss des bereits erheblichen Wasserdrucks
auf dem Boden des Genfersees entwöhnt, von ihren Schwimmborsten
einen richtigen[S. 9] Gebrauch zu machen, sie hat ihre Füsse niemals im
Schwimmen geübt und hat so, trotz den vorhandenen Schwimmborsten,
die zum Schwimmen notwendigen kräftigen Bewegungen dauernd verlernt.
Es scheint also auch ein richtiger Gebrauch der sonst zum Schwimmen
hinreichend ausgerüsteten Füsse vorausgesetzt werden zu müssen,
damit die Schwimmborsten eine dem Wasser hinreichenden Widerstand
entgegensetzende Fläche bilden und so die Fortbewegung bewirken können.
Wir wenden uns nun wieder der Gestalt unserer Piona zu. Während
die gewölbte Oberseite derselben dem beobachtenden Blick ausser den
deutlich wahrnehmbaren dunklen Augenpunkten wenig bietet, wird er,
nachdem man die Milbe auf den Rücken gelegt hat, bei Betrachtung der
Unterseite von einigen besonderen Organen gefesselt.
Auf der Bauchfläche bemerkt man nämlich vier von einander getrennt
stehende härtere Hautplatten, und an jeder sind zwei Füsse
eingelenkt[I]. Bei näherer Betrachtung ergiebt es sich, dass jede der
Platten aus zwei mit einander verschmolzenen Plättchen zusammengesetzt
ist und dass jedes der acht Plättchen die Gelenkhöhle für das Hüftglied
je eines der acht Füsse trägt. Wir nennen die Plättchen die Hüftplatten
der Füsse oder die Epimeren. Unsere Piona flavescens lässt
erkennen, dass die Hüftplatten für die beiden vierten Füsse, rechts
und links, die umfangreichsten sind. Das ist nicht immer der Fall.
Überhaupt finden wir in der Ausbildung und gegenseitigen Gruppierung
dieser Epimeren eine so ausserordentliche Mannigfaltigkeit, dass nicht
mit Unrecht die hieraus sich ergebenden Unterschiede zur schärferen
Trennung der Gattungen verwendet worden sind.
Zwischen den Epimeren der beiden vorderen Füsse befindet sich der
Mundapparat, in der Regel gestützt durch eine vermutlich als Unterlippe
zu deutende, erhärtete Platte. Diese Platte trägt zunächst die meist
fünfgliedrigen Taster.
[S. 10]
Dieselben dienen wohl hauptsächlich dem Gefühlssinn, werden aber
auch zum Festhalten der Beute benutzt. Die Mundöffnung selbst wird
in der Regel in der Tiefe eines oben offenen, eine kurze Halbröhre
darstellenden kopfähnlichen Anhangs gefunden, in welchem sich als
wesentlichstes Mundwerkzeug die Kaukiefer eingelassen finden. Diese
Kiefer bilden einen wichtigen Anhalt zur Unterscheidung grösserer
Abteilungen unter den Süsswassermilben, indem sie bei einigen Gattungen
eingliedrig sind und die Gestalt eines kräftigen nach vom gerichteten
Stachels haben, bei anderen dagegen aus zwei Gliedern bestehen, von
denen das zweite einer mit der Spitze nach oben gewendeten Klaue oder
Kralle gleicht. Ganz derselben Verschiedenheit begegnet man auch bei
derjenigen Gruppe der Landmilben, welche ich bereits oben als die
den Süsswassermilben am nächsten stehende bezeichnet habe, bei den
Trombididen.
Was für Gliedmassen oder Teile ausserdem noch zu dem Mundapparat
gehören, ist noch durch weitere Beobachtungen genauer zu erforschen.
Es giebt nach Ansicht einiger Beobachter noch ein zweites Kieferpaar,
welches dieselben in gewissen verhärteten Stäbchen entdeckt zu haben
glauben, während andere diese Deutung jener Stäbchen leugnen. Es ist
daher auch die verwandtschaftliche Stellung der Milben durchaus noch
nicht allgemein festgestellt. Die einen, z. B. G. Haller,
wollen sie zu den krebsartigen Tieren ziehen, indem das Vorhandensein
einer grössern Anzahl von Kiefern dafür geltend gemacht wird. Die
anderen sind der Meinung, dass die Milben den spinnenartigen Tieren
zuzuzählen sind. Sicherlich kann hierüber lediglich aus anatomischen
Gründen ein sicheres Urteil nicht gezogen werden, vielmehr muss
die Entwickelungsgeschichte ein entscheidendes Wort mitsprechen.
Vielleicht gelingt es zunächst aus den an Gamasiden noch anzustellenden
Beobachtungen, hierüber mehr Licht zu verbreiten.
Auf der Unterseite unserer Milben begegnen wir nun noch der
Geschlechtsöffnung und der Afteröffnung. Umfangreich, wenigstens beim
weiblichen Geschlechte, ist die erstere, meist punktförmig klein die
zweite, jedoch fehlt letztere nirgends, wie irrtümlich[S. 11] behauptet
worden. Die Geschlechtsöffnung ist bei den allermeisten Gattungen
von eigentümlichen, entweder napfartigen oder porenartigen Gebilden
begleitet, welche meist neben ihr zu Gruppen vereinigt auf besonderen
verhärteten Plättchen aufgestellt oder in die weiche Haut eingebettet
sind, in einzelnen Fällen aber auch auf der innern Fläche der die
Öffnung schliessenden Klappen ihren Platz gefunden haben. Die Abbildung
dieser Gebilde, wie sie sich beim Männchen und Weibchen von Nesaea
fuscata finden, in Fig. 3i und k, giebt wohl eine
hinreichend deutliche Vorstellung davon. Stehen diese sogenannten
Haftnäpfe neben der Geschlechtsöffnung, so dürften sie kaum noch ihrem
Zweck als Haft- oder Tastorgan genügen, finden sie sich dagegen auf der
innern Deckklappenfläche, so ist ihre Funktion noch unbeeinträchtigt.
Die einzelnen Gattungen der Wassermilben zeigen in der Zahl, Anordnung
und Grösse dieser Näpfe eine so ausserordentliche Verschiedenheit,
dass sie hierdurch häufig mit grosser Leichtigkeit von einander
unterschieden werden können. Es gehören diese Haftnäpfe zu einer
ganz besonders charakteristischen Eigentümlichkeit gerade unserer
Süsswassermilben, so dass es gerechtfertigt erscheint, noch einen
Augenblick bei ihnen zu verweilen. Es sind offenbar umgebildete
Oberhautporen. Betrachtet man nämlich die Oberfläche einer
Süsswassermilbe genauer, so findet man dieselbe mit einer Anzahl
regelmässig verteilter grösserer Porenöffnungen versehen, welche
meistens zu kleinen, häufig aber auch sehr umfangreichen Hautdrüsen
führen. Bei sehr vielen Milben ist die nächste Umgebung einer solchen
Pore verhärtet, so dass dort eine kleine Platte in die Haut eingelassen
scheint, auch wird die Öffnung regelmässig von einer Haarborste
begleitet. Diese Hautdrüsen mögen wohl eine Flüssigkeit absondern,
welche anderen Wassertieren Ekel erregt, so dass sie die Wassermilben
nicht verspeisen mögen. Hat man doch beobachtet, dass unsere Milben von
den Fischen verschmäht werden. Welchen Vorrat an solcher Flüssigkeit
diese Hautdrüsen, wenn sie enorm entwickelt sind, enthalten, lässt
sich bei den Hartmilben (Arrenurus) erkennen. Wird eine solche
in Spiritus gelegt, so fährt nach kurzer Zeit der Drüseninhalt wie
ein langer sich[S. 12] kräuselnder Faden aus der Pore hervor, so dass die
ganze Milbe völlig eingewickelt wird. Nun ist es wohl nicht falsch
geurteilt, wie bereits oben angedeutet wurde, wenn wir annehmen, dass
die Haftnäpfe auf der Unterseite des Hinterleibes ursprünglich solche
Hautdrüsenöffnungen gewesen sind. Die ursprünglichen Drüsengebilde sind
aber in Haftorgane umgewandelt und dementsprechend hat sich die Öffnung
samt der Platte, in welcher sie steht, umgestaltet. Meistens bemerkt
man noch deutlich innerhalb des Umrisses eines solchen Napfes die
Porenöffnung. In anderen Fällen ist sie aber auch schon geschwunden,
was darauf hindeutet, dass das Organ seine Funktion wieder eingebüsst
hat.
Wenn ich soeben bemerkte, dass diese Haftnäpfe eines der wesentlicheren
Merkmale unserer Süsswassermilben darstellen, so ist damit dennoch
nicht gesagt, dass es unter ihnen nicht auch solche gäbe, die überhaupt
dieser Organe entbehrten. Die Mannigfaltigkeit der Formen, die wir
allerwärts im Tierreiche finden und die dem ordnenden Zoologen oft
so viele Schwierigkeiten entgegenstellt, findet sich auch bei den
winzigen, das süsse Wasser belebenden Acariden. Die Kräfte der Natur
lassen sich in kein Schema und System zwingen, sondern gestalten die
Formen nach den vorhandenen Lebensbedingungen aus. So mag es denn wohl
sein, dass die Masse der das süsse Wasser bevölkernden Milben aus
mehreren verschiedenen Hauptstämmen erwuchs, deren Ursprung wir nicht
mehr deutlich unterscheiden können. Der eine Stamm, der namentlich
alle schwimmenden Süsswassermilben umfasst, hat Haftnäpfe auf der
Bauchfläche erzeugt, der andere, zu denen einige kriechende Gattungen
zu rechnen wären, hat solche Näpfe nicht hervorgebracht. Es ist auch
bemerkenswert, dass die ersten Larvenstadien, also die eben dem Ei
entschlüpften Jungen, keine Haftnäpfe besitzen.
Im Vorhergehenden sind die wesentlichsten Organe und die
hauptsächlichsten Erscheinungen der äussern Gestalt kurz berührt. Es
wird später sich noch Gelegenheit bieten, manches, was jetzt übergangen
werden musste, so z. B. die besonderen Kennzeichen und Abzeichen
der Männchen im Gegensatz zu den Weibchen, zur[S. 13] Sprache zu bringen
und so das Bild der äussern Gestalt zu vervollständigen. Auch die
Beschaffenheit der äussern Körperhaut, ob sie hart oder weich ist, ob
glatt oder mit Fortsätzen versehen, hat wohl für die Beurteilung der
äussern Gestalt im weitern Sinne Bedeutung, kann jedoch in einer nur
kurz schildernden Darstellung bloss andeutungsweise berührt werden.
Ebenso ist es mit dem Grössenverhältnis der Glieder zum Rumpfe, indem
nur erwähnt werden kann, dass der Rumpf wie bei Atax gegen
die ungemein grossen Füsse fast verschwindet, während bei anderen
Gattungen, wie beispielsweise Axona, die Füsschen kaum über den
Rand des Rumpfes hervorragen. Dies Alles und manches Andere würde einer
Spezialgeschichte der Hydrachniden Stoff zu ausführlichen Betrachtungen
geben. Wir aber verlassen die äussere Gestalt und wenden uns einer
weitern wichtigen Angelegenheit zu, der Darstellung der inneren Organe.
Die inneren Organe sind in neuester Zeit mehrfach eingehend beobachtet
und beschrieben worden. Wir finden in der Regel eine sehr feine, nach
hinten sich erweiternde Speiseröhre, deren vorderer Abschnitt zu einem
Saugapparat umgewandelt ist, indem Muskelstränge sich an der obern und
untern Röhrenfläche ansetzen, welche diesen Teil des Organs abwechselnd
erweitern und verengern können. Der Magen ist sehr umfangreich und
besitzt bis fünf blindsackartige Ausstülpungen, deren Oberfläche mit
einer Schicht von Leberzellen ausgerüstet ist. Der Enddarm ist in der
Regel wiederum ein Kanal von ziemlicher Enge. Die Speiseröhre steht in
ihrem vordersten Abschnitt in Zusammenhang mit den nicht unerheblichen
Speicheldrüsen. Diese sind in der Regel in mehrfacher Anzahl vorhanden,
führen jedoch immer nur jederseits zu einer einzigen Ausflussöffnung.
Der Enddarm seinerseits nimmt in seinem letzten Abschnitt den
Ausführungsgang der charakteristischen Exkretionsdrüse auf, welche
bei allen Milben zu einer so ausserordentlichen Entwickelung gelangt
ist. Diese Drüse liegt oberhalb des Magens und besitzt einen längeren
Hauptstamm, welcher sich etwa in der Mitte des Rückens in zwei Äste
gabelt. Da der Inhalt dieser Exkretionsdrüse merklich durch die Haut
der Mehrzahl der Hydrachniden[S. 14] hindurchschimmert, so hat sie von jeher
die Aufmerksamkeit der Beobachter erregt, aber auch solche, die auf die
Färbung der Milben ein allzugrosses Gewicht legten, oft irregeführt, da
je nach der Füllung der Farbeneindruck ein sehr verschiedener sein kann.
Das Atmungssystem wird bei den Hydrachniden durch zwei in
ausserordentlich zahlreiche Tracheenfäden auseinanderfahrende
Tracheenstämme dargestellt, welche in einem Paar von Luftlöchern
ausmünden. Diese letzteren liegen zwischen den Einlenkungsstellen der
Kiefer in einer kleinen Platte. Der Hauptstamm jeder Trachee ist mit
einem Spiralfaden versehen, während die zarten Tracheenfäden selbst
keine solchen besitzen.
Das Tracheensystem tritt auch bei den Hydrachniden gerade so wie bei
allen anderen Milben, wo es überhaupt zur Ausbildung gelangt, erst nach
der ersten Häutung auf, die sechsfüssigen Larven besitzen noch keine
Andeutung davon. Die Luftatmung der Wassermilben giebt uns hier nun
Veranlassung, einen kurzen Blick auf andere luftatmende Wassertiere zu
werfen.
Vergleicht man das Betragen der hierher gehörigen verschiedenen, dem
niedern Tierreich angehörenden Bewohner des süssen Wassers, so bemerkt
man bald einen sehr in die Augen fallenden Unterschied. Die Wasserkäfer
z. B. und die Mehrzahl der Wasserwanzen vermögen nur kurze Zeit zu
tauchen. Immer wieder müssen sie die zum Atmen nötige Luft unmittelbar
aus der über dem Wasser stehenden Atmosphäre schöpfen und deshalb
häufig die Oberfläche aufsuchen. Auch vielen Mückenlarven geht es nicht
besser, sie fahren unruhig bald in die Höhe, bald in die Tiefe. Dagegen
sind die in das Wasser eingewanderten Milben völlig und ausschliesslich
Wassertiere geworden. Sie bleiben stets unter der Oberfläche, trotzdem
dass sie ein ausgebildetes Luftatmungssystem besitzen. Sie hüllen
sich auch nicht etwa, wie zahlreiche Uferkäfer und die merkwürdige
grosse Wasserspinne (Argyroneta aquatica), in einen dichten
Mantel von Luft, den sie unausgesetzt mit sich führen und immer
wieder erneuern, sondern leben gerade wie die Larven von zahlreichen
Libellen, Frühlingsfliegen und Mücken nur und allein im Wasser. Ihr
Luftröhrensystem ist trotzdem, dass sie niemals[S. 15] die Oberfläche
aufsuchen, mit Luft durchaus angefüllt. Es weist uns diese Beobachtung
auf einen wohl noch nicht ganz aufgeklärten Naturvorgang. Die Frage,
auf welche Weise die Luft in die Atmungskanäle der Süsswassermilben
gelangt und wie sie sich, nachdem sie etwa durch Atmen verbraucht
worden ist, wieder erneuert, ist es, welche dabei noch der Lösung
harrt. Unter ganz ähnlichen Verhältnissen atmen die durchsichtigen
Larven der bei uns häufigen Büschelmücke (Corethra plumicornis).
Bei dieser hat der bekannte Zoologe A. Weissmann zuerst
beobachtet, dass in den beiden grossen Paaren von Luftsäcken, welche
dem Tier zur Aufrechterhaltung des Gleichgewichts beim Schwimmen zu
dienen scheinen, die Luft während eines bestimmten Zeitpunktes der
Entwickelung ganz von selbst auftritt, ohne dass in der Wandung jener
Blasen auch nur die geringste Öffnung vorhanden wäre oder irgend
eine Verbindung derselben mit der Oberhaut des Tieres bestünde. Die
Luftfüllung tritt plötzlich auf und verbleibt dann in den Blasen. Bei
den Wassermilben, welche nach der ersten Häutung zahlreiche Tracheen
besitzen, könnte es vielleicht ebenso sein, indem die Luft sich ganz
von selbst innerhalb der vorher mit Flüssigkeit angefüllten Luftkanäle
bildet. Denn es ist in der That schwer denkbar, dass die im Wasser
verteilte Luft sich in die feine Ausmündungsstelle der Luftkanäle
eindrängen könne, um von da aus in die zuletzt überaus feinen Fäden
zu gelangen. Ein Saugapparat ist bisher an den Atmungsröhrchen noch
nicht beobachtet und allein ein solcher könnte die Luft dazu vermögen,
die Flüssigkeit aus den Luftröhren zu verdrängen. Genug, wir erwarten
von der Zukunft hier, wie noch für so manche Vorkommnisse, eine
befriedigende Erklärung der Thatsachen.
Die Hydrachniden sind getrennten Geschlechtes. Die samenerzeugenden
Organe des Männchens sind wohl in der Regel paarig und haben
einen gemeinsamen Ausführungsgang, welcher mit einem besondern
Begattungsorgan versehen ist. Die Eierstöcke der Weibchen sind zwar
auch paarig, jedoch sind sie mit ihren Enden derart verschmolzen,
dass sie ein einziges ringartiges Organ darstellen; die beiden
Eileiter sind jedoch zunächst getrennt und bilden erst kurz vor der
Geschlechtsöffnung einen kugeligen Uterus.
[S. 16]
Das Nervensystem besteht in einem zentralen sogenannten Schlundganglion
und den von diesem ausgehenden Nervensträngen. Das Schlundganglion
wird der Länge nach von der Speiseröhre durchsetzt, wodurch es in ein
oberes und ein unteres geteilt wird, die indes so nahe an einander
gerückt sind, dass sie eine gemeinsame Masse bilden, an welcher keine
Schlundkommissur auffindbar ist[II].
[II] Dr. v. Schaub, „Anatomie von Hydrodroma“, p. 29.
Vom obern Schlundganglion gehen die Nerven zu den Augen und
den Mundteilen, von dem untern dagegen zu den Füssen und den
Geschlechtsorganen. Die Nervenfäden spalten sich übrigens häufig und
lösen sich mit Ausnahme der Augennerven in eine Fülle von überallhin
sich verbreitenden Nervenfasern auf.
Das Nervensystem möge uns noch einen Augenblick länger fesseln. So
klein und zum Teil winzig unsere Süsswassermilben auch sind, so
entbehren sie doch der die Aussenwelt auffassenden Sinne nicht.
Gesichts- und Gefühlssinn haben sogar eine hohe Entwickelungsstufe
erreicht. In der Regel finden wir bei den Hydrachniden zwei Paare
von Augen. Diese stehen entweder weit von einander getrennt, wie bei
jener oben genauer besprochenen Piona, oder sie sind auf der
Mitte des Vorderrückens dicht an einander gestellt, so dass sie an
die Augen der Weberknechte (Phalangidae) erinnern. Mag nun
die Anordnung die eine oder die andere sein, jedesmal findet sich in
einem Milbenauge eine Linse, welche das Licht nach einem hinter ihr
gelegenen Punkte bricht, und eine becherförmig gestaltete Netzhaut
mit dunklem Pigment, welche die Lichteindrücke zur Empfindung bringt.
Der Sehnerv leitet dieselben dann dem oben erwähnten Gehirn zu. Ob
nun die Milbe wohl einen Gesamteindruck von ihrer Umgebung durch
ihre Augen gewinnt? Jedenfalls kommt bei Beantwortung dieser Frage
der allgemeine Stand des Seelenlebens in Betracht und nicht bloss
die physikalische Vollkommenheit des Sehapparats. Dass sie aber
scharfe und deutliche Eindrücke durch ihren Gesichtssinn erhalten,
scheint mir aus der grossen Raschheit hervorzugehen, mit welcher die
Mehrzahl der Süsswassermilben,[S. 17] welche des Schwimmens kundig sind,
entgegenstehenden Hindernissen ausweichen. Allerdings wird hier, wie
auch sonst wohl vielfach im Tierreiche, der Gesichtssinn offenbar durch
den sehr entwickelten Tastsinn unterstützt. Bei unseren Milben sind
Tastorgane in ganz ähnlicher Weise, wie bei den übrigen Gliedertieren,
über den ganzen Körper verbreitet und ragen als Haartaster über die
Oberfläche des Leibes hervor. Man wird wohl nicht irregehen, wenn
man die in grosser Regelmässigkeit, wenn auch meist in ziemlich
spärlicher Anzahl auf der Haut der Tierchen allenthalben zerstreuten,
meist kurzen Haarborsten als ebenso viele Tastorgane ansieht. Ebenso
ist ein grosser Teil der auf den Beinen und namentlich den Tastern
befindlichen Haarborsten zu den Fühlhaaren zu rechnen. An zahlreichen
grossen Borsten, welche die ersten Füsse schmücken, hat man die
Einrichtung solcher Fühlhaare genauer untersucht. Die Haarborste steht
in einer kleinen Pore der Oberhaut; sie ist selbst zumteil hohl und
mit lebendiger Substanz ausgefüllt, welche durch die Pore hindurch
mit dem Innern des Milbenkörpers in Zusammenhang steht. Unmittelbar
unter das in der Porenöffnung stehende Ende der Borste hat sich ein
Nervenfaden hinbegeben und dort ein kleines Knötchen gebildet, welches,
sobald durch irgend eine äussere Veranlassung die Borste gedrückt
oder gezerrt wird, durch das untere Borstenende gestreift und in
Mitleidenschaft gezogen werden muss. Hierdurch kommt die Tastempfindung
und zugleich auch jedenfalls eine Ortsbestimmung zu stande. Die
Oberhaut des Tierchens ist, da sie aus einer festen widerstandsfähigen
Masse gebildet ist, an sich einer Empfindung nicht fähig. Dieser
Mangel wird durch jene Tastborsten fast völlig ausgeglichen. Man hat
übrigens auch in dem der Haut von innen anliegenden Zellengewebe
zahlreiche Nervenelemente entdeckt und so ist es auch möglich, dass ein
an irgend einer Stelle auf die weiche und nachgiebige Hautfläche bei
den weichhäutigen Hydrachniden ausgeübter Druck auch ohne besondere
Tasthaare eine Gefühlsempfindung auslöst.
In ganz besonderer Weise sind, wie sich wohl vermuten lässt, die Taster
mit Gefühlshaaren ausgerüstet. Es ist dies um so[S. 18] notwendiger, als die
meist weit vom Munde und auf der Stirnwölbung aufgestellten Augen die
Umgebung des Mundes nicht zu übersehen vermögen. Von den Mundteilen
kann also nur nach bestimmten Gefühlseindrücken gehandelt werden. Diese
werden durch dicht aufgestellte Tasthaare vermittelt.
Ähnlich ist auch die Unterseite der Füsse und zwar namentlich ihrer
Endglieder mit zahlreichen kurzen Haaren versehen, welche beim
Schreiten und wohl auch beim Schwimmen, wie ferner beim Ergreifen der
Beute die Tastempfindungen veranlassen.
So haben die unscheinbaren und auf den ersten Blick willkürlich über
den Körper verstreuten Haarborsten einen wichtigen Beruf.
Ob die Wassermilben Gehörs- und Geruchsempfindungen besitzen, ist nicht
beobachtet. Geschmacksempfindungen werden sie ganz gewiss haben, da es
höchst wahrscheinlich ist, dass sie ihre Beute auch durch den Geschmack
unterscheiden.
Ob man nun auch von einem seelischen Leben unserer Geschöpfe sprechen
darf? Einen bestimmten Charakter hat jedenfalls jede Wassermilbe im
Vergleich mit anderen. Darunter kann ich hier freilich nur die Art
und Weise ihres Betragens verstehen. Die Schnelligkeit, mit welcher
die Bewegungen ausgeführt werden; die Feindschaft oder Freundschaft,
um diese Worte hier zu gebrauchen, welche den übrigen Mitgliedern
einer und derselben Art entgegengebracht werden; die Gewandtheit, mit
welcher eine solche Milbe einer drohenden Gefahr zu entrinnen sucht,
alles dieses, namentlich das Zuletzterwähnte, lassen den Schluss
gerechtfertigt erscheinen, dass wir bei diesen kleinen Geschöpfen ein
verhältnismässig hoch entwickeltes Seelenleben voraussetzen dürfen.
Wenn sich nun auch diese seelischen Regungen wohl zumeist auf den
Erwerb der Nahrung, auf Sicherstellung des einzelnen Tieres und auf
Erhaltung der Art richten, so ist doch das Vorhandensein derselben von
ausserordentlichem Interesse und fordert zu immer neuen Beobachtungen
auf, damit auch für diese niedrigen Geschöpfe die Tiefe und Ausdehnung
des ihnen verliehenen seelischen Lebens immer mehr bekannt werde.
[S. 19]
Wir wenden uns nun einer kurzen Besprechung der hauptsächlichsten
Gattungen zu.
An der Hand der oben gegebenen Beschreibung von Piona flavescens
und unter Benutzung der beigefügten Abbildung in Fig. 1 ist jeder
leicht imstande sich eine hinreichend genaue Vorstellung von der
Gestalt einer Hydrachnide überhaupt zu machen. Jedoch wird der
Naturfreund, der es unternimmt, die häufigeren Mitglieder unserer
Tierfamilie in den stehenden und fliessenden Gewässern seiner Heimat
genauer zu betrachten, bald bemerken, dass er in der That recht
verschiedenartige Geschöpfe in sein Aquarium bringt. Wir wollen ihm in
dem Nachfolgenden einige Winke für das Unterscheiden der gesammelten
Tierchen geben, verweisen aber zu einer eingehenderen Betrachtung
auf unsern Anhang. Vieles lässt sich schon mittels einer schärferen
Lupe deutlich genug erkennen, ganz wird es indessen nicht ohne die
Zuhilfenahme eines Mikroskops abgehen. Und gerade die mikroskopische
Betrachtung der hier uns beschäftigenden, zumteil recht kleinen
Geschöpfe entfaltet eine solche Fülle zierlicher Formen vor unseren
Augen, dass wir unwillkürlich von der Begierde ergriffen werden, immer
neue Gestalten einer solchen zu unterwerfen.
Die im Anhang gegebene Tabelle nimmt ihre Merkmale nicht mit Rücksicht
auf die gegenseitige Verwandtschaft der Hydrachniden, sondern mit
Rücksicht auf Deutlichkeit und leichtes Auffassen. Hier dagegen möchte
ich versuchen, die hauptsächlichsten Gruppen herauszuheben.
Betrachtet man die klaren Wasser eines Grabens oder kleinen Teiches
aufmerksam, so geschieht es wohl, dass man ein linsengrosses,
blutrotes, kugelförmig aufgeschwollenes Geschöpf in kräftigem Zuge
durch die stille Flut eilen sieht, lebhafte Wendungen bald hierin
bald dorthin ausführend. Ein schneller Griff mit dem Fangnetz und die
Schwimmerin ist in Gefangenschaft geraten. Wir entlassen sie in eine
bereitgehaltene flache Schale und können sie nun mit blossem Auge
deutlich erkennen und prüfen. Entweder streckt sie den vierten Fuss
jederseits beim Schwimmen unbeweglich nach hinten, oder sie macht auch
mit ihm, also mit sämtlichen[S. 20] acht Füssen lebhafte Schwimmbewegungen.
Im erstern Falle haben wir eine unserer ansehnlichsten und häufigsten
Süsswassermilben vor uns, einen Repräsentanten der oben schon erwähnten
Gattung Eylaïs[III]. Die Milbe ist eine arge Räuberin und
überfällt mit ihren kräftigen Füssen und den zwar ausserordentlich
kleinen, aber überaus kräftigen Kiefern, was ihr in den Weg kommt.
Sie bildet eine ganz besondere Abteilung der Hydrachniden für sich,
da bei ihr die Mundöffnung und die damit zusammenhängende Ausbildung
der Kiefer eine ganz eigentümliche geworden ist. Wie eine sorgfältige
Betrachtung des Mundes mit dem Mikroskop lehrt, hat die ganze Gestalt
und Anordnung desselben eine überraschende Ähnlichkeit mit einer
Saugscheibe. Man bemerkt eine kreisrunde Platte, eingerahmt durch einen
Kranz zierlicher Haarborsten, in der Mitte die winzige Mundöffnung
tragend, aus welcher die schwärzlichen Spitzen der Kieferendglieder in
Gestalt zweier scharfer Zähnchen etwas hervorgeschoben werden können.
Gegen diese Zähnchen drücken die kräftigen Taster und Füsse ihre Beute,
welche alsbald die tödliche Wunde empfängt. Betrachten wir das in
ausgewachsenem Zustande bis 4 mm lange Tier von oben her, so
wird man auf dem Vorderrücken, und zwar ziemlich nahe der Mittellinie
vier schwarze, dicht bei einander stehende Augenpunkte gewahr werden.
Lösen wir mit einem scharfen Messer die dieselben tragende Hautstelle
ab und richten nun das Mikroskop auf sie, so erblicken wir die zu den
einzelnen Augen gehörigen Linsen, jede in eine besondere dickwandige
Hautkapsel eingeschlossen. Es giebt nur wenige Hydrachniden, bei denen
die Augen in der angegebenen Art und Weise angeordnet und geschützt
sind. Auffallend ist es und mag hier gleich erwähnt werden, dass die
eben dem Ei entschlüpften jungen Eylaïs weit von einander
getrennt aufgestellte Augenpunkte führen, dass also die soeben
erwähnte eigentümliche und von der Regel abweichende Augenstellung
erst in späteren Stadien ihres Lebens und zwar nach der ersten Häutung
beobachtet wird. Es ist dies eine Thatsache, welche bei Beurteilung
der[S. 21] Verwandtschaft unserer Gattung auch mit Landmilben nicht ohne
Bedeutung ist.
Hatte die gefangene und in die Glasschale entlassene Milbe, wie eine
kurze Betrachtung bald ergeben wird, auch am vierten Fusse jeder Seite
dichte Büschel von Schwimmborsten, so werden wir eine Vertreterin
einer zweiten wichtigen Gruppe von Süsswassermilben vor uns haben,
und zwar der Gruppe, nach welcher die ganze Familie ihren Namen
bekommen zu haben scheint. Sie ist dann ein Mitglied der Gattung
Hydrachna[IV], welche in mehreren sehr stattlichen Arten unsere
Gewässer bevölkert.
Die Gattung Hydrachna ist ausgezeichnet durch ihre sehr
eigenartige Mundbildung. Keine andere Süsswassermilbe hat einen
ähnlich gebauten, zu einem Stechorgan umgestalteten Schnabel, welcher
sanft gekrümmt in ansehnlicher Länge zwischen den kurzen Tastern
hervorragt. Dieser Schnabel wird durch die Unterlippe gebildet und
stellt eine oben offene Halbröhre dar, ist selbst scharf zugespitzt,
dennoch aber nicht als eine gefahrdrohende Waffe anzusehen, so wenig
wie die beiden haarscharfen messerartigen eingliedrigen Kiefer,
welche in jener Rinne auf- und abgeschoben werden können. Die Milbe
scheint nicht von tierischer Nahrung zu leben, sondern benutzt ihr
umfangreiches Stechorgan dazu, um Pflanzenstiele anzubohren. Ihre Eier
wenigstens legt sie in Höhlungen, welche sie mittels ihres Schnabels
in Blattstiele von Wasserpflanzen eingebohrt hat. Der kugelförmig
aufgetriebene Körper wird von den weit nach vorn gerückten Füssen nicht
besonders schnell durch das Wasser getragen, er scheint sogar häufig
durch sein Gewicht einen hemmenden Einfluss auszuüben, wenigstens
wird oftmals die Unterseite des Tieres beim Schwimmen von oben her
sichtbar. Auch Hydrachna besitzt, gerade wie Eylaïs,
Augen, welche paarweise in eine mit wulstigen Rändern versehene,
harte Kapsel eingeschlossen sind, jedoch stehen die beiden Augenpaare
weit von einander entfernt. Die Bauchfläche trägt jederseits von der
Geschlechtsöffnung eine Platte mit zahlreichen Haftnäpfen, jedoch sind
letztere klein und unansehnlich.
[S. 22]
Im Gegensatze zu den beiden soeben erwähnten Gattungen, denen in
gewissem Sinne noch zwei andere beigesellt werden können, nämlich
Hydrodroma und Limnochares[V], weil sie sechsfüssige
Larven von ausgesprochenem Trombidium-Charakter besitzen, bilden
die dann noch übrigen etwa 24 Hydrachniden-Gattungen eine grössere
Abteilung für sich, da sie viel Verwandtschaftliches zeigen und daher
zunächst auch noch zusammengefasst werden können.
Das Hauptmerkmal dieser Gruppe dürfte wohl darin bestehen, dass die
sechsfüssige Larve einen eignen Typus hat, den ich den Nesaea-Typus
nenne, und welcher durch die Abbildung in Fig. 3a S. 39
dargestellt ist. Da jedoch diese Larven nicht immer leicht zu
beschaffen sind, so müssen wir wohl damit zufrieden sein, die Milben
nach der Form der Kiefer näher zu bezeichnen. Als Beispiel für die
ganze eben in Rede stehende Süsswassermilbenabteilung gilt die
oben näher beschriebene und abgebildete Piona flavescens.
Dieselbe besitzt Kiefer, wie sie in Fig. 3b zur Anschauung
gebracht sind. Dieselben sind zweigliedrig und das letzte Glied
besitzt die Gestalt einer Klaue, wonach die ganze Kieferart den Namen
klauenförmiger Kiefer erhalten hat. Dieselben werden von der Milbe so
getragen, dass die Klauen mit ihrer Spitze nach oben schlagen. Die
ganze Menge der hierhergehörigen Milben zerfällt in zwei grössere
Heerlager, welche wir am einfachsten nach ihrer äussern Körperhülle
in sogenannte Hartschwimmer und Weichschwimmer zu trennen haben.
Hartschwimmer sind solche Süsswassermilben, deren Körper durch eine
feste, allseitig geschlossene wahre Panzerhaut geschützt ist, während
die Weichschwimmer eine solche Verhärtung der Haut nicht aufzuweisen
haben. Bemerkenswert ist es allerdings, dass auch die jüngsten, dem Ei
entschlüpften Larven der Weichschwimmer, soweit sie zur Beobachtung
gekommen sind, eine wenigstens auf dem Rücken ziemlich weitgehende
Hautverhärtung aufzuweisen haben, welche sich aber in späteren Stadien
wieder verliert. Diese Beobachtung wird zur Beurteilung der wahren
verwandtschaftlichen Verhältnisse künftighin wohl nicht ganz ausser
Acht gelassen werden können.
[S. 23]
Zu den Hartschwimmern gehört namentlich die ausserordentlich
artenreiche Gattung Arrenurus[VI]. Es scheint keinen Teich oder
See zu geben, für welchen sich nicht eine eigentümliche Art dieser
merkwürdigen Gattung aufweisen liesse. Sie ist ganz besonders der
Aufmerksamkeit wert, weil es nach den bisherigen Beobachtungen den
Anschein hat, als wäre sie besonders der Veränderlichkeit unterworfen.
Grünrote, braune, ja ganz bunte Arten derselben wimmeln häufig durch
einander und bereiten, wenn man nur Weibchen vor sich hat, dem
Untersucher fast unüberwindliche Schwierigkeiten. Diese verschwinden
aber sogleich, sobald die Männchen mit ihren so ganz eigentümlich
gebauten Hinterleibsanhängen zu Gebote stehen.
Es bietet sich hier eine passende Gelegenheit, den geschlechtlichen
Unterschieden in der äussern Gestalt, welche bei Arrenurus in
ausserordentlich hohem Masse zu Tage treten, etwas Aufmerksamkeit zu
widmen. Es erscheint wunderbar, dass innerhalb einer und derselben
Tiergruppe diese Unterschiede in so ungleichem Masse ausgebildet
sind, denn während sie bei der eben erwähnten Gattung in einem höchst
bedeutenden Grade vorhanden sind, treten sie bei vielen anderen, die
mit ihr unter nahezu gleichen Bedingungen leben, eigentlich völlig
zurück. Eine Erklärung dieser Thatsachen ist bis jetzt nicht gut zu
erwarten gewesen. Vielleicht lüftet eine fortgesetzte Beobachtung den
über dieser ganzen Frage der geschlechtlichen Formverschiedenheiten
noch ruhenden Schleier. Jedenfalls ist bei Arrenurus diese
Verschiedenheit der Geschlechter am weitesten fortgeschritten und hat
ausser den Füssen, wo sie auch bei anderen Gattungen beobachtet wird,
noch den Rumpf ergriffen, indem ausser ansehnlichen kegelförmigen
Fortsätzen auf dem Rücken namentlich der Hinterrand des Leibes in
mannigfacher Weise umgestaltet ist. In den Abbildungen Fig. 2 S. 24
haben wir einige Formen solcher Schwanzanhänge zur Anschauung gebracht.
Es lassen sich dabei offenbar ganz bestimmte typische Gestalten, die
alsdann in freier Weise variieren, unterscheiden.
[S. 24]
Fig. 2. a Arrenurus calcarator.b Arrenurus caudatus.c Arrenurus albator.
So unterscheiden wir eine langgestreckte Schwanzform, wie sie unter
Fig. 2b von Arrenurus caudatus abgebildet ist, von einer
fischschwanzähnlichen, bei welcher die hinteren Seitenecken in zwei
ansehnliche Zipfel ausgezogen sind, zwischen denen in einer mittleren
Abteilung oft sonderbar gestaltete Fortsätze und Blättchen auffallen.
Ein Beispiel hierfür ist Fig. 2c, Arrenurus albator.[S. 25]
Eine dritte Form bietet im ganzen an sich unbedeutendere Anhänge,
welche keine leicht bestimmbare Gestalt besitzen und daher der
Beschreibung Schwierigkeiten entgegenstellen. Einen solchen Anhang
zeigt Fig. 2a von Arrenurus calcarator.
Diese Anhänge wie überhaupt die besonderen geschlechtlichen
Eigentümlichkeiten der Gestalt treten erst nach der letzten Häutung
des Tieres hervor und sind auch dann nicht gleich voll entwickelt,
sondern scheinen erst nach und nach ihre endgültige Form anzunehmen.
Ausser durch ihren Schwanzanhang sind die Männchen von Arrenurus
noch durch eine den Weibchen abgehende Bewaffnung des vierten Fusses
jeder Seite ausgezeichnet. Beiden Geschlechtern gemeinsam ist jedoch
die bereits oben berührte starke Ausstattung der hinteren Füsse mit
Schwimmborsten, welche es ihnen ermöglicht, mit ausserordentlich
kräftigen Bewegungen das Wasser zu durcheilen. Ihre Nahrung besteht
vornehmlich aus den kleinsten Bewohnern des süssen Wassers, den
Rädertierchen, Daphniden etc., sie sind leicht in Gefangenschaft zu
halten, selbst in sehr kleinen Aquarien. Drollig nehmen sich beim
Schwimmen die Arten mit einem langen Schwanzanhang aus, indem die
verhältnismässig schwachen Füsse es nicht verhindern können, dass der
langgestreckte Hinterleib gleich einem Pendel fortwährend hin und her
schwankt, was dem Schwimmen einen ungeschickten Anschein verleiht.
Die Grösse der bei uns vorkommenden Arrenurus-Arten ist
ausserordentlich verschieden und schwankt zwischen 2 und ½ mm.
Neben dieser sehr artenreichen Gattung Arrenurus sind noch
einige weniger in die Augen fallende harthäutige Hydrachniden
erwähnenswert, so die kleinen bunten Vertreter der Gattung
Axona[VII]. Sie sind zwar mit unbewaffnetem Auge kaum
wahrzunehmen, zeichnen sich aber einesteils durch ihre lebhafte
Färbung, anderseits aber auch durch die ganz besonders merkwürdige
Gestalt, welche dem vierten Fusse des Männchens eigentümlich ist, aus.
Offenbar ist dieses anhangsreiche Glied dem Männchen gegeben,[S. 26] um das
Weibchen festhalten zu können, doch mag es auch ein blosser Schmuck
sein, der die Weibchen fesselt und anzieht, wenn man auch zweifelhaft
sein könnte, ob diese winzigen Tiere ein so hohes geistiges Leben
entfalten, dass man von Freude am Schmuck bei ihnen reden kann. Dass
jedoch die Milben an und für sich auch tieferen Gefühlsbewegungen nicht
unzugänglich sind, tritt sehr deutlich bei einer kleinen Landmilbe
(Cheyletus) hervor, welche ihre in Häufchen zusammengelegten
Eier nicht nur bis zum Ausschlüpfen der Jungen nicht verlässt, sondern
dieselben auch gegen Angriffe tapfer verteidigt. Wenn dies auch das
einzige mir bis jetzt bekannte Beispiel eines höheren geistigen Lebens
bei unserer Tierfamilie ist, so ist es gerade hinreichend, um überhaupt
ein solches bei derselben zu beweisen.
Es bleibt nun noch als letzte Gruppe diejenige zu erwähnen übrig,
welche ich im Gegensatze zu den soeben erwähnten Hartschwimmern als die
Weichschwimmer bezeichnet hatte.
Gemeinsames Kennzeichen ist bei den erwachsenen Tieren dieser Gruppe
eine weiche Körperhaut. Die zahlreichen Gattungen derselben bieten
sonst allerdings sehr mannigfaltige Verschiedenheiten, doch dürfte es
selbst dem geübteren Beobachter ziemliche Schwierigkeiten bereiten,
diese Unterschiede zu bemerken.
Als Repräsentant dieser Gruppe muss wiederum jene oben ausführlicher
erwähnte und auch abgebildete Piona dienen, und es mag
genügen, hier auf sie zu verweisen. Nur einen Punkt möchte ich auch
bei dieser Gruppe besonders hervorheben, nämlich noch einmal die
Gestaltverschiedenheiten, welche man zwischen Männchen und Weibchen
derselben Art beobachtet. Hier sind es besonders und vor allem
die Füsse, welche bei beiden Geschlechtern in auffallender Weise
verschieden gestaltet sind, indem die Männchen mancherlei besondere
Anhänge und Umformungen einzelner Fussglieder aufzuweisen haben.
Wie bei zahlreichen Männchen von Insekten sind nämlich die Füsse hier
in Fangorgane umgestaltet, um die flüchtigen Weibchen festzuhalten.
Allerdings findet dies auch nicht bei allen hierhergehörigen Gattungen
statt, auch ist für zahlreiche Arten überhaupt[S. 27] das Männchen noch
nicht beobachtet, wo es aber bekannt geworden ist, da dient zugleich
auch die besondere Gestalt der Füsse zu einer verhältnismässig
leichten Unterscheidung der oft zahlreichen Arten. So muss es als
feststehend gelten, dass die in unseren Gewässern neben der Gattung
Arrenurus am häufigsten vorkommende Gattung Nesaea bei
allen ihren zahlreichen Arten im männlichen Geschlecht ein umgeformtes
drittes und viertes Fusspaar besitzt. Jeder Fuss des dritten Paares
hat ein keulenförmig gestaltetes Endglied, an welchem auch die Kralle
eine von der gewöhnlichen abweichende Gestalt besitzt, wie sie Fig.
3g von Nesaea fuscata zeigt. Weiter ist, was auch
schon bei geringer Vergrösserung bemerkt werden kann, das drittletzte
Glied an den Füssen des vierten Fusspaares hufeisenförmig eingebogen
und trägt an den Rändern der Einbuchtung einen Kranz starrer und
auffallender Haarborsten (Fig. 3h).
Einer andersgearteten Umgestaltung des vierten Fusses begegnet man bei
den Piona-Männchen. Es würde jedoch zu weit führen, wenn hier noch mehr
in Einzelheiten eingegangen würde.
Jedoch noch einer besonderen Erscheinung, welche bei Männchen und
Weibchen einer kleinen Gruppe von Gattungen beobachtet wird, kann ich
nicht unterlassen Erwähnung zu thun, weil sie zeigt, wie sonderbar oft
die Richtung zu sein scheint, in welcher die Umformung der Gestalt
fortschreitet. Hier handelt es sich auch um eine Eigentümlichkeit der
Füsse. Es ist Regel, dass die Hydrachniden an sämtlichen acht Füssen
deutliche und wohl ausgebildete Krallen tragen. Die Abbildung Fig.
3d zeigt eine solche von Nesaea fuscata. Jene kleine
Gruppe von Milben, zu denen unter anderen die in unseren Gewässern
häufig gefundene Gattung Limnesia[VIII] gehört, hat nun diese
Krallen an den beiden vierten Füssen durchaus eingebüsst. Hier
endigt das letzte Fussglied mit einer stumpfen kegelförmigen Spitze.
Allerdings beobachtet man leicht, dass bei zahlreichen Wassermilben
die Krallen an den vierten Füssen ungleich kleiner sind als namentlich
am zweiten und dritten[S. 28] Fusspaar aber sie sind stets gut ausgebildet
und zeigen auch die für die Gattung charakteristische Form. Wie lässt
es sich hier nun erklären, dass sie bei Limnesia völlig fehlen.
Nur als Vermutung könnte angeführt werden, dass Limnesia das
vierte Fusspaar niemals zum Festklammern des Körpers benutzt, sondern
stets in schwingende Bewegung setzt, sobald sie vom Schwimmen ausruht,
vielleicht um das Wasser um den Leib in Zirkulation zu bringen.
Im Anschluss an die soeben, wenn auch nur in flüchtigen Umrissen
gegebene Übersicht der Hauptformen unserer Süsswassermilben möge ein
kurzes Wort über ihren Aufenthalt, ihre Verbreitung und allgemeine
Lebensweise folgen. Zwar sind auch über diesen Punkt die Beobachtungen
nur wenig umfassend, aber so weit sie ein Urteil zulassen, darf
man wohl sagen, dass die Hydrachniden stehende klare Gewässer den
fliessenden vorzuziehen scheinen. Auch trifft man in grösseren
Wasserbecken, deren Ufer durch die offenbar von Wind und Wellenschlag
herrührenden zerstörenden Einflüsse des Wassers mit absterbenden
Pflanzenresten bedeckt, auch häufig mit moderndem Schlamm überzogen
sind, viel seltener Milben an, als in den kleinen mit dichtem
Wasserpflanzengebüsch durchsetzten Weihern und Teichen. Hier, wo die
kleinen Kruster, wie Daphniden und Cyclopiden, ihr Wesen treiben, wo
die Mücken und zarten Netzflügler ihre Eier massenhaft ablegen, wo
zahllose Infusorien an den Wasserpflanzen auf- und niederfahren, da
finden unsere zumeist vom Raube lebenden Hydrachniden ein geeignetes
Jagdgebiet, welches sie in allen Stadien ihrer Entwickelung in meist
rastloser Eile durchlaufen, den Beobachter in Erstaunen setzend über
die Ausdauer und Kraft ihrer Muskeln, welche, am Tage und oft auch
des Nachts angestrengt, dennoch nicht ermüden und in gleichmässiger
Schnelligkeit den Körper von Ort zu Ort führen. Doch ist es nicht nur
die Reichhaltigkeit der Nahrung, es ist auch die bald höher steigende
Temperatur solcher stehenden Gewässer, welche offenbar unseren Milben
sehr angenehm ist. Beobachtet man doch, dass, wenn im Hochsommer die
kleinen Wasserbecken bis auf zwanzig und mehr Grad erwärmt werden,
die Scharen, namentlich der Arrenurus-Arten, ganz ausserordentlich[S. 29]
anwachsen. Wie die Ameisen im Sonnenbrand nur um so rastloser ihren
Zwecken und Pflichten nachgehen und für den Beschauer ein Schwindel
erregendes Gewimmel hervorbringen, so jagen sich die roten, grünen und
bunten Hartschwimmerarten durch die untergetauchten Wasserpflanzen
und fallen massenhaft dem Sammler ins Netz. Übereinstimmend hiermit
ist die von einem Beobachter gemachte Bemerkung, dass die höher im
Gebirge gelegenen Teiche gewöhnlich arm an Wassermilben sind, weil
die Temperatur derselben selbst im Sommer eine verhältnismässig
niedrige ist. Dass die Milben allerdings auch in kaltem Wasser gut
zu leben vermögen, beweist der Umstand, dass man schon sehr früh
im Jahre, wenn das Eis noch auf dem Wasser steht, reichliche Beute
findet und zwar nicht bloss erwachsene Tiere, sondern solche auf allen
Entwickelungsstadien. Daher ist es wohl möglich, dass die niedrigere
Temperatur der Gebirgsteiche die den meisten Milben zur Nahrung
dienenden Kruster, sowie andere zartere Geschöpfe, die von ihnen
verfolgt werden, nicht recht zur Entwickelung kommen lässt, so dass
das Fehlen zahlreicher Milben erst hieraus zu erklären wäre. Diese
letztere Ansicht wird vielleicht durch eine Beobachtung unterstützt,
welche aus südlicheren Gegenden stammt, wo überhaupt wohl auch in
kühlerem Wasser reicheres Leben zu finden ist, so dass auch Milben
darin nicht zu darben brauchen. So hat der französische Naturforscher
Th. Barrois während einer der Erforschung der Azoren gewidmeten
Reise eine Hydrachnide beobachtet, über deren Lebensgewohnheiten er
sich folgendermassen auslässt: „Ich fand diese Art stets in rasch
fliessendem, wenig tiefem Wasser der Quell- und Sturzbäche, welche
von den Bergen herabkommen, um entweder in einen See, was indessen
nicht oft vorkommt, oder sogleich ins Meer sich zu ergiessen, und
deren Bett zahlreiche Kieselsteine enthält. Obgleich die Tiere sehr
gute Schwimmer sind, so habe ich sie niemals mit dem Netz gefangen;
sie leben vielmehr auf der Unterseite der Steine, wo sie in Gruppen
von fünf, sechs, zehn und noch mehr zusammen sich in die Löcher der
basaltigen Laven festsetzen, um nicht von dem Strome mit fortgerissen
zu werden. Die Temperatur dieser Bäche und Flüsschen[S. 30] ist sehr niedrig
und steigt auch im August und September höchstens auf 15½°.
Die Verbreitung dieser Art in vertikaler Richtung bietet grosse
Unterschiede. Ich habe sie fast unmittelbar am Meer gesammelt und
auch in einer Höhe von 800 Metern“. Hierbei wird noch eines besonders
merkwürdigen Umstandes wie folgt gedacht: „Obwohl die beobachtete
Milbe in grosser Menge in gewissen Giessbächen lebt, welche sich in
Seen ergiessen, so findet sie sich in diesen Seen selbst niemals. Zur
Erklärung dieses gewiss auffallenden Vorkommnisses lässt sich Folgendes
etwa anführen: Unsere Hydrachnide liebt sehr flache, sprudelnde,
reine Gewässer. Man trifft sie niemals in Lachen. Nun sind die Seen
ruhig und führen weniger klares und reines Wasser als die Bäche, denn
die darin befindlichen Steine sind meist mit einer mehr oder weniger
dicken Schicht von Schlamm bedeckt, welchen man sehr selten an den von
den Milben besetzten Lavaschlacken der Bäche findet. Vor allem muss
man aber in dem Temperaturunterschied der Seen und Bäche den Grund
für die Abwesenheit der Milben in den ersteren suchen. Denn in dem
See steigt die Wassertemperatur wohl um 9° höher als in dem Zufluss.
Der schroffe Wechsel der Temperatur wird denjenigen Milben, welche
von dem Bache mit in den See hinabgerissen werden, verderblich, denn
sie sind überhaupt sehr empfindlicher Natur. Es ist mir mehrere Male
bei meinen Ausflügen vorgekommen, dass ich versucht habe, sie lebend
heimzubringen, aber fast regelmässig fand ich sie trotz aller Vorsicht
tot vor“. So empfindlich wie die soeben angeführte Bewohnerin der
Azoren sind nun freilich die meisten unserer Hydrachniden nicht. Zumal
gegen erwärmtes Wasser zeigen, wie schon erwähnt, die meisten der
unsere Kleingewässer, namentlich die Teiche und Weiher, bewohnenden
Milben eine starke Widerstandsfähigkeit. Jedoch wird es noch immer
ausgedehnter Beobachtungen bedürfen, um die Einflüsse der Temperatur
auf das Leben unserer Wassertiere genauer kennen zu lernen.
Mit den Azoren ist wohl die von Europa fernste Station, auf welcher
Süsswassermilben beobachtet worden sind, genannt worden. Steht es
überhaupt mit der Kenntnis der Acariden in den aussereuropäischen[S. 31]
Ländern ziemlich schlecht, so sind die Gebiete, in denen man sich nach
den im süssen Wasser lebenden Milben umgesehen hat, im wesentlichen
in Europa zu suchen und auch da sind noch die meisten Strecken
unerforscht. Es folgt fast naturgemäss aus diesem Umstande, dass
das Wissen über diese Tiergruppe ein in jeder Beziehung durchaus
lückenhaftes sein muss. Allerdings scheinen ja, und das gilt auch
von sehr zahlreichen Landmilben, die einzelnen Gattungen und Arten
sehr grosse Verbreitungsgebiete zu besitzen, aber dennoch ist noch
überall das Fehlen gewisser an anderen Orten vorkommender Formen ausser
Zweifel, und es trifft auch hier die Wahrnehmung zu, dass der Süden an
Formen reicher ist, als der Norden.
Halten wir eine Überschau ab über die Gegenden, welche überhaupt
nach Süsswassermilben durchsucht sind, so sind zu nennen zahlreiche
schwedische Gewässer (durchforscht von C. J. Neuman), ein
Teil der norditalienischen (d. v. J. Canestrini und A.
Berlese), die Schweizer Seen (d. v. G. Forel und G.
v. Haller), die nordöstlichen Gebiete Frankreichs (d. v. J.
Barrois) und manche Gebiete Deutschlands (d. v. C. L. Koch,
Koenike, Kramer).
Wie es aber bei einer erst beginnenden Erforschung einer Tierklasse
fast natürlich ist, haben sich die meisten Beobachtungen zunächst
auf die äussere Erscheinung der Hydrachniden gerichtet, die
Lebensbeziehungen dagegen sind zum grösseren Teil noch übersehen
worden. Dennoch lässt sich Einiges auch bereits jetzt hierüber sagen.
Zunächst hat sich wohl als unzweifelhaft ergeben, wie auch schon
weiter oben betont worden ist, dass die Hydrachniden auf Grund ihrer
Entwickelungsgeschichte in zwei Gruppen gesondert werden können, welche
sich durch die Lebensweise ihrer sechsfüssigen ersten Larven ergeben.
Die eine Gruppe besitzt Larven, welche Gestalt und Lebensweise der
Trombidiumlarve zeigen, die anderen nicht. Man kann es nämlich wohl
als Regel aufstellen, vorbehaltlich freilich einer erst in Zukunft zu
gewinnenden ganz allgemeinen Bestätigung, dass die trombidiumartigen
Milben während ihrer ersten Jugend vom Blute anderer Tiere, namentlich
der[S. 32] Insekten, leben und sich daher an solche ansaugen. Erst nachdem
sie die erste Häutung überstanden haben, führen sie nicht mehr ein
parasitisches, sondern ein freies Leben. Solche Lebensweise führen nun
die Jungen von Hydrachna, Eylaïs und Limnochares
und, soweit ich sehe, auch von Hydrodroma, einer schön
scharlachroten, ziemlich ansehnlichen Wassermilbe. Die Beobachtungen
sind bei den soeben namhaft gemachten Acariden noch keineswegs in
gleichem Masse vollständig, ganz abgeschlossen dürften sie vielmehr
nur bei der Gattung Hydrachna sein, jedoch unterliegt es keinem
Zweifel mehr, dass bei allen die Übereinstimmung in der Hauptsache der
Entwickelung, der parasitischen Lebensweise, eine sehr weitgehende ist.
Die Jungen von Hydrachna bleiben von dem Augenblick des
Ausschlüpfens aus dem Ei im Wasser, dagegen steigen die der drei
anderen Gattungen möglichst bald an die Oberfläche und machen
ausgiebigen Gebrauch von ihren Rennbeinen, indem sie mit einer fast
staunenswerten Geschwindigkeit auf dem Wasser und den Pflanzen des
Ufers auf und ab eilen, um Insekten zu suchen, welche sie besteigen
können. Die Larven von Limnochares nehmen z. B. die ebenfalls
auf der Wasseroberfläche lebenden Schreitwanzen zu erwünschten
Nährtieren und bohren sich in die weichen Chitinskelettpartien ein.
Bei den Larven von Hydrodroma bot sich noch ein anderes
Schauspiel, als ich Gelegenheit hatte, einen auskriechenden Schwarm von
jungen Tieren zu verfolgen. Nachdem sie an die Oberfläche des Wassers
emporgekommen waren und ich die Tierchen sammeln wollte, um sie an
Blattläusen sich festsaugen zu lassen, musste ich zu meinem Erstaunen
bemerken, dass sie die Fähigkeit besassen in mächtigen Sprüngen
fortzuhüpfen. In kurzer Zeit war daher der ganze Schwarm zerstreut und
den Blicken völlig entschwunden.
Die sechsfüssigen Larven der übrigen Hydrachniden leben, im Gegensatze
zu den eben namhaft gemachten, frei schwimmend im Wasser und nähren
sich vom Raube. Sie besitzen daher eine dieser Lebensweise angepasste
Gestalt, welche zwar ebenfalls von der künftigen des erwachsenen Tieres
abweicht, aber namentlich in[S. 33] der Bildung der Füsse ganz verschieden
ist von den Larven obiger Arten. Eine bemerkenswerte Ausnahme von
dieser Lebensweise beobachten wir nur bei einigen Arten der Gattung
Atax[IX]. Von den bisher bekannten sieben oder acht Arten dieser
Gattung führen etwa drei oder vier ein vollständig freies Leben,
während die anderen Arten sich einer ausschliesslich parasitischen
Lebensweise ergeben haben. Diese Arten hausen von Generation zu
Generation zwischen den Weichteilen der Teich- und der Malermuschel und
verlassen ihr Wohntier wahrscheinlich nur während ihrer Larvenzeit,
um ein neues aufzusuchen. Dabei sei gleich erwähnt, dass durch das
parasitische Leben sich eine bestimmte Veränderung ihres Organismus
eingestellt hat, wenigstens möchte ich dieselbe damit in ursächlichen
Zusammenhang bringen. Während nämlich die frei lebenden Hydrachniden
sämtlich, soweit die Beobachtungen reichen, ein wohl ausgebildetes
Atmungssystem haben, fehlt den parasitischen Atax-Arten jede Spur von
Tracheen und der mit den Luftlöchern in Verbindung stehende Hauptstamm
ist auf ein ausserordentlich geringfügiges Stückchen zurückgebildet,
so dass man fast von einem vollständigen Fehlen desselben reden
kann. Ob es nun der Aufenthalt in einer so schleimigen und luftarmen
Flüssigkeit, wie sie die Muscheln erfüllt, mit sich gebracht hat, dass
das Atmungsorgan ausser Thätigkeit gesetzt wurde und daher verkümmerte,
darüber eine bestimmte Ansicht auszusprechen scheint noch nicht der
rechte Zeitpunkt gekommen zu sein.
Wenn uns die parasitischen Atax-Arten schon darauf führten, von der
Auswanderung von einem Wohntier auf ein anderes zu reden, so dürfte es
an der Zeit sein, überhaupt die Verbreitung der Süsswassermilben aus
ihren doch meist eng begrenzten heimatlichen Wasserbecken in andere
etwas näher zu betrachten. Ich komme dabei auf die Beobachtungen
des Dr. Th. Barrois, die er auf den Azoren gemacht hat, noch
einmal zurück. Er traf dort zwei Gattungen an und unter diesen eine,
welche ja allerdings allgemein verbreitet ist, deren Arten aber
nicht gerade zu den[S. 34] allergewöhnlichsten gehören. Er fand stets
sehr zahlreiche Exemplare der diesen beiden Gattungen angehörigen
Milben in den dortigen Bächen an. Dies hatte wohl seinen Grund darin,
dass die Gewässer der vulkanischen Azoren in der ersten Periode
des Bestehens dieser aus dem Ozean aufgetauchten Inseln wohl noch
völlig unbevölkert gewesen sind. Erst nach und nach werden dorthin
verschlagene Süsswasserbewohner von dem neuen Gebiet Besitz genommen
und frei von aller Konkurrenz sich rasch vermehrt haben. Wie lässt es
sich wohl erklären, dass sich auch die immerhin zarten Wassermilben
dabei beteiligten? Unter allen Umständen wird die Verbreitung
derselben dorthin auf mechanische Ursachen zurückzuführen sein. Durch
irgend einen Besucher oder Bewohner eines schon von ihnen besetzten
Süsswasserbeckens muss ihre Überführung in ein von ihnen noch nicht
bewohntes vermittelt worden sein, sobald der natürliche Lauf der
Gewässer auch bei Überschwemmungen die neue Heimat mit der alten
nicht in Verbindung zu setzen vermochte. Als Dr. Barrois, um
über die Verpflanzung seiner Sperchon-Arten nach den Azoren ins Klare
zu kommen, die dortigen Gewässer aufmerksam durchforschte, fand er
ausser den Milben vor allen Dingen zahlreiche Wasserwanzen von der
Gattung Corixa vor. Die Wasserwanzen können, wie sämtliche
entwickelte Wasserinsekten, lange Stunden ausserhalb des Wassers
leben, denn sie sind ja eigentlich Lufttiere und gehen nur um ihrer
Nahrung willen ins Wasser. Sie machen auch grosse Flüge, namentlich
bei Nacht, um aus einem Becken in ein anderes zu gelangen, wobei sie
wahrscheinlich einem scharfen und sicheren Geruch folgen. Solche
fliegende Wasserinsekten werden unter Umständen ganz ebenso, wie
Schmetterlinge und Heuschrecken, vom Winde erfasst und aufs Meer
hinausgetrieben, wobei sie wohl meist zu Grunde gehen, gelegentlich
aber auch einsam daliegende Inseln erreichen, deren Gewässer sie dann,
wenn es gerade mit befruchteten Eiern beladene Weibchen waren, nun
mit ihrer Nachkommenschaft bevölkern können, vorausgesetzt, dass die
Lebensbedingungen sonst ihrem Körper entsprechen. Auf diese Art und
Weise werden die Wasserwanzen wahrscheinlich nach den Azoren gekommen
sein.
[S. 35]
Nun haben schon häufig Beobachter bemerkt, dass die zur Larvenruhe
gelangten noch unentwickelten Milben sich an solche Wasserwanzen, wie
eben beschrieben, anheften, um daselbst ihre Verwandlung unter dem
Schutze der alten Larvenhaut durchzumachen. So kann es leicht gekommen
sein, dass eine Wasserwanze, welche solche Larven von Wassermilben
an sich trug, vom Winde nach den Azoren verschlagen wurde. Wenn
dann die angeheftete Wassermilbenlarve während der gewiss längeren
Reise nicht ganz austrocknete, so wird sie, nachdem ihr Träger eine
neue Heimat in irgend einem Gewässer der Azoren gefunden hatte,
dort nach einiger Zeit ausschlüpfen und, wenn Männchen und Weibchen
gleichzeitig hinübergetragen worden sind, ihre Art in den Flüsschen
der Azoren weiter fortpflanzen müssen. Nun hat Dr. Barrois
bemerkt, dass die in Larvenruhe verfallenen Larven des ersten Stadiums
gegen Austrocknung mehr Widerstand entgegensetzen, als es der sonst
sehr zarte Körper der Hydrachniden vermag, und dass daher obige Art
und Weise die wahrscheinlichste sei, wenn man sich die Verbreitung
der Süsswassermilben über das Meer hinüber erklären will. Gewiss
ist dies wohl eine Möglichkeit, aber doch keineswegs eine so einzig
gültige, dass man annehmen müsste, die ersten Larven der Wassermilben
hätten überhaupt die Gewohnheit angenommen, sich namentlich an
Süsswasserinsekten festzusetzen, um dort ihre Larvenruhe abzumachen,
und diese Gewohnheit bringe es nun mit sich, dass die während der
Larvenruhe gegen das Austrocknen besser gewappneten Tiere von den
ausfliegenden Wasserinsekten auch dorthin transportiert würden,
wohin sie auf gewöhnlichem und ihrer Organisation entsprechendem
Wege nicht gelangen könnten. Es sind die Möglichkeiten, eine weit
entfernte Station zu erreichen, so mannigfaltige, dass es unserem
Ermessen nach noch nicht thunlich ist, zu betonen, dass es die auf
den Azoren gefundene Corixa ist, welche zu der Zeit, wo sie
selbst durch Zufall die Azoren erreichte, auch die Sperchon-Art mit
nach dem neuen Aufenthaltsorte hinüberbrachte. Es ist ja gewiss, dass
im Verlauf der Jahrtausende, welche seit dem Auftauchen der Azoren aus
dem Meere verstrichen sind, oftmals eine Corixa[S. 36] atomaria von
Portugal hinübergeführt sein kann und dass immer einmal wieder ein
Sperchon glandulosus an ihr angeheftet gewesen sei. Wenn man
diesen Umstand aber immer wieder durchdenkt, so wird man immermehr zu
der Überzeugung sich hinneigen, dass diese Möglichkeit ja allerdings
wohl vorliegt, dass aber ebensosehr auch Wasservögel, welche, wie
Barrois in einer Anmerkung selbst hervorhebt, an ihren Füssen
und Schnäbeln so häufig Reste von Pflanzen aus süssem Wasser mit sich
führen, die Vermittler sein können, ja, dass es nicht bloss Larven zu
sein brauchen, welche in die neue Heimat hinübergeführt worden sind,
Larven, die der Gefahr der Unfruchtbarkeit ausgesetzt sind, sondern in
den feucht bleibenden vegetabilischen Resten im Schnabel und an den
Zehen können die erwachsenen Tiere ebensogut der neuen Heimat zugeführt
werden. Zudem sind auch nicht alle Beobachter darin übereinstimmend,
dass die Wassermilben, wenn sie ihrem feuchten Elemente entnommen sind,
dem Eintrocknen so leicht erliegen. Claparède, der bedeutende
Genfer Naturforscher, hat vielmehr an der die Teichmuschel parasitisch
bewohnenden Atax-Art gefunden, dass sowohl Muttertiere als Eier
sehr lebenskräftig sind. In den Muscheln, die wochenlang ausserhalb
des Wassers in der Dürre gelegen und halb ausgetrocknet dem Tode
entgegengehen, fand er die Milben zwar durch Verdunstung erstarrt,
jedoch beim ersten Wasserzusatz sehr schnell wieder lebendig und die
Eier entwickelungsfähig werden.
Es ist also nicht ausgeschlossen, dass die Besiedelung der mitten
im Meere gelegenen Inseln mit Süsswassermilben unmittelbar durch
Überführung fortpflanzungsfähiger Individuen geschehen sei und
noch geschieht. Es wird sogar die Verbreitung der das süsse Wasser
bewohnenden Milben im allgemeinen überhaupt so gedacht werden müssen,
dass erwachsene Weibchen, nicht unbefruchtete und unfruchtbare Larven,
auf mechanischem Wege von Wasserbecken zu Wasserbecken getragen werden
und so ihre Art in Gebieten, wo dieselbe noch nicht vertreten war,
heimisch machen.
[S. 37]
Es bleibt uns nun endlich noch ein wichtiger Gegenstand zur Besprechung
übrig, nämlich die Entwickelung des Einzeltieres vom Ei bis zum
erwachsenen Zustand.
Für jedes Lebewesen ist es eine der wichtigsten Aufgaben, seine
Art durch Nachkommen zu erhalten. Daher ist für den aufmerksamen
Naturfreund die Fortpflanzung und Entwickelung des Einzeltieres ein
Gegenstand des grössten Interesses.
Bei den Milben ist nun gerade die Entwickelungsgeschichte noch sehr des
weiteren Studiums benötigt, jedoch ist namentlich für die Hydrachniden
das Beobachtungsmaterial nicht ganz unerheblich, so dass es gelingen
wird, ein Bild des Entwickelungsganges einer Wassermilbe in grossen
Zügen zu entwerfen.
Ich wähle zwei Beispiele, nämlich die Entwickelung der in unseren
heimischen Gewässern häufigsten kleinen rotbraunen Wassermilbe,
Nesaea fuscata Koch, welche für die Mehrzahl aller anderen als
Muster dienen kann; daran mag sich die schon seit langer Zeit bekannte
Entwickelung der kugeligen Wassermilbe, Hydrachna globosa Dugès,
anschliessen.
Die Eierchen der rotbraunen Wassermilbe werden in Häufchen bis zu
dreissig und mehr an Wasserpflanzen oder an Steinen des Wassergrundes
gelegt. Es trifft sich wohl, dass man ein Weibchen bei seinem Geschäft
der Eiablage genauer beobachten kann. Es legt die Eier in lockeren
Haufen, dabei meist rückwärts schreitend und sie in schneller Folge
aus dem Körper hervorstossend. Die roten Eierchen sind dabei von einer
sehr dünnen weisslichen Schicht umgeben. Dieses ist die klebrige
Kittsubstanz, welche jedem Ei mitgegeben wird. Schnell quillt dieselbe
im Wasser auf, und während die letzten Eier gelegt werden, hat sich
bei den ersten bereits ein breiter weisser Hof um dieselben gebildet.
Nach Verlauf einiger Stunden sind die Zwischenräume zwischen den Eiern
völlig von der Kittsubstanz ausgefüllt, die Eierchen ruhen jetzt, zu
einem einzigen Häufchen verschmolzen, unter einer im Wasser vollends
erhärtenden Hüllschicht.
Nach gemessener Zeit hat sich aus dem ursprünglichen Eiinhalt der
sogenannte Embryo gebildet, zu gleicher Zeit ist aber[S. 38] auch noch
eine zweite, innere Eihaut entstanden, welche sich nach und nach
bedeutend ausdehnt und sich, weil sie innerhalb der harten äusseren
Eischale keinen genügenden Raum findet, bald in viele Falten legt.
Jetzt platzt die harte äussere Eihaut, die zweite dehnt sich durch
Aufsaugen von Wasser sehr rasch aus und glättet sich vollkommen. So
geschieht es, dass der Beobachter ein neues, sehr viel grösseres Ei
aus dem ursprünglich gelegten hervorgehen sieht. Die Gestalt desselben
ist in Fig. 3f dargestellt. Ein solcher Vorgang, dass aus
einem Ei nicht ein entwickeltes Tier mit freibeweglichen Gliedmassen
hervortritt, sondern ein zweites Ei, ist bisher bei den Milben nicht
selten beobachtet worden, jedoch scheint er nur bei den Acariden
verbreitet zu sein und zwar bei den trombidiumartigen Milben ganz
besonders. So habe ich ihn bei den oben bereits einmal erwähnten
Cheyletus vorgefunden. Hier entwickelt das Ei eine innere
Eihaut, welche an einer bestimmten Stelle einen scharfen Stechapparat
besitzt, der zur bestimmten Zeit die alte Eihaut durchsticht,
um dem sogenannten zweiten Ei den Austritt zu gestatten. Bei
Nesaea fallen die alten Eischalen einfach ab, indem sie durch
die aufschwellende noch in der zweiten Haut befindliche junge Milbe
gesprengt werden.
Ist nun die Larve innerhalb dieser zweiten Eihaut entsprechend
entwickelt, so sprengt sie dieselbe, was etwa nach vierzehn Tagen
geschieht. Für die ausgeschlüpften Larven ist es aber keine leichte
Arbeit, ganz frei zu werden, denn noch trennt die erhärtete
Oberflächenschicht der Kittsubstanz die Tierchen von ihrem Element.
Durch die unruhige Bewegung zahlreicher Füsse wird die innere Masse
der Hüllsubstanz bald zerbröckelt, so dass die immer zahlreicher
ausschlüpfenden Larven durch einander kriechen und drängen; aber noch
ist ein Ausweg nicht gefunden.
Fig. 3. a Erste Larve von Nesaea fuscata, von der Bauchfläche aus
betrachtet — b Mundschnabel derselben, von der Seite, mit Taster (α) und
Kiefer (β) — c Fusskralle derselben Larve — d Fusskralle der
erwachsenen Nesaea — e Zweite Larve von Nesaea fuscata —
f Aus der gesprengten ersten Eischale tritt das zweite eiförmige Stadium
hervor. Der Embryo mit deutlichem Rücken- und Bauchschild ist von dem Apoderma
umhüllt — g Endglied des dritten Fusses vom erwachsenen Männchen der
Nesaea fuscata — h Vierter Fuss desselben Männchens — i
Weibliche Geschlechtsöffnung von Nesaea fuscata mit den Haftnäpfen —
k Männliche Geschlechtsöffnung von Nesaea fuscata mit den
Haftnäpfen.
Endlich hat eine auch glücklich die äussere Hüllschicht an einer
besonders nachgiebigen Stelle durchbrochen und enteilt in die freie
Flut. Dies glückt zuletzt allen, aber es geht manche Stunde darüber
hin, bis die letzte Milbe dem Gefängnis entronnen ist. Und vielleicht
würde es noch länger dauern, ja mancher gar nicht gelingen, wenn nicht
die Taster unserer jungen Milben auf dieser[S. 40] ersten Entwickelungsstufe
eine von der späteren Gestalt sehr abweichende und für die Sprengung
eines entgegenstehenden Hindernisses sehr geeignete Form und Bewaffnung
hätten (Fig. 3b). Sie sind sehr dick und nehmen eine starke
Muskulatur auf, so dass sie sehr kräftige Bewegungen zulassen. Am
vorderen Ende des letzten der sehr gedrungenen Glieder bemerkt man
eine grosse, stark gekrümmte Hakenkralle, die wie eine Hippe in die
Hüllmasse der Eier einsetzen und dieselbe aufreissen kann. Auf späteren
Entwickelungsstufen werden die Taster schlank und jene Krallen sind
ganz verschwunden. Das Tierchen bedarf ihrer später nicht mehr. Nicht
unerwähnt darf ich hier allerdings lassen, dass es auch Nesaea-Arten
giebt, bei denen die sechsfüssigen Larven die Freiheit nicht mehr
gewinnen, sondern noch unter der Kitthülle ihre nächste Häutung
durchmachen.
Wie es bei den Milben als Regel anzusehen ist — Ausnahmen sind
ungemein selten —, hat unsere junge Larve (Fig. 3a) nur
sechs Füsse, jederseits drei, mit denen sie das Wasser lebhaft tritt,
um schnell vorwärts zu eilen. Die grossen, als schwarze Pünktchen
hervortretenden Augen bestimmen Richtung und Ziel der kräftigen
Schwimmbewegung.
Wendet man eine hinreichende Vergrösserung an, so bemerkt man bald,
dass diese erste, jüngste Larve auch noch in anderer Hinsicht, nicht
nur mit Rücksicht auf die Taster, gegen die späteren Larvenstufen
recht hervorstechende Verschiedenheiten zeigt, welche schon nach der
nächsten Häutung verschwinden. So ist der ganze Rücken durch eine
vorn breitere, hinten etwas zugespitzte härtere Deckplatte geschützt
(Fig. 3f). Eine solche findet sich bei den erwachsenen Tieren
zahlreicher Wassermilbenarten auch nicht mehr andeutungsweise; bei
anderen ist ein geringfügiger Rest übrig geblieben (Hydrodroma)
und bei einer nur geringen Anzahl Arten ist die Verhärtung der
Rückenhaut eine für alle Lebensstufen dauernde.
Die Unterseite unserer jungen Larven (Fig. 3a) ist von der der
späteren Larvenstufen und der erwachsenen Tiere zwar auch verschieden,
doch finden sich die auch später beobachteten[S. 41] Verhältnisse im
allgemeinen hier wieder vor. Auffallend ist dagegen, dass die den
Wassermilben so eigentümlichen sogenannten Haftnäpfe hier noch völlig
fehlen. So stellt die erste jüngste Larvenform eine sehr eigenartige
Entwickelungsphase dar und es kommt in der That bei dem Übergang zur
zweiten Larvenform zu einer in jeder Beziehung sehr durchgreifenden
Gestaltveränderung.
Etwa fünf oder sechs Tage, nachdem das Tierchen sich aus der Eihülle
befreit hat, stellen sich die Vorboten dieser Verwandlung ein. Die
Milbe fühlt wohl selbst, dass mit ihr etwas Bedeutsames vorgehen soll
und hängt sich, um die inneren Entwickelungsvorgänge ungestört ablaufen
zu lassen, mit ihren Tastern an irgend einem geschützten Orte fest an.
Die Glieder werden bewegungslos und die innere Körpersubstanz zieht
sich aus ihnen zurück, eine rundliche von der alten Haut umschlossene
eiartige Masse bildend. Ähnliches wiederholt sich später ebenso bei
der zweiten noch eintretenden Häutung und wird auch bei der grossen
Mehrzahl der Land und Wasser bewohnenden Milben beobachtet, wenn es
auch nicht durchaus bei sämtlichen zu einer völligen Zurückziehung des
Körperinhalts aus den Gliedern kommt.
Es gestaltet sich während der jetzt eben in Rede stehenden Larvenruhe
unter der alten Haut eine neue Milbe aus. Man beobachtet deutlich die
neuen Glieder durch die durchsichtige, nun bloss noch einen Schutz für
das zarte neue Geschöpf abgebende bisherige Haut hindurch. Nach wenigen
Tagen ist es soweit, dass das eingeschlossene Tier die Hülle sprengen
kann. Statt einer sechsfüssigen Acaride tritt eine achtfüssige aus
der Larvenhaut hervor (Fig. 3e). Und während vorher die sechs
Füsse ausserordentlich dünn und verhältnismässig kurz waren, fällt
jetzt die Länge der acht neuen Füsse auf. Dieselbe übersteigt um ein
Bedeutendes die Gesamtlänge des Rumpfes und giebt dem ganzen Tier etwas
Schlankes und Zierliches. Das Rückenschild ist bis auf die letzte Spur
verschwunden und auf der Unterseite sieht man ausser den völlig anders
geformten grossen Hüftplatten noch jederseits von der kaum angedeuteten
Geschlechtsöffnung ein schräg gestelltes Plättchen, welches zwei
deutlich ausgebildete kreisrunde Haftnäpfe trägt. Noch[S. 42] ein Blick auf
die Füsse belehrt uns, dass nicht nur reichliche Schwimmborsten die
mittleren Glieder derselben schmücken, zwar noch nicht so zahlreiche
wie beim erwachsenen Tier, aber doch schon genau ebenda aufgestellt
wie bei diesem, sondern dass auch die Krallen eine ganz veränderte
Form bekommen haben. Fig. 3c zeigt die Kralle der ersten
Larve, während Fig. 3d diejenige aller späteren Lebensstufen
darstellt, besser als eine Beschreibung es zu geben vermag. Die grösste
Veränderung hat aber der Mundabschnitt erfahren. Der lange Schnabel ist
verschwunden und die Taster sind langgestreckt und schlank, auch fehlt
ihnen die scharfe Kralle, welche den ersten Larven so eigentümlich
ist. Einem genaueren Beobachter entgeht auch nicht, dass die grosse
Pore, welche bei letzterer zwischen den Hüftplatten des ersten und
zweiten Fusses stand, jetzt ganz verschwunden ist. Dies hängt wohl
mit der auffallendsten der inneren Veränderungen zusammen. Jetzt hat
nämlich die junge Milbe zahlreiche Luftröhren bekommen, welche alle
von zwei am Grunde der Kiefer ausmündenden Hauptstämmen ausgehen. Mit
dieser Larvenform, welche in Fig. 3e abgebildet ist, ist für
die Hydrachniden die letzte Stufe vor der ganz ausgebildeten Form
erreicht. Es ist dies keineswegs Gesetz in dem ganzen Reiche der
Acariden, dass zwischen Ei und erwachsenem Tier nur zwei Larvenstadien
eingeschaltet sind. Vielmehr zeigen zahlreiche Gattungen, namentlich
aus den Familien der Hartmilben und Tyroglyphus-artigen Milben drei
und mehr Larvenstadien zwischen Ei und erwachsenem Tier, so dass
es ein interessantes Gebiet der Forschung ist, entweder die Gründe
einer so eingreifenden Verschiedenheit der Entwickelungsgeschichte
innerhalb der Familie der Acariden darzulegen oder durch richtigere
Auffassung der bestehenden Verhältnisse eine Übereinstimmung der
scheinbar verschiedenen Entwickelungsformen zu erkennen. Bemerkenswert
ist es jedenfalls, dass die Trombidium-Arten eine mit den Hydrachniden
im wesentlichen übereinstimmende Entwickelungsweise zeigen, so dass
auch hierdurch deren nahe Verwandtschaft bestätigt wird. Doch kehren
wir noch einmal zu unserer Larve zurück. Diese war zunächst dadurch
ausgezeichnet, dass die Füsse[S. 43] im Vergleich zum Rumpf ausserordentlich
lang, wenn auch zierlich sind. Dieses Verhältnis ändert sich jedoch
bald, indem die Larve durch sehr reichliche Nahrungsaufnahme schnell
und erheblich wächst, so dass der Rumpf mächtig anschwillt, während die
Gliedmassen unverändert dieselbe Grösse beibehalten und so allmählich
immer mehr gegen den Rumpf zurücktreten.
Bald tritt nun eine neue Pause in der Entwickelung ein. Die Milbe
sucht ein Versteck und versinkt in einen neuen Zustand völliger
Regungslosigkeit, während welcher sich innerhalb der Larvenhaut die
vollständig ausgebildete Milbe entwickelt. Entschlüpft dann der
Larvenhaut das endgültig fertiggestellte Tier, so fällt auch wieder
vor allem der Unterschied in der Längenentwickelung der Füsse auf.
Aber auch in vielen anderen Punkten ist an dem neuen Geschöpfe eine
wesentlich andere Form und Bildung zu bemerken. So sind die Weibchen
nunmehr von den Männchen deutlich zu unterscheiden, während bis
zur letzten Larve ein äusserlicher Unterschied der männlichen und
weiblichen Larven nicht festgestellt werden konnte. Es sind aber
auch abgesehen von diesem Unterschiede in beiden Geschlechtern die
Schwimmborsten an den Gliedern namentlich der hinteren beiden Fusspaare
ausserordentlich viel zahlreicher geworden. Endlich findet man statt
der wenigen Haftnäpfe auf der Unterseite des Hinterleibes nunmehr deren
zahlreiche jederseits in einem Häufchen vereinigt (Fig. 3i u.
k).
Ist die erwachsene Milbe nach der letzten Häutung noch verhältnismässig
klein, so bewirkt eine reichliche Nahrungszufuhr bald ein ansehnliches
Wachstum, so dass es namentlich Weibchen bis zu 1½ mm Länge
giebt, während die Männchen meist erheblich kleiner bleiben. Überhaupt
ist bei den Milben das Männchen meistens ziemlich viel kleiner als
das Weibchen, selbst dann, wenn letzteres nicht durch die zahlreichen
allmählich heranwachsenden Eier aufquillt.
Wie lange eine erwachsene Milbe zu leben vermag, ist nur selten
Gegenstand des Experiments gewesen. Ich selbst habe Larven von
Nesaea fuscata einen ganzen Winter hindurch gehalten, ohne dass
sie sich verwandelt hätten. Andere Beobachter haben ähnliche[S. 44] Resultate
bemerkt. Es ist wohl möglich, dass eine Milbe der erwähnten Art mehrere
Jahre hindurch ihr Leben erhalten kann.
Den Beschluss dieser Darstellungen aus der Entwickelungsgeschichte
der Süsswassermilben mag die Lebensgeschichte der Hydrachna
globosa machen. Dieselbe ist schon von Dugès in den dreissiger
Jahren unseres Jahrhunderts entdeckt und seitdem oftmals bestätigt
worden. Diese Wassermilbe ist, wie bereits oben gesagt, insofern
bemerkenswert, als aus ihren Eiern junge Larven schlüpfen, welche von
den entsprechenden der Hart- und Weichschwimmer, also auch denen von
Nesaea und Atax völlig verschieden sind, dagegen mit
den Larven der auf dem Lande lebenden Trombidien grosse Ähnlichkeit
besitzen. Eine derartige Larve, welche aber nicht zu Hydrachna,
sondern zu einer Gattung gehört, welche den Namen Hydrodroma
erhalten hat, ist in Fig. 4 abgebildet. Sie zeichnet sich durch
eine mächtige Entwickelung der Kiefer aus und führt Taster, welche
unmittelbar an die Taster aller trombidiumartigen Landmilben erinnern,
dadurch, dass das letzte Tasterglied an dem untern Ende des vorletzten
Gliedes seitlich und nicht am obern Ende eingelenkt ist. Die Füsse
sind mit Krallen bewehrt, welche sich in dieser Form ebenfalls
bei zahlreichen Landmilben, selbst bei Panzermilben (Oribatiden)
wiederfinden. Diese kleinen scharlachroten Larven scheinen unter
allen Umständen ein parasitisches Leben führen zu müssen. So heften
sich die Hydrachna-Larven an Insekten, welche im Wasser leben, fest.
Sehr bevorzugt wird die graue Wasserwanze, an welcher man im Frühjahr
häufig die schon über stecknadelkopfgrossen Parasiten mit ihrem Kopfe
in die weicheren Hautpartien an den Gelenken eingebohrt findet. Da die
völlig ausgewachsene Wassermilbe schon im Juni beobachtet wird, so
wird bereits Anfang Juli das Ausschlüpfen aus den Eiern stattfinden.
Die jungen Milben machen ihre erste Larvenzeit teils freilebend, so
lange nämlich, bis sie ein geeignetes Nährtier gefunden haben, teils
als Parasiten angeheftet an letzterem durch. Diese letztere Periode
erstreckt sich durch den Herbst bis zum Frühjahr. Während dieser Zeit
nimmt die Grösse des Tieres durch unausgesetzte Nahrungszufuhr dauernd
zu, so dass Larven bis zu 2 mm[S. 45] Länge an den Nährinsekten
angetroffen werden. Der Hinterleib dehnt sich dabei allein aus,
während die Gliedmassen keine Grössenzunahme erfahren. Gewissermassen
gestaltet sich das Tier zu einer Vorratskammer von Nährstoffen um,
welche nun im Frühjahr zur Neubildung der zweiten Larve verwendet
werden. In dem aufgetriebenen Hinterleibe bemerkt man, wie in einem
zugeschnürten Beutel ruhend, nunmehr die völlig anders gestaltete neue
Milbe. Dieselbe hat acht Füsse statt der früheren sechs und zeigt ein
verändertes Kopfsegment, ist überhaupt in keiner Weise vergleichbar der
ersten Larve.
Fig. 4.
Erste Larve von Hydrodroma rubra.
Diese zweite Larve hat bereits im wesentlichen die Gestalt des
erwachsenen Tieres, zunächst aber noch nicht dessen Grösse. Durch[S. 46]
schnelles Wachstum erreicht sie dieselbe aber bald und begiebt sich an
einen Ort, wo sie die zweite Larvenruhe durchmachen kann. Zu diesem
Zweck entleert sie einen Tropfen ihres Speichelvorrats und heftet
dadurch ihren langen Schnabel an die Unterlage fest an. Die Glieder
verlieren ihre Beweglichkeit und der Leibesinhalt gestaltet sich noch
einmal zu einem neuen Tiere um. Nach etwa zehn Tagen durchbricht
letzteres die schützende Haut der ruhenden Larve. Nunmehr ist das Tier
ausgebildet und erfährt nur noch ein Grössenwachstum.
Anhang.
Tabelle zur Bestimmung der bis jetzt bekannt
gewordenen Gattungen der Hydrachniden.
1.
Die Augen nahe bei einander, der Mittellinie des
Körpers sehr genähert
2.
Die Augen in zwei weit von einander getrennte
Gruppen gesondert, dem Seitenrande des Körpers genähert
3.
2.
Sämtliche Füsse ohne Schwimmborsten
Limnochares, Latr.
Nur das vierte Fusspaar ohne Schwimmborsten
Eylaïs, Latr.
3.
Die Mandibeln eingliedrig, stechborstenartig, in
einem schnabelartigen Mundrohr laufend
Hydrachna, Müller
Mandibeln zweigliedrig
4.
4.
An den Füssen keine Schwimmborsten
5.
An den Füssen Schwimmborsten
6.
5.
Auf dem Rücken zahlreiche polygonale, einander sehr
genäherte erhärtete Felder
Thyas, C. L. Koch.
Eine einzige Panzerplatte bedeckt den Rücken
Aturus, Kramer.
Der Rücken durchaus weichhäutig
Bradybates, Neuman.
[S. 47]
6.
Die Körperhaut in beiden Geschlechtern überall
erhärtet
7.
Auf dem Rücken des Männchens eine denselben fast
vollständig bedeckende poröse Platte, welche dem Weibchen fehlt
Forelia, Haller.
Auf dem Vorderrücken bei beiden Geschlechtern eine
nur wenig umfangreiche vierzipfelige Stirnplatte
Hydrodroma, C. L. Koch.
Der Rücken bei beiden Geschlechtern durchaus
weichhäutig
9.
7.
Am vierten Fusse jeder Seite keine Krallen
Marica, C. L. Koch.
Am vierten Fusse jeder Seite Krallen
8.
8.
Jederseits nur drei Haftnäpfe
Axona, Kramer.
Jederseits sehr zahlreiche kleine, porenartige
Haftnäpfe
Arrenurus, Dugès.
9.
Die Taster endigen scherenförmig
Diplodontus, Dugès.
Die Taster endigen nicht scherenförmig
10.
10.
Die Epimeren sämtlicher Füsse jeder Seite bilden
ein zusammenhängendes Feld
11.
Die Epimeren der Füsse jeder Seite sind in zwei von
einander getrennte Gruppen gesondert
14.
11.
Der Körper bedeutend mehr lang als breit, oval
abgerundet
Pseudomarica, Neuman.
Der Körper kreisrund
12.
Der Körper hinten fast gradlinig abgestutzt
Acercus, C. L. Koch.
12.
Die Haftnäpfe fehlen ganz
Pontarachna, Philippi.
Jederseits drei Haftnäpfe
13.
Jederseits zahlreiche Haftnäpfe
Midea, Bruzelius.
13.
Die Haftnäpfe auf der Innenfläche der
Geschlechtsdeckklappe
Lebertia, Neuman.
Die Haftnäpfe auf der äussern Oberfläche der
Geschlechtsdeckklappe
Mideopsis, Neuman.
14.
Am vierten Fusse ohne Krallen
15.
Am vierten Fusse Krallen
16.
15.
Jederseits drei Haftnäpfe auf der Innenfläche der
Geschlechtsklappe
Teutonia, Koenike.
Jederseits drei Haftnäpfe auf der Aussenfläche der
Geschlechtsklappe
Limnesia, C. L. Koch.
[S. 48]
16.
Jederseits drei Haftnäpfe auf der Innenfläche der
Geschlechtsklappe
Sperchon, Kramer.
Die vorhandenen Haftnäpfe neben dem Geschlechtshof
in die Haut eingelassen, oder auf Platten angebracht
17.
17.
Jederseits drei Haftnäpfe
18.
Jederseits sechs oder viele Haftnäpfe
21.
18.
Am Vorderrande des vorletzten Tastergliedes ein
stumpfer zapfenartiger Fortsatz neben dem fünften Gliede
Piona, C. L. Koch.
Am Vorderrande des vorletzten Tastergliedes kein
zapfenartiger Fortsatz
19.
19.
Die Epimere des vierten Fusses jeder Seite am
hintern Rande in der Mitte in eine längere Spitze nach hinten ausgezogen
Hydrochoreutes, C. L. Koch.
Die Epimeren des vierten Fusses jederseits hinten
geradlinig abgeschnitten
20.
20.
Die Haftnäpfe klein, den Geschlechtshof völlig
einrahmend
Megapus, Neuman.
Die Haftnäpfe verhältnismässig gross, auf
besonderen, neben dem Geschlechtshof stehenden Plättchen eingelassen
Hygrobates, C. L. Koch.
21.
Die Krallen mit blattartig erweiterter Basis;
Männchen mit besonders umgestaltetem dritten und vierten Fuss
Nesaea, C. L. Koch.
Die Krallen nicht mit blattartig erweiterter Basis,
Männchen mit nicht besonders umgestalteten Füssen
Atax, Bruzelius.
[S. 49]
Litteratur.
1) Otto Friedrich Müller, Hydrachnae, quas in aquis Daniae
palustribus detexit, descripsit, pingi et tabulis aeneis XI incidi cur.
Lipsiae 1781.
2) Pierre André Latreille, Précis des caractères génériques des
Insectes. Paris 1796.
3) Antoine Dugès, Recherches sur l’ordre des Acariens en général
et la famille des Trombidiés en particulier. Premier mémoire. Annales
des Scienc. nat. Paris 1834.
4) Carl Ludwig Koch, Deutschlands Crustaceen, Myriapoda und
Arachniden, Heft 1–40. Regensburg 1835–41. Desselben Verfassers
Uebersicht des Arachnidensystems. Nürnberg 1842.
5) Ragnar Magnus Bruzelius, Beskrifning öfver Hydrachnider, som
förekomma inom Skåne. Akad. Abhandl. Lund 1854.
6) Edouard Claparède, Studien an Acariden. Zeitschrift für
wissensch. Zoologie von Siebold und Kölliker, XVIII. Bd. Leipzig 1868.
7) A. Croneberg, Ueber den Bau der Hydrachniden. Zool. Anzeiger
von V. Carus. Leipzig 1878, Jahrg. I, Nr. 14.
8) C. J. Neuman, Om Sveriges Hydrachnider, med 14 Taflor. Kongl.
Svenska Vetenskaps-Akad. Handlingar, Bd. 17, 1879.
9) G. Haller, Die Hydrachniden der Schweiz, mit 4 Tafeln. Bern
1882.
10) F. Koenike, Ueber das Hydrachniden-Genus Atax. Bremen 1881.
[S. 50]
11) F. Koenike, Revision von A. Leberts Hydrachniden des
Genfersees. Zeitschr. f. wiss. Zool. Bd. XXXV, 1881.
12) F. Koenike, Zwei neue Hydrachniden vom Isergebirge.
Zeitschr. f. wiss. Zool. Bd. XLIII.
13) F. Koenike, Eine neue Hydrachnide aus dem Karraschsee bei
Deutsch-Eylau. Schriften der naturforschenden Gesellschaft zu Danzig.
N. F. VII. Bd.
14) F. Koenike, Ein neues Hydrachniden-Genus (Teutonia). Archiv
f. Naturgesch. 1890, Bd. 1.
15) F. Koenike, Einige neubenannte Hydrachniden. Abh. des
naturw. Vereins in Bremen, Bd. IX.
16) Robert Schaub, Über die Anatomie von Hydrodroma. Ein Beitrag
zur Kenntnis der Hydrachniden (mit 6 Tafeln). Sitzungsberichte der
kaiserl. Akad. der Wissenschaften in Wien. Math.-naturw. Klasse, Bd.
XCVII, 1888.
17) Robert Schaub, Über marine Hydrachniden nebst einigen
Bemerkungen über Midea (mit 2 Tafeln). Sitzungsberichte der kaiserl.
Akad. der Wissenschaften in Wien. Math.-naturw. Klasse. Bd. XCVIII,
1889.
18) Th. Barrois, Matériaux pour servir à l’étude de la faune des
eaux douces des Azores. I. Hydrachnides. Lille 1887.
19) Th. Barrois et R. Moniez, Catalogue des Hydrachnides
etc. Lille 1887.
20) Th. Barrois, Note sur la dispersion des Hydrachnides. Revue
biolog. du Nord de la France, T. I. 1888–1889.
[S. 51]
Kerfe und Kerflarven des süssen Wassers, besonders der stehenden
Gewässer.
Von Dr. E. Schmidt-Schwedt in Berlin.
[S. 53]
Vorbemerkung. Der folgende Aufsatz will nur Skizzen aus
der Kerfwelt des süssen Wassers geben; dass dabei ganz wesentlich die
Kerfe und Kerflarven der stehenden Gewässer berücksichtigt worden
sind, ist schon in der Überschrift angedeutet worden. Ich habe diese
Beschränkung geübt, um mich in den Schilderungen fast ganz auf eigene
Anschauung, auf eigene Beobachtungen stützen zu können. Auch der
Kundige, so hoffe ich, wird darin, besonders in biologischer Hinsicht,
manches Neue finden; in jedem einzelnen Falle es als solches kenntlich
zu machen oder zu abweichenden Angaben in der Litteratur besonders
Stellung zu nehmen, schien mir in einem Aufsatze wie dem vorliegenden
überflüssig.
Einleitung.
Wenn von dem zahllosen Heer der Kerfe die Rede ist, denkt wohl fast
Jeder zunächst nur an die augenfälligen Vertreter derselben, welche dem
Luftmeer angehören und auf Blatt und Blüte, wie auf und in der Erde
ihr Wesen treiben: dass auch dem Wasser Vertreter dieses „luftigen“
Volkes in reichlicher Menge angehören, der Gedanke liegt wohl Vielen
fern. Wer dann unter kundiger Leitung besonders ein pflanzenreiches
stehendes Gewässer etwa im Mai mit dem Netz[X] untersuchen will,
der dürfte wohl staunen[S. 54] über das reiche Insekten- und vornehmlich
Insektenlarvenleben derselben und wenn er einiges mit nach Hause nimmt
zu genauerer Untersuchung und Beobachtung der Lebenserscheinungen,
könnte sich leicht ein dauerndes Interesse für diese Tierwelt
daraus entwickeln. Derjenige nun, welcher seine Aufmerksamkeit den
Wasserinsekten zuwenden will, kann zeitig im Frühjahr mit seiner
Thätigkeit beginnen. Wenn kaum einige Zeit das Eis unserer Gräben und
Teiche den wieder kräftiger werdenden Sonnenstrahlen gewichen ist,
beginnt schon ein grosser Teil der im Wasser lebenden Insekten und
Insektenlarven ihr gewohntes Treiben. Geraume Zeit also, bevor der
erste Schmetterling, die erste Grabbiene (Andrena), die erste
Pollenia (Fliegenart) erscheint, bietet ein Ausflug nach Teichen und
Gräben auf Wasserinsekten reichliche Ausbeute. Ja, manche Vertreter
unserer biologischen Gruppe scheinen eine Winterruhe kaum zu halten;
wenigstens habe ich mehrfach selbst im Januar Larven der Käferfliegen
unter dem Eise ihrer Lebensaufgabe, dem Frassgeschäft, obliegen und
Schwimmkäfer wie Wasserwanzen sich ebendort tummeln sehen. Es ist eben
das Wasser, die Urheimat alles Lebendigen, für alle Lebewesen, die
in seinen Schoss sich begeben haben, in vieler Hinsicht ein ungleich
freundlicheres Element als das Luftmeer. Die besonders im Frühjahr
sprungartigen Veränderungen in der Wärme der Luft, die allen Tieren,
welche zu früh ihre Winterplätze verlassen haben, leicht verderblich
werden, gleicht das Wasser mässigend aus und die trüben Regen- und
Schneeschauer des Frühjahrs hält es seinen Inwohnern sicher vom
Leibe. Bei den nicht zu hohen Wärmegraden bietet sogar das Wasser
im Frühjahr denen unter seinen Bewohnern, welche zur Atmung nicht
die atmosphärische Luft, sondern die im Wasser gebundene benutzen,
besonders günstige Daseinsbedingungen, da der Luftgehalt des Wassers
dann wegen der geringen Wärme grösser ist als im Sommer[XI]. Das
reichste Insektenleben zeigen die Gewässer wohl im Mai; die Zahl
der Larven, welche den grösseren[S. 55] Bestandteil der Insektenwelt des
Wassers ausmachen, ist dann auf ihrem Höhepunkt. Zum Hochsommer
nimmt deren Zahl wieder ab. Nicht wenige der dem Wasser angehörigen
Insektenlarven überwintern freilich selbst in diesem Zustand, z. B.
einzelne Schwimmkäferlarven, zahlreiche Mücken-, Köcherfliegen- und
Eintagsfliegenlarven.
