*** START OF THE PROJECT GUTENBERG EBOOK DIE TIER- UND PFLANZENWELT DES SüSSWASSERS. ERSTER BAND. ***
Anmerkungen zur Transkription
Der vorliegende Text wurde anhand der Buchausgabe von
1891 so weit wie möglich originalgetreu wiedergegeben. Typographische
Fehler wurden stillschweigend korrigiert. Ungewöhnliche und heute
nicht mehr verwendete Schreibweisen bleiben gegenüber dem Original
unverändert; fremdsprachliche Ausdrücke wurden nicht korrigiert.
Fußnoten wurden direkt nach dem betreffenden Absatz
eingefügt, Literaturhinweise als Endnoten dagegen am Ende eines jeden
Kapitels, geordnet nach deren Auftreten im Text. Darüberhinaus haben
einige der Autoren dort weitere nummerierte Literaturhinweise angefügt,
welche durch Zahlen gefolgt von runden Klammern gekennzeichnet werden.
Botanische und zoologische Bezeichnungen werden meist in
kursiver Schrift dargestellt, Personennamen meist
gesperrt. Dies wird
allerdings nicht durchweg konsistent gehandhabt; in der vorliegenden
Bearbeitung wurde dies auch nicht harmonisiert.
Abhängig von der im jeweiligen Lesegerät installierten
Schriftart können die im Original gesperrt
gedruckten Passagen gesperrt, in serifenloser Schrift, oder aber sowohl
serifenlos als auch gesperrt erscheinen.
Die Tier- und Pflanzenwelt des Süsswassers.
Die Tier- und Pflanzenwelt des Süsswassers.
Einführung in das Studium derselben.
Unter Mitwirkung von
Dr. C. Apstein (Kiel), S. Clessin (Ochsenfurt), Prof. Dr. F. A. Forel
(Morges, Schweiz), Prof. Dr. A. Gruber (Freiburg i. Br.), Prof. Dr.
P. Kramer (Halle a. d. S.), Prof. Dr. F. Ludwig (Greiz), Dr. W. Migula
(Karlsruhe), Dr. L. Plate (Marburg), Dr. E. Schmidt-Schwedt (Berlin),
Dr. A. Seligo (Danzig), Dr. J. Vosseler (Tübingen), Dr. W. Weltner
(Berlin) und Prof. Dr. F. Zschokke (Basel)
herausgegeben
von
Dr. Otto Zacharias,
Direktor der Biologischen Station am Grossen Plöner See in
Holstein
Erster Band.
Mit 79 in den Text gedruckten Abbildungen.
Leipzig
Verlagsbuchhandlung von J. J. Weber
1891
Alle Rechte vorbehalten.
[S. v]
Vorwort.
Der Zweck dieses Werkes ist klar im Titel desselben ausgesprochen. Es
soll zur Einführung in die Lebewelt des Süsswassers dienen und auf
möglichst kurzem Wege die Bekanntschaft mit denjenigen Pflanzen- und
Tierformen vermitteln, welche am häufigsten in unseren Tümpeln, Teichen
und Seen vorkommen. Es galt in erster Linie ein Orientierungsbuch
für den Anfänger zu schaffen und aus diesem Grunde ist von Seiten
des Herausgebers und seiner Herren Mitarbeiter eine thunlichst
gemeinverständliche Darstellungsweise angestrebt worden. Niemals ist
aber, wie der sachkundige Leser finden wird, bei Verfolgung dieses
Zieles dem wissenschaftlichen Charakter dieses Werkes Abbruch
geschehen.
Vollständig freilich in dem Sinne, dass alle Hauptgruppen der
einheimischen Wasserfauna zur Berücksichtigung gelangt wären, ist
unsere „Einführung“ nicht. Man wird die Infusionstiere, die Hydren und
die höheren Würmer vermissen. Indessen ist über diese drei Gruppen in
der Fachlitteratur sehr viel leichter Aufschluss zu[S. vi] erhalten, als
hinsichtlich der anderen Vertreter der Süsswasserfauna. Das Nämliche
gilt von den Bryozoen (Moostierchen), worüber das treffliche und
mit zahlreichen schönen Tafeln ausgestattete Werk von Prof. K.
Kräpelin („Die deutschen Süsswasserbryozoen“. Hamburg 1887) sowie
die neuere Publikation von Dr. Fr. Braem („Untersuchungen über
die Bryozoen des süssen Wassers“. Kassel 1891) jede nur wünschenswerte
Auskunft giebt.
Unser Buch wendet sich vorwiegend an Solche, welche sich
wissenschaftlich-praktisch und nicht etwa bloss litterarisch mit der
pflanzlichen und tierischen Bewohnerschaft der Seen beschäftigen
wollen. So geartete Leser und Beurteiler der nachfolgenden Kapitel
werden auch abzuschätzen vermögen, dass die Herausgabe eines nur
einigermassen umfassenden Werkes über die Flora und Fauna des
Süsswassers mit mannigfachen Schwierigkeiten verbunden war, unter denen
die Gewinnung von geeigneten Mitarbeitern obenan stand. In letzterer
Hinsicht sind aber meine Bemühungen erfolgreich gewesen.
Die zur Erläuterung des Textes beigefügten Illustrationen wurden
grösstenteils nach Originalzeichnungen der bei der Herausgabe
mitbeteiligten Herren hergestellt, und sie sind (mit Ausnahme der bloss
schematischen) von vollendeter Naturtreue. Der Opferwilligkeit der
Verlagshandlung ist es zu danken, dass mit der Beigabe von Abbildungen
nicht gespart zu werden brauchte.
Biologische Station am Plöner See (Holstein). Ende Juni 1891.
Dr. Otto Zacharias.
[S. vii]
Inhaltsverzeichnis.
Seite
I.
Allgemeine Biologie eines
Süsswassersees. Von Prof. Dr. F. A. Forel
in Morges (Schweiz).
Tiere und Pflanzen der Uferzone. — Die Flora und Fauna des Seegrundes.
— Der „organische Filz“. — Aufzählung der Tiefenbewohner
des Genfersees. — Die pelagischen Tier- und Pflanzenformen. — Mikroorganismen.
— Die chemische Zusammensetzung des Wassers. — Übergang
der organischen Materie von einem Lebewesen zum andern. — Betrag
der jährlichen Abfuhr von organischen Stoffen aus dem Genfersee durch
die Rhône. — Ersatzquellen für diesen Verlust. — Schlussfolgerungen
Einleitende Bemerkungen. — Fundstätten der Algen. — Das Vorkommen
der verschiedenen Arten nach Jahreszeit und Wasserbeschaffenheit.
— Orientierung über die 5 Hauptgruppen: 1. Spaltalgen (Schizophyceae),
2. Kieselalgen (Bacillariaceae oder Diatomaceae), 3. Grünalgen (Chlorophyceae),
4. Braunalgen (Melanophyceae), 5. Rotalgen (Rhodophyceae). —
Kurze Charakteristik der den Algen nahestehenden Armleuchtergewächse
(Characeae). — Torfmoose, Wasserfarne und Schachtelhalme
Zur Biologie der phanerogamischen
Süsswasserflora. Von Prof. Dr. Fr. Ludwig
in Greiz.
Die allgemeinen Bedingungen des höheren Pflanzenlebens im Wasser.
— Einzelne Beispiele von untergetauchten Wasserpflanzen: das gemeine
Hornblatt (Ceratophyllum demersum), die Blasenpflanze (Aldrovandia
vesiculosa), der Wasserschlauch (Utricularia vulgaris), die Sumpffeder
(Hottonia palustris), das Tausendblatt (Myriophyllum), das Laichkraut
(Potamogeton). — Schwimmgewächse: Wasserlinsen, Seerosen, die
Wassernuss (Trapa), der Wasserknöterich (Polygonum). — Die Luftpflanzen
unserer Gewässer: der Froschlöffel (Alisma Plantago), das
Pfeilkraut (Sagittaria sagittifolia), die Schlangenwurz (Calla palustris),
die Wasserschwertlilie (Iris Pseudacorus), die Rohrkolben (Typhaceae). —
Die pflanzlichen Parasiten der Wassergewächse
Ein Wurzelfüsser des Süsswassers in Bau
und Lebenserscheinungen. Von Prof. Dr. A. Gruber
in Freiburg i. Br.
Charakteristik einiger Wurzelfüsser (Difflugia acuminata, D. spiralis,
D. urceolata, Centropyxis aculeata, Hyalosphenia papilio, Arcella vulgaris,
Cyphoderia ampulla, Quadrula symmetrica und Euglypha alveolata). —
Ausführliche Schilderung der Euglypha. — Vermehrung derselben durch
Teilung. — Beschreibung der dabei stattfindenden karyokinetischen Vorgänge.
— Die Einkapselung (Encystierung) der Euglypha. — Der Kunsttrieb
derselben und ihrer Verwandten. — Beweis für die Einheit der
belebten Natur
Die Flagellaten (Geisselträger). Von
Dr. W. Migula in Karlsruhe.
Stellung dieser Organismengruppe auf der Grenze zwischen Tier und
Pflanze. — Einzeln lebende und koloniebildende Flagellaten. — Bau
und Entwickelungsgeschichte von Volvox aureus (Kugeltierchen). — Die
Volvocineen-Gattungen Eudorina, Pandorina, Stephanosphaera und
Gonium. — Euglena viridis. — Anthophysa vegetans. — Die Dinobryen. —
Die Gattung Ceratium
Die Süsswasserschwämme (Spongilliden).
Von Dr. W. Weltner in Berlin.
Erkennen und Auffinden derselben. — Geschichtliches. — Äussere
Beschaffenheit (Form, Grösse, Färbung, Konsistenz und Geruch) der
Spongilliden. — Anatomie und Histiologie: das Skelett und der Weichkörper.
— Physiologie: geschlechtliche und ungeschlechtliche Fortpflanzung,
Atmung, Nahrungsaufnahme, Verdauung, Wachstum und
Bewegung. — Kurze Schilderung der einheimischen Arten: Euspongilla
lacustris, Spongilla fragilis, Trochospongilla erinaceus, Ephydatia
Mülleri, Eph. fluviatilis, Eph. bohemica und Carterius Stepanowi. —
Schlüssel zur Bestimmung der europäischen Formen. — Verbreitung
der Süsswasserschwämme. — Parasiten und Kommensalen derselben. —
Das Sammeln, Konservieren und Untersuchen der Spongilliden
Die Strudelwürmer (Turbellaria).
Von Dr. O. Zacharias in Plön (Holstein).
Anatomische und histiologische Orientierung über die beiden Hauptgruppen
der Süsswasserturbellarien: Rhabdocoela und Dendrocoela. —
Das Genus Bothrioplana als Verbindungsglied zwischen diesen Gruppen.
— Bothrioplana silesiaca. — Kurze Beschreibung einiger Rhabdocöliden-Spezies:
Macrostoma viride, Microstoma lineare, Stenostoma leucops,
Stenostoma unicolor, Catenula lemnae, Mesostoma viridatum und Vortex
truncatus. — Präparationsmethode. — Geographische Verbreitung
Die Rädertiere (Rotatoria).
Von Dr. L. H. Plate in Marburg.
Fundstätten für Rädertiere. — Eingehende anatomische Analyse des
Krystallfischchens (Hydatina senta). — Vergleichende Schilderung der
Morphologie der Rotatorien: die Körpergestalt, die Körperhaut, der
Räderapparat, die Muskulatur, das Nervensystem, der Verdauungskanal,
die Exkretionsorgane, die Klebdrüsen, der Keimdotterstock und die Eibildung.
— Die männlichen Rotatorien. — Einige Bemerkungen über die
Biologie der Rädertiere. — Überblick über das System der Rotatorien
Die Krebsfauna unserer Gewässer.
Von Dr. J. Vosseler in Tübingen.
Überblick über das System der Krustaceen. — Die Entomostraken:
a) Freilebende Copepoden. — Kurzer geschichtlicher Rückblick. —
Körperform und Gliedmassen. — Histologie der Haut und die Häutung.
— Nervensystem und Sinnesorgane. — Verdauungskanal. — Leber und
Niere. — Blut und Blutkreislauf. — Atmung. — Muskulatur. — Fortpflanzungsorgane
und geschlechtlicher Dimorphismus. — Embryonale und
postembryonale Entwickelung. — Gattungen und Arten. — Biologische
Bemerkungen: Die Nahrung und deren Aufnahme. — Feinde. —
Fundstellen. — Verbreitung in horizontaler und vertikaler Richtung. —
Anpassung an die Verhältnisse der Ufer-, pelagischen und Tiefenregion
unserer Seen. — Passive Wanderung. — Entstehung der Tiefenfauna. —
b) Parasitische Copepoden. — Die durch den Parasitismus bedingten
Veränderungen in der Körperform und Umbildung der Gliedmassen. —
Innere Organisation: Nervensystem und Sinnesorgane, Herz, Darm und
Geschlechtsorgane. — Zwergmännchen. — Vorkommen. — Gattungen und
Arten. — c) Kiemenschwänze: Beschreibung des Argulus. — Schaden
und Nutzen der Entomostraken. — d) Phyllopoden, Cladocera: Gestaltung
des Körpers und der Gliedmassen. — Anatomie der Eingeweide,
Geschlechtsorgane und des Nervensystems. — Eier (Dauereier) und Entwickelung.
— Gattungen und Arten. — e) Branchiopoda: Beschreibung des
Apus und Branchipus und deren Lebensweise. — f) Ostracoden: Schale
und Gliedmassen. — Anatomie. — Fortpflanzung. — Lebensweise. —
g) Malacostraken.Isopoden: Körperform und Gliedmassen von Asellus.
— Darm und Kaumagen. — Leber. — Herz. — Kiemen. — Nervensystem
und Sinnesorgane. — Brutraum. — Auf dem Lande lebende
Asseln. — h) Amphipoden: Gliederung und Form des Körpers. — Gliedmassen
mit Kiemenanhängen. — Darm mit Leber. — Nervensystem. —
Herz und Gefässe. — Fortpflanzung. — Vorkommen. — i) Decapoden:
Morphologie des Körpers und der Gliedmassen von Astacus. — Äussere
Kennzeichen der Geschlechter. — Nervensystem und Sinnesorgane. —
Ohr. — Häutung. — Herz und Blutgefässe. — Kiemen. — Verdauungskanal
mit Kaumagen, Kalkansammlungen und Leber. — Geschlechtsorgane
und Fortpflanzung. — Einige biologische Bemerkungen über
Nahrung und Wiederergänzung verloren gegangener Gliedmassen
Ein See ist ein Mikrokosmus, eine Welt, die sich selbst genügt, in
welcher das Lebensspiel der verschiedenen Organismen sich hinreichend
im Gleichgewicht hält, um ein stabiles Verhältnis zwischen den
ausgeschiedenen und nutzbar gemachten Stoffen zu bilden, ohne dass
die Zusammensetzung des Mediums durch die in ihm wohnenden Wesen eine
Veränderung erlitte. Tiere und Pflanzen, höhere und niedere Organismen,
leben da gleichzeitig mit einander, jedes nach seiner Art, und gemäss
den ihm eigentümlichen Funktionen; jedes findet in dem Medium, von
dem es umgeben ist, die zur Lebensfristung notwendigen Elemente,
und jede Gruppe von Wesen vervielfältigt sich in Individuen, die um
so zahlreicher sind, in je grösserer Fülle die ihr unentbehrlichen
Elemente vorhanden sind.
Auf der andern Seite ist ein Süsswassersee kein ganz geschlossenes
Bassin, kein verschlossenes Gefäss. Vielmehr steht er in Verbindung
mit der übrigen Welt, sei es durch die atmosphärische Luft, welche
einen unaufhörlichen Austausch von Gasen mit ihm unterhält, sei es
durch seinen Abfluss, der ihm Wasser mit Substanzen in gelöstem und
ungelöstem Zustand entführt, sei es durch seine Zuflüsse, die ihm neue
Stoffe zuleiten. Er nimmt also an dem grossen Kreislauf der Materie
teil, der zwischen den verschiedenen Regionen des Erdkörpers besteht,
ebensogut in der organischen, wie in der anorganischen Welt. Zugang
und[S. 4] Verlust von Stoffen modifizieren die biologischen Verhältnisse des
Mikrokosmus; sie müssen konstatiert und ergründet werden, wenn man das
Spiel des Lebens in einer so abgegrenzten Welt verstehen, wenn man den
Kreislauf der Materie innerhalb der beschränkten Reihe von Wesen, die
den See bevölkern, bestimmen will.
Unter diesem doppelten Gesichtspunkt betrachten wir einen
Süsswassersee. Die vorliegende Studie soll als Einleitung in die Fragen
der allgemeinen und speziellen Biologie dienen, welche man in den
ferneren Abteilungen dieses Buches ausgeführt findet.
In erster Linie ist ein See ein beschränkter Raum, in welchem die
Lebewesen in angemessenen Proportionen sich entwickeln, derart, dass
ihre Ernährungsfunktionen in einer stabilen Weise im Gleichgewicht
bleiben. Während das Spiel des animalischen Lebens zuletzt zur
Oxydation der organischen Stoffe führt, welche zeitweilig in die
Körper der Tiere behufs Ernährung ihrer Gewebe eindringen, wird die
Kohlensäure reduziert durch das Spiel des vegetabilischen Lebens
infolge der dabei vorherrschenden Funktionen des Chlorophylls und
gleichartiger Substanzen, und sie wird in Stoffe verwandelt, die
den animalischen Organismen assimilierbar sind. Das Tier scheidet
Kohlenwasserstoffverbindungen und Stickstoff aus, nach dem Stande der
maximalen Oxydation und unter einer Form, welche es ihnen erlaubt, sich
in der sie umgebenden Flüssigkeit aufzulösen (Kohlensäure, Harnstoff,
gallenartige Substanzen etc.). Die Pflanze absorbiert diese durch das
Tier ausgeschiedenen Stoffe und eignet sie sich unter der Form des
Protoplasmas und der Kohlenwasserstoffverbindungen an. Die Pflanze
nährt sich also von den durch das animalische Leben aufgelösten Stoffen
ihrer Umgebung; das Tier hingegen von den Stoffen, die sich in den
Geweben der Vegetabilien bilden.
Wenn wir nun in einem abgegrenzten Raum Tiere und Vegetabilien in
normalen Verhältnissen zusammen antreffen, so halten sich diese beiden
entgegengesetzten Arten von Lebewesen im Gleichgewicht, und ein Reich
entspricht, jedes auf seine Rechnung, den[S. 5] biologischen Bedingungen
des andern. Das ist auch in einem See der Fall; die beiden organischen
Reiche sind hier repräsentiert durch reich entwickelte Tier- und
Pflanzengesellschaften, welche wir nach ihrem Wohnort und nach den
Verhältnissen des Raumes, denen sie unterworfen sind, in drei Gruppen
teilen können: Gesellschaften am Ufer, in der tiefen Region, und in
solche, die das freie Wasser bewohnen.
Die littoralen Gesellschaften der Tier- und Pflanzenwelt
befinden sich in der Zone, die sich dem Ufer entlang hinzieht, rings
um den See vom eigentlichen Uferrande, bis zu einer Tiefe von 5–25
m, je nach der Grösse des Sees. Je grösser der See ist, desto
tiefer steigt die Uferregion hinab.
Die Eigentümlichkeiten des Raumes, welche diese Region
charakterisieren, sind: felsiger, mit Kieseln bedeckter, sandiger oder
schlammiger Boden, beleuchtetes Wasser mit veränderlicher Temperatur,
je nach den Jahreszeiten; schwacher Druck; beträchtliche Bewegungen,
die durch Wellen oder Wasserströmungen hervorgerufen werden. Es ist
dies die Gegend, die vermöge der Veränderlichkeit des Bodens und der
wechselnden Verhältnisse der Wasserbewegung am reichlichsten mit
Abwechselung bedacht ist. Hier sind die Veränderungen der Temperatur
und des Lichtes am stärksten; hier können Pflanzen und Tiere die
verschiedenartigste Umgebung finden, welche die mannigfaltigen
Bedürfnisse der verschiedenen Typen befriedigt. Hier ist auch die
Pflanzen- oder Tierwelt am reichlichsten nach Menge und Abwechselung
vorhanden.
Man trifft hier, was die Pflanzen anlangt, alle Gruppen von
Wasserpflanzen, die fähig sind, sich dem lakustrischen Leben
anzupassen. Ohne eine genaue Aufzählung geben zu wollen, gruppiere ich
sie folgendermassen:
die grossen Gramineen und Cyperaceen (Schilfrohr und Cypergras),
deren straffe Stengel durchs Wasser hindurch aufsteigen, um die
Kronen ihrer Blüten und Blätter in der freien Luft sehen zu lassen;
[S. 6]
die Wälder der grossen Phanerogamen, Nymphaeaceen, Potamogetaceen,
Halorageen, Ceratophylleen, Hydrocharideen etc., deren lange,
biegsame Stengel und zierliche Blätter schattige und pittoreske
Büsche bilden, welche sich bis zur Oberfläche des Wassers erheben;
die dichten und dunkeln Rasen der Characeen. Dieselben bilden
gegen das Eindringen der Fische fast unzugängliche Schutzorte. Die
Teppiche von festen Algen (Chladophora, Ulothrix,
Leptothrix, Oscillarieen), welche die unter Wasser
befindlichen Steine und Hölzer mit einem samtweichen, grünlichen,
dichtanliegenden Überzug bekleiden. Die inkrustierenden, in
Kalkwasser lebenden Algen, Zonotrichia calcivora,
Hydrocoleum calcilegum, bilden eine sehr interessante
Abwechselung in diesem Teppich; unter ihrer Inkrustation stellen
die Kalksteine vertiefte Skulpturen dar, deren Deutung noch ein
ungenügend gelöstes Problem ist;
die niederen freien Algen, Diatomaceen, Desmidiaceen,
Vaucheriaceen, Palmellaceen etc., die mit einer leichten,
unbeständigen Schichte die verschiedensten Körper unter Wasser
bedecken und ihnen eine im allgemeinen bräunliche Farbe geben;
endlich die schwimmenden Algen: Protoderma,
Conferva, Pandorina etc., welche auf einmal in gewissen
Jahreszeiten und an gewissen Stellen erscheinen, um schliesslich
wieder zu verschwinden, unter Bedingungen, welche die Botaniker uns
noch nicht genug erklärt haben.
Von Tieren findet man hier alle die, welche das lakustrische Leben
annehmen: Wasservögel, Reptilien, Amphibien, Fische, Insekten,
Hydrachniden, Entomostraken, Mollusken, Würmer, Hydren, Spongien, bis
hinab zu den untersten Protozoen. Das ist die klassische „lakustrische
Fauna“ der alten Schriftsteller, die einzige, welche man vor den
Untersuchungen der letzten dreissig Jahre kannte. Diese Fauna, deren
Arten von einem See zum andern ziemlich verschiedene Varietäten
darbieten, kann man wieder in[S. 7] partielle oder lokale Faunen einteilen,
im wesentlichen, je nach der Bodenbeschaffenheit und nach der
Vegetation, die der See besitzt. Man kann so unterscheiden die Fauna
der Felsenwände, die der mit Geröllen bedeckten Ufer, ferner die Fauna
des Sandbodens, des Schlammes, die des im Wasser wachsenden Waldes, die
des Rasens von Characeen etc. Ich mache hier nicht den Versuch, die
Arten und Gattungen aufzuzählen, welche diese allgemeine und lokale
Tierwelt bilden; ich verweise auf die ausführlichen Beschreibungen,
welche die gelehrten Mitarbeiter dieses Werkes geben werden.
Ich beschränke mich darauf, zunächst den unumgänglichen Charakter zu
bestimmen, welchen die littorale Fauna darbieten muss im Vergleich
mit anderen Faunen der lakustrischen Welt. Die Ufertiere, welche viel
beweglicheren und abwechslungsreicheren Bedingungen unterworfen sind
als diejenigen von anderen Regionen, sind und müssen in Wirklichkeit
viel stärker, thätiger und widerstandsfähiger sein. Sie setzen sich
am Boden oder auf untergetauchten Körpern fest, oder sind fähig,
sich zeitweilig festzusetzen; oder sie wissen sich zurückzuziehen an
verborgene und geschützte Plätze während der Wellenströme, welche
die Ufergegend aufwühlen. Ich mache sodann auf die bedeutende
Veränderlichkeit und den sehr speziellen Lokalcharakter der littoralen
Gesellschaften aufmerksam; sie sind verschieden von einer Station zur
andern und von einem See zum andern, je nach den örtlichen Bedingungen.
In der Uferregion findet man nicht nur den grössten Reichtum an
Lebewesen, sondern auch die grösste Zahl von verschiedenen Typen und
die grösste Mannigfaltigkeit innerhalb der Typen.
Endlich ist es die einzige Region eines Süsswassersees, wo die Flora
durch Pflanzen von hohem Wuchs vertreten ist.
Die biologischen Gesellschaften der tiefen Region, die
Tiefsee-Fauna und -Flora, wohnen auf und in dem Seeboden,
in der gesamten Mulde vor der Uferregion, das heisst in Tiefen,
welche 5–25 m übersteigen, je nach der Grösse des Sees. In
dieser Region ist der Boden überall lehmig oder schlammig,[S. 8] weich,
ohne Rauhigkeiten oder feste und harte Körper; bei bemerkenswerter
Einförmigkeit in der physikalischen Zusammensetzung zeichnet er sich
durch keine Abwechselung aus, ausser etwa an einigen sehr seltenen
Stellen, und in einigen Seen nur durch felsige, vertikale Wände oder
durch erratische Blöcke. Abgesehen von diesen Ausnahmsfällen ist
die Gesellschaft, welche dort wohnt, wesentlich und einzig limikol
(schlammbewohnend). Es herrscht eine absolut und relativ beinahe
vollständige Ruhe ohne mechanische und molekulare Bewegungen, ohne
Wellen, ohne wesentliche Strömungen, ohne Licht, ohne Wärme. Die
Veränderungen der Temperatur sind hier entweder gleich Null oder nur
sehr schwach. Die Dunkelheit ist mehr oder weniger vollständig; es
giebt keine direkte Verbindung mit der äusseren Luft; der See zeigt
hier keine periodischen Veränderungen, weder für einzelne Jahreszeiten
noch für das ganze Jahr. Nächst dem Meeresgrund ist der Grund eines
Sees die am wenigsten bewegte Gegend, die man auf dem Erdball finden
kann.
In dieser Region konstatieren wir eine noch ziemlich reichliche
Tierwelt, die mehr oder weniger allen Typen der Süsswassertiere
angehört. Als Beispiel will ich eine Übersicht über die Tierarten
anführen, die man aus der Tiefe des Genfersees kennt, ohne der
Endoparasiten zu gedenken.
Fische
14
Wirbeltiere
14
Insekten
3
Spinnen
9
Crustaceen
16
Arthropoden
28
Gasteropoden
4
Lamellibrancheen
2
Mollusken
6
Anneliden
4
Nematoden
3
Cestoden
1
Turbellarien
18
Bryozoen
1
Rädertierchen
3
Würmer
30
Hydroiden
1
Cölenteraten
1
[S. 9]
Infusorien
10
Rhizopoden
19
Cilio-Flagellaten
2
Protozoen
31
Das sind mehr als hundert Tierarten.
Ihr Reichtum vermindert sich in dem Masse, als die Tiefe zunimmt; aber
wir haben solche, die am Grunde der tiefsten Seen aufgefischt worden
sind: im Genfersee in einer Tiefe von 309 m (Forel), im Comersee
bei 415 m (Asper), im Baikalsee bei 1370 m (Dybowski).
Der Ursprung dieser Tierwelt ist zu suchen in der Ansiedelung von
Individuen, welche sich in diese tiefe Region verirrt haben, die durch
einen Zufall weit von ihrem gewöhnlichen und ursprünglichen Wohnort
weggeführt wurden und die, weil widerstandsfähig gegen diese immerhin
starke Veränderung ihrer Lebensbedingungen, sich vermehrt und eine
Stammkolonie gebildet haben. Die Armut des Mediums und seine relative
Ruhe haben die ursprünglichen Typen ziemlich bedeutend modifiziert und
kleinere, schwächere Varietäten geschaffen. Was den Ursprung dieser
Tierwelt in der Tiefe betrifft, so findet man denselben für die weitaus
überwiegende Mehrzahl in der littoralen Fauna des Sees selbst, für
einige Arten in der Fauna der unterirdischen Wasser, der Höhlungen des
Festlandes, dessen Verbindungen mit dem See dadurch bewiesen werden.
Die Tiefen-Fauna der verschiedenen Seen hat gleichartigen
allgemeinen Charakter, ein Ergebnis der Gleichartigkeit des Mediums,
das in allen Seen beinahe identisch ist; ihre spezielle Zusammensetzung
wechselt von einem See zum andern, und richtet sich nach den Ufertypen,
von denen sie herstammen, oder nach den lokalen Zufällen, welche die
Bevölkerung in die untere Region gebracht haben.
Die Pflanzenwelt der tiefen Region ist sehr wenig entwickelt.
Die Rasen der Characeen hören auf in einer Tiefe von 20–25 m,
an der Grenze der littoralen und der tiefen Region. In der obern Zone
dieser letzteren bis zu 100 m abwärts, findet man noch an[S. 10] der
Oberfläche des Schlammes eine bräunliche Schicht von niederen Algen
(Palmellaceen, Diatomeen, Oscillarien), welche den von mir so genannten
organischen Filz darstellen und welche unter der Wirkung des
schwachen Lichtes, das in diese Schichten eindringt, noch ein gewisses
Reduktionsvermögen besitzen. Die felsigen Partien der tiefen Region
sind bisher nur an einem Punkte studiert worden, nämlich an der
unterseeischen Moräne von Yvoire im Genfersee in einer Tiefe von 60
m. Wir haben hier ein aquatisches Moos gefunden, Thamnium
alopecurum, Schimper, var. Lemani J. B. Schnetzler,
sehr reich chlorophylliert und in schönem Wachstum. Es ist sehr zu
wünschen, dass analoge Forschungen an anderen Orten gemacht werden, wo
Felsenwände vorkommen, damit man durch Erfahrung entscheiden kann, ob
es wirklich ein Ausnahmefall ist, wie man bisher geglaubt hat; oder
ob die Abwesenheit von grünen Pflanzen, die in einer Tiefe von mehr
als 25 m allgemein zu sein scheint, einzig von der schlammigen,
inkonsistenten Beschaffenheit des Bodens abhängt und sich durch das
seltene Vorkommen von festen Körpern, auf welchen sich die fraglichen
Moose festsetzen könnten, erklären liesse.
Die pelagische Gesellschaft bewohnt die allgemeine, unbestimmte,
unbegrenzte Masse des Sees, von der Oberfläche an bis zum Grund, vom
Rande der Uferregion bis in die Mitte des Sees, in seiner ganzen
Ausdehnung, soweit er nicht in unmittelbarer Berührung mit dem Ufer
oder dem Grunde steht. Diese Gesellschaft besteht aus Schwimmtieren
und schwebenden Algen. Diese Organismen gehören einer kleinen Zahl von
Arten an; aber die Zahl der Individuen ist enorm. Von Tieren sind es
einige Arten von Fischen (Coregonen), Entomostraken, Rädertierchen,
Cilio-Flagellaten, Rhizopoden; von Pflanzen einige grüne Algen und
Diatomeen.
In der eigentlichen pelagischen Region ist die oberste Schicht, welche
mit der Luft am meisten in Berührung steht, auch die am reichsten
bevölkerte. Doch haben die Untersuchungen von Asper, Imhof und eigene
Beobachtungen bewiesen, dass die pelagische Fauna auch in den grössten
Tiefen unserer Seen noch gut vertreten ist. Es giebt keine Region, wo
das Leben gänzlich aufhört.
[S. 11]
Einige dieser Wesen, speziell die Entomostraken, zeigen tägliche
vertikale Wanderungen; bei Nacht kommen sie an die Oberfläche des
Wassers herauf, bei Tag steigen sie von der helleren Zone wieder in die
unteren, finsteren Schichten hinab. Dort finden sie in der Dunkelheit
Schutz gegen die Jagd, welche ihre Feinde auf sie machen.
Die Verhältnisse des Mediums der pelagischen Region variieren mit
der Tiefe hinsichtlich des Drucks, der Bewegung des Wassers, der
Temperatur und des Lichtes. Eine einzige, überall sich gleichbleibende
Eigenschaft ist die Abwesenheit von festen Körpern, welche etwa
den Organismen einen Punkt zum Festsetzen oder einen Zufluchtsort
gewähren könnten. Von diesem Leben in einem unbegrenzten Medium
ohne feste Körper rührt es her, dass fast alle pelagischen Wesen
schwimmen oder im Wasser schweben; indessen haben wir einige an
den Ort gebundene Tiere zu nennen, nämlich unter anderen einige
Vorticellen, die sich an schwimmende Algen oder an Pflanzenreste
heften, welche von der Oberfläche des Wassers abgetrieben haben, sowie
einige Ektoparasiten der Fische und der pelagischen Entomostraken. Es
folgt aus der Durchsichtigkeit des Wassers in der pelagischen Region,
dass die meisten pelagischen Organismen, die Entomostraken und die
Rädertierchen insbesondere, durchsichtig sind; für gewisse Arten ist
die Durchsichtigkeit absolut. Man kann annehmen, dass dieser Mangel
an Farbe durch natürliche Zuchtwahl in einer Reihe von Generationen
erworben wurde; er ist in der That ein sehr wirksames Mittel, sich
gegen die Verfolgung von seiten der fleischfressenden Tiere zu
schützen; für letztere wiederum ist dieser Mangel an Farbe insofern
nützlich, als sie dadurch der Aufmerksamkeit ihrer Beute entgehen.
Während die littoralen Gesellschaften in jedem See ihre
Eigentümlichkeiten haben und aus verschiedenen Arten und Varietäten
zusammengesetzt sind, je nach den verschiedenen lokalen Verhältnissen
an jeder Station, und während die Tiefseefauna, gemäss ihrer Herkunft
von den littoralen Organismen, in jedem See lokalen Charakter hat,
eine Folge der absoluten Unabhängigkeit ihrer Differenzierung,
die sich auf dem Grunde eines jeden Sees als in einem speziellen[S. 12]
„Schöpfungszentrum“ vollzogen hat, zeigen die pelagischen
Gesellschaften einen sehr ausgesprochenen kosmopolitischen Charakter.
Man findet die gleichen Arten, die gleichen Varietäten in allen
Gewässern des Kontinents wieder. Die einzige Verschiedenheit, welche
man konstatieren kann, ist das Fehlen von einigen Typen in gewissen
Seen. Es ist wahrscheinlich, dass diese weite Verbreitung der
pelagischen Organismen durch passive Migrationen von einem See zum
andern erklärt werden muss, die von einem Transport durch die Federn
und Füsse und in den Gedärmen der Wandervögel herrühren. Während diese
auf der Oberfläche des Wassers schwimmen, nehmen sie die grösstenteils
sehr widerstandsfähigen Keime von pelagischen Tieren und Pflanzen auf
und tragen sie auf ihren periodischen Wanderungen durch die Lüfte
von See zu See. Das Fehlen von einigen Arten in gewissen Seen muss,
wenn es sich nicht durch eigentümliche Verhältnisse des Mediums, bei
Bergseen z. B. durch die hohe Lage, erklären lässt, als das Resultat
von zufälligen Umständen bei dieser Art der Besiedelung durch passive
Wanderung angesehen werden.
Ausser diesen Tier- und Pflanzengesellschaften mit einer
verhältnismässig höhern Organisation, die sich so in drei Gruppen
teilen, nach den Verhältnissen der Regionen, welche sie bewohnen, haben
wir noch im Wasser der Seen das Leben von elementaren Mikroorganismen
zu verzeichnen, Mikroben von der Gruppe der Schizomyceten, Pilze,
Bakterien, Vibrionen etc. Man trifft sie überall, in allen Schichten
von der Oberfläche bis zum Seegrund, vom Ufer bis in die Mitte des
Sees, in den littoralen, pelagischen und Tiefengewässern; in dieser
Hinsicht unterscheiden sich die Wasser der Seen nicht von den anderen
freien Gewässern, wo die Mikroben schwärmen. Ihre Zahl ist hier sehr
beträchtlich, doch weniger gross als in den meisten anderen natürlichen
Gewässern. Während man in dem Quellwasser der Ebene oft ihre Keime nach
tausenden zählt, ist ihre Zahl in dem helleren Wasser des Genfer- oder
Zürichersees von den Herren Fol, Dunant und Cramer durchschnittlich auf
36–38 Keime oder Kolonien pro Kubikcentimeter Wasser bestimmt worden.
[S. 13]
Die biologische Funktion dieser Mikroben, ihre Rolle in der lebenden
Welt besteht darin, die Zersetzung, die Auflösung der animalischen
und vegetabilischen Kadaver, die nicht direkt von den Tieren verzehrt
werden, zu bewirken. Sie sind so die Agenten der Verwesung und bewirken
die Umbildung der organischen unauflöslichen Materie in Substanzen, die
im Wasser lösbar sind.
Es ist schwer, in allgemeiner Weise anzugeben, welches die nächsten
Glieder dieser Umsetzung sind; die letzten Glieder sind Kohlensäure,
Ammoniak, Salpetersäure und Stickstoff.
Wir haben also im Wasser eines Sees zahlreiche und verschiedene
biologische Gesellschaften, Tiere und Pflanzen, höhere Organismen und
Protisten, die neben einander leben, absorbieren und ausscheiden, die
aber alle sich von den von ihren Nachbarn ausgestossenen Produkten
nähren. Alle assimilieren, jedes in seiner Art, die zur Ernährung
der Gewebe nötigen Stoffe; alle stossen mit ihren Exkretionen die
Residuen ihres Ernährungslebens aus. Was so vielen verschiedenen Wesen
gestattet, neben einander zu leben und im gleichen Medium gleichzeitig
zu existieren, ohne dessen Vorräte zu erschöpfen, das ist die wichtige
Thatsache, dass ihre Produkte und Bedürfnisse entgegengesetzt sind und
zwischen den verschiedenen Gruppen sich im Gleichgewicht halten. Was
von den einen ausgeschieden wird, ist für das Leben der anderen nötig.
Die Residuen des Ernährungsprozesses der einen Gruppe werden nutzbar
für den Ernährungsprozess der andern. Die proportionelle Verteilung
der verschiedenen Typen von animalischen und vegetabilischen Wesen
regelt sich von selbst durch einen automatischen Prozess: Ein Überfluss
von Ernährungsstoffen begünstigt die überreichliche Entwickelung
von Wesen, welche sich dieselben nutzbar machen können; ein Defizit
solcher Materien führt infolge der Not eine Verminderung der nämlichen
Organismen herbei.
Das biologische Gleichgewicht ist in einem See also dadurch
möglich, dass die verschiedenen Arten, die ihn bewohnen, verschiedene
Typen der entgegengesetzten organischen Reiche[S. 14] repräsentieren. In
der littoralen und in der pelagischen Region haben wir gleichzeitig
Repräsentanten von beiden Reichen, Pflanzen und Tiere; vermöge dieses
Gleichgewichtes finden die Ernährungsfunktionen der einen wie der
anderen dort hinlänglich, was sie brauchen. Nicht ganz so, scheint es,
verhält es sich in der Tiefenregion, speziell in den unteren Schichten
von 100 m an abwärts, wo wir keine Pflanzen mehr kennen. Wie
können in diesen Tiefenregionen die zur Ernährung nötigen Elemente
für die noch reiche Fauna, die dort wohnt, sich erneuern? Eine reiche
und genügende Quelle dieser Ernährung zeigt sich in den Kadavern der
animalischen und vegetabilischen Organismen der pelagischen Region,
welche in die Tiefe fallen und auf den Grund des Sees hinabkommen. Wir
haben Anzeichen davon in der enormen Zahl von chitinösen Häuten der
pelagischen Entomostraken, welche sich im Schlamm der grossen Tiefen
zeigen. Die organischen Reste des Ufers und diejenigen, welche durch
die Zuflüsse in den See hineingetrieben werden, sinken, nachdem sie
durch die Wellen und Strömungen auf der Oberfläche umhergetrieben
worden sind, von selbst in die grossen Tiefen hinab und tragen so
ebenfalls zur Erneuerung des Nahrungsstoffes für die Tiefenfauna bei.
Wie in dem Tierkörper die verschiedenen Gewebe der verschiedenen Organe
aus der Lymphe des Blutes die zu ihrer Ernährung notwendigen Stoffe
ziehen und der Lymphe die Produkte ihrer Desassimilation geben, so ist
in einem See das Wasser das Medium, in welchem alle diese Reaktionen
des Ernährungsprozesses für die darin wohnenden Organismen vor sich
gehen. Die chemische Zusammensetzung dieses Wassers bietet
also ein grosses Interesse dar. Als Beispiel will ich diejenige des
Genfersees geben, die bis jetzt am besten studiert ist.
Ein Liter Wasser vom Genfersee enthält in aufgelöstem Zustand:
In Gasform:
Sauerstoff
6.65
cc
Stickstoff
14.69
„
Kohlensäure
2.85
„
[S. 15]
An festen Substanzen:
Natrium- und Kaliumchlorid
1.8
mg
Schwefelsaures Natrium
15.0
„
Schwefelsaures Ammoniak
Spuren
Schwefelsaures Calcium
47.9
„
Salpetersaures Calcium
1.0
„
Kohlensaures Calcium
73.9
„
Kohlensaures Magnesium
17.0
„
Kieselsäure
3.7
„
Thonerde und Eisenoxyd
1.9
„
Organische Materie, Verluste
11.9
„
total
174.1
mg
Hinsichtlich der organischen Stoffe, welche durch übermangansaures Kali
nachweisbar sind, sind die beiden äussersten Ziffern bei verschiedenen
Analysen, die von verschiedenen Autoren gemacht wurden, 5.6 und 15.1
mg; die durchschnittliche Ziffer ist 10 mg pro Liter.
Das Studium der allgemeinen physischen Beschaffenheiten des Sees,
dessen Resultat durch chemische Analyse von Brandenburg und
Walter bestätigt ist, hat uns gezeigt, dass das Wasser der
tieferen Schichten des Sees dieselbe Zusammensetzung hat, wie das an
der Oberfläche; die festen und gelösten Substanzen bewegen sich hier
in gleichen Proportionen; die Gase sind hier ein wenig reichlicher;
besonders giebt es ein wenig mehr Sauerstoff: 7.08 und beträchtlich
mehr Kohlensäure: 5.28 cc pro Liter (J. Walter).
Berechnet man nach der Bunsenschen Formel die Quantität des
Gases, welches das Wasser vermöge seines einfachen Kontaktes mit
der atmosphärischen Luft in gelöstem Zustand auf der Höhe des
Wasserspiegels des Genfersees enthält, so kommt man zu folgenden
Ziffern — pro Liter —
Sauerstoff
Stickstoff
Kohlensäure
bei + 5° C.
7.3 cc
13.6 cc
0.6 cc
bei + 20° C.
5.7 „
10.7 „
0.3 „
[S. 16]
Das Wasser des Lac Leman ist also in einem Zustande der Sättigung an
Sauerstoff und Stickstoff und enthält einen beträchtlichen Überschuss
an Kohlensäure.
Wir haben somit in einem Süsswassersee ein flüssiges Medium, bestehend
aus reinem Wasser, welches aufgelöst enthält:
mineralische Stoffe: diese sind für den Ernährungsprozess
der Organismen von noch nicht genügend aufgeklärter Bedeutung; wir
können sie beiseitelassen;
Gase, unter anderen: Sauerstoff, der zur Atmung der
Tiere und Pflanzen dient, und Kohlensäure, welche von den Pflanzen
aufgenommen und zerlegt wird;
aufgelöste organische Substanzen, nach dem Verhältnis von
10 mg pro Liter. — Was sind diese organischen Stoffe? Ihre
Natur kann variieren nach den zufälligen Umständen, die sie erzeugt
haben; wir können annehmen, dass es wesentlich sind:
stickstoffhaltige Substanzen: Albuminoide, Harnstoff, Kreatin
und analoge Produkte;
alle Zwischenstufen der Verwesung der stickstofffreien und der
stickstoffhaltigen Stoffe, welche sie zu den letzten Produkten
führen: Kohlensäure, Ammoniak, Salpetersäure und Stickstoff.
In diesem, also sehr zusammengesetzten Medium finden Pflanzen und
Tiere die Elemente ihrer Nahrung; anderseits lösen sich darin die
Ausscheidungs- und Zersetzungsprodukte der organischen Wesen auf.
Die organische Substanz, in ihrer allgemeinsten Bedeutung genommen,
läuft so durch die Körper der verschiedenen Wesen hindurch. Sie wird
von ihnen aus dem Vorrat geschöpft, den man unerschöpflich nennen
kann und der von dem sie umgebenden Wasser geliefert wird. Sie wird
diesem Vorrate zurückerstattet, nachdem sie für einige Zeit in den
Geweben von Tieren und[S. 17] Pflanzen assimiliert worden ist. Wir können die
aufeinanderfolgenden Phasen dieser Zirkulation der Materie in der Reihe
der Wesen folgendermassen charakterisieren:
Ausgangspunkt: Organischer Stoff, aufgelöst in Wasser: ternäre
und quaternäre Substanzen, Kohlensäure, Sauerstoff.
1. Phase: Organisation der Materie. Die Pflanzen absorbieren
die Kohlensäure und reduzieren sie durch Assimilation in der Form von
Kohlenwasserstoffverbindungen, besonders von Cellulose und Stärke. Sie
absorbieren die stickstoffhaltige Materie und assimilieren dieselbe in
der Form des Protoplasmas; die Protisten absorbieren die organische
Materie und assimilieren sie in ihren Geweben; die Tiere absorbieren
durch ihren Verdauungsapparat die im Wasser aufgelöste organische
Materie, welche einen Teil ihrer Nahrung bildet. Alle lebenden Wesen
absorbieren Sauerstoff für ihre Atmung.
2. Phase: Übergang der Materie von einem Wesen zum andern: Die
Pflanzen sind die Nahrung der pflanzenfressenden Tiere, die kleinen
Tiere die Nahrung der fleischfressenden Tiere; die Reste von Pflanzen
und Tieren diejenige der Omnivoren. Die organisierte Materie geht also
von einem Wesen zum andern und macht eine Reihe von Inkarnationen
durch, ehe sie in die fundamentale Masse der toten Materie zurückkehrt.
3. Phase: Auflösung. Alle lebenden Organismen geben direkt
oder indirekt die Stoffe ihrer Gewebe an das sie umgebende Medium ab.
Lebende Pflanzen und Tiere werfen ihre exkrementellen Ausscheidungen,
welche lösbar sind, ab; tote Pflanzen und Tiere, die nicht von den
Omnivoren verzehrt worden sind, gehen infolge der Verwesung in Stoffe
über, die im Wasser löslich sind.
Diese dritte Funktion führt uns zu einem Endpunkt zurück, nämlich
auf die im Wasser aufgelöste organische Materie, die schon unser
Ausgangspunkt gewesen ist. Wir haben also einen geschlossenen Kreis,
einen Cyklus: die organische Materie zirkuliert in der Reihe der
Lebewesen.
[S. 18]
Ein solcher Prozess kann sich in einem geschlossenen Raum vollziehen
und sich ins Unendliche fortsetzen. Man kennt diese Süsswasser-
oder Salzwasseraquarien, bestehend in einem dicht verschlossenen
Glasbehälter, in welchem einige tierische und pflanzliche Organismen
zusammen den vollständigen Kreislauf ihres Lebens durchmachen. Ich habe
solche unter den Händen meines Kollegen und Freundes Prof. G. Du
Plessis, damals in Lausanne, mehrere Jahre hinter einander bis zur
Vollkommenheit funktionieren sehen.
Aber ein See ist durchaus nicht ein von allen Seiten geschlossenes
Gefäss. Er steht vor allem in Beziehung zur Atmosphäre und ein Teil der
organischen Materien, welche er einschliesst, zerstreut sich in die
darüber liegende Luft: Kohlensäure, das letzte Produkt der Respiration
der Organismen, und Sumpfgas sind der Endpunkt der Verwesung der
organischen Materien. Ein Teil dieser zwei Gase bleibt aufgelöst
im Wasser; aber ein anderer Teil verdunstet in die Atmosphäre. Die
Kohlensäure ist, wie wir bereits gesehen, im Seewasser überreichlich
vorhanden. Dieser Überfluss geht an der Oberfläche des Wassers in die
Atmosphäre über. Anderseits löst sich das Methan, in wie geringer
Menge es sich auch vorfindet, im Wasser auf, wo es oxydiert und in
Oxydationsprodukte höhern Grades verwandelt wird. Der Beweis dafür ist
der Umstand, dass es in der Analyse der Wassergase nirgends angeführt
wird. Wenn es sich aber in zahlreichen Bläschen loslöst, entquillt es
an der Oberfläche des Sees und verliert sich in die Atmosphäre.
Durch diesen Prozess geht eine gewisse Menge organischer Stoffe
verloren und ein mit der Atmosphäre in Verbindung stehender See würde
schliesslich der für die lebenden Wesen notwendigen Nahrungsmittel
ermangeln. Zwar enthält das in den See fallende Regenwasser, wie wir
gleich sehen werden, eine gewisse Quantität organischer Stoffe, durch
welche ein Teil dieser Verluste ersetzt werden würde. Allein wenn es
nur diesen Zufluss gäbe, so würde der Vorrat äusserst gering und die
biologische Bevölkerung eine sehr beschränkte sein.
[S. 19]
Hier haben wir den zweiten Faktor des allgemeinen biologischen
Gleichgewichtes einzuführen, nämlich die Thatsache, dass der See an der
allgemeinen Zirkulation der Stoffe zwischen den verschiedenen Regionen
des Erdballs teilnimmt. Der See hat einen Abfluss und einen Zufluss
und er erhält und verliert durch dieselben eine bedeutende Quantität
organischer Stoffe; er steht in Verbindung mit der Atmosphäre und
erhält von ihr Gasstoffe und giebt sie ihr.
Jeder Süsswassersee hat einen Abfluss als notwendige Bedingung
seiner Existenz. In einem See ohne Abfluss konzentrieren sich die
Gewässer durch Verdunstung und ihr Salzgehalt steigt, bis die Salze
sich krystallisieren: Ein See ohne Abfluss ist ein Salzsee. Ein
Süsswassersee hat also einen Abflusskanal, der Tag für Tag, jahraus,
jahrein eine Menge aufgelöster und unlösbarer organischer Stoffe
ableitet. Ist es der Mühe wert, sie in Anschlag zu bringen?
Ich werde diese Frage beantworten, indem ich mich auf die Angaben über
den Genfersee stütze.
Die Wassermenge, die durchschnittlich im Jahre durch die Rhône bei Genf
abfliesst, beträgt ungefähr 10000 Millionen Kubikmeter. In einem so
bedeutenden Volumen enthält diese Wassermenge noch sehr beträchtliche
Stoffmassen, wie verdünnt diese auch sein mögen.
Das Wasser enthält durchschnittlich pro Liter 5 cc aufgelöste
Kohlensäure. Für die 10 Milliarden cm macht das 50 Milliarden
Liter Gas aus. Da das Gewicht eines Liters Kohlensäure 2 g
beträgt, so stellt diese Quantität ein Totalgewicht von 100000 Tonnen
Kohlensäure, resp. 28000 Tonnen Kohlenstoff vor.
Im Durchschnitt finden sich in einem Liter dieses Wassers 10 mg
organische durch übermangansaures Kali oxydierbare Stoffe vor.
Mithin enthalten die 10 Milliarden cm Wasser 100000 Tonnen
organische Stoffe, die darin aufgelöst oder im Zustande lebenden oder
abgestorbenen Staubes sind.
Dieses Wasser enthält durchschnittlich 36 Mikroben pro cc,
das macht für die ganze Wassermasse 380000000000000 Mikroben.[S. 20] Die
Zahl ist ungeheuer, aber ihr Gewicht sehr klein. Wäre die Behauptung
übertrieben, dass ihrer eine Million auf ein Milligramm geht? Wenn
dem so wäre, so würden die unter dieser Form dem See abfliessenden
organischen Stoffe nicht 380 Tonnen überschreiten.
Dieses Wasser enthält Tiere und Pflanzen. Um deren Zahl zu schätzen,
setze ich voraus, ihre Bevölkerungsdichtigkeit entspreche derjenigen
der pelagischen Region; indem ich mich auf die Beobachtungen Imhofs
stütze, finde ich durch Berechnungen, die ich ihrer Länge wegen hier
nicht vorbringen kann, dass jeder Kubikmeter durchschnittlich 400
Mikrozoen und 8000 Mikrophyten enthält. Wenn wir annehmen, diese
Mikroorganismen haben ein mittleres Gewicht von etwa 0.01 mg, so
erhalten wir ein Totalgewicht von 840 Tonnen für den Abfluss.
Demnach würde die Rhône jährlich aus dem Genfersee abführen:
100000
Tonnen
Kohlensäure,
100000
„
oxydierbare organische Stoffe,
380
„
Mikroben,
840
„
Mikroorganismen,
somit im ganzen mehr als 200000 Tonnen. Dem muss noch folgendes
hinzugefügt werden: der Staub und die organischen Reste, die
abgestorbenen, im Wasser des Abzugskanals schwimmenden oder an das
Ufer geworfenen Tierkörper und Pflanzenteile; die von Menschen und
von fischfressenden Vögeln gefangenen Fische; die Insekten, die im
Larvenzustand im See sich entwickelt und denselben als vollkommene
Insekten verlassen haben, um von den insektenfressenden Säugetieren und
Vögeln (Fledermäusen und Schwalben) verzehrt zu werden. Um alles zu
sagen, fügen wir noch hinzu, dass eine gewisse Quantität organischer
Stoffe, die im Lac Leman sehr gering, in anderen Seen aber bedeutender
ist, durch Fossilisation im neuen Alluvium, das sich auf dem Grunde des
Sees bildet, absorbiert wird.
Diese Berechnungen sind nur annähernd richtig; auch beschränke ich mich
darauf, zu sagen, dass die im Wasser aufgelösten und[S. 21] suspendierten,
lebenden oder abgestorbenen organischen Stoffe, die jährlich
hauptsächlich durch den Abfluss dem Genfersee entzogen werden, mehrere
100000 Tonnen betragen.
Nun ist, wie wir sogleich erörtern werden, die Zusammensetzung des
Wassers eines grossen Sees eine stabile; auch führen die Monate
und Jahre keine Variationen in derselben herbei; die abfliessenden
organischen Stoffe werden folglich anderweitig ersetzt.
Welches sind nun die Quellen, welche dem See die hunderttausende von
Tonnen organischer Stoffe ersetzen, die der Abfluss ihm jährlich
entführt?
Er erhält dieselben:
durch die atmosphärische Luft, nämlich:
Sauerstoff, der sich im Wasser der Oberfläche auflöst und daselbst
den Zustand der Sättigung erhält. Derselbe wird durch Diffusion und
hauptsächlich durch die thermische und mechanische Konvektion von den
oberen Schichten des Wassers in die tieferen geführt. Durch seine
Berührung mit der Luft und die Ausscheidung der chlorophyllhaltigen
Gewächse ist das Seewasser mit Sauerstoff gesättigt: es ist
lufthaltiges Wasser. Da die Lösbarkeit des Sauerstoffes im Wasser
grösser ist, als die des Stickstoffes, so ist die im Wasser
aufgelöste Luft an Sauerstoff reicher als die atmosphärische und
enthält ⅓ Sauerstoff und ⅔ Stickstoff.
Die im Regenwasser aufgelösten organischen Stoffe, welche nach dem
Studium des Herrn A. Levy am Observatorium zu Montsouris bei
Paris wie folgt geschätzt werden können: Auf ein Liter kommen:
Ammoniak
2.3
mg
Salpetersäure, salpetrige Säure
0.9
„
Nitrate und Nitrite
Organische durch übermangansaures
Kali oxydierbare Stoffe (Staub etc.)
49.0
„
Wenn wir annehmen, es falle im Durchschnitt jährlich eine Schicht
von 0.9 m meteorisches Wasser, so stellt dies[S. 22] folgende
Quantitäten dar, welche ich unter zwei Formen angebe: nämlich die
Wassermenge, die auf einen Quadratkilometer, und die, welche auf
die ganze Oberfläche des Genfersees fällt, um mich an das bis jetzt
benutzte Beispiel zu halten.
Pro qkm:
Auf dem Genfersee:
Ammoniak
2.1
Tonnen
1200
Tonnen
Salpetersäure etc.
0.8
„
460
„
Oxydierb. organ. Stoffe
44.1
„
25500
„
Durch den Transport fester Stoffe, die durch die Atmosphäre
herbeigeführt werden, nämlich: der organische Staub, die
vegetabilischen Körperteile, die Vögel und Insekten, welche im
See ertrinken oder ihre Exkremente da fallen lassen; die Eier der
Luftinsekten, aus welchen die Wasserlarven entstehen, etc.
Durch die Zuflüsse gelangen in den See mineralisches Alluvium und
organische Stoffe. Letztere befinden sich:
im Zustande aufgelöster Stoffe. Das Wasser der Zuflüsse ist das
Waschwasser des ganzen zum Seebecken gehörenden Flussbettes, das
Wasser, welches den Boden von den löslichen Stoffen, namentlich von
den Abfällen des vegetabilischen und animalischen Lebens befreit. Es
reisst das Wasser der Aborte und Kloaken der Städte, Fabriken, das
durch die Humussubstanzen gebräunte Wasser der torfigen Moräste mit.
Im Rhônewasser im Wallis hat Buenzod ein auflösliches Residuum
von 23 cg per Liter gefunden, während das Wasser des Lac
Leman nur 17.5 cg enthält. Da die mikroskopische Analyse des
Seeschlammes keine Spur von krystallinischen Niederschlägen gezeigt
hat, ist es wahrscheinlich, dass der Unterschied (5.5 cg auf
den Liter) grösstenteils durch organische Stoffe gebildet wird.
In unaufgelöstem Zustande als schwebende Materie: Tierleichen und
Wasser- und Landpflanzen oder deren Teile. Diese Zufuhr ist sehr
bedeutend. Ich kenne aber kein Mittel, sie zu schätzen und in Zahlen
anzugeben.
[S. 23]
Auf diese Weise erhält der See eine genügende Quantität organischer und
anorganischer Stoffe, um die durch Gasdiffusion, durch den Abflusskanal
und die Fossilisation verlorenen zu ersetzen.