[X] Ich benutze dazu mit Vorliebe wenig tiefe Netze aus
weissen Rosshaaren. Dieselben bieten besonders den Vorteil, dass der
Stoff im Wasser nicht quillt, also die Lücken gleich weit bleiben und
dass ferner Algenschleim u. dergl. nicht daran haften bleibt.
[XI] Auch bei den Ausflügen, die dem Fange unserer Tiere
gelten, macht sich geringe Wärme als günstiger Umstand bemerkbar;
ungleich leichter als an warmen Sommertagen bringt man die Tiere ohne
Abgang an Toten an kühlen Frühlingstagen heim.
Ist nun auch die Zahl der Insekten und besonders der Insektenlarven,
welche im Wasser leben, eine recht erkleckliche, so ist doch
anderseits zu betonen, dass die eigentliche Stärke dieser Grossmacht
des Tierreichs auf der Erde, in der Luft liegt und — was für unsere
Betrachtungen wichtiger ist — dass das Verhältnis der Insekten zum
Wasserleben im wesentlichen ein gleiches ist wie das der Säugetiere
zu demselben. Ich will damit sagen, dass hier wie dort der Grundtypus
der Klasse in seiner ganzen Gestaltung auf das Landleben hinweist
(wie etwa umgekehrt der der Fische und Krebse auf das Wasserleben)
und dass die im Wasser lebenden Vertreter, bei den Säugetieren z. B.
Seeotter, Seehund, Walfisch, zwar mehr oder minder weit gehende
Abänderungen von jenem Grundtypus zeigen, die eben im Zusammenhange
mit ihrem besonderen Aufenthaltsorte stehen, aber doch immer noch
deutlich genug erkennen lassen, dass solche Eigentümlichkeiten erst
etwas nachträglich Hinzugekommenes sind, oder vom Standpunkte der
Abstammungslehre aus: Die Vorfahren dieser im Wasser lebenden Tiere
waren Landtiere; erst nachträglich haben sich die Anpassungen, welche
in Beziehung zum Wasserleben stehen, herausgebildet. Zwei Gebiete der
Lebensthätigkeit nun werden bei der Gegenüberstellung von Land- und
Wasserleben besonders berührt und also auch die denselben dienenden
Organe: Atmung und Bewegung[XII]. Diesen beiden Funktionen
und ihren Organen werden wir also im Folgenden in erster Linie unsere
Aufmerksamkeit zuzuwenden haben.
[XII] An dritter Stelle wären die Sinnesorgane in Betracht zu
ziehen, doch muss ich dieselben, da dies ein völlig unbebautes Gebiet
ist, im Folgenden ausser Betracht lassen.
Bei den Säugetieren ist die Zahl der Ordnungen, aus denen Vertreter
dem Wasser angehören, nicht eben gross, wenige Raubtiere, Nagetiere
und die beiden Ordnungen der Robben und Wale;[S. 56] ein gleiches trifft
für die Insekten zu, wenn man die entwickelten Tiere im Auge hat —
es sind dann nur Käfer und Schnabelkerfe (Wanzen) zu nennen. Anders
gestaltet sich aber das Bild, wenn man, wie billig, auch die Larven
berücksichtigt. Dann ist nur eine Ordnung im Wasser nicht vertreten,
nämlich die der Hymenopteren, d. h. der Verwandten von Biene, Wespe
und Blattwespe. Um nun die stattliche Reihe aller dieser Vertreter in
eine übersichtliche Anordnung zu bringen, dürfte es sich empfehlen, die
natürlichen Gruppen des Insektenvolkes, die Ordnungen, zu Grunde zu
legen.
Käfer.
1. Taumelkäfer (Gyriniden).
Zu den wenigen Vertretern der Wasserinsekten, welche sich auch dem
flüchtigen Auge des Spaziergängers aufdrängen, gehören die mässig
grossen (bis 7 mm), nicht eben zahlreichen Arten der Taumel-
oder Drehkäfer (Gyriniden). In vielfach durch einander geschlungenen
Linien schwimmen sie blitzschnell bei schönem Wetter gesellig auf
der Oberfläche stehender wie fliessender Gewässer dahin. Diesen
Tummelplatz, die Oberfläche des Wassers, teilt mit ihnen unter den
Wasserinsekten in gleich ausgesprochener Weise nur noch eine Familie
der Schnabelkerfe, die Wasserläufer (Hydrodromici), die
auf langen Beinen zur Überraschung des Zuschauers auf dem Wasser
dahinlaufen[XIII].
[XIII] Auf der Oberfläche des Wassers trifft man nicht selten
auch verschiedene Fliegenarten, besonders solche aus der Familie der
Dolichopiden und Musciden (Ephydrinen). Ebenso leicht, wie sie auf dem
Wasser dahin laufen, fliegen sie auch wieder von dort fort; Anpassungen
an das Leben an diesem Aufenthaltsort wie bei Taumelkäfern und
Wasserläufern sind mir nicht bekannt. — An vierter Stelle könnte man
hier vielleicht noch die winzigen Springschwänze (Poduren) nennen, die
sich bisweilen in der Nähe des Ufers in grossen Scharen ansammeln und
dann einer Masse Schiesspulver gleichen, dessen Körner auf rätselhafte
Weise in hüpfende Bewegung versetzt sind.
Unterscheiden sich die Taumelkäfer von den Wasserläufern schon dadurch,
dass sie an der Oberfläche dahinschwimmen, nicht auf ihr laufen,
so kommt dazu noch weiter, dass sie auch, was jene nie thun, leicht in
das Wasser hinabtauchen, sei es nun, um sich der Ungunst des Wetters
oder drohender Gefahr zu entziehen oder um Beute zu erhaschen. Während
der Käfer auf der[S. 57] Oberfläche schwimmt, scheint er gleichzeitig die
Umgebung über wie unter Wasser zu mustern; wenigstens
weist darauf eine seltsame Einrichtung seiner Augen hin. Jedes
derselben ist nämlich durch eine breite Chitinleiste in einen nach
oben und einen nach unten gewendeten Teil geschieden, so dass man bei
unseren Tieren von vier Netzaugen sprechen kann.
Die Ausgestaltung ihrer Beine zum Schwimmen ist vollkommener als bei
irgend einem anderen Käfer und zwar gilt das in annähernd gleichem
Grade für Mittel- wie Hinterbeine, während die Vorderbeine, welche
nur zum Greifen und Festhalten benutzt werden, armartig verlängert
sind. An den Mittel- und Hinterbeinen sind besonders die Schienen
und vier ersten Fussglieder flossenartig verbreitert und dabei
etwas schaufelartig ausgehöhlt; der Aussenrand ist überdies mit
Schwimmborsten besetzt. Schwimmen, besonders schnelles Schwimmen mit
den Gliedmassen, ist eine ansehnliche Arbeitsleistung und stellt
daher an die Festigkeit des Körperbaues, vornehmlich hinsichtlich der
Anfügung der Gliedmassen, hohe Anforderungen. Wer den Bau unseres
Käfers auf diesen Gesichtspunkt hin untersuchen will, wird ihn in
Einklang mit jenen Forderungen finden. Besonders die Hinterhüften
sind auffallend gross und fest mit dem Brustskelett verwachsen;
an der Mittelbrust, von deren Hüftteilen annähernd Gleiches gilt
wie von den Hinterhüften, fällt besonders der grosse Mittelteil
(Mesosternum) auf, dessen Ausdehnung einen Schluss erlaubt auf
die auch in diesem Brustring entwickelten Muskelmassen.
Die Anpassung der Beine an das Schwimmen ist eine so ausgeprägte, dass
die Käfer auf dem Lande ebenso unbeholfen sind wie manche Vertreter der
im Wasser lebenden Säugetiere. Wollen sie jedoch von einem Gewässer zum
anderen wandern, so stehen ihnen wie allen Wasserkäfern dazu die Flügel
zu Gebote.
Fig. 5[XIV]. 1 Larve von Gyrinus striatus Oliv. Von den vier Endhaken sind nur
zwei abgebildet. Tr Tracheenkiemen — 2 Larve von Dytiscus
marginalis — 3 Larve von Acilius sulcatus — 4 Larve von
Hyphydrus ovatus. 1 und 4 nach Schiödte, 2 und 3 nach Schiödte und Rösel
kombiniert.
[XIV] Die vergrösserten Figuren für die Zinkographie sind
von Herrn Kupferstecher Ettel, Berlin, dem ich dafür zu Dank
verbunden bin, teils nach vorhandenen Abbildungen, teils nach meinen
Handzeichnungen, teils nach der Natur gezeichnet worden; im letzteren
Falle ist stets Zeichnung und Objekt von mir verglichen worden. Im
ersten Falle ist stets der Schriftsteller genannt worden, dem die Figur
entlehnt wurde, in den beiden letzten ist angegeben worden: nach der
Natur.
Sind die Gyriniden hinsichtlich ihrer Bewegung ausgesprochene
Wassertiere, so bieten sie dagegen rücksichtlich der Atmung nichts
Besonderes dar. Befinden sie sich an der Oberfläche, also mit der
Rückseite ganz an der Luft, so atmen sie wie gewöhnliche Landkäfer;
tauchen sie unter, so sieht man am Hinterende stets[S. 58] ein kleines
Luftbläschen glänzen. Ganz anders verhalten sich in diesem Punkte
die Larven der Taumelkäfer. Diese besitzen (Fig. 5, 1 Larve von
Gyrinus striatus) jene eigenartigen Atmungsorgane, welche[S. 59]
einem Teil der Insektenlarven des Wassers zukommen und mit dem Namen
Tracheenkiemen belegt worden sind. Wie die gewöhnlichen Kiemen
vermögen diese Organe dem Wasser den absorbierten Sauerstoff zu
entnehmen, enthalten jedoch nicht einen lebhaften Blutstrom, sondern
verzweigte Luftröhren (Tracheen), welche von zartem Protoplasma umgeben
sind. Bei den Gyrinidenlarven sind diese Organe seitlich bewimpert (s.
Fig. 5, 1); jeder Hinterleibsring trägt an der Seite je ein Paar dieser
Kiemenfäden, der letzte aber vier derselben, die nach hinten gerichtet
sind. In schlechtem, d. h. sauerstoffarmem Wasser habe ich unsere
Larve mehrfach den Hinterleib auf- und niederschwingen sehen, während
sie sich mit den Beinen festhielt. Offenbar dienten diese Bewegungen
zur schnelleren Erneuerung des für die Atmung nötigen Wassers. Durch
gleichartige Bewegungen des Körpers, also ebenso wie die Egel unserer
Gewässer, vermag auch die Larve ziemlich schnell durch das Wasser zu
schwimmen; dabei gewinnen auch wohl die Wimpern der Tracheenkiemen ihre
Bedeutung. An der Gestalt und Verteilung dieser Organe, an den sechs
Brustfüssen und an den vier gekrümmten Chitinhaken am Hinterleibsende
(in unserer Figur sind nur zwei davon sichtbar) sind unsere Larven
leicht kenntlich.
So häufig übrigens die Taumelkäfer sind, so selten habe ich — und
wie es scheint auch Andere — die Larven angetroffen. Vermutlich
halten sich dieselben in Schlupfwinkeln, etwa unter Steinen am Boden
der Gewässer, auf; dafür sprechen ihre Atmungsorgane, die sie davon
befreien, wie andere Wasserkäferlarven zur Atmung an die Oberfläche
kommen zu müssen, die zarte Beschaffenheit des Hinterleibes und
der Besitz der gekrümmten Haken am Körperende, die anscheinend zum
Festhalten in Gängen oder an Gegenständen dienen. Die Larven leben
übrigens wie die Käfer vom Raube. Zur Verpuppung verlassen sie, wie das
fast für alle Käferlarven des Wassers gilt, dieses Element und gehen an
das Land.
2. Schwimmkäfer (Dytisciden).
Ungleich reicher an Arten als die nur in zwei Gattungen mit zwölf Arten
vertretenen Taumelkäfer ist die Schar der eigentlichen[S. 60] Schwimmkäfer
(Dytisciden). Unter den im Wasser lebenden Käfern sind sie schon
durch die fadenförmigen, auch im Wasser stets frei getragenen Fühler
gekennzeichnet. Nahezu 150 deutsche Arten werden unterschieden, für die
zwölf Gattungen aufgestellt worden sind. Die ansehnlichsten Vertreter
unserer Familie sind die Arten der Gattung Dytiscus und
Cybister (über 2½ cm gross); unter ihnen wieder ist
wohl der Gelbrand, Dytiscus marginalis, diejenige Art, welcher
man am häufigsten begegnet[XV].
[XV] Im Netz gebärdet er sich gewöhnlich sehr unruhig; fasst
man ihn, so sucht er sich durch Entleerung seines Unrats und einer
milchigen Flüssigkeit, die dem Vorderbrustring entquillt, zu befreien.
Der Gelbrand hält sich wie alle seine Familiengenossen, sofern
er nicht fliegend das bisher bewohnte Gewässer mit einem anderen
vertauscht, nur in, nie auf dem Wasser auf, dort seinem
räuberischen Treiben nachgehend. In nicht zu langen Zwischenräumen
muss er jedoch, um zu atmen, an die Oberfläche kommen. Die Hauptmenge
der Atemöffnungen liegt aber an der Rückseite des Hinterleibes unter
den Flügeln und daher steckt der Käfer, das Kopfende schräg nach unten
haltend, nur das Ende des Hinterleibes heraus und lüftet dabei ein
wenig die Flügeldecken. Bei der geringsten Beunruhigung fährt er sofort
in die Tiefe. Eine schnelle Erledigung des in dieser Lage offenbar
gefährlichen Atemgeschäftes wird nun ermöglicht durch ungewöhnlich
grosse, mit vollkommenen Schutzvorrichtungen gegen Staub versehene
Luftlöcher; besonders die dem Körperende näheren sind im Gegensatz zu
dem bei Landkäfern üblichen Verhalten durch Grösse ausgezeichnet. Sind
sie es doch auch, welche hier zunächst in Anspruch genommen werden.
Durch die Luft, welche sich in den Tracheen und unter den Flügeln
befindet, wird übrigens der Körper im Verhältnis zu seinem Rauminhalt
so leicht, dass der Käfer infolgedessen im Wasser emporsteigt und also
sich anstrengen muss, nicht wenn er zur Oberfläche will, sondern
umgekehrt, wenn er zur Tiefe hinab will. Den Weg dahin nimmt er
meist nicht in einfach senkrechter Richtung, sondern fast immer etwas
schräg; er benutzt so den Widerstand, welchen seine breite Gestalt der
Wirkung[S. 61] des Auftriebes entgegenstellt, wie der Arbeiter, welcher das
Fass nicht einfach hebt, sondern auf der Schrotleiter emporrollt.
Einer bemerkenswerten, auf die Atmung bezüglichen Beobachtung sei hier
noch gedacht, die ich zuerst bei Cybister Roeselii machte: die
Haut der Bauchseite dieses Käfers ist so durchsichtig, dass an dem
auf den Rücken gelegten Tier die Luftröhren, besonders die stärkeren,
querverlaufenden Stämme unter den Ringen des Hinterleibes und eine
grosse Tracheenblase der Brust deutlich zu erkennen sind. An ihnen
kann man nun ohne Schwierigkeit wahrnehmen, wie zum Auspressen und
Wiedereinsaugen der Luft ihre Wandungen abwechselnd zusammen und
wieder aus einander gehen. Das erstere geschieht infolge besonderen
Muskeldruckes, das Öffnen dagegen infolge der Spannkraft der
Luftröhrenwände.
Zum Schwimmen benutzt der Gelbrand fast nur die Hinterbeine, deren
Schienen und Fussglieder mit Schwimmhaaren versehen sind, und zwar
beim Männchen mit zwei Reihen, beim Weibchen mit einer. Wie ein
geschulter Schwimmer bewegt der Käfer die in ihren Fussgliedern nach
oben gekrümmten Beine dieses Paares stets gleichzeitig. Von auffälliger
Grösse sind wiederum die fest mit dem Körper verwachsenen
Hüften, die bei der ersten Betrachtung ein Teil der Brust zu sein
scheinen; eigenartig ist die Befestigung der übrigen Beinteile daran,
nämlich derart, dass den Schenkeln und Schienen nur die Bewegung
von vorn nach hinten möglich ist. Die Mittelbeine sind zwar auch
mit einigen Schwimmborsten versehen, ihre Bedeutung beim Schwimmen
besteht aber nur darin, dass sie mithelfen, wenn nötig, die Richtung
des Körpers zu ändern. Ihre Hüften sind klein, kugelig, die ganze
Mittelbrust ist an der Unterseite von der Hinterbrust fast verdrängt,
so dass dadurch die Unterseite des Käfers ein charakteristisches
Gepräge erhält. Dass die geringe Ausbildung der Mittelbrust in
Beziehung steht zu ihrer geringen Muskelmasse und diese wieder zu
der geringen Inanspruchnahme der Mittelbeine, braucht hier wohl nur
angedeutet zu werden. Die Vorderbeine sind fast ganz in den Dienst
des Mundes getreten; sie halten die Beute und führen sie dem Munde
zu. In sprechender Weise kommt die Arbeitsteilung unter den[S. 62] Beinen
zum Ausdruck, wenn man dem Tier etwa einen Regenwurm bietet. Mit den
Vorderbeinen ihn haltend, schwimmt der Käfer mit den Hinterbeinen einem
Ruhepunkte zu, wobei die Mittelbeine die oben angegebene Nebenrolle
spielen; angelangt, hält er sich mit den Mittelbeinen fest, während
die Hinterbeine in einer eigentümlich gekrümmten Haltung stehen
und die Vorderbeine den Mundteilen Handlangerdienste leisten. Der
Unterschied in der Thätigkeit der drei Beinpaare prägt sich auch in
der Krallenbildung aus; nur an den Hinterbeinen sind die Krallen in
ziemlichem Grade verkümmert.
In Beziehung zum Schwimmen steht ferner offenbar die feste
Ineinanderfügung von Kopf, Brustschild (Halsschild) und den übrigen
Körperteilen, wobei besonders auch der Dorn an der Unterseite der
Vorderbrust, der sich in eine Rinne der Mittelbrust legt, zu beachten
ist, und ebenso die Verwachsung der ersten Bauchringe: Durch alle
diese Punkte wird erreicht, dass beim Schwimmen die Teile sich nicht
hin und her bewegen und dass keine unnötigen vorspringenden Teile die
Reibung erhöhen. Zum Durchschneiden des Wassers ist so der flache
Körper trefflich eingerichtet. Als Besonderheit bleibt endlich zu
erwähnen, dass beim Männchen die drei ersten Fussglieder der Mittel-
und Vorderbeine in ausgeprägter Weise zu Haftorganen umgebildet
sind; die der Vorderbeine bilden eine rundliche Scheibe, von deren
mit Saugnäpfchen versehenen Haaren sich besonders zwei durch Grösse
auszeichnen. Dass infolge der Elastizität der Wandung die einmal
angedrückten Teile eben nach dem Prinzip von Saugnäpfen ohne weiteres
Zuthun des Käfers haften, lehrt ein Versuch mit den toten Tieren.
Drückt man die feuchte Haftscheibe etwa an eine Glasplatte, so bleibt
nicht nur beim Aufheben dieser Platte der Käfer daran hängen, sondern
man kann auch noch einen zweiten und dritten an den ersten anhängen.
Die Weibchen anderseits zeigen einen eigentümlichen Dimorphismus,
insofern nämlich, als die eine Reihe derselben mit gefurchten, die
andere mit glatten Flügeldecken versehen ist.
Die Larven der Gattung Dytiscus (Fig. 5, 2) sind im Frühjahr
in unseren stehenden Gewässern eine recht häufige Erscheinung.[S. 63]
Abgesehen von den Larven der Libellen, die wegen der bald entwickelten
Flügelansätze nicht mit Käferlarven verwechselt werden können, sind
sie wohl mit denen von Cybister die grössten Insektenlarven
des süssen Wassers; erwachsen messen sie etwas über 5 cm. Als
Schwimmkäferlarve ist sie kenntlich durch die grossen sichelförmigen,
auf der Innenseite nicht gezähnten Oberkiefer, durch den Mangel
einer eigentlichen Mundöffnung, durch den platten Kopf, ferner
durch die Zweizahl der Krallen an den drei Brustfüssen und endlich
durch die stärkere Chitinisierung der Rückenseite — nur der
letzte Hinterleibsring ist ringsum stärker chitinisiert[XVI]. Als
unterscheidend für die Larven unserer Gattung wäre neben der
Grösse hervorzuheben, dass zwischen den vier längeren Fühlergliedern
anscheinend je ein kürzeres eingeschaltet ist, dass die beiden letzten
Leibesringe seitlich mit stärkeren Schwimmhaaren versehen sind und dass
die beiden am Körperende befindlichen Anhänge gross und stark befiedert
sind.
[XVI] Vgl. dazu den Versuch einer Tabelle über die Kerflarven
am Ende dieses Aufsatzes.
Wie der Käfer muss die Larve zum Atmen an die Oberfläche kommen; die
beiden einzigen thätigen Luftlöcher[XVII] liegen am Ende des letzten
Ringes. Die beiden Körperanhänge flach auf dem Wasser ausbreitend,
hängt die Larve beim Atmen in S-förmiger Krümmung an der Oberfläche.
[XVII] Die sieben anderen Stigmenpaare des Hinterleibes und
die beiden Paare der Brust sind geschlossen und besonders an jüngeren
Tieren schwer wahrzunehmen; eine Bedeutung haben sie nur noch bei
den Häutungen der Larve, indem sie dann als Anheftepunkte der alten
Tracheen dienen, die aus den neugebildeten herausgezogen werden müssen.
Wird sie beunruhigt, so schnellt sie sich durch einen kräftigen Schlag
des Hinterleibes ein gut Stück in die Tiefe. Diese Zuhilfenahme
des Abdomens bei der Bewegung ist übrigens ein charakteristischer Zug
vieler Insektenlarven gegenüber dem entwickelten Tier. Schwimmt die
Larve ruhiger im Wasser umher, so benutzt sie dazu in gleichmässiger
Weise die drei mit Schwimmhaaren versehenen Beinpaare, die Beine
desselben Paares aber meist nicht wie der Käfer gleichzeitig bewegend.
Die Larve führt ein ausgesprochen räuberisches Leben, dabei frisst sie
ihre Beute nicht, sondern saugt derselben die Säfte aus;[S. 64] besonders
oft habe ich sie so Kaulquappen aussaugend beobachtet. Wie schon oben
erwähnt, fehlt eine eigentliche Mundöffnung; aber die Oberkiefer haben
auf der Innenseite eine durch Ineinandergreifen der Ränder geschlossene
Rinne, welche nach der Spitze zu offen ist und nach dem Grunde zu in
den Verdauungskanal mündet. Will man die Larven zur Verpuppung bringen,
so darf man es nicht an Nahrung und einer Gelegenheit, an das Land zu
kommen, fehlen lassen.
Ist hier auch nicht der Ort, auf die Besonderheiten der anderen
Gattungen allgemeiner einzugehen, so möchte ich mir doch nicht
versagen, auf den einzigen, nicht eben häufigen Käfer unserer Familie
hinzuweisen, der bei Berührung Töne von sich giebt — er benutzt
dazu die Flügeldecken und letzten Hinterleibsringe —, und der auch
sonst eine eigenartige Stellung in der Familie einnimmt: Pelobius
hermanni, und ferner auf die beiden Gattungen Haliplus und
Cnemidotus, deren kleine Arten bei abweichender Hüftenbildung
der Hinterbeine nicht regelrecht schwimmen, sondern pudeln.
Zur Bestimmung der Käfer stehen zahlreiche Bücher zur Verfügung; die
Bestimmung der Larven bietet dagegen grosse Schwierigkeiten[1][XVIII]; ja
viele von ihnen sind erst durch die hervorragenden Arbeiten Schiödtes
bekannt geworden. Ich versuche hier noch einige der häufigeren,
wenigstens der um Berlin häufigeren Larven kenntlich zu machen.
[XVIII] Die Buchstaben verweisen auf die Litteratur am Schluss
des Aufsatzes.
Leicht zu kennzeichnen ist die nicht seltene Larve von Acilius
sulcatus (Fig. 5, 3), die erwachsen etwa 3 cm misst. Der
erste Brustring ist etwa dreimal so lang als in der Mitte breit; die
mittleren Hinterleibsringe sind stark verbreitert, die beiden letzten
Ringe seitlich mit Schwimmhaaren versehen, die beiden Anhänge dagegen
unbewimpert. Der Kopf ist durch schwarze Flecke in der Mitte und an den
Seiten ausgezeichnet; die Fühler haben auch hier abwechselnd längere
und kürzere Glieder. Die ersten Jugendstadien der Larve sind übrigens
fast ganz schwarz.
[S. 65]
Gewissermassen ein Gegenstück zu der Larve von Acilius bildet
die bis 2.2 cm lange Larve von Colymbetes fuscus. Der
erste Brustring derselben ist nämlich 1½mal so breit als lang und
breiter als die Mitte des sich nach hinten verjüngenden Abdomen. Nur
der letzte Ring trägt einen seitlichen Haarbesatz, dagegen sind die
beiden Schwanzanhänge gleichmässig und beiderseitig bewimpert. Die
Fühler sind einfach viergliedrig. Sehr nahe schliessen sich hieran die
Larven von Agabus und Ilybius. Larven dieser Gruppe habe
ich vielfach in einzelnen Exemplaren auch überwinternd angetroffen.
Unter den kleineren, d. h. bis 1 cm grossen Larven ist die
von Hyphydrus ovatus (Fig. 5, 4) durch die lange, schmale,
schnabelartige Verlängerung des Kopfes nach vorn auffällig. Das
Körperende läuft in drei Anhänge bez. Fortsätze aus, von denen der
mittlere haarlos ist.
Einen ähnlichen, aber weit kürzeren und breiteren Stirnfortsatz
besitzen die noch kleineren Larven von Hydroporus
parallelogrammus.
Endlich erwähne ich wegen ihrer Eigenart die kleinen Larven von
Cnemidotus, die an Brust und Hinterleib mit fadenförmigen,
gegliederten Tracheenkiemen versehen sind und wie die von
Haliplus nur je eine Kralle an den Beinen besitzen, und die nach
Schiödte mit wahren, d. h. bluterfüllten Kiemen ausgestatteten Larven
von Pelobius.
3. Kolbenwasserkäfer (Hydrophiliden).
Ein bemerkenswertes Gegenstück zu den Schwimmkäfern bilden in fast
allen Punkten die Arten der dritten dem Wasser angehörigen Familie,
der Kolbenwasserkäfer oder Hydrophiliden. Als Vertreter wollen wir
auch wegen seiner Grösse (3.5 bis 4.5 cm) den pechschwarzen
Wasserkäfer (Fig. 6, 5 S. 66) wählen, der treffend hie und da beim
Volke Wasserkuh heisst.
[S. 66]
Fig. 6.
5 Hydrophilus piceus von unten. T Unterkiefertaster — F
Fühler — MH Grenze von Mittel- und Hinterbrust. Die Beine der rechten
Seite sind bis auf die Hüften entfernt — 6 Larve von Hydrous caraboïdes —
7 Kopf der Larve von oben. Ok Oberkiefer — Uk Unterkiefer —
Ul Unterlippe — F Fühler — 8 Eiergehäuse mit Blatt, das letztere
etwas abgebogen. — 5 und 8 nach der Natur, 7 nach Schiödte, 6 nach Schiödte und
Rösel kombiniert.
Kolbenwasserkäfer werden unsere Käfer wegen der Gestalt ihrer
Fühler genannt; wenn man freilich dieses unterscheidende Kennzeichen
an unserem Vertreter oder an irgend einem seiner Verwandten, so lange
er im Wasser ist, aufsuchen will, kann einem Seltsames begegnen: die
Fühler scheinen doch fadenförmig, nicht kolbenförmig zu sein.
Die Lösung ist: der Käfer trägt im Wasser nie wie die Dytisciden
die Fühler vorgestreckt, sondern nach unten[S. 67] und hinten in die an
der Unterseite haftende Luftmasse zurückgelegt; dafür aber sind die
Kiefertaster so auffallend lang, dass sie leicht für Fühler gehalten
werden können. Palpicornes wurden deswegen auch die Käfer
genannt. Sobald sie freilich aus dem Wasser genommen werden[XIX],
pflegen sie die Fühler aus dem Versteck hervorzustrecken. Es macht den
Eindruck, als ob er wie alle seine Verwandten in Fühlern und Tastern
für Luft und Wasser besondere Sinnesorgane hätte.
[XIX] Beim Gelbrand hatten wir bei dieser Gelegenheit
seiner nicht eben sauberen Bemühungen zu gedenken sich zu befreien.
Hydrophilus scheint ausser den lebhaften Bewegungen der mit
Dornen versehenen Beine, die zusammenwirken mit einem scharfen
Stachel der Hinterbrust, noch ein besonderes Mittel zu gleichem Zweck
anzuwenden. Ergriffen, etwa mit der Pincette an den Beinen, und zwar
nur dann, lässt er einen knirschenden Ton hören. Fasst man den Käfer
mit starker Pincette am Kiel der Mittelbrust, so sieht man, wie er bei
der Erzeugung des Tones den Hinterleib zugleich in der Richtung von
hinten nach vorn und von oben nach unten bewegt. Man kann bei toten wie
lebenden Tieren einen gleichartigen Ton erzeugen, indem man entweder
den Brustkiel niederdrückt oder mit der Pincette den Hinterleib von
hinten nach vorn und etwas nach dem Rücken zu bewegt. Über die Lage der
tonerzeugenden Stellen vermag ich trotzdem keine sicheren Angaben zu
machen.
Atmung. Zur Atmung kommt der pechschwarze Wasserkäfer wie der
Gelbrand zur Oberfläche, jedoch hierbei einen charakteristischen
Unterschied gegen den letztern zeigend. Nicht das Ende des Hinterleibes
steckt er aus dem Wasser, sondern den Kopf bringt er an die Oberfläche,
beugt ihn zur Seite, so dass die behaarte Stelle an der Hinterseite
der Augenwölbung die Oberfläche berührt, und legt dann die konkave
Seite der behaarten Glieder der Fühlerkeule, diese zwischen dem
ersten und zweiten Gliede umbiegend, hier von aussen an, so dass ein
röhrenförmiger Zugang für die Luft zu der behaarten und unbenetzten
Unterseite gebildet wird. Durch die nun beginnenden Pumpbewegungen
des Tieres wird der Körper im Wasser gehoben und gesenkt. In der
seidenartigen Behaarung der Unterseite, die sich in der ganzen
Breite bis zum ersten Hinterleibsringe einschliesslich und von da
an den Seitenrändern erstreckt (Fig. 6, 5), wird die Luft zu den
Stigmen des Hinterleibs fortgeleitet. Doch dürften die Luftlöcher
in der Verbindungshaut zwischen Vorder- und Mittelbrust, welche der
Zugangsstelle der Luft soviel näher liegen, bei unserem Käfer die
wichtigere Rolle spielen. Darauf weist wenigstens der Umstand[S. 68] hin,
dass sie, ganz abweichend von den Verhältnissen beim Gelbrand, die
Hinterleibsstigmen an Grösse übertreffen. Für das Festhalten der Luft
an der Unterseite — der Käfer trägt unter Wasser also einen den
grössten Teil des Körpers einnehmenden „Silber“-überzug — ist auch
die Eigentümlichkeit von Bedeutung, dass die Flügeldecken über den
Hinterleib nach unten vorragen.
Schwimmt der Gelbrand eigentlich nur mit den Hinterbeinen, so benutzt
der Kolbenwasserkäfer dazu Mittel- und Hinterbeine. Die Beine desselben
Paares bewegt er dabei abwechselnd (er pudelt), so dass der Körper
beim Schwimmen hin und her „wackelt“. Eine Hilfe, trotzdem die
Richtung zu halten, gewährt wohl die dach- bez. kielförmige Gestaltung
der Unterseite des Hinterleibes. Die ganze Bewegung ist ungleich
schwächlicher als die des Gelbrandes. Dem entspricht auch die schmale
Gestalt der Hinterhüften, die überdies nicht wie beim Gelbrand mit der
Hinterbrust fest verwachsen, sondern ihr beweglich eingefügt sind.
Die Beteiligung der Mittelbeine am Schwimmen zeigt sich nicht nur in
ihrer Behaarung, sondern auch darin, dass die Mittelbrust nicht wie
beim Gelbrand durch die Hinterbrust von der Unterseite verdrängt ist.
Ähnlich wie der Hinterleib ist auch die Brustunterseite gekielt; der
Kiel der Hinterbrust geht nach hinten in einen langen und scharfen
Schutzdorn aus.
Übrigens sind auch bei unserem Käfer die Vorderbeine der Männchen
ausgezeichnet: das letzte Tarsenglied ist beilförmig verbreitert.
Anderseits findet sich bei einem Teil der Weibchen am vordern Teil des
Seitenrandes der Flügeldecken ein leistenartiger Vorsprung.
Nahrung. Gewöhnlich werden die Hydrophiliden kurzweg als
Pflanzenfresser bezeichnet und das würde den Gegensatz zu den
fleischfressenden, räuberischen Dytisciden recht scharf machen. Doch
ist jene Bezeichnung nicht ohne Einschränkung zutreffend; oft habe ich
Hydrophilus piceus und ebenso den nah verwandten mittelgrossen
(1.5 bis 1.75 cm) Hydrous caraboïdes tote oder auch nur
matte Wassertiere auffressen sehen. Anderseits ist schon auf Grund der
Untersuchung der unverdauten Bestandteile seiner Nahrung[S. 69] deren häufige
Herkunft aus dem Pflanzenreiche nicht zu bezweifeln. Man bezeichnet ihn
also wohl am besten als Allesfresser.
Alles, was wir bisher an dem Vertreter der Kolbenwasserkäfer
kennen gelernt haben, weist darauf hin, dass irgend eine nähere
verwandtschaftliche Beziehung der beiden Käferfamilien zu einander
nicht besteht, und jeder weitere Untersuchungspunkt bestätigt diese
Auffassung. Können wir auch auf Einzelheiten hier nicht weiter
eingehen, so sei doch wenigstens kurz erwähnt, dass neben den
schon erwähnten Punkten (Bildung der Fühler, Brust, Beine etc.)
besonders auch die Bildung der Mundteile und die Flügeladerung für
diese Auffassung spricht. In allen diesen Punkten stellen sich die
Dytisciden, soweit nicht die Anpassung an das Leben im Wasser in
Frage kommt, unmittelbar neben die ebenfalls räuberischen Laufkäfer
des Landes und mit ihnen thun das, wenngleich nicht ebenso nahe, die
Taumelkäfer; dagegen muss für die Hydrophiliden der Anschluss bei ganz
anderen Familien der Landkäfer gesucht werden.
Entwickelung. In nachdrücklicher Weise wird diese Ansicht auch
durch alles, was auf die Entwickelung Bezug hat, gestützt. Schon
in der Eiablage thut sich ein Gegensatz zwischen Hydrophiliden und
Dytisciden kund: während die Dytisciden keine besondere Fürsorge
irgend welcher Art für die Eier und die ausschlüpfenden Larven
treffen, gilt das für die Hydrophiliden in auszeichnender Weise. Die
Arten unserer Gattung und mehrerer anderer fertigen aus einer Masse,
die in Fäden aus röhrenartigen Vorsprüngen des Hinterleibsendes
tritt, ein weisses, rundliches, ansehnliches Gespinst (Fig. 6, 8),
das an einer Seite in einen nach oben gerichteten schornsteinartigen
Fortsatz ausläuft. Im Innern liegen die grossen länglichen Eier und
bringen die ausgeschlüpften Larven geschützt die erste Zeit zu. Bei
Hydrophilus ist das Gespinst der Unterseite eines Blattes
angeklebt, bei Hydrous ist das Blatt ringartig um das Gespinst
befestigt. Wer diese Masse nicht schon von Ansehen kennt, wird leicht
in die Versuchung kommen, vom Ufer aus sie für Papierstücke zu halten,
die in das Wasser gefallen sind.
[S. 70]
Die anfangs bräunlichen, später schwärzlichen, zum grösseren Teile
weichhäutigen Larven[XX] unterscheiden sich weitgehend von denen der
Schwimmkäfer. Als systematisch wichtiges Kennzeichen ist zu betonen,
dass die Beine nicht Fuss und Klaue gesondert haben und die Beine
nicht wie fast immer bei den Schwimmkäferlarven mit zwei Klauen,
sondern nur mit einer solchen versehen sind. Stärker fällt in die
Augen der Unterschied der Mundteile: die Unterlippe ist gross und
vorstehend (bei den Schwimmkäfern bis auf die Taster sehr winzig),
die Oberkiefer (Fig. 6, 7) nicht einfach sichelförmig, sondern auf
der Innenseite mit Höckern (bei den Verwandten mit starken Zähnen)
versehen. Die Nahrung besteht nämlich auch bei diesen Larven aus
erbeuteten Wassertieren, z. B. Schnecken; dieselben werden aber nicht
durch die Oberkiefer ausgesogen, sondern vor der Mundöffnung mit
den Oberkiefern zermalmt, und dann die Säfte durch die Mundöffnung
aufgesogen, während die Chitinteile vor derselben liegen bleiben. Durch
diese eigentümliche Art der Nahrungsaufnahme wird die Gewohnheit der
meisten Hydrophiliden-Larven bedingt (für die von Hydrophilus
selbst habe ich es noch nicht beobachtet), die Beute ausserhalb des
Wassers zu tragen, den Kopf dann so zu heben, dass die Mundöffnung
und die davorliegende Beute gerade nach oben gerichtet ist, und in
dieser eigentümlichen Haltung die Beute zu verzehren. Die nicht
benutzbaren Chitinteile werden zuletzt bei Seite geworfen. Im Wasser
würde bei dieser Weise der Nahrungsaufnahme ein grosser Teil der
Säfte der Beute verloren gehen und viel Wasser in den Verdauungskanal
aufgenommen werden; in der Luft dagegen gelangen nur zum Schluss
einige Luftblasen mit in den Verdauungskanal. Dieselben fallen bei
mikroskopischer Betrachtung kleinerer durchsichtiger Larven leicht auf;
ihre Herkunft konnte ich mir lange nicht erklären. Die Bewegung der
Larven ausserhalb des Wassers erinnert übrigens durch das abwechselnde[S. 71]
Strecken und Verkürzen der vordern und hintern Körperhälfte lebhaft an
die Bewegungsart vieler madenförmiger Zweiflüglerlarven. Mit dieser
Bewegungsart hängt augenscheinlich auch die Weichhäutigkeit des
Hinterleibes und der beiden letzten Brustringe bis auf kleine Teile des
Rückens sowie die in der Abbildung nicht wiedergegebene Faltenbildung
des Hinterleibes zusammen.
[XX] Die Larven von Hydrophilus piceus habe ich in
erwachsenem Zustande nicht zu häufig angetroffen; ungleich häufiger
die von dem schon erwähnten nahe verwandten Hydrous caraboïdes.
Deswegen ist auch das Gespinst und die Larve dieses Käfers Fig. 6, 6
und 7 in die Abbildung aufgenommen worden.
Zur Atmung dienen der Larve wiederum nur zwei Stigmen am Ende des
Hinterleibes; dieser Körperteil wird also auch hier an die Oberfläche
gebracht. Zum Schwimmen dienen die oben und unten stark bewimperten
Beine; die vier hinteren werden dabei anders als beim Käfer, nämlich
gleichzeitig bewegt. Bei den Larven von Hydrous caraboïdes
trägt der Hinterleib (Fig. 6, 7) an den Seiten grössere bewimperte
Anhänge, die an die Tracheenkiemen der Gyrinus-Larve erinnern. Doch
können die Anhänge hier nicht als Tracheenkiemen angesprochen werden,
da ihnen ein reicher entwickeltes Luftröhrensystem fehlt. Auch vom
Blutstrom werden sie, wenigstens bei älteren Larven, nicht durchzogen.
Ihre Bedeutung liegt vielleicht in der Oberflächenvergrösserung für
mediane Schwimmbewegungen des Hinterleibes. Für gewöhnlich sieht man
freilich auch unsere Larve nur mit den Beinen schwimmen und zwar
auffälligerweise mit allen gleichzeitig.
Zur Verpuppung gehen die Larven der Hydrophiliden ebenso wie die der
Dytisciden ans Ufer, um dort eine geschützte Stelle über dem Wasser
aufzusuchen.
Die übrigen
Hydrophiliden.
Ziemlich zahlreich sind die Verwandten des pechbraunen Wasserkäfers,
wenn auch nicht so zahlreich wie die des Gelbrandes; die Artenzahl der
Hydrophiliden ist etwa halb so gross wie die der Dytisciden. An Grösse
kommt unserem Käfer, von der zweiten Art seiner Gattung Hydrophilus
aterrimus abgesehen, keine Art auch nur annähernd gleich; die
nächst grösste, oben mehrfach genannte Art geht schon auf 17 mm
herab, die folgenden (Hydrobius-Arten) meist auf 7–8 mm
und die meisten anderen erreichen nicht mehr 5 mm Länge. Mit
Ausnahme von Hydrous caraboïdes, den[S. 72] Arten von Hydrobius
und Berosus besitzt keine Art mehr bewimperte Beine und die
Fähigkeit zu schwimmen. Die Mehrzahl der Arten kriechen also nur
an den Pflanzen unter Wasser umher; losgerissen treiben sie hilflos
zur Oberfläche und suchen dann, den Bauch nach oben, mühsam wieder
eine Pflanze oder das Ufer zu erreichen[XXI]. Auch die genannten Arten
benutzen übrigens keineswegs immer ihre Schwimmfähigkeit, um von einer
Stelle zur andern zu kommen, wie das für die Dytisciden gilt, sondern
oft genug sieht man sie, besonders die Arten von Hydrobius, an
Gegenständen im Wasser umherkriechen.
[XXI] Einige Arten, welche der Familie zugerechnet werden,
leben sogar beständig ausserhalb des Wassers.
Die Fähigkeit, willkürlich Töne hervorzubringen, habe ich ausser bei
Hydrophilus noch bei Hydrous caraboïdes, Spercheus
emarginatus, Hydrobius oblongus und Berosus luridus
beobachtet[XXII]. Besonders die beiden letzten Arten knirschen bei
Berührung bez. nach derselben sehr laut und regelmässig; bei
Hydrous habe ich die Töne mehrfach abends gehört, als eine
Schale mit mehreren Tieren dieser Art unmittelbar vor mir stand. Auf
Berührung hat er dagegen nicht geantwortet.
[XXII] Bei Spercheus wurde, wie ich beobachten konnte,
bei der Erzeugung des Tones der Hinterleib nicht in der Mittellinie des
Körpers, sondern seitlich bewegt.
Abgesehen von diesen beiden Punkten stimmen jedoch auch die übrigen
im Wasser lebenden Verwandten mit unserem Vertreter in fast allen
biologischen Punkten ebenfalls überein, so besonders in Fühler-,
Kiefer-, Brustbildung. Sie zeigen auch z. B. unter Wasser stets nur die
Taster, erst an der Luft die Fühler; so nehmen sie ferner stets auf
der Unterseite die Atemluft mit in das Wasser und zwar ist bei allen
ausser der Gattung Hydrophilus die ganze Unterseite mit Luft
bedeckt. Auch sorgen alle in besonderer Weise für die ausschlüpfenden
Larven, wenn auch nicht genau in der Weise wie Hydrophilus.
Für Hydrous wurde das Gespinst für die Eier schon erwähnt.
Ähnliche mit hornartigem Ansatz versehene Gespinste in freilich viel
kleinerem Massstabe werden von Helephorus gefertigt. Ein
Weibchen des Helephorus aquaticus z. B.,[S. 73] das ich für sich in
einer kleinen Schale hielt, lieferte Ende Mai solche Gespinste an Kork
angeklebt in Mehrzahl mit ungefähr je zehn Eiern. Hydrobius und
Philhydrus kleben ihre weissen, pantoffelförmigen, „ungehörnten“
Gespinste im ersten Frühjahr an den Blättern stehender Pflanzen fest;
besonders oft habe ich sie in Mehrzahl an Grasblättern getroffen.
Die Weibchen von Spercheus und Helochares tragen die
verklebten Eierpakete unter dem Hinterleibe mit sich herum, sie mit den
Hinterbeinen und den vorstehenden Rändern der Flügeldecken haltend.
Beide sind im Mai um Berlin recht häufige Erscheinungen. Die Larven
der Mehrzahl dieser Arten kann man, wie ich vielfach erprobt habe, aus
den gekennzeichneten Gespinsten oder Paketen leicht erhalten. Deswegen
glaube ich darauf verzichten zu sollen, noch die Unterschiede der
Larven der einzelnen Arten anzugeben. Sie besitzen übrigens in den
Mundteilen, der Beinbildung (Fussglied und Klaue nicht gesondert),
Zahl der Fussklauen, Weichhäutigkeit des Hinterleibes alle die oben
geschilderten Merkmale. Hervorgehoben sei nur, dass nach Schiödte
die mir unbekannte Larve von Berosus spinosus mit sieben Paar
fädiger Tracheenkiemen und die von Philhydrus testaceus mit
fünf Paar Afterbeinen ähnlich denen der Raupen, jedoch in anderer
Verteilung, versehen ist.
4. Käfer und Käferlarven
anderer Familien.
a) Parnus.
Mit den kleineren Hydrophiliden findet man oft an Wasserpflanzen
einen 5 mm grossen Käfer bräunlicher Farbe, Parnus
prolifericornis, der ebenso wenig zu schwimmen vermag wie diese
Hydrophiliden, der ebenso wie jene die Luft zur Atmung an der
Aussenseite des Körpers mit unter Wasser nimmt, aber nicht allein
an der Unterseite, sondern wie die Wasserspinne an der ganzen
Körperoberfläche. Als Mittel, diese Lufthülle zu halten, dient ihm
wie der Spinne ein seidenartiger Haarüberzug des Körpers. Biologisch
schliesst er sich ganz den kleineren Hydrophiliden an; Fühlerbildung
und andere morphologische Eigenschaften[S. 74] weisen ihm dagegen mit wenigen
anderen Genossen, die teilweise ebenfalls das Wasser lieben, eine
besondere systematische Stellung an[XXIII].
[XXIII] Die Larve unserer Art ist meines Wissens noch nicht
bekannt, die von Parnus auriculatus fand Hh. Th. Beling in
feuchter Erde. S. Verh. d. Zool.-botan. Vereins in Wien 1882.
b) Cyphon-Larven.
Recht häufig trifft man schon zeitig im Frühjahr in Gräben mit
verwesenden Blättern in Gemeinschaft mit den Larven der Stechmücke
eine Käferlarve, die offenbar keiner der oben genannten Familien
angehört. Wie die Zucht des Tieres ergab, gehört sie zu einer Art der
Gattung Cyphon; die Käfer selbst halten sich gern auf Blättern
in der Nähe des Wassers auf. Die zarthäutige, dunkle Larve wird bis
1 cm lang und ist von oben nach unten stark zusammengedrückt;
als gutes Kennzeichen dient die für Larven ganz ungewöhnliche Länge
der Fühler, welche bei sehr zahlreichen Gliedern länger als die Hälfte
des Körpers sind, und der Mangel an Hinterleibsanhängen irgend welcher
Art. Zur Atmung kriecht die Larve zur Oberfläche; die beiden thätigen
Luftlöcher liegen auch hier am Körperende. Die Mundteile lassen sie als
Pflanzenfresser erkennen. Zur Verpuppung geht auch sie an das Ufer.
Die Larve einer andern Art, die sich durch lichte Färbung von der
obigen unterscheidet, fand ich im Juni besonders an den Blättern des
Froschbiss.
c) Donacia-Larven und -Puppen.
Zum Schluss sei endlich der ganz eigenartigen biologischen Verhältnisse
wegen der Larven und Puppen der Schildkäfer (Donacia und
Haemonia) gedacht. Die bleichen, bis 1½ cm langen Larven
leben im Schlamm an den Wurzeln verschiedener Wasserpflanzen, der
Seerosen, Igelkolben, Schachtelhalme, so dass man ohne besonderes
Suchen ihrer kaum gewahr wird. Da sie ohne Tracheenkiemen oder
eigentliche Kiemen und von sehr träger Bewegung sind, da ferner ihre
derbe Haut und Gestalt den Gedanken an Hautatmung ausschliesst, so
sind sie dem Beobachter hinsichtlich[S. 75] ihrer Atmung zunächst ein
Rätsel. Die Lösung der Atemaufgabe ist bei ihnen so seltsam wie
möglich: sie benutzen die Luft, welche in den stets reich entwickelten
Luftgängen von Wasserwurzeln vorhanden ist. Ich habe mich bemüht[XXIV]
den Nachweis dafür zu führen, dass die Larven dazu die beiden
sichelförmigen braunen Anhänge am Ende des Hinterleibes benutzen, die
nichts anderes seien als eigentümlich einseitig über die Körperhaut
verlängerte Stigmenränder. Diese Anhänge werden, wie unmittelbare
Beobachtung und das Vorhandensein der entsprechenden paarigen Narben
an den Wurzeln ergab, in die Pflanze eingedrückt, durch den Druck des
Pflanzengewebes werden zwei Längsspalten an der Rückseite der Anhänge
geöffnet und nun die Luft eingesogen. Zur Ausatmung dürften die beiden
kurzen Stigmenöffnungen an der Basis der Anhänge dienen. Auch zur
Nahrung dienen den Larven, wenigstens denen von D. crassipes,
die Wurzeln. Zur Zeit, wenn die Verpuppung naht, fertigt die Larve ein
elliptisches Gehäuse, das der Wurzel angeklebt ist, beisst hier ein
Loch in die Wurzel, so dass die ausströmende Luft das Wasser aus dem
Gehäuse verdrängt, und schliesst nun völlig das Gehäuse, um so von Luft
umgeben der Umwandlung zur Puppe und zum Käfer entgegenzugehen. Diese
Gehäuse trifft man an Wurzeln und Rhizomen ungleich häufiger als die
Larven, welche leicht beim Herausziehen der Wurzel abgestreift werden.
Im Gehäuse ist dann entweder noch die Larve, oder schon die Puppe,
nicht selten auch der meist farbenprächtige Käfer vorhanden. Frisst
sich endlich der Käfer durch das Gehäuse durch, so steigt er infolge
der Luftschicht, welche seiner kurzseidenhaarigen Unterseite anhaftet,
an die Oberfläche. Man trifft ihn nicht selten beim Fischen zwischen
den Blättern und Binsen der Oberfläche an. Die Eier werden an den
Blättern bestimmter Pflanzenarten abgelegt. Donacia crassipes
benutzt die Blätter der Seerose und zwar werden die Blätter an einer
Stelle durchbissen und die Eier in zwei Bogenreihen auf der Unterseite
neben diesem Loch angeklebt.
[XXIV] S. Berliner entomologische Zeitschrift Bd. XXXI, S.
325–334, und Bd. XXXIII, S. 299–308.
Durch die Zartheit und Ungeschütztheit der Flügel sind die Zweiflügler
selbst, d. h. die Mücken und Fliegen, von dem Leben im Wasser[XXV] so
gut wie völlig ausgeschlossen; von den Larven und Puppen derselben
aber lebt eine recht erkleckliche Anzahl im Wasser, und diese Zahl
wächst besonders noch an, wenn dabei auch diejenigen Fliegenlarven
mitgerechnet werden, welche sich in Lachen kleinsten Umfangs aufhalten
und halb ein Wasser-, halb ein Luftleben führen. Von den letzteren soll
im folgenden ganz abgesehen werden; auch dann werden wir noch gezwungen
sein, uns auf die häufigeren und typischen Formen zu beschränken.
[XXV] Der Dipteren, welche auf dem Wasser angetroffen
werden, wurde schon oben beim Taumelkäfer kurz gedacht. Eine winzige
Mücke, Clunio adriaticus, lebt seltsamerweise nach v.
Frauenfeld bei Triest unter dem Meeresspiegel an Miesmuscheln.
Vorweg sei der Merkmale gedacht, an denen wir Larven des Wassers
als Dipteren-Larven zu erkennen vermögen.
Zunächst fehlen ihnen stets echte, d. h. gegliederte Beine, die sonst
bei allen Insektenlarven des Wassers vorhanden sind. Bei einigen
Gattungen der Mücken (z. B. Chironomus, Tanypus,
Simulia) kommen zwar sogenannte falsche Beine vor, nämlich
solche, die ungegliedert und mit einer Gruppe von Haken versehen sind,
aber selbst abgesehen von diesem Unterschied der Bildung lässt schon
die Stellung derselben, ein Paar am ersten Brustring und meist ein Paar
am Hinterleibsende, eine Verwechselung dieser Larven mit denen
anderer Ordnungen nicht zu. Da die Verwandlung der Zweiflügler wie
die der Käfer eine vollkommene ist, also den Larven jede Spur
von Flügelansätzen fehlt und auch oft der Kopf wenig deutlich
ist, so haben manche von ihnen ein recht wurmähnliches Aussehen.
Im Zweifelfalle wird stets der Besitz von Tracheen oder von einem
chitinösen Kopfskelett mit Kiefern oder von einer Schlundkapsel mit
Chitinhaken die Dipteren-Larve kenntlich machen.
Fig. 7.
9 Puppe von Culex pipiens — 10 Larve von Mochlonyx culiciformis —
11 Larve von Dixa amphibia. a Luftlöcher — 12 Larve von
Chironomus plumosus. f1, f2 Die
beiden Paare fussartiger Fortsätze mit zahlreichen Chitinhaken — a
Schlauchförmige Anhänge — r Rektaldrüsen — 13 Puppe von Simulia
ornata — 14 Larve von Stratiomys spec. 10, 11, 12, 13 nach Meinert —
9 nach E. Köhne — 14 nach der Natur.
Unter den Dipteren-Larven des Wassers nimmt fraglos die Gruppe
der „Eucephalen“ (Fig. 7, 9–13), d. h. derjenigen, welche[S. 77]
einen deutlichen Kopf mit Fühlern, Augen und ordentlichen Kiefern
besitzen, und für welche die Larve der gemeinen Stechmücke ein
gutes Beispiel ist, den ersten Rang ein. Ihnen werde ich die mit
fadenförmigen Tracheenkiemen versehene Schnakenlarve (Phalacrocera[S. 78]
replicata) anschliessen. Die nächst wichtige Gruppe bilden die
Larven der Waffenfliegen (Fig. 7, 14), gekennzeichnet durch den
einziehbaren, unansehnlichen Kopfteil, durch die deutliche Gliederung
des Körpers und den Kranz von Atemhaaren am Ende des Hinterleibes. An
letzter Stelle werde ich endlich die Larven der Schwebfliegengattung
Eristalis anzuführen haben, denen ein Kopfabschnitt fehlt, deren
Körpergliederung wenig deutlich ist und die mit einem fernrohrartig
einziehbaren Atemanhang versehen sind.
1. Eucephale Zweiflüglerlarven
(Mückenlarven).
Unter allen unseren Insektenlarven giebt es vielleicht keine zweite,
die so leicht zu haben, so leicht zu halten und in ihrer Entwickelung
zu verfolgen ist, und deren Verwandlung dabei einen so überraschenden
Eindruck macht, wie die unserer Stechmücken-Arten[XXVI] (Culex).
Im März, gleich nach dem Auftauen der Gewässer, bei mildem Wetter oft
auch im Dezember und Januar, trifft man in Gräben mit stehendem Wasser,
besonders wenn dieselben vermodernde Laubblätter enthalten, die Larven
der Gattung Culex oft in ausserordentlicher Menge an. Sie sind
um diese Zeit meist noch winzig, etwa 3–4 mm lang, wachsen aber
schnell heran. Zur Aufzucht hat man nur nötig ein Glas oder eine Schale
mit Wasser, zur Nahrung einige vermodernde, untersinkende Blätter. Die
Bälge der Larven findet man nach den bald eintretenden Häutungen im
Wasser schwimmen, den Kopf und Atemanhang derselben von schwärzlicher
Farbe, die übrigen Teile recht durchsichtig. Ist dann nach einigen
Wochen oder, wenn man später grössere Larven nahm, oft nach einigen
Tagen, die Verpuppung eingetreten, so kann man nach einer Frist von
neun bis zehn Tagen das Ausschlüpfen der Mücke erwarten.
[XXVI] Bis zu gewissem Grade teilen alle eucephalen Larven
diese Eigenschaften mit den Culex-Arten.
Zu erkennen sind die Larven der Gattung Culex unter den anderen,
mit deutlichem Kopf versehenen Zweiflüglerlarven unschwer 1) durch
die Verwachsung der Brustringe, deren Zahl noch[S. 79] durch drei grössere
Borstenbündel angedeutet ist, und 2) durch den langen, zur Atmung an
der Oberfläche benutzten Fortsatz des achten Hinterleibsringes. Das
Ende des Leibes geht also scheinbar zweiteilig aus; den einen Gabelast
bildet der eben genannte, von den beiden Hauptluftröhrenstämmen
durchzogene Fortsatz, den andern das eigentliche Körperende mit dem
After (vgl. Fig. 7, 10). An diesen stehen vier ausstreckbare, zarte
Fortsätze (Fig. 7, 12 r), die sogenannten Rektaldrüsen; es
stehen ferner am letzten Segment in der Mittelebene des Körpers nach
unten gerichtet in deutlicher Reihe die wichtigsten Schwimmborsten.
Bewegung. Wie freilich die Bewegung zu stande kommt, lässt
sich, wenn sie recht schnell ausgeführt wird, kaum erkennen; purzelnd
schiesst dann das Tier bald dahin, bald dorthin vorwärts. Erst bei
langsamerer Bewegung unterscheidet man das Wie derselben, so z. B.,
wenn das Tier zur Atmung zur Oberfläche kommt und fast dieselbe
erreicht hat, dann zeigt sich, dass der Hinterkörper mit seiner in der
Mittelebene des Körpers stehenden Borstenreihe rechts und links schlägt
und so den Körper, das Afterende voran, vorwärts treibt. Dem entspricht
nun, dass am Vorderteil die Borsten seitlich gerichtet
sind. Eine eigenartige Fortbewegung kann man ausserdem oft wahrnehmen,
wenn die Larven an den Wänden den Algenansatz benagen: durch die
Bewegung der Mundteile selbst rücken sie dann ziemlich schnell an der
Wand fort.
Atmung und Nahrung. Dass die Larven zum Atmen an die Oberfläche
kommen und dass die vereinigten Atemöffnungen an dem Fortsatz des
achten Segmentes liegen, ist schon oben angedeutet worden. Die
Zeit, in welcher sich dieses Emporkommen wiederholt, ist ziemlich
kurz. Die Tiere hängen dabei in etwas schräger Körperhaltung an der
Oberfläche; bei richtiger Haltung des Auges kann man die Einsenkung
der Oberflächenschicht an der betreffenden Stelle deutlich wahrnehmen.
Schliessen sie die Atemöffnungen, so sinken sie langsam an den
Boden. Dort beginnen sie wieder ihre Nahrung, besonders vermodernde
Pflanzenstoffe, zu verarbeiten. Tritt übrigens irgend eine Beunruhigung
bei der Atmung ein, so purzeln sie nach allen Richtungen fort.
Bisweilen heben[S. 80] die Larven auch, wenn sie an der Oberfläche hängen,
ihren Kopf bis zu derselben empor und drehen sich, indem sie lebhaft
die Kiefer bewegen, nach dem Prinzip der Rückwirkung, kreisend um den
Atemfortsatz.
Puppe (Fig. 7, 9). Während bei den Käfern die Larven stets zur
Verpuppung das Wasser verlassen, bleibt bei den Dipteren, deren Larven
im Wasser leben, auch die Puppe im Wasser. Dadurch werden auch bei den
Puppen eigenartige Verhältnisse hinsichtlich der Atmung und Bewegung
bedingt. Die Puppen der „eucephalen“ Larven werfen alle, wie das bei
den Insekten ja die Regel ist, bei einem grossen Teil der Dipteren aber
gerade nicht zutrifft, die letzte Larvenhaut völlig ab; es lassen also
ihre Puppen (s. Fig. 7, 9 u. 13) wie die Schmetterlingspuppen deutlich
bereits Kopf mit Augen, Fühlern und Mundteilen, Brust mit Flügeln und
Beinen und endlich den Hinterleib unterscheiden. Flügel, Beine und
Fühler sind aber nicht wie bei den Schmetterlingspuppen mit dem übrigen
Körper verklebt, sondern frei.
Die Puppen von Culex atmen wie die Larven die Luft direkt,
müssen also entweder stets an der Oberfläche hängen oder die
Fähigkeit haben, sich zu derselben zu bewegen. Das letztere ist
der Fall und so besitzen sie eine für echte Insektenpuppen ganz
überraschende Beweglichkeit. Dabei ist die Art der Bewegung plötzlich
eine ganz andere als bei den Larven: zwar ist auch hier der
Hinterleib der treibende Teil, wie ja auch bei anderen Puppen, z. B.
Schmetterlingspuppen, derselbe durch Beweglichkeit ausgezeichnet
ist, derselbe schlägt aber nicht seitlich, sondern in
der Mittelebene, und ist im Zusammenhang damit am Ende mit
zwei seitlich gestellten Platten versehen. Atemöffnungen sind zwei
vorhanden, nicht aber am Hinterleib, sondern an der Rückseite
der Brust, am Ende zweier leicht beweglicher, trichterförmiger
Anhänge[XXVII].[S. 81] Fragezeichen ähnlich hängen die Puppen zur Atmung an
der Oberfläche.
[XXVII]Palmén behauptet in seiner wichtigen Arbeit
über die Morphologie des Tracheensystems (Leipzig 1877) S. 64
auffallenderweise, dass diese Anhänge keine Öffnungen haben. Ich fand
keine Schwierigkeit, mich an Längsschnitten der Anhänge vom Gegenteil
zu überzeugen; als Luftsieb — die Staubteilchen dort beweisen es —
dienen Haare, die in zierlicher Weise nach Art von Strebepfeilern
emporragen, gruppenweise zusammenneigen und oben verwachsen sind.
Leichter noch gewinnt man die Überzeugung von der Endöffnung dieser
Anhänge, indem man eine Puppe auf ausgeschliffenem Objektträger unter
Wasser bringt, und mit einem Deckglas bedeckt; bei leichtem Druck auf
das letztere sieht man dann eine grosse Luftblase aus den Enden der
Anhänge hervorquellen und, wenn man rechtzeitig wieder aufhört, beim
Nachlassen des Druckes wieder in den Anhang zurücktreten.
Es möge gleich hier vermerkt werden, dass bei allen Puppen dieser
Gruppe sich gleiche oder homologe Anhänge an der Rückseite zwischen
Vorder- und Mittelbrust befinden und dass im Zusammenhang damit
die Bewegung der Puppe stets durch Schlagen des Hinterleibes in
der Mittelebene erfolgt. Die Erklärung dafür dürfte in folgender
Eigentümlichkeit der Entwickelung liegen: Will das Insekt der
Puppenhaut entschlüpfen, so platzt in der ganzen Unterordnung,
welcher unsere Tiere nach dem System von Brauer zugehören, die
Puppenhaut in einem Tförmigen Spalt an der Rückseite der
Brust. Diese muss sich daher bei unseren Puppen zu der Zeit an der
Oberfläche des Wassers befinden, also muss der ganze Bau der Puppe
und ihre immerhin beschränkte Bewegungsweise derart sein, dass die
Rückseite der Brust an die Oberfläche gebracht wird. Die mit Luft
erfüllten Atemtrichter halten dann zuletzt, auch ohne Bewegung des
Hinterleibes, die Puppe in der rechten Stellung an der Oberfläche. —
Das Ausschlüpfen der Mücke und die Erhärtung ihrer Teile erfolgt in
recht kurzer Zeit, in einer bis zu einigen Minuten[XXVIII]. Dass der
ganze Entwickelungsschritt schnell vollzogen wird, ist der gefahrvollen
Lage wegen, in der sich das Tier währenddem befindet, von Wichtigkeit.
Nicht nur die Feinde im Wasser und in der Luft bereiten ihm dann
Gefahr, auch jeder Windstoss kann verderblich werden. Gern warten daher
die Tiere die ruhige, feuchtwarme Luft nach einem Gewitterregen ab.
[XXVIII] Genauere Notizen habe ich von unserer Art darüber
nicht zur Verfügung. Bei einer Puppe von Chironomus plumosus,
aus der die Mücke, während ich dies niederschrieb, ausschlüpfte,
dauerte der Akt vom ersten Platzen der Haut bis zum ersten Auffliegen
der Mücke nicht ganz 20 Sekunden!
Der Laich bildet nach allgemeiner Angabe eine flache runde Scheibe,
die in der Mitte etwas ausgehöhlt ist und deswegen auf dem Wasser
schwimmt. Ich selbst habe das Ablegen der Eier und den Laich noch
nicht beobachten können; so oft ich Laich von der beschriebenen Form
auffand, erwies er sich doch stets bei[S. 82] weiterer Entwickelung als der
von Corethra plumicornis. Stechmücken, die im Aquarium in Menge
ausgeschlüpft waren, haben niemals bei längerer Gefangenschaft, wie ich
das bei anderen Gattungen beobachtet, den Laich abgelegt. Selbst als
ich solche Mücken mit verdünntem Honig und Blut fütterte und wochenlang
unter einer Glasglocke bei Gegenwart einer kleinen Schale mit Wasser
hielt, erhielt ich keinen Laich.
Anopheles sp. und Dixa (Fig. 7, 11). Biologisch
schliessen sich an die Larven von Culex am nächsten die von
Anopheles und Dixa an; beide atmen durch zwei Stigmen
(Fig. 7, 11 a) am achten Hinterleibsring und beide sind
Pflanzenfresser. Schwimmen sie, so geschieht das gleichfalls durch
seitliches Schlagen des Hinterleibes. Von Culex unterscheiden
sie sich durch den Mangel eines Atemfortsatzes und biologisch dadurch,
dass sie sich ganz vorwiegend an der Oberfläche aufhalten, meist
auf dem Rande schwimmender Blätter oder Gegenstände ruhend. Die
schwärzliche Larve von Dixa nimmt dabei stets die Gestalt eines
lateinischen U an und schiebt sich in dieser Haltung ziemlich
schnell durch abwechselnde Bewegung der beiden Schenkel des U
fort, die entsprechend mit Borsten versehen sind. Weder Kopf noch
Körperende, sondern die Mitte des Tieres schreitet hier also voran:
ein seltsamer Anblick. Den Kopf nach oben und hinten zurückgeschlagen,
wirbelt sich die Larve durch die Bewegung der Mundteile die Nahrung zu.
Dem gegenüber ist die Larve von Anopheles durch lichtere Färbung
und einfache gerade Körperhaltung gekennzeichnet; überdies sind bei
Anopheles die drei Brustringe wie bei Culex verwachsen,
bei Dixa wenigstens der erste deutlich frei. Wie die Larve
von Dixa wirbelt auch sie sich die Nahrung durch die Bewegung
der Mundteile zu; auch sie hält dabei die Unterseite des Kopfes nach
oben, erreicht dies aber durch eine Drehung des Kopfes von 180° um
die Längsachse. Die Puppen gleichen in ziemlichem Grade denen von
Culex.
Einen zweiten Typus der mit deutlichem Kopf versehenen Mückenlarven
stellt die viel untersuchte Larve von Corethra plumicornis
dar. Dieselbe liebt pflanzen- und deswegen tierreiche[S. 83] stehende
Gewässer mit grösseren Stellen freien, klaren Wassers. Um Berlin
ist sie eine recht häufige Erscheinung; sie scheint auch stets in
grosser Zahl zu überwintern. Gekennzeichnet wird sie fast hinlänglich
durch die Bemerkung, dass sie von glasartiger Durchsichtigkeit ist;
nur die beiden Luftblasenpaare im vorderen und hinteren Teile des
Körpers, sowie gewöhnlich der gelbliche bis schwach gelbrote Darm
machen sie wahrnehmbar. Anfangs entgeht sie wohl Jedem oft genug,
wenn sie ruhig im Netz liegt. Als weitere Kennzeichen seien hier noch
folgende genannt: ein Atemfortsatz am achten Hinterleibsring ist
nicht vorhanden, ebenso wenig Luftlöcher, der Kopf ist schnabelförmig
verlängert, die Mundteile und selbst die Fühler sind zum Rauben
eingerichtet.
Atmung und Bewegung. Da bei der Larve von Corethra
vom Tracheensystem nur zwei paarige lufterfüllte Anschwellungen vorn
und hinten entwickelt sind, thätige Luftlöcher und auch Kiemen völlig
fehlen, so kann nur Hautatmung für dieselbe angenommen werden.
Gestützt wird diese Annahme in hohem Masse durch die ausserordentlich
zarte und durchsichtige Haut des Tieres, die sie zu einem so beliebten
Objekt für mikroskopische Betrachtung macht. Die Larve kommt also
auch nicht zur Oberfläche; in horizontaler Lage steht sie oft lange
Zeit still im Wasser, um sich dann plötzlich durch einen seitlichen
Schlag fortzuschleudern und zwar anscheinend in völlig regelloser
Weise. Der wirksame Teil ist auch hier der hintere Körperabschnitt;
der neunte Hinterleibsring trägt zahlreiche, lange, in der Mittelebene
stehende, fiederförmig bewimperte Schwimmhaare. Nährt sich die Larve
von Culex von Pflanzenstoffen, so ist die von Corethra
durchaus räuberischer Natur; zartere Wassertiere, ja selbst kleinere
Larven der eigenen Art bilden ihre Nahrung.
Puppe. Ungleich ähnlicher als die Larven sind die Puppen
von Culex und Corethra. Auch die Corethra-Puppe
bewegt sich durch mediane Schläge des Hinterleibes, der am Ende
ebenfalls besondere, seitlich gestellte Schwimmplatten trägt. Auch
die Anhänge der Vorderbrust finden sich wieder, aber sie sind nicht
wie bei der[S. 84] Puppe von Culex trichterförmig offen und dienen
nicht der Atmung[XXIX]. Mehrfach habe ich 30–40 Puppen in einem
höheren Becherglase vor mir stehen gehabt, ohne dass in Stunden
auch nur eine an die Oberfläche zur Atmung gekommen wäre. Mehrere
Centimeter tief unter der Oberfläche schweben sie. Erst wenn sie dem
Ausschlüpfen sich nähern, wenn zwischen Puppen- und Mückenhaut sich
eine Luftschicht ansammelt, steigen sie infolge der Verringerung
des spezifischen Gewichtes dauernd empor. Aber auch dann habe ich
nie die Oberflächenschicht des Wassers an der betreffenden Stelle
unterbrochen gesehen, wie das sonst bei richtiger Stellung des Auges
stets zu sehen ist, wenn Insekten oder Insektenlarven zur Atmung an die
Oberfläche kommen. Die Bedeutung der lufterfüllten Anhänge am Rücken
der Vorderbrust kann also bei Corethra nur darin liegen, dass
sie der Puppe ohne weiteres besonders beim Ausschlüpfen die geeignete
Körperstellung geben. Das Ausschlüpfen der Mücke vollzieht sich auch
hier in recht kurzer Zeit.
[XXIX] Vgl. auch Palmén, „Morphologie des
Tracheensystems“.
Laich. Dass die abgelegten schwarzen Eier eine flache Scheibe
bilden, erwähnte ich schon bei Culex. Nicht selten habe ich
dieselben im Freien angetroffen und auch von ausgeschlüpften Mücken
erhalten, denen durch eine übergestülpte Glasglocke das Fortfliegen
verwehrt war. Die Larven schlüpfen, wenn man nur das Wasser der Schale
durch untergetaucht lebende Pflanzen, z. B. Riccien, frisch erhält,
bald aus der Laichmasse aus.
In merkwürdiger Weise vermittelt den Übergang von Culex zu
Corethra die Larve von Mochlonyx culiciformis (Fig. 7,
10). In Körpergestalt, in Durchsichtigkeit hält sie die Mitte zwischen
jenen beiden Gattungen. Wie bei Corethra finden sich zwei Paar
Luftröhrenanschwellungen vorn und hinten; aber es sind auch noch die
beiden freilich schwachen Längsstämme des Luftröhrensystems vorhanden
und lufterfüllt, die wie bei Culex in einen Anhang des achten
Hinterleibsringes ausgehen. Und nun wird dieser „Atemfortsatz“ doch
wieder nicht wie bei Culex zur Atmung benutzt; die Larven
schweben ebenso wie die von Corethra in horizontaler[S. 85] Lage
im Wasser und niemals habe ich sie an die Oberfläche kommen sehen.
Auch die Mundteile zeigen eine Bildung zwischen denen von Culex- und
Corethra-Larven; freilich stehen sie denen der letzteren näher. In der
räuberischen Lebensweise stimmen sie mit diesen völlig überein. Die
Puppe schliesst sich der von Corethra nahe an. Die schon von
De Geer beobachtete Larve scheint lange Zeit der Aufmerksamkeit
entgangen zu sein. Ich habe sie um Berlin seit einer Reihe von Jahren
vielfach gefunden und zwar stets mit Larven von Culex zusammen.
Chironomus und Tanypus. Einen dritten Typus stellen
die Larven der Gattungen Chironomus und Tanypus dar,
die nicht der Oberfläche oder den mittleren Schichten der Gewässer,
sondern hauptsächlich dem Boden der Gewässer, und zwar sowohl
fliessender wie stehender, angehören. In stehenden Gewässern sind
wohl die blutroten Larven von Chironomus plumosus (Fig. 7, 12)
mit die häufigsten und auffälligsten. Gekennzeichnet sind die Larven
dieser beiden Gattungen durch den Besitz von winzigen Punktaugen
und von je zwei sogenannten falschen Beinen am Vorderbrustring und
am Körperende (Fig. 7, 12 f1, f2) — die Beine
sind ungegliedert und mit einer Gruppe von Chitinhaken versehen —,
ferner durch das Fehlen der Atemöffnungen und eines ausgebildeten
Luftröhrensystems. Die Larven der beiden Gattungen unterscheiden
sich unter einander vornehmlich dadurch, dass bei den Larven von
Tanypus die drei Brustringe mehr oder minder zu einer Masse
verwachsen, bei denen von Chironomus aber frei sind. Überdies
besitzen manche Chironomus-Larven, z. B. die von Ch. plumosus,
vier schlauchförmige, zarte Auswüchse an der Bauchseite des achten
Hinterleibsringes (Fig. 7, 12 a).
Atmung. Die Larven von Chironomus leben, wie schon
angegeben wurde, auf dem Boden, wo sie sich mit Hilfe einer
Schleimmasse, die unterhalb der Mundöffnung austritt, aus den Schlamm-
oder Sandteilchen Röhren zum Schutz und Aufenthalt bauen[XXX]. Thut
man z. B. in eine Schale mit Larven von[S. 86]Chironomus plumosus
etwas Sand, so nimmt der Boden in kurzer Zeit ein Aussehen an, welches
im Kleinen an das Bild einer Wiese erinnert, an deren Oberfläche
zahlreiche Maulwürfe ihre Gänge gezogen haben. Zur Wasseroberfläche
kommen die Larven nicht; ihre Atmung ist wie die von Corethra
Hautatmung. Die Auswüchse am achten Hinterleibsring der Larve von
Ch. plumosus sowie die vier Drüsen um den After unterstützen
wohl die Atmung, da sie von einem lebhaften Blutstrom, in dem
eigenartige, fadenförmige Blutkörperchen treiben, durchzogen werden[XXXI].
[XXX] Ob das auch für einzelne Arten von Tanypus gilt,
muss ich dahingestellt sein lassen.
[XXXI] Dieser Blutstrom wird leicht bei ungünstiger Lage
der Larve auf dem Objektträger unterbrochen und entgeht dann dem
Beobachter. Auch mir war er früher entgangen; erst jüngst, als
ich durch eine Äusserung von Hrn. Weltner, der Larven von
Chironomus auf dem Boden tiefer Seen gefunden hatte, zu erneuter
Beobachtung veranlasst wurde, nahm ich denselben wahr.
Hält man Larven unserer Gattungen in sauerstoffarmem Wasser, so sieht
man oft, wie sie sich mit den Haken der „falschen“ Vorderfüsse an
einigen Gespinstfäden, welche sie am Gefäss befestigt haben, festhalten
und nun lebhaft in medianen, wellenförmigen Bewegungen das Wasser
schlagen, offenbar, um sich beständig frischeres Atemwasser zuzuführen.
Bewegung und Nahrung. Am Boden bewegen sich die Larven
von Chironomus mit Hilfe der vorderen und hinteren falschen
Beine den Spannerraupen ähnlich fort; im Wasser schwimmen sie, wenn
man es so nennen darf, recht ungeschickt ohne feste Richtung durch
Sförmige Krümmungen des Körpers[XXXII]. Als Nahrung dienen
ihnen die Schlammteile des Bodens oder richtiger die darin enthaltenen
organischen Bestandteile.
[XXXII] Bei einer Tanypus-Larve, wohl T. monilis,
beobachtete ich Schwimmen durch wellenförmige Bewegungen in der
Mittelebene des Körpers.
Puppe. Auch während und nach der Verpuppung bleiben die Tiere in
den Schlammröhren; dabei ist die Larvenhaut oft noch nicht vollständig
vom Hinterleibsende der Puppe abgestreift. Als gutes Kennzeichen
der Puppen von Chironomus ist die eigenartige Ausbildung
der Vorderbrustanhänge zu erwähnen; diese sind nicht einfach und
trichter- oder keulenförmig wie bei den früheren Gattungen und bei
Tanypus, sondern bestehen aus einer grossen Menge zarter Fäden,
die von feinen Tracheenröhren durchzogen[S. 87] sind und deswegen silberweiss
erscheinen (vgl. Fig. 7, 13). Sie sind also nach ihrer Bildung als
Tracheenkiemen anzusprechen. Die Puppe zeigt übrigens lebhafte
Atembewegungen, und zwar ein Schwingen in der Mittelebene des Körpers.
Zur Zeit, wenn das Ausschlüpfen naht, steigt sie an die Oberfläche,
wobei die lufthaltigen Brustanhänge ihr den Weg zeigen.
Laich. Unter allen Mücken ist Chironomus diejenige,
deren Laich ich am häufigsten angetroffen habe. Die länglichen,
schwach bräunlichen Eier liegen in wurstförmiger, durchsichtiger
Gallertmasse. Wenig auffällig, entgehen sie leicht der Wahrnehmung.
Ihre Zugehörigkeit lässt sich leicht dadurch feststellen, dass man die
Larven in besonderer Schale mit Wasserpflanzen ausschlüpfen lässt.
Simulia. Biologisch und auch wohl morphologisch schliessen
sich an die Larven von Chironomus und Tanypus die der
Simulia-Arten an. Dieselben leben ausnahmslos in fliessenden
Gewässern. In der näheren Umgegend von Berlin kenne ich sie nur aus der
Panke oberhalb Schönhausen und aus der Wuhle, die bei Köpenick in die
Spree mündet. Sie sind leicht daran kenntlich, dass zwar ein vorderes,
weit verwachsenes falsches Beinpaar wie bei Chironomus und
Tanypus vorhanden ist, dagegen das hintere fehlt, und dafür ein
Kranz von Haken am Ende des im hinteren Abschnitt verdickten Abdomens
steht. Auch sie sind Hautatmer und kommen nie an die Oberfläche.
Einen recht seltsamen Anblick gewährt ihre Bewegungsweise. Mit einer
Gespinstmasse, welche ähnlich der der Raupen aus einem Vorsprung
oberhalb der Unterlippe austritt, ziehen sie am Boden einige kurze
Fäden, greifen in diese mit dem Hakenkranz der Vorderfüsse, krümmen den
Körper zu einer Schleife, greifen nun mit dem hinteren Hakenkranz in
diese Fäden, lassen dann vorn los, strecken sich, spinnen neue Fäden
u. s. w. So bewegt sich die Larve ausgesprochen spannermässig. Hält man
sie im tief ausgeschliffenen Objektträger oder einer andern derartigen
Vorrichtung (angekitteter Glasring), so kann man das Ziehen der Fäden
gut[S. 88] beobachten[XXXIII]. Durch dieses Mittel sind sie befähigt, sich
im strömenden Wasser an Wasserpflanzen und Steinen festzuhalten.
Oft richten sie sich, mit dem hinteren Hakenkranz in die Spinnfäden
greifend, senkrecht zur Oberfläche des Blattes und lassen dann ihren
Strudelapparat am Munde spielen, der ihnen mit dem entstehenden
Wasserwirbel die winzigen Nahrungsbestandteile zuführt.
[XXXIII] Auch bei Tanypus-Larven habe ich diese
Bewegungsweise, wenn auch nicht so ausgeprägt wie bei Simulia,
beobachtet.
Puppe. Die sehr gedrungene Puppe (Fig. 7, 13) lebt in einem
tütenförmigen Gehäuse, das seitlich an Wasserpflanzen mehr oder
minder tief befestigt ist. Ihre Vorderbrustanhänge gleichen denen von
Chironomus, doch sind die Fäden weit weniger zahlreich. Erst zum
Ausschlüpfen steigt sie zur Oberfläche.
Rückblick. Alles in allem genommen, zeigen also die „eucephalen“
Mückenlarven der Gewässer und deren Puppen eine grosse Mannigfaltigkeit
der biologischen Verhältnisse. Atmen die Larven von Culex,
Dixa und Anopheles Luft in gewöhnlicher Weise an der
Oberfläche, so sind die von Mochlonyx, Corethra,
Chironomus, Tanypus, Simulia Hautatmer; bei
einzelnen Arten von Chironomus findet sich dem Anschein nach
nebenbei Atmung durch echte schlauchförmige Kiemen[XXXIV]. Sind
die Larven von Culex, Anopheles, Mochlonyx,
Corethra ausschliesslich oder fast ausschliesslich Schwimmer
und zwar unter seitlicher Bewegung des Hinterleibes, so zeigt die
von Dixa, die U-Larve, daneben eine seltsame,
oben beschriebene Art des Fortschiebens, die Chironomus-Larven ein
unregelmässiges Schwimmen durch Sförmige Krümmungen,
die Tanypus-Larven ein Schwimmen durch mediane Bewegungen, beide
Gattungen ausserdem und Simulia eine spannerartige Bewegung
unter Benutzung von Hakenkränzen am Vorder- und Hinterende des Körpers
und teilweise (Simulia, Tanypus) von Gespinstfäden. Als
Nahrung dienen den meisten Larven Pflanzenstoffe, Mochlonyx
und Corethra aber sind ausgesprochene Raubtiere. Leben die
Larven von Dixa und Anopheles fast ausschliesslich an
der Oberfläche, so die von Chironomus und Tanypus meist
in[S. 89] Schlammröhren am Boden, bisweilen in ansehnlicher Tiefe, die von
Simulia an allen möglichen Gegenständen des Wassers; tummeln
sich Culex-Larven durch alle Schichten hindurch, so bevorzugen die von
Mochlonyx und Corethra die mittleren Wasserschichten.
Dazu kommt dann noch das plötzliche Umspringen der Bewegungsarten bei
der Verpuppung der Mehrzahl der Larven und die Übereinstimmung aller
Puppen in dem Besitz besonderer Anhänge am Rücken der Vorderbrust und
in der Bewegungsart: zwei Punkte, die ich schon oben biologisch zu
deuten suchte.
[XXXIV] Auffälligerweise fehlt die Atmung durch Tracheenkiemen
ganz.
Es kann hiernach nicht überraschen, dass sich die Larven im Anschluss
an diese Unterschiede oft scharf nach der Art der Gewässer in ihrem
Vorkommen sondern. Während ich z. B. in der oberhalb Schönhausen stark
fliessenden Panke nur Larven von Simulia, Chironomus und
Tanypus fand, zeigte sich dicht daneben ein Wiesentümpel mit
Laubblättern, aber ohne nennenswerten Pflanzenwuchs nur von zahlreichen
Culex-Larven bevölkert, und ein naher Graben mit stehendem Wasser und
dichtem Pflanzenwuchs die schwarzen Larven von Dixa und die
helleren von Anopheles. Um Corethra zu beherbergen,
schien er wegen des Mangels an grösseren pflanzenfreien Stellen nicht
geeignet zu sein. Es haben, möchte man sagen, die verschiedenen
biologischen Gruppen die Gewässer unter sich geteilt, oder richtiger
ausgedrückt: es haben sich die Gattungen nach den verschiedenen
Lebensbedingungen der Gewässer unter einander differenziert.
Phalacrocera replicata. Den eucephalen Mückenlarven schliesse
ich am besten die Larve einer Schnake, Phalacrocera replicata,
an, die schon De Geer bekannt war. Sie scheint seitdem
wenig aufgefunden zu sein, wahrscheinlich jedoch nicht wegen ihrer
Seltenheit, sondern weil sie den Blicken so leicht entgeht. Ich habe
sie um Berlin fast überall gefunden, sobald der Pfuhl oder Teich am
Grunde reichlich mit Wassermoosen bedeckt war. An solchen lebt und
frisst sie; den älteren blattlosen Stengeln sieht sie recht ähnlich
und wegen der Trägheit ihrer Bewegungen übersieht man sie doppelt
leicht. Sie ergänzt in merkwürdiger Weise in einer Hinsicht die
Mückenlarven; keine derselben hat Tracheenkiemen,[S. 90] unsere Schnakenlarve
ist auf der Rückseite mit ziemlich starren, gegabelten, fadenförmigen
Tracheenkiemen versehen. Der mangelhaft ausgebildete „Kopf“ ist klein
und kann völlig eingezogen werden. Das Hinterende des Körpers trägt
zwei grosse, gekrümmte Chitinhaken.
Die Puppe besitzt wie die der eigentlichen Mücken zwei
Vorderbrustanhänge und lässt Kopf, Flügel, Beine u. s. w. deutlich
erkennen. Mit zwei Paaren von Fortsätzen an den hinteren Körperringen
— ausserdem stehen noch zwei andere Paare dort — hält sie sich an
Pflanzen fest und streckt die Anhänge zur Atmung heraus. Von der
Verpuppung bis zum Ausschlüpfen der ansehnlichen Schnake verfliessen
nur wenige Tage.
Larven der Waffenfliegen. Von anderen Zweiflüglerlarven sind mir
als einigermassen häufig und auffällig[XXXV] nur noch die unter einander
recht ähnlichen Larven der Waffenfliegen (Stratiomyden) (Fig. 7, 14)
entgegengetreten. Sie sind deutlich gegliedert und mit undeutlichem,
einziehbarem Kopfteil versehen, aus dem zwei seitliche „Fressspitzen“
hervortreten. Am Ende des Körpers steht ein zierlicher Kranz von
Haaren, den die Larven auszubreiten und zusammenzulegen vermögen.
Inmitten dieses Kranzes liegt nämlich der Eingang zu einer Art Vorhof
der Luftlöcher der Larve; wollen die Tiere atmen, so strecken sie
das Körperende an die Oberfläche und breiten den Haarkranz aus. Ihre
Bewegung ist recht unbeholfen; durch lebhaftere Krümmungen kommen sie
langsam in ziemlich regelloser Weise vorwärts. In Übereinstimmung
mit ihrer Atmungsweise und mit ihrer schwachen Bewegungsfähigkeit,
leben sie nur an der Oberfläche stehender pflanzenreicher Gewässer.
Trifft man übrigens etwa im Mai, Juni Larven an der Oberfläche, die
andauernd bewegungslos bleiben, so — hat man es ziemlich sicher mit
einer Puppe zu thun. Sicherheit kann man über diesen Punkt
leicht[S. 91] gewinnen dadurch, dass man die „Larve“ am Ende öffnet; im
zutreffenden Falle ist nämlich die Haut an den Enden hohl und mehr in
der Mitte liegt die zarte, ziemlich fliegenähnliche Puppe. Es wird also
bei der Verpuppung die letzte Larvenhaut nicht abgestreift, sondern
das Tier hat sich innerhalb derselben zur Puppe umgebildet. Erst beim
Ausschlüpfen der Fliege wird auch diese Larvenhaut gesprengt und zwar
in einem Tförmigen Spalt. Von den echten sogenannten
Tonnenpuppen der grossen Menge der Fliegen unterscheiden
sich übrigens diese Puppen wesentlich dadurch, dass erstens die
Larvenhaut bei der Verpuppung weiter keine Veränderung der Form erfährt
und dass das Aufspringen nicht in einem Ringe, sondern in einem
Tförmigen Spalt erfolgt.
[XXXV] Bisweilen habe ich auch in Gräben mit
vielem Algenschleim und wenig sauberem Wasser die bekannten
Rattenschwanzlarven der Schlammfliege angetroffen. Kenntlich sind
die zarthäutigen Larven an dem Mangel eines Kopfes und an dem
fernrohrartig ausstreckbaren Atemrohr am Ende des Körper. Ziemlich
ähnlich sehen ihnen wegen eines gleichen Atemanhangs die Larven von
Ptychoptera (Schnake); doch ist der Kopf deutlich, wenn auch
einziehbar, und die Puppe eine echte Schnakenpuppe (Mumienpuppe)
mit einem sehr verlängerten Vorderbrustanhang. — Wegen mehrerer
Musciden-Larven und -Puppen des Wassers sei hier nur auf G.
Gerke, „Verh. des Vereins für naturw. Unterhaltung zu Hamburg“,
1876 ff., verwiesen.
Die Schmetterlinge selbst sind ebenso und aus gleichem Grunde von dem
Leben im Wasser ausgeschlossen[XXXVI], wie die Fliegen; von den Larven und
Puppen dagegen leben einige auch aus dieser Insektengruppe im Wasser.
[XXXVI] Dabei muss freilich von den flügellosen Weibchen
des Acentropus niveus abgesehen werden, welches nach
Litteraturangaben mit dem Rücken nach unten auf der Wasseroberfläche
umherschwimmt.
Die interessanteste und nach meinen Beobachtungen häufigste dieser
Wasserraupen ist die von Paraponyx stratiotata; ich habe
dieselbe um Berlin fast nirgends vermisst, wo ihre Haupt-Nährpflanze,
die stachlige Wasseraloë (Stratiotes alioïdes), wächst. Ihren
Raupencharakter bekundet die bis auf Kopf und ersten Brustring
zarthäutige Larve durch den Besitz von Afterbeinen mit Hakenkränzen
am dritten bis sechsten und letzten Hinterleibsringe neben den sechs
gegliederten Brustbeinen sowie durch die Bildung der Mundteile.
Dass sie aber ein wohlangepasstes Wassertier ist, zeigt das Fehlen
von offenen Atemlöchern und die Ausbildung von fadenförmigen
Tracheenkiemen, welche die Seiten und den Rücken vom zweiten Brustring
an einnehmen[XXXVII]. Oft findet man die Raupe zwischen zwei Blättern der
Wasseraloë, die lose durch Fäden zusammengeheftet sind;[S. 92] anscheinend
dient solche Stelle dann als Zufluchtsort. In vollkommnerer Weise
spinnt sich die Raupe zur Zeit der Verpuppung ein geschlossenes,
dichtes Gehäuse, das den Blättern fest angeklebt ist. Schneidet man ein
solches Gehäuse auf, so findet man die Puppe nicht in Wasser,
sondern in Luft liegend. Sie hat auch nicht mehr Tracheenkiemen,
sondern Luftlöcher, an denen nur auffällig ist, dass die vorderen auf
einer Erhöhung liegen. Woher stammt nun die Luft im Puppengehäuse,
das doch unter Wasser hergestellt ist? Ich habe an andrer Stelle[XXXVIII]
wahrscheinlich zu machen gesucht, dass die Luft ähnlich wie bei den
Puppengehäusen von Donacia aus der Pflanze stammt und dass diese
unter Druck aus dem Innern der Pflanze hervortretende Luft das Wasser
aus dem noch nicht völlig geschlossenen Gehäuse verdrängt. Man findet
nämlich einerseits die Blätter dort, wo ein Puppengehäuse angeklebt
ist, bis zu ihren grossen Luftgängen angefressen und anderseits ist
eine andere Weise als die angedeutete, das Gehäuse mit Luft zu füllen,
unter den gegebenen Verhältnissen kaum vorstellbar[XXXIX]. Ist das Gehäuse
völlig fertig, so sind übrigens die Frasslöcher zu den Luftgängen
zugesponnen.
[XXXVII] Auch die Wasserraupen von Acentropus niveus sind
nach den Angaben in der Litteratur mit Tracheenkiemen versehen. Aus
eigener Beobachtung kenne ich dieselben nicht.
[XXXIX] Für die ebenfalls kiementragende Wasserraupe einer
brasilianischen Cataclysta-Art hat freilich W. Müller
eine andere Weise, das Puppengehäuse mit Luft zu füllen, beschrieben.
Eine zweite Gruppe von ziemlich häufigen Wasserraupen gehört den
Gattungen Hydrocampa und Cataclysta an. Äusserlich
sind diese Zünsler-Larven den weiter unten zu behandelnden
Phryganiden-Larven recht ähnlich, da sie sich aus zwei
zurechtgebissenen Blattstücken, die von H. nymphaeata z. B. aus
den Blättern der Seerose oder des Laichkrautes oder (Cataclysta)
aus zahlreichen verklebten Wasserlinsen ein Schutzgehäuse herstellen
und damit im Wasser herumkriechen. Sobald man freilich die Larve
aus diesem Versteck herauskriechen sieht, zeigt sich auch abgesehen
von den Hinterleibsbeinen ein wichtiger Unterschied gegen die
Phryganiden-Larven: die Raupen besitzen keine Tracheenkiemen, sie
sind vielmehr bis auf den Kopf und ersten Brustring mit einer im
Wasser silberglänzenden Luftschicht umgeben und ebenso ist auch das
Gehäuse mit Luft[S. 93] erfüllt. Sie sind also einfache Luftatmer und auf
die Oberfläche des Wassers angewiesen. Die Puppe liegt gleichfalls in
diesem Luftbett.
Andere Arten von Wasserraupen, deren Gattungszugehörigkeit mir unsicher
geblieben ist (vielleicht Scirpophaga?), habe ich mehrfach
in ausgefressenen Binsen- oder Schachtelhalmstücken getroffen; das
Verfahren zur Herstellung des Schutzgehäuses war also ein recht
einfaches. Zur Verpuppung fand ich solche Gehäuse an den Stengeln von
Wasserpflanzen in senkrechter Stellung angeklebt und die abstehende
Öffnung verschlossen, ähnlich dem, was man bei den Gehäusen der
Phryganiden beobachtet.
Während in den drei obigen Ordnungen diejenigen Kerfe, die entweder im
entwickelten Zustande oder als Larve im Wasser leben, einen kleinen
oder selbst verschwindenden Teil ihrer Gruppengenossen ausmachen,
ändert sich das plötzlich bei den Netzflüglern in auffälliger Weise.
Die entwickelten Tiere meiden zwar wie die Fliegen und Schmetterlinge
das Wasser, aber unter den Larven lebt die Mehrzahl der freilich
nicht artenreichen Ordnung im Wasser, und zwar stellt die Gruppe der
Köcherfliegen (Phryganiden) die Hauptmasse derselben. Welcher Art
ein Gewässer auch immer sein mag, ob stark fliessend oder stehend,
ob pflanzenreich oder pflanzenarm, ob mit Sand- oder Schlammboden:
fast immer wird man Larven dieser Gruppe darin finden und oft in sehr
auffälliger Menge. Durch einen eigentümlichen Zug in ihrer Lebensweise
machen sie sich auch dem Laien auffällig. Die meisten von ihnen
kitten sich nämlich aus diesem oder jenem Stoff ein köcher- oder
röhrenförmiges Schutzgehäuse, welches sie mit sich herumschleppen und
in das sie sich bei Beunruhigung sogleich zurückziehen. Sie nehmen
dazu feinem Sand, gröbere Kieskörner, Schneckenhäuser mit oder ohne
Bewohner, Gras und breitere Pflanzenblätter, abgebissene Stücke von
Pflanzenstengeln, oder alte Holzstücke etc., und legen diese Teile der
Länge nach oder der Quere nach (Fig. 8, 16 u. 17) oder in spiraliger
Anordnung an einander. Wegen ihrer Auffälligkeit[S. 94] haben sie auch vom
Volke besondere Namen erhalten: Sprottwürmer oder Sprocken heissen sie
z. B. in einzelnen Teilen der Mark.
Fig. 8.
15 Limnophilus rhombicus — 16 Larvengehäuse von vorn — 17 von oben — 18
Puppe von der Bauchseite. 16 und 17 nach Pictet, 15 und 18 nach der Natur.
Ob ein aufgefundenes Tier zu den Phryganiden-Larven gehört oder nicht,
darüber wird auch beim Anfänger selten ein Zweifel sein. In den bei
weitem meisten Fällen wird man die gewöhnliche[S. 95] Larvenform (Fig. 8,
15) vor sich haben, die durch den Besitz des Gehäuses und von drei
Brustbeinpaaren, durch die stärkere Chitinisierung des ganzen Kopfes
und mehr oder minder ausgedehnter Teile der Brust, sowie endlich durch
die fadenförmigen Tracheenkiemen und die beiden Endhaken des zarten
Hinterleibes gekennzeichnet ist[XL]. So leicht nun die Zugehörigkeit
einer Larve zu unserer Gruppe festzustellen ist, so schwierig und
umständlich ist anderseits die Unterscheidung der zahlreichen
Gattungen, von den Arten ganz zu schweigen. Ich werde also im folgenden
ganz davon Abstand nehmen und nur an einem Vertreter der Gruppe — es
möge die Larve von Limnophilus rhombicus[XLI] dazu dienen —
die biologischen Verhältnisse und die damit in Beziehung stehenden
morphologischen Eigenschaften zu schildern versuchen.
[XL] Über die in Gehäusen lebenden Wasserraupen, d. h. Larven
von Schmetterlingen, vergleiche man oben den dritten Abschnitt, über
Phryganiden-Larven ohne Gehäuse weiter unten.
[XLI] Dass die betreffende Larve zu Limnophilus
rhombicus gehört, habe ich freilich nicht durch Zucht festgestellt,
sondern ich schliesse es nur aus dem Vergleich mit den Zeichnungen und
Angaben bei Pictet (Fig. 8, 16, 17).
Unsere Larve lebt besonders in Pfühlen und Gräben mit stehendem
Wasser. Das Gehäuse (Fig. 8, 16 u. 17) besteht in der Regel aus
dünnen Stengelstücken, die quer zur Längsachse verklebt sind, und
zwar schliessen je vier bis fünf einmal den Umfang; im Querschnitt
ist also die Aussenseite des Futterals vier- bis fünfeckig, wobei
die Seiten noch über die Ecken verlängert sind. Nicht selten benutzt
die Larve daneben auch Moosstücke oder Früchte oder Steinchen und
Schneckenhäuser, die Queranordnung jedoch, soweit möglich, beibehaltend.
Der Kopf und die beiden ersten Brustringe oben sind ebenso wie die
Beine, von denen das erste Paar besonders kräftig, wenn auch kurz ist,
stark hornig; diese Teile sind nämlich beim Fressen und Umherkriechen
mehr oder minder ausserhalb des Futterals, dürfen also nicht zart sein.
Über die helleren und dunkleren Flecke dieser Teile verweise ich auf
die Abbildung (Fig. 8, 15). Der dritte Brustring hat auf der Oberseite
sechs dunkle, mit Haaren besetzte Hornflecke und ebenso ist die Haut
oberhalb der Hüften stärker chitinisiert. Die Mundteile, besonders
auch die mit der[S. 96] Unterlippe verwachsenen Unterkiefer ähneln denen der
echten Raupen; auf die besondere Gestalt dieser Teile bei unserer Art
ohne Abbildung näher einzugehen — für die Unterscheidung der Gattungen
ist das wichtig — erscheint zwecklos. Wie bei den Raupen liegt
übrigens der Ausgang der Spinndrüse in der Mitte der Unterlippe. Fühler
fehlen wie bei fast allen Phryganidenlarven vollständig und die Augen
sind nur winzige Punktaugen.
An dem zarten, gelbweissen Hinterleib hebt sich in der Mitte als
dunklerer Streifen das Rückengefäss ab, dessen von hinten nach vorn
fortschreitende Kontraktionen schon mit unbewaffnetem Auge zu verfolgen
sind. An den Seiten zieht sich je eine dichte Reihe dunkler Haare
hin, die sogenannte Seitenlinie. Teils unmittelbar über und unter
derselben, teils etwas weiter nach oben und unten stehen vom zweiten
Hinterleibsringe an und zwar nahe dem vordern und hintern Rande die
haarförmigen, weissglänzenden Tracheenkiemen (s. Fig. 8, 15). Um die
Zahl und Verteilung derselben kurz darzustellen, hat sich meines
Wissens Klapalek zuerst besonderer Schemata bedient; ich gebe
hier ein nach seiner Weise gebildetes Schema für unsere Larve.
Oben.Sl. Unten.
3
3
3
3
3
3
II
3
3
3
3
3
3
III
3
2
3
3
2
3
IV
3
2
3
2
1
3
V
2
3
2
2
VI
2
2
2
1
VII
2
1
VIII
Zum Verständnis desselben braucht wohl nur bemerkt zu werden, dass
Sl. die Seitenlinie, die römischen Ziffern die Hinterleibsringe
und[S. 97] die arabischen Ziffern die Anzahl der Kiemenfäden bedeuten, welche
teils nahe dem Vorder-, teils nahe dem Hinterrande, und zwar nahe
der Seitenlinie oder weiter nach oben oder unten stehen. Am ersten
Hinterleibsringe stehen keine Tracheenkiemen, dagegen in der Mitte
oben und an jeder Seite je ein vorstülpbarer Fleischzapfen; am letzten
Hinterleibsring befinden sich seitlich zwei kurze derbe Chitinhaken und
in der Mitte in einem länglichen Spalt der After. Mit den Chitinhaken
des Hinterleibsendes halten sich die Tiere recht erfolgreich im
Gehäuse fest. Erwachsen kriechen die Larven langsam im Wasser umher;
jugendliche Phryganiden-Larven habe ich fast immer — ob freilich
solche unserer Art dabei waren, ist ungewiss — daneben lebhaft mit
Hilfe der langen, stark bewimperten Hinterbeine umherschwimmen sehen.
Solche jugendliche, wohl eben dem Ei entschlüpfte Larven sind übrigens
von den älteren noch durch den Mangel an Tracheenkiemen unterschieden,
so dass sie ganz auf Hautatmung angewiesen sind.
Dass die Atmung bei den älteren Larven durch Tracheenkiemen und
daneben wohl durch die zarte Hinterleibshaut geschieht, ist oben schon
angedeutet worden. In Beziehung zur Atmung dürfte auch die sogenannte
Seitenlinie, d. h. der beiderseitige Wimpersaum des Hinterleibes
stehen, wenigstens wenn sie stärker entwickelt ist. Bei durchsichtigen
Phryganiden-Gehäusen sieht man nämlich oft, besonders wenn das Wasser
sauerstoffarm ist, die Larve den Hinterleib in der Mittelebene
schwingen, höchst wahrscheinlich, um das den Körper umspülende
Wasser schneller zu erneuern. Die Seitenlinie macht nun, indem sie
den Hinterleib verbreitert, dieses Verfahren sicher wirksamer. Auch
die Fleischzapfen des ersten Hinterleibsringes finden dabei ihre
Verwendung; der Körper wird durch sie mehr in der Mitte des Gehäuses
gehalten, ein allseitig den Körper umspülender Wasserstrom also wohl
dadurch erleichtert. Wenn übrigens das Wasser recht schlecht zur Atmung
wird, so sieht man die Larven vorn weit aus dem Gehäuse kommen und nun
lebhaft in gleicher Richtung wie im Gehäuse das Wasser schlagen.
[S. 98]
Dass die Phryganiden-Larven wie die Raupen vorherrschend
Pflanzenfresser sind, kann man leicht beobachten; doch lässt sich auch
unschwer feststellen, dass sie keineswegs Fleischkost, lebende und tote
Tiere, deren sie habhaft werden können, verschmähen. Hat man einer
Larve das Gehäuse genommen, so ist man nicht sicher, ob dieselbe nicht
in kurzer Zeit von ihres Gleichen angefressen wird.
Zur Verpuppung spinnt unsere Larve wie die meisten anderen
Phryganiden-Larven das Gehäuse an Wasserpflanzen oder Steinen etc.
fest, schliesst die Öffnungen durch ein Gitterwerk von Fäden, denen
noch Stengelstückchen angeklebt sind, so dass einerseits Feinde dadurch
abgehalten werden und anderseits das Atemwasser hindurchspülen kann.
Die Puppe (Fig. 8, 18) bietet mit ihren frei abstehenden Fühlern,
Beinen und Flügeln, mit den stark auffälligen Augen und gekreuzten,
hakenförmigen Oberkiefern, mit den weissen, fädigen Tracheenkiemen und
der schwarzen, mächtig entwickelten Seitenlinie am Hinterleib einen
seltsamen Anblick dar. Die nach dem Geschlecht verschiedene Gestaltung
der Hinterleibsanhänge und die Zahl und Verteilung der Dorne an den
Beinen geben zusammen mit dem Kiemenschema wichtige Anhaltspunkte zur
Unterscheidung der Gattungen und Arten. Dass auch hier die Seitenlinie
gleiche Bedeutung für die Atmung hat wie die oben beschriebene, scheint
mir zweifellos. Die Atmungsbewegungen der Puppen kann man unschwer
beobachten.
Hält man die Puppen im Zimmer, so steht Einem unmittelbar vor
dem Ausschlüpfen der Köcherfliege noch eine seltsame Beobachtung
bevor. Die Puppen kommen eines Tages aus dem Gehäuse, indem sie die
Oberkiefer zum öffnen benutzen, und schwimmen mit dem bewimperten
zweiten Beinpaar oder kriechen mit Hilfe der beiden vorderen Beinpaare
lebhaft umher, bis sie eine geeignete Stelle an der Oberfläche für
das Ausschlüpfen gefunden haben. Die ausgeschlüpften Insekten, denen
die Oberkiefer der Puppen fehlen, sehen so schmetterlingsartig und im
besondern mottenartig aus, dass sie von Laien wohl stets für[S. 99] Motten
gehalten werden. Das Fehlen des Saugrüssels, die Aderung der Flügel
und die Faltung der Hinterflügel können zur Unterscheidung von den
Schmetterlingen dienen.
Der Laich der Phryganiden — den unserer Art im besondern kenne ich
nicht — bildet meist eine grosse, oft ringförmige Schleimmasse, in der
die Eier eingebettet liegen; leicht kann derselbe mit Schneckenlaich
verwechselt werden. —
Ist diese Larvenform der Köcherfliegen auch die unbedingt
vorherrschende im Tieflande, so möchte ich doch nicht ganz den
Vertreter des zweiten unter den Phryganiden-Larven vorhandenen
Typus übergehen, welchen ich um Berlin mehrfach, besonders auch
an Stratiotes, angetroffen habe. Nach Vergleich mit den
Abbildungen und Angaben von Klapalek gehört diese Larve zur
Gattung Polycentropus (Unterfamilie Rhyacophilidae).
Der auffälligste Unterschied der ziemlich durchsichtigen, etwas
grünlich und rötlich gefärbten Larve gegen die des ersten Typus bildet
das Fehlen eines Gehäuses und der Tracheenkiemen. Die Larve ist ein
Hautatmer[XLII].
[XLII] Es ist bemerkenswert, dass keine Neuropteren-Larve des
Wassers durch Luftlöcher atmet, während das in allen früheren Ordnungen
ein recht häufiger Fall war.
Es fehlen auch die Seitenlinie und die Fleischzapfen des ersten
Hinterleibsringes, was nach unserer obigen Deutung beim Fehlen
des Gehäuses ohne weiteres verständlich ist. Die Chitinhaken
am Unterleibsende stehen dagegen auf langen, zweigliedrigen
„Nachschiebern“; die Larven gebrauchen dieselben besonders, wenn
sie beunruhigt werden und sich dann unter medianen Schwingungen des
Hinterleibes rückwärts in Sicherheit zu bringen suchen[XLIII].
[XLIII] Eine derartige Rückwärtsbewegung der Larven, sobald
sie beunruhigt werden, scheint im ganzen Gebiet der Netzflügler,
gleichgültig ob die Larven im Wasser leben oder nicht, die allgemeine
Regel zu sein.
Mangelt den Larven auch das Gehäuse, so heften sie doch mit Hilfe der
Spinnmasse an Blättern einzelne Gegenstände derart fest, dass ein
gedeckter Gang entsteht, der ihnen als Zufluchtsort dient. Zur Zeit der
Verpuppung wird dann eine festere Schutzhülle hergestellt. Die Puppe
ist mit Tracheenkiemen und Seitenlinie versehen.
[S. 100]
Eine Mittelstellung zwischen den beiden obigen Typen nehmen die
Larven der Gattung Hydropsyche ein, die in stärker fliessenden
Gewässern, z. B. hier in der Panke, an und unter den Steinen des
Bodens leben. Sie haben ebenfalls kein Gehäuse, welches sie mit
sich herumtragen, sondern fertigen sich aus grösseren und kleineren
Sandkörnern Gänge an den Steinen, dagegen besitzen sie büschelförmige
Tracheenkiemen. Abweichend von der oben beschriebenen Verteilung
der Kiemen bei Limnophilus, stehen dieselben hier nur auf
der Unterseite von Brust und Hinterleib. Die Nachschieber sind
von ähnlicher Länge wie bei Polycentropus und durch ein
starkes, auffälliges Borstenbüschel an der Ansatzstelle der Krallen
ausgezeichnet.
Zur Verpuppung wird auch von diesen Larven ein besonderes Schutzgehäuse
hergestellt.
Die übrigen Neuropteren.
In den übrigen Familien der Netzflügler giebt es nur noch vereinzelte
Gattungen, deren Larven im Wasser leben, nämlich die Gattungen
Sialis, Sisyra und Osmylus. Häufiger habe ich
davon nur die Sialis-Larven angetroffen und zwar stets am Boden
stehender, mit Schlammgrund versehener Gewässer. Man muss schon mit
dem Netz etwas Schlamm mitfassen, wenn man die Larven fangen will.
Die Ähnlichkeit mit den Phryganiden-Larven ist nicht zu verkennen.
Nur der Kopf, die Beine und die Brustringe sind stärker chitinisiert,
der Hinterleib ist zarthäutig und an den sieben ersten Ringen mit
je einem Paar verhältnismässig starker, gefiederter und
gegliederter Tracheenkiemenfäden versehen. Das Ende des
Hinterleibes läuft in eine einzelne kräftige, gefiederte Endborste aus.
Ausser der abweichenden Bildung der Tracheenkiemen und des
Hinterleibsendes kann zur Unterscheidung von den Phryganiden-Larven
auch der Umstand dienen, dass der Kopf deutliche, mehrgliedrige
Fühler trägt. Die Mundteile, besonders die Oberkiefer, sind zum
Raube eingerichtet. Als Kiemenatmer brauchen die Larven nicht an die
Oberfläche zu kommen; auf dem Schlammboden kriechen sie nach Beute
umher. Beunruhigt schwimmen sie ganz nach Art[S. 101] der Phryganiden-Larven,
die ihres Gehäuses beraubt wurden, durch mediane Schwingungen des
Körpers rückwärts.
Zur Verpuppung gehen die Larven wie die Käferlarven an das Ufer;
die Puppe ruht dann dort im Moose. Das ausgeschlüpfte Insekt, die
„Schlammfliege“, findet man im Mai und Juni in der Nähe von Gewässern
an Pflanzen träge ruhend, leicht kenntlich an den auch mit zahlreichen
Queradern versehenen, eigentümlich grauen Flügeln. Die schwärzlichen
Eier werden in regelmässigen dichten Reihen an Pflanzen oberhalb des
Wassers abgelegt. —
Die kleinen, etwa 4 mm langen Larven von Sisyra leben in
Süsswasserschwämmen. Ausgezeichnet sind dieselben durch vielgliedrige
Fühler und sehr lange, gebogene, aus Ober- und Unterkiefer
zusammengesetzte Saugzangen. Fühler wie Saugzangen übertreffen den
Kopf um das Mehrfache an Länge. Die Hinterleibsringe tragen auf der
Unterseite je ein Paar gegliederter Tracheenkiemen und auf der
Rückseite je vier mit Borsten versehene Auswüchse.
Die Larven des Bachameisenlöwen (Osmylus) lieben stark
strömendes Wasser; in Gebirgsbächen unter Steinen ist nach den
Litteraturangaben ihr Aufenthalt. Ich habe sie noch nicht angetroffen.
Wie bei den Netzflüglern fast die ganze Familie der Köcherfliegen[XLIV]
in ihren Larven dem Wasser angehört, so unter den Geradflüglern die
drei Familien der Wasserjungfern (Odonaten oder Libelluliden), der
Eintagsfliegen (Ephemeriden) und Afterfrühlingsfliegen (Perliden). Mit
einigen anderen Geradflüglern wurden sie früher ihrer Flügelbildung
wegen der vorigen Ordnung zugezählt und werden bisweilen heute noch
Pseudoneuroptera genannt. Im Larvenheer der Gewässer nehmen ihre
Larven einen hervorragenden Platz ein.
[XLIV] Nur die Larven der Gattung Enoicyla sind davon
ausgenommen. Dieselben leben zwischen Moos.
Fig. 9.
19 Larve von Aeschna, die Unterlippe Ul nach Cloëon
vorschnellend — 20 Larve von Libellula — 21 Larve von Agrion — 22
Larve von Calopteryx — 23 Larve von Cloëon dipterum. 19–22 nach
der Natur; Haltung von 19 und 23 nach Rösel, 23 nach Eaton.
Die Zugehörigkeit einer Wasserlarve zu einer dieser drei Familien ist
sichergestellt, wenn sie neben beissenden Mundteilen und drei Paar
grosser, meist kräftiger Brustbeine an Mittel- und Hinterbrust[S. 102] meist
recht deutliche Flügelansätze (Fig. 9, 19–23, Fig. 10, 24) besitzt[XLV].
Unsere Tiere machen nämlich kein Puppenstadium durch, d. h. ein
Ruhestadium, in dem weder Nahrung aufgenommen wird[S. 103] noch lebhaftere
Ortsbewegung stattfindet und in dem der grosse Entwickelungsschritt von
der Larve zum vollkommenen Insekt geschieht; solche Larven erhalten die
Flügel schrittweise mit den verschiedenen Häutungen.
[XLV] Nur den jüngsten Larven fehlen diese Flügelansätze. Die
Übereinstimmung in den anderen Punkten mit den ungleich häufigeren
älteren Stadien lässt sie trotzdem unschwer erkennen.
Die Larven der Libelluliden und Ephemeriden finden sich sowohl in
stehendem wie fliessendem Wasser, die der Perliden habe ich nur in
fliessendem Wasser angetroffen. Die Larven der Libellen sind unter
den anderen Larven der Ordnung leicht kenntlich an der armartig
vorstreckbaren Unterlippe (Fig. 9, 19), die der Ephemeriden an den
blatt- und fadenförmigen Tracheenkiemen, die nur an den Seiten des
Hinterleibes stehen, und an den drei Afterborsten (Fig. 9, 23). Die
Larven der Perliden (Fig. 10, 24) besitzen keines dieser besonderen
Kennzeichen; besitzen sie Tracheenkiemen, so stehen sie meist auf der
Unterseite der Brustringe.
1. Die Larven der Libelluliden oder
Odonaten.
Unter den Libellenlarven der stehenden Gewässer treten dem Beobachter
ohne weiteres drei Typen entgegen, die der Agrion-,
Libellula- und Aeschna-Gruppe. Die ersteren (Fig. 9,
21) sind langgestreckt und schwächlich; am Ende des Hinterleibes mit
drei ziemlich langen blattartigen Tracheenkiemen versehen. Die hierher
gehörigen Agrion- und Lestes-Larven unterscheiden
sich dadurch, dass der Mittelteil der Unterlippe bei Lestes
fast stielrund, dagegen bei Agrion fast ebenso breit wie
der Endabschnitt ist. Die Larven der Libellula-Gruppe (Fig.
9, 20) sind ohne solche äussere Tracheenkiemen, dafür besitzen sie
aber eigenartige innere Tracheenkiemen, nämlich an den Wandungen des
Enddarmes. Die Hinterbeine sind länger als der gedrungene Hinterleib
und der Vorderabschnitt der Unterlippe ist so ausgehöhlt, dass er
in der Ruhelage die Mundteile von vorn wie eine hohle Hand bedeckt.
Dieser Gruppe gehören auch die Larven der Gattungen Cordulia
und Epitheca an. Die Larven endlich der Aeschna-Gruppe
(Fig. 9, 19) stimmen in den Atemorganen mit denen von Libellula
überein, dagegen ist hier die Unterlippe im Vorderabschnitt einfach
flach und der schlanke Hinterleib ist länger als die Hinterbeine.
[S. 104]
Alle unsere Larven sind also Tracheenkiemen-Atmer, aber in
verschiedener Weise. Die Larven der beiden letzten Typen ziehen
willkürlich das Atemwasser durch den mit Schutzwehren versehenen After
ein und stossen es wieder aus. Diese Atembewegungen werden kräftig,
aber unregelmässig ausgeführt; oft sieht man minutenlang umsonst nach
dem ein- und austretenden Wasserstrom. Agrion- und Lestes-Larven
besitzen natürlich derartige Atembewegungen nicht, dagegen sieht
man nicht selten, dass sie, mit den Beinen sich festhaltend, durch
seitliches Schlagen des Hinterleibes das Atemwasser erneuern. Eine
recht eigentümliche Thatsache bei allen diesen Larven ist nun, dass an
der Brust Luftlöcher vorhanden sind, deren Ränder für gewöhnlich dicht
aneinander liegen, die aber geöffnet werden können. Beweis dafür ist
die Erscheinung, dass aus diesen Stigmen, wenn man die Larven[XLVI] in
Alkohol oder heisses Wasser wirft, eine Reihe von Luftblasen aufsteigen.
[XLVI] Nur bei den jüngsten Larven tritt das nicht ein. —
Dass die Stigmen nicht verwachsene Ränder haben, stellte ich auch an
Querschnitten derselben fest.
Welche Bedeutung können nur diese Luftlöcher haben? Ich vermöchte nur
ungeprüfte Vermutungen als Antwort auf diese Frage zu geben.
Bewegung. Bei allen unseren Larven dienen die Atembewegungen
zugleich zum Schwimmen. Lange Zeit freilich kriechen oft die Larven am
Boden oder an Wasserpflanzen umher. Wollen Agrioniden-Larven schwimmen,
so treiben sie sich durch seitliches Schlagen des Hinterleibes mit
seinen Kiemenblättern vorwärts. Seltsamer aber nimmt sich das Schwimmen
der anderen Larven aus: mit angelegten Beinen sieht man sie plötzlich
durch das Wasser schiessen, ohne dass man zunächst zu erkennen vermag,
wie das bewirkt wird. Genauere Beobachtung zeigt — ein sandiger
Grund leistet dabei gute Dienste —, dass die Larven mit grosser Kraft
das Wasser aus dem After ausstossen und dadurch nach dem Prinzip
des Rückstosses vorwärts schiessen. Hebt man die Larve im rechten
Augenblick ausser Wasser, so kann man das ausgestossene Wasser oft drei
bis vier Spannen weit fliegen sehen.
[S. 105]
Der Ernährung nach sind alle diese Larven Raubtiere und zwar erbeuten
sie die Tiere durch plötzliches Vorstrecken der Unterlippe. Bietet man
einer Aeschna-Larve z. B. Ephemeriden- und Mückenlarven, so zeigt sich
zugleich dabei, dass die Beute erst wahrgenommen wird, wenn sie sich
zu bewegen beginnt: eine Bemerkung, die für fast alle Räuber unter den
Wasserinsekten und Wasserlarven zu gelten scheint.
Andere Libelluliden-Larven. Von anderen Libelluliden-Larven, die
ich nur in fliessenden Gewässern, nicht wie die obigen in beiderlei
Gewässern, angetroffen habe, nenne ich noch zur Ergänzung die von
Calopteryx (Fig. 9, 22), der prachtvoll dunkelblauen oder
grünen Libelle der Ufer, und von Gomphus. Die ersteren bilden
einen eigenartigen Typus dadurch, dass sie sowohl Schwanzkiemen wie
innere Kiemen haben. Kenntlich sind sie vor allem dadurch, dass sie
bei ähnlicher Gestalt wie die Agrioniden-Larven Fühler besitzen, die
weit länger als der Kopf sind und deren Grundglied mehrfach länger und
stärker als die übrigen Glieder zusammengenommen ist. — Die Larven von
Gomphus schliessen sich am nächsten denen von Aeschna an;
die Unterlippe ist vorn ebenfalls flach, der Darm mit Kiemen versehen;
das unterscheidende Merkmal liegt vornehmlich in dem auffallend breiten
und grossen Endglied der Fühler.
2. Ephemeriden-Larven.
Die Larven der Eintagsfliegen (Fig. 9, 23) sind in stehenden wie
fliessenden Gewässern eine recht häufige Erscheinung; aber fast immer
gehörten die Tiere, welche ich um Berlin in stehenden Gewässern fing,
einer Art an: Cloëon dipterum. Erwachsen sind dieselben,
die drei langen, befiederten Schwanzanhänge nicht mitgerechnet,
etwa 1 cm lang. Die drei letzten Hinterleibsringe sind ohne
Kiemenblättchen, das viertletzte hat ein einfaches rundliches
Blättchen, die sechs vorhergehenden Ringe aber je zwei solcher
Kiemenblätter. Unschwer erhält man das Insekt selbst, welches nach der
Zweizahl der Flügel und nach den Schwanzborsten leicht zu bestimmen
ist. Die den Ephemeriden einzig zukommende[S. 106] Eigentümlichkeit, dass das
der Larvenhaut entschlüpfte, mit beweglichen Flügeln versehene Insekt
(Subimago genannt) sich noch einmal häutet, beobachtete ich
nicht immer, anscheinend fehlte den Tieren dazu häufig die Kraft; sie
starben vorher ab.
Durch den Besitz der blattartigen Tracheenkiemen ist die Atemart
unserer Larven hinreichend bezeichnet; zu erwähnen bleibt nur, dass
dieselben zur Erneuerung des Wassers oft schnell hintereinander auf-
und niederbewegt werden, ohne dass sich etwa das Tier vom Platze
bewegt. Für gewöhnlich gehen die Larven mit den gut entwickelten
Beinen am Boden einher; bei der geringsten Beunruhigung jedoch fahren
sie jäh durch das Wasser dahin. Durch kräftige mediane Bewegungen des
Hinterleibes mit seinen befiederten Anhängen schnellen sie sich fort.
Ihre Nahrung besteht aus Pflanzenkost; wie erfolgreich sie dieselbe
verarbeiten, zeigt sich z. B. darin, dass eine Schale mit Cloëon-Larven
und Pflanzen sich am Boden bald mit den Resten guter, gesegneter
Verdauung füllt.
Ausser den Cloëon-Larven habe ich in kleineren stehenden Gewässern nur
noch hie und da die von Caenis luctuosa angetroffen. Dieselben
sind leicht daran zu erkennen, dass das erste Paar Kiemenblättchen zu
Schutzdecken für die folgenden umgewandelt ist und dass die letzteren
blattartig, aber am Rande mit zarten Fortsätzen versehen sind. Die vier
letzten Hinterleibsringe sind von jenen Schutzplatten nicht bedeckt.
Ungleich reicher als in stehenden Gewässern sind die Ephemeriden-Larven
in fliessenden entwickelt; reicher und mannigfaltiger gestalten sich
dort auch die Lebensbedingungen derselben. Manche von ihnen (z. B.
Ephemera und Palingenia) graben sich mit den kräftigen
Vorderbeinen Gänge in das Ufer; andere wieder, wie die der alten
Gattung Baëtis, drücken sich bei eigenartig platter Gestalt
dicht den Steinen an und vermögen sich so im Strome zu halten; noch
andere kriechen, wohl an ruhigeren Stellen, auf alles Andere Verzicht
leistend, im Schlamme umher[XLVII]. Recht mannigfach ändert dabei auch die
Gestalt der Tracheenkiemen ab.[S. 107] Näher hierauf einzugehen, muss ich mir
schon aus äusseren Gründen versagen[5].
[XLVII] Die Cloëon-Larven würden zu denen gehören, welche
Pictet als Schwimmer bezeichnet.
3. Larven der Perliden oder
Afterfrühlingsfliegen.
Die Larven der Perliden scheinen stehende Gewässer ganz zu meiden; auch
schwächer fliessende sind arm an ihnen, wogegen reissende Gebirgsbäche
ihre rechte Heimat sind.
Die einzige Larve, welche ich in schwach fliessenden Gräben um Berlin
angetroffen habe, ist die von Nemura variegata (Fig. 10, 24 S.
109). Als Orthopteren-Larve ist sie, wenigstens in den älteren Stadien,
an den Flügelansätzen des zweiten und dritten Brustringes kenntlich;
von den Ephemeriden-Larven, mit denen sie noch am ersten verwechselt
werden könnte, unterscheidet sie sich erstens durch den Mangel an
Tracheenkiemen und zweitens dadurch, dass sie nicht drei, sondern nur
zwei lange Anhänge am Hinterleibsende hat. Innerhalb der Familie gehört
die Larve zu denen, bei welchen das zweite Glied des dreigliedrigen
Fusses kleiner ist als das erste.
Die Larve besitzt keine Tracheenkiemen; wie atmet sie denn? Ich habe
sie weder jemals zur Atmung an die Oberfläche kommen sehen, noch
spricht die Derbheit ihrer Haut und ihre Grösse für Hautatmung, so dass
mir die Atemverhältnisse der Larve rätselhaft sind.
Eigenartig nimmt sich die Bewegung der Larve aus. Bei keiner bisher
genannten Larve sind die Beine so kräftig und lang zugleich, wird der
Hinterleib so wenig auf dem Boden nachgeschleppt, ist die Bewegung
ein so flinkes Gehen wie bei dieser Larve. Ihre Lebensweise ist eine
räuberische.
Die Ähnlichkeit zwischen der älteren Larve und dem entwickelten Insekt
ist recht auffallend; der Schritt von dem einen zum andern Zustand
scheint deswegen auch recht leicht und schnell zu geschehen. Brachte
ich Larven in feuchter Schachtel mit Moos zusammen heim, so fand ich am
nächsten Tage meist mehrere davon schon ausgeschlüpft.
Von den übrigen Larven der Familie, die sich alle einander recht
ähnlich sehen, möchte ich nur noch die grösseren Perla-Larven[S. 108]
erwähnen, die durch Tracheenkiemen an der Unterseite der Brust
ausgezeichnet sind[XLVIII].
[XLVIII] Zuerst bei Tieren dieser Familie wurde die überraschende
Thatsache festgestellt, dass auch entwickelte Kerfe noch Tracheenkiemen
besitzen. Palmén hat dann diese Erscheinung als allgemeiner
verbreitet nachgewiesen.
Fig. 10. 24 Larve von Nemura variegata — 25Notonecta glauca,
von der Bauchseite — 26Corisa striata — 27Naucoris
cimicoides — 28Ranatra linearis — 29 Binsenstück mit
zahlreichen Eiern von Ranatra — 30 Ein einzelnes Ei von
Ranatra, vergrössert.
Schlussbemerkung. Sowohl am Anfang dieser wie der vorigen
Ordnung hob ich hervor, dass die Anzahl der Wasserlarven innerhalb
der Ordnung ungleich grösser als bei den früheren ist. Dazu kommt
nun noch ein Anderes. Bei den Larven der Käfer, Zweiflügler und
Schmetterlinge war die Anpassung an das Wasserleben, besonders
hinsichtlich der Atmung, eine recht verschiedene; eine Reihe von Arten
atmete noch durch Luftlöcher, daneben fand sich mehr oder minder
ausgedehnt Atmung durch die Haut, echte Kiemen oder Tracheenkiemen.
Ungleich einheitlicher, geschlossener erscheinen darin die Larven
der Neuropteren und amphibischen Orthopteren: fast alle sind sie
Tracheenkiemenatmer, selten finden sich daneben Hautatmer und nie
einfache Luftatmer. Ferner sind sie einerseits alle nach ihrer
Organisation ausgesprochene Wassertiere, und anderseits werden die
einzelnen Gruppen den verschiedenartigen Bedingungen des Wasserlebens
in oft recht eigenartiger Weise gerecht. Das Alles legt die Frage nahe:
Ist das Verhältnis dieser Larven zum Wasserleben nicht ein ganz anderes
als das der Käfer-, Zweiflügler-, Schmetterlings- und — wie wir gleich
hinzufügen wollen — Wanzen-Larven? Diese verglichen wir den Seehunden
und Walen, d. h. wir hielten für recht wahrscheinlich, dass sie von
Vorfahren abstammen, die samt ihren Larven in der Luft lebten; sind
jene Larven dagegen nicht vielleicht echte, ursprüngliche Wassertiere.
Besonders für die Ephemeriden ist wegen der unvollkommenen Verwandlung,
wegen der einfachen Bildung der Mundteile und weil sich die Bildung
der Flügel bei ihrem ersten Auftreten mit der der Tracheenkiemen
vergleichen lässt, schon öfters ausgesprochen worden, dass sie dem
Urtypus der Insekten ziemlich nahe stehen dürften; leiten sie sich
vielleicht unmittelbar von einer im Wasser lebenden Stammform ab? Sind
sie also den Amphibien im Kreise der Wirbeltiere zu[S. 109] vergleichen?
Diese Frage näher zu erwägen[XLIX], ist hier nicht der Ort, doch möchte
ich hier kurz auf den einen Punkt hinweisen,[S. 110] dass man im Falle der
Bejahung annehmen müsste, die Bildung eines Tracheensystems und zwar
eines geschlossenen, das für die Bewegung der Luft im Körper so
ungünstige Bedingungen bietet, sei bei Tieren, die unter Wasser leben,
an Stelle der Kiemen entstanden!
[XLIX] Verwiesen sei hier besonders auf die für die Frage
wichtigen Untersuchungen und Erörterungen von Palmén
(„Morphologie des Tracheensystems“).
Die Schnabelkerfe oder Rhynchoten lassen sich hinsichtlich ihres
Verhaltens zum Wasser recht wohl mit den Käfern vergleichen; wie bei
diesen leben unter den Schnabelkerfen nicht nur eine Reihe von Larven
im Wasser, sondern auch entwickelte Kerfe, die in beiden Ordnungen
freilich nicht völlig an diesen Aufenthalt gebannt sind. Ferner gilt
auch in dieser Ordnung wie in jener[L] der Satz, dass, wenn das
entwickelte Tier diesem Element angehört, auch die Larve im Wasser
lebt. Endlich stellen sich, wie schon bei den Käfern erwähnt wurde,
beide Ordnungen auch darin neben einander, dass einige ihrer Arten
auf der Wasseroberfläche leben: neben den sich auf der Oberfläche
herumtummelnden Gyriniden finden sich die nicht minder auffälligen
Wasserläufer, welche innerhalb der Ordnung eine besondere kleine
Familie bilden.
Am häufigsten trifft man auf stehenden Gewässern die Arten der Gattung
Hydrometra an. Der Körper ist ebenso wie die beiden hinteren
Beinpaare auffallend lang und dünn, die Vorderbeine, zwischen denen
der ziemlich lange, mit Stechborsten versehene Rüssel liegt, ungleich
kürzer. Eigenartig ist der Eindruck, den ihr Stehen und ihre Bewegung
auf dem Wasser macht. Mit den wagerecht gestreckten Fussgliedern der
Mittelbeine und zeitweise der Vorderbeine, sowie den Schienen und
Fussgliedern der Hinterbeine ruhen sie auf der Wasseroberfläche, die
an der betreffenden Stelle eingedrückt erscheint, wie bei einer auf
der Oberfläche schwimmenden Nähnadel. Durch ein feines, lufthaltiges
Haarkleid der Beine ist die Gefahr des Einsinkens gemindert. Bei der[S. 111]
Bewegung gebrauchen die Tiere vornehmlich die Mittelbeine, dieselben
gleichzeitig bewegend; stossweise gleiten sie auf der Oberfläche
ziemlich schnell dahin[LI].
[LI] Die Gattung Velia, welche schattige, fliessende
Gewässer liebt, schreitet oder läuft dagegen auf dem Wasser.
Die Nahrung der Hydrometriden besteht aus schwächeren Insekten,
besonders wohl auch aus eben ausgeschlüpften Mücken, die sie mit ihrem
Stech- und Saugrüssel aussaugen. An den Larven fällt besonders die ganz
ausserordentliche Verkürzung des Hinterleibes[LII] auf; hiervon und von
dem Mangel ausgebildeter Flügel abgesehen, gleichen sie, besonders auch
in der Lebensweise, sehr den entwickelten Kerfen.
[LII] Sonst ist bei Insektenlarven der Hinterleib der am
kräftigsten entwickelte Körperteil. Das ist nach der Lebensaufgabe des
Larvenstadiums ohne Weiteres verständlich. Hier wird die Abweichung von
der Regel durch die Bewegungsweise auf der Oberfläche und die damit
verbundenen Anforderungen bedingt.
Die übrigen Schnabelkerfe nun, welche dem Wasser angehören, führen ein
Leben ähnlich dem der Dytisciden und Hydrophiliden; einige von ihnen,
nämlich die Gattungen Notonecta (Fig. 10, 25), Plea,
Corisa (Fig. 10, 26), Naucoris (Fig. 10, 27), sind
lebhafte Schwimmer, andere dagegen, Nepa und Ranatra
(Fig. 10, 28), kriechen mehr den Hydrophiliden ähnlich an den
Wasserpflanzen umher. Alle aber kommen, den Käfern gleich, zum Atmen an
die Oberfläche, im einzelnen dabei freilich verschiedene Einrichtungen
und Weisen zeigend. Systematisch sind sie übrigens auch dadurch
zusammengehalten, dass sie neben dem gegliederten Saugrüssel und den
beiden ungleichartig ausgebildeten Flügelpaaren ausserordentlich kurze
Fühler besitzen, die sie verborgen am Unterkopf tragen.
Unter den Schwimmern ist der ansehnlichste und mit auch der
häufigste Notonecta glauca, in kennzeichnender Weise
Rückenschwimmer genannt. Fassen wir ihn, wenn er aus dem Netz
genommen werden soll, nicht vorsichtig, so lernen wir wahrscheinlich
auch durch seinen empfindlichen Stich verstehen, weshalb er hie und da
Wasserbiene heisst.
[S. 112]
Die Eigentümlichkeit, mit dem Bauch nach oben zu schwimmen[LIII],
kennzeichnet unser Insekt hinreichend. In Beziehung zu dieser
Schwimmart steht auch die Färbung des Tieres: nicht der Rücken ist, wie
das bei den meisten Wasserinsekten die Regel, dunkel gefärbt, sondern
der Bauch, während unter Wasser die Rückseite durch anhaftende Luft
lebhaft weiss erscheint. Nachdrücklich wird hierdurch in ähnlicher
Weise wie durch die Färbung der Flundern die Auffassung gestützt, dass
die dunklere, dem Boden ähnliche Färbung der Rückseite und die lichtere
der Bauchseite so vieler Wassertiere, z. B. so vieler Fische, eine
Schutzfärbung ist.
[LIII] Nur bei der kleinen, 2 mm grossen Verwandten,
Plea minutissima, findet sich die gleiche Schwimmart.
Als Ruder sind nur die stark verlängerten kräftigen Hinterbeine
thätig, die an Schienen und Fussgliedern mit zwei Reihen Schwimmhaaren
versehen sind; ihre Endkrallen sind verkümmert. Als ordentlicher
Schwimmer gebraucht das Tier die Ruder gleichzeitig. Wie es Vorder- und
Mittelbeine verwendet, sieht man leicht, wenn man auf die Oberfläche
etwa eine Fliege wirft. Meist nimmt der vornehmlich nach oben schauende
Rückenschwimmer die Beute schnell wahr, schiesst jählings darauf los
und packt sie mit armartig gekrümmten Vorder- und Mittelbeinen, um
schwebend sie auszusaugen.
Zur Atmung streckt er, den Schwimmkäfern ähnlich, das Hinterleibsende
aus dem Wasser; man möchte also hier auch grosse und ansehnliche
Luftlöcher vermuten. Die Luftlöcher des Hinterleibes jedoch sind
auffallend klein; grosse, mit zarten Schutzhaaren versehene Luftlöcher,
welche fraglos für die Atmung in erster Linie in Betracht kommen,
liegen dagegen ziemlich verborgen seitlich an der Brust, etwas nach
hinten und bauchwärts von der Ansatzstelle der Hinterflügel, ferner in
der Verbindungshaut der Vorder- und Mittelbrust, sowie zwischen Mittel
und Hinterbrust und zwar am Rande der Unterseite. Die Luft aber wird
von dem Hinterleibsende nach diesen Stigmen in eigentümlicher Weise
geleitet. Es ist nämlich der Bauch in der Mitte gekielt (Fig. 10, 25)
und wiederum an den Rändern erhaben, sodass zwei seitliche, freilich
flache Rinnen entstehen. Über diesen Rinnen stehen je zwei Haarreihen,
eine[S. 113] vom Aussenrande und eine von der Mitte her und unter solchem
Haardach wird die Luft in den beiden Rinnen von hinten her zur Brust
und zwischen Haaren derselben weiter zu den Stigmen fortgeleitet. Nicht
selten sieht man die Hinterbeine, Geigenbogen vergleichbar, über den
Hinterleib hinfahren, um die Luft in der einen oder anderen Richtung
fortzuschieben. Hin und wider klappen auch an der Oberfläche die
Haarreihen aus einander. Die Aussenränder der drei letzten Ringe sind
übrigens noch mit nach aussen gerichteten Haarreihen versehen und diese
ruhen beim Atmen auf der Oberfläche; auch die Bürste zur Reinigung der
Schwimmhaare befindet sich anscheinend hier.
Laich und Larven. Im Zimmeraquarium habe ich die Eier des
Rückenschwimmers mehrfach schon im März erhalten; sie waren einzeln an
lebenskräftigen, grünen Blättern abgelegt. Die ausschlüpfenden Larven
lassen die Zugehörigkeit zu dem entwickelten Insekt in allen Punkten
bis auf die weissliche Färbung und den Mangel der Flügel erkennen. In
Menge trifft man die Larven während der Sommermonate in den Gewässern
an, dem Raube obliegend wie die entwickelten Kerfe. Ende Juni thun
die ersten den letzten Schritt in der Entwickelung, der bei den
Schnabelkerfen im Vergleich z. B. zu den Käfern recht klein ist. Dass
hier kein Ruhe- d. h. Puppenstadium nötig ist, um die letzte Häutung
vorzubereiten, ist ohne Weiteres verständlich.
Mit Notonecta und Plea werden bisweilen die zahlreichen
und teilweise recht häufigen Arten der Gattung Corisa zu einer
Familie vereinigt. Ist auch eine gewisse oberflächliche Ähnlichkeit
vorhanden, so erweist sich doch Corisa bei eingehenderer
Betrachtung so verschieden von den beiden anderen, dass eine derartige
Vereinigung recht gewaltsam erscheint. Ich beschränke mich hier darauf,
hervorzuheben, dass die Corisa-Arten, unter denen C. Geoffroyi
(fast 1,5 cm messend) die grösste, aber nicht häufigste ist,
nicht mit dem Bauch, sondern mit dem Rücken nach oben schwimmen, dass
sie ferner nicht das Hinterleibsende zur Atmung an die[S. 114] Oberfläche
bringen, sondern den Vorderkörper, und zwischen Kopf und Vorderbrust
oder Vorder- und Mittelbrust, die dabei von einander gebogen werden,
die Luft einsaugen. Besonders grosse Luftlöcher liegen in der
Gelenkhaut der Vorder- und Mittelbrust, an den Seiten der Mittelbrust
und scheinbar an denen der Hinterbrust. Im Wasser erscheint die ganze
Unterseite mit einer silberglänzenden Luftschicht bedeckt. Natürlich
fehlen nun auch die eigentümlichen Luftrinnen an der Bauchseite des
Hinterleibes.
Von den Beinen ist wiederum das letzte Paar zu Schwimmbeinen
umgestaltet. Das Schwimmen erfolgt regelrecht unter gleichzeitiger
Bewegung beider Beine. Eine eigenartige, schaufelförmige Ausbildung
zeigt das Vorderbeinpaar. Es dient mit dem Schnabel zusammen bei
einzelnen Arten (oder bei allen?) als Musikinstrument. Die ziemlich
laute und anhaltende „Musik“ habe ich übrigens bei den Tieren, welche
ich hielt, stets erst am Abend gehört. Die Tiere hielten sich unter
Wasser mit den Mittelbeinen fest und geigten mit den Vorderbeinen über
den Schnabel. Deutlich liess sich die Gleichzeitigkeit des Tones mit
der Bewegung der Vorderbeine beobachten.
Wie alle Schnabelkerfe des Wassers leben auch die Corisa-Arten vom
Raube; aber sie dürften bei der Eigenart ihrer Mundteile nicht nur
zu saugen, sondern auch kleinere, festere Sachen zu zerkleinern
vermögen[LIV]. Häufig habe ich sie an winzigen Mückenlarven gesehen. Der
Schnabel weicht wesentlich von dem des Rückenschwimmers schon durch
die auffallende Kürze und Breite ab. Die Hauptmasse desselben wird
freilich, wie ich im Gegensatz zu den meisten mir bekannten Angaben,
welche gleichmässige Beteiligung der Ober- und Unterlippe an der
Schnabelbildung behaupten, fand, hier wie bei den anderen Rhynchoten
von der nach oben zusammengelegten und gegliederten Unterlippe
gebildet, in der die kurze Oberlippe und die vier Stechborsten
liegen[LV].
[LIV] Vergl. hierzu die Angaben von Geise, „Die
Mundteile der Rhynchoten“ im Archiv für Naturgeschichte 1886.
An der Bildung der Mundteile sind auch die im Mai ausnehmend häufigen
Larven als Corisa-Larven leicht zu erkennen. Dieselben bieten vor
der ersten Häutung in ihrer Atmung eine Abweichung sowohl von den
Notonecta-Larven als auch von den eigenen entwickelten Zuständen. Sie
kommen nämlich nicht dazu an die Oberfläche, sondern atmen durch die
zarte Hautoberfläche. Das Tracheensystem ist zu dieser Zeit an manchen
Teilen der Haut sehr fein und reich verzweigt, aber es besitzt, wie bei
dem durchsichtigen Körper mit Hilfe des Mikroskops leicht festzustellen
ist, keine thätigen Luftlöcher. Oft sah ich solche Larven sich mit
den Hinterbeinen frisches Wasser zufächeln, während sie sich mit den
Mittelbeinen festhielten. Larven mit Flügelansätzen zeigen freilich
schon die Atmungsweise der entwickelten Tiere[LVI].
[LVI] Die sehr ähnliche Gattung Sigara, besonders durch
die Zahl der Fühlerglieder unterschieden, habe ich bisher nur in
grösseren Gewässern, nicht in Gräben und Teichen gefunden.
3. Nepa, Ranatra, Naucoris.
Den dritten Typus der im Wasser lebenden Schnabelkerfe bilden die
beiden, je nur eine Art umfassenden Gattungen Nepa und
Ranatra (Fig. 10, 28). Beide sind kenntlich an den langen,
aus zwei seitlichen Halbrinnen bestehenden Atemröhren am Körperende
und an den ausgeprägten, nach vorn gerichteten Raubbeinen des ersten
Brustringes. Die Schiene derselben kann gegen den Schenkel wie die
Schneide eines Taschenmessers gegen den Griff umgeschlagen werden; die
Krallen am Ende fehlen, der eingliedrige Fuss selbst sieht dagegen
krallenartig aus. Hinsichtlich der Beinbildung bildet die Gattung
Naucoris (Fig. 10, 27) den Übergang zu Nepa; sonst
freilich weist sie nicht viel Übereinstimmung mit derselben auf.
Nepa cinerea, der Wasserskorpion, und Ranatra linearis
unterscheiden sich von einander leicht durch die Gestalt; der
erstere ist breit, verhältnismässig kurz und von oben nach unten
platt gedrückt, während Ranatra am besten gekennzeichnet
wird durch den Namen, welchen ihr jüngst einer meiner Schüler gab:
Strohhalmwanze. Wer sie nicht kennt, dürfte sie in der That oft[S. 116]
genug jener Ähnlichkeit wegen, da sie überdies meist bewegungslos mit
abgestreckten Beinen im Netz liegt, übersehen.
Mit der Schwimmbefähigung und Schwimmneigung beider ist es nicht
sonderlich bestellt; die Mittel- und Hinterbeine, welche dabei
gebraucht werden, sind nur schwach bewimpert. Sie bewegen jedoch die
Beine desselben Paares gleichzeitig und Ranatra, dessen Beine
ausserordentlich lang sind, kommt schwimmend ziemlich schnell fort.
Die Lieblingsstellung beider Tiere ist übrigens, sich schräg abwärts
gerichtet an einer Wasserpflanze mit den beiden letzten Beinpaaren zu
halten, so dass die Atemröhre eben zur Oberfläche emporragt, und so
bewegungslos auf Raub zu lauern. Blitzschnell erfassen sie mit ihren
Vorderbeinen vorbeischwimmende Tiere, auch winziger Grösse, z. B.
Daphnien. Einmal gefasst, entwindet sich keine Beute so leicht diesen
Zangen. Bisweilen sah ich Ranatra eine Daphnia aussaugen
und gleichzeitig mit jedem Vorderbeine eine neue Beute halten[LVII].
[LVII] Die Mundteile der Nepiden bieten übrigens eine
morphologische Besonderheit dar; die Unterlippe derselben besitzt am
vorletzten Schnabelgliede eingliedrige Taster im Gegensatz zu der
allgemeinen Angabe, dass den Rhynchoten eigentliche Lippentaster
fehlen. Das hatte bereits Savigny erkannt und abgebildet.
Seine Abbildung ist in Lehrbüchern immer wieder kopiert worden,
aber anscheinend ohne Verständnis und Nachuntersuchung, denn die
Taster sind in den Abbildungen recht undeutlich geworden und in den
Erklärungen und Texten völlig verschwunden. Burmeister, der
auch die Abbildung von Savigny bringt, hat sogar auf das Fehlen der
Unterlippentaster hin eine besondere Deutung der Gliederung des
Rüssels aufgestellt, die oft wiederholt wurde, und Huxley
(„Wirbellose Tiere“) zieht mit deswegen in Frage, ob die Mundteile
der Rhynchoten mit denen der anderen Insekten homologisiert werden
können. Aufallenderweise giebt auch Geise, der die Mundteile der
Schnabelkerfe so genau untersucht hat, an, dass bei Nepa und
Ranatra die Lippentaster fehlen. Vgl. meine Mitteilungen über
Mundteile der Rhynchoten und die Stigmen derselben in d. Sitzungsber.
der Gesellschaft naturforschender Freunde zu Berlin, 1891.
Die Eier legen beide Wanzen im Mai an abgestorbenen schwimmenden Binsen
u. dergl. ab und zwar so, dass das eigentliche Ei in die Pflanze
eingesenkt ist und nur die eigenartigen, fadenförmigen Anhängsel der
Eier (bei Ranatra zwei, bei Nepa mehrere) hervorragen
(Fig. 10, 29 u. 30). Von Ranatra habe ich mehrfach die Eier
im Mai zu vielen Hunderten angetroffen. Die jungen Larven schlüpfen
nach kurzer Zeit aus; die Zugehörigkeit zum entwickelten Tier ist bei
ihnen wie bei Nepa ohne Weiteres zu erkennen. Es fehlen ihnen
zunächst nur die Flügelansätze und die Atmungsweise ist eine etwas
andere. Bei den entwickelten[S. 117] Kerfen liegen die einzig offenen Stigmen
am Ende des Hinterleibes, am Grunde der Atemröhre; die übrigen Stigmen
desselben sind auf der Bauchseite zwar vorhanden, aber geschlossen und
nicht in Thätigkeit; bei den Larven sind dagegen an der Bauchseite
zwei gleichartige Haarrinnen vorhanden wie bei Notonecta, und
unschwer erkennt man auch, dass von der kurzen Atemrinne aus —
eine zweiteilige Atemröhre ist noch nicht ausgebildet — die
Luft in diesen seitlichen Gängen zu den dort liegenden Luftlöchern
fortgeleitet wird. In allen anderen Punkten aber, in der Bildung der
Mundteile, im Schwimmen, in der gewöhnlichen Körperhaltung, sind die
jungen Tiere ein getreues Abbild der alten.
Schlussbemerkungen.
Reich und mannigfaltig ist also, wie wir sehen, das Kerf- und besonders
Kerflarvenleben der süssen Gewässer entwickelt. Naturgemäss
drängt sich da zum Schluss die Frage auf: Wie verhält sich dazu
die Kerfwelt des Meeres? Ist dort ein ähnlicher Reichtum an Formen
vorhanden; sind es verwandte und gleiche Formen wie die des süssen
Wassers? Die Auskunft auf solche Fragen muss für Jeden zunächst
überraschend sein: Von Kerfen und besonders Kerflarven des Meeres ist
so gut wie gar nicht die Rede. Ausser einer Gattung der Wasserläufer
(Halobates, Meerwanze) giebt es nur ganz vereinzelte Meerestiere
unter den Kerfen und Kerflarven. Dass der Salzgehalt des Meerwassers
die Ursache dieser auffallenden Erscheinung sein sollte, ist, auch
schon wegen der Fauna der salzigen Gewässer des Binnenlandes, nicht
wohl anzunehmen. Vielleicht ist dieselbe im folgenden zu suchen. Die
fast nie ruhende Bewegung der Meeresoberfläche, besonders auch näher
der Küste, macht zunächst allen Kerfen und Larven, die zur Atmung an
die Oberfläche kommen, oft auf lange Zeit das Atmen und also das Leben
im Meere unmöglich; aber auch für diejenigen Larven, bez. Puppen,
welche durch Kiemen oder Tracheenkiemen atmen und die also zunächst
in der schützenden Tiefe verbleiben können, kommt früher oder später
der Zeitpunkt, wo[S. 118] sie zur Verpuppung oder zum Ausschlüpfenlassen des
Insektes das Wasser verlassen müssen, und welche Schwierigkeit bietet
sich dann für die schwachen Lebewesen durch die Brandung das Ufer
und weiter eine Stelle zu erreichen, an der sie nicht eine spätere
Welle oder die steigende Flut erreiche. Wie dem nun auch sein möge,
jedenfalls bilden die Kerfe hierin einen bemerkenswerten Gegensatz zu
der verwandten Klasse der Kruster, den echten Wassertieren, welche im
Meere so ungleich reicher als im süssen Wasser entwickelt sind.
Anhang.
Tabelle zur annähernden Bestimmung der im Wasser lebenden
Kerflarven[LVIII].
Mit je einem kräftigen Chitinhaken an zwei kürzeren
oder längeren Afterbeinen am Hinterleibsende. Fühler meist fehlend, selten
vorhanden und dann zweigliedrig, winzig. Mit oder ohne fadenförmige
mehrreihige Kiemen. Mit oder ohne Gehäuse
[LVIII] Die weitere Unterscheidung der unten folgenden Gruppen
siehe teilweise im Text.
[LIX] Die eingeklammerten Zahlen weisen auf den Gegensatz hin.
[LX] Die jüngsten Larven sind zwar ohne Flügelansätze, aber
durch ihre sonstige Ähnlichkeit mit den ungleich häufigeren
älteren Larven, die solche Ansätze haben, leicht kenntlich. Übrigens
lassen sich die Larven der vier ersten Gruppen auch abgesehen von
den Flügelansätzen durch die oben gekennzeichneten Merkmale der
Mundbildung, der Kiemen und Endanhänge von den folgenden Larvengruppen
unschwer unterscheiden. Zu Hilfe kann man noch nehmen, dass die Larven
dieser vier Gruppen alle Netzaugen und grosse, wohl entwickelte Beine
mit fast immer (oder immer?) teilweise gegliederten Tarsen haben und
dass Brust- und Hinterleibsringe meist deutlich in ihrer Bildung von
einander unterschieden sind.
[LXI] Die Gattungen sind unschwer durch die Ähnlichkeit mit den
erwachsenen Tieren zu bestimmen.
[LXII] Kiemen steht in der Tabelle der Kürze wegen statt
Tracheenkiemen.
[LXIII] Meines Wissens kommen nur bei Philhydrus testaceus
Afterbeine vor und zwar stehen sie am dritten bis siebenten
Hinterleibsring.
[LXIV] Hierher gehören eigentlich auch wegen ihrer Kiemen
Berosus (Hydrophilide), Cnemidotus und Pelobius
(Dytisciden). Von den Gyriniden, Sialis und Sisyra
unterscheidet sich Cnemidotus dadurch, dass auch an den
Brustringen Kiemen stehen, Pelobius durch bluterfüllte echte
Kiemen an der Unterseite der Brust und Berosus durch die
besondere Oberkiefer- und Beinbildung (S. 24) der Hydrophiliden.
[S. 121]
Litteratur.
[1] Zum Bestimmen der im Wasser lebenden Käfer wie der
Käfer überhaupt ist zu empfehlen der betreffende Band der „Fauna
austriaca“, nämlich Redtenbacher: „Die Käfer“. Wien 1858. Die
Litteratur über die Käferlarven und Käferpuppen ist übersichtlich
zusammengestellt in M. Rupertsberger, „Biologie der Käfer
Europas“. Linz 1880. Für unsern Zweck sind fraglos am wichtigsten die
Arbeiten Schiödtes, welche in der Naturhistorik. Tidsskrift von
Kröyer, Kopenhagen, erschienen sind und zwar in den Jahrg. 1862,
1864, 1867, 1872. Der Text ist teilweise dänisch, teilweise lateinisch;
die wichtigste Auskunft geben übrigens schon die vorzüglichen
Abbildungen. Nicht unterlassen möchte ich, gleich hier auf die unser
ganzes Gebiet behandelnden älteren Werke von Rösel v. Rosenhof,
Réaumur und De Geer hinzuweisen.
Rösel, „Monatlich herausgegebene Insektenbelustigungen“. 1746
bis 1761.
de Réaumur, Mémoires pour servir à l’histoire des Insectes.
Paris 1734–42.
de Geer, Mémoires pour servir à l’histoire des Insectes.
Stockholm 1752–78. (Deutsch von Götze. Nürnberg 1778–83.)
[2] Eine umfassende Zusammenstellung der Litteratur über die
Metamorphose der Dipteren giebt Fr. Brauer, „Die Zweiflügler
des kaiserlichen Museums zu Wien“ (Denkschr. d. k. k. Akad. d. Wiss.
Bd. 47. Wien 1883). Dort sind auch die Larven systematisch gruppiert
und kurz geschildert. Für die eucephalen Larven ist wegen seiner
vortrefflichen Abbildungen besonders zu empfehlen: Fr. Meinert,
„Eucephale Myggelarver“ (Vidensk. Selsk. 6 Räkke naturvidensk. og math.
Ath. III 3). Kopenhagen 1886.
[S. 122]
[3] Abbildungen der Wasserraupen finden sich in v.
Praun, „Abbild. u. Beschreib. europäischer Schmetterlingsraupen“.
Herausgegeben von E. Hoffmann, 1874. Vergl. auch
Sorhagen, „Kleinschmetterlinge der Mark“.
[4] Für Phryganiden, Ephemeriden und Perliden sind mit
Vorteil zu benutzen die Werke von F. J. Pictet, 1) Recherches
pour servir à l’histoire et à l’anatomie des Phryganides. Genf-Paris
1834. 2) Histoire naturelle des insectes neuroptères. Genf-Paris
1841–1843.
Ferner ist für beide Ordnungen zu empfehlen: Fr. Brauer,
Neuroptera austriaca. Wien 1857. Eine Anzahl Beschreibungen und Muster
für genaue Beschreibung der Larven und Puppen der Phryganiden findet
man in: Klapalek, „Metamorphose der Trichopteren“. Prag 1888.
Für die Bestimmung der entwickelten Insekten ist besonders zu
nennen: Mc. Lachlan, A monograph revision and synopsis of the
Trichoptera of the European Fauna. London 1874–80.
[5] Die Ephemeriden nebst ihren Larven sind sehr eingehend
behandelt in: Eaton, A revisional monograph of recent
Ephemeridae. (Transact. of the Linnean Society.) London. Zoology. N. S.
3. 1888. Das Werk enthält zahlreiche Tafeln über die Larven.
[6] Zur Bestimmung der Gattungen der Wasserwanzen dürften
die allgemeinen systematischen Handbücher ausreichen, z. B. das von
Ludwig-Leunis. Zur Unterscheidung der Arten sind die Arbeiten
von Fieber, besonders „Hemiptera europaea“, zu empfehlen.
[S. 123]
Die Mollusken des Süsswassers.
Von S. Clessin in Ochsenfurt.
[S. 125]
Unsere Gewässer, von der kleinsten Pfütze bis zu den grössten Seen und
von der spärlichsten Quelle bis zu den wasserreichsten Flüssen, werden
von Mollusken verschiedener Gattungen bewohnt. Aber obwohl die in den
Gewässern vorkommenden Arten meist in reicher Individuenzahl auftreten,
fallen sie dennoch dem nicht geübten Beobachter nicht so leicht ins
Auge und es bedarf selbst der gewandte Sammler in der Regel besonderer
Instrumente, um lebende Mollusken in grösserer Anzahl zu fangen. Leere
Gehäuse werden dagegen oft in reicher Menge an gewissen Lokalitäten
angeschwemmt gefunden.
Die Mollusken spielen im Haushalte der Natur eine wichtige Rolle, indem
sie faulende Pflanzenstoffe, welche in den Gewässern sich ansammeln,
verzehren und dadurch die Wasser rein erhalten. Die Mehrzahl der
Arten werden ihrer geringen Grösse und verborgenen Lebensweise wegen
leicht übersehen, doch beherbergen unsere Gewässer auch grosse, recht
ansehnliche Arten, namentlich aus der Klasse der Muscheltiere, die
bezüglich ihrer Entwickelungsgeschichte noch besonderes Interesse
bieten.
Einteilung der Mollusken.
Die im Wasser lebenden Conchylien gehören zwei sehr verschiedenen
Klassen an. Die eine besteht aus Tieren, welche einen Kopf mit Fühlern
und Augen haben, die gewöhnlich an der Basis der Fühler sitzen, im
übrigen aber jenen der Landschnecken, der[S. 126] Klasse der Gasteropoden
oder Bauchfüssler ähnlich sind. Sie haben mit wenig Ausnahmen (Genus
Ancylus Geoff. und Velletia Gray[LXV]) eine gewundene
Schale, und ist das Gehäuse zuweilen mit einem Deckel verschlossen.
Die zweite Klasse die Muscheltiere, Bivalven oder Zweischaler, haben
keinen Kopf und keine Augen; das Tier besteht nur aus einem sackartigen
Körper, dessen unterer, ausdehnbarer Teil als „Fuss“ die Bewegung
vermittelt. Den Körper umhüllen auf jeder Seite zwei buchblattartig
am Rücken angeheftete Kiemen und wird das ganze Tier von einem Mantel
umschlossen, dessen Ränder entweder ganz frei bleiben, oder teilweise
zusammengewachsen sind; im letztern Falle hat der Mantel einen Schlitz
am Vorderteile des Tieres zum Durchgange des Fusses und eine Öffnung
für die Anal- und Atemröhre. Die Schalen der Bivalven sind nicht
gewunden, sondern bestehen aus zwei gleichgrossen Klappen, die durch
ein elastisches Band, das Ligament, verbunden sind und sich nur wenig
öffnen können. Das Tier ist an den gegenüberstehenden Enden durch zwei
sehr starke Muskeln (die Schliessmuskeln), welche zugleich das Öffnen
der Schalen regeln, und durch einen kleinen Wirbelhaftmuskel an die
Schale angeheftet sind.
[LXV] Bezüglich der Arten und Genera verweise ich auf meine
Werke: „Deutsche Excursions-Mollusken-Fauna“. 2. Aufl. Nürnberg 1884,
Bauer & Raspe, und „Molluskenfauna von Österreich-Ungarn und der
Schweiz“. Nürnberg 1890, Bauer & Raspe.
Die meisten Süsswassermuscheln sind freibeweglich; nur eine Art unserer
heimischen Arten heftet sich durch einen Byssus (einen Büschel spröder
Haare) an anderen Gegenständen an (Dreissena polymorpha Pallas
Fig. 11).
Fig. 11. Dreissena polymorpha.Fig. 12. Ancylus fluviatilis mit Tier.Fig. 13. Valvata piscinalis mit Tier.
Die Genera der nicht gedeckelten Süsswasserschnecken sind durchweg
Lungenatmer. Sie haben sehr verschiedene Formen, indem der Modus
des Aufwindens der Umgänge sehr mannigfaltig ist. Die meisten Arten
haben eine rechtsgewundene Schale, nur die Genera Amphipeplea,
Physa und Aplexa[S. 127] winden ihre Umgänge nach links. Ferner
besitzen die Genera Limnaea, Physa, Aplexa,
Amphipeplea ein mehr oder weniger erhöhtes Gewinde, nur
Genus Planorbis rollt seine Umgänge in platter Form auf, für
welche der Name „Tellerschnecke“ sehr bezeichnend ist. Die Genera
Ancylus (Fig. 12) und Velletia haben eine napfförmige
Schale, von denen diejenige des ersteren Genus einer Jakobinermütze
sehr ähnlich ist; bei beiden beschränkt sich die Windung des Gehäuses
auf eine geringe Neigung des Wirbels nach rechts oder links. Die
meisten Arten haben eine rauhe Schale, an welcher die Zuwachsstreifen
deutlich erkennbar sind. Nur Genus Physa und Aplexa haben
glatte, glänzende Gehäuse.
Die Deckelschnecken, mit Ausnahme des Gen. Vivipara, bestehen
aus kleinen Arten. Alle sind mit Kiemen zur Wasseratmung ausgerüstet.
Gewöhnlich bleiben die Kiemen in der Kiemenhöhle verborgen, nur Genus
Valvata (Fig. 13) besitzt die Fähigkeit, die federförmige Kieme
auszustülpen und frei hervortreten zu lassen.
Das Gewinde ist bei diesen Schnecken ein kreiselförmiges (Gen.
Vivipara und Valvata) oder ein mehr oder weniger
getürmtes, nur Gen. Neritina und Lithoglyphus haben ein
kurzes, wenig hervortretendes Gewinde und eine starke Schale und weite
Mündung; die Oberfläche der Arten des Gen. Neritina ist mit
hübschen netzartigen Zeichnungen bedeckt.
Die frei beweglichen Muscheln gehören zwei Familien an. Die grossen
Arten gehören in die Familie der Najaden. Diese haben offenen Mantel,
zwei gleichgrosse Kiemen und an der Mundöffnung jederseits zwei
Mundlappen; der Mantel ist an seinem Rande am Hinterteile mit einem
Kranze dunkelgefärbter Papillen besetzt. Die Kiemen dienen zugleich
als Brutbehälter, haben gitterförmige Abteilungen, die, wenn Brut
vorhanden, mit einer ungeheuren Anzahl von Embryonen besetzt sind.
Die kleineren Zweischaler der Familie Cycladidae haben
geschlossenen Mantel, und je eine Anal- und Atemröhre, welche mehr oder
weniger über den Mantelrand hervortritt.
[S. 128]
Wohnorte und Gewohnheiten.
Die ungedeckelten Wasserschnecken der Familie der Limnaeiden halten
sich den grössten Teil des Tages auf dem Grunde der Gewässer im
Schlamme auf, und ziehen die meisten Arten stehende Gewässer vor;
nur wenige Spezies finden sich in fliessenden Wassern, für welche
sie wegen ihrer dünnen, zerbrechlichen Schale wenig geeignet sind.
In fliessenden Wassern kommt in der Regel nur Limnaea peregra
vor. — Die übrigen Limnaea-Arten, die Spezies der Gen. Physa,
Aplexa, Amphipeplea und Planorbis bewohnen nur
stehende Gewässer.
Fig. 14. Aplexa hypnorum.Fig. 15. Vivipara vera Fr.
Die Limnaeen (Limnaea stagnalis, auricularia,
ovata, palustris) steigen bei heiterem, warmem Wetter
an Wasserpflanzen an die Oberfläche des Wassers und kriechen, die
Fusssohle nach oben gerichtet, das Gehäuse untergetaucht, ebenso sicher
dahin, als wenn sie an einem festen Gegenstande kröchen. Wahrscheinlich
saugt sich die Sohle an der auf dem Wasser aufstehenden Luftsäule an,
da die Tiere sich jederzeit plötzlich zu Boden fallen lassen können. —
Das Aufsteigen der Limnaeen an die Oberfläche wird mit dem Bedürfnisse
der Tiere, Luft zu atmen, in Verbindung gebracht, da die Limnaeiden
mit Lungen ausgerüstet sind, während die mit Kiemen versehenen
Deckelschnecken nie an die Oberfläche kommen. Die Limnaeen haben jedoch
dieses Bedürfnis nur bei heiterem Wetter und bei erhöhter Temperatur
des Wassers ihrer Wohnorte.
Eine Aplexa-Art (Apl. hypnorumFig. 14) besitzt sogar die
Fähigkeit, plötzlich vom Grunde des Wassers an die Oberfläche
aufzutauchen, von wo sie sich nach einigen Sekunden ebenso schnell
wieder zu Boden fallen lassen kann.
Die gedeckelten Wasserschnecken der Genera Vivipara (Fig. 15),
Bythinia und Valvata leben im Schlamme der Gewässer,
und zwar meist in stehenden, höchstens in sehr langsam fliessenden
Wassern. —[S. 129] Die Arten der Genera Neritina und jene der Familie
der Melaniiden leben nur in bewegtem Wasser, in welchem sie sogar
stark flutende Stellen bevorzugen, für welche die Neritinen durch ihr
kaum hervortretendes Gewinde und ihre weite Mundöffnung vorzugsweise
geeignet erscheinen, weil sie den Fluten wenig Fläche darbieten. — Die
Bythinellen (Fig. 16) finden sich nur in Quellen an Steinen sitzend;
die Vitrella-Arten kommen ausschliesslich in Höhlengewässern
vor. — Velletia lacustris lebt in stehendem, Ancylus
fluviatilis und verwandte Arten nur in fliessendem Wasser. Doch
findet sich erstere zuweilen auch in Bächen, während Ancylus-Arten auch
in Seen vorkommen.
Fig. 16. Bythinella austriaca Frf.
Die Muscheln stecken am Grunde der Gewässer im Schlamme, in dem sie
fast völlig eingebettet sind, so dass nur das hintere Ende frei
ins Wasser hervorragt. Sie saugen das Wasser durch die Mundöffnung
am vordern Ende der Muschel ein, lassen dasselbe durch den Körper
zirkulieren und stossen es durch die Atemöffnung am hintern Ende wieder
aus. Wenn man eine Muschel rasch aus dem Wasser nimmt, schliesst sie
ihre Schalen und das Wasser spritzt dann, oft in ziemlich lebhafter
Weise, durch die Atemöffnung aus. Beim Einblick in helles nicht
tiefes Wasser kann man die im Schlamme steckenden Muscheltiere leicht
bemerken. Man gewahrt jedoch nur die mit Cirren am hintern Mantelrande[S. 130]
besetzte Atem- und Analöffnung. Schiebt man vorsichtig ein Rütchen in
diese Öffnung, so schliesst das Tier die Schalen, und die Spitze des
Rütchens wird mit eingeklemmt. Mit dem Rütchen lässt sich dann die
Muschel aus dem Schlamme ziehen, wenn man dieselbe fangen will.
Die Muscheln (s. Fig. 17) heften sich in fliessendem Wasser mit dem
ausgestreckten Fusse in den unter der Schlammschicht befindlichen
festen Boden. Ihre Bewegungsfähigkeit ist eine sehr geringe,
und ihr ruckweise erfolgender Marsch erstreckt sich nur auf 1–2
m Länge. Derselbe wird durch Ausstrecken und Einziehen des
Fusses bewerkstelligt; bei letzterer Operation wird die Muschel
nachgeschleift, wobei sie im Schlamme eine Furche zurücklässt, an
welcher man die Länge und Richtung des Marsches erkennen kann. Die
kleineren Muscheln der Familie der Cycladen leben ebenfalls frei
beweglich im Schlamme. Nur eine Art unserer Süsswassermuscheln,
Dreissena polymorpha, heftet sich durch einen Byssus an
andere im Wasser liegende feste Gegenstände an, und wechselt dann
ihren Standort nicht mehr bis zu ihrem Tode. Die Muscheln sitzen
oft in ganzen Klumpen zusammen und verstopfen beispielsweise leicht
Wasserleitungsröhren, wenn sie in selbe gelangen. — Das Festsitzen
dieser durch ihre dreieckige Form auffallenden Muschel ist die
Veranlassung zur Verschleppung in die nord- und westeuropäischen
Gewässer geworden: Ursprünglich in den Flüssen heimisch, welche ins
Schwarze Meer münden, wurde sie durch Schiffe, an deren Planken sie
sich angehängt hatte, an die Küsten der Ost- und Nordsee, sowie des
Atlantischen Meeres verschleppt, und gelangte von hier wieder durch
Flussschiffe in alle grösseren ins Meer mündenden Flüsse, von welchen
sie in deren Nebenflüsse vordrang. Durch den Donau-Main-Kanal war
es ihr sogar möglich, die Wasserscheide zwischen Rhein und Donau zu
überschreiten und in die obere Donau zu gelangen, wo ich im Jahre 1868
das erste Exemplar fand. Einige Jahre später wurde sie bei Deggendorf
beobachtet und so wird sie nun sicher die Donau abwärts wandern, bis
sie wieder das Schwarze Meer, ihren Ausgangspunkt, erreicht hat.
Fig. 17. Anodonta mutabilis v. cellensis Chem.
[S. 131]
Entwickelung und Alter der Mollusken.
Die meisten Wasserschnecken sind Zwitter; wenigstens die nicht
gedeckelten Arten, also insbesondere die Limnaea- (Fig. 18),
Planorbis-, Physa-, Ancylus-, Velletia- und Amphipeplea-Arten. Bei
Limnaea peregra habe ich mehrfach beobachtet, dass ganze
Ketten, 6–8 Individuen, bei der Begattung zusammenhingen. — Bei den
Deckelschnecken, wenigstens bei Vivipara, lassen sich männliche
und weibliche Formen unterscheiden, ebenso nach Hazay[LXVI] bei
den Muscheln der Familie der Najaden. Ich halte dies jedoch noch immer
für sehr zweifelhaft, bis weitere verlässliche Beobachter und Anatomen
dieses Verhältnis bestätigt haben.
[LXVI] „Mollusken-Fauna von Budapest“. Kassel 1881, Theodor
Fischer.
Die Wasserschnecken legen Eier; nur bei den Vivipara-Arten entwickeln
sich die jungen Individuen schon im Muttertiere, so dass sie bereits
mit einem etwa aus zwei Umgängen bestehenden Gehäuse ausgestossen
werden. Auch die Arten der Familie Cycladidea stossen ihre
Jungen schon als fertige Muscheln aus.
Die Schnecken und Muscheln sind schon fortpflanzungsfähig lange bevor
sie ausgewachsen sind. Die im Mai ausgekrochenen Jungen der Limnaea-
und Planorbis-Arten begatten sich noch im selben Herbste, obwohl sie
ein Alter von 3–4 Jahren erreichen.
Fig. 18. Lim. palustris v. corvus.
Die Eier werden in Schnüren oder in Paketen an Steinen, Wasserpflanzen
oder häufig sogar auf die Gehäuse anderer Individuen derselben Art
abgesetzt, so z. B. bei Limnaea ampla. Hazay hat auf den
Gehäusen dieser Art 8–12 Eierschnüre gefunden, so dass das Tier nur
mühsam sich fortbewegen konnte. — Limnaea auricularia setzt
20–25 mm lange, 7–8 mm breite raupenförmige Eierschnüre
ab, welche 80–150 Eier enthalten, die kugelrund sind und 1 mm
Durchmesser haben. Der Eidotter ist weisslichgelb und wird während
der Furchung hellweiss. — Limnaea stagnalis-variegata Hazay
setzt eine Eierschnur von 45–55 mm Länge[S. 132] ab, die 110–180
Eier enthält; die Eierchen sind länglich-oval und 1½–2 mm
gross. Der Dotter ist strohgelb, das Eiweiss wasserhell; Aplexa
hypnorum legt den Laich in ganz flachen rundlichen Scheiben von
4–7 mm Durchmesser und ⅔ mm Dicke, mit den Enden
gegeneinandergeheftet, ab. Die Zahl der Eier wechselt zwischen 20–50.
— Planorbis corneus legt ebenfalls eine 25 bis 30 mm
lange, 5 mm breite, glatte, an den Enden zusammengeheftete
Eierschnur ab. Zahl der Eier 45–70.
Die Entwickelung des Embryo beansprucht bei Gen. Limnaea
gewöhnlich 20, bei Planorbis und Physa nur 15, bei
Bythinia 25 Tage. Je nach der Temperatur des Wassers wird
der Entwickelungsprozess beschleunigt und verzögert. Hazay
hat beobachtet, dass bei Laich der Limnaea palustris var.
Clessiniana die Embryonen sich schon in 12 Tagen entwickelten.
Die jungen Tiere wachsen ziemlich rasch und erlangen vier bis sieben
Umgänge schon im ersten Jahre, jenachdem sie mehr oder weniger
frühzeitig im Jahre als Laich abgesetzt wurden. Das grösste Wachstum
entfällt auf das erste und zweite Jahr und nimmt dasselbe dann von Jahr
zu Jahr ab. Im Herbst und Winter erfolgt nicht das geringste Wachstum.
Während der letzten Wachstumsmonate wird der letzte frische Anbau des
Gehäuses verdickt und die Mündung verstärkt.
Die Lebensdauer der Limnäen erstreckt sich im höchsten Falle auf 4–5
Jahre; nur wenige erreichen jedoch dieses Alter. Die Jahre, welche
die Mollusken zum Ausbau ihres Gehäuses brauchen, lassen sich an den
Jahresabsätzen deutlich erkennen, da diese Tiere gleich den Insekten,
Lurchen etc. einen Winterschlaf halten. Schon im Hochsommer wächst das
Gehäuse, dessen Weiterbau im Frühjahr sofort nach dem Erwachen aus dem
Winterschlafe, meist im Monat April, beginnt, nicht mehr weiter; die
Zeit bis zum Eintritt der Winterruhe wird dazu benutzt, die Mündung
des Gehäuses durch Ablage einer Schmelzschicht zu verstärken, damit
dieselbe beim Einbohren in den Schlamm nicht beschädigt wird. Die
Jahresabsätze sind daher an den Gehäusen, durch die meist nach aussen
durchscheinenden Verstärkungsschichten, leicht zu[S. 133] erkennen, und lassen
sich aus der Zahl dieser Absätze die Jahre, die das Tier bis zur
Vollendung des Gehäuses braucht, ablesen. Die Limnäen weisen zwei bis
drei solcher Absätze, unter Umständen sogar deren vier, auf. Limnaea
peregra hat in der Regel nur drei; ich habe jedoch auch aus höheren
Lagen im Gebirge stammende Gehäuse dieser Art mit vier Jahresabsätzen
gefunden, so dass anzunehmen wäre, dass die kürzere Sommerperiode
höher gelegener Lokalitäten die Lebensdauer verlängert. — Limnaea
auricularia und ovata sterben meistens schon im zweiten
Jahre ab, Limnaea palustris (Fig. 18) gewöhnlich im dritten.
Planorbis corneus, marginatus und carinatus
vollenden ihre Gehäuse im dritten und leben selten länger als 3–3½
Jahre; Planorbis albus, spirorbis und alle kleineren
Arten dieses Genus sterben in der Regel schon im zweiten Jahre.
Amphipeplea glutinosa lebt nur ein Jahr.
Unter allen Wasserschnecken werden die Limnäen am meisten von
Schmarotzertieren gequält, so dass die allermeisten derselben meist
schon, bevor die Schale ausgewachsen ist, zu Grunde gehen. Hazay
sagt hierüber folgendes: Keine einzige der Limnäen, welche das dritte
und vierte Lebensjahr erreicht hat, bleibt von denselben verschont; in
diesem Alter fallen alle denselben, wie einer allgemein herrschenden
Alterskrankheit, zum Opfer. Im zweiten Lebensjahre bereits finden sich
einzelne Sporocisten an dem Darm und der Leber als längliche gelbe
Schläuche vor, im dritten Lebensjahre sind dies schon massenhafte
Schlauchbündel, welche alle inneren Organe bedecken, die ganze Leber
erfüllen, langsam Herz und Lungenwand durchsetzen, so dass endlich
das Tier absterben muss. Dieser Zustand der Tiere macht sich durch
auffallende Trägheit und durch eine starke gelbe Färbung bemerkbar.
Zieht man solche Tiere aus dem Gehäuse, so erscheint unter der Haut
das ganze Innere des Körpers als gelbe Masse, alle Organe sind von
Sporocistenbündeln belegt und von der Leber ist keine Spur mehr
vorhanden.
Die Vivipara-Arten setzen keinen Laich ab; die Eier entwickeln
sich im Muttertiere bis zu Gehäusen von 11 mm Länge und 7
mm Breite, welche etwa 3½ Umgänge zählen (Vivipara
hungarica Hazay, l. c. p. 91). Die Schalen solch junger Tiere[S. 134]
haben eine dichte Spiralstreifung und sind die Streifen mit an einander
gereihten rundlichen Wärzchen besetzt, von denen manche kurze Borsten
tragen. Hazay fand im Uterus des Weibchens der genannten Art
sechsundvierzig schon mit Schale und Binde versehene junge Tierchen und
sehr viele Eier in allen Stadien der Entwickelung. — Nach demselben
Autor sind die Tiere getrennten Geschlechtes und lassen sich die
Geschlechter an der Form der Schale gut unterscheiden. — Die Schalen
ausgewachsener Tiere erreichen sieben Umgänge und erlangen dieselben
ein Alter von 8–10 Jahren.
Die Arten der Familien Valvatidae und Hydrobiidae setzen
Laich ab. Sie erreichen ein Alter von 2–3 Jahren. Neritina- und
Lithoglyphus-Arten können nach Hazay ein Alter von 5 Jahren
erreichen.
Die Muscheln der Familie der Najaden sind wahrscheinlich Zwitter,
obwohl mehrere Autoren männliche und weibliche Formen an den
Muscheln (vorzugsweise an der mehr aufgeblasenen Form der Schalen)
unterscheiden wollen. Da nämlich die Kiemen als Brutbehälter für die
Eier dienen und strotzend mit denselben gefüllt werden, wird die
Muschel sehr aufgetrieben, während jene Muscheln, die keine Eier in
die Kiemen bringen und aus irgend welchem Grunde vielleicht nicht
fortpflanzungsfähig sind, wenig aufgeblasene Schalen behalten. —
Die Anodonta- und Unio-Arten produzieren ganz enorme Massen von
Eiern, die aus den Ovarien in die Kiemen gelangen und hier die ersten
Stadien ihrer Entwickelung durchmachen. Bei Anodonta anatina
wurden 120000, bei An. cygnea sogar 400000 Eier gezählt. Die
Einlagerung solch grosser Massen von Eiern kann nicht auf einmal
erfolgen. Dieselben werden allmählich, je nach ihrer Entwickelung,
eingeführt und zwar füllen sich die mittleren Fächer der Kiemen
zuerst, denen dann die gegen die Enden der Muschel zu gelegenen
folgen. — In den Kiemen entwickeln sich die Eier zu Larven, wozu
sie nach Hazay je nach den Temperaturverhältnissen 2–3
Monate brauchen. — Die Eihülle wird erst gesprengt, wenn sich die
eigentümlich gestaltete Larvenschale vollkommen ausgebildet hat.
Dieselbe[S. 135] ist von dreieckiger Form und besitzt in der Mitte der
Bauchseite einen kleinen Höcker (Fig. 19). Ist die Eihülle entfernt,
so bilden sich an den Larven Byssusfäden, mit denen sich die in einem
Kiemenfache befindlichen Individuen derart verwickeln, dass sie wie
aneinandergeheftet erscheinen. Die zusammenhängenden Larvenklumpen
werden vom Muttertiere ausgestossen, und fallen in den Gewässern
zu Boden, wo die Byssusfäden der Larven im Wasser flottieren. Die
Byssusfäden verfangen sich an langsam über dem Schlamme schwimmenden
Fischen, hängen sich an dieselben an, bilden an den Fischen kleine
Cysten, in welchen sie sich so lange aufhalten, bis die junge Muschel
soweit ausgereift ist, dass sie nun ohne Schutz, allein ihre weitere
Entwickelung finden kann. Hazay hat an folgenden Fischarten die
Cysten von Najaden gefunden: Perca fluviatilis L., Acerina
cernua L., Acerina Schraetzer L., Cottus gobio
L., Squalius cephalus L., Leuciscus virgo Heck.,
Rhodeus amarus Blain., Tinca vulgaris Cuv., Carassius
vulgaris Nils. und Cyprinus carpio L. — Die Zeit, während
welcher sie als Schmarotzer an Fischen leben, beträgt nach Braun
70–73 Tage.
Fig. 19.
Junge Muschel von Anodonta zur Zeit, wenn dieselbe aus den
Kiemen ausgestossen wird.
(Vergrössert.)Fig. 20.
Junge Muschel von Unio batavus im ersten Lebensjahre.
Es ist sehr wahrscheinlich, dass die Existenz der Muscheln durch das
Vorhandensein von Fischen bedingt ist, da sich in stehenden Wassern,
welche keine Fische beherbergen, sich auch keine Najaden finden. —
Bei diesem Verhältnisse der beiden so verschiedenen Tierklassen ist es
gewiss gerechtfertigt, dass auch die grossen Muscheln den Fischen einen
Gegendienst erweisen. Es finden sich nämlich in denselben in der innern
Kieme Fischembryonen vor, die hier ihre Entwickelung erfahren.
Hazay fand Mitte April bei kiementrächtigen Anodonten (bei
fast jedem zweiten der untersuchten Tiere) in den inneren Kiemen 4–8,
ebenso bei Unio pictorum 5–16 Fischembryonen, die möglicherweise
von den Muschellarven sich genährt haben. Bei der grossen Anzahl der
Eier, welche die Fische absetzen, ist es wahrscheinlich,[S. 136] dass diese
Fischembryonen nur durch einen glücklichen Zufall zwischen die Kiemen
der Muscheln geraten. Es ist leider noch nicht festgestellt, ob gewisse
Fischarten nur auf diesem Wege ihre Embryonen-Stadien durchzumachen
haben. Hazay hat ferner beobachtet, dass die Fische sich in
den Muscheln bis zu voller Ausbildung aufhalten, und dass die jungen
Fischchen ausserhalb der Muschel sich ganz nahe der Atemöffnung der
Muschel halten und durch dieselbe in die Muschel hineinschlüpften.
Die Fortpflanzungsfähigkeit tritt bei Genus Unio im dritten, bei
Genus Anodonta im dritten oder vierten Lebensjahre ein.
Die Muscheln erreichen ein verhältnismässig hohes Alter, welches sich
auf zehn bis zwölf Jahre erstrecken kann, was sich aus der Zahl der
Jahresabsätze leicht erkennen lässt. Die ersten Jahresabsätze sind
durch breite Zwischenräume getrennt, die mit fortschreitendem Alter
immer schmäler werden, und ist bei Muscheln höheren Alters der Rand
der Schalen ein häutiger, während jüngere Muscheln (Fig. 20) scharfe
Ränder haben. Häufig verändern dieselben mit zunehmendem Alter ihre
Formen derart, dass sie mit der jugendlichen Gestalt nur noch wenig
Ähnlichkeit haben.
Die Muscheln, insbesondere die Arten des Genus Anodonta, werden
häufig von einer Milbenart geplagt, die auf den Mantelhäuten des Tieres
lebt und sich vom Blute der Muschel nährt. Diese Milbe, Limnocharis
Anodontae Pfr., kriecht, sich langsam fortschleppend, auf den
schlüpfrigen Häuten des Muscheltieres herum, an welchen sie sich
mittels der Krallen an den Füssen und den Spitzen der Palpen festhält.
Die Eier befestigt sie an der Mantelhaut und zwar gewöhnlich dem
Hinterteil näher. C. Pfeiffer fand in einer Muschel 30–50
solcher Tiere. Das Muscheltier wird mager, unfruchtbar und seine
Kiemenblätter sind schlaff und befinden sich in einem der Verwesung
ähnlichen Zustande.
Die Arten der Cycladeen (Gen. Sphaerium, Calyculina und
Pisidium) sind Zwitter und gebären lebendige Junge, die beim
Abstoss aus dem Muttertiere schon eine verhältnismässig beträchtliche
Grösse erreicht haben. Die Zahl der Jungen eines Tieres ist deshalb
auch eine geringe und wird selten 20 übersteigen. Die[S. 137] Calyculinen
sind einjährige Tiere, die nur überwintern, wenn sie im Spätsommer
ausgestossen werden. Die Sphaerien und Pisidien haben eine Lebensdauer
von zwei bis vier Jahren.
Anpassungsfähigkeit der Mollusken.
Es giebt wohl keine Tierklasse, welche sich mehr an die Beschaffenheit
ihrer Wohnorte anzupassen im stande ist, als jene der Mollusken.
Die Ursache dieser Erscheinung liegt in der eigenartigen Bildung
des Gehäuses, an welches das Tier bei den Schnecken nur durch den
Spindelmuskel, bei den Bivalven durch die zwei Schliess- und den
Wirbelhaftmuskel gebunden ist. Die älteren Umgänge der ersteren
oder die älteren Schichten der letzteren fallen deshalb alsbald den
zersetzenden Einflüssen der Umgebung zum Opfer und können nicht mehr
nachgebildet werden; dennoch ist die Schale ein unentbehrlicher Teil
des Tieres, welcher den weichen Körper desselben gegen die schädigenden
Einflüsse der Umgebung schützen muss, und dessen Zertrümmerung den Tod
des Tieres zur Folge hat.
Das Gehäuse verändert sich nach den eigenartigen Verhältnissen des
Wohnortes bezüglich der Färbung, Dickschaligkeit, ja sogar bezüglich
der Form, und der Kenner wird an den Merkmalen der Schalen mit
Sicherheit auf die Beschaffenheit der unmittelbaren Umgebung schliessen
können. Die Schalen werden durch die Ausscheidungen des Mantels
gebildet, und zwar wird die Oberhaut nur vom Saume desselben, die Kalk-
und Perlmutterschichten aber, welche dieselben widerstandsfähig machen,
von den übrigen Teilen desselben ausgeschieden werden. Die Thätigkeit
des Mantels, beziehungsweise die Fähigkeit Kalkteile auszuscheiden,
erlischt erst mit dem Tode des Tieres. Die Dickschaligkeit der Gehäuse
ist deshalb als Merkmal des Alters zu betrachten. Besonders ist die
Schale der Wassermollusken Veränderungen ausgesetzt, und zwar weit
mehr als jene der Landmollusken, welche Thatsache ihren Grund in den
Eigentümlichkeiten des Mediums, in dem sie leben, nämlich des Wassers,
findet. Die chemische Zusammensetzung des[S. 138] Wassers gestattet weit
grössere Verschiedenheiten, als jene der Luft, und ebenso ist die
Zusammensetzung des Schlammes, die Bewachsung der Gewässer u. s. w.
weit veränderlicher und steht zu den Wassermollusken in engerem
Verhältnis, als die Bodenbeschaffenheit und der Pflanzenwuchs zu den
Landconchylien. Es muss deshalb den Wassermollusken ein viel grösserer
Spielraum der Variation eingeräumt werden, als den Landschnecken. Die
Schaffung einer Menge neuer Arten von Wasserschnecken und Muscheln,
wie sie zurzeit, vorzugsweise von französischen Autoren, beliebt wird,
ist deshalb unbedingt zu verwerfen. Wer die Wassermollusken längere
Zeit im Freien beobachtet, wird sehr bald zur Überzeugung kommen,
dass fast jeder einzelne Fundort derselben eigenartige, mehr
oder weniger ausgeprägte Abweichungen vom Typus der bezüglichen Art
erzeugt, und dass es geradezu zu den allergrössten Seltenheiten gehört,
zwei ziemlich übereinstimmende Formen von verschiedenen Fundorten
zu konstatieren. Ja, sogar derselbe Fundort erzeugt bei geänderten
Verhältnissen andere Varietäten, und oft genug finden sich verschiedene
Formen ein und derselben Art an sich berührenden Stellen desselben
Gewässers, wenn die Beschaffenheit des Grundes, die Strömung des
Wassers, die Bewachsung u. s. w. sich ändert. So kommen in den grossen
Seen der Voralpen Schnecken und Muscheln mit ausgeprägtem Seecharakter
und solche, welche nicht oder kaum von jenen zu unterscheiden sind,
die in Sümpfen leben, neben einander vor, und zwar jenachdem die
bezüglichen Wohnplätze bei seichtem Wasser und mangelnder Bewachsung
der vollen Wirkung des Wellenschlages ausgesetzt sind, oder die Ufer in
sumpfige Stellen übergehen.
Manche Arten ziehen fliessendes, andere stehendes Wasser vor; nur
wenige Spezies jedoch bewohnen ausschliesslich das eine oder das
andere, obwohl hie und da auch einmal eine Art sich an ihr nicht
zusagenden Stellen halten kann. Velletia lacustris ist eine nur
in stehenden, sumpfigen Gewässern sich aufhaltende Spezies; dennoch
habe ich sie in einem kleinen Bächlein bei Jettingen im Mindelthal
(Bayern) mit Ancylus fluviatilis zusammen an Steinen sitzend
gefunden. Allerdings war dieses Bächlein aus[S. 139] kleinen sumpfigen
Pfützen, welche durch kurze Strecken raschfliessenden Wassers mit
steinigem Grunde verbunden war, zusammengesetzt.
Fig. 21. Limnaea stagnalis Typ.Fig. 22. Limnaea stagnalis v. producta Colb.Fig. 23. Limnaea stagnalis v. borealis Brgt.Fig. 24. Limnaea stagnalis v. turgida Mke.
Die Veränderlichkeit der Schalen ist bei den Wassermollusken eine sehr
beträchtliche. Als Beispiel für dieselbe möge Limnaea stagnalis
(Fig. 21) herausgegriffen werden. Diese Art findet sich als var.
subulata West. in sehr schlanker Form, mit langem, spitzem Gewinde
und wenig erweitertem letzten Umgange; als var. producta Colb.
(Fig. 22) mit ähnlichem Gewinde, aber sehr aufgeblasener letzter
Mündung. Beide Varietäten finden sich in wenig mit Wasserpflanzen
durchwachsenen Altwassern oder Weihern mit nicht sumpfigem Boden. In
stark mit Wasserpflanzen besetzten stehenden Wassern, welche mehr mit
faulenden Pflanzenstoffen gemischten Schlamm am Grunde haben, bilden
sich Formen mit kürzerem Gewinde und aufgeblasenem letzten Umgange:
var. turgida Mke. (Fig. 24), roseolabiata Wolf, und
ist bei dieser Varietät die Spindel meist rosenrot gefärbt. Ähnlich
gestaltet ist var. borealis Brgt. (Fig. 23), nur nehmen die
Umgänge rascher an Breite zu und ist deshalb das Gewinde weniger spitz
ausgezogen. Gehäuse mit sehr verkürztem Gewinde und starker Schale
(var. lacustris, Fig. 26 S. 140) finden sich an den Ufern der
grossen Seen, wo die Tiere, dem Wellenschlage ausgesetzt, sich an
den Steinen oder am Boden[S. 140] festklammern müssen, und wo ihnen in der
Nahrung sehr viel Kalk geboten wird. Zwischen diesen Varietäten finden
sich Zwischenformen aller Art, so dass sich fast eine fortlaufende
Reihe allmählicher Übergänge zwischen den extremsten Formen herstellen
lässt. Gerät L. stagnalis bei Hochwasser in des Pflanzenwuchses
entbehrende Lachen am Ufer grosser Flüsse, so bilden sich sogenannte
„Hungerformen“; die Tiere verkümmern, wachsen langsam und nehmen
deshalb die Umgänge gleichförmiger zu; als solche Hungerformen mögen
var. arenaria Colb. (Fig. 25) und var. aquarii Colb.
gelten.
Fig. 25. Limn. stagnalis v. arenaria Colb.Fig. 26. Limn. stagnalis v. lacustris Stud.
Fig. 27. Limn. tumida Held.
Ähnlich wie L. stagnalis verhalten sich die übrigen Arten
des Genus; ja es haben sich aus Limnaea auricularia G. im
Laufe der Jahre in den Seen sogar eigenartige festschalige Arten
(Limn. tumida Held [Fig. 27] und Limn. rosea Gall.)
ausgebildet, welche für ihre Wohnorte charakteristisch geworden
sind, und welche durch die Unregelmässigkeit ihrer Formen sozusagen
die Eigentümlichkeiten ihrer Wohnorte dokumentieren. Limnaea
peregra, welche sich auch in fliessenden Wassern findet, ist
die formenreichste aller Arten ihres Genus, weil sie eben in beiden
Gattungen unserer Gewässer vorkommt.
Die Planorbis-Arten mit ihrer flachen, tellerförmigen Schale, die
sehr leicht Verwerfungen der Windungsebene ausgesetzt sind, finden
sich nahezu ausschliesslich in mit Wasserpflanzen durchwachsenen
stehenden Wassern, wo sie an den Stengeln und Blättern der Pflanzen
herumkriechen. Die verhältnismässig grosse platte Schale, in welcher
ein kleines Tier steckt, bietet den Fluten eine zu grosse Fläche dar,
die denselben zu leicht zum Opfer fallen und an Orte transportiert
werden würde, wo sie nicht die ihrer Organisation entsprechenden
Verhältnisse[S. 141] findet. Deshalb fehlen in den grossen Alpenseen alle
Arten mit Ausnahme von Plan. albus, welche nur vier, höchstens
fünf Umgänge erreicht. Wie hart übrigens der Kampf ums Dasein sich für
diese Art gestaltet, beweist die Verwerfung der Umgänge, welche bei ihr
in den Seen zur Regel wird (Plan. deformis Hartm.).
Aber auch an anderen Arten des Genus finden sich zuweilen Verwerfungen
der Schalenfläche in grosser Zahl. Ich habe einmal Plan.
contortus L. in einer ausgetrockneten Pfütze fast durchaus mit
verkrüppelter Schale gesammelt. Die Tiere hatten sich beim Verschwinden
des Wassers in den Boden eingewühlt und sich hier so lange lebend
erhalten, bis sich die Pfütze wieder mit Wasser füllte, und die
Tiere wieder aus ihren Schlupfwinkeln hervorkriechen konnten. Beim
Verkriechen in den Schlamm wurden die noch weichen, in der Bildung
begriffenen Ansätze der Umgänge beschädigt und aus ihrer normalen Lage
gedrängt und dadurch wurde die Fläche des Gehäuses uneben und die
Gewinde verschoben sich.
Der interessanteste Fall von massenhaften Gehäuseverkrüppelungen der
sonderbarsten Art wurde von Prof. Piré in Magné (Belgien)
beobachtet. Hier fand sich in einem kleinen Teiche massenhaft Plan.
marginatus Drap. (Fig. 28) vor. Die ganze Oberfläche des Teiches
war aber mit Wasserlinsen bedeckt, die einen dichten Filz bildeten. Die
Tiere mussten sich durch denselben durchwinden, um an die Oberfläche
zu kommen und um dort Luft zu atmen. Beim Durchwinden durch den
Linsenfilz wurden aber die weichen, neugebildeten Umgänge abgestreift
und aus der normalen Windungsebene gedrängt, und es bildeten sich
nicht nur kegelförmige Gehäuse, sondern auch solche, bei welchen die
Umgänge nach ganz verschiedenen Richtungen gedrängt wurden, und welche
die sonderbarsten unregelmässigsten Formen[S. 142] annahmen (Fig. 29). In
Linsenfilzen, welche in einem Kübel mitgenommen wurden, fanden sich am
anderen Morgen alle normal gewundenen Exemplare tot am Boden des Kübels
liegend, während die skalariden und abnorm gestalteten munter an der
Oberfläche des Wassers herumkrochen.
Da die Arten des Genus Planorbis bezüglich ihrer Wohnorte auf
einen engeren Kreis beschränkt sind, ist die Formveränderlichkeit
der einzelnen Arten auch bei weitem keine so grosse, als bei den
Limnaea-Arten, nur Plan. corneus macht in dieser Beziehung eine
Ausnahme.
Ganz besonders für flutendes Wasser gebaut sind: Die Ancylus-Arten, die
durch den breiten Fuss, mit dem sie sich an Steine u. s. w. anklammern
können, und die mützenförmige Schale den Wellen am leichtesten
Widerstand leisten können; ferner die Neritina-Spezies, die mit weiter
Mündung und dem wenig hervortretenden Gewinde von der Natur zum
Aufenthalt im flutenden Wasser besonders ausgestattet wurden. — Die
Färbung und Zeichnung der Oberfläche der Neritina-Arten wechselt sehr
mannigfaltig nach der Beschaffenheit des Wassers; doch liegen noch
keine genaueren Beobachtungen in dieser Richtung vor.
Die grösste Anpassungsfähigkeit besitzen die Muscheln der Familie
der Najaden. Jeder Fundort derselben hat bezüglich der chemischen
Zusammensetzung des Wassers, des Schlammes und des Bodens, in dem
die Muscheln stecken, der physikalischen Verhältnisse des Wassers
u. s. w. eine unbeschränkte Zahl von Eigentümlichkeiten, welche die
Schalenbildung beeinflussen und an derselben ihren Ausdruck finden.
Selbst an einander stossende Fundorte erzeugen ganz verschiedene
Formen, jenachdem der Boden steinig oder schlammig, jenachdem
der Schlamm ein erdiger oder humusreicher ist. Werden, wie es in
grösseren Flüssen häufig vorkommt, durch Hochfluten Muscheln aus
Altwassern oder Abschnitten mit stehendem Wasser in das Flussbett
versetzt, so entstehen notwendigerweise Mischformen. Leider werden
diese Verhältnisse viel zu wenig beachtet; gewöhnlich werden nur neue
Varietäten beschrieben, ohne dass man den Umständen nachforscht,
welche[S. 143] dieselben veranlasst haben. In den Beiträgen zur Molluskenfauna
der bayrischen Seen (Corresp.-Blatt zoolog.-mineral. Ver. Regensburg
1873–75) habe ich den Versuch gemacht, die eigenartigen Formen der
Seemuscheln aus der Beschaffenheit ihrer Wohnorte zu erklären, aber ich
habe bisher wenig Nachfolger gefunden.
Im allgemeinen ziehen die Anodonta-Arten stehende Gewässer vor. Nur
in diesen finden sie ihre volle Entwickelung (als var. cygnea
L.). — In fliessenden Wassern findet sich meist nur die kleine,
gewissermassen verkümmerte Varietät var. anatina L., welche
durch schmale Jahresansätze, dunkle Färbung der Oberhaut u. s. w.
charakterisiert ist. Die grösste Form, var. cygnea L., findet
sich in Weihern mit erdig-schlammigem Boden. Diese Varietät zeichnet
sich durch rundlich-eiförmige Gestalt, durch die feste Schale, die
lebhaft gefärbte Epidermis und reines glänzendes Perlmutter aus; sie
erreicht bis 190 mm Länge. Anodonta rostrata Kok. ist
die Varietät, welche sich in Altwassern mit tiefem Humusschlamm am
Grunde bildet. Die Muscheln haben eine verlängerte Gestalt mit breitem
abgestutzten Hinterteile, eine meist dunkle Färbung der Epidermis und
mehr oder weniger fettfleckiges Perlmutter. Bei zunehmender Versumpfung
der Altwasser werden die Muscheln dünnschaliger, die Wirbel werden
kariös, von Insekten angebohrt, das Perlmutter wird noch schmutziger,
die Tiere verlieren die Fähigkeit sich fortzupflanzen, und sterben
deshalb bald an dem betreffenden Wohnorte völlig aus. Ich suche den
Grund dieser Erscheinung in der Überhandnahme der Humussäure im
Bodenschlamm. Auch mit zunehmendem Alter verändern die Anodonten
ihre Umrissform. Die jungen Muscheln haben in der Regel mehr eine
rundliche Gestalt und scharf hervortretendes Schild und Schildchen.
Später wird dieselbe länglicher, die vortretenden Ecken verschwinden
und es bilden sich die Varietäten: cellensis, rostrata,
ponderosa und anatina. Nur in Flüssen erhält sich
zuweilen die rundlich-eiförmige Gestalt var. piscinalis Nils.
länger, wenn auch hier die Ecken des Schildes und Schildchens mehr
zurücktreten.