Wenn man aber die Verschiedenheiten dieser Bezugsquellen, ihre
gegenseitige Unabhängigkeit, ihre Variabilität, den völligen Mangel
an Wechselbeziehungen zwischen Zufluss und Abfluss erwägt, so
scheint es zuerst, dass die chemische Zusammensetzung des Seewassers
sehr verschieden sein muss, dass die Jahreszeiten, Jahrgänge und
Zufälle Veränderungen herbeiführen müssen. Es ist das jedoch nicht
der Fall. Wir besitzen zahlreiche Analysen des Genferseewassers,
darunter zwölf vollständige, allgemeine, und etwa hundert partielle,
die zu verschiedenen Zeiten, von verschiedenen Chemikern nach
verschiedenen Methoden und zu verschiedenen Zwecken vorgenommen
worden sind. Kompetente Fachmänner, welche diese Analysen prüften,
haben festgestellt, dass dieselben hinsichtlich des Wesentlichen
übereinstimmen. Die hie und da zu Tage tretenden Verschiedenheiten sind
lokale, und verschwinden bald durch Diffusion oder durch mechanische
Mischung der grossen Wassermasse.
Die Ursache dieser fortdauernden Gleichmässigkeit der Zusammensetzung
muss in der ungeheuren Grösse der erforschten Wassermasse gesucht
werden. Der Genfersee misst ungefähr 89000 Millionen cm.
Wenn wir den Inhalt dieses Wasserbeckens mit irgend einer Substanz
um ein Milligramm pro Liter ändern wollten, so müssten wir 89000
Tonnen à 1000 kg von dieser Substanz hineingiessen oder dem
See entziehen. Nun vermögen keine auch noch so mächtigen Kräfte,
ausgenommen ein Kataklysmus, irgend eine Substanz in so grosser Menge
auf unregelmässige Weise dem See zuzuführen. Dieselbe Beweisführung
muss für die meisten Seen gelten; denn obschon das Volumen der Gewässer
des Lac Leman im Verhältnis zur Ausdehnung seines Zuflussbeckens ein
grosses ist, so liegt doch in diesem Verhältnis der beiden Quantitäten
nichts Ausserordentliches.
[S. 24]
Wir müssen also annehmen, dass die Zusammensetzung des Seewassers eine
konstante, unveränderliche ist; dass die Tiere und Pflanzen in einem
Medium leben und sterben, das in chemischer Beziehung stets dasselbe
bleibt.
Wie die chemische Zusammensetzung des Ozeans fast immer und überall
dieselbe ist, so ist auch die chemische Zusammensetzung eines
Süsswassersees eine unveränderliche.
Anderseits ist es wahrscheinlich, ja gewiss, dass es von einem See zum
andern ziemlich grosse Verschiedenheiten geben kann, welche von der
petrographischen Natur des zum See gehörigen Flussbeckens herrühren
oder von dem Kulturstand des den See umgebenden Landes und der
Verhältnisse, welche dieser Kulturzustand bedingt, oder von welchen er
bedingt wird (Humusbestand und Klima).
Der Vorrat an organischen Stoffen im See erneuert sich also durch
das Hinzukommen neuer Materien, welche die weggeführten ersetzen.
Es ist klar, dass der grösste Teil der Stoffe durch den Abfluss dem
See entzogen wird. Das gestattet uns annähernd die Intensität eines
solchen Stoffwechsels im Lac Leman, der uns als Beispiel gedient,
zu berechnen. Die Wassermasse dieses Sees beträgt 89000 Millionen
cm; die Wassermasse, die jährlich durch die Rhône bei Genf
abfliesst, beträgt etwa 10000 Millionen; die jährlich abfliessende
Wassermasse ist also ungefähr der neunte Teil der Totalmasse; es wird
somit durch den Abfluss jährlich ungefähr ein Neuntel des Vorrates an
organischen Stoffen entzogen. Da noch die Stoffe in Rechnung gebracht
werden müssen, die in der Atmosphäre aufgehen oder die im Alluvium
fossilisiert werden, so können wir sagen, dass die organischen Stoffe
höchstens sieben oder acht Jahre im See verweilen, um den lokalen
Kreislauf unter den verschiedenen ihn bewohnenden Wesen zu vollenden,
bevor sie in den grossen Cyklus der allgemeinen Weltzirkulation
zurückkehren.
Ein See stellt uns also nach dem Dargelegten ein beschränktes, mit
Wasser gefülltes Becken dar, das, obschon[S. 25] es im Vergleich mit dem
Meerwasser süss ist, doch auf je ein Liter
150–250
mg
aufgelöste mineralische Salze,
10
„
organische Stoffe,
20–25
cc
Gase
enthält. Dieses Wasser enthält ausserdem schwebenden organischen und
mineralischen Staub, dessen Menge vom Wasserstand der Zuflüsse und von
ihrer Natur (Gletscherbäche, Moorwasser etc.) abhängt.
Diese Materien bilden einen Vorrat, der durch die atmosphärischen
Niederschläge und die Gewässer der Zuflüsse erhalten wird; ein Teil
dieser Stoffe wird durch den Abfluss entzogen oder verliert sich in der
atmosphärischen Luft. Allein Zufuhr und Abfuhr heben sich auf und die
Zusammensetzung des Wassers bleibt immer dieselbe.
Dieser Vorrat dient zur Ernährung zahlreicher und mannigfaltiger
Organismen, welche den beiden Reichen der organischen Welt angehören,
den verschiedenen Typen: von den Wirbeltieren und Dikotyledonen an bis
zu den Protozoen, Algen, Protisten und den Mikroben.
Diese verschiedenen Typen zusammen lebender Wesen absorbieren
organische Stoffe und bilden neue; durch die Wechselbeziehungen
entgegengesetzter Funktionen ergänzen sie sich in der Konsumtion und
Restitution der Vorratssubstanzen gegenseitig. In dieser Hinsicht ist
ein See ein Mikrokosmos, eine abgeschlossene Welt, die sich selbst
genügt. Aber zugleich greift er mittels seiner Zuflüsse und seines
Abflusses in die allgemeine Kreisbewegung des Erdballes ein. In dieser
Hinsicht ist der See nichts weniger als isoliert, sondern gehört mit
zum Ganzen des Universums.
Indem wir uns auf das obige, über die allgemeine Biologie Gesagte
stützen, ziehen wir folgende Schlüsse:
[S. 26]
1. Der organische Stoff vollzieht seinen Kreislauf unter den
verschiedenen Wesen verschiedener Typen, welche im beschränkten Raume
eines Süsswassersees neben einander leben.
2. Dieser dem See angehörende organische Stoff ist nicht absolut und
für immer in diesem verhältnismässig kleinen Raume lokalisiert, sondern
er tritt als Glied in den grossen Cyklus des allgemeinen Kreislaufes
ein, welcher die verschiedenen Regionen des Erdballes durch die Ströme,
den Ozean und die Atmosphäre verbindet.
[S. 27]
Die Algen.
Von Dr. W. Migula in Karlsruhe.
[S. 29]
Das Wasser ist die Heimat des organischen Lebens. Alle Thatsachen
deuten darauf hin, dass die ersten lebenden Wesen im Wasser auftraten,
und dass erst sehr langsam mit der fortschreitenden Entwickelung der
Organismen eine Besiedelung des Landes begann. So müssen wir auch
voraussetzen, dass wir im Wasser die am einfachsten gebauten Organismen
antreffen werden, wenn sich irgendwelche Nachkommen jener ersten
Wesen erhalten haben. Dies ist thatsächlich der Fall. Die niedersten
Lebensformen beider Reiche gehören dem Wasser an, und je tiefer wir
in den Kreis dieses Lebens hinabsteigen, um so ähnlicher werden sich
die Wesen, um so schwieriger wird es, Tier und Pflanze aus einander zu
halten. Wir stehen dann schliesslich vor einer Gruppe von Wesen, welche
sowohl der Zoologe wie der Botaniker für seine Wissenschaft in Anspruch
nimmt und welche beweisen, dass das gesamte organische Leben der Erde
nur von einer Wurzel getragen wird, aus welcher sich wie zwei mächtige
Stämme Tierreich und Pflanzenreich entwickelt haben.
Man kann deshalb auch nicht gut davon reden, was eher auftrat, Tier
oder Pflanze; es waren eben jene einfachsten Wesen, in denen sich noch
die Eigenschaften beider vereinigen. Freilich muss man annehmen, dass
es zunächst diejenigen waren, welche die Fähigkeit besassen, aus den
anorganischen Stoffen, dem Wasser, der Kohlensäure, den anorganischen
Stickstoffverbindungen und dem Sauerstoff, ihren Körper aufzubauen, und
diese stehen im grossen und ganzen eben um dieser Eigenschaft willen
dem Pflanzenreich[S. 30] näher. Denn Organismen, welche diese Fähigkeiten
nicht besitzen, konnten erst dann auftreten, wenn ihnen durch jene
organische Stoffe bereitet waren, welche ihnen zur Nahrung dienen
konnten, und diese Eigenschaft kommt im allgemeinen den Tieren zu.
Will man also nur ganz allgemein reden, so müssen die Pflanzen eher
existiert haben als die Tiere.
Die einfachsten Pflanzen werden durch zwei grosse vielfach durch
Berührungspunkte verbundene Klassen repräsentiert, durch Pilze
und Algen. Da aber die ersteren durch ihr Unvermögen, sich
von anorganischen Stoffen zu ernähren, den Tieren gleichen und
schon die Anwesenheit anderer Wesen voraussetzen, müssen wir in den
Algen diejenigen Organismen suchen, welche den Ausgangspunkt
für das organische Leben der Gegenwart bilden, will man sich nicht
auf zu gewagte Phantasien über die untergegangenen Urwesen einlassen.
Die Algen zeigen auch den grössten Formenreichtum und die
überraschendste Vielgestaltigkeit unter den Pflanzen unserer Gewässer,
so mannigfach auch deren Schmuck mit Blütenpflanzen sein mag. Sie sind
eigentliche Wasserpflanzen und nur wenige vermögen auch in feuchter
Luft an nassen Felsen oder zwischen Moos zu gedeihen, noch weniger
sind wirkliche Landpflanzen, die auch auf Dächern, Rinde und trockenen
Steinen fortkommen. Aber wo auch nur immer sich Wasser angesammelt hat,
in Bächen, Pfützen oder Seen, sind auch Algen zu finden, nur ist die
Vegetation derselben je nach der Beschaffenheit des Wassers und nach
der Jahreszeit verschieden zusammengesetzt.
Man kann an ein und demselben Ort das ganze Jahr hindurch Algen
sammeln und wird fast jeden Monat andere Arten finden. Ein torfiger
Wiesengraben zeigt im Frühjahr, wenn Schnee und Eis eben verschwunden
sind, reiche Entwickelung von goldbraunem Schaum an der Oberfläche
des Wassers, der unter dem Mikroskop die zierlichen, mit bräunlichen
Körnern oder Platten gefüllten Kieselpanzer der Diatomeen
in zahlloser Menge erkennen lässt. Ein wenig später findet sich an
derselben Stelle vom Boden aufsteigend ein dünner grüner Schleim, der
sich allmählich verdichtet, an die Oberfläche steigt und zahlreiche
kleine Gasblasen[S. 31] festhält. Fährt man dann mit dem Spazierstock hinein,
so bleiben sicher eine Anzahl äusserst dünner, glatter und schlüpfriger
Fäden hängen, welche über ihre Zugehörigkeit zu den Zygnemaceen
keinen Zweifel lassen und mikroskopisch durch ihre eigenartigen
Chlorophyllkörper leicht von anderen Algen zu unterscheiden sind.
Steigt die Sonne höher und fallen ihre heissen Strahlen senkrechter
auf den sinkenden Wasserspiegel des Grabens, so verschwinden die
Zygnemaceen und machen anderen Algen Platz: grünen, nicht schleimigen
Flöckchen aus der Familie der Conferven, deren Chlorophyll
die ganze Zellwand auskleidet. Allmählich treten zwischen den Fäden
derselben die zierlichen einzelligen Desmidieen auf, welche umsomehr
zur Herrschaft gelangen, je herbstlicher es draussen auf den Fluren
wird. So wechseln an demselben Standort Vertreter aller Familien die
Herrschaft, während zu gleicher Zeit andere Algen nur vereinzelt
zwischen den Individuen des gerade besonders entwickelten Geschlechtes
vorkommen.
Doch nicht nur die Jahreszeit, auch die Beschaffenheit des Wassers
übt einen gewaltigen Einfluss auf das Gedeihen der verschiedenen
Arten. Während die grünen Fadenalgen sowie die meisten Diatomeen nur
in frischem, unverdorbenem Wasser zu existieren vermögen, ziehen
die blaugrünen Oscillarien fauliges, mit verwesenden organischen
Stoffen erfülltes Wasser vor. Manche Gattungen wie Spirogyra,
Oedogonium, Bulbochaete lieben stehendes oder nur
schwach fliessendes Wasser, andere wie Lemanea, Cladophora
glomerata und einige Diatomeen befinden sich in reissenden
Gebirgsbächen, an Wehren oder Wasserfällen am wohlsten. Auch giebt es
Algen, welche wesentlich von der Temperatur abhängen; gewisse Arten
der blaugrünen Cyanophyceen leben nur in heissen Quellen, wie die
Lyngbya thermalis in den Geysern Islands und den Schlammvulkanen
Italiens, wogegen Hydrurus irregularis in der heissen Jahreszeit
verschwindet, aber vom Herbst an den ganzen Winter hindurch und auch
noch im kühleren Frühjahr auftritt. Die chemische Zusammensetzung
des Wassers spielt ebenfalls eine wichtige Rolle in Bezug auf das
Gedeihen der einen oder andern Art. Andere Arten leben in Gräben der
Torfmoore,[S. 32] als in den Bächen lehmiger Wiesen, andere in den Tümpeln
auf sandigem Boden. In den frischklaren Gebirgsseen tritt eine andere
Algenflora auf als in den wärmeren Gewässern der Ebene und das süsse
Wasser birgt andere Formen als die Salzlachen des Binnenlandes.
Manche der niedersten Spaltpflanzen scheinen das Vorhandensein von
Schwefelverbindungen zu ihrem Leben durchaus nötig zu haben, während
wieder andere nur in eisenhaltigen Gewässern ihr Gedeihen finden.
Die Verhältnisse in der Zusammensetzung des Teich- und Flusswassers
sind ja so mannigfaltig, dass sich die Ansprüche der Algen nach sehr
verschiedenen Richtungen hin entwickeln konnten, wenn sie sich die in
der Natur gegebenen Bedingungen möglichst zu Nutze machen und sich
ihnen anpassen wollten.
Die Orientierung über die Hauptgruppen ist, von einzelnen Fällen
abgesehen, bei den Algen nicht schwer. Sie bilden auch keine so
einheitliche Klasse wie etwa die Moose, sondern werden wesentlich
nur durch den einfachen zelligen, noch nicht deutlich in Stengel und
Blätter gegliederten Bau und durch das Vorhandensein von Chlorophyll
oder einer seiner Modifikationen zusammengehalten und von den höher
organisierten Pflanzen und den chlorophyllfreien Pilzen unterschieden.
Abgesehen von einigen zweifelhaften Meeresbewohnern lassen sich fünf
grosse Gruppen aufstellen, welche sich wesentlich durch Merkmale der
Fortpflanzung, Gestalt und Färbung unterscheiden. Sie lassen sich kurz
folgendermassen charakterisieren:
1. Schizophyceae, Spaltalgen. Färbung blaugrün, spangrün,
orange, rot, violett, aber niemals rein chlorophyllgrün. Sehr einfach
organisierte Wesen, deren Fortpflanzung und Vermehrung durch einfache
Querteilung der Zellen, bei manchen ausserdem noch durch Bildung von
Dauersporen auf ungeschlechtlichem Wege erfolgt.
2. Bacillariaceae oder Diatomaceae, Kieselalgen.
Die Färbung ist eine gelb- oder goldbraune und wird hervorgerufen
durch runde oder plattenförmige Chromatophoren im Innern der von einem
Kieselpanzer umgebenen Zelle. Fortpflanzung durch Kopulation
zweier Individuen, Vermehrung durch Zweiteilung, indem[S. 33] die beiden
Schalen wie die Teile einer Schachtel auseinanderweichen und sich
zwischen ihnen zwei neue Schalen bilden.
3. Chlorophyceae, Grünalgen, Algen im engeren Sinne. Sie
besitzen fast stets rein chlorophyllgrüne Färbung, sind aber sonst sehr
verschiedenartig gestaltet und variieren auch namentlich in Bezug auf
die Fortpflanzung, welche sowohl geschlechtlich wie ungeschlechtlich
sein kann. Sehr häufig kommen beide Fortpflanzungsarten neben einander
vor. Eine Vermehrung findet oft in ausgiebiger Weise durch Zweiteilung
der Zellen statt.
4. Melanophyceae, Braunalgen, durchweg Meeresbewohner
mit stets zweigeisseligen geschlechtlichen und ungeschlechtlichen
Schwärmzellen, meist braun oder schwarzbraun gefärbt.
5. Rhodophyceae, Rotalgen. Ebenfalls zum grössten
Teil Meeresbewohner und nur durch wenige Gattungen im süssen Wasser
vertreten. Sie sind rot, oft prachtvoll gefärbt, einige der im
süssen Wasser vorkommenden Arten haben noch einen andern grünen oder
blaugrünen vorherrschenden Farbstoff. Fortpflanzung ungeschlechtlich
und geschlechtlich, in allen Fällen durch ruhende Zellen.
1. Die Schizophyceen oder
Spaltalgen.
Wo sich in verdorbenem Wasser schmutzig graugrüne oder dunkel
stahlblaue Flocken an der Oberfläche ansammeln oder an feuchten Mauern
ein rötlicher gallertartiger oder schleimiger Überzug entsteht, wo
sich auf Teichen spangrüne Wasserblüten bilden oder bräunlichgrüne
Gallertkugeln schwimmen, in den Torflachen des Hochmoores wie in
den Seen der Ebene, an nassen Felswänden wie auf feuchter Erde und
zwischen Moos treten uns die Spaltalgen entgegen. Viele Arten, wie
die echten Oscillarien, bedürfen grösserer Mengen organischer Stoffe
zu ihrem Gedeihen und treten deshalb vorzugsweise in verdorbenem
Wasser, in Abzugsgräben von Fabriken und ähnlichen Orten auf, einige
sind genügsamer und stellen wesentlich dieselben Forderungen an ihr
Nährsubstrat, wie andere Algen auch.
[S. 34]
Es sind sehr einfach organisierte Geschöpfe. Teils bilden sie nur
einzelne Zellen, welche in festerem oder lockerem, oder auch in
gar keinem Zusammenhange mit einander stehen, teils bleiben sie zu
verschiedenartig gestalteten Zellfäden verbunden. Die Membran der
Zellen ist meist sehr dünn, aber in vielen Fällen, wenigstens in den
äusseren Schichten, sehr quellbar und zur Gallertbildung geneigt.
Neuerdings hat man auch sehr kleine und zwar mehrere Zellkerne in
den Zellen nachgewiesen. Die Färbung scheint nicht an bestimmte
Chromatophoren gebunden, sondern im Plasma verteilt zu sein, sie kann
sehr verschieden, rot, blau, braun, blaugrün, spangrün, violett, in den
verschiedensten Nüancen, aber niemals rein chlorophyllgrün sein. Die
Vermehrung erfolgt durch Querteilung der Zellen und Loslösung derselben
oder einer Gruppe von Zellen, wenn diese einen gemeinschaftlichen
Verband darstellen, seien es Fäden oder nur lose verbundene
Einzelzellen. Solche losgelöste Fadenstücke, welche man Hormogonien
nennt, bilden dann neue Familien, wenn man diesen Ausdruck auf einen
selbständigen Zellkomplex von sehr verschieden innigem Zusammenhange
ausdehnen will.
Neben diesen rein vegetativen Zwecken dienenden und in der Mehrzahl
vorhandenen Zellen kommen auch noch seltener bei manchen Arten zwei
andere Arten von Zellen vor, die Grenzzellen oder Heterocysten
und die Dauerzellen oder Dauersporen. Die ersteren unterscheiden
sich von den vegetativen Zellen leicht durch die bedeutendere Grösse,
durch einen geringeren Gehalt an plastischen Stoffen und durch
abweichende, meist sehr viel hellere Färbung. Sie haben das Vermögen
verloren, sich zu teilen und bilden gewissermassen Grenzpfähle
zwischen den vegetativen Zellen; welchen Zweck sie erfüllen, ist nicht
bekannt. Die Dauerzellen oder Dauersporen, auch kurz Sporen genannt,
weichen ebenfalls von den vegetativen Zellen in der Gestalt ab, nur
sind sie im Gegensatz zu den Heterocysten reicher an Plasma und in
der Regel dunkler gefärbt. Auch ihre Membran ist meist stärker, so
dass sie schädlichen äusseren Einflüssen besser widerstehen können
als die vegetativen Zellen. Sie dienen denn auch dazu, unter[S. 35]
ungünstigen Verhältnissen die Art zu erhalten und fortzupflanzen. Tritt
beispielsweise grosse Dürre ein und trocknen die Pfützen aus, welche
von Dauerzellen bildenden Spaltalgen bewohnt waren, so gehen wohl die
vegetativen Zellen zu Grunde, die Dauerzellen bleiben aber am Leben,
treten in einen Ruhezustand ein und entwickeln, wenn sich die Pfützen
wieder füllen, neue Pflänzchen.
Der Farbstoff, welcher den Spaltalgen eigen ist, wurde Phycochrom
und daher die ganze Gruppe Phycochromaceen genannt; derselbe besteht
wesentlich aus dem blauen Phycocyan (daher Cyanophyceen) und dem gelben
Phycoxanthin, welches dem Blattgelb (Xanthophyll) ähnlich ist, daneben
scheint aber noch ein dem Chlorophyll sehr ähnlicher grüner Farbstoff
vorhanden zu sein. Ausser diesem das Innere der Zellen erfüllenden
Farbstoff finden sich aber noch andere, welche den Hüllen selbst eigen
sind und meist eine gelbbraune oder rote Färbung verleihen, über deren
Eigenschaften man aber noch so gut wie gar nichts weiss. Wenn wir uns
unter den Spaltalgen umsehen, so finden wir eine grosse Einförmigkeit;
nur geringe Unterschiede trennen die Gruppen und die Artenkenntnis
ist grösstenteils eine recht zweifelhafte. Am übersichtlichsten ist
wohl die nachstehende Einteilung der Spaltalgen in Hauptgruppen oder
Familien, jenachdem man unter ihnen noch besondere Unterabteilungen
bildet oder nicht.
1. Coccogene Spaltalgen, Chroococcaceae. Die Zellen
trennen sich nach der Teilung von einander und bleiben entweder
völlig ohne Zusammenhang oder stehen nur in äusserlichem durch
die Gallertbildung der Membran bedingtem Zusammenhang (Fig. 1
a–c). Die wichtigsten Gattungen sind folgende:
Chroococcus (Fig. 1 a), Zellen rund oder eckig, einzeln,
zu zwei oder vier in eine nicht zerfliessliche Gallerte eingebettet,
in welcher man zwar bei manchen Arten Schichtungen, aber keine
Einschachtelungen erkennen kann. Die Gattung Gloeocapsa (Fig.
1 b) unterscheidet sich von der vorhergehenden durch stets
runde Zellen und durch Zellfamilien, in welchen die Zellmembran der
Mutter um die Tochterzellen stets erhalten bleibt, so dass vollständige
Einschachtelungen entstehen. Viele Arten besitzen eine sehr lebhafte
rote oder violette[S. 36] Färbung dieser Gallertschichten, während der Inhalt
der Zelle selbst mehr spangrün gefärbt ist. Ganz ähnlich ist die
Gattung Gloeothece, nur sind hier die Zellen länglich. Diesen
beiden Gattungen entsprechen Aphanocapsa und Aphanothece
vollständig, nur sind hier die Hüllmembranen nicht in einander
eingeschachtelt, sondern bilden eine homogene Gallerte. Die Gattung
Merismopedia bildet Zellfamilien von Tafelform, je vier Zellen
stehen näher zusammen (Fig. 1 c). Clathrocystis bildet
rundliche Zellen, welche durch vergallertende Membranen zu kleinen
Hohlkugeln verbunden bleiben. Sie bildet, ebenso wie die sehr ähnliche
Gattung Polycystis, oft spangrüne Wasserblüten, welche den
Fischen verderblich werden können.
2. Nematogene Spaltalgen. Die Zellen bleiben zu längeren oder
kürzeren Fäden vereinigt. Die Oscillariaceen repräsentieren
unter ihnen den niedrigsten Stand. Bei ihnen ist noch keinerlei
Unterschied zwischen den einzelnen Zellen vorhanden, sie besitzen
weder Heterocysten noch Dauersporen und beide Enden des Fadens
sind[S. 37] gleichartig ausgebildet. Unter ihnen ist wieder die Gattung
Oscillaria (Fig. 1 d) am einfachsten organisiert, es
sind einfache scheidenlose spangrüne, blaugrüne oder violette Fäden,
in denen man die Querwände der einzelnen Zellen oft nur undeutlich
erkennt. Von ihr unterscheidet sich die Gattung Beggiatoa
(Fig. 1 n) durch das Fehlen des Farbstoffes. Beide Gattungen
sowie die verwandte korkzieherartig gedrehte Spirulina (Fig.
1 m) besitzen Bewegungsvermögen, eine kriechende mit Drehung
um die Längsachse und die Krümmungen des Fadens verbundene Bewegung,
welche sie befähigt, an den Wänden von Glasgefässen etc. in die Höhe
zu steigen. Die Oscillarien sind Bewohner unreinen Wassers, und wo
man sie findet, kann man ohne weiteres darauf schliessen, dass in dem
Wasser irgendwelche Fäulnisprozesse stattgefunden haben, und dass es
für den Gebrauch als Trinkwasser ungeeignet ist. Bei zwei anderen
Gattungen dieser Gruppe finden sich Scheiden um den Faden, welche aus
den vergallertenden Aussenwänden der Zellmembranen entstehen. Bei
Chamaesiphon ist die Scheide sehr zart, die Pflänzchen bestehen
aus wenigen undeutlich begrenzten Zellen und sitzen oft massenhaft auf
anderen Fadenalgen auf (Fig. 1 k). Bei Lyngbya (Fig.
1 l) sind sie sehr stark und oft infolge der Einlagerung von
Eisenocker gelb gefärbt. Die in eisenhaltigem Wasser sumpfiger Gräben
auftretenden rostroten Flocken gehören den oft schon abgestorbenen
Fäden der Ockeralge, Lyngbya ochracea an, welche allerdings viel
feinere Fäden bildet als die in Fig. 1 l abgebildete Art.
Die Nostocaceen stehen schon wesentlich höher; zwar bilden auch
sie nur einfache unverzweigte Fäden, deren Basis von der Spitze nicht
verschieden ist, aber es sind schon Grenzzellen vorhanden; und auch
Dauersporen kommen den meisten Arten auf der Höhe ihrer Entwickelung
zu. Die Gattung Nostoc bildet rosenkranzförmige Ketten, welche
aus je einer Anzahl vegetativer Zellen zwischen zwei Heterocysten
zusammengesetzt sind (Fig. 1 h). Eine Anzahl solcher
Perlschnuren liegen dann in einer bestimmt geformten meist kugeligen
Gallerte eingebettet, welche wieder aus den quellbaren Membranen der
Zellen entstehen. So tritt uns Nostoc im[S. 38] Wasser wie an feuchten
Felsen oder Hohlwegen im Walde nicht selten entgegen und hat wenigstens
teilweise eine Rolle bei der Sternschnuppengallerte gespielt, während
diese letztere auch, und wohl hauptsächlich, auf die ausserordentlich
quellbaren Elemente der Eierstöcke weiblicher irgendwie zerstückelter
Frösche zurückzuführen ist. Die Gattung Anabaena unterscheidet
sich von Nostoc nur dadurch, dass ihre Fäden nicht in
Gallertklumpen zusammengelagert sind. Wir kultivieren unabsichtlich
eine Art derselben in unseren botanischen Gärten, die Anabaena
Azollae, welche die hohlen Blätter des kleinen ausländischen
Wasserfarns Azolla caroliniana fast regelmässig bewohnt. Eine
andere hierher gehörige Alge Aphanizomenon oder Limnochlyde
Flos aquae (Fig. 1 f) mit grossen cylindrischen Dauerzellen
bildet kleine zusammenhängende Flöckchen, welche auf der Oberfläche des
Wassers schwimmen und oft eine dichte Wasserblüte hervorrufen. In der
Gattung Cylindrospermum stehen die Grenzzellen terminal, daneben
die meist cylindrisch gestalteten Dauerzellen.
Höher organisiert sind die Stigonemaceen, welche bereits
verzweigte Fäden bilden, wie der in Fig. 1 i abgebildete
Hapalosiphon. Die Verzweigung entsteht dadurch, dass der
Zellfaden nicht bloss an den Enden, sondern auch in der Mitte wächst,
d. h. dass sich die Zellen auch hier teilen und eine Spannung der
Gallertscheide bewirken, die schliesslich an einer Stelle reisst und
die Zellen hervordringen lässt, welche nun wieder sich teilen und
dadurch einen Zweig bilden. Am höchsten stehen die Rivularien,
welche bereits einen deutlichen Gegensatz zwischen Basis und Spitze
zeigen, wie eine in Fig. 1 g abgebildete Gruppe junger
Rivulariafäden erkennen lässt.
Wie schon erwähnt zerfällt der Thallus der fadenbildenden
Spaltalgen auf einer bestimmten Entwickelungsstufe in eine
Anzahl kurzer Fadenstücke, Hormogonien, welche eine Zeitlang
Bewegungsfähigkeit besitzen und umherwandern, um sich ein neues Heim
zu suchen. Dabei wählen sie mit Vorliebe Orte zu ihrem Aufenthalt,
die ihnen gegen aussen Schutz gewähren, leere Insekten oder
Crustaceenschalen, grosse abgestorbene Pflanzenzellen, oder irgendwie[S. 39]
hohle Organe lebender Pflanzen, wie wir sie in den Schwimmblasen der
Utricularia und bei mehreren Torf- und Lebermoosen finden.
Auch aus den kugeligen Zellenfamilien der Chroococcaceen können sich
Kugelsegmente loslösen und zu neuen Familien heranwachsen, wodurch alte
Kolonien ein ganz durchlöchertes Aussehen gewinnen.
Das Interesse, welches die Spaltalgen uns erwecken, wird vorzüglich
noch durch zwei Punkte vermehrt, durch ihre unbestreitbar nahe
Verwandtschaft zu den Spaltpilzen und durch die Rolle, welche sie im
Flechtenthallus spielen.
Die Spaltpilze kommen wesentlich in denselben Formen vor, und nur
die Farbe der Zellen geht ihnen ab. Wir haben aber in vielen Fällen
so genau dieselbe Anordnung der Zellen, wie bei Merismopedia,
bei Leptothrix und ähnlichen, dass die Gattungen sowohl bei
Spaltalgen wie bei Spaltpilzen aufgenommen wurden und man zu diesen
die farblosen, zu jenen die gefärbten Formen stellt. Aus anderen
Gründen, deren Erörterung hier zu weit führen würde, ist man noch mehr
gezwungen, in beiden Gruppen die nächsten Verwandten zu erblicken und
nur Zweckmässigkeitsgründe lassen es erwünscht erscheinen, sie noch aus
einander zu halten.
Dass die als Gonidien bezeichneten grünen Zellen des Flechtenkörpers
wirklich nur Algenzellen sind, wird wohl von niemandem mehr ernstlich
bezweifelt, der sich irgendwie eingehend mit diesen Fragen beschäftigt
hat. Nur einige ältere Flechtensystematiker können sich noch nicht zu
dieser Ansicht bekehren, denn sie wollen die Selbständigkeit derjenigen
Pflanzen, welchen sie soviel Zeit und Arbeit gewidmet haben, nicht
gern aufgeben. Thatsächlich sind die Flechten aber keine einheitlichen
Organismen, sondern durch ein höchst eigentümliches und interessantes
Zusammenleben von Pilzen und Algen entstandene Pflanzengebilde. Indem
die Sporen gewisser Pilze auf eine Algenvegetation fallen, treiben
sie Hyphen, mit welchen sie die Zellen umspinnen und vollständig
einschliessen. Die Algen nehmen übrigens, wie es scheint, dabei
durchaus keinen Schaden, sondern befinden sich ganz wohl dabei, teilen
sich in demselben Verhältnis wie die Hyphen, wachsen und liefern diesen
die[S. 40] nötigen organischen Stoffe, während sie anderseits von ihnen ihre
Feuchtigkeit und ihre anorganischen Baustoffe den Pilzen verdanken.
Dieses als Symbiose bezeichnete seltsame Zusammenleben zweier
so ganz verschiedener Organismen hat die Förderung jedes einzelnen
ohne Benachteiligung des anderen zum Zweck. Allerdings sind dabei die
Pilze vollständig auf die Algen angewiesen, denn ihre Sporen gehen zu
Grunde, wenn sie nicht die entsprechenden Algen finden, während die
letzteren ebensogut auch ohne Pilze leben können. Das ist ja gerade
einer der Hauptbeweisgründe für die Algennatur der Flechtengonidien,
dass man dieselben Zellen auch ausserhalb des Flechtenthallus kennt,
und dass man sie hier zu selbständigen Gattungen und Arten erhob,
während sie in der Flechte durchaus nur unselbständige Teile des
Thallus sein sollten. Übrigens ist es nicht unmöglich, dass ein
grosser Teil der als „Raumparasitismus“ im Tier- und Pflanzenreich
bezeichneten Erscheinungen auf wirkliche Symbiose zurückzuführen
ist, bei welcher sowohl Wirt als Gast ihre Rechnung finden würden
und sich gewissermassen zu gegenseitiger Unterstützung verbunden
haben. Die meisten der bei der Flechtenbildung beteiligten Algen
gehören den Cyanophyceen an, einige den Grünalgen, besonders der
Gattung Protococcus; aber auch grüne Fadenalgen treten im
Flechtenthallus auf, so eine Cladophora in der tropischen
Gattung der Fadenflechten Coenogonium.
Die Spaltalgen bilden mit den Spaltpilzen zusammen eine gegen die
übrigen Pflanzen streng abgeschlossene Gruppe, ohne jeden vermittelnden
Übergang. Mit dem Tierreich werden sie fast unmerklich und ohne
dass eine scharfe Grenze gezogen werden kann, durch die Gruppe der
Monadinen verbunden.
2. Bacillariaceen.
Höher organisiert als die Spaltalgen, stehen sie doch allen anderen
Gruppen der Algen als ein streng abgeschlossenes Ganzes gegenüber und
zeigen überhaupt eine ganz isolierte, durch keinerlei Beziehungen zu
anderen Organismen vermittelte Stellung im Reiche der lebenden Wesen.
[S. 41]
Unter dem Mikroskop sind sie leicht erkennbar; ihr durch einen
Kieselpanzer geschützter Zellinhalt wird aus farblosem Protoplasma
gebildet, in welchem in bestimmter Stellung braune Körner oder Platten
auftreten, deren Farbstoff, Diatomin genannt, das Chlorophyll vertritt.
Weit mehr fällt aber der Kieselpanzer selbst in die Augen, da er meist
eine feine Zeichnung trägt, welche die Diatomeen oder Bacillarien zu
den zierlichsten Geschöpfen macht. Diese Zeichnung tritt besonders
schön bei manchen marinen Arten auf, in Form von sich kreuzenden
Liniensystemen oder dicht aneinanderschliessenden Sechsecken; sie
sind bei den in Fig. 2 (S. 42) abgebildeten Formen (ebenso wie der
Zellinhalt) weggelassen, weil die Abbildungen sonst zu grossen Raum
beansprucht haben würden. Die Linien liegen bei manchen Arten so nahe,
dass es nur den besten Mikroskopen gelingt, sie aufzulösen, d. h.
getrennt von einander deutlich sichtbar zu machen; deshalb werden
gewisse Diatomeen wie Pleurosigma angulatum und Surirella
gemma zur Prüfung der besten Objektive verwendet.
Die überaus zierlichen Kieselalgen verdienen übrigens wegen ihrer
Schönheit und Mannigfaltigkeit eine eingehendere Beobachtung auch
von solchen, denen irgend eine naturwissenschaftliche Beschäftigung
erwünscht ist und welche sich im Besitz eines Mikroskopes befinden,
oder sich ein solches anschaffen können (vergl. die Anmerkung am
Schluss). Der Formenreichtum ist ein ausserordentlicher und wenn bei
den Spaltalgen die Einförmigkeit der ganzen Gruppe auffiel, so tritt
uns bei den Kieselalgen eine Vielgestaltigkeit entgegen, wie sie
ausgeprägter kaum in einer anderen Pflanzenklasse vorkommt. Die Figur
2 giebt uns einen Überblick über die verschiedenen Formen, welche
unsere süssen Gewässer bewohnen; freilich konnte nur ein kleiner Teil
berücksichtigt werden.
In Nr. 1 tritt uns eine der vielen schwer unterscheidbaren Arten der
Gattung Pinnularia entgegen. Sie sind fast symmetrisch gebaut
und mit deutlicher, in der Regel starker Streifung, welche auch in der
Figur angegeben ist. Sehr ähnlich ist Navicula von kahnförmiger
Gestalt (Nr. 2), die Streifung ist aber sehr zart und oft nur mit
den besten Linsen zu erkennen. Die Streifen sind[S. 42] hier aus dichten
reihenförmigen Punkten gebildet. Bei der Gattung Stauroneis (Nr.
3) ist ein deutliches helles Kreuz erkennbar, welches die ebenfalls
kahnförmige Zelle am Längs- und Querdurchmesser trägt. Eigentümlich
gebogen sind die Zellen der Gattung Pleurosigma (Nr. 4)
und nur nach einer Richtung symmetrisch die stark gestreiften von
Cymbella (Nr. 5).
Fig. 2.
Kieselalgen, Bacillariaceen. 1. Pinnularia — 2. Navicula — 3.
Stauroneis — 4. Pleurosigma — 5. Cymbella — 6.
Amphora — 7. Gomphonema — 8. Nitschia — 9. Surirella
— 10. Synedra — 11. Epithemia — 12. Meridion — 13.
Fragillaria — 14. Diatoma — 15. Melosira — 16.
Campylodiscus (a von der Seite, b von oben). Stark
vergrössert.
Bei Amphora (Nr. 6) bildet die Streifung zwei eigentümliche
Bänder, während andere Teile des Kieselpanzers ungestreift bleiben.
Bei der Gattung Gomphonema sitzen die einzelnen Zellen auf
Gallertstielen, welche ein vielfach verzweigtes Bäumchen darstellen
(Nr. 7).[S. 43] Eigentümliche ovale oder verzogene Zellen werden durch die
Gattung Surirella (Nr. 9) repräsentiert, Synedra bildet
meist lange, nadelförmige Zellen, welche oft wie die Speichen eines
Rades zusammensitzen (Nr. 10). Die Gattung Epithemia (Nr. 11)
erinnert etwas an Cymbella, ist jedoch schon durch die Streifung
leicht unterschieden. Dann giebt es eine Gruppe von Diatomeen, deren
Zellen zu Fäden verbunden bleiben, wie bei Fragillaria,
Diatoma, Meridion, Melosira (Nr. 12–15), noch
andere bilden schildförmige, mehr oder weniger gebogene Platten, wie
Campylodiscus (Nr. 16). Aber auch nur einigermassen genaue
Beschreibung der deutschen Gattungen zu liefern, ist an diesem Ort
wegen des Formenreichtums und der Vielgestaltigkeit der Diatomeen
unmöglich. Dagegen soll uns noch ein Blick in das Leben dieser
zierlichsten aller Geschöpfe vergönnt sein.
Der Kieselpanzer einer Diatomee ist kein einheitliches Gebilde, sondern
besteht aus zwei sehr ähnlichen Hälften, die sich nur durch eine
geringe Grössendifferenz unterscheiden, sonst aber, namentlich in der
Zeichnung, vollständig übereinstimmen. Diese beiden Hälften sitzen in
einander wie die Teile einer Schachtel, was man bei günstigen Objekten
direkt sehen kann. Nimmt die Zelle an Volumen zu, so kann dies nur
dadurch geschehen, dass die beiden Teile etwas auseinanderweichen, da
ja die starren Kieselschalen ein Wachstum in die Länge oder Breite
verhindern. Endlich kommt bei dieser Volumenzunahme der Zelle aber
ein Stadium, in welchem die beiden Hälften oder Schalen nicht mehr
ineinandergreifen, sondern die Zelle nicht mehr vollständig bedecken
und einen schmalen Streif Plasma zwischen ihren Rändern freilassen.
Dann bilden sich an dieser Stelle zwei neue Schalen, von denen sich
die eine der grösseren, die andere der kleineren der alten Schalen
ebenso einfügt, als diese es ursprünglich waren, und aus der einen
Diatomee sind bei diesem Vorgange zwei geworden, welche in jeder
Beziehung dem Mutterindividuum gleichen, nur ist die eine um die Dicke
einer Schale kleiner als jene. Eine derartige Verkleinerung muss immer
erfolgen, weil die Kieselschalen starr sind und sich die jüngere Schale
immer[S. 44] der ältern einfügt. Bei weiteren Teilungen werden die jüngeren
Individuen mit der jüngern Schalenhälfte immer kleiner und wir sehen
oft von derselben Art Exemplare, die um mehr als das Doppelte in der
Länge von einander abweichen. Die Verkleinerung findet aber auch
naturgemäss ihre Grenze; ist die Grösse der Individuen bis auf ein
bestimmtes Mass herabgesunken, so teilen sie sich nicht weiter, sondern
es erfolgt eine Art Regeneration durch einen Vorgang, den man als
Auxosporenbildung bezeichnet hat.
Die Auxosporenbildung tritt in drei verschiedenen Modifikationen auf;
entweder findet eine wirkliche Befruchtung durch die Verschmelzung
zweier Individuen statt, oder es findet nur eine Berührung statt,
oder endlich ein einziges Individuum schickt sich dazu an. Dieser
letzte Vorgang ist der einfachste, er ist am häufigsten bei Melosira
beobachtet worden. Eine der Zellen eines Fadens treibt unter rascher
Volumenzunahme die beiden Schalen ohne sich zu teilen aus einander,
tritt zwischen diesen teilweise hervor und bildet eine Kugel, welche an
zwei Punkten noch in den alten Schalen stecken bleibt, aber mehr als
den doppelten Durchmesser als diese hat. Erst jetzt teilt sich diese
„Auxospore“ und umgiebt sich mit Kieselschalen, welche zunächst von
denjenigen einer Melosira noch in der Gestalt wesentlich abweichen,
da sie Hälften einer nicht ganz regelmässigen Kugel sind. Aber schon
die beiden nächsten Schalen, welche sich innerhalb der Auxospore
entwickeln, nehmen die normale cylindrisch-schachtelförmige Gestalt an
und die beiden ersten aus der Auxospore entstehenden Zellen tragen je
eine halbkugelige und eine normale Schale. Die folgenden Individuen
sind wieder mit normalen Schalen versehen, nur zwei Nachkommen
behalten immer eine normale und eine Kugelschale. Bei einer Anzahl
anderer Diatomeen ist wenigstens eine Berührung zweier Individuen zur
Auxosporenbildung erforderlich. Die Schalen derselben weichen dann aus
einander, die Plasmamassen treten hervor und nehmen ausserordentlich
rasch an Volumen zu, während sich die zarte, nicht kieselhaltige
Membran dieser Plasmamassen stark dehnt. Erst wenn ein Wachstum nicht
mehr stattfindet,[S. 45] scheiden sich die Kieselschalen aus und die beiden
so entstandenen Zellen haben dann eine mehr als doppelte Länge wie
die Mutterzellen. Die Einzelheiten bei diesem Vorgang sind nach den
Gattungen verschieden und oft recht kompliziert. Noch andere Diatomeen
lassen ihre aus den Schalen ausgetretenen Plasmamassen wirklich
verschmelzen, wodurch ein der Konjugation der Spirogyren ähnlicher
Prozess herbeigeführt wird. Die konjugierte Plasmamasse wächst dann
bedeutend in die Länge und scheidet eine einzige zusammenhängende und
die ganze Zelle umschliessende Kieselmembran aus, innerhalb deren sich
die beiden Schalen einer neuen Zelle entwickeln. Dieselben werden durch
die Volumenzunahme der Zelle auseinandergetrieben und sprengen dann die
kieselhaltige harte Membran der Auxospore, wodurch die neue Zelle frei
wird und sich in normaler Weise weiter teilt. In allen Fällen hat die
Auxosporenbildung nur den Zweck, aus der allmählich zu klein gewordenen
Generation eine neue, grössere zu bilden, sie dient nicht ähnlichen
Zwecken wie etwa die Dauersporen der Schizophyceen, sondern es tritt
zwischen Auxosporenbildung und der Entwickelung neuer Zellen keinerlei
Ruhezustand ein.
Eigentliche Dauerzustände, welche durch besondere Zellen repräsentiert
werden, kommen bei den Diatomeen nicht vor, dagegen können Zellen unter
günstigen Verhältnissen auch eine längere Austrocknung ertragen, ohne
abzusterben. Das Plasma zieht sich dabei in eine Ecke der Kieselschalen
zurück und umgiebt sich mit einer Membran, bei Eintritt günstiger
Lebensbedingungen die normalen Funktionen wieder aufnehmend.
Eine eigentümliche, aber noch nicht aufgeklärte Eigenschaft der
Diatomeen ist ihre Bewegungsfähigkeit, welche ihnen jedoch nur
zukommt, wenn sie festem Substrat aufliegen, sie kriechen also an
demselben herum. Diese Thatsache macht es wahrscheinlich, dass die
Bewegung durch Plasmafäden vermittelt wird, welche durch Öffnungen
in den Kieselpanzern oder zwischen den Schalen hervortreten und sich
dem Substrat anheften; man hat jedoch bisher noch keinerlei derartige
Bewegungsorgane wahrnehmen können. Die[S. 46] Bewegung selbst ist eine
gleitende, oft ruckweise, meist genau der Längsrichtung des Körpers
folgend.
Die Diatomeen sind die einzigen Algen, welche sich seit der Zeit ihres
Auftretens auf der Erde in ihren Kieselschalen unverändert erhalten
haben, so viel Jahrtausende auch über ihre Grabstätten dahingerauscht
sind. Ihre Schalen haben auch so manche Proben bestanden, bei
denen nur wenig andere Geschöpfe nicht der völligen Vernichtung
anheimfielen. Von kleinen mikroskopischen Tieren verschluckt, wurden
die Diatomeen hier zum ersten Male verdaut, gerieten mit diesen in den
Magen von grösseren Weichtieren und mussten den Prozess zum zweiten
Male durchmachen. Zum dritten Male wurde ihnen das Los zu teil,
wenn diese Weichtiere von Fischen verspeist wurden, um mit diesen
zum vierten Mal von Seevögeln verdaut zu werden. Nichtsdestoweniger
passierten ihre Schalen unverändert den Darmkanal der Vögel und im
Guano finden sich dieselben noch in schönster Erhaltung. Bei solcher
Widerstandskraft ist es kein Wunder, dass sich die Diatomeen, wo sie
sich in Masse entwickelten, auch noch heute an diesen Orten fossil
vorfinden, nur das ist thatsächlich wunderbar, dass diese kleinsten
Organismen — durchschnittlich gehen 10 bis 20 Millionen Individuen auf
1 Kubikmillimeter — meilenweite und viele Fuss tiefe Lager bilden,
wie bei uns in Deutschland namentlich in der Lüneburger Heide,
bei Berlin und bei Königsberg. Die als Tripel, Mergel,
Kieselguhr und Infusorienerde bezeichneten Fossilien sind wesentlich
nichts anderes als die Kieselschalen der Diatomeen, deren zierliche
Form unter dem Mikroskop sich sofort verrät.
3. Die Chlorophyceen.
Die Grünalgen, Chlorophyceen, Algen im engern Sinne, sind im
wesentlichen hinreichend durch die rein grüne Farbe gegenüber den
anderen Gruppen charakterisiert. Gestalt und Fortpflanzungsweise sind
so mannigfach, dass sie sich schlecht zu einem Merkmal der ganzen
Gruppe verwenden lassen und selbst nahe verwandte Gattungen zeigen
hierin die grössten Verschiedenheiten.
[S. 47]
Zunächst finden wir eine Gruppe grüner Algen, welche in der
Fortpflanzung mit dem einen Modus derselben bei den Diatomeen
übereinstimmen, so dass sie, aber gewiss entschieden unrichtig, mit
jenen zu einer grossen Ordnung „Zygosporeae“ von manchen
Forschern vereinigt werden. Indem nämlich zwei (oder zuweilen auch
mehr) Zellen ihren Inhalt zusammenfliessen lassen, entsteht eine
als „Zygospore“ bezeichnete Zelle, welche dazu bestimmt ist, nach
einer Ruheperiode zu keimen und eine neue Generation zu entwickeln.
Es ist also eine Dauerzelle und unterscheidet sich schon hierdurch
sehr wesentlich von den Auxosporen der Diatomeen, welche letztere
nur als ein Mittel zur Vergrösserung der Zellen dient und keinerlei
Eigenschaften besitzt, wie sie den Sporen der Kryptogamen im
allgemeinen zukommen. Ausserdem kommt es nur bei einem Teile der
Diatomeen zu einer wirklichen Vereinigung der Plasmakörper zweier
Zellen, wie wir oben gesehen haben. Es ist deshalb besser, man
lässt diese Gruppe grüner Algen, welche man als Conjugatae
zusammenfasst, als unterste Ordnung der Chlorophyceen bestehen.
Eine zweite Ordnung wird durch eine Anzahl einzelliger Algen gebildet,
welche teilweise einen Übergang zu den Flagellaten erkennen lassen
und aus teils freien, teils zu Kolonien vereinigten rundlichen oder
eiförmigen meist ziemlich kleinen Zellen bestehen. Man hat sie als
Protococcoideae bezeichnet. Ihre Fortpflanzung ist sehr
verschiedenartig und oft höchst kompliziert.
Die Siphoneae oder Schlauchalgen zeichnen sich durch
einen einzelligen fadenförmigen oder verästelten Bau aus; sie sind im
süssen Wasser nur durch zwei Gattungen vertreten.
Reich an Formen ist die vierte Ordnung der Grünalgen, die
Confervoideae. Sie bilden einen fadenförmigen, oft reich
verzweigten, vielzelligen Thallus.
Nach dieser kurzen Charakteristik der Hauptgruppen sollen die einzelnen
eine etwas eingehendere Besprechung erfahren, da sich ja in der Klasse
der Grünalgen die interessantesten und mannigfaltigsten Formen,
die verschiedenartigsten Verhältnisse der Fortpflanzung und die
merkwürdigsten Lebenserscheinungen finden.
[S. 48]
Die Conjugaten bilden eine Ordnung, welche zwei nahe verwandte Familien
vereinigt. Gemeinsam ist ihnen besonders die Art der geschlechtlichen
Fortpflanzung, die Konjugation, ferner die eigentümliche Anordnung des
Chlorophylls innerhalb der Zelle in Bändern, Platten, Sternen oder
anderen Formen, stets aber abweichend von derjenigen bei den anderen
Grünalgen, ferner im weitesten Sinne die Einzelligkeit. Jede Zelle ist
befähigt für sich allein zu existieren und sich zu vermehren, selbst
wenn sie normalerweise nur mit anderen zusammen zu Fäden verbunden
vorkommt. Die Zellen eines Fadens sind auch sämtlich gleichartig und
es ist kein Gegensatz zwischen Basis und Spitze ausgebildet. Eine sehr
ausgiebige Vermehrung geschieht durch Zweiteilung der Zellen, die bei
diesen Algen besonders gut zu beobachten ist.
Die Desmidieen, die eine der beiden Familien, zeigen äusserst
zierliche Formen, wie sie in Fig. 3 a–m dargestellt
sind. Sie bewohnen einzeln unsere Wiesengräben, Bäche und Flüsse,
zahlreicher und oft geradezu massenhaft die Gewässer der Torfmoore,
und erreichen im Spätsommer und Herbst ihre grösste Entwickelung. Die
Zellen sind symmetrisch, bei vielen mit einer Einschnürung in der
Mitte, meist einzeln, seltener zu leicht zerreissenden Fäden verbunden.
Das Chlorophyll ist entweder sternförmig um zwei stärkehaltige Körper
auf die beiden Hälften der Zelle verteilt, zentral, d. h. der Wand
nicht anliegend, oder tritt in Form von Platten, welche ebenfalls
der Wand nicht anliegen, oder endlich in mehr oder weniger spiralig
gerollten der Wand anliegenden Bändern auf. Bei der Vermehrung treten
die beiden Zellhälften an der Einschnürung aus einander und zwischen
ihnen entwickelt sich an jeder eine neue Zellhälfte, die ursprünglich
rundlich und glatt ist und erst allmählich die Form der andern
Zellhälfte bei weiterem Wachstum erhält (vergl. Fig. 3 e).
Bei der als Kopulation bezeichneten geschlechtlichen Fortpflanzung
tritt der Inhalt zweier Zellen aus der Membran hervor und vereinigt
sich zu einer einzigen, den Mutterzellen durchaus unähnlichen Zelle,
welche den Namen Zygospore führt (vergl. Fig. 3 f Zygospore von
Micrasterias) und einen Ruhezustand darstellt; bei der Keimung[S. 49]
entwickeln sich aus ihr eine bis vier, meist zwei Keimzellen, aus
denen dann wieder durch Teilung die gewöhnlichen Zellen entstehen. Die
Desmidieen besitzen ein geringes vom Licht abhängiges Bewegungsvermögen.
Fig. 3.
Conjugaten a–m. Desmidiaceen: aDesmidium, Faden
von der Seite — b Dasselbe, Faden auf dem Querschnitt — cEuastrum — dMicrasterias denticulata — e Dasselbe
in der Teilung begriffen — f Zygospore desselben — hMicrasterias crenata — gCosmarium — iStaurastrum — kXanthidium — lPenium —
mClosterium — n–r Zygnemaceen: nZygnema — oSpirogyra — pZygnema in
Kopulation, 1–7 bezeichnen die verschiedenen Stufen derselben bis zur Bildung
der Zygospore (7) — q und r Zygosporen von Zygnemaceen.
Eine Beschreibung der einzelnen Gattungen lässt sich an dieser
Stelle nicht geben; für die Abbildung wurden einige der zierlichsten
Desmidieen gewählt, in denen der Inhalt der Zellen wegen der geringen
Vergrösserung nur angedeutet oder ganz weggelassen wurde.
Die zweite Familie, die Zygnemaceen, wird aus fadenförmigen
unverzweigten Algen gebildet, deren Chlorophyll zu spiraligen Bändern
(Spirogyra), Doppelsternen (Zygnema, Zygogonium),
axilen Platten (Mongeotia, Mesocarpus) zusammentritt.
Ihre geschlechtliche Fortpflanzung ist derjenigen der Desmidieen
sehr ähnlich, nur kopulieren meist Zellen verschiedener Fäden mit
einander, indem sie bei leiterförmiger Kopulation Ausstülpungen gegen
einander treiben (wie dies Fig. 3 p in verschiedenen Stadien
der Kopulation dargestellt ist), oder indem sich die Zellen bei
knieförmiger Kopulation[S. 50] direkt an einander legen, was durch eine
Biegung der Fäden gegen einander bewirkt wird. Selten kopulieren zwei
neben einander liegende Zellen ein und desselben Fadens (seitliche
Kopulation), indem sie ebenfalls Ausstülpungen gegen einander treiben.
Haben sich die Ausstülpungen an einander gelegt, so beginnt eine
Veränderung in den Zellen, der Inhalt ballt sich zusammen, indem Wasser
ausgestossen wird, und derjenige der einen Zelle fliesst in die andere
hinüber, um sich mit dem Inhalt dieser zu vereinigen, oder beide
Plasmamassen treten aus den Zellen hervor und treffen sich zwischen
den Fäden. Im erstem Falle entsteht das Produkt der geschlechtlichen
Vereinigung — die Zygospore — in einer der Mutterzellen, im andern
zwischen den beiden Mutterzellen. Die Zygosporen der Zygnemaceen sind
weit weniger mannigfaltig als die der Desmidiaceen; auch sie machen
eine Ruheperiode durch, nach welcher sie keimen. Die Zygnemaceen finden
sich im ersten Frühjahr, sowie nur das Eis unsere Teiche und Tümpel
verlassen hat, überall in stehenden Gewässern als schleimige hellgrüne
Flocken oder Fadenmassen.
Beide Familien entwickeln keine beweglichen Fortpflanzungszellen,
wie die meisten übrigen Grünalgen; einige, namentlich die grösseren
Spirogyreen und Desmidieen, lassen aber im Innern der Zelle deutlich
die Bewegung des Plasmas erkennen. Bei Closterium findet sich
am Ende der beiden halbmondförmig gekrümmten Zellhälften ein farbloses
Bläschen, in welchem eine Anzahl kleiner (aus Gips? bestehender)
Körnchen sich sehr lebhaft bewegen (Fig. 3 m). Überhaupt
bieten die Zellen der Conjugaten viele interessante Eigenheiten und
sie sind auch, besonders die Gattung Spirogyra, ein sehr
vielfach benutztes Objekt der Pflanzenphysiologen, weil die Grösse
und Einfachheit der Zellen leichter eine eingehende Untersuchung
ermöglicht. Besonders zu Studien über den Zellkern liefern sie ein
vorzügliches Material und wir finden da, um eine Eigentümlichkeit zu
erwähnen, bei Spirogyra beispielsweise einen grossen, flach
scheibenförmigen, spindelförmig erscheinenden Zellkern, welcher mit an
den Wänden befestigten Plasmafäden in der Mitte der Zelle aufgehängt
ist.
[S. 51]
Die zweite Ordnung der Grünalgen, die Protococcoideae, fasst
eine Gruppe sehr verschiedenartiger Organismen zusammen. Gemeinsam ist
ihnen die Einzelligkeit und die Entwickelung von Schwärmzellen. Eine
der drei hierhergehörigen Familien, die Volvocineen, zeigen eine solche
Verwandtschaft zu den Flagellaten, einer Gruppe der niedersten Tiere,
dass sie von den Zoologen einfach mit jenen vereinigt werden, und auch
in diesem Buche unter jenen abgehandelt werden sollen[I]. Scheiden wir
also an dieser Stelle die später in einem eigenen Kapitel eingehend
behandelten Volvocineen hier aus, so bleiben zwei sehr ähnliche
Familien übrig, Protococcaceae und Palmellaceae.
[I] Der Verfasser hält jedoch an der Ansicht fest, dass die
Volvocineen ebensogut zu den Algen gestellt werden können, zu welchen
sie unbestritten nahe Verwandtschaft zeigen.
Die Protococcaceae sind einzellige Algen, denen eine Teilung der
Zellen zum Zweck einer vegetativen Vermehrung abgeht, welche dagegen
eine ungeschlechtliche Fortpflanzung durch Schwärmsporen (Zoosporen)
und eine geschlechtliche durch Kopulation von Schwärmsporen besitzen.
Die Zellen bleiben dabei entweder zu Familien vereinigt oder sie
lösen sich los und werden frei. Einen Vertreter der ersteren, das
Wassernetz Hydrodyction utriculatum wollen wir als Beispiel
wählen, dabei aber zugleich bemerken, dass wir hier einen viel
komplizierteren Entwickelungsprozess vor uns haben, als bei den
meisten übrigen Gattungen der Familie. Das Wassernetz (Fig. 4
a–d S. 52) bildet ein wirkliches Netz, dessen Maschen
von einer Anzahl cylindrischer Zellen umschlossen sind, welche zu
je drei an einem Punkte zusammentreffen (a). Die Chlorophyll
führende grüne von Vacuolen unterbrochene Plasmaschicht kleidet die
ganze Zellwand aus, ballt sich aber, wenn es zur Fortpflanzung kommt,
in sehr kleine, zunächst ebenfalls der Wand anliegende Plasmaportionen
zusammen. Die Fortpflanzung geschieht nun entweder geschlechtlich
oder ungeschlechtlich. Im letztern Falle sind die Plasmaportionen, in
welche der Zellinhalt zerfällt, grösser; anfangs eckig, runden sie
sich allmählich ab (b) und bilden sich zu Schwärmsporen um,
welche eiförmige Gestalt und zwei Geisseln besitzen. Auch zeigt sich
ein Gegensatz zwischen dem geisseltragenden[S. 52] Ende, welches durchsichtig
und hell, und dem geissellosen, welches grün und mit körnigem Plasma
erfüllt ist. Man bemerkt auch ferner einen kleinen roten Punkt in
dem hellen Teil, den Pigmentfleck, über dessen Bedeutung man nur
Vermutungen hegt, vielleicht stellt er den Anfang eines Sinnesorganes
vor. Diese[S. 53] Zellen, Makrogonidien genannt, schwärmen in der Mutterzelle
einige Zeit umher, kommen aber allmählich zur Ruhe, indem sie sich
zu gleicher Zeit in derselben Weise ordnen wie die Zellen des
Mutternetzes und ein kleines diesem sehr ähnliches Netz bilden,
welches, ohne dass durch Teilung neue Zellen darin entstehen, wächst
und dem Mutterzellnetz völlig gleich wird. Die umhüllende Membran der
Mutterzelle zerfliesst schon sehr früh in dem umgebenden Wasser (vergl.
Fig. 4 c).
Fig. 4.
Protococcoiden. a–f Protococcaceen: a–dHydrodyction utriculatum: a Teil eines Zellnetzes — b
Beginnende Makrogonidienbildung, Teil einer Zelle, stärker vergrössert —
c Stück einer Zelle, in welcher sich die Makrogonidien zu einem neuen
Netz an einander lagern, stärker vergrössert — d Zur Ruhe gekommener
Schwärmer (vergl. Text) — e Verschiedene Formen von Scenedesmus —
fPediastrum — g–h Palmellaceen: gCosmocladium — hRaphidium (e–h sind nur
Umrisszeichnungen).
In derselben Weise wie die Makrogonidien, nur in viel grösserer Anzahl,
bilden sich die Mikrogonidien, welche viel kleiner sind und vier
Geisseln besitzen. Sie verlassen aber die Mutterzelle und schwärmen
im Wasser umher, kopulieren und verlieren allmählich ihre Bewegung,
indem sie zu rundlichen grünen Zellen werden, welche sich allmählich
vergrössern. Diese Zellen stellen einen Ruhezustand dar, aus welchem
sich nach einiger Zeit wenige zweigeisselige Schwärmzellen entwickeln,
welche ebenfalls austreten, umherschwärmen und ihre Bewegung verlieren,
aber sich nicht zu runden, sondern zu eigentümlich eckigen mit
unregelmässigen Auswüchsen versehenen Zellen (d) entwickeln,
in denen wieder die oben beschriebenen Makrogonidien entstehen und zu
einem kleinen Netz zusammentreten.
So vielgestaltig ist der Entwickelungsgang bei den übrigen
Protococcaceen in der Regel nicht, wir besitzen jedoch von den meisten
noch keine genügende Kenntnis desselben. Von den übrigen Gattungen der
Familie finden sich häufiger Pediastrum, dessen in einer Ebene
gelagerte Zellen zu zierlichen, meist mit kleinen Hörnchen verzierten
Täfelchen zusammentreten (f), Scenedesmus, dessen
Zellen zu vier und acht mit der Längsseite aneinandergelagerte Reihen
bilden und ebenfalls häufig Hörnchen oder Dornen zeigen (e)
und Protococcus, welcher einfache nicht zusammenhängende runde
Zellen bildet und häufig den grünen pulverigen Überzug von Baumstämmen,
Holzzäunen und Schindeldächern bildet, aber in der heissen Jahreszeit
verschwindet.
Die Palmellaceen sind äusserlich oft sehr schwer von den Algen
der vorigen Familie zu trennen, unterscheiden sich aber in[S. 54] ihrer
Entwickelungsgeschichte sehr wesentlich von jenen durch die Fähigkeit,
sich durch einfache vegetative Zweiteilung zu vermehren, und durch
den Mangel einer geschlechtlichen Fortpflanzung. Sie zeigen ebenfalls
sehr zierliche Formen, wie die in Fig. 4 abgebildeten Vertreter dieser
Familie: Cosmocladium (g) und Raphidium (h)
darthun.
Eine andere Ordnung einzelliger Grünalgen wird durch die
Siphoneen (Schlauchalgen) gebildet, welche jedoch nur in
wenig Arten im süssen Wasser auftreten. Sie haben eine fadenförmige,
langgezogene, reich verzweigte, oder eine auf verästelter Basis ruhende
blasenförmige Gestalt, ohne dass sich jedoch zwischen den Ästen oder
Verzweigungen Querwände in der Zelle fänden. Ihre Fortpflanzung
geschieht auf geschlechtlichem und ungeschlechtlichem Wege. Die Gattung
Vaucheria findet man in Gräben oder auf feuchter, schattiger
Erde, zarte Fadenpolster bildend, die Gattung Botrydium
namentlich auf überschwemmt gewesenen schlammigen Flächen als kleine
stecknadelkopf- bis hanfkerngrosse schwarzgrüne Kügelchen.
In der vierten Ordnung treten uns die höchstentwickelten Formen der
Grünalgen, die Confervoideae, entgegen. Auch bei ihnen herrscht
in Gestalt und Fortpflanzungsweise die grösste Mannigfaltigkeit.
Jedenfalls sind es stets mehrzellige Algen, deren Zellen meist in
Fäden, oft mit Verzweigung, seltener in flächenförmigen Lagern
angeordnet sind. Ihre Fortpflanzung ist noch nicht bei allen Gattungen
bekannt, doch entwickeln wahrscheinlich alle bewegliche Schwärmzellen,
welche teils ungeschlechtlicher Natur sind, teils zu mehr oder weniger
deutlich ausgeprägten Geschlechtszellen werden.
Die einfachsten Formen aus dieser Ordnung bilden die eigentlichen
Conferven, welche einfache Zellfäden bilden, sich von den Zygnemaceen
aber leicht dadurch unterscheiden, dass bei ihnen das Chlorophyll
gleichmässig der Zellwand anliegt. Bei der Gattung Conferva hat
man bisher nur ungeschlechtliche Fortpflanzung durch Schwärmsporen
wahrgenommen, während bei der sehr ähnlichen Ulothrix ausser
dieser auch eine geschlechtliche Fortpflanzung insofern auftritt, als
zwei kleinere Schwärmsporen mit einander[S. 55] verschmelzen (kopulieren)
und das Produkt der Kopulation eine Dauerzelle ist, welche eine
Ruheperiode durchmacht ehe sie keimt. Verzweigte Zellreihen finden
wir bei den artenreichen Cladophoren, deren Entwickelungsgeschichte
noch sehr wenig bekannt ist, trotzdem dass sie zu den häufigsten
Süsswasseralgen gehören. Namentlich die Gattung Cladophora
selbst, deren Artenstudium beinahe für sich allein eine Wissenschaft
bildet und welche das Hieratium der Kryptogamen ist, findet sich
in ihren verschiedenen Formen überall, und in Quellen und reinen
Bächen werden wir häufig der überaus zierlichen etwas schlüpfrigen
Draparnaldia begegnen. An Schilf und Holzstücken setzen
sich gern kleine grüne Kugeln oder dem Geweih eines Hirsches
ähnlich verzweigte gallertartige Körper fest, welche der Gattung
Chaetophora angehören. Auch das im Gebirge allbekannte
Veilchenmoos oder der Veilchenstein, Chroolepus Jolithus, gehört
hierher, obgleich seine braunrote Farbe gar nicht zu den Grünalgen
zu passen scheint. Dies kommt aber daher, dass neben dem Chlorophyll
noch ein anderer Farbstoff, der in überwiegender Menge vorhanden ist,
den Zellen die Färbung verleiht. Der Veilchengeruch, den diese Alge
ausströmt, besitzt unter Umständen auch noch nach Jahren eine ziemliche
Intensität.
Von den übrigen Familien dieser Ordnung wollen wir noch die
Oedogoniaceen einer eingehenderen Berücksichtigung unterziehen, weil
ihre Fortpflanzung und Entwickelungsgeschichte von besonderem Interesse
ist und man die Vorgänge dabei auch verhältnismässig genau kennt. Die
Gattung Oedogonium zeigt einfache unverzweigte Zellfäden, in
denen schon eine sehr eigentümliche Zellteilung auftritt. Es bildet
sich nämlich in der Zelle ein ringförmiger Wulst von dem Stoff der
Zellwand, welcher unter dem Mikroskop allerdings nur an beiden Seiten
der Zelle sichtbar ist (Fig. 5, 1 a). Plötzlich reisst die
Membran der Mutterzelle an der Stelle dieses Wulstes auf und die Zelle
streckt sich um das Doppelte in die Länge, der Wulst verschwindet,
indem er zu einer anfangs sehr dünnen Zellwand für das neuzugewachsene
Zellstück wird (Nr. 1 b). Erst dann bildet sich zwischen
der alten und der neuen Zellhälfte eine Scheidewand. Der Riss der
Mutterzellmembran bleibt übrigens[S. 56] dauernd sichtbar und oft sieht man
Zellen, die ein förmliches Schraubengewinde zu tragen scheinen. Die
ungeschlechtliche Fortpflanzung geschieht in der Weise, dass sich der
Inhalt einer vegetativen Zelle zusammenballt, abrundet und, indem sich
die Zelle durch einen kreisförmigen Riss (Nr. 2) öffnet, austritt.
Sofort beginnt er vermöge eines Wimperkranzes (vergl. Nr. 3) sich
zu drehen und davonzuschwimmen. Schliesslich setzt er sich irgendwo
fest, entwickelt eine Haftscheibe am unteren Ende und wächst zu einem
Zellfaden aus.
Fig. 5.
Entwickelung von Oedogonium (vergl. Text).
Wesentlich verwickelter ist die geschlechtliche Fortpflanzung und
hier kommen in einer Gattung so viel Verschiedenheiten vor, wie wohl
sonst im ganzen Pflanzenreich nicht wieder. Das Wesentliche des
Vorganges ist, dass eine ruhende weibliche durch eine bewegliche
männliche Fortpflanzungszelle durch Verschmelzung befruchtet wird. Die
Verschiedenheiten werden durch die Art der Entwickelung der männlichen
Zelle bewirkt. Wir wollen dabei einen der Fälle ins Auge fassen und
auf die beobachteten Abweichungen hinweisen. Die männlichen Zellen
entwickeln sich in Antheridien, welche entweder mit den weiblichen
Zellen in demselben Faden entstehen (monoecische Arten) oder in
besonderen Fäden (dioecische Arten), welche wiederum entweder normal
sind und neben den Antheridien auch gewöhnliche vegetative Zellen
enthalten, oder nur aus wenigen Zellen von abweichender Gestalt
bestehen und dann als sogenannte Zwergmännchen sich auf den weiblichen
Zellen oder in der Nähe derselben finden (Nr. 1 d). Diese[S. 57]
Zwergmännchen entwickeln sich aus Schwärmsporen, welche ähnlich wie
die ungeschlechtlichen gebildet werden und diesen auch ganz ähnlich,
nur etwas kleiner sind. Diese Schwärmsporen schwärmen eine Zeit lang
umher, setzen sich dann in unmittelbarer Nähe der weiblichen Organe
fest und werden zu den Zwergmännchen, welche mit oder ohne eine
etwas längere Fusszelle eine geringe Anzahl flach scheibenförmiger
Zellen, Antheridien, entwickeln, in denen je zwei männliche
Fortpflanzungszellen „Spermatozoiden“ entstehen (Nr. 4 a,
b).
Die weiblichen Geschlechtsorgane, Oogonien, sind runde, angeschwollene
Zellen, welche zwischen den vegetativen liegen (1 c). Zur Zeit
ihrer Empfängnisfähigkeit bildet sich ebenso wie bei den vegetativen
Zellen ein kreisförmiger Riss und die Zelle klappt auf; es tritt jedoch
noch eine sehr dünne gewölbte mit einem kleinen Loch versehene Membran
(1 e) hervor, welche die Eizelle abschliesst. Zu gleicher Zeit
öffnet sich das Antheridium und entlässt die Spermatozoiden, von denen
eins durch das Loch der erwähnten zarten Membran schlüpft und unter
bohrender Bewegung sich mit dem Ei vereinigt. Hierauf umgiebt sich die
Eizelle mit einer starken oft stacheligen Membran (5) und macht eine
Ruheperiode durch. Wenn es zur Keimung kommt, reisst die Sporenmembran
durch und der in eine Schleimhülle gebettete Inhalt tritt hervor, um
sich in vier Schwärmzellen zu teilen (6), welche ausschwärmen, sich
nach einiger Zeit festsetzen und neue Oedogonienfäden entwickeln. So
kompliziert ist die Entwickelungsgeschichte eines so einfachen Wesens!
Wir konnten bei dem geringen zu Gebote stehenden Raum nur einzelne
interessante Vorgänge eingehender betrachten, aber wenn auch wesentlich
anders, finden sich nicht minder verwickelte Prozesse bei anderen
Gattungen, wie Sphaeroplea und Coleochaete, die hier
übergangen werden müssen.
[S. 58]
4. Die Rhodophyceen.
Nur ein überaus kümmerliches Bild von der farbenprächtigen Algengruppe
wird uns durch die wenigen Vertreter des süssen Wassers zu teil,
und diese sehen oft noch nicht einmal rot aus sondern braun,
schwärzlich, violett oder grün. Die in schnellfliessenden Gebirgsbächen
auftretende Gattung Lemanea zeigt Formen, die man äusserlich
feinen im Wasser geschwärzten Würzelchen vergleichen möchte,
Batrachospermum, schleimige reich verzweigte Fadenmassen von
sehr zierlichem Aussehen unter dem Mikroskop und meist blaugrüner oder
grauvioletter Farbe, Chantransia, deren Arten vielleicht nur
Jugendzustände von Batrachospermum sind, ist der vorigen Gattung
ähnlich, nur weit einfacher gebaut und nur Hildebrandtia zeigt
leuchtend purpurrote Überzüge auf Steinen. Das sind, von einigen sehr
seltenen abgesehen, unsere Süsswassergattungen.
Bei den Rhodophyceen finden sich zweierlei Arten der Fortpflanzung,
eine ungeschlechtliche und eine geschlechtliche. Die erstere
findet in der Weise statt, dass sich gewöhnlich vier in Tetraden
zusammen liegende Zellen zu Sporen (Tetrasporen) umbilden. Bei
der geschlechtlichen Fortpflanzung bilden sich weibliche Zellen
(Carpogonien) und männliche rundliche Fortpflanzungszellen, welche
beide unbeweglich sind. Das Carpogon entwickelt noch einen Halsteil
(Trychogyne), an welchen die männlichen Zellen vom Wasser herangespült
werden, festhaften und die Befruchtung vollziehen. Hierauf sprossen aus
dem Carpogon Zellfäden, welche an ihrem Ende ebenfalls unbewegliche
Fortpflanzungszellen abgliedern, diese keimen dann und entwickeln
neue Pflänzchen. Bei unseren Süsswasserarten sind diese Vorgänge aber
noch nicht ganz erforscht, und da dieselben gegenüber den anderen
Algengruppen an Zahl und Formenreichtum weit zurückbleiben, so mag
dieser kurze Hinweis genügen.
[S. 59]
Den Algen nahestehend, aber höher entwickelt und in manchen Punkten
den höheren Kryptogamen ähnlich, tritt uns noch eine seltsame Gruppe
echter Wassergewächse in den Armleuchtern oder Charen
entgegen. Die Armleuchter unterscheiden sich von den Algen durch die
grosse Regelmässigkeit ihres Aufbaues sowie durch die hochentwickelte
Fortpflanzung. Sie bilden in der Regel kleine Büsche von wenigen
Stengeln, welche oft reich verzweigt sind und in gewissen Abständen
quirlständige, wieder mit einfacheren Blättchen versehene „Blätter“
tragen. An den Blättern (bei einer Gattung auch in den Blattachseln)
stehen die Fortpflanzungsorgane: die weiblichen Sporenknöspchen, die
männlichen Antheridien genannt. Die Sporenknöspchen bestehen aus einer
Eizelle und fünf schlauchförmigen Hüllzellen, welche die erstere
umgeben und an der Spitze noch bei den eigentlichen Charen je eine,
bei den Nitellen je zwei zu einem fünf- oder zehnzelligen Krönchen
zusammentretende Zellen abgliedern. Ist die Eizelle empfängnisfähig
geworden, so fällt das Krönchen ab oder die Zellen desselben weichen
auseinander, so dass die männlichen Fortpflanzungszellen freien Zutritt
zu den weiblichen erhalten. Die männlichen Fortpflanzungszellen
(Spermatozoiden) sind spiralig gewundene Fäden, welche an ihrem
vorderen Ende zwei lange Geisseln tragen und denen höherer
Kryptogamen, besonders denen der Torfmoose ausserordentlich ähnlich
sind. Sie entstehen in sehr kompliziert gebauten Antheridien von
roter Farbe und kugeliger Gestalt. Bei der Befruchtung dringen die
aus dem klappenförmig aufspringenden Antheridium ausschwärmenden
Spermatozoiden in das Sporenknöspchen durch die auf oben angegebene
Weise entstandene Öffnung ein und verschmelzen wahrscheinlich mit
der Eizelle; das letztere hat man noch nicht beobachtet. Hierauf
umgiebt sich die Eizelle mit einer holzigen Schale, auch häufig
noch mit einem Kalkmantel und macht eine oft jahrelange Ruheperiode
durch, ehe sie keimt. Aus der Spore entwickelt sich zunächst ein sehr
einfacher Vorkeim, der nach unten feine, einfache, weisse Fäden aus
langgestreckten Zellen, Wurzeln, entsendet, nach oben einen grünen
Stengel entwickelt, welcher nur einen sehr einfachen Blattquirl trägt.
An Stelle des einen Blattes in diesem[S. 60] Quirl tritt der Stengel der
eigentlichen Charenpflanze auf, welche zwar sehr einfach, aber sehr
regelmässig gebaut ist. Zwischen je zwei Blattquirlen tritt eine
einzige oft sehr lange und dicke Zelle, die Internodialzelle, auf,
dann folgt eine flache und darauf wieder eine Internodialzelle. Diese
erleiden ausser weiterem Wachstum keine Veränderung mehr, während
die erwähnte flache Zelle sofort wieder eine Anzahl Zellen an ihrer
Peripherie bildet, aus denen die Blätter entstehen. An den Blättern
wiederholt sich derselbe Wechsel zwischen Internodial- und Knotenzellen
und die letzteren gliedern wieder peripherische Zellen ab, aus denen
dann die stets einzelligen Blättchen entstehen. Aus den Knoten der
Stengel und Blätter entsteht auch ihre Berindung, indem schon in sehr
jugendlichem Alter der Sprosse vom Knoten aus aufwärts und abwärts
eine Anzahl röhrenförmiger Zellen wachsen, welche ungefähr in der
Mitte des Internodiums zusammentreffen und dasselbe rings umhüllen.
Aus gewissen kurzbleibenden Zellen der Berindung wachsen auch manchmal
Stacheln hervor, welche die Pflanze oft ganz behaart erscheinen lassen
(z. B. Chara crinita). Bei der Gattung Nitella fehlt die
Berindung, auch die Blätter sind anders gebaut. Eine ungeschlechtliche
Fortpflanzung fehlt den Charen, nur eine vegetative Vermehrung durch
Wurzelknöllchen oder verschiedenartige aus den Knoten hervortretende
Sprosse ist vorhanden.
Ein besonderes Interesse bieten die Armleuchter durch die Strömung
des Protoplasmas in den Zellen, welche man hier deutlicher als
irgend sonst im Pflanzenreich beobachten kann, was zumteil mit der
Grösse der Zellen zusammenhängt. Diese gehören nämlich zu den grössten
überhaupt bekannten Zellen, denn sie werden bei manchen Arten (z. B.
die ersten Internodialzellen von Tolypella prolifera) bis 20
cm lang und bis 2 mm dick. Im Innern der bei der Grösse
der Zellen dünnen Zellwand findet sich zunächst eine eng anliegende
ruhende Protoplasmaschicht, welche die reihenweise angeordneten
Chlorophyllkörner trägt. Auf diese folgt eine Protoplasmaschicht,
welche sich in einem geschlossenen, in der Längsrichtung der Zellen
liegenden Ringe bewegt und zwar so, dass der Strom auf der einen Seite
der Zelle aufsteigt, am oberen[S. 61] Ende derselben umbiegt und auf der
anderen Seite wieder absteigt, um am unteren Ende wieder umzubiegen.
Die Plasmaschicht führt eine Anzahl heller Zellkerne und kleinere
Körnchen mit sich und bewegt sich bald langsamer, bald schneller, je
nach Temperatur und Beleuchtung und wohl auch nach dem Alter der
Zellen. Besonders schön ist diese Plasmaströmung (Rotation) bei den
nicht berindeten Nitellen zu sehen, weil die berindeten Charen ausser
den Rindenzellen auch noch sehr häufig eine dicke Kalkinkrustation
zeigen, welche den Zellinhalt verdeckt. Die Plasmaströmung hat denn
auch die Gattung Nitella zu einem wertvollen Objekt für die
Pflanzenphysiologie gemacht, ohne dass man jedoch trotz vieler
Untersuchungen über diesen Punkt vollständige Aufklärung erhalten hätte.
Die Characeen sind eine zwar artenarme aber formveränderliche und
schwierige Pflanzengruppe, welche ziemlich isoliert im Pflanzenreiche
dasteht, von den Systematikern bald bei den Algen, bald bei den Moosen
untergebracht wird. Vielleicht ist es am besten, sie als eigene Gruppe
zwischen beide zu stellen. Sie finden sich besonders im süssen Wasser,
wo sie oft vollständig den Grund der Seen überziehen. Einzelne Arten,
so Ch. foetida, entwickeln einen widerwärtigen Geruch, den man
beim Austrocknen von Charen bewohnter Tümpel oft auf weite Entfernungen
wahrnimmt.
Von den anderen Wasserkryptogamen mögen zunächst noch die Torfmoose
erwähnt werden, welche wenigstens gewissen Wasseransammlungen ihren
Charakter verleihen, besonders den Tümpeln der Hochmoore. Hier
bilden sie nicht nur ein dichtes Polster um die Wasserfläche herum,
sondern fluten auch noch in dem Wasser selbst. Sie sind von den
anderen Moosen leicht durch die eigentümliche Bildung der Blätter zu
unterscheiden, denn diese bestehen nicht wie bei den letzteren aus
gleichartigen Zellen, sondern aus grossen luftführenden weissen, schief
rhombischen, durch spiralige Leisten verdickten und mit Öffnungen
nach aussen versehenen Zellen und aus dazwischen liegenden schmalen
chlorophyllhaltigen. Die Fortpflanzung der Torfmoose, wie die der
Moose[S. 62] überhaupt, ist durch einen sogenannten Generationswechsel
charakterisiert. An der Moospflanze entstehen die Geschlechtsorgane
in „Blüten“, bald männliche, Antheridien, und weibliche, Archegonien,
zusammen, bald getrennt. Im Grunde des Archegoniums findet sich eine
Eizelle, welche durch die den beschriebenen Spermatozoiden der Charen
ähnlich gestalteten, aus den Antheridien entstandenen Samenkörperchen
befruchtet werden. Die befruchtete Eizelle entwickelt sich zu der
zweiten, ungeschlechtlichen Generation, der Mooskapsel. Diese bringt
in ihrem Innern eine Anzahl unbefruchtete Sporen hervor, aus denen
sich bei der Keimung Vorkeime entwickeln und an diesen wieder die
geschlechtliche Generation, die Moospflanzen.
Bei den höher stehenden Farnpflanzen oder Pteridophyten, von denen
eine Gruppe, die Wasserfarne (Hydropteriden), recht
eigentliche, obwohl seltene Wasserbewohner sind, findet sich ebenfalls
ein deutlich ausgesprochener Generationswechsel; nur ist hier die
geschlechtliche Generation ein unscheinbares kleines lebermoosartiges
Gebilde, an welchem die ungeschlechtliche hochentwickelte Pflanze, das
eigentliche Farnkraut, entsteht, also ganz entgegengesetzt den Moosen.
Von den bei uns heimischen Wasserfarnen ist am häufigsten Salvinia
natans, ein zierliches auf dem Wasser schwimmendes Gewächs, welches
entfernt einem Akazienblatt ähnlich ist. Die Salvinia trägt zwei
Arten von Blättern, zwei Reihen laubartige, nach oben gekehrte und eine
dritte Reihe sehr fein zerschlitzter in das Wasser hineinragender,
welche man sich schwer entschliessen kann, nicht für Wurzelbüschel
anzusehen. Viel seltener und in Deutschland nur in Schlesien und am
Rhein heimisch ist die dort Wasserklee oder Vierklee genannte
Marsilia quadrifoliata, welche in der That einem Stock nur
Vierblätter tragenden Klees ausserordentlich ähnlich ist.
Auf dem Grunde unserer Seen lebt noch eine dritte Gattung in wenigen
Arten, Isoetes, welcher ebenso wie die zerstreut durch
Deutschland in Wiesengräben vorkommende Pillularia an kleine
Binsenbüsche erinnert.
[S. 63]
Nicht eigentlich zu den Wassergewächsen gehören die
Schachtelhalme, aber eine Anzahl Arten lieben die Ufer von
Teichen und Sümpfen und verleihen ihnen so einen bestimmten Charakter.
Damit sind die Wasserkryptogamen, abgesehen von den Pilzen, erschöpft
und wenn die höher organisierten, die Moose und Farne, vermöge ihrer
Grösse weit mehr in die Augen fallen und die Flora eines Wassers
mehr zu bestimmen scheinen, so verschwindet ihre Zahl doch gegen den
Reichtum an Formen, und an Schönheit gegenüber den mikroskopischen
Algen.
[S. 64]
Litteratur.
Für Süsswasseralgen sind folgende allgemeine Werke von besonderer
Wichtigkeit:
1. Rabenhorst, Flora Europaea Algarum aquae dulcis et
submarinae.
2. Kützing, Species Algarum.
3. „ Tabulae phycologicae.
4. De Toni, Sylloge Algarum (bis jetzt Chlorophyceen
erschienen).
5. Hansgirg, Prodromus.
6. Kirchner, Algenflora von Schlesien.
7. Falkenberg, Algen in Schenk, Handbuch der Botanik.
Für Diatomeen sind eine grosse Anzahl, aber meist sehr teuere Werke
erschienen, dem Anfänger mögen Rabenhorsts Süsswasserdiatomeen (mit 10
Tafeln) genügen. Um die mikroskopische Technik zu erlangen, studiert
man am besten Strassburgers Botanisches Praktikum (1884).
[S. 65]
Zur Biologie
der phanerogamischen Süsswasserflora.
Von Prof. Dr. Friedrich Ludwig.
[S. 67]
Die Beziehungen der Pflanzenwelt zu ihrer belebten und unbelebten
Umgebung sind so mannigfaltige, die Anpassungen so ungeahnt bis ins
einzelne gehend, dass trotz der grossen Zahl der Forscher, die
sich in der Gegenwart auch hier, wie auf allen Gebieten geistiger
Arbeit, zusammengefunden haben, um diese Beziehungen aufzudecken,
fortgesetzt neue Entdeckungen zu verzeichnen, neue Fragen zu
beantworten, neue Probleme zu lösen sind. Wir erinnern nur an die
merkwürdigen gegenseitigen Anpassungen der beiden grossen Familien
der Feigen und der Feigenwespen[1], an die Ameisenpflanzen[2], die
Milbenpflanzen mit ihren Milbenhäuschen (Acarodomatien)[3], die
mannigfachen Eigenschaften, welche die Pflanzen im Zusammenleben mit
den Schnecken und anderen omnivoren niederen Tieren und als Schutz
gegen dieselben erhalten haben[4], an die höchst eigentümliche Art
der Ernährung unserer Waldbäume und vieler anderen Pflanzen durch die
Vermittelung der Pilze (Mycorrhizen der Cupuliferen, Ericaceen etc.),
die Stickstoffernährung der Hülsenfrüchtler durch die Wurzelknöllchen
bildenden Bacillen (Bacillus radicicola Beyerinck, Franks
Rhizobium), an die fleischverdauenden Pflanzen, an die Beziehungen
zwischen gewissen Wirts- und Schmarotzerfamilien und -Arten, wie sie
z. B. bei der Gruppe der Rostpilze so mannigfach zu Tage treten (die
Gattung Phragmidium ist auf die Rosaceen, die Ravenelia
auf Leguminosen beschränkt, Gymnosporangium bildet die erste
Generation nur auf Pomaceen, die zweite und dritte auf Coniferen aus
etc.), — der grossen Kapitel von den Anpassungen der Blumen und Blüten
an Insekten,[S. 68] Vögel, Wind etc., von den Verbreitungsmitteln der Früchte
und Samen, den Schutzmitteln gegen allerlei schädigende Einflüsse, die
ganz besondere Bearbeitung gefunden haben, nicht zu gedenken. Immerhin
hat man eine stattliche Anzahl solcher Beziehungen der Pflanzen
zu der übrigen organischen und unorganischen Welt aufgedeckt, die
Fäden gefunden, welche die verschiedensten Naturkörper einheitlich
verknüpfen, zu einem Naturganzen zusammenschlingen — soweit es sich
um Landtiere und Landpflanzen handelt. Nicht so weit ist man in die
Lebensgeheimnisse der Wasserwelt eingedrungen.
Das Pflanzenleben im Wasser ist doch an ganz andere Bedingungen
geknüpft. Luft und Erde mit ihren Organismen und unorganischen
Bestandteilen sind es allein, mit und in welchen die gewöhnliche
Landpflanze zu leben hat. Eine dritte Hülle unseres Planeten, die
Wasserhülle, mit all ihren abweichenden physikalischen und chemischen
Eigenschaften, mit einer ganz anderen Lebewelt, die teils der Pflanze
von Nutzen ist, teils von ihr abgewehrt werden muss, ist das Element
der Wasserpflanzen. Dazu kommt, dass auch das Luftmeer über dem Wasser
und der Erdboden unter dem Wasser andere physikalische Eigenschaften
und ein anderes Tierleben besitzt als Luft und Boden im Trocknen. Das
grössere spezifische Gewicht des Wassers macht gewisse Einrichtungen
der Landpflanzen, welche der Festigung dienen, überflüssig, indem das
Wasser einen guten Teil der Last der Pflanzenorgane bei Wasserpflanzen
trägt. Das Verhalten des Wassers zur Wärme ist der Pflanzenwelt
besonders günstig. Die Temperaturveränderungen erfolgen allmählich,
nicht plötzlich wie die der Luft und des trocknen Bodens, und vermöge
seiner Eigenschaft, bei + 4° C. den kleinsten Raum einzunehmen oder
die grösste Dichtigkeit zu haben, gefriert das Wasser nur an der
Oberfläche, in der Tiefe eine Temperatur von über 0° bewahrend, welche
auch dem Boden der Gewässer zu gute kommt. Im Wasser, am und im Boden
der Gewässer, können daher Gewächse überwintern, welche am Lande im
Winterfrost zugrundegehen würden. Daher sind auch die eigentlichen
Wasserpflanzen ausdauernd, mit Ausnahme der uferbewohnenden[S. 69]Tännelarten (Elatine) und des Nixenkrautes
(Najas flexilis und minor); und auch von solchen Arten,
die in einer besondern einjährigen Landform vorkommen, wie dem
Wasserhahnenfuss, dem schwimmenden Igelkolben etc., ist
die Wasserform ausdauernd, auch im Winter flutend.
Die Löslichkeit für die Bestandteile der atmosphärischen Luft ist eine
ungleiche und das Wasser nimmt von ihnen nur 2–3% Sauerstoff auf,
während das Luftmeer davon etwa 21% enthält; dagegen hat das Wasser
einen hohen Kohlensäuregehalt, auch die Bodengase haben eine wesentlich
andere Zusammensetzung auf dem Wassergrund als am Lande. Schliesslich
sind die Beleuchtungsverhältnisse im Wasser, das einen guten Teil der
Sonnenstrahlen absorbiert, andere als in der Luft.
Diese und ähnliche Verhältnisse müssen in dem Bau und der Entwickelung
der Wasserpflanzen zum Ausdruck gekommen sein und thatsächlich werden
uns die wesentlichsten Unterschiede zwischen Land- und Wasserpflanzen
von diesem Gesichtspunkte aus verständlich.
Von vornherein ist im Wasser ein fünffaches Pflanzenleben denkbar: 1.
Das Leben im Boden der Gewässer (Schlammpflanzen). 2. Das Leben in
Boden und Wasser (submerse Wasserpflanzen mit Wurzeln). 3. Das Leben im
Wasser (nichtwurzelnde submerse Wasserpflanzen). 4. Das Leben in Wasser
und Luft (nichtwurzelnde Schwimmpflanzen und nichtwurzelnde submerse
Pflanzen, welche ihren Blütenstand über Wasser zur Entwickelung
bringen) und 5. Das Leben in allen drei Medien zugleich (wurzelnde
Pflanzen, welche ihren Blütenstand über Wasser entwickeln und die
eigentlichen Sumpf- und Uferpflanzen). Von Schlammpflanzen,
welche in dem an toter organischer Substanz reichen, lockeren, auch im
Winter nicht gefrierenden Boden unserer Gewässer leben, sind bisher
wohl nur die niedersten Pilzformen bekannt geworden, welche aber
zumteil bedeutende Veränderungen an der Erdoberfläche hervorgebracht
haben und die Zusammensetzung der Nährstoffe für die phanerogamischen
Süsswassergewächse nicht unwesentlich beeinflussen. Erinnert sei
an die Spaltpilze der Cellulosegärung (Bacillus Amylobacter
etc.), die Bildner des Sumpfgases, durch dessen Wirkung[S. 70] der Gips
unter Entwickelung von Schwefelwasserstoff in kohlensauren Kalk
umgewandelt wird, ferner an die „Schwefelbakterien“ (Beggiatoa,
Clathrocystis rosea-persicina, Ophidomonas etc.), welche
den Schwefelwasserstoff der Cellulosegärung zu Schwefel, den sie in
Form kleiner amorpher Kügelchen in der Zelle aufspeichern, und letztern
sodann zu Schwefelsäure oxydieren. Durch die letztere verwandeln sie
die Karbonate wieder in Sulfate und geben freie Kohlensäure an das
Wasser ab. Auch die von Winogradski als „Eisenbakterien“ bezeichneten
Bildner des Sumpf- und Wiesenerzes u. a. gehören hierher[5].
Die phanerogamischen Wassergewächse gehören den übrigen Abteilungen an,
deren Lebensverhältnisse, soweit sie sicher bekannt sind, im folgenden
an einigen ausführlicheren Beispielen etwas eingehender behandelt
werden sollen.
Die submersen
(untergetauchten) Wasserpflanzen.
Die Ernährung der Landpflanzen geht bekanntlich im grossen und ganzen
in der Weise vor sich, dass das Wasser des Bodens mit den darin
gelösten Nährsalzen von der Wurzel nach den chlorophyllhaltigen
Geweben des Blattes emporgehoben und mit der aus der Atmosphäre
durch die Blätter aufgenommenen Kohlensäure zu den ersten
organischen Verbindungen (Stärke etc.) verarbeitet, „assimiliert“
wird. Die Bewegung des Wassers wird dabei wesentlich durch die an
der Blattfläche, besonders auch durch die in den Spaltöffnungen
der untern Blattseite stattfindende Verdunstung und hierdurch
ausgeübte Saugkraft auf die darunter gelegenen Elemente bewirkt.
Dieser „Transspirationsstrom“ geht durch die Gefässe von der Wurzel
durch die Hauptachse zu den seitlichen Auszweigungen und den
Blättern an ihnen, während die Kohlensäure durch die Spaltöffnungen
in das „Schwammparenchym“ des Blattes gelangt. Das Blatt zeigt
in der Regel unter chlorophyllfreier Epidermis oben das zu den
Beleuchtungsverhältnissen in der Luft in Beziehung stehende sogenannte
Pallisadengewebe, unten,[S. 71] innerhalb der von den Spaltöffnungen
durchsetzten Epidermis, das Schwammgewebe mit den Atemhöhlen.
Bei den submersen Wasserpflanzen kommt dieser Transspirationsstrom,
mit ihm der strenge Gegensatz von Haupt- und Nebenachse und das
Hervortreten der Gefässe in Wegfall.
Die Spaltöffnungen fehlen den untergetauchten Blättern.Die
Aufnahme der im Wasser gelösten Kohlensäure sowohl, wie der
anorganischen Nährsalze, geschieht direkt durch die dünnwandige
chlorophyllhaltige Epidermis[6]. Die Wurzeln dienen in der
Hauptsache da, wo sie vorhanden sind, als Haftorgane, haben
ihre Rolle der Nahrungsaufnahme verloren (besitzen keine Wurzelhaare
mehr etc.), in vielen Fällen fehlen sie jedoch gänzlich. Da die ganze
grüne Blattmasse an ihrer Oberfläche durch Diffusion die Aufnahme
der flüssigen Nahrung und den Gasaustausch besorgt, muss bei ihr
das Prinzip der äussern Oberflächenvergrösserung durch
tiefgehende Zerteilung und Verzweigung, das den Grundbau der ganzen
höhern Pflanze (Verzweigung von Wurzel und Stamm) im Gegensatz zu dem
der höheren Tiere beherrscht, besonders zur Geltung kommen. Spaltet
man einen Würfel oder einen andern Körper fortgesetzt, so kann man
bekanntlich, ohne das Gesamtvolumen zu ändern, die Oberfläche bis zu
jeder beliebigen Flächengrösse hinauf wachsen lassen. Derartig ist auch
die Vergrösserung der mit der spärlichen Sauerstoffmenge im Wasser
haushaltenden Atmungsorgane (Kiemen) unserer Wassertiere. Thatsächlich
ist nun den submersen Wasserpflanzen mit wenigen Ausnahmen und nicht
allein ihnen, sondern auch den untergetauchten Wasserblättern der
Pflanzen der vierten und fünften Gruppe eine weitgehende Zerschlitzung
des Blattes bis zu feinen haar- oder borstenförmigen Abschnitten
gemein. So z. B. den Arten des Hornblattes (Ceratophyllum),
des Tausendblattes (Myriophyllum), des Wasserschlauches
(Utricularia), der Sumpffeder (Hottonia), der
Aldrovandie. So sind wenigstens die untergetauchten Blätter haarförmig
fein geteilt bei den Wasserhahnenfussarten (Batrachium), bei
der Rebendolde (Oenanthe[S. 72] Phellandrium etc.). In anderen Fällen
sind die dünnen Blätter zwar einfach, aber schmallinealisch, so z. B.
bei der Wasserpest (Elodea), der Vallisnerie (Vallisneria
spiralis), den Wassersternen (Callitriche) u. a. Die feine
Zerteilung etc. sichert zugleich das Blatt vor einem Zerreissen durch
Wasserströmungen und Wassertiere.
Betrachten wir des weitern die untergetauchte Wasserpflanze in ihren
Beziehungen zu ihrer Umgebung an einigen besonderen Beispielen.
Fig. 6.
Befruchtung des Gemeinen Hornblattes (Ceratophyllum demersum: a
Männlicher Blütenstand — b Hüllkelch desselben nach der Entleerung —
c Ein Staubgefäss (vergrössert) — l Auftrieb — d Oberer
Teil desselben (Auftrieb) — e Pollenkörner — f Weibliche Blüte
(schwach vergrössert) — g Griffel — h i Narbenfläche).
Das Gemeine Hornblatt (Ceratophyllum demersum) ist
eine in Teichen, langsam fliessenden Gewässern und am Rande von
Seen weit verbreitete, wurzellose, frei im Wasser flutende Art mit
quirlig stehenden, wiederholt gabelspaltigen, feingeteilten Blättern
an verzweigtem runden Stengel, der an der Spitze fortwächst, vom
untern Ende aus absterbend. Es ist die einzige Pflanzengattung
des Süsswassers, welche streng hydrophil ist, d. h. zur
Übertragung des Pollens vom Staubgefäss zur Narbe des Stempels des
Wassers bedarf, während von Meerespflanzen eine Anzahl hydrophiler
Pflanzen bereits seit längerer Zeit bekannt ist. Als ich vor zehn
Jahren auf Veranlassung meines Freundes Hermann Müller
die Bestäubungsverhältnisse unserer Süsswasserpflanzen und ihre
Anpassungen an das Wasser und gewisse wasserbewohnende Insekten einem
eingehenderen Studium unterzog[7], da war über diese — wie H. Müller
sagt — „bis dahin auffallend vernachlässigte Gruppe“ noch so gut wie
gar nichts bekannt. Ceratophyllum demersum (Fig. 6) zeigte
mir einen in wunderbarer Weise der Wasserbefruchtung angepassten
Mechanismus. Männliche und weibliche Blüten stehen, kaum gestielt,
getrennt in verschiedenen Blattwirteln ordnungslos durcheinander
(nur scheinen zu unterst die weiblichen Blüten zu überwiegen). Die
männlichen staubgefäss- und pollenreichen Blüten sind in beträchtlich
grösserer Zahl vorhanden, als die weiblichen. Letztere enthalten in
einem anliegenden vielzipfeligen Kelche einen ovalen Fruchtknoten
mit einem den Kelch um das vier- bis fünffache überragenden hakig
nach unten gekrümmten Griffel, der sich nach der Spitze zu allmählich
verschmälert. Der letztere ist nirgends warzig, doch sondert seine[S. 73]
ganze Unterseite einen Klebstoff ab und fungiert als Narbe. — Der
männliche Blütenstand besteht aus 12–16 sehr kurz gestielten Antheren,
die von einer vielteiligen Hülle umgeben sind. Die einzelnen Teile der
letztern sind linealisch, gestutzt, meist zweidornig. Die Staubgefässe
bestehen im unteren, dem kurzen Stiele aufsitzenden Teile aus zwei
seitlich sich längs öffnenden Pollenkammern und im oberen Drittel aus
lockerem lufthaltigen Gewebe, an der Spitze mit zwei nach der Mitte
zu gekrümmten Dörnchen, zwischen denen meist noch eine schwärzliche,
mehr oder weniger[S. 74] gerade höckerige Drüse sich befindet. Diese
Spitzenanhängsel des pollenerzeugenden Apparates (ebenso wie die sie
einschliessenden Dornspitzen) kommen in fast gleicher Weise an den
Enden der Hüllblätter und Laubblätter vor (vgl. Fig. 6 k). Sie
sind nach neueren Untersuchungen[8]tanninhaltig und bilden
nach Stahl[4] ein wirksames Schutzmittel gegen Wasserschnecken,
wohl auch gegen andere Wassertiere. Zur Befruchtung stehen sie in
keiner Beziehung. Anders verhält es sich mit dem aus lockerem
lufthaltigen Gewebe bestehenden Antherenfortsatz, den ich als
„Auftrieb“ bezeichnet habe. Derselbe macht das ganze
Staubgefäss spezifisch leichter als Wasser und treibt dasselbe, wenn
es aus der Blüte losgelöst wird, nach der Oberfläche des Wassers.
Die rundlichen oder länglichen nur von einer zarten
Haut umgebenen Pollenkörner haben genau das spezifische
Gewicht des Wassers, so dass sie in jeder beliebigen Tiefe
suspendiert bleiben. Dieses verschiedene spezifische Gewicht der
Pollenkörner und des gesamten pollenerzeugenden Apparates, zusammen
mit dem Verhalten der starrblättrigen Hülle, bestimmt den eigentlichen
Pollentransport. Die Hüllblätter haben nämlich das Bestreben,
sich nach innen zu biegen — an entleerten Blütenständen sind sie
aufrecht —, so dass die Staubgefässe zur Zeit ihrer völligen
Ausbildung keinen genügenden Platz mehr haben. Zur Zeit der Dehiscenz
werden die letzteren daher aus der Hülle herausgepresst und
schwimmen unter der Wirkung des Auftriebes nach oben, bis sie
die Wasseroberfläche erreicht haben, oder, was häufiger geschieht,
zwischen den hakigen Blättern der oberen Stengelglieder zurückgehalten
werden. Während dieser Aufwärtsbewegung werden die Antheren
entleert, wobei die durch den Auftrieb bedingte Vertikalstellung
des Staubgefässes besonders günstig ist, und verbreiten sich —
weil vom spezifischen Gewicht des Wassers — über den ganzen vom
pollenerzeugenden Apparat bestrichenen Raum. Wird so schon das Wasser,
in welchem Ceratophyllum wächst, überall von dessen grossen
Pollenkörnern (40–50 µ breit und 50–75 µ lang)[S. 75] erfüllt, so kommt der
Verbreitung der letzteren resp. einer Fremdbefruchtung durch dieselben
nächst den Bewegungen des Wassers selbst und den passiven Bewegungen
des Ceratophyllum durch Wassertiere ein anderer Umstand zu statten
— die Eigenbewegung des Ceratophyllumstammes, die besonders
in ruhigem, stehendem Wasser (in welchem ich meine Beobachtungen
anstellte) nicht unterschätzt werden darf. Dieselbe wurde zuerst von
E. Rodier[9] nachgewiesen. Die jungen (blütentragenden) Internodien
haben eine vom Lichte unabhängige, komplizierte Bewegung,
welche einen besonderen Fall der Darwinschen Circumnutationen[10]
darstellt, sich aber von der Mehrzahl dieser durch die Weite und
die verwickelte Art der Bewegung unterscheidet. Im allgemeinen
biegen sich die Stämme früh von rechts nach links und am
Nachmittag in entgegengesetzter Richtung. Zuweilen werden in
sechs Stunden Winkel von 200°, in einem Fall wurde sogar in drei
Stunden ein Winkel von 220° zurückgelegt. Zudem führen die Zweige um
ihre Wachstumsachse Torsionsbewegungen aus. Die Biegung der
Stämme ist schliesslich eine ganz eigentümliche, sie beginnt an
der Spitze und pflanzt sich von da in abnehmender Stärke
nach unten fort, während die Rückwärtsbewegung unten
beginnt und oben endigt, so dass die letzten Internodien kurz
vor ihrer Zurückbewegung zuweilen mit der Achse einen spitzen Winkel
bilden. Berücksichtigt man noch, dass der Pollen in ausserordentlich
reichlicher Menge erzeugt wird, so dürfte nach alledem eine
erfolgreiche Fremdbefruchtung der — wie ich glaube etwas vor den
Antheren entwickelten — weiblichen Blüten gesichert sein. — Herm.
Beyer[11] spricht von einer gleichzeitigen Bewegung der
weiblichen Blüten nach der Oberfläche, an welcher der vorher nicht
entleerte Teil des Pollens umherschwimmt, doch habe ich eine solche
Bewegung nicht wahrgenommen.
Die eigentümliche Befruchtungsweise der Ceratophyllumarten
(bei den übrigen deutschen Arten C. submersum und C.
platyacanthum wird sie in gleicher Weise angegeben) steht allem
Anschein nach in der Mitte zwischen der der Seegräser einer-[S. 76]
und der Hydrocharidaceen anderseits. Bei den Seegräsern
(Zostera, Cymadocea, Halodule etc.) besteht
der Pollen aus algenähnlichen bis zwei Linien langen
Schläuchen, welche, vom spezifischen Gewicht des Wassers,
direkt in dieses entleert werden und von den band- oder
hakenförmigen Narben aufgefangen die Befruchtung bewirken; auch
bei dem Nixenkraut (Najas), dessen Blütenverhältnisse
Jönssen und Magnus näher studiert haben, kommen ähnliche
Verhältnisse vor; die Pollenkörner sind aber hier nach Jönssen
durch ihren Gehalt an Stärkekörnern schwerer als das Wasser und sinken
nach unten, um von den Fangapparaten der tiefer sitzenden weiblichen
Blüten aufgefangen zu werden (man vergleiche aber die diese Auffassung
berichtigende Bemerkung von Magnus, nach dem erst die ausgekeimten
Pollenkörner verbreitet werden). Durch den Mangel der Exine,
der äusseren schützenden und oft mit Stachelfortsätzen besetzten
Haut der Pollenkörner und das hohe spezifische Gewicht
der letzteren stehen die Hornblattgewächse diesen submers
blühenden Seegräsern nahe, während sie durch die sich
loslösenden aufsteigenden Staubgefässe an die Hydrocharideen
erinnern. Von letzteren ist die Vallisnerie (Vallisneria
spiralis), die seit Jahrhunderten bekannte und schon vielfach
poetisch und prosaisch geschilderte Wasserblume, wie eine im
indischen Ozean heimische Verwandte (Enhalus acoroides), am
charakteristischsten. Die ganzen männlichen Blüten lösen sich los
und schwimmen während der Dehiscenz an der Wasserfläche umher,
während die weiblichen Blüten auf langem schraubenförmigen Stiele
die Oberfläche erreichen, um hier von den im Winde hin und her
getriebenen Staubblüten die Pollenkörner aufzunehmen. Nach geschehener
Befruchtung schraubt sich ihr Stiel wieder auf den Boden zurück, um
hier die Früchte zu reifen. Auch bei der Wasserpest (Elodea
canadensis), deren weibliche Form im Jahr 1836, nach Irland kam[12]
und sich von da aus mit anfangs erschreckender Schnelligkeit überall
in Europa ausbreitete, steigen in der nordamerikanischen Heimat die
männlichen Blüten zur Höhe, um an der Oberfläche ihre Pollenmassen[S. 77]
in die auf gestrecktem Fruchtknoten an die Oberfläche gelangten
Stempelblüten auszuschütten. Bei Hydrilla verticillata, einer
einheimischen Verwandten der Elodea, ist der Bestäubungsvorgang noch
nicht ermittelt, wohl aber ein ähnlicher. Bei der Verwandten unseres
Laichkrautes, der Meerstrands-Ruppie (Ruppia spiralis),
streckt sich der Stiel des weiblichen Blütenstandes ähnlich wie der
Blütenstiel der Vallisnerie schraubig zur Oberfläche, aber nur die aus
den Antheren austretenden Pollenkörner gelangen infolge ihres geringen
spezifischen Gewichtes zur Oberfläche, um dort herumschwimmend die
weibliche Narbe zu erreichen.
Die Verbreitung der hakigen Früchte der Hornblattarten geschieht
ohne Zweifel durch Wassertiere (Wasservögel, Wasserratten
etc.), wie auch die leicht zerbrechlichen Zweige, die durch Fische
und andere, auch über Land wandernde Tiere abgebrochen und an ihren
hakigen Blättern verschleppt werden, leicht und schnell weiterwachsen,
so dass in manchen Gegenden das Hornblatt eine der Wasserpest ähnliche
Verbreitung erreicht hat.
Zwei weitere submerse Wasserpflanzen, der Wasserschlauch
(Utricularia) und die ihm biologisch verwandte
Aldrovandia, verdienen nach den Hornblattgewächsen ohne Zweifel
zunächst unsere Beachtung.
Fig. 7.
Blatt der Venusfliegenfalle (Dionaea muscipula).
Die Blasenpflanze (Aldrovandia vesiculosa), deren
eigentliche Heimat die Gewässer des Südens von Südfrankreich und
Italien bis Indien und Australien sind, findet sich zerstreut in
Vorpommern, der Mark Brandenburg und Schlesien,
ferner in den Etschsümpfen bei Bozen in Tirol und im Bodensee. Der
submerse dünne Stengel wird kaum 30 cm lang und verästelt
sich wenig; er ist dicht mit wirtelständigen Blättern besetzt, deren
Stiel gegen das Ende breiter wird und in vier bis sechs steifen
Vorsprüngen mit kurzen Borsten endet. Die dazwischen befindliche
Spreite selbst erscheint, wenn sie geschlossen, blasenförmig.
Abgesehen von den blasigen Blättern erinnert die Aldrovandie im
Habitus an das Ceratophyllum; die starren Blattstacheln des letzteren
sind wohl auch der Funktion nach den spitzen Seitenfortsätzen am
Grunde der Aldrovandiablasen gleich. In einer anderen Hinsicht
steht aber[S. 78] die letztere der prächtigen Venusfliegenfalle
(Dionaea muscipula) sehr nahe, ja sie kann als eine kleine
im Wasser wachsende Dionaea bezeichnet werden. Es ist bekannt,
dass diese hübsch blühende Sumpfpflanze Carolinas (Fig. 7), welche
in unseren Gewächshäusern nicht selten zu finden ist, auf breitem
Blattstiel ein zweilappiges Blatt trägt, dessen Teile klappenartig
zusammenschlagen, wenn sich ein Tier auf die Blattfläche setzt.
Der Londoner Kaufmann Ellis, welcher die Dionaea benannte,
hatte bereits beobachtet, dass auf jeder der beiden Blatthälften
drei starre Haare befindlich sind, und gemeint, dass beim
Zusammenschlagen des Blattes, dessen Randstacheln dann den Fingern der
zum Gebet gefaltenen Hände ähnlich ineinandergreifen, Fliegen nicht nur
gefangen, sondern durch die sechs messergleichen Spitzen durchbohrt
würden, wie einst die Verbrecher von der „eisernen Jungfrau“ in der
Folterkammer zu Nürnberg. Dies war ein Irrtum. Später fand man, dass
die sechs Spitzen die Tentakeln des Blattes sind, deren Berührung ein
ausserordentlich schnelles Schliessen des ganzen Blattes zur Folge
hat, die aber selbst dabei den Klingen eines Taschenmessers ähnlich
sich zusammenlegen. Das Blatt der Dionaea selbst kann man zwicken,
stechen, schütteln, mit Wasser übergiessen, ohne dass es sich
bewegt, stösst man aber einen der sechs Tentakeln mit einem Strohhalm
leise an, so schliesst sich das Blatt im Nu[13]. Wie bei uns
Fliegen gefangen werden, so werden in der Heimat an dem natürlichen[S. 79]
Standort der Dionaea auch zahlreiche grosse und kleine Käfer, Spinnen,
Skolopender gefangen und — verzehrt; denn die Wissenschaft hat
nunmehr unumstösslich festgestellt, dass die Dionaea und ihre ganze
Sippe der Sonnentaugewächse, nebst einer Reihe anderer Familien
„Fleischfresser“ sind, Tiere fangen, durch Ausscheidung
einer unserem Magensaft ähnlichen Flüssigkeit verdauen und die
Verdauungsflüssigkeit als Nahrung aufsaugen.
Fig. 8.
Blasenpflanze (Aldrovandia vesiculosa). a Ganze Pflanze (nach
Schlechtendal-Schenk) — b Blattwirtel (vergr.) — c Offenes,
flachgedrücktes Blatt (b und c nach Cohn-Darwin).
Bei den Landsonnentaugewächsen bewegen sich nur die Blattdrüsen,
während sich bei der Dionaea das ausserordentlich reizbare Blatt
selbst bewegt. Unsere Aldrovandia (Fig. 8), die gleichfalls zu
den Sonnentaugewächsen zählt, verhält sich nun, wie Stein 1873
entdeckte und hernach Cohn und andere festgestellt haben[14],
ganz wie die Dionaea. Das zweilappige Blatt, dessen Mittelrippe an der
Spitze mit einer kurzen Borste endigt, öffnet sich bei warmem Wetter
etwas weiter, bei uns gewöhnlich aber nur so viel als die[S. 80] beiden
Klappen einer lebenden Muschel. Lange gegliederte Haare auf der
Mittelrippe und dem konkaven Teil der Blattlappen sind sensitiv wie
die sechs Dionaeahaare. Eine Reizung derselben bewirkt ein
augenblickliches Zusammenklappen des Blattes. Larven und andere
Wassertiere, besonders Krustentiere, werden nach den Untersuchungen
von Cohn in grosser Menge gefangen und verzehrt. Neben den
erwähnten in der Nähe der Mittelrippe und auf dieser selbst
befindlichen längeren gegliederten Tentakeln ist nämlich das
Blatt in dem der Mittelrippe zu gelegenen konkaven Teile
(der eigentlichen Oberseite des Blattes) mit gestielten farblosen
Drüsen, die denen des Dionaeablattes gleichen (aber einfacher
als diese sind), dicht besetzt (vgl. die Figur). Die Ähnlichkeit
mit den entsprechenden genauer untersuchten Verhältnissen bei der
Fliegenfalle, wie auch Versuche mit Fleischaufguss und verschiedene
Beobachtungen machen es mehr als wahrscheinlich, dass sie es sind,
welche eine wahre verdauende Flüssigkeit absondern und später die
verdaute Substanz aufsaugen. Der äussere und breitere Teil
des Lappens am Aldrovandiablatte ist flach und sehr dünn und
wird nur aus zwei Zellschichten gebildet (der konkave Teil). Seine
Oberfläche (Innenfläche) trägt keine Drüsen, aber an ihrer Stelle
kleine vierspaltige kreuzförmige Trichome. Zwei der schräg
auseinanderlaufenden Arme derselben sind gegen die Peripherie gerichtet
und zwei gegen die Mittelrippe. Ein schmaler Rand des flachen äusseren
Teiles jedes Lappens ist einwärts gebogen, so dass, wenn die Lappen
geschlossen sind, die äusseren Oberflächen der eingefalteten Teile in
Berührung kommen. Der Rand trägt eine Reihe sehr zarter Spitzen, welche
aber nicht wie die peripherischen Spitzen bei Dionaea Verlängerungen
der Blattscheibe, sondern nur Hautgebilde sind. Sie, wie besonders die
vierteiligen Trichome, dürften nach Darwin dazu dienen, faulende und
zerfallene tierische Substanz aufzusaugen, welche von der
Konkavität des Blattes abfliesst. Es würde, falls sich diese Ansicht
bestätigt, hier der merkwürdige Fall vorliegen, dass verschiedene Teile
eines und desselben Blattes sehr verschiedenen Zwecken dienen: der eine
zu[S. 81] wahrer Verdauung, ein anderer zur Aufsaugung der Fäulnisprodukte
der nicht verdauten tierischen Kadaver. Die Aussenseite des
Aldrovandiablattes ist mit sehr kleinen zweiarmigen Papillen besetzt,
die den achtstrahligen über Spreite und Blattstiel verbreiteten
Papillen von Dionaea und den Papillen auf dem Blatt des gemeinen
Sonnentaues analog sind. Für sie hat Darwin bei Drosera nachgewiesen,
dass sie nicht abzusondern, wohl aber zu absorbieren vermögen. Sie
könnten bei unserer Blasenpflanze die aus den Blasen ausgeflossenen
Zersetzungsprodukte, welche nicht verdaut wurden, noch aufnehmen.