Die bisher aufgeführten Varietäten haben eine sehr ausgedehnte
Verbreitung, so dass man sie fast mehr als Standorts-Formen, denn[S. 144]
als Varietäten betrachten könnte, da sie sich in mehr oder weniger
übereinstimmender Weise überall entwickeln, wo sie die ihre Form
bedingenden Verhältnisse finden. Die grossen Alpen- und Voralpenseen
der bayrischen Hochebene und der Schweiz erzeugen dagegen, ihren
eigentümlichen physikalischen Verhältnissen u. s. w. entsprechend,
eigenartige Formen, von denen sogar fast jeder See eine oder mehrere
ihm eigentümliche Varietäten enthält. Die Seevarietäten erreichen
meist nur eine geringe Grösse, haben hellgefärbte Epidermis, starke
Schale, reines Perlmutter und ist meistens das Vorderteil durch dicke
Ablagerungen der Perlmutterschicht ausgezeichnet. Dieselben bilden
sich aber gewöhnlich nur an solchen Stellen der Ufer, die bei seichtem
Wasser und mangelnder Bewachsung der vollen Wirkung des Wogenschlages
ausgesetzt sind. Hier kann sich keine tiefere Schlammschicht, die den
Muscheln Schutz gewährt, anhäufen und werden deshalb dieselben oft
genug von den Wellen mit lebendem Tiere aufs Trockene geworfen, wo sie
verschmachten und Vögeln zur Beute fallen. Ich möchte als Beispiel
einer solchen Seemuschel hier nur An. callosa aus dem Chiemsee
erwähnen, da die Aufzählung aller mir aus den verschiedenen Seen
bekannt gewordenen zu weit führen würde.
Fig. 30. Unio pictorum.
Auch die Arten des Genus Unio nehmen in den Seen besondere
Formen an. Ich habe beobachtet, dass die Muscheln von Unio
pictorum (Fig. 30) und batavus, wenn sie im tieferen Wasser
in einer hohen Schlammschicht sich aufhalten, ein sehr verlängertes
Hinterteil erhalten, weil sich dieselben im festen Boden festklammern,
dabei aber mit dem Hinterteile aus dem Schlamme hervorzuragen[S. 145]
suchen, um die Atemröhre freizuhalten. Das Hinterteil wird dadurch
bei fortwährendem Strecken des Tieres verlängert, und krümmt sich
dabei oft mehr oder weniger nach abwärts, so dass die Muschel eine
etwas hakenförmige Gestalt annimmt, wie es bei Unio arca Held
aus dem Chiemsee und Unio platyrhynchus Rossm. (Fig. 31) aus
dem Wörthsee der Fall ist. Merkwürdigerweise kommt in den bayrischen,
Schweizer und wahrscheinlich auch in den oberösterreichischen
Seen nur Unio pictorum vor, während Unio batavus in
denselben fehlt, obwohl diese letztere Art in den zufliessenden
Bächen reichlich vorhanden ist. In den Schweizer Juraseen findet sich
Unio tumidus, die in allen im Alpengebiete liegenden grossen
Wasserbecken nicht vorkommt. Unio batavus dagegen ist auf die
Kärntner- und Juraseen beschränkt.
Fig. 31. Unio platyrhynchus Rossm.
Die Mollusken der Tiefenfauna.
Den Untersuchungen Dr. Forels[LXVII], welcher die Tiefenfauna der
grossen Schweizer Seen untersucht hat, verdanken wir die Kenntnis,
dass auch die Klasse der Mollusken zu derselben ihr Kontingent stellt,
und dass sich auch in den grössten Tiefen derselben noch einzelne
Arten von Schnecken und Muscheln aufhalten. Unter den ersteren sind
sogar Lungenatmer des Genus Limnaea, welche im seichten Wasser
bei heiterem, warmem[S. 146] Wetter die Gewohnheit haben, an die Oberfläche
des Wassers aufzusteigen, was aus einer Tiefe von 2–300 m zur
Unmöglichkeit wird.
[LXVII] Matériaux pour servir à l’étude de la Faune profonde du
lac Léman. Lausanne 1874.
Dr. Forel teilt die Fauna der Seen in drei Abteilungen:
Die Uferfauna; sie umfasst die Tiere, welche sich an der
Oberfläche des Wassers und in einer Tiefe bis zu 5 m aufhalten.
Die pelagische Fauna, welche jene Tiere umfasst, die
entfernt von den Ufern oder untergetaucht im Wasser leben.
Die Tiefenfauna, welche den Seeboden von 25–30 m an
abwärts bewohnt.
Die Uferfauna enthält Arten aller Genera unserer heimischen
Süsswasserconchylien, welche unter Umständen eigenartige Varietäten
bilden, die wir in vorhergehenden Abschnitten schon erwähnt haben.
Die pelagische Fauna entbehrt der Mollusken. Die Tiefenfauna dagegen
besitzt noch einige Arten der Genera Limnaea, Vivipara,
Valvata und Pisidium, die ich sämtlich, soweit
sie mir bis jetzt bekannt wurden, in meiner „Molluskenfauna von
Österreich-Ungarn und der Schweiz“ S. 768–791 beschrieben habe.
Die eigentümlichen Verhältnisse am Seeboden, geringe Temperatur des
Wassers (wenig um 4° C. schwankend), der grosse Druck der
Wassersäule, die sehr spärliche Nahrung, welche ihnen der Schlamm des
Seebodens bietet, geben die Veranlassung, dass die in so grossen Tiefen
lebenden Conchylien nur kleine, verkümmerte, unscheinbare Arten sind,
die sich naturgemäss von den Arten der Uferfauna abgezweigt haben
müssen. Diese Tiefseemollusken führen ein kümmerliches Dasein. Da die
Temperatur des Wassers das ganze Jahr über eine sehr gleichförmige ist
und der Wechsel der Jahreszeiten das geringe Wachstum der Schalen nicht
unterbricht, fehlen die Marken der Jahresabsätze; die Epidermis ist
sehr dünn und stösst sich, trotz der fast völligen Ruhe des Wassers,
leicht ab; die Muscheln bleiben dünnschalig und leichtzerbrechlich.
Eine Art Pisidium fragillimum Cless. aus dem Silvaplaner See
besitzt eine so dünne Schale, dass jede Berührung an[S. 147] derselben einen
Eindruck zurücklässt. Jeder der bisher untersuchten Seen beherbergt
wenigstens eine eigentümliche Art des Genus Pisidium. Limnäen
wurden nur im Genfersee beobachtet; Vivipara (eine Art) findet
sich im Gardasee; Valvata-Arten (drei) gehören nur der Tiefseefauna des
Genfer- und des Gardasees an.
Höhlen-Mollusken.
In den Kalkgebirgen, vorzugsweise in den Juraformationen, finden
sich ausgedehnte Höhlen, in denen Bäche und im Karstgebiete sogar
Flüsschen auf weite Strecken unterirdisch dahinfliessen. Es ist
selbstverständlich sehr schwierig, diese unterirdischen Wasserläufe
auf ihre Fauna zu untersuchen; gewöhnlich sind die in denselben
lebenden Mollusken nur in leeren Gehäusen im Geniste oberirdischer
Bäche und Flüsse zu bekommen. Nur in seltenen Fällen ist es
geglückt, lebende Tiere zu erbeuten, welche sämtlich einem für die
Höhlenfauna eigentümlichen Genus Vitrella Cless. (Fig. 32),
Bythiospeum Brgt., angehören. Die Untersuchungen derselben
durch Wiedersheim[LXVIII]
und Rougemont[LXIX] haben ergeben,
dass diese Höhlen-Mollusken blind sind, ebenso wie die Tiere anderer
Tierklassen, welche die gleichen Aufenthaltsorte bewohnen. Die
Tiere, welche seit vielen Generationen nur im Dunkeln leben, haben
die Augen nicht mehr nötig. Rougemont hat lebende Tiere der
Vitrella (Rougemonti Cless.) aus dem Brunnen des
Anatomiegebäudes in München heraufgepumpt, welche gleichfalls augenlos
waren.
[LXVIII] Beiträge zur Kenntnis der württ. Höhlenfauna.
[LXIX] Etudes des faunes des eaux privées de lumière.
Fig. 32. Vitrella pellucida.
Die unterirdischen Wasserläufe werden fast ausschliesslich von
Vitrella-Arten bewohnt, von denen jeder derselben eine ihm
eigentümliche Art zu besitzen scheint. Ausser diesen kleinen,
zierlichen, am meisten an das Genus Hydrobia erinnernden
Deckelschnecken fand sich in der Uracher Höhle (schwäbische Alp in
Württemberg)[S. 148] eine Ancylus-Art, Ancylus fluviatilis, und in
Krainer Höhlen einige Valvata-Spezies. Diese Arten sind kleine,
verkümmerte Formen mit dünner, farbloser Schale und geben dieselben
somit wieder ein merkwürdiges Beispiel von der Anpassungsfähigkeit der
Mollusken.
Die Perlenmuschel.
Die kalkarmen Bäche unserer Urgebirgsformationen beherbergen eine
grosse und sehr dickschalige Muschel, Margaritana margaritifera
(Fig. 33), welche einen sehr wertvollen Schmuck, nämlich die Perlen,
liefert. Die Muscheln stecken oft in sehr grosser Anzahl in sandigen
Stellen so völlig im Sande eingesenkt, dass nur an den flottierenden
Cirren der Atemöffnung sich das Vorhandensein der Muscheln erkennen
lässt.
Die Perlenmuscheln haben in ausgewachsenem Zustande eine nierenförmige
Gestalt und erreichen eine Länge von 120 mm. Sie haben eine
dunkle, fast schwarze Oberhaut, sind in der Regel um die Wirbel stark
zerfressen und ihr Perlmutter ist gewöhnlich durch schmutzig-gelbe
Fettflecken verunziert.
Fig. 33. Margaritana margaritifera.
Die Erzeugung von Perlen vollzieht sich in dem Raume zwischen Mantel
und Schale und muss die Perle hier frei beweglich bleiben, so dass
sie ständig in rollender Bewegung erhalten wird. Eine Perle bildet
sich nur dann, wenn ein kleiner fremder[S. 149] Körper, ein Sandkörnchen,
ein Stückchen eines abgestorbenen Schmarotzertieres u. s. w. an die
erwähnte Stelle gerät. Der Druck, welchen dieser fremde Körper auf die
äussere Mantelfläche ausübt, veranlasst eine stärkere Ausscheidung
des Perlmutterstoffes, welcher sich in Schichten um denselben legt
und allmählich den fremden Körper umhüllt. Die Entstehung der Perlen
ist also gewissermassen eine zufällige, und deshalb kommt nach
Beobachtungen aus den bayrischen Perlenbächen auf etwa 95–100 Muscheln
nur eine Perle. Aber nicht einmal alle Perlen sind brauchbar und
wertvoll, sondern nur jene, welche weisse Farbe und schönen Glanz
haben. Man unterscheidet drei Klassen brauchbarer Perlen und zwar 1.
Klasse: ganz helle, weisse Perlen von schönstem Glanze; 2. Klasse:
weisse Perlen von minder vollkommenem Glanze; 3. Klasse: sogenannte
Sandperlen, welche noch so viel Glanz und weisse Farbe besitzen, um
verwertet werden zu können. Eine gute Perle ersten Ranges kommt nach
v. Hessling[LXX] auf 2701, eine Perle mittlerer Qualität auf 2215
und eine schlechter Qualität auf 103 Muscheln. Ausser diesen weissen
Perlen finden sich aber auch, und in grösserer Häufigkeit als diese,
solche von brauner und von schwarzer Farbe. Zusammengesetzte Perlen von
Stäbchenform sind sogar häufig zur Hälfte braun, zur anderen Hälfte
schwarz gefärbt. Die dunklen Perlen werden als „unreif“ bezeichnet,
obwohl diese Benennung durchaus nicht zutreffend ist, weil auch grosse
Perlen die dunkle Farbe behalten. Die Ursache dieser Erscheinung ist
jedenfalls in der Nahrung der Tiere zu suchen, welche ja auch das
fettfleckige, unreine Perlmutter der Schalen erzeugt. Die aus der
Urgebirgsformation kommenden Gewässer haben in der Regel eine dunkle
Färbung, welche durch eine starke Beimischung von Humussäure erzeugt
wird, und dieses die Bildung wertvoller Perlen sehr beeinträchtigende
Verhältnis wird sich wohl nicht beseitigen lassen.
[LXX]Theod. v. Hessling: „Die Perlmuscheln und ihre
Perlen“. Leipzig 1859. Ich bin im ganzen den Ausführungen dieses Autors
gefolgt.
Versuche, um auf künstlichem Wege Perlen zu erzeugen, beziehungsweise
durch Einschieben kleiner Kügelchen u. s. w.[S. 150] zwischen Mantel und
Schale das Tier zur Perlenbildung zu veranlassen, haben keine günstigen
Resultate ergeben.
Die Ernte der Perlenbäche gilt in Deutschland durchaus als
Staats-Regal; sie wird aber gewöhnlich an Private verpachtet, welche
das Fischen und die Behandlung der Muscheln ohne jede Kenntnis der
Eigentümlichkeiten derselben betreiben und dadurch den Bestand an
Perltieren arg schädigen.
[S. 151]
Die deutschen Süsswasserfische und ihre Lebensverhältnisse.
Von Dr. A. Seligo in Heiligenbrunn bei Danzig.
[S. 153]
Herrscher im Wasser ist der Fisch. Es giebt kaum irgend einen
Wasserorganismus, der ihm nicht direkt oder indirekt Nutzen bieten
muss. Das Wasser ist auch ausschliesslich das Element, in welchem die
Fische dauernd zu leben vermögen. Zwar können nicht wenige Fische
ausserhalb des Wassers eine mehr oder minder kurze Zeit am Leben
bleiben, — es giebt, besonders in den Tropen, sogar Fischarten, welche
freiwillig an das Land gehen[LXXI], — aber auch in diesen Fällen kann
das Luftmeer nur vorübergehend mit dem Wasser vertauscht werden, und
die Fische müssten zu Grunde gehen, wenn man ihnen die zeitweilige
Rückkehr in das Wasser verwehrte.
[LXXI] z. B. die Labyrinthfische und Salarias scandens
Ehrenberg.
Ist das Vorhandensein des Wassers die erste Lebensbedingung des
Fisches, so ist in zweiter Linie die Beschaffenheit des Wassers in
Betracht zu ziehen. In dieser Beziehung sind namentlich die mittlere
Wärme des Wassers, sein Luftgehalt und sein Gehalt an anderen gelösten
Stoffen für die Arten der Fische als Lebensbedingungen massgebend.
Das natürliche Wasser kommt auf der Erdoberfläche nirgends in
chemischer Reinheit vor. Der grösste Teil des irdischen Wassers, das
Meerwasser, enthält bekanntlich etwa 3.5% an Kochsalz und zahlreichen
anderen Salzen in Lösung. Da die meisten Fischarten des Meeres im
süssen Wasser bald sterben, anderseits die Süsswasserfische[S. 154] im
Meerwasser meist nicht lange aushalten, so scheidet die Stärke des
Salzgehaltes im Wasser, dessen die Fischarten bedürfen, diese in
Süsswasserfische und Meerfische.
Eine Anzahl von Fischarten ist allerdings im stande, von Zeit zu
Zeit den Aufenthalt in der einen Wasserart mit dem in der andern zu
vertauschen. Diese als Wanderfische bezeichneten Arten folgen bei dem
Wechsel ihres Lebenselementes dem mächtigen Fortpflanzungstriebe.
Das süsse Wasser enthält in der Regel noch 0.004 bis 0.02%
Salze, meist Kalksalze, in Lösung. Bringt man die Fische in ganz
salzfreies destilliertes (wenn auch lufthaltiges) Wasser, so tritt der
Tod in wenigen Stunden ein, indem die Gewebe der vom Wasser direkt
bespülten Organe, namentlich der Kiemen, quellen und funktionsunfähig
werden[7]. Eine geringe Menge im Wasser gelöster Salze ist also
für das Leben auch der Süsswasserfische nötig, welche den
ausschliesslichen Gegenstand dieser Schilderung bilden werden.
Der unbeschränkten Ausbreitung der Süsswasserfische stehen im
allgemeinen die Grenzen der von ihnen bewohnten Gewässer, nämlich das
feste Land und das Meer, entgegen. Das letztere wird nicht nur von den
eigentlichen Wanderfischen, sondern auch von einigen anderen Arten,
welche gegen den Salzgehalt minder empfindlich sind, gelegentlich
passiert[LXXII] und dient daher ausnahmsweise zur Verbreitung solcher
Arten[8]. Auch das Land, welches die Flusssysteme trennt, ist
keine absolute Schranke für die Fische. Die Übertragung der Fischeier
durch Wasservögel und Landtiere, Überschwemmungen niedriger Teile der
Wasserscheiden, unter Umständen auch dauernde geologische Veränderungen
der letzteren ermöglichen die Verbreitung der Fischarten aus einem
Flusssystem in ein benachbartes. Hierzu kommen die allmählichen
Veränderungen, welchen die Konturen des Festlandes im Laufe der
geologischen Perioden unterworfen sind und welche die weitläufige
Trennung von[S. 155] ursprünglich eng verbundenen Landmassen bewirken, während
sie anderseits Länder, die von einander entfernt gelegen haben, durch
Landbrücken mit einander verbinden können. Dies sind die Umstände,
welche die Verbreitung der Fischarten des süssen Wassers herbeizuführen
pflegen.
[LXXII] Es kommt auch vor, dass einzelne Seefische, welche den
Aufenthalt im Süsswasser vertragen, sich gelegentlich in die Ströme
verirren und in diesen weit aufwärts schwimmen, z. B. die Flunder,
die Lamprete. Indessen sind diese Fische nicht zur Süsswasserfauna zu
rechnen, vielmehr als Meerfische zu betrachten.
Das Gebiet, mit dessen Süsswasserfischen wir uns hier zu beschäftigen
haben[9], möge so begrenzt sein, dass es die Flusssysteme,
welche vom Rhein bis zur Memel an den Südküsten der Nord-
und Ostsee münden, sowie das Donaugebiet[10] umfasst.
Ausserhalb des so umschriebenen Gebietes liegt von deutschen
Ländern nur der zum Etschgebiet gehörige Teil von Tirol[11]. Das so
umgrenzte deutsche Fischgebiet gehört bezüglich seiner Fischarten
dem europäisch-nordasiatischen Gebiete an, liegt also nach
Sclaters[12] zoogeographischer Einteilung in der paläarktischen
Region. Unter den Fischarten des Gebietes sind daher am stärksten
vertreten die Familien der in dieser Region so verbreiteten Cypriniden
und Salmoniden.
Der Ursprung dieser Familien ist ein fast entgegengesetzter zu nennen.
Die Cypriniden bilden etwa ein Dritteil aller bekannten
Süsswasserfische der Gegenwart. Günther[13] nimmt an, dass
sie ihren Ursprung in der Alpenregion genommen haben, welche die
gemässigten und tropischen Teile Asiens scheidet. Von hier breiteten
sie sich nach Norden und Süden, nach Osten und Westen aus. Australien
nebst Celebes und die übrigen ozeanischen Inseln, sowie Südamerika
wurden von ihnen nicht erreicht. In der Gegend unseres Gebietes fanden
sie sich schon in der Tertiärzeit vor. Die Salmoniden dagegen
scheinen ihren Ursprung im kalten Norden genommen und während der
Eiszeit sich in einzelnen Vertretern weit nach Süden verbreitet zu
haben. Die meisten Arten finden sich auch jetzt in den nördlichen
Teilen unserer Hemisphäre und auch die Arten unseres Gebietes
beschränken sich fast durchgehends auf kühle Gegenden der Gewässer.
Nicht gering an Zahl sind in unserem Gebiet auch die Vertreter der
Familie der Perciden. Diese Familie ist weit verbreitet im
Süsswasser und in den Meerküstengegenden aller[S. 156] Regionen. Ihre Reste
findet man in den Ablagerungen seit der Tertiärzeit.
Die Zahl der im Gebiet vertretenen Familien der Fische beträgt
vierzehn, aus ihnen gehören hierher vierzig Gattungen mit siebzig bis
achtzig Arten. Die überwiegende Zahl gehört, wie überall im Süsswasser
zu den Knochenfischen.
Aus der Familie der Perciden kommen vor die Gattungen der
Barsche, Zander, Streber und Kaulbarsche. Der Flussbarsch
(Perca fluviatilis L.)[LXXIII] ist nicht nur durch das ganze Gebiet
verbreitet, sondern findet sich durch ganz Europa, Nordasien und
Nordamerika. Er ist bei uns einer der gemeinsten Fische, und fehlt
kaum in irgend einem Tümpel. Der Zander oder Schill
(Lucioperca sandra C.) ist ein östlicher Fisch, welcher sich
von Osteuropa aus nach Westen verbreitet zu haben scheint. Er findet
sich ursprünglich nicht im Rheingebiet und im Gebiet der Weser.
Obwohl er in den Seen, in denen er vorkommt, vortrefflich wächst und
daher durchaus nicht als ausschliesslicher Flussfisch bezeichnet
werden kann, so findet man ihn doch in zahlreichen von den von ihm
bewohnten Hauptströmen weit abgelegenen Seen desselben Flussgebietes
nicht, was darauf schliessen lässt, dass seine Ausbreitung spät nach
dem Ende der Eiszeit, wenn auch während des Bestehens der Verbindung
zwischen Elbe, Oder und Weichsel erfolgt ist. Eine nahe verwandte
Art ist L. volgensis Pall., die sich in der Donau und ihren
grossen ungarischen Nebenflüssen, sowie in den übrigen Flüssen des
pontisch-kaspischen Gebietes findet. Ganz auf die Donau, bezw. auf
das pontische Gebiet beschränken sich die Arten der Streber
(Aspro zingel C. und A. streber Syb.), welche
gelegentlich auch in den Zuflüssen der obern Donau gefunden werden.
Von den Kaulbarschen[S. 157] ist der gemeine Kaulbarsch (Acerina
cernua L.) durch das Gebiet, durch ganz Mitteleuropa und Sibirien
verbreitet. Auch er fehlt bei uns kaum in irgend einem Gewässer. Der
ihm verwandte Schrätzer (Acerina schrätzer L.) dagegen
findet sich ausschliesslich in den Zuflüssen des Schwarzen Meeres, also
auch im Donaugebiet.
[LXXIII] Da für die vorliegende Abhandlung nur ein im Verhältnis
zu dem weiten Umfange des Themas geringer Raum zur Verfügung gestellt
werden konnte, so musste auf die Beschreibung der einzelnen Fischarten
sowie auf Abbildungen verzichtet werden. Man findet mehr oder weniger
ausführliche Beschreibungen in den im Litteraturverzeichnis angeführten
Werken von Heckel und Kner, von Siebold, Benecke u. a.; neuerdings sind
mehrere Werke[14] erschienen, welche eine zum Bestimmen der Fischarten
bequeme Übersicht der Hauptmerkmale bieten, sowie Auszüge aus dem
Beneckeschen Werke[15], welche die meisten deutschen Fische in guten
Abbildungen geben.
Während die Perciden ziemlich gleichmässig im Salzwasser und in
den süssen Gewässern verbreitet sind, gehört die Familie der
Cottiden fast ausschliesslich dem Meere an. Ein Vertreter
dieser Familie, der Kaulkopf (Cottus gobio L.), lebt
auch in den süssen Gewässern der paläarktischen Region, während er im
Meere nur in der salzarmen Ostsee östlich von Gotland vorkommt. Wenig
verschieden von ihm ist der C. poecilopus Heck.[16], welcher
sich in den Gewässern der Karpathen aufhält, sonst nur noch aus den
Pyrenäen bekannt ist. Gleichfalls aus den Küstengegenden des Meeres
stammt die Familie der Stichlinge (Gasterosteiden), welche
meist Bewohner des Seewassers wie des Süsswassers sind. Wir haben in
unseren süssen Gewässern zwei Stichlingsarten, von denen die kleinere
(Gasterosteus pungitius) vornehmlich in den süssen Gewässern der
Küstengegenden vorkommt, die grössere (G. aculeatus) auch die
mehr im Binnenlande belegenen Gewässerteile bewohnt, ohne dass beide
Arten indessen sich ausschliessen. Man unterscheidet bei beiden Arten
je zwei Varietäten, von denen die eine (trachurus) an den Seiten
des Schwanzes ebenso wie an den Körperseiten Knochenplatten trägt und
längere Stacheln besitzt, während die zweite (leiurus) kleinere
Stacheln und einen unbewehrten Schwanz hat. Beide Stichlingsarten
sind durch das ganze Gebiet mit Ausnahme der Donau und ihrer Zuflüsse
verbreitet. Über unser Gebiet hinaus findet sich der kleine Stichling
an allen Küsten der Nordmeere, der grosse Stichling durch ganz
Europa mit Ausnahme des pontischen Gebietes, sowie in Algier und in
Nordamerika[17].
Zu einer echten Seefischfamilie, den Dorschen (Gadoiden), gehört
ferner ein anderer unserer verbreitetsten Fische, die Aalquappe
oder Rutte (Lota vulgaris C.), welche sowohl in Flüssen[S. 158]
und Bächen, als auch in tieferen Seen der paläarktischen Region sich
überall verbreitet findet. Als einzigen Vertreter einer überaus
zahlreichen Familie des Süsswassers, der Siluriden, besitzen
wir den Wels oder Schaiden (Siluris glanis L.),
der durch Osteuropa bis zum Rhein und in Nordasien verbreitet ist.
Die Siluriden bilden nach Günther ein Vierteil aller bekannten
Süsswasserfische. Ihre Heimat ist anscheinend in Ostindien zu suchen;
von dort haben sie sich durch die süssen Gewässer fast aller Gegenden,
besonders aber in den Tropen, verbreitet.
Von Cypriniden[18] besitzt unsere Fauna, abgesehen von
lokalen Varietäten und Bastardformen, etwa dreissig Arten. Man hat die
zahlreichen Arten der Cypriniden zu Gruppen zusammengestellt. Zu der
Gruppe der Cypriniden gehören der Karpfen, die Karausche, die Barben
und die Gründlinge.
Der Karpfen (Cyprinus carpio L.) stammt anscheinend
aus Südosteuropa, wo er im Gebiete des Pontus und des Caspisees bis
weit nach Mittelasien hinein wild lebt. In Europa, neuerdings auch in
Nordamerika ist er durch die Fischzucht jetzt weit verbreitet. Die
Teichwirtschaft, welche in Böhmen sich besonders stark entwickelt hat,
hat mehrere Varietäten erzeugt. Zu diesen gehört der Lederkarpfen,
welcher keine Schuppen trägt, der Spiegelkarpfen (C. rex
cyprinorum), welcher an jeder Körperseite nur eine Reihe sehr
grosser Schuppen trägt, der blaue Karpfen, der Goldkarpfen (Carpe
d’or), dessen rötlicher Schimmer nach Carbonnier von der
Lachsfarbe seines Fleisches herrührt, der galizische Karpfen u. a.
Die Karausche (Carassius vulgaris Nils.) ist über das
ganze Gebiet wie überhaupt in der paläarktischen Region verbreitet.
Sie bewohnt stehende und langsam fliessende Gewässer mit weichem
Grunde. Als Giebel bezeichnet man im Gegensatz zu der hochrückigen
sog. Seekarausche die schlankeren Formen, welche sich in kleinen
Gewässern entwickeln. Eine nahe verwandte Karauschenart, vielleicht
nur eine Abart unserer gewöhnlichen Karausche, ist der aus Japan und
China stammende Goldfisch (Carassius auratus), der in
zahlreichen Varietäten (Teleskopfisch, Schleierfisch) jetzt auch in
Europa gezogen wird[19].
[S. 159]
Zu den artenreichsten Gattungen der Süsswasserfische gehören die
Barben, von denen man etwa 200 meist tropische Arten kennt. In
unserem Gebiet ist allverbreitet nur die auf Mitteleuropa beschränkte
Flussbarbe (Barbus fluviatilis Ag.), die Seen und Flüsse
bewohnt. Eine andere Art (Barbus Petenyi Heck.) ist in den
Karpathenflüssen, auch in der Weichsel gefunden worden. Neuerdings
glaubt man sie auch in der Lohe, einem Oderzufluss, aufgefunden zu
haben[20]. Der Gründling (Gobio fluviatilis C.) ist über
ganz Europa verbreitet, wo er in fliessenden und stehenden Gewässern
vorkommt. Eine verwandte Art, Gobio uranoscopus Ag., bewohnt die
Nebenflüsse der Donau, sowie einzelne Gewässer der obern Weichsel.
Aus der Gruppe der Rhodeina kommt bei uns ein typischer
Vertreter, der Bitterling (Rhodeus amarus Bl.), vor, der
über ganz Europa verbreitet ist.
In reicherer Zahl finden sich in Deutschland die Abramidina,
zu denen die Bressenarten, der Blei, Rapen, Uklei, die Ziege und
das Moderlieschen gehören. Der Bressen oder Brachsen
(Abramis brama L.) findet sich in ganz Mitteleuropa, mit
Ausnahme der Alpen. Ebenso verbreitet ist die Zärthe (A.
vimba L.), doch scheint sie vom Rheingebiet ausgeschlossen zu sein.
Einen viel engeren Verbreitungsbezirk haben der Seerüssling
(A. melanops H.) und der Pleinzen (A. sapa),
welche auf das pontische Gebiet beschränkt sind. Die Zope (A.
ballerus) findet sich in den Unterläufen der Ströme und in den
grossen Seen im Gebiet allenthalben. Sehr gemein in Seen und Flüssen
ist der Blei oder Güster (Blicca Björkna L.), der
in Mittel- und Nordeuropa vorkommt. Seltener, aber in den grösseren
Gewässern des Ostens ebenfalls überall verbreitet, ist die Ziege
oder der Sichling (Pelecus cultratus L.); westlich von
der Oder scheint dieser Fisch zu fehlen. Dagegen ist der Uklei
oder die Laube (Alburnus lucidus Heck.) über ganz
Mitteleuropa bis nach Frankreich verbreitet. Nicht weniger verbreitet,
aber selten und vielfach übersehen ist der Schneider (A.
bipunctatus Bl.). Dagegen ist die nahe verwandte Mairenke
(A. mento Ag.) auf das pontische[S. 160] Gebiet beschränkt. Der
Rapen oder Schied (Aspius rapax Ag.) und
das Moderlieschen, Mutterlosken oder Motken
(Leucaspius delineatus Sieb.) sind dagegen im Gebiete überall zu
finden.
Die grosse Gruppe der Leuciscina enthält ebenfalls mehrere Arten
des Gebietes, die Plötzen, Rotaugen, Döbeln, Orfen, Schleien, Nasen,
Elritzen und Strömer. Die Plötze (Leuciscus rutilus L.)
ist im ganzen Gebiet wie in ganz Mittel- und Nordeuropa verbreitet und
einer der gemeinsten Fische. Dagegen ist der Frauennerfling
(L. virgo Heck.) auf die Donau beschränkt, während der
Frauenfisch (L. Meidingeri Heck.) zu jenen Bewohnern
der tiefen Alpenseen gehört, welche nur zur Zeit der Laichablage
gefangen werden können, sonst aber ihr Leben in unzugänglichen Tiefen
verbringen. Eine ähnliche Verbreitung wie die Plötze hat das mit ihr
oft verwechselte Rotauge oder die Rotfeder (Scardinius
erythrophthalmus L.). Auch die Orfe (Idus melanotus
Heck.) ist über das ganze Gebiet verbreitet. Eine schöne Varietät
derselben ist die Goldorfe (var. miniatus). Der Döbel
oder Aitel (Squalius cephalus L.) und der Häsling
oder Hasel (S. leuciscus L.), sowie die Elritze
oder Pfrille (Phoxinus laevis Ag.) sind ebenfalls im
Gebiete, namentlich in fliessenden Gewässern, überall zu finden.
Sehr sporadisch trifft man dagegen den Strömer (Telestes
Agassizii Val.) an, der ausser im Rhein und in den Zuflüssen
der Donau neuerdings auch in einem kleinen Oderzufluss am Zobten
aufgefunden ist[21]. Die Nase (Chondrostoma nasus L.)
ist ein osteuropäischer Fisch, welcher in den Flussgebieten der Nordsee
mit Ausnahme der Elbe (wie der Zander) fehlt. Eine andere Nasenart,
C. Genei Bon., welche im allgemeinen auf Südeuropa beschränkt
ist, wird von Siebold auf Grund eines gelegentlichen Vorkommens
zur Fauna des Rheins gerechnet.
Die Schleie (Tinca vulgaris C.) ist ein in ganz Europa
verbreiteter Fisch weichgründiger Gewässer.
Zweifelhaft ist es, ob man zu den Cypriniden auch die kleine Gruppe
der Acanthopsiden oder Schmerlen zu rechnen[S. 161] hat, die sich
namentlich durch ihre knöcherne Schwimmblasenhülle und durch den Bau
ihrer Unterschlundknochen von den ihnen sonst nahestehenden Cypriniden
unterscheiden. Aus unserer Fauna gehören hierher die Steinbeisser,
Schlammpeitzker und Schmerlen. Der Steinbeisser (Cobitis
taenia L.) findet sich durch ganz Europa, die Schmerle
(C. barbatula L.) auch in Asien, der Schlammpeitzker
(C. fossilis L.) endlich in Asien und dem östlichen Europa mit
Einschluss unseres Gebietes.
Eine ganz isolierte Stellung nimmt die kleine Familie der Umbriden
ein, welche nur aus zwei Süsswasserarten besteht, von denen die eine
im mittleren Nordamerika, die andere, der Hundsfisch (Umbra
Crameri Müll.), in einigen Nebengewässern der unteren Donau
(Neusiedler See, Plattensee u. a.) und anscheinend auch in anderen
Teilen des Pontischen Gebietes sich vorfindet.
Die Familie der Salmoniden[22] ist bei uns durch fünf
Gattungen vertreten, deren Arten grossenteils zur Varietätenbildung
neigen, sodass eine Übereinstimmung unter den Fischkundigen über die
Abgrenzung der Arten in mehreren Fällen noch nicht erzielt ist. Die
meisten Arten sind als Sport- und Speisefische hochgeschätzt und werden
als Edelfische bezeichnet. Die Gattungen, welche hierher gehören, sind:
die Maränen oder Renken, die Aesche, der Stint, die Saiblinge und die
Forellen und Lachse.
Es ist schon erwähnt, dass die Salmoniden sich anscheinend von
Norden her verbreitet haben, dass sie noch jetzt im Norden die
stärkste Artentwickelung besitzen und in unserem Gebiet meist kühle
Wohnplätze aufsuchen. Solche finden die Bewohner der Seen in den
sehr tiefen Seen der Alpen und in einigen norddeutschen Seen, in
Tiefen, in welchen beständig eine Temperatur von nur 2–6° C.
herrscht. Grösstenteils Bewohner solcher Seen sind die Maränen oder
Coregonen. Nüsslin[23] hat die Arten derselben nach der
Form der Schnauze und der Bezahnung der Kiemenbögen geschieden; nach
diesem System hat man zu unterscheiden: den Nordseeschnepel
(Coregonus oxyrhynchus L.), einen Wanderfisch, der die
Nordsee bewohnt und ihre Ströme zur Laichzeit aufsucht, die
kleine Maräne (C. albula L.), die in den[S. 162] tieferen
Seen der baltischen Seenplatte von Holstein bis nach Russland
hinein, sowie in den skandinavischen Seen lebt, den Kilch
oder Kropffelchen (C. hiemalis Jur.) aus der Tiefe
des Bodensees und des Ammersees, den Ostseeschnepel (C.
lavaretus L.), der die Ostsee bewohnt und in den Buchten und Haffen
derselben laicht, die Madümaräne (C. maraena Bl.) aus dem
Madüsee in Pommern, die Bodenrenke (C. fera Jur.) aus den
tiefen schweizerischen, oberösterreichischen und bayrischen Seen, die
Pulsseemaräne (C. generosus Peters)[24] aus dem Pulssee
in der brandenburgischen Neumark[25] (Ostseeschnepel, Madümaräne,
Bodenrenke und Edelmaräne werden von Anderen für Varietäten einer Art
gehalten)[26], ferner den Blaufelchen (C. Wartmanni
Bl.) aus den tieferen nordalpinen Seen, die Traunseemaräne
(C. Steindachneri Nüssl.), Pfäffikoner Maräne (C.
Sulzeri Nüssl.) aus dem Traunsee bezw. Pfäffikoner See, und den
Gangfisch (C. macrophthalmus Nüssl.) aus dem Bodensee
(die drei letztgenannten Arten werden von anderer Seite für Varietäten
des Blaufelchen gehalten).
Die Aesche (Thymallus vulgaris Nils.) ist ein anderer
Salmonide, der kleine, raschfliessende Flüsse im ganzen Gebiete bewohnt
und über dasselbe hinaus durch Europa verbreitet ist; verwandte Formen
finden sich in Nordasien und Nordamerika. Der Stint (Osmerus
eperlanus L.) findet sich an den Küsten des nördlichen Teiles des
Atlantischen Ozeans und in dessen Zuflüssen, in denen er laicht. Im
Rhein ist er nicht beobachtet worden. In einigen norddeutschen Seen
kommt er ebenfalls vor, ohne zum Meere zu wandern.
Die naheverwandten Gattungen der Saiblinge, Lachse und Forellen hat
man nach Siebolds Vorgange nach der Bezahnung des in der
Gaumendecke liegenden Pflugscharbeins unterschieden. Das Pflugscharbein
der Saiblinge (Salmo) hat eine bezahnte Platte, aber einen
unbezahnten Stiel; das der Lachse (Trutta) hat einen bezahnten
Stiel bei unbezahnter Platte, während bei den Forellen (Trutta)
sowohl Stiel wie Platte bezahnt sind. Der Saibling (Salmo
salvelinus L.) bewohnt die tiefen Gebirgsseen Mittel- und
Nordeuropas. Der ebenfalls zu den Saiblingen gerechnete Huchen[S. 163]
(S. hucho L.) kommt ausschliesslich im Donaugebiete vor. Der
Lachs (Trutta salar L.) bewohnt den nordatlantischen
Ozean, mit Ausschluss des Schwarzen Meeres und des Mittelmeeres,
und steigt in die Flüsse, welche sich in die von ihm bewohnten
Meere ergiessen, zum Laichen auf. Die Forellen unterscheidet man
als Bachforelle (T. fario L.), Seeforelle
(T. lacustris L.) und Meerforelle (T. trutta
L.). Die beiden letzteren Arten sind ursprünglich wohl Abarten der
Bachforelle[26], aber durch verschiedene Lebensweise und körperliche
Abweichungen von ihr unterschieden. Während die Bachforelle in
raschfliessenden Bächen im ganzen Gebiete lebt, bewohnt die Seeforelle
die tiefen Gebirgsseen, die Meerforelle die Nord- und Ostsee; alle
drei Arten laichen aber ausschliesslich in Bächen, in welche die See-
und die Meerforelle zu diesem Zweck aufsteigen. Die Meerforelle hat
also eine ähnliche Lebensweise wie der Lachs, mit dem sie deshalb oft
verwechselt wird.
Die Familie der Hechte (Esociden) ist bei uns durch den
allbekannten, in allen süssen Gewässern Europas, Nordasiens und
Nordamerikas lebenden Esox lucius L. vertreten.
Aus der Familie der Heringe (Clupeiden) sind zwei Wanderfische
zu unserer Fauna zu rechnen, der Maifisch (Alosa vulgaris
C.) und die Finte (Alosa finta C.). Ersterer bewohnt
die Küstengegenden im nördlichen Atlantischen Ozean, die letztere
verbreitet sich noch mehr südlich und östlich bis zum Nil. Beide
besuchen zum Laichen die Süsswasserströme. Die Weichsel wird indessen
nur von der Finte besucht. Zur Familie der Muräniden gehört
unser Aal (Anguilla vulgaris Flem.), der in allen Flüssen
lebt, die in den Nordatlantischen Ozean gehen, mit Einschluss des
Mittelmeergebietes, mit Ausschluss aber der Pontischen Flüsse.
Wenden wir uns nun von den Knochenfischen zu den Ganoiden, so
finden wir die Familie und Gattung der Acipenserinen[27] in
mehreren Arten vertreten. Der Stör (Acipenser sturio
L.) ist ein Wanderfisch und bewohnt den Nordatlantischen Ozean
mit Ausschluss des Mittelmeeres und seiner Nebenmeere, also auch
des Schwarzen Meeres. Das letztere wird dagegen von mehreren
verwandten Arten bewohnt. Es sind dies der Glattdick[S. 164] (A.
glaber Heck.), der Scherg (A. stellatus Pall.),
der Dick (A. schypa Güldenst.), der Waxdick
(A. Güldenstädtii Brandt) und der Hausen (A.
huso L.). Alle diese Störe wandern zur Laichzeit in die
Flüsse, die letztgenannten in die Donau und die anderen Ströme des
Pontusgebietes, der Stör in die europäischen und amerikanischen Flüsse
seines Wohngebietes, um hier zu laichen. Mehr Standfisch ist der
Sterlet (A. ruthenus L.), welcher die Pontischen Flüsse,
ausserdem aber auch die in das Eismeer mündende Düna bewohnt und das
Meer in der Regel nicht aufsucht.
Aus der Ordnung der Cyclostomen endlich, deren von den übrigen
Fischen völlig abweichender Bau Anlass gegeben hat, diese Tiere
von der Klasse der Fische ganz auszuschliessen, gehören zu unseren
Süsswasserfischen zwei Vertreter der Familie der Petromyzontiden, das
Flussneunauge (P. fluviatilis L.), ein Wanderfisch,
der zum Zweck der Laichablage aus dem Meere in die Süsswasserströme
wandert, wo seine Larve mehrere Jahre lang aufwächst, und das
Bachneunauge (P. Planeri Bl.), welches sein ganzes Leben
in Bächen und kleinen Flüssen zubringt. Beide Arten sind durch die
ganze arktische Region verbreitet.
Überblickt man die jetzige Ausbreitung unserer Fischarten, so lassen
sich zwei Hauptrichtungen der Verbreitung erkennen, eine aus Nordwesten
bezw. Nord und West kommende, und eine aus Südost bezw. Süd und
Ost kommende. Der ersteren ausschliesslich gehören die Fische des
nordatlantischen Küstengebietes an: die Stichlinge, Stint, Lachs,
Nordseeschnepel, Meerforelle, Maifisch, Finte, Aal und Stör. Alle
diese Fische sind von dem Gebiet des Schwarzen Meeres (Donaugebiet)
ausgeschlossen, während demselben ausschliesslich angehören:
Wolgazander, die Streber, Schrätzer, Seerüssling, Pleinzen, Mairenke,
Frauennerfling, Hundsfisch, Huchen, Glattdick, Dick, Scherg, Waxdick
und Hausen. Eine kleine Reihe von anderen Fischen weist auf allmähliche
Verbreitung von Ost nach West hin: der Wels, der seine Westgrenze
im Rhein hat, die Zärthe, die westlich der Weser sich nicht findet,
der Zander und die Nase, deren Verbreitung nach Westen mit der Elbe
abschliesst, endlich die Ziege, welche nicht über die Oder hinaus nach
Westen geht.[S. 165] Noch andere Fische finden sich im Gebiet stellenweise,
so aus dem Süden eingewandert der Barbus Petenyi, der Gobius
uranoscopus, der Strömer, das Chondrostoma Genei, aus
dem Osten der Sterlet, ferner an gewisse Örtlichkeiten gebunden der
karpathische Kaulkopf, der Ostseeschnepel, endlich die auf die tiefen
Seen beschränkten Arten: der Frauenfisch, die kleine Maräne, die
übrigen Felchen- und Renkenarten, der Seesaibling und die Seeforelle.
Die übrigen Fische sind dem ganzen Gebiete gemeinsam. Eine kleine
Zahl von ihnen ist über die ganze arktische Region (Nordasien, Europa
und Nordamerika) verbreitet: Barsch, Aalquappe, Hecht, Fluss- und
Bachneunauge. Einige finden sich allgemein im Norden der alten Welt:
Kaulbarsch, Kaulkopf, Karausche und Bachforelle. Die übrigen gehören
dem europäischen Gebiete nördlich von den Alpen an und sind teilweise
bis Asien hinein verbreitet, nämlich die meisten Cypriniden, die
Acanthopsiden sowie die Aesche.
Haben wir uns in dem bisherigen über die in unserm Gebiet vorkommenden
Arten und ihre Verbreitung orientiert, so wenden wir uns nun zur
Betrachtung der Lebensverhältnisse derselben, welche wir am besten an
der Hand ihres Körperbaues[28] und ihrer Organe kennen lernen.
Die äussere Körperdecke, die Haut, ist wie bei den höheren
Wirbeltieren eine doppelte, indem sie aus der Oberhaut oder Epidermis
und der Lederhaut oder dem Corium zusammengesetzt ist. Die Oberhaut
besteht aus einer mehrschichtigen Lage von Zellen, deren äusserste
Schichten zerfallen und in Gemeinschaft mit dem Schleim einzelner
grosser Drüsenzellen die Oberfläche der Haut schlüpfrig machen. Die
unter der Oberhaut liegende Lederhaut besteht aus Bindegewebsfasern
und ist meist sehr zähe (Aal). In taschenförmigen Vertiefungen dieser
Haut liegen die Schuppen[29], Hornplättchen, welche sich meist
dachziegelartig decken und einen dichten Schutzpanzer bilden (Perciden,
Cypriniden, Salmoniden, Clupeiden, Hecht). Bei dem Hundsfisch ist
auch der Kopf mit Ausnahme der Schnauze mit Schuppen bedeckt, während
derselbe bei den übrigen Fischen frei von Schuppen ist. Bei manchen
Fischen sind die Schuppen so fein und die Oberhaut so dick, dass[S. 166]
die Schuppen nicht ohne Weiteres erkannt werden können (Aal, bei dem
sie in Zickzacklinien liegen, Aalquappe, Schleihe, Schmerlen). Die
Stichlinge tragen an Stelle der Schuppen an den Seiten schmale dünne
Knochenschienen, welche zusammenhängende Seitenpanzer bilden. Der
Körper der Störe ist mit starken, mit scharfen Höckern versehenen
Hautknochen als wirksamem Schutz besetzt. Ganz ohne Hautbewehrung sind
von unseren Fischen die Kaulköpfe, der Wels und die Neunaugen.
Der Silberglanz, welchen die meisten Fische zeigen, wird dadurch
hervorgerufen, dass die Oberfläche der Lederhaut (bezw. die Innenseite
der Schuppen) mit einer Lage von mikroskopisch kleinen, krystallartig
geformten Plättchen bedeckt ist, welche neben Kalk auch Guanin[30]
enthalten[LXXIV]. Letzterer Stoff findet sich auch in der glanzlosen Haut
der Neunaugen[31]. Bei manchen Fischen bringen die Glanzkörperchen,
namentlich zur Laichzeit, schöne Interferenzfarben hervor (Stichling,
Bitterling u. a.). Sie werden in der Farbenwirkung unterstützt durch
die Farbzellen (Chromatophoren)[32], welche in der Lederhaut
liegen. Die meisten Farbzellen sind mit schwarzem Farbstoff, viele auch
mit rotem oder gelbem Farbstoff gefüllt. Die letzteren, Zooerythrin
und Zoofulvin, kommen nach Krukenberg[33] auch bei den
Vögeln vor, fehlen aber eigentümlicherweise ganz bei allen anderen
Wirbeltierklassen. Die Farbzellen der Fischhaut haben in hohem Grade
die Fähigkeit, sich bald fast punktförmig zusammenzuziehen, wodurch
sie fast unsichtbar werden, bald sich wieder zu sternförmigen, weit
ausgebreiteten Körpern auszudehnen und damit ihre Farbe zur Wirkung
zu bringen. Auf diese Weise kann die Farbe der Fische sich der ihrer
Umgebung anpassen und dadurch den Fisch vor Verfolgern schützen
oder die Wachsamkeit seiner Beute täuschen. Dieser Farbwechsel ist
von der eigenen Lichtempfindung des Fisches abhängig; geblendete
Fische zeigen nach Pouchet[34] diese Farbanpassung nicht.
Bei manchen Fischarten, namentlich Cypriniden (Karausche, Schleihe,
Orfe, Barbe, Plötze), finden sich Varietäten,[S. 167] denen die schwarzen
Farbzellen ganz oder stellenweise fehlen, während die roten und gelben
stark entwickelt sind. Diese Varietäten werden oft als Zierfische in
Parkteichen gezogen. In seltenen Fällen sind auch die roten Farbstoffe
nicht entwickelt (Albinismus), oder die Silberglanzkörperchen fehlen
(Alampia). Normal fehlen die Glanzkörper beim Stint.
[LXXIV] Der Schuppenglanz mancher Cypriniden wird im grossen rein
gewonnen zur Herstellung der Farbe künstlicher Perlen.
Seine Stütze erhält der Fischkörper durch die ihn der Länge nach
durchziehende Wirbelsäule. Bei den Knochenfischen besteht
dieselbe aus durchbohrten bikonkaven cylindrischen Knochenstücken,
den Wirbeln, deren Innenräume durch die elastische Chorda ausgefüllt
sind. Nach oben und unten setzen sich an jeden Wirbelkörper
paarweise knöcherne Fortsätze an, die Rücken- und Bauchstrahlen. Die
Rückenstrahlen jedes Wirbelkörpers bilden einen Kanal, indem sie an
ihren oberen Enden mit einander verschmelzen. In dem so gebildeten
Kanal an der Oberseite der Wirbelsäule liegt das Rückenmark. Die
Bauchstrahlen verschmelzen nur im Schwanzteil des Fischkörpers mit
einander zu einem Kanal, der die grossen Blutgefässe des Schwanzes[LXXV]
aufnimmt. Im Vorderteil des Körpers bilden sie als Rippen die Stützen
der Seitenwände der Leibeshöhle.
[LXXV] Diese Blutgefässe sticht man beim Schlachten grosser
Fische an, die man durch Verblutung töten will.
Die Körperform der Fische ist entweder eine seitlich mehr
oder minder zusammengedrückte, oder mehr spindelförmig bis walzig.
Erstere Form zeigen am stärksten ausgeprägt der Bressen und die
Seekarausche, letztere der Aal und die Neunaugen, sowie die Aalquappe,
der Schlammpeitzker, der Wels, der Kaulkopf, lauter Fische, die
vorzugsweise am Grunde der Gewässer leben und sich gelegentlich auf
demselben schlängelnd bewegen. Die hauptsächliche Bewegungsart
unserer Fische ist aber das Schwimmen im freien Wasser, und hierzu
ist der Fischkörper nicht nur selbst in geeigneter Weise geformt,
sondern auch mit besonderen Anhängen versehen, den Flossen. Die
Flossen sind gebildet durch Häute, welche durch eingelagerte bewegliche
knöcherne Spangen ausgespannt werden können, etwa wie ein Schirm oder
ein mit Zeug bezogener[S. 168] Fächer. Man unterscheidet paarige Flossen,
welche an den Seiten des Körpers stehen, und unpaare Flossen, welche in
der Mittellinie des Rückens und des Schwanzes stehen.
Die paarigen Flossen sind meist zu zwei Paaren vorhanden, welche man
nach ihrer gewöhnlichen Stellung als Brustflossen und Bauchflossen
unterscheidet; sie entsprechen den Gliedmassen der höheren Wirbeltiere.
Sie sind an Knochen befestigt, welche bei den Brustflossen mit
dem Kopf, bei den Bauchflossen unter einander verbunden sind. Sie
fehlen ganz den Neunaugen, während der Aal nur Brustflossen, keine
Bauchflossen hat. Die unpaaren Flossen unterscheidet man je nach
ihrer Lage als Rückenflossen, Schwanzflossen und Afterflossen.
Rückenflosse und Afterflosse sind dadurch am Fischkörper befestigt,
dass jeder Strahl der Flosse scharnierartig verbunden ist mit einer
Knochenschiene von kreuzförmigem Querschnitt, welche im Körper des
Fisches liegt und sich je an einen Rückenstrahl bezw. Bauchstrahl der
Wirbelsäule anlehnt. Rückenflossen sind in Zweizahl vorhanden bei den
Perciden, Aalquappe, Kaulkopf und den Neunaugen. Bei den Stichlingen
sind die ersten Strahlen, ebenso wie die Bauchflossen, zu einzeln
stehenden, teilweise zackigen Stacheln umgewandelt, welche als Waffen
dienen. Bei den Salmoniden findet sich hinter der Rückenflosse eine
kleine sogenannte Fettflosse, welche diese Familie von allen anderen
einheimischen Fischen leicht unterscheiden lässt. Diese Fettflosse wird
als ein Überbleibsel aus dem Larvenstadium des Fisches betrachtet;
sie hat keine festen knöchernen Strahlen, wie die übrigen Flossen,
sondern an deren Stelle nur weiche hornige Fäden, wie alle Flossen der
Neunaugen, der Haie und Rochen und der eben ausgeschlüpften Jungen der
Knochenfische. Die Afterflosse fehlt nur den Neunaugen. Sie beginnt
stets kurz hinter der Afteröffnung. Die Schwanzflosse liegt bei den
meisten Fischen mit dem grössten Teil unterhalb der Wirbelsäule. Die
trotzdem vorhandene Symmetrie des Schwanzes wird dadurch ermöglicht,
dass die letzten Wirbelkörper zu einem Stiel verschmolzen und so nach
oben gebogen sind, dass die Unterseite der Wirbelsäule und die an sie
sich ansetzenden Schwanzflossenstrahlen nach hinten gerichtet sind.
[S. 169]
Sehen wir uns nun nach den Muskeln um, welche die Bewegungen
des Fisches bewirken, so finden wir zunächst zu beiden Seiten der
Wirbelsäule starke Fleischmassen, welche leicht einen eigentümlichen
Bau erkennen lassen. An jeden Wirbelkörper setzen sich beiderseits
Muskelplatten an, welche hohlkegelförmig gewölbt sind, so dass ein
Querschnitt durch den Fischkörper mehrere hinter einander liegende
Muskelplatten ringförmig blosslegt. Die vier an einer Stelle der
Wirbelsäule sich ansetzenden Muskelplatten bilden einen Muskelabschnitt
(Myokamma). Die Muskelabschnitte sind unter sich durch dünne
Bindegewebshäute (Ligamente) getrennt[LXXVI], die einzelnen Muskelbündel
liegen in den Muskelabschnitten in der Längsrichtung des Fisches, ihre
Enden setzen sich daher nicht an Knochen, wie die meisten Muskeln
der höheren Wirbeltiere, sondern an Ligamente an (interligamentale
Muskulatur). Die Muskelplatten bilden zusammen zwei grosse
Seitenmuskeln, welche den grössten Teil des Fischkörpers einnehmen
und die Bewegungen des Schwanzes bewirken. Neben den Seitenmuskeln
treten die Muskeln des später zu betrachtenden Kopfes und die der
Flossen an Umfang sehr zurück. Die Muskulatur der unpaaren Flossen
besteht aus zahlreichen kleinen Muskelzügen, welche einerseits an die
inneren Halteknochen der Flossen, anderseits an die Flossenstrahlen
sich ansetzen (interosteale Muskulatur) und die letzteren aufrichten,
niederziehen und seitwärtsbiegen können. Bei den paarigen Flossen wird
die Ausbreitung und die fächelnde Bewegung, welche dieselben ausführen,
durch stärkere Muskelbündel bewirkt, welche ebenfalls an den inneren
Gerüstknochen dieser Flossen befestigt sind. Wie kommt nun mittels der
Flossen und ihrer Muskeln die Ortsbewegung zu stande[35]? Die
Flossen, die paarigen sowohl wie die unpaaren, sind für sich allein
nicht im stande, den Fisch schwimmend zu erhalten, wie durch Versuche
festgestellt ist. Die paarigen Flossen halten den Fisch, wenn er im
freien Wasser[S. 170] schwimmend steht, im Gleichgewicht, auch wirken sie mit
bei Wendungen und bei der Rückwärtsbewegung sowie beim plötzlichen
Aufhalten. Die unpaaren Flossen können durch leichte wellenförmige
Bewegungen eine langsame Vorwärtsbewegung des Fisches bewirken. Das
rasche Schwimmen der Fische dagegen wird durch Ruderschläge des
Hinterleibes bewirkt, dessen Fläche, um den Widerstand des leicht
ausweichenden Wassers zu erhöhen, durch Aufrichten der unpaaren
Flossen vergrössert werden kann, wobei die Schwanzflosse hauptsächlich
als Steuer dient. An diesen Bewegungen, welche durch abwechselnde
Kontraktionen der beiden Seitenmuskeln vor sich gehen, nimmt der
hintere Teil des Körpers teil, welcher durch eine durch die Vorderenden
der Rückenflosse und der Afterflosse gelegte Ebene abgegrenzt wird. Der
Körperteil vor dieser Ebene enthält die Leibeshöhle mit ihren
Organen der Blutzirkulation, der Verdauung und der Fortpflanzung. Ein
Zwerchfell trennt die kleine Brusthöhle, welche das Herz
enthält, von der geräumigen Bauchhöhle.
[LXXVI] Beim Erwärmen des toten Fischkörpers lösen sich diese
Häute unter Leimbildung auf, sodass die einzelnen Muskelplatten sich
von einander trennen und leicht schollenartig auseinanderfallen. In den
Ligamenten liegen namentlich bei den Perciden und Cypriniden feine,
spitze Stützknochen, Fleischgräten, welche sich als Knochen beim Kochen
nicht auflösen.
Das Herz[36] liegt, vom Herzbeutel umschlossen, dicht
hinter dem Kopfe. Es besteht aus der muskulösen Herzkammer und der
dünnwandigen Vorkammer, die durch ein Klappenventil getrennt sind. Der
Vorkammer schliesst sich der Sinus venosus an, der das Venenblut
aufnimmt und der Vorkammer zuführt. Aus der Herzkammer entspringt, mit
einer Anschwellung (Bulbus aortae) beginnend, die Kiemenarterie,
welche das venöse Blut aus dem Herzen in die Kiemen führt. Aus den
Kiemen sauerstoffreich zurückkehrend sammelt sich das Blut in der
grossen Körperarterie (Aorta descendens), aus welcher es sich
in die Organe des Körpers verteilt. Das hier gebrauchte Blut wird zur
Ausscheidung der nicht gasförmigen Stoffwechselprodukte durch die Leber
und die Niere geführt. Das Produkt der Leber[37], die Galle,
sammelt sich in der Gallenblase und gelangt aus dieser in den Darm.
Das Produkt der Nieren, der Harn, wird durch die Harnkanälchen
in die beiden Harnleiter und sodann in der Regel zunächst in eine
Erweiterung des gemeinsamen Endteils derselben, die Harnblase, geführt,
von wo er durch einen Ausführungsgang hinter dem After nach aussen
gelangt.[S. 171] Bei den Fischen liegen die Nieren an der Decke der
Bauchhöhle als zwei dicht an der Wirbelsäule durch die ganze Länge der
Bauchhöhle sich erstreckende, grossenteils mit einander verschmolzene,
dunkelrote, sehr weiche Organe. Sie dienen bei den Fischen nicht allein
zur Harnabsonderung, sondern sie sind daneben, wie auch wahrscheinlich
die zwischen den Eingeweiden liegende Milz[38], die
Vermehrungsstätten der roten Blutkörperchen[39].
Die Organe der Ernährung und ihre Hilfsorgane liegen teils in der
Mundhöhle, teils in der Bauchhöhle. Die Mundhöhle nimmt den
unteren Teil des Kopfes ein, während in dem oberen, in einer Kapsel aus
Knorpel (Neunauge, Stör) oder Knochen (bei den meisten Knochenfischen),
das Gehirn eingebettet ist. Die Knochen, welche den vorderen Teil des
Bodens des Hirnschädels bilden, sind das Dach der Mundhöhle, deren
Seitenwände und Boden teils aus den Kiemenbögen und den ihnen homologen
Knochenbögen, den Unterschlundknochen und dem Zungenbein, teils aus
den Kieferknochen, und aus ihren häutigen und muskulösen Verbindungen
bestehen. Die Öffnungen zwischen den Kiemenbögen führen in die
Kiemenhöhlen, welche nach aussen durch die mehr oder minder beweglichen
Kiemendeckel geschlossen sind.
Alle Knochen der Mundhöhle sind mit einander beweglich verbunden,
sodass die Mundhöhle bedeutender Erweiterung fähig ist. Je nach der
Nahrung des Fisches sind die Gestalt der Mundöffnung und die Bezahnung
der Mundknochen verschieden. Die Perciden, Kaulköpfe, Aalquappe, Wels,
auch manche Cypriniden, wie Rapen, Döbel, Orfe, ferner die Aesche, der
Stint, die Salmo- und Trutta-Arten, Hecht und Aal haben ein breites,
weit aufsperrbares Maul und, mit Ausnahme der Cypriniden, auf mehr
oder minder zahlreichen Knochen desselben teils nur feine Zähnchen,
die in grosser Zahl mehr oder minder dicht beisammen stehen (Sammet-,
Bürsten-, Hechelzähne, z. B. beim Aal, Barsch, Wels), teils zwischen
diesen noch grössere Fangzähne (Zander, Hecht, Lachs, Forelle)[40].
Diese Fische sowie die Stichlinge nähren sich ausschliesslich, wie
Lachs, Hecht, Wels und Zander, oder teilweise von Fischen, welche
sie, auch wenn sie über die Mundhöhle[S. 172] hinausragen, mittels der Zähne
festhalten können. Die meisten Cypriniden, Maränen und Clupeiden haben
dagegen eine kleine rundliche oder mehr hohe als breite Mundöffnung,
welche am Ende einer rüsselartig vorstreckbaren, häutigen Röhre liegt,
die oben von den halbringförmigen Zwischen- und Oberkieferknochen,
unten von dem Unterkiefer gestützt wird. Mittels dieses Saugrüssels
schlürfen diese Fische ihre aus kleinen niederen Tieren bestehende
Nahrung ein, nach welcher sie teils im freien Wasser, teils an den
festen Gegenständen in demselben, den Pflanzen, Steinen, dem Holzwerk,
oder auf dem Grunde suchen. Sie finden hier kleine Crustaceen aus den
Ordnungen der Cladoceren, der Ostracoden, der Copepoden, ferner die das
Wasser bewohnenden Larven vieler Insekten, namentlich der Mücken und
Eintagsfliegen, auch Würmer, Rädertiere, kleine Weichtiere, nehmen wohl
auch die schleimigen Massen der Kieselalgen und den für sie allerdings
unverdaulichen Mulm zerfallener Pflanzenteile, Sand und Schlamm
ein, verschonen auch nicht, wie hier gleich erwähnt sein mag, Eier
und Brut von Fischen, selbst nicht die eigene Nachkommenschaft. Man
unterscheidet die letztgenannte Gruppe von Fischen als Kleintierfresser
oder Friedfische von den ersterwähnten Raubfischen[41].
Während die Kiemenspalten der breitmäuligen Fische ziemlich weit
und nur mit weitläufig gestellten Zähnen versehen sind, sind die
Kiemenspalten der engmäuligen Fische eng, meist kurz, die Kiemenbogen
sind an der Innenseite mit je zwei Reihen dicht gestellter Stäbchen
besetzt, welche in einander greifend einen reusenartigen Verschluss
bilden, durch den wohl das in die Mundhöhle aufgenommene Wasser, nicht
aber die feinkörnige Nahrung in die Kiemenhöhle entweichen kann. Am
Gaumen vieler Cypriniden findet sich ein muskulöser Wulst, welcher
Sinnesorgane (Schmeckbecherchen) enthält. Jede Berührung dieses Organs
bringt eine Anschwellung der berührten Stelle hervor. Es scheint auch
durch seine Kontraktionen beim Aufsaugen der Nahrung mitzuwirken.
Am hinteren Abschluss der Mundhöhle haben die Cypriniden, welche
sonst ganz zahnlos sind, auf den Unterschlundknochen stumpfe aber
starke Zähne, deren bei den einzelnen Arten[S. 173] verschiedene Form ein
vorzügliches Mittel zur Abgrenzung und Erkennung der Arten ist. Diesen
Zähnen gegenüber steht am Gaumen eine harte Knorpelplatte. Zwischen
den Zähnen und der Gaumenplatte wird die Nahrung zerdrückt. Die Zähne
werden bei den Cypriniden nach v. Siebold jährlich in der
Laichzeit abgestossen und erneuert.
Abweichend von dem Maul der übrigen Fische ist das der Neunaugen
gebaut. Es ist eine am vorderen Körperende gelegene Saugscheibe, welche
mit mehreren zahntragenden Hornplatten ausgestattet ist. Es dient dem
Fisch vornehmlich dazu, sich an feste Gegenstände oder an seine Beute
anzusaugen. Die letztere wird dann mittels der Zähne angebohrt und
ausgesaugt.
Das durch die Kiemenspalten abfliessende Wasser gelangt in die
Kiemenhöhlen, in welchen sich die Kiemenbögen befinden. Die
Kiemenbögen bestehen aus gebogenen rinnenförmigen Knochenplatten mit
nach unten gekehrten Rinnen. In diesen Rinnen laufen die Blutgefässe,
welche das Blut aus dem Herzen in die Kiemen leiten, und andere, die
das Blut aus den Kiemen dem Körper zuführen. In den Kiemenblättchen,
welche an jedem Bogen in zwei Reihen dicht gedrängt stehen, tritt
das kohlensäurehaltige Blut mit dem sauerstoffhaltigen Wasser in
Gasaustausch, das Blut tritt sauerstoffreich in den Körper zurück,
während das verbrauchte Atemwasser unter dem Kiemendeckel durch die
Kiemenöffnung abfliesst und durch neues aus der Mundhöhle ersetzt
wird. In der Regel sind vier Paar Reihen von Kiemenblättchen auf jeder
Seite des Kopfes vorhanden. An der Innenseite des Kiemendeckels sitzt
häufig noch eine sogenannte Nebenkieme (z. B. beim Stör), welche aber
funktionslos ist. Die Acanthopsiden (Schlammpeitzker, Steinbeisser
und Schmerle) können im Notfalle auch durch den Darm die Atmung
vollziehen, indem sie Luft einschnappen und durch den Darm treten
lassen, wobei sie einen quietschenden Ton erzeugen. Die anderen Fische
atmen dagegen hauptsächlich durch die Kiemen. Manche Fische, welche
einen sehr fest schliessenden Kiemendeckel haben, wie der Aal und
viele Cypriniden, können stundenlang, selbst tagelang ausserhalb des
Wassers zubringen, ohne zu ersticken, indem die Wassermenge,[S. 174] welche
in den nach aussen fest geschlossenen Kiemenhöhlen zurückgehalten
ist, genügt, um die Aufnahme der durch den Mund eingeschnappten Luft
durch die Kiemen zu vermitteln. Andere Fische, namentlich die mit sehr
weiten Kiemenspalten und kurzen Kiemendeckeln versehenen Salmoniden,
sterben aus dem Wasser genommen sehr bald ab. Bei den Neunaugen weichen
auch die Kiemen von denen der anderen Fische ab. Sie sind nicht an
Kiemenbögen befestigt, sondern bestehen jederseits in sieben Säckchen,
welche durch ein festes Knorpelgerüst gestützt werden, und im Innern
mit zahlreichen Kiemenfalten, welche die Stelle der Kiemenblättchen
vertreten, bekleidet sind. Das Atemwasser wird aus der Mundhöhle durch
einen besonderen Längskanal, welcher mit entsprechenden Seitenöffnungen
versehen ist, zugeführt, und fliesst aus jedem Kiemensäckchen
durch eine besondere Öffnung nach aussen ab. Die sieben äusseren
Kiemenöffnungen samt dem Auge und dem einfachen Nasenloch sollen dem
Fisch seinen Namen „Neunauge“ gegeben haben.
Ausser den Kiemen atmen die Fische auch durch die Haut, wie A. von
Humboldt und Provençal[42] durch Versuche nachwiesen.
Kohlensäure wirkt nach diesen Forschern tödlich auf die Fische,
während Stickstoff und Wasserstoff, wie bei den höheren Wirbeltieren,
indifferent sind. In luftlosem (ausgekochtem) Wasser starben die
eingesetzten Fische nach 1¾ bis 4 Stunden. Nach den genannten
Untersuchungen berechnete Treviranus, dass die Schleihe,
deren Sauerstoffbedürfnis für gering zu halten ist, für je 100 Gran
Körpergewicht in 100 Minuten 0.01 cbcm Kohlensäure erzeugt,
während Säugetiere das fünfzigfache an Kohlensäure produzieren.
Das verschiedene Sauerstoffbedürfnis ist anscheinend, neben dem
Wärmebedürfnis, eine der Hauptursachen der Verteilung der Fischarten
auf die einzelnen Gegenden eines und desselben Flussgebietes. Der
Sauerstoffgehalt einer Wassermenge ist teils von der Temperatur, teils
von der Menge der Stoffe bezw. der Organismen abhängig, welche den
Sauerstoff zu absorbieren vermögen.
Die Temperatur übt einen wesentlichen Einfluss auf die Fähigkeit
des Wassers, Luft aufzunehmen, aus. Eine Wassermenge, welche bei
5° C. 100 Raumteile Sauerstoff aufnimmt, kann[S. 175] davon bei
20° nur etwa 79.2 Teile in Lösung halten. Man stelle sich vor, der
Sauerstoffgehalt der von den Landtieren eingeatmeten Luft nehme an
einer Örtlichkeit um etwa ⅕ ab, und man wird es natürlich finden,
dass die Lebewelt an dieser Örtlichkeit eine abweichende ist.
Viel bedeutender noch kann die Verminderung des Sauerstoffs durch
die Einwirkung oxydierbarer Substanzen werden. Untersuchungen des
Themsewassers in der Umgegend von London haben ergeben, dass das Wasser
dieses Flusses, welches etwa 5 Meilen oberhalb London bei Kingston 7.4
cbcm Sauerstoff im Liter enthält, dicht bei London davon nur 1.5
cbcm, nach dem Durchgange durch die Riesenstadt, bei Woolwich,
sogar nur 0.25 cbcm, also nur eine Spur Sauerstoff, den 30.
Teil von seinem Gehalte in der minder verunreinigten Flussgegend,
enthält[43].
Bei so enormer Verminderung des Gehaltes an Atemluft kann es nicht
in Verwunderung setzen, wenn die mit leicht oxydierbaren organischen
Substanzen gefüllten Abwässer grosser Städte und industriereicher
Gegenden den Bestrebungen zur Vermehrung und Veredelung des
Fischbestandes ein kaum zu überwindendes Hindernis entgegensetzen[44].
Nicht so jäh und verderblich, wie die Abfuhrstoffe der Städte
und Fabriken, aber sicher auch von erheblicher Wirkung auf den
Sauerstoffgehalt des Wassers sind die Reste abgestorbener Lebewesen,
der organische Mulm, welcher durch die Regen- und Schneewässer aus
dem Niederschlagsgebiet des Flusssystems dem Wasser desselben auf
seinem Laufe zum Meer in immer steigender Menge zugeführt wird. Auch
das Gefälle und die Bodenbeschaffenheit des Flussbettes sind von
Einfluss: Ein über Kiesbänke und Steine rauschender Bach bietet seinem
Wasser mehr Gelegenheit zur Sauerstoffaufnahme, als ein träges, tiefes
Gewässer, in dem noch dazu der hineingeschwemmte Mulm sich ablagert.
Die Wirkung des Sauerstoffmangels im Wasser ist für die Fische eine
doppelte: Nicht nur mangelt den Organismen, Tieren wie Pflanzen, die
notwendige Lebensluft, sondern es nimmt auch die Bildung schädlicher
Stoffe, besonders des betäubenden Sumpfgases, zu.
[S. 176]
Unter diesen Verhältnissen ist es verständlich, dass
sauerstoffbedürftige Fische sauerstoffarme Gewässer vermeiden.
Anderseits bedürfen manche Fische einer gewissen Wärme des Wassers
(z. B. der Karpfen), damit ihre Lebensfunktionen, Ernährung und
Fortpflanzung, zur Thätigkeit angeregt werden. Solche Fische sind von
dauernd kühlen Gewässern ausgeschlossen.
Man hat gefunden, dass viele Arten der Fische gemeinsame
Lebensbedürfnisse haben, so dass sie in Flussstrecken, welche eine
gewisse Beschaffenheit haben, leben können. A. Fritsch[45]
hat zuerst die Flussregionen Böhmens, welche charakteristische
Fischgesellschaften enthalten, unterschieden und nach ihren
Hauptfischen benannt. M. von dem Borne[9] hat diese Methode
für die deutschen Gewässer durchgeführt, und sie ist jetzt allgemein
angenommen. Man unterscheidet:
die Forellenregion, mit Bachforelle, Elritze, den Kaulköpfen,
Schmerle, Döbel,
die Aeschenregion, mit Aesche, Barbus Petenyi, Gründling,
Bachneunauge[46] (im obern Teile dieser Region liegen die
Laichstellen der Lachse),
die Barbenregion, mit Barbe, Huchen, Nase, Rapen, Zärthe,
Schneider, Häsling, Karpfen, Quappe, Bitterling, Mairenke, Streber,
Strömer, Motken,
die Bressenregion (Bleiregion), mit Bressen, Blei, Wels, Orfe,
Rotauge, Schleihe, Karausche, Aal.
Manche Fische finden sich in allen Regionen, wie Hecht, Barsch, Plötze,
Stichling. Auf eine einzige der angeführten Regionen beschränkt ist
kein Fisch, vielmehr werden die benachbarten Regionen auch häufig
aufgesucht.
Etwas abweichend von den fliessenden Gewässern verhalten sich die Seen,
deren Lebensverhältnisse wesentlich von ihrer Tiefe abhängen. Man
unterscheidet hier flache Bressenseen, Seen von mehr als 20 m
Tiefe, in welchen sich die kleine norddeutsche Maräne aufhält, und
Seen von über 50 m Tiefe, in welchen, je nach ihrer Tiefe,
verschiedene Maränenarten, der Seesaibling und die Seeforelle leben.
[S. 177]
Bei der Wichtigkeit des Atmungsprozesses spielt die Funktionsfähigkeit
der Kiemen eine grosse Rolle. Sobald ihre Oberfläche durch Trockenheit
abstirbt oder sobald sie sich mit einem dichten Belag von Fremdkörpern
bedeckt, sind die Fische einem raschen Erstickungstode ausgesetzt: die
Ursache, weshalb Trübungen des Wassers von den Fischen gemieden werden
und ihnen, wenn sie dauernd, z. B. durch Fabrikwässer, verursacht
werden, den Tod bringen können.
Während das in die Mundhöhle aufgenommene Wasser durch die
Kiemenspalten abfliesst, gelangt die Nahrung durch den trichterförmigen
Schlund in den eigentlichen Darmtractus[47]. Der Darm der
Fische ist mehr oder minder gewunden und bei den Cypriniden ein
Schlauch von fast überall gleicher Weite. Bei den übrigen Fischen ist
eine mehr oder minder ausgeprägte magenartige Erweiterung vorhanden.
Immer ist der Magendarm durch eine ringförmige Einschnürung, die
Pförtnerklappe, vom Mitteldarm getrennt. Die mit einem Magen versehenen
Fische, mit Ausnahme des Welses, des Hechtes und des Hundsfisches,
haben auf der Grenze zwischen Magen und Mitteldarm Blindschläuche,
wenige bei den Perciden, den Kaulköpfen und Stichlingen, viele bei
den Salmoniden und der Aalquappe. Der Enddarm ist im Vergleich mit
dem Mitteldarm weit. Der Darm der Neunaugen ist gerade und ohne
Anhänge, der des Störs trägt an der Innenwand eine spiralig verlaufende
Hautleiste (Spiralklappe). Von den dem Darm anhängenden Drüsen ist die
Leber schon erwähnt. Die zweite bei den Wirbeltieren sonst vorkommende
Verdauungsdrüse, das Pankreas, fehlt den meisten unserer Fische; es ist
bis jetzt nur nachgewiesen bei Schmerle, Stichling, Hecht, Barsch, Aal
und Forelle, meist stark zerstreut zwischen den Eingeweiden[48].
Die verdauende Flüssigkeit der Fische wird daher, da auch
Speicheldrüsen fehlen und die Galle nur bei der Aufnahme der Fette
durch den Körper mitwirkt, meist ausschliesslich vom Darm geliefert.
Dafür vermag aber auch nicht nur der Magendarm, wie bei den höheren
Wirbeltieren, sondern jeder Abschnitt, selbst die[S. 178] kurze Speiseröhre
der mit Magen versehenen Fische, Pepsin abzusondern, den Stoff,
welcher in Verbindung mit der bei den Fischen reichlich vorhandenen
Säure die Eiweissstoffe für die Aufnahme in den Körper geeignet
macht (peptonisiert[49]). Von dem Pepsin der höheren Wirbeltiere
unterscheidet sich das Pepsin wenigstens der Forelle und des Hechtes
dadurch, dass es selbst bei 0° noch verdauend wirkt[50]. Die Auflösung
der Nahrung beginnt schon in der Speiseröhre und braucht erst im
Enddarm mit dem Auswerfen der unverdauten Nahrungsteile aufzuhören. Am
kräftigsten ist die Verdauung bei den Fischfressern, minder intensiv
bei den Kleintierfressern. Die Absonderung der Nahrungsflüssigkeit
erfolgt wahrscheinlich von allen Zellen der Darmschleimhaut, die je
nach ihrem Reifestadium ein verschiedenes Aussehen haben können[51].
Von grosser Wichtigkeit, namentlich für die Fütterung der Teichfische,
ist, dass die Fische rohe Stärke, das Hauptprodukt der meisten
Pflanzen, fast gar nicht zu verdauen vermögen, während gequollene
(gekochte) Stärke verdaut wird[52]. Diese Thatsache bietet die
Erklärung zu der erst neuerdings gehörig gewürdigten Erscheinung, dass
die Fische sich fast gar nicht von Pflanzenstoffen, sondern meist von
Tieren nähren, so dass in der Regel die von den Pflanzen erzeugte
Nahrung erst in den Körper eines niedern Tieres aufgenommen sein muss,
bevor sie zur Ernährung der Fische dienen kann. Eine nur scheinbare
Ausnahme hiervon bilden die Kieselalgen (Diatomeen), welche man oft
in Menge in dem Fischdarm findet. Diese in grosser Menge im Wasser
auftretenden mikroskopischen Pflänzchen, welche eine Hauptnahrung
vieler niederer Wassertiere bilden, erzeugen nicht, wie die meisten
anderen Pflanzen, Stärke, sondern an deren Stelle Öl, das als Fett den
Verdauungssäften der Fische zugänglich ist. Ähnlich verhält sich die
Algengattung Vaucheria, die also den Fischen ebenfalls direkt
Nahrung liefern könnte.
Als Grund für das Fehlen der stärkeartigen Stoffe in der Fischnahrung
hat man angeführt, dass die Fische ebenso wie die Amphibien es nicht
nötig haben, ihren Körper mit diesen wärmeerzeugenden Kohlehydraten
gewissermassen zu heizen, da ihr Körper[S. 179] seine Wärme von dem ihn
umgebenden Wasser erhält. Immerhin bewirkt auch bei den Fischen die
Respiration eine Erwärmung des Körpers über die Temperatur des Wassers.
Dieselbe beträgt nach Broussonet bei kleinen Fischen ½ bis
⅔°, beim Aal ¾°, beim Karpfen 1°. Despretz fand bei 10.83°
C. Wassertemperatur die Körperwärme von zwei Karpfen zu 11.69°,
von zwei Schleihen zu 11.54° C.
Änderungen in der Wasserwärme haben auch den grössten Einfluss auf
die Lebensfunktionen der Fische, namentlich auf die Ernährung und
Fortpflanzung. Während die meisten unserer Fische bei steigender Wärme
laichen (Sommerlaicher: die Perciden, Kaulköpfe, Stichlinge, Wels,
Cypriniden, Acanthopsiden, Hecht, Clupeiden, Störe und Neunaugen),
legen andere ihre Eier bei sinkender Wärme ab (Winterlaicher: Aalquappe
und die Salmoniden ausser Stint, Huchen und Aesche). Manche Arten
bedürfen einer bestimmten Mindestwärme, um laichreif zu werden,
namentlich die Karpfen, welche nicht in Wasser unter 19° C.
laichen. Auch das Nahrungsbedürfnis ist abhängig von der
Wasserwärme. In kaltem Wasser können die Fische wochenlang, ja manche
Arten monatelang ohne Nahrung bestehen. Viele Cypriniden nehmen im
Winter auch im Freien keine Nahrung zu sich; der Karpfen z. B. frisst
nur, wenn das Wasser mindestens 9° C. Wärme hat, dabei verliert
er während der Zeit, in welcher er nicht frisst, nur etwa 3–5 % seines
Körpergewichtes. Bei der Bachforelle und anderen Raubfischen nimmt zwar
die Fresslust im Winter ab, hört aber nicht völlig auf. Die Forelle
lässt auch im Sommer in der Ernährung nach, wenn die Wassertemperatur
über 25° C. steigt.
Bei höher steigender Wärme sterben die Fische. Der Karpfen verträgt
eine Höchsttemperatur von 32–35° C., der Barsch eine Temperatur
von 28° C.[43]. Rasche Abkühlung vertragen viele Fische
ebenfalls nicht, allmähliches Sinken der Temperatur dagegen hat keinen
schädlichen Einfluss auf unsere Fische, welche in eiskaltem Wasser
lebend bleiben, ja, soweit sie geringe Ansprüche an Luftversorgung
machen (Schleihe, Karausche), auch einfrieren können, wenn nur die
Eiskälte nicht über die Eigenwärme des Fisches siegt[S. 180] und wenn durch
die letztere eine Wasserschicht um den Fisch flüssig erhalten bleibt
(Johannes Müller).
Bei direkter Einwirkung des Frostes auf den Fischkörper erstarren die
Fische zunächst, können aber, wenn die Frostwirkung nicht stundenlang
dauert, zuweilen wieder belebt werden. Beim Durchfrieren des
Fischkörpers sterben die Fische dagegen natürlich ab[44].
Die Menge der von den Fischen aufzunehmenden Nahrung richtet sich
während der Hauptfresszeit in erster Linie nach der Menge der
vorhandenen geeigneten Nahrungsmittel.
Die Fische können mit geringen Mengen von Nahrung erhalten werden,
haben dann aber ein entsprechend geringes Wachstum und zeigen oft auch
in ihrer Körperform Abweichungen von dem normalen Aussehen ihrer Art
(so ist die als Giebel bekannte Abart der Karausche eine Hungerform,
— den Fischzüchtern ist es längst bekannt, dass Fische mit kurzem
Kopf rascher gewachsen und deshalb geeigneter zur Zucht sind, als
Fische mit gestrecktem Kopf). Anderseits können sie sehr grosse Mengen
von Nahrung aufnehmen und dementsprechend wachsen. Man weiss aus
guten Beobachtungen, dass ein Karpfen am Ende des ersten Jahres bei
ungünstiger Nahrung nur wenige Gramm, bei günstiger Nahrung gegen 1
Kilo wiegen kann, und dass das Gewicht der Hechte im ersten Herbst
ihres Lebens zwischen 100 Gramm und mehreren Pfund schwanken kann.
Bei der Fütterung verbraucht die Forelle zur Zunahme um einen
Gewichtsteil fünf bis acht Gewichtsteile Futterfleisch, während der
Karpfen schon aus drei bis vier Gewichtsteilen in gleichem Masse
eiweisshaltigen Futters einen Gewichtsteil Körperzunahme gewinnt.
Man benützt bei der Fütterung der Fische hauptsächlich eiweissreiche
Futterarten, wie Fleischmehl, Leguminosensamen, Malztreber u. a.
Eine Anzahl von Fischarten erreicht in der Regel nur eine bestimmte
geringe Grösse; man kann diese Fische als Zwergfische bezeichnen.
Dahin gehören Kaulbarsch, Kaulkopf, Stichling, Gründling, Bitterling,
Uklei, Moderlieschen, Elritze, kleine Maräne, Stint,[S. 181] die Neunaugen.
Die meisten anderen Fische wachsen entsprechend ihrer Ernährung sehr
ungleichmässig. So erreicht die Bachforelle in den nahrungsarmen
Gebirgsbächen selten ein Gewicht von einem Pfund, während sie in
Teichen, Flüssen und Seen (z. B. im Weitsee bei Berent in Westpreussen)
es zu einem Gewicht von 12 Kilo bringen kann. Karpfen und Hechte hat
man bis 25 Kilo schwer, Welse noch bedeutend grösser gefangen. In engen
Gefässen bleibt das Wachstum der Fische beschränkt, wie die konstante
Grösse der in engen Gläsern gehaltenen Goldfische zeigt.
Über dem Darm und seinen Anhängen und Drüsen liegt die
Schwimmblase, ursprünglich eine Ausstülpung des Darmes, bald
mit dem Anfangsteil desselben durch einen im Alter oft geschlossenen
Kanal verbunden (Physostomi: Siluriden, Cypriniden, Salmoniden,
Hecht, Hundsfisch, Clupeiden, Aal, Stör), bald ohne Ausführungsgang
(Perciden, Stichlinge, Aalquappe). Die Schwimmblase fehlt dem
Kaulkopf und den Neunaugen. Sie ist ein häutiger Sack, prall gefüllt
mit einer Gasmischung aus Stickstoff und Sauerstoff in wechselnden
Mengen und etwas Kohlensäure. Diese Gase gelangen nicht etwa durch
den zuweilen vorhandenen Ausführungsgang in die Schwimmblase, sondern
sie werden von der Innenfläche derselben, wo sich oft Anhäufungen
feiner Adern, sogenannte Wundernetze, finden, ausgeschieden. Man hält
die Schwimmblase deshalb für sein Homologon der Lunge der höheren
Wirbeltiere; dem ist jedoch widersprochen worden, weil sie oberhalb,
nicht unterhalb des Darmkanals wie die Lunge, liegt[53]. Auch die
Funktion der Schwimmblase wird verschieden gedeutet. Vielfach hält man
sie für einen hydrostatischen Apparat, der den Körper im Gleichgewicht
erhält und zugleich geeignet ist, durch Kontraktionen das Steigen und
Sinken des Fisches zu regeln. Indessen glaubt Charbonnel-Salle
nachgewiesen zu haben, dass die Druckschwankungen der Schwimmblase
keinen Einfluss auf das spezifische Gewicht des Fisches und auf
sein Auf- und Niedertauchen ausüben[LXXV]. Ursprünglich[S. 182] war sie
vielleicht ein Sauerstoffreservoir (nach Biot[54] hat man bei
Tiefenfischen bis 87% Sauerstoff in der Schwimmblasenluft gefunden).
Sie steht mit dem Hörorgan in Verbindung und wird wohl auch andere
Druckwirkungen als die Schallwellen zur Empfindung bringen helfen.
[LXXV] Platzt einem Fisch die Schwimmblase und verbreitet
sich die nicht mehr unter Druck stehende Luft derselben in der
Leibeshöhle, so kann der aufgeblähte Fisch nicht mehr die Oberfläche
verlassen und schwimmt auf dem Rücken. Diese Erfahrung spricht gegen
Charbonnel-Salles Ansicht.
Neben den Organen des Darmtractus liegen in der Bauchhöhle die
Fortpflanzungsorgane der Fische, deren Produkte bei den
männlichen Fischen als Milch, bei den weiblichen als Rogen bezeichnet
werden. Danach heissen die Männchen auch Milchner, die Weibchen
Rogner. Unsere Fische sind sämtlich getrennten Geschlechtes; nur bei
einzelnen Individuen ist, wie in allen Tierklassen, gelegentlich
Hermaphroditismus nachgewiesen.
Die Männchen sind bei den Fischen oft kleiner[55] und häufig seltener
als die Weibchen. Unter den Steinbeissern hat man sogar nur 10%
Männchen gefunden[56].
Das Geschlecht ist bei manchen Fischen, besonders zur Laichzeit,
auch äusserlich durch die Beschaffenheit der Geschlechtsteile oder
durch sekundäre Geschlechtsmerkmale erkennbar. Die weiblichen Fische
zeigen in der Laichzeit einen durch die reifen Eier gewölbten Bauch,
während die Männchen schlank bleiben. Die Geschlechtsöffnung, welche
hinter dem After liegt, befindet sich beim Weibchen oft an der Spitze
einer kegelförmigen Erhöhung, die in der Laichzeit gerötet ist und
beim Bitterling zu einer 3–4 cm langen Legeröhre auswächst.
Unsere einheimischen Süsswasserfische sind sämtlich ovipar, d. h.
die Weibchen legen Eier ab; die Eier werden erst nach dem Austreten
befruchtet. Da eine innere Befruchtung der Eier anscheinend nie
erfolgt, so fehlen den Männchen besondere Organe für die innere
Begattung, ihre Geschlechtsöffnung liegt meist in der Tiefe einer
flachen Rinne. Sekundäre Geschlechtscharaktere finden sich namentlich
als lebhafte Färbung bei den Männchen vieler Fischarten, meist nur in
der Brunstzeit. So glänzen namentlich die Männchen der Stichlinge und
der Bitterlinge während der Laichzeit in bunten Farben. Bei anderen
Fischen werden die immer vorhandenen Farben des Körpers und der Flossen
lebhafter. Bei den meisten Cypriniden tritt an den Männchen in der
Laichzeit[S. 183] an den Seiten und teilweise auch am Kopfe auf jeder Schuppe
ein weisses hartes Höckerchen auf, das aus Oberhautzellen besteht
und nach der Laichzeit wieder verschwindet. Bei den Coregonen finden
sich ähnliche Hautwarzen zur Laichzeit bei beiden Geschlechtern als
Brunstmerkmale. Beim männlichen Schleih ist der zweite Strahl der
Bauchflossen stark verbreitert, verdickt und gekrümmt, gleichzeitig
ist das Stützskelett dieser Flossen etwas verstärkt. Eine ähnliche
Abweichung findet sich bei den männlichen Steinbeissern am zweiten
Strahl der Brustflossen.
Die Geschlechtsreife der weiblichen Fische tritt in der Regel im
dritten Jahre ein, seltener später oder schon im zweiten Jahre. Die
Männchen werden oft schon im zweiten Jahre laichreif. Manche Individuen
(bei den Cypriniden, Lachsen, Forellen, Stören, Aalen) bleiben ganz
unfruchtbar. Da diese Exemplare besonders fett und wohlschmeckend sind,
so hat man die Fische auch, und zwar mit dem gewünschten Zuchterfolge,
kastriert.
Die Neunaugen machen nach ihrem Ausschlüpfen aus dem Ei noch
eine lange dauernde Metamorphose durch[46]. Die Larven, Querder
(früher für eine besondere Art, Ammocoetes branchialis,
gehalten) sind blind und leben im Grunde der Bäche. Gegen den Herbst
des vierten oder fünften Jahres beginnt die Verwandlung in die Form
der erwachsenen Neunaugen, nach deren Ausbildung die Flussneunaugen,
etwa 20 cm lang, in die See hinabwandern, von wo sie nach
mehreren Jahren laichreif zurückkehren, während die Bachneunaugen ihre
Laichreife in ihren Heimatsbächen erreichen können. Die Neunaugen
laichen nur einmal in ihrem Leben, nach dem Laichen sterben sie ab.
Bei den Stören und den meisten Knochenfischen sind die Eierstöcke
und Hoden[47] paarig vorhanden, bei den Perciden und Acanthopsiden
einfach. Die Hoden besitzen besondere Ausführungsgänge, welche sich
vereinigen und gemeinsam mit den Harnleitern hinter dem After münden.
Auch die Eierstöcke besitzen meist Ausführungsgänge, welche
nach ihrer Vereinigung zwischen dem After und der dahinter liegenden
Harnöffnung nach aussen münden. Diese mit Ausführungsgängen versehenen
Eierstöcke sind im übrigen[S. 184] geschlossene Säcke, in deren Falten die
Eier sich entwickeln. Die Salmoniden, Acanthopsiden und der Aal haben
dagegen Eierstöcke, welche nur gefaltete Platten darstellen, aus denen
die Eier nach ihrer Reife in die Bauchhöhle fallen. Aus dieser treten
sie durch eine besondere Öffnung, welche die gewöhnliche Lage hat, ins
Freie. Bei den Salmoniden finden sich beiderseits vom After offene
oder nur durch Rudimente angedeutete sogenannte Abdominalporen, durch
welche die Bauchhöhle nach aussen mündet. Huxley[57] hält
diese Poren, ebenso wie ähnlich gelegene Spalten in der Leibeswand
beim Stör, für rudimentäre Eileiter, während Weber[58]
sie für rudimentäre Segmentalgänge erklärt (also den Mündungen der
Segmentalorgane der Anneliden entsprechend). Anderer Art als diese
Mündungen der Bauchhöhle sind die Öffnungen, welche beim Wels hinter
den Brustflossen sich finden und nicht in die Bauchhöhle, sondern in
Taschen, die in der Haut liegen, münden. Auch über die Bedeutung dieser
Öffnungen weiss man nichts Gewisses.
Die Samenfäden der Fische bestehen aus einem meist rundlichen
Köpfchen, welches den Kern enthält, und einer fädlichen Geissel von
etwa 0.05 mm Länge. Die Eier der einheimischen Fische sind
fast kugelig und mit mehreren Hautschichten umschlossen, welche von
zahlreichen Poren durchsetzt sind. An einer Stelle findet sich eine
etwas vertieft gelegene Verdünnung der Eihäute, die Mikropyle, durch
welche in der Regel der Samenfaden in das Ei tritt. Die Eier der
Salmoniden-Arten sind verhältnismässig gross, das Lachsei hat 5–7
mm Durchmesser. Die übrigen Fischeier haben nur 1–3 mm
Durchmesser.
Die Ablage und Befruchtung der Eier findet an dem Orte statt,
wo die Eier die ihnen zusagenden Entwickelungsbedingungen finden. Die
meisten Eier werden klebend, sobald sie in das Wasser kommen, indem
die äusserste Schicht der äusseren Eihaut, der Zona radiata,
entweder gleichmässig aufquillt oder beim Quellen in Zotten oder Fäden
zerreisst[59]. Auf diese Weise kleben die Eier an den Gegenständen
fest, auf die sie fallen, namentlich Pflanzen, Steine, Baumwurzeln,
Kies, je nach der Örtlichkeit, die die Fische[S. 185] zum Laichen aufsuchen.
Die Hechte, die am frühsten im Jahr laichen, legen ihren Laich auf
dem Grase überschwemmter Wiesen ab, die Cypriniden, die sich auf den
Laichplätzen in Scharen zusammenfinden, meist auf Wasserpflanzen,
andere auf Kiesbänken im strömenden Wasser oder am Abhang des
Ufergrundes der Seen. Die Coregonen lassen ihre Eier entweder in
die Wassertiefe sinken oder sie streuen sie an die auf Mergelboden
wachsenden Armleuchtergewächse, auf Ceratophyllum oder ähnliche starre
Wasserpflanzen. Häufig tritt bei manchen Arten nach dem Laichen eine
starke Sterblichkeit ein, z. B. bei den Maifischen.
Besondere Eigentümlichkeiten in ihren Laichverhältnissen zeigen
einige Fischarten, welche teils im Meer, teils im Süsswasser
leben. Dahin gehört namentlich der Aal[60]. Die Eierstöcke dieses
Fisches wurden zuerst von Mondini[61] entdeckt, später von
Rathke[47] von Neuem aufgefunden. Es sind zwei weisse,
gekräuselte Bänder, die sogenannten Manchettenorgane, welche zu
beiden Seiten der Schwimmblase von vorn bis hinten sich erstrecken.
Betrachtet man ein Stückchen dieser Bänder unter dem Mikroskop (bei
50–100facher Vergrösserung), so sieht man ausser vielen ungleich
grossen Fettzellen die durchsichtigen runden Eier, jedes mit einem
„Keimbläschen“ (dem Kern) im Innern. Diese Eier sind zu mehreren
Millionen in einem weiblichen Aal enthalten, werden aber im Süsswasser
nicht grösser als etwa ¼–⅓ mm im Durchmesser. Da man
männliche Individuen unter den Aalen lange Zeit nicht fand, so hielt
man die Aale für Zwitter. Man glaubte auch die Hoden in Fettwulsten
neben dem Ovar gefunden zu haben. Erst Syrski[62] fand
1874, dass beim Aal die Geschlechter getrennt sind, indem er in den
männlichen Aalen die Hoden (Lappenorgane) nachwies. Man weiss nun, dass
die Aale in den süssen Gewässern meist Weibchen sind, dass dieselben
im fünften oder sechsten Lebensjahre in die See wandern, dass sie im
Brackwasser die Männchen finden und dass die ausgewachsenen Individuen
beider Geschlechter sodann in der Tiefe des Meeres verschwinden,
ohne wiederzukehren. Darüber hinaus ist unsere Kenntnis von der
Fortpflanzung der Aale noch nicht gekommen. Ein Versuch, erwachsene
Aale in grossen[S. 186] Fischkästen in der Ostsee laichreif werden zu lassen,
führte auch zu keinem Resultat[63]. Man nimmt daher an, dass die
Aale in der Tiefe des Meeres[LXXVI] (die der Ostsee in der Nordsee)
ähnlich den übrigen Fischen den Laich ablegen und befruchten, und zwar
im Winter, und dass sie dann absterben. Im Frühjahr kommen im März oder
April die jungen Aale in ungeheuren Massen als fingerlange, schlanke,
durchsichtige Tierchen an die Küsten, und ziehen an diesen entlang
in die Ströme und bis in deren kleinste Nebenwässer. Sie wandern
immer gegen die Strömung und nur nachts, besonders bei warmem Wetter;
am Tage halten sie sich an ruhigen Stellen, zwischen Kies, unter
Steinen, im Kraut auf. Sie wachsen während ihrer Wanderung. Kommen sie
an Mühlenwehre oder andere Stauwerke, so suchen sie an schadhaften
Stellen, an denen ein wenig Wasser herabrieselt, aufzusteigen. Ihre
klebrige Oberhaut und ihr gelenkiger, dünner Körper begünstigt diese
Kletterversuche, so dass ein Teil von ihnen in der Regel die nicht zu
hohen und nicht ganz festgeschlossenen Stauwerke zu überwinden vermag.
Erst die kalte Jahreszeit scheint ihren Wanderungen ein Ziel zu setzen,
doch hat man noch bei ein- und zweijährigen Aalen Wanderungen gegen das
strömende Wasser beobachtet.
[LXXVI]Fritsch nimmt an, dass die Laichstellen in der
Gegend von Süsswasserquellen liegen, aus welchem Grunde, ist leider
nicht gesagt[64].
Ein anderer interessanter Wanderfisch ist der Lachs. Im allgemeinen
bewohnt er das Meer und steigt aus diesem in das Süsswasser auf,
um hier laichreif zu werden. Ausnahmsweise hat man gefunden, dass
Lachse auch im Meere laichreif geworden sind[65]. Anderseits haben
Fritsch[66] und Metzger[67]
beobachtet, dass
Lachsmilchner, ohne das Süsswasser verlassen zu haben, schon im zweiten
Herbst ihres Lebens laichreif geworden sind und die Eier der in ihre
Wohnbäche aufgestiegenen Lachsrogner befruchtet haben. Doch sind
beide Fälle vermutlich nur Ausnahmen. Die Zeit des Eintritts in die
Stromsysteme ist sehr verschieden. In das kurische Haff (Memel) geht
der Lachs im Mai, in die in der Nähe der Weichsel in die Danziger Bucht
mündende Rheda steigt er am stärksten im Juli auf, während er in die
Weichsel[S. 187] selbst fast ausschliesslich im Herbst eintritt. In der untern
Oder findet sich der Lachs im August und September. Im Rhein[68]
unterscheidet man nach der Zeit ihres Auftretens und nach ihrem
Körperzustande den St. Jakobsalm, der in Holland um Jakobi (25. Juli)
in den Rhein tritt und etwa 1½ k schwer und 40–50 cm
lang ist, und den Wintersalm, der viel grösser und schwerer ist und um
Mitte September in Holland erscheint.