Haben organische Körper den Reiz verursacht, so schliesst sich die der
klaffenden Austerschale vergleichbare Tierfalle der Aldrovandia nur auf
kürzere Zeit, 1–1½ Tage bei den Beobachtungen von Stein, während sie
bei organischen Körpern geschlossen bleiben bis dieselben verdaut sind.
Eine besondere Beachtung bei der Venusfliegenfalle, wie
bei der Blasenpflanze, verdient noch die fast momentane
Schliessbewegung der Blattklappen, die auch in der Tentakelbewegung
einiger exotischen Sonnentauarten beobachtet worden ist. Von zwei
australischen Sonnentauarten (Drosera pallida und Drosera
sulfurea) wie von einer indischen (D. lunata) und mehreren
afrikanischen Spezies (besonders von D. trinervis) wird
berichtet, dass sie ihre Blätter mit grosser Rapidität über Insekten
schliessen.
Bei unserer einheimischen Drosera anglica Huds. hat v.
Klinggraeff[15] sogar beobachtet, dass sich mehrere Blätter an
dem Fang grösserer Tiere zugleich beteiligen, eine Beobachtung, die
ich nach den Befunden an getrockneten Exemplaren einer australischen
Spezies (dieselbe wurde mir als D. binata übersandt, weicht
aber von der von Darwin behandelten Spezies durch nur einfach
gegabelte Blätter ab) auch für diese glaube behaupten zu können. H.
von Klinggraeff schildert einen derartigen Fang eines Schmetterlings
in folgender Weise: ... „Nach kurzer Zeit bogen sich mehrere Tentakeln
zusammen und klemmten den das Blatt berührenden Aussenrand des
Unterflügels ein, hielten ihn so fest, dass bei dem heftigen Flattern
derselbe einriss, der Schmetterling sich aber nicht[S. 82] befreien konnte.
Bei dem Flattern wurde ein anderes Blatt mit dem Oberflügel berührt
und, jedenfalls dadurch gereizt, bog sich dasselbe langsam gegen
den Schmetterling hin, bis es den Körper desselben erreichte und
umschlang. Während dessen hatte auch das erste fangende Blatt sich um
den Schmetterling geschlungen, so dass dessen Bewegungen zuletzt ganz
aufhören mussten. Meistens sah ich Schmetterlinge, die nur von zwei
Blättern umschlungen waren, an einigen Exemplaren jedoch nahmen drei,
auch vier Blätter an der Umschlingung teil“. Die zahlreichsten Opfer
waren immer Papilio Daplidice, wie man sich auch an den vielen
am Boden liegenden auf der Unterseite grünlich marmorierten Flügeln
überzeugen konnte, dann P. Rapae; auch von einem muskelkräftigen
Perlmutterfalter, Argynnis Latonia, wurde ein Exemplar gefangen.
Diese Bewegungen der ganzen Blätter wie auch der Tentakeln
nach der gereizten Stelle hin bei den Sonnentauarten sowohl, wie
die rapiden Bewegungen der Fliegenfalle und Aldrovandia, geschehen
durch eine eigentümliche Fortleitung des Reizes — die
nach der hübschen Entdeckung Darwins unter dem Mikroskop durch die
Zusammenballung („Aggregation“) des roten Saftes von Zelle zu
Zelle direkt beobachtet werden kann. Der Aggregation gehen
indes, wie Burdon-Sanderson, durch Darwin veranlasst (1874),
aufgefunden hat, elektrische Ströme voraus. Das ungereizte Blatt
ist oben positiv, unten negativ elektrisch, bei der Reizung wird jedoch
ein umgekehrter Strom erzeugt. Bezüglich der sonst sehr komplizierten
elektrischen Erscheinungen, welche den Reiz des Dionaeablattes
fortleiten und auch bei den Sonnentauarten etc. eine wichtige Rolle
spielen, sei hier auf die bis in die neueste Zeit fortgesetzten
Veröffentlichungen der Resultate von Burdon-Sanderson[16] selbst
verwiesen.
Die Bestäubung der Aldrovandia geschieht offenbar über Wasser durch
Insekten, doch ist Näheres über die Bestäubungsvermittler der
unscheinbaren, durch über das Wasser emporragenden Stiel getragenen
Blüten nicht bekannt geworden, zumal die Pflanze in Europa selten
zu blühen scheint. Die Früchte werden unter Wasser gereift. Den Bau
des Samens und die Keimung hat[S. 83]S. Korschinsky[17] neuerdings
genauer beschrieben. Der Keimling, welcher beim Auskeimen den besondern
Deckel der Samenhülle hervorstösst, hat noch ein rudimentäres
Würzelchen und rudimentäre Erstlingsblätter. Es deutet dies darauf
hin, dass die später wurzellose Pflanze, welche in der Keimung
noch grosse Ähnlichkeit mit den Landformen der Sonnentaugewächse hat,
verhältnismässig spät ihr Wasserleben angefangen hat, während bei
der folgenden Gattung, dem Wasserschlauch, der Keimling durchaus
ungegliedert, ohne Wurzelanlage ist.
Fig. 9.
Wasserschlauch (Utricularia). aUtricularia intermedia —
b Blatt (vergr.) — c Blattzweig von U. vulgaris — d
Blase davon (vergr.) — e Aufgeschnittene Blase (stärker vergr.) —
f Kleines Stück der Innenseite der Blase von U. neglecta mit den
vierteiligen Fortsätzen (s. Text) (a nach v. Schlechtendal, d und
f nach Darwin).
[S. 84]
Die weitverbreiteten, in unseren Gewässern häufigen Arten des
Wasserschlauchs (Utricularia), welche (mit einer
fleischfressenden Landpflanze, dem Fettkraut, Pinguicula)
der Familie der Lentibulariaceen angehören, haben in biologischer
Hinsicht viel mit der Aldrovandia gemein. Die häufigste der
sechs bekannten deutschen Arten ist der gemeine Wasserschlauch
(Utricularia vulgaris). Bei ihr tragen die langen, zarten,
verzweigten Stengel, welche wurzellos vom untern Ende aus absterben,
haarförmig zerschlitzte Blätter in wechselständiger, mehr oder minder
zweizeiliger Anordnung. Das Blatt des gemeinen Wasserschlauchs
(Fig. 9) teilt sich in zwei grössere mittlere und zwei kleinere
seitliche Abschnitte, von denen jeder sich mehrfach fiederteilig
oder gabelspaltig in feine cylindrische Zipfel auflöst. An letzteren
treten die bekannten Blasen auf, welche von älteren Autoren als
Schwimmblasen gedeutet wurden, jedoch lediglich dem Fang und der
Verdauung kleiner Wassertiere dienen. Cohn[18] hat 1875 zuerst eine
Darstellung der merkwürdigen Art und Weise gegeben, auf welche die
Utricularia kleine Wassertiere zu fangen vermag, Darwin hat
sodann diese Beobachtungen bestätigt und erweitert. Seitdem haben
sich viele Forscher mit den Wasserschlaucharten in morphologischer
und biologischer Hinsicht beschäftigt, zuletzt am erfolgreichsten M.
Büsgen[19].
Die Blasen (Utricula) haben die Grösse kleiner
Pfefferkörner, sind inwendig hohl mit einer Öffnung an der Seite,
die durch eine von oben herabhängende Klappe verschlossen ist; vor
der Öffnung befinden sich schleimige Härchen, von denen bereits Cohn
vermutet, dass sie den Köder für die Wasserinsekten enthalten. Zwei
borstliche Anhängsel an der Stirn geben dem ganzen Gebilde eine
merkwürdige Ähnlichkeit mit einem Wasserfloh (Daphnia
pulex), wie die Blasen der Aldrovandia grosse Ähnlichkeit
mit Muschelkrebschen etc. haben. Cohn sagt: „Scharenweise
gehen die kleinen Wasserkrebse der gefährlichen Lockung nach,
heben dabei unversehens die einwärts leicht sich zurückschlagende
Klappe; sobald sie aber ins Innere der Blase geraten, verschliesst
die Klappe, die nach aussen sich nicht öffnen lässt, ihnen den
Rückweg. Hierdurch lassen sich jedoch andere[S. 85] nicht abhalten, bald
darauf dem gleichen Schicksal zu verfallen, und ich habe 1874,
wo ich zuerst diese Beobachtung machte, in einzelnen Blasen eine
ganze Menagerie von kleinen Wasserkrebsen, Mückenlarven und anderen
Wassertierchen eingeschlossen gefunden, die vergeblich den Ausweg
aus ihrem grünen Kerker suchten; sie alle waren nach wenigen Tagen
dem Tode rettungslos verfallen; später findet man nur ihre leeren
Schalen, die Weichteile sind völlig aufgezehrt“. Cohn fand ausser
zahlreichen Krebschen (Daphnia, Cypris, Cyclops etc.) noch Naïs
elinguis, Planaria u. a. Würmer, Blattläuse von Wasserpflanzen
(Stratiotes), Rädertierchen, Infusorien und Wurzelfüsser gefangen.
Büsgen teilt mit, dass bei seinen Untersuchungen eine mässig grosse
Pflanze während eines anderthalbstündigen Aufenthaltes in Wasser,
in welchem sich viele Wasserflöhe (Daphnia) befanden, in einer
einzigen Blase zwölf Daphniden einfing. Eine andere Pflanze trug an
jedem Blatte durchschnittlich sechs Blasen. Nur ganz vereinzelte
derselben waren leer. Die meisten waren dicht erfüllt mit Exemplaren
von Chydorus sphaericus. Im ganzen hatte die kleine, etwa 15
cm lange Pflanze mit ihren fünfzehn entwickelten Blättern
etwa 270 ziemlich grosse Krebschen zu sich genommen. Dass
der Wasserschlauch sogar Fischbrut fängt und in Fischteichen
nicht unbedenklich zu dulden ist, berichtet H. N. Mosely[20]. Als ein
Bekannter von ihm eine Pflanze in ein Glasgefäss brachte, in welchem
sich zahlreiche junge Rochen befanden, die vor kurzem ausgebrütet
waren, fand er, dass viele derselben in den Öffnungen der Blasenfallen
gefangen wurden und daselbst verendeten. Mosely brachte nun selbst
ein frisches Exemplar des Wasserschlauches in ein Gefäss mit frischen
jungen Fischen und Laich und fand nach etwa sechs Stunden mehr als
ein Dutzend Fische in Gefangenschaft. Die meisten waren am Kopf
gefasst, der bis zur Hinterwand in die Blase hineingedrungen war, bei
anderen war ein grosser Teil des Schwanzes verschluckt. Drei oder
vier Beispiele wurden beobachtet, in denen ein Fisch mit seinem Kopfe
von einer Blasenfalle und mit dem Schwanze von einer benachbarten
verschluckt war, ein Fall, der mit dem von Klinggraeff[15] bei dem[S. 86]britischen Sonnentau (Drosera anglica) beschriebenen
analog ist und die Frage nahelegt, ob die verzweigten Stirnborsten,
die „Antennen“, nach deren Aufrichten (bei jungen Blasen sind sie über
den Eingang nach unten gekrümmt) erst „die Falle gestellt ist“, nicht
doch auch eine Reizbarkeit besitzen. Mosely fand beim Durchschneiden
der fischgefüllten Blasen die Gewebe des Fisches in einer mehr oder
weniger schleimigen Verflüssigung, wohl infolge seiner Zersetzung,
die vierfiedrigen Fortsätze der Blasendrüsen reichten in die
schleimige, halbflüssige, tierische Substanz hinein und schienen sehr
viel körnige Substanz zu enthalten, jedenfalls das Ergebnis einer
Resorption. Ch. Darwin kam bereits im Anfang seiner Utricularia-Studien
(1874) zu dem Resultat, dass die Utricularia ein Aasfresser
und nicht Fleischfresser ist; so schreibt er[21] am 7. Juli
1874 an J. Hooker: ... „Die Blasen fangen eine Menge Entomostraceen
und Insektenlarven. Der Mechanismus zum Fangen ist ausgezeichnet. Es
findet sich aber vieles, was wir nicht verstehen können. Nach dem, was
ich heute gesehen habe, vermute ich stark, dass sie nekrophag
ist, d. h. dass sie nicht verdauen kann, sondern zerfallende
Substanz absorbiert“, und am 18. Sept. 1874 an Lady Dorothy
Nevill: „... Denn Utricularia ist ein Aasfresser und nicht streng
genommen fleischfressend wie Drosera“. Auch später kommt Darwin[14]
zu dem Resultat, dass die Blasen eine Verdauungsflüssigkeit nicht
ausscheiden, wohl aber Zersetzungsprodukte wie auch fauliges
Wasser und Ammoniaksalze absorbieren und zwar mittels der
vierarmigen Haare, die allein dem Blaseninnern eigen sind und den
gleichgestalteten Haaren der Aldrovandia entsprechen dürften, wie die
echten fleischverdauenden Trichome des Fettkrautes (Pinguicula)
nur den farblosen Digestionsdrüsen der Konkavität der Aldrovandia
entsprechen. „Wir können auch hiernach verstehen“, sagt Darwin bei
Aldrovandia, „wie eine Pflanze durch den allmählichen Verlust einer
der beiden Fähigkeiten (Fleischverdauung und Aasverdauung) nach und
nach der einen Thätigkeit angepasst werden kann, mit Ausschluss der
andern; und es wird später gezeigt werden, dass zwei Gattungen,[S. 87]
nämlich Pinguicula und Utricularia, die zu derselben Familie gehören,
diesen zwei verschiedenen Funktionen angepasst worden sind.“ Büsgen
schildert auf Grund seiner neueren Untersuchungen den Fang bei dem
Wasserschlauch folgendermassen: „Die Antennen und sonstigen von der
Blase nach verschiedenen Seiten ausstrahlenden drüsenlosen langen
Haare bilden eine Art von Leitstangen, auf welchen man sehr oft kleine
Cypridinen nach der Blasenmündung hin wandern sieht. Dort angelangt,
treffen sie die den Eingang umstehenden Köpfchenhaare, welche aus
einer mehr oder minder langen Stielzelle, einer kurzen, besonders
dickwandigen Halszelle und endlich einer etwas dickeren, länglichen
oder runden Kopfzelle zusammengesetzt sind. In der letzteren bestehen
die inneren Schichten der Membran aus einer glänzenden Masse, die
sich mit Jod und Schwefelsäure blau färbt und mit Kalilauge stark
aufquillt, wobei das Protoplasma von der Spitze des Haares her nach
der Basis der Kopfzelle stark zusammengedrückt wird. Stellenweise
findet man die äusserste Membranschicht durch die beschriebene Masse
blasig aufgetrieben. Schon früh erscheint die ganze Kopfzelle von einem
Schleim umgeben, der in reinem Wasser nur sehr schwer sichtbar ist,
mit Methylviolett aber leicht nachgewiesen werden kann, da er sich
mit diesem Reagens hellviolett färbt. Manchmal findet man neben dem
Schleim am Grunde der Kopfzelle eine häutige, faltige Manschette. Aus
dieser und den vorerwähnten Beobachtungen ist zu schliessen, dass der
Schleim einer innern Membranschicht entstammt, die zu einer bestimmten
Zeit aufquillt und die Cuticula sprengt; eigentümlicherweise besitzen
aber auch die mit Schleim und Manschette versehenen Kopfzellen unter
einer festen Membranschicht jene glänzende, quellungsfähige Substanz
und anscheinend auch eine Cuticula. Es müssen diese Bildungen, wenn
obiger Schluss über die Entstehung des Schleimes richtig ist, sehr
rasch regeneriert werden, was übrigens auch sehr im Interesse der
Pflanze liegt, da der letztere als Köder dient“. — Dass es der Schleim
ist, welchem die Kruster nachgehen, schliesst Büsgen daraus, dass sich
dieselben sehr bald an ins Wasser geworfenen Samen mit verschleimender
Aussenschicht ansammeln.[S. 88] Bei dem Besuch der Knöpfchenhaare öffnet
sich die Klappe meist ganz plötzlich mit weitem Spalt, um den
vorwitzigen Gast verschwinden zu lassen und vom nächsten Augenblick
wieder dieselbe Lage anzunehmen. Dieses Öffnen lässt sich jedoch ohne
die Annahme von Reizerscheinungen aus den Elastizitätsverhältnissen
der Klappe erklären. Die Tiere scheinen in ihrer Falle einen
Erstickungstod zu erleiden; wenigstens sah Büsgen noch nach 24
Stunden, Cohn noch nach 6 Tagen die gefangenen Tiere im Innern
umherschwimmen. Die in den Blasen sich findenden symbiotischen
Fäulnis-Bakterien dürften sodann bei dem Wasserschlauch die
Sekretion der Verdauungsflüssigkeit der echten carnivoren Pflanzen
ersetzen.
Auch für die Blatthöhlungen der Blätter unserer Schuppenwurz
(Lathraea squamaria) und der Alpenbartsie glaubt
Scherffel (Mitt. d. bot. Inst. zu Graz, Heft II) gezeigt zu haben,
dass die von Kerner und von Wettstein[22] beschriebenen rhizopodoiden,
die Drüsenzellwand durchbrechenden Protoplasmafortsätze, die Jost
für Wachsausscheidungen hielt, aus Bakterien bestehen, so dass diese
Pflanzen gleichfalls Aaspflanzen wären. Büsgen hat bei den Kopfhaaren
des Wasserschlauchs diese rhizopodoiden Fortsätze gleichfalls gefunden
und ist geneigt, sie als aus Bakterien bestehend zu betrachten. Ob es
sich aber hier wie bei der Schuppenwurz und in den Blattbechern der
Weberkarde nicht doch um Elemente des Protoplasmas, nämlich die
nach den Untersuchungen von Fayod[23] jedem Protoplasma eigenen, sich
auch durch die Scheidewände der Zellen erstreckenden Elemente handelt,
die er Spirifibrillen und Spirosparten nennt, bedarf jedenfalls noch
der Untersuchung.
Fig. 10. A Schlauchtragendes Blatt von Genlisea, etwa dreimal vergr.
l Oberer Teil der Blattspreite — b Schlauch — n Hals —
o Mündung desselben — a Schraubig gewundene Arme (Enden
abgebrochen). B Teil der Innenfläche des in den Schlauch führenden
Halses, stark vergrössert, die Reussenhaare und Digestionsdrüsen zeigend. (Nach
Darwin.)
Den Beweis, dass unsere Wasserschlaucharten thatsächlich von
dem Krebsfange leben, hat Büsgen endgültig durch
vergleichende Kulturversuche an gefütterten und nicht gefütterten
Pflanzen erbracht, wie er für die echten Fleischfresser schon
früher von Reess, Kellermann etc. erbracht wurde. Der Zuwachs
der gefütterten Pflanzen war etwa der doppelte von dem der nicht
gefütterten u. s. f. — Im tropischen[S. 89] Amerika und Asien leben auch
Wasserschlaucharten auf dem Lande oder auf dem Moos der Bäume,
aber auch hier von den Tieren ihres Wohnorts. Utricularia
nelumbifolia kommt in Brasilien auf den hochgelegenen Felsen der
Orgelberge vor, aber sie wird hier nur in dem Wasser gefunden, welches
sich auf dem Grunde der Blätter einer grossen Tillandsia
ansammelt, sie gelangt ausser durch Samen durch Ausläufer von einem
Tillandsiabecken ins andere. Ihre Blasen fangen in gleicher Weise
Wassertiere, wie die unserer Wasserschlaucharten. Darwin fand bei
neun Utriculariaarten, die[S. 90] er untersuchte, allenthalben die Blasen
mit Tieren und Tierresten erfüllt. Von verwandten Gattungen sei hier
noch die merkwürdige Genlisea ornata (Fig. 10) aus Brasilien
erwähnt, bei welcher das schlauchtragende Blatt die Beute nicht mittels
einer elastischen Klappe, sondern durch eine einer Aalfalle ähnliche,
wenngleich kompliziertere Vorrichtung fängt. Die schmale Blattscheibe
trägt nämlich am Ende eine Blase, die sich in eine etwa fünfzehnmal so
lange Röhre fortsetzt. Seitlich der Mündung der letztern entspringt
auf jeder Seite ein aus einem spiralig gerollten, linealen Blattzipfel
gebildeter Cylinder. Diese Seitenröhren wie die Hauptröhre sind
mit langen, nach abwärts gerichteten Borsten (Reussen) und mit den
vierzelligen Drüsen der Utricularia bekleidet. In Seitenhälsen und im
Hauptrohre fanden sich Überreste von Würmern und Gliedertieren.
Unsere Wasserschlaucharten sind ausgeprägte Insektenblütler, die
ihre Blütentrauben mit gelben, auffälligen Blüten auf langem Stiel
über Wasser senden. Die Blumenkrone, deren Bau und Entwickelung
zuerst Buchenau[24] genauer untersucht und deren Bestäubungseinrichtung
Hildebrand[25] erläutert hat, birgt in einem Sporn den Honig zur
Anlockung der Insekten. Ein Insekt muss, wenn es zur Gewinnung des
Honigs seinen Rüssel unter die Oberlippe steckt, zuerst mit seiner
Oberseite einen die Staubgefässe überragenden, mit der papillösen Seite
nach unten umgebogenen Narbenlappen berühren, so dass dieser, wenn
das Insekt schon vorher eine Blüte besucht hat, mit Pollen derselben
behaftet wird, sodann die Staubgefässe, die es von neuem mit Pollen
behaften. Der Pollenlappen ist, ähnlich der in der Blüte der an
Flüssen und Bächen Deutschlands jetzt weit verbreiteten amerikanischen
Gauklerblume (Mimulus luteus) und weniger anderen Blumen,
reizbar, sodass er bei der Berührung sofort nach oben umklappt
und so eine Übertragung des Blütenstaubes derselben Blüte auf die
Narbe beim Rückzug des Insektes unmöglich macht.[26] Die ausgeprägte
Honigblume wird vermutlich durch Hymenopteren und Schmetterlinge
bestäubt, doch ist hier wie bei den meisten Insektenblumen unserer
Gewässer über die Bestäubungsvermittler näheres nicht bekannt. —
Die Früchte der Utricularien[S. 91] werden nicht, wie dies bei den meisten
echten Wasserpflanzen der Fall ist, unter Wasser gereift, sondern, wie
auch bei der Wasserfeder (Hottonia) und Lobelie (Lobelia
Dortmanni), über Wasser. Die vielsamigen Kapseln streuen ihren
Samen ins Wasser aus, wo er an der Oberfläche weit verbreitet und von
wo er durch Wassertiere auch von Gewässer zu Gewässer übertragen wird.
Eine Verbreitung der vegetativen Organe durch die Larven der
Köcherfliegen (Phryganiden) hat Gilbert[27] beobachtet. Nach ihm
werden die Winterknospen des mittlern Wasserschlauchs (Utricularia
intermedia) von diesen Insekten zum Larvenköcher verwendet, wodurch
sie an andere Stellen gelangen, an denen sie zu weiterer Entwickelung
kommen können.
Gegen Tierfrass besitzen die Wasserschlaucharten spitze
Stacheln und chemische Schutzmittel. Mit Alkohol oder heissem Wasser
ausgelaugte Pflanzenstücke wurden von kleinen Krustern bei einem
Versuch Büsgens bald gefressen, während frische Pflanzenteile gänzlich
verschont blieben. Der schützende Stoff schien Gerbstoff in einer
schwachen Lösung zu sein, da bei Behandlung mit doppeltchromsaurem
Kali eine nicht auswaschbare Färbung des Zellinhaltes auftrat. Nur die
Stachelhaare färbten sich intensiv braun.
Wie die Wasserschlaucharten hinsichtlich der Blüteneinrichtung eine
grosse Übereinstimmung mit ihren Verwandten auf dem Lande (dem
gleichfalls tierfangenden, aber fleischverdauenden, nicht aasfressenden
Fettkraut, Pinguicula) haben, so stimmt auch eine weitere
submerse Wasserpflanze, die Wasser- oder Sumpffeder (Hottonia
palustris), welche aber zur Familie der Primulaceen gehört (Fig.
11 S. 92) mit unseren Schlüsselblumenarten in Bezug auf die
Fortpflanzungsorgane völlig überein, während sie im übrigen ganz die
abweichenden Vegetationsorgane der bisher betrachteten Wasserpflanzen
hat. Es giebt kaum einen grelleren Gegensatz zwischen Land- und
Wasserpflanze als es gerade der zwischen der Schlüsselblume (und
anderen Primulaceen) und der Wasserfeder ist — dort grosse,
einfache, ganze Blätter, die dicht dem Wurzelstock entspringen, hier
die feingeteilten Wasserblätter des Hornblattes, der Tausendblätter[S. 92]
(Myriophyllum) etc., an wurzellosem, nur anfänglich im Schlamm
steckenden Stengel.
Fig. 11.
Heterostylie der Wasserfeder (Hottonia palustris). a Langgrifflige
Blüte — b Narbenpapillen derselben — c Kurzgrifflige Blüte —
d Narbenpapillen bei gleicher Vergrösserung wie in b. (Nach H.
Müller.)
Zur Blühzeit steigt die Pflanze zur Oberfläche auf und streckt ihren
terminalen Blütenstand über das Wasser empor. Die Sumpffeder ist,
wie unsere Schlüsselblumen, ausgeprägt heterostyl-dimorph, d. h. es
kommen zweierlei, wie männliche und weibliche Stöcke zusammengehörige
Sorten von Stöcken vor, deren eine lauter langgrifflige Blüten mit
tiefstehenden Staubgefässen trägt, während in den Blüten der andern die
Griffel kurz, tief in der Blütenröhre versteckt sind, die Staubgefässe
aber weit aus derselben hervorragen. Die Verschiedenheit zwischen
den beiden Formen hat bereits die Aufmerksamkeit des Vaters der
heutigen Blütenbiologie, Christian Konrad Sprengel, im Jahre 1793
erregt, welcher sagt, dass er die Existenz der zwei Formen nicht
für zufällig halte, obschon er ihren Zweck nicht erklären könne.
Bekanntlich hat Darwin[28] die Bedeutung dieser Zwiegestaltigkeit[S. 93]
bei Primeln erkannt und durch Versuche festgestellt, dass erfolgreich
allein die Bestäubung der langen Griffel durch die Pollen der grossen
Staubgefässe der andern Form und die der kurzgriffligen Narben der
letztern durch die Staubkörner aus den Staubgefässen der erstern
wirkt, dass mit anderen Worten die Bestäubung zwischen den Organen
gleicher Höhe einzutreten hat, wenn sie vollen Erfolg haben
soll. Andere als diese „legitimen“ Bestäubungsweisen (also
„illegitime“) haben bei vielen heterostylen Pflanzen überhaupt keinen,
bei anderen Arten nur einen schwachen Erfolg. Nach der Entdeckung der
heterostylen Pflanzen, zu denen zahlreiche Arten, von Wasserpflanzen
noch Arten der Gattungen Villarsia, Menyanthes — unser
dreiblättriger Fieberklee — und Polygonum, gehören, hat man
noch die verwandten Gruppen der tristylen oder lang-, kurz- und
mittelgriffligen Pflanzen und die rechts- und linksgriffligen entdeckt.
Zu den ersteren gehört der an den Ufern unserer Gewässer wachsende
Färbeweiderich (Lythrum Salicaria) nebst vielen ausländischen
Sauerkleearten etc. Die Staubgefässe bilden in derselben Blüte zwei
Höhensätze, in Bezug auf welche bei den dreierlei Stöcken der Griffel
lang, kurz oder mittellang ist. Bei den rechts- und linksgriffligen
Pflanzen (Verwandte unserer Kartoffel, wie Solanum rostratum,
gehören dazu) sind zweierlei Stöcke da, in deren Blüten der Griffel
entweder links oder rechts von dem Staubgefässbüschel steht und nach
entgegengesetzter Seite gebogen aus der Blüte hervorsieht. Auch hier
gilt das Gesetz der legitimen Bestäubung, d. h. des erfolgreichen
Zusammenwirkens der Organe gleicher Lage. Dieses Zusammenwirken wird ja
unter gewöhnlichen Umständen das natürliche sein, wenn gewisse Insekten
alle Blüten in gleicher Weise besuchen, indem sie dann den Blütenstaub
an einer Stelle absetzen, an welcher sie ihn in den anderen Blüten
an ihrem Körper aufgenommen haben. Ein honigsuchendes Insekt
wird also bei unserer Wasserfeder in der That am Vorderkörper
den Blütenstaub der tiefgelegenen Staubbeutel, am Hinterleib
den der langen Staubgefässe mit sich führen; mit letzterem kann es
aber nur die Narbe der langen Griffel, mit ersterem die der kurzen[S. 94]
Griffel bei seiner gewöhnlichen Art des Besuchs erlangen.
Anders ist es mit pollensuchenden Insekten. Sie haben
keinen Grund, die kurzen Griffel zu besuchen, während beim Besuch der
kurzen Staubgefässe der staubbeladene Kopf gelegentlich die Narbe der
langen Griffel berühren wird. Stellen wir uns nun vor, dass bei der
Ausbildung einer Blüteneinrichtung nur oder vorwiegend honigsuchende
Insekten mitgewirkt haben, so würden Formen der Heterostylie zu
stande gekommen sein, wie sie sich thatsächlich häufig finden, bei
denen illegitime (künstliche) Bestäubung fast gänzlich
wirkungslos ist, während da, wo pollensuchende Insekten neben
den honigsuchenden regelmässige Gäste sind, eine Heterostylie
entstanden sein wird, bei welcher die legitime Bestäubung zwar die
erspriesslichste ist, von den illegitimen Bestäubungen aber die
der langen Griffel immerhin jener nahekommt. So ist es nach den
künstlichen Befruchtungsversuchen von John Scott und Herm. Müller[26]
bei der Sumpffeder, bei welcher nach Herm. Müller die Bestäubung
durch pollenfressende Fliegen in ausgedehntem Masse ausser durch
saugende Hautflügler (Pompilus viaticus) und Fliegen
(Empis, Eristalis, Rhingia) bewirkt wird. Bei den Versuchen Müllers
ergaben die zwei legitimen Verbindungen mit den zwei illegitimen
zusammen verglichen Samenkörner im Verhältnis von 100 zu 61. Es war die
mittlere Zahl der Samenkörner auf die Kapsel bei
1.
legitimer Bestäubung der langgriffligen Form
91.4,
2.
illegitimer Bestäubung der langgriffligen Form
von einer verschiedenen Pflanze
77.5,
3.
legitimer Bestäubung der kurzgriffligen Form
66.2,
4.
illegitimer Bestäubung der kurzgriffligen Form
mit Pollen dieser Form von einer verschiedenen Form
18.7.
Sehr deutlich sprechen die Resultate für die schädliche Wirkung der
Inzucht bei Kreuzung der Blüten ein und desselben Stockes und bei
Selbstbefruchtung. Die mittlere Samenzahl war hierbei nur 15.7 bei den
langgriffligen und 6.5 (wie bei Scott) bei den[S. 95] kurzgriffligen Blüten
anstatt 77.5 und 18.7 (bei illegitimer Kreuzung). Die ungleiche Grösse
der Narbenpapillen bei den grossen und kleinen Griffeln stimmt bei der
Sumpffeder ebenso wie bei anderen Heterostylen überein mit der
ungleichen Grösse der Pollenkörner in den oberen und unteren Antheren
(die letzteren kleiner), für welche Verschiedenheit die bisherigen
Erklärungsversuche nach neueren Versuchen nicht auszureichen scheinen.
Den frei im Wasser flottierenden Arten submerser Gewächse,
welche wurzellos sind, schliessen sich solche in grösserer Zahl an,
die unter gewöhnlichen Verhältnissen zwar ein Wasserleben führen, aber
am Boden festgewachsen sind. Bei ihnen ist die Wurzel in den
meisten Fällen nur Haftorgan. Sie haben zumeist die Fähigkeit, bei
Wassermangel auch in der Luft längere oder kürzere Zeit zu vegetieren,
oder gar eine besondere Luftform zu bilden. Eine Umbildung der
Blattorgane, welche der neuen Lebensweise angepasst (Verbreiterung
und Verkürzung der Blattflächen, Auftreten von Spaltöffnungen etc.),
tritt oft sehr bald, zuweilen sogar an einzelnen Blattteilen, die
künstlich in dem neuen Medium gehalten werden, ein. Solche Landformen
sind von Schenk z. B. für das Tausendblatt (Myriophyllum)
beschrieben, deren Arten sonst mit ihren feingeteilten Blättern ein
ausgeprägtes Wasserleben führen. — Die Myriophyllumarten
besitzen ähnlich wie die Wasserpest, die Hornblattarten,
die Wasserschlaucharten eine sehr üppige vegetative Vermehrung,
abgebrochene Zweige, die durch Strömung oder Tiere verschleppt sind,
entwickeln sehr bald wieder eine weitverbreitete Vegetation am Grunde
des Gewässers. Solche üppig sich vermehrende Arten kommen an manchen
Orten überhaupt selten zur Blüte. Das Ährentausendblatt
(Myriophyllum spicatum) trägt in seinen in die Luft
emporgestreckten Blütenständen in den Achseln kleiner Brakteen zu
oberst männliche, unten weibliche Blütenstände, welche sich viel
früher als die ersteren entwickeln (die Pflanze ist proterogynisch).
Der platte, leicht verstäubende Pollen, die grossen, an dünnen
Staubfäden lebhaft im Winde flatternden Antheren, wie auch die stark
höckerigen Narben kennzeichnen die Pflanze als windblütig[S. 96]
(anemophil), während bei dem quirlblättrigen Tausendblatt dadurch,
dass die Blüten in den Blattwinkeln sitzen, die Anpassung an den
Pollentransport durch den Wind vermindert ist. Es kommen aber wie bei
den anderen Myriophyllumarten (M. alternifolium habe ich nicht
untersucht) auch unter Wasser normale (nicht wie Hermann Müller in
seinem Referat meiner Arbeit[7] vermutete[29] kleistogame) Blüten zur
Entwickelung, so dass hier vermutlich Übergänge von der Windbefruchtung
zur Wasserbefruchtung vorliegen. Nach dem Blühen tauchen auch bei M.
spicatum die Ähren unter Wasser, um hier ihre Früchte zu reifen.
Zu den submersen Gewächsen, die aber in einigen ihrer Arten Übergänge
zu den Schwimmpflanzen bilden, indem sie bei diesen andersgestaltete,
an der Oberfläche ausgebreitete oder schwimmende Blätter entfalten,
gehören ferner die Wassersternarten und die Laichkräuter,
deren submerse Blätter aber nicht haarförmig zerteilt, sondern
einfach, schmallinealisch etc. sind. Die völlig submersen Wassersterne
bilden auf dem Boden der Gewässer eine sehr üppige, dichte, hellgrüne
Vegetation, die an die der kleinen Armleuchtergewächse (Nitella)
erinnert, während die schwimmenden breitblätterigen Blattrosetten die
Oberfläche den Wasserlinsen ähnlich überkleiden. Die einfache Blüte
besteht aus einem starren Staubgefäss oder aus einem zweifächerigen
Fruchtknoten mit kurzen Griffeln. Auch sie wird gewöhnlich für
windblütig gehalten, doch dürften die an der Oberfläche schwimmenden
Rasen mit ihren starren Staubfäden und spärlicher Pollenproduktion eher
die später zu erörternden Eigentümlichkeiten der Wasserlinsen
besitzen. Die zahlreichen untergetauchten Blüten haben
Wasserbefruchtung. Die Gruppe Pseudocallitriche, zu welcher
unser Herbstwasserstern (Callitriche autumnalis) gehört,
besitzt, wie die früher besprochenen Wasserblütler, Pollenkörner
ohne äussere Zellhaut (Exine), sind nach Jönsson ölhaltig
und leichter als Wasser, steigen daher nach der Oberfläche zu,
wobei sie die Befruchtung vollziehen können, es wechseln nicht selten
mehrere weibliche und männliche Stengelglieder mit einander ab; bei der
Gruppe der Eucallitrichen dagegen, die auch wohl als Varietäten
ein und[S. 97] derselben Art, des Frühjahrswassersterns (Callitriche
verna) betrachtet werden, sind die Pollenkörner noch von einer
äussern etwas höckerigen derben Haut umkleidet und an den Antheren ist
die den Luftblüten eigentümliche Faserschicht vorhanden, welche bei
dem Aufspringen der Staubbeutel eine wichtige Rolle spielt.
Zieht man die frischgrün beblätterten Zweige der Wassersterne oder
anderer submerser Wasserpflanzen (Hornblatt, Tausendblatt etc.) aus
dem Wasser, so findet man zwischen dem feinen Blattgewirr ein buntes
Tierleben von Wasserasseln, Flohkrebsen, Milben, Käfern, Larven,
Schnecken etc., das wohl hauptsächlich durch die reiche Entwickelung
des Sauerstoffs bei der Assimilation herbeigezogen wird. Es ist
kaum anders zu erwarten, als dass dieses reiche Tierleben, mit
welchem diese Wasserpflanzen in steter Berührung sind, mehr als alle
Landpflanzen, dem pflanzlichen Organismus zu mancherlei Anpassungen
Veranlassung gegeben hat. Schutzvorkehrungen gegen Tierfrass sind
thatsächlich mehrfach bekannt geworden, wie später noch hervorgehoben
werden wird. Ob aber die Pflanze Glieder dieser Tierwelt sich
zu Nutzen gemacht hat und mit ihnen im Bunde gegenseitiger
Förderung lebt ist noch kaum untersucht worden und verdient
seitens der Süsswasserforschung besondere Beachtung. Hier sei nur
hervorgehoben, dass diesen Tieren sicherlich die Reinhaltung
der assimilierenden Blattflächen zuzuschreiben ist und dass
es mir immer geschienen hat, als ob an Standorten, wo ein besonders
reiches Tierleben sich bemerkbar machte, die submersen Pflanzenorgane
frischer grün und reiner aussahen, als an Orten wo diese fehlte. An den
letzteren sind die Blätter und Blattzipfel oft völlig eingehüllt und
unkenntlich gemacht von zahlreichen Algen (Kieselalgen, wie Exilaria,
Gomphonema etc.), Wasserpilzen (Chytridiaceen), Infusorien (Vorticellen
etc.), welche ja bekanntlich manchem der erwähnten Tiere zur Nahrung
dienen.
Das Hauptkontingent liefern zur Wasserflora die Laichkräuter
(Potamogeton), von denen etwa 50 Arten überhaupt und etwas weniger als
die Hälfte aus den süssen Gewässern Deutschlands,[S. 98] Österreichs und der
Schweiz[30] bekannt sind. Sie weichen durch ihre einfachen Blätter wie
durch mannigfache Variationen von den völlig untergetauchten Formen
bis zu den ausgeprägtesten Schwimmpflanzen, wie auch schliesslich
dadurch ab, dass sie ein am Boden kriechendes sehr ausgebreitetes
Rhizom haben. Im fliessenden Wasser treiben sie, ähnlich wie andere
festwurzelnde Flussgewächse (Flusshahnenfuss etc.), lange Laubtriebe,
welche durch besondere mechanische Elemente in ihrem anatomischen
Aufbau der hohen Inanspruchnahme durch die Wasserströmung völlig
gewachsen sind. In stehenden Gewässern sind die Stengelglieder oft sehr
verkürzt. Nach der Meinung von Schenk stammen die Laichkräuter
von Landpflanzen ab, die sich erst der amphibischen, gegenwärtig
nicht mehr erkennbaren, Lebensweise und sodann der Lebensweise der
Schwimmpflanzen, zuletzt aber der submersen Lebensweise angepasst
haben. Es würde dann unser gemeines schwimmendes Laichkraut
(Potamogeton), welches nur ausgeprägte Schwimmblätter besitzt,
der Urtypus unserer jetzt lebenden Laichkrautarten sein. Hierfür
spricht auch dessen Verbreitung. Nach A. de Candolle[31] giebt
es nur 19 Phanerogamen, die sich über mehr als die halbe Erdoberfläche
verbreitet haben, zu denen z. B. das einjährige Straussgras
(Poa annua), die Krötenbinse (Juncus bufonius),
unsere Brennesseln (Urtica dioica und U. urens),
Gänsefussarten (Chenopodium album und Ch.
murale), eine Taubnessel (Lamium amplexicaule), der
schwarze Nachtschatten (Solanum nigrum), die gemeine
Sternmiere (Stellaria media), die Gänsedistel
(Sonchus oleraceus), das Hirtentäschel (Capsella
bursa pastoris) und das schwimmende Laichkraut von
bei uns gemeinen Pflanzen gehören. Das letztere ist aber die
einzige kosmopolitische Wasserpflanze, alle übrigen sind
Landgewächse und zwar Ruderalpflanzen. Die übrigen Vertreter des
Laichkrautstammes Heterophylli (Potamogeton oblongus,
P. fluitans, P. spathulatus, P. rufescens,
P. gramineus, P. nitens, P. Hornemanni) sind
der submersen Lebensweise bereits mehr oder weniger angepasst,
indem sie oft nur wenige oder gar keine Schwimmblätter mehr zeigen,
während ihre untergetauchten Blätter[S. 99] zwar den anatomischen Bau,
aber noch nicht die Gestalt echter Wasserblätter haben. Bei den
Gruppen der Homophylli und Enantiophylli[30] fehlen bereits die
Schwimmblätter gänzlich und es bahnt sich der Übergang von den
breitblätterigen zu den sehr schmalblätterigen Arten an, der in den
Gruppen Chloëphylli und Coleophylli echt submerser Laichkräuter
(Potamogeton pusillus, trichoides etc.) sein Extrem
erreicht. Eine gleiche Stufenleiter der Anpassung ergiebt sich auch
bezüglich der Blüteneinrichtungen. Das schwimmende Laichkraut
und seine nächsten Verwandten sind ausgeprägte Windblütler, die
ihre Blüten in dicht gedrängter Ähre an langer dicker Spindel über
das Wasser heben. Das kleine Laichkraut hat bereits nur noch
kleine, meist vierblütige Ährchen und das Haar-Laichkraut in
jeder Blüte nur noch einen Fruchtknoten (sonst vier). Dazwischen und
von dem letztern weiter finden sich Übergangsformen bis zu solchen,
welche den submersen Wasserpflanzen sehr nahe kommen, deren Blüten
nur aus einem Staubgefäss, oder einem Fruchtknoten bestehen. P.
pectinatus erhebt einen Blütenstand überhaupt nicht mehr über
Wasser, sondern besitzt lange dünne schwimmende Blütchen, die zur
Fortpflanzungszeit nur wenig über das Wasser emporragen. Ob bei unserer
zweiten Gattung der Laichkräuter, der Zanichellie, bereits eine
echte Wasserbestäubung eintritt nach Art der der Meerstrands-Ruppie,
bedarf noch der nähern Untersuchung, ist aber nicht unwahrscheinlich,
zumal auch nach Fritzsche dem Pollenkorn die äussere Hülle
fehlt. Die Windblütigkeit der Laichkräuter, deren Blüten oft dicht an
der Wasseroberfläche stehen, wird nach v. Kerner[32] unterstützt
durch einen Ausschleuderungsmechanismus der Blüte, durch welchen
der Blütenstaub in die Luft gelangt.
Schwimmgewächse.
Wie die untergetauchten Blätter, so sind auch die Schwimmblätter
der Wasserpflanzen völlig ihren Lebensverhältnissen angepasst. Die
schwimmenden Blätter sind nie zerteilt, stets einfach, ganzrandig
rundlich oder elliptisch länglich u. dergl., auf der Oberseite
lederartig, schwer benetzbar, im anatomischen Bau eine dreifache[S. 100]
Anpassung verratend, eine mechanische zum Schutz gegen die an der
Oberfläche am heftigsten wirkenden Bewegungen des Wassers und gegen
die heftigere Wirkung der auffallenden Regentropfen, eine Anpassung
an die schwimmende Lebensweise und an die intensivere Einwirkung des
direkten Sonnenlichtes. Dementsprechend zeigt das Schwimmblatt an der
Oberseite, unter einer chlorophyllfreien wasserhaltigen Epidermis, die
assimilierenden grünen Zellen als Palissadenparenchym ausgebildet.
Unter diesem sind grössere lufthaltige Intercellularräume, welche das
Blatt schwimmend erhalten. Die Spaltöffnungen, welche zur Ermöglichung
des Transpirationsstromes bei den Schwimmpflanzen — im Gegensatz zu
den submersen Pflanzen — vorhanden sind, finden sich hier allein
auf der Oberseite des Blattes, die mit der Luft in Berührung steht,
während sie bei den Luftblättern zur Verhinderung einer zu starken
Transpiration in der Epidermis der Unterseite zur Ausbildung gelangen.
Die zur Assimilation nötige Kohlensäure wird hier hauptsächlich aus der
Luft, wohl aber auch durch Diffusion von der dem Wasser aufliegenden
Blattunterseite besorgt. Da, wo Blattstiele vorhanden sind, zeigen
diese die besondere Eigentümlichkeit, dass sie ihr Längenwachstum genau
nach der Tiefe des Wassers einrichten, so dass das Blatt immer zum
Schwimmen kommt.
Die Wasserlinsengewächse, von denen nur eine Art (Lemna
trisulca) bei uns submers vorkommt, gehören zu unseren einfachsten
Schwimmpflanzen, die gleichzeitig durch ihre Kleinheit, wie durch ihr
massenhaftes Auftreten — sie bedecken oft weithin die ganze Fläche
der Gewässer — auffallen. In unseren Gewässern finden sich nur fünf
Arten, von denen die kleinste, die wurzellose Wasserlinse
(Wolffia arrhiza) nur die Grösse von 1 mm erreicht —
nur die in Bengalen lebende Wolffia microscopica, zweifellos
die kleinste Phanerogame, ist noch winziger — gar keine Wurzeln
erzeugt und in einzelnen Sprossen (Stengelgliedern) vorkommt, während
die übrigen Arten ein oder mehr Adventivwurzeln nach abwärts treiben,
welche mehr als zur Nahrungsaufnahme zur Erhaltung der horizontalen
Lage dienen dürften. Es sind dies ausser der untergetauchten
Wasserlinse (Lemna trisulca), welche bereits[S. 101] erwähnt
wurde, noch die buckelige (L. gibba) und grosse
Wasserlinse (Spirodela polyrrhiza) mit mehreren Wurzeln,
welche gesellig mit der auch häufig in ganz reinen Beständen wachsenden
kleinen Teichlinse (L. minor), unserer häufigsten Art,
vorkommen. Hegelmaier[33], auf dessen Monographie (die auch von
Engler in seinen „Natürlichen Pflanzenfamilien“ hauptsächlich
zu Grunde gelegt ist) wir hier besonders hinweisen müssen, weil es
unmöglich ist, im Rahmen dieser Abhandlung die obwaltenden Verhältnisse
auch nur einigermassen genau zu schildern, hat 19 Arten von
Wasserlinsen unterschieden, welche über den ganzen Erdkreis verbreitet
sind:
1) Die wurzellosen Wolffioideen, bei denen auf dem Rücken des Sprosses
in kleinen Grübchen (zwei bei Wolffia Welwitschii, eine bei den übrigen
Arten) die einfachen Blüten mit einem Staubgefäss und Stempel (resp.
einer männlichen und einer weiblichen Blüte) entspringen und sich
auch sonst durch die Sprossverhältnisse auszeichnen. Sie zerfallen
in die beiden Untergattungen Wolffia und Wolffiella, mit je acht
und vier Arten, von denen letztere durch ihre abnorme Gestalt mit
länglich-pfeilförmigen Hauptsprossen ausgezeichnet sind.
2) Bei den bewurzelten Lemnoideen, unter denen ausser durch die
Sprossverhältnisse noch durch das Vorhandensein von vielen Wurzeln
Spirodela [mit zwei Arten: Sp. oligorhiza (im indisch-malayischen
Gebiet) und unserer L. polyrrhiza] von Lemna abgegrenzt wird, kommen
die Blütenstände (Blüten) mit je zwei Staubblüten(gefässen) und einem
weiblichen Blütenstempel in seitlichen Taschen zur Entwickelung. Von
Lemna sind ausser den einheimischen Arten noch zwei unserer kleinen
Wasserlinse nahe stehende Arten bekannt. Die Wolffiaarten scheinen
am meisten dem Wasserleben angepasst zu sein, da bei ihnen auch an
der Oberseite wenige, bei Wolffiella sogar gar keine Spaltöffnungen
vorhanden sind und die Blüten ganz auf dem Rücken stehen.
Fig. 12.
Wasserlinsen (Lemnaceen). ALemna minor (b Blüte) —
B Blütenstand — C Stempel (vergrössert) — D Pollenkorn
nach 610facher Vergrösserung — ELemna gibba — F Frucht
derselben (das Operculum o wird bei der Keimung abgehoben) — GLemna trisulca — HWolffia arrhiza, a in natürlicher
Grösse, b schwach vergrössert, c im Durchschnitt, um den dem
Spross eingesenkten Blütenstand zu zeigen — MWolffia repanda (von
Loanda) — JWolffia hyalina (aus Ägypten) — KWolffiella
gladiata (aus Mexiko) — LWolffiella oblonga (aus Chile)
(Hc, J, K, L, M nach Hegelmaier).
Bezüglich der Bestäubungsverhältnisse der Wasserlinsen, die
zu den interessantesten Vorkommnissen der Blütenbiologie überhaupt
gehören, liegen zumteil scheinbar widersprechende[S. 102] Beobachtungen
vor, die aber recht wohl zugleich richtig sein können. Hat doch
die Blütenbiologie mehrfach gezeigt, dass eine Pflanze z. B. in
einer Gegend typisch proteranderisch, in einer andern, besonderen
Verhältnissen entsprechend, proterogynisch ist. Ich werde zunächst die
Verhältnisse schildern, wie ich sie bei unserer gemeinen Wasserlinse
um Greiz typisch getroffen habe und zwar in einem ruhig gelegenen
Teiche sowohl, wo sie vom Mai bis in den Juli hinein blühte, wie auch
im Zimmer. Der monöcische[S. 103] Blütenstand besteht hier entweder aus einem
höher stehenden kurzgriffeligen Stempel und zwei tiefer stehenden,
wie jener nach oben gerichteten Staubgefässen mit nicht allzulangem
Filament und gelben Antheren, oder Stempel und Staubgefässe kommen an
verschiedenen Stellen des Thalloms hervor, von einer unregelmässig
zerreissenden Hülle umschlossen. Die beiden Staubgefässe entwickeln
sich nach einander, aber längere Zeit bevor der Stempel hervorbricht.
Proterandrische Dichogamie und Stellung schliessen demnach auch
Selbstbestäubung aus. Ebenso ist es undenkbar, dass der Wind bei
der Kürze der starren Sexualorgane und der geringen Pollenmenge, die in
den beiden Staubblüten erzeugt wird, unmittelbar über dem dicht durch
Lemnarasen bedeckten Wasserspiegel bei der Übertragung des Pollens
eine Rolle spielt. Von den bekannten zoidiophilen Pflanzen weicht die
Wasserlinse ab durch den gänzlichen Mangel eines gefärbten Perigons
oder anderer auffälligen Anlockungsmittel der Blüten; trotzdem
glaube ich behaupten zu können, dass Lemna minor ausgeprägt
zoidiophil ist — und zwar angepasst den auf der Oberfläche des
Wassers sich umhertreibenden Wassertieren (Hydrometriden etc.).
Die Pollenkörner sind stachelig mit zahlreichen Protuberanzen
besetzt, wie bei den ausgeprägtesten Entomophilen (z. B.
Malva, Cucurbita, Compositen). Die Pollenkörner haben einen
Durchmesser von etwa 26 µ, ihre Stacheln eine Länge von etwa 1 µ,
sodass sie einerseits leicht dem Körper der über die Staubgefässe
streichenden Insekten, anderseits der etwas konkaven Narbenscheibe am
Ende des Griffels anhaften, während sie durch den Wind wohl kaum von
der dehiszierenden Anthere losgerissen werden könnten. Die auf den
Lemnarasen oder zwischen ihnen umherkreisenden Kerfe müssen anderseits
sowohl mit den Staubgefässen als den Narben in Berührung kommen, ohne
dass sie besonders darauf aufmerksam gemacht werden. Die Pflanze hat
es hier nicht nötig, besondere Lockmittel zu gebrauchen, ohne
allen Aufwand und ohne eine andere Gegenleistung, als etwa die
Gewährung eines festen Untergrundes, erreicht sie dasselbe, was
die „Blumen“ durch[S. 104] Entwickelung von Farbenpracht, von Honigsaft
und Wohlgeruch, die zuweilen nur unberufene Gäste anlocken,
erzielen. Trelease[34] hatte später (ehe er indessen
meine Arbeit[7] kannte) vermutet, dass die Lemnarasen zusammengedrängt
würden und dabei sich wechselseitig befruchteten, an seinen Exemplaren
waren die Blütenstände proterogynisch. Im Zimmer dehiszierte das erste
Staubgefäss drei Tage nachdem die Narbe empfängnisfähig geworden
und ebenso lange nachher das zweite Staubgefäss des Blütenstandes.
Auch Hegelmaier hatte L. minor als proterogynisch
bezeichnet und Spirodela polyrrhiza. George Engelmann[35]
hat aber bei letzterer gleichfalls Proterandrie beobachtet.
Federico Delpino bestätigt meine Beobachtungen und Deutung der
Blütenentwickelung; vermutet aber, dass Schnecken bei der Befruchtung
in Betracht kommen. Er sagt[36]: „A rinforzare la interpretazione di
Ludwig noi potremmo addurre il singolare appianamento e allivellamento
di caulomi, antere e stimmi; per il che per manifesta la designazione
a pronubi striscianti e perambulanti. Così questa rara associazione di
caratteri biologici, efficiente un apparecchio florale reptatorio, si
riproduce in questa minuscole pianticelle. E non deve far meraviglia,
poi chè le lemnacee senza dubio appartengono alla famiglia delle
aroidee, presso la quale tanto frequentemente accorono apparecchi
reptatorii. Ludwig non menziona fra i pronubi le chioccioline
aquatiche, eppura noi congetturiamo que queste sia noi principali
ausiliarii della dicogamia delle lemne“. — Henry Gillmann[37]
berichtet von der grossen Wasserlinse (Sp. polyrrhiza),
die er in einem Gefäss mit Wasser beobachtete, dass sich um 4 Uhr
15 Min. nachmittags die Stamina entfalteten, in der Nacht aber sich
zurückbewegten (but closed at night). Am andern Morgen um 7 Uhr
entfalteten sich wieder zunächst drei Blütenstände, bis 7 Uhr 45 Min.
18, und bis 8 Uhr 30 Min. hatten sich 30 Blüten geöffnet.
Die von Delpino vermutete Beteiligung der Schnecken bei der
Pollenübertragung der Lemnaceen, wie auch die von andrer Seite
vermutete bei einer Aroidee, der Schlangenwurz (Calla
palustris), einer Sumpf- und Uferpflanze unserer Teiche, verdient[S. 105]
noch besondere Beachtung der Botaniker und Zoologen, da über die
Schneckenbefruchtung (Malacophilie) der Pflanzen viel hin und
her gestritten worden ist, die bisherigen Beobachtungen aber zur
Entscheidung der Frage nicht ausreichen. Auffällig und bemerkenswert
bleibt es immerhin, dass diese beiden nach Delpino u. a.
malacophilen Pflanzen in ihren Blattorganen besondere Schutzmittel
gegen Schneckenfrass haben. Nach Stahl[4] bilden die
Raphiden (Bündel von beidseitig sehr spitzen Nadeln des
Kalkoxalates) eins der wirksamsten Schutzmittel gegen Schneckenfrass.
Bei der gemeinen Schlangenwurz hat die Wirkung dieser Raphiden
in den Blättern schon der alte Tabernaemontanus verspürt. Er sagt von
ihr: „Am Anfang wo man sie kauet scheint sie ungeschmackt zu sein,
aber bald darauf zwackt sie die Zungen, gleich als stäche man sie
mit den allerkleinsten Dörnern“. Auch die Zellen der Wasserlinsen
sind vollgepfropft von den spitzen Nadelbüscheln des oxalsauren
Kalkes.Nur Wolffia macht auch in dieser Hinsicht eine
Ausnahme, indem sie keine Raphiden besitzt. Dagegen
scheint ein brauner Farbstoff in Oberhaut und Grundgewebe wie
die Gerbsäure bei den Hornblattgewächsen bei ihr zu wirken; denn
nach Stahls Versuchen verschmähen auch die Wasserschnecken die
gerbstoffreichen Wasserpflanzen wie die Hornblattgewächse, ferner die,
gleichzeitig durch Kalkinkrustation geschützten Laichkräuter
und andere (Hippuris, Hydrocharis, Trapa etc.), sowie sie die durch
ätherische Öle und Bitterstoffe (Bitterklee und Kalmus) oder
Verkieselung der Zellwände geschützten Gräser etc. verschonen.
Die eigentümliche Symbiose verschiedener Wasserlinsen mit
Nostocaceen (Hydrocytium), die der regelmässigen Symbiose
von Anabaena in den Azollablättern und des Nostoc bei Gunnera und
Cycadeen sowie bei gewissen Lebermoosen analog ist, ist gleichfalls
als Schutzmittel für die Wasserlinsen gedeutet worden. Trotzdem
haben auch die Wasserlinsen ihre Feinde aufzuweisen aus anderen
Abteilungen der Lebewelt, Pilzen (Chytridiaceen) und Insekten. Eine
Fliege Hydrellia albilabris Mg. höhlt die Wasserlinsen aus trotz
ihrer Raphiden und Nostockolonien.
[S. 106]
Bei den Wasserlinsen drängt sich uns am ersten die Frage auf nach
dem Verbleiben der Schwimmpflanzen im Winter. Im Herbste noch
sehen wir unsere Teiche von einem dichten scheinbar immergrünen
Lemnateppich bedeckt, im Frühjahr ist aber keine Spur mehr davon zu
sehen, die Oberfläche ist rein und klar. Wo ist die dichte grüne Decke
hingekommen und wo kommt sie in der wärmeren Jahreszeit eben so schnell
wieder her?
Während die Hornkrautarten, die Wasserpest und einige
andere völlig submerse oder an der Oberfläche blühende Pflanzen im
Wasser ganz überwintern, sinken andere zu Boden und
bilden besondere Winterknospen (Hibernacula), die
sich im Frühjahr loslösen (die übrige Pflanze stirbt ab) und an die
Oberfläche schwimmen. Zu ihnen gehören die Wasserschlaucharten
(Utricularia), die Wasserfeder (Hottonia),
die Blasenpflanze (Aldrovandia), welche jedoch in
ihrer südlichen Heimat überwintert. Noch andere überwintern durch
ihre Rhizome, wie der Kalmus, die Wasserrosen,
der Knöterich und das schwimmende Laichkraut.
Winterknospen bilden noch der Froschbiss (Hydrocharis
morsus ranae), die Krebsschere (Stratiotes aloides),
die quirlblättrige Hydrille (Hydrilla verticillata), das
kleine Laichkraut (Potamogeton pusillus) etc. Bei manchen
Laichkrautarten, z. B. dem krausen Laichkraut (Potamogeton
crispus), wandeln sich einzelne Seitenzweige zu besonderen
Winterästen um, die sich infolge ihrer starren Beschaffenheit leicht
von der Mutterachse loslösen, zu Boden sinken und daselbst überwintern.
Dieses Laichkraut überwintert auch durch sein im Boden befindliches
Rhizom, während wieder andere Laichkrautarten wie Potamogeton
pectinatus L. ebenso wie das Pfeilkraut (Sagittaria
sagittifolia), Froschlöffel (Alisma), Binsen
(Scirpus palustris) u. a. Knollen bilden und im Herbst bis auf
diese absterben. Die Überwinterung unserer Wasserlinsen geschieht
nach der Beschreibung von Hoffmann und Hegelmaier
bei der wurzellosen Wasserlinse (Wolffia arrhiza) bei uns — in
wärmeren Gegenden sind keine besonderen Vorkehrungen nötig — durch
Sprosse, die sich von den Sommersprossen wenig, hauptsächlich durch
massenhafte Anhäufung[S. 107] von Reservestoffen, unterscheiden. Sie sinken
im Herbst infolge dieser Aufspeicherung von Reservestoffen und der
dadurch bedingten Zunahme des spezifischen Gewichtes zu Boden, um im
Frühjahr wieder emporzusteigen. Auch die grosse Teichlinse
(Spirodela) bildet besondere Wintersprosse, welche aber von den
Sommersprossen durch geringere Grösse und fast nierenförmige Gestalt
unterschieden sind, keine Lufthöhlen bilden, einen bis 2 mm
langen Sprossstiel haben, durch den sie sich scharf abgrenzen, und nur
zwei bis drei sehr kurze Wurzeln treiben. Die Spaltöffnungen bleiben
bei den Wintersprossen, so lange sie auf dem Boden liegen, geschlossen.
Im Frühjahr sprossen aus ihnen die Sommersprossen hervor, brauchen die
Reservestoffe auf und steigen, indem sie Lufthöhlen bilden, an die
Oberfläche. Die Ausbildung der Wintersprosse geschieht hier auch im
Zimmer. Die eigentlichen Lemnaarten: die gemeine Wasserlinse und
die untergetauchte Wasserlinse (L. trisulca) können einen
hohen Kältegrad vertragen, bilden daher keine besonderen Wintersprosse.
Die überwinternden Sprosse trennen sich nur bei Abschluss der
Vegetationsperiode meist in jugendlichem Zustand von ihren verwesenden
Muttersprossen los und harren so den Winter aus, Lemna minor
verbleibt dabei am Wasserspiegel so lange er nicht zufriert.
Zu den prächtigsten Bewohnern unserer Seen, Teiche und Flüsse gehören
die Seelilien oder Teichrosen, die Nymphaeaceen, eine Familie
der Wassergewächse, welche in einer früheren geologischen Epoche
auf dem Höhepunkt ihrer Entwickelung stand. Unsere wenigen jetzt
lebenden europäischen Arten geben uns nur noch ein mattes Bild von der
bunten, prächtigen Nymphaeaceenflora der Braunkohlenzeit. Der Graf
von Saporta[38] sagt darüber: „Man muss nach Ägypten, Nubien,
an die Gewässer von Senegambien und die überschwemmten Savannen von
Guyana oder an die Lagunen von Indien und China gehen, um auch dann
noch abgeschwächte Beispiele von dem zu finden, was in Europa in der
oligocänen Zeit die Seelilien waren. Nicht allein Nelumbium Buchii
Ett. vom Monte Promina und die Fragmente von Wurzelstöcken,
welche Heer auf der Insel Wight beobachtete, bezeugen die[S. 108]
Gegenwart von europäischen oligocänen Lotosblumen. Die eigentlichen
Nymphaeen (Nymphaea parvula Sap., N. Charpentieri Mr.)
beweisen nicht allein die Existenz von Pflanzen, doppelt so gross als
unsere weisse Seelilie (N. alba); es gab auch in dem damaligen
Europa Gattungen oder Sektionen von Gattungen, die heute ausgestorben
sind, deren Charaktere wir nur in sehr unvollkommener Weise
analysieren können, die sich aber hinlänglich von unseren heutigen
Arten unterscheiden, um uns glauben zu lassen, dass ihre Blumen uns
überraschen und unsere Bewunderung erregen würden, wenn es uns möglich
wäre, sie zu betrachten. Der erste dieser tertiären Typen ist in den
Gipsen von Aix vertreten (Nymphaea gypsorum Sap.), ein anderer
in Saint-Zacharie (N. polyrrhiza Sap.), ein dritter, wie es
scheint, in dem Aquitan von Manosque (N. calophylla Sap.).
Ein Bruchstück seiner Früchte mit Lappen von Blumenblättern umgeben,
beweist, dass er gefüllte Blumen hatte, die wenigstens doppelt so
gross als diejenigen unserer heutigen Seelilien und nach einem ganz
anderen Plan konstruiert waren.....“. In unseren Gewässern finden
sich gegenwärtig nur noch die beiden Gattungen der gelben und
weissen Seerose (Nuphar und Nymphaea), von denen
die erstere fünf grosse gelbe Kelchblätter und zahlreiche winzige zu
Nektarien umgestaltete Blumenblätter besitzt, während die letztere
weisse nektarienlose Blumenblätter und grüne Kelchblätter hat. Von den
gelben Teichrosen sind bei uns zwei Arten (Nuphar luteum und
N. pumilum), von den weissen gleichfalls zwei Arten (Nymphaea
candida und N. alba) mit verschiedenen Spielarten und
Bastarden bekannt. Man kennt im Ganzen gegen 52–53 lebende Arten der
Nymphaeaceen, die sehr verschiedene biologische Anpassungen zeigen.
Die (zwei) Arten von Nelumbo (Unterfamilie Nelumbonoideae) strecken
ihre schildförmigen Blätter hoch über das Wasser empor und reifen auch
die Früchte über Wasser, ihre Blüten sind gelblich oder rosenfarben.
In der Unterfamilie Cabomboideae haben dagegen die (vier) Arten von
Cabomba schildförmige, unten ausgerandete Schwimmblätter
und vielteilige untergetauchte Blätter von ähnlicher Gestalt und
Zerteilung wie unser gemeiner Wasserhahnenfuss. Bei der[S. 109] Gattung
Brasenia (eine Art, Brasenia purpurea) sind Schwimmblätter
und Wasserblätter schildförmig elliptisch. Die dritte Unterfamilie
der Nymphaeoideae umfasst 1) die Barclayeae mit der Gattung
Barclaya (drei Arten) mit fünfblättrigem Kelche und oberständiger,
walzig-röhrenförmiger Blumenkrone, der innen die zahlreichen
abwärtsgebeugten Staubgefässe eingefügt sind. Die Blätter der B.
longifolia sind länglich, gestielt, denen unseres Wasserknöterichs
nicht unähnlich; 2) die Nuphareae mit der Gattung Nuphar (sieben
Arten); 3) die Tetrasepaleae mit den Gattungen Nymphaea (etwa 32
Arten), Euryale (eine Art) und Victoria (zwei bis drei Arten). Die
Gattung Victoria, deren bekannteste Art Victoria regia mit ihren
riesigen Schwimmblättern von 1 m Durchmesser und Blüten von 2–4
dm Durchmesser in den ruhigeren Nebenflüssen des Amazonenstromes
oft meilenweit die Wasserfläche bedeckt, und die Gattung Euryale haben
bestachelte Stengel und Blätter. In der Gattung Nymphaea selbst sind
die Arten der Untergattungen Lotos (z. B. die afrikanische Nymphaea
Lotus, die sich bereits auf alten ägyptischen Denkmälern vielfach
dargestellt findet und mit der N. thermalis der ungarischen
Thermen identisch ist) und Hydrocallis (mit grüngelblichen
Blüten) mit glattem Blütenstaub versehen und blühen nur des Nachts.
Die Nymphaea Amazonum öffnet ihre Blüte nur etwa 20–30 Minuten
in den ersten Morgenstunden. Die übrigen Arten der Untergattungen
Xanthantha (mit gelber Blüte), Castalia (mit fünf Arten, worunter
unsere einheimischen), Brachyceros und Anecephya sind Tagblüher,
meist mit kurzstacheligen oder warzigen Pollenkörnern versehen. Die
blaue Lotos der Ägypter Nymphaea coerulea hat blaue, die den
Ostindiern heilige Nymphaea stellata und die australische N.
gigantea haben blau rosig und weiss aussehende Blüten.
Die Blütenbiologie unserer einheimischen Seerosen ist bereits mehrfach
studiert worden. Bei unserer weissen Teichrose (Nymphaea alba)
öffnet sich die Blüte des Morgens gegen 7 Uhr und schliesst sich
nachmittags gegen 4 Uhr, dabei birgt sie ihre zarten Blüten (wohl gegen
die Kälte der Nachtluft) unter Wasser. Linné (Disquis. de sexu
plantarum 1760) sagt:
[S. 110]
Nymphaea alba quotidie mane ex aqua tollitur, floremque dilatat adeo ut
meridiano tempore tres omnino pollices pedunculo aquam superemineat.
Sub vesperam penitus clausa et contecta demergitur. Circa horam enim
quartam post meridiem contrahit florem, agitque sub aqua omnem noctem,
quod nescio an cuiquam per bis mille annos notatum sit, id est inde
a Theophrasti aevo, qui hoc observavit in Nymphaea Loto.... Scripsit
autem Theophrastus, hist. plant. IV, 10. de Loto eaquae sequuntur:
„In Euphrate caput floresque mergi referunt, atque descendere usque
in medias noctes: tantumque abire in altum, ut ne de missa quidem
manu capere sit: diluculo dein redire et ad diem magis. Sole oriente
jam extra undas emergere floremque patefacere, quo patefacto amplius
insurgere, ut plane ab aqua absit alte“. — Idem prorsus mos est
nostrae Nymphaeae albae.
Die Seerosen sind zoidiophil, der Bestäubung durch Tiere angepasst,
da sie aber, wie wir oben hervorhoben, die Höhe ihrer Entwickelung
bereits in einer früheren geologischen Epoche erreicht hatten, wird
man heutzutage nicht mehr mit Sicherheit behaupten können, welcher
Abteilung der Tiere sie ihre Bestäubungsverhältnisse angepasst haben.
Für einige Arten scheinen die geeigneten Bestäuber ausgestorben zu
sein, so dass dieselben jetzt auf Selbstbestäubung angewiesen sind.
So soll nach Caspary[39] in der Unterabteilung Hydrocallis der
Gattung Nymphaea die Bestäubung vor Aufbruch der Blüten stets mit
eigenem Blütenstaub erfolgen, wobei 10000 bis 30000 Samen, die aufs
beste keimen, gebildet werden. Auch Euryale ferox befruchtet
sich stets selbst, oft bei ganz geschlossener, ja völlig unter Wasser
bleibender Blüte. Gegenwärtig scheinen unsere Seerosen
auf die Käfer und Fliegen angewiesen zu sein. Nach Delpinos
Vermutung werden Nymphaea alba und Victoria regia von
Cetonien (Rosenkäfern) und Glaphyriden befruchtet. Bei Nuphar sondert
die Unterseite der reduzierten Blumenblätter Honig ab, während die
Kelchblätter durch Ausdehnung ihrer Fläche und ihre gelbe Farbe die
Rolle der Blumenblätter übernommen haben. Unsere Nuphar wie Nymphaea
sind proterogynisch und ist z. B. bei Nymphaea die Narbe nur am[S. 111] ersten
Tage des 3–7 Tage währenden Blühens empfängnisfähig, so dass Bestäubung
ohne Insekten nicht erfolgt.[40]
Sprengel fand bei der gelben Teichrose Blumenkäfer in den
Blüten und Hermann Müller sah dieselben ausser von Meligethes
von Schilfkäfern (Donacia dentata) und Fliegen (Onesia
floralis) besucht. Neuerdings hat der amerikanische Biologe
Charles Robertson[41] den Bestäubungsverhältnissen der Seerosen
ein besonderes Interesse zugewandt. Nach ihm ist Nelumbo und
NymphaeaPollenblume, während NupharPollen
und Nektar deren Besuchern darbietet. Jordan giebt
jedoch bei Nymphaea alba vor den introrsen Staubgefässen (nach
innen) gelegene flache Honigdrüsen an.[42]Robertson fand
bei Nelumbo lutea als Hauptbestäuber Hautflügler, besonders
Andreniden (Halictus) und Schwebfliegen, bei Nuphar
advena sowohl im Staate Illinois (im August) wie in Florida
(im Februar) Halictus pectoralis (Andrenide), Helophilus
divisus (Schwebfliege), Donacia piscatrix (Schilfkäfer),
Trelease fand in Madison gleichfalls Halictus pectoralis
und Donacia piscatrix. Bei Nymphaea tuberosa fand
Robertson acht Andreniden, zwei Syrphiden, eine Bombylide,
besonders häufig auch hier Halictus pectoralis, letztere
auch bei Nymphaea alba. Piccioli fand bei dieser
gleichfalls Donacia, so dass als besonders regelmässige
Bestäubungsvermittler der Seerosen Schilfkäfer (Donacia
dentata in Deutschland, D. piscatrix in Nordamerika),
Halictusarten (H. pectoralis in Nordamerika) und
Fliegen bisher beobachtet worden sind.
Die Blüten der Seerosen scheinen nicht selten eine Todesfalle für
Insekten zu sein, so fand Delpino tote Insekten bei Nymphaea
alba und nahm an, dass sie durch die durch einen starken Geruch
bemerklichen Ausdünstungen der Pflanze getötet worden seien.
Planchon (Flore des serres et des jardins 1850) denkt an eine
Anhäufung von Kohlensäure in dem Blütenkessel. Robertson fand
zuweilen in den Nelumboblüten tote Hummeln (Bombus virginicus)
und Fliegen, die durch die Petala eingeschlossen waren und in
ihrem Gefängnis erstickt zu sein schienen. Dagegen fand er in dem
Blütenbassin von Nymphaea tuberosa Halictus occidentalis[S. 112]
ertränkt. Auch A. Bacon (Bull. Torr. Bot. Club V, 51) berichtet,
dass er in den Blüten der Nymphaea odorata gefangene und
getötete Insekten eingeschlossen gefunden habe.
Fig. 13.
Luftgänge und Sternhaare aus dem Stengelquerschnitt einer Teichrose.
(Nach Engler.)Fig. 14. a Same von Villarsia nymphaeoides — b Frucht von Bidens
tripartitus — c Frucht von Trapa natans — d Blüte von
Scirpus lacustris — e von Scirpus palustris mit
widerhakigen Perigon-Borsten — f Frucht von Typha latifolia.
a–e der Verbreitung durch Tiere, f der Windverbreitung
angepasst; c zugleich „Ankerklette“. (Nach Hildebrand und
Huth.)
Die von Lufthöhlen durchzogenen Blatt- und Blütenstiele (Fig. 13) und
Wurzelstöcke sind bei unseren Teichrosen (überhaupt bei Victoria,
Euryale, Nymphaea, Nuphar, bei letzterer Gattung nur die Wurzelstöcke
nicht) im Innern mit vielästigen Sternhaaren mit körniger Oberfläche
versehen, welche in die Zwischenzellräume hineinragen. Man bemerkt
dieselben schon mit blossem Auge deutlich, wenn man ein kurzes Stück
aus dem Blattstiel ausschneidet und durch dasselbe hindurch sieht.
Nach den Untersuchungen Stahls sind solche Feilenhaare
ein Schutzmittel gegen Schneckenfrass und die Zerstörungen
anderer omnivoren Tiere. Nun scheinen Schnecken in der That gerne
das Innere grüner Stengel auszufressen. (In meinem Garten wurden die
Stengel der Kaiserkrone mehrere Jahre hindurch durch Nacktschnecken
abgefressen, die den Stengel dicht an der Erde durchfressen und
dann aushöhlen, während die Versuche, an den Blättern zu fressen,
wohl[S. 113] wegen des hohen Raphidengehaltes derselben, bald aufgegeben
wurden.) Bei anderen Wasserpflanzen, welche einen solchen Schutz
ihres Stengelmarkes nicht haben, werden die Stengel nicht selten
ausgehöhlt und zu Grunde gerichtet. So leben von dem Mark des
Wasserliesches (Butomus umbellatus) die Larve einer
Fliege (Agromyza confinis), in dem des Igelkolbens etc.
die Larve von Faltern (Orthothaelia sparganella, Nonagria
Sparganii) und fressen die Stengel aus, wenn nicht Blesshühner,
Rohrdommeln und Störche, welche Raupen und Puppen sehr geschickt
herausholen, oder Schlupfwespen (Ichneumon divisorius) Einhalt
thun. Abgesehen von einer Blattlaus (Aphis Nymphaeae, die
sich auch an verschiedenen anderen Wasserpflanzen, dem Froschbiss,
Froschlöffel, Wasserliesch etc. einfindet), einigen Käfern (Donacia
crassipes, Donacia Menyanthidis, Galeruca Nymphaeae)
und Falterraupen (Nymphula nymphaealis verpuppt sich unter
einem der oberen Blattfläche angeleimten Blattstückchen, der Falter
ist im Mai und Juni in der Nähe der Teichrosen häufig) haben unsere
Teichrosen wohl wenig Feinde oder nutzlose Gäste (Bryozoën etc.)
aus der Tierwelt aufzuweisen. Auch gegen Pilzschmarotzer scheinen
sie gefeit zu sein. Ein durch dieses Vorkommen bemerkenswerter
Rostpilz (Aecidium nymphaeoides)[II] ist sehr wenig verbreitet.
Die Verbreitungsmittel der Wassergewächse sind nicht selten
derartige, dass sowohl eine Weiterverbreitung innerhalb desselben
Gewässers, als auch eine weitere Verbreitung von Gewässer zu Gewässer
möglich ist. In ersterer Hinsicht ist es von Vorteil, wenn die
Samen oder Früchte zu schwimmen vermögen (wobei sie durch
Wasserströmung und Wind verbreitet werden), später aber
ein grösseres spezifisches Gewicht als das Wasser
erlangen und zu Boden sinken[S. 114] (wenn sie nicht wurzellosen
Pflanzen zugehören). Bei der Verbreitung von Gewässer zu Gewässer
kommen, wenn wir von Überschwemmungen absehen, hauptsächlich Wind und
Tiere als Verbreitungsfaktoren in Betracht. Bei den Schwimmgewächsen
spielen besonders die letzteren eine hervorragende Rolle und unter
ihnen besonders die geflügelten Wassertiere, Vögel und Wasserkäfer.
Bei unseren weissen Teichrosen sind die Samen mit einem
Samenmantel versehen, so dass sie nach dem Platzen der Frucht an
der Wasserfläche, durch die zwischen ihnen und dem Samenmantel
enthaltene Luft gehalten, umherschwimmen können. Dieser
Schwimmmantel umgiebt den Samen lose als weissliche Hülle.
Zunächst sind nach dem Auseinanderfallen der Fruchtwände die Samen
zu einem schleimigen Klumpen vereinigt, der sich aber schliesslich
auflöst, so dass die Samen sich frei umherbewegen. Zuletzt vergeht
auch der Samenmantel, die Luftblasen entweichen und der Same fällt
vermöge seiner Schwere zu Boden[43]. Anders verhält sich die
gelbe Teichrose. Bei ihr findet sich die Vorrichtung zur
Wasserverbreitung nicht an den Samen, sondern sie liegt in einer
besonderen Konstruktion der Fruchtwände. Zur Reifezeit löst sich
die Frucht gleichfalls von ihrem Stiele ab, aber die Samen werden dabei
nicht sogleich frei, sondern es geschieht, wie es Hildebrand
schildert, etwas dem Verfahren ähnliches, welches man einschlägt, wenn
man eine Apfelsine in einzelne halbmondförmige Teile zerlegt. Von der
äusseren Fruchtwand löst sich nämlich nur die äussere grüne Schicht
los, während die innere mit den Scheidewänden der Frucht in Verbindung
bleibt. Die Scheidewände spalten sich dann bald von aussen beginnend
in je zwei Lamellen, wodurch jene halbmondförmigen Scheiben entstehen,
gebildet aus einer festen Aussenhaut, welche die zahlreichen schweren
Samen in einen Schleim eingebettet umschliesst. Diese Scheiben sinken
nicht unter, weil in dem Schleime ihres Inneren zahlreiche Luftblasen
entstanden sind. Erst später, wenn die Scheiben länger umhergeschwommen
sind, löst sich die äussere Hülle auf, die Luftblasen entweichen
aus dem Schleime und die Samen werden auf[S. 115] den Grund des Wassers
ausgesäet. Der Samenmantel der Nymphaea fehlt ganz. Noll hat darauf
hingewiesen, dass die Wasserhühner die eigentlichen Verbreiter
der Samen der Teichrosen von Teich zu Teich sind. Die Früchte sind ihre
Lieblingsspeise. Bei dem Verzehren derselben bleiben die klebrigen
Samen den Vögeln an Haaren und Schnäbeln haften und werden, wie
Noll fand, von ihnen verschleppt. Durch Wasservögel werden
auch unsere Wasserlinsen weiter verbreitet. Nicht im Wasser
schwimmende Samen und Früchte (Fig. 14) kletten sich den[S. 116] Wasservögeln
häufig an und die Verbreitung vieler Wasserpflanzen ist von
den Wanderungen und der Zugrichtung der Wasservögel abhängig.
So verdanken Villarsia- und Limnanthemumarten den
Wasservögeln ihre weite Verbreitung. Leersia oryzoides ist nach
Ebelings Vermutung durch Steissfüsse, Enten,
Wasserhühner aus Südeuropa bis zur norddeutschen Küste
verbreitet worden. Unsere WasserbinsenScirpus lacustris
und Scirpus palustris, sind mit einem aus sechs Borsten
bestehenden Perigon mit rückwärts gerichteten Stacheln versehen.
Ähnliche Klettvorrichtungen sind bei Rhynchospora etc., ferner
vom Zweizahn (Bidens) bekannt. Bidens cernuus ist
— jedenfalls durch Vögel verschleppt — aus Europa zu Anfang dieses
Jahrhunderts nach Nordamerika gekommen. B. pilosus wanderte
aus Nordamerika nach Teneriffa und Neuseeland, B. leucanthus
aus Mittelamerika nach Madeira und der Insel Mauritius und der in
Nordamerika heimische B. bipinnatus ist jetzt in Tirol eine
wahre Landplage geworden. Die mit gefiedertem Flugapparat versehenen
Samen vom Rohrkolben und andere den Wind als Transportmittel
benutzende Wasserpflanzen haben gleichfalls noch das Vermögen eine Zeit
lang an der Oberfläche zu schwimmen. — Bei einer Anzahl tropischer
Wassergewächse keimen die Samen erst, nachdem sie eine gewisse Zeit
ausgetrocknet waren[45]; es kann dies mit ihrem Vorkommen in Gewässern
zusammenhängen, die regelmässig einen Teil des Jahres austrocknen.
Möglich wäre es aber auch, dass diese, von einem vorherigen Austrocknen
bedingte Keimfähigkeit der Samen in Zusammenhang stände mit dem
Transport der Samen von Gewässer zu Gewässer durch das Trockene
hindurch.
[II] Nachdem das Obenstehende niedergeschrieben, ist die
Zugehörigkeit des Aecidium nymphaeoides DC. zu Puccinia
Scirpi DC. auf Scirpus lacustris sowie die der Aecidien auf
Hippuris und Sium latifolium zu Uromyces lineolatus
Desm. auf Scirpus maritimus erwiesen worden. Immerhin
bleibt noch eine ganze Anzahl von Rostpilzen auf den Binsen- und
Schilfpflanzen übrig, deren Zugehörigkeit zu denen anderer Wasser- und
Sumpfpflanzen noch zu ermitteln ist — eine der vielen Aufgaben, welche
an den Süsswasser-Stationen, wie sie Dr. O. Zacharias ins Leben
gerufen hat (vergl. das 15. Kapitel dieses Werkes), ins Auge gefasst
werden können.
Eine Schwimmpflanze, welche nicht nur, wie die Seerosen, die Zeit
ihrer hauptsächlichsten Entfaltung an der Erdoberfläche hinter sich
hat, vielmehr jetzt im Aussterben begriffen zu sein scheint,
ist die Wassernuss (Trapa natans). Nathorst und
Carlsen[46], welche die Verbreitung dieser Pflanze in Schweden
untersucht haben, fanden ihre Früchte in vielen Seen Schwedens[S. 117]
verbreitet, in denen die Pflanze jetzt gar nicht mehr wächst oder
jetzt sehr selten geworden ist, in dem Schlamm des Bodens. Die Pflanze
scheint gewöhnlich mit der weissen Seerose gelebt zu haben und
es ist daher zweckmässig, in der Nähe dieser auch die Seen anderer
Länder nach den Trapafrüchten, die zudem einen Reichtum von Varietäten
zur Schau tragen, zu durchsuchen.
Nach Huth[44] sind die hakigen Früchte als „Ankerkletten“ zu
betrachten (welche ein Festhaken des Keimlings am Boden bewirken) — im
Gegensatz zu den Verschleppungskletten des Zweizahns (Bidens)
etc.
Die submersen Blätter, welche sich neben den Schwimmblättern
finden, besitzen eine Reihe von Eigentümlichkeiten, welche anderen
submersen Blättern abgehen. Die obersten von ihnen besitzen sowohl
Luftspalten als Wasserspalten von charakteristischer Anordnung.
Luft- und Wasserspalten sind sonst bei Wasserpflanzen sehr selten.
Sie finden sich nach De Bary noch auf den Samenlappen von
Batrachium, den Laubblättern von Callitricheen, Hippuris, Hottonia.
Die sehr feinfiedrig geteilten grünen submersen Blattorgane sind
Wasserwurzeln, welche gewissermassen an die grünen von Fritz
Müller beobachteten Luftwurzeln anderer Pflanzen erinnern. Eine
eingehendere Darstellung dieser Verhältnisse wie auch der Keimung hat
Wittrock[47] gegeben. — In Deutschland findet sich die (mit
einer anderen Wasserpflanze Isnardia und den landlebigen Weidenröschen
und Circaeaarten) zu den Nachtkerzengewächsen (Onagraceae)
gehörige Wassernuss sehr zerstreut. Orte ihres Vorkommens sind ausser
den seenreichen Gegenden Norddeutschlands z. B. der Niederrhein, Seen
und Teiche um Dessau, bei Plothen und Drebra bei Gera.
Die Arten des Wasserhahnenfusses (Batrachium), deren
systematische Bearbeitung z. B. bei J. Freyn[48] nachzusehen
ist, zeigen merkwürdige Übergänge von dem Landleben und amphibischen
Leben zu echten Schwimmpflanzen und typisch submersen Arten. Von
den acht in Nord- und Mitteldeutschland vertretenen Arten ist nur
Batrachium hederaceum eine echte Schwimmpflanze, die lauter
eigentliche Schwimmblätter besitzt. Ihm steht eine in[S. 118] Sizilien,
Asturien, England etc. vorkommende Art B. caenosum nahe.
Die submersen Arten bilden wie die Tausendblattarten sehr
ausgedehnte Wiesen unter Wasser oder weithin flutende Rasen in unseren
Flüssen. Am häufigsten sind B. divaricatum in stehenden
Gewässern mit nur zerschlitzten (untergetauchten), flächenförmig
ausgebreiteten, kreisrunden Blättern, die meist einmal dreiteilig sind
mit weiterhin wiederholt gabelspaltigen Zipfeln, B. fluitans,
welches vorwiegend in Flüssen und Bächen wächst, dreiteilig
vielspaltige, sehr langhin ausgedehnte Wasserblätter, nur selten
aber nierenförmige oder geschlitzte Schwimmblätter besitzt und die
gemeinste und vielgestaltigste, allen möglichen Lebensverhältnissen
angepasste Art B. aquatile. Letztere erzeugt im Wasser nur
zuweilen Schwimmblätter und zwar zur Blütezeit. Askenasy[6] ist
durch Versuche zu der Ansicht gelangt, dass ein typisches Schwimmblatt
sich bilde, wenn das einer Blüte gegenüber stehende Blatt in einer
bestimmten Periode in die Luft emporgehoben wird. Besonders merkwürdig
sind die Luftformen der Hahnenfussarten in kleinen Tümpeln etc.,
die sonst sehr dicht mit Wasserpflanzen besetzt sind. Sie haben
ausser submersen zerschlitzten Blättern und echten Schwimmblättern
typische Luftblätter (mit den Spaltöffnungen an der unteren Seite)
etc. Eigentliche Landformen kommen beim völligen Austrocknen der
Sümpfe bei fast allen drei Arten vor, sie haben aber bei B.
divaricatum nur die typischen zerschlitzten Blätter. Der Übergang
der einen in die andere biologische Form erfolgt meist schnell
bei Eintritt anderer Lebensverhältnisse. — Ähnliche amphibische
Wasserpflanzen sind z. B. der Tannenwedel (Hippuris),
die Tännelarten Elatine etc. — Die Blüteneinrichtung des
gemeinen WasserhahnenfussesB. aquatile ist von
Herm. Müller (l. c.) beschrieben worden. Die Honigblumen,
zu deren Saftdrüsen die als Saftmal dienende gelbgefärbte Basis
der Blumenblätter führt, werden besonders von Schwebfliegen
(Eristalisarten, Helophrilus florens, Chrysogaster
viduata), kleineren und grösseren Musciden, Bienen (Apis
mellifica), Hummeln (Bombus terrestris) bestäubt. Ein von
H. Müller beobachteter Käfer (Helodes Phellandri) frisst
die Blumenblätter und Staubgefässe. Bei hohem Wasserstand bleiben die
Blüten[S. 119] geschlossen unter Wasser und befruchten sich selbst. Ob solche
kleistogamische Blüten, wie sie sich bei dieser mit dem Landhahnenfuss
(Ranunculus) doch nahe verwandten und wohl von diesem abstammenden
Gattung von Batrachium finden, auch sonst bei echten Hydrophyten
vorkommen, oder ob hier überall die früher erörterten Anpassungen an
eine typische Wasserbefruchtung eingetreten sind, bedarf noch eines
besondern Studiums. H. Müller war nach brieflichen Mitteilungen, welche
er an mich richtete, geneigt, eine weitere Verbreitung kleistogamischen
Blühens auch bei Wasserpflanzen anzunehmen. Das Vorkommen einer gross-
und kleinblütigen Form, wie es bei vielen Landpflanzen (Stellaria
graminea, Cerastium arvense, Thymian, Gundermann etc.) bekannt ist
— in Verbindung mit einer Trennung in Zwitterblüten und weibliche
Blüten, ist z. B. von Kirchner beobachtet worden, welcher unter
den normalen Stöcken (Blüte 20–27 mm im Durchmesser) solche
mit Blüten von 3–4 mm Durchmesser und wenigen Staubgefässen
fand. Der letztgenannte Forscher hat auch die Blüteneinrichtung des
B. divaricatum näher beschrieben. Beyer[49] hat die spontanen
Bewegungen der Staubgefässe des gemeinen Wasserhahnenfusses, dessen
Blüte „auf Unterbestäubung eingerichtet ist“, in ihrem gesetzlichen
Ablauf näher studiert. (Bei Ranunculus auricomus hatte schon
Konrad Christian Sprengel die Staminalbewegung beobachtet.)
Von amphibischen Schwimmpflanzen sei hier nur noch des
Wasserknöterichs (Polygonum amphibium) mit einigen Worten
Erwähnung gethan. Der Gegensatz einer besondern Landform und Wasserform
tritt bei ihm besonders hervor. Wächst Polygonum amphibium
im Wasser, so bildet es die Form natans mit langgestielten,
breitlanzettlichen, am Grund herzförmigen Schwimmblättern von
lederartiger Konsistenz der Blätter und Stengel. Bei der Landform
P. terrestris sind dagegen die dem Wurzelstock entspringenden
Stengel aufrecht von unten auf mit schmallanzettlichen, festsitzenden
Blättern besetzt, deren Fläche nicht glatt, sondern runzlig ist.
Während bei der Wasserform der Luftkanal stärker entwickelt und
dadurch das spezifische Gewicht[S. 120] des Stengels herabgesetzt ist,
bildet die Landform zur Erreichung der für sie unentbehrlichen
Biegungsfestigkeit ausserhalb des Bastgewebes (Phloëms)
besondere mechanische Zellen, ein Skelett, im Stengel aus[50].
Das ganze Hautgewebe hat zum Zweck der Transpiration und Durchlüftung
besondere Umgestaltungen erfahren. Besonders beachtenswert ist aber,
dass die Landform besondere Schutzmittel gegen ungeflügelte
aufkriechende Tiere hat. Kerner[51] hebt es besonders hervor,
dass den Wasserpflanzen, wie Alisma, Butomus, Sagittaria,
Hottonia, Utricularia, Villarsia, Nuphar, Nymphaea, Hydrocharis,
Stratiotes, welche durch das umgebende Medium vor aufkriechenden
Tieren (Ameisen, Raupen etc.) geschützt sind, die besonderen
Schutzmittel der Blüte fehlen, welche bei den Landpflanzen in so
reichem Masse zur Entwickelung gelangt sind. „Sehr lehrreich in
dieser Beziehung“, sagt Kerner, „ist das Verhalten des Polygonum
amphibium. Die schön rosenroten Blüten dieser Pflanze sind zu
kleinen Cymen vereinigt und diese bilden eine dichte, cylindrische,
ährenförmige, sehr reiche Inflorescenz von 2.5 bis 3.5 cm
Länge und 1 bis 1.2 cm Breite. Die Blätter des Perianthiums
sind fast bis zum Grunde getrennt; der Fruchtknoten ist von einem
fleischigen, roten, fünflappigen, nektarabsondernden Becher umgeben
und der Grund der Blüte auch reichlich mit Nektar erfüllt. Die mit der
Basis des Perianthiums verwachsenen Pollenblätter sind sehr kurz und
die Pollenbehälter bleiben in der Tiefe der Blüte geborgen; die zwei
Griffel des Gynaeceums sind dagegen sehr lang und ragen sogar über die
Blätter des Perianthiums hinaus. Während der Anthese beträgt die Länge
des Perianthiums 4 mm, die obere Weitung kaum 3 mm. Da
der Nektar im Blütengrunde durch kein besonderes Gebilde am Perianthium
geschützt ist, so erscheint er selbst kleinen Insekten zugänglich und
wird von diesen auch gern abgeholt. Bei den angegebenen Dimensionen
der Blüte können aber selbst sehr kleine anfliegende Insekten
nicht vermeiden, dass sie beim Abholen des Nektars zuerst an die über
das Perianthium vorstehenden und[S. 121] etwas spreizenden Narben und dann
an die dicht über dem Nektar befindlichen Pollenbehälter streifen,
und da die Blüten proterandrisch sind, so wird selbst durch sehr
kleine anfliegende Insekten, welche mehrere Blüten und Blütenstände
nach einander besuchen, Kreuzung der Blüten (bald Geitonogamie, bald
Xenogamie) veranlasst. Die von unten her kommenden flügellosen,
aufkriechenden kleinen Insekten würden sich aber nicht die Mühe
nehmen, über den obern Rand des Perianthiums an den aus der Apertur
hervorragenden Narben vorbei zum Blütengrunde vorzudringen, sondern
auf dem kürzesten und für sie bequemsten Wege von unten durch die
tiefen, die Perigonzipfel trennenden Spalten sich den Nektar holen.
Sie würden daher auch eine Belegung der Narben nicht veranlassen und
es würde somit der Nektar geopfert, ohne dass zugleich der Vorteil der
Allogamie erreicht wäre. Da zudem bei Polygonum amphibium infolge der
Dichogamie und der gegenseitigen Lage der Blütenteile eine Autogamie
(Selbstbestäubung) unmöglich ist, würde durch den Besuch solcher
aufkriechenden kleinen Insekten das Entstehen von Früchten überhaupt
gänzlich vereitelt werden. Zu den Blüten der im Wasser wachsenden
Stöcke des Polygonum amphibium können nun sehr kleine ankriechende
Insekten auch nicht kommen. Wie aber dann, wenn das Wasser abgelaufen
ist und Polygonum amphibium aufs Trockene gesetzt wird? — Da ist
es nun sehr merkwürdig, dass sich in solchem Falle besondere
Schutzmittel ausbilden, welche der im Wasser wachsenden Pflanze bisher
fehlten. Es entwickeln sich nämlich dann aus der Epidermis sowohl
der Blätter als der Stengel eine Unzahl horizontal abstehender,
im Mittel 0.7 mm langer Trichomzotten („Drüsenhaare“), die
insbesondere an dem Stengelteile, welcher durch eine Inflorescenz
abgeschlossen ist, so dicht als nur möglich gestellt sind und deren
kugelige Schlusszellen einen klebrigen Stoff secernieren, so dass sich
die Achse, welche die Inflorescenz trägt, ganz schmierig anfühlt.
Jene kleinen, flügellosen aufkriechenden Insekten, welche den Nektar
rauben möchten, ohne dabei den Vorteil einer Kreuzung der Blüten zu
vermitteln, können über diese klebrige Achse nicht emporkommen,[S. 122] sie
würden an derselben wie an Leimspindeln kleben bleiben.“ Die Landform
hat also dasselbe Schutzmittel gegen unberufene Blütenbesucher wie
die Pechnelke (Lychnis viscaria) und viele andere Pflanzen.
Wird der Standort des Polygonum aber wieder einmal unter Wasser
gesetzt, so entwickelt dasselbe Rhizom wieder die Wasserform ohne
diese Trichomzotten, aber mit allen Anpassungen an das Wasserleben.
— Übrigens dürfte Kerner bei den oben beschriebenen Exemplaren nur
die langgrifflige Form des Wasserknöterichs vor sich gehabt haben,
daneben aber eine kurzgrifflige (wie bei der Wasserfeder und dem
Fieberklee) existieren; wenigstens hat Kirchner[52], wie bereits
angedeutet wurde, in der Landform einen neuen Fall des Vorkommens
der Heterostylie entdeckt. Ob bei dem Wasserknöterich eine ähnliche
Blühfolge besteht, wie ich sie für den gemeinen Wiesenknöterich
beschrieben habe[53], hat ebenso besonderes Interesse (weil diese Art
nicht heterostyl ist), wie die Frage, welches der Insektenkreis
der Bestäubungsvermittler der Landform und welches der der Wasserform
ist. Doch ist man zur Beantwortung dieser wie tausend anderer ebenso
einfacher und doch wichtiger Fragen zur Biologie der Wasserpflanzen
bisher nicht gekommen.
Die Luftpflanzen
unserer Gewässer.
Die Luftpflanzen unserer stehenden und fliessenden Gewässer sind
stets im Boden festgewurzelt, haben zumeist auch einen sehr
kräftig entwickelten Wurzelstock, wie z. B. der Kalmus, die
Schwertlilien etc., zeigen im übrigen aber noch deutliche
Anpassungen an das Wasserleben. Wir können zwei Hauptformationen
unterscheiden, die Schilfgewächse und die unter deren Schutz
(gegen Wind und Wasserströmung) gestellten, die Wasserfläche nur wenig
überragenden Sumpfpflanzen. Beginnen wir mit den letzteren,
die sehr verschiedenen Abteilungen, zumeist aber den Monokotyledonen
angehören.
Am tiefsten in die Gewässer steigen von höheren Gewächsen (z. B.
neben der den Kreuzblütlern angehörigen Brunnenkresse, den[S. 123]
Wasserehrenpreisarten etc.) noch einige Doldenpflanzen, wie der
Wassermerk (Sium, Berula), die Sumpfdolde
(Helosciadium), die Rebendolde (Oenanthe fistulosa
und aquatica) und der Wasserschierling (Cicuta
virosa) hinein. Besonders Oenanthe aquatica ist mit röhrigem
Stiel (wie auch der noch durch Gift geschützte Wasserschierling)
und mit haarförmigen, sehr fein zerteilten Wasserblättern versehen
und kann in sehr tiefem Wasser nur die letzteren untergetaucht
ausbilden, ein völlig submerses Wasserleben führend. Auch die
Froschlöffelgewächse (Alismaceen) bilden noch besondere
Wasserblätter aus und führen gewöhnlich ein Wasserluftleben oder
unter besonderen Verhältnissen sogar ein Wasserleben. Nur der
schwimmende Froschlöffel, der von Buchenau, dem Bearbeiter
dieser Pflanzenfamilie, unter die besondere Gattung Elisma
(Elisma natans) gebracht wird — Buchenau beschreibt zehn
Gattungen mit 45–48 Arten von Alismaceen —, ist eine echte
Schwimmpflanze mit Schwimmblättern und submersen Blättern. Die
beiden verbreitetsten Arten unserer Flora sind der gemeine
Froschlöffel (Alisma Plantago) und das Pfeilkraut
(Sagittaria sagittifolia), welche beide mit ihren zierlichen
Blattbüscheln und hübschen Blütenständen die Ufer unserer Teiche und
Flüsse schmücken. Beide bilden in tiefem Wasserstand submerse Formen
mit schmallinealischen Blättern (forma graminifolia), die
von Linné sogar mit einer Vallisnerie verwechselt werden konnten,
auch der hahnenfussblättrige Froschlöffel (Echinodorus
ranunculoides) verhält sich so. Unser Alisma Plantago,
dessen Blütenstand sich mit mathematischer Regelmässigkeit in
die Luft erhebt (nach den Messungen und Abbildungen von Xaver
Pfeifer[54] träte das Verhältnis des goldenen Schnittes besonders
häufig in dem Aufbau dieser Pflanze zutage), besitzt zierliche kleine
Zwitterblüten mit sechs (2 × 3) vor den Kelchblättern stehenden
Staubgefässen, während der Blütenstand des Pfeilkrautes aus
grösseren, rötlichweissen Blüten getrennten Geschlechts besteht,
von denen die oberen männlich, die unteren weiblich mit vielen
Staubgefässen bezügl. Stempeln in spiraliger Anordnung versehen sind.
Beide Pflanzen sind der Bestäubung durch Vermittlung der Insekten[S. 124]
angepasst (sind entomophil). Nach Hermann Müller sind bei Alisma
Plantago die pollenübertragenden Insekten Schwebfliegen
(Eristalis, Syritta, Ascia, Melanostoma, Melithreptus). Bei Elisma
natans kommen nach Hildebrand auch kleistogamische Blüten vor[55].
Die Verbreitung der Samen geschieht durch den Wind (mit teilweiser
Anpassung an die nächstgelegene Verbreitung durch das Wasser), bei der
ostindischen Gattung Limnophyton besitzen die Samen zwischen der
verholzten Innenschale (Endocarpium) und der Aussenschale Lufthöhlen
(Schwimmvorrichtung); Caldesia parnassifolia kommt bei uns
überhaupt nicht zur Fruchtbildung, sondern pflanzt sich nur durch
Brutknospen fort. — Die gleichfalls im Schutz der Schilfpflanzen
wachsende Schlangenwurz (Calla palustris), welche bei uns
zweimal blüht (im Mai und September), wäre besonders in Bezug auf die
Bestäubungsweise zu untersuchen. Ob diese durch Raphiden geschützte
Pflanze durch Schnecken oder Insekten befruchtet wird oder, wie Kerner
u. A.[56] vermuten, einer Befruchtung durch Regen und Tau angepasst
ist, ist bisher nicht festgestellt worden (vgl. die Bemerkung bei
Lemna). Sie hat rote Beerenfrüchte (Verbreitung durch Vögel!).
Fig. 15.
Schematische Querschnitte durch Blätter der Schilfgewächse. a
Schwertlilie — b Kalmus — c Ästiger Igelskolben — d
Einfacher Igelskolben — e Wasserveilchen oder Blumenbinse — f
Breitblättriger Lieschkolben — g Schmalblättriger Lieschkolben — h
Waldbinse.
Ein hervorragendes Interesse beanspruchen noch die
Schilfgewächse, welche neben dem Röhricht der Schachtelhalme
und Binsen der Wasserlandschaft ihr eigentümliches Gepräge
verleihen. Sie gehören alle den Monokotyledonen an, und da zudem
ihre Arten bereits in den Tertiärformationen auftreten, ist es
wahrscheinlich, dass sie eine der ältesten Anpassungen an das
Luftwasserleben darstellen. In den Luftströmungen und besonders
gegen die Bewegung des Wassers, wie sie besonders bei heftigeren
meteorologischen Katastrophen (Wolkenbrüche, Überschwemmungen etc.),
die in der Vorzeit noch weit mächtiger und häufiger eingetreten
sein mögen als jetzt, vorkommen, sind die schmallinealischen
(schwertförmigen), sehr elastischen, leichten und dabei biegungsfesten
Blätter der Schilfgewächse (die Wind und Regen eine minimale
Angriffswaffe darbieten) besonders widerstandsfähig. Ihr ganzer Aufbau
(Fig. 15), das von Lufthöhlen durchzogene Blatt (dem meist auch der
Blütenschaft gleichgestaltet ist) mit seiner verdickten Mittelrippe,[S. 125]
den oft parallel gestellten Querwänden, die wie Strebepfeiler wirken
(bei Iris, Typha, Scirpus silvaticus), deuten darauf hin. Mächtige,
weit verbreitete Rhizome, welche am Grunde festgewurzelt sind,
sichern diesen Pflanzen eine dauernde Existenz durch Sommer und
Winter und eine rasche Ausbreitung über ein grosses Areal. Besondere
Vorkehrungen[57] schützen die jugendlichen Teile gegen Verletzungen
durch die Bodenteile (tütenförmige Niederblätter bei Glyceria etc.).
So schildert Warming[58] die besondere Entwickelung der Rhizome
von Phragmites communis. Die neuen, am Grunde der Muttersprosse
entspringenden Sprosse senken sich tief in die Erde, ehe sie sich
umbiegen und nach oben wachsen. Das ganze unterirdische Stengelsystem
wird so immer weiter in die Erde gebracht, bis es eine gewisse
Normaltiefe hat, und es entsteht in wenigen Monaten ein sehr reich und
weit verzweigtes Rhizom. — In welcher Weise die von der mittleren
Windrichtung beeinflusste Verwachsung der Teiche, Seen und Flüsse durch
solch üppige Rhizombildung vor sich geht und durch sie ein Weiterrücken
der Flussläufe bewirkt wird, hat kürzlich M. J. Klinge[59] in
einer besonderen Abhandlung ausgeführt. Haben flutende, schwimmende
und untergetauchte Gewächse, die sich am günstigsten entwickeln an
Stellen, die von[S. 126] der Stromrichtung nicht getroffen sind, im Verein mit
dem zwischen ihnen abgelagerten Detritus soweit vorgearbeitet, dass
Butomus umbellatus, Sagittaria sagittifolia, Glyceria, Acorus, Arundo,
Phragmites, Scirpus lacustris etc. geeigneten Boden finden, so arbeiten
diese Gewächse durch Massenentwickelung darauf hin, das Gefälle der
Flüsse durch Überwachsen ganz zu heben, um für ihre Sippschaft weiten
Raum zu schaffen. Der Fluss sucht der Pflanze seitlich auszuweichen und
zwar meist unter dem Winde.
Zu den Schilfgewächsen gehören zunächst von ausgeprägten
Windblühern die Lieschkolben oder Rohrkolben (Typhaceen),
deren kolbenförmiger Blütenstand zuoberst die männlichen, unten die
weiblichen Blüten trägt. Den eigentümlichen Blütenstand, welcher einem
Lampencylinder-Putzer nicht unähnlich ist, haben Dietz[60]
und Kronfeld[61] näher untersucht. Die Blüten stellen die
denkbar einfachsten Monokotyledonenblüten dar, aus zwei Staubfäden
oder einem Stempel bestehend, an dessen Grund haarähnliche Gebilde
stehen. Sie bilden bei den weiblichen Blüten später den Flugapparat
der Früchtchen, die aber auch zu schwimmen vermögen. Im Herbst und
Frühjahr treiben sich die letzteren in mächtigen wolligen Massen auf
und an den Gewässern umher. Gegen Tierfrass sind die Lieschkolben
durch Büschel von Nadeln des Kalkoxalats (Raphiden) geschützt. Von
zwölf bekannten Arten finden sich bei uns verbreitet der breite
und der schmalblättrige Lieschkolben (Typha latifolia
und angustifolia), selten der kleine Lieschkolben
(T. minima). Ihnen verwandt sind sodann die gleichfalls
windblütigen Igelkolben (Sparganiaceen), deren männliche Blüten
in zahlreichen endständigen Köpfchen der einfachen oder verästelten,
ährig angeordneten Inflorescenzen drei langfädige Staubgefässe haben,
während die gleichfalls kugeligen weiblichen Köpfchen am Grunde sitzen
und 1–2 Griffel haben. Die Haargebilde fehlen und könnten die Früchte
des stacheligen Fruchtstandes wohl eher durch Tiere verbreitet werden.
Der Blütenstand ist hier wie bei den Typhaceen proterandrisch, so dass
der Wind nur Fremdbestäubung bewirkt. Am häufigsten ist Sparganium
ramosum und S. simplex, während S. minimum und
S. affine, die[S. 127] häufig flutende Formen ausbilden und echte
Schwimmpflanzen darstellen können, seltenere Arten sind.
Von anderen Gattungen, die das Wasserleben vereinigt und in Bezug
auf die vegetativen Organe gleichgestaltet hat, die aber in
Bezug auf die Biologie der Blüte sehr verschiedene Anpassungen —
nämlich die der nahe verwandten Landpflanzen — besitzen, mögen hier
noch die Blumenbinsen (Butomus), die Schwertlilien
(Iris) und der Kalmus (Acorus), welche alle
drei zoidiophil sind, Erwähnung finden. Die prächtigen Blütendolden
des Wasserveilchens (Butomus umbellatus), welche in
dem schwertförmigen Blattwerk einen ebenso fremdartigen Eindruck
auf den Beschauer machen, wie die gelben, grossen Blumen der
Wasserschwertlilie (Iris Pseudacorus) mit ihren
blattähnlichen Griffeln, werden durch Insekten bestäubt. Sprengel
und H. Müller fanden sie ausgeprägt proterandrisch, während sie A.
Schulz[62], der gleichfalls ihre Blüteneinrichtung beschreibt, homogam
oder schwach proterandrisch fand. Am nächsten ist unserem
Wasserveilchen die ausländische Gattung Tenagocharis mit
langgestielten eilanzettförmigen Staubblättern verwandt, während eine
andere Butomacee Lemnocharis (ebenso wie die Alismaceen Echinodorus und
Hydrocoleis) auffallende Ähnlichkeit mit Nymphaea und der Gentianee
Limnanthemum nymphaioides haben. Die Wasserbinsengewächse, welche
überhaupt den Froschlöffelgewächsen nahe stehen, haben wie diese
Milchgefässe, welche eine Ölemulsion (Schutzmittel?) enthalten. — Der
Kalmus (Acorus Calamus) ist dagegen durch ätherische
Öle geschützt, ein Umstand, der ebenso wie die Zugehörigkeit zu den
Schilfpflanzen um so mehr auffällt, als die ihm verwandte (gleichfalls
zu den Arongewächsen gehörige) Schlangenwurz (Calla palustris),
die die Blattform unserer Zimmer-Calla (C. aethiopica) teilt,
Raphiden zum Schutz hat.
Der grünlichgelbe Blütenkolben des Kalmus gehört nach den Vermutungen
Delpinos zu den malacophilen (der Schneckenbefruchtung angepassten)
Blüteneinrichtungen. Bei uns bringt der Kalmus, der sich sehr
rasch durch sein Rhizom vermehrt, überhaupt keine Früchte, er ist
„selbst steril“, adynamandrisch, wie dies[S. 128] z. B. die Fliegenfalle
(Apocynum), gewisse Reiherschnabelarten (Erodium
macrodenum) u. a. Pflanzen sind. Die letzteren bringen nie Früchte,
wenn die Bestäubung zwischen Stöcken vollzogen wird, welche von
demselben Rhizom abgezweigt sind, setzen wohl aber bei Kreuzung zweier
Stöcke verschiedenen Ursprungs Früchte an. Unsere Kalmuspflanzen sollen
samt und sonders von einem Rhizom abstammen, das 1574 von Clusius in
Wien eingeführt worden ist. Verbreitet ist der Kalmus ausser in Europa
in dem tropischen und extratropischen Ostasien, Ostindien, auf der
Insel Bourbon und in Nordamerika und da trägt er Beerenfrüchte. Wäre
daher die Adynamandrie des Kalmus auf ähnliche Ursachen zurückzuführen
wie bei anderen bekannten Pflanzen, so würde ein Experiment dies leicht
bestätigen, das indessen bisher noch nicht gemacht worden ist. Man
brauchte nämlich dann nur von einem andern Erdteil Kalmuspflanzen in
unsere Teiche mit einzusetzen und — falls die Bestäubungsvermittler
fehlten — den Blütenstaub zwischen den Stöcken verschiedener Herkunft
wechselweise zu benutzen, um Früchte zu erzielen — ein Versuch, den
ich begonnen habe. In Japan giebt es noch einen schmalblättrigen Kalmus
(Acorus graminifolius).
Bei unserer Wasserschwertlilie ist es zur Ausbildung
zweier an verschiedene Insekten angepassten Blütenformen gekommen,
indem bei der einen Form die Narbenlappen der drei äusseren Blätter
der Blumenkrone anliegen, bei der andern von ihr abstehen. Die
Bestäubungsvermittler der ersten Blütenform ist eine Schwebfliege
(Rhingia rostrata), die der zweiten sind Hummeln
(Bombus vestalis, B. agrorum, B. hortorum). Ein
ähnlicher Fall von Dientomophilie wie der hier erwähnte, über den
bei Hermann Müller (Befr. d. Bl. p. 68, 69) des Näheren nachzusehen
ist, ist nur noch für Aconitum Lycoctonum, eine Landpflanze,
durch Aurivillius nachgewiesen worden[63]. Aurivillius fand im
mittlern Schweden von dieser Pflanze gleichfalls zweierlei Stöcke,
von denen die Blüten der einen einen kurzen stärkern, fast
geraden stumpfern, die der anderen einen engen, nach hinten
verschmälerten und aufwärts gebogenen[S. 129] Sporn haben. Während die
erstere Form von Schmetterlingen besucht wird, sind diese von
dem Besuch der zweiten Blütenform gänzlich ausgeschlossen. Letztere
wird denn auch regelmässig von Hummeln (Bombus hortorum und
B. consobrinus) bestäubt. — An der Schilfbildung unserer
Gewässer beteiligen sich noch mannigfach andere Pflanzengruppen, so
besonders die grösseren Riedgrasarten und Gräser (Phragmites,
Arundo, Glyceria) und die flachblättrigen Binsen
(Scirpus), von denen häufig Verwandte submers und flutend
auftreten (Scirpus fluitans, Glyceria etc.). Doch
soll auf sie wie auch auf die eigentlichen Wasserbinsen (Scirpus
lacustris etc.) hier nicht näher eingegangen werden.
Die Schutzmittel der Schilfpflanzen gegen die Tierwelt sind schon
früher erörtert worden. Trotz derselben findet sich in dem Röhricht
und Schilf noch ein reiches Tierleben, das jedoch nur unwesentliche
Schädigungen der Pflanzenwelt des Wassers herbeiführt. Auch die
pflanzlichen Parasiten thun bei uns den Wasserpflanzen nur geringen
Schaden. Dieselben bestehen zumeist aus dem Wasserleben angepassten
Pilzen — Chytridiaceen bei Acorus, Iris etc.
(Cladochytrium tenue) — der Brandpilzgattung Doassansia
(bei Butomus, auch bei Alisma, Potamogeton), den
allenthalben verbreiteten Ascomyceten und den Rostpilzen
(auf Iris Puccinia Iridis und Uromyces Iridis, auf Acorus Calamus
Uromyces pyriformis). Von anderen Wasserpflanzen beherbergen ausser
den bereits erwähnten Seerosen z. B. noch Hippuris (Aecidium
Hippuridis, dessen Teleutosporen auf Uferhalbgräsern zur
Entwickelung kommen), Limosella (Uromyces Limosellae), Cicuta
virosa (Puccinia Cicutae virosae), Hydrocotyle, Oenanthe,
Nasturtium, Veronica Anagallis, Polygonum amphibium u. a. Rostpilze.
Von den Rostpilzen sind besonders diejenigen Arten von Interesse,
welche Beziehungen zwischen den Wasserpflanzen und gewissen
Landpflanzen unterhalten, indem sie wirtwechselnd auftreten, in
der ersten Generation (Aecidium) meistens Landpflanzen,
in der zweiten gewisse Wasserpflanzen befallend. So
verursachen Rostpilze, welche[S. 130] zuerst die Lysimachia vulgaris,
Cineraria palustris, Achillea Ptarmica etc. erkranken
machen, in ihren weiteren Generationen Krankheiten verschiedener
Riedgräser etc. unserer Teiche und die Krankheiten der
Landhahnenfussarten und Ampferarten haben weitere
Rostkrankheiten des gemeinen Schilfrohres (Phragmites communis)
zur Folge.
Litteratur und Anmerkungen.
[1] Vergl. meinen Aufsatz in Naturw. Wochenschrift von Dr.
H. Potonié, Bd. II, 1888, p. 113–115, 123–125, 159: „Die Feigen
und ihre Liebesboten“.