Die Geschlechtsorgane sind beim Eintritt in das Süsswasser noch wenig
entwickelt, der Eidurchmesser etwa 0.5 mm gross, das Gewicht
der Geschlechtsorgane beträgt kaum 0.5 % des Körpergewichts, während
dieselben im reifen Zustande fast ein Viertel des Körpergewichtes
ausmachen. Im Süsswasser entwickeln sie sich allmählich und zwar
ausschliesslich auf Kosten der Rumpfmuskulatur, welche nicht nur
verhältnismässig ärmer an Fett und Eiweissstoffen wird, sondern auch
an absolutem Gewicht stark abnimmt, weil der Lachs im Süsswasser keine
Nahrung aufnimmt[69], sondern ganz auf Kosten seiner im Meere gut
genährten Organe sein Leben fristet. Das Fleisch der im Süsswasser sich
so entwickelnden Fische wird daher immer schlechter und verliert seine
rote Farbe. Gleichzeitig treten sekundäre Geschlechtscharaktere auf.
Die Haut verdickt sich namentlich am Kopfe und Rücken schwartenartig
und wird dunkler, die Männchen bekommen rote Flecken an den Seiten,
namentlich auf den Kiemendeckeln, an der Spitze ihres Unterkiefers
entwickelt sich ein knorpeliger Haken, der bei grösseren Tieren so
stark wird, dass er das Schliessen des Maules hindert. Endlich, bei den
im Herbst ins Süsswasser gelangten Fischen, meist erst im Herbst des
folgenden Jahres, tritt die völlige Laichreife ein. Die Fische suchen
nun, wie Fritsch[70] schildert, die seichten Stellen der Bäche
auf, am liebsten oberhalb stärkerer Strömungen, dort wo das Wasser sich
zu brechen anfängt. Hier wirft das Weibchen durch Schwanzbewegungen die
Kiesel des Bachgrundes zur Seite und stellt dadurch eine seichte Grube
her, die Laichgrube. Nach einiger Zeit stellen sich bei der Laichgrube
täglich morgens und abends das Weibchen und ein oder mehrere Männchen
ein. Sie liegen oft still in den Laichgruben, so dicht neben einander,
dass[S. 188] man Männchen und Weibchen mit dem (gabelförmigen) Fischspeer
zugleich spiessen kann. Beim Laichgeschäft streicht das Weibchen, indem
es den Bauch auf dem Grunde der Grube reibt, die Eier ab; das Männchen
steht etwa 1 m stromaufwärts und lässt die Milch in das Wasser,
die Milch strömt zu den Eiern und bewirkt die Befruchtung. So setzen
sie es mehrere Wochen lang fort, wenn sie nicht gestört werden.
Nach dem Laichen, in Böhmen beim Fallen des ersten Schnees,
verschwindet der Laichlachs. Er ist dann ganz abgemagert, mit schlaffem
Körper, und völlig kraftlos und wird, wenn er nicht abstirbt, von
der Strömung abwärts zum Meere getragen. Nach Miescher kommt
der männliche Lachs in der Regel zwei- bis dreimal in mehrjährigen
Zwischenräumen, der weibliche ein- bis zweimal zur Laiche in den Rhein.
Unter den kleinen Jakobsalmen sind wenige Weibchen zu finden; man
schliesst daraus, dass die Weibchen oft erst später reif werden, als
die Männchen.
Die aus den Eiern schlüpfenden Lachse werden als Salmlinge bezeichnet.
Sie halten sich in der Regel nur bis zum nächsten Hochwasser in dem
Bach, in dem sie geboren sind, auf, und gehen mit dem Hochwasser als
etwa handlange Fische in das Meer hinab. So lange sie in den Bächen
bleiben, leben sie hier ähnlich wie die verwandten Salmoarten, sind
aber nicht so lichtscheu, wie die Forellen, sondern stehen gern im
fliessenden Wasser gegen die Strömung gerichtet.
Die Forellen haben ähnliche Laichverhältnisse wie die Lachse. Die
Meerforellen und Seeforellen wandern zur Laichzeit stromaufwärts,
letztere gegebenen Falles auch stromabwärts, um in geeigneten Bächen
auf Kies- oder Sandgrund zu laichen.
Alle einheimischen Salmoniden mit Ausnahme des Stint, des Huchen und
der Aesche, laichen im Herbst oder Winter, und die Jungen schlüpfen
erst im Beginn des Frühjahres aus den Eiern. Diese bleiben also
monatelang unausgebrütet, ohne jeden Schutz, der Vernichtung durch
zahlreiche tierische Feinde, durch Wasserschimmel, durch die Abwässer
der Fabriken ausgesetzt.
[S. 189]
Um diese als Delikatesse gesuchten Edelfische nun vor der Ausrottung
zu bewahren, hat man sich in grossem Massstabe und mit viel Erfolg
eines Hilfsmittels bedient, welches ihnen den Kampf ums Dasein mit den
übrigen Wassertieren erheblich erleichtert, der sogenannten künstlichen
Fischzucht[71], durch welche nicht nur die Vermehrung der Salmoniden
in den deutschen Seen, Flüssen und Meeren, sondern auch die vieler
anderer Fische in stehenden und fliessenden Gewässern, sowie die
Übertragung von Fischarten in neue, von ihnen nicht bewohnte Gewässer
in zahlreichen Fällen erzielt ist.
Unter „künstlicher Fischzucht“ versteht man zunächst die Befruchtung
und Erbrütung von Fischeiern unter Zuthun des Menschen. Ein Detmolder
Landwirt, Jakobi, erfand diese Methode in der Mitte des vorigen
Jahrhunderts, doch wurde sie wegen der mühsamen Bedienung des im
Bache stehenden Brutkastens selten angewendet. In Norwegen, Russland,
namentlich aber in Frankreich wurden später ähnliche Verfahren
entdeckt. Professor Coste in Paris interessierte sich dafür,
und auf seine Veranlassung wurde 1852 von der französischen Regierung
die Brutanstalt in Hüningen bei St. Ludwig im Elsass gegründet, welche
1871 vom Deutschen Reiche übernommen wurde. Von Deutschen ist zuerst
in München ein Bruthaus für künstliche Fischerbrütung angelegt.
Inzwischen ist die Methode besonders in Amerika, in neuerer Zeit auch
in Deutschland weitergebildet und sehr vervollkommnet worden[72], und
jetzt zählen die Anstalten für künstliche Fischzucht in Deutschland
nach hunderten.
Zur künstlichen Befruchtung werden die Eier des reifen Weibchens,
welche lose im Ovar, bezw. in der Bauchhöhle liegen, durch gelindes
Streichen herausgedrückt und in einer Schale aufgefangen, eine kleine
Menge Samen, welcher in gleicher Weise aus den Hoden eines Männchens
herausgestrichen ist, wird auf die Eier gebracht und durch vorsichtiges
Umrühren zwischen die Eier verteilt, Wasser hinzugegossen und dann
die Schale mit ihrem Inhalt für kurze Zeit sich selbst überlassen.
Die Samenfäden fangen ihre Schwärm- und Bohrbewegungen an, sobald sie
mit dem Wasser[S. 190] in Berührung kommen. Die Eier vieler Fische schwellen
durch Wasseraufnahme auf und saugen dabei gewissermassen die Samenfäden
in sich ein. Auf diese Weise werden alle reifen Eier, die sich in
der Schale befinden, befruchtet (während bei der Laichablage in der
freien Natur eine grosse Menge der abgesetzten Eier unbefruchtet
bleibt) und sind dann entwicklungsfähig. Sie können jetzt sofort in
die freien Gewässer gebracht und dort an geeigneten Stellen, d. h. an
solchen, an welchen die Fische ihrer Art laichen würden, ausgesetzt
werden. Allein der Fischlaich ist im Freien unzähligen Gefahren
ausgesetzt. Fast alle Wassertiere, welche ihn bewältigen können,
stellen ihm nach, ungünstige Witterung tötet ihn, Wellenschlag wirft
ihn auf das Land. Deshalb ist es besser, ihn so lange als möglich
unter Obhut zu behalten, ihn künstlich zu erbrüten. Man bringt ihn
in Brutapparate. Die Brutapparate sind fast durchgehends so
eingerichtet, dass in ihnen die Eier von fliessendem, klarem, reinem,
aber sauerstoffreichem und gleichmässig kühlem Wasser bespült werden.
In Jakobis Brutkiste lagen sie auf Kies, die Kiste hatte auf
den Schmalseiten Gitter und war so in einen Bach gestellt, dass das
Wasser durch die Gitter über die Eier floss. Coste legte die
Eier auf einen Glasrost, der in einem Gefäss stand. Solche Gefässe
stellte er staffelförmig über einander, so dass das Wasser, das in das
höchststehende geleitet war, aus diesem in das nächst tiefere floss
etc. Später wandte man Siebe aus Metall oder Thon an. Die Amerikaner
scheinen zuerst Drahtgeflechte zur Aufnahme der Eier angewandt zu
haben. Dies ist jetzt die übliche Unterlage. Man lässt das Wasser
entweder seitlich an den Eiern vorbeifliessen, oder man richtet die
Apparate so ein, dass das Wasser von unten her durch die Unterlage
und dann durch die Eierschichten strömt, wodurch das Wasser am
besten ausgenutzt wird. In anderen Apparaten werden die Eier nicht
ruhend, sondern schwebend ausgebrütet, indem in das kelchförmige
oder cylinderförmige Brutgefäss ein kräftiger Wasserstrahl von unten
her eingeleitet wird, welcher die Eier in die Höhe trägt; nach oben
hin verteilt sich der Wasserstrom und verliert an Kraft, die Eier
geraten in das ruhigere Wasser an den Gefässwänden, sinken hier[S. 191]
durch ihre Eigenschwere hinab und werden von dem Wasserstrom unten
sofort in erneutem Spiel in die Höhe getrieben[73]. Diese Apparate
sind besonders für kleinere Eier, wie die der Hechte und Coregonen,
geeignet. Sie haben unter anderem den besondern Vorteil, dass die
abgestorbenen Eier, welche etwas leichter werden als die lebenden,
sich von den letzteren absondern und bei etwas verstärktem Strome von
selbst mit dem durchgeleiteten Wasser abschwimmen können. Man nennt sie
deshalb Selbstausleser.
Eine dritte Methode ist die Erbrütung in Eisschränken, in welchen
die Eier nur von dem tropfenweise herabrinnenden Schmelzwasser des
über ihnen angebrachten Eises feucht und kühl gehalten werden. Diese
Eisbrutschränke dienen auch zum Transport von Eiern, welche wochenlang
unterwegs sein müssen. Man hat in ihnen die Eier fremder Fischarten
über die Ozeane in neue Gebiete, selbst über den Äquator hinaus,
eingeführt, z. B. den Lachs der nördlichen Hemisphäre in australische
Gewässer.
Leider ist die Methode der künstlichen Befruchtung und Erbrütung nur
bei einer beschränkten Zahl von Fischarten praktisch anwendbar. Der
erste Mangel, den diese Methode hat, liegt in der Notwendigkeit,
dass die Laichfische in dem gerade zur Befruchtung geeigneten
Laichreifestadium zur Hand sein müssen. Einige Fische können dieses
Stadium in der Gefangenschaft erreichen. Viele andere aber werden,
auch kurz vor der Laichreife eingefangen, in engen Behältern nicht
laichreif. Bei anderen Fischarten (Karpfen, Bressen, Stör und den
meisten anderen Sommerlaichern) quillt, wie oben erwähnt, die Eihaut
bei der Berührung mit Wasser zu einer klebrigen Substanz auf; diese
Eier ballen sich zu zähen Klumpen zusammen, wenn sie abgestrichen sind,
so dass die Zuführung von Wasser und Sauerstoff zu den im Innern des
Klumpens gelegenen Eiern und dadurch ihre Erbrütung unmöglich gemacht
wird. Man kann diese Eier indessen unmittelbar nach der Befruchtung
auf Wacholder, Moos oder Wasserkräuter vorsichtig in dünner Schicht
verteilen und sie in flachen Körben in stillem, warmem Wasser ausbrüten
lassen; die Jungen schlüpfen dann nach einigen Tagen aus und gelangen
durch die Korböffnungen in das Gewässer, in[S. 192] dem der Korb steht, worin
sie, wenn das Gewässer fischleer ist, so lange aufwachsen, als sie
Nahrung finden.
Die Eier der meisten Salmoniden sind sehr geeignet zur künstlichen
Erbrütung. Forellen und Lachse werden in Behältern reif, und ihre Eier
sowie die der Coregonen und Aeschen kleben nur wenig. Gerade diese
Salmoniden bedürfen aber auch des besondern Schutzes in hohem Masse.
Ihre Laichzeit fällt, bis auf die der Aesche, in den Herbst und den
Winter, die Entwickelungsdauer der Eier ist eine sehr lange, und die
Eier sind gross und oft lebhaft gefärbt. Dabei ist die Zahl der Eier,
welche die Salmoniden produzieren, nicht so bedeutend, wie die vieler
im Sommer laichender Fische. Deshalb ist es nötig, den Laich dieser
Edelfische so gut als möglich zu schützen und ihn künstlich befruchtet
in Bruthäusern ausschlüpfen zu lassen.
Wie auf die meisten anderen Lebensverhältnisse der Fische, so ist
auch auf die Länge der Entwickelungszeit der Eier die Wasserwärme
von grösstem Einfluss. Je kälter das erbrütende Wasser ist, um
so längere Zeit muss es auf die Eier einwirken, bevor dieselben
ausschlüpfen[LXXVII]. Die Eier vieler Sommerlaicher vertragen dabei nur
schlecht die Kühle und sterben im Freien oft ab, wenn das Wasser auf
die Dauer kalt bleibt. Die Eier der Salmoniden dagegen vertragen
kaltes Wasser sehr gut, am kräftigsten entwickeln sich die Fische
aus Eiern, die in eiskaltem Wasser gebrütet sind. Man unterscheidet
während der Brütung der Salmoniden-Eier zwei Hauptperioden. Die erste
reicht von der Befruchtung bis zum Sichtbarwerden der schwarzen
Augenpupillen des Embryo, und dauert für Lachse und Forellen bei einer
Durchschnittstemperatur des Wassers von 5° C. etwa 3½ Monate;
die zweite umfasst die Zeit bis zum Ausschlüpfen, sie dauert bei Lachs
und Forelle etwa 2 Monate bei der angeführten Wassertemperatur[74].
Die Eier der Coregonen haben eine kürzere Entwickelungsdauer.[S. 193] In
der zweiten Entwickelungsperiode sind alle Salmoniden-Eier ziemlich
widerstandsfähig; sie können dann, kühlgehalten, auf weiche Unterlage
gebettet und mit dieser fest verpackt, weithin versandt werden und
wochenlange, im Eisschranke sogar monatelange Reisen überdauern. Nach
dem Ausschlüpfen schwärmen die jungen Coregonen wie die Brut der
meisten anderen Fische sogleich frei umher, obwohl die Brustflossen
noch gar nicht und die Bauchflossen erst als Stummel entwickelt
sind. Die Brut der Forellen und Lachse dagegen ist mit einem grossen
Dottersäckchen beschwert, das ihr am Bauche hängt und bei den
Schwimmbewegungen anfangs hinderlich ist, weshalb diese Tierchen in
den ersten Wochen, ohne sich viel zu bewegen, am Boden der Brutgefässe
liegen. Erst wenn der Dottervorrat eingesogen ist, haben auch sie freie
Beweglichkeit erlangt. Dann ist es Zeit, sie in die freien Gewässer zu
bringen.
[LXXVII] Man muss aus den Resultaten der Untersuchungen von
Barfurth (Jahresbericht des Rheinischen Fischereivereins 1888)
allerdings schliessen, dass die Entwickelungsdauer der Salmoniden
nicht genau in dem Verhältnis verkürzt wird, in welchem man die
Wassertemperatur erhöht, dass vielmehr die Eier in wärmerem Wasser mehr
Wärme verbrauchen als in kälterem Wasser.
Die Erfolge, welche bis jetzt durch die künstliche Fischzucht
erzielt sind, sind recht erhebliche. Zunächst ist mit ihrer Hilfe
der Bestand an Lachsen nachweisbar vermehrt worden. Der Lachs
hat seines hohen Preises wegen und weil er verhältnismässig sicher
zu fangen ist, grosse Bedeutung für die Fischerei, und zwar
sowohl für die Binnenfischerei als auch für die Meeresfischerei
(wenigstens in der Ostsee). Seit 1879 hat sich nach der holländischen
Verkaufsstatistik[75] der Ertrag des Lachsfanges in den
Rheinmündungen etwa verdoppelt, und diese günstige Änderung wird mit
Recht auf die Aussetzung zahlreicher Brutmengen in die Nebenbäche des
Rheins zurückgeführt. Ebenso ist der Lachsbestand in der Ems und der
Weser, in denen er sehr zurückgegangen war, allem Anschein nach wieder
durch Bruteinsetzungen gehoben worden. In der Elbe ist der Lachsbestand
besonders durch künstliche Lachszucht in Böhmen vermehrt worden. Im
Odergebiet liegen die Laichplätze der Lachse in einigen Nebenflüsschen
der Netze, der Drage und der Küddow, wahrscheinlich, weil den Lachsen
der Zutritt zu den Quellgebieten der Oder in Schlesien durch die
grossen Wehre bei Breslau seit langen Zeiten[S. 194] abgeschnitten ist, und
an diesen Wehren leider noch keine Fischwege angebracht sind. In der
Weichselmündung hat sich der Lachsfang infolge der Brutaussetzung in
Galizien und Westpreussen ebenfalls deutlich vermehrt.
Ein dem Lachs nach Körperform, Grösse und Lebensweise sehr ähnlicher
Fisch ist die Meerforelle. Auch mit ihr sind durch künstliche
Fischzucht, namentlich in den holsteinischen Auen, vorzügliche
Resultate erzielt worden.
Die stärkste Vermehrung durch künstliche Fischzucht dürfte der
Bachforelle zu teil werden, von der alljährlich mehrere
Millionen künstlich erbrüteter Jungfische zur Besetzung von Zuchtbächen
und Teichen benutzt werden, um als 2–3jährige Fische zum Verbrauch
ausgefischt zu werden.
Wie oben auseinandergesetzt ist, gelingt es nicht bei allen Fischen,
den Laich zur künstlichen Erbrütung zu verwenden. Wo dies unbequem
oder unmöglich ist, ist man für die geschützte Vermehrung der Fische
darauf angewiesen, die Fische das Laichgeschäft auf natürliche Weise
in ablassbaren Bassins, Teichen oder ähnlichen Behältern, deren Inhalt
man in seiner Gewalt hat, vollziehen zu lassen und die gewonnene
Fischbrut in geeignetem Alter wie die durch künstliche Erbrütung
gewonnene zu verwenden. Die Teichzucht gehört deshalb auch zur
künstlichen Fischzucht, und um so mehr, als die künstlich erbrüteten
Fische zweckmässigerweise zuerst einen Sommer über in einem Teich oder
einem ablassbaren Graben gezogen und erst, wenn sie hier zu kräftigen
Fischchen herangewachsen sind, in die freien Gewässer übertragen werden.
Besonders häufig wird der Karpfen in Teichen gezogen[76].
Am geeignetsten für die Karpfenzucht sind flache Teiche. Da aber
die Karpfen in diesen im Winter unter dem Eise leicht ersticken, so
nimmt man sie im Herbst aus solchen Teichen in der Regel heraus und
bringt sie in kleine, tiefe, durchströmte Teiche (Winterheller).
Da eine gleiche Zahl Karpfen je nach ihrem Alter und ihrer Grösse
mehr oder minder grosse Teichflächen[S. 195] beansprucht, so setzt man die
einzelnen Jahrgänge in besondere Teiche zusammen, deren Grösse ihrem
Nahrungsbedürfnis entspricht. Die für das Ablaichen und die erste
Entwickelung der Brut bestimmten Teiche heissen Streichteiche. Aus
ihnen überträgt man die Jungen in die sogenannten Streckteiche, wo
sie 1–2 Jahre aufwachsen, bis sie zur Erreichung der Grösse, in der
sie verkauft werden (meist 1–1¼ Kilo schwer), in die sogenannten
Abwachsteiche kommen, in denen man ihnen meist kleine Raubfische
(Hechte, Zander, Forellen) beigiebt, damit die etwa von frühreifen
Karpfen erzeugte Brut sogleich beseitigt wird und nicht den zum
Auswachsen bestimmten Karpfen das Futter schmälert. Man nimmt an,
dass die Fische um so wohlschmeckender sind, je rascher sie gewachsen
sind. Aus diesem Grunde und weil ein rasches Wachstum, ein möglichst
gründliches Ausnützen des vorhandenen Futters für den Züchter offenbar
von Vorteil ist, bemüht man sich, möglichst schnellwüchsige Karpfen zur
Zucht zu nehmen. Man erreicht dies einerseits dadurch, dass man nur
die am besten gewachsenen unter den zur Laichreife gelangten Karpfen
in die Streichteiche nimmt, anderseits, indem man die Karpfenbrut
schon im ersten Sommer ihres Lebens durch reichliche Nahrung und
Schutz vor Feinden zu kräftigen Tieren erzieht. Ein Verfahren, um dies
zu erreichen, ist von dem schlesischen Fischzüchter Dubisch
erfunden. Man nimmt danach die Karpfenbrut schon acht Tage nach
ihrem Ausschlüpfen mit Gazenetzen aus dem Teich und bringt sie in
andere Teiche, so dass etwa 25000 auf den Hektar Teichfläche kommen.
Schon nach vier Wochen fängt man sie abermals heraus und überträgt
sie in andere Teiche, so dass nur 1000 Stück im Hektar enthalten
sind. Im zweiten Frühjahr bringt man sie in eine dritte Klasse von
Streckteichen, in welchen nur 500 im Hektar sich befinden. Endlich im
dritten Frühjahr bringt man sie in Abwachsteiche und setzt in diesen
auf den Hektar 200 Karpfen. Man erzielt auf diese Weise pro Hektar
Teichfläche etwa 120 Karpfen von etwas über 1 Kilo Schwere, was einem
jährlichen Ertrag von etwa 162 Mark aus jedem Hektar entspricht.
[S. 196]
Mit Hilfe der künstlichen Erbrütung von Eiern und der durch das
Dubisch-Verfahren vervollkommneten Teichwirtschaft hat man auch eine
Anzahl ausländischer Fischarten in Deutschland eingeführt, die man
zwar noch nicht gut als der deutschen Fauna angehörig betrachten kann,
die aber teilweise doch einmal eine Rolle in unserer Tierwelt werden
spielen können.
Man sollte bezüglich der Einführung neuer Fischarten sein Augenmerk
zunächst auf die Gegend wenden, von wo die meisten unserer
einheimischen Fischarten herstammen, nach Osteuropa und Nordasien,
da die Fische dieser Gegenden sich ohne Zweifel in ähnlichen
Lebensverhältnissen befinden, wie unsere einheimischen. Man hat aus
dieser Gegend den Sterlett in die norddeutschen Ströme einzuführen
versucht; leider sind die Fische bezw. Eier auf dem Transport meist zu
Grunde gegangen, wahrscheinlich nur infolge von Zufälligkeiten.
Am leichtesten gelingt der Transport der sich langsam entwickelnden
Eier der Salmoniden. Man hat in dieser Form nicht nur europäische
Lachse und Forellen nach Tasmanien und Neuseeland[77], Forellen und
Maränen[78] nach Amerika gebracht, sondern auch eine ganze Anzahl
von Salmoniden Amerikas in Deutschland eingeführt[79]. Von diesen
sind der Bachsaibling (Salmo fontinalis Gemminger) aus den
Bächen Nordamerikas und die Regenbogenforelle (Salmo irideus,
Livingston Stone) aus dem Höhenlande Kaliforniens weit verbreitet und,
der erstere in rasch fliessenden Bächen, die selbst der Forelle zu
reissend sind, die letztere auch in Teichen, vortrefflich gediehen.
Auch einige amerikanische Sommerlaicher sind durch die Bemühungen des
Fischzüchters M. von dem Borne in Deutschland verbreitet. Es
sind dies der Schwarzbarsch (Grystes nigricans Günther) und der
Forellenbarsch (Grystes salmoides Gü.), zwei Fische der Barben-
und Bleiregion, denen ein vorzüglich feines Fleisch zugeschrieben wird
und die bezüglich ihrer Lebensbedingungen, wenigstens was die Reinheit
des Wassers betrifft, anspruchsloser als die feineren einheimischen
Tafelfische sind[80], — ferner, erst neuerdings eingeführt, der
Steinbarsch, Centrarchus aeneus C., aus dem[S. 197] Mississippi[81],
und der Zwergwels, Amiurus catus Jord. u. Gilb., aus den
flacheren Gewässern dieses Gebietes[82].
Auch innerhalb unseres Gebietes haben mehrere Fischarten mit Hilfe der
Fischzucht oder der aus derselben gezogenen Erfahrungen eine weitere
Verbreitung erhalten, als ihnen von Natur zukommt. Dahin gehört der
Karpfen, der seit alter Zeit in ganz Deutschland gezüchtet wird, und
der Zander, welcher in die Gebiete des Rheins, der Ems und der Weser,
sowie in zahlreiche norddeutsche Seen, in denen er bisher fehlte,
künstlich eingeführt ist.
Vorzügliche Resultate sind mit der Verbreitung der Aalbrut
erreicht worden, die in den Mündungen des Po und der Flüsse der
französischen Ozeanküste in Menge gefangen wird. Direktor Haack
bringt jährlich grosse Massen davon nach Hüningen, von wo die jungen
Tiere in feuchtes Kraut verpackt bequem mit der Post versendet werden.
Zahlreiche Gewässer, welche die Aalbrut auf ihrer Wanderung nicht
aufsuchen kann, sind auf diese Weise mit Aalen bevölkert worden. Auch
ist ein gross angelegter Versuch gemacht worden, das Donaugebiet, das
wie alle Flussgebiete des Schwarzen Meeres den Aal bisher nicht besass,
mit diesem Fisch zu besetzen. Zahlreiche junge Aale sind in dem oberen
Donaugebiet ausgesetzt worden und die Tiere wuchsen dort gut auf. Der
grösste Teil der eingesetzten Brut bestand indessen aus Weibchen. Um
die Fortpflanzung zu sichern, wurde daher eine grosse Zahl erwachsener
Aalmännchen aus der Nordsee in das Schwarze Meer gebracht. Man hofft
dadurch den Fortbestand der Aale im Gebiete des Schwarzen Meeres
gesichert zu haben.
Auf die Entwickelung der Fische im Ei kann hier des Raummangels wegen
nicht eingegangen werden[83].
Einige Fische behüten ihre Eier und Jungen eifrig vor den
Nachstellungen der Feinde, so der Zander, der Schwarzbarsch, der
Stichling, der die Eier in ein Nest aus Pflanzen legt, das mit aus
den Harnkanälen stammenden Schleimfäden[84] befestigt ist, der
Kaulkopf, der ebenfalls ein Nest im Bachgrunde bereitet, und zwar[S. 198] sind
es meist die Männchen, welche Wache halten und selbst die Weibchen den
abgelegten Eiern nicht nahekommen lassen. Der weibliche Bitterling
legt seine Eier mit Hilfe seiner langen Legeröhre in die Kiemenräume
der Teichmuscheln (Unioniden), wo sie bis zum Ausschlüpfen geschützt
bleiben (während anderseits die Teichmuscheln bekanntlich ihre
Larvenzeit als Hautparasiten der Fische und anderer Wasserwirbeltiere
zubringen).
Werden die Fische an der Laichablage verhindert, so werden die
Geschlechtsprodukte resorbiert, oft unter Krankheitserscheinungen[85].
Wenige Worte seien noch dem Nervensystem und den Sinnesorganen der
Fische gewidmet.
Das Gehirn[86] liegt in der Schädelkapsel unter einer dicken
Fetthülle, es ist im Verhältnis zur Körpermasse und zu den übrigen
Teilen des Nervensystems klein. Auffallend ist, dass für das Grosshirn,
an welches bei den höheren Wirbeltieren Bewusstsein und Wille
gebunden erscheinen, bei dem Fisch noch gar keine bestimmte Funktion
hat nachgewiesen werden können[87]. Döbeln, deren Grosshirn
herausgenommen war, zeigten in keiner Weise eine Abweichung ihrer
Lebensthätigkeiten.
Das Auge[88] wird von der Körperhaut überzogen, welche an
dieser Stelle völlig durchsichtig ist. Die darunter liegende Hornhaut
(Cornea) ist sehr flach, die Linse dagegen fast kugelrund. Die
Fische gelten deshalb für kurzsichtig; für den Hecht ist eine Sehweite
von etwa 65 cm nachgewiesen worden[89]. Die Regenbogenhaut
(Iris) ist metallglänzend infolge der Einlagerung von
Glanzkörperchen, wie sie auch sonst den Glanz der Fische verursachen.
Übrigens ist die Regenbogenhaut nicht kontraktil, wie bei den höheren
Wirbeltieren. Eigentümliche Bildungen des Fischauges sind die
sogenannte Glandula chorioidea, ein in der Nähe des Eintritts
des Sehnervs liegender, an Blutgefässcapillaren reicher Körper, und
eine zweite Falte der Gefässhaut (Chorioidea) des Auges, welche
die Nervenhaut (Retina) des Auges durchsetzt und an der Linse
mit einer Verdickung, der Campanula Halleri, endet.
[S. 199]
Das Hörorgan der Fische[90] liegt seitlich vom Gehirn im Grunde
des Schädels in der Nähe des Hinterendes desselben, oft in einer
besonderen Ausbuchtung des Schädels. Ein äusseres Ohr kommt bei den
Fischen nicht vor. Das Hörorgan besteht aus mehreren häutigen Teilen,
nämlich dem Säckchen (Sacculus), das eine Ausstülpung zeigt, die
der Schnecke der höheren Wirbeltiere entspricht und einen oder mehrere
Gehörsteine (Otolithen) von zahnschmelzartiger Substanz enthält, und
dem Utriculus oder Alveus communis, an welchem sich bei
den Neunaugen zwei, bei den übrigen Fischen drei „halbzirkelförmige
Kanäle“ befinden, die zuweilen in Erweiterungen ebenfalls Otolithen
enthalten. Im Sacculus endigen die Nervenfasern des Hörnervs
in Sinneszellen, welche am äusseren Ende je eine kleine starre Borste
tragen. Ähnliche in Haare ausgehende Sinneszellen finden sich auch
in den sogenannten Seitenorganen. An den Seiten der Fische,
mit Ausnahme der Neunaugen, findet sich nämlich eine Reihe von Poren,
welche die Schuppen durchsetzen und die „Seitenlinie“ bilden. Sie
führen in Kanäle, welche unter der Haut liegen. In diesen Kanälen[91]
liegen im Grunde von Vertiefungen kleine Erhebungen, welche aus
ebensolchen Sinneszellen bestehen, wie sie sich in dem Cortischen Organ
des Hörapparates finden. Man nimmt daher an, dass diese Organe, welche
keinem der Sinne der höheren Wirbeltiere entsprechen, eine ähnliche
Funktion haben wie das Hörorgan, indem die Seitenorgane zwar nicht die
kurzen, rasch verlaufenden Schallwellen, aber andere Druckwirkungen,
wie die Bewegungen anderer Körper im Wasser, empfinden sollen.
Schon bei Erwähnung der Schwimmblase ist hervorgehoben worden, dass
diese mit Luft prall gefüllte Blase bei den meisten unserer Fische
mit dem Gehörorgan in Verbindung steht. Diese Verbindung wird bei
den Perciden und Clupeiden dadurch hergestellt, dass Verlängerungen
der Schwimmblase dicht an den Utriculus herantreten, bei den
Cypriniden und dem Wels durch eine Reihe von kleinen Knochen, deren
Reihe einerseits die Schwimmblase, anderseits einen Verbindungskanal
der Vestibula beider Seiten berührt. Auf diese Weise dient
gewissermassen der[S. 200] ganze Mittelkörper, soweit in ihm die Schwimmblase
liegt, als eine Art äusseren Ohrs. Die Prallheit und Elastizität,
welche die Schwimmblase dem Fischkörper giebt, mag auch zur Folge
haben, dass die Druckwahrnehmungen der Seitenorgane deutlicher
empfunden werden.
Die einander verwandten Empfindungen des Geschmacks und des
Geruches werden bei den meisten Fischen anscheinend nur durch ein
Sinnesorgan, das der Nase der höheren Wirbeltiere entspricht,
wahrgenommen. Indessen kann man, wie J. Müller hervorgehoben
hat, deshalb den Geruch der Fische mit dem Geschmack derselben nicht
identifizieren, wiewohl die Geruchsstoffe im Wasser gelöst sind, denn
auch bei den Luftwirbeltieren muss sich der Geruchsstoff erst im
Wasser der Nasenschleimhaut lösen, um empfunden zu werden. Besondere
Geschmacksorgane hat F. E. Schulze in der Mundhöhle einiger
Cypriniden entdeckt, wie schon bei der Beschreibung dieses Körperteils
erwähnt ist. Das Geruchsorgan[92] kommt allen Fischen zu, es ist bei
den Neunaugen einfach, bei den anderen Fischen doppelt vorhanden. Es
liegt jederseits zwischen den Augen und dem Mund in einem längeren oder
kürzeren Hautkanal, welcher am Vorderende oft in eine häutige Röhre
verlängert ist, die geschlossen und geöffnet werden kann, sodass es im
Belieben des Fisches liegt, während des Schwimmens einen Wasserstrom
durch das Organ gehen zu lassen und mittels desselben zu prüfen, oder
dasselbe ruhen zu lassen. Das Geruchsorgan selbst ist eine regelmässig
gefaltete Schleimhaut, in welcher die Sinneszellen liegen[93].
Das Tastgefühl scheint mehr oder minder um den Mund konzentriert zu
sein, wo sich bei vielen Fischen längere oder kürzere „Barteln“,
Tastfäden, befinden, wie bei Aalquappe, Wels, Karpfen, Barbe,
Gründling, Schleihe, den Acanthopsiden und Stören.
Die Sinnesorgane, die in vielen Punkten von denen der Landwirbeltiere
verschieden sind und in ihrer Funktionsweise dem Wasserleben
entsprechen, leiten den Fisch zu den Stätten im Wasser, deren er
jeweilen bedarf. In der Hauptnährzeit, im Sommer, verteilen[S. 201] sich die
Süsswasserfische fast stets weit in den Gewässern, jeden zugänglichen
Winkel nach der ihnen geeigneten Nahrung durchsuchend. In zwei
Zeitperioden sind die Fische dagegen wählerisch in ihrem Aufenthalt:
zur Laichzeit und zur Zeit des herabgeminderten Nahrungstriebes, im
Winter. Zur Laichzeit suchen sie die für die Entwickelung der Eier
und der ausgeschlüpften jungen Brut geeignetsten Wassergegenden auf,
Kiesberge, Pflanzenrasen, Röhricht, schwimmendes Pflanzengewirr,
überströmte Kiesbänke u. s. w. Im Winter ziehen sie sich an
geschützte Stellen, meist in die Tiefe des Wassers, zurück. Einige
Arten scheinen sich in den weichen Grund einzuwühlen, besonders in
flacheren Gewässern, welche bis in die Nähe des Grundes gefrieren.
Andere Arten suchen reinen, schlammfreien Grund, Sandflächen, auf
(Bressen, Karpfen), oder sie halten sich an der Oberfläche des offenen
Wassers und stehen auch unter dem Eise hoch über dem Grunde (Uklei).
Nicht nur die Friedfische, auch die Raubfische, namentlich die
jüngeren Generationen, suchen geeignete Stellen zur Überwinterung auf
(Barsch, Wels).[94] Da nun die besonderen Bedürfnisse der Art in weiten
Wasserstrecken oft nur an wenigen Stellen erfüllt sind, so sammeln
sich an diesen Stellen die Fische einer Art in grossen Scharen. Die
Laichzeit und die Zeit der Winterruhe sind daher für die praktische
Fischerei von Bedeutung, da man die Fische dann verhältnismässig
leicht in Menge mit Netzen umstellen und fangen kann. Die Erhaltung
der Fischart erfordert es allerdings, dass die laichenden Fische weder
gefangen, noch auch nur gestört werden, und dass man den Massenfang
auf den Winter beschränkt, eine Forderung, welcher der Fischer, im
Gegensatze zum Fischzüchter, nicht immer gern nachkommt.
Ohne Zweifel ist es der wissenschaftlichen Fischkunde vorbehalten,
unterstützt von einer gründlichen Kenntnis der Natur der Gewässer und
aller ihrer Bewohner, die nur scheinbar entgegengesetzten Bestrebungen
der Fischzüchter und der Fischfänger zu vereinigen und unter Förderung
rationeller Fangmethoden sowohl wie der Fischzucht die Grundsätze
einerseits einer zweckmässigen Fischereigesetzgebung, anderseits der
praktischen Verwertung der[S. 202] Gewässer durch Fischzucht völlig sicher
und unangreifbar zu ermitteln. Bis jetzt liegen Theorie und Praxis
der Fischerei noch sehr im Argen; die Fangmethoden sowohl wie die
Gesetzesvorschriften „schleppen von Geschlecht sich zu Geschlechte“,
während andere Zweige der Wasserverwertung, namentlich die der
Landwirtschaft und der Industrie, mächtig vorwärtsgeschritten sind und
der Fischerei kaum das Dasein gönnen.
[8]K. Moebius und F. Heincke, Die Fische der
Ostsee. 1883.
[9]E. Th. E. von Siebold, Die Süsswasserfische von
Mitteleuropa. 1863.
Marcus Eliezer Bloch, Oekonomische Naturgeschichte der Fische
Deutschlands. 1782–84.
Berthold Benecke, Fische, Fischerei und Fischzucht in den
Provinzen Ost- und Westpreussens. 1880.
Cuvier et Valenciennes, Histoire naturelle des poissons.
22 Bde. 1828–1848.
A. Günther, Catalogue of fishes of the British Museum. 8 Bde.
1859–1870.
Agassiz, Histoire naturelle des poissons d’eau douce de l’Europe
centrale. 1842.
Wittmack, Beiträge zur Fischereistatistik des Deutschen Reiches
u. s. w. 1875.
Max von dem Borne, Die Fischereiverhältnisse des Deutschen
Reiches u. s. w. 1882.
[10]Heckel und Kner, Die Süsswasserfische der
Oesterreichischen Monarchie. 1858.
[11]C. Heller, Die Fische Tirols und Vorarlbergs.
1871.
[12]Sclater, Über den gegenwärtigen Stand unserer
Kenntnis der geographischen Zoologie. 1857.
[S. 204]
[13]A. Günther, Handbuch der Ichthyologie, übersetzt
von G. v. Hayek. 1886.
[14]G. Henschel, Praktische Anleitung zur Bestimmung
unserer Süsswasserfische. 1890.
E. Schulze, Fauna piscium Germaniae. 1890.
[15]Benecke, Die westpreussischen Fische. 5 Tafeln.
1887.
Gemeinfassliche Belehrung über die Süsswasserfische des Elbgebietes.
Schriften des Sächsischen Fischereivereins Nr. 1. 1884.
[16]Nowicki, Fauna und Verbreitung der Fische
in den Gewässern Galiziens. Mitteilungen des Österreichischen
Fischereivereins, 2. Jahrg., S. 149.
[17]F. Heincke, Untersuchungen über Stichlinge.
Ofversigt af K. V. Akad. Förhandling. Jahrg. 46, Nr. 6. 1889. S. 399.
[18]Heckel, Abbildungen und Beschreibungen der
Fische Syriens nebst einer neuen Klassifikation und Charakteristik
sämtlicher Gattungen der Cyprinen. Stuttgart 1843.
[41]Šusta, Die Ernährung des Karpfen und seiner
Teichgenossen. 1888.
[42]Provençal et Humboldt in Schweiggers
Journal für Chemie und Physik, Bd. 1, S. 84.
[43]Miller, mitgeteilt in Roscoe-Schorlemmer,
Ausführl. Lehrbuch der Chemie, Bd. 1, S. 205.
[44]C. Weigelt, Schädigung der Fischerei durch
Abwässer der Fabriken. Archiv für Hygiene, Bd. 3, 1885.
König, Die Verunreinigung der Gewässer. 1887.
[45]Fritsch, Arbeiten der zool. Sektion der
Landesdurchforschung von Böhmen. Prag 1872 (Arch. d. naturw.
Landesdurchforschung von B., 2. Bd., 4. Abt., S. 112 u. f.).
[46]August Müller, Über die Entwickelung der
Neunaugen. Joh. Müllers Archiv 1856, S. 334.
[47]H. Rathke, Über den Darmkanal und die
Zeugungsorgane der Fische. Neueste Schr. d. Naturforschenden Ges. in
Danzig, 1. Bd., 3. Heft, 1824.
[48]Cajetan, Beitrag zur Lehre von der Anatomie und
Physiologie des Tractus intestinalis der Fische. 1883.
[S. 206]
[49]Decker, Zur Physiologie des Fischdarms (in:
Festschrift für Kölliker). 1887.
[50]A. Fick, Über d. Magenferment kaltblütiger Tiere.
Verhandl. d. physikal.-medic. Ges. zu Würzburg. N. F. 2. Bd.
[51]Edinger, Zur Kenntnis der Drüsenzellen d. Magens:
Arch. f. mikrosk. Anatomie, Bd. 17.
Trinkler, Bau der Magenschleimhaut. Arch. f. mikrosk. Anatomie,
Bd. 24.
[52]Richet et Morrut, De quelques faits
relatifs à la digestion gastrique des poissons. Comptes rend. Bd. 90.
[59]C. K. Hoffmann, Ontogenie der Knochenfische.
Verhandl. d. k. Acad. van Wetenschapen i. Amsterdam, Bd. 21.
[60]Jakoby, Der Fischfang in der Lagune von Comacchio
nebst einer Darstellung der Aalfrage. 1880.
[61]Carlo Mondini, De anguillae ovariis. De
Bononiensi scientiarum et artium instituto atque academia commentarii.
T. 6. Bon. 1783.
[62]Syrski in: Abhandl. d. Kaiserl. Akademie d.
Wissensch. in Wien. 1874.
[63]Moebius, Ber. über Untersuchungen des Zustandes
der Geschlechtsdrüsen weiblicher und männlicher Aale. 5. Bericht d.
Komm. zur wissensch. Untersuchung der deutschen Meere, 1887, S. 127.
[64]Fritsch, Erwägungen über die Aussetzung
von Lachsen und Aalen in das Donaugebiet. Mitth. d. Österreich.
Fischereivereins 1884.
[65]Day in: Proceed. zool. Societ. London 1884, Bd.
1, S. 17.
[66]Fritsch, Untersuchungen über die Biologie und
Anatomie des Elblachses. Mitteil. d. Österr. Fischereivereins 1885, S.
99.
[67]Metzger, Beiträge zur Statistik und Kunde der
Binnenfischerei d. Preussischen Staates. 1880.
[S. 207]
[68]Miescher und Glaser, Statistische
und biologische Beiträge zur Kenntnis des Rheinlachses im Rhein. In:
Ichthyolog. Mittheilungen aus der Schweiz. 1880.
[69]Barfurth, Nahrung und Lebensweise der Salme,
Forellen und Maifische. Archiv f. Naturgeschichte 1875.
[70]Fritsch in: Circ. d. Deutschen Fischereivereins,
1882, S. 46.
[71]M. von dem Borne, Benecke und
Dallmer, Handbuch der Fischzucht und Fischerei. 1886.
[72]Zenk, Über Brutapparate für Salmoniden.
Bayerische Fischereizeitung 1881–1883.
[73]Weiss, Anleitung zur Handhabung des
Brutapparats u. s. w. Allgem. Fischereiztg. 1889, S. 141.
[74]Metzger, Fischerei und Fischzucht in den
Binnengewässern. In: Loreys Hdb. der Forstwissenschaft, Bd. 1, 1887.
[75]ten Houten und de Raadt, Zufuhr von
Lachsen am Kralingsche Veer. Deutsche Fischereiztg. 1888, S. 23.
[76]Horak, Die Teichwirthschaft mit besonderer
Rücksicht auf das südliche Böhmen. 1869.
Niklas, Lehrbuch der Teichwirthschaft. 1880.
[77]A. Nicols, The acclimatisation of the
Salmonidae at the Antipodes. 1882.
[78]Smiley in: Bull. Unit. Stat. Fisherie
Commission, Bd. 5, S. 468.
[79]von Behr, Fünf amerikanische Salmoniden in
Deutschland. Circ. d. Deutschen Fischereivereins. 1882.
von dem Borne, Sechs amerikanische Salmoniden in Europa. Neudamm
1890.
[80]von dem Borne, Der Schwarzbarsch und der
Forellenbarsch in Deutschland. 1888.
[81]von dem Borne, Der amerikanische Steinbarsch in
Deutschland. 1890.
[82]von dem Borne, Der amerikanische Zwergwels in
Deutschland. 1890.
[83]Kupffer, Die Befruchtung des Forelleneies.
Allg. Fischereizeitung, 1886, S. 2.
K. E. von Baer, Über die Entwickelungsgeschichte der Fische.
1835.
His, Untersuchungen über das Ei und die Eientwickelung bei
Knochenfischen. 1873.
Michal Girdwoyn, Pathologie des poissons. Traité des maladies,
des monstrosités et des anomalies des oeufs et des embryons. Paris
1880. Text und einige Figuren auch in der Deutschen Fischereizeitung,
1881, S. 413 u. f., 1882, S. 2 u. f.
S. auch Nr. 53.
[S. 208]
[84]Moebius in: Arch. f. mikroskop. Anat., Bd. 25,
S. 554.
[85]Barfurth, Biologische Untersuchungen über die
Bachforelle. Arch. f. mikroskop. Anatomie, Bd. 27.
[86]Rabl-Rückhard, Das Gehirn der Knochenfische
und seine Anhangsgebilde. Arch. f. Anat. und Physiol., Anatom. Abt.
1883.
L. Stieda, Über die Deutung der einzelnen Teile des
Fischgehirns. Zeitschr. f. wissensch. Zoologie, Bd. 23.
[87]Steiner, Die Funktionen des
Zentralnervensystems und ihre Phylogenese. 2. Abt.: Die Fische. 1888.
[88]Berger, Beiträge zur Anatomie des Sehorgans
der Knochenfische. Morphol. Jahrb., Jahrg. 8, Bd. 1.
[89]Hirschberg i. Arch. f. Anatomie und
Physiologie, Physiol. Abt., 1882, S. 493.
[91]Leydig, Über die Schleimkanäle der Knochenfische.
Müllers Arch. für Anatomie und Physiologie, 1860.
Leydig, Über das Organ eines sechsten Sinnes. 1868.
F. E. Schulze, Über die Sinnesorgane der Seitenlinie bei den
Fischen und Amphibien. In: Arch. f. mikroskop. Anatomie, Bd. 6, 1870.
[92]Dogiel, Bau des Geruchsorgans bei Fischen und
Amphibien. 1886.
[93] Bulletin of Unit. Stat. Fisherie Commission, Bd. 5, S.
142.
[94]Knauthe, Erfahrungen über das Verhalten von
Amphibien und Fischen gegenüber Kälte. Zool. Anzeiger, 1891, S. 1 u.
f.
[S. 209]
Die Parasiten
unserer Süsswasserfische.
Von Prof. Dr. Fr. Zschokke in Basel.
[S. 211]
Es ist eine gewöhnlich kaum beachtete Thatsache, dass der Körper des
Fisches den Angriffen verschiedenartigster Schmarotzer ausgesetzt
ist. Eine stattliche Zahl von durch Bau und Gestalt weit von einander
abweichenden Parasiten sucht und findet am und im Fisch bleibende
oder vorübergehende Wohnung und Nahrung. Es sind Geschöpfe, die den
verschiedensten Stämmen des Tierreichs angehören, Protozoen, Mollusken,
Krebse, Plattwürmer, Rundwürmer, Blutegel. Nur ein gemeinschaftliches
Band verbindet sie, ein biologisches, dieselbe Lebensweise, das
Schmarotzertum; nach Abstammung, Struktur, Entwickelungsgeschichte
gehen sie weit aus einander. Allerdings hat die Angewöhnung an dieselbe
Lebensweise, an den Parasitismus, die ihm unterworfenen Geschöpfe in
derselben Richtung modifiziert, und so zwischen ursprünglich einander
fernstehenden Tieren Ähnlichkeiten im Bau und im Lebensgang in
sekundärer Weise geschaffen.
Die den Fisch heimsuchenden Schmarotzer sind übrigens recht
unscheinbare Wesen. Zu ihrer niedrigen Lebensweise passt es ja am
besten, wenn sie weder durch besondere Grösse, noch Form und Farbe
die Aufmerksamkeit auf sich ziehen. Der Laie macht wohl gelegentlich
die Beobachtung, dass ein Lachs mit gierigen Blutegeln überdeckt ist,
oder sieht, wie sich der Leibeshöhle eines frisch geöffneten Karpfens
ein breiter, hässlicher Riemenwurm entwindet; der eine oder andere
Fisch wird wohl auch mit Recht oder Unrecht beschuldigt, dem Menschen
gegenüber die Rolle eines[S. 212] Zwischenträgers schmarotzender Würmer zu
spielen. Weiter geht die Kenntnis der Fischparasiten kaum. Das mit dem
verächtlichen Namen Schmarotzer belegte Geschöpf fesselt nicht durch
sein Äusseres und stösst geradezu ab durch seine Lebensweise.
Und doch wäre der Fischparasit eingehenden Studiums so sehr würdig.
Schon vom rein praktischen, medizinischen Standpunkt aus verdienen
die ungebetenen Gäste der Fische unsere volle Aufmerksamkeit. Hat
uns doch die neuere Forschung gelehrt, dass einer der verbreitetsten
Bandwürmer, der durchaus nicht unbedenkliche Bothriocephalus
latus, gerade durch die gesuchtesten Tafelfische des süssen Wassers
auf den Menschen übertragen wird. Für andere Würmer liegt die Vermutung
nahe, dass sie auf ähnlichem Wege, eingekapselt in Zwischenwirte
aus der Klasse der Fische, in unsern Körper eingeschmuggelt werden.
Die Notwendigkeit unsere Feinde zu kennen, um uns ihrer entledigen,
sie von uns fernhalten zu können, weist uns also schon gebieterisch
darauf hin, Bau und Eigenschaften der Fischparasiten zu ergründen und
ihrem oft so verwickelten Lebensgang zu folgen. Ausgerüstet mit den
nötigen Kenntnissen dürfte es uns wohl auch gelingen, Fischepidemien
parasitärer Herkunft einzudämmen, die unter den Bewohnern unserer
Gewässer, jenem nicht gering anzuschlagenden Teil des nationalen
Gutes, zahlreiche Opfer fordern. Ausser rein praktischen Erwägungen
medizinischer und nationalökonomischer Art müssen uns aber auch
wissenschaftliche Gesichtspunkte und Ziele beim Studium der
Fischparasiten leiten. Die Frage, welchen Einfluss übt die parasitische
Lebensweise auf ursprünglich freilebende Geschöpfe aus, wie werden
im Laufe ungezählter parasitischer Generationen Bau und individuelle
Geschichte des früher nicht schmarotzenden Tieres verändert, tritt uns
vor allen anderen schwerwiegend entgegen. Der Parasitismus führt ja
zu einer vollkommenen Umgestaltung und Neuschöpfung in Anatomie und
Entwickelungsgeschichte. Und um diese Neuschöpfung ihrer Entstehung
nach würdigen zu können, bietet uns gerade die Parasitenfauna der
Fische Gelegenheit und Material. Manche Schmarotzer beziehen den Fisch
nur temporär, um Nahrung aufzunehmen, sonst führen[S. 213] sie eine freie
Lebensweise. Andere bewohnen ihn wohl stationär, begnügen sich aber
damit, als Ektoparasiten die Oberfläche des Wirtes zu bewohnen und
dort ihren Unterhalt zu suchen, ohne jemals innere Organe zu besetzen.
Wieder andere werden Entoparasiten, siedeln sich aber in offenen mehr
oder weniger leicht zugänglichen Teilen, den Kiemen, dem Darmkanal,
an. Zahlreiche endlich liegen eingekapselt in allseitig geschlossenen
Organen, den Muskeln, den Augen, der Schwimmblase.
Grad und Dauer des Schmarotzertums sind bei den Fischparasiten höchst
verschieden. Manche schmarotzen während des ganzen Lebens, bei
anderen sind freie Stadien in mehr oder weniger reichem Masse in die
Lebensgeschichte eingestreut. Vom nur gelegentlich parasitierenden
Blutegel, von der jungen Muschel, die sich nur kurze Zeit vom Fisch
herumtragen lässt, bis zum Bandwurm im Darmkanal des Hechtes oder des
Lachses und dem Spulwurm aus dem Barsch, typischen, entoparasitischen
Gestalten, stossen wir auf mancherlei Zwischenstufen. Und gerade in
diesen verschiedenen Stadien, mit ihrem durch den Parasitismus mehr
oder weniger veränderten Bau und der ebenfalls verschieden stark
beeinflussten Lebensgeschichte, sehen wir noch Punkte, Stationen,
die von der Natur auf dem Wege berührt worden sind, als sie aus dem
ursprünglich freien Geschöpf den Parasiten schuf. Wir können den Weg
verfolgen, der vom freien Vorfahr zum parasitierenden Enkel führt. So
kann es uns klar werden, welch tiefen Einfluss die Angewöhnung an eine
so spezielle und niedrige Lebensweise, wie sie der Parasitismus nun
einmal ist, auf tierische Organisation und tierisches Leben ausübt.
Mit dem zunehmenden Schmarotzertum treten anatomische Vereinfachungen
und Umbildungen allmählich ein; Organsysteme werden rudimentär
und verschwinden zuletzt. Die Entwickelungsgeschichte weist oft
Komplikationen auf und schlägt Umwege ein, die ihre Erklärung einzig in
der Angewöhnung an die neue Lebensweise und in der Erfüllung der von
dieser gestellten Bedingungen finden. Das Studium der Parasiten führt
so den Forscher ein in die interessantesten und weitgehendsten Fragen
der Biologie. Die Fischschmarotzer aber, in ihrer bunten Formenfülle[S. 214]
und mit ihrer mannigfaltigen Lebens- und Entwickelungsgeschichte bieten
zur Behandlung dieser Probleme ein reiches Material.
Noch manche andere Frage wird sich auf diesem Gebiete stellen und
teilweise wenigstens beantworten lassen. Wie beeinflussen sich
gegenseitig Wirt und Gast? Wie passt sich der Parasit den ihm vom
Träger gebotenen Bedingungen an und welche Eigenschaften werden
infolge dieser Anpassung erworben? Wie findet sich auf der andern
Seite der Fisch mit den ihn bewohnenden Würmern ab; erwirbt nicht auch
er gewisse Eigenschaften, um die Eindringlinge abzuhalten, oder die
einmal Eingedrungenen bis zu einem gewissen Grad unschädlich zu machen?
Welche Veränderungen erleidet der Fischkörper unter dem Drucke der
Parasiteninvasion?
Interessant wird es auch sein, die bis jetzt kaum berührte Frage in
Fluss zu bringen, welchen Einfluss die Lebensweise, speziell die
Ernährungsweise des Fisches auf die Zusammensetzung der ihn bewohnenden
Parasitenfauna ausübt. Pflanzenfresser werden andere Formen von
Schmarotzern beherbergen als Fleischfresser, Schlammbewohner andere als
frei pelagisch schwimmende Geschöpfe. Der Fisch des schnellfliessenden
Stroms, des Sees, des Teiches wird von einander verschiedene Elemente
in seiner Parasitenfauna aufweisen. Besonders eigentümlich werden sich
diese Verhältnisse gestalten bei den Wanderfischen, die bald das süsse
Wasser, bald die salzige See aufsuchen und in beiden Medien eine oft
recht verschiedene Lebensweise befolgen.
Eine mehr oder minder starke Vermischung von Meer- oder
Süsswasserparasiten wird sich in diesem Falle wohl nachweisen
lassen. Reine Meerformen werden weit hinauf in die Flüsse getragen,
Süsswasserschmarotzer dagegen dem Meer zugeführt. Wieder wird uns die
verschiedene Mischung der beiden Parasitenelemente im Wanderfisch, das
Überwiegen des einen oder anderen, Rückschlüsse auf die Lebensweise
des Wirtes gestatten. Die Parasitenfauna wird so bis zu einem gewissen
Grade zum Spiegelbild der Gewohnheiten ihres Trägers.
[S. 215]
Von diesen Erwägungen geleitet ist eine erste kleine Arbeit über die
Parasitenfauna des Rheinlachses erschienen, deren Resultate geeignet
sind, die vorangehenden theoretischen Betrachtungen zu stützen. Sie
mögen deshalb hier in kurzen Zügen skizziert werden.
Der Rheinsalm, so bewiesen es His und Miescher, und so
wussten es auch schon längst die Lachsfischer, nimmt vom Aufsteigen
aus dem Meer bis er verlaicht hat, niemals Nahrung zu sich. Als
Folge dieses Fastens im Süsswasser durfte wohl erwartet werden, dass
die Parasitenfauna von Trutta salar typisch marinen Anstrich
zeige. Mit dem Ausschluss der Nahrungsaufnahme ist für jedes Geschöpf
ja gleichzeitig die Hauptinfektionsquelle mit parasitischen Würmern
verstopft.
Von 45 untersuchten Lachsen aus dem Rhein waren 42 mit Würmern
besetzt, nur drei erwiesen sich als vollkommen parasitenfrei. Elf
Arten Schmarotzer konnten bestimmt werden; mehrere fanden sich nur in
einem der geprüften Fische, andere öfters, der Spulwurm Agamonema
capsularia Dies. sogar in 35 Wirten. Parasitenarm waren in den
45 Lachsen die offenen Organe. Im Darmkanal fand sich kein einziger
Schmarotzer unterhalb der Appendices pyloricae. Die meisten Parasiten
waren in verschiedenen Organen eingekapselt, seltener lagen sie wohl
geborgen in den Falten des Schlundes und in den Pylor-Anhängen. Es
erinnert dies an das Verhalten gefangener Meerfische, die im Aquarium
verhältnismässig rasch ihre Darmschmarotzer verlieren. Man hat ja
auch geradezu behauptet, viele Wanderfische ziehen ins Süsswasser,
um sich ihrer Parasiten zu entledigen. Die Schmarotzerfauna der 45
Rheinlachse hatte einen fast reinen marinen Charakter. Die Hauptmasse
der gefundenen Würmer gehörte nach Arten und Individuen äusserst
typischen Meerformen an. Auch die von mir noch nicht nachgewiesenen
Lachsparasiten sind wesentlich Gäste von Meerfischen. Ein einziger der
gefundenen Schmarotzer gehört neben den Wanderfischen ausschliesslich
dem süssen Wasser an. So spiegelt die Zusammensetzung der
Parasitenfauna die eigentümliche Gewohnheit des Rheinlachses wider, im
Süsswasser keine Nahrung aufzunehmen. Sie unterscheidet sich scharf von
der[S. 216] Schmarotzerwelt anderer Wanderfische, deren Nahrungsbedürfnis im
Fluss nicht aufhört.
Über diese Verhältnisse mag die folgende Zusammenstellung näheren
Aufschluss geben.
Verbreitung der parasitischen Würmer der
Wanderfische:
Name des Fisches:
Zahl der ihn
bewohn. Spezies
parasit. Würmer:
Davon typisch für
den betreffenden
Fisch:
Nur in
Wanderf.
vorkommend:
Ausser in
Wanderfischen:
auch in
marinen
Fischen:
auch in
Süsswasserfisch.:
in mar. u.
Süsswasserfisch.:
1. Trutta salar, Lachs
20
7
7
7
1
5
2. Trutta trutta, Lachsforelle
15
1
1
1
8
5
3. Osmerus eperlanus, Stint
17
10
11
1
4
1
4. Coregonus oxyrhynchus, Schnäpel
6
0
1
0
4
1
5. Alausa vulgaris, Maifisch
7
3
4
1
0
2
6. Alausa finta, Finte
4
1
2
0
0
2
7. Anguilla vulgaris, Aal
25
10
10
5
5
5
7. Petromyzon fluviatilis, Neunauge
4
2
2
0
1
1
Besonders typisch für die Würmerwelt des Rheinlachses ist das Auftreten
der sogenannten Tetrarhynchen, eigentümlicher, mit vier starken,
hakentragenden Rüsseln bewehrter Bandwurmlarven, die sich häufig in den
verschiedensten Knochenfischen des Meeres einnisten, um mit ihnen in
den Verdauungstractus der Haifische zu gelangen und dort zu erwachsenen
Bandwürmern, Rhynchobothrien, auszuwachsen.
Eine in weit vorgeschrittener Vorbereitung sich befindende zweite
Arbeit über die Lachsparasiten, der hier nicht vorgegriffen werden
kann, wird, auf reiches Material gestützt, den marinen Charakter
der den grossen Wanderfisch bewohnenden Würmer noch viel greller
hervortreten lassen. Sie wird auch Ermittelungen über nordische und
schottische Lachse enthalten, die im Süsswasser dem Fasten nicht so
strenge zu huldigen scheinen wie ihr Vetter vom Rhein und bei denen
infolgedessen die parasitologischen Verhältnisse sich wesentlich anders
gestalten.
[S. 217]
Den relativ grossen Parasitenreichtum verdanken die Fische
verschiedenen Umständen: zunächst wohl ihrer Eigenschaft als
Wasserbewohner. Neben zahlreichen entoparasitisch die inneren Organe
aufsuchenden Schmarotzern kann auch mancher Ektoparasit an der
Aussenfläche des Wassertiers sich ansiedeln, der nur im feuchten
Element sein Leben zu fristen vermag. So wird denn die Oberfläche
des Fisches, die Haut, sowie die verhältnismässig freiliegenden, von
einem fortwährenden Wasserstrom bespülten Kiemen von manchem Wurm als
geeigneter Standort gewählt.
Auch die so mannigfache Ernährungsweise wird den Fischen manchen
Schmarotzer verschaffen. Mit der Nahrung dringt ja der grosse Haufe
ungebetener Gäste in den tierischen Körper ein. Die Parasitenfauna
einer Tiergruppe, die Pflanzen- und Fleischfresser umschliesst, wird
den verschiedenen Nahrungsstoffen gemäss ebenfalls mannigfaltig
ausfallen.
Mit dem Atmungswasser können ferner Jugendstadien von Schmarotzern in
das Innere des Fisches gelangen. Der Invasion ist somit eine neue,
bei anderen Geschöpfen geschlossene Pforte geöffnet. Manche Parasiten
werden auch aktiv eindringen, an wenig geschützten Körperstellen
mittels spezieller Apparate sich einbohren.
Die äusseren Lebensbedingungen, unter denen der Fisch steht, seine
Beziehungen zu anderen Lebewesen sind endlich sehr mannigfaltiger
Natur. Mit der grösseren Vielseitigkeit dieser Beziehungen steigert
sich im allgemeinen auch die Möglichkeit und Gelegenheit einer
Infektion. Von zahlreichen anderen Tieren verfolgt und verzehrt
wird sich der Fisch als Zwischenträger von Parasiten wohl eignen.
In ihm stellen sich zahlreiche Jugendstadien von parasitierenden
Würmern ein, die mit dem Fisch sicher in den Darmkanal eines weitern
Wirtes übertragen werden sollen, um dort zum geschlechtsreifen Tier
heranzuwachsen. So gelangen die Larven des breiten Bandwurms mit
dem Fleisch von Fischen in den Verdauungstractus des Menschen. Aber
auch als Hauptwirt von Parasiten ist der Fisch berufen eine grosse
Rolle zu spielen; mit den zahllosen kleinen Geschöpfen, die ihm zur
Nahrung dienen, können auf leichte Weise eingekapselte Larven in ihn
eingeschmuggelt werden. Die[S. 218] Fische erscheinen durch ihre Beziehungen
zu anderen Geschöpfen besonders geeignet, bald als Zwischenträger von
jungen Würmern, bald als definitive Wirte der geschlechtsreifen Form zu
figurieren.
So vereinigen sich manche Umstände, um den Fischkörper zu einer
richtigen Parasitenherberge zu machen. Einer Invasion ist Thür und Thor
geöffnet. Die Oberfläche des Wirtes und die nach aussen offenen Organe
werden vorzugsweise von geschlechtsreifen Schmarotzern, geschlossene
Körperteile von jungen Stadien bewohnt. Kein Organ bleibt aber
gelegentlich verschont. Zwölf Fischarten des Genfersees beherbergten 37
verschiedene Parasiten, die sich folgendermassen im Körper einquartiert
hatten:
Bewohntes Organ:
Zahl der Arten:
Ösophag und Magen
2
Darm
15
Rectum
2
Pylor-Anhänge
2
Leber
6
Milz
1
Schwimmblase
1
Harnblase
1
Auge
1
Muskulatur
2
Kiemen
2
Leibeshöhle
1
Peritonäum und Darmwand
8
Die Zahl der bis jetzt bekannten Parasiten der Süsswasserfische aus
dem Kreise der Würmer ist schon eine sehr beträchtliche, sie dürfte
kaum unter 250 zurückbleiben; sie steigt jährlich an und nichts lässt
voraussetzen, dass die diesbezüglichen Listen so bald als vollständig
geschlossen betrachtet werden können. Dem momentanen Stand unserer
Kenntnisse über die Vertretung von schmarotzenden Würmern im Körper der
verbreitetsten Fische des süssen Wassers dürfte etwa folgende Tabelle
entsprechen:
[S. 219]
Name der Fische:
Zahl der bei ihm vorkommenden Parasitenarten:
Saug-
würmer.
Band-
würmer.
Faden-
würmer.
Kratzer.
Total.
Barsch
12
8
9
4
27
Kaulbarsch
8
2
5
3
18
Zander
7
1
2
3
13
Groppe
3
2
0
2
7
Stichling
4
4
6
3
17
Aalraupe (Quappe)
6
9
6
4
25
Wels
2
2
6
3
13
Karpfen
8
1
2
4
15
Gründling
3
2
3
5
10
Barbe
7
3
1
5
16
Aitel
7
2
4
2
15
Rotauge
8
3
5
3
19
Hasel
7
2
3
4
16
Elritze
5
1
4
3
13
Schleihe
2
5
3
4
14
Laube
3
4
3
3
13
Schmerle
3
2
6
2
13
Schlammpeitzger
3
0
3
0
6
Saibling
4
9
1
1
15
Lachs
6
15
7
2
30
Forelle
4
2
4
7
17
Stint
6
4
9
2
21
Schnäpel
4
2
3
1
10
Felchen
4
7
1
1
13
Aesche
7
2
5
2
16
Hecht
10
6
7
3
26
Maifisch
4
1
4
3
12
Aal
14
3
12
8
37
Neunauge
5
1
3
0
9
Die vorangehenden Zahlen bedürfen kaum eines weiteren Kommentars. Sie
sind wiederum geeignet, die Abhängigkeit der Parasitenfauna von der
Lebensweise des Wirtes mit Beispielen zu[S. 220] belegen. Am reichsten an
Schmarotzern sind die grossen Räuber und die omnivoren Fische, denen
sich jeden Augenblick eine Infektionsgelegenheit bietet. In ihrem Darm
wohnt eine reiche Fauna von geschlechtsreifen Band- und Saugwürmern,
die in die Nahrung eingekapselt leicht in den definitiven Wirt gelangt
sind.
Bei den Karpfen, die mehr an pflanzliche Kost gewöhnt sind oder
höchstens kleine Tiere verzehren, sinkt die Zahl der Parasiten
bedeutend. Die Band- und Saugwürmer spielen hier eine untergeordnete
Rolle, während die Kratzer, deren Jugendstadium in kleinen Krustaceen
eingeschlossen liegt, mehr in den Vordergrund treten. Am reichsten
und buntesten gestaltet sich die Parasitenfauna des Aals, dessen
mannigfaltige Lebens- und Ernährungsweise ihn auch vielfacher
Infektionsgefahr aussetzen wird. Fleisch- und Pflanzenfresser unter
den Fischen beherbergen eine ziemlich verschiedene Schmarotzerwelt;
die letzteren dienen oft als Zwischenwirte für die Parasiten der
ersteren. In Fischen mit gemischter Ernährungsweise mengen sich
auch die Bestandteile der beiden Faunen. Wahrscheinlich wird in ein
und derselben Fischart in den verschiedenen Monaten des Jahres eine
Veränderung der Parasitenwelt nach Auswahl der sie bildenden Arten und
nach Zahl der Individuen sich nachweisen lassen. Die Beobachtungen über
diese Schwankungen sind noch äusserst lückenhaft und lassen uns nicht
einmal vorläufige Schlüsse ziehen. Immerhin glaubte ich im Reichtum der
Schmarotzerfauna der grossen Räuber, Hecht, Quappe, Forelle, Saibling,
während des ganzen Jahres keine wesentliche Veränderung zu bemerken.
In den Karpfen dagegen vermehren sich die Parasiten nach Arten und
Individuen im ersten Frühjahr, nachdem der Fisch seine Winterruhe mit
einem aktiven Leben vertauscht hat. Der Barsch ist besonders im März
bis Mai reichlich infiziert. Weitere Studien über die Saisonverteilung
der parasitierenden Würmer wären sehr erwünscht, auf ein reiches und
sorgfältig beobachtetes Material sich stützend würden sie sicher zu
interessanten biologisch-faunistischen Resultaten führen.
Dass ein Fisch Parasitenträger ist, scheint fast als der normale
Zustand zu betrachten zu sein. Von 382 nach dieser Richtung[S. 221] geprüften
Bewohnern des Genfersees war kaum ein Drittel parasitenfrei.
Lönnberg öffnete in Schweden 870 Fische des süssen und salzigen
Wassers; 564 trugen Parasiten, 306 waren nicht infiziert. Cestoden
beherbergten 128 Individuen, Trematoden 96, Kratzer 294, Nematoden
205. Oftmals bewohnen die Schmarotzer ihren Wirt in gewaltiger
Individuenfülle. Ihre Gegenwart kann so für das infizierte Tier kaum
gleichgültig sein, schon aus dem naheliegenden Grunde nicht, weil die
Parasiten eine Menge Nahrungsstoffe verzehren, die in des fischlichen
Organismus hätten aufgenommen werden sollen. Die Pylor-Anhänge
von Forelle, Saibling, Hecht, Aesche sind meist vollgepfropft
von unzählbaren Exemplaren eines Bandwurms (Bothriocephalus
infundibuliformis Rud.). In einem Hecht von fünf bis sechs Kilo
fand ich mehr als dreihundert Exemplare dieses Schmarotzers, von 28 bis
35 cm Länge; ungezählte hunderte von jungen Würmern waren über
die Schleimhaut des ganzen Darmes zerstreut. Grosse Massen desselben
Cestoden bevölkern in der Regel auch den pylorischen Darmteil des
Lachses, wie ja die Pylor-Anhänge überhaupt der Lieblingsstandort
vieler Fischparasiten sind. Die Würmer finden dort neben sehr
reichlicher Nahrung sichere Wohnung. Auch ein anderer grosser Bandwurm
(bis 50 cm lang), der Triaenophorus nodulosus Rud.,
stellt sich oft in 80 bis 100 Exemplaren im Hecht ein. Ein Saugwurm,
Distoma nodulosum Zeder, bewohnt in grösster Menge den Darm
des Barsches; ein anderes Distoma, D. folium Olfers,
füllt oft buchstäblich die ganze Harnblase der Groppe an, so dass
dieses Organ prall aufgetrieben erscheint; die Kapseln eines larvären
Saugwurms, des Tetracotyle Percae, durchsetzen fast regelmässig
in grösster Menge die Wandungen der Schwimmblase der Barsche. Die
beiden Kratzer Echinorhynchus proteus Westrumb. und E.
angustatus Rud. bewohnen das Eingeweide ihrer zahlreichen Wirte oft
in hunderten von Individuen. Von den Spulwürmern tritt in bedeutender
Zahl auf der Kappenwurm, Cucullanus elegans Zeder, ein für
den Barsch äusserst charakteristischer Parasit; im Hecht findet sich
oft zahlreich die Ascaris acus Bloch, in der Leibeshöhle
der verschiedensten Fische spiralig[S. 222] eingerollt, und allen Organen
angeheftet, die noch geschlechtslose Agamonema capsularia Dies.
Diese wenigen Beispiele mögen das massenhafte Auftreten einer
Parasitenspezies im Fischkörper illustrieren. Es sei nur noch darauf
hingewiesen, dass auch die Larve des breiten Bandwurms des Menschen in
ein und demselben Wirt sich massenhaft einstellt. So schälte ich aus
Leber, Milz, Nieren, Wandungen des Darmkanals einer Seeforelle über
200 Finnen des Bothriocephalus latus. Es erklärt sich dadurch
leicht die gelegentliche Masseninfektion des Menschen mit dem breiten
Bandwurm. Roux trieb einer einzigen Person über 90 Exemplare
gleichzeitig ab.
Die verschiedensten Arten parasitischer Würmer können übrigens
nebeneinander in ein und demselben Wirt leben. Dass ein Fisch
gleichzeitig fünf bis acht Schmarotzerformen beherbergt ist keine
Seltenheit. Im Lachs ist oft der pylorische Darmabschnitt mit
Bandwürmern angefüllt, während zahlreiche Individuen des für diesen
Fisch typischen Distoma varicum im Schlund neben verschiedenen
Spulwürmern leben und die Darmwandungen reichlich gespickt sind mit
Larven von Tetrarhynchen, Bothriocephalen und aufgerollten Agamonemen.
Barsch, Hecht, Aalraupe, Saibling geben oft wahre Parasitenherbergen
ab, in denen gleichzeitig acht bis zehn Arten Schmarotzer, meist in
grosser Individuenzahl, als ungern gesehene Gäste die verschiedensten
Organe bewohnen. Die zahlreichen Parasiten dürften das Leben des
Fisches in seiner kühlen Wasserheimat kaum so angenehm und wohlig
gestalten, wie man es sich etwa vorstellt.
Ist schon der Süsswasserfisch von sehr zahlreichen Würmern heimgesucht,
so trifft das noch in bedeutend höherem Masse für seine marinen
Verwandten zu, die in weit grösseren Verhältnissen leben und unter dem
Einflusse viel mannigfaltigerer äusserer Lebensbedingungen stehen.
Die Parasitenfauna der Meerfische ist mindestens ebenso reich an
Individuen und sehr viel mannigfaltiger an äusserst verschiedenartigen
Formen, als die der Süsswasserbewohner. Wenige Angaben mögen wenigstens
einen kurzen Vergleich der[S. 223] bezüglichen Verhältnisse gestatten.
Lönnberg suchte Parasiten in 342 Meerfischen; 241 davon waren
infiziert, 101 parasitenfrei. 78 trugen Bandwürmer, 39 Saugwürmer,
28 Kratzer, 114 Spulwürmer. In Neapel waren von 257 Fischen nur 74
parasitenlos. Von 72 Arten erwiesen sich 53 als infiziert, bei 34
waren sämtliche untersuchte Exemplare mit Würmern besetzt. Es wurden
in den 257 Fischen 77 Parasitenarten — 38 Bandwürmer, 16 Saugwürmer,
3 Kratzer und 20 Spulwürmer — gefunden, während 382 Fische des
Genfersees nur 35 Schmarotzer aus dem Kreise der Würmer (11 Bandwürmer,
11 Saugwürmer, 3 Kratzer, 10 Spulwürmer) beherbergten. Auch im
Meere ist die Parasitenfauna der grossen Räuber, Haie und Rochen,
viel reicher und aus anderen Elementen zusammengesetzt, als die der
kleineren Knochenfische. Nur sechs Parasitenarten waren Selachiern und
Teleosteern gemeinsam, erstere waren ausserdem von 34, letztere von 36
Schmarotzern heimgesucht.
Es wurden auf ihre Parasiten geprüft:
Zahl der Arten
Zahl der Individuen
Zahl der Parasitenarten
Selachier:
20
96
40
Teleosteer:
34
160
42
Ganoiden:
1
1
1
Die Hauptmasse der Parasiten von Rochen und Haien setzt sich aus
geschlechtsreifen Bandwürmern und teilweise aus Saugwürmern zusammen,
während die Teleosteer vorzüglich Spulwürmer, Kratzer, Saugwürmer und
larväre Bandwürmer beherbergen.
Nach diesen allgemeinen und einleitenden Auseinandersetzungen sollen
eine Anzahl der häufigsten und praktisch wie wissenschaftlich
wichtigsten Schmarotzer unserer Süsswasserfische spezieller besprochen
werden. Bau und besonders Entwickelungsgeschichte vieler, ja der
meisten Formen ist uns noch unbekannt. Doch werden wir in dem, was
durch die Arbeit der Forscher durchsichtig und zugänglich gemacht
worden ist, manche spezielle Illustration zu den vorangehenden
allgemeinen Betrachtungen über die Parasiten der Fische des Süsswassers
finden.
[S. 224]
Die an und in unseren Süsswasserfischen schmarotzenden Krustaceen
und die Muschellarven (Glochidien), welche die Oberfläche der Fische
während einer gewissen Zeit parasitisch bewohnen, haben an anderer
Stelle dieses Werkes bereits Berücksichtigung gefunden. Hier wäre es
somit nur die Aufgabe, der Fischgäste aus dem Kreise der Würmer zu
gedenken. Sie rekrutieren sich aus sehr verschiedenen Abteilungen des
Wurmreiches. Vertreten sind unter diesen Schmarotzern die Blutegel
(Hirudinei), die Spulwürmer (Nematodes), die Kratzer
(Acanthocephali) und zwei Gruppen der Plattwürmer, die
Saugwürmer (Trematodes) und die Bandwürmer (Cestodes).
Kleine Blutegel schmarotzen häufig und oft in ziemlich bedeutender Zahl
auf der Oberfläche karpfenartiger Fische, Karpfen, Schleihen, Barben;
sie sind aber auch im Schlunde der Hechte und an der Körperbedeckung
des Rheinlachses angetroffen worden. Bei letzterem Wirt scheinen sie
Exemplare zu bevorzugen, die infolge langer Wanderschaft wundgeriebene
Stellen an Bauch und Flanken aufweisen. Die Würmer gehören zur Gattung
Piscicola, aus der Gruppe der Rüsselegel. Ausser einer vorderen,
kleineren und hinteren, doppelt so grossen Haftscheibe besitzen sie
in der Mundhöhle einen kräftigen, vorstreckbaren Rüssel. Acht Augen
sind paarweise auf dem vorderen Saugnapfe verteilt. Der gestreckte,
cylindrische Körper verschmälert sich nach vorn nur schwach. Die
Farbe ist gelblich oder aschgrau mit feinen braunen Punkten, und
einzelnen Reihen hellerer elliptischer Flecke. Nach anatomischen
Unterschieden des Magendarms und der Geschlechtsorgane wurden zwei
Arten, Piscicola geometra L., und P. respirans Troschel,
unterschieden. Der Parasitismus der Fischegel darf nur als ein
temporärer bezeichnet werden. Es werden die Würmer nach Leydig
häufig frei schwimmend, oder zu mehreren an der Unterfläche der Steine
dicht zusammensitzend angetroffen. Ist dem Nahrungsbedürfnis durch
Aufnahme von Fischblut für einmal wieder Genüge gethan, so verlässt die
Piscicola den Wirt, sich so in der Lebensweise scharf von dem
Krebsegel (Branchiobdella) unterscheidend, der sein ganzes Leben
auf dem Flusskrebs schmarotzend zubringt. Die[S. 225] kleinen, gelbroten,
längsgestreiften Eier der Piscicolen sollen auf der Aussenfläche der
Fische befestigt werden.
Fig. 34. Cucullanus elegans aus dem Barsch.
Die weite Gruppe der Nematoden oder Fadenwürmer, zum grössten Teil aus
Parasiten verschiedensten Grades zusammengesetzt, liefert auch für die
Süsswasserfische ein stattliches Kontingent von Schmarotzern. Es sind
alles cylindrische, gestreckte, von einer derben Cuticula umhüllte
Würmer, mit endständigem Mund und etwas vor dem Hinterende liegendem
After. Der meist wohl entwickelte Darmkanal zerfällt in der Regel in
drei bis vier mehr oder weniger differente Abschnitte; die Geschlechter
sind beinahe immer getrennt.
Es ist fast unmöglich, den Darmkanal eines Flussbarsches zu öffnen,
ohne in den Pylor-Anhängen in oft sehr beträchtlicher Zahl kleine
Rundwürmer (♀ 12–18 mm lang, ♂ 5–8 mm) anzutreffen,
die durch ihre gelbe oder selbst grell rote Farbe sich von den sonst
farblosen Parasiten auffallend unterscheiden. Die lebhaft beweglichen
Tiere sind auch in anderen Fischen — Hecht, Forelle, Quappe, Felchen,
Zander, Kaulbarsch — zu Hause. Was sie ganz besonders auszeichnet,
ist die starke, kappenförmige, hornige Mundkapsel, der sie auch
ihren Namen, Kappenwürmer (Cucullanus elegans Zed.) (Fig. 34)
verdanken. Der merkwürdige, eine weite Mundhöhle umschliessende Apparat
besteht im wesentlichen aus einer längsgestreiften Kapsel. Sie steht
jederseits in Beziehung mit zwei kleineren, nach hinten gerichteten
Hornstäben. Am Vorderrand ist die Kapsel durch vier dreieckige
Ansatzstücke verstärkt. Nach hinten wird der ganze Apparat durch einen
Chitinring abgeschlossen. Männchen und Weibchen weichen in äusserer
Erscheinung und innerem Bau nicht[S. 226] unbeträchtlich von einander ab, eine
bei Nematoden gewöhnliche Erscheinung.
Fig. 35.
Larve von Cucullanus elegans aus Cyclops.
Die Entwickelungsvorgänge im befruchteten Ei der Kappenwürmer sind
von Bütschli zum Gegenstand einer Untersuchung gemacht worden.
Leuckart verfolgte das weitere Schicksal der Embryonen, die
schon im Muttertier die zarten Eihüllen verlassen und oft zu tausenden
die Geschlechtswege anfüllen. Trächtige Weibchen fand ich während
des ganzen Jahres. Die Jungen sind etwa 0.4 mm lang, äusserst
beweglich; ihr Darm ist noch einfach, ohne weitere Einteilung, der
komplizierte Mundbecher fehlt oder ist nur durch einen Chitinzahn
vertreten; das Hinterende läuft in einen langen, spitzen, lebhaft
schlagenden Schwanz aus (Fig. 35). In diesem Zustande durch eine
starke Cuticula genügend geschützt, leben die Tierchen während Wochen
im Wasser. Früher oder später wird ein Zwischenwirt bezogen, in der
Regel ein kleiner Krebs, aus der Gruppe der Cyclopiden, seltener
Insektenlarven. Die jungen Parasiten werden durch die Mundöffnung
aufgenommen, oft in grosser Zahl. Leuckart zählte in einem
Cyclops nicht weniger als 34 Eindringlinge. Im Zwischenwirt
durchbrechen sie mit ihrem Bohrzahn die Darmwand und gelangen so in die
Leibeshöhle, wo sie nach wenigen Tagen eine Häutung eingehen, die ihren
Bau wesentlich verändert. Der Bohrzahn, der ja seinen Zweck erfüllt
hat, ist verschwunden, der Schwanz ist kürzer geworden; der Ösophag
zeigt eine erste Andeutung der zukünftigen Gliederung in Muskel- und
in Drüsenmagen. In der Folge entwickelt sich allmählich der chitinöse
Mundbecher zu seiner typischen Gestalt. Eine neue Häutung tritt
ein; das Nervensystem und der Hautmuskelschlauch erscheinen in der
definitiven Form, während die Geschlechtsorgane noch wenig entwickelt
sind und das Hinterleibsende dem des erwachsenen Tieres noch unähnlich
aussieht. Auch Grösse und Skulptur des Mundbechers weichen noch von den
bezüglichen Verhältnissen beim erwachsenen Tier ab. Im Sommer braucht
der Entwickelungsgang des Cucullanus bis zu diesem Punkte[S. 227] nur
wenige Tage, im Winter dagegen mehrere Wochen. Damit ist aber auch die
Grenze der im Zwischenwirt möglichen Ausbildung erreicht. Jetzt müssen
die Kappenwürmer eingeschlossen in die Cyclopiden auf den definitiven
Wirt, den Barsch, übertragen werden. Mit der Nahrung nimmt der Fisch
gleichzeitig die jungen Schmarotzer auf. Im Magen des Hauptwirtes
freigeworden, beziehen die Kappenwürmer die Pylor-Anhänge, wachsen dort
rasch heran, um nach einer weiteren Häutung bald die Geschlechtsreife
zu erreichen. Schon zehn bis vierzehn Tage nach der Infektion findet im
Darm des Barsches die Begattung der Würmer statt.
Einen ähnlichen den Stempel des Parasitismus tragenden
Entwickelungsgang mit Übertragung des jungen Schmarotzers auf einen
Zwischenwirt, in dem längere Zeit ein latentes Leben geführt werden
kann, bis wiederum mit der Nahrung der Parasit in den definitiven Wirt
eingeschmuggelt wird, durchläuft ein bekannter Fadenwurm des Hechts,
die Ascaris acus Bloch.
Fig. 36.
Larve von Ascaris acus (Trichina cyprinorum) aus den
Weissfischen.
Schon seit längerer Zeit sind aus vielen karpfenähnlichen Fischen
kleine Nematoden bekannt, die wenig umfangreiche (bis 1 mm
lange) Kapseln der Mesenterien und des Lebergewebes bewohnen. Im
Genfersee sind die betreffenden, höchstens 2 mm langen Würmer
speziell in der Laube (Alburnus lucidus) häufig. Verwandte
Arten sind von v. Linstow in der Quappe und Aesche eingekapselt
gefunden worden. Die schlanken Tierchen zeichnen sich durch einen
weiten Mund mit kräftigem Bohrzahn und einen drüsigen, vom Pharynx sich
abzweigenden Blindsack aus. Sie sind geschlechtslos. Diesing
legt ihnen in seinem Hauptwerk „Systema Helminthum“ fälschlich den
Namen Trichina cyprinorum bei (Fig. 36). Offenbar sind sie vom
Darm her in Leber und Mesenterien der Weissfische eingewandert. Im
Zwischenwirt können sie wohl längere Zeit ein latentes Leben führen,
eine für die Erhaltung der parasitischen[S. 228] Spezies wichtige Eigenschaft.
Die Dauer der Möglichkeit einer erfolgreichen Übertragung auf den
Hauptwirt wird so verlängert und der Verbreitung des Schmarotzers
dadurch kräftiger Vorschub geleistet. Während seines Aufenthalts im
Zwischenträger gehen mit dem jungen Wurm keine weiteren Veränderungen
vor sich; er wächst bis zu einem gewissen Grade, ohne eine Metamorphose
durchzumachen, ein Verhalten, das von dem des Cucullanus
bedeutend abweicht. Mit den Weissfischen, der Lieblingsnahrung des
Hechtes, gelangen die Nematoden in den Magen dieses grossen Räubers
des Süsswassers. Leuckart fand sie dort wieder eingekapselt
in den Wandungen des Verdauungstractus, dann aber auch frei in den
verschiedensten Grössen und Entwickelungsstadien Magen und Darm
des Wirtes bevölkernd. Von der Trichina cyprinorum bis zur
ausgewachsenen, geschlechtsreifen Ascaris acus liessen sich alle
wünschbaren Übergangsformen nachweisen. Der Bohrzahn geht verloren;
die drei für Ascaris charakteristischen Lippen erscheinen am
Umfange der Mundöffnung; die Geschlechtsorgane wachsen schlauchförmig
aus. Dagegen bleibt im erwachsenen Tier jener eigentümliche Blindsack
des Ösophagus bestehen, der schon die junge Larve in den Weissfischen
auszeichnet und so junge und alte Form leicht auf einander zurückführen
lässt.
Ascaris acus Bloch ist kein seltener Gast im Darm und auch in
der Leibeshöhle von Hecht, Quappe und Forelle. In jüngeren Stadien,
frisch aus den Weissfischen stammend, lebt sie zunächst eingekapselt
in den Wandungen des Verdauungstractus ihrer Wirte, wie dies
Leuckart beobachtete. Nachdem sie eine bestimmte Grösse erreicht
hat, bricht sie nach dem Darm oder der Leibeshöhle durch. Länge des ♀
bis 40 mm, des ♂ bis 30 mm. Die Mundlippen sind stark
entwickelt, Cuticula deutlich quer gestreift. Wie bei fast allen
Parasiten werden Eier in grösster Menge gebildet. An diesen Ascariden
des Hechts hat Bunge durch eine Reihe interessanter Experimente
nachzuweisen verstanden, dass sie vier bis sechs mal 24 Stunden in
vollkommen sauerstofffreien Medien leben können.
[S. 229]
Eingekapselte, geschlechtslose Nematoden sind in den verschiedensten
Organen von Fischen des süssen und salzigen Wassers eine ganz
gewöhnliche Erscheinung. Über ihre Zugehörigkeit zu erwachsenen
Formen aber sind wir bis jetzt in den wenigsten Fällen genügend
aufgeklärt. Äusserst verbreitet und oft in ungeheuerer Zahl auftretend
ist in sehr vielen Fischarten die Agamonema capsularia Dies.,
die wahrscheinlich mit der unter ähnlichen Umständen vorkommenden
Ascaris capsularia Rud. und der Filaria piscium Rud.
zusammenfallen dürfte. Es liegen die Würmer spiralig aufgerollt und
von einer leichten Kapsel umschlossen an und in den verschiedensten
Organen: Leber, Nieren, Milz, Geschlechtsorganen, im Peritonäum, auf
der Aussenfläche des gesamten Verdauungstractus, besonders zwischen
den Appendices pyloricae. Aus ihrer Kapsel befreit, schwimmen
die glänzenden, äusserst beweglichen, schlanken Würmer sehr lebhaft im
Wasser. Es gelang mir in zahlreichen Versuchen Exemplare aus dem Lachs
Monate lang in kleinen Wassermengen am Leben zu erhalten, nie aber
nach vollkommener Austrocknung dieselben durch Anfeuchtung wieder zu
beleben. Länge bis 20 mm, Tegument glatt, keine Seitenmembranen,
Vorderende etwas verschmälert, drei kleine rudimentäre Lippen. Der
Ösophag trägt auch hier wieder ein kleines seitliches Caecum. Keine
Geschlechtsorgane. Früher wurden diese „Filarien“ fälschlich zum
Entwickelungscyklus gewisser die marinen Fische bewohnender Cestoden
(Tetrarhynchen) gezogen! Wahrscheinlich erreicht Agamonema
capsularia die Geschlechtsreife in grossen Raubfischen. Wenigstens
fand ich im Magen eines Haifisches freie Agamonemen neben halbverdauten
Resten eines Meeraals, ein Fisch, der mit larvären Würmern oft besetzt
ist. Neben den Cysten der Agamonemen liegen in manchen Organen
vieler Süsswasserfische noch Kapseln anderer Nematoden. Kapsel und
Insasse aber unterscheiden sich durch Lage, Form, Bau leicht von
den soeben beschriebenen Bildungen. Sie gehören wohl verschiedenen
geschlechtsreifen Arten von Fadenwürmern an, doch ist ihr Zusammenhang
mit erwachsenen Formen nur sehr wenig klargestellt.
[S. 230]
An die Klasse der Nematoden schliesst man gewöhnlich die Gruppe der
Kratzer, Acanthocephalen oder, wie sie nach ihrer einzigen Gattung
genannt wird, der Echinorhynchen an. Es sind dies ohne Ausnahme
typische Darmschmarotzer. Ihr Bau verrät sofort den Parasitismus durch
den mit kräftigen Haken bewaffneten, in eine Scheide zurückziehbaren
Rüssel, der als starker Haftapparat dient, und durch die völlige
Abwesenheit des Verdauungssystems. Die Nahrungssäfte werden aus dem
Wirt osmotisch durch die ganze Körperoberfläche aufgenommen. Im
gestreckten, oder eiförmigen Körper liegen, von einem aus Quer- und
Längsfasern zusammengesetzten Muskelschlauch umschlossen, hauptsächlich
die kompliziert gebauten Geschlechtsorgane. Die Geschlechter sind
getrennt, die Fruchtbarkeit erfährt auch hier eine für den Parasiten
so wünschenswerte starke Steigerung. Im Grunde der Rüsselscheide
entwickelt sich das Nervensystem in Gestalt eines Ganglions mit
davon ausstrahlenden Nerven; in die Leibeshöhle ragen noch zwei
eigentümliche, schlauchartige Bildungen, die Lemnisken, denen man die
Funktion von Exkretionsorganen zuschreibt. Die Lebensgeschichte der
Echinorhynchen ist durch das Schmarotzertum stark beeinflusst.
Kratzer im Darme von Süsswasserfischen sind eine recht häufige
Erscheinung. Besonders verbreitet in allen Fischen aus der Familie
der Karpfen, aber auch in Barsch, Quappe, Hecht, Aal, Stör, Forelle,
Aesche, Groppe, Saibling, Kaulbarsch findet sich der Echinorhynchus
proteus Westrumb. Oft ist der Enddarm des Wirtes von den rötlichen
oder gelblichen Schmarotzern prall angefüllt. Die Länge der Tiere
beträgt bis 30 mm; vom Januar bis Juni nahm sie bei den
Exemplaren aus dem Genfersee stetig zu. Äusserlich zeichnet sich die
Art durch einen langen, schmalen, unbewehrten Hals aus, der an der
Übergangsstelle zum Rüssel eine durchaus charakteristische, kugelige,
als Bulla bezeichnete Bildung trägt. Der Rüssel selbst ist nach
Hamanns neuester, schöner Arbeit mit 23 bis 25 Querreihen
von Haken bewaffnet, die drei verschiedenen Formen angehören. Eine
verwandte, von Hamann zuerst genau festgestellte Art, der E.
Linstowi, besitzt nur zehn Hakenreihen mit zweierlei Formen der
Haftgebilde.[S. 231] Gesamtzahl der Haken für E. proteus 230–250, für
E. Linstowi 60. Die Grösse der Exemplare von E. proteus
richtet sich nach der Grösse des bewohnten Fisches, ein Verhältnis, das
für Wirt und Gast bei parasitischen Geschöpfen überhaupt oft gültig
ist. In der Jugend sind die Echinorhynchen noch kaum angeheftet; ältere
Individuen versenken Rüssel und Hals tief in die Schleimhaut des
Fisches.
Die reifen, bereits embryonenhaltigen Eier fallen, nachdem das
Ovarium geplatzt ist, in die Leibeshöhle; sie werden von einem
eigentümlichen, sich fortwährend öffnenden und schliessenden Teil
des Geschlechtsapparats, der Uterusglocke, aufgeschluckt und von
da durch die Leitungsapparate an die Aussenwelt abgegeben. Von
einzelnen Kratzern werden so Millionen sehr kleiner Eier in der
freien Natur zerstreut. Ihre Entwickelungsfähigkeit bleibt lange
Zeit eine ungestörte. Die Embryonen sind in drei starke Hüllen von
verschiedener Natur eingeschlossen. Ihre Gestalt ist die eines
schlanken Kegels mit abgerundeten Enden; das breitere, ventral
scheibenartig abgeflachte Vorderende trägt einen aus zehn bis zwölf
stilettförmigen Borsten zusammengesetzten Stachelapparat, der durch
eine spezielle Muskulatur bewegt werden kann. Leuckart hat
schon vor längerer Zeit nachgewiesen, dass die Eier unseres E.
proteus in den Darm des weit verbreiteten, gewöhnlichen Flohkrebses
(Gammarus pulex) gelangen müssen. Die Eihülle und endlich auch
die Darmwand des Zwischenwirtes wird von den Embryonen mit Hilfe ihres
Stachelapparates durchbrochen; in der Leibeshöhle des Krebses läuft
die weitere Entwickelung ab. Noch drei Wochen bleiben die Bewegungen
im Zwischenwirt recht lebhafte; nach und nach werden sie langsamer,
um endlich ganz aufzuhören. Der embryonale Hakenkranz ist inzwischen
verloren gegangen; die Tiere liegen wie Puppen von Insekten, von ihrer
äusseren, derben Haut wie von einer Kapsel umgeben und mit eingezogenem
Rüssel in der Leibeshöhle des Gammarus. Die weitere Entwickelung
des scheinbar ruhenden Körpers ist eine höchst komplizierte. Aus
einer zentralen Körnermasse der Larve, dem sogenannten Embryonalkern,
geht die Hauptmasse der Organe,[S. 232] Nervensystem, Rüsselscheide,
Geschlechtsapparate, hervor, während die Körperwandung mit den
Lemnisken, dem eigentümlichen, sie durchziehenden Lakunensystem und
dem Muskelschlauch den peripherischen Teilen des Embryos den Ursprung
verdankt.
Eingeschlossen in den Zwischenwirt müssen die jungen Echinorhynchen
in den Verdauungstractus der ihnen als Hauptwirte zusagenden Fische
gelangen. Dort werden sie frei, fixieren sich, ihre volle Grösse wird
erreicht und nach definitiver Ausbildung der Geschlechtsorgane wird die
Begattung vollzogen.
Hamann hat indessen die Beobachtung gemacht, dass die
Lebensgeschichte noch kompliziertere Bahnen einschlagen kann. In
manchen Fischen, Stichling, Barbe, Groppe, Gründling, besonders
massenhaft aber in der Elritze (Phoxinus laevis) fand der
Göttinger Zoologe der Oberfläche der Leber angeheftet regelmässig
kugelige bis eiförmige Gebilde von etwa 2 mm Durchmesser.
Sie bestehen aus einer von der Leber aus gebildeten Kapsel, die die
orangefarbene Larve einer Echinorhynchus-Art umschliesst. Diese jungen
Tiere aus der Leibeshöhle der kleinen Fischarten entsprechen in
jeder Beziehung den Jugendstadien von Echinorhynchus proteus
aus dem Flohkrebs. Sie müssen unbedingt als zum Entwickelungsgang
des besprochenen Kratzers gehörend betrachtet werden. Es gelang
Hamann sogar, aus den in Fischen gefundenen Larven in der
Forelle den typischen Echinorhynchus proteus gross zu ziehen.
Also kann ein Fisch, z. B. die Elritze, so gut wie der Flohkrebs zum
Zwischenwirt eines Kratzers werden, der selbst wieder Raubfische
bewohnt. Die genannten kleinen Fische können übrigens gleichzeitig
Haupt- und Zwischenwirt des Echinorhynchus proteus sein. Sie
beherbergen ihn als geschlechtsreifen Wurm massenhaft im Darmkanal,
gleichzeitig findet er sich ganz konstant, oft in der Zahl von
zwanzig Exemplaren, als geschlechtslose Larve der Leber und den
Mesenterien angeheftet. So sucht ja auch die Trichine den Menschen in
zwei verschiedenen Entwickelungsstadien heim, als geschlechtsreifer
Darmbewohner und als in den Muskeln ruhende, unreife Larve.
Hamann nimmt an, dass die Elritzen u. s. w. gelegentlich
Flohkrebse verschlucken, in deren Darm frisch aufgenommene, noch[S. 233] nicht
geöffnete Eier von E. proteus liegen. Die Embryonen gelangen
nun im Fischdarm zum Ausschlüpfen, sie durchbohren die Wandungen
des Verdauungstractus und benützen den Fisch an Stelle des von ihm
verschlungenen Krebses als Zwischenwirt. Ein Raubfisch wie die Forelle
ist also einer doppelten Infektionsgefahr ausgesetzt, sie kann den
Echinorhynchus vom Fisch oder vom Krebs beziehen. Für die
Verbreitung des Parasiten aber wird die geschilderte Vermehrung der
Zahl von Zwischenwirten von Bedeutung und Vorteil sein.
Fig. 37. Echinorhynchus angustatus.
Kaum weniger verbreitet als der Echinorhynchus proteus
ist ein anderer Kratzer zahlreicher Süsswasserfische, der E.
angustatus Rud. Als seine gewöhnlichsten Wirte dürfen wohl Barsch,
Hecht, Quappe, Karpfen und Forelle bezeichnet werden. In gewissen
Bächen, die den Zwischenwirt des Wurmes, die Wasserassel (Asellus
aquaticus), häufig beherbergen, ist auch der Darm der Forellen
regelmässig mit hunderten der Schmarotzer angefüllt. Der gestreckte,
spindelförmige oder cylindrische Körper besitzt nur einen kurzen Hals.
Der walzenförmige Rüssel trägt 8–20 Hakenreihen, deren Elemente von
zweierlei verschiedener Form sind. Bei mittelgrossen Tieren mit 15
Reihen beträgt die Hakenzahl 120 (Fig. 37).
Es kamen mir keine Exemplare von E. angustatus aus
Süsswasserfischen zu Gesicht, deren Gesamtlänge 25 mm überstieg.
Die Lebensgeschichte dieses Kratzers kopiert die für E. proteus
näher geschilderten Vorgänge. Wie dort bedarf die Larve bis zu ihrer
vollständigen Ausbildung acht bis zehn Wochen Zeit. Die Eier des
Parasiten werden von der Wasserassel in grosser Menge aufgenommen. Im
Gegensatz aber zu den bereits geschilderten Verhältnissen gelangen
die ausschlüpfenden Embryonen von E. angustatus schon in
der Drüsenschicht des Darmes ihres Zwischenträgers, die sich in
beträchtlicher Dicke zwischen Chitinwand und[S. 234] Muskelhaut legt, zur
Ruhe. Hier liegen sie in grosser Zahl bewegungslos. Die äussere Form
der Larven verändert sich; innere, komplizierte Umbildungen finden
statt; die Körpergrösse nimmt zu. Endlich fallen die jungen Schmarotzer
infolge von in der Wandung des Darmes eintretenden Veränderungen
passiv in die Leibeshöhle des Zwischenwirtes. Von der Blutflüssigkeit
des Krebses umspült, und so reichlich ernährt, wächst der Schmarotzer
verhältnismässig rasch unter allmählicher Gestaltveränderung. Es
entsteht auf verwickeltem Wege der junge Echinorhynchus, der mit
der Assel auf den Hauptwirt (Fische) übertragen werden muss, um dort
seine Geschlechtsreife zu erlangen.
Für einen andern Kratzer, den E. clavaeceps Zed., hat
Villot den Zwischenwirt in der wasserbewohnenden Larve eines
weit verbreiteten und massenhaft auftretenden Netzflüglers, der
Sialis lutaria L., entdeckt. Der junge Wurm liegt eingekapselt
im Fettkörper des Insekts; er sieht in Form und Bau dem erwachsenen
Echinorhynchus sehr ähnlich, doch sind die Geschlechtsorgane
noch unentwickelt. Zufällig finden sich die Larven des E.
clavaeceps auch in einem Blutegel (Nephelis octoculata)
und in Wasserpulmonaten (Limnaeen); doch scheint die Sialis-Larve
der richtige Zwischenträger zu sein. Ihr wird vom Hauptwirt des
betreffenden Kratzers, der Schmerle (Cobitis barbatula), eifrig
nachgestellt. E. clavaeceps ist übrigens auch kein seltener Gast
zahlreicher Karpfenarten.
Die formenreiche Gruppe der Trematoden oder Saugwürmer umschliesst
ausschliesslich parasitierende Wesen. Doch leben sie bald als
Entoparasiten in den inneren Organen ihres Wirtes, bald heften sie
sich ektoparasitisch auf seiner Oberfläche an. Es sind meist flache,
blattförmige, ungegliederte Würmer, die als Haftapparate Saugnäpfe
und oft daneben noch Hakengebilde besitzen. Eine Leibeshöhle fehlt;
der gegabelte, oft vielfach verzweigte Darmkanal endigt immer blind.
Über dem Schlunde liegt ein Doppelganglion, das mehrere Nervenstämme,
speziell zwei nach hinten ventral verlaufende, abgiebt. Röhriges, meist
durch einen unpaarigen hinteren Porus ausmündendes Exkretionssystem,
das mit zahlreichen[S. 235] Wimpertrichtern im Parenchym beginnt. Die
Trematoden sind fast ausschliesslich Zwitter mit sehr komplizierten
Geschlechtsapparaten. Ihre Lebensgeschichte ist oft äusserst
verwickelt. Sie setzt sich besonders bei den entoparasitischen Formen
für ein und dieselbe Art aus mehreren, auf verschiedene Weise aus
einander hervorgehenden Generationen zusammen (Generationswechsel).
Freie Stadien wechseln mit parasitischen; oft werden mehrere Wirte
durchlaufen.
Von den entoparasitischen Formen, die mit höchstens zwei Sauggruben,
nie mit Haken bewehrt sind, stellt besonders die artenreiche Gattung
Distoma zahlreiche Schmarotzer für die Süsswasserfische.
Stossich kennt 170 Distomeen aus etwa zweihundert Fischarten;
davon parasitieren etwa vierzig Arten in Süsswasser- und Wanderfischen.
Sie bewohnen oft in sehr grosser Individuenzahl hauptsächlich die
verschiedenen Abschnitte des Verdauungstractus, vom Schlund bis
zum Enddarm. Doch fehlen sie auch nicht in anderen, nach aussen
offenstehenden Organen, wie z. B. in der Harnblase (Distoma
folium Olfers bei Groppe, Hecht, Forelle, Aesche, Saibling). In
Körpergestalt, Grösse und Bau weisen die Distomeen der Süsswasserfische
gar mancherlei Verschiedenheiten auf. Als Extreme seien erwähnt das
D. tereticolle Rud. aus dem Schlund von Hecht, Aalraupe,
Forelle, Saibling etc. mit gestrecktem bis 50 mm langem,
halbcylindrischem Körper, und das schon angeführte D. folium
aus der Harnblase mancher Fische, dessen Leib eine flache Scheibe von
kaum 2 mm Durchmesser bildet, von der ein Halsteil sich scharf
abhebt. In zahlreichen Fischen lebt das D. appendiculatum Rud.,
ausgezeichnet durch einen in den Körper rückziehbaren Schwanzteil; im
Schlunde des Lachses stösst man oft auf das auch sonst verbreitete
D. varicum Zeder mit drehrundem, geknicktem Körper, konkaver
Rücken- und konvexer Bauchfläche. In den Karpfen lebt das durch seinen
kugeligen Bauchsaugnapf ausgezeichnete D. globiporum Rud.
Fig. 38.
Flimmerembryo von Distoma laureatum.
Über die Lebensgeschichte der die Süsswasserfische bewohnenden
Distomeen ist uns fast nichts bekannt. Sie wird wohl wie diejenige
verwandter Formen durch mehrere provisorische Wirte[S. 236] führen und für ein
und dieselbe Art aus verschiedenen Generationen zusammengesetzt sein.
Parasitische und freie Zustände werden mit einander wechseln, bis sich
endlich ein junges geschlechtsloses Distoma entwickelt hat,
das eingeschlossen in einen letzten Zwischenwirt, wie beim Leberegel
unserer Rinder und Schafe, in den Darm des Hauptwirtes gelangt. Die
Fruchtbarkeit der in Frage stehenden Saugwürmer ist eine gewaltige.
Bei dem abscheulichen Ausbeutungssystem, das den Parasitismus nun
einmal charakterisiert, können Stoffe zur Ausarbeitung der Eier in
Hülle und Fülle aufgenommen werden. So wird der weitern Verbreitung
der schmarotzenden Spezies starker Vorschub geleistet. Die mit einem
kleinen Schalendeckel versehenen Eier werden mit dem Kot des Wirtes
an die Aussenwelt abgegeben. Vom Distoma laureatum Zed. der
Salmoniden berichtet uns v. Willemoes-Suhm, dass den ins Wasser
gelegten Eiern nach Verlauf von 34 Tagen ein grosser, bewimperter
Embryo mit schwarzem Augenfleck entschlüpfte (Fig. 38). Bei Distoma
globiporum ist nach demselben Gewährsmann der länglich-runde
Embryo ebenfalls stark bewimpert. Er besitzt Seitengefässe und trägt
an einem Ende ein saugnapfartiges Gebilde, aus dessen Mitte sich ein
konischer Zapfen erhebt. Der Embryo von D. folium ist herzförmig
mit rudimentärem Mundnapf, Seitengefässen und Flimmerüberzug. Auch
die erste Jugendform von D. nodulosum aus dem Magen von
Hecht, Zander, Barsch etc. zeigt ähnlichen Bau. In seinem schwarzen
Pigmentfleck ist deutlich eine rundliche Linse erkennbar. Der Embryo
von D. tereticolle besitzt im Exkretionsapparat deutliche
Flimmerläppchen. Über die weiteren Schicksale all dieser bewimperten,
freischwimmenden Jugendstadien der Süsswasserdistomeen wissen wir
leider fast gar nichts. Speziell ist es uns sehr oft unbekannt, in
welchen Zwischenwirt eingeschlossen sie endlich in den Darmkanal des
definitiven Wirtes übertragen werden. In manchen Fällen mag der letzte
Zwischenwirt dem Kreis der Mollusken angehören. So lebt die noch
geschlechtslose Larve von[S. 237]D. globiporum Rud. in dünnwandigen
Cysten weitverbreiteter Wasserschnecken (Limnaea stagnalis,
Planorbis marginatus, Succinea putris etc.). Es können
übrigens auch die Süsswasserfische die Rolle von Zwischenträgern von
Trematoden gegenüber anderen Geschöpfen — Raubfischen, Wasservögeln —
spielen.
In der grossen Mehrzahl der Individuen des Flussbarsches ist
die Wandung der Schwimmblase durchsäet mit kleinen, kugeligen,
starkwandigen, opalescierenden Cysten. Sie umschliessen larväre
Trematoden mit gut entwickeltem Mundnapf, an dessen Seite zwei
bohnenförmige Organe — Drüsenausmündungsstellen — liegen. Auch ein
hinterer Bauchnapf lässt sich nachweisen. Der Leib ist reichlich mit
Kalkkörperchen durchsetzt; er umschliesst einen kurzen gegabelten
Darm. Es sind diese Larven mit dem Namen Tetracotyle belegt
worden. Ähnliche junge Saugwürmer siedeln sich nicht selten im Auge
verschiedener Fische, Barsch, Quappe, Zander, Karpfen, an, wo sie als
Diplostomum beschrieben worden sind. Eine Kapsel fehlt ihnen.
v. Nordmann fand in der Linse einer Quappe 290 solcher Würmer,
im Glaskörper 157.
Es ist in hohem Grade wahrscheinlich gemacht worden — siehe ausser
Leuckart die Arbeit von Brandes —, dass diese
Tetracotylen und Diplostomen, die ausser Fischen auch Mollusken,
Amphibien, Reptilien, Vögel und Säugetiere infizieren, im Darme
des Hauptwirtes — Reptilien, Säugetiere und besonders Vögel —
sich zu geschlechtsreifen Vertretern der Familie der Holostomiden
entwickeln. Der bewimperte Embryo dieser Trematoden sucht einen
passenden Zwischenwirt auf und reift in ihm zur völlig ausgebildeten
Holostomiden-Larve heran, die, mit dem Zwischenträger in den Darm
des definitiven Wirtes übergeführt, dort zum geschlechtlichen Tier
auswächst. Ein Generationswechsel wäre in dieser Lebensgeschichte
ausgeschlossen, dagegen wird immer ein Zwischenwirt aufgesucht.
Der Zwischenträger fällt nun bei den sofort zu besprechenden
ektoparasitischen Saugwürmern ebenfalls fort.
Die an der Aussenfläche von Wassertieren, speziell Fischen, lebenden
Polystomeen bedürfen zu ihrer Festsetzung weit kräftigerer Haftorgane
als die entoparasitischen Distomeen. Ist doch für sie[S. 238] die Gefahr,
vom Wirte abgestreift zu werden, eine immer drohende. So wird denn
bei ihnen nicht nur die Zahl der Saugnäpfe bedeutend vermehrt,
sondern es werden dieselben in ihrer Funktion oft noch durch
mannigfaltige Chitinhaken und Borsten unterstützt. Bei diesen mit der
freien Aussenwelt während des ganzen Lebens in Beziehung stehenden
Schmarotzern sind auch Sinnesorgane, besonders Augen, keine Seltenheit,
Bildungen, die bei den Entoparasiten unter den Trematoden höchstens
während der frei schwärmenden Jugendzeit auftreten. Die Polystomeen
bilden meistens nur wenige, aber grosse und oft mit verschiedenen
Anhängen versehene Eier aus. Die denselben entstammenden Embryonen
entwickeln sich am häufigsten ohne Generationswechsel und Wanderungen
zum definitiven Geschlechtstier. Die Eier werden oft mit ihren
Fortsätzen am Wirt befestigt, aus ihnen entspringt ein äusserlich und
innerlich schon hoch differenzierter Embryo. Das für ihn unnötige
Wimperkleid ist schwach, oder gar nicht entwickelt; dagegen besitzt
er schon einfache Haftorgane, die sich meistens später komplizieren.
Die Distomeen liefern ungemein zahlreiche, einfache Embryonen, von
denen wenigstens einige alle mannigfaltigen Wendungen des Geschicks
des entoparasitischen Entwickelungsganges glücklich überstehen werden;
die Polystomeen entlassen aus wenigen, aber dotterreichen Eiern einige
Junge, die im ganzen schon den Eltern ähnlich sind. Zuweilen werden
indessen auch weitergehende Metamorphosen durchlaufen.
In manchen Fällen bietet der Lebensgang der ektoparasitischen
Saugwürmer trotz der direkten Entwickelung Verhältnisse, die zum
eigentümlichsten gehören, was uns tierische Geschichte lehrt. Speziell
unter den Parasiten der Süsswasserfische stossen wir in dieser
Beziehung auf höchst seltsame Vorgänge.
Zeller hat uns in seinen schönen Arbeiten sowohl mit Bau,
als mit Lebensgeschichte des Diplozoon paradoxum Nordm., des
fremdartigen Doppeltieres, bekannt gemacht. Es ist ein Trematode, der
häufig an den Kiemen zahlreicher Süsswasserfische — Elritze, Quappe,
Groppe etc. — schmarotzt. Im ausgewachsenen, geschlechtsreifen
Zustand erscheint der Körper des Diplozoon als ein[S. 239] x- oder
kreuzförmiges Gebilde, das, wie wir sehen werden, durch die dauernde
und enge Verwachsung von zwei ursprünglich getrennten Wurmleibern
entstanden ist. Die Hinterenden besitzen zwei grosse, in vier Gruben
geteilte Haftscheiben, zu denen sich noch vier starke Klammerpaare
gesellen (Fig. 39).
Fig. 39. Diplozoon paradoxum von den Kiemen der Elritze.Fig. 40.
Ei von Diplozoon paradoxum.
Fig. 41.
Flimmerembryo von Diplozoon paradoxum.
Beide das Diplozoon zusammensetzenden Tiere sind gleich,
hermaphrodit, bringen Eier hervor und befruchten sich gegenseitig. Mit
dem Eintritt der kälteren Jahreszeit hört die Eierbildung auf, die
weiblichen Organe existieren dann nur andeutungsweise. Bei steigender
Temperatur (künstlich oder natürlich) beginnt die Ausarbeitung
der Eier sofort wieder, die Geschlechtsorgane entwickeln sich und
treten in Thätigkeit. Die Eibildung und Befruchtung kann nun in
allen Einzelheiten verfolgt werden. Das reife, hochgelb gefärbte,
gedeckelte Ei ist 0.28 bis 0.30 mm lang; seine Schale besitzt
zunächst an einem Pol einen kurzen, schnabelartigen Fortsatz, der
allmählich zu einem langen, sich aufrollenden und erstarrenden
Faden auswächst (Fig. 40). Am siebenten bis achten Tag werden die
Eier aus dem erweiterten Ende des Ovidukts ausgestossen. In reinem
Wasser läuft die weitere Entwickelung rasch ab. Nach acht Tagen
umschliesst die Eischale einen deutlichen Embryonalkörper, der sich
zuerst schwach, dann immer kräftiger bewegt. Am fünfzehnten Tag etwa
wird der Deckel des Eies abgeworfen, der Embryo schwimmt leicht im
Wasser. In diesem Zustand ist das[S. 240] junge Tier, was sich schon durch
die Eischale erkennen liess, mit zwei dorsal liegenden Augen und einem
Wimperbesatz ausgestattet. Die Augen sind Schälchen eines bräunlichen
Pigments, die ein helles, kugeliges, linsenartiges Körperchen
umschliessen. Das ganze Körperparenchym ist reichlich mit glänzenden,
formveränderlichen Kügelchen durchsetzt. Am Vorderende liegt der Mund
mit zwei eigentümlichen seitlichen Saugnäpfen, das Hinterende trägt
zwei Klammern und zwei kleine Angeln. Im Innern des Körpers erscheint
der stark muskulöse, in einen einfachen Darm führende Schlundkopf
(Fig. 41). Die Bewegungen dieser jungen Tierchen sind äusserst
mannigfaltig und ausgiebig. Bietet sich indessen im Verlauf von sechs
Stunden keine Gelegenheit zur Fixierung auf den Kiemen eines Fisches,
so gehen die Diplozoon-Embryonen zu Grunde. Hat die Festsetzung aber
stattgefunden, so verwandelt sich der Embryo zu einem Geschöpf, das
als kiemenbewohnender Parasit schon von Dujardin unter dem
Namen Diporpa beschrieben worden ist. Der unnötig gewordene
Schwimmapparat, der Wimperbesatz, geht verloren; die Augen, die
gleichfalls keinen Dienst mehr zu leisten haben, zerfallen, der Darm
füllt sich mit dem Wirt entzogenen Stoffen. Im Juli und August findet
man die Kiemen der Elritzen oft mit hunderten von Diporpen besetzt.
Es sind diese Wesen von lanzettförmiger, abgeplatteter Körpergestalt;
ihre Bauchfläche trägt etwas unterhalb der Mitte einen kleinen
Saugnapf, am Rücken liegt, noch etwas kleiner als der Bauchnapf,
ein zapfenförmig hervortretendes Gebilde. Das Kopfende des Tieres
verlängert sich rüsselartig, an ihm liegt zentral ein quergestellter
Mund; seitlich öffnen sich zwei Saugnäpfe in die Mundhöhle. Der Darm
beginnt seitliche Ausstülpungen zu treiben. In jeder Körperhälfte liegt
ein Hauptstamm des Exkretionssystems. Am Hinterende ist schon das erste
Haken- und Klammerpaar befestigt. In diesem Zustand auf den Kiemen
schmarotzend und allmählich an Grösse zunehmend, können die Diporpen
Wochen und Monate lang leben. Nie aber wird das isolierte Einzeltier
geschlechtsreif,[S. 241] es muss der Geschlechtsreife die enge, definitive
Verwachsung zweier Diporpen zum Diplozoon vorausgehen, wie
dies schon v. Siebold richtig erkannte. Die Vereinigung findet
immer so statt, dass jedes der beiden sich konjugierenden Individuen
mit seinem Bauchnapf den Rückenzapfen des Gefährten umfasst. Die
notwendige Folge dieser Verbindungsweise wird eine Kreuzung der beiden
Exemplare und so die Annahme der typischen Diplozoongestalt sein.
Eine Trennung findet nicht mehr statt, die Tierkörper verwachsen an
den Berührungsstellen innig; aus zwei ursprünglich getrennten
Individuen, hervorgegangen aus zwei verschiedenen Eiern, ist ein
Geschlechtstier entstanden. Das Diplozoon wächst nun noch
weiter; die vorderen Saugnäpfe gehen verloren, am Hinterende legen
sich ein zweites, drittes und viertes Klammerpaar und entsprechende
Sauggruben an, die übrigens oft schon in älteren Diporpen teilweise
vorgebildet sind. Endlich erscheinen in beiden ursprünglichen Tieren
die Geschlechtsorgane.
Eine ziemlich umfangreiche Gruppe von Kiemenparasiten der Fische
sind die Gyrodactyliden, charakterisiert durch ihre grosse terminale
Schwanzscheibe und den kräftigen Hakenapparat. Im süssen Wasser
sind diese sehr kleinen Schmarotzer in vielen Arten hauptsächlich
auf verschiedenen Cypriniden, sowie Barsch, Kaulbarsch, Zander zu
Hause. Die Gattung Gyrodactylus besitzt zwei Kopflappen,
acht bewegliche Pharynxstacheln, zwei starke Haken inmitten der
Schwanzscheibe und zahlreiche kleine Häkchen am Umfange derselben.
Kaum minder gut mit Haftapparaten ausgestattet ist die verwandte Form
Dactylogyrus. Sie trägt vier Kopflappen, zahlreiche kleine
Randhaken, neben zwei grösseren Klammern auf der Scheibe. Oft stellt
sich noch eine zweite, zentrale Haftplatte ein.
Die merkwürdige Geschichte von Gyrodactylus elegans Nordm. hat
uns G. Wagener geschildert. Das junge Tier macht im Innern der
Mutter seine vollständige Metamorphose durch und wird lebend, nach
Gestalt und Bau dem Muttertier durchaus ähnlich geboren, während z. B.
die verwandte Gattung Dactylogyrus Eier zur Welt bringt. Bevor
aber der junge Gyrodactylus frei wird, umschliesst[S. 242] er selbst
bereits wieder einen Sprössling, so dass drei Generationen in einander
eingeschachtelt liegen. Das Muttertier umfasst die auf geschlechtlichem
Wege erzeugte, aber noch nicht geschlechtsreife Tochter; diese birgt
einen Keimling, der wie angenommen wird einer übrig gebliebenen
Furchungskugel des Eies, aus dem das Tochtertier hervorging,
seinen Ursprung verdankt. Tochter- und Enkelindividuum wären somit
gleichzeitig aus der Masse gleichgestalteter und gleichaltriger
Embryonalzellen des ursprünglichen Eies hervorgegangen.
Unter dem Namen Sphyranura Osleri haben Ramsay Wright und
Macallum jüngst einen interessanten ektoparasitischen Trematoden
von der Haut des grossen Salamanders Necturus lateralis
Raf. aus den Seen Nordamerikas beschrieben. Die fragliche Form
schiebt sich anatomisch und embryologisch zwischen Polystomum
und Gyrodactylus ein. Ein weiterer Kiemenbewohner der
Süsswasserfische ist die Gattung Tetraonchus.
Wie die Saugwürmer gehören auch die Bandwürmer (Cestoden) der grossen
Abteilung der Plattwürmer an. Es fehlt sogar nicht an Formen, die
morphologisch und embryologisch als Übergangsstufen sich zwischen die
beiden parasitierenden Gruppen der Plathelminthen einschieben und
es so unmöglich machen, Trematoden und Cestoden durch eine scharfe
Grenzlinie zu scheiden. Letztere haben sich offenbar aus ersteren
herausentwickelt, sind aus ihnen durch Anpassung an immer schärfer
ausgeprägtes Schmarotzertum entstanden. Von Ektoparasitismus ist hier
keine Rede mehr. Alle Bandwürmer bewohnen die inneren Organe ihrer
Wirte. Unter dem Drucke des intensiver werdenden Schmarotzertums sind
manche bei den Trematoden noch mehr oder weniger entwickelte Organe und
Organsysteme als unnötig vollkommen zurückgebildet worden. Sinnesorgane
und Verdauungssystem sind spurlos verschwunden. Das schwach entwickelte
Nervensystem besteht in der Regel aus zwei seitlichen Längsstämmen,
die im vorderen Körperende, dem sogenannten Kopf, durch einfache
oder mehrfache Kommissuren verbunden sind. Gut ausgebildet ist das
zweiseitig angelegte, röhrig gebaute Exkretionssystem, dessen feinste[S. 243]
Verzweigungen im Körperparenchym mit zahlreichen Flimmerläppchen
endigen. In der Regel ist der Bandwurmkörper mehr oder weniger deutlich
segmentiert, gegliedert. Sein vorderster, die Haftorgane in Gestalt
mannigfaltiger Haken und Saugnäpfe tragender Teil wird als Kopf oder
Scolex bezeichnet; auf ihn folgen nach hinten in weiten Grenzen
schwankender Zahl die Glieder, Ringe oder Proglottiden. Jede Proglottis
umschliesst männliche und weibliche Geschlechtsorgane. Oft sind die
Proglottiden, nachdem sie eine gewisse Entwickelung durchgemacht
haben, nur noch lose mit einander verbunden; sie lösen sich sogar
von der allgemeinen Kette ab, um selbständig als Einzelindividuen
weiter zu leben. Das hat zur Auffassung des Bandwurmkörpers als
einer aus einem Hafttier (Scolex) und zahlreichen Geschlechtstieren
(Proglottiden) zusammengesetzten Kolonie geführt. Doch giebt es auch
Bandwürmer ohne jeden kolonialen Charakter. Die Glieder sind fest und
dauernd verbunden, die äussere Segmentierung verwischt; die beiden
Geschlechtsapparate kommen in gewissen Fällen im Körper überhaupt
nur in der Einzahl vor. Zwischen Einzelindividualität und Stocknatur
des Cestodenleibs lässt sich keine Grenze ziehen. Die den Trematoden
am nächsten stehenden Formen zeigen von Segmentierung noch keine
Spur; die Gliederung tritt zuerst schüchtern, dann immer deutlicher
hervor und führt zuletzt sogar zur Selbständigkeit der einzelnen
Segmente. Für alle verschiedenen Stufen liefern gerade die bei den
Fischen schmarotzenden Formen treffliche Beispiele. Auch in der
vielfach verschlungenen Lebensgeschichte der Cestoden spiegelt sich
ihr parasitärer Charakter wieder. Wanderungen und Wirtswechsel sind
allgemein verbreitet; freie Lebensstadien spielen eine viel geringere
Rolle als bei den Trematoden. Sie sind in den Entwickelungsgang der
verschiedenen Bandwürmer in verschieden reichem Masse eingestreut.
Der Körper der Süsswasserfische bietet relativ zahlreichen und
mannigfaltigen Cestoden Herberge, sei es als Zwischenwirt, sei es als
Träger der definitiven Geschlechtsform. Die eigentlichen Taenien,
Cestoden, die für die höheren Wirbeltiere und speziell auch den
Menschen eine ganze Anzahl von Arten stellen, sind im[S. 244] Süsswasserfisch
nicht gerade häufig. Barsch, Kaulbarsch, im Genfersee auch Quappe,
Felchen, Hecht, Forelle, Saibling beherbergen in ihrem Darm oft in
grosser Zahl die Taenia ocellata Rud., mit vier seitlich und
einem endständig am Scolex liegenden Saugnäpfen. T. longicollis
Rud. ist für die Salmoniden charakteristisch. In den Pylor-Anhängen
und dem Dünndarm der befallenen Fische leben oft hunderte dieses
Schmarotzers. Länge bis 25 cm, fünf Saugnäpfe. In Stichling und
Barsch ist die mit vier kleinen, aber starken Saugnäpfen bewehrte T.
filicollis Rud. nicht selten, während die Cypriniden sowie Felchen
und Quappe nicht allzu häufig durch die T. torulosa Batsch
infiziert werden. Bau und Entwickelung all dieser Fischtaenien sind uns
fast völlig unbekannt[LXXVIII].
[LXXVIII]O. v. Linstow hat uns jüngst über den Bau
derselben einiges mitgeteilt; eine weitere Arbeit über Cestoden der
Süsswasserfische wird von einem meiner Schüler vorbereitet.
Durch sein massenhaftes Auftreten in allen Salmoniden fällt der
Bothriocephalus infundibuliformis Rud. auf. Er fehlt auch nicht
in Barsch und Hecht. Sein Scolex trägt — wie dies für die Gattung
Bothriocephalus charakteristisch ist — zwei nur mässig starke
Sauggruben. Hunderte von Exemplaren des bis 40 cm langen
Wurmes sind kräftig in den Pylor-Anhängen des Wirtes befestigt.
Salm, Saibling und Forelle scheinen am meisten unter der Überzahl
der Schmarotzer leiden zu müssen. In manchen Meerfischen und in der
Quappe wird der B. infundibuliformis ersetzt durch die verwandte
Form B. rugosus Rud. Seine Länge geht bis 38 cm; er
ist ausgezeichnet durch randständige Geschlechtsöffnungen, während
dieselben sonst bei den meisten Bothriocephaliden auf der Fläche der
Glieder liegen.
An die Saugwürmer schliesst sich morphologisch und anatomisch eng
ein nicht seltener Darmschmarotzer der verschiedenen Karpfenarten,
der Nelkenwurm (Caryophyllaeus mutabilis Rud.) an. Der
Wurmkörper ist vollkommen ungegliedert, sein oft wie eine Nelkenblüte
gefalteter, hakenloser Vorderrand dient als einziges Fixationswerkzeug.
Männlicher und weiblicher Apparat, jüngst von Saint-Remy
genauer beschrieben, sehen denen der Trematoden ähnlich[S. 245] und bleiben
wie dort in der Einzahl. Kompliziert ist das Exkretionssystem mit
seinen zahlreichen geschlängelten Längskanälen und wohl ausgebildeten
Wimpertrichtern. Im Gegensatz zu den meisten Cestoden scheint
die Entwickelung des Nelkenwurms relativ einfach zu sein. Eine
Metamorphose ersetzt den Generationswechsel; dagegen ist die Wanderung
durch den Zwischenwirt beibehalten. Wahrscheinlich lebt der junge
Caryophyllaeus in dem unter dem Namen Tubifex rivulorum
bekannt gewordenen Ringelwurm und wird mit ihm in den Darm der
Cypriniden übertragen, wo er die Geschlechtsreife erreicht.
Schon etwas mehr dem Typus der Cestoden nähert sich der „Becherkopf“
(Cyathocephalus truncatus Pallas), der sich mit seinem
eigentümlichen, becher- oder trichterförmigen Scolex sehr fest in
den Pylor-Anhängen von Hecht, Barsch, Felchen, Forelle, Quappe,
Saibling ansaugt. Der bis gegen 40 mm lange Leib besteht aus
wenigen, fest verbundenen und undeutlich gegeneinander abgesetzten
Proglottiden, die flächenständig männlichen und weiblichen Porus,
sowie die Uterusöffnung tragen. Durch den Bau der Geschlechtsorgane
scheint sich der Cyathocephalus der Gruppe der Bothriocephaliden
anzuschliessen. Leider ist seine Anatomie erst unvollkommen, seine
Lebensgeschichte gar nicht bekannt.
Traten uns in den bis jetzt geschilderten Fällen die Fische
ausschliesslich als Wirte des ausgewachsenen, geschlechtsreifen
Bandwurms entgegen, so sind doch auch genügend Beispiele bekannt,
wo der Fisch die Rolle des Zwischenträgers spielt und in seinen
Organen die Larven des Cestoden birgt. In einem gut beschriebenen
Entwickelungsgang wird Haupt- und Zwischenwirt der Gruppe der
Süsswasserfische entnommen. Es betrifft dies den Triaenophorus
nodulosus Rud., der als Bandwurm von bedeutender Länge (bis
½ m) den Darm des Hechtes oft in recht beträchtlicher
Zahl bewohnt. Seltener bezieht er im geschlechtsreifen Zustand
Aesche, Forelle und Barsch. In den Fischen des Genfersees gehört
der Triaenophorus zu den gewöhnlichsten Erscheinungen. Sein
undeutlich abgesetzter Kopf trägt neben zwei schwachen Sauggruben zwei
Paar starker Chitinhaken, die durch ihre dreizackige Gestalt an[S. 246] die
starke Bewaffnung mancher Schmarotzer der Meerfische (Calliobothrien)
erinnern. Äusserlich ist der Bandwurmkörper trematodenhaft kaum
merklich gegliedert; innerlich dagegen ist die Segmentierung scharf
ausgedrückt durch die sich regelmässig in grosser Zahl folgenden,
nach dem Typus der Bothriocephalen gebauten Geschlechtsorgane. Im
Jugendzustand soll der Triaenophorus eingekapselt in der
Leber der Beutefische des Hechtes, hauptsächlich der Cypriniden,
liegen. Doch fand ich ihn wenigstens für den Genfersee am häufigsten
im Barsch, seltener im Hecht, am seltensten und nur in schwachen,
kleinen Exemplaren in Forelle und Aesche. Bemerkenswert bleibt
immerhin die Thatsache, dass auch hier wieder alle angeführten
Fischarten Haupt- und Zwischenträger der nämlichen schmarotzenden
Spezies werden können. Einmal traf ich den jungen Parasiten auch
im Lachs. Die Triaenophorus-Larven liegen meistens eingebettet in
erbsengrosse dickwandige Bindegewebecysten der Leber ihres Wirtes.
Seltener bewohnen sie Milz, Peritonäum und Muskeln. In der Leber ist
die Larvenzahl oft sehr bedeutend — nach eigener Erfahrung bis 36 —,
so dass sich die Gegenwart von hunderten von ausgewachsenen Würmern
im Hechtdarm wohl erklärt. Haken und Sauggruben sind im Jugendzustand
schon wohl ausgebildet, zudem folgt auf den Kopf ein sehr langer (bis
25 cm) Wurmkörper, der reich an Kalkkörperchen ist und sich
in der engen Cyste in den mannigfaltigsten Knäueln und Windungen
aufwickelt. Der bandförmige Anhang ist übrigens bestimmt, nach der
Übertragung auf den definitiven Wirt zum guten Teil verloren zu gehen.
Auf den Hauptträger übergeführt werden die Triaenophorus-Larven wohl
sehr bald geschlechtsreif. Ihre reifen Eier gelangen in ungezählten
Mengen mit dem Kot des Fisches ins Wasser. Ihnen entschlüpfen schon
nach sechs bis acht Tagen bewimperte, mit sechs provisorischen Haken
bewehrte Embryonen, die auf der Suche nach einem Zwischenwirt munter
umherschwimmen. Ob aber die oben aufgezählten Fische schon vom
Embryo bezogen werden, oder ob ihnen der Parasit in vorgerückterer
Entwickelung durch einen ersten, wirbellosen Zwischenwirt übertragen
wird, ist noch fraglich.
[S. 247]
Die Leibeshöhle der karpfenartigen Fische beherbergt oft
langgestreckte, bandförmige, schwach segmentierte Schmarotzer, ohne
deutliche Haftapparate. Ihre Zahl in einem Wirt ist oft ziemlich
beträchtlich; ihre Länge kann sehr bedeutend werden — kenne ich doch
aus einem Rötel ein Exemplar von 83 cm Länge und 2 cm
Breite. Kein Wunder, dass unter dem Drucke des wachsenden Wurmes die
von ihm mannigfaltig umschlungenen Organe des Fisches verkümmern.
Meist tritt tödliche Peritonitis ein. Schwere Fischepidemien, die
viele Karpfenteiche entvölkerten, sind einzig auf die Gegenwart
dieser Riemenwürmer oder Liguliden zurückzuführen. Es ist also
schon praktisches Interesse, wenn wir die Lebensgeschichte der
Ligula simplicissima Rud., die uns im ganzen und grossen von
Donnadieu geschildert worden ist, verfolgen. Wir wissen nun,
dass die Schmarotzer aus der Leibeshöhle der Karpfen nichts anderes
sind denn aussergewöhnlich grosse Larven von Bandwürmern. Sie wachsen
schon im Zwischenwirt, eben den Fischen, zu ungewöhnlicher Länge aus
und bilden auch schon die Geschlechtsorgane vor. Doch treten diese erst
in Funktion, wenn der kaltblütige Zwischenträger mit einem warmblütigen
Hauptwirt vertauscht wird. Dazu bietet sich am besten Gelegenheit,
nachdem die Ligula, durch die unter dem Drucke immer dünner
werdenden Bauchdecken des Fisches hindurchbrechend, ins Wasser gelangt,
in dem sie längere Zeit frei leben kann, ein für einen Parasiten
nicht gewöhnliches Verhältnis. Forel hat freie Exemplare von
Ligula in den grossen Tiefen des Genfersees gefischt. Jetzt
muss der noch nicht geschlechtsreife Wurm von einer Ente oder einem
anderen Wasservogel aufgenommen werden. Liguliden von mindestens 10
cm Länge, ja sogar Bruchstücke grösserer Exemplare werden im
Entendarm schon nach Verlauf von 24 Stunden vollkommen reif; kleinere
Tiere werden ausgeworfen. Die beiden Sauggruben treten deutlicher
hervor; die Segmentierung des Körpers prägt sich etwas schärfer
aus; der Leib streckt sich; in den in grosser Zahl sich regelmässig
folgenden Geschlechtsapparaten beginnt die Eibildung und Befruchtung.
Durch spezielle in jeder Proglottis sich wiederholende Uterusöffnungen
werden die von einer harten,[S. 248] gedeckelten Schale umschlossenen Eier
in den Darm des Wirtes und von dort, gemäss der echt parasitischen
Fruchtbarkeit der Ligula, in ungeheueren Mengen an die
Aussenwelt abgegeben. Geschlechtsreife Liguliden verweilen übrigens
nur kurze Zeit im Darme ihres Trägers, schon nach zwei bis drei Tagen
sollen sie nach Donnadieu ausgestossen oder verdaut werden. Im
Wasser dagegen können die ausgestossenen Würmer bis zehn Tage lang
weiterleben.
Fig. 42.
Ei von Bothriocephalus latus.
Die Eier in Wasser gebracht entlassen nach acht bis vierzehn Tagen
einen flimmernden, sechshakigen, lebhaft schwärmenden Embryo —
eine Jugendform, die ja überhaupt bei Plattwürmern verbreitet ist,
welche wasserbewohnende Zwischenwirte aufzusuchen haben. Nach
Donnadieus Erfahrungen muss der Embryo von den Karpfen
in den Darmkanal aufgenommen werden. Er wird die Wandungen des
Verdauungstractus durchbrechen und, vielleicht nach vorübergehender
Einkapselung in der Leber, die Leibeshöhle des Fisches beziehen.
Wenige Wochen nach der Infektion mit flimmernden Embryonen beherbergt
der Körperraum der Karpfen schon typisch ausgebildete Liguliden, von
6–12 mm Länge. Die Entwickelungsgeschichte der Riemenwürmer
ist ausgezeichnet durch zahlreich eingestreute freie Stadien und die
weitgehende Differenzierung der Larve schon im Zwischenwirt. Es bedarf
das junge Tier nur noch eines kurzen Aufenthaltes im warmen Darm des
Hauptwirtes, um reife Eier zu liefern. Eine Infektion der Karpfen wird
natürlich am besten durch Ausschluss der Hauptwirte — der Wasservögel
— von den Teichen verhindert.
In Bau und Lebensgeschichte schliesst an die Ligula
unmittelbar der Schistocephalus dimorphus Crepl. an. Er lebt
als bandförmige, bereits gegliederte Larve in der Leibeshöhle des
Stichlings, als reifer Bandwurm im Darm der Wasservögel. Es sei endlich
noch erwähnt, dass in der Gallenblase und zwischen den Darmzotten
der Schleihen zwei verschiedene Larven von Bandwürmern aufgefunden
worden sind, die sich durch manche Eigentümlichkeiten auszeichnenden
Gyporhynchen. Nach ihrem Übertritt in den Darm der Reiher entwickeln
sie sich zur Taenia macropeos Wedl und T. unilateralis
Rud.
[S. 249]
Fig. 43.
Flimmerembryo von Bothriocephalus latus.Fig. 44.
Larve von Bothriocephalus latus aus verschiedenen
Süsswasserfischen.
Aus den Süsswasserfischen bezieht endlich auch der Mensch einen
häufigen Bewohner seines Darmkanals, den Grubenkopf (Bothriocephalus
latus Brems.). Dieser längste aller in unserem Körper vorkommenden
Bandwürmer — kann er doch bis 10 m erreichen — kommt
besonders in weiter Verbreitung in Länderstrichen vor, die reich
an Süsswasserseen sind. Es lässt sich das nach der Natur seiner
Zwischenwirte ja zum voraus erwarten. So treffen wir ihn häufig in
der Westschweiz, im Seengebiet Oberitaliens, in den weiten an das
baltische Meer grenzenden, zu Deutschland und Russland gehörenden
Seebezirken, in Polen, in gewissen Teilen Russlands, in Schweden, in
Japan. Auch am Ufer des Starnbergersees hat sich ein Infektionsherd
gebildet. Bis vor wenigen Jahren war die Herkunft des so häufigen
und nicht unbedenklichen Schmarotzers unbekannt; das Verdienst, den
Schleier über diesem Dunkel gelüftet zu haben, gehört in erster Linie
Prof. M. Braun. Doch bleiben auch heute noch weite Lücken
in der Kenntnis der Lebensgeschichte des Bothriocephalus
auszufüllen. Aus der Uterusöffnung der einzelnen Bothriocephalenglieder
treten die gedeckelten, ovalen Eier, die auch bei diesem Wurm, dank
der parasitischen Lebensweise, und zu Gunsten der Verbreitung der
schmarotzenden Spezies, in Übermenge gebildet werden (Fig. 42). Im
Wasser zerstreut entwickelt sich in ihnen ein Embryo nach Vorgängen,
die jüngst von Schauinsland verfolgt worden sind. Er verlässt
die Eischale je nach den äusseren Temperaturbedingungen und der Masse
des das Ei bespülenden Wassers früher oder später. Im Sommer findet
das Ausschlüpfen schon zwei bis vier Wochen nach der Eiablage statt;
unter ungünstigen Umständen (im Winter) können aber acht und mehr
Monate vergehen, bis der junge Embryo frei wird. Er ist ähnlich wie
das erste Jugendstadium von Ligula und Triaenophorus,
Würmer, die übrigens auch im anatomischen Bau des erwachsenen Tieres
dem Bothriocephalus nahekommen, mit sechs Haken bewaffnet
und schwimmt im Wasser mit Hilfe eines langen Wimperpelzes (Fig.
43 S. 250). Das freie[S. 250] Leben der Embryonen kann bis eine Woche
dauern, dann sinken die Tierchen zu Boden und streifen gewöhnlich
die Flimmerbekleidung ab, um sich noch eine Zeit lang kriechend zu
bewegen. Was nun mit den jungen Bothriocephalen geschieht, ist uns
völlig unbekannt. Hier öffnet sich eine weite Lücke in unserem Wissen.
Es hat in keiner Weise gelingen wollen, die sechshakigen Embryonen
direkt auf Fische zu übertragen; ebensowenig ist es geglückt, niedere
wirbellose Wasserbewohner mit den Flimmerembryonen zu infizieren. So
steht die Frage noch vollkommen offen, ob die Süsswasserfische den
ersten und einzigen Zwischenwirt für den Grubenkopf abgeben, oder
ob die Embryonen, etwa wie die ihnen nach Bau und Lebensweise so
ähnlichen frühesten freien Jugendstadien der Trematoden, zunächst in
einen wirbellosen Zwischenwirt eindringen müssen, um dort weitere
Veränderungen durchzumachen.
Wie dem auch sei, wir finden die jungen Bothriocephalen erst wieder
unter ganz veränderter Gestalt im Leibe mancher Süsswasserfische.
Von Braun ist 1883 zunächst der Hecht als Zwischenträger
des Grubenkopfes erkannt worden, seitdem haben Parona und
Grassi in Italien, Ijima in Japan, ich in Genf und
Basel die Bothriocephalus-Finnen in einer ganzen Reihe von Fischen
der Süsswasserseen entdeckt und meist mit gutem Infektionserfolg auf
den Menschen übertragen. Die Zwischenwirte des breiten Bandwurms
sind fast ausschliesslich Räuber, eine Thatsache, die vielleicht
darauf hindeutet, dass ihnen der junge Wurm bereits in einen ersten
Träger eingeschlossen übermittelt wird. Als Wirte der Finnen von
B. latus kennen wir heute den Hecht, die Quappe, den Barsch,
die Forelle, die Aesche, den Saibling, die Seeforelle, sowie die
beiden japanischen Lachsarten Onchorhynchus Huberi und
O. Perryi, eine Liste, die sich mit der Zeit noch bedeutend
vergrössern dürfte. Im Lachs sind Finnen von B. latus, entgegen
der[S. 251] Annahme von Küchenmeister, nicht gefunden worden. Auch die
Flussfische sind häufig mit dem Jugendstadium unseres Schmarotzers
besetzt; wenigstens fand ich die Finnen in Forellen und Hechten aus dem
schnellfliessenden Rhein und der reissenden Aare.
Die Finnen sind gestreckt 8 bis 25 mm lang, 1 bis 3 mm
breit, von schwach abgeflachtem Körper. Vorn trägt das Tier einen
konischen Kopfaufsatz, der in zwei seichten Sauggruben schon die
künftigen Haftapparate des erwachsenen Bothriocephalus aufweist.
Oft ist übrigens das Vorderende in den Körper eingestülpt. Der Leib
ist solid, ungegliedert, höchstens die Kutikula zeigt eine starke
Querrunzelung. Gegen hinten wird der Leib in völlig gestrecktem
Zustand etwas schmäler. Auffallend ist der grosse Reichtum von im
Körper zerstreuten Kalkkügelchen (Fig. 44). Die Finnen liegen,
von dünnwandigen Kapseln umschlossen, oft in bedeutender Zahl in
den verschiedensten Organen des Fisches. Speziell häufig treten
sie auf in den Wandungen des Schlundes, in der Leber, in Milz,
Nieren, Geschlechtsorganen und endlich, für die Übertragung wohl am
wichtigsten, in der Rückenmuskulatur. Oft trifft man sie auch frei
wandernd in der Leibeshöhle. Die Zahl der Finnen in einem Fisch
kann recht bedeutend werden (50–100); auf das massenhafte Vorkommen
derselben in einer Seeforelle ist schon hingewiesen worden. Im
Wasser leben die Würmchen längere Zeit weiter. Solche Jugendstadien
in den menschlichen Verdauungstractus gebracht entwickeln sich zum
typischen breiten Bandwurm, zum Bothriocephalus latus. Es ist
dies besonders durch Experimente an Dorpater und Genfer Studenten
bewiesen worden. Das Wachstum des Kettenwurms im menschlichen Darm
ist ein ungemein rasches; es beträgt unter günstigen Umständen 6 bis
8 cm im Tag, so dass der Schmarotzer bald nach Metern gemessen
und seine eng verbundenen Proglottiden[S. 252] nach tausenden gezählt werden
können. Nach drei Wochen ungefähr, von der Infektion an gerechnet,
werden wieder reife Eier abgegeben.
Wir erwerben den breiten Bandwurm durch den Genuss ungenügend
zubereiteter Fische. In den Ostseeprovinzen spielt der Hecht vorzüglich
die traurige Rolle des Zwischenträgers; in Genf ist der Hauptsünder
die Quappe. Sie gelangt massenhaft auf den Markt, ihre Leber wird ganz
leicht gebacken als Leckerbissen verzehrt, gerade dieses Organ aber
beherbergt fast regelmässig Bothriocephalus-Finnen. Auch der Barsch
muss in dieser Hinsicht stark angeschuldigt werden.
Ausser den Larven von B. latus wohnen in vielen
Süsswasserfischen noch die Finnen anderer Grubenkopfarten. Im Lachs
speziell fand ich gelegentlich fünf verschiedene Formen solcher
jugendlicher Würmer, die wohl mehreren Arten angehörten, von denen
aber weder morphologisch noch experimentell eine auf den breiten
Bandwurm des Menschen bezogen werden konnte. Die Art und Weise ihres
Vorkommens im Fisch war analog den für die Jugendstadien von B.
latus beschriebenen Verhältnissen. Die Bothriocephaliden scheinen
vorzugsweise Fische als Zwischenträger zu benutzen.
So gewinnt die Annahme Leuckarts, dass ein anderer Grubenkopf,
der Bothriocephalus cordatus Lt., der in Grönland häufig den
Darm von Hund, Seehund, Walross, zufällig den des Menschen bewohnt,
durch Fische in seine Wirte eingeschmuggelt werde, sehr an Gewicht.
[S. 253]
Litteratur.
1) G. Brandes, Die Familie der Holostomiden. Zoolog.
Jahrbücher. Abt. f. Systematik, Geographie u. Biologie der Tiere. Bd.
5.
2) M. Braun, Zur Entwickelungsgeschichte des breiten
Bandwurmes. Würzburg 1883.
3) O. Bütschli, Zur Entwickelungsgeschichte des Cucullanus
elegans. Zeitschr. f. wiss. Zoologie. Bd. 26. 1876.
4) G. Bunge, Weitere Untersuchungen über die Atmung der Würmer.
Zeitschr. f. physiol. Chemie. Bd. 14. 1889.
5) C. M. Diesing, Systema Helminthum.
6) Donnadieu, Contributions à l’histoire de la Ligule. Journal
de l’anatomie et de la physiologie. 1877.
7) F. Dujardin, Histoire naturelle des Helminthes.
8) O. Hamann, Die Nemathelminthen. Erstes Heft: Die
Acanthocephalen. 1891.
9) F. Küchenmeister, Die Finne des Bothriocephalus latus und
ihre Übertragung auf den Menschen.
10) R. Leuckart, Die Parasiten des Menschen und die von ihnen
herrührenden Krankheiten. 1. und 2. Auflage.
11) O. v. Linstow, Compendium der Helminthologie. Hannover
1878.
12) O. v. Linstow, Nachtrag dazu. Die Litteratur der Jahre
1878–1889.
13) E. Lönnberg, Helminthologische Beobachtungen von der
Westküste Norwegens. Erster Teil: Cestoden. Svenska Vet. Akad.
Handlingar. Bd. 16. Afd IV.
[S. 254]
14) E. Lönnberg, Bidrag till Kännedomen on i Sverige
förekommande cestoder. Ibid. Bd. 14, IV.
15) F. Miescher-Rüsch, Statistische und biologische Beiträge
zur Kenntnis vom Leben des Rheinlachses im Süsswasser. Ichthyologische
Mitteilungen aus der Schweiz zur internationalen Fischereiausstellung
zu Berlin 1880.
16) A. Moquin-Tandon, Monographie de la famille des Hirudinées
avec atlas. Paris 1846.
17) E. Parona, Intorno la genesi del Bothriocephalus latus
(Brems.) e la sua frequenza in Lombardia. Torino 1887.
18) M. Stossich, I Distomi dei pesci marini e d’acqua dolce.
Trieste 1886.
19) M. Stossich, Appendice al mio lavoro i Distomi dei pesci
marini e d’acqua dolce. Trieste 1888.
20) G. Saint-Remy, Recherches sur la structure des organes
génitaux du Caryophyllaeus mutabilis. Revue biologique du Nord de la
France. T. 2. 1889–90.
21) A. Villot, Echinorhynchus clavaeceps. Note sur son
organisation et son développement. Bulletin de la société des sciences
naturelles du Sud-Est.
22) R. v. Willemoes-Suhm, Helminthologische Notizen. II.
Zeitschr. f. wiss. Zool. Bd. 20. 1870.
23) E. Zeller, Untersuchungen über die Entwickelung des
Diplozoon paradoxum. Ibid. Bd. 22. 1872.
24) E. Zeller, Über den Geschlechtsapparat des Diplozoon
paradoxum. Ibid. Bd. 46. 1888.
25) F. Zschokke, Recherches sur l’organisation et la
distribution zoologique des vers parasites des poissons d’eau douce.
Archives de Biologie. 1884.
26) F. Zschokke, Helminthologische Bemerkungen. Mitteilungen a.
d. zool. Station zu Neapel. Bd. VII.
27) F. Zschokke, Der Bothriocephalus latus in Genf.
Centralblatt f. Bakteriol. u. Parasitkunde. Bd. 1. 1887.
28) F. Zschokke, Ein weiterer Zwischenwirt des Bothriocephalus
latus. Ibid. Bd. 4. 1888.
29) F. Zschokke, Über Bothriocephalenlarven in Trutta salar.
Ibid. Bd. 7. 1890.
[S. 255]
Über
die quantitative Bestimmung des Plankton im Süsswasser.
Von Dr. C. Apstein in Kiel, Zoolog. Institut.
[S. 257]
In seinem Werke[103][LXXIX] „Über die Bestimmung des Planktons oder
des im Meere treibenden Materials an Pflanzen und Tieren“ bezeichnet
Hensen mit Plankton „Alles, was im Wasser treibt“.
Es sind also darunter alle Organismen, sowohl Pflanzen wie Tiere,
zu verstehen, die willenlos der Wellenbewegung und den
Strömungen folgen. Der Begriff ist enger gefasst als „Auftrieb oder
pelagische Organismen“, denn zu letzteren gehören beispielsweise
die freischwimmenden Fische, die aber nicht zum Plankton zu zählen
sind, da sie vermöge ihrer Grösse und Stärke nicht machtlos im
Wasser treiben, dagegen bilden die schwimmenden Fischeier und junge
Fische einen Teil des Plankton. Ebenso könnte es scheinen, als ob
die Copepoden ausscheiden müssten, die bekanntlich einer ziemlich
energischen Bewegung fähig sind; doch ist dieselbe zu wenig ausgiebig,
um den bewegenden Agentien irgend einen nennenswerten Widerstand
entgegensetzen zu können.
Schon im Jahre 1884 hat Hensen in seiner Arbeit[102]: „Über das
Vorkommen und die Menge der Eier einiger Ostseefische, insbesondere
der Scholle, des Flunder und des Dorsch“ ein Problem in Angriff
genommen, an dessen Lösung niemand vor ihm gedacht hatte, nämlich die
quantitative Bestimmung der im Meere treibenden Organismen.
[S. 258]
Nur einmal ist, unabhängig von Hensen[LXXX], jedoch nach diesem,
dieselbe Art der Forschung versucht worden von Asper und
Henscher[95] 1886, welche die Organismen aus einer bekannten
Wassermenge sammelten und dann die Zahl der Individuen bestimmten, die
sich in einem Tropfen Flüssigkeit, von denen fünfzehn auf 1 ccm
gingen, befanden. Dadurch erhielten sie einen ungefähren Überblick über
die Menge der pelagischen Tiere in dem untersuchten Süsswasserbecken.
Doch ist dieser Versuch nicht in eine Reihe zu stellen mit den
sorgfältigen Methoden der Hensenschen Forschungen. Hensen blieb nicht
bei den einfachen quantitativen Bestimmungen stehen, sondern zeigte
auch, warum diese von so hoher Wichtigkeit sind. In letzter Linie kam
es ihm darauf an, den Stoffwechsel des Meeres näher kennen zu lernen.
Bekanntlich vermögen nur die chlorophyllführenden Wesen, also Pflanzen
und Peridineen, von denen letztere noch von manchen zu den Flagellaten
gerechnet werden, aus anorganischen Stoffen organische Verbindungen
herzustellen; die Kraft, durch die diese Umbildung geschieht, liefert
das Sonnenlicht. Dieses dringt bis höchstens 400 m in das Wasser
ein und soweit finden wir auch nur chlorophyllhaltige Organismen.
Dieser Fähigkeit wegen können wir auch die pflanzlichen Wesen mit
Einschluss der Peridineen Nahrungsproduzenten[96] nennen.
Ihnen stehen die Nahrungskonsumenten gegenüber, welche die oben
erwähnte Fähigkeit nicht besitzen, also ganz von den Pflanzen abhängig
sind; es sind, um es kurz zu sagen, die Tiere. Wo keine Pflanzen
sind, können also auch keine Tiere sein! Da, wie oben erwähnt,
die Sonne die Kraft den Pflanzen giebt, organische Verbindungen zu
bereiten, so hätten wir ein Mass für die Produktion des Wassers an
belebter Substanz, wenn wir alle unter einer bekannten Oberfläche —
als Einheit 1 qm — vorhandenen Pflanzen bestimmen[S. 259] könnten.
In dieser Richtung hat Hensen[LXXXI] zur ersten Orientierung eine
Anzahl analytischer Gewichtsbestimmungen gemacht, denen wir folgende
Daten entnehmen.
[LXXX]Hensen hat schon im Jahre 1885 mehrere
Mitteilungen über sein Verfahren veröffentlicht, so in den
Sitzungsberichten des physiologischen Vereins in Kiel, in den
Mitteilungen des Vereins schleswig-holsteinischer Ärzte, im 8.
Jahresbericht des Centralfischereivereins für Schleswig-Holstein, dann
1886 im Tagesberichte der Naturforscher-Versammlung in Berlin.
Ein ganzer Fang (aus der Ostsee) vom Februar 1885, der reich an einer
Diatomeenart (Rhizosolenia) war, enthielt unter 1 qm
Oberfläche 1608.3 ccm Plankton. Davon wurden 70 ccm
weiter verarbeitet. Diese enthielten 0.18 gr organische
Substanz = 42.1% und 0.2575 gr = 57.9% Asche. Der ganze Fang
würde also 4.296 gr organische Substanz geliefert haben. Dies
ist sehr wenig, aber da die Diatomeen so zahlreich waren, so ist der
beträchtliche Aschenanteil durch die Kieselsäureskelette verständlich.
Zu ferneren Analysen wurden einzelne Bestandteile des Plankton
verwendet, so die Ceratien und Copepoden. Von ersteren fanden sich in
7.25 ccm Fang 12.45 Millionen. Diese enthielten 0.389 gr
organische Substanz = 96.05% und nur 0.016 gr Asche = 3.94%.
Auf 1 Million Ceratien käme also 0.031245 gr organische Substanz.
Von Copepoden wurde einmal Rhinocalanus gigas zur
Bestimmung benutzt. 4 ccm davon = 321 Stück ergaben 0.0527
gr = 99.4% organische Substanz und nur 0.0003 gr = 0.6%
Asche.
Diese Zahlen benutzt Hensen weiter, wobei hervorzuheben ist,
dass alle Angaben nur Minimalzahlen sind. Hensen hat nun
nach Versuchen[LXXXII] — denen er selbst nur einen vorläufigen Wert
beimisst — berechnet, dass ein Copepod zu seiner Ernährung täglich 12
Ceratien, das ist pro Jahr 4370 Ceratien bedarf. Da nun auf 1 qm
Oberfläche (also einer Wassersäule, die 1 qm zur Grundfläche und
die Tiefe des Wassers[LXXXIII] zur Höhe hat) ungefähr 1 Million Copepoden
lebt, so bedürfen diese zur Nahrung 4370 Millionen Ceratien, welche
4370 × 0.031245 = 133.35 gr organische Substanz liefern.
Die Jahresproduktion an Diatomeen[LXXXIV] berechnet Hensen auf 6570
ccm pro Quadratmeter Oberfläche, diese enthalten 14.8 bis 17.7
gr organische Substanz. Es würden also in Summa[S. 260] pro Jahr von
Diatomeen und Ceratien 133 + 17 gr = 150 gr organische
Substanz pro Quadratmeter Oberfläche erzeugt werden.
Nach Berechnungen von Rodewald erzeugt 1 qm bebauten
Landes (in Form von Heu) 179 gr organische Substanz. Es ist also
die Produktion des Plankton nur um 20% geringer als die der gleichen
Fläche Ackerlandes[LXXXV]. Da jedoch für die Berechnungen des Wassers nur
Minimalzahlen genommen sind, so wäre es möglich, dass in der That die
Produktion des Wassers gleich ist der des Landes. Hiermit ist also ein
Mass für die Ertragsfähigkeit des Wassers gewonnen und zugleich ein
Ausdruck für die belebende Wirkung des Sonnenlichtes.
[LXXXV] Dasselbe erwähnt
Seligo[107] in seinen
„Hydrobiologischen Untersuchungen“. Er sagt: „Wie eine Wiese ist die
Wasserfläche gleichmässig bewachsen. Allerdings liegen die Pflänzchen
normal nicht dicht an einander, aber dafür beschränkt sich ihre
Anwesenheit und ihr Gedeihen nicht auf die Wasseroberfläche, sondern
die oberen Wasserschichten bis zu mehreren Metern Tiefe sind davon
durchsetzt, sodass die Gesamtmenge der unter einem bestimmten Teil der
Oberfläche wachsenden Pflänzchen ungefähr so viel Pflanzenmenge sein
dürfte, wie auf einer gleichgrossen Fläche einer dünn bewachsenen Wiese
sich findet“.
Die oben gewonnenen Zahlen lassen sich jedoch noch weiterhin benutzen,
z. B. für praktische Zwecke der Fischerei, wie Heincke[101]
dargethan hat. Jedoch würde es uns zu weit führen, auf alle diese
interessanten Berechnungen weiter einzugehen, wir müssen auf die
Originalwerke verweisen.
Ein Punkt ist jedoch noch von Wichtigkeit, der es klarlegen soll, dass
die Planktonmethode zu den vorhin dargelegten Schlüssen berechtigt.
Haben wir ein Recht, von der Beobachtung, die wir aus einem kleinen
Wasserquantum gewonnen haben, auf die Zusammensetzung des Plankton
eines ganzen Wasserbeckens zu schliessen? Da hat sich nun gezeigt bei
Untersuchungen in der Ostsee, dass die Verteilung des Plankton eine
ziemlich[LXXXVI] gleichmässige ist. Diese gleichmässige Verteilung
ist von Hensen in seiner Arbeit[102]: „Über das Vorkommen und
die Menge der Eier einiger Ostseefische, insbesondere der Scholle,
des Flunder und des[S. 261] Dorsch“ so erklärt. Trifft ein Stoss, z. B. eine
Welle, ein schwimmendes Ei, so können zwei Fälle in Betracht kommen:
erstens, ist der Stoss senkrecht, so wird das Ei in der Richtung
des Stosses fortbewegt; zweitens, ist der Stoss unter einem Winkel
auf das Ei gelangt, so schiebt eine Komponente des Stosses das Ei
weiter, während die andere es dreht. So werden die Eier nach und nach
auseinandergetrieben, wenn sie auch an einer Stelle der Oberfläche sich
befanden, da ein Stoss nicht alle Eier in derselben Richtung trifft.
Was für die Eier gilt, ist auch für die anderen Organismen des Plankton
anzuwenden. So wird durch die Wellen, welche wie eine Schüttelbewegung
wirken, die Zerstreuung besorgt. Diese Thatsachen hat Hensen
auch noch direkt durch Experimente (siehe die gleiche Arbeit) erhärtet,
indem er mehrere versilberte Glaskugeln, die so beschwert waren, dass
sie gerade noch schwammen, in das Wasser versenkte und nach einiger
Zeit wahrnahm, dass sie weit auseinandergetrieben waren.
[LXXXVI] In einer neuen Arbeit Hensens[104]:
„Einige Ergebnisse der Plankton-Expedition der Humboldt-Stiftung“ heisst es:
„... Die Expedition ging von der rein theoretischen Ansicht aus, dass
in dem Ozean das Plankton gleichmässig genug verteilt sein müsse, um
aus wenigen Fängen über das Verhalten sehr grosser Meeresstrecken
sicher unterrichtet zu werden, und diese Voraussetzung hat sich weit
vollständiger bewahrheitet, als gehofft werden konnte“.
Nachdem wir gesehen haben, was für weitgehende Schlüsse Hensen
mit Hilfe seiner Planktonmethode, d. h. der Art und Weise der Gewinnung
und Verarbeitung eines Planktonfanges, zu ziehen imstande war, wollen wir diese
Methode[LXXXVII] näher ins Auge fassen.
[LXXXVII] Diese Methode hat in neuester Zeit von E. Häckel
eine sehr absprechende Beurteilung erfahren; da jedoch schon von
anderer Seite die Missverständnisse und Entstellungen der Häckelschen
Schrift richtiggestellt sind, so gehe ich nicht weiter auf dieselbe
ein, sondern verweise auf die Arbeiten Brandts[102] und Hensens.
Ich möchte daher den Leser bitten, mich auf einer Exkursion zu
begleiten und nachher den Arbeiten am Lande beizuwohnen. Wir besteigen
einen Dampfer und haben Zeit, ehe wir an Ort und Stelle anlangen, die
Ausrüstung zu der „Planktonfahrt“ in Augenschein zu nehmen.
Vor allem fällt uns das grosse Vertikalnetz[LXXXVIII]
auf (Fig. 45 S. 262): An ihm können wir drei Teile unterscheiden, erstens das
eigentliche konische Netz (A), dann den ebenfalls konischen
Aufsatz (B) und drittens den Blecheimer (Fig. 48 S. 265).
[LXXXVIII] Ich habe in folgendem die Masse des Netzes nach denen
von der Plankton-Expedition auf dem Atlantischen Ozean verwendeten
Apparaten wiedergegeben. Da, wo auf Süsswasserseen kein Dampfer zur
Verfügung steht, müssten die Netze verkleinert werden, denn vom Boote
aus ist es unmöglich, mit den fast 2 m hohen Netzen zu fischen.
[S. 262]
Das eigentliche Netz ist folgendermassen gebaut:
An einem starken Eisenringe (S), der 90 cm Durchmesser
hat, ist innen eine 5 cm grosse Falte von Barchent befestigt
(r), deren äusserer Teil eine grössere Anzahl Knopflöcher
trägt. Von hier soll nach dem untern Ringe (K) das weiterhin
zu beschreibende Netzzeug ausgespannt werden. Dieser untere Ring
(K) trägt drei gabelartige Vorsprünge, an welchen durch
Überfallschrauben der Blecheimer befestigt werden kann. Auf diesem
Ringe ist ein zweiter Ring (K1) durch mehrere mit Ösen
versehene Schrauben (m) befestigt.
Fig. 45.
Das Gazenetz (i) trägt an seinem obern Rande einen
Leinwandstreifen (l), welcher mit Knöpfen versehen ist, die in
die oben erwähnten Knopflöcher[S. 263] der Barchentfalte hineinpassen[LXXXIX],
unten ist das Netz an einen Barchentring angenäht, welcher an dem
Messingringe (K) durch mehrere Schrauben (n) befestigt
werden kann. Damit das feine Gazenetz nicht allein den Druck der
filtrierenden Wassermasse auszuhalten hat, ist an seiner Aussenseite
ein einfaches Fischernetz (f) ausgespannt, das oben an dem Ringe
S mit Bindfaden angebunden ist und unten zwischen die beiden
Ringe K und K1 geklemmt wird. Die beiden erwähnten
Netze sind aber zu schwach, den an den Ringen K und K1
hängenden Eimer zu tragen, es sind daher zwischen S und den
Schraubenösen (m) einige starke Schnüre (g) ausgespannt.
[LXXXIX] Es ist daher jederzeit möglich, das Netz herabzunehmen
und auszuwaschen.
Bei der Wahl des Netzzeuges handelt es sich einerseits darum,
dass die Löcher möglichst fein und gleichmässig sind, damit auch die
kleineren Organismen nicht hindurchgehen können, anderseits auch darum,
dass die Fäden des Gewebes nicht quellen und sich nicht verschieben
können. Diese Bedingungen werden allein durch die Müllergaze erfüllt,
die in mehreren Sorten in den Handel kommt, und sich durch die Grösse
der Löcher unterscheidet. Diese Gaze ist aus Seidenfäden verfertigt und
wird in Mühlen zur Trennung des Mehles nach der Korngrösse benutzt.
Dieses Gewebe (Müllergaze Nr. 20) ist von solcher Feinheit, dass auf
einen Quadratcentimeter Fläche 5926 Löcher kommen[XC], von denen
jedes eine Seitenlänge von 0.053 mm hat. Mit diesem Netzzeug
werden fast alle Organismen gefangen, nur wenige Diatomeen, die mit
ihrer Längsachse auf ein Loch treffen, werden hindurchschlüpfen. Einen
grossen Vorzug besitzt dieses Gewebe noch durch seine grosse Glätte, es
bleiben einmal wenig Organismen daran hängen, dann auch fasert es nicht
aus, so dass der Fang nicht durch Fäden verunreinigt wird.
Ein aus diesem Seidengewebe gefertigtes Netz soll nun zwischen den
beiden Ringen so ausgespannt werden, dass es keine Falten schlägt. Es
ist dazu nötig, ein Muster zu entwerfen[XCI], nach dem[S. 264] das Netzzeug
zugeschnitten wird. Bei dem vorliegenden Netze beträgt der Radius des
obern Ringes R = 45 cm, der des untern r = 10
cm, die Mantelhöhe des abgestumpften Kegels i = 150
cm.
[XCI] Die Berechnung weicht von der von Hensen im Planktonwerk
S. 6 angeführten ab, da statt der Sehnen der Winkel an der Spitze
benutzt ist.
Vervollständige ich den abgestumpften Kegel (Fig. 46), so kann ich mit
Hilfe der Mantelhöhe des abgeschnittenen Stückes (x) den Winkel
an der Spitze (α) (Fig. 47) berechnen.
Fig. 46.
Es ist x : x + i = r : R. Daraus
folgt
x = r iR − r.
Nach unserem Beispiel erhalten wir für x = 42.9 cm.
Denken wir uns jetzt den Kegelmantel aufgerollt (Fig. 47), so ist
A B = U = 2 R π
und
C D = u = 2 r π.
Fig. 47.
Es muss sich verhalten der Umfang des Kreises, den ich mit dem Radius
x (= 2 x π) schlagen kann, zu u, wie 360° : α.
Also
2 x π2 r π
= 360°α
Daraus folgt
xr
= 360°α
In unserem Beispiel erhalten wir für α = 83.9°.
Ebenso wäre die Rechnung, wenn wir für u : U genommen
hätten, dann hätten wir statt x aber x + i setzen
müssen. Um nun nach vorstehenden Zahlen ein Muster zu zeichnen,
verfahren wir so, dass wir uns den Winkel α = 83.9° konstruieren
und von dem Scheitelpunkte desselben mit den Radien x = 42.9
cm und (x + i) = 192.9 cm Kreisbogen
schlagen, dann geben die Strecken dieser Bogen zwischen den Schenkeln
des Winkels den obern und untern Umfang des Netzzeuges an. Nach diesem
Muster wird dann das Gazenetz ausgeschnitten, wobei berücksichtigt
werden muss, dass bei AC und BD das Netz aneinandergenäht
wird, zu welchem Zweck eine Einschlagskante bleiben muss. Die[S. 265] Nähte
müssen natürlich nach aussen kommen und ausserdem darf nur eine ganz
feine Nadel verwendet werden, da jeder Nadelstich dem feinen Netzzeug
gegenüber ein grosses Loch darstellt.
Der Aufsatz des Netzes besteht aus einem 1 cm dicken
Eisenringe (a) von 36 cm Durchmesser, der mit dem obern
Netzringe (S) durch drei starke 60 cm lange Eisenstangen
(d) verbunden ist, die oben in Haken (c) zur Befestigung
des Taues auslaufen. Zwischen den beiden Ringen, jedoch unter
den verbindenden Eisenstangen, ist ein Barchentmantel[XCII] ausgespannt.
Dieser Aufsatz ist von grosser Wichtigkeit für die Brauchbarkeit des
Netzes. Wird das Netz auf den Grund des Wasserbeckens hinabgelassen,
so würden, wenn dieser Netzteil fehlen sollte, Schlamm und Organismen
von dem Boden in das Netz geraten können. Bei dieser Einrichtung
jedoch stösst höchstens der Eisenring (S) auf den Schlamm auf,
und dieser wird, wenn auch etwas aufgewirbelt, doch nicht die obere
Netzöffnung erreichen und den Fang verunreinigen können. Ferner dient
dieser Aufsatz auch als Reservoir, wenn bei stürmischem Wetter der
Fang aus dem Netz hinaufgespült wird. Dann auch wird durch die Öffnung
des Aufsatzes (= 1000 qcm) ein nur geringer Wasserstrom in das
Netz hineingelangen und durch die gegen 26mal so grosse Netzwand fast
vollständig filtriert werden können.
[XCII] Derselbe wird ebenfalls nach den obigen Formeln
konstruiert.
Fig. 48.
Der Eimer (Fig. 48) ist ein cylindrisches Blechgefäss
(Eisenblech), dessen Boden nach der Mitte zu abfällt und hier eine
durch eine Schraube verschliessbare Öffnung (u) trägt. Der
obere Rand des Gefässes ist erhaben (n) und trägt drei
Überfallschrauben (m), die in die oben erwähnten drei
gabelartigen[S. 266] Fortsätze des untern Netzringes (Fig. 45k)
hineinpassen. In der Mitte ist der Blechcylinder noch von einem Reif
(q) umgeben. Zwischen diesen beiden Reifen (q und
n) ist die eine Seite der Eimerwand herausgenommen und durch
Müllergaze (o) verschlossen. Der ganze Eimer steht auf sechs
Füssen (f), die durch einen Ring (r) verbunden sind.
Neben dem Vertikalnetz kommt hauptsächlich der Filtrator (Fig.
49) in Betracht. Dieser stellt einen Metallcylinder dar, dessen
Wände durch Müllergaze ersetzt sind mit Ausnahme von einigen Stützen
(s), die unten durch einen sehr flachen Reif (K)
verbunden sind. Die Müllergaze wird zwischen dem obern und untern Ringe
(R und K) und den Stützen (s) folgendermassen
ausgespannt. In dem obern Ringe (R) befindet sich ein zweiter
Ring (R′), der an den erstern angeschraubt werden kann;
zwischen beide wird die Gaze eingeklemmt, ebenso geschieht dies bei
dem untern Ringe (K), auf den ein zweiter Ring (K′)
passt, und schliesslich auch bei den Stützen (s), an welche
von innen die Gaze durch Metallplatten angedrückt wird. Indem man den
obern und untern Rand der[S. 267] Gaze zwischen den beiden Ringen einspannt,
die seitlichen Ränder aber zwischen einer Stütze und ihrer innern
Platte, hat man den Vorteil, dass an dem Netzzeuge des Filtrators kein
Nadelstich nötig ist.
Der Filtrator trägt an jeder Seite einen dreikantigen Vorsprung
(v), der dazu dient, den oben beschriebenen Apparat mit Hilfe
eines Bügels (b) und einer Überfallschraube (a) auf
eine Glasplatte (G) fest anzudrücken, so dass unter dem Ringe
(K′) kein Wasser entweichen kann, sondern dasselbe alles durch
das Netzzeug filtrieren muss. Da die beiden unteren Ringe (K
und K′) sehr flach sind, so wird nur sehr wenig Wasser im
Apparat zurückbleiben, welches man auch durch vorsichtiges Neigen des
Filtrators nach einer Seite zum Ablaufen bringen kann.
Ferner sind wir mit einigen weithalsigen Stöpselgläsern versehen,
die mit Konservierungsflüssigkeit gefüllt sind, und von denen jedes
zur Aufnahme eines Fanges bestimmt ist. Am bequemsten ist die
Anwendung der Pikrinschwefelsäure, die folgende Zusammensetzung hat:
300
Raumteile
Wasser,
100
„
einer konzentrierten wässerigen Lösung von Pikrinsäure,
2
„
von konzentrierter Schwefelsäure.
Fig. 49.
Der Fang kommt, wie weiter unten gezeigt werden soll, in diese Mischung
und kann in derselben bleiben, wenn die Bearbeitung sogleich vor sich
gehen soll. Muss der Fang jedoch einige Zeit stehen, so empfiehlt es
sich, nachdem die Organismen zu Boden gesunken sind, die überschüssige
Säure abzugiessen und durch Alkohol von 60% zu ersetzen, der mehrmals
gewechselt werden sollte. Jedoch muss das mit der grössten Vorsicht
geschehen, da noch zahlreiche Diatomeen in der Flüssigkeit suspendiert
sind und sich sehr langsam absetzen. Bleiben die Organismen zu lange
in der Säure, so werden die Kalksalze aus einigen ausgezogen und noch
anderweitige Veränderungen bewirkt, die Tiere werden schliesslich ganz
weich und sind für nachfolgende Bearbeitung wenig geeignet.
[S. 268]
Ebenso wirksam ist eine konzentrierte Lösung von Sublimat; bei dieser
tritt nur der Übelstand ein, dass man sehr lange mit Wasser[XCIII]
auswaschen muss, um eine nachfolgende Ausscheidung nadelförmiger
Sublimatkrystalle zu verhindern; natürlich muss das Wasser später durch
Alkohol ersetzt werden.
[XCIII] Statt dessen kann man Jodalkohol benutzen, den man so
lange wechseln muss, bis die Färbung des Jods nicht mehr verloren geht.
Wie andere Fixierungsmittel sich bewähren, kann ich nicht angeben,
glaube aber, dass Osmiumsäure (event. Chromosmiumessigsäure) sehr gut
anwendbar ist. Der Fang wird in ein Glas, in dem etwas Wasser sich
befindet, gebracht und darauf einige Tropfen Osmiumsäure zugesetzt.
Nach einigen Minuten oder sobald alle Organismen getötet sind, wird das
Glas mit Wasser gefüllt, dann lässt man die Organismen sich absetzen,
decantiert und setzt schliesslich Alkohol zu.
Von kleineren Apparaten sind noch vorhanden ein Spatel, eine
Spritzflasche, eine Giesskanne und einige Papierzettelchen, die mit
Bemerkungen versehen und in die Fanggläser gelegt werden können.
Unterdessen sind wir an dem Punkte angelangt, an dem wir die
Untersuchung auszuführen gedenken. Das Schiff hält still und das
Fischen kann beginnen. Das Tau, an dem das Vertikalnetz
befestigt ist, wird über eine Rolle, die an einem galgenartigen,
eisernen Gestelle angebracht ist, gelegt. Der freie Schenkel dieses
sogenannten „David“ ragt über die Schiffswand hinaus und erleichtert,
da die ganze Vorrichtung um ihre Achse drehbar ist, das Einholen des
Netzes. Das Netz, an dem der Eimer angeschraubt ist, wird in das
Wasser hinabgelassen, zuerst langsam, damit das Netzzeug angefeuchtet
wird, dann etwas schneller. Die Hand, die das Tau leitet, kann
leicht den Zug des sinkenden Netzes spüren und daher auch den Moment
wahrnehmen, in welchem der Netzring auf dem Boden aufstösst. Um
letzteres jedoch zu verhindern, ist es ratsam, vorher zu loten, dann
kennt man die Wassertiefe und auch die Bodenbeschaffenheit, da sich[S. 269]
Grundproben an dem unten mit Talg versehenen Lote eindrücken. Das
Tau des Vertikalnetzes trägt in Entfernungen von je 1 m bunte
Läppchen und alle 10 m anders gefärbte. Man kann also das Tau
langsam ablaufen lassen, da man die gelotete Tiefe ablesen kann und
kein Aufstossen zu befürchten braucht. Sobald das Netz in der Tiefe
angelangt ist, wird es mit mittlerer Geschwindigkeit von ½–¾
m pro Sekunde senkrecht aufgezogen. Die Geschwindigkeit hängt
von der Filtrationsgrösse[XCIV] des Netzzeuges ab. War z. B. das Netz 20
m hinabgelassen, so können wir, da die Netzöffnung 0.1 qm
beträgt, die Wassermenge, die durch das Netz gegangen sein sollte,
berechnen, sie ist 20×0.1 qm = 2 cbm gross. In Wahrheit
ist jedoch etwas weniger Wasser filtriert worden, nämlich nur 1.8
cbm, wie Versuche von Hensen ergeben haben. 2 cbm
würden durch den Netzring gehen, wenn kein Netz daran hinge, so wird
aber durch den Widerstand des Netzzeuges jener Bruchteil (10%) über den
Netzring abfliessen. Sobald das Netz über dem Wasserspiegel angelangt
ist, wird dasselbe von aussen mit Wasser beworfen. Dadurch wird das
dem Netze anhaftende Material in den Eimer hinabgespült, durch dessen
filtrierende Fläche das überschüssige Wasser abläuft. Nun befinden sich
alle Organismen im Eimer und zwar in einer verhältnismässig kleinen
Wassermenge, die nun weiter zur Verarbeitung kommt.
Der Eimer wird vom Netze gelöst, die Schraube, die sich in der Röhre
am Boden des Eimers befindet, herausgedreht, so dass der Inhalt in
eine darunter gestellte Flasche gelangt. Aus der Flasche kann man dann
nach und nach die Masse in den Filtrator giessen. Diese Methode ist
sicherer, als wenn man den Fang direkt aus dem Eimer in den Filtrator
bringt, da bei schwankendem Schiffe leicht etwas vorbeilaufen kann.
Das Wasser sickert nun allmählich durch die Gazewände des Filtrators
durch, und zwar verschieden schnell, je nach der Beschaffenheit des
Fanges. Sind viel Diatomeen oder Nostocaceen (Limnochlide)[S. 270] vorhanden,
so hat der Fang ein schleimiges Aussehen und filtriert sehr langsam.
Sind dagegen Copepoden oder Peridineen am zahlreichsten, so läuft
das Wasser sehr schnell ab, da sich die Poren des Netzzeuges nicht
verstopfen. Nachdem auf diese Weise die Organismen ziemlich vollständig
vom Wasser befreit sind, wird die Glasplatte, denn auf dieser hat
sich jetzt der Fang niedergesetzt, unter dem Filtrator hervorgenommen
und mit Spatel und Spritzflasche werden die Tiere und Algen in die
Konservierungsflüssigkeit gebracht, in der sie bis zur Verarbeitung
bleiben oder, wie oben auseinandergesetzt ist, in Alkohol übertragen
werden. In jedes Glas wird ein Zettelchen gelegt, auf dem der Fundort,
das Datum, die Tiefe des Planktonzuges, die Konservierung, die
Temperatur des Wassers und allenfalls noch die Windrichtung angegeben
sind.
Die nun folgenden Arbeiten, die die quantitative Bestimmung des
Fanges bezwecken, werden ausgeführt, nachdem wir zu Hause angelangt
sind. Zuerst wird das Volumen des Fanges festgestellt. Zu dem
Zwecke wird der Inhalt eines einen Fang enthaltenden Glases in einen
Messcylinder entleert. Nach und nach sinken die Organismen zu Boden,
die grösseren schneller, die kleinen langsamer. Unter letzteren sind
namentlich die Diatomeen zu erwähnen, diese setzen sich, wenn sie
in grösseren Mengen vorkommen, so langsam ab, dass nach tagelangem
Stehen noch immer eine Volumenverringerung zu beobachten ist. Befinden
sich grosse Tiere im Fange, so muss bei diesen das Volumen besonders
bestimmt werden und dieses geschieht am besten und genauesten durch
„Verdrängung“, d. h. das Tier wird in einen Messcylinder
hineingebracht, in dem sich eine bekannte Menge Flüssigkeit (Wasser
— oder Alkohol, wenn der Fang in Alkohol war —) befindet, dann
kann man durch das Steigen der Wasseroberfläche das Volumen des
Körpers bestimmen. Meist hat sich die Masse in 24 Stunden so weit
abgesetzt, dass das Volumen derselben abgelesen werden kann. Da die
Volumenbestimmung zum Vergleiche der einzelnen Fänge unter sich dienen
soll, so ist es zweckmässig, allen Fängen die gleiche Zeit zum Absetzen
zu lassen und zwar genügen dazu 24 Stunden.
[S. 271]
Nachdem so das Volumen des Fanges festgestellt ist, wird zur
Zählung der Organismen geschritten. Es ist selbstverständlich,
dass nicht alle Individuen des Fanges gezählt werden können, das
beweisen schon folgende Zahlen, die ich Zählungen Hensens[XCV]
entnehme: Hensen fand im Oktober 1884 in 1 cbm Ostseewasser
(Kieler Bucht) 13 Millionen Ceratium tripos, und im März 1885
ebenda 102 Millionen Rhizosolenia semispina, und wenn wir gar
lesen, dass im September sich im Stettiner Haff in ½ cbm
Wasser 9983 Mill. Fäden von Limnochlide[XCVI] fanden, dann ist es klar,
dass diese Zahlen auf anderem Wege gewonnen sind, als durch Zählung
jedes einzelnen Individuums. Die sinnreich von Hensen erdachte
und angewendete Methode ist folgende:
[XCVI] Die letzte Zahl entnehme ich einem Zählungsprotokolle
von Herrn Geheimrat Hensen, das er mir freundlich für diese
Arbeit überliess und das am Ende des Kapitels sich abgedruckt findet.
Die folgenden Zahlen sowie Betrachtungen beziehen sich auf dieses
Protokoll. Siehe auch Hensen[105].
Von dem Fange wird die überschüssige Pikrinschwefelsäure abgegossen
und dann Wasser so viel zugesetzt, bis sich die Masse gut
durcheinanderschütteln lässt. Befindet sich der Fang in Alkohol, so
muss der Alkohol durch Wasser erst ausgewaschen werden, was mehrere
Tage in Anspruch nimmt. Nehmen wir an, dass nach der Verdünnung
das Volumen 500 ccm betrage, so ist es klar, dass sich in 1
ccm die verschiedenen Organismen nicht in der gleichen Zahl
finden. Während wir vielleicht eine Leptodora finden, befinden
sich in demselben Volumen gegen 20 Millionen Limnochlide. Um letztere
zählen zu können nehmen wir von dieser ersten Verdünnung 1 ccm
ab und verdünnen ihn auf 1000 ccm, dann haben wir in dieser
zweiten Verdünnung in jedem Kubikcentimeter nur 200000001000 = 20000
Fäden der Alge. Von dieser Verdünnung können wir ⅒ ccm,
der 2000 Fäden enthalten würde, bequem zählen. In dieser Wassermasse
würden wir aber keinen einzigen der selteneren Organismen finden, daher
dürfen wir, wenn wir diese zählen wollen, die Verdünnung nicht so weit
treiben, sondern vielleicht 1 ccm der ersten Verdünnung nur auf
100 oder 10 ccm verdünnen, für[S. 272] die ganz seltenen werden wir
aber die erste Verdünnung selbst zur Zählung benutzen.
Da, wie wir gesehen haben, sich in 1 ccm Flüssigkeit noch
Millionen von Organismen vorfinden können, so muss das Entnehmen einer
bestimmten Menge von Flüssigkeit durch ganz besondere Vorkehrungen
geschehen; denn das Abmessen in einem Messcylinder kann für diesen
Zweck nur ganz rohe Werte geben. Es sind daher von Hensen
besondere Stempelpipetten[XCVII] (Fig. 50) konstruiert worden,
die ganz Vorzügliches leisten. Solch ein Instrument besteht aus einem
kräftigen Glasrohr (B), das unten ganz eben abgeschliffen ist.
In diesem Rohr bewegt sich ein Stempel, der abwechselnd aus Kork-
(h) und Metallplatten (i) zusammengesetzt ist, die durch
zwei Schrauben fest an einander gedrückt werden. An diesen Stempel
ist ein massiver Metallcylinder (m) angeschraubt, der genau
in die Glasröhre hineinpasst. Von diesem Cylinder wird nun so viel
Metall ausgeschliffen, dass zwischen ihm und dem Glasrohr (B)
genau ein bestimmtes Volumen bleibt, z. B. 1 ccm. Dies wird
so bewerkstelligt, dass zuerst ein Teil aus dem Metallcylinder
herausgenommen wird. Dann wird die Pipette gewogen, hierauf wird die
Höhlung mit Quecksilber gefüllt und wieder gewogen. Da man nun das
Gewicht eines Kubikcentimeters Quecksilber kennt, so kann man genau
den Punkt treffen, wo die Höhlung im Stempel 1 ccm fasst.
Es sind von diesen Stempelpipetten sechs verschiedene Grössen zum
Gebrauche nötig, nämlich zu 0.1; 0.2; 0.5; 1; 2.5; 5 ccm. Diese
Pipetten werden so angewendet, dass sie mit vorgestossenem Stempel
in ein durch einen durchbohrten Kork verschlossenes Glas mit starken
Wandungen (A), in dem die Flüssigkeit sich befindet, von der
ein Teil entnommen werden soll, hineingestellt werden (siehe Fig.
50). Die Masse wird durch kräftiges Schütteln aufgerührt, und sobald
sich die Organismen möglichst gleichmässig verteilt haben, wird das
Glasrohr B niedergestossen; dann ist zwischen dem Glasrohr
und dem Stempel m ein genau bekanntes Volumen Flüssigkeit
eingeschlossen. Ehe man[S. 273] jedoch diese Flüssigkeitsmenge entleert, ist
es nötig, den unteren Rand des Glasrohres mit Fett zu bestreichen, da
sonst leicht ein Tropfen daran hängen bleiben kann. Nach Entleerung
des Volumens wird dann noch mit einigen Tropfen Wasser nachgespült,
so dass man sicher sein kann, dass keine Organismen zurückgeblieben
sind. Dieses abgemessene Volumen wird dann zur Verdünnung benutzt resp.
gezählt.
Haben wir uns eine genügende Verdünnung hergestellt, dann kann die
Zählung beginnen. Hierzu wird das Zählmikroskop[XCVIII]
benutzt. Dieses Mikroskop zeichnet sich durch seinen Objekttisch aus.
Dieser ist so gross, dass er Glasplatten von 11½ × 10 cm
fassen kann, und was die Hauptsache ist, er ist durch zwei Schrauben
sowohl von vorn nach hinten, als seitwärts verschiebbar. Auf den
rahmenförmigen Objekttisch werden Glasplatten aufgelegt, die fein mit
dem Diamanten liniiert sind und zwar hat jede Platte ein bestimmtes
Liniensystem. Wählt man die passende Vergrösserung, so kann man im
Gesichtsfelde zwei parallele Linien laufen sehen, und wenn man an einer
seitlichen Schraube dreht, so bewegt sich die Glasplatte[S. 274] langsam
weiter, wobei man immer den Raum zwischen denselben Linien im Auge
behalten kann. Ist man am Ende eines Zwischenraumes angelangt, so
wird mit Hilfe der anderen Schraube der Objekttisch senkrecht zu der
vorherigen Richtung um einen Zwischenraum weiter gedreht und dann in
diesem die Beobachtung weiter fortgesetzt. So kann man allmählich die
ganze Platte mit dem Mikroskop untersuchen und ist sicher, dass kein
Punkt übersehen ist.
Bringen wir nun auf eine liniierte Glasplatte ein bestimmtes Mass
einer Verdünnung, so können wir die Zahl der einzelnen Organismen,
die sich in diesem Volumen befinden, bestimmen. Die Verdünnung wählt
man am besten so, dass man von der häufigsten Spezies nie mehr als
3000 und nie weniger als 1000 auf der Platte hat. Würde es sich nur um
eine Spezies handeln, so wäre die Zählung leicht auszuführen.
Man brauchte nur die Platte allmählich zu durchsuchen und jedes
Individuum, das in das Gesichtsfeld kommt, zu zählen, dann wüsste
man, wie viel Organismen auf der Platte sind und könnte, da man die
Verdünnung kennt, die Summe der Organismen im ganzen Fange berechnen.
Hätten wir z. B. eine Verdünnung von 1 : 10 angewendet und 1 ccm
Verdünnung durchgezählt und fanden 146 Coscinodiscen, dann wären im
ganzen Fange (von 500 ccm) 146 × 10 × 500 = 730000 Coscinodiscen
vorhanden.
Handelt es sich jedoch um mehrere Spezies, so kann man diese nicht im
Kopfe getrennt zählen. Doch auch hier hat Hensen Rat geschafft.
Da in einem Fange 30–50 verschiedene Spezies von Tieren und Pflanzen
vorhanden sind, so werden an einem Setzerkasten, der ebenso viel
Fächer enthält, die Namen der vorhandenen Organismen angebracht, für
jede Spezies ein Fach. Untersucht man jetzt eine Platte, so werden die
mannigfaltigen Organismen nicht mehr gezählt, sondern sobald irgend
einer im Gesichtsfelde sich blicken lässt, wird für ihn ein Pfennig
(Spielmarke etc.) in sein betreffendes Fach gelegt. So kann man leicht
eine Platte, auf der sich 50 verschiedene Arten durcheinandergemengt
befinden, zählen.[S. 275] Auf den ersten Platten werden die Diatomeen, die
meist am zahlreichsten in einem Fange vorhanden sind, gezählt, andere
Organismen natürlich auch berücksichtigt. Zuerst wird ein stark
verdünnter Teil des Fanges genommen, da trotzdem genug Individuen
auf die Platte kommen. Die Vergrösserung muss anfangs sehr stark
sein, etwa 200, zum Zählen der Diatomeen und anderer Algen. Auf die
Platte kommt nur 0.1 ccm Flüssigkeit, die mit der betreffenden
Stempelpipette abgemessen wird. Für die starke Vergrösserung bildet
diese geringe Wasserschicht aber immerhin noch ein Hindernis alle
Organismen zu sehen; hat man das Mikroskop auf die Oberfläche der
Platte eingestellt, so entgehen einem die Organismen, die an der
Oberfläche der Flüssigkeit sich befinden. Daher ist es vorteilhaft, die
Diatomeen trocken zu zählen. Es wird zu diesem Zwecke ein bestimmtes
Volumen Flüssigkeit auf eine Platte gebracht und diese dann der Wärme
eines heizbaren Objekttisches oder eines Ofens ausgesetzt, damit die
Flüssigkeit verdunstet; dann sind die Diatomeen auf der Platte in einer
Ebene ausgebreitet und können nicht so leicht übersehen werden. Da
die Mischungen und Verdünnungen nie ganz genau sein können, so wird
natürlich die Zählung jeder neuen Platte etwas abweichende Resultate
ergeben, es fragt sich daher, wie lange eine Spezies gezählt werden
muss; wann solch ein Grad von Genauigkeit erreicht ist, um von den
wenigen Zählungen auf die quantitative Zusammensetzung des ganzen
Fanges schliessen zu können. Im allgemeinen lässt sich sagen, dass
es bei den häufigsten Formen genügt, wenn man einen Bruchteil (z. B.
⅒) der Quadratwurzel sämtlicher Individuen zählt. Haben wir (siehe
Protokoll) auf der ersten Platte für Melosira 27 Fäden gefunden
und wissen wir, dass die durchzählte Wassermasse der 5000000. Teil
von dem ganzen Fange ist, so würden wir nach dieser ersten Zählung
schliessen, dass 135000000 Melosira im Fange sein werden, daraus
nehmen wir ⅒ der Quadratwurzel = 1162. Haben wir also mindestens
1162 Melosira gezählt, so können wir diese aus den Zählungen
ausscheiden, d. h. wir brauchen sie nicht mehr mitzuzählen.
[S. 276]
Um die Genauigkeit zu finden, bis zu welcher die Zählung erfolgen
muss, führt Hensen[XCIX] noch folgende Erwägung an. Nachdem
einige Zählungen gemacht sind, zieht man aus diesen das Mittel.
Denken wir nun, dass noch eine Zählung hinzugekommen wäre und diese
mit der am meisten abweichenden übereinstimmen würde, und nähmen
wir dann aus diesen das Mittel, so genügen die Zählungen, wenn das
Resultat sich nicht mehr als um 5% ändert. Im Protokoll finden wir für
Melosira die Zahlen 382, 396, 396, Summe 1174, Mittel daraus
391. Käme noch eine Zählung hinzu und zwar 382, so wäre die Summe 1556,
Mittel daraus 389.
Das Resultat weicht also nur um 0.6% ab, die Zählung ist genau genug,
kann also unterbrochen werden; jedoch ist es stets besser, mehr
Platten zu zählen, als zu wenig.
Haben wir eine genügende Genauigkeit erreicht, so können wir die
Diatomeen beim Zählen überspringen und schwächere Verdünnung und
schwächere Vergrösserung zur Zählung benutzen. Für seltenere Formen
wird schliesslich die erste Verdünnung benutzt und von dieser 1
ccm, zuletzt 2.5 durchzählt, was meist sehr schnell geht, da
man nur mit sehr schwachen Vergrösserungen zu arbeiten braucht und nur
wenige Tiere zu zählen hat.
Die einzelnen Zählungen werden notiert und zwar in Form eines
Protokolles. Ein solches Protokoll ist im Anhange beigegeben
und aus diesem die Einrichtung zu ersehen. Folgendes möge noch zur
Erläuterung desselben erwähnt sein. In der linken obern Ecke findet
sich Datum und Ort des Fanges, hier also: 13. September 1887, Stettiner
Haff. Über sämtliche Fänge wird ein Journal geführt, es bedeutet J.
No. 1 = Journal No. 1. Daselbst finden sich die näheren Daten, die bei
Erlangung des Fanges als wichtig notiert wurden, wie die Tiefe des
Fanges (hier 5 m), die Temperatur des Wassers und der Luft,
Windrichtung,[S. 277] Beschaffenheit des Fanges, ob locker, flockig, schnell
absetzend; letztere Aufzeichnungen sind wichtig, da sie, wie wir oben
gesehen haben, schon einen Einblick in die Zusammensetzung des Fanges
erlauben.
In dem Protokolle sehen wir einige Vertikalreihen, darauf
Horizontalreihen. Betrachten wir zuerst die Vertikalreihen. In der
ersten Kolumne mit der Überschrift „Art der Untersuchung“ steht überall
feucht, d. h. alle Platten, die durchzählt worden sind, enthielten die
Organismen in Wasser suspendiert. Den Gegensatz bilden die trockenen
Platten, die, wie oben erwähnt wurde, meist zum Zählen der Diatomeen
verwendet werden. In unserem Fange waren Diatomeen in verhältnismässig
geringer Anzahl vorhanden, Melosira, Coscinodiscus und
Bacillaria zusammen etwa 100 Millionen, diesen standen von
anderen Algen allein Limnochlide mit 9653 Millionen gegenüber. Letztere
würden beim Trocknen bis zur Unkenntlichkeit geschrumpft sein und daher
die Zählung vereitelt haben, während die kieselschaligen Diatomeen
nicht nur ihre Form behalten, sondern auch leichter in trockenem
Zustande zu bestimmen sind.
In der zweiten Vertikalreihe sind die Vergrösserungen angegeben,
bei denen die einzelnen Platten gezählt worden sind. Das stärkste
der angewendeten Objektive (Vergrösserung 200) hatte einen solchen
geringen Abstand von der Glasplatte oder von der Flüssigkeitsmenge
(0.1 ccm), die sich auf der Platte befand, dass es nur
der gleichmässigen und leichten Verschiebung des Objekttisches
zuzuschreiben ist, dass das Objektiv nicht in das Wasser eintauchte.
Dadurch ist einerseits einer weiteren Erhöhung der Vergrösserung ein
Ziel gesetzt, anderseits würde die Zählung einer Platte mit noch
stärkeren Objektiven viel längere Zeit in Anspruch nehmen und die
Augen übermässig anstrengen. Beiläufig will ich noch erwähnen, dass
die Zählung solch einer Platte etwa drei Stunden dauert[C]. Nach und
nach nahm die Vergrösserung, bei der gezählt wurde, ab bis auf 22;
mit letzterer wurden nur noch Leptodora,[S. 278] Milben und ein nicht
bestimmtes Rädertier gezählt. Die anderen Organismen waren entweder
schon zu zahlreich auf der Platte und lagen infolgedessen zu dicht an
einander, oder es war schon die genügende Zahl gezählt, oder endlich
reichte die Vergrösserung nicht mehr aus, die kleineren Organismen
genau und schnell zu erkennen. Aus letzterem geht hervor, dass man nie
ein schwächeres Linsensystem anwenden darf, ehe nicht alle Organismen
genügend gezählt sind, die mit diesem System nicht mehr genau erkannt
werden können.
[C] Die genaue und bis in die Einzelheiten gehende Zählung
eines Fanges nimmt etwa vierzehn Tage in Anspruch bei vierstündiger
Arbeitszeit.
In der dritten Reihe ist die Grösse der Verdünnung angegeben.
Aus dem oben Gesagten erklären sich die Angaben leicht. 1:1000 heisst
also, dass 1 ccm der ersten Verdünnung mit 999 ccm Wasser
verdünnt wurde, so dass das Gesamtvolumen 1000 ccm = 1 Liter
war. Man richtet sich am besten solche Messflaschen ein, die 1000, 500,
200, 100, 80 ccm halten, und benutzt dazu verschieden grosse
Kochflaschen, die eine abgemessene Flüssigkeitsmenge so aufnehmen
können, dass diese gerade noch in den Hals der Flasche hineinragt,
dort bringt man mit dem Diamant eine Marke an. Dann hat man für jede
Verdünnung sogleich eine Flasche bereit. Zu den letzten Zählungen
ist die erste Verdünnung genommen worden, es wurden aber auch nur
die grössten Tiere gezählt, so auf einer Platte, die 2.5 ccm
Flüssigkeit enthielt, nur Hyalodaphnia Kahlbergensis, Daphnia
longispina, Sida crystallina, Leptodora hyalina,
Milben und das oben erwähnte Rädertier. Es waren im ganzen (in No.
27) 94 Individuen, so dass, trotz der grossen Flüssigkeitsmenge, die
Zählung nur ungefähr eine halbe Stunde in Anspruch nahm.
Die nächste Spalte enthält die „Nummern“ der gezählten Platten.
Meist genügen 22–24 Platten; in unserem Fange waren aber die grossen
Formen selten, so dass, um einen einigermassen sicheren Einblick in
die Massenhaftigkeit ihres Vorkommens zu erhalten, mehrere Platten
allein für sie verarbeitet werden mussten (Platte 26–31). Die
fortlaufenden Nummern der Platten sehen wir wieder als Kopfzahlen bei
den Horizontalreihen, zu denen wir weiter unten übergehen werden.