[2] Referate über die bisher erschienenen Abhandlungen über
Myrmecophilie gab ich im Biologischen Centralblatt Bd. IV–IX.
[3]Axell Lundström, Pflanzenbiologische Studien II.
Die Anpassungen der Pflanzen an Tiere. Mit 4 Taf. Upsala 1887.
[4]Ernst Stahl, Pflanzen und Schnecken. Eine
biologische Studie über die Schutzmittel der Pflanzen gegen
Schneckenfrass. Jen. Zeitschr. f. Naturw. u. Med. Bd. XXII. N. F. XV.
Jena 1888 (126 S.).
[5] Vgl. auch Seligo, Hydrobiologische Untersuchungen.
I. Zur Kenntnis der Lebensverhältnisse in einigen westpreussischen
Seen. Schriften d. Naturforsch.-Ges. zu Danzig. N. F. Bd. VII, H. 3.
1890 (47 S.).
[6] Das wichtigste Werk über die Biologie der Wassergewächse,
das mehrfach benutzt wurde, ist das von H. Schenck, Die Biologie
der Wassergewächse. Bonn 1886 (162 S. u. 2 Taf.). Von demselben Verf.
erschien „Vergleichende Anatomie der submersen Wassergewächse“ (mit 10
Taf.). Bibliotheca botanica Heft I.
[7]F. Ludwig, Über die Bestäubungsverhältnisse einiger
Süsswasserpflanzen und ihre Anpassungen an das Wasser und gewisse
wasserbewohnende Insekten. Kosmos V., 1881, S. 7–12. Mit 17 Holzschn.
[S. 131]
[8]John E. Klercker, Sur l’anatomie et le
développement de Ceratophyllum. Meddelanden från Stockholms Högskola.
No. 26 m. Bihang t. k. Svenska Vet-Akad. Handl. 1884.
[9]E. Rodier, Sur les mouvements spontanés et
reguliers d’une plante aquatique submergée, le Ceratophyllum demersum.
Compt. rend. 1877, T. LXXXIV, No. 18, 30. Apr. Seconde note sur les
mouvements etc. ibid. T. LXXXV, No. 20, 12. Nov. u. Sep.-Abdr. (in
Bordeaux erschienen).
[10]Charles Darwin, Ges. Werke in deutsch. Übers. Bd.
XIII.
[11]Herm. Beyer, Die spontanen Bewegungen der
Staubgefässe und Stempel. Wehlau 1888 (Beil. zum Progr. d. kgl. Gymn.
Ostern 1888, Progr. No. 18, 56 S.).
[12]Egon Ihne, Geschichte der Einwanderung von
Puccinia Malvacearum und Elodea canadensis. 18. Ber. d. Oberhess. Ges.
f. Nat. u. Heilkunde. Giessen 1879.
[13]Ferd. Cohn, Die Pflanze. Breslau 1882, XI.
Insektenfressende Pflanzen, S. 341–366.
[15]H. von Klinggraeff, Schmetterlingsfang der Drosera
anglica Huds. Naturf.-Ges. zu Danzig. N. F. Bd. VII, H. 3. 1890.
[16]Burdon Sanderson, Die elektrischen Erscheinungen
am Dionaeablatt. Biol. Centralbl. II. 1882, S. 481–500, IX. 1889, S.
1–14, Bot. C. XXXVIII 1889, S. 701–708.
[17]S. Korschinsky, Über die Samen der Aldrovandia
vesiculosa. Bot. Centralbl. XXVII, 302–304, 334–335, m. 1 Taf.
[18]Ferd. Cohn, Beiträge zur Biologie der Pflanzen I,
3, p. 71. 1875.
[19]M. Büsgen, Über die Art und Bedeutung des
Tierfangs bei Utricularia vulgaris. Ber. d. D. Bot. Ges. 1888. VI, p.
LV-LXIII.
[20]H. N. Mosely, Nature. Vol. XXX, 1884, p. 81.
Naturforscher 1884, S. 276.
[21]Ch. Darwin, Ges. Werke (Leben u. Briefe III.) Bd.
XVI, S. 315.
[22]A. v. Kerner und R. v. Wettstein, Über die
rhizopodoiden Verdauungsorgane tierfangender Pflanzen. Sitzber. d. k.
Akad. d. Wiss. Wien 1886 I. Bd. XCIII; vgl. Biol. Centralbl. VI, p.
484.
[23]V. Fayod, Über die wahre Struktur des lebendigen
Protoplasmas und die Zellmenbranen. Naturw. Rundschau V. 1890 (Vorläuf.
Mitteil.).
[26]Herm. Müller, Die Befruchtung der Blumen durch
Insekten und die gegenseitigen Anpassungen beider. Leipzig 1873. — The
fertilisation of flowers. London 1883 (564 S.).
[27]Gilbert, Reproduction végétative de l’Ultricularia
intermedia. Bull. Soc. bot. belg. Extr. Justs Bot. Jahresber. XI.
(1883), S. 55.
[30]v. Schlechtendal, Langethal u. Schenk,
Illustr. Flora von Deutschland. Neue Aufl. von Hallier besorgt.
[31]Julius Wiesner, Biologie der Pflanzen (III. T. d.
wissensch. Botanik). Wien 1889 (305 S. m. 60 Illustr. u. 1 Karte).
[32]A. v. Kerner, Über explodierende Blüten. V. K. k.
bot. zool. Ges. Wien, XXXVII, 1887, S. 28 ff. Bot. Centralbl. Bd. XXX,
1887, S. 180 ff.
[33]Hegelmaier, Monographie der Lemnaceen. Leipzig
1868. Über die Fructifikationsteile von Spirodela, Bot. Ztg. 1871;
danach die Lemnaceen in Englers Monographie (Engler u.
Prantl, Die natürl. Pflanzenfamilien 1890).
[34]William Trelease, On the Structures which favor
Cross-fertilizization in several Plants. Proceed. of the Bost. Soc.
of Nat. Hist. Vol. XXI, 1882, March 15, p. 410–440 (m. 52 Fig. auf 3
Taf.).
[35]George Engelmann, Spirodela. Bull. Torr. Bot.
Club Nov. 1870 I, p. 42–43; Anthers of Lemnae (l. c. März 1871 II, p.
10–11).
[36]Federico Delpino, Rivista botanica dell’ anno
1881. Milano 1882, S. 33.
[37]Henry Gillmann, Lemna polyrrhiza again discovered
in flower on the Detroit river. Amer. Naturalist. XV, 1881, Nov., p.
896–897.
[38] Graf von Saporta, Die Pflanzenwelt vor dem
Erscheinen des Menschen. Deutsch von Karl Vogt. Braunschweig
1881.
[39] Vgl. Casparys Bearb. d. Nymphaeaceen in Potoniés
illustr. Flora von Nord- u. Mitteldeutschl. Berlin 1889. S. 249–251.
[40]Engler und Prantl, Die natürlichen
Pflanzenfamilien. Leipzig 1888.
[41]Charles Robertson, Bot. Gaz. Vol. XIV, S. 122–125,
297–298. — W. Watson, Notes on nymphaeas. Gardeners Chronicle
1884, XXI, S. 87–88. (Das Öffnen u. Schliessen zu bestimmter Tageszeit
kann durch künstliche Belichtung oder Verdunkelung nicht abgeändert
werden.) — Die Bestäubungseinrichtungen der Nymphaeaceen sind auch von
A. Schulz (Beiträge zur Kenntnis der Bestäubungseinrichtungen u.
Geschlechtsverteilung bei d. Pflanzen. I. Bibl. bot. Heft X, II. Bibl.
bot. Heft XVII) beschrieben worden.
[S. 133]
[42]K. Fr. Jordan, Die Stellung der Honigbehälter und
Befruchtungswerkzeuge in den Blumen. Doktordiss. Halle 1886.
[43]F. Hildebrand, Die Verbreitungsmittel der
Pflanzen. Leipzig 1873.
[44]E. Huth, Die Klettpflanzen mit besonderer
Berücksichtigung ihrer Verbreitung durch Tiere. Bibl. bot. Heft IX.
Cassel 1887 (36 S. u. 78 Fig.).
[45]F. Ludwig, Über durch Austrocknen bedingte
Keimfähigkeit der Samen einiger Wasserpflanzen. Biol. Centralbl. VI, S.
229.
[46]A. G. Nathorst, Untersuchungen über das frühere
Vorkommen der Wassernuss Trapa natans. Bot. Centralbl. XXVII, 1886, S.
271–274.
[47]V. B. Wittrock, Einige Beiträge zur Kenntnis der
Trapa natans. Bot. Centralbl. XXXI, 1887, S. 352–357, 387–389.
[48]Potonié, Illustrierte Flora von Nord- u.
Mitteldeutschland. Berlin 1889, S. 241–242.
[50] Vgl. Volkens, Beziehungen zu Standort und
anatomischem Bau der Vegetationsorgane. Jahresber. d. kgl. bot. Gart.
Berlin, Bd. III, 1884.
[51]A. Kerner, Ritter von Marilaun, Die Schutzmittel
der Blüten gegen unberufene Gäste. Wien 1876, S. 22, 23.
[52]O. Kirchner, Neue Beobachtungen über die
Bestäubungseinrichtungen einheimischer Pflanzen. Stuttgart 1886.
[53]F. Ludwig, Biologische Notizen. I. Das Blühen von
Polygonum Bistorta. D. Bot. Monatsschr. VI, 1888, S. 4 ff.
[54]Fr. Xav. Pfeifer, Der goldene Schnitt und dessen
Erscheinungsformen in Mathematik, Natur u. Kunst. Augsburg 1885. —
Vgl. auch meinen Aufsatz in d. wiss. Rundsch. d. Münchener N. N, 1889,
No. 84.
[55]F. Hildebrand, Die Geschlechterverteilung bei den
Pflanzen. Leipzig 1867.
[56]Otto Kuntze, Die Schutzmittel der Pflanzen gegen
Tiere und Wetterungunst etc. Leipzig 1877, S. 80–82.
[S. 134]
[57] Vgl. H. Ortmann, Beiträge zur Kenntnis
unterirdischer Stengelgebilde. Doktordissert. Jena 1886 (weitere
Litteratur). — C. Müller, Der Bau der Ausläufer von Sagittaria
sagittaefolia. Sitzungsber. d. Ges. Naturforsch. Freunde zu Berlin.
1884.
[58]E. Warming, Über die Keimpflanzen von Phragmites
communis. Bot. Centralbl. XXI, S. 156 ff.
[59]M. J. Klinge, Über den Einfluss der mittleren
Windrichtung auf das Verwachsen der Gewässer nebst Betrachtung
anderer von der Windrichtung abhängiger Vegetationserscheinungen
im Ostbalticum. Englers bot. Jahrbücher XI, 1889, S. 264–313. Bot.
Centralbl. XLII, S. 25.
[60]Sandor Dietz, Die Blüten- und Fruchtentwickelung
bei den Gattungen Typha und Sparganium. Biblioth. bot. Heft 5, 1887
(mit 3 Taf.). — Bot. Centralbl. XXVIII, 1886, S. 26–30, 56–60.
[61]Kronfeld, Über den Blütenstand des Rohrkolbens.
Sitzungsber. d. k. Akad. d. W. Wien 1886, I. Abt, Bd. XVI, S. 78–109.
[62]A. Schulz, Bibl. bot. H. 10. Cassel 1888, S. 96–97
(vgl. unter 40).
[63] Vgl. F. Ludwig, Ein neuer Fall verschiedener
Blütenformen bei Pflanzen der nämlichen Art etc. Biol. Centralbl. VI,
1887, S. 737–739.
[S. 135]
Ein Wurzelfüsser
des Süsswassers in Bau und Lebenserscheinungen dargestellt.
Von Prof. Dr. A. Gruber.
[S. 137]
Noch ist der Schleier, der über die erste Entstehung des Lebens auf
unserer Erde gezogen ist, nicht gelüftet. Seit der Mensch angefangen
hat, über seine Umgebung nachzudenken, sind mythische und mystische
Vorstellungen, sind wissenschaftliche Spekulationen und Theorien
entstanden und vergangen und auch heute haben wir für die Frage, wie
die lebende organische Substanz entstanden sei, keine Antwort zu geben.
Wohl aber sind wir durch die Fortschritte der Wissenschaft der Lösung
einer sich daran schliessenden zweiten Frage nahegetreten, nämlich der,
wie die ersten lebenden Organismen ausgesehen haben mögen.
Die erste Anwendung des Mikroskops im 17. Jahrhundert lehrte den
Menschen in den Gewässern der Erde Wesen von ungeahnter Kleinheit
erkennen, die Begründung der Zellenlehre im Anfange unseres
Jahrhunderts verschaffte denselben ihre richtige Bedeutung als
Elementarorganismen und die ungeahnte Vervollkommnung der optischen
Hilfsmittel in unseren Tagen gestattet uns einen tiefen Einblick in den
einfachen und doch so komplizierten Bau ihres Körpers und lässt uns
an ihnen die Lebensäusserungen der Materie auf ihrer niedersten Stufe
erkennen.
In diesem Reiche der Urorganismen oder Protisten stehen wohl zu
unterst die Schleimtiere, die Wurzelfüsser oder wie ihr Name lauten
mag, kurz diejenigen Organismen, deren Protoplasmaleib, einer einzigen
Zelle an Wert entsprechend, keine feste Gestalt besitzt, sondern
regellos nach allen Richtungen des Raumes auszufliessen vermag. Ich
will die Streitfrage nicht weiter berühren,[S. 138] ob wirklich noch heute
in den Abgründen des Oceans ausgedehnte Mengen organischen Schleimes
leben oder nicht, ob der seinerzeit auch in der populären Litteratur
viel besprochene Bathybius, beziehungsweise ob ein ihm nahestehender
Organismus wirklich existiert, es mag uns die sichere Thatsache
genügen, dass wir viele und ganz unzweifelhafte Lebewesen kennen,
die eben jener Vorstellung von der niedersten Lebensstufe vollkommen
entsprechen. Im Meere sind die Wurzelfüsser schon lange bekannt, weil
dort auch dem blossen Auge sichtbare, von fester Schale umgebene Formen
leben. Die Reste solcher Schalen haben sich aus den ältesten Zeiten
unserer Erdgeschichte in grossen Massen erhalten und schon im Altertum
sind z. B. die Nummuliten bekannt, wenn auch nicht erkannt gewesen.
Aber auch das süsse Wasser, mit dessen Bewohnern dieses Buch
sich beschäftigt, beherbergt Wurzelfüsser in grosser Zahl und
mannigfacher Gestaltung. Schon im Jahre 1755 hat der wackere Rösel
von Rosenhof[64] die ersten Amöben aufgefunden und in seiner
„Insektenbelustigung“ beschrieben und abgebildet.
Die Beschreibung ist, obgleich sie sich natürlich nur auf die äussere
Gestalt und Formveränderung bezieht, trefflich und, wie alles im
Röselschen Buch, anmutend und auch für uns Fortgeschrittenere
noch belehrend.
Die Amöbe ist nichts weiter als ein kleiner Protoplasmatropfen, von
keiner festeren Haut umgeben und aller denkbaren Formveränderung
fähig, bald in gleichmässigem schneckenartigen Kriechen oder, besser
gesagt, wie der Regentropfen an der Fensterscheibe gleitend, bald
sich zusammenkugelnd oder hie und da Fortsätze aussendend und wieder
einziehend. Es giebt kaum etwas Wunderbareres, als die mannigfaltigen
Bewegungen dieses Tröpfchens lebender Materie unter dem Mikroskope zu
verfolgen. Von hohem Interesse ist es, dass man in der jüngsten Zeit
dahin gekommen ist, dieselbe amöboide Bewegung an leblosen Objekten zu
beobachten[65]. Man stellte verschiedenartig gemischte Schmierseifen-
oder Olivenölschäume dar und sah Tropfen derselben gerade wie Amöben
bis sechs Tage lang ununterbrochen strömen. Ist es also[S. 139] gelungen,
künstliche Amöben im Laboratorium darzustellen? Das natürlich nicht,
denn all die komplizierten Lebenserscheinungen, von welchen wir
weiterhin zu sprechen haben werden, fehlen diesen Schaumtropfen, sie
lehren uns aber, wie die so rätselhaften Bewegungserscheinungen der
niedersten Lebewesen auf bekannte mechanische Vorgänge zurückgeführt
werden können.
Eine grosse Anzahl der Süsswasser-Amöben umgiebt ihren weichen Körper
mit einer schützenden Schale, die zwar nicht so kompliziert ist, wie
bei vielen der Meeres-„Foraminiferen“, aber zierlich und in ihrer
Entstehung interessant genug. Diese Schalenträger sind es, welchen
ich diesen kleinen Aufsatz widmen möchte. Sie leben vorzugsweise in
den stehenden Gewässern, teils auf dem Grunde, teils an lebenden oder
abgestorbenen Pflanzenteilen hinkriechend. Umstehendes Gruppenbild
(Fig. 16 S. 140) zeigt einen Tropfen irgend eines Weihers oder
Sumpfes mit drei Algenfäden, auf welchen verschiedene Vertreter der
Wurzelfüsser sich festgeheftet haben, alles bei starker Vergrösserung.
Da sehen wir rechts unten (Fig. 16, 2) das spitz zulaufende Gehäuse der
Difflugia acuminata, ganz aus Sandkörnern aufgebaut, zwischen
denen auch längliche Diatomeenschalen angeklebt sind, aus der Mündung
treten die Plasmafortsätze aus, mittels deren das Tier sich fortbewegt.
Noch zwei andere Difflugia-Arten zeigt unser Bild: Die eigentümlich
gewundene D. spiralis (Fig. 16, 3) und links oben (Fig. 16,
1) die D. urceolata, deren Gehäuse wie eine zierliche Urne
regelrecht gebaut ist. Ebenfalls eine Schale aus Fremdkörpern, zugleich
versehen mit stachelartigen Fortsätzen, besitzt die Centropyxis
aculeata (Fig. 16, 5). Im Gegensatz zu den genannten Arten ist bei
der Fig. 16, 6 abgebildeten Hyalosphenia papilio das Gehäuse
ein Ausscheidungsprodukt, ein feines Häutchen, so durchsichtig, dass
man den Plasmakörper deutlich darin liegen sieht, mit feinen Fäden in
der Schale aufgehängt. Nun giebt es aber auch Formen, bei denen die
Schale zwar auch vom Tier selbst erzeugt wird, aber nicht im ganzen,
sondern in einzelnen Stücken, die, wie wir später sehen werden,
kunstvoll an einander gefügt sind. Bei der Arcella vulgaris
(Fig. 16, 4), einer der häufigsten Arten in unseren Gewässern, ist[S. 141]
die linsenförmige Schale aus winzigen Prismen zusammengesetzt, so
dass dieselbe bei nicht ganz starker Vergrösserung fein punktiert
erscheint. Ebenso aus ganz kleinen Bestandteilen besteht das Gehäuse
der zierlichen Cyphoderia ampulla (Fig. 16, 8), während bei
Quadrula symmetrica (Fig. 16, 7) und Euglypha alveolata
(Fig. 16, 9) die einzelnen Schalenbestandteile leichter zu erkennen
sind; bei der erstern sind es quadratische Plättchen, bei der andern
annähernd kreisrunde Scheiben, welche die zarte Hülle zusammensetzen.
Ich könnte noch eine grosse Reihe von Formen aufführen, die den auf
der Tafel dargestellten mehr oder weniger ähnlich sind und die, so
weit unsere Kenntnisse diesen Schluss erlauben, über die ganze Erde
verbreitet sind. Dieselben Arten nämlich, die wir in Europa kennen,
sind zumteil schon in Nord- und Südamerika, Asien, Afrika, Australien
und in den arktischen Ländern gefunden worden; auch vertikal scheint
für ihre Verbreitung kaum eine Grenze zu ziehen, denn in der Schweiz
fanden sie sich noch in 8000′, in Nordamerika sogar in 10000′ Höhe
unverändert vor[66]. Es ist mir aber hier nicht um eine Aufzählung
aller bisher entdeckten Formen zu thun, ich möchte mich begnügen eine
einzige zu beschreiben, diese soll aber in ihren feinsten Einzelheiten
untersucht und in allen ihren Lebensvorgängen verfolgt werden. Dass
eine derartige Spezialisierung ihren hohen Wert besitzt, hoffe ich dann
am Schlusse nachweisen zu können.
Fig. 16.
Wurzelfüsser des süssen Wassers in einem Wassertropfen an Algenfäden sitzend
bei starker Vergrösserung. 1 Difflugia urceolata — 2 Difflugia
acuminata — 3 Difflugia spiralis — 4 Arcella vulgaris von der
Seite und von oben — 5 Centropyxis aculeata — 6 Hyalosphenia
papilio — 7 Quadrula symmetrica — 8 Cyphoderia ampulla — 9
Euglypha alveolata.
(Das Grössenverhältnis der hier abgebildeten Arten zu einander entspricht nicht
genau der Wirklichkeit.)
Unter allen ihren Verwandten in der Gruppe der schalentragenden
Wurzelfüsser des süssen Wassers ist die Euglypha alveolata
wohl am genauesten beschrieben, ja man kann wohl sagen, dass heute
kaum ein Organismus, Tier oder Pflanze, besteht, der so vollständig
in Gestalt und Lebensweise erforscht wäre. Anno 1841 wurde das Tier
von Dujardin zum ersten Mal, später auch von Ehrenberg
kenntlich dargestellt, und seither haben wohl mehr als fünfzehn
Forscher sich damit beschäftigt[67]. Kein Wunder also, wenn die
Erkenntnis eines so einfach gebauten Organismus weit gefördert werden
musste.
Der Körper der Euglypha besteht, wie derjenige aller Wurzelfüsser,
aus einer kleinen Menge von Protoplasma, welches mehr oder[S. 142] weniger
körnig erscheint und an dem zunächst bei schwächerer Vergrösserung
keinerlei Strukturen zu erkennen sind; ein starker lichtbrechender
kugeliger Körper tritt aber auch jetzt schon ziemlich deutlich hervor,
das ist der Kern, und damit haben wir die erste wichtige Thatsache
festgestellt, dass die Euglypha den Formwert nur einer Zelle besitzt.
Wir können also nicht erwarten, in ihrem Körper Organe im Sinne der
höheren Tiere und Pflanzen aufzufinden, denn ein Organ besteht ja schon
an sich aus einer Vielheit von Zellen. Gehen wir in der Betrachtung des
einzelligen Geschöpfes von aussen nach innen vor: Der protoplasmatische
Körper der Euglypha steckt in einem überaus zierlichen, etwa
tonnenförmigen Gehäuse (Fig. 17) von winzigen Dimensionen: Bei der
gewöhnlichen Form nämlich ist der Längsdurchmesser nur ⁶⁄₁₀₀ mm
und der Querdurchmesser ³⁄₁₀₀ mm im Mittel.
Trotz dieser Winzigkeit gestatten uns unsere heutigen Instrumente,
die einzelnen Bestandteile, aus welchen die Schale aufgebaut ist,
genau zu erkennen: Es sind konvex-konkave Plättchen aus einer dem
Chitin ähnlichen, vielleicht von Kieselsäure imprägnierten Substanz.
Die Plättchen, deren konvexe Seite nach aussen gekehrt ist, decken
sich dachziegelartig und da diejenigen Stellen, wo sie über einander
greifen, dunkler erscheinen, so macht es den Eindruck, als wenn die
Schale polygonal gefeldert wäre. Diejenigen Plättchen, welche vorn
die Öffnung umgeben, sind nicht rund, sondern laufen in eine Spitze
aus und sind an ihrem freien Rande fein gezähnelt. Bei einer andern
Euglypha-Art stehen noch zwischen den Schalenplättchen ab und zu spitze
Stacheln.
Fig. 17.
Ein leeres Gehäuse von Euglypha alveolata.Fig. 18.
Eine Euglypha alveolata im ausgebildeten Zustande (etwa 700mal vergr.).
HZ Hintere, hyaline Zone — KZ Mittlere, Körnchenzone — AZ
Vordere, alveoläre Zone — Ch Die Protoplasmamaschen (Cyto-Hyaloplasma) —
Cch Die Maschenräume (Cyto-Chylema) — Cm Die Körnchen
(Cyto-Mikrosomen) — ps Die Pseudopodien — n Der Kern mit dem
Kernkörperchen ncl — CV Die kontraktile Vacuole — sp Die
Reserve-Schalenplättchen — nk Nahrungskörper — e Exkretkörnchen.
Die Schale der Euglypha wird vom Protoplasmakörper meist nicht
vollkommen ausgefüllt, sondern es bleibt an der Seite ein freier Raum
übrig.
Vorn aus der Mündung (Fig. 18) strahlen feine Fortsätze des Plasmas
(ps), die sogenannten Scheinfüsschen oder Pseudopodien,[S. 143]
aus, deren wurzelartige Verzweigungen dem Namen Wurzelfüsser oder
Rhizopoden den Ursprung gegeben haben. Sie müssen zwei wichtige
Funktionen vermitteln, erstens die Ortsbewegung und zweitens die
Nahrungsaufnahme. Die Nahrung (nk) besteht meistens aus kleinen,
einzelligen Algen, Diatomeen und dergleichen. Die Wurzelfüsschen
erkennen mittels des Tastgefühls die ihnen zusagende Beute, umfliessen
dieselbe und führen sie dann dem Körper zu. Dies geschieht dadurch,
dass das Plasma des betreffenden Fortsatzes einfach zurückfliesst und
den Nahrungskörper im Strome mitreisst. Betrachtet man den vordern
Abschnitt des Körpers, aus dem die Scheinfüsschen ihren Ursprung
nehmen (AZ), bei starker Vergrösserung, so erkennt man,
dass das Plasma nicht eine homogene Masse darstellt, sondern dass
es aus einem Maschenwerk besteht. Die Maschen (Ch) werden
von hyalinem Plasma gebildet, in welchem winzig kleine Körnchen
(Cm), die sogenannten Mikrosomen, suspendiert sind, und die
Maschenräume (Cch) von einem dünnflüssigeren Safte erfüllt.
Die Körnchen ermöglichen es, ein fortwährendes Strömen des Plasmas
zu erkennen. Durch diesen Strom wird auch die aufgenommene Nahrung
im Körper verschoben und zwar gelangt sie in den mittlern Abschnitt
(KZ) des Körpers, der sich durch ein engeres Maschenwerk
und grossen Körnerreichtum auszeichnet. Während wir an den ersten
Abschnitt im wesentlichen[S. 144] die Funktionen der Bewegung gebunden sehen,
dürfen wir den mittleren Teil des Körpers als die Ernährungs- oder
Stoffwechselzone bezeichnen. Eine scharfe Trennung besteht übrigens
zwischen den beiden Abteilungen nicht, sondern der Übergang ist
ein ganz allmählicher. Die Körner, welche die Zone oft ganz dicht
erfüllen, hat man mit dem Namen Exkretkörner (e) belegt und
sieht sie als Endpunkte des Stoffwechsels an. Dass sie sowohl wie
die Nahrung gerade an den mittlern Körperteil gebunden sind, dies
berechtigt uns, diesem Teil des Plasmas eine verdauende Funktion
zuzuschreiben. In demselben Abschnitt liegt auch ein Gebilde, welches
man mit dem Stoffwechsel in Zusammenhang gebracht hat, nämlich die
sogenannte kontraktile, oder pulsierende Vacuole (CV). Es ist
ein Bläschen, welches in regelmässigen Pulsationen sich leert und
wieder füllt. Man hat angenommen, dass durch diese bei den Urtieren
sehr verbreitete Blase die Endpunkte des Stoffwechsels nach aussen
befördert würden; es ist dies auch nicht unwahrscheinlich, die
Hauptbedeutung der kontraktilen Vacuole liegt aber offenbar darin, das
in den Körper eindringende Wasser wieder zu entfernen, es wird mit
andern Worten dadurch eine kontinuierliche Aufnahme sauerstoffhaltigen
Wassers ermöglicht und in dieser Weise die Atmung befördert. Demnach
wäre an den mittlern Körperabschnitt die Assimilation, die Exkretion
und die Respiration gebunden. Ebenfalls wieder ohne scharfe Grenze
geht diese Zone in den hintersten Teil des Körpers über (HZ),
der fast hyalin erscheint, weil hier das Maschenwerk noch feiner
und das Lichtbrechungsvermögen von Maschen und Mascheninhalt fast
dasselbe ist. Hier liegt der Zellkern (n) als helle Kugel,
in welcher ein dunkleres Korn, das Kernkörperchen (ncl),
eingeschlossen liegt. Die Forschungen der neuesten Zeit haben uns mit
der unumstösslichen Thatsache bekannt gemacht, dass der Kern im Leben
der Zelle eine ausserordentlich wichtige Rolle zu spielen hat und
dass bei der Befruchtung und Fortpflanzung er das allein Wesentliche
und Beherrschende ist. Gerade die Urtiere sind es auch gewesen, bei
welchen entscheidende Versuche über die Bedeutung des Kernes angestellt
worden sind[68]. Man hat solche Organismen mittels scharfer Instrumente
zerschnitten und hat nun[S. 145] gefunden, dass eine Regeneration, ein Ersatz
verloren gegangener Teile nur dann eintritt, wenn das abgetrennte
Stück den Kern oder wenigstens einen Teil desselben noch enthält;
die kernlosen Stücke dagegen regenerieren sich nicht, sondern gehen
über kurz oder lang zu Grunde. Mit anderen Worten, die Zelle und
ebenso der einzellige freilebende Organismus sind nicht im stande,
Neubildungen hervorzubringen, wenn die Zellsubstanz nicht auch mit
Kernsubstanz versehen ist. Bei denjenigen Wurzelfüssern, deren Schale
ein Ausscheidungsprodukt ist, wie bei den marinen Foraminiferen, hat
man Stücke der Schale ausgebrochen und auch diese wurden nur dann
durch neue Ausscheidungen ersetzt, wenn ein Kern vorhanden war[67 d].
Weiter hat man gefunden, dass der Kern allein das Substrat enthält, an
welches bei der Fortpflanzung der Organismen die Vererbungstendenzen
gebunden sind. Die Eigenschaften der Eltern gehen nur durch den Kern
der Fortpflanzungszellen auf die Kinder über. Ich werde darauf noch
zurückzukommen und es mag zunächst genügen, darauf hingewiesen zu
haben, dass wir im Kerne der Euglypha den wichtigsten Bestandteil des
Tieres erkannt haben. Er ist an sich ein Mikrokosmos und es darf uns
nicht Wunder nehmen, dass wir, je schärfer unsere Instrumente werden,
immer kompliziertere Differenzierungen in ihm erkennen. Um den Kern der
Euglypha, also ebenfalls im hinteren Abschnitte des Körpers, finden
sich zur Zeit der vollkommenen Reife eine Menge kleiner Körperchen
gelagert (sp), die nichts anderes sind als Schalenplättchen,
ganz gleich denjenigen, welche das Gehäuse des Tieres zusammensetzen.
Wir werden gleich sehen, wie dieselben zur Verwendung gelangen.
Bei allen zu weiterem Leben fähigen Urtieren, wie bei jeder Zelle,
tritt nach reichlicher Ernährung ein Moment ein, wo der Körper das
ihm eigentümliche höchste Mass erreicht hat und dann erfolgt die
Vermehrung durch Teilung. Bei Euglypha, wo die Schale eine weitere
Ausdehnung des Körpers nicht gestattet, muss die Grenze des Wachstums
in einer bestimmten Konzentration des Plasmas gesucht werden. Ist diese
erreicht, dann teilt sich die Euglypha und zwar in folgender höchst
merkwürdigen Weise: Es[S. 146] werden zunächst die Scheinfüsschen eingezogen
und an ihrer Stelle tritt ein Klumpen Plasma aus der Schalenmündung
aus, der immer grösser und grösser wird. Zugleich geraten jene um
den Kern her gelagerten Reserve-Schalenplättchen in Bewegung und wir
sehen sie in stetem Flusse, eins hinter dem andern, aus der Mündung
heraus in den ausgetretenen Plasmazapfen hereinwandern. Wie von einer
unsichtbaren Hand geleitet, werden sie an die Oberfläche dieses immer
mehr und mehr anwachsenden Zapfens geschoben und so an einander
gelagert, dass sie sich weit übergreifend dachziegelförmig decken und
der ganze Spross dadurch ein tannenzapfenförmiges Gebilde darstellt
(Fig. 19). Der Spross schwillt aber immer mehr an und zwar so lange,
bis er genau die Grösse des ursprünglichen Tieres erreicht hat, und
dann hat das Plasma die Schalenplättchen so weit auseinandergedrängt,
dass sie sich nicht mehr und nicht weniger decken, als diejenigen des
alten Gehäuses. Dabei zeigt es sich, dass genau so viele Plättchen
vorhanden waren, als zum Aufbau der neuen Schale nötig sind, und
ferner, dass die ersten Plättchen, welche austraten, die gezähnelten
Randplatten sind, welche genau mit denjenigen des alten Gehäuses
ineinandergreifen (Fig. 20).
Fig. 19.
Euglypha in Teilung (etwa 700mal vergr.). Ein Teil des Protoplasmas ist
ausgetreten, die Reserveplättchen werden hinausgeschoben und beginnen die neue
Schale zu bilden. Der Kern befindet sich im sogen. Sonnenstadium der Teilung.
Fig. 20.
Euglypha in Teilung. Die neue Schale ist fertig. Kern im Stadium der
sogen. Sternform.Fig. 21.
In der alten Schale ist fast nur noch Plasma der hintern Zone. Im Kern
haben sich die Schleifen gespalten.
Fig. 22.
Durchschnürung des Kerns.Fig. 23.
Der Teilungsprozess ist beendigt; die beiden Schalen und ihr Inhalt sind
vollkommen kongruent; an den Mündungen beginnen Pseudopodien auszutreten.
Buchstabenerklärung wie bei Fig. 18.
Oberflächlich betrachtet scheinen nun zwei vollkommen kongruente
Euglyphen aus der einen ursprünglichen entstanden; dem ist aber noch
nicht so, denn die neu entstandene enthält ja noch keinen Kern und
noch kein pulsierendes Bläschen. Dieses tritt vielmehr erst ganz am
Schlusse des Teilungsprozesses auf, nachdem es auch im Muttertier
verschwunden gewesen war. Der Kern hat[S. 147] von dem Moment an, wo das
Plasma aus dem Muttergehäuse auszutreten begann (Fig. 19), auffallende
Veränderungen gezeigt, die seine Teilung einleiteten; nachdem die neue
Schale sich gebildet hat, ist auch er in zwei Teile zerfallen, unter
Erscheinungen, die ich noch eingehender besprechen werde. Nun wandert
die eine Hälfte des geteilten Kernes in den Tochterspross hinein,
während die andere im hinteren Ende des Muttertieres zurückbleibt.
Aber[S. 148] auch damit sind die beiden Hälften noch nicht vollkommen
gleichwertig, denn der Tochterspross enthält fast nur Plasma der
ersten und zweiten Zone (Fig. 21 u. 22), während das der dritten Zone
im alten Gehäuse zurückgeblieben ist. Damit nun eine gleichmässige
Verteilung aller Plasmaelemente bewerkstelligt werde, beginnt eine
lebhafte zirkulierende Strömung von einer Schale zur andern, wie wenn
in dem lebenden Brei herumgerührt würde. Die Strömung[S. 149] hält so lange
an, bis jedes Individuum seinen gleichen Anteil an Plasma erhalten
hat, ja sogar die Nahrungskörper, welche zur Zeit der Teilung sich in
der Euglypha befanden, werden annähernd gleich auf die beiden Hälften
verteilt. Mittlerweile haben auch die beiden Kerne ihre normale
Struktur und Lage angenommen und in jedem Tiere pulsiert wieder eine
contraktile Vacuole, jetzt erst sind beide Hälften einander kongruent
(Fig. 23). Bald sieht man zwischen den Randplatten feine Scheinfüsschen
sich hervordrängen, die Mündungen lösen sich von einander ab und die
beiden Euglyphen gehen selbständig ihre Wege. Dies ist die merkwürdige
Vermehrungsweise der Euglypha alveolata. Wir haben aber noch eine
Beschreibung der Kernteilung nachzuholen mit Berücksichtigung der
inneren Vorgänge im Kern. Trotz der Kleinheit des Objektes — der
Durchmesser des Kernes beträgt nur etwa ⁸⁄₁₀₀₀ mm — sind
dieselben doch genau bekannt: Die Teilung wird dadurch eingeleitet,
dass in dem bisher homogen erscheinenden Kerne eine feinmaschige
Struktur sichtbar wird (Fig. 24); zwischen den Maschen ist das
Kernkörperchen noch deutlich zu sehen. Aus der maschigen wird eine
faserige Struktur (Fig. 25) und bald erkennt man in den Fasern
regelmässige Fäden, welche einen dicht verschlungenen Knäuel bilden.
Während der Kern stetig an Umfang zunimmt, werden die Fäden dicker und
werden dann in eine grössere Anzahl unter sich gleich langer Stücke
zerlegt (Fig. 26); das Kernkörperchen verschwindet und die einzelnen
Fadenstücke biegen sich, so dass sie allmählich eine V-förmige Gestalt
annehmen.
[III] Die Figuren 24–32 sind nach Präparaten bei etwa
1200maliger Vergrösserung entworfen. Die Figuren 18–32 sind Kopien nach
Schewiakoff 67 m.
Fig. 24[III].
Netzstruktur des Kerns.Fig. 25.
Faseriger Bau des Kerns.Fig. 26.
Der Knäuel löst sich in Fadenstücke auf, die sich umzubiegen beginnen.
[S. 150]
Fig. 27.
Kern in der Sonnenform.Fig. 28.
Kern im Anfangsstadium der Sternform.Fig. 29.
Die Sternform; von den Polkörperchen aus strahlen die feinen Fäden der
Kernspindel zu.
Von einer im Mittelpunkt der Kernkugel wirkenden Kraft werden all
diese gebogenen Fadenstücke oder „Schleifen“ mit ihrer Winkelspitze
nach dem Zentrum gerichtet; man nennt dies die „Sonnenform“ (Fig. 27).
Auch im umgebenden Zellplasma macht sich die Anziehungskraft bemerkbar
und erhält ihren Ausdruck in feinen Linien, welche radiär auf den
Umfang des Kernes zustrahlen (Fig. 19). Während des Sonnenstadiums
macht der Kern allerlei Bewegungen und verliert dann die Kugelform,
um sich an den Polen abzuflachen (Fig. 28). Zugleich erscheinen hier
zwei neue Attraktionszentren und zwar in Gestalt kleiner kegelförmiger
Körper, der sogenannten Polkörperchen oder, wie sie neuerdings genannt
werden, Centrosomen. Ihnen streben jetzt die Linien im umgebenden
Plasma zu (Fig. 29) und die Anziehungskraft im Mittelpunkt des Kernes
hört auf; die Schleifen, welche in der auf die Sonnenform folgenden
sogenannten „Sternform“ (Fig. 28 u. 29) regelmässig um den Äquator
angeordnet waren, werden nun nach den Polen gezogen, sie kehren
sich um und wenden die Spitze vom Zentrum ab. Während der Kern nun
in umgekehrter Richtung sich am Äquator abflacht, haben sich feine
blasse Fäden ausgebildet, welche von Pol zu Pol ziehend die zierliche
sogen. Kernspindel darstellen (Fig. 29). Diese Spindelfäden sind es,
welche die Schleifen den Centrosomen zuleiten. Vorher aber spielt
sich der für die Kernteilung bedeutendste Vorgang ab, die Spaltung
der Schleifen: Der ganze Kernfaden und damit auch die aus ihm
entstandenen Schleifen besteht nämlich aus kleinsten Körnchen, die[S. 151]
in regelmässigen Abständen hintereinander aufgereiht liegen. Alle
diese Körnchen werden mit einem Male der Länge nach zerteilt und so
die ganze Schleife längsgespalten; damit erhalten wir im Kerne also
doppelt so viel Schleifen als vorher (Fig. 30). Von jeder gespaltenen
Schleife aber gleitet nun die eine Hälfte links, die andere rechts
vom Äquator weg oder, anders ausgedrückt, die eine nach dem Nord-,
die andere nach dem Südpol den Spindelfasern entlang. Das Resultat
ist klar: Die färbbare Kernsubstanz, denn diese ist es, welche die
Schleifen bildet, wird auf das genaueste in zwei Teile zerlegt, die
uns quantitativ als Hälften des Ganzen erscheinen. Jede Hälfte ist für
einen der Tochterkerne bestimmt. Nachdem diese sorgfältige Zerteilung
der Kernsubstanz beendet, zieht sich der Kern im Äquator immer mehr
zusammen, die Schleifen drängen sich um die Pole her, die blassen
Spindelfäden reissen durch (Fig. 31) und endlich schnürt sich der
Kern in zwei Hälften aus einander (Fig. 32 S. 152). Nun machen die
Tochterkerne rückwärts dieselben Veränderungen wieder durch, die wir
am Mutterkern ablaufen sahen, bis sie wieder die Gestalt und Struktur
des sogenannten „ruhenden Kerns“ angenommen haben. Damit ist der
Teilungsvorgang vollendet. Die Teilung der Euglypha bedeutet ihre
Vermehrung. Bei höheren Tieren und Pflanzen geht der Vermehrung die
Befruchtung voraus. Sie besteht in der Verschmelzung[S. 152] zweier Zellen
verschiedenen Ursprungs, der Ei- und Samenzelle. Hier müsste sie
also in der Verschmelzung zweier ganzer Individuen bestehen. Dies
ist auch bei den meisten Urtieren der Fall und auch bei Euglypha ist
ein solcher Befruchtungsakt beobachtet worden[67 l]: Zwei Tiere legen
sich mit ihren Schalenmündungen neben einander, aus der doppelten
Menge von Reserveplättchen entsteht eine neue Schale, in welcher
die zwei Plasmakörper und die zwei Kerne in eins verschmolzen zu
liegen kommen. Es ist also durch Vermischung zweier Individuen ein
einziges entstanden. In welchen Zwischenräumen diese „Kopulation“
eintritt, wie viele Teilungen hintereinander folgen können bis wieder
eine Befruchtung eintritt, darüber sind für unsere Art noch keine
Beobachtungen angestellt worden.
Fig. 30.
Umordnung und Spaltung der Schleifen.Fig. 31.
Stadium der Tochtersterne; die Spindelfasern reissen in der Mitte
durch.Fig. 32.
Durchschnürung des bisquitförmig gewordenen Kerns.Fig. 33.
Eine Euglypha mit Stacheln im encystierten Zustande. D Diaphragma —
a. s. äussere Schale — i. s. Innere Schale — C
Cystenhülle.
Noch haben wir nicht alle Lebensvorgänge der Euglypha verfolgt: Bei
ihrem Aufenthalt in den oft seichten stehenden Gewässern kann es
nicht ausbleiben, dass die Euglypha der Gefahr des Austrocknens oder
Einfrierens ausgesetzt ist. Davor weiss sie sich wie die meisten
übrigen Urtiere und Urpflanzen durch die sogenannte Einkapselung oder
Encystierung zu schützen (Fig. 33): Sie zieht[S. 153] die Pseudopodien ein
und schliesst das Gehäuse vorn durch einen Deckel, das Diaphragma
(D) ab; ausserdem baut sie sich aus den Reserveplättchen
innerhalb ihres Gehäuses (a. s.) noch ein zweites, kleineres,
bei welchem die Plättchen weiter übereinandergreifen und das vorn
geschlossen ist (i. s.). Innerhalb der zweiten Schale zieht sich
nun der Protoplasmakörper zu einer Kugel zusammen und um diese wird
erst die eigentliche Cystenhaut (C) ausgeschieden. In diesem
Zustande ist die Euglypha vor Trockenheit und Frost geschützt.
Die Encystierung ist aber auch ein wichtiges Mittel zur Verbreitung
der Art. Mit dem Staube werden solche Cysten weit fortgetrieben,
Wasservögel und Wasserinsekten tragen sie im Schlamm, der da und
dort an ihrem Körper haften bleibt, mit sich weg in entfernte
Gewässer. Ja es scheint, dass die Spitzen und Stacheln, welche manche
Süsswasserrhizopoden an ihrer Schale haben (s. Fig. 16) und welche, wie
schon erwähnt, auch bei Euglypha vorkommen (Fig. 17), als Haftapparate
anzusehen sind, um den Transport zu begünstigen. Dieser passiven
Wanderung verdanken die Urtiere offenbar ihre weite Verbreitung[69],
darauf beruht es, dass gerade die schalentragenden Wurzelfüsser
des süssen Wassers, so weit man bis jetzt weiss, alle sogenannte
kosmopolitische Arten sind[70].
Will die Euglypha sich aus ihrer Cyste wieder befreien, so löst sie
zunächst die Cystenhülle auf, dann sprengt sie die innere Schale, so
dass die Plättchen wieder lose umherliegen, zerreisst das Diaphragma
und teilt sich auf die oben beschriebene Weise, indem nun die Plättchen
der inneren Schale gleich für das Tochtergehäuse benutzt werden[67 k].
Wir haben nun die Euglypha in ihrem ganzen Lebenslauf verfolgt,
wir haben gesehen, wie sie sich bewegt, frisst, verdaut, wie der
Stoffwechsel vor sich geht, wir kennen die Zusammensetzung und
Entstehung ihrer zierlichen Schale, wir erkennen die feinere Struktur
des Plasmas, wissen, wie die Vermehrung, die Befruchtung, die
Einkapselung vor sich geht; ja wir können in die feinsten Vorgänge der
Kernteilung eindringen; in dem Kerne, der selbst nur ⁸⁄₁₀₀₀ mm
gross ist, erkennen wir noch zahlreiche Fäden, in diesen[S. 154] Fäden sehen
wir noch Körner und wir sehen diese Körner sich noch teilen. Das ist
das Höchste, was wir mit unseren heutigen Mikroskopen leisten können,
aber — wird der Leser sagen — das ist auch der höchste Grad der
Spitzfindigkeit. Wohl, mag dem so sein, aber wir lernen doch sehr viel
dabei: Das Plasma ist der Träger des Lebens, und wie bei manchen Zellen
der höheren Tiere gelingt es hier bei der Euglypha dem bewaffneten
Auge, zu erkennen, dass dieser Sitz des Lebens, entsprechend seiner
Vielseitigkeit, ein viel komplizierteres Gefüge hat, als man bisher
geahnt. Das Maschengerüst mit seinen grösseren und kleineren von
Zellsaft erfüllten Waben, mit seinen feinen und feinsten Körnchen,
die Vorgänge bei der Kernteilung lassen uns ahnen, dass man auch
in dem kleinsten Plasmaklümpchen einen Mikrokosmos von unendlicher
Kompliziertheit voraussetzen muss.
Das Wachstum aller höheren Tiere und Pflanzen beruht auf der Vermehrung
der Zellen, welche ihren Körper zusammensetzen. Die Zellen sind es,
welche durch Nahrungsaufnahme wachsen und sich teilen, durch ihre
Teilung die Gewebe und Organe und schliesslich den ganzen Organismus
vergrössernd. Die Teilungen der Zellen aber gehen unter eigentümlichen
Veränderungen am Kerne — der Kernmitose — vor sich; die färbbare
Substanz des Kernes löst sich in Fäden auf, diese zerfallen in
Schleifen, welche sich um den Äquator anordnen, dann sich spalten und
den Polen zurücken. Und nun lehrt uns die Euglypha, dass dieselben
Phänomene auch auf der niedersten Stufe der Lebewesen sich abspielen,
wenn es sich um Zellteilung, beziehungsweise hier um Teilung des
Individuums handelt.
Die neuesten Forschungen haben uns die unumstössliche, wunderbare
Thatsache gelehrt, dass es sich bei der Befruchtung jedes vielzelligen
Organismus wesentlich um eine Kernverschmelzung handelt, Eikern und
Samenkern vereinigen sich und zwar sind es nur wenige Kernschleifen
des weiblichen und ebensoviele gleich grosse des männlichen Kernes,
welche dabei zusammentreten. In diesen Schleifen, winzigen, nur bei
stärkster Vergrösserung wahrnehmbaren Körpern, müssen also alle jene
tausenderlei Eigenschaften enthalten[S. 155] sein, welche sowohl vom Vater
wie von der Mutter auf den entstehenden Organismus vererbt werden. Es
ist also ein gar kostbares Material diese Kernsubstanz und es darf uns
nicht wundernehmen, dass ein so überaus feiner Mechanismus besteht, um
die Verteilung desselben zu bewerkstelligen. Dass auch schon bei den
einzelligen Wesen eine so genaue Verteilung der im Kerne enthaltenen
Potenzen stattfindet, lehrt uns abermals die Euglypha alveolata. Dabei
ist noch eine wichtige Thatsache hervorzuheben: Es zeigen sich nämlich
bei der Kernteilung der Euglypha gewisse Eigentümlichkeiten, welche
sich sonst nur bei den Kernteilungen der niedersten vielzelligen Tiere,
der niedersten Pflanzen und der Eier der höheren Tiere vorfinden[67 m].
Diese Ähnlichkeit im wichtigsten Lebensprozess dieser Zellen lehrt
uns mit unzweifelhafter Sicherheit, wie nahe die niedersten Pflanzen
und Tiere den Urtieren noch stehen und wie die höheren Tiere in ihrer
ersten Entwickelungsstufe im Ei ihren früheren Zusammenhang mit den
Urorganismen, resp. ihre Abstammung von diesen verraten.
Die Befruchtung ist, wie gesagt, eine Vereinigung männlicher und
weiblicher Kernsubstanz, mit anderen Worten, es vermischen sich
dabei von zwei verschiedenen Individuen die Charaktere, denn diese
sind ja in der Kernsubstanz enthalten. Eine neuere Theorie sucht
darin, wie mir scheint, mit Recht das wesentliche Moment des
Befruchtungsvorganges[71]. Durch diese Vermischungen entstehen neue
Kombinationen von Eigenschaften und dieser bedarf die Natur, um die
Organismen den sich stets verändernden äusseren Lebensbedingungen
angepasst zu erhalten. Bei den einzelligen Organismen beruht
dieser Vermischungsprozess meistens auf einer Verschmelzung zweier
ganzer Individuen und nichts erläutert dies besser, als die oben
beschriebene Kopulation der Euglypha, wo zwei Tiere vollkommen in eins
zusammenfliessen.
Man hat die Urtiere Organismen ohne Organe genannt; denn sie sind im
stande alle diejenigen wesentlichen Funktionen zu verrichten, welche
bei höheren Tieren an einen oft sehr komplizierten Mechanismus gebunden
sind. Sie tasten und empfinden ohne Nervensystem, bewegen sich ohne
Muskulatur, fressen und verdauen[S. 156] ohne Magen und Darm, atmen und
besorgen den Stoffwechsel ohne Lunge und Niere. Ja die Euglypha lehrt
uns, dass sie sogar kompliziertere Lebensthätigkeiten bekunden, dass
sie ohne Gehirn und Nerven eine Art von Instinkt besitzen können, die
Baukunst. Die Schale ist ja nicht im ganzen ein Ausscheidungsprodukt,
sondern nur die Schalenplättchen werden ausgeschieden und diese
müssen, wie wir sahen, bei der Teilung sorgfältig Stück für Stück
zusammengefügt und kunstvoll so angeordnet werden, dass das Ganze
die richtige Form erhält. Dieselben uns ungeformt erscheinenden
Plasmateile, welche den Nahrungskörper ergreifen und hereinziehen,
wissen die Schalenplatten aus dem alten Gehäuse heraus und an ihren
richtigen Platz zu führen; keine geübte Hand könnte dies sorgfältiger
und sicherer thun. Dasselbe Plasma weiss sich der Plättchen auch
noch auf andere Weise zu bedienen, wenn es sich darum handelt, gegen
drohende Austrocknung eine schützende Cyste zu bilden.
Noch wunderbarer erscheint uns dieser Kunsttrieb bei den schon
mehrfach erwähnten Verwandten der Euglypha, welche ihre Gehäuse aus
Fremdkörpern aufbauen. Die zierlichen, mannigfach gestalteten Schalen
der Difflugia-Arten, deren einige unsere Figur 16 aufweist, bestehen
aus Sand-, meistens Quarzkörnchen. Diese Sandkörnchen liest die
Difflugia in ihrer Umgebung zusammen, zieht sie in ihren Körper hinein
und speichert sie da so lange auf, bis sie zur Teilung schreitet.
Diese verläuft ganz wie bei der Euglypha, nur dass die Difflugia die
schwierige Aufgabe hat, aus ganz unregelmässigem Material eine neue
Schale herzustellen, und doch bringt sie es so gut fertig, dass das
neue Gehäuse vollkommen dem alten gleicht. Man hat den sinnreichen
Versuch gemacht[68 d], solche Wurzelfüsser in kleine Aquarien zu
bringen, in welchen der Boden mit zerstossenen Glassplittern bedeckt
war, und siehe da, auch die Glasstückchen wurden aufgenommen und die
Tiere, welche sich teilten, erzeugten Schalen, welche den alten zwar
vollkommen gleich waren, aber statt aus Sand aus Glas bestanden.
Es giebt höhere Organismen, welche ganz denselben Bautrieb besitzen,
wie diese Difflugien, ich meine die Larven der Phryganiden,[S. 157] die in den
meisten stehenden und fliessenden Gewässern vorkommen. Auch sie bauen
köcherartige Gehäuse aus allerlei Fremdkörpern zusammen, gerade wie
die Wurzelfüsser, aber um zu demselben Ziele zu gelangen, gebrauchen
sie einen komplizierten physiologischen Mechanismus, der in ihrem
hochentwickelten Nervensystem, in ihrer Muskulatur, ihrem Hautskelett
u. s. w. besteht.
Wie ist es aber möglich, fragen wir uns, dass ein einzelliges Wesen
ohne jeglichen nervösen Organe zu so hohen Leistungen befähigt ist.
Setzt ein derartiger Kunsttrieb nicht psychische Fähigkeiten voraus und
wo haben diese ihren Sitz?
Ausgedehnte vorzügliche, „psycho-physiologische Protistenstudien“,
die erst vor kurzem veröffentlicht wurden[68 e], haben sich eingehend
mit der Beantwortung dieser Frage beschäftigt. Die Urtiere wurden
in ihrem Verhalten auf die verschiedenartigsten Reize, Licht-,
mechanische, akustische, chemische Reize u. s. f., untersucht und
zwar in unverletztem Zustande und auch nach künstlich beigebrachten
Verletzungen, Entfernung des Kerns etc. Das Resultat aller solcher
Versuche ist, dass von einem nervösen Zentrum im Protistenkörper
keine Rede sein kann, dass die nervöse Potenz eine diffuse ist; jedes
Protoplasma-Elementarteilchen ist ein selbständiges Zentrum und
hat seine eigene selbständige Psyche. Eine Psyche im höheren Sinne
existiert freilich noch nicht und alle Bewegungen der Urtiere müssen
als willenlose Reflexbewegungen aufgefasst werden. Das Scheinfüsschen
der Difflugia berührt ein Sandkörnchen, der Reiz veranlasst dasselbe
sich zusammenzuziehen und dabei wird es den Fremdkörper, falls er nicht
zu schwer ist, mit sich reissen. Auf diese Weise sammelt es willenlos
sein Schalenmaterial. Aber trotz dieser rein mechanischen Erklärung
bleibt der Vorgang wunderbar genug. Bedenken wir nur das eine, dass die
Reize, welche Nahrungskörper auf die Pseudopodien ausüben, diese in
derselben Weise reagieren lassen wie die Berührung eines Sandkörnchens,
dass aber nachher beim Schalenbau Nahrungspartikeln und Schalenmaterial
doch auseinandergehalten werden. Was also bei der Phryganidenlarve
tausende von Zellen, in der Arbeit sich[S. 158] teilend, erreichen, schafft
hier eine einzige Zelle in derselben Weise aus sich selbst.
Man lasse mich hier noch einen zwar etwas drastischen, aber belehrenden
Vergleich anstellen: Der Längsdurchmesser der Euglypha beträgt etwa
⁶⁄₁₀₀, der Querdurchmesser ³⁄₁₀₀ mm, somit erhalten wir für das
Tier einen Kubikinhalt von 54 oder rund 50 Millionstel Kubikmillimeter.
Nehmen wir nun als Kubikinhalt eines der lebenden Tierriesen, z. B.
eines Elefanten, etwa 3 Kubikmeter an, so wären dies 3 Milliarden
Kubikmillimeter, d. h. gegenüber den 50 Millionstel Kubikmillimetern
der Euglypha 60000 Milliarden mal mehr. Wir finden also bei der
Euglypha dieselben Lebensäusserungen, wenn auch sehr vereinfacht, wie
bei dem 60000 Milliarden mal grösseren Elefanten; wir finden bei der
Teilung der Euglypha im Kerne dieselben eigentümlichen Vorgänge, wie
im Kerne einer der Myriaden von Zellen, welche den Riesenleib des
Elefanten zusammensetzen. Ist das nicht ein überwältigender Beweis für
die Einheit der belebten Natur? Ich meine, es gehöre schon ein hoher
Grad von Blasiertheit dazu, wenn man gegenüber solchen Thatsachen nicht
immer wieder von bewunderndem Staunen ergriffen wird. Beschleicht uns
nicht dasselbe Gefühl von der Unendlichkeit der Natur, als wenn wir in
einer klaren Nacht zum Sternenhimmel aufsehen und uns sagen, dass alle
die Tausende von Fünkchen Welten sind, so gross und grösser als unsere
eigene?
So führt uns das Studium des Kleinen und des Kleinsten im Kleinen
zum Verständnis des Grossen und Grössten. Denn je weiter wir in den
Zusammenhang der Organismen einzudringen vermögen, je mehr wir alle
Erscheinungen auf gemeinsame Gesetze zurückführen können, desto
einheitlicher und damit desto grösser erscheint uns die Schöpfung. Die
Spezialisierung, welche sich in der heutigen Forschung so sehr geltend
macht, artet nicht in Spitzfindigkeit aus, sondern leitet zu grossen
Resultaten, sobald wir sie richtig anzuwenden wissen.
Man macht der heutigen Naturforschung so oft den Vorwurf, dass sie es
sei, welche den materialistischen Zug, der durch unsere Zeit geht,
verschuldet habe und befördere. Ich glaube dies nicht,[S. 159] sondern
finde vielmehr, dass gerade die heutige Naturauffassung, die bei allem,
was sie schafft, das Auge auf die Entstehung und Entwickelung des
Ganzen gerichtet hat, am wenigsten eines idealen Zuges entbehrt.