[S. 279]
Wir überspringen einige Spalten und sehen uns die letzte mit der
Überschrift „Gebrauchtes Mass“ an. Die Zahlen dieser Rubrik
besagen, eine wie grosse Wassermenge jedesmal zur Untersuchung benutzt
worden ist. Aus der ersten Zeile ersehen wir, dass 0.1 ccm
gezählt wurde und zwar — wie aus den daneben stehenden Reihen
hervorgeht — von der Verdünnung 1:1000 bei einer Vergrösserung von
200 auf feuchter Platte. Die Flüssigkeitsmengen werden mit den oben
beschriebenen Stempelpipetten abgemessen und auf die Platte übertragen.
Es sind, wie das Protokoll ausweist, alle dort erwähnten Grössen in
Anwendung gekommen, mit Ausnahme von 5 ccm, die nur zum Abmessen
der Volumina zwecks der Verdünnung gedient hat (siehe Platte 21–25,
auch 10–20).
Kehren wir nun zu der alten Reihenfolge zurück, so treffen wir die
Spalte, die die Wahren Masse enthält. Diese unterscheiden
sich insofern von dem „Gebrauchten Mass“, als sie nicht angeben,
wie viel Flüssigkeit auf jeder einzelnen Platte durchzählt wurde,
sondern der wievielte Teil diese Flüssigkeitsmenge von der ganzen
betreffenden Verdünnung ist. Wie wir diese Zahlen erhalten, ergiebt
sich am leichtesten an der Hand unseres Protokolls: Bei Platte 1 haben
wir 1 ccm der ersten Verdünnung auf 1000 ccm (zweite
Verdünnung) gebracht; würde ich hiervon 1 ccm abnehmen, so wäre
dieser der 0.001. Teil der ganzen zweiten Verdünnung oder des einen
Kubikcentimeter der ersten Verdünnung, den ich für die zweite benutzt
habe. Zur Zählung ist aber nur 0.1 ccm verwendet, dieser ist
dann nur der 0.0001. Teil der ganzen zweiten Verdünnung, enthält also
auch nur den 0.0001. Teil der Organismen der ganzen zweiten Verdünnung
resp. des 1 ccm der ersten Verdünnung, von dem die zweite
Verdünnung hergestellt ist.
Bei Platte 10 haben wir die Verdünnung 10:100, also 0.1; davon 0.2
ccm genommen, erhalten wir 0.02 als wahres Mass.
Bei No. 21 haben wir 30:100, also 0.3, ein Kubikcentimeter davon also
0.3 wahres Mass.
Dieses wahre Mass ist nun wichtig für die Berechnung des Koeffizienten.
In der Rubrik „Berechnung“ ist dieselbe ausgeführt.[S. 280] Bei der
Besprechung des wahren Masses sahen wir, wie wir z. B. bei Platte 1
fanden, dass die gezählten Organismen auf dieser Platte den 0.0001.
Teil der in 1 ccm enthaltenen Wesen bilden. Beziehen wir aber
die gezählte Zahl auf das ganze Flüssigkeitsvolumen von 500 ccm
(erste Verdünnung), so haben wir nur 5000.0001 gezählt, das ist der
50000001 = 5000000. Teil. Finden wir also auf der ersten Platte in 0.1
ccm der Verdünnung 1:1000 2024 Limnochlide-Fäden, so wissen wir,
dass wir in dem ganzen Fange 2024 × 5000000 Limnochlide ungefähr
werden finden müssen, was 10120 Millionen ergeben, eine Zahl, deren
Fehler durch weitere Zählungen eingeschränkt wird.
Haben wir eine Spezies während mehrerer Platten gezählt und sehen
wir, dass wir abbrechen können[CI], dann handelt es sich darum,
den Koeffizienten für die Summe der gezählten Individuen zu
finden. Zu dem Zwecke addieren wir die wahren Masse aller der Platten,
auf denen diese Spezies beobachtet wurde, und verfahren wie oben
für eine Platte angegeben ist. Wir haben z. B. für Limnochlide fünf
Platten gezählt (1–5) und finden die Zahlen 2024, 1835, 2048, 1954,
1792, Summe 9653. Die Summe der wahren Masse für diese fünf Platten
ist 5 × 0.0001 = 0.0005, dann erhalten wir 5000.0005 = 1000000 als
Koeffizienten der Summe. Die gezählten Individuen 9653 bilden also den
1000000sten Teil aller im Fang vorhandenen Limnochlide, das ergiebt
9653000000 Limnochlide. Habe ich Spirogyra erst von der zwölften
Platte bis zur zwanzigsten gezählt, so summieren für diese Alge nur die
wahren Masse dieser Nummern, also Summe 12–20 = 0.51, und wir erhalten
den Koeffizienten 980.
Durch die letzteren Betrachtungen sind wir nun schon bei den
Horizontalreihen angelangt.
In einer Spalte derselben stehen die Namen der Organismen, die sich in
dem gezählten Fange befanden. Rechts davon sind[S. 281] dann die Ergebnisse
der Zählung jeder einzelnen Platte angegeben und zwar in der Rubrik,
deren Kopfzahl der betreffenden Platte entspricht. Hört man auf,
einen Organismus mitzuzählen, so steht in der Rubrik der betreffenden
Platte ein Fragezeichen. Wird auf einer Platte von einer Spezies kein
Individuum gefunden, so steht natürlich eine Null. Ist dagegen eine
Spezies beobachtet, aber auf einigen Platten nicht mitgezählt, wie
z. B. bei Spirogyra 1–11, so wird auch hier ein Fragezeichen
gesetzt.
In den beiden letzten Spalten sind zuerst die Summen der gezählten
Individuen jeder Spezies angegeben, dann die Platten, auf denen
diese Organismen gezählt wurden.
Um nun die Gesamtsumme der in dem Fang vorhandenen Tier- und
Pflanzenindividuen zu finden, brauchen wir nur die Summe der
gezählten Organismen mit dem Koeffizienten, welcher der
angewendeten Plattenzahl entspricht, zu multiplizieren. So haben wir
bei Limnochlide 9653 Fäden gezählt, und zwar auf Platte 1–5, der
Koeffizient der Platten 1–5 ist 1000000, also sind im ganzen Fange
9653000000 Limnochlide-Fäden vorhanden.
Diese Gesamtsumme „Gezählte Masse“ steht in einer Rubrik vor den
Namen, damit man mit diesen das Endresultat sogleich übersieht.
Vor der letzteren Rubrik finden wir eine solche mit der Überschrift
„Ganze Masse“. Wie wir oben gesehen haben, wird nicht die ganze
Wassersäule filtriert, die dem Querschnitte des Netzes und der Tiefe
des Wassers, bis zu der das Netz herabgelassen wurde, entspricht,
sondern ein kleiner Teil fliesst über den Netzrand ab. Hensen
hat deshalb für jedes Netz den Filtrationskoeffizienten[CII] berechnet,
der besagt, mit welcher Zahl man Volumen oder Anzahl der Organismen
eines Fanges multiplizieren muss, um die wirklichen Werte zu finden,
wenn die ganze Flüssigkeitssäule filtriert worden wäre. In unserem
Falle war der Koeffizient 1.034. Von Leptodora waren z. B. 371
Individuen im Fange. In der Wassersäule von 0.1 qm Querschnitt
und 5 m Höhe waren aber[S. 282] 371 × 1.034 = 384 Individuen vorhanden.
In Folgendem habe ich aber die Zahlen der gezählten Masse benutzt.
Nachdem wir in Vorhergehendem die Hensensche Methode der quantitativen
Untersuchung des Plankton kennen gelernt haben, erübrigt es noch, auf
die qualitative Zusammensetzung des Süsswasserplankton[CIII] einzugehen.
Jedoch muss ich noch ein paar Worte vorausschicken. Die Planktonmethode
ist bis jetzt fast nur auf das Meer angewendet worden, nur einmal ist
bei einer Fahrt in der östlichen Ostsee von Hensen auch ein Fang
im Stettiner Haff[105], also im Süsswasser, gemacht worden. Die Fänge
wurden, wie wir oben gesehen haben, sofort konserviert, daher kommt es,
dass manche Organismen bei nachfolgender Zählung ganz unkenntlich sind,
da sie sich beim Töten zusammengezogen haben; dieses ist namentlich der
Fall bei Rädertieren[CIV]. Es ist daher nötig, dass an Ort und Stelle
auch lebendes Material untersucht wird, denn, wenn dieses bestimmt ist,
sind die Organismen leicht in konserviertem Zustande wiederzuerkennen.
Dieses ist nun bei einer grösseren Fahrt sehr schwer ausführbar, da
sich auf dem Schiff, namentlich bei bewegter See, schwer oder gar nicht
mikroskopieren lässt. Anders verhält sich die Sache, wenn das Institut,
in dem die Arbeiten ausgeführt werden sollen, direkt am Wasser liegt,
da kann quantitative und qualitative Bestimmung Hand in Hand gehen, so
ist das hier in Kiel, und auch in der unlängst von Zacharias
errichteten Süsswasserstation zu Plön der Fall. Es müssen daher in
den Protokollen vorläufige Namen für die unbestimmten[CV] Organismen
gesetzt werden und späterer Zeit überlassen bleiben, das Versäumte bei
Gelegenheit an Ort und Stelle nachzuholen. Folgendes ist ferner auch
noch von Bedeutung. Es genügt nicht, einen Fang zu beliebiger
Zeit zu machen, sondern es müssen die Planktonfahrten in bestimmten
Zeitabschnitten (alle zwei oder vier[S. 283] Wochen)[CVI] unternommen werden,
dann erhält man erst einen Einblick in die wahre Zusammensetzung des
Plankton, das in fortwährendem Werden und Vergehen begriffen ist.
Dieses wäre eine sehr dankbare Arbeit für eine Süsswasserstation.
[CIII] Von Häckel[100] S. 21 Limnoplankton genannt.
[CIV] Ausgenommen von diesen sind nur die gepanzerten
(Loricata), die trotzdem leicht zu erkennen sind.
[CV] Leider war der Fang aus dem Stettiner Haff, da er als
ausgebraucht betrachtet wurde, weggeschüttet, so dass auch nachträglich
keine nähere Untersuchung vorgenommen werden konnte. Es empfiehlt sich
daher, das Material stets aufzubewahren.
[CVI] So werden die Untersuchungen in der Kieler Bucht von
Prof. Brandt[97] seit September 1888 ausgeführt und noch
fortgesetzt.
Nach dem Gesagten ist es klar, dass die folgenden Darlegungen wenig
positives bringen können, es kann nur gezeigt werden, in welcher Art
ein Fang oder eine fortlaufende Reihe solcher verwertet werden können.
Ich beabsichtige also nur ein Beispiel zu der oben erläuterten
Methodik zu geben.
Beginnen wir mit den Algen, der Urnahrung, so fallen uns die
Vertreter zweier Ordnungen durch ihr massenhaftes Auftreten auf,
es sind die Diatomeen und Schizophyceen. Weniger zahlreich sind
Protococcoideen, von denen Pediastrum, Gleocystis und
Scenedesmus quadricaudatus gefunden wurden, und Konjugaten, die
durch Spirogyrafäden vertreten sind.
Was die Diatomeen anbelangt, so müsste man glauben, dass sie
wegen ihrer grossen Zahl, trotz des geringen Anteils an organischer
Substanz, eine wichtige Nahrung für die pelagischen Tiere bilden.
Jedoch werden sie, wie Hensen[CVII] beobachtet hat, von allen
Tieren verschmäht. Dagegen erwähnt Seligo[107] in seinen
Hydrobiologischen Untersuchungen, dass die Diatomeen die Hauptnahrung
mehrerer Tierarten[CVIII] bilden. Leider sagt er nicht, für welche. Ihre
Bedeutung ist also in anderer Richtung zu suchen. Alle Diatomeen haben
eine Vegetationsperiode, d. h. sie vermehren sich zu einer bestimmten
Zeit ganz enorm, um dann wieder allmählich oder fast plötzlich zu
verschwinden. Letzterer Umstand ist durch das Bilden von Dauersporen,
die bei vielen Meeresdiatomeen beobachtet sind, leicht erklärlich,
da die Spore alsbald zu Boden sinkt. Ob bei Süsswasserdiatomeen die
gleichen Verhältnisse vorkommen, kann ich nicht angeben. Die Sporen
enthalten eine sehr[S. 284] konzentrierte organische Nahrung, die den Tieren
der Tiefenregion wohl zu gute kommt.
[CVIII] Bei meinen Untersuchungen hiesiger Süsswasserseen habe
ich den Darm von Hyalodaphnien und Bosminen dicht mit Melosirazellen
angefüllt gefunden (1891).
Da jede Diatomee aus zwei Schalen besteht, so muss man jede
vollständige Diatomee als 2 zählen, jede allein liegende Schale als
1, und dann die erhaltene Summe durch 2 dividieren, dann erhält man
die Zahl der Zellen. Bei Melosira würde dieses Verfahren zu
viel Zeit in Anspruch nehmen, man zählt daher nur die Fäden und
stellt bei einer Zahl von Fäden die Zellenzahl fest. In unserem
Fange fanden sich pro 1 qm[CIX] 984614400 Fäden, von denen
20 = 246 Zellen enthielten, es wären also im ganzen 12110757120 Zellen
vorhanden gewesen. Ohne Berechnung der Zellen kann man Melosira
nicht mit anderen Diatomeen vergleichen. Von Melosira kamen
zwei Arten vor, nämlich M. granulata und eine andere, nicht
näher bestimmte. An 226 Fäden war erstere mit 208, letztere mit 18
beteiligt. Coscinodiscus fand sich in 6440890 und Bacillaria
paradoxa in 7388640 Individuen. Letztere Diatomee zeigt bei ihrer
Zählung ziemlich abweichende Resultate, da mehrere Zellen an einander
liegen und so Platten bilden, andere sind auseinandergefallen, so
dass, wo einmal eineBacillaria gefunden wird, ein ander
Mal eine Platte, die aus mehreren Individuen besteht, sich
vorfindet. In geringerer Anzahl war eine Surirella-Art vorhanden,
nämlich 477570 pro 1 qm. Gegen Melosira tritt aber die
Summe aller übrigen Diatomeen weit zurück.
[CIX] Wenn die Zahlen in folgendem auf 1 qm Oberfläche
berechnet sind, so bedeutet dies stets eine Wassersäule vom Querschnitt
1 qm und einer Tiefe von 5 m.
Die zweite Hauptgruppe der Algen bildeten die Schizophyceen.
Einzelne von diesen verursachen zu Zeiten die sogen. „Wasserblüte“. Sie
sollen den Fischen verderblich werden, jedoch ist die Ursache davon
noch unbekannt.
Vor allen trat Limnochlide flosaquae Ktz. hervor, es wurden
von ihr unter 1 qm Oberfläche 96530000000 Fäden gefunden, von
denen 20 = 294 Zellen enthielten, so dass wir fast 1½ Billion Zellen
erhalten. Die filtrierte Wassermenge betrug fast 0.5 cbm = 500[S. 285]
Millionen Kubikmillimeter, es fanden sich also in 1 cmm Wasser
2838 Zellen = 142 Fäden. Ihre Bedeutung für den Stoffwechsel ist mir
nicht klar, es wäre aber möglich, dass die Sporen dieser Alge ebenfalls
(siehe Diatomeen) den Tieren der Tiefenregion zur Nahrung dienen
könnten.
Neben Limnochlide kamen noch mehrere Arten von Chroococcaceen vor.
Sie sind im Protokoll einfach mit Coccus bezeichnet und mit
einem Beinamen versehen, der auf die Form der Kolonie oder der
einzelnen Individuen Bezug hat. Sie gehörten meist der Gattung
Polycistis an, der sogenannte feinkörnige Coccus war
P. ichthyoblabe. Dagegen ähnelte Coccus 1 (viereckig)
mehr einer Merismopedia.
Die in Pikrinsäure konservierten Algen lassen aber nicht ihre
natürliche Färbung erkennen und sind dann nicht mehr bestimmbar.
Zusammen fanden sich 109824460 Kolonien, die einzelnen Individuen sind
nicht zu zählen.
Zur Urnahrung müssen wir ferner, wie in der Einleitung erwähnt ist, die
Peridineen rechnen. Es waren im Fange drei Arten vorhanden, Ceratium
tripos, C. fusus und Peridinium divergens. Diese drei
Peridineen sind echte Meeresbewohner und es muss daher ihr Vorkommen im
Süsswasser[CX] sehr auffallen. Ich vermutete daher zuerst, dass sie
von früheren Fängen aus der westlichen Ostsee am Netze hängen geblieben
waren, dieses war aber nicht der Fall, da, wie ich nachher erfuhr, ein
ganz neues Netz zum Fischen verwendet war. Es müssen also sich die
Peridineen dem Leben im Süsswasser angepasst haben. Der Salzgehalt im
Stettiner Haff war so gering, dass er mit den Instrumenten nicht mehr
gemessen werden konnte. Von den Peridineen, die bisher im Süsswasser
gefunden wurden, sind keine beobachtet.
[CX]Hensen[105] führt auch an, dass Ceratium
tripos von Pringsheim nahe bei Berlin gefunden ist, also im
Süsswasser (S. 74).
Von den Protozoen finden wir im Plankton die Tintinnen. Dieses
sind die niedrigststehenden Tiere, die durch einen Mund geformte
Nahrung aufnehmen. Worin diese Nahrung besteht, ist[S. 286] nicht bekannt,
Hensen[CXI] vermutet darunter der geringen Grösse der Tintinnen
wegen noch kleinere Wesen, als bis jetzt nachgewiesen sind, und die
regelmässig durch das Netzzeug hindurchschlüpfen. Tintinnen sind in
zwei Arten im Fange vorhanden gewesen. Tintinnus ventricosus,
der auch im Meere zahlreich vorkommt, war sehr häufig, auf 1 qm
Oberfläche 1586040 Tiere, noch mehr war T. borealis Hensen[CXII]
zu finden, eine neue Art, die eine Länge von nur 0.048 mm hat,
und an Chaetoceros und anderen Organismen festsass. Von ihm
waren 2881960 Individuen unter 1 qm Oberfläche.
[CXII]Hensen[105] hier auch Beschreibung und Figur.
Von Rädertieren wurden sechs Arten gefischt. Drei davon gehörten
zu der Gattung Anuraea, die anderen drei konnten nicht bestimmt
werden, da sie zu sehr kontrahiert waren, es befand sich darunter eine
sehr grosse durchsichtige Art[CXIII], von der nur 3180 unter 1 qm
Oberfläche lebten.
Die beiden anderen unbestimmten Rotatorien waren sehr zahlreich zu
finden, das eine in der Zahl von 776240, das andere von 3203200 unter
derselben Oberfläche. Von Anuraea fanden sich A. aculeata
Ehbg. mit 203840, quadridentata mit 722180 und foliacea
Ehbg. mit 58240 Individuen. Ausserdem 2567380 Rädertiereier.
Von Daphniden wurden sechs Arten gefunden. Drei von diesen sind
allgemein als pelagisch bekannt, nämlich Leptodora hyalina
Lilj., Hyalodaphnia Kahlbergensis Schödler und Chydorus
sphaericus O. Fr. M., während die drei übrigen bei den
bisherigen Untersuchungen von Süsswasserbecken meist als littorale
Formen nachgewiesen wurden. Daphnia longispina Leyd. ist
jedoch auch von Zacharias[109] in den 6–8 m tiefen
Mansfelder Seen pelagisch gefunden, ebenso Sida crystallina
von Forel[99] in Schweizer Seen. Da die Daphniden vielen
Süsswasserfischen zur Nahrung dienen, so ist ihre Bestimmung von hohem
praktischen Interesse, namentlich da sie sich in Bezug auf ihren Gehalt
an organischer Substanz[S. 287] ähnlich wie die Copepoden verhalten dürften,
für die wir oben 99.4% gefunden haben. Wenn wir in unserem Fange unter
einem Quadratmeter Oberfläche 1826100 Daphniden finden, so können wir
uns leicht ein Bild von ihrer Bedeutung für die Fischzucht machen.
Mit Hilfe der Planktonmethode würde sich ein sehr klares Bild über
den Entwickelungsgang dieser Tiere finden lassen. Bei fortgesetzten
Untersuchungen könnte man das Auftreten der Daphniden im Frühjahr
feststellen, sowie dessen Abhängigkeit von der Wassertemperatur, ferner
würde sich die rapide Zunahme gegen den Sommer hin zeigen, wobei es
von Wichtigkeit wäre, die Zahl der Eier und Embryonen im Brutraume
zu berücksichtigen, so dass sich für jede Art eine Durchschnittszahl
ergeben würde. Zugleich liesse sich das Erscheinen der verschiedenen
Arten in den einzelnen Monaten finden. Im Spätsommer würden dann
die Männchen hinzukommen und schliesslich würde man die Cladoceren
verschwinden sehen, nachdem man die Bildung der Dauereier beobachtet
hätte.
Es würden sich alle diese Verhältnisse zahlenmässig klarlegen lassen
und einen präzisen Ausdruck liefern für die bisherigen Ausdrücke, wie
„im Frühjahr nach Schmelzen des Schnees massenhaft auftretend“.
Die Copepoden, deren Bedeutung wir schon oben gesehen haben,
bilden einen anderen Hauptbestandteil des Plankton. Sie waren unter
1 qm Oberfläche mit 697480 vertreten. Dazu kommen noch die
Larvenformen mit 786520.
In dem Protokoll sind die einzelnen Arten nicht getrennt aufgeführt
worden. Es schien anfangs nicht möglich, während des Zählens die
verschiedenen Spezies auseinanderzuhalten, und mit Hilfe der bisherigen
Diagnosen ist dieses auch nicht auszuführen. Nachdem es sich aber
herausgestellt hat, dass in einem Fange selten mehr als sechs Arten
(bei Zählungen der Copepoden des Plankton im Kieler Hafen) vorhanden
sind, ist die getrennte Zählung versucht worden und hat sich auch
durchführen lassen, da jede Spezies irgend ein bestimmtes Merkmal
besitzt, an dem sie sofort erkannt werden kann. Man kann schliesslich
noch weiter[S. 288] gehen und auch die Geschlechter getrennt zählen. Neben
den ausgewachsenen Copepoden werden dann die Larven berücksichtigt.
Diese nach der Spezies zu zählen wird wohl fürs erste kaum geschehen
können, da die Entwickelungsreihen vom Ei bis zum erwachsenen Tier nur
erst für sehr wenig Formen festgestellt sind, bei genauem Studium und
einiger Ausdauer liesse sich dieses vielleicht auch ausführen. Ebenso
müssten die Eiersäckchen mit der durchschnittlichen Zahl der Eier
berücksichtigt werden.
Nach den erwähnten Untersuchungen von Hensen nähren sich die
Meerescopepoden von Peridineen. Die des Süsswassers müssen aber
andere Nahrung zu sich nehmen, denn nach unserem Fange standen 697480
Copepoden, ohne Larven, nur 122090 Peridineen zur Verfügung, die nach
den Hensenschen Berechnungen nur 10000 Copepoden genügen würden.
Nach Claus[98] leben sie von pflanzlichem und tierischem
Detritus, Vosseler[108] hat dasselbe beobachtet, meint jedoch,
dass noch Infusorien sich beigesellen. Ob dieses aber auch die Nahrung
der pelagischen Copepoden ist, wäre noch experimentell festzustellen.
Von Hydrachniden wurde Nesaea elliptica Kram. in
erwachsenen und jugendlichen Formen gefunden. Ihre immerhin
beträchtliche Zahl von 2400 pro 1 qm ist bemerkenswert. Ob
sie in dem Haushalt der Natur irgend eine Rolle spielen, vermag ich
nicht anzugeben. Eigentümlich ist das pelagische Vorkommen, obgleich
Zacharias[110] in norddeutschen Seen die Milben nur littoral
gefunden hat. Dagegen erwähnt auch Nordquist[106] in seinem
Aufsatz über die pelagische und Tiefseefauna finnischer Seen pelagische
Hydrachniden.
Von Mollusken wurden nur Muschellarven zahlreich gefangen,
Schneckenlarven fehlten. Es fanden sich 40770 auf den Quadratmeter
Oberfläche. Nimmt man für eine Muschel 1 qcm Bodenfläche an,
so würden das auf 1 qm immer nur 10000 ausmachen, der Raum ist
aber viel zu gering bemessen. Von den Larven kann also im günstigsten
Falle nur ¼ am Leben bleiben und diese müssten den Boden dann dicht
überziehen. Das ist aber nicht wahrscheinlich. Ob die Larven zu einer
Spezies gehören, ist bei[S. 289] der Zählung nicht berücksichtigt worden, die
Art selbst zu bestimmen ist bis jetzt auch nicht möglich, würde sich
aber bei speziellen Studien gewiss ausführen lassen. Es würde das ein
Licht auf die Zeit und die Dauer des Schwärmens der Larven werfen.
Es konnte in Vorhergehendem nur meine Aufgabe sein, dem Leser die
Methodik zur quantitativen Bestimmung des Plankton im Süsswasser zu
erklären; etwas Näheres über die Organismen des Plankton zu sagen, war
nach dem einen Süsswasserfange noch nicht möglich. Es wäre zu wünschen,
dass die Hensensche Methode auch in einem grösseren Landsee angewendet
würde, interessante und wichtige Ergebnisse würde sie liefern, wie das
schon der Fall bei ihrer Anwendung im Meere gewesen ist. Endlich möchte
ich nochmals auf die epochemachende Arbeit Hensens hinweisen,
die so viel des interessanten bietet, worauf ich nur hinweisen konnte,
oder das ich wegen Raummangel ganz übergehen musste.
Anmerkung: Der Fang, nach dem dieses Protokoll berechnet ist,
wurde mit einem Planktonnetze von 0.1 qm Öffnung gemacht;
es müssten also, da die Tiefe des Netzzuges 5 m betrug, 0.5
cbm Wasser durch das Netz filtriert sein; in Wahrheit aber nur
0.45 cbm (siehe oben bei Netze). Wollen wir die Anzahl der
Organismen unter einem Quadratmeter Oberfläche kennen, so müssen obige
Zahlen mit 10 multipliziert werden, da die Öffnung des Netzes, also die
Grundfläche der Wassersäule 0.1 qm beträgt.
[95]Asper und Heuscher, Neue Zusammensetzung
der pelagischen Organismen in: Zool. Anzeiger 1886, Bd. 9, S. 448.
[96]Brandt, Über die biologischen Untersuchungen
der Plankton-Expedition in: Verhandl. der Gesellschaft für Erdkunde zu
Berlin 1889, Heft 10.
[97]Brandt, Häckels Ansichten über die
Plankton-Expedition in: Schriften des Naturwissenschaftl. Vereins f.
Schleswig-Holstein, Bd. VIII, Heft 2. Kommissionsverlag von Homann in
Kiel.
[98]Claus, Anatomie und Entwickelung der Copepoden
in: Arch. f. Naturgeschichte 1858, Bd. 1.
[99]Forel, Faunistische Studien in den
Süsswasserseen der Schweiz. Zeitschrift für wissenschaftliche Zoologie
1878, Bd. 30 Suppl.
[100]Häckel, Plankton-Studien. Vergleichende
Untersuchungen über die Bedeutung und Zusammensetzung der pelagischen
Fauna und Flora. Jena 1890.
[S. 293]
[101]Heincke, Die Untersuchungen von Hensen über die
Produktion des Meeres an belebter Substanz in: Mitteilungen der Sektion
für Küsten- und Hochseefischerei, No. 3–5, März bis Mai 1889.
[102]Hensen, Über das Vorkommen und die Menge der
Eier einiger Ostseefische, insbesondere der Scholle, des Flunder
und des Dorsch in: 4. Bericht der Kommission zur wissenschaftlichen
Untersuchung der deutschen Meere zu Kiel für 1877–81. Berlin 1884.
[103]Hensen, Über die Bestimmung des Planktons oder
des im Meere treibenden Materials an Pflanzen und Tieren in: 5. Bericht
der Kommission zur wissenschaftlichen Untersuchung der deutschen Meere
zu Kiel 1887, S. 1–106.
(Oben kurz als „Planktonwerk“ zitiert.)
[104]Hensen, Einige Ergebnisse der Plankton-Expedition
der Humboldt-Stiftung in: Sitzungsberichte der Königlich Preussischen
Akademie der Wissenschaften zu Berlin.
Sitzung d. physikal.-mathemat. Klasse vom 13. März 1890.
[105]Hensen, Das Plankton der östlichen Ostsee und des
Stettiner Haffs in: 6. Bericht der Kommission zur wissenschaftlichen
Untersuchung der deutschen Meere in Kiel 1890.
105a: Hensen, Die Plankton-Expedition und Häckels Darwinismus.
Kiel, Lipsius und Tischer 1891.
[106]Nordquist, Über die pelagische und Tiefsee-Fauna
finnischer Seen in: Zoolog. Anzeiger 1887, Bd. 10, S. 339 und 358.
[107]Seligo, Hydrobiologische Untersuchungen in:
Schriften der Naturforschenden Gesellschaft zu Danzig, N. F., Bd. VII,
Heft 3. 1890.
[S. 294]
[108]Vosseler, Die freilebenden Copepoden
Württembergs. Stuttgart 1886.
[109]Zacharias, Fauna des süssen und salzigen Sees bei
Halle a. d. S. in: Zeitschrift für wissenschaftliche Zoologie 1888, Bd.
46.
[110]Zacharias, Zur Kenntnis der pelagischen
und littoralen Fauna norddeutscher Seen in: Zeitschrift für
wissenschaftliche Zoologie 1887, Bd. 45, S. 255.
[S. 295]
Die Fauna des Süsswassers in ihren Beziehungen zu der des Meeres.
Von Dr. Otto Zacharias in Plön (Holstein).
[S. 297]
Unter den vielen bemerkenswerten Thatsachen, welche die in neuerer
Zeit mit so grossem Eifer betriebene Erforschung der Binnenseen zu
Tage gefördert hat, ist das unzweifelhafte Vorkommen mariner Tiere
im süssen Wasser eine der interessantesten. Besonders waren es
italienische Seen, in denen man zuerst jene überraschende
Entdeckung machte. So beherbergt z. B. der weitab vom Meere
gelegene Gardasee drei Fischspezies, welche marinen Gattungen
angehören: 1) einen heringsartigen Fisch (Alosa finta), zu
dessen nächsten Verwandten die sog. „Maifische“ zählen; 2) eine
Meergrundel (Gobius) und 3) einen Schleimfisch (Blennius
vulgaris). Ausserdem kommt in demselben Wasserbecken ein Krebs
(Palaemonetes) vor, von dem der bekannte Berliner Zoologe E.
v. Martens sagt: „Er steht unserer Ostseegarneele (Palaemon
squilla) nahe, unterscheidet sich aber von ihr durch geringere
Grösse und durch die Gestalt des Schnabels“. Auch in den Kraterseen
von Albano und Nemi kommt dieser kleine Krebs zugleich mit der schon
erwähnten Blennius-Art vor.
Unter den schweizerischen Seen ist es der von Genf (Lac
Léman), welcher in den Muschelkrebschen Acanthopus resistans
(= Cytheridea lacustris Sars) und Acanthopus elongatus
(= Limnicythere relicta Lillj.) zwei Tierformen enthält, die
der marinen Gruppe der Cytheriden sehr nahe verwandt sind und deren
Anwesenheit im Süsswasser uns daher überrascht.
[S. 298]
Die skandinavisch-finnischen Seebecken besitzen ebenfalls
in ihrer Fauna eine Anzahl von Krustern (Mysis relicta,
Pontoporeia affinis, Idotea entomon u. s. w.), welche
Vertreter von im Meere lebenden Gattungen sind.
Ganz ähnliche Thatsachen liegen für die grossen kanadischen Seen
in Nordamerika vor. Wir begegnen dort den nämlichen Krebsen wie in
Skandinavien und ausserdem noch zwei Fischen (Triglops-Arten),
welche weit mehr die Charaktere von Meeres- als diejenigen von
Süsswasserfischen besitzen.
Diese Befunde, welche sich aus anderen Seengebieten leicht vermehren
liessen, haben in der Folge dazu geführt, die Theorie der sogenannten
„Reliktenseen“ aufzustellen. Darunter versteht man solche Seen,
welche für die Reste einer ehemaligen Meeresbedeckung angesprochen
werden. Man fühlte sich befugt, diesen Ursprung hauptsächlich
denjenigen Wasserbecken beizumessen, in denen man die oben angeführten
Krustaceen und Fische (bezw. andere zu marinen Gattungen gehörige
Tiere) vorgefunden hatte. Ausser Stande oder nicht daran gewöhnt, das
Vorhandensein solcher Fremdlinge auf eine andere Weise zu erklären, als
dadurch, dass dieselben Überbleibsel (Relikte) einer vormaligen, an
Ort und Stelle heimisch gewesenen Meeresfauna seien, zog man hieraus
den weiteren Schluss, dass in einer nicht sehr weit zurückliegenden
Periode der Erdgeschichte eine mehrmalige Andersverteilung von Land und
Wasser stattgefunden haben müsse, wobei Einsenkungen der Festländer mit
Meerwasser angefüllt oder Fjorde direkt vom Meere abgesperrt worden
wären, so dass in den so entstandenen Seebecken gewisse marine Spezies
zurückblieben, und dem durch Regengüsse sich immer mehr aussüssenden
Wasser allmählich angepasst wurden.
Diese Ansicht war sehr lange Zeit in Geltung, und zum Teil ist sie
es auch noch heute. Aber bei näherer Prüfung dieses „faunistischen
Arguments“ für den marinen Ursprung einer Anzahl von Binnenseen
zeigt es sich, dass dasselbe weder vor der geologischen noch vor der
zoologischen Kritik Stand hält.
[S. 299]
In letzterer Hinsicht hat es sich nämlich herausgestellt, dass es
eine grosse Anzahl von Tieren giebt, welche ebenso gut im süssen
wie im Brack- oder Salzwasser leben können. Zunächst ist hierbei an
die allbekannten Wanderfische (Lachs, Aal, Scholle u. s. w.)
zu erinnern, die sich gleich gut im Meere wie in den Flussläufen
aufzuhalten vermögen. Dann bieten aber auch die Mollusken
bemerkenswerte Beispiele dafür dar, dass manche Arten einen recht
erheblichen Wechsel des Salzgehalts im Wasser vertragen können. So
lebt eine kleine Meerschnecke (Hydrobia ulvae) in dem beinahe
ganz süssen Wasser der inneren Ostsee; sie ist aber ebenso zahlreich
in der Nordsee zu finden. Neritina fluviatilis, eine Bewohnerin
grosser Flüsse und Binnenseen, wurde 1887 von Prof. M. Braun
auch in der Wismarer Bucht angetroffen. Noch anpassungsfähiger ist
aber die weitverbreitete Wandermuschel (Dreyssena polymorpha).
Ursprünglich nur in Südosteuropa, namentlich im kaspischen Meere
vorkommend, ist sie durch den Verkehr in den Schiffahrtskanälen
seit 1825 von einem Flusssystem zum anderen (über Ostpreussen) nach
Norddeutschland eingewandert, und hat sich von da flussaufwärts
verbreitet, so dass sie nunmehr in der Saale bei Halle, im Neckar bei
Heilbronn und im Rhein bei Basel angetroffen wird. Diese Verschleppung
geschieht sehr leicht, weil sich die Muschel mittels ihrer Byssusfäden
(vergl. diesen Band S. 126 u. 130) an Flosse und Lastkähne anheftet und
auf solche Art als blinder Passagier weite und bequeme Reisen machen
kann. Sie vermag im Brackwasser ebenso gut auszudauern wie in rein
süssen Gewässern.
Was die Krebstiere anlangt, so sind dieselben der Mehrzahl nach
allerdings streng in Süss- und Salzwasserbewohner geschieden, aber es
giebt unter letzteren auch Formen, wie z. B. Mysis vulgaris,
die in fast vollkommen süssem Wasser zu existieren vermögen. Auf der
Westerplatte bei Danzig fand ich diese eigentlich dem Meere angehörige
Art in einem nur Spuren von Salz enthaltenden Tümpel.
Parasitische Kruster, welche auf Aalen, Lachsen und Stören schmarotzen,
sind gegen den Wechsel von Meer- und Flusswasser[S. 300] ganz unempfindlich.
Von den spaltfüssigen Krebsen (vergl. Band I dieses Werkes S. 349)
soll Diaptomus castor, der in kleinen Lachen und Teichen des
Binnenlandes lebt, auch an der Ostseeküste vorkommen.
Von den Hohltieren (Cölenteraten) vermag der See-Keulenträger
(Cordylophora lacustris) ebenso gut im Brackwasser wie im
gewöhnlichen Flusswasser sein Leben zu fristen. Unser kleiner
Süsswasserpolyp (Hydra) stirbt dagegen sehr bald, auch wenn er
nur in ganz schwaches Salzwasser gebracht wird.
Medusen als Süsswasserbewohner waren bis in die neueste Zeit
herein gänzlich unbekannt. Da entdeckte Dr. J. Kennel in
vollkommen ausgesüssten Strandseen auf der Insel Trinidad (1882) eine
winzige Spezies dieser echten Meerestiere. Mit Recht hebt anlässlich
dieses wichtigen Fundes der genannte Forscher hervor, dass dem
Vorkommen einer Qualle im Süsswasser gegenüber nicht einzusehen sei,
weshalb irgend einem anderen Meeresbewohner die Möglichkeit eines
Wechsels seines Lebenselements, bezw. der Übergang aus dem Salzwasser
in das Süsswasser verschlossen sein sollte.
Die plötzliche Versetzung von Meerestieren in gewöhnliches
Brunnen- oder Flusswasser erweist sich nach den bisherigen Erfahrungen
für die meisten als todbringend. Nicht so aber — wie die Experimente
des Franzosen Beudant zeigen — eine allmählich vorgenommene
Verdünnung des Meerwassers mit gewöhnlichem Wasser. Auf die letztere
Weise gelang es, zahlreiche Arten von marinen Weichtieren an fast
vollständig ausgesüsstes Seewasser zu gewöhnen. Allerdings wird die
Beweiskraft dieser Versuche dadurch geschmälert, dass bei denselben die
Frage unberücksichtigt geblieben ist, ob die betreffenden Mollusken,
welche für sich selbst den Aussüssungsprozess gut überstanden, nun auch
fähig gewesen wären, sich in dem neuen Medium fortzupflanzen. Von der
Auster wissen wir z. B., dass dieselbe in erwachsenen Individuen, ohne
Schaden zu erleiden, einen Aufenthalt im süssen Wasser verträgt. Aber
trotzdem wollen keine Austernbänke in der salzarmen Ostsee fortkommen,
woraus zu schliessen sein dürfte, dass es die junge Brut[S. 301] ist, die
einen stärkeren Salzgehalt zu ihrem Gedeihen nötig hat, als er in jenem
grossen Binnenmeere zu finden ist.
Dem gegenüber kann nun freilich der Umstand angeführt werden, dass wir
auf Grund geologischer Erwägungen die heutigen Süsswassermollusken von
marinen Voreltern herleiten müssen, welche nach dem Auftauchen der
Kontinente aus dem Urmeere in die Flussläufe einwanderten und hier sich
veränderten Lebensbedingungen anbequemten. Eine andere Entstehungsweise
für die gegenwärtigen Bewohner unserer süssen Gewässer vermögen wir
überhaupt nicht anzunehmen, und eben darum müssen auch die Mollusken
unserer binnenländischen Wasserwelt als die Nachkommen von Schnecken
und Muscheln des Meeres betrachtet werden.
Ein Beispiel dafür, wie dies einstmals vor sich gegangen sein mag,
haben wir an den Verhältnissen, welche der Ortoire-Fluss im Süden der
Insel Trinidad noch heute darbietet. Hier wird, nach Kennels
Beobachtungen[111], die Einwanderung von Meerestieren durch die
Thatsache begünstigt, dass die schwache Strömung täglich zwei Mal durch
die Flutwelle zum Stehen gebracht wird, und dass dann der Übergang
aus dem Meerwasser in das brackische und süsse ein ausserordentlich
allmählicher ist. In bedeutender Höhe des Flusslaufes (12 engl.
Meilen von dessen Mündung entfernt) und weit oberhalb der Grenze des
Brackwassers fand Kennel förmliche Anhäufungen von Tieren,
denen man sonst nur im Meere begegnet; so namentlich mächtige Bänke
von einer Miesmuschelart, frei schwimmende marine Borstenwürmer, und
einige Spezies von Seekrebsen — also eine unleugbare Meeresfauna im
süssen Wasser. Es besteht natürlich nicht der Schatten eines Zweifels
darüber, dass alle jene Tiere zuerst mit der Flut in den Ortoire-Fluss
hineingeraten sind, und sich hier — weil sie den allmählichen Wechsel
im Salzgehalt auszuhalten vermochten — dauernd angesiedelt haben. In
ganz analoger Weise haben wir uns auch die erstmalige Einwanderung
von Meeresbewohnern in das süsse Wasser der Flüsse und der damit in
Verbindung stehenden Seen geschehen zu denken.
[S. 302]
Prof. Milnes Marshall hat unlängst[CXVI] geltend gemacht, dass
viele das Meer bewohnende wirbellose Tiere schon deshalb unfähig wären
in das Süsswasser einzuwandern, weil sie das Ei als sehr kleine,
bewimperte Larven verlassen, welche ganz ausser Stande seien, gegen
irgend welche Strömung anzukämpfen. Es könnten daher — nach seiner
Ansicht — nur solche Formen, welche sich von dem freischwimmenden
Larvenzustand emanzipiert haben und welche das Ei in ansehnlicher
Grösse und Stärke verlassen, dem Leben im Süsswasser angepasst
werden. Diese Erklärung mag im allgemeinen wohl das Richtige treffen;
indessen zeigt uns die neuerdings von E. Korschelt[120] zum
Gegenstande einer speziellen Untersuchung gemachte Entwickelung von
Dreyssena polymorpha, dass diese ursprünglich marine Muschel das
freischwimmende Larvenstadium (Trochophora) beibehalten hat.
Dieser Nachweis ist von hohem Interesse. Die Larven sind sehr klein und
sie machen beim ersten Anblick den Eindruck von Infusorien. Wenn sich
das Mundsegel (Velum) in stark wimpernder Bewegung befindet,
könnte man sie auch für kleine Rädertiere halten. Die Schwärmzeit
dieser winzigen Wesen beläuft sich auf etwa acht Tage. Korschelt
konstatierte, dass sie im Tegeler See bei Berlin ungefähr Ende Juni
erscheinen und die oberen Wasserschichten in Menge bevölkern. Im
Grossen Plöner See waren sie nach meiner eigenen Wahrnehmung in
diesem Sommer (1891) während der Zeit vom 5. bis 10. Juli ebenfalls
massenhaft vorhanden, und zwar durchweg im freien Wasser des ganzen,
mächtigen Sees bis zu 2 m Tiefe. Prof. F. Blochmann
hat auch im Warnowflusse (bei Rostock) Dreyssena-Larven angetroffen,
und hiermit wird eine Erklärung für die schnelle Verbreitung dieser
Muschel gegeben. Denn offenbar können jene winzigen Wimperlarven
innerhalb der achttägigen Schwärmzeit ausserordentlich weit von der
Strömung fortgetragen werden. Das erstaunlich üppige Gedeihen der
Dreyssena in manchen Gewässern zeigt übrigens, dass sich die
zarten Larven derselben in unseren Flüssen und Binnenseen sehr wohl
befinden müssen.
[CXVI] „Über Rekapitulation in der Embryologie.“ Rede zur
Eröffnung der biologischen Sektion in der British Association
(Jahresversammlung) zu Leeds. 1890.
[S. 303]
Handelt es sich um die Erklärung des Vorhandenseins von Tieren marinen
Charakters in solchen Seebecken, die heutzutage nicht mehr mit einem
Flusssystem zusammenhängen, sondern eine völlig isolierte Lage haben
(wie z. B. zahlreiche schwedische und finnische Seen), so hat man sich
vor Augen zu halten, dass die hydrographischen Verhältnisse Nord- und
Mitteleuropas am Schlusse der Eiszeit ganz andere waren, als sie jetzt
sind.
Die Flussläufe weiter Länderstrecken sind gegenwärtig nur schwächliche
Abbilder früher ungleich wasserreicherer und breiterer Stromrinnen.
Hindernisse, welche sich heute in Gestalt von Wasserfällen und
Stromschnellen der Tiereinwanderung entgegenstellen, waren ehedem
überhaupt nicht oder doch nur in geringerem Massstabe vorhanden.
Zwischen jetzt getrennten Flusssystemen bestanden Verbindungskanäle;
seeartige Erweiterungen der Flüsse existierten in weit grösserer
Anzahl als unter den heutigen erdgeschichtlichen Verhältnissen und
bildeten Etappen für die Wanderung der im Süsswasser sich ansiedelnden
Meeresfauna. In den breit ausgewaschenen Thälern des baltischen
Landrückens erkennen wir noch deutlich die alten Verbindungen der jetzt
getrennten, ehemals aber zu einem gewaltigen Urstromsystem vereinigten
ostdeutschen Flüsse, durch welche die Gewässer der Weichsel, der Oder
und der Elbe vereinigt zur Nordsee abflossen.
Es ist ein Verdienst des Greifswalder Professors der Erdkunde Rud.
Credner, diese Momente zuerst nachdrücklich betont und zum
Gegenstande einer umfangreichen Monographie gemacht zu haben[112],
aus welcher der Zoolog die Mahnung schöpfen kann, dass er auf blosse
Tierfunde hin nicht berechtigt ist, irgend einen See für den Rest
einer vorzeitlichen Meeresbedeckung anzusehen. Nur wenn aus den
geologischen Verhältnissen zugleich mit hervorgeht, dass wir
es wirklich mit einem abgesperrten Fjord oder einer einstmaligen
Meeresbucht zu thun haben, liegt Sicherheit dafür vor, dass wir in der
anwesenden marinen Fauna keine späteren Einwanderer, sondern wirklich
von der Meeresflut selbst hierher getragene und nach dem Rückzuge des
Wassers in loco verbliebene Lebewesen vor uns haben. Ein See, der die
vollgültige[S. 304] geologische Legitimation für seinen marinen Ursprung
aufzuweisen vermag, ist dann ein echter Reliktensee im Gegensatz zu den
vielen anderen ebenso bezeichneten Wasseransammlungen, die zu einer
marinen Tierwelt nur dadurch gekommen sind, dass an das salzfreie
Element bereits gewöhnte Meerbewohner aktiv oder passiv in sie
einwanderten.
Mit dem Ausdruck „passive Wanderung“ benennt man die verschiedenen
Arten von Verschleppung kleinerer Tiere, welche durch grössere
Organismen, die dabei als Transporteure thätig sind, bewirkt wird. So
wird z. B. ein asselartiges Krebstier (Idotea entomon), welches
als ein Hauptrepräsentant der Reliktenfauna zahlreicher skandinavischer
Seen aufgeführt wird, durch Störe, an die es sich anheftet, in den
Flüssen Sibiriens weite Strecken stromaufwärts gebracht. In ähnlicher
Weise sind auch Sturmwinde und wandernde Sumpfvögel wirksam, insofern
sie kleinere Tiere des Meeres oder deren Eier und Jugendformen (mittels
Transports durch die Luft) dem Süsswasser oder zunächst salzärmeren und
der Aussüssung unterworfenen Gewässern zuführen.
Ich war in hohem Grade verwundert, als ich bei einer im Sommer 1884
vorgenommenen Durchforschung der bekannten Hochseen des Riesengebirges
in jedem dieser beiden isoliert gelegenen Wasserbecken eine
Monotus-Art antraf. Hierunter sind Strudelwürmer zu verstehen,
die einer im Meere lebenden Gattung angehören. Überraschender Weise
ist dieser Riesengebirgs-Monotus nahe verwandt mit einer im
Sunde zahlreich vorkömmlichen Spezies, welche in der Litteratur unter
dem Namen Monocelis spinosa Jensen aufgeführt wird. Später
entdeckte man die nämliche Spezies, für die ich die Bezeichnung
Monotus lacustris in Vorschlag gebracht habe, auch im Peipus
(Russland) und in mehreren schweizerischen Seen[113]. Charakteristisch
für die Biologie dieses Tierchens ist der Umstand, dass sein Vorkommen
auf grosse und kühltemperierte Wasserbecken beschränkt erscheint. In
kleineren Teichen oder Tümpeln, die sich leicht erwärmen, ist es bis
jetzt nicht aufgefunden worden.
[S. 305]
Diese Vorliebe des Süsswasser-Monotus für kalte Seen, und die
bereits hervorgehobene Eigentümlichkeit, dass derselbe mit einer
marinen Spezies des Nordens in nächster Verwandtschaftsbeziehung
steht, lassen die Hypothese nicht ungerechtfertigt erscheinen,
dass wir es in dem eigentümlichen, kälteliebenden Tiere mit einem
überlebenden Bewohner jener zahlreichen Schmelzwasserseen zu thun
haben, die sich am Ausgange der Eiszeit bildeten, und die sowohl
unter sich als auch mit dem nördlichen Meere durch natürliche
Zwischenkanäle in Verbindung standen. In ein derartig zusammenhängendes
System von grösseren und kleineren Wasseransammlungen konnte eine
anpassungsfähige Turbellarienform des Meeres leicht einwandern und
eine grosse Verbreitung erlangen. Sie vermochte aber andernteils,
wenn die einzelnen Seen aus Mangel an Wasserzufuhr verdunsteten, nur
an solchen Örtlichkeiten auszudauern, welche annähernd die nämlichen
Lebensbedingungen darboten, wie die von den Schmelzwässern der
nordischen Eisströme gebildeten Gletscherseen. Auf solche Art erklärt
sich auch die merkwürdige sporadische Verbreitung des Monotus
lacustris am ungezwungensten, und es ist damit gleichzeitig
motiviert, dass ich dieses Tier in einer früheren Publikation als eine
fremdartige Erscheinung in unserer Süsswasserfauna bezeichnet habe.
Natürlich halte ich diese Erklärung zunächst für hypothetisch, aber sie
ist, im Anschluss an die oben mitgeteilten anderweitigen Thatsachen,
für den Augenblick entschieden annehmbar[114].
Ein namhafter italienischer Naturforscher, Prof. Pietro Pavesi
in Pavia, tritt mit grossem Eifer auch für den marinen Ursprung
der sogenannten „pelagischen“ Fauna unserer Landseen ein, insofern
er die beiden Hauptvertreter dieser vorwiegend aus kleinen Krebsen
bestehenden Tierschwärme (Leptodora und Bythotrephes)
für Meeresformen erklärt, welche die Gewohnheit, im Süsswasser zu
leben, angenommen haben. Da sich für Bythotrephes (Fig. 76 des
ersten Bandes) in einem kleinen ozeanischen Krebse (Podon) ein
in morphologischer Beziehung verwandtes Geschöpf nachweisen lässt,
und da Leptodora (Fig. 75 in Band I) ihrer Organisation nach
völlig isoliert unter den Süsswasserkrustern dasteht, so mag Pavesis[S. 306]
Ansicht für diese beiden pelagischen Spezies zu Recht bestehen. Weshalb
aber die anderen Bewohner des freien Wassers unserer Binnenseen,
deren nahe Verwandtschaft mit den Uferspezies sofort in die Augen
fällt, gleichfalls mariner Herkunft sein sollen, dies ist schwer
ersichtlich. Noch unbegreiflicher aber ist Pavesis Schlussfolgerung,
dass diejenigen Seen, in denen sich eine pelagische Fauna konstatieren
lässt, immer wirkliche Reliktenseen, d. h. Überbleibsel einer
vormaligen Meeresbedeckung, sein sollen. Wäre diese Schlussweise
zulässig, so müsste auch der Bremer Stadtgraben (in welchem 1838 die
vielberufene Leptodora hyalina von Dr. Focke entdeckt
wurde) mit zu den echten Reliktenseen gezählt werden, was wohl Niemand
im Ernste verlangen wird. Und ebenso hätte man das Recht, manche mit
Wasser angefüllte Ziegelei-Ausschachtungen, weil in denselben einige
pelagische Rädertierspezies vorkommen, für abgesperrte Fjorde eines
vorzeitlichen Ozeans zu halten. Dies wäre aber einfach absurd. Prof.
Pavesi lässt aus Liebe zu seiner Theorie der „Fauna relegata“
ganz ausser Acht, dass die kleinen Süsswassertiere (oder deren Eier)
durch wandernde Sumpfvögel leicht von See zu See verschleppt werden,
sodass sie, wie durch Imhof erwiesen ist, selbst bis in die
hochgelegensten Alpenseen hinauf verbreitet sind. Auf dem Wege solcher
passiven Wanderungen — wie man es nennt — werden jene
Organismen auch in die zahllosen binnenländischen Tümpel und Seen
übergeführt, ohne dass letztere zu irgend einer Zeit mit dem Meere in
direkter Verbindung gestanden haben.
Positive Beweise dafür, dass Schwimmvögel zur Verbreitung von kleinen
Wasserorganismen beitragen können, liegen mehrfach vor. F. A.
Forel wurde 1876 durch eine Beobachtung von Alois Humbert
in Genf zu der Überzeugung gebracht, dass mindestens die kleinen
Kruster auf solche Weise gelegentlich verpflanzt werden. Humbert
hatte nämlich Wintereier von Cladoceren (Wasserflöhen) dem
Gefieder von wilden Enten und Tauchern anhängend gefunden. Diese
einzige Wahrnehmung warf mit einem Male Licht auf das sonst rätselhafte
Vorhandensein von zahlreichen Spezies niederer Organismen in völlig
isolierten Wasseransammlungen. In welcher[S. 307] Menge solche Wintereier am
Schlusse des Sommers vorkömmlich zu sein pflegen, darauf wirft eine
Schilderung Licht, die von Prof. G. Asper und J. Heuscher
seinerzeit gegeben worden ist[115]. „Als wir“ — so heisst es in
derselben — „am 27. Juli 1886 am oberen Ende des Fählensees (Schweiz)
Steine umwenden wollten, um die darunter sich aufhaltenden Tiere zu
sammeln, trafen wir den ganzen Ufersaum etwa einen halben Meter breit
mit einer dunklen Schicht bedeckt. Die ins Wasser eingetauchte Hand
wurde beim Herausziehen schwarz durch eine Unzahl kleiner Körperchen,
die hartnäckig anhafteten. Es waren die Ephippien (Eiersättel) einer
Daphnie, sehr wahrscheinlich solche von D. longispina. Sie waren
im Trockenen kaum von der Haut wegzubringen, lösten sich dagegen sehr
leicht ab, wenn man die Hand wieder ins Wasser tauchte. Die Körperchen
zeigten keine Adhäsion fürs Wasser, sie blieben trocken wie die Federn
der Schwimmvögel und flottierten an der Oberfläche. Der scharf über den
See streichende Wind hatte wohl einen bedeutenden Teil der zerstreuten
Eier an das obere Ufer getrieben. Die ungemein weite Verbreitung der
genannten Spezies kann uns hiernach nicht in Erstaunen setzen. Denn wie
viele Tausende von Eiern bleiben an den Füssen der Rinder hängen, die
hier und dann anderwärts zur Tränke gehen; wie leicht kleben sie an der
Brust jedes Vogels fest, der ins Wasser geht, oder auch an der Gemse,
die hier ihren Durst stillt.“
Was die Cyclops- oder Diaptomus-Arten betrifft (vergl.
das IX. Kapitel des ersten Bandes), die fast niemals in einer
grösseren Wasserlache fehlen, so scheint es, dass dieselben eine
nahezu vollständige Austrocknung vertragen können. J. Vosseler
bemerkt darüber in einer älteren Publikation[116] folgendes: „Mehrere
Male waren einige meiner Fundorte trocken gelegt und bis zu einer
Tiefe von 1–1½ Fuss kein feuchtes Erdreich mehr zu finden. Kaum stand
jedoch über dem trockenen Schlamm etwas Wasser, so war dies alsbald
wieder von Cyclopiden belebt“. Tiere von solcher Lebenszähigkeit werden
also sicherlich auch, wenn sie auf das Gefieder eines Vogels geraten,
einen weiten Transport durch die Luft auszuhalten vermögen. Übrigens
würden die den weiblichen[S. 308] Spaltfusskrebsen anhängenden Eier auch dann
entwickelungsfähig bleiben, wenn das Muttertier unterwegs zu Grunde
gehen sollte.
Neben den Vögeln spielen aber, wie Dr. W. Migula gezeigt hat,
auch die Wasserkäfer eine bedeutsame Rolle bei der Verbreitung der
kleinen und zum Teil mikroskopischen Süsswasserorganismen. Der Genannte
fand nämlich, dass Eudorina elegans, Pandorina morum,
Scenedesmus obtusus und sonstige Algen durch derartige
Käfer verschleppt und in andere Wasserbecken verpflanzt werden.
Migula fasst das Ergebnis seiner Untersuchungen in folgendem
Passus zusammen[117]: „Da die Wasserkäfer (besonders des Nachts)
ihren Aufenthalt häufig wechseln und nachweisbar oft weit entfernte
Gewässer aufsuchen, so vermitteln sie gewiss in allen jenen Fällen
die Verbreitung der Algen, wo es sich um kleine Lachen und Tümpel
handelt, die wohl für Wasserkäfer, aber nicht für Wasservögel von
Interesse sind. Das konstante Vorkommen von Algen an den Körperteilen
von Wasserkäfern lässt sogar darauf schliessen, dass diesen bei
dem Transport von Mitgliedern der niederen Flora eine grössere
Rolle zukommt, als den Wasservögeln oder der strömenden Luft. In
Wirklichkeit verhält es sich wahrscheinlich so, dass die Luft kleinste
und der Austrocknung widerstehende Formen verbreitet, Wasservögel den
Transport nach weit entfernten Gegenden vermitteln und Wasserkäfer
in ausgedehnter Weise für die Ausbreitung einer Spezies innerhalb
enger räumlicher Grenzen thätig sind“. Dass mit den Algen zugleich
auch eingekapselte Protozoen, Eier von kleinen Würmern u. s. w.
transportiert werden können, wird Niemand als etwas Unwahrscheinliches
betrachten.
Auf die ebenfalls weit verbreiteten Wassermilben (Hydrachniden)
scheint der Modus einer Überführung derselben von einem Gewässer
zum andern überhaupt nicht anwendbar zu sein, weil diese Tiere für
das Trockenwerden sehr empfindlich sind und ausserhalb des Wassers
schnell zu Grunde gehen. Um so dringlicher erhebt sich hiernach die
Frage, auf welche Weise diese spinnenartigen Wesen in die grossen
und kleinen Seebecken hineingelangen, wo wir sie vorfinden. Über
diesen Punkt hat uns unlängst der französische[S. 309] Naturforscher Th.
Barrois[118] aufgeklärt. Derselbe entdeckte nämlich, dass es nicht
die erwachsenen Individuen, sondern die durch einen dicken Chitinpanzer
geschützten Puppen der Wassermilben seien, welche der passiven
Wanderung unterworfen werden, insofern sich dieselben an den Leibern
verschiedener Wasserwanzen (Nepa, Notonecta) festheften
und so von diesen fliegenden Insekten auf weite Entfernungen hin
transportiert werden. Barrois zeigte durch das Experiment, dass
Wasserwanzen viele Stunden auf dem Trockenen ausdauern können, ohne
dass die Entwickelungsfähigkeit jener Puppen darunter leidet. Letztere
werden also auch dann ungefährdet bleiben, wenn die Wasserwanzen, ihrer
Gewohnheit folgend, während der Nacht von einem Teiche zum anderen
fliegen. Auf solche Art gelangen nun zahlreiche zum Ausschlüpfen reife
Larven von Hydrachniden an weit entfernte Wohnplätze und verbleiben
dort für immer, nachdem sie die Puppenhülle gesprengt und verlassen
haben. Ihr weiteres Wachstum vollzieht sich in dem einen Gewässer so
gut wie in dem anderen, und daher kommt es, dass wir selten in einem
Graben oder Tümpel vergeblich nach Hydrachniden suchen.
Andere überall vorkommende Tiere, wie z. B. die kleinen
Süsswasser-Oligochäten (Nais, Chaetogaster
etc.), sind mit zahlreichen Büscheln von Hakenborsten ausgerüstet,
und dies führt auf den Gedanken, dass sie durch diese Borsten in
der Vornahme passiver Wanderungen stark begünstigt werden. Manche
Turbellarien besitzen, wie wir früher sahen, sogenannte
„Klebzellen“ in der Nähe des hinteren Körperendes, und höchst
wahrscheinlich dienen dieselben gelegentlich ebenfalls dazu, ihren
Besitzern eine Luftreise zu ermöglichen. Jene Zellen sind einer so
energischen Thätigkeit fähig, dass man die betreffenden Strudelwürmer
oft eher zerreissen, als von ihrer Befestigungsstelle loslösen kann.
Eine ganz vorzügliche Geeignetheit zur Ausführung passiver Wanderungen
müssen wir übrigens auch bei vielen Spezies von Wasserschnecken
und Muscheln voraussetzen, denn diese Mollusken sind selbst
noch in manchen Gebirgsseen anzutreffen.[S. 310] Fand doch A. Brandt
selbst in dem 1904 m hohen Goktschai (Armenien) Limnäen, Planorbis
carinatus und Pisidien vor. Das Pisidium fossarinum
konstatierte A. Wierzejsky in 21 Seen der Hohen Tatra, und einer
Notiz Imhofs zufolge ist die nämliche Muschel sogar noch auf dem
Splügen zu finden.
Nach einer wertvollen Beobachtung Darwins scheint hauptsächlich
den ganz jungen Schnecken das Vermögen zu weiten Wanderungen
beizuwohnen, wie sich aus folgender Stelle des Kapitels über
geographische Verbreitung in der „Entstehung der Arten“ ergiebt.
Darwin sagt dort: „Wenn eine Ente sich plötzlich aus einem mit
Wasserlinsen bedeckten Teiche erhebt, so bleiben oft einige dieser
kleinen Pflanzen auf ihrem Rücken hängen, und es ist mir vorgekommen,
dass, wenn ich einige Wasserlinsen aus einem Aquarium ins andere
versetzte, ich ganz absichtslos das letztere mit Süsswassermollusken
des ersteren bevölkerte. Doch ist ein anderer Umstand vielleicht noch
wirksamer. Ich hängte einen Entenfuss in einem Aquarium auf, wo viele
Eier von Wasserschnecken auszukriechen im Begriffe waren, und fand,
dass bald eine grosse Menge der äusserst kleinen Schnecken an dem
Fusse umherkrochen und sich so fest an demselben anklebten, dass sie
kaum abgeschabt werden konnten, obwohl sie in einem etwas vorgerückten
Alter freiwillig davon abgefallen wären. Diese frisch ausgeschlüpften
Mollusken lebten an dem Entenfusse in feuchter Luft 12–20 Stunden lang,
und während dieser Zeit kann eine Ente oder ein Reiher mindestens
600–700 englische Meilen weit fliegen, um sich dann in einem Sumpfe
oder Bache niederzulassen“.
Im Anschluss an diese Mitteilung berichtet Darwin noch über den
merkwürdigen Fall, wo ein Wasserkäfer (Dytiscus) mit einer ihm
anhaftenden Napfschnecke (Ancylus) gefangen wurde. Über andere,
nicht weniger interessante Vorkommnisse, welche speziell die passiven
Wanderungen von Muscheln betreffen, berichtet Charles Darwin in
einem Aufsatze der „Nature“ vom Jahre 1882.
Die kosmopolitische Verbreitung vieler Protozoen, hauptsächlich
diejenige der Difflugien und Arcellen (die fast nirgends
fehlen, wo etwas Feuchtigkeit vorhanden ist), geschieht vorwiegend
durch den[S. 311] Wind, wenn er über die Böden ausgetrockneter Tümpel
hinfegt. Doch wird es auch vorkommen, dass manche Spezies mit den
Schlammklümpchen, die an den Schwimmfüssen wilder Enten u. s. w. hängen
bleiben, einen Ortswechsel erfahren. Besondere Anpassungen scheinen bei
diesen niederen Organismen sehr selten nachweisbar zu sein.
Doch ist mir gelegentlich eine Difflugia im Riesengebirge zu
Gesicht gekommen, welche im Umkreise der weiten Wölbung ihres Gehäuses
acht stachelartige Fortsätze besitzt, von denen jeder noch eine
gekrümmte Spitze trägt, die sich wie eine winzige Kralle ausnimmt.
Jedes Exemplar der von mir in nassen Moospolstern (Sphagnum)
gesammelten Difflugien (siehe Band I, Fig. 16) zeigt konstant die
geschilderte Eigentümlichkeit, während sie im übrigen fast ganz mit
der von Leidy[119] beschriebenen Difflugia corona
übereinstimmt. Ich erblicke in der Riesengebirgs-Difflugia ein
interessantes Beispiel dafür, dass auch bei Protozoen gelegentlich
spezialisierte Haftorgane zur Ausbildung gelangen, die offenbar dazu
dienen können, passive Wanderungen zu erleichtern. —
Nach allen vorausgegangenen Ausführungen und Erörterungen bietet also
das Auftreten von marinen Tierformen im Süsswasser gar keine Gewähr
dafür, dass diese Wesen an Ort und Stelle selbst den Anpassungsprozess
von dem einen Medium ans andere vollzogen haben. Vielmehr ist es, wie
einige der mitgeteilten Thatsachen zeigen, in den weitaus meisten
Fällen als das Wahrscheinlichere zu betrachten, dass jene Spezies von
marinem Habitus durch aktive oder passive Einwanderung in die jetzt
von ihnen bewohnten Binnenseen gelangt sind. Nur wenn in überzeugender
Weise durch den geologischen Befund erhärtet werden kann, dass
die bezüglichen Seen wirkliche (aber im Laufe der Zeit ausgesüsste)
Meeresabschnitte sind, kann von der Existenz einer eigentlichen
Reliktenfauna in ihnen die Rede sein. Von den mehr als hundert Seen,
in denen Tiere von marinem Charakter gefunden worden sind, leisten nur
sehr wenige der obigen Bedingung Genüge. Echte Reliktenseen aber sind
z. B. zahlreiche Wasserbecken des mittlern und südlichen Schweden.
[S. 312]
Litteratur.
[111]J. v. Kennel, Biologische und faunistische
Notizen aus Trinidad. Arbeiten aus dem Zool.-anatom. Institut in
Würzburg, 1883.
[112]R. Credner, Die Reliktenseen. Petermanns
Mitteilungen 1887.
[113] Vergl. G. Duplessis-Gouret, Essay sur la faune
profonde des Lacs de la Suisse, 1885.
[114] Vergl. Fr. Zschokke, Die Tierwelt der
Hochseen. Verhandl. der Deutschen Zoolog. Gesellschaft auf der 1.
Jahresversammlung von 1891.
[115]G. Asper und J. Heuscher, Zur
Naturgeschichte der Alpenseen. Jahresbericht der St. Gallisch. Naturw.
Gesellschaft, 1885–86.
[116]J. Vosseler, Die freilebenden Copepoden
Württembergs etc., 1886.
[117] Vergl. W. Migula, Die Verbreitungsweise der
Algen. Biolog. Zentralblatt, 8. Bd., No. 17, 1888.
[118]Th. Barrois, Note sur la dispersion des
Hydrachnides. Revue biologique du Nord de la France. T. I. 1888–89.
[119]Leidy, Freshwater Rhizopoda of North-America,
1879.
[120]E. Korschelt, Die Entwickelung von Dreyssena
polymorpha. Sitzungsber. der Gesellsch. naturf. Freunde in Berlin,
Jahrg. 1891.
[S. 313]
Über
die wissenschaftlichen Aufgaben biologischer Süsswasser-Stationen.
Von Dr. Otto Zacharias in Plön (Holstein).
[S. 315]
Auch ausserhalb der Fachkreise dürfte es ziemlich allgemein bekannt
sein, dass ich vor einigen Jahren (1888) die Errichtung einer
besonderen Anstalt zum Zwecke eingehender Untersuchungen über die Tier-
und Pflanzenwelt des Süsswassers angeregt und in ihrer Notwendigkeit
begründet habe. Es geschah dies durch einen Aufsatz in No. 269 des
„Zoologischen Anzeigers“. Seitdem sind drei Jahre verflossen und in der
wissenschaftlichen sowohl wie in der Tagespresse ist der betreffende
Vorschlag vielfach erörtert worden. Namhafte Zoologen und Botaniker
zollten meinem Plane sogleich Beifall und bestärkten mich in meinem
Vorhaben; andere, nicht minder ausgezeichnete Forscher nahmen aber
das Projekt mit Zurückhaltung auf. Dies ist der gewöhnliche Gang
der Dinge, sobald es sich um eine Neuerung handelt. Meistenteils
werden in einem solchen Falle auch noch absprechende Stimmen laut;
diese Regel bestätigte sich jedoch meinen Bestrebungen gegenüber
nicht. Im Gegenteil gesellte sich zu den beistimmenden Kundgebungen
alsbald noch der weitere günstige Umstand, dass wohlhabende Fach-
und Privatleute das Projekt in freigebigster Weise durch Geldspenden
förderten. Hierdurch und durch das wahrhaft liberale Entgegenkommen
des Bürgermeisters[CXVII] und der Stadtgemeinde von Plön ist es mir
schliesslich gelungen, meine Pläne zu verwirklichen, und gegenwärtig
erhebt sich am Nordufer des Grossen Plöner Sees — in [S. 316]unmittelbarster
Wassernähe — ein stattliches, villenähnliches Gebäude, welches
eine hinlängliche Anzahl von Räumlichkeiten umfasst, in denen
wissenschaftliche Untersuchungen mit derselben Bequemlichkeit
vorgenommen werden können wie in den biologischen Laboratorien
kleinerer Universitäten.
Von Seiten der Preussischen Staatsregierung wurde dem neubegründeten
Institute in der Folge auch eine finanzielle Beihilfe (zunächst auf
fünf Jahre) zu teil, sodass ein recht glücklicher Anfang für das
lediglich durch Privat-Initiative ins Werk gesetzte Unternehmen zu
verzeichnen gewesen ist.
Der Studien-Aufenthalt in dieser ersten „Biologischen
Süsswasser-Station“ ist Jedem gestattet, der die zum selbständigen
Arbeiten erforderlichen Vorkenntnisse mitbringt. Insbesondere
freilich sind die fünf vorhandenen Arbeitsplätze für Naturforscher
von Fach bestimmt, welche am Grossen Plöner See zoologische,
pflanzenphysiologische oder auf das Fischereiwesen bezügliche
Beobachtungen anstellen wollen. Für alle diese Zwecke sind in der
Station die geeigneten Hilfsmittel (Fahrzeuge, Fanggerätschaften,
Mikroskope, Reagentien und Aquarien) vorhanden.
Wer davon unterrichtet ist, mit welch interessanten Lebensformen
uns die letztjährigen Durchforschungen unserer heimatlichen Tümpel,
Teiche und Seen bekannt gemacht haben, der wird die Nachricht von der
Begründung einer Dauerstation zur näheren Untersuchung jener Organismen
mit aufrichtiger Genugthuung begrüssen. Die Umgebung von Plön ist in
vorzüglicher Weise für diesen Zweck geeignet, insofern das Thal des
Schwentine-Flusses, in welchem das freundliche Städtchen gelegen ist,
fast lediglich aus einer Aneinanderreihung von Wasserbecken besteht,
von denen die kleinsten so gross sind wie unsere ansehnlichsten
mitteldeutschen Seen. Hier ist also ein weites Feld für faunistische
und biologische Forschungen eröffnet, d. h. für Studien, welche die
Feststellung der verschiedenen Tier- und Pflanzenorganismen des
Süsswassers und die Ermittelung von deren Existenzbedingungen zum Ziel
haben.
Im Hinblick auf den Reichtum an Lebewesen, welchen das Meer in seinem
Schosse birgt, waren Viele von der Ansicht[S. 318] beherrscht, dass es
sich wohl erst gar nicht verlohne, Zeit und Kraft an die Gewässer
des Binnenlandes zu verschwenden. So wurde die Süsswassertierwelt
allmählich zum Aschenbrödel der wissenschaftlichen Zoologie degradiert,
und wer sich wirklich noch damit abgab, lief Gefahr, von seinen für das
Salzwasser schwärmenden Fachgenossen als ein nicht ganz ebenbürtiges
Mitglied der Forschergilde betrachtet zu werden. Glücklicherweise
giebt es aber zu jeder Zeit Leute, die den Mut haben, allgemeinen
Vorurteilen zu trotzen, und so hat auch die Süsswasserfauna in den
jüngstverflossenen zwei Jahrzehnten ihre Freunde und Bearbeiter
gefunden. Männer wie F. A. Forel, G. Asper und E.
Imhof in der Schweiz, P. Pavesi in Italien, A.
Fritsch, B. Hellich und W. Vavra in Österreich,
O. Nordquist in Finnland, Jules Richard und Jules de
Guerne in Frankreich (zahlreicher anderer nicht zu gedenken) haben
mit bewundernswerter Unermüdlichkeit dem Studium der Wassertierwelt
obgelegen und Erfolge erzielt, deren wissenschaftliche Bedeutung von
Niemand mehr übersehen oder in Abrede gestellt werden kann. Ich selbst
habe während des Zeitraumes von 1884 bis 1889 die Fauna der nord- und
mitteldeutschen Seen, sowie diejenige der Eifelmaare durch eingehende
Untersuchungen festgestellt. Durch eben diese Forschungen sind wir mit
vielen neuen Arten von kleinen Krebstieren (Entomostraken) bekannt
geworden, haben den Reichtum unserer Gewässer an schwimmenden und
schlammbewohnenden Würmern, an Schnecken, Muscheln, Moostieren und
einzelligen Lebewesen (Protozoen und niederen Algen) kennen gelernt,
sind in die bunte Gesellschaft der Wassermilben und Wasserkerbtiere
eingedrungen, deren Gewimmel hauptsächlich die seichtere Uferzone
belebt — kurz, wir haben einen umfassenden Überblick über die
mannigfaltige Bewohnerschaft unserer binnenländischen Seebecken
erlangt, die bisher nur Fische und „Gewürm“ (als deren Nahrung) zu
enthalten schienen. Unsere vermehrte Kenntnis erstreckt sich aber
nicht nur auf die einzelnen Gattungen und Arten der äusserlich
unscheinbaren Wasserfauna, sondern auch mit auf die Art und Weise, wie
jede Spezies ihren besonderen Lebensverhältnissen angepasst ist, wie
sie sich ernährt und ihren[S. 319] Platz im Kampfe ums Dasein behauptet, was
für Mittel ihr zur räumlichen Ausbreitung verliehen sind und welcher
Zusammenhang zwischen der Bevölkerung des Seegrundes und derjenigen
der oberflächlichen Wasserschichten (bezw. der Uferzone) besteht. Aber
mit Gewinnung dieser Einsicht sind wieder zahlreiche neue Probleme
aufgetaucht, welche sich auf die Ursachen der Veränderlichkeit, die
Wirkung der Isolierung, den mutmasslichen Einfluss des „äusseren
Mediums“ u. dergl. beziehen, sodass es niemals an Arbeit für zahlreiche
Forscher auf diesem Gebiete fehlen kann.
Der Hauptvorteil eines dicht am Seeufer gelegenen und mit
allen Einrichtungen der modernen Forschungstechnik versehenen
Stationsgebäudes besteht augenscheinlich darin, dass man auf solche
Weise in den Stand gesetzt wird, alle Chancen des Wetters und der
Beleuchtungsverhältnisse beim Einsammeln der Untersuchungsobjekte
wahrzunehmen, und dass sich einem in beständiger Wassernähe die
Möglichkeit zu zahlreichen Beobachtungen darbietet, welche auf nur
gelegentlichen Ausflügen an dieses oder jenes Wasserbecken — aus
Mangel an Zeit und Ruhe — überhaupt nicht gemacht werden können.
Ich denke da in erster Linie an die Erforschung der Zusammensetzung der
sogenannten pelagischen Süsswasserfauna (des Limnoplanktons) in den
verschiedenen Jahreszeiten, und an die sehr wünschenswerte Klarstellung
der Beziehungen dieser merkwürdigen Tiergesellschaften zu den übrigen
Bewohnern des betreffendes Sees, besonders auch ihr Verhältnis zu
den Fischen, von denen einige, wie man glaubt, vorwiegend in ihrer
Ernährung auf gewisse pelagisch lebende, d. h. beständig im freien
Wasser sich aufhaltende Krebstiere angewiesen sind. Im Grossen Plöner
See besteht jene Fauna pelagica nach meinen Ermittelungen (von
1886 und 1891) aus folgenden Spezies:
Dazwischen kommt auch noch in grossen Mengen ein zur
Flagellaten-Gattung Dinobryon gehöriges Wesen (vergl. Fig. 35
im ersten Bande dieses Werkes) und das ebenfalls zu den Geisselträgern
gehörige Ceratium hirundinella vor. Ein feines Schwebnetz aus
Müllergaze, mit dem wir vom Boote aus bloss zehn Minuten lang die
oberflächlichen Wasserschichten der Seenmitte abfischen, enthält nach
Ablauf dieser kurzen Zeit einen förmlichen Brei auf seinem Grunde,
welcher lediglich aus den soeben namhaft gemachten Krebs-, Rädertier-
und Flagellaten-Spezies besteht.
Die nähere Erforschung der Lebens- und Ernährungsweise dieser
pelagischen Tiere, welche in staunenswert grosser Individuenzahl unsere
Binnenseen bevölkern, wäre — wie schon betont — eine sicher zu
wichtigen Aufschlüssen führende Arbeit, welche von einer biologischen
Süsswasserstation in Angriff genommen werden könnte. Freilich würden
zur Bewältigung einer solchen Aufgabe keineswegs nur Wochen und Monate,
sondern zweifellos mehrere Jahre erforderlich sein. Was wir bis jetzt
über die Biologie jener rastlos schwimmenden Wesen wissen, ist durch
die verschiedensten Forscher bei Gelegenheit von Ferienreisen, in
Sommerfrischen u. s. w., wodurch die Betreffenden zufällig in die Nähe
grösserer Süsswasseransammlungen gelangten, festgestellt worden. Hin
und wieder (ich erinnere nur an die ausgezeichneten Forschungen Prof.
Aug. Weismanns über Daphniden) sind solchen Gelegenheitsstudien
die schönsten und weittragendsten Resultate zu verdanken gewesen.
Aber eben darum, weil sich solche Untersuchungen schon öfters als im
hohen Grade lohnend erwiesen haben, erscheint es geboten,[S. 321] dieselben
fortzusetzen und sie so zu organisieren, dass wertvolle Ergebnisse
nicht bloss vom Zufall abhängen, sondern vielmehr mit einiger
Sicherheit erwartet werden können.
Eine andere Frage vom allgemeinsten wissenschaftlichen Interesse
wäre die nach der Winterfauna unserer Landseen, d. h. eine
Ermittelung derjenigen Tiere, welche während der Kältemonate unter
der Eisdecke im Wasser ausdauern und weiter leben, während andere
beim Eintritt der niedrigen Temperatur hinsterben, nachdem sie
den Fortbestand ihrer Art durch die Produktion und Ablage von
Dauer-Eiern (vergl. Bd. I S. 367) gesichert haben. Dass
verschiedene Infusorienspezies, Spaltfusskrebschen und mancherlei
Würmer in unseren Teichen während des Winters zu finden sind, weiss man
schon seit längerer Zeit; aber auf diese wenigen Thatsachen beschränkt
sich gegenwärtig unsere Kenntnis, sodass es angezeigt wäre, sich einmal
näher darüber zu unterrichten, welche Tiere (bezw. niedere Pflanzen)
es denn sind, deren Lebensfunktionen unter der Einwirkung von Kälte
so gut wie gar keine Beeinträchtigung erfahren. Diese Aufgabe könnte
gleichfalls auf das Programm einer nahe am Seeufer befindlichen Station
gesetzt werden, und sie wird wohl auch einer solchen reserviert
bleiben, da es vollkommen unthunlich ist, derartige Untersuchungen
ausserhalb des Bereichs einer den Forscher sowohl wie das von ihm
aufgefischte Material vor Frost schützenden Unterkunft vorzunehmen.
Ein reiches und anziehendes Arbeitsgebiet für den in unmittelbarer
Seenähe stationierten Zoologen würde selbstredend auch die
Beobachtung der Wasserinsekten und der Larvenzustände von solchen
Landkerbtieren sein, welche ihre Eier ins Wasser ablegen. Es ist
nicht zu bezweifeln, dass Studien dieser Art, wenn sie auf eine
grössere Anzahl verschiedener Objekte ausgedehnt werden, interessante
Aufschlüsse in allgemein biologischer Hinsicht zu liefern im stande
sind. Ich erinnere hier nur an die schöne Untersuchung, welche Dr.
E. Schmidt-Schwedt unlängst (1887) über Atmung der Larven und
Puppen des Schilfkäfers (Donacia crassipes) veröffentlicht
hat[121], und an die auf den Gehäusebau der Phryganiden-Larven (vergl.
diesen Band S. 94 und ff.)[S. 322] bezüglichen Beobachtungen der bekannten
Naturforscherin Fräul. Marie v. Chauvin.
Ein nicht minder grosses Interesse würde die Erforschung jener
eigentümlichen Fortpflanzungsverhältnisse darbieten, welche bei
einigen unserer verbreitetsten Süsswasserturbellarien (Stenostoma
leucops, Microstoma lineare) abwechselnd in der Form
von geschlechtlicher und ungeschlechtlicher Vermehrung auftreten.
Man weiss zwar, dass zu Beginn der kalten Jahreszeit die letztere
an die Stelle der ersteren tritt, aber man ist noch sehr wenig
darüber informiert, durch welche histologischen Vorgänge es zu einer
Hervorbildung männlicher und weiblicher Zeugungsorgane in den bis
dahin geschlechtslos gewesenen Würmern kommt, die sich nur auf dem
Wege der Querteilung (vergl. Bd. I, S. 259) fortpflanzten. Dasselbe
Problem liegt auch in betreff gewisser Anneliden, z. B. beim gemeinen
Wasserschlängelchen (Nais) vor, und es wäre im hohen Grade
wertvoll, über den Modus der geschlechtlichen Differenzierung in beiden
Würmergruppen genauere Angaben zu erhalten. Dass wir solche noch
vermissen, liegt an der Schwierigkeit der Materialbeschaffung. Befindet
man sich nicht in nächster Nachbarschaft eines Sees oder grösseren
Teiches, so ist es ganz unmöglich, den rechten Moment wahrzunehmen, um
die genannten Tiere in den geeigneten Stadien einzusammeln.
In solchen und ähnlichen Fällen hängt der Fortschritt unseres Wissens
im wesentlichen nur von der rechtzeitigen und bequemen Erlangung
der Beobachtungsobjekte ab. Und das ist der Hauptpunkt, welchen ich
bei Motivierung der Notwendigkeit von permanenten Stationen für die
Erforschung der Süsswasser-Lebewelt nicht oft genug betonen kann.
Faunistische Exkursionen in irgend einer Seengegend sind ganz gewiss
für die Erweiterung unserer Kenntnis der Wasserfauna von Wert; aber
wer eine derartige ambulante Forschungsthätigkeit längere Zeit
hindurch betrieben hat, wird wissen, dass man dabei eigentlich
niemals zur Ruhe kommt. Man schwelgt bei solchen Ausflügen häufig in
einer herzerquickenden Fülle von Material, hat aber unterwegs höchst
selten so viel Zeit, um sich der Bearbeitung[S. 323] desselben mit der
erforderlichen Musse zu widmen. Infolgedessen konserviert man möglichst
zahlreiche Objekte und kehrt mit einer grossen Menge von Gläschen
nach Hause zurück. Hier findet nun erst die eingehende Besichtigung
der verschiedenen Funde statt, wobei man aber in der Regel die wenig
erfreuliche Wahrnehmung macht, dass man von der einen Materialsorte
viel zu viel, von der anderen aber leider lange nicht genug angesammelt
hat. Wäre man an Ort und Stelle in der Lage gewesen, umfassendere
Studien vorzunehmen, so würde bei demselben Zeit- und Kraftaufwande
ein belangreicheres Resultat zu verzeichnen gewesen sein. Auch diese
Erfahrung, mit der ich gewiss nicht ganz allein stehe, spricht klar
für die Nützlichkeit von Dauerstationen, wenn es sich um das Studium
unserer Süsswasserfauna handelt. Dasselbe gilt natürlich auch im
Hinblick auf die lakustrische Pflanzenwelt.
Dass indessen auch faunistische Exkursionen, wenn sie mit Eifer und
Gründlichkeit ausgeführt werden, nach verschiedenen Richtungen hin
Neues zutagefördern können, dafür legt eine unlängst publizierte
Arbeit von Prof. M. Braun („Die Turbellarien Livlands“, 1885)
beredtes Zeugnis ab. Ebenso liefert die bekannte Abhandlung Dr. K.
Ecksteins über die Rädertiere der Umgebung von Giessen eine
schlagende Bestätigung für die beherzigenswerte Mahnung: „Sieh, das
Gute liegt so nah’ ....“ Auch durch meine eigenen Arbeiten über die
niedere Fauna einheimischer Seebecken und Teiche hoffe ich den Beweis
erbracht zu haben, dass in unseren süssen Gewässern noch mancherlei
Neues zu entdecken ist. Ich brauche in diesem Bezug nur an die schon
erwähnte Auffindung einer den Monotiden nahestehenden Turbellarie
in den Hochseen des Riesengebirges zu erinnern, deren Anwesenheit
später in verschiedenen schweizerischen Seen und neuerdings (1890)
durch Prof. Fr. Zschokke auch im See von Partnun (auf der
Rhätikonbergkette) nachgewiesen wurde. Von nicht geringerem Interesse
war die Entdeckung mehrerer Vertreter der ausserordentlich merkwürdigen
Turbellarien-Gattung Bothrioplana, welche sich ebenfalls als
Folge der von mir unternommenen Ausflüge an die Riesengebirgsteiche
ergab. Hierzu kommt noch die[S. 324] Erbeutung mehrerer neuer Kruster- und
Hydrachniden-Arten in den nord- und mitteldeutschen Wasserbecken bei
Gelegenheit meiner Studienreisen in den Jahren 1885 und 1886. Besonders
weise ich aber auch auf den von Dr. W. Weltner erst kürzlich
konstatierten und bisher gar nicht vermuteten Reichtum der Spree an
Spongillen (vergl. Bd. I, 6. Kapitel) hin und auf die umfassenden
Ermittelungen W. Vavras über die Verbreitung der Ostracoden
(Muschelkrebse) in Böhmen[122].
Solche Exkursionen werden auch fernerhin nicht zu entbehren sein,
namentlich wenn es sich um vergleichende Untersuchungen über
die Fauna verschiedener Landseen handelt. Für Studien dieser Art kann
dann eine permanente biologische Süsswasserstation, welche in einem
seenreichen Gebiet gelegen ist, ein recht fruchtbarer Mittelpunkt
werden. Man wird von einem solchen Zentrum aus vielleicht auch die
Frage nach den äusseren physikalischen Ursachen der Veränderlichkeit
mancher Organismengruppen in Angriff nehmen können, und möglicherweise
mit der Zeit nachzuweisen im stande sein, warum der eine See in dieser,
der andere in jener Weise auf die Gestalt der in ihm lebenden Wesen
abändernd einwirkt. Augenblicklich wissen wir über die Faktoren,
welche hier in Betracht kommen, so gut wie nichts. Und doch ist der
Einfluss der jedesmaligen Lokalität auf manche Organismengruppen mit
ausreichender Sicherheit erwiesen. Clessin hat diese Thatsache
schon vor einem Jahrzehnt für die Mollusken festgestellt, und er nimmt
zur Erklärung derselben „die Anpassung an gegebene Verhältnisse“ in
Anspruch. Es wird nicht überflüssig sein, in den Zusammenhang dieses
Kapitels eine Stelle einzuschalten, die der Leser bereits auf S. 138
dieses Bandes vorgefunden hat. Sie ist aber besonders dazu geeignet,
das, was wir hier besprochen, zu illustrieren. Clessin fasst
das Resultat seiner reichen Erfahrung in folgenden Zeilen zusammen:
„Wer die Wassermollusken längere Zeit im Freien beobachtet, wird sehr
bald zu der Überzeugung kommen, dass fast jeder einzelne Fundort
eigenartige, mehr oder weniger ausgeprägte Abweichungen vom Typus der
bezüglichen Art erzeugt, und dass es geradezu zu den allergrössten
Seltenheiten[S. 325] gehört, zwei ziemlich übereinstimmende Formen an
verschiedenen Fundorten zu konstatieren. Ja sogar der nämliche Fundort
erzeugt bei geänderten Verhältnissen andere Varietäten, und oft genug
finden sich verschiedene Formen einer und derselben Art an sich
berührenden Stellen desselben Gewässers, wenn die Beschaffenheit des
Grundes, die Strömung des Wassers, die Bewachsung u. s. w. sich ändert.
So kommen in den grossen Seen der Voralpen Schnecken und Muscheln mit
ausgeprägtem Seecharakter und solche, welche nicht oder kaum von jenen
zu unterscheiden sind, die in Sümpfen leben, neben einander vor, und
zwar jenachdem die bezüglichen Wohnplätze bei seichtem Wasser und
mangelnder Bewachsung der vollen Wirkung des Wellenschlags ausgesetzt
sind, oder die Ufer in sumpfige Stellen übergehen“.
J. Vosseler hat auf den gleichen Einfluss der chemischen und
physikalischen Unterschiede unserer Gewässer auf den Habitus, die
Färbung und Gliedmassengrösse bei spaltfüssigen Krebsen hingewiesen.
So existiert z. B. in den Maaren der Eifel ein Copepode, der
augenscheinlich dem Cyclops agilis Koch nahesteht, aber kürzere
Antennen, schwächer entwickelte Mundteile, längere Schwimmbeine
und eine sehr gestreckte Schwanzgabel besitzt. Vosseler hat
diesen von mir aufgefundenen Krebs näher untersucht und ihn seines
beschränkten Vorkommens wegen Cyclops maarensis genannt.
Höchstwahrscheinlich ist diese neue Spezies in den Maaren selbst
entstanden und stellt eine interessante Lokalform dar, welche für ihre
Bildung den Cyclops agilis als Ausgangsform gehabt hat.
Im Müskendorfer See bei Konitz in Westpreussen fand ich 1886
zahllose Exemplare einer merkwürdigen Varietät der Hyalodaphnia
cucullata, deren Kopfteil sichelartig gekrümmt und ventralwärts
stark herabgebogen ist. Diese Form (var. nov. procurva Poppe)
kommt lediglich in dem genannten See vor[123] und ist anderwärts bis
jetzt nicht aufgefunden worden. Manche Abweichungen geringern Grades
vom Typus der Art sind für gewisse Fundorte überhaupt charakteristisch.
[S. 326]
So variiert beispielsweise die bekannte Dinoflagellaten-Spezies
Ceratium hirundinella O. Fr. M. von einem See zum andern
hinsichtlich der Panzerbreite und der Hörnerlänge. Wahrnehmungen
hierüber habe ich hauptsächlich bei meiner Durchforschung der
westpreussischen Seen gemacht. Um dieselbe Zeit etwa konstatierte Prof.
G. Asper ähnliche Gestaltungsdifferenzen zwischen den Ceratien
des Thalalpsees und denen des Züricher Sees, wovon er in seiner
Abhandlung über die Naturgeschichte der Alpenseen berichtet[124].
Das pelagische Rädertier Anuraea longispina, welches eine sehr
weite Verbreitung besitzt, variiert nicht bloss hinsichtlich der
Mächtigkeit seiner langen (nadelförmigen) Panzerfortsätze, sondern auch
in der Form des Körperquerschnittes, der gewöhnlich ein Kreissegment
darstellt, häufig aber auch vollkommen dreieckig ist. In Westpreussen
zeigten oft sogar benachbarte Seen langdornige Anuräen, die in der
angegebenen Weise von einander verschieden waren. Nach Asper
ist ein nicht minder verbreitetes Rotatorium, Anuraea aculeata,
ebenfalls bedeutender Variation unterworfen, welche sich aber
vorzugsweise nur auf die Felderung und Skulptur des Panzers erstreckt.
Ähnliche Abweichungen hat Imhof bei Anuraea cochlearis
Gosse angetroffen und die weitgehendsten davon mit besonderen
Speziesnamen (A. intermedia und A. tuberosa) bezeichnet.
Leptodora hyalina, der pelagische Krebs par excellence, zeigt
an seinen verschiedenen Fundorten nicht bloss Verschiedenheiten der
Körperlänge, sondern auch solche, welche die Grösse des Auges, die
Entwickelung des ersten Paares der Schwimmfüsse und die Geräumigkeit
des Brutraumes betreffen. Die gleichen Wahrnehmungen habe ich an
Polyphemus pediculus, einem Kruster der Uferzone, gemacht, der
in klaren und kühlen Bergseen grösser und farbenprächtiger zu werden
scheint, als in den seichteren Gewässern der Ebene.
Nach Anführung dieser Beispiele, welche noch durch Beobachtungen von
A. Wierzejski über die Unbeständigkeit der Artcharaktere bei
Spongilla lacustris vervollständigt werden könnten[125], wird
es einleuchten, dass auch die Süsswasserfauna Stoff zur[S. 327] Diskussion
des Speziesproblems zu liefern im stande ist. Durch eine vergleichende
Untersuchung bestimmter Mitglieder der Wassertierwelt aus verschiedenen
Seen dürfte sich im Laufe der Zeit etwas Genaueres über die Richtung
der Abweichungen und über deren Betrag bei einzelnen Arten ergeben.
Schliesslich möchte ich aber auch einen ganz praktischen
Gesichtspunkt geltend machen, welcher die Errichtung von ständigen
Beobachtungsstationen in der Nähe von grösseren Süsswasserseen
wünschenswert erscheinen lässt. Dies ist nämlich unsere noch sehr
ungenügende Einsicht in die Ernährungs- und sonstigen Lebensbedingungen
der Fische. Auf diesem Felde ist noch sehr viel zu thun, um
für die Bewirtschaftung unserer Seen und Teiche rationelle Grundlagen
zu schaffen. Ein guter Anfang dazu ist von dem österreichischen
Fischzüchter Josef Susta in Wittingau gemacht worden durch
dessen bekannte Untersuchungen über die Ernährung des Karpfens[126].
Aber nicht bloss die Umstände, welche das Gedeihen der Fische
begünstigen, sondern auch deren natürliche Feinde und die Ursachen
solcher Krankheiten, welche gelegentlich eine Massensterblichkeit
unter denselben hervorrufen — alles dies ist der näheren Erforschung
wert und würdig. Aber die zahlreichen Fragen und Probleme, die wir
im Vorstehenden als zum Programm der Thätigkeit einer Biologischen
Süsswasserstation gehörig bezeichnet haben, sind unmöglich von einem
einzigen Forscher zu bewältigen, sondern es müssen sich mehrere zu
diesem Zwecke verbünden, und es bedarf hinsichtlich mancher Aufgaben
längerer Zeiträume (oft vieler Jahre), um sie in befriedigender Weise
zu lösen. Hierüber macht man sich in Laienkreisen häufig recht falsche
Vorstellungen, und ich nehme deshalb in diesem Werke, welches seiner
Tendenz nach für weitere Kreise bestimmt ist, Gelegenheit, allzu
sanguinischen Hoffnungen vorzubeugen.
Das Plöner Stationsgebäude liegt, wie schon erwähnt, unmittelbar am
Grossen Plöner See und die umgebende Naturszenerie ist so beschaffen,
dass ein Zeitungsberichterstatter[CXVIII] davon gesagt hat:
„Ein König könnte sich keinen herrlicheren Fleck der Erde
auswählen, wenn er, der Welthändel müde, glückliche Tage im
Vollgenusse eines grandiosen Landschaftsbildes verleben wollte“.
Für die hier vorzunehmenden Forschungen ist die „herrliche“ Lage
selbstverständlich ganz gleichgültig, aber der See ist durch seine
Grösse (50 qkm = 20000 preussische Morgen) und durch seinen
Organismenreichtum besonders dazu geeignet, ein Arbeitsfeld für
zoologische und pflanzenphysiologische Untersuchungen zu bilden. Dazu
kommt noch die Nachbarschaft anderer grosser Wasserbecken (Kleiner
Plöner See, Trammersee, Behlersee, Dieksee, Kellersee, Grosser und
Kleiner Eutiner See, Ukeleisee u. s. w.), sodass hierdurch zugleich die
denkbar günstigste Gelegenheit zur Vornahme von faunistischen Ausflügen
gegeben ist. Den Verkehr auf den einzelnen Seen vermitteln grosse
Segel- und Ruderboote. Der Biologischen Station steht ausserdem noch
die Benutzung eines Petroleum-Schraubenbootes[CXIX] zur Verfügung,
welches eine ansehnliche Fahrgeschwindigkeit (10–15 km pro
Stunde) besitzt.
[CXIX] Daimlers Patent (geliefert von der Firma Meyer u.
Remmers in Hamburg).
Das Stationshaus ist ein zweistöckiges Gebäude, welches ausser den
erforderlichen Arbeitsräumen (Laboratorium, Experimentierzimmer
und Bibliothek) auch die Wohnung für den Direktor enthält. Im
Erdgeschoss sind die Aquarien untergebracht, welche durch eine
Röhrenleitung mit fliessendem Wasser aus dem See gespeist werden
können. Der Mikroskopiersaal hat dreiflügelige grosse Fenster und die
Arbeitstische sind mit vorzüglichen Instrumenten aus der Optischen
Werkstätte von C. Zeiss in Jena ausgerüstet. Bei aller
Bescheidenheit ihrer Einrichtung besitzt die Plöner Station, wie man
sieht, doch Alles, was zur Ausführung von mikroskopisch-anatomischen
und entwickelungsgeschichtlichen Arbeiten erforderlich ist. Mehr
ist nicht versprochen worden und zu einer luxuriösern Ausstattung
wären auch die Mittel nicht vorhanden gewesen. Vom 1. April 1892 ab
werden die Arbeitsplätze in der Biologischen Station zu Plön für
süsswasserfreundliche Zoologen und Botaniker[127] benutzbar sein.
[S. 329]
Mit Genugthuung übermittele ich am Schlusse dieses Kapitels dem
Leserkreise unseres Buches die Nachricht, dass der bekannte und
verdienstvolle österreichische Zoolog, Prof. Anton Fritsch in
Prag, neuerdings gleichfalls eine stabile Station für Erforschung
der Süsswasserfauna ins Leben gerufen hat. Dieselbe hat ihren Stand
am Unterpocernitzer Teiche bei Bechovic in Böhmen. Es ist ein
festgebautes, hübsches Häuschen, welches ausser einem Arbeitszimmer
von 12 qm Fläche noch einen kleinen Wohnraum von 6 qm
enthält. Dieses Forscherheim hat ein Privatmann, Béla Freiherr
v. Derschenyi, im Interesse der durch Fritsch so tüchtig
geförderten Kenntnis der Wassertierwelt Böhmens auf eigene Kosten
erbauen lassen. Überdies benutzt der Prager Forscher (schon seit
Juni 1888) zu seinen Studien noch eine ortswechselnde Station in
Gestalt eines zusammenlegbaren hölzernen Häuschens, welches in 2½
Stunden aufgestellt und in 1½ Stunden wieder abgebrochen werden
kann[128]. Diese „fliegende Station“ steht jetzt am Gatterschlager
Teich bei Neuhaus, und hier ist besonders der Assistent des Prof.
Fritsch, Herr W. Vavra, während des verflossenen Jahres thätig
gewesen. Unter Anderem wurde in diesem Teiche unlängst ein neuer zu den
Cytheriden gehöriger Muschelkrebs entdeckt, der vorläufig den Namen
Limnicythere stationis erhalten hat. Es ist der kleinste bisher
bekannte Vertreter seiner Gattung.