Dem heutigen Forscher, obgleich er das Wunder nicht mehr anerkennt,
ist die Empfindung für die Grossartigkeit der Natur nicht verloren
gegangen, nein, er muss dieser mit noch grösserer Bewunderung
gegenüberstehen, als seine Vorgänger, denen eine naivere Vorstellung
von der Schöpfung die eigentlich belebende Seite des Forschens
versagte. Es geht ein hoher, idealer Zug durch die Naturforschung in
unseren Tagen, und in der heutigen, weitausschauenden Richtung gelehrt,
muss sie ein wichtiges Moment für die Erziehung werden, nicht nur für
den Arzt, der ohne sie zum Handwerker herabsinkt, zum Spezialisten
im schlechten Sinne des Wortes, sondern für jeden, der auf Bildung
Anspruch macht.
Leider versündigen sich Unverstand und Missverstand gar zu oft an Natur
und Forschung. Zu öfteren Malen hat man in den periodischen Wahlkämpfen
die extremste Partei predigen hören, die Ziele der Sozialdemokratie
seien in der Natur begründet, die Descendenzlehre sei ihre Stütze!
Gerade das Gegenteil ist der Fall: Nichts von allgemeiner Gleichheit
gestattet der Kampf ums Dasein, das Recht des Stärkeren wird die Losung
sein, so lange die Erde Lebewesen trägt. Danach freilich müssen wir
Menschen streben, dass es bei uns kein Faustrecht sei, sondern ein
Geistesrecht!
Ist es nicht auch eine falsche Naturauffassung, die heute eine
weitverbreitete Richtung in der Kunst beherrscht, wo der Künstler
nur dadurch an die Natur sich anlehnen zu können glaubt, dass er
das Hässliche, oder zum mindesten das Langweilige und Nichtssagende
darstellt?
Sollen wir die Descendenzlehre daran schuld sein lassen, dass
Zola den Atavismus als erklärendes Prinzip für seine
Verbrecherromane herbeizieht? Vererben sich denn nur die Laster,
vererben sich denn nicht auch die schönen und edlen Eigenschaften im
Menschen nach denselben Gesetzen? Ist denn das Schöne nicht auch Natur?
Der Forscher soll an der Hand der nackten Thatsachen[S. 160] die Wahrheit
suchen und darf auch vor dem Widerlichen nicht zurückbeben. Auch der
Künstler soll Wahres schaffen, aber ihm ist ja die Wahl gelassen
in den unendlichen Schätzen der Natur und warum greift er dann das
Hässliche heraus? Wer im Sommer 1889 die grosse Internationale
Kunstausstellung in München besucht hat, den empfing gleich am Eingang
die bekannte grosse Marmorgruppe von Fremiet, der Gorilla, der
ein Weib entführt. Was kann es Widerlicheres geben als diesen Affen
und diese Situation! Mögen die Motive, welche den Künstler zu diesem
Werke geführt, sein, welche sie wollen, das ist kein Vorwurf für ein
wahres Kunstwerk; ihm gebührt die grosse goldene Medaille nicht! Hat
dazu der Künstler sein grosses Talent, hat er dazu seine wunderbare
technische Fertigkeit, hat er dazu Zeit, Arbeit und Geld angewendet, so
ist dies eine Verirrung! Wenn aber der Forscher denselben hässlichen
Gorilla darstellt und beschreibt und daran nachweist, wie er als
Glied einer langen Kette von Organismen sich einfügt, oder wie er
als letzter Rest einer grossen Reihe von Vorfahren auf unserer Erde
lebt, wie hier Eigentümlichkeiten seines Körperbaues, dort Äusserungen
seines Intellekts zu wichtigen Vergleichen anregen, dann entkleidet er
das Tier seiner Hässlichkeit, statt angeekelt uns abzuwenden, kehren
wir uns ihm mit Interesse zu; da steht der Forscher in der idealen
Auffassung über dem Künstler. Unverstand und Geistlosigkeit müssen
freilich überall auf falsche Wege führen; wo aber der Verstand die
Natur zu erkennen strebt, da giebt es keinen falschen „Naturalismus“
und keinen „Materialismus“, wo der Geist die Materie belebt.
[S. 161]
Litteratur.
[64]August Johann Rösel von Rosenhof: Der monatlich
herausgegebenen Insecten-Belustigung Dritter Theil. Nürnberg 1755.
[65] S. darüber Bütschli: Über die Struktur des
Protoplasmas in: Verhandlungen d. naturhist. med. Ver. zu Heidelberg.
N. F. IV. Bd. 3. Heft. Sitzung vom 3. Mai und 7. Juni 1889.
[66] S. hierüber Bronn: Classen und Ordnungen des
Tierreichs; Bütschli: Die Protozoen. I. Abt. p. 228 u. f.
Heidelberg und Leipzig 1880–82.
[67] Die hauptsächlichste Litteratur über Euglypha ist
folgende:
a) Dujardin: Histoire naturelle des Zoophytes infusoires. Paris
1841.
b) Ehrenberg: Verschiedene Schriften 1841–1872. Übersicht der
seit 1847 fortgesetzten Untersuchungen über das von der Atmosphäre
unsichtbar getragene Leben in: Abh. der Berliner Akad. aus d. J. 1871.
Berlin 1872.
c) Perty: Zur Kenntniss kleinster Lebensformen nach Bau,
Funktionen, Systematik, mit Spezialverzeichniss der in der Schweiz
vorkommenden. Bern 1852.
d) Carter: On Fresh-water Rhizopoda of England and India in:
Annals and Magazine of Natural History. London 1884.
e) Hertwig u. Lesser: Über Rhizopoden und denselben nahestehende
Organismen in: Archiv für mikroskopische Anatomie. Bd. X. Suppl. 1874.
f) F. E. Schulze: Rhizopodenstudien III in: Archiv f. mikr.
Anat. Bd. XI. 1875.
g) Leidy: Fresh-water Rhizopoda of North-America in: United
States geological survey of the territories. Vol. XII. 1879.
h) Gruber: Der Theilungsvorgang bei Euglypha alveolata in:
Zeitschr. für wissenschaftliche Zoologie. Bd. 35. 1881.
i) Gruber: Die Theilung der monothalamen Rhizopoden in:
Zeitschr. f. wiss. Zoologie. Bd. 36. 1882.
k) Gruber: Kleinere Mitteilungen über Protozoenstudien in:
Berichte der naturforschenden Gesellschaft zu Freiburg i. Br. Bd. II.
Heft 3. 1886.
l) Blochmann: Zur Kenntnis der Fortpflanzung von Euglypha
alveolata in: Morphologisches Jahrbuch. Bd. 13. 1887.
m) Schewiakoff: Über die karyokinetische Kernteilung von
Euglypha alveolata in: Morphol. Jahrb. Bd. 13. 1887.
[S. 162]
[68] Die hauptsächlichsten Arbeiten über künstliche Teilung
bei Urtieren sind:
a) Nussbaum: Über die Teilbarkeit der lebendigen Materie. I. Die
spontane u. künstliche Teilung der Infusorien in: Archiv f. mikr. Anat.
Bd. 26.
b) Gruber: Beiträge zur Kenntnis der Physiologie u. Biologie der
Protozoen in: Ber. d. naturf. Ges. zu Freiburg i. Br. Bd. I. 1886.
c) Balbiani: Recherches expérimentales sur la mérotomie des
infusoires ciliés in: Recueil zoologique. Tome 5. Fasc. 1. Genève-Bâle
1888.
f) Hofer: Experimentelle Untersuchungen über den Einfluss
des Kerns auf das Protoplasma in: Jenaische Zeitschr. für
Naturwissenschaft. N. F. Bd. 17. 1889.
[69] Ich habe in den Torfmooren des Schwarzwaldes, oberhalb
des Höllenthales, fast alle die Rhizopoden nachgewiesen, die Leidy[67 g]
in den Gewässern Nordamerikas aufgefunden hat (s. darüber das
Grossherzogtum Baden, Lieferg. 2. Die Tierwelt. pg. 136. Karlsruhe
1883).
[70] Über die „passive Migration“ s. O. Zacharias:
Bericht über eine zoologische Exkursion an die Kraterseen der Eifel in:
Biologisches Centralblatt. Bd. 9. No. 2–4. Erlangen 1889.
[71]Weismann: Die Bedeutung der sexuellen
Fortpflanzung für die Selektionstheorie. Jena 1886. (S. auch
Weismann: Bemerkungen zu einigen Tagesproblemen. Biol.
Centralblatt. Bd. X. No. 1. 1890).
[S. 163]
Die Flagellaten
(Geisselträger).
Von Dr. W. Migula in Karlsruhe.
[S. 165]
Zu den niedersten Organismen, auf der Grenze zwischen Tier und Pflanze
stehend, gehört die interessante Abteilung der Flagellaten. Mit
ihren nächsten Verwandten, den Sarkodinen, Sporozoen und Infusorien,
bilden sie die Gruppe der Protozoen, aber wenn sie auch mit
jenen zusammengefasst werden und vielfache Berührungspunkte mit ihnen
und mit gewissen Algen zeigen, so sind sie in ihren Eigenschaften
doch so scharf charakterisiert, dass sie, von schwierigen Fällen
abgesehen, leicht zu erkennen sind. Vor allem zeichnen sie sich durch
den Besitz von Geisseln aus. Es sind dies sehr feine peitschenförmige
Plasmafäden, durch deren schwimmende oder schlagende Bewegung sie
sich fortbewegen. Hierdurch unterscheiden sie sich auch von den
drei anderen genannten Abteilungen der Protozoen, denen derartige
Bewegungsorgane nicht zukommen. Denn die Infusorien bewegen sich mit
Hilfe von Cilien, haarförmigen, viel kürzeren Gebilden, welche in
grosser Anzahl vorhanden sind, während die Geisseln oft die Länge des
Körpers übertreffen und stets nur in geringer Anzahl — 1, 2, 3, 4
oder 5 — auftreten. Die Sarkodinen mit den Rhizopoden (vergl. Kapitel
4), Heliozoen und Radiolarien dagegen bewegen sich im wesentlichen
durch ein eigentümliches mit fortwährender Formveränderung verbundenes
Hinfliessen des Körpers, und die Sporozoen haben in ihrem erwachsenen
Zustande überhaupt keine Bewegungsorgane. Der Körper der Flagellaten
ist sehr verschiedenartig gestaltet, stets einzellig, oft nackt,
d. h. ohne feste Membran[S. 166] oder Zellwand. Dagegen finden sich bei
einzelnen wieder sehr starke feste Hüllen um den zarten Zellleib,
welche mitunter sehr zierlich gezeichnet sind. Der Zellinhalt ist
entweder farblos oder bräunlich oder chlorophyllgrün gefärbt und der
Farbstoff ist dann gewöhnlich an plattenförmige Träger gebunden. Ein
Zellkern ist wohl stets vorhanden, daneben oft noch andere feste
Plasmakörperchen von zumteil noch unbekannter Bedeutung. Ebenso finden
sich im Innern des Plasmas Hohlräume, welche mit Zellsaft gefüllt sind
und von denen häufig zwei in bestimmten Zwischenräumen abwechselnd
sich zusammenziehen und allmählich wieder wachsen; man hat sie
pulsierende Vakuolen genannt. In der Regel findet sich auch noch ein
(seltener zwei) roter kleiner Punkt in der Zelle, der Augenfleck oder
Pigmentfleck.
Die Geisseln stehen in geringer Zahl zusammen an einem Punkte des
Körpers, gewöhnlich am Vorderende, sie sind gleich oder ungleich lang,
bei einigen von einem schwer sichtbaren feinen Mantel umgeben. Bei
der Fortbewegung wird das Wasser mit der ganzen Fläche der Geissel
gepeitscht und diese dann, ähnlich wie beim Schwimmen, zurückgezogen.
Die Fortpflanzung geschieht nur bei einigen — soweit bisher bekannt
— auf geschlechtlichem Wege durch Kopulation zweier Individuen,
oder wie bei Volvox durch die Befruchtung einer ruhenden Eizelle
durch ein bewegliches Spermatozoid. Häufig ist eine Vermehrung durch
Querteilung oder durch Längsteilung beobachtet. Vielen, vielleicht
allen Flagellaten kommt die Fähigkeit zu, bei Eintritt ungünstiger
Verhältnisse in einen Ruhezustand überzugehen, indem sie sich
encystieren, d. h. unter Zusammenziehung des Körpers zu rundlichen
Massen sich mit einer festen Membran umgeben. Sie können in diesem
Zustande längere Austrocknung vertragen und zugleich ist es ein
Mittel zur Verbreitung der Flagellaten. Denn wenn dieselben in dem
Ruhezustande auf dem Boden ausgetrockneter Pfützen liegen, so genügt
ein Windhauch, um sie emporzuheben und als feine Staubkörnchen
fortzuführen. So gelangen sie vielleicht in einen Wassertümpel, wo sie
ihre schützende Hülle sprengen und zu neuem Leben erwachen.
[S. 167]
Oft bleiben die Zellen nach der Teilung in mehr oder weniger festem
Zusammenhang zu verschiedenartigen Kolonien vereinigt. Manche Arten
leben parasitisch in anderen Organismen, viele bewohnen fauliges
Wasser, manche treten nur in ganz reinen Wässern auf. Nach zwei
Richtungen hin zeigen sie sehr enge Beziehungen zum Pflanzenreich: zu
den Spaltpflanzen durch ihre einfachsten Vertreter, die Monadinen, und
zu den einzelligen Grünalgen, den Protococcoiden, durch die Volvocineen
und Chlamydomonadinen.
Dies sind im Umriss die allgemeinen Eigenschaften einer Gruppe von
Organismen, welche von den Botanikern und Zoologen als strittiges
Gebiet betrachtet werden und welche man deshalb, wenigstens teilweise,
sowohl in Lehrbüchern der Zoologie wie in solchen der Botanik behandelt
findet. Bei der Mannigfaltigkeit und dem Reichtum der grossen Gruppe,
bei der Verschiedenheit der Entwickelung selbst nahe verwandter Formen
und — last not least — bei der geringen Kenntnis, welche wir trotz
vieler und vorzüglicher Arbeiten von den Flagellaten besitzen, mag
es genügen, wenn wir einige interessante Vertreter dieser Abteilung
herausgreifen und einer eingehenderen Beschreibung unterziehen.
Wer sich für diese schwierigen und nur mit den besten Mikroskopen
erfolgreich zu untersuchenden Organismen interessiert, sei auf die
Litteraturangabe am Schluss verwiesen.
Zu den am höchsten entwickelten Flagellaten sind unbedingt die
Volvocineen zu rechnen, welche in etwa sieben Arten in unseren
Gewässern vorkommen. Wir wollen uns vorzugsweise an die Gattung
Volvox und an die häufigere der beiden Arten, an Volvox aureus
(Volvox minor Stein) halten.
Fig. 34.
Volvocineen. 1 Volvox aureus Ehrbg.: a Kolonie mit fünf
Tochterkugeln (Parthenogonidien) und drei bereits befruchteten Eizellen
(Oosphaeren), b Kolonie mit zwei Parthenogonidien, elf unbefruchteten
Eizellen, zwei männlichen Kolonien, c Teil einer Kugel, stärker
vergrössert, mit einer Zelle, welche sich teilt und den Anfang einer
Tochterkolonie darstellt, d reife Oosphaere, e Teilungszustand
einer sehr jungen Volvoxkolonie; die Zellen schliessen noch dicht zusammen,
f männliche Kolonie mit Antheridien, g Antheridien von der Seite,
h Spermatozoiden — 2 Pandorina Morum — 3 Gonium pectorale
(1 a und b schwach, alle übrigen Figuren stark, zumteil sehr
stark vergrössert).
Das Kugeltierchen, wie es von Leeuwenhoek, der es schon
vor 200 Jahren beobachtet hat, genannt wurde, oder unser Volvox
bildet kugelige oder eiförmige Kolonien von etwa ½–¾ mm
Durchmesser, doch kommen sowohl grössere wie kleinere vor (vergl.
hier und in der folgenden Darstellung Figur 34 S. 168 und Erklärung).
Es sind Hohlkugeln, welche auf der Oberfläche eine grosse Anzahl
kleiner grüner Zellen in regelmässigen Abständen tragen. Die Zellen
werden durch eine farblose Gallertmasse zusammengehalten, welche[S. 168]
aus den verschleimenden Membranen der Einzelzellen besteht und daher
nur eine wenige Mikromillimeter (= ¹⁄₁₀₀₀ mm) dicke Schale
bilden, während der Inhalt der Kugel aus Wasser besteht. Die Zellen
sind etwa ⁵⁄₁₀₀₀ mm dick, etwas eiförmig gestaltet, mit dem
grösseren Durchmesser dem Kugelradius parallel. Es lässt sich in
ihrem Innern wenigstens eine pulsierende Vakuole (vielleicht zwei)
erkennen, ein sehr kleiner, aber deutlicher roter Pigmentfleck,
eine chlorophyllgrüne, die Zellwand zu etwa ⅔ auskleidende Platte
und darunter ein oft undeutlicher Zellkern nebst einigen kleineren
Körnchen. Jede Zelle trägt an ihrem nach aussen stehenden Ende
zwei Geisseln, welche die ganze Kolonie wie mit einem Wimperkleid
umgeben erscheinen lassen und durch deren Schwingung eine[S. 169] drehende
Fortbewegung der ganzen Kolonie erzeugt wird. Zwischen den einzelnen
Zellen laufen in der Gallertmasse noch Verbindungsfäden, welche mehr
oder minder regelmässige Dreiecke auf der Kugeloberfläche bilden. Die
Zahl der Zellen einer solchen Volvoxkugel ist sehr verschieden, sie
schwankt zwischen 200 bis 2000, kann aber die Zahlen nach unten und
oben auch noch wesentlich überschreiten.
Diese eben beschriebenen Zellen besitzen einen rein vegetativen
Charakter, d. h. sie dienen nur der Aufnahme und Verarbeitung der
Nährstoffe und allen den Lebensthätigkeiten des Organismus, welche
nicht mit der Fortpflanzung zusammenhängen. Für sich allein, auch
losgetrennt von der Kolonie, würden sie zwar noch eine Zeit lang
ihr Leben fristen, aber schliesslich zu Grunde gehen, ohne für die
Erhaltung der Art zu sorgen. Diese Funktion übernehmen ganz anders
gestaltete Zellen, welche übrigens aus Zellen hervorgehen, welche
den vegetativen ursprünglich gleich gestaltet sind. Aber um diese
Verhältnisse zu übersehen, müssen wir auf die Entstehung der Zellen
etwas näher eingehen. Bei den Volvocineen finden sich Dauerzustände
(Sporen), welche den Winter über ruhen und im Frühjahr sich zu neuen
Volvoxkugeln entwickeln. Eine solche Spore (1 d) besitzt eine
ziemlich dicke farblose Membran und bei völliger Reife gelbroten
körnigen, mit Reservestoffen erfüllten Inhalt. Die erste Veränderung,
welche im Frühjahr in ihr vorgeht, ist, dass die gelbrote Farbe des
Zellinhaltes einer grünen Platz macht; die Zelle nimmt dann Wasser auf
und sprengt die harte Membran. Vorher teilt sich der Zellinhalt in
zwei und diese weiter in vier, diese wieder in acht Zellen; letztere
entwickeln sich zu Schwärmzellen, welche den Ausgangspunkt für die
Entwickelung der Volvoxkolonien bilden. Die Schwärmzellen teilen
sich in einen vielzelligen Körper unter stetem Wachstum, bleiben
aber dicht zusammen. Erst wenn die Zellen sich nicht mehr teilen,
beginnt eine Differenzierung sich bemerkbar zu machen; einige Zellen
sind etwas grösser und mit den anderen, wenn sie auseinanderrücken,
durch zahlreichere Fäden verbunden. Sobald nämlich die Zellteilungen
aufgehört haben, fangen die Membranen der einzelnen[S. 170] Zellen an, zu
quellen, wodurch diese auseinandergedrängt werden. Nun beginnen
einige wenige von Anfang an etwas grössere, aber ebenfalls in den
Gallertmantel der Kugeloberfläche eingebettete Zellen heranzuwachsen
und sich zu teilen in ganz gleicher Weise, wie das eben beschrieben
wurde. Je grösser diese Zellkomplexe werden, desto mehr rücken sie in
das Innere der Kugel, lösen sich schliesslich ganz von der Oberfläche
los und schwimmen frei in dem Hohlraum umher. In dem einen Falle
gestalten sich aus ihnen Kugeln, welche den Mutterkugeln vollkommen
gleichen und auch ihrerseits wieder junge Kugeln im Innern entwickeln,
so dass man thatsächlich sehr oft Grossmutter, Mutter und Enkel in
einander eingekapselt sieht. Die Kolonien schwimmen frei im Innern der
Kugel umher, bis sie an dem einen Pol der Mutterkugel ausschlüpfen
und ein selbständiges Dasein beginnen. Alle Kugeln, welche auf
diese Weise entstehen und sich aus Zellen der Kugeloberfläche ohne
Befruchtung entwickeln, hat man Parthenogonidien genannt. Ihre Zahl
innerhalb einer Mutterkugel ist sehr verschieden: 2–12, selten mehr;
neben ihnen können aber auch noch Zellen oder Zellfamilien ganz
anderer Natur auftreten und mit der Beschreibung derselben betreten
wir das Gebiet der geschlechtlichen Fortpflanzung, welche bei Volvox
sehr hoch ausgebildet ist. Zunächst entwickeln sich einige der
Zellen der Mutterkugel, ohne sich weiter zu teilen, zu sehr grossen
weiblichen Geschlechtszellen, den Oogonien, welche dunkler grün sind
und ein körniges Plasma zeigen. Sie sind leicht mit sehr jugendlichen
Parthenogonidien zu verwechseln, aber durch den Mangel an Teilungen von
diesen unterschieden. Schliesslich lösen sie sich ebenfalls aus dem
Zellverbande heraus und treten in den Hohlraum der Kugel. Noch andere
Zellen der Mutterkugel erfahren zwar ähnlich den Parthenogonidien
Teilungen, aber sie entwickeln in der Regel keine neuen
Parthenogonidien; es teilen sich vielmehr nach dem Auseinanderrücken
der Zellen einige derselben, nachdem sie stärker herangewachsen sind,
in ein Bündel länglicher Zellen, welche jedoch zunächst von der
gemeinsamen Hülle ihrer Mutterzelle umschlossen bleiben, mit welcher
sie auch aus dem Zellverbande austreten. Die Hülle ist nach der[S. 171] einen
Seite haubenförmig erweitert und man sieht in dieser Erweiterung eine
Anzahl sehr feiner Fäden in Bewegung, welche man als die Geisseln
der bündelförmig gruppierten Zellen erkennt. Diese Zellen lösen sich
schliesslich von einander und suchen die Oogonien auf; sie tragen zwei
lange Geisseln am Vorderende ihres länglich-eiförmigen, gebogenen und
membranlosen Körpers und an der Geisselbasis einen deutlichen roten
Pigmentfleck. Ihre Färbung ist ein mattes Ockergelb, sie weichen also
auch hierin nicht unbedeutend von den übrigen Zellen des Volvox ab.
Sobald diese Spermatozoiden frei sind, treiben sie sich umher und
suchen zu den in derselben oder in anderen Kugeln eingeschlossenen
Oogonien zu gelangen, an welche sie sich unter eigentümlich bohrenden
Bewegungen anlegen und wahrscheinlich mit ihnen verschmelzen. Nach
dieser Befruchtung umgiebt sich die Eizelle mit einer dicken Membran
und macht eine Ruheperiode durch, nach welcher sie sich wieder in der
beschriebenen Weise zu Volvoxkugeln entwickelt.
Dies sind in groben Zügen Bau und Entwickelungsgeschichte unseres
Volvox, welche jedoch eine solche Fülle von interessanten Einzelheiten
bieten, auf die hier nicht näher eingegangen werden konnte, dass dieses
Wesen immer wieder Gegenstand eingehender Untersuchungen ist. So finden
wir namentlich in dem Auftreten geschlechtlicher und ungeschlechtlicher
Kolonien eine grosse Variabilität. Es kommen Volvoxkugeln vor, welche
nur Parthenogonidien enthalten, solche, welche neben diesen auch
Oogonien, solche, welche letztere allein oder mit Androgonidien, d. h.
neben den vegetativen ausschliesslich männliche Zellen enthalten, und
solche, welche letztere allein oder mit Parthenogonidien zusammen in
sich tragen. Noch komplizierter wird aber die Sache dadurch, dass die
Parthenogonidien selbst wieder alle diese Kolonienformen in gleicher
Weise in sich bergen können, so dass hierdurch ausserordentlich
verwickelte Verhältnisse entstehen. Auch in Bezug auf die männlichen
Kolonien können sehr grosse Verschiedenheiten herrschen, doch würde
eine Erörterung derselben an dieser Stelle zu weit führen.
[S. 172]
Der grössere Volvox globator unterscheidet sich wesentlich
durch die nicht runden, sondern mehr dreieckigen vegetativen Zellen
und durch die stachelige Sporenmembran. Beide Arten kommen meist
unter einander vermischt, selten rein vor. Sie lieben kleine, lichte,
dünn mit Binsen oder Rohr bestandene Sumpflöcher, sind aber in ihrem
Auftreten sehr unbeständig und wählerisch. An Orten, wo man sie Jahr
für Jahr regelmässig gefunden hat, bleiben sie plötzlich aus und
erscheinen auch nicht wieder, während sie sich anderseits wieder
oft dort in Masse ansiedeln, wo man sie kaum vermuten dürfte. So
trat plötzlich in einem Aquarium, welches von mir mit einer Nitella
bepflanzt war, seit zwei Jahren im Zimmer stand und mit Leitungswasser
gefüllt wurde, Volvox aureus in solcher Menge auf, dass man
mit jedem Tropfen Kolonien herausholen konnte. Aber ebenso verschwand
er daraus und trat nicht wieder auf, ohne dass irgend welche Tiere
vorhanden waren, die seine Vernichtung hätten herbeiführen können.
Er bildet nämlich eine Lieblingsspeise kleiner Crustaceen, besonders
der Cypris- und Cyclopsarten, aber auch Kaulquappen und Mückenlarven
stellen ihm begierig nach. Wo derartige Organismen in grösserer Zahl
auftreten, verschwindet der Volvox auch im Freien sehr bald und kann
vollständig vernichtet werden, da auch die schon befruchteten Eizellen,
die Sporen, gefressen werden. Im Zimmer lässt sich Volvox nicht leicht
züchten; er zeigt manchmal ganz plötzlich eine Neigung einzugehen,
ohne dass man irgend einen Grund dafür finden könnte. Am besten hält
er sich in grossen Gefässen, welche nur von oben Licht erhalten und
eventuell an den Seitenwänden durch schwarzes Papier undurchsichtig
gemacht werden müssen. Hier kann man einige Wasserlinsen, am besten
Lemna polyrrhiza, hineinthun, dass sie etwa ⅓ der Oberfläche,
nicht mehr, bedecken. Die Volvox-Kolonien gewähren dem Beobachter,
auch wenn er nur eine Lupe anwendet, so viel Genuss, dass es sich wohl
lohnt, dieselben zur leichtern und bessern Beobachtung im Zimmer zu
kultivieren.
Die Ernährung des Volvox geschieht auf rein pflanzliche Weise; die
einzelnen Zellen haben vermöge ihres Chlorophyllgehaltes die Fähigkeit
zu assimilieren, d. h. aus anorganischen Stoffen, aus der[S. 173] Kohlensäure
und dem Sauerstoff der im Wasser gelösten Luft, aus dem Wasser und
seinen Bestandteilen an Salzen ihren Organismus aufzubauen. Es kommt
ihnen als Produkt der Assimilation die Stärke im Inhalt der Zellen zu,
und auch die Art der Befruchtung und Keimung der Sporen hat Volvox mit
vielen zweifelhaften Algen gemein. Dagegen nähert sich der Organismus
den Flagellaten besonders dadurch, dass die Beweglichkeit der Zellen
während des ganzen eigentlichen Lebens erhalten bleibt. Wir sehen in
den Volvocineen eben Wesen, welche mit dem gleichen Rechte als Tiere
und als Pflanzen angesehen werden können und welche sowohl für den
Botaniker wie für den Zoologen von gleicher Wichtigkeit sind, weil sie
die Gemeinsamkeit des Organischen von neuem bestätigen.
Zu den Volvocineen gehören noch folgende vier durch grosse Zierlichkeit
ausgezeichnete Gattungen:
Eudorina mit der einzigen Art E. elegans. Die Familie
wird gebildet aus 16 oder 32 Zellen, welche nicht bloss vegetativ
sind, sondern auch unter Umständen geschlechtliche Funktionen besitzen
können. Sie sind weit grösser als bei Volvox, etwa ²⁰⁄₁₀₀₀ mm
dick, während die ganze Kolonie im Durchschnitt nur ⅒ mm
im Durchmesser hat. Sonst sind die Zellen denen von Volvox sehr
ähnlich: mit pulsierenden Vakuolen, Pigmentfleck und je zwei Geisseln.
Die ungeschlechtliche Vermehrung erfolgt durch Teilung jeder Zelle
in 16 oder 32 Zellen, welche wieder eine neue Kolonie bilden; die
geschlechtliche durch Oogonien, welche von Spermatozoiden befruchtet
werden. Zu Oogonien bilden sich sämtliche Zellen um, mit Ausnahme
derjenigen vier, welche an dem bei der Bewegung nach vorn gerichteten
Teil stehen. Diese werden zu Antheridien, in denen sich je 64 hellgrüne
Spermatozoiden entwickeln.
Pandorina mit der einzigen Art P. Morum (Fig. 34, 3).
Die Familien haben eine rundlich eiförmige Gestalt und bestehen
aus 16, 32 oder 64 dicht zusammen liegenden Zellen von beinahe
herzförmiger Gestalt und von einem gemeinsamen Gallertmantel umgeben.
Die ungeschlechtliche Vermehrung geht ganz in derselben[S. 174] Weise vor
sich, wie bei Eudorina; die geschlechtliche weicht dagegen in sehr
erheblicher Weise ab. Aus jeder Zelle entstehen nämlich in der Regel
acht Schwärmzellen, welche ausschlüpfen und umherschwärmen, bis
sie auf gleiche Zellen von anderen Pandorinakolonien treffen. Mit
diesen kopulieren sie dann (zu zweien), indem der Inhalt der beiden
Schwärmzellen vollständig verschmilzt. Die aus der Kopulation beider
entstehende Zelle, die Zygote, macht dann eine Ruheperiode durch und
teilt sich bei der Keimung in 1–3 grosse Schwärmzellen, welche wieder
zu Zellfamilien werden.
Ähnlich verhält sich die Entwickelung der in den hellen
Wasseransammlungen in Felsplatten etc. vorkommende Stephanosphaera
pluvialis mit acht lang-spindelförmigen Zellen. Eine andere
Gattung (Gonium) bildet kleine Zelltäfelchen und zwar G.
sociale aus vier, G. pectorale aus sechzehn, seltener acht
oder vier Zellen bestehend. Die Membranen oder Einzelzellen sind bei
ausgewachsenen Exemplaren stark gequollen und durch gegenseitige
Berührung eckig; sie lassen sechzehn dreieckige Räume und einen
viereckigen zwischen sich frei. Um das ganze Täfelchen befindet
sich eine sehr dünnflüssige Gallerthülle, aus welcher die Geisseln
hervorragen. Die ungeschlechtliche Fortpflanzung findet in derselben
Weise wie bei Eudorina statt; eine geschlechtliche ist bisher nicht
beobachtet.
Wie wir im vorstehenden gesehen haben, findet innerhalb ein und
derselben Familie eine solche Verschiedenheit in der geschlechtlichen
Fortpflanzung statt, dass sich hierauf allein ein System nicht
begründen liesse, wollte man nicht sonst eng verwandte Formen ganz
auseinanderreissen. Es wird aber zugleich auch einleuchtend sein, dass
bei dem Umfang und dem Zweck dieser Abhandlung ein genaueres Eingehen
auf die Entwickelungsgeschichte unmöglich ist, und dass wir nur noch
kurz dieselbe berühren können. Wichtiger scheint es dagegen, noch die
Lebensweise und die Eigenschaften einiger Organismen zu betrachten,
welche hierin von den oben beschriebenen Volvocineen abweichen.
Die Volvocineen gehören zu den kolonienbildenden Flagellaten, wie
wir gesehen haben; wesentlich gleichgestaltete, aber einzeln lebende
Organismen sind die Chlamydomonadinen, deren geschlechtliche[S. 175]
Fortpflanzung mit der von Pandorina Ähnlichkeit hat. Unter ihnen
ist besonders Chlamydococcus pluvialis bekannt, welcher nach
heftigen Gewitterregen im Gebirge klares Wasser der Felsklüfte oft
vollständig rot oder grün färbt und auch manchem Touristen eine
bekannte Erscheinung in diesem Zustande sein dürfte. Auch diese Familie
kann mit dem gleichen Rechte zu den Algen gezogen werden; ja sie steht
diesen sogar noch näher als die erstbeschriebene.
Von wesentlich anderem Bau ist ein Organismus, welcher Schmutzlachen,
Pfützen auf lehmigen Wegen etc. oft intensiv grün färbt, die Euglena
viridis, welche wir als Vertreter einer eigenen Gruppe hier näher
schildern wollen. Sie besitzt einen mehr oder weniger langgestreckten
spindelförmigen Körper mit einer langen Geissel am Vorderende, zwei
pulsierende Vakuolen und einen deutlichen roten Augenfleck. Bis auf
einen Teil am Vorderende enthält sie zahlreiche kleine scheibenförmige
Chlorophyllkörnchen direkt unter der Oberfläche. Ausserdem finden wir
hier an der Geisselbasis eine Mundöffnung, welche zur Aufnahme fester
Nahrung dient. Diese und ähnliche Formen, namentlich aber die farblosen
— denn es giebt in dieser Gattung auch chlorophyllfreie —, leben von
Bakterien oder anderen sehr kleinen Organismen, welche sie mit der
Geissel gegen die Mundöffnung schleudern. Wir haben hier Organismen von
zweifellos tierischem Bau und Leben, welche aber dennoch Chlorophyll
enthalten und also wie die Pflanzen auch aus anorganischen Stoffen
Nahrung ziehen und bilden können. Wir erkennen dies auch noch daran,
dass man bei manchen thatsächlich Stärke gefunden hat, ein Produkt,
welches bekanntlich bei der Assimilation der Pflanzen entsteht. Sie
tritt dann in einer Form auf, welche es zweifellos macht, dass sie in
dem Organismus selbst entstanden und nicht etwa von aussen mit der
Nahrung aufgenommen worden ist.
Noch in einem andern Punkte bieten uns die Euglenen interessante
Beobachtungsobjekte dar; ihr Körper ist metabolisch, d. h. er besitzt
die Fähigkeit, durch Kontraktion seine Gestalt zu verändern und
die gewöhnlich lang-spindelförmigen Wesen nehmen oft Kugelform an.
Gewöhnlich werfen die Euglenen die Geissel ab, ehe sie in dieses
Stadium eintreten; ihre Bewegungen werden dann[S. 176] kriechend und sind mit
fortwährender Gestaltveränderung verbunden. Ihre Fortpflanzung erfolgt
in einem Ruhezustande durch Teilung, auch sind Zustände bekannt, welche
eine längere Austrocknung ertragen können, ohne dabei zu Grunde zu
gehen. (Vergl. Fig. 35, 1 a–c.)
Fig. 35.
1 Euglena viridis, a im schwimmenden geisseltragenden, b
und c im kriechenden geissellosen Zustand, A Augenfleck, m
Mundstelle — 2 Anthophysa vegetans, a ein Zweig mit fünf Köpfchen,
b ein stark vergrössertes Einzelindividuum — 3 Dinobryum
Sertularia, a kleine Kolonie, b stärker vergrössertes
Einzelindividuum — 4 Mastigamoeba, a in amoeboidem, b in
flagellatenartigem Zustand — 5 Bodo caudatus — 6 Ceratium
Hirundinella.
Sämtlich stark vergrössert.
Nahe verwandt der Gattung Euglena ist die Gattung Colacium, jedoch
durch ihre Lebensweise von jener verschieden. Die Colacien setzen
sich nämlich auf kleinen Wassertieren fest, entwickeln einen
Gallertmantel und Gallertfuss, verlieren die Geissel und teilen sich.
Die Tochterzellen trennen sich jedoch nicht von einander, sondern
bleiben zu Kolonien vereinigt; sie sind eigentlich nur kolonienbildende
Euglenen.
Ebenfalls Kolonien bildend, aber von anderem Bau ist die Gattung
Anthophysa. Sie gehört zu der Gruppe der Heteromonadina,[S. 177] welche
sich durch die Entwickelung von einer Hauptgeissel und einer oder
zwei kürzeren Nebengeisseln auszeichnet. Bei der vorliegenden
Art, Anthophysa vegetans, ist nur eine Nebengeissel
vorhanden. Die einzelnen Organismen sind klein, etwa ¹⁄₃₀ mm
lang, kegelförmig, am Vorderende plötzlich abgestutzt und mit einem
kurzen, spitzen Anhängsel, nach unten zu in einen spitzen Stiel
verschmälert. Sie besitzen eine kontraktile Vakuole, einen Zellkern
und eine Mundöffnung zur Aufnahme fester Nahrung, ein Pigmentfleck
fehlt, ebenso Chromatophoren. Eine Anzahl dieser Einzelindividuen
sitzen kopfförmig auf einem Gallertstiel auf, welcher meist wiederholt
gabelig verzweigt ist und an jedem Ende ein derartiges Köpfchen trägt.
Diese Köpfchen können sich loslösen und umherschwärmen; sie zerfallen
dabei oft in die einzelnen Zellen, so dass schliesslich nur die reich
verzweigten Gallertstiele zurückbleiben. Diese sind in der Jugend
rein weiss und durchsichtig; im Alter färben sie sich durch Aufnahme
von Ocker gelb bis dunkelbraun. Der Organismus scheint namentlich in
eisenhaltigem Wasser häufig zu sein, wo er mit einer Alge, Lyngbya
ochracea (Ockeralge), zusammen vorkommt. Die alten, stark mit Ocker
durchtränkten und inkrustierten Stiele der Flagellaten und die ebenso
aussehenden Fäden der Alge sind dann oft nur zu unterscheiden, wenn
man durch Salzsäure den Eisenbelag löst. Wo beide in grösserer Menge
vorkommen, kann man mit Sicherheit auf einen grösseren Eisengehalt des
Wassers schliessen. Die Einzelzellen der Anthophysa können übrigens
auch unter Umständen einen amöbenartigen Zustand annehmen; sie
verlieren dann Geissel und Form, senden Pseudopodien aus und kriechen
ganz wie jene umher. Eine Fortpflanzung ist bisher nur durch Teilung
der Zellen beobachtet worden (Fig. 35, 2a–b).
Zu derselben Hauptgruppe gehört auch die interessante
Gattung Dinobryon, welche in zwei Arten in unseren
Süsswasseransammlungen vorkommt. Die Dinobryen besitzen ebenfalls
eine Haupt- und eine Nebengeissel, einen Augenfleck, 1–2 kontraktile
Vakuolen, einen Zellkern und zwei schwach bläulichgrün gefärbte
plattenförmige Chromatophoren. Ihre Gestalt ist eiförmig, bald mehr
länglich, bald mehr rundlich, was sie aber noch ganz besonders
auszeichnet, ist[S. 178] das eigentümliche Gehäuse, welches jedes Einzeltier
umgiebt. Dasselbe besteht aus einer dünnen, farblosen Haut von
becherförmiger Gestalt, welches sich am Hinterende zuspitzt, am
Vorderende aber abgestutzt und geöffnet ist. Am Grunde dieser Gehäuse
sitzen die Zellen und strecken ihre Geisseln aus denselben hervor. Bei
der Fortpflanzung teilt sich die Zelle in zwei Tochterindividuen, von
denen das eine am Grunde des Gehäuses zurückbleibt, das andere sich an
dessen Mündung ansiedelt und ein neues Gehäuse entwickelt. So entstehen
allmählich frei im Wasser umherschwimmende, verzweigte Kolonien, welche
aussehen, als ob eine Anzahl Becher in einander geschoben wäre. Die
Organismen können auch in einen Ruhezustand eintreten; sie verlassen
dann ihr Gehäuse, verlieren die Geisseln, runden sich ab und umgeben
sich mit einer dicken Membran. Sehr ähnlich dieser Gattung ist das
ebenfalls in unseren süssen Wässern und oft in Menge vorkommende
Epipyxis utriculus, dessen Individuen sich jedoch nach ihrer
Teilung von einander trennen und keine Kolonien mehr bilden (Fig. 35, 3
a–b).
Ein Übergang der Flagellaten zu den Rhizopoden wird durch eine
Anzahl Formen vermittelt, welche man unter dem gemeinsamen Namen der
Rhizomastiginen zusammenfasst. Unter ihnen ist namentlich die mit
einigen Arten in unseren Gewässern vertretene Gattung Mastigamoeba von
Interesse. Sie kommt in zwei wesentlich von einander verschiedenen
Zuständen vor, als Flagellate und als Amöbe, welche in einander
übergehen können. Gewöhnlich tritt sie in der letztern Form auf, bildet
dabei mehrere bis zahlreiche (auch verzweigte) Pseudopodien, welche sie
bald einzieht, bald an anderen Stellen des eiförmigen Körpers wieder
hervorstreckt. In diesem Zustande kriecht sie an festen Gegenständen
umher und schwimmt nicht, obgleich sie auch als Amöbe eine Geissel
besitzt. Zuweilen ziehen jedoch die Individuen ihre amöbenartigen
Fortsätze ein, so dass nur die eiförmige Zelle selbst mit der Geissel
erscheint, mit vereinzelten Höckern oder Auswüchsen, welche die
Stelle der eingezogenen Pseudopodien andeuten. Jetzt kriechen sie
auch nicht mehr, sondern bewegen sich schwimmend wie Flagellaten
durch das Wasser. In der Nahrungsaufnahme liegt ebenfalls eine
Annäherung[S. 179] an die Rhizopoden: die Pseudopodien umfliessen nämlich,
ganz wie bei diesen, die verschiedenen als Nahrung dienenden Körper
und diese werden entweder mit den Pseudopodien eingezogen, oder der
Körper der Mastigamoeba fliesst selbst nach dem Gegenstande hin. Die
anderen hierher gehörigen Gattungen sind sehr ähnlich organisiert und
unterscheiden sich durch den Besitz von zwei Geisseln oder auch durch
das Fehlen derselben im Amöbenzustande (Fig. 35, 4 a–b).
Eine Gruppe der Flagellaten trägt den Namen der Heteromastigoda, weil
sie sich durch zwei verschieden gestaltete Geisseln auszeichnen. Beide
stehen am Vorderende des Körpers, aber nur eine und zwar die kleinere
ist auch nach vorn gerichtet, während die andere zurückgebogen ist
und bei der Bewegung nachgezogen wird. Es sind kleine farblose Wesen,
welche zuweilen amöbenartige Fortsätze entwickeln, aber ihre Nahrung
nicht durch Umfliessen der Gegenstände, sondern durch Aufnahme an
einer bestimmten Stelle, der Mundstelle des Körpers, die zuweilen zu
einem deutlichen Munde ausgebildet ist, zu sich nehmen. Hierher gehört
die Gattung Bodo, welche ovale oder längliche Formen enthält, deren
längere nach hinten gerichtete Geissel sehr häufig zur Anheftung an ein
Substrat dient. Bei dieser Gattung ist auch eine Vereinigung zweier
Individuen beobachtet, welche zur Bildung eines Ruhezustandes führt.
Aus dieser ruhenden Zelle gehen dann eine Anzahl neuer Individuen
hervor. Einige Arten dieser Gattung kommen im süssen Wasser vor,
eine (vielleicht aber auch mehrere) schmarotzen in anderen Tieren,
beispielsweise im Darm von Eidechsen. Die frei im Wasser lebenden Arten
ernähren sich meist auch parasitisch, indem sie andere Flagellaten oder
Algen anbohren, mit ihrem spitzen Vorderende eindringen und aussaugen.
So sind besonders Chlamydomonadinen, Volvocineen, Protococcaceen und
Palmellaceen den Angriffen des Bodo caudatus ausgesetzt, und
wo diese Art in Kulturen einmal sich eingestellt hat, sind letztere
auch in der Regel verloren. Denn ihre natürlichen Feinde, grössere
Infusorien, ferner Cypris- und Cyclopsarten, welche in unserem
Süsswasser der Überhandnahme dieser parasitischen Flagellaten Einhalt
thun, verzehren auch die[S. 180] Algen, und da sie leichter zu entfernen sind
als die mikroskopischen Organismen und ebenfalls die Algen vernichten
würden, sucht man sie, wenn irgend möglich, sorgsam wegzufangen, und so
bleibt dann den kleinen Räubern freier Spielraum (Fig. 35, 5).
Noch mag darauf hingewiesen werden, dass es eine Anzahl sehr einfacher
Flagellatenformen giebt, welche eine entschiedene Verwandtschaft zu
den Bakterien zeigen. Es ist dies beispielsweise mit der Gattung Monas
der Fall, welche gewissen geisseltragenden Bakterien sehr ähnlich ist.
Ebenso kann die Gattung Oikomonas mit ihnen verglichen werden. Die
Flagellaten enthalten aber einen Zellkern und kontraktile Vakuolen,
welche den Bakterien entschieden abgehen. Manche grosse Formen, deren
Zugehörigkeit zu den Bakterien jedoch sehr fraglich ist, besitzen nach
Bütschlis neuen Untersuchungen allerdings kleine Körperchen
im Zellinhalt, welche Zellkernreaktionen zeigen. So sind denn die
Flagellaten einerseits mit den Tieren, anderseits mit den Pflanzen nahe
verwandt und durch deutliche Übergänge verbunden; viele Formen zeigen
durchaus tierische Eigenschaften, viele mehr pflanzliche, und man
würde sie dem Pflanzenreich unbedingt einordnen müssen, wenn sie nicht
durch vollständige Formenreihen den echt tierischen Flagellaten weit
näher ständen, als den nächsten pflanzlichen Organismen. Noch heute
vertritt der eine Forscher die durchaus pflanzliche Natur gewisser
Organismen, während ein anderer sie unbedingt zu den Tieren stellt und
ein dritter beiden Wissenschaften, der Zoologie und der Botanik, die
Berechtigung zuerkennt, sie als Übergangsformen und Endglieder gewisser
Entwickelungsreihen in ihre Systeme aufzunehmen.
Mit dieser Schilderung verlassen wir das Gebiet der eigentlichen
Flagellaten und wollen uns noch Formen zuwenden, welche zu einer mit
diesen nahe verwandten Gruppe, den Dinoflagellaten, gehören. Auch sie
besitzen zwei Geisseln, welche wie bei den Heteromastigoden verschieden
ausgebildet sind, sie zeigen meist nur einen Zellkern im Innern und
ebenso wie gewisse Flagellaten Chromatophoren, aber, soweit bekannt,
keine kontraktilen Vakuolen. Von besonderer Wichtigkeit ist die
harte Schale, welche die Zelle[S. 181] umgiebt und, abgesehen von einigen
Übergangsformen, welche hier nicht berücksichtigt werden können, der
ganzen Gruppe ein einheitliches Gepräge verleiht. Diese Schalenhülle
zeigt ausser verschiedenartigen Grübchen oder Erhöhungen eine quer
verlaufende Furche, in welcher die eine der beiden Geisseln liegt und
durch sehr kurze wellige Bewegungen den Anschein erweckt, als ob in der
Furche eine Anzahl Wimpern in Thätigkeit wären, was man auch bis vor
kurzer Zeit thatsächlich geglaubt hat.
Wir wollen eine in unseren Seen nicht seltene Gattung Ceratium
mit vielen oft sehr schwer unterscheidbaren Formen auswählen, um an
ihr die Eigenschaften der Gruppe näher kennen zu lernen. Ceratium
Hirundinella (Fig. 35, 6) zeigt einen mit verschiedenen, in Gestalt
und Ausbildung recht variierenden Fortsätzen versehenen Körper, welcher
eine Quer- und eine Längsfurche trägt. Die letztere ist bei der Gattung
überhaupt sehr entwickelt und fast ebenso breit wie lang, so dass
sie kaum noch den Namen einer Furche verdient. Im Gegensatz dazu ist
die Querfurche sehr schmal und läuft um den ganzen Körper herum. Auf
jeder Seite der Querfurche setzen eine Anzahl Täfelchen die harte
Zellhülle zusammen, von denen am Hinterende drei zu den erwähnten
Fortsätzen oder Hörnern auswachsen. Während aber am Hinterende jede
der drei Täfelchen ein eigenes Horn entwickeln, bilden drei Täfelchen
des Vorderendes gemeinsam ein meist sehr langes Horn, welches am obern
Ende eine Öffnung, die Apikalöffnung, trägt. Die Körperhülle, welche,
wie erwähnt, aus einer Anzahl kleiner Täfelchen zusammengesetzt ist,
zeigt ein poröses Aussehen und ist ausserdem mit kleinen Stacheln
besetzt. Im Innern der Zelle finden sich zahlreiche gelbbraune oder
bräunlichgrüne Chromatophoren, welche die Färbung des Organismus
bedingen. Ältere Schalen nehmen allerdings ebenfalls eine gelbe oder
bräunliche Färbung an. Die Fortpflanzung der Ceratien erfolgt auf
verschiedene Weise. Einmal ist es sicher, dass sich die Individuen auch
im beweglichen Zustande durch Zweiteilung vermehren, indem die feste
Hülle gesprengt wird und an dieser Stelle eine Einschnürung des Körpers
und schliesslich eine Loslösung der beiden[S. 182] Teile von einander erfolgt.
Wahrscheinlich entwickeln sich dann die fehlenden Zellhälften aus
den beiden Teilen in normaler Weise mit ihren Umhüllungen. Aber auch
in einem Ruhezustande, in welchen die Ceratien zeitweilig eintreten,
erfolgt eine Teilung, jedoch hier in mehrere Individuen. Ob eine
geschlechtliche Fortpflanzung vorkommt, lässt sich bis jetzt nicht mit
Sicherheit angeben, sie ist aber für einige Formen der Dinoflagellaten
wahrscheinlich. Die Bewegung der hierhergehörigen Formen ist derjenigen
der eigentlichen Flagellaten fast gleich. Die Ceratien nehmen keine
feste Nahrung auf und vermögen wohl mittels ihrer Chromatophoren
auch überhaupt aus anorganischen Verbindungen wenigstens teilweise
diejenigen Stoffe zu bereiten, welche sie zu dem Aufbau ihres Körpers
brauchen. Bei anderen Dinoflagellaten scheint aber die Aufnahme fester
Nahrung sicher vorzukommen. Da manche der Ceratien eine für Flagellaten
ziemlich ansehnliche Körpergrösse besitzen (bis ½ mm), so ist
es wohl möglich, sie unter Umständen mit blossem Auge zu erkennen, wenn
man das Ceratien haltende Wasser etwa in einer flachen Schicht auf
einem Porzellanteller vor sich hat.
Schliesslich mag noch darauf hingewiesen werden, dass eine Anzahl
mariner Dinoflagellaten zum Leuchten des Meeres beitragen, während
allerdings noch einige andere Organismen die Hauptrolle dabei spielen.
Dies ist der Fall bei der ebenfalls zu den Flagellaten im weitesten
Sinne gehörenden Gattung Noctiluca, welche mit noch einer andern
marinen Gattung zusammen die Abteilung der Cystoflagellaten ausmacht.
Auch einige Bakterienarten nehmen unter den mikroskopischen Organismen
teil an der Erzeugung des Meerleuchtens.
[S. 183]
Litteratur.
Die drei wichtigsten Werke über Flagellaten sind:
1. Ehrenberg, Die Infusionsthiere als vollkommene Organismen.
1838.
2. Stein, Der Organismus der Infusionsthiere. 1878.
3. Bütschli, Protozoen. 1880.
Weniger ausführlich, aber mit recht guten Abbildungen versehen und
für die Bestimmung unserer Süsswasserformen in den meisten Fällen
ausreichend ist Kirchner u. Blochmann, Die mikroskopische
Pflanzen- und Tierwelt des Süsswassers. 1885 und 1886. II. Teil.
Auch Eiferth, Die einfachsten Lebensformen des Tier- und
Pflanzenreiches, ist zu empfehlen. — Diese letzteren Werke geben zwar
nicht die Fülle des Materials und die künstlerischen Abbildungen der
obigen, aber sie sind wesentlich billiger und nicht jeder ist in der
Lage, sich Steins prachtvolles Infusorienwerk anzuschaffen.
Die spezielle Litteratur ist bei Bütschli zusammengestellt; Angaben
über die mikroskopische Technik finden sich bei Blochmann und Eiferth.
[S. 185]
Die Süsswasserschwämme.
Von Dr. W. Weltner in Berlin.
[S. 187]
1. Erkennen und Auffinden.
Fährt man an einem windstillen Tage auf einem unserer Seen oder Teiche
langsam am Rande des Schilfrohres hin, welches die Ufer oft auf lange
Strecken bis zu einer Wassertiefe von einem Meter und mehr einsäumt,
so gewahrt man an den Rohrstengeln nicht selten lebhaft grün gefärbte,
graue oder graubraune Massen. Sie sitzen fast immer nur an den
abgestorbenen Schilfstengeln, welche nach dem Rohrschnitt im Herbste
im Wasser stehen geblieben sind und sich noch Jahre lang an der Wurzel
aufrecht erhalten. Jene Massen am Rohr bieten ein verschiedenes Äussere
dar. Sie erscheinen entweder als dünne Überzüge oder als dickere
Krusten, oft sogar als dicke Klumpen, oder aber sie sind baumförmig
verzweigt und können in dieser Gestalt eine Länge von mehreren Metern
erreichen. Auch in unseren Flüssen, Kanälen und Flusshäfen sehen
wir sie, hier am Ufer an den Steinen, an lebenden Baumwurzeln, an
untergesunkenen Holzstücken, oder an den Wänden der Schleusen und
Mühlenzuflüsse, an Brückenpfählen oder an Mauern (selbst innerhalb der
Städte) wachsend. Aber wir brauchen sie nicht einmal in ihrem Elemente
mit den Augen zu suchen, wir finden sie auch, freilich meist zerrissen
und zerbröckelt, wenn wir an trüben, windigen Tagen im unklaren Wasser
mit einem Ketscher an Pflanzen, Steinen oder an ins Wasser geratenen
Gegenständen hinfahren. Selten findet man sie an Körpern angewachsen,
welche im Wasser flottieren. Ja, man hat sie sogar ohne irgend welches
sie tragende Substrat an der Wasseroberfläche[S. 188] schwimmend angetroffen;
in diesem Falle haben wir es aber sicher mit Stücken zu thun, die von
ihrer Unterlage losgerissen wurden.
Nehmen wir die auf die eine oder andere Weise erhaltene Masse in die
Hand und betrachten wir sie genauer. Es bietet sich uns eine mehr oder
weniger schleimige Substanz dar, die einen ganz eigentümlichen Geruch
besitzt. An der Oberfläche bemerken wir viele feine Spitzen, sehr
viele kleine Poren und eine geringe Anzahl grössere Löcher. Schon der
Unkundige erkennt an dieser Oberflächenbeschaffenheit eine Ähnlichkeit
mit dem Badeschwamm. Bemühen wir uns, ein unverletztes Stück
aufzufischen und möglichst schnell in ein Glas mit Wasser zu setzen!
Bald sehen wir, wie einige in das Wasser gestreute Karminkörnchen,
wenn sie in die Nähe der grösseren Löcher geraten, hier plötzlich weit
fortgetrieben werden.
Diese Massen, welche auf den ersten Blick so wenig Einladendes bieten
und die wohl viele von uns, ohne sie näher zu betrachten, schon gesehen
haben, sind Schwämme, und zwar im süssen Wasser die einzigen
Vertreter dieses in allen Meeren sehr verbreiteten Tierkreises. Um
so mehr haben diese Süsswasserschwämme oder Spongilliden schon seit
langer Zeit das Interesse der Naturforscher erweckt, und sie haben
infolgedessen ihre eigene Geschichte in der Wissenschaft. Man kann von
ihnen nicht sagen, wer sie entdeckt hat, da sie bei ihrem ungemein
verbreiteten Vorkommen und ihrer Häufigkeit wohl von jeher gekannt
waren. Es hat sehr lange gedauert — eine Spanne von 140 Jahren —,
ehe man ihre wahre Natur erkannte, und diese konnte überhaupt erst
aufgedeckt werden durch das Studium des innern Baues, ein Verdienst,
welches sich Robert Edmund Grant erwarb, als er im Jahre
1826 seine Untersuchungen über die Organisation von „Spongilla
friabilis“ bekannt gab (Edinburgh Phil. Journal, Vol. XIV,
p. 270–284. 1826).
[S. 189]
2. Geschichtliches.
Der erste, welcher über unsere Schwämme überhaupt geschrieben hat, war
Joh. Ray (Rajus), der 1686 in seiner Historia plantarum
einen Süsswasserschwamm beschreibt. Die erste Abbildung lieferte
Leonard Plukenet 1691. Seit ihrer ersten Erwähnung bis zu
Pallas (1766) für Pflanzen gehalten, werden sie von den späteren
Autoren bis zu Grant hin für Pflanzen, für Tierpflanzen,
Pflanzentiere und endlich für Tiere angesehen. Die einen trennen sie
von den Meeresschwämmen, die Mehrzahl vereinigt sie jedoch mit ihnen.
Selbst noch nach den gründlichen Untersuchungen Grants wurde
ihnen eine pflanzliche Natur zugeschrieben, wozu die grüne Farbe und
die „Samenkörner“ (gemmulae) allerdings verlocken mussten.
Trotzdem dass nach Grants Arbeit einige wichtige Entdeckungen
aus der Lebensgeschichte unserer Schwämme in den folgenden zehn
Jahren gemacht wurden, gebührt doch erst Dujardin (1838) die
Ehre, einen bedeutenden Schritt in der Erkenntnis der Organisation
der Spongilliden gethan zu haben, indem er die Zusammensetzung aus
amöboiden Zellen und aus Wimperzellen lehrte. Die grössten Verdienste
um die Kenntnis der Süsswasserspongien erwarben sich dann Carter
(1847 bis jetzt) und Lieberkühn (1856–70), der eine ein Schüler
Grants, der andere der Assistent Joh. Müllers. Ihren
Arbeiten sind zahlreiche andere gefolgt, die wir den nachfolgenden
Zeilen über die Naturgeschichte der Spongilliden zu Grunde legen. Dabei
mögen eine Anzahl eigener Beobachtungen mit einfliessen.
3. Äussere Beschaffenheit.
Nachdem wir oben schon die Merkmale besprochen haben, an denen man
einen Süsswasserschwamm erkennt, müssen wir noch einiges über den
Habitus dieser Organismen hinzufügen.
a) Form und Oberflächenbeschaffenheit.
Die Gestalt jeder jungen Spongillide — mag sie sich aus einer Larve
oder einer Gemmula entwickelt haben — ist gewöhnlich ein flacher,
seltener ein hoher Kegel. Bei dem weiteren Wachstum[S. 190] nimmt der Schwamm
zunächst immer die Gestalt einer flachen Scheibe an und wächst erst
danach in die Dicke. War das den jungen Schwamm tragende Substrat
keine gerade Ebene, sondern ein dünner cylinderförmiger Gegenstand,
ein Pflanzenstengel, ein Bindfaden, ein Eisendraht etc., so umwächst
der Schwamm seine Unterlage und nimmt erst dann an Dicke zu. Mit
einem Wort, er passt sich zunächst an seine Unterlage an. Erst
wenn eine gewisse Grösse erreicht ist, kommen die beiden für die
Süsswasserschwämme eigentümlichen Gestalten zum Vorschein. Die einen
beginnen fingerförmige Fortsätze zu treiben, die sich bei weiterem
Wachstum verzweigen, so dass endlich baum- oder strauchförmige Massen
entstehen (Euspongilla lacustris), die anderen bleiben in
der Regel zeitlebens krustenförmig, ihre Oberfläche ist mehr oder
weniger uneben oder mit spitzigen Zapfen oder gerundeten Wülsten oder
blattförmigen Erhebungen versehen, und wenn längere Fortsätze an ihnen
sichtbar sind, so rührt diese scheinbar selbständige Verzweigung
von der Unterlage her. In Fig. 44 (S. 219) haben wir einen solchen
Schwamm abgebildet, der einen verästelten Baumzweig überzogen hat;
einen gleichen Fall hat auch Retzer[72] wiedergegeben. Alle
diese krustenförmigen Spongilliden gehören den Gattungen Spongilla,
Trochospongilla und Ephydatia an. Von diesen im allgemeinen gültigen
Regeln haben wir indessen zwei Ausnahmen zu verzeichnen. Es kommen
nämlich auch Exemplare von Ephydatia Mülleri vor, die verzweigte
Massen bilden, doch sind das Seltenheiten. Ferner tritt die sonst
stets verzweigte Euspongilla lacustris unter gewissen Umständen
unverästelt auf, wenn nämlich diese Art an stärker fliessenden Stellen
sich angesiedelt hat; hier unterbleibt dann die Bildung der Fortsätze.
Ein solches Exemplar sehen wir in Fig. 38 (S. 212). Es wurde, einen
Wollenfaden umwachsend, an der Oberfläche der Spree in Berlin unter
einer Brücke gefunden und zwar an einer Stelle, an welcher starke
Strömung herrschte.
Linné war der Meinung, die verzweigte Form fände sich mehr in
Seen, die krustenförmige mehr in Flüssen. Pallas glaubte das
Gegenteil annehmen zu müssen, machte aber die richtige[S. 191] Beobachtung,
dass die verzweigte Form gewöhnlich nach oben wächst und bei starker
Wasserströmung ihre Äste in horizontaler Richtung entsendet. Indessen
finden wir sowohl die verzweigten als die unverzweigten Formen in
Seen und Flüssen; während aber eine verzweigte Spongilla in ruhigen
Gewässern auch mehr in der Nähe der Oberfläche des Wassers wächst
und ihre Zweige eine bedeutende Länge erreichen und hier selbst bei
senkrecht gestellter Unterlage nach oben streben können, hält sich
dieselbe Art in stärker fliessendem Wasser in der Tiefe auf, in welcher
die Strömung weniger stark ist; wenn sie hier aber an der Oberfläche
wächst, so verlaufen ihre Äste in horizontaler Richtung. So wächst
in der Spree in Berlin, welche hier durch ihren enormen Reichtum an
Spongillenexemplaren ausgezeichnet ist, die verzweigte Euspongilla
lacustris mit Vorliebe am Grunde des Flusses, während sich die
unverzweigten massigen Formen mehr an der Oberfläche auf jedem festen
Gegenstand angesiedelt haben.
b) Die Grösse.
Die Süsswasserschwämme gehören unter den Spongien zu den grösseren
Formen. Die verzweigte Euspongilla lacustris geht sogar über
das gewöhnliche Mass hinaus, sie wird über einen Meter lang. Von den
unverzweigten Arten erreichen wohl Ephydatia fluviatilis und
Mülleri den grössten Umfang. Wir haben einmal ein Exemplar von
19 cm Länge und 8½ cm Dicke gefunden und man trifft
öfters rasenförmige Stöcke dieser Arten an, welche bei geringer Höhe
einen halben Meter breit werden. Siehe auch Potts[73].
c) Die Farbe.
Es wird gewöhnlich angenommen, dass die am Lichte wachsenden
Spongilliden grün seien, während andere, welche vom Lichte abgewandt
unter Brücken oder unter Steinen wachsen, farblos, d. h. weiss,
gelblich oder grau sind. Es ist das im allgemeinen richtig.
So findet man selbst Exemplare, die je nach der Beleuchtung auf
der einen Seite ein grünes, auf der andern Seite ein gelbliches[S. 192]
Kolorit zeigen. Hält man solche Schwämme in Aquarien, so ergrünt
auch die früher gelblich gefärbte Seite. Es fehlt aber nicht an
Beobachtungen, dass am Lichte wachsende Spongilliden auch farblos
sind. Carter fand solche in Bombay, Potts in Amerika
und Weber[74][75]
auf Sumatra. Die von Weber gefundenen
Ephydatia fluviatilis waren nur an gewissen Körperpartien grün,
aber diese grüne Färbung hat, wie wir später sehen werden, eine ganz
andere Ursache, als das grüne Kolorit unserer Spongilliden, welches von
mikroskopisch kleinen in den Zellen des Schwammkörpers liegenden grünen
Körperchen herrührt, denen man den Namen Zoochlorella parasitica
(Brandt) gegeben hat. Nun kann dieses grüne Pigment auch durch
ein anderes vertreten sein; Carter fand, dass die graue bis
lilagleiche Farbe der Spongilla cinerea von ebenso gefärbten
Körnchen in den Zellen herrühre und wir fügen dem ein weiteres Beispiel
hinzu. Bei Berlin lebt in einem See untermischt mit anderen grün
gefärbten Arten eine stets grau oder braun kolorierte Spongilla
fragilis. Diese Art lässt sich daher von allen dort lebenden
Spongilliden sofort an ihrer Farbe erkennen. Die braune Färbung jenes
Schwammes rührt von gelbbraun tingierten Körnchen her, die in denselben
Zellen liegen, welche bei den anderen Arten die grünen Zoochlorellen
beherbergen. Die Spongilla fragilis trägt aber nicht immer
eine braune Farbe, sie ist an anderen Orten weiss, gelblichweiss oder
grün (Potts). Von Ephydatia fluviatilis beschreibt
Lankester[76] blasslachsfarbene, hellgrüne und hellbraune
Exemplare. Aus alledem geht hervor, dass die Süsswasserschwämme
verschiedene Pigmente führen und dass selbst ein und dieselbe Art
verschieden koloriert auftritt.
d) Die Konsistenz.
Die Konsistenz unserer Spongilliden hängt, wie bei den meisten
Schwämmen, von der Struktur des Skelettes ab. Wir unterscheiden danach
zweierlei. Bei den einen ist die Masse der Kittsubstanz, welche die
einzelnen Nadeln zu einem Gerüste verbindet, mächtig entwickelt,
bei den anderen ist sie sehr gering, dafür ist aber das kieselige
Skelett mächtiger entfaltet. Diejenigen Süsswasserschwämme[S. 193] mit viel
Kittsubstanz setzen dem Zerreissen mehr Widerstand entgegen, als
solche mit gering entwickelter Kittmasse. Dagegen sind die Arten mit
einem Gerüste, welches wesentlich aus Kieselnadeln besteht und wenig
Spongiolinmasse führt, viel härter als die schwieriger zerreissbaren.
Es giebt daher im allgemeinen zwei Sorten unter unseren Schwämmen,
solche, welche weich anzufühlen und schwieriger zerreissbar sind
(Eusp. lacustris), und solche, die eine gewisse Härte zeigen
und sich leicht zerbrechen lassen (Ephyd. fluviatilis). Unter
den letzteren kommt, was den Grad der Härte anlangt, noch der Wuchs
in Betracht. Diejenigen, welche als minder dicke Krusten Baumzweige
überziehen (Fig. 44 S. 219), sind weniger hart als die, welche kompakte
Massen bilden (Fig. 42 S. 218). Es giebt unter den zuletzt genannten
klumpenförmige Stücke, welche in ihrer Härte einem Zimokka-Badeschwamm
nichts nachgeben. — Was den Weichteil der Süsswasserschwämme angeht,
so scheint derselbe bei allen krustenförmigen Arten gleich zähflüssig
zu sein, während er bei der verzweigten Eusp. lacustris
dünnflüssiger ist. Von einigen älteren Autoren wird angegeben, dass der
Schleim dieser Art nach dem Herausziehen des Schwammes aus dem Wasser
bald abtropfe, ein Verhalten, welches wir nie beobachten konnten. —
Im trockenen Zustande sind alle unsere Spongilliden sehr brüchig und
zerfallen vollständig, wenn man sie zwischen den Fingern reibt. Auf
diese Eigenschaft weisen die alten Namen fragilis und dergl.
mehr hin.
e) Der Geruch.
Man hat den Geruch eines frisch aus dem Wasser gezogenen Schwammes
fischig, schlammig, moderartig, auch jodähnlich genannt. Allein diese
Vergleiche passen nicht genau, der Geruch ist ein unserem Schwamme ganz
eigentümlicher.
4. Anatomie und Histiologie.
Als im Jahre 1875 F. E. Schulze die erste seiner zahlreichen
Untersuchungen über den Bau und die Entwickelung der Spongien
veröffentlichte, begann eine neue Periode in der Kenntnis der Schwämme.
Obwohl vor jenen Untersuchungen kein anderer[S. 194] Schwamm anatomisch
und histiologisch so genau zergliedert worden war wie gerade unser
Süsswasserschwamm (Lieberkühn, Carter), so gaben doch
erst Schulzes Arbeiten das richtige Verständnis für die
Organisation auch unseres Tieres, weshalb wir auch hier auf denselben
fussen, ohne in all den einzelnen Fällen immer wieder den Namen dieses
Forschers anzuführen.
Soweit bekannt, giebt es keine Süsswasserschwämme ohne ein aus
Kieselnadeln gebildetes Skelett. Wir werden also passend die beiden
einen Schwamm zusammensetzenden Teile, das Skelett und den Weichteil,
gesondert besprechen.
a) Das Skelett.
Dasselbe besteht aus dem festen Gerüste und den lose und ohne Ordnung
im Weichteil liegenden sogenannten Fleischnadeln. Das Gerüst wird aus
schlanken, fast immer leicht gekrümmten, glatten oder bedornten Nadeln,
welche an beiden Enden zugespitzt sind, aufgebaut. In ihrem Innern
zeigen sie einen an beiden Enden geschlossenen Kanal, den Zentralkanal,
in welchem man an geeigneten Präparaten einen feinkörnigen Zentralfaden
erkennt. Die diesen umhüllende Kieselsubstanz ist nicht homogen,
sondern lässt abwechselnd Lagen von Kieselsäure und Spongiolinsubstanz
erkennen. Diese Nadeln legen sich nun zu mehreren zu einem Bündel
zusammen, an welches sich ein zweites, dann ein drittes und so fort
anschliesst, welche alle durch eine Kittmasse (Spongiolinsubstanz)
mit einander verbunden sind. So kommen festere Stäbe zustande, die
an der Basis des Schwammes mächtiger sind als an der Oberfläche.
Wir nennen sie Hauptfasern. Sie allein würden dem überaus zarten
Weichteil noch keinen genügenden Halt geben; sie sind deshalb durch
Querbrücken, Verbindungsfasern, gestützt, welche nur aus wenigen, oft
nur aus einer einzigen Nadel bestehen. So kommt ein netzförmiges, auf
einer basalen Spongiolinplatte angekittetes Skelettwerk mit Maschen
von der verschiedensten Gestalt und Grösse zustande. Schon in dem
wenige Stunden alten Schwamm macht sich die Tendenz bemerkbar, die
jungen Nadeln, deren jede stets in einer Zelle gebildet wird, zu
einem Netzwerk zusammenzufügen.[S. 195] Der lockere Weichteil kann desselben
nicht entbehren. Auch an einem schon grossen Schwamm sehen wir immer
noch neue Nadeln entstehen, die durch eine oder mehrere kugelige
Verdickungen in dem mittleren Teile als solche meist kenntlich sind.
Noll hat die Bildung der Spikula verfolgt und glaubt, dass die
Zelle (Silikoblast), in welcher die Nadel entsteht, zu deren weiteren
Ausbildung auch genüge, obwohl dagegen spricht, dass die ausgewachsene
Nadel um sehr vieles grösser ist als die junge. Auch der Umstand, dass
man den Nadelsträngen seitlich anliegend viele Zellen sieht, welche
gar keine Nadel in sich bergen, weist darauf hin, dass die Spikulae
nachträglich durch besondere Zellen wachsen. Was die Kittsubstanz
anlangt, so verdankt sie ebenfalls besonderen Zellen (Spongoblasten)
ihre Entstehung (Noll). — Unter den Nadeln eines
Süsswasserschwammes kommen fast stets Abnormitäten mannigfaltigster
Art vor; es können zwei Nadeln unter verschiedenem Winkel vollständig
mit einander verwachsen, das eine Ende einer Nadel kann gegabelt oder
umgebrochen oder abgerundet sein; eine Nadel kann in der Mitte in
verschiedenen Winkeln geknickt werden, kurzum es sind dieser Gestalten
so viele, dass man einige Tafeln Abbildungen zusammenstellen könnte.
Man verwechsele aber hiermit nicht jene kleineren in der Mitte kugelig
angeschwollenen Nadeln, welche nur die Jugendzustände der Gerüstnadeln
sind und häufig als Fleischnadeln angesprochen wurden.
b) Der Weichkörper.
Das Fleisch oder Parenchym unseres Schwammes besteht aus einer inneren
Masse und einer äusseren Haut, welche dieselbe wie ein Sack einhüllt.
Diese Haut ruht auf den Enden der aus dem Innern kommenden Hauptfasern
des Skelettes und lässt unter sich einen grossen kontinuierlichen
Hohlraum, den Subdermalraum, frei, der nur durch die Skelettbalken und
gelegentlich durch nadelfreie Gewebszüge unterbrochen wird. Die Haut
enthält mikroskopisch kleine Löcher, die Einströmungsporen, deshalb
so genannt, weil durch sie das Wasser in den Schwamm einströmt. Am
Boden der grossen sackförmigen Höhle zwischen Haut und Innerm bemerkt[S. 196]
man mit dem blossen Auge grössere Löcher, es sind die Öffnungen von
Kanälen, die sich in den Schwamm hinein erstrecken, sich vielfach
verzweigen und seitlich angelagert kleine kugelige Gebilde tragen, die
man wegen ihrer Zusammensetzung aus wimpernden Zellen Wimper- oder
Geisselkammern genannt hat. Sie münden in die eben besprochenen Kanäle,
die wir Einströmungskanäle nennen, durch eine Anzahl sehr kleiner
Poren, Einströmungsporen der Kammern. Auf der abgewendeten Seite trägt
eine solche Geisselkammer ein grosses Loch, die Ausströmungspore, durch
welche sie sich in einen Kanal öffnet, der im allgemeinen weiter ist
als die Verzweigungen der Einströmungskanäle. Jene weiteren Kanäle
sammeln sich zu noch grösseren und münden endlich in ein einziges
grosses Rohr, das Kloakenrohr. Die zwischen den Geisselkammern
und diesem Rohr gelegenen Kanäle sind die Ausströmungskanäle.
Das Kloakenrohr durchsetzt den unter der äusseren Haut liegenden
Hohlraum mit einer eigenen geschlossenen Wand und öffnet sich an
der Oberfläche des Schwammes mit einem runden, von einem Hautsaum
umgebenen Loch, dem Oskulum, oder zeigt ein über der Oberfläche des
Schwammes hervorstehendes weissliches Röhrchen, das Oskular- oder
Auswurfsrohr, welches nichts anderes als eine Fortsetzung der äusseren
Haut ist. An Stelle des Kloakenrohres sehen wir an dünneren Stellen
krustenförmiger Schwämme und auf den Zweigen der verästelten Form
besondere Rinnen von oft sternförmiger Gestalt eingegraben; an dem
Boden dieser Furchen fallen grössere Löcher auf, welche die Enden der
grossen Ausströmungskanäle darstellen. Über den Furchen zieht sich
die äussere Haut hinweg, die an dieser Stelle einzelne grosse Poren
oder Röhren trägt und in der Mitte meist ein grösseres Auswurfsrohr
zeigt. Diese Rinnen oder das sternförmige Rinnensystem sind nur
eine besondere Form des Endabschnittes der Kloakenhöhle. An grossen
Schwammexemplaren tritt noch eine besondere Modifikation solcher
Kloakenhöhlen auf. Wir sehen nämlich hier an der Schwammoberfläche
grosse, öfters bis 1 cm breite und tiefe Löcher, an deren
Boden man in die Öffnungen der ausströmenden Kanäle gelangt. Die so
verschieden gestalteten Kloakenhöhlen sind[S. 197] stets an der Stelle,
an welcher sie den grossen unter der Haut hinziehenden Hohlraum
durchsetzen, von einer eigenen, soliden Wand begrenzt. Dadurch entsteht
ein für sich geschlossenes einführendes Kanalsystem, welches von den
Einströmungsporen der Haut bis zu den Geisselkammern reicht, und ein
für sich abgegrenztes ausführendes System, von den Kammern beginnend
und mit den Auswurfsöffnungen endend. Verfolgen wir jetzt den Lauf des
Wasserstromes weiter. Durch die Poren in den einführenden Subdermalraum
gelangt, strömt das Wasser durch die an dessen Boden liegenden Löcher
in die einführenden Kanäle und gelangt durch deren Verzweigungen in
die Geisselkammern. Diese sind es, in welchen wir die treibende
Kraft für den beständigen Wasserstrom im Schwamme zu erblicken haben.
Von ihnen gelangt das Wasser durch ihre grossen Ausströmungsporen in
die ableitenden grösseren Kanäle, und durch diese in das Kloakenrohr
oder in die verschieden gestalteten Kloakenhöhlen und fliesst
schliesslich durch das Oskulum oder dessen Röhren nach aussen ab. Wir
sehen, dass die Bezeichnung Oskulum garnicht für jene Löcher passt,
welche vielmehr die Enden eines Kloakenrohres sind und gerade das
entgegengesetzte von dem bedeuten, was ihr Name anzeigt. Deshalb hat
auch schon Grant statt Oskulum die Bezeichnung Kloakenmündung
vorgeschlagen, allein der erstere Name ist nun einmal gang und gäbe
geworden und lässt sich auch gebrauchen, wenn man mit Vosmaer
das Wort in Verbindung mit cloacae bringt. Die Kloakenhöhlen und
die sternförmigen Kloakenbezirke zeigen am Schwamme nur sehr selten
eine regelmässige Anordnung. So fanden wir einmal eine Spongilla
fragilis, an welcher die Kloakenhöhlen sämtlich in einer Reihe
übereinander lagen, während die ganze übrige Oberfläche frei davon war.
Wir wollen hier auch noch bemerken, dass bei den verzweigten Formen
die Oskula nie an den Spitzen der Zweige oder der Zapfen liegen, wie
es bei vielen marinen Schwämmen der Fall ist. Bei diesen führt dann
das auf dem Gipfel gelegene Ausströmungsloch in eine grosse zentrale
Kloakenhöhle, die sich in den Zweigen von Eusp. lacustris nie
findet.
[S. 198]
Wir gehen jetzt zur Betrachtung der Gewebeschichten unseres Tieres
über. Die Aussen- und Innenseite der Haut, der Ausströmungsröhren,
ferner des Septums, welches den grossen Subdermalraum von dem
Endabschnitt der Kloakenhöhle trennt, diese selbst, der Boden des
Subdermalraumes und endlich alle Kanäle sind von einem dünnen
einschichtigen Lager platter, polygonaler Zellen ausgekleidet. Eine
besondere Gestalt gewinnen diese Zellelemente in der Umgebung der
Hautporen und der Ausströmungsöffnung der Geisselkammern. Sie haben
hier die Form einer Mondsichel von geringerer oder grösserer Breite.
Besonders die Poren, dann aber auch die Ein- und Ausströmungsöffnungen
der Kammern sind veränderliche Gebilde, die entstehen und vergehen
können. Besonders an den Hautporen lässt sich verfolgen, wie sie in
einer Zelle als Loch entstehen, ein solches Loch vergrössert sich
bald, bis von der Zelle nur noch ein schmaler Ring übrig bleibt. An
anderen Stellen sieht man, wie sich an der weit geöffneten Pore die sie
umgrenzende sichelförmige Zelle verbreitert, bis endlich von der Pore
nur noch ein kleines Loch übrig bleibt; auch dieses kann geschlossen
werden und die Pore ist verschwunden. — Die zwischen dem ein- und
ausführenden Kanalsystem eingeschobenen Geisselkammern bestehen
aus langgestreckten, radiär um die Höhle der Kammer angeordneten
Zellen. Sie sind durch eine zwischen den Zellen liegende Substanz von
einander getrennt, oder sie stossen eng zusammen und platten sich auch
gegenseitig ab. Jede Zelle trägt einen langen hyalinen Kragen und eine
lange beständig schlagende Geissel; durch die Gesamtheit der Bewegungen
aller Geisseln wird der Wasserstrom erzeugt. Es ist interessant, dass
manchen im Winter gesammelten Schwämmen die Geisselkammern fehlen
können. — Zwischen den Platten- und Geisselzellen bleibt nun eine
Schicht, welche, wie bei allen Schwämmen, auch hier an Mächtigkeit
die eben genannten Zelllager bei weitem übertrifft. Es ist die
Bindesubstanzschicht, welche aus einer hyalinen Substanz mit amöboiden
Zellen besteht. Unter den Zellen sieht man sofort zwei Sorten, die
einen haben einen Inhalt von eng aneinander liegenden, annähernd gleich
grossen Körnchen, die[S. 199] anderen führen ungleich grosse, weniger dicht
liegende Körner. Bei den grün gefärbten oder braunen Schwämmen enthält
stets nur die zuletzt genannte Sorte das grüne oder braune Pigment.
Das grüne Kolorit der Schwämme wird durch kleine rundliche Körperchen
in diesen Zellen hervorgebracht, welche Chlorophyll enthalten. Während
aber die einen dasselbe für tierischen Ursprungs halten, betrachten
die anderen jene Körper als einzellige Algen (Zoochlorella
parasitica Brandt). Ausser den genannten Zellen finden sich noch
sehr langgestreckte Formen, die den kontraktilen Faserzellen anderer
Spongien entsprechen. Der Siliko- und Spongoblasten haben wir schon
oben gedacht und einige andere Zellformen, die sich nur zu gewissen
Zeiten finden, werden wir sogleich kennen lernen.
5. Physiologie.
Die Süsswasserschwämme pflanzen sich auf geschlechtlichem und
ungeschlechtlichem Wege fort.
a) Die geschlechtliche
Fortpflanzung.
Sie findet bei uns in den Monaten Mai bis in den September statt.
Während man die männlichen Keimstoffe, die Spermatozoen, nur vom Mai
bis in den August findet, trifft man die Eier zu allen Jahreszeiten
an, sie kommen aber im Winter nur vereinzelt vor und werden dann nicht
entwicklungsfähig. Die Spongilliden sind getrennten Geschlechts und
zwar tritt bei ihnen zuerst die Entwickelung der Spermatozoen und
später die Reifung der Eier auf. Ein äusserer Unterschied in der Grösse
oder der Gestalt der männlichen und weiblichen Exemplare existiert
nicht, wenn es auch vorkommen mag, dass kleinere auf beweglicher
Unterlage angesiedelte Schwämme [Keller[77]] männlich sind.
Die Spermatozoen entstehen durch fortwährende Teilung einer Zelle
der Bindesubstanzschicht, deren Kern die Köpfe und deren Plasma die
Schwänze der Samenfäden liefert. Auch die Eier sind Abkömmlinge solcher
Zellen. Fiedler hat die Sperma- und Eientwickelung genau
verfolgt und Maas hat die Bildung des jungen Schwammes aus der[S. 200]
Flimmerlarve, welche aus dem sich furchenden Eie entsteht, in klarer
Weise auseinandergesetzt.
b) Die ungeschlechtliche
Fortpflanzung.
Der aus einer Larve oder einer Gemmula entstandene junge Schwamm
zeigt ein einziges zusammenhängendes Kanalsystem, welches mit einem
einzigen Oskulum oder mit einem sternförmigen Ausströmungsbezirk nach
aussen mündet. Wir nennen einen solchen Schwamm ein Individuum. Durch
weiteres Wachstum sehen wir ein zweites Oskulum an einer anderen Stelle
auftreten, bald bilden sich neue und wir haben nun eine durch Knospung
entstandene Kolonie vor uns. — Ob eine andere Fortpflanzung, nämlich
durch Teilung, wie es Laurent behauptet, wirklich vorkommt,
bleibt zweifelhaft; sicher aber ist, dass gewaltsam losgetrennte,
grössere Stücke eines Süsswasserschwammes wie bei anderen Spongien
fortwachsen, wenn sie sich auf einer geeigneten Unterlage festsetzen
können.
Zur Herbstzeit besonders findet man in den Spongilliden kleine,
gelbliche oder gelbbraune, annähernd kugelige Gebilde von der Grösse
eines Senfkornes: die Gemmulae. Während man weder über die erste
Entstehung noch über die Herkunft einzelner Teile dieser Gebilde
vollkommene Klarheit hat, sind wir über den Bau der ausgebildeten
Gemmula ziemlich gut orientiert. Sie besteht aus einer Hülle mit einer,
seltener mit mehreren Öffnungen und einem aus Zellen zusammengesetzten
Keim oder Kern. Es ist behauptet worden, dass in dem Keime Stärke
enthalten sei. Allein schon Lieberkühn hat das bestritten
und sicher sind jene stark lichtbrechenden Körner der Keimzellen
nicht Amylum. Dagegen wissen wir durch die Untersuchungen von
Carter, Keller[78], Ray Lankester,
Brandt
und Wierzejski[79], dass in den Süsswasserschwämmen Amylum und
amyloide Substanz vorkommt.
Die Hülle der Gemmula zeigt eine innerste dicke Membran (innere
Chitinmembran, innere Kutikula), welche die Höhle der Gemmula
umschliesst, auf diese Membran folgt eine Kruste (Belegmembran,
Luftkammerschicht), die entweder fein blasig aussieht[S. 201] oder sehr
deutlich zellig erscheint und Luft enthalten soll. In dieser zweiten
Schicht stecken die für die einzelnen Arten der Spongilliden
charakteristischen Nadeln (Belagsnadeln), deren Formen wir später
kennen lernen werden. Bei einigen Arten ist die jene Nadeln
beherbergende Schicht noch durch eine dritte Schicht (äussere Kutikula,
äussere Chitinmembran) abgeschlossen; solche Gemmulae sind dann glatt,
während andere, die der Membran entbehren, rauh erscheinen. Unter
den Belagsnadeln kommen oft abnorm gebildete Formen vor, besonders
bei Schwämmen, welche zu einer raschen, unzeitigen Gemmulabildung
veranlasst wurden [Wierzejski[80]].
Fragen wir zunächst: welchen Zweck haben die Gemmulae? Es wird
gewöhnlich angegeben, dass unsere Süsswasserschwämme gegen den Herbst
hin unter Bildung von Gemmulae absterben. Es ist das im allgemeinen
richtig. Man findet also von den meisten unserer Arten nur im Winter
die Gemmulae, ihr Weichteil ist zerfallen. Jene überwintern und im
Frühlinge kriecht aus ihnen der Inhalt aus und entwickelt sich zu einem
neuen Schwamm. Es ist also die Aufgabe der Gemmulae, den Schwamm, der
als solcher den Winter nicht überstehen würde, über diese Jahreszeit
hinwegzubringen. Ganz ähnlich ist es in den Tropen. Während der
Regenzeit sind Lachen, Bäche und Flüsse mit ausgebildeten Spongilliden
erfüllt, tritt dann die Trockenperiode ein, so entwickelt der Schwamm
Gemmulae, welche Monate und Jahre lang [Carter, Potts,
Lendenfeld[81]] der sengenden Hitze ausgesetzt bleiben können,
um später, wenn sie wieder vom Wasser bedeckt sind, zu neuen Schwämmen
zu erstehen. In den Gemmulae sehen wir also Anpassungserscheinungen an
die äusseren Lebensbedingungen. Es ist einleuchtend, dass Schwämme,
die dem Eintrocknen oder Einfrieren ausgesetzt sind, sich durch
besondere Vorrichtungen dagegen schützen müssen. Anderseits wird es
möglich sein, dass Spongilliden, die jahraus jahrein unter denselben
Bedingungen leben, der Gemmulae entbehren können. Diesen Gedanken
findet man zuerst ausgesprochen bei W. Marshall[82] und in der
That giebt es solche Süsswasserschwämme ohne Gemmulae. Dybowski
hat an den bis 100 m tief im Baikalsee lebenden Lubomirskien
nie Gemmulae[S. 202] gefunden. Lieberkühn gab an, dass in der Spree
in Berlin Schwämme überwintern, ohne vollständig in Gemmulae zu
zerfallen; Metschnikoff hat dies in Russland bestätigt,
Potts hat in Amerika eine Anzahl ähnlicher Fälle beobachtet und
Wierzejski fand solche Schwämme (nach brieflicher Mitteilung)
in Galizien. In allen diesen Fällen überwintert der Schwamm mit seinem
Weichteil, in dem aber immer mehr oder weniger Gemmulae gebildet
worden sind. Dass es aber auch bei uns Schwämme giebt, die überhaupt
nicht mehr zur Gemmulation schreiten, ist neuerdings bewiesen worden;
an der im Tegeler See lebenden Ephydatia fluviatilis kommen
nach Beobachtungen, die sich über einen Zeitraum von sechs Jahren
erstrecken, Gemmulae überhaupt nicht mehr vor (Weltner). Auch
scheint es [Marshall[82], Potts, Hinde], dass
anderwärts grosse Schwammexemplare durch ununterbrochenes Fortwachsen
während ein oder mehrerer Jahre zustandekommen.
Wir kennen also auch bei uns einige Ausnahmen von der Regel, dass alle
Süsswasserschwämme im Herbst unter Gemmulabildung zerfallen. In unserer
Zone scheint nur bei Ephydatia fluviatilis die Überwinterung des
Weichteils vorzukommen, während alle anderen einheimischen Schwämme zum
Winter absterben und nur ihre Gemmulae gefunden werden. — Auch ist
es nicht richtig, dass die Gemmulae sich bloss im Winter finden. Sie
kommen auch an verschiedenen Schwämmen, Ephydatia fluviatilis
und Mülleri, in den Sommermonaten vor und finden sich bei
ersterer Art neben männlichen und weiblichen Keimstoffen (Götte,
Weltner).
Über die Entwickelung des Keimes zum jungen Schwamm liegen nur wenig
übereinstimmende Nachrichten vor. In Anbetracht, dass dieser Gegenstand
einer erneuerten Untersuchung bedarf, unterlassen wir weitere
Auseinandersetzungen.
c) Atmung, Nahrungsaufnahme,
Verdauung und Exkretion.
In der Physiologie der Spongien sind diese Fragen am wenigsten
aufgeklärt. Offenbar geschieht die Atmung während des beständig den
Schwamm durchlaufenden Wasserstromes und es wird auch durch diesen
Strom zugleich die Nahrung herbeigeführt. Wenn[S. 203] andere Schwämme
durch besondere Pigmente atmen (Mereschkovsky), so müssen
weitere Untersuchungen zeigen, in wie weit solche Pigmente bei den
Spongilliden verbreitet sind. Es ist aber bisher noch nicht mit
Sicherheit entschieden, wo im Schwamme geatmet wird. Dasselbe gilt von
der Verdauung. Aus den zahlreichen Fütterungsversuchen, welche man
mit Farbstoffkörnchen bei Schwämmen und zwar zuerst bei Spongilliden
gemacht hat, geht hervor, dass es die Geisselkammerzellen sind, welche
die Nahrung aufnehmen (Carter, Lieberkühn, Heider,
Metschnikoff, Lendenfeld), wenn auch an einzelnen
Schwämmen beobachtet wurde (Metschnikoff, Topsent),
dass gerade diese Zellen von Karmin frei blieben. Die ausgedehnten
Fütterungsversuche Lendenfelds mit verdaulichen Stoffen
zeigen, dass die Geisselzellen diese aufnehmen, zerteilen und an die
Wanderzellen abgeben. Welcher Art ist nun die Nahrung der Schwämme? Es
sind wahrscheinlich zerfallene organische Stoffe, welche mit dem Wasser
in den Schwamm eingeführt werden. Die nicht brauchbaren Stoffe werden
von den Kragenzellen später wieder ausgeschieden, die brauchbaren
werden in mehr oder weniger assimiliertem Zustande an die Zellen der
Zwischenschicht, welche jedenfalls den Nahrungstransport besorgen,
abgegeben; auch die Exkretion dürfte von den Geisselzellen besorgt
werden [Lendenfeld[83]]. Diese Anschauung gilt auch für unsere
Schwämme.
Dennoch muss es möglich sein, dass sich die Spongilliden auch
von lebenden Infusorien und anderen Protozoen ernähren. Denn
Lieberkühn und nach ihm Metschnikoff sahen, wie in die
Spongillide geratene Protozoen dort verdaut wurden. Gewöhnlich findet
man aber in einem Süsswasserschwamm keine grösseren Organismen, es
sei denn, dass diese als Parasiten in ihm leben (s. unten). Auch die
Thatsache, dass die Süsswasserschwämme in dem fliessenden Wasser der
Städte, in welches Abfälle der unglaublichsten Art geraten, äusserst
üppig entwickelt sind, während man sie in Teichen mit klarem Wasser in
geringer Anzahl trifft, spricht für Lendenfelds Anschauung.
[S. 204]
d) Wachstum und Lebensdauer.
Im allgemeinen scheint den Spongien ein schnelles Wachstum eigen zu
sein und aus den spärlichen Angaben, die betreffs dieser Frage bei den
Spongilliden vorliegen, zu schliessen, trifft das auch für diese zu.
Schon Eper gab 1794 an: „Ihr Wachstum ist sehr geschwinde“.
Carter sah in Bombay eine Spongillide in noch nicht drei
Monaten einen Durchmesser von drei Zoll erreichen. Ein energisches
Wachstum unserer Schwämme findet jedenfalls im Frühjahr statt, wenn
das Wasser wärmer zu werden beginnt. Auch geht die Entwickelung der
jungen Schwämme aus den Gemmulae schnell von statten und man sieht, wie
solche aus den im Skelett liegenden (Eusp. lacustris) oder in
einer basalen Schicht abgelagerten (Sp. fragilis, Trochosp.
erinaceus) Gemmulae entstandenen Schwämme in kurzer Zeit bis zur
Fortpflanzung eine bedeutende Grösse erreichen. — Potts ist der
Ansicht, dass der aus einer Gemmula entstandene Schwamm bis zur
Zeit der wieder eintretenden Gemmulation — also vom Frühling bis zum
Herbst — eine Grösse erreicht hat, um nun zwölf oder mehr Gemmulae
zu bilden. Kommen von diesen nur die Hälfte im nächsten Frühlinge
aus, so soll der aus ihnen entstandene Schwamm am Ende des zweiten
Jahres so gewachsen sein, dass er wenigstens sechs mal so gross als im
ersten Jahre ist. So würde in wenigen Jahren ein Schwamm von mehreren
Zoll Durchmesser zustandekommen. — Die Grösse, welche die aus Larven
entstandenen Schwämme im ersten Jahre erreichen, ist sehr verschieden
und richtet sich nach der Zeit, wann die Larve aus dem Mutterkörper
ausschwärmte. So werden Larven, welche sich schon im Juni festgesetzt
haben, bis zur Zeit, zu welcher die aus ihnen entstandenen Schwämme
unter Gemmulabildung absterben, also im September und Oktober, zu
grösseren Exemplaren angewachsen sein, als solche Larven, welche erst
im August entstanden waren. In der That finden wir denn auch im Herbst
unter den einjährigen Schwämmen Exemplare der verschiedensten Grösse.
Die kleinsten sind kaum 2 mm gross, andere über 2 cm.
Diese grösseren können aber durch Verwachsen mehrerer Exemplare,[S. 205] die
dicht bei einander sassen, entstanden sein. Jedenfalls werden alle
diese aus Larven entwickelten Schwämme in demselben Jahre nicht mehr
geschlechtsreif. Sie zerfallen im Herbst in Gemmulae und man kann
leicht beobachten, wie die kleinsten Exemplare eine einzige Gemmula,
die grösseren zwei, drei Gemmulae u. s w. bilden.
Man sieht aus diesen Angaben, wie wenig wir über die
Wachstumsschnelligkeit und Grösse, welche die gemmulae erzeugenden
Schwämme erreichen, wissen. Nicht viel anders steht es mit den
perennierenden Schwämmen. Wir haben schon oben erwähnt, dass
die grossen Spongillidenexemplare, welche man gefunden hat —
Lubomirskia im Baikalsee, Uruguaya im Uruguay und andere
(s. Potts) —, wahrscheinlich durch ununterbrochenes Wachstum
während einer längeren Zeitdauer zustandekommen. Auch die im Tegelsee
lebenden grossen Exemplare von Ephyd. fluviatilis entstehen
offenbar in derselben Weise. Das Wachstum dieser perennierenden Art
ist während des Winters sehr gering. Bringt man ihnen zu dieser Zeit
grössere Wunden bei, so verwachsen diese zwar während des Winters,
irgend welche bedeutendere Grössenzunahme findet jedoch nicht statt.
Ähnliches hat schon Lamouroux von den Spongien überhaupt
angegeben.
Diesen Auseinandersetzungen steht die Ansicht gegenüber, dass die
Schwämme durch den Prozess der Fortpflanzung dem Tode geweiht sind
(Laurent, Götte). Es ist allerdings richtig, dass
man Süsswasserschwämme zur Zeit der geschlechtlichen Fortpflanzung
absterben sieht. Die Ausbildung der Keimstoffe und ihrer Ernährung
durch die mütterliche Spongillide zerstört zumteil und schwächt dessen
Gewebe. Allein es ist ebenso richtig, dass andere Exemplare auch
nach der vollendeten Ausbildung der Geschlechtsprodukte weiterleben
(Weltner). Absterbende Schwämme trifft man zu jeder Jahreszeit
an, und bei der perennierenden Form haben wir den Tod gerade nach
Überstehung der Winterzeit häufiger gesehen. Da man nun im Sommer
stets nur entweder männliche oder weibliche Schwämme und keine Neutra
antrifft, und alle diese Schwämme im Sommer nach der Fortpflanzung[S. 206]
absterben müssten, so erklärt man sich nicht, wie man zu jeder Zeit
Exemplare von der verschiedensten Grösse findet.
e) Bewegung.
In seinen vorzüglichen Beobachtungen über die Bewegungserscheinungen
der Süsswasserschwämme teilt Lieberkühn dieselben ein in
solche, welche die einzelne Zelle betreffen, und solche, welche der
ganze Schwamm oder ein grösserer Teil desselben ausführen. Zu den
ersteren gehört die amöboide Beweglichkeit der Zellen im Schwammkörper,
die Kontraktion und die Neubildung gewisser in diesen Zellen und
den Geisselzellen vorkommenden mit Flüssigkeit gefüllten Alveolen,
die Zusammenziehung und Ausdehnung der kontraktilen Faserzellen,
das Vergehen und Entstehen der Poren, die Bewegung der Geisseln der
Kragenzellen und der Spermatozoen. Lieberkühn berichtet über die
Bewegungserscheinungen der amöboiden Zellen in ausführlicher Weise.
„Die Bewegungen“, sagt er, „sind äusserst langsam und fast niemals
direkt sichtbar. Es entsendet eine Zelle lange spitze Fortsätze,
welche ihren Durchmesser bedeutend übertreffen, eine andere, entfernt
liegende Zelle schickt ihr gleiche, eben so lange entgegen; es dringen
auch Körnchen in die entsandten Fortsätze hinein; bald verschwinden
die Fortsätze wieder und treten neue an einer andern Stelle der Zelle
hervor, dabei ändert die Zelle selbst beständig ihre Gestalt; wenn sie
kugelig war, wird sie eiförmig oder vieleckig, oder breitet sich in
eine dünne Scheibe aus; die Kerne von zwei Zellen nähern sich bisweilen
so, dass man glaubt, sie gehören einer Zelle an, und rücken alsdann
bald wieder aus einander; oft sieht man auch nur lange und breite
Streifen im Gewebe, welche sich spalten und wieder vereinigen, ohne
dass man eine Zelle aufzufinden vermag, zu der sie gehören.“ Wir müssen
es uns versagen, alle die ausgezeichneten Beobachtungen mitzuteilen,
welche Lieberkühn uns überliefert hat. Nach einer später
anzugebenden Methode kann man sich geeignetes Material beschaffen,
an welchem sich alle diese Beobachtungen wiederholen lassen, und wir
empfehlen die[S. 207] Spongillide als ein sehr dankbares Objekt zum Studium
der Bewegungserscheinungen.
Was die Bewegung einzelner Teile des Schwammes anlangt, so beginnen wir
mit dem Oskularrohr, weil die Gestaltsveränderungen desselben schon
den früheren Beobachtern aufgefallen sind: Dutrochet (1828) und
Bowerbank (1857) beschreiben dieselben in ausführlicher Weise.
Aber Lieberkühn erst wies nach, dass diese Veränderungen auf die
Bewegung der Zellen zurückzuführen sind. Ein solches Oskularrohr sieht
man tagelang unverändert an derselben Stelle, während es zu anderen
Zeiten in wenigen Minuten verschwindet und nach geraumer Zeit wieder
entsteht; oder es wird an einer anderen Stelle ein neues gebildet.
Eine schon vorhandene Röhre kann sich gabeln und jedes Röhrchen erhält
ein Ausströmungsloch. Die Zusammenziehung einer Oskularröhre geschieht
sehr langsam. Nur auf einen plötzlichen Reiz mit der Nadel oder durch
bedeutende Temperaturveränderung des Wassers, durch Alkohol, Säuren
etc. geschieht dieselbe sehr schnell. Deshalb werden beim Abtöten der
Süsswasserschwämme in Alkohol die Ausströmungsröhren stets bis fast
zum Verschwinden gebracht. — Der im Leben ausgedehnte röhrenförmige
Fortsatz zeigt eine ziemlich glatte Oberfläche; indem er sich
zusammenzieht, wird die Wandung zusehends dicker und höckerig durch die
zusammengedrängten Zellen, deren Grenzen jetzt deutlich sichtbar sind.
Die Kontraktion kann soweit gehen, dass die ganze Röhre die Gestalt
eines Zellenhaufens annimmt oder gänzlich verschwindet. Die kürzeste
Zeit der Zusammenziehung ist eine Minute; es ist aber die Kontraktion
meistens nur eine vorübergehende und die Ausdehnung ist der bleibende
Zustand. Ganz dasselbe gilt für die Zellen selbst und Lieberkühn
vergleicht ihre Zusammenziehung und Ausdehnung mit der des Muskels.
Die besprochene Verlängerung und Verkürzung der Röhre ist nicht zu
verwechseln mit einem ganz anderen Vorgang, der auf Wachstum beruht.
Indem Zellen aus dem Schwamminnern in die Röhre einwandern, kann sich
dieselbe verlängern oder auch verdicken. Wir haben daher bei den
Bewegungserscheinungen[S. 208] einzelner Teile zwischen blosser Bewegung und
Bewegung verbunden mit Wachstum zu unterscheiden.
Ähnliche Bewegungen sehen wir auch an der äusseren Haut des
Schwammkörpers, die von Carter und Lieberkühn genauer
geschildert worden sind.
Auch die Art und Weise, in welcher sich kleine, aus dem Schwammkörper
geschnittene Stückchen auf eine Glasplatte anheften, rechnet
Lieberkühn hierher, es gehört aber diese Erscheinung ebenso
wie die Anheftung der schwimmenden Larve an ihre Unterlage oder wie
die des aus der Gemmula kriechenden Keimes unter die zuerst genannten
Bewegungen. Das diesbezügliche findet man bei Lieberkühn,
Carter, Götte und Maas.
Es kommt sogar bei unserem Schwamm eine Bewegung des ganzen Körpers
vor. Es sind freilich nur junge Spongilliden, bei denen diese
Erscheinung beobachtet wird. Lieberkühn sah, wie sich ein 2½
Monate alter Schwamm beständig hin und her bewegte, ohne eigentlich
vom Platze zu rücken. Aber an einem jungen Schwamme konstatierte er,
wie sich derselbe von seiner Unterlage ablöste und an einer anderen
Stelle festsetzte. Auch Marshall[84] hat über eine solche
Ortsveränderung Mitteilung gemacht.
6. Systematik der einheimischen
Arten.
Die erste Unterscheidung der Süsswasserschwämme geschah nach
ihrer äusseren Gestalt. Wie wir aber gesehen haben, kann man nur
eine einzige Art (Eusp. lacustris) und diese auch nur im
ausgewachsenen Zustande an ihrer busch- oder baumförmigen Gestalt
erkennen. Man hat sich daher genötigt gesehen, die Gestalt der Skelett-
und Gemmulaenadeln zur systematischen Unterscheidung zu benutzen
(Ehrenberg, Lieberkühn). Ausser diesen Skelettelementen
hat man neuerdings auch den Bau der Gemmulaschale zur Erkennung
benutzt. In der That bietet die Beschaffenheit der genannten Teile
die einzige Möglichkeit, die Arten von einander zu unterscheiden,
wenn man nicht gewisse histiologische Besonderheiten herbeiziehen
will. — Nun sind aber die Gemmulae,[S. 209] die hauptsächlich zur Erkennung
der einzelnen Arten dienen, nicht integrierende Bestandteile des
Spongillidenkörpers und man ist deshalb öfters in die Verlegenheit
gesetzt, einen Süsswasserschwamm nicht bestimmen zu können. Man thut
daher gut, bei der Bestimmung oder beim Sammeln von Schwämmen sich von
dem Vorhandensein der Gemmulae zu überzeugen.
Wir geben im folgenden eine kurze Beschreibung der deutschen Arten
und wollen auch die beiden übrigen europäischen Arten, die sich
wohl bei näherer Nachforschung auch in Deutschland finden werden,
berücksichtigen.
Familie Spongillidae, Süsswasserschwämme.
Skelett aus einachsigen Kieselnadeln bestehend, welche durch Spongiolin
zu einem netzförmigen Gerüst verbunden sind. Bei den einheimischen
Arten unterscheidet man an denselben Haupt- und Verbindungsfasern.
Die Spongiolinsubstanz hüllt entweder die Nadelzüge vollständig ein
oder sie ist schwach entwickelt und verkittet nur die Enden der Nadeln
mit einander. Fleischnadeln vorhanden oder fehlend. Die kugeligen
Geisselkammern münden seitlich in ausführende Kanäle, welche nach ihrer
Vereinigung zu weiteren Bahnen endlich in eine einzige grosse Höhle,
die Kloakenhöhle, sich vereinigen oder getrennt von einander in einen
unmittelbar unter der äusseren Haut liegenden Ausströmungsbezirk von
oft sternförmiger Gestalt sich ergiessen. Ausser der geschlechtlichen
Fortpflanzung kommt noch eine ungeschlechtliche durch innere Keime
(Gemmulae) vor. Sie leben mit einer einzigen Ausnahme nur im süssen
oder im brackischen Wasser. Kosmopolitisch.
Das System und die folgende Beschreibung der Arten ist entlehnt
aus Vejdovskys Darstellung in dem Werke von Potts; den
Beschreibungen haben wir einige Bemerkungen hinzugefügt.
A. Subfamilie Spongillinae (Carter).
I. Genus Spongilla (Autt.).
a) Subgen. Euspongilla (Vejdovsky).
1. Euspongilla lacustris (Autt.).
[S. 210]
b) Subgen. Spongilla (Wierzejski).
2. Spongilla fragilis (Leidy).
B. Subfamilie Meyeninae (Carter).
II. Genus Trochospongilla (Vejdovsky).
3. Trochospongilla erinaceus (Ehrenberg).
III. Genus Ephydatia (Lamouroux).
4. Ephydatia Mülleri (Lieberkühn).
5. Ephydatia fluviatilis (Autt.).
6. Ephydatia bohemica (F. Petr).
IV. Genus Carterius (Potts).
7. Carterius Stepanowi (Dybowsky).
A. Unterfamilie Spongillinae
(Carter).
Gemmulae entweder einzeln oder in Gruppen vereinigt, gewöhnlich
mit einer Luftkammerschicht umgeben, in welcher an beiden Enden
zugespitzte, fast stets gedornte Nadeln liegen.
I. Gattung Spongilla
(Autt.).
Mit langen, glatten Skelettnadeln und kurzen, geraden oder gekrümmten,
glatten oder rauhen Fleischnadeln. Gemmulae entweder nackt oder mit
einer äusseren Luftkammerschicht, in welcher die Belagsnadeln entweder
tangential oder radiär oder ganz unregelmässig liegen.
a) Untergattung Euspongilla (Vejdovsky).
Gemmulae immer einzeln im Schwamme liegend.
1. Euspongilla lacustris (Autt.).
Der Schwamm bildet gewöhnlich baum- oder buschförmig verzweigte
Massen auf einer krustenförmigen Basis von geringerer (Fig. 36)
oder grösserer (Fig. 37 S. 212) Ausdehnung. Unter Umständen, z. B.
an stark fliessenden Stellen, kommt es nicht zur Ausbildung der
charakteristischen fingerförmigen Fortsätze und Zweige, sodass
klumpenförmige Massen entstehen (Fig. 38). Kleinere Exemplare sind
einfach krustenförmig, ebenso gestaltete Exemplare[S. 211] von nicht
unbeträchtlicher Grösse findet man selbst in ruhigen Gewässern um
Teichrohrstengel gewachsen; an solchen Exemplaren in ruhigem Wasser
tritt aber früher oder später die Bildung von Ästen auf. Die Farbe
ist grasgrün, gelblich, grauweiss oder braun.[S. 212] Das Skelett besteht
aus Gerüst- und Fleischnadeln. Die Gerüstnadeln sind gerade oder
leicht gekrümmt, scharf, aber allmählich zugespitzt und glatt. Sie
sind zu Bündeln mit einander vereinigt, welche durch stark entwickelte
Spongiolinsubstanz ganz eingehüllt werden und lange, starke Stäbe
bilden (Hauptfasern), welche die Zweige der Länge nach durchziehen
und nach der Peripherie derselben dünnere Äste aussenden. An ihren
Enden laufen diese Hauptfasern dünn aus. Die stärkeren Bündel in
der Achse eines Zweiges am Schwamme bestehen aus 20–30 vollständig
in der Kittsubstanz liegenden Nadeln. Diese Längsfaserzüge oder
Hauptfasern sind durch wenige kurze Nadelbrücken mit einander
verbunden, welche in unregelmässigen Abständen von einander entfernt
sind. Diese Verbindungsfasern sind meist nur an ihren Enden durch
Spongiolinsubstanz an die[S. 213] Hauptfasern angekittet oder sie liegen
ganz in der Kittmasse eingeschlossen. Bei solchen Exemplaren,
welche einfache Krusten bilden, stehen die Hauptfasern senkrecht
auf der Unterlage. — Durch den geschilderten Bau des Gerüstes
erlangt der Schwamm eine gewisse Festigkeit und ist schwieriger
zerreissbar als Sp. fragilis und Eph. fluviatilis.
— Die Spongiolinsubstanz ist in Kalilauge unlöslich, wodurch sich
Eusp. lac. von allen anderen Arten unterscheiden lässt
(Dybowski). — Die Fleischnadeln sind in wechselnder Anzahl
vorhanden; sie können in einem Schwamm in ungeheuerer Menge auftreten
(var. Lieberkühnii Noll), in anderen häufig, in noch anderen
sehr sparsam sein. Sie sind meist leicht gekrümmt und vollständig
mit feinen Dörnchen besetzt, selten sind sie glatt. Die Gemmulae
treten in verschiedenen Formen auf. Es giebt nackte, d. h. einer
äusseren Luftkammerschicht entbehrende, die nur wenige oder gar keine
Nadeln tragen. Andere Gemmulae sind mit einer dünneren oder dickeren
Luftkammerschicht bedeckt, welche nach aussen durch eine deutliche
Membran abgeschlossen ist, in anderen Fällen fehlt dieselbe. In dieser
Luftkammerschicht liegen die Belagsnadeln entweder in radiärer oder
in tangentialer Lage auf der Gemmula oder sie sind auf ihr ohne alle
Ordnung zerstreut. Sie stellen kurze, weniger oder mehr, mitunter bis
zum Kreise gekrümmte und mit dicken Dornen versehene Spikula dar, und
sind nur selten ganz glatt. Die Gemmulae liegen im ganzen Schwamme
zerstreut, dessen Weichteil nach ihrer Ausbildung vollständig zu Grunde
geht, während das Skelettgerüst mit den Gemmulae in seinen Maschen oft
den Winter hindurch erhalten bleibt. In anderen Fällen zerfällt auch
dieses. — In stehendem und fliessendem Wasser in ganz Deutschland. Es
scheint die gemeinste Art zu sein und ist auch im finnischen Meerbusen
im Brackwasser gefunden worden (Dybowski).
Fig. 36. Euspongilla lacustris (Autt.). ½ nat. Grösse. Nach dem Leben. An
einem Mauersteine. (19. Juli 1890 gefunden in der Spree, Berlin:
Jannowitzbrücke, Stadtbahnbögen.)
Fig. 37. Euspongilla lacustris (Autt.). Nach dem Leben. ½ natürlicher Grösse.
(6. Juni 1890 Tegelsee.)Fig. 38. Euspongilla lacustris. Auf einem dicken Wollenfaden gewachsen, der an
einer Seite an einem Brückenpfahl eingeklemmt an der Oberfläche des Wassers
flottierte. Spree, Berlin: Waisenbrücke, 26. Aug. 1886. ½ natürlicher
Grösse. (Alkoholexemplar.)
Als eine Lokalform von dieser Art betrachten wir die von Retzer
beschriebene „Spongilla rhenana“. Dieser bisher nur am
Faschinengesträuch im Altrhein bei Eggenstein unweit Karlsruhe
gefundene Schwamm überzieht als dünne Kruste Holzstücke, Gesträuch
und dergl. und sendet wenige, kleine Fortsätze aus oder wächst auch
an manchen Stellen zu dicken Klumpen an. Seine[S. 214] Farbe ist grün. Die
Skelettnadeln sind gerade oder leicht gebogen und gehen entweder
plötzlich in eine scharfe oder allmählich in eine weniger scharfe
Spitze über. Sie bilden zu Bündeln vereinigt ein dichtes Netz.
Die Gemmulae haben die Form und Grösse derjenigen von Eusp.
lacustris, der Porus zeigt aber einen breiten flachen Trichter.
Auf der inneren Hülle der Gemmulaschale liegt eine sehr dünne
Luftkammerschicht, in welcher die Zellen in zwei- bis dreifacher Lage
übereinander liegen. In dieser Schicht sind die Belagsnadeln gewöhnlich
tangential, wenige radiär angeordnet. Die sehr variable Gestalt lässt
drei Hauptformen unterscheiden. Die häufigsten sind solche, welche
den Skelettnadeln ähneln, andere, in geringer Anzahl vorhandene, die
sich auch einzeln im Weichteil finden, sind glatt und in gleichen,
nicht sehr grossen Abständen an den Enden zweimal geknickt, drittens
giebt es leicht gebogene, in der Mitte verdickte Nadeln. Die Gemmulae
liegen überall im Schwamm zerstreut. Bemerkenswert ist, dass an ihnen
Nebenpori vorkommen, deren drei bis sechs gesehen wurden. Ungefähr jede
zehnte Gemmula hat neben dem Hauptporus einige seitliche Trichter.
Dieser aus Vejdovsky und Retzer entnommenen Beschreibung
fügen wir nur hinzu, dass der Schwamm auch grössere, verzweigte Äste
treibt und dass unter den doppelt geknickten Gemmulaenadeln auch
fein bedornte vorkommen. — Vejdovsky hat diese Retzersche
Art beibehalten. Wierzejski[80] betrachtet dieselbe nur als
eine „lokale Form, vielleicht eine Abnormität der Euspongilla
lacustris“. Wir schliessen uns der Auffassung, dass man es hier mit
einer Lokalform zu thun habe, an.
b) Untergattung Spongilla (Wierzejski).
Zwei bis dreissig Gemmulae liegen in einer stark entwickelten, deutlich
zelligen Luftkammerschicht eingebettet oder die Gemmulae bilden eine
pflastersteinartige Kruste in ebenso gestalteter Luftkammerschicht an
der Basis des Schwammes. In der Luftkammerschicht liegen rauhe und
glatte Nadeln zerstreut.
Der Schwamm ist nie verzweigt und scheint meist eine glatte Oberfläche
zu haben. Die Farbe ist weisslich, hellgrau, graubraun, braun,
seltener grün. Die Skelettnadeln sind fast gerade oder nur leicht
gebogen, scharf zugespitzt und glatt. Die Spongiolinsubstanz ist
schwach entwickelt, der Schwamm ist daher sehr leicht zerreissbar. Die
Belagsnadeln der Gemmulae bilden eine dichte Kruste auf ihr, sie sind
gerade oder gekrümmt und tragen viele kleine Dornen. Sie übertreffen
gewöhnlich an Länge und Dicke diejenigen von Eusp. lacustris.
Die kleinen, rundlichen Gemmulae tragen ein verlängertes, gewöhnlich
etwas gebogenes Porusrohr, welches aus der dicken Luftkammerschicht
hervorragt. Die grossen Zellen der Luftkammerschicht sind radiär um die
Gemmulae geordnet. Diese erscheinen in zwei Formen, die an der Basis
des Schwammes liegenden sind flach, die im Schwammkörper in Gruppen
zu zwei oder drei bis dreissig und mehr vereinigten Gemmulae bilden
kugelige oder halbkugelige Massen.
Lebt in stehendem und fliessendem Wasser und gehört in Deutschland zu
den gemeineren Arten.
B. Unterfamilie Meyeninae (Carter.)
Gemmulae gewöhnlich einzeln. Sie sind von einer Luftkammerschicht
umgeben, in welcher Amphidisken (Stäbe mit einer Querscheibe an jedem
Ende