An die Errichtung solcher eigens dem Studium der Tier- und Pflanzenwelt
des Süsswassers gewidmeter Forschungstationen ist merkwürdigerweise
erst in allerneuester Zeit gedacht worden, obgleich dieselben Gründe,
welche für die Anlage mariner Stationen zum Zwecke biologischer
Studien sprechen, sich auch für lakustrische Observatorien ins Feld
führen lassen. Das Weitere wird nun die Erfahrung und der Erfolg
lehren. Da, wo etwas Neues ins Werk gesetzt wird, tauchen stets auch
einige Pessimisten auf, welche Erwägungen darüber anstellen, ob
es sich wohl auch verlohnen werde, die süssen Gewässer in der von
Fritsch und mir inaugurierten Weise zu durchforschen. Besonders
giebt es unter den Praktikern, d. h. unter den Fischzüchtern und
Fischerei-Interessenten,[S. 330] Leute, welche in erster Linie die Frage
des „Verlohnens“ auf der Zunge haben, ohne manchmal auch nur einen
blassen Schimmer von den Aufgaben zu haben, welche durch die Thätigkeit
einer Süsswasserstation in Angriff genommen werden sollen. Derartigen
Leuten empfehle ich folgenden Ausspruch Prof. Anton Fritschs
zur Beherzigung: „Eine genaue Kenntnis dessen, was der Teich in
seinem Wasser beherbergt, ist die Grundbedingung für dessen rationelle
Bewirtschaftung“. Es ist zu hoffen, dass die Richtigkeit dieses
Satzes in immer weiteren Kreisen zur Anerkennung gelange, und dass
auch von massgebender Seite das Streben der Naturforscher gebührende
Würdigung und Unterstützung finde.
[S. 331]
Litteratur.
[121] Berl. Entomolog. Zeitschrift, 31. Bd., 1887, S. 325–334.
[122] Vergl. Wenzel Vávra, Monographie der Ostracoden
Böhmens. Archiv d. naturwiss. Landesdurchforschung von Böhmen, 8. Bd.,
No. 3.
[123] Eine Beschreibung der Müskendorfer Hyalodaphnia hat
Poppe geliefert in: O. Zacharias, Zur Kenntnis der pelagischen
und littoralen Fauna norddeutscher Seen. Zeitschr. f. wiss. Zoologie,
45. Bd., 2, 1887.
[124]G. Asper und J. Heuscher, Zur
Naturgeschichte etc. Jahresber. der St. Gallischen Gesellschaft,
1885–86.
[125]A. Wierzejski, Beitrag zur Kenntnis der
Süsswasserschwämme. Verhandl. d. k. k. zool.-botan. Gesellsch. in Wien,
1888.
[126]J. Susta, Die Ernährung des Karpfens und seiner
Teichgenossen. Mit 2 Tafeln. 1888.
[127] Vergl. F. Ludwig, Die botanischen Aufgaben der
von O. Zacharias geplanten lakustrischen Station. Biolog. Zentralbl.,
9. Bd., No. 13, 1889.
[128]Anton Fritsch, Die Stationen zur Durchforschung
der Süsswasserfauna. Wiener Landwirtschaftl. Zeitung, 1891.
[S. 333]
Das Tierleben auf Flussinseln und am Ufer der Flüsse und Seen.
Von Fr. Borcherding in Vegesack.
[S. 335]
Wie im Wasser selbst, so regt sich auch an den Ufern unserer Flüsse,
Weiher und Seen ein mannigfaltiges Tierleben, auf welches am Schlusse
dieses Werkes noch ein musternder Blick geworfen werden soll. Auf
zahlreichen faunistischen Ausflügen hat der Verfasser dieses Kapitels
Gelegenheit gehabt, die bunte Gesellschaft der in der Nähe des Wassers
und in demselben sich aufhaltenden Säugetiere, Vögel, Reptilien,
Amphibien, Fische, Mollusken und Insekten zu beobachten und den
eigentümlichen Reiz zu empfinden, den das Leben und Treiben dieser
verschiedenartigen Wesen auf den Freund der Natur ausübt.
Wie schön und fast nur einer poetisch angehauchten Schilderung
zugänglich ist das einsame, waldumgürtete oder von Schilf umkränzte
Ufer eines grossen, breit dahinfliessenden Stromes, oder der ebenso
geschmückte Saum eines im Sonnenschein glitzernden Sees. Wie
vielseitig, buntfarbig und anregend ist das Bild, welches sich hier
an einem herrlichen Frühlingsmorgen oder an einem warmen Sommerabend
unseren Augen darbietet. Überall regt sich vielgestaltiges Leben.
Schreiten wir bei einbrechender Dämmerung über eine Flussinsel oder
fahren wir mit einem Boote geräuschlos am Ufer entlang, so können wir
häufig mehrere Fledermaus-Arten beobachten, und zwar meistenteils
die langohrige Fledermaus, Plecotus auritus Geoffr.,
die frühfliegende Fledermaus, Vespertilio noctula K.
u. Bl., und die Teichfledermaus, Vespertilio dasycneme
Boie.[S. 336] Unermüdlich schwirren diese fluggewandten Tierchen unter dem
Schutze der Dunkelheit durch die Luft, um ihren stets regen Appetit
mit den erbeuteten Fliegen, Mücken, Käfern und Nachtfaltern zu
befriedigen. Eine gleich erfolgreiche Insektenvertilgung üben ausser
den Fledermäusen wohl nur noch Maulwürfe, Talpa europaea
L., und die Wasserspitzmäuse, Crossopus fodiens, aus.
Einzeln erscheint an den Seen und Flüssen auch der Fuchs,
Canis vulpes L., um sich nach einem leckern Entenbraten
umzusehen. Es ist höchst anziehend, den Meister Reineke unbeobachtet
auf seinen Jagdzügen belauschen zu können. Mit eingezogenen Beinen,
mehr über den Boden und durchs Rohr wegkriechend als gehend, die Rute
vornehm hinter sich herschleppend, dieselbe nur dann und wann bald
etwas rechts, bald etwas links bewegend, schleicht er sich durchs Gras
und Gebüsch unvermerkt der Stelle zu, von wo der Entenruf herübertönt,
aber immer gegen den Wind, denn er weiss nur zu gut, wie unangenehm den
Enten seine Witterung ist.
Das schädlichste Säugetier, welches die Ufer der Flüsse und Seen
bewohnt, ist unstreitig der Fischotter, Lutra vulgaris
Erxl. Trotz der vielen Nachstellungen von Seiten der Fischer und Jäger
hat dieser arge Räuber sich in den letzten Jahren in unserm Nordwesten
ganz bedeutend vermehrt. Die grossen Rohr- und Weidendickichte an
den Ufern der Flüsse und Seen und auf den grösseren und kleineren
Flussinseln geben ihm solch sichere Verstecke, dass er sehr leicht
dem Jäger entgeht. Zudem ist der Tisch immer reichlich für ihn
gedeckt; er braucht deshalb nicht, wie der Fuchs, die Nähe der
menschlichen Wohnungen aufzusuchen. Welchen ungeheuren Schaden der
Otter der Fischzucht zufügt, mag aus folgendem Beispiel erhellen. Ein
ausgewachsener Otter gebraucht zu seiner täglichen Nahrung 2 kg
Fische. Das macht für ein Pärchen ohne Jungen in einem Jahre 2 mal 2
mal 365, also 1460 kg. Ist der Fischotter in sonst fischreichen
Flussgebieten häufig, so sieht man aus den angeführten Zahlen, wie
stark dann der Fischbestand durch ihn dezimiert werden muss.
[S. 337]
Ein weiteres auch recht schädliches Mitglied der Familie der
Mustelinae, Foetorius putorius K. u. Bl., verirrt sich
zum Glück nur einzeln an unsere Gewässer. Vor einigen Sommern erhielt
Verfasser ein prächtiges Männchen vom Iltis, im Volksmunde „Ilk“, „Elk“
oder „Ülk“ genannt, welches auf einer Weserhalbinsel erlegt worden
war. Sodann mag noch ein für Deutschland sehr seltener Wasserbewohner
erwähnt werden, der Nörz, Wasserwiesel oder auch wohl Sumpfotter
genannt, Putorius lutreola K. u. Bl. Dieser wegen seines
wertvollen Pelzes eifrig verfolgte Marder ist im östlichen Europa
ziemlich häufig, dagegen gehört er in Deutschland zu den grössten
Seltenheiten. Der Nörz bewohnt mit Vorliebe die bewaldeten Ufer der
Flüsse, ist aber vereinzelt auch in der Ebene angetroffen worden. Vor
einigen Jahren wurde im Blocklande an der Wümme (unweit Bremen) ein
Exemplar erlegt.
Dass aus der Ordnung der Nager nur allzu häufig an den Gewässern
die Wanderratte, Mus decumanus Pall., anzutreffen ist,
mag ebenfalls nicht unerwähnt bleiben. Auch die Wasserratte,
Arvicola amphibius Lacep., findet sich an den Ufern der
kleineren Gewässer nicht selten; in unserm Nordwesten recht häufig in
schwarzer Färbung. Zu einer wahren Landplage wird in manchen Jahren
die gemeine Feldmaus, Arvicola arvalis Pall., welche
in den Marschen und an den Deichen oft zu vielen Tausenden erscheint
und dort grossen Schaden verursacht. Einige wenige Zahlen mögen ein
Bild von ihrem massenhaften Auftreten geben. Im Amte Elsfleth an der
Weser wurden im Jahre 1880–1881 347571 Mäuse eingeliefert und 20284.80
Mark an Prämien dafür bezahlt; im Amte Brake nördlich von Elsfleth,
ebenfalls an der Weser gelegen, wurden 1880 158913 Mäuse eingeliefert
und an Prämien 12237.85 Mark bezahlt; 1881 wurden ebendaselbst 338781
Mäuse eingeliefert und 19127.31 Mark an Prämien dafür bezahlt. Als
Prämie wurde im „Oldenburgischen Mäuseverbandsbezirke“ je nach der
Häufigkeit der Mäuse 2, 5, 10 oder 20 Pfennige für das Stück bezahlt.
In diesem Sommer (1891) war die Feldmaus nur vereinzelt anzutreffen;
der lange und strenge Winter mit dem hohen Wasserstande hat stark unter
ihnen aufgeräumt, stärker[S. 338] als in den reichsten Jahren Mäusefänger,
Bussarde, Weihen, Raben und Füchse es vermögen.
Auf den Aussterbeetat ist wohl in unserm Vaterlande der Biber,
Castor Fiber L., gesetzt. Nur wenig bekannte Kolonien finden
sich in Deutschland, in denen er sich einstweilen, wenn auch nur
in geringer Zahl, noch erhalten hat. Die bedeutendste ist zwischen
Magdeburg und Wittenberg an der Elbe; auch an der Havel, Oder und
Weichsel finden sich noch vereinzelte schwach bewohnte Kolonien. Leider
wird diesem seltenen Nager seines kostbaren Pelzes wegen gar zu sehr
nachgestellt und trotz des Regierungsschutzes, der ihm in letzter Zeit
zu teil geworden ist, wird er in nicht zu ferner Zeit zu denjenigen
Tieren Deutschlands gehören, die aufgehört haben zu leben, und einzelne
Ortschaften, Gründe u. s. w. werden nur noch mit ihrem Namen an das
frühere Vorkommen dieses stattlichen Nagers erinnern.
In die Flüsse, besonders die der Nordsee, steigt bei hohen Fluten mit
starken Nordweststürmen vereinzelt auch der Seehund, Phoca
vitulina L., hinauf und kommt dann den Fischern zuweilen ins Garn.
Häufiger findet sich in unseren Flüssen Phocaena communis Cuv.,
der Tümmler oder Braunfisch; denselben kann man bei Springfluten
oft in der Nähe der Städte beobachten, wie er in kurzen Zwischenräumen
sich an die Oberfläche des Wassers begiebt, um im nächsten Augenblicke
wieder in die Tiefe zu verschwinden.
Versetzen wir uns in Gedanken um etwa 200 Jahre zurück, so finden
wir sogar einen Walfisch im Weser- und Lesumflusse. Es war
Hyperoodon rostratus Pontop., welcher damals in der Lesummündung
oberhalb Vegesack gefangen wurde. Das Exemplar befindet sich im Bremer
Museum.
Beendigen wir hiermit unseren Streifzug, auf welchem wir uns
ausschliesslich nach den Säugetieren, die an und in den Gewässern
vorkommen, umgesehen haben, so gewahren wir, obwohl wir nicht jeden
Säuger, der sich uns auf unseren Exkursionen am Wasser zeigen könnte,
angeführt haben, dass trotzdem die Zahl der Arten eine ziemlich
geringe bleibt. Ganz anders gestaltet sich[S. 339] dagegen das Bild, wenn wir
einen neuen Beobachtungsgang unternehmen und uns nun der Vogelfauna
der süssen Gewässer zuwenden. Da hat fast jede Jahreszeit ihr
eigenartiges Gepräge. Im Frühjahre finden wir ausser den ansässigen
und heimkehrenden Brutvögeln viele durchziehende Wanderer, die für
kurze Zeit Rast an den Gewässern und auf den Inseln machen. Im
Sommer und Herbste sehen wir ausser den alten Brutvögeln die junge
Nachkommenschaft in den verschiedenartigsten Kleidern. Auf dem
Herbstzuge kommen noch Hunderte von Gästen hinzu, welche der Vogelwelt
(Ornis) eines bestimmten Gebietes oft ein ganz fremdartiges Aussehen
verleihen.
Halten wir zunächst eine systematische Umschau unter denjenigen Vögeln,
welche an den Ufern der Flüsse und Seen und auf den kleinen und
grösseren Flussinseln ihre Wohnungen eingerichtet haben, und fassen
wir dann die Gäste, welche sich zur Frühjahrs- und Herbstzugszeit
an unseren Gewässern bald längere, bald kürzere Zeit aufhalten,
etwas näher ins Auge. Als Brutvögel treffen wir aus der Ordnung der
Raubvögel zuerst zwei Weihenarten an, Circus aeruginosus Sav.,
die Rohr- oder Sumpfweihe, und Circus cineraceus
Mont., die Wiesenweihe. Beide werden im Volksmunde gewöhnlich
„Grashoafk“ genannt. Wo dichtes Rohr- und Weidengestrüpp auf wenig
belebten Flussinseln und auf den einsamen Groden der Flüsse sich
findet, da kann man mit ziemlicher Sicherheit den Horst der einen oder
anderen Weihe erwarten. Derselbe befindet sich im dichtesten Gestrüpp
am Boden und ist nur dann mit Sicherheit aufzufinden, wenn man die
Alten, welche an ihrem schwebenden Fluge, den langen, spitzen Flügeln
und dem ziemlich langen Schwanze leicht von den Bussarden, Habichten
und Milanen zu unterscheiden sind, beobachtet und sich genau die Stelle
merkt, an welcher sie niedergehen. Dieses Ausspionieren muss jedoch
mit der grössten Vorsicht geschehen; denn glaubt sich der Beobachter
schlau, so ist der Beobachtete doch in vielen Fällen noch gewitzigter
und hat ersteren oft viel eher bemerkt, als derselbe ihn. Viel leichter
ist der Horst aufzufinden, wenn die Weihen Junge haben; dann braucht
man nur aus möglichster Ferne[S. 340] das Männchen, bei welchem man sehr
leicht mit einem guten Glase die Beute in den Fängen erkennen kann, zu
beobachten. Ist es in der Nähe des Horstes angelangt, so erscheint mit
einem lauten, scharfen „kirrr“ über der Rohrfläche das Weibchen, aber
in bedeutend geringerer Flughöhe als ersteres. Ist das Männchen über
der Gattin angekommen, so wirft sich letztere geschickt auf den Rücken
in dem Augenblicke, in welchem der Gatte die Beute fallen lässt. Mit
grosser Sicherheit greift das Weibchen dieselbe auf und eilt raschen
Fluges dem Horste zu. Das Männchen streicht sofort von dannen, um neue
Beute heranzubringen. Das ist der günstigste Augenblick, um sicher
den Nistplatz auszukundschaften, denn wir brauchen uns nur genau den
Platz zu merken, an welchem das Weibchen sich niederlässt. Ist das
Glück uns in dieser Weise günstig gewesen, so finden wir im hohen Grase
oder Rohre auf dem Boden ein ziemlich grosses mit trockenem Grase und
Rohr ausgepolstertes Nest, in welchem sich drei bis fünf hungrige,
gelbgraue Junge befinden. In dem Horste der Wiesenweihe findet man
Mitte Mai etwa vier bis fünf weisse, etwas ins bläuliche übergehende
Eier. Im Neste der Rohrweihe trifft man zur selben Zeit vier bis fünf
grünlich weisse Eier. Befestigt man über dem Neste ein gutes Schlagnetz
und entfernt sich dann möglichst weit, um im dichten Rohr eine gute
Deckung zu suchen, so dauert es gewöhnlich nicht lange, bis das
Weibchen zum Horste zurückkehrt, allerdings zuerst nur, um auszukunden,
ob alles wieder in gewohnter Ordnung ist. Doch es lässt sich noch nicht
sogleich nieder, sondern in weitem Bogen umkreist es einige Mal die
nähere und weitere Umgebung des Nistplatzes. Bald ist aber die Furcht
vor der Gefahr geschwunden, die Liebe zu den ängstlich kreischenden
Jungen ist grösser, es streicht zum Horste, lässt sich nieder und —
sitzt gefangen unter dem Schlagnetze. Schwieriger ist es, des Männchens
habhaft zu werden, da letzteres selten das Füttern besorgt und noch
seltener zum Horste geht. In den ersten Stunden nach dem Verlust der
Gattin lässt es nur die Beute aus der Luft ins Nest fallen und eilt
wieder fort, um neue Nahrung zu beschaffen; doch endlich ist auch bei
ihm die Liebe zu seinen Jungen,[S. 341] die ihn durch ihr Geschrei auf ihren
Hunger und ihre Einsamkeit aufmerksam machen, vollständig erwacht;
es lässt sich nieder, um das Amt der Gattin zu übernehmen. Aber das
Netz ist wieder aufgestellt worden, es schlägt abermals zu und nun
sitzt auch der Gatte gefangen bei seinen Jungen. Nicht immer glückt
ein solcher Jagdzug, viel Geduld und Vorsicht gehört dazu. — Die
Nahrung der Weihen besteht aus jungen Vögeln, welche aus den am Boden
befindlichen Nestern geraubt werden, und aus Mäusen. Der Nutzen, den
sie durch Vertilgung der letzteren gewähren, wird wohl reichlich durch
den Schaden, den sie durch Zerstören der jungen Vogelbrut verursachen,
aufgehoben. Die Wiesenweihe erscheint im April, die Rohrweihe im März;
beide verlassen uns im Oktober.
Von den Eulen treffen wir an unseren Gewässern als Brutvogel dann
und wann Otus brachyotus Boie, die Sumpfohreule,
im Volksmunde „Moorule“ genannt. Häufiger ist sie nur in reichen
Mäusejahren zu beobachten. Den Horst findet man, allerdings nicht
leicht, auf den alten Weidenköpfen, die sich stellenweise an den
Flussläufen finden. Einzeln entdeckt man ihn auch im langen Grase oder
im Rohrdickichte. Im Neste finden sich Anfang Mai vier bis sechs fast
runde weisse Eier. Die Nahrung dieses nächtlichen Räubers besteht fast
ausschliesslich aus Mäusen, und muss die Sumpfohreule daher zu den
nützlichen Vögeln gerechnet werden.
Ein häufiger Bewohner unserer Inseln und Flussufer ist der
Kuckuck, Cuculus canorus L. Nach der Meinung der
Landleute ist derselbe im Sommer Kuckuck, im Winter „Stothoafk“. Die
Erklärung dieser irrigen Meinung ist sehr leicht. Im Frühjahre und
Sommer findet sich der „Stothoafk“, Astur nisus K. u. Bl., nicht
in der Nähe der menschlichen Wohnungen, sondern in den dichten Wäldern
bei seinem Brutplatze. Der Kuckuck lässt dann aber überall seinen
Ruf ertönen. Im Herbst, wenn der Kuckuck längst über alle Berge ist,
erscheint aber der Sperber in der Nähe der menschlichen Wohnungen.
Da etwas Ähnlichkeit im Gefieder der beiden besteht, findet obige
Fabel leicht Glauben bei der Landbevölkerung. Als Pflegeeltern seiner
Brut wählt der Kuckuck[S. 342] sich die Rohrsänger, die gelbe Bachstelze,
ja auch einzeln das Blaukehlchen. Letztere besorgen die Pflege mit
der grössten Gewissenhaftigkeit und oft sogar mit Aufopferung der
eigenen Jungen, welche vor diesem gefrässigen Stiefbruder zurückstehen
müssen. Verfasser dieses hatte Gelegenheit, in der Marsch einen
vollständig flüggen jungen Kuckuck zu beobachten, der sich durch sein
klägliches „zirrk, zirrk“ bemerkbar machte, wie dieser grosse Bursche
sich von seinen Pflegeeltern, Budytes flava Cuv., mit grosser
Behaglichkeit noch füttern liess. Der Kuckuck erscheint Ende April und
verweilt bis Anfang September.
Dem hinsichtlich seines Gefieders schönsten und an die Ornis der
Tropen erinnernden Vogel, Alcedo ispida L., Eisvogel,
begegnen wir häufig an den Flüssen und auf den Inseln, welche steile,
lehmige Ufer haben. Regungslos sitzt dieser prächtige Geselle auf einem
über die Wasserfläche hinhängenden Zweige, seinen Blick unverwandt
nach unten gerichtet; plötzlich stösst er ins Wasser, um im nächsten
Augenblicke wieder auf der Oberfläche mit einem erbeuteten Fische im
Schnabel zu erscheinen, welcher in wenigen Sekunden gierig verschlungen
wird. Dieses interessante Schauspiel kann man oft in ganz kurzer
Zeit mehrmals beobachten. Sein Nest, welches sich immer an den steil
aufsteigenden Wänden der Flussufer befindet, ist am Ende einer etwa
einen Meter langen Röhre angelegt, und dort findet man auf Fischgräten,
wenigen Hälmchen u. dgl. Mitte April sechs bis sieben glänzend weisse
Eier.
Einen ähnlichen Brutplatz, wie den des Eisvogels, wählt sich auch
aus der Familie der Schwalben die Uferschwalbe, Sand-, auch
Bergschwalbe genannt, Cotyle riparia L. An den schroffen Stellen
der Flussufer, an welchen die Geest unmittelbar an das Wasser tritt,
kann man zur Frühjahrszeit Hunderte dieser geschickten Insektenjäger
aus- und einfliegen sehen. Das Nest befindet sich am Ende einer
oft zwei Meter langen, wagerechten Röhre, ist mit Federn weich
ausgepolstert und enthält Ende Mai oder Anfang Juni fünf bis sechs
schneeweisse Eier. Die Uferschwalbe erscheint Anfang Mai und verlässt
uns Anfang oder Mitte September.
[S. 343]
An den Gewässern der gebirgigen Gegenden unseres Vaterlandes finden
wir aus der Familie der Wasseramseln den Wasserstaar, auch
Wasseramsel oder Wasserschmätzer genannt, Cinclus aquaticus
Bechst. Ich hatte zu verschiedenen Malen Gelegenheit, diesen munteren
halb Wasser- halb Singvogel in seinem Elemente zu beobachten, so
unter anderen an den Berlebecker Quellen, an den Ilsefällen u. a. m.
Bald watet er bis an den Hals durchs Wasser, bald steht er regungslos
auf einem erhöhten Steine, um im folgenden Augenblicke ins Wasser zu
stürzen; bald läuft er am Boden des Gewässers hurtig dahin, bald fliegt
er durch herabstürzende Wasserfälle, wobei ihm sein dichtes Gefieder
von grossem Vorteil ist. Das einzige Nest, welches der Verfasser
Gelegenheit hatte näher zu betrachten, befand sich hinter einem
Wasserfalle in einer kleinen Felshöhle; obwohl auch seitlich dahin zu
gelangen war, nahm das kecke Tierchen jedesmal seinen Weg durch die
herabströmende Wassermenge. In dem Neste befinden sich im April vier
bis fünf weisse Eier.
Der schönste Vogel, welcher unsere Flussufer und Inseln bewohnt,
ist unstreitig das Blaukehlchen, Cyanecula suecica
Brehm. Wo dichtes Weidengebüsch an unserm Weserufer oder auf den
Platen[CXX] vorhanden ist, da findet man auch dieses prächtige
Tierchen, jedoch halten sie sich meistens sehr verborgen und laufen
zwischen dem dichten Gestrüpp einher, aber ein klares, reines „fied,
fied“ oder ein kurzes „täck, täck“ verrät bald ihr Vorhandensein.
Das Nest dieses schönen Sängers ist immer recht versteckt angelegt
und es gehört zu den grössten Seltenheiten, ein solches aufzufinden,
da es auf unseren Weserplaten immer im dichtesten Gestrüpp, oft hart
am Ufer in Weidenstümpfen sich befindet. Selten fliegt der Vogel vom
Neste, um dadurch seinen Brutplatz zu verraten, sondern gewöhnlich
springt er unbemerkt auf den Boden und läuft, ohne einen Laut von
sich zu geben, im Gestrüpp davon. Es ist mir erst zweimal geglückt,
ein solches Nest aufzufinden; [S. 344]dasselbe ist ziemlich kunstvoll gebaut
und sitzt gewöhnlich in einer kleinen Vertiefung; es besteht aussen
meist aus trockenen Grashalmen, innen dagegen vorwiegend aus der Wolle
der Salix-Arten. In dem schön gerundeten tiefen Neste liegen gegen
Ende April fünf schön grüngraue Eier, welche mit einzelnen zierlichen
rotbraunen Tüpfelchen und Pünktchen bedeckt sind. Das Blaukehlchen
erscheint schon Ende März mit dem Hausrotschwänzchen zusammen und
verlässt seinen Brutplatz Ende August, streicht aber dann noch bis Ende
September umher. In den letzten Tagen des September oder Anfang Oktober
verlässt es uns; einzelne Nachzügler dagegen verweilen oft bis Ende
Oktober in unserm Gebiete.
[CXX] „Platen“ heissen in Norddeutschland die kleinen mit
Rohr und Weiden bewachsenen Flussinseln.
Mit dem Blaukehlchen zusammen an fast denselben Lokalitäten treffen
wir das Braunkehlchen, Pratincola rubetra Koch, und
einzeln auch die Braunelle, Accentor modularis Cuv.,
an. Beide haben mit ihm die versteckte Lebensweise sowie die Art des
Nestbaues gemein; beide verlassen auch lautlos ihre Nester und eilen
unter dem Gesträuch davon. Nur wenn die Vögel ihre Brut gefährdet
sehen, lassen sie ihre Klagetöne hören und verraten allerdings dadurch
dem aufmerksamen Beobachter, dass man sich unmittelbar beim Neste
befindet, und trotzdem hält es manchmal recht schwer, dasselbe zu
entdecken. Verfasser dieses Kapitels, welcher sich am 23. Mai 1891
unmittelbar bei einem solchen Neste befand, gebrauchte über eine
halbe Stunde, um dasselbe endlich in einem dichten Grasbüschel, kaum
einen halben Meter von ihm entfernt, zu gewahren. In dem sauber mit
Tierhaaren ausgepolsterten Neste befanden sich vier grünlich blaue,
fein rot punktierte Eier. Das Nest der Braunelle aufzufinden ist mir,
trotz eifrigen Suchens, bislang noch nicht gelungen. Das Braunkehlchen
erscheint Mitte April und verlässt uns Ende September. Die Braunelle
dagegen kommt oft schon Mitte März und verschwindet erst gegen Mitte
Oktober.
Die „Kuckucksamme“, Curruca cinerea Lath., ist ebenfalls
nicht selten auf den Flussinseln anzutreffen. Die Dorngrasmücke
führt den oben genannten Namen daher, weil der Kuckuck sehr gern
sein Ei in ihr Nest legt und ihr auch die Erziehung seines[S. 345]
Sprösslings überlässt Schon am frühen Morgen mit Sonnenaufgang lässt
die Dorngrasmücke ihr munteres Lied ertönen, und sie ist einer von
denjenigen Vögeln, welche am längsten singen, ja oft vernimmt man noch
Anfang August ihren fröhlichen Gesang. Mitte April trifft dieser Vogel
bei uns ein und verlässt uns wieder Mitte September.
An den Flussufern und auf den Inseln beobachtet man ziemlich häufig den
Fitis-Laubvogel, Weidenzeisig, Phyllopneuste trochilus
Bp. Sehr bald verrät sich dieses zutrauliche Vögelchen durch sein
angenehm klingendes „hüid, hüid, hoid, hoid“. Es lässt sich auch
ganz in der Nähe beschauen, aber desto schwieriger ist es, sein Nest
aufzufinden. Dasselbe befindet sich am Boden, meist im dichten Gestrüpp
oder Gewirr; es ist vollständig überwölbt und man sieht nur ein kleines
seitliches Loch. Anfang Mai findet man in demselben fünf bis sieben
kleine, weisse, rötlich gefleckte Eier, an deren stumpfen Ende sich die
Makeln und Pünktchen dichter als am spitzen Ende gruppieren. Dieser
niedliche Sänger verweilt bei uns von Anfang April bis Ende September.
War das Vorkommen der bis jetzt angeführten Sänger an den Flussufern
und auf den Inseln kein ausschliessliches — auch ausserhalb dieser
Gebiete werden dieselben angetroffen —, so wollen wir jetzt eine
Gruppe kennen lernen, welche ausschliesslich ihre Heimat an den
Ufern der Flüsse, auf den Inseln und Groden haben. (Unter „Groden“
versteht man die mit Weiden und anderm Gestrüpp bewachsenen
Aussendeichsländereien, welche auch stellenweise Grasflächen haben.)
Es ist dies die Gattung der Rohrsänger. Schon ihr Name sagt uns
deutlich genug, wo wir diese Bewohner aufzusuchen haben. Nicht
weniger als sechs Arten dieser Gattung bewohnen das in Frage kommende
Gebiet unseres Vaterlandes. Ausserdem giebt es davon noch eine Reihe
Spezies, welche mehr dem südlichen Europa angehören. Für uns kommen
in erster Linie unsere heimatlichen Sänger in Betracht. Es sind
folgende: Der Schilfsänger, Calamoherpe phragmitis Bp.;
der Binsen-Rohrsänger, Calamoherpe aquatica Degland;
der Sumpf-Rohrsänger,[S. 346]Calamoherpe palustris Boie; der
Heuschrecken-Rohrsänger, Calamoherpe locustella Penn.;
der Teich-Rohrsänger, Calamoherpe arundinacea Boie, und
endlich der Drossel-Rohrsänger, Calamoherpe turdoides
Meyer. Erstere werden im Volksmunde gewöhnlich „Reitmeeschen“,
auch „Rohrsperlinge“ genannt; der letztere heisst gewöhnlich die
„Rohrdrossel“. Fast alle erscheinen bei uns Anfang Mai und bleiben
bis Anfang September. Nicht alle sind gleich häufig anzutreffen;
einzelne dagegen, wie die Rohrdrossel, der Schilfsänger, der Sumpf-
und Teich-Rohrsänger, sind sogar ziemlich häufig. Sie entziehen sich
aber durch ihre versteckte Lebensweise im Röhricht und durch die
Lokalitäten, an denen sie leben, sehr oft dem Auge des Beobachters
und daher ist manche Spezies vielleicht viel häufiger als an manchen
Orten allgemein angenommen wird. Der Drossel-Rohrsänger ist sofort
durch seinen weit vernehmbaren Gesang, der etwa: „düi, düi, düi, karre,
karre, karre, kei, kei, kei, kerr, kerr, kerr, karra, karra, kied“
klingt, zu erkennen. Der Gesang von Calamoherpe palustris Boie
ist dem flötenden Gesange des Spottvogels, Ficedula hypolais
Schlegel, nicht unähnlich und daher ein sicheres Erkennungszeichen
des Sumpf-Rohrsängers, da der Spottvogel in den Gebieten, wo die
Sumpf-Rohrsänger leben, nicht zu Hause ist. Beim Schilfsänger,
C. phragmitis Bp., besteht der Gesang fast ausschliesslich
aus einem langen, wohlklingenden, flötenartigen Triller. Der Gesang
beim Teich-Rohrsänger, C. arundinacea Boie, gleicht mehr
einem Geplapper der sehr rasch hinter einander ausgesprochenen
Silben „terr, terr, terr, tri, tri, tri, zerrr, zerrr, zerrr, zäck,
zäck, zäck“ u. a. m. Eine annähernde Ähnlichkeit hat der Gesang
des Binsen-Rohrsängers, C. aquatica Degland, mit dem des
Schilfsängers; doch kommen in dem melodischen Triller sehr häufig
Töne wie „jüpp, jüpp, jüpp, jüpp, tütt, tütt, tütt, tütt“ vor. Den
eigentümlichsten Gesang, wohl richtiger Geschwirre genannt, hat der
Heuschreckensänger, C. locustella Penn. Mit aufgeblasener
Kehle, am Boden zwischen den Rohr- und Weidenstengeln dahinlaufend,
bringt er nur einen wie „sirrrrrrrirrr“ klingenden lange anhaltenden
Ton hervor. Für einen aufmerksamen Beobachter ist es nicht allzu[S. 347]
schwer, das Vorkommen der einen oder andern Art nach dem Gesange
festzustellen, und um so bequemer, als das Eindringen in die im und
am Wasser belegenen Rohrfelder sowie auf die mit Schlick bedeckten
Groden, in welchen sich die Rohrsänger ebenfalls gern aufhalten, mit
grossen Schwierigkeiten und Anstrengungen verknüpft ist. Auch im
Nestbau gleichen sich unsere Rohrsänger mehr oder weniger. Zwischen
drei bis vier bei einander stehenden Rohrstengeln an oder über der
Wasserfläche, zwischen Nesselpflanzen oder zarten Weidenruten, zwischen
starken Grashalmen oder anderen Pflanzenstengeln befestigen sie
kunstvoll ihr Nest und zwar so, dass immer zwei oder drei Stengel durch
die Seitenwandungen des Nestes hindurch gehen. Das schön gebaute Nest
wächst mit den Pflanzen in die Höhe und ist so bei Hochwasser vor dem
Überschwemmtwerden geschützt.
Aus der Familie der Motacillidae halten sich am Gewässer und
in der Nähe desselben die drei bekannten Bachstelzenarten auf.
Motacilla alba L., die weisse Bachstelze, auch Quäksteert oder
Wippsteert genannt; die graue Bachstelze, Motacilla sulphurea
Bechst., und die gelbe Bachstelze oder Kuhstelze „gäle Quäksteert“,
Motacilla flava Cuv. Obwohl man die erstere auch entfernt vom
Wasser antrifft, so schlägt sie doch mit Vorliebe ihre Wohnung unter
Brücken, an Mühlen und auf den in der Nähe des Wassers stehenden
Weidenstümpfen auf. Die im nördlichen Deutschland seltene graue
Bachstelze hält sich fast ausschliesslich an Bächen, Quellen, überhaupt
an fliessenden Gewässern auf und baut auch ihr Nest stets in die Nähe
des Wassers, in Höhlen, unter Brücken, in Felslöchern u. dgl. m.
Die gelbe Bachstelze dagegen bewohnt die freien, von menschlichen
Wohnungen fern liegenden Weiden auf den Inseln und an den Flussufern;
mit Vorliebe diejenigen Weiden, auf welchen Vieh weidet. Das Nest
derselben findet man nicht selten in Carices-Büscheln. Die weisse
Bachstelze erscheint in unserm Nordwesten oft schon im Februar — nach
dem verflossenen harten Winter 90/91 wurden die ersten Bachstelzen
erst am 16. März 91 beobachtet — und verlässt uns Oktober. Die weisse
Bachstelze ist nach dem Staar unser erster Frühlingsbote.[S. 348] Die gelbe
Bachstelze trifft bei uns Anfang April ein und verweilt bis Mitte
September.
In Gemeinschaft mit den gelben Bachstelzen trifft man ziemlich häufig
den Wiesenpieper, Anthus pratensis L. Er wählt zu seinem
Brutplatze dieselben Lokalitäten wie die Bachstelzen. Gewöhnlich
erscheint er bei uns im März und bleibt oft bis November. Einzelne
bleiben sogar in gelinden Wintern ganz bei uns. Charakteristisch und
dabei ein sicheres Erkennungszeichen der Art ist ihr Verhalten beim
Gesange, welcher etwa „witje, witje, witje, zick, zick, zick, jück,
jück, jück, tirrrirrr“ lautet. Plötzlich erhebt sich der Pieper singend
einige Meter in die Höhe und fällt ebenso rasch wieder zur Erde, um
auf einem Carex- oder Scirpus-Büschel den Gesang fortzusetzen oder zu
vollenden.
Von den Lerchen kommt für unser Gebiet nur die durch ihren
jubilierenden herrlichen Gesang bekannte Feldlerche, Alauda
arvensis L., insoweit in Betracht, als sie ziemlich häufig als
Brutvogel auf den Flussinseln anzutreffen ist. Sie erscheint oft schon
Mitte Februar und verkündet dann durch ihre schmetternden Lieder das
Neuerwachen des Frühlings. Sie verlässt uns Ende Oktober oder zu Beginn
des November.
Von den Ammern suchen auch einige zu ihrem Brutplatze die Nähe
der Gewässer auf. So findet sich auf den Weserinseln gar nicht selten
die Grauammer, Emberiza miliaria Bp. Ihr Nest ist meist tief
versteckt in den Grasbüscheln angelegt; auf dem Festlande dagegen wählt
sie mit Vorliebe die Getreidefelder zu ihrem Nistplatze. Wenn man im
Frühjahre die grösste unserer Ammern auf irgend einem Weidenstumpfe
sitzen sieht und das ziemlich eintönige „zick, zick, zick, sirrrr“
hört, so kann man sicher sein, dass in gar nicht zu grosser Entfernung
sich das Nest befindet. Fast ausschliesslich an Gräbenufern nistet die
bekannte Goldammer, „Gälgöschen“, Emberiza citrinella
L. Der Gesang „si, si, si, si, siiiih“ unterscheidet sie sofort, ohne
dass man den Vogel zu sehen braucht, von der Grauammer. Stets schlägt
ihren Wohnsitz die Rohrammer, Rohrsperling oder Reithlüning,
Emberiza schoeniclus L., in der Nähe der Gewässer auf. Das sehr
verborgene[S. 349] Nest findet man in Carex-Büscheln. In demselben trifft man
Anfang Mai fünf bis sechs rötlich weisse Eier, deren ganze Schale mit
braunroten Strichen und grösseren Flecken bedeckt ist. Während die
beiden erstgenannten Ammern den Winter über bei uns bleiben und uns
nur einzeln bei sehr strenger Kälte verlassen, wohl auch nur etwas
südlicher streichen, um bei der nächsten gelinden Witterung sich wieder
bei uns einzustellen, verlässt uns die Rohrammer gewöhnlich schon
Ende September, erscheint aber im März wieder auf der Bildfläche. In
gelinden Wintern bleiben dann und wann auch wohl einzelne Pärchen hier.
Ganz einzeln trifft man auch den Hänfling, „Grauiserken“,
Fringilla cannabina Bp., auf den Flussinseln an, welche viel
hohes Gestrüpp haben. Auch der Feldsperling, Weidenspatz,
Passer montanus Koch, ist als Brutvogel auf den Inseln zu
finden, wo alte Weidenstümpfe, die er zu seinem Nistplatze wählt,
stehen. Der Hänfling ist den ganzen Winter hindurch auf den Inseln,
Platen und an den Flüssen anzutreffen, wenn kein Schnee fällt; ein Teil
derselben verlässt uns im September und kehrt im ersten Frühjahre, im
März, zurück. Der Feldspatz ist im ganzen Gebiete Standvogel.
Selbst die Ordnung der Hühner ist auf den grösseren, mit langem Grase
bewachsenen Platen durch das Rebhuhn, Perdix cinerea
Briss., vertreten. Wenn man am frühen Morgen oder gegen Abend in einem
Boote an einer solchen Insel entlang fährt, so vernimmt man bald aus
der nächsten Nähe, bald wieder aus weiterer Ferne den wohlbekannten
Ruf des Rebhahnes „girrräk“. Noch häufiger hört man des Abends von
den Inseln und Groden das „röärp, röärp“, den gewöhnlichsten Ruf des
Wachtelkönigs, Crex pratensis Bechst., herübertönen.
Obgleich dieser Vogel an den Flussufern und auf den Inseln ziemlich
häufig ist — man kann an warmen Abenden oft drei, vier und mehr zu
gleicher Zeit rufen hören —, bekommt man denselben doch sehr selten zu
Gesicht, da er gewöhnlich im Grase rasch fortläuft, ohne aufzufliegen.
Selbst vor dem suchenden Hunde fliegt er nicht eher auf, bis er in
die Enge, etwa an einen Graben, getrieben wird. Das Nest[S. 350] findet man
sehr selten und schwer. Die prächtigsten Gelege — zehn bis zwölf Eier
von schöner graugelber Farbe mit vielen rötlich braunen Flecken —,
die in meinen Besitz gekommen sind, verdanke ich ausschliesslich den
Grasmähern. Der „Snarrentar“, wie er im Volksmunde nach seinem Rufe
genannt wird, erscheint bei uns gewöhnlich Anfang Mai und verschwindet
Anfang September.
Von den Gallinulidae treffen wir weiter auf den Inseln und an
den Flussufern als Brutvögel das punktierte Rohrhuhn, Rallus
porzana L.; die Wasserralle, Rallus aquaticus L.;
das grünfüssige Rohrhuhn, Gallinula chloropus Lath.,
und das schöne, grosse Blässhuhn, Fulica atra L., an.
Diese echten Wasserbewohner sind in unserm ganzen Nordwesten an und
auf den grossen Flussinseln, in den undurchdringlichen Rohr- und
Schilffeldern der Groden an den grösseren und kleineren Seen fast
überall anzutreffen, aber äusserst schwer zu beobachten, da sie
dem Auge des Naturbeobachters durch geschicktes Tauchen oder durch
Verschwinden zwischen dem dichten Schilf und Rohr auszuweichen wissen.
Es ist interessant, diese gewandten Schwimmer und Taucher in ihrem
nassen Elemente beobachten zu können. Hat man sich unbemerkt mit dem
Boote in irgend eine gedeckte Bucht oder in ein Rohrfeld gelegt, so
erscheint oft, wenn das Glück günstig ist, in unmittelbarer Nähe des
Bootes einer dieser munteren Vögel, aber im nächsten Augenblicke,
kaum dass wir Zeit hatten, ihn auch nur halbwegs ins Auge zu fassen,
verschwindet er in der kühlen Flut, schwimmt eine grosse Strecke unter
der Wasserfläche fort, um oft in ganz entgegengesetzter Richtung
wieder zu erscheinen; oft nimmt er sogar seinen Weg unter dem Boote
durch, und im nächsten Augenblicke ist er auch schon wieder fort.
Glaubt er sich erst in sicherer Entfernung, dann kann man ihn auch
längere Zeit auf der Wasserfläche beobachten. Oft sieht man dann die
ganze Familie, Alt und Jung, im bunten Durcheinander, bald ruhig
dahinschwimmend, bald Tauchübungen anstellend, bald über die glatte
Wasserfläche dahinlaufend; aber nie lassen sie dabei die nötige
Vorsicht ausser Acht; bei dem geringsten verdächtigen Geräusch
verschwindet die ganze Gesellschaft im nächsten Rohrfelde[S. 351] und lässt
sich für lange Zeit nicht wieder blicken. Auch im Bau ihrer Nester,
welche sehr versteckt und meist nach dem Wasser hin in den Rohr- und
Binsenfeldern angebracht sind, gleichen sie sich. Auf niedergebogenem
Rohr, oft auf halb schwimmendem Gestrüpp sind die ziemlich kunstlosen
Nester angelegt. Die Zahl der Eier beträgt gewöhnlich acht bis zwölf.
Die Gallinulidae erscheinen in unserm Gebiete Anfang Mai und
verlassen uns Mitte September.
Von den Charadriidae bewohnt die Flussinseln und Groden
als Brutvogel der allen bekannte Kiebitz, Vanellus
cristatus Meyer und Wolf, welcher leider von Jahr zu Jahr an
Zahl bedenklich abnimmt. Die Ursache dieser von Jahr zu Jahr sich
steigernden Abnahme dieses nützlichen Vogels liegt in dem leidigen,
unvernünftigen Eiersammeln, welches einzig den Zweck hat, den Gaumen
des Gourmands zu kitzeln. Allerdings tragen auch ein gut Teil Schuld
die Entwässerungsanstalten, welche in jedem Frühjahre die nassen
und sumpfigen Wiesen trocken pumpen und es dadurch den Eiersammlern
ermöglichen, auf die Poller und höher gelegenen Stellen, die Brutplätze
des Kiebitzes, zu gelangen, um auch dort ihr Zerstörungswerk mit Erfolg
zu betreiben. Der Kiebitz gehört zu unseren ersten Frühlingsboten; er
erscheint gewöhnlich Mitte März und verlässt uns Ende September oder
Anfang Oktober. In milden Jahren erscheint er oft schon im Februar
und bleibt einzeln bis in den November. Manchmal aber, wenn sie sich
zu früh hergewagt haben und Kälte und Schnee zurückkehren, müssen sie
wieder flüchten und auch dabei gehen viele zu Grunde. Am 23. Februar
dieses Jahres beobachtete Verfasser einen Trupp von etwa 50 Stück,
welche sich trotz des vergangenen strengen Winters in unserm Nordwesten
eingefunden hatten; als aber nach einigen Tagen von neuem Kälte eintrat
und ziemlicher Schnee fiel, zogen sie auf einige Zeit wieder südlicher.
Mit dem Kiebitz erscheint und verschwindet fast zur selben Zeit der
Flussregenpfeifer, Charadrius fluviatilis Bechst. Dieser
hurtige, behende Geselle bewohnt mit Vorliebe die kahlen, kiesigen Ufer
der Inseln und Flüsse und legt auch dort sein Nest an, welches[S. 352] sehr
leicht übersehen werden kann. In einer kleinen Vertiefung fast ohne
alle Unterlage liegen auf kiesigem Grunde drei bis vier graugelbe,
mit dunkelgrauen Punkten und Strichen ausgestattete Eier, welche in
ihrer Färbung so sehr der Umgebung ähneln, dass man oft, wenn man das
Nest gefunden, es auch schon wieder aus den Augen verloren hat und von
neuem suchen muss. So ging es am 18. Mai dieses Jahres dem Verfasser,
der, kaum zwei Fuss vom Neste entfernt, es erst nach längerem scharfen
Umhersehen wieder entdeckte.
Der Rotschenkel, Totanus calidris Bechst., welcher
sich in grosser Zahl als Brutvogel an unseren Nordseeküsten findet,
wird auch einzeln als solcher an den Ufern der Flüsse und auf den
Flussinseln beobachtet. Er erscheint bei uns Mitte Mai und verlässt
uns Anfang September. Häufiger als den Rotschenkel kann man an den
sandigen Ufern als Brutvogel den Flussuferläufer, Actitis
hypoleucos Brehm beobachten, derselbe erscheint im März und
bleibt bis zum Oktober. Auf den feuchten, kurzgrasigen Wiesen unserer
Flussniederungen stellt sich in ziemlich grosser Individuenzahl
der durch seine possierlichen Kapriolen bekannte Kampfhahn,
Machetes pugnax Cuv. ein. Stundenlang kann man, wenn ein
Gestrüpp oder ein Grabenufer uns die nötige Deckung giebt, diesem
tollen Treiben des Streithahns oder Streitvogels zusehen. Bei diesen
sogenannten „Kämpfen“, die besonders zur Paarungszeit häufig sind,
nimmt er die wunderbarsten Stellungen ein, sträubt sein Gefieder bald
so und im nächsten Augenblick wieder anders. Sie stürzen auf einander
los, vorwärts, rückwärts, und scheinbar mit einer solchen Wut, dass
der uneingeweihte Beobachter glauben muss, keiner verlasse lebend den
Kampfplatz; indessen scheint es mehr eine Spiegelfechterei zu sein,
denn sie lassen kaum Federn dabei und nach einiger Zeit, wenn sie des
Kampfes müde sind, eilen sie vergnügt von dannen, um sich für ein
späteres Turnier, deren man täglich mehrere beobachten kann, wieder
zu stärken. Das Nest, welches im Bau sowohl wie im Aussehen dem des
Kiebitzes ähnlich ist, befindet sich in einer kleinen Vertiefung, einer
Kuhspur oder dergleichen, und ist mit wenig Grashälmchen ausgelegt.[S. 353]
Nach beendigtem Paarungsgeschäfte verlassen uns schon meistens die
Männchen und ziehen an die Meeresküste, während die Weibchen und Jungen
bis zum September an ihren Brutplätzen verweilen. Im März treffen sie
wieder zusammen an letzteren ein.
Auf den feuchten, mit kleinen Wasserläufen durchzogenen Wiesen der
Inseln der Aussendeichsländereien und Groden, auf den Dobben und
Platen treffen wir in ziemlicher Zahl als Brutvögel die einfache
Bekassine, auch „Häwelamm“ oder „Bäwerbuck“ genannt, Scolopax
gallinago L., und die Doppelbekassine, Scolopax major
L., an. Unter Dobben versteht man beweglichen Moorboden, der oben durch
eine Grasnarbe bedeckt und ziemlich fest, weiter nach unten aber noch
weich ist. Beim Betreten solchen Bodens bewegt sich die ganze Fläche
und der auf diesem Boden Unbekannte bricht sehr leicht ein; es bedarf
einer besondern Geschicklichkeit, darüber hinwegzugehen. Pferde, welche
ihn betreten wollen, bekommen Holzschuhe angeschnallt, und die Räder
der Wagen werden, damit sie nicht einschneiden und dann versinken, mit
dicken, gedrehten Strohseilen umwickelt. An warmen Frühlingsabenden
macht sich die einfache Bekassine durch ihr eigentümliches Gemecker,
dem einer Ziege nicht unähnlich, daher „Häwelamm“, „Himmelsziege“
genannt, welches sie nur im Fluge vernehmen lässt, bemerkbar. An
einzelnen Moorseen ist dieser, von Feinschmeckern sehr geschätzte
Vogel oft zu hunderten anzutreffen, so am Balk-See, einem inmitten des
unwirtlichen Moores bei Cadenberge unweit der niederelbischen Bahn
gelegenen grossen Moorsees, dessen Ufer von grossen Dobbenfeldern
gebildet werden. Auf solch sumpfigen Wiesen in der Mitte eines
Carex-Busches findet sich das sehr schwer zu entdeckende Nest. Die
gemeine Bekassine erscheint bei uns oft schon im März und bleibt bis
zum November. Die Doppelbekassine kommt erst im April und geht schon im
September.
Anfang Mai erscheint ebenfalls auf den feuchten Flussniederungen die
Pfuhlschnepfe, Limosa melanura Leisler, um dort ihr
Brutgeschäft zu verrichten; sie verlässt uns gewöhnlich[S. 354] schon Anfang
September. Auch die grosse Rohrdommel, Botaurus stellaris
Briss., „Iprump“ nach ihrem unheimlich klingenden Rufe „üü — prump“ so
genannt, ist als vereinzelter Brutvogel der Flussinseln aufzuzählen. Es
hält schwer, dieses stattliche Tier zu Gesicht zu bekommen, da es durch
eine eigentümliche List sich dem Auge des Beobachters zu entziehen
weiss. Bemerkt die Rohrdommel einen Feind in ihrer Nähe, so richtet
sie sich gerade auf, zieht den Hals ein, streckt Kopf und Schnabel
senkrecht in die Höhe und bleibt in dieser Stellung unbeweglich stehen,
bis die Gefahr vorüber ist. In dem Weidengebüsch, in welchem sie sich
gern aufhält, gleicht sie in dieser Stellung, wobei ihr die Färbung
ihres Gefieders, welche mit der Umgebung grosse Ähnlichkeit hat, sehr
zu statten kommt, täuschend einem alten abgebrochenen Weidenstumpfe.
Die Rohrdommel erscheint im April auf ihrem Brutplatze und zieht Ende
September wieder von dannen.
Aus der Ordnung der Schwimmvögel treffen wir zunächst an unseren
süssen Gewässern einige Entenarten als Brüter an. Da mag zuerst die
Löffelente, Rhynchaspis clypeata L., erwähnt werden,
welche im dichten Rohr, umgeben von Wasser, ihr verstecktes Nest
anlegt; ferner die Knäckente, Anas querquedula L., die
Krickente, Anas crecca L., und die gemeine wilde Ente,
auch Stockente genannt, Anas boschas L., gehören zu den
häufigeren Arten der Entensippschaft, welche an den süssen Wassern
brüten. Die Enten erscheinen auf dem Frühjahrszuge meist im März und
ziehen Oktober wieder fort. Einzelne Exemplare von boschas und
crecca trifft man fast den ganzen Winter an den Gewässern an und
diese verlassen uns nur dann, wenn auch die letzten offenen Stellen der
Flüsse und Seen mit einer Eisdecke verschlossen sind.
An dem in Betracht kommenden Gebiete trifft man von den
Pelecanidae im Binnenlande einzeln die Kormoran-Scharbe,
Halieus Carbo Ill., als Brutvogel an. Im Nordwesten Deutschlands
ist nur eine Kolonie dieses der Fischerei sehr schädlichen Vogels
bekannt und zwar im Lüneburgischen an der Elbe. Der Kormoran legt
seinen Horst, entgegen der Gewohnheit der anderen Schwimmvögel,[S. 355] auf
Bäumen an, benutzt aber gewöhnlich die Nester der Reiher und Raben für
sich, wobei sich häufig ein hartnäckiger Kampf zwischen diesen und
jenen entspinnt. Trotz seiner grossen Schwimmfüsse bäumt der Kormoran
sehr geschickt auf und weiss sich ganz sicher auf den Ästen zu benehmen.
Wenn der eigentliche Aufenthaltsort der Seeschwalben und
Möven auch das salzige Meer, die Watten und die Inseln des
Meeres sind, so giebt es doch eine Reihe Arten davon, welche vorziehen
im Binnenlande an den süssen Gewässern sich aufzuhalten und dort
ihr Heim einzurichten. Es sind dies die Küstenmeerschwalbe,
Sterna macrura Naum.; die Flussmeerschwalbe, Sterna
hirundo L.; die kleine Seeschwalbe, Sterna minuta
L., und die schwarze Seeschwalbe, Sterna nigra Briss.,
und von den Möven die Lachmöve, Larus ridibundus L. Ihre
Nester, welche, fast ohne jegliche Unterlage, nur aus einer kleinen
Vertiefung bestehen, finden sich auf den kiesigen, sandigen Stellen an
den Gewässern und sind sehr schwer von der Umgebung zu unterscheiden,
da die Plätze gewöhnlich alles Pflanzenwuchses entbehren. Im Winter
bleiben immer einzelne dieser geschickten Segler bei uns, wenn auch
als gewöhnlich anzunehmen ist, dass sie im März in grösseren Scharen
erscheinen und uns im Oktober verlassen.
Endlich mögen noch zwei Brutvögel der Binnengewässer aufgezählt
werden. Es sind dies der grosse Lappentaucher, auch
Kronentaucher genannt, Podiceps cristatus L., und der kleine
Lappentaucher, welcher im Volksmunde den etwas derben Namen
„Pärködel“ führt, Podiceps minor L. Ersterer ist auf fast
allen unseren Seen ein gemeiner Brutvogel. Schon von ferne hören wir
seinen lauten Ruf „koar, koar, koar“ über die Wasserfläche zu uns
herüberschallen, ehe man den geschickten Schwimmer und Taucher zu
Gesicht bekommt. Er ist ein äusserst scheuer und schlauer Vogel, der
seinen Beobachter immer in respektabler Entfernung hält; kommt man ihm
trotzdem unvermerkt zu nahe, so verschwindet er plötzlich unter der
Wasserfläche und erscheint weit weg nur mit dem Kopfe über derselben,
um schon im nächsten Augenblicke von neuem zu verschwinden. Dieses
Experiment[S. 356] wiederholt er so häufig, bis er sich wieder sicher fühlt.
Einen Jäger lässt er daher sehr selten in Schussnähe kommen. Wird der
Taucher angeschossen, und ist der Schuss nicht gleich tödlich, so
taucht er fort und hält sich am Grunde des Gewässers am Rohr und Schilf
fest, um nicht wieder an die Oberfläche zu kommen. Binsenschneider
am Dümmersee brachten dem Verfasser, welcher sich zu der Zeit am See
aufhielt, ein Exemplar des Kronentauchers, welcher am Vormittage von
einem Herrn angeschossen war, ohne dass er ihn bekommen hatte. Das
Exemplar hatten dieselben beim Schneiden mit herauf gebracht. Es hatte
noch die Binse, an welcher es sich festgehalten hatte, im Schnabel,
und dieselbe auch im Todeskampfe nicht losgelassen. Der kleine Taucher
ist ebenso geschickt im Schwimmen und Tauchen, wie sein grösserer
Vetter. Das Nest der Taucher ist fast immer frei auf der Wasserfläche
zwischen Rohr befindlich und an einzelnen Stengeln befestigt, damit es
vom Winde nicht fortgetrieben wird. Die Nester sind äusserst schwer
aufzuspüren, da man nur von der Wasserseite mit dem Boote an die Rohr-
und Binsenfelder gelangen kann, in welchen sie angelegt sind.
Im vorhergehenden ist versucht worden, dem Naturbeobachter in kurzen
Zügen ein Bild zu entwerfen von den Brutvögeln, welche derselbe auf
Flussinseln, an den Ufern der Flüsse und Seen aufzufinden vermag.
Es ist damit aber nicht beabsichtigt, ein genaues, vollständiges
Verzeichnis aller an den in Frage kommenden Lokalitäten brütenden
Vögel zu geben; im wesentlichen sind die häufigeren und am meisten ins
Auge fallenden Arten berücksichtigt, welche den Nordwesten unseres
Vaterlandes bewohnen. Im östlichen und südlichen Deutschland finden
sich noch einzelne Arten, welche nicht erwähnt worden sind. Auch im
folgenden hat Verfasser vorwiegend die Bewohner der Gewässer unseres
Nordwestens im Auge gehabt. Wenn bei manchen Vögeln Angaben über
Brutzeit, über Eintreffen und Verschwinden an ihren Nistplätzen gemacht
worden sind, so beziehen sich dieselben ebenfalls auf Beobachtungen,
welche im Nordwesten angestellt worden sind.
[S. 357]
Ist schon das Gesamtbild unserer befiederten Freunde zur Brutzeit ein
buntes und mannigfaltiges, so gestaltet es sich noch viel reicher
im Frühjahrs- und Herbstzuge, ja manchmal erhält das Bild ein ganz
fremdartiges Aussehen; denn da erscheinen an unseren süssen Gewässern
Gäste, die sonst nur im hohen Norden, am Meere oder anderen uns fern
liegenden Orten anzutreffen sind. Es soll im folgenden versucht
werden, dem Naturfreunde auch davon ein kleines, also nicht auf
Vollständigkeit Anspruch machendes Bild zu entwerfen. Durchwandern wir
im Geiste noch einmal die Vogelwelt und beginnen wir von neuem mit den
Raubvögeln, so finden wir gar nicht selten zur Zugzeit auf den grossen
Wasserflächen den Seeadler, Haliaëtos albicilla Leach,
der besonders häufig erscheint, wenn viele wilde Gänse zur Herbstzeit
in den Flussniederungen sich länger aufhalten. Im Volksmunde wird er
deshalb gewöhnlich mit „Goosarnt“ bezeichnet. Der Fischadler,
Pandion haliaëtos Less.; der Bussard, Buteo
vulgaris Bechst.; die Gabelweihe, Milvus regalis
Briss.; selbst der Wanderfalke, Falco peregrinus L.;
der Baumfalke, Falco subbuteo L.; der Turmfalke,
Falco tinnunculus L.; selbst der Hühnerhabicht, Astur
palumbarius Briss., und der Sperber, Astur nisus K.
und Bl., halten sich vorübergehend mit Vorliebe auf den Flussinseln und
Flussniederungen auf, weil ihnen dort der Tisch reichlich und bequem
gedeckt ist; ganz besonders finden sie zur Herbstzeit dort an Staaren
und Feldmäusen reichliche Nahrung.
Der Mauersegler, Cypselus apus L.; die Haus- und
Rauchschwalbe, Hirundo urbica L. und rustica
L., halten sich, bevor sie fortziehen, eine Zeit lang an unseren
Flüssen und in den Niederungen auf, um sich zu grossen Scharen dort
zu versammeln. Ende September oder Anfang Oktober kann man Tausende
dieser leicht beschwingten Flieger an den oben erwähnten Lokalitäten
antreffen, bis sie auf einmal über Nacht fortgezogen sind.
Während des Herbstzuges sind auf den Platen die Blaumeise,
Parus coeruleus L., der Krammetsvogel, Turdus
pilaris L., und die Weindrossel, Turdus iliacus
L., gar nicht selten anzutreffen. Selbst der grosse Würger,
Lanius excubitor L.,[S. 358] erscheint einzeln auf den alten
Weidenstümpfen, um sich nach einem leckeren Mahle umzusehen. Der
Volksmund hat ihn sehr treffend mit „Radäkster“ bezeichnet. In seinem
Äussern hat er eine entfernte Ähnlichkeit mit der Elster und er macht
an freistehenden Zweigen oft radförmige Bewegungen. Dieser im Fliegen
ziemlich ungeschickte Räuber sucht seine Beute zu überlisten. Er sitzt
bald auf den Zweigen, bald hängt er mit halbausgebreiteten Flügeln
unter denselben, bald macht er radförmige Bewegungen um dieselben.
Die kleinen arglosen Vögel werden durch dieses wunderliche Gebahren
herangelockt, setzen sich auf die benachbarten Sträucher, oft ganz in
seine Nähe, um diesem tollen Treiben zuzusehen, und im unbemerkten
Augenblicke werden sie von diesem arglistigen Räuber gefangen. Seine
erwischte Beute spiesst er oft auf Dornen, um sie dann stückweise und
nach Bedarf zu verzehren.
Auch die Familie der Finken schickt ihre Vertreter an die
Gewässer. Auf dem Frühjahrszuge bemerken wir den Flachsfinken,
Acanthis linaria Bp.; zur Herbstzeit treffen wir den
Berghänfling, Linota montium Bp.; ebenso den
Stieglitz, den muntersten und schönsten seiner Sippschaft,
Carduelis elegans Steph.; den ungeschickten, im Baue etwas
plumpen Grünling, Chlorospiza chloris Bp.; selbst
„Jochen“, der wohl mit Unrecht so sehr gescholtene Strauchdieb und
Gassenbube, der Sperling, Passer domesticus Koch, dem
sogar der Ausrottungskrieg angekündigt werden soll, mischt sich unter
die Gäste auf den Inseln und Platen. Wenn man aber diesen, allerdings
bei vielen gehassten munteren und kecken Burschen während der Brütezeit
beobachtet, wie er von Zweig zu Zweig, von Blatt zu Blatt, von Blüte
zu Blüte die Obstbäume absucht, um die heisshungrigen Jungen zu
befriedigen, die er fast ausschliesslich mit Kerbtieren und deren
Larven füttert, dann muss man ihm doch wohl etwas freundlicher gesinnt
werden, denn der Nutzen, den er dadurch unseren Obstgärten zuwendet,
ist jedenfalls ein recht bedeutender und hebt gewiss einen grossen Teil
seines Schadens auf. Nehmen sie zu sehr überhand und fügen sie später
den Kornfeldern und Erbsenäckern zu grossen Schaden zu, so möge man sie
im Herbste[S. 359] dezimieren, aber jedenfalls nicht zur Brutzeit, denn dann
gerade stiften sie Nutzen. Ebenso thöricht ist es, die junge Brut zu
zerstören, die gerade der Kerbtiere zu ihrer Nahrung bedarf.
Zu Tausenden und Abertausenden sind die Staare, Sturnus
vulgaris L., im Spätsommer und zur Herbstzeit in den
Flussniederungen, in den Rohrfeldern an Flüssen und Seen nach
beendigtem Brutgeschäfte anzutreffen. Gegen Abend sieht man sie in
dichten Wolken, bald nahe über dem Boden, bald hoch durch die Lüfte
umherziehen, dann in die grossen Rohrfelder einfallen, um dort ihre
Nachtruhe zu halten.
Ganz buntfarbig ist zur Herbstzugzeit das Bild der Sumpf- und
Schwimmvögel. Da treffen wir den nordischen Kiebitzregenpfeifer,
Squatarola helvetica Cuv.; den Goldregenpfeifer,
Charadrius pluvialis L.; den Halsbandregenpfeifer,
Pluvialis hiaticula Briss.; den gravitätisch einherstolzierenden
Austernfischer, Haematopus ostralegus L.; die rasch
über die Sandflächen dahintrippelnden Wasserläufer, Totanus
glottis Bechst., Totanus fuscus Leisl., Totanus
glareola Temm., Totanus ochropus Temm.; die zierlichen,
blitzschnellen Strandläufer, Tringa subarquata Temm.,
Tringa alpina L., Tringa minuta Leisl. und Tringa
Temminckii Leisl. Auch die stumme Bekassine, Telmatias
gallinula Boie, gesellt sich hinzu.
Truppweise in Flügen zu dreien und vieren besuchen im August und
September auch die „Unwährsvögel“, Numenius arquata Latham, ihr
Flussrevier. Im Volksmunde führt dieser Vogel den Namen „Gütvoagel“
nach seinem eintönigen Rufe, der etwa wie „tlaüd, tlaüd“ klingt,
oder „Unwährsvoagel“; denn meistens, wenn diese Vögel abends ziehen
und ihren weitklingenden Ruf ertönen lassen, giebt es schlechtes
Wetter, Unwetter. Ganz besonders zahlreich erscheinen zur Zugzeit
die Gänse, Anser cinereus W. u. M., die Graugans, und
Anser segetum Bechst, die Saatgans — vereinzelt findet
sich auch darunter die Blässgans, Anser albifrons Gm.,
und die Ringelgans, Bernicla brenta Pall. — in den
Flussniederungen und es wird in manchen Jahren eifrig[S. 360] Jagd auf die
sehr scheuen Tiere gemacht. Doch selten wird in den Ebenen die Jagd mit
Erfolg gekrönt, da die ausgestellten Posten den Jäger gewöhnlich viel
zu früh wittern. Ehe derselbe zum Schuss kommen kann, geht die ganze
Schar auf und davon. Günstiger ist der Erfolg, wenn mehrere Jäger sich
verabreden und eine solche Fläche, auf welcher sich ein Gänseschwarm
niedergelassen hat, unbemerkt umstellen können; dann treten plötzlich
an der einen Seite die Jäger vor, die Gänse streichen sofort nach
der entgegengesetzten Richtung ab und kommen nun den dort versteckt
stehenden andern in die Schusslinie.
Sehr zahlreich erscheinen auch im Herbst die Vertreter der
Entenfamilien. Da können wir an unseren Gewässern beobachten
die schöne Brand-, Fuchs- oder Höhlenente,
Vulpanser tadorna Pall.; die Schnatterente, Anas
strepera L.; die Spiessente, Anas acuta L.; die
Pfeifente, Anas Penelope L.; die Tafelente,
Fuligula ferina L.; die Reiherente, Fuligula
cristata Ray; die Bergente, Fuligula marila L.; die
schöne Schellente, Glaucion clangula K. u. Bl.; die
Eisente, Harelda glacialis Leach, und die stattliche
Trauerente, Oidemia nigra Flemm. An manchen Flüssen und
Inseln werden zur Zugzeit in eigens dazu angelegten Entenfängen oder
in Entenhütten Hunderte dieser schmackhaften Schwimmvögel erlegt und
liefern den Uferbewohnern eine nahrhafte und billige Fleischspeise.
Auch der kleine Säger, Mergus albellus L.; der
grosse Säger, Mergus merganser L., und der mittlere
Säger, Mergus serrator L., erscheinen zur Herbstzeit
auf den Binnengewässern. Einzelne Möven, Rissa tridactyla
Leach, die dreizehige Möve; Larus canus L., die
Sturmmöve; Larus argentatus Brünn., die Silbermöve
— letztere fast nur im Jugendkleide —; ganz vereinzelt die grosse,
stattliche Mantelmöve, Larus marinus L., und die
mittlere Raubmöve, Lestris pomarina Temm., halten
sich vorübergehend an unseren Binnengewässern auf. Und endlich
trifft man auch noch den rotkehligen Taucher, Eudites
septentrionalis Ill.; den Polartaucher, Eudites
arcticus Ill., sowie den gehörnten Lappentaucher oder
arktischen Steissfuss, Colymbus cornutus L.,[S. 361] zur Herbstzeit auf
den grösseren, mit Rohr bewachsenen Binnengewässern und Flüssen an.
Nachdem wir so in der Ornis Umschau gehalten haben nach dem, was
der Naturfreund an Flüssen und Seen, auf Inseln und Groden von
der befiederten Welt antreffen kann, wollen wir im Folgenden die
Kaltblüter Revue passieren lassen. Die Klasse der Reptilien
ist bald abgefertigt, denn hier kann es sich höchstens um ein Tier
handeln, welches in dem oben bezeichneten Gebiete vorkommt. Es ist
die europäische Sumpf-Schildkröte, Emys europaea Gray,
welche in den Seen und Flussgebieten des baltischen und karpatischen
Höhenrückens in Sachsen, Schlesien, Mecklenburg, Brandenburg
u. s. w. gelegentlich, aber nicht gerade sehr häufig, vorkommt. Ihre
Verbreitung im nördlichen Deutschland mag eine grössere sein, als
augenblicklich allgemein angenommen wird, da sie sich durch ihre
versteckte Lebensweise — am Grunde der Gewässer — dem Beobachter
entzieht. Meistens wird in den bekannten Gebieten ihr Vorkommen durch
frei auf der Wasseroberfläche schwimmende Fischblasen verraten. Ihre
Nahrung besteht vorwiegend aus Fischen und ist sie daher der Fischzucht
sehr schädlich. Die Vermehrung geschieht durch Eier, welche das
Weibchen ausserhalb des Wassers im Mai in selbstgegrabenen kleinen
Gruben ablegt. Aus dem Ei entwickelt sich das vollkommene Reptil ohne
Metamorphose.
Die Amphibien gehören sämtlich dem Süsswassergebiete an,
wenigstens zur Paarungszeit und in ihrem Kaulquappenzustande. Von
den fünf deutschen Froscharten, dem braunen Grasfrosch,
Rana fusca Rösel; dem Moorfrosche, Rana arvalis
Nilsson; dem grünen Wasserfrosche, Rana esculenta
L.; dem Seefrosche, Rana fortis Boulenger, und dem
Springfrosche, Rana agilis Thomas, ist der letztere
im nördlichen Deutschland noch nicht nachgewiesen worden. Ihren
Laich legen sie klumpweise ab und sind auch nur dann, während der
Begattungszeit, zahlreich bei einander und leicht zu fangen. Sie lassen
sich bei der Begattung, ohne sich zu lösen, aus dem Wasser heben, ja
zwei Pärchen von Rana arvalis, welche Verfasser in Spiritus
abtötete, liessen auch[S. 362] im Tode nicht von einander und zieren jetzt in
dieser Stellung die städtischen Sammlungen in Bremen.
Bedeutend schwieriger ist die Geburtshelferkröte, Alytes
obstetricans Wagl., zu beobachten. Diese Art lebt das ganze Jahr
hindurch in tiefen Erdlöchern, aus denen sie spät abends zum Vorschein
kommt, und nur auf ganz kurze Zeit begiebt sich das Männchen ins
Wasser, um die um die Hüften gewickelte Eierschnur, aus welcher dann
sehr bald die jungen Larven ausschlüpfen, abzustreifen. Am leichtesten
findet man ihre Verstecke, wenn man abends sich genau den Ort merkt,
von welchem der helle, flötenartig klingende Ruf, der an den Ton,
welcher durch Anschlagen an eine Glasglocke hervorgebracht wird,
erinnert, herüberschallt, und nun am folgenden Tage die Erdhöhlen,
Spalten, Steinhaufen u. s. w. in dem betreffenden Reviere untersucht.
Auch die Knoblauchkröte, Pelobates fuscus Wagl., ist
schwer zu beobachten, da sie den Tag über versteckt in der Erde lebt
und nur zur Nachtzeit hervorkommt. Zur Paarungszeit im April geht sie
ins Wasser und man trifft dann, wenn man einen solchen Laichplatz
einmal ausgekundschaftet hat — sie wählen alljährlich denselben
Laichplatz wieder —, immer eine grössere Anzahl beisammen, aber fast
immer am Boden der Gewässer. Ihren Laich setzen sie schnurweise ab.
Die Larven dieser Art sind von allen Anuren-Larven die grössten und
erreichen etwa die Länge eines Decimeters. Schon an den Larven, die
mit denen keiner anderen Art verwechselt werden können, lässt sich die
Spezies identifizieren.
Die beiden Feuerkröten, Bombinator pachypus Bonap., mit
gelbem Bauch und schwarzen Flecken, und Bombinator bombinus
L., mit blauschwarzem Bauch und fast zinnoberroten Flecken, legen wie
die Frösche ihren Laich klumpweise ab, sind auch fast ausschliesslich
Wassertiere. Man findet sie mehr in gebirgigen Gegenden als in der
Ebene, besonders in lehmhaltigen Tümpeln, welche sie, sobald sie sich
beobachtet sehen, durch Aufwühlen des Bodens trübe machen und sich
dadurch dem Auge des Beobachters entziehen.
[S. 363]
Die bekannten Vertreter aus der Familie der Bufoniden sind die
gemeine Kröte, volkstümlich „Ütze“, auch „Pädde“ genannt,
Bufo vulgaris Laur.; die Kreuzkröte, Bufo calamita
Laur., und die Wechselkröte, Bufo variabilis Pall. Nur
zur Laichzeit trifft man sie zahlreich beisammen in stehenden Gewässern
und vernimmt dann schon aus der Ferne ihren Unkengesang. Die Laichzeit
beginnt oft schon im April und dauert gewöhnlich nur kurze Zeit;
einzeln trifft man sie auch noch im Mai in der Begattung. Mit Vorliebe
wählen sie mit Rohr bewachsene Tümpel und Gräben und man kann in
denselben den Laich, welcher in Schnüren abgesetzt wird, um die Stengel
geschlungen sehen. Bei Bufo vulgaris erreichen die Laichschnüre
oft eine Länge von einigen Metern, die einzelnen Eier (oft mehrere
Tausend) sind etwas schräg gestellt und dreireihig angeordnet. Beim
Absetzen der Eier wird die Schnur bedeutend ausgedehnt und erscheint
sie dann oft zwei-, ja sogar nur einreihig. Die jungen Kaulquappen
schlüpfen gewöhnlich nach vierzehn Tagen aus und man trifft sie dann
zu Tausenden im Wasser umherschwimmend. Die Eier in der 3–4 m
langen Laichschnur von Bufo variabilis Pall. sind spiralig
angeordnet und erscheinen daher bei einer oberflächlichen Betrachtung,
als ob sie regelmässige Dreiecke bildeten. Bufo calamita Laur.
setzt eine zweireihige dünne Laichschnur ab.
Aus der Abteilung der Discodactylia gehört schliesslich noch
hierher der gemeine Laubfrosch, Hyla arborea L. Dieses
muntere und zierliche Tier trifft man gewöhnlich nur einzeln auf den
Blättern von Bäumen und Sträuchern an. Er weiss seine Farbe aber dem
Blatte, auf welchem er sitzt, so anzupassen, dass er in den meisten
Fällen von dem Beobachter übersehen wird. Anfang Mai jedoch, zur
Paarungszeit, trifft man sie in grösserer Zahl bei einander und mit
Vorliebe wählen sie alljährlich denselben Laichplatz wieder. Es sind
gewöhnlich flache, etwas lehmige Tümpel, in denen sie ihren Laich,
wie die echten Frösche, klumpweise absetzen, und auch nach beendeter
Laichzeit trifft man sie dann noch einige Tage an den Ufern im Grase
oder auf dem unmittelbar am Wasser stehenden Gesträuch.
[S. 364]
Lassen wir nun auch noch die Urodelen, welche ebenfalls zur
Laichzeit unsere süssen Gewässer bewohnen, im Geiste an uns
vorüberziehen. Wir bemerken da zuerst den stattlichsten aller
Molche, den Feuersalamander, Salamandra maculosa Laur.
In den feuchten gebirgigen Teilen unseres Vaterlandes trifft man
diesen langsamen, plumpen Molch zur Abendzeit gar nicht selten an.
Viel schwieriger ist er in unseren Ebenen, von wo auch einzelne
Aufenthaltsorte bekannt sind, aufzufinden. Bei uns treffen wir ihn
nur in den allersumpfigsten dunkeln Wäldern und dort nur unter oder
in alten halb vermoderten Baumstümpfen. Das Verbreitungsgebiet im
Nordwesten ist ein eigentümliches. Bekannt ist er z. B. aus einzelnen
feuchten Waldungen der Geest, welche auf der Wasserscheide zwischen
Ems und Weser sich befinden. Aus dem Gebiete zwischen Weser und Elbe
dagegen ist bislang kein Fundort bekannt. Es ist möglich, dass sein
nächtliches und verstecktes Leben in den feuchten Wäldern, denn nur
diese kommen für unsere Ebene in Betracht, ihn bislang noch vor dem
weiteren Auffinden in unserm Nordwesten bewahrt hat. Aus den Eiern,
welche in Waldtümpeln im Mai abgesetzt werden, schlüpfen die fast
völlig ausgebildeten Larven aus.
Eine ebenfalls eigentümliche Verbreitung haben die Tritonen speziell
in unserm Nordwesten. Die vier bekannten deutschen Arten sind sämtlich
hier vertreten. Es sind der grosse Kammolch, Triton
cristatus Laur.; der kleine Wassermolch, Triton
taeniatus Schneider; der Alpenmolch, Triton alpestris
Laur., und der Schweizermolch, Triton helveticus
Razoumowsky. Die beiden ersten finden sich zur Laichzeit im April fast
in jedem lehmigen Graben und Tümpel. Triton alpestris Laur.
dagegen ist nur ziemlich häufig an einzelnen Orten der Oldenburger
Geest; zwischen Weser und Elbe ist er nicht sicher nachgewiesen;
aus dem Hamburger Gebiete dagegen ist er wieder bekannt. Noch
eigentümlicher ist die Verbreitung unseres Triton helveticus
Razoum. Von demselben hat Verfasser nur einen Fundort, und zwar auf der
rechtsseitigen Wesergeest, aufzufinden vermocht, obgleich zur Laichzeit
seit fast zwei Dezennien viele Lokalitäten nach diesem[S. 365] seltenen Gaste
durchsucht sind. Der fadenförmige Schwanzanhang, sowie die starken
Schwimmhäute der Hinterfüsse des Männchens und die Mittelleiste
des Rückens statt des Flossensaumes bei den anderen Arten sind so
charakteristische Unterscheidungsmerkmale, dass er bei aufmerksamem
Beobachten gar nicht mit den anderen verwechselt werden kann. Die
Tritonen legen ihre Eier gewöhnlich im April, in milden Frühjahren
oft schon im März, einzeln an Pflanzen, Laub, Stengeln u. dergl. im
Wasser ab. Die Larven haben ihre volle Entwickelung gewöhnlich Ende
des Sommers erreicht, alsdann verlieren sie auch die äusseren Kiemen,
oft trifft man aber auch noch im Winter und im folgenden Frühjahre
vollständig ausgebildete Tritonen mit den äusseren Kiemen. Verfasser
traf in einem Tümpel fast alljährlich zur Laichzeit im April 6
bis 10 cm lange Exemplare von Triton cristatus Laur. mit
noch vollständig ausgebildeten äusseren Kiemen.
Den deutschen Süsswasserfischen und ihren Lebensverhältnissen
ist in diesem Bande schon eine eingehende Abhandlung gewidmet. Wenn
im folgenden auch noch wieder von den Fischen die Rede sein soll,
so geschieht es nur zu dem Zwecke, dem Naturbeobachter auch hier
ein Bild einer lokalen Fischfauna des süssen Wassers zu geben, um
daran zu zeigen, welche Arten an bestimmten Lokalitäten zu finden
sind. Zuerst möge das Bild einer Flussfischfauna, sodann das eines
Geestsees und endlich das eines Moorsees folgen. Der Fischreichtum
in unserer Weser — ich habe dabei das Gebiet etwa eine Meile unter-
und oberhalb Vegesack im Auge — ist ein ziemlich bedeutender. Von
den Barschen finden sich ziemlich häufig der Flussbarsch, Perca
fluviatilis L., sowie der Kaulbarsch, Acerina cernua Cuv.
In grossen Mengen ist im Frühjahre in allen Buchten der gemeine
Stichling, Gasterosteus aculeatus L., zu beobachten. Ebenso
wird recht häufig die schmackhafte Quappe, Lota fluviatilis
Bl., gefangen. Von den Pleuronectae ist Platessa flesus,
der kleine Wasserbutt, fast nur im ersten Jugendstadium, etwa 2–4
cm gross, vorhanden. Einzeln kommen auch den Weserfischern
der Karpfen, Cyprinus carpio L., die Karausche, Carassius
vulgaris L., die Schleihe, Tinca vulgaris Cuv., und[S. 366] die
Barbe, Barbus fluviatilis Cuv., ins Netz. Letztere gehört
allerdings mehr dem oberen Flussgebiete der Weser an. Zwischen den
einzelnen Schlengen und in den flachen Ausbuchtungen ist sehr häufig
der zierliche Gründling, Gobio fluviatilis Cuv., zu finden. Zu
den gewöhnlichsten und am meisten von den Fischern gefangenen Fischen
gehören der Alander, Leuciscus Idus Selys-Long., der Rotflossen,
Leuciscus erythrophthalmus Valenc., der Döbel, Leuciscus
Dobula Valenc., das Rotauge, Leuciscus rutilus Agassiz,
der Bresen, Abramis Brama L., und der Schnäpel, Abramis
vimba L. Auch wird einzeln der Lachs, Trutta salar L.,
und die Lachsforelle, Trutta trutta L., gefangen. In grossen
Zügen erscheint im Januar, Februar und auch noch im März der Stint,
Osmerus eperlanus Art. An Individuenzahl hat in den letzten
Jahren der Tielemann, Corregonus oxyrrhynchus L., bedeutend
abgenommen, ebenso der Maifisch, Alosa vulgaris Cuv. Letzterer
wird jetzt viel weiter weserabwärts gefangen. Häufige Bewohner der
Weser dagegen sind der Hecht, Esox lucius L., und der Aal,
Anguilla anguilla L. — Alljährlich wird in einzelnen, oft
recht stattlichen Exemplaren auch der Stör, Acipenser sturio
L., gefangen. Von den Rundmäulern sind beide Neunaugen, die Lamprete,
Petromyzon marinus L., und das Flussneunauge, Petromyzon
fluviatilis L., zu den Weserbewohnern zu rechnen. Erstere ziemlich
selten, letztere dagegen häufig. Gefangen werden die Neunaugen vom
Dezember bis zum Februar.
Zum Vergleich möge nun eine Aufzählung der Fische folgen, welche im
Zwischenahner Meere, einem auf der Geest gelegenen grossen
See, leben. Bekannt aus diesem „Meere“ sind: Der Barsch, Perca
fluviatilis L.; der Kaulbarsch, Acerina cernua Cuv.; der
Stichling, Gasterosteus aculeatus L., und der kleine Stichling,
Gasterosteus pungitius L.; die Quappe, Lota fluviatilis
Bl.; der Schlammpeitzger, Cobitis fossilis L.; der Karpfen,
Cyprinus carpio L.; die Karausche, Carassius vulgaris
L.; die Schleihe, Tinca vulgaris Cuv.; der Gründling, Gobio
fluviatilis Cuv.; der Alander, Leuciscus Idus Selys-Long.;
der Rotflossen, Leuciscus erythrophthalmus Valenc.; das Rotauge,
Leuciscus rutilus Agass.; der Brassen, Abramis Brama L.;
der Weissfisch,[S. 367]Abramis Blicca Agass.; der Lachs, Trutta
salar L.; die Lachsforelle, Trutta trutta L.; der Stint,
Osmerus eperlanus Art.; der Hecht, Esox lucius L., und
der Aal, Anguilla anguilla L.
Und endlich ein Verzeichnis der Fische des Balk-Sees, eines
mitten im Moore gelegenen 800 Morgen grossen Wasserbeckens. Häufig
werden dort der Sandard, Lucioperca sandra Cuv.; Perca
fluviatilis L., der Barsch; Lota fluviatilis Bl., die
Quappe; Tinca vulgaris Cuv., die Schleihe; Abramis brama
L., der Brassen; Esox lucius L., der Hecht; Anguilla
anguilla L., der Aal, und vereinzelt auch Cyprinus carpio
L., der Karpfen, gefangen.
Was die Mollusken anlangt, so ist denselben in diesem Bande
schon ein längerer Aufsatz gewidmet. Im folgenden soll deshalb dem
Naturbeobachter nur gezeigt werden, was er im und am Süsswasser von
dieser Tierklasse auffinden kann. Von den Nacktschnecken findet man
auf den Inseln der Flüsse, an den Ufern unter Genist und Steinen
die schwarze Theerschnecke, Arion empiricorum Fér.,
Arion fuscus Müller, den gefrässigen Limax agrestis
L. und den zierlichen braunen Limax brunneus Drap. Unter
feuchten und faulenden Weidenblättern lebt die glänzende Hyalina
nitida Müller. An den Schlengen Helix hispida L.; Helix
rubiginosa Zgl.; Helix liberta West. und besonders
häufig da, wo Aster salicifolius Scholler wächst, Helix
arbustorum L. In Gesellschaft mit Hyalina trifft man
Cionella lubrica Müll.; Cionella lubricella Zgl.;
Cionella acicula Müll.; Pupa antivertigo Drap. und
Carychium minimum Müll. An den Pflanzen am Wasser ist sehr
häufig Succinea putris L. (die Bernsteinschnecke) mit ihren
zahlreichen Varietäten, an Pflanzen im Wasser ebenso häufig Succinea
Pfeifferi Rossm. und am Boden Succinea oblonga Drap. Von
den eigentlichen Wasserschnecken findet der Naturfreund eine
ganze Reihe: zunächst die zahlreichen Limnaeen, von der grossen
Limnaea stagnalis L. bis zur kleinsten Limnaea truncatula
Müll., dann die eigenartig gewundenen Tellerschnecken,
Planorbis; die Napfschnecken, Ancylus; ferner eine
Reihe von Paludinen, Bythinien, Bythinellen und Valvaten, darunter in
der Weser und Elbe die seltene Valvata[S. 368] fluviatilis Colbeau,
ferner Neritina fluviatilis L. Von den Lamellibranchien
gehören die sämtlichen Unionen, Margaritana, Anodonten, Sphaerien und
Pisidien hierher und endlich die seit 1828 in Deutschland bekannte,
aus dem Osten eingewanderte Dreyssena polymorpha Pall. Es
würde zu weit führen, die sämtlichen Süsswasserschnecken und Muscheln
hier eingehender zu behandeln und es möge genügen, im obigen auf
die zahlreichen Vertreter dieser Gruppe hingewiesen zu haben. Eine
besondere Aufmerksamkeit verdient die neuerdings aus dem Osten
eingewanderte Süsswasserschnecke, Lithoglyphus naticoides Fér.,
welche vor einigen Jahren zuerst in der Weichsel aufgefunden wurde,
dann etwas später auch in der Umgegend von Berlin und Küstrin entdeckt
ist und schliesslich auch bei Rotterdam nachgewiesen ist. Bislang
war sie nur bekannt aus dem südöstlichen Deutschland und aus den
Flussgebieten des Schwarzen Meeres.
Werfen wir nun noch einen flüchtigen Blick auf die Vertreter der
niederen Tierklassen, so kann man sich nicht genug wundern über die
Menge und Mannigfaltigkeit der niederen Geschöpfe. In einem kleinen
Raume, den das Auge bequem mit einem Male übersehen kann, welche
Vielfachheit der Gestalten, welches Spiel der Farben, welche Emsigkeit
in ihrem Leben und Weben, welch ein Huschen und Flattern, welch
ein Springen und Hüpfen an Blüte und Strauch, auf Stein und Boden.
In dem von der Flut eben zurückgelassenen Geniste wimmelt es von
verschiedenen Käferarten. Hier sieht man die hurtigen Tachys,
dort huschen von dannen Nebria- und Elaphrus-Arten;
hier über Pflanzenstengel und Gestein, bald drüber, bald drunter
weg, eilen Blethisa, Bembidium, Panagaeus und
manch andere Arten der kleinen und kleinsten Läufer. Dort erscheinen
an der Oberfläche des Wassers Dytisciden und Hydrophiliden. Hier
tummeln sich im buntesten Durcheinander die lebendigen und nimmer
rastenden Gyriniden. Auf den Blüten der Aster und am Weidengesträuch
schwirren die Canthariden und an ebendenselben Pflanzen kriechen
die bedächtigen Curculioniden. Auf den Blüten der Umbelliferen
tummeln sich die graziösen Cerambiciden. Hier wiegen sich auf
schwankendem[S. 369] Rohr die metallisch glänzenden Donacien und dort auf
beweglichen Grashalmen schaukeln sich die oft in den herrlichsten
Farben schillernden Chrysomeliden. Auf den Blüten von Cirsium
oleraceum Scop. gaukeln bunte Falter. Raschen Fluges von Blüte zu
Blüte, nirgends lange verweilend, eilen die Sesien dahin; dort sitzt
bedächtig an einer Weide, die Flügel übereinandergeschlagen und den
Abend erwartend, der Weidenbohrer, Cossus ligniperda F., und
neben ihm läuft hastig bald auf bald ab am Stamme der Moschusbock,
Aromia moschata L. Hier schwirren über die Wasserfläche die
schlanken Agrion-Arten, dort sieht man bald am Rohr sitzend,
bald raschen Fluges dahinschwirrend die stattlichen Aeschna-
und Libellula-Arten. Über dem Wasser, besonders an warmen
Sommerabenden, treiben Hunderte von Fliegen und Mücken ihr lustiges
Spiel. Über die Wasserfläche eilen die flinken, langbeinigen
Hydrometriden und unter Steinen treiben die lichtscheuen Myriapoden
ihr Wesen, daneben hüpfen von Blatt zu Blatt, von Stein auf Stein
die bunten Thysanuren. Überall regt sich Leben, am Rohr und im Gras,
auf Blüte und Strauch, auf der Wasserfläche und am Grunde des nassen
Elements, im Geniste und auf dem Sande des Ufers; ein Leben, welches
den Naturfreund fortdauernd mit Wissbegier erfüllt und ihn anspornt,
sich den noch unerforschten Problemen desselben mit Eifer zu widmen.
Druck von J. J. Weber in Leipzig.
*** END OF THE PROJECT GUTENBERG EBOOK IE TIER- UND PFLANZENWELT DES SüSSWASSERS. ZWEITER BAND. ***

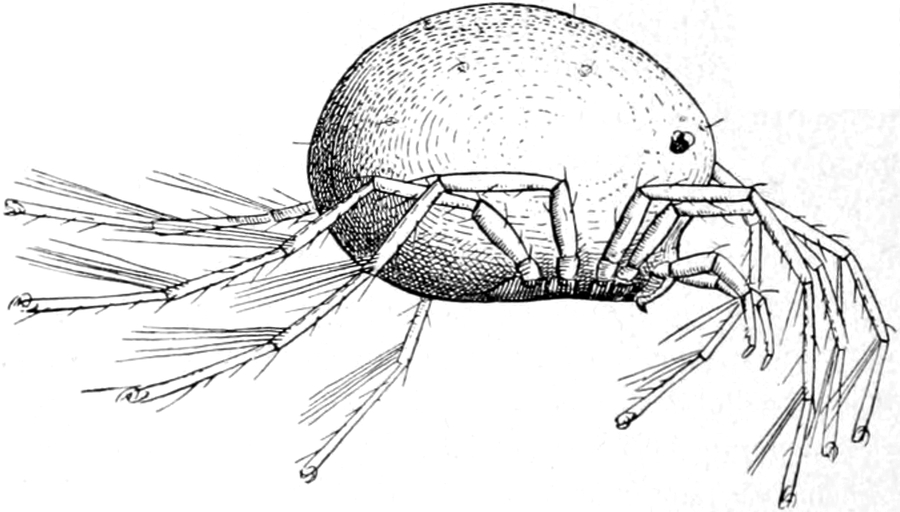
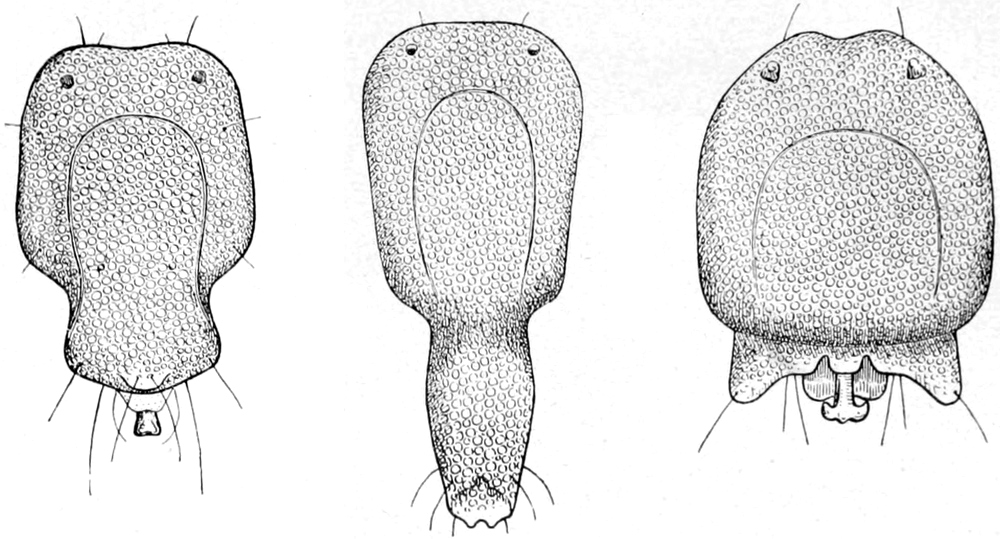




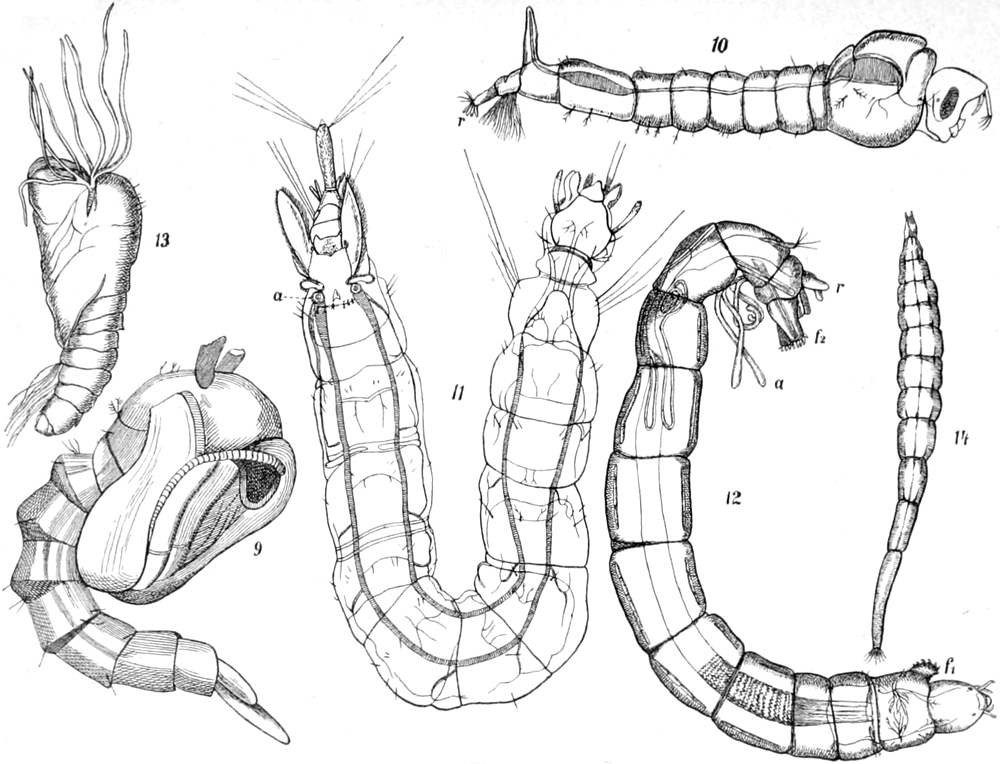
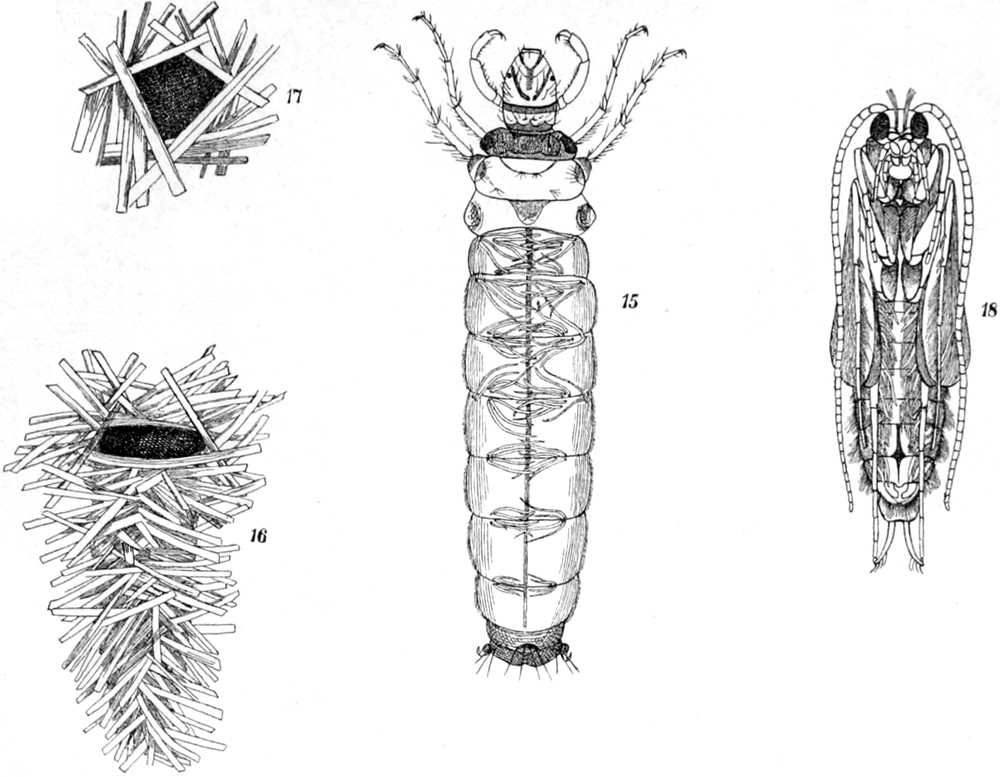















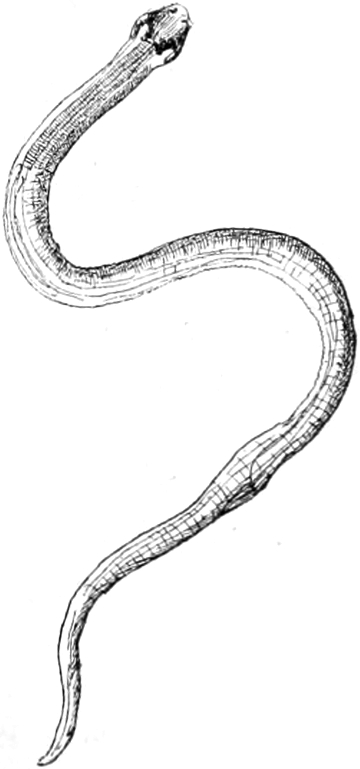




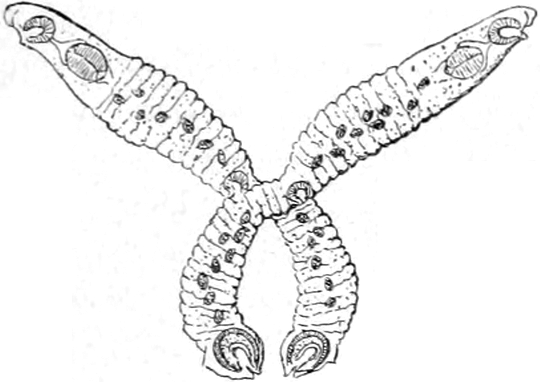





{kind=link}
{kind=link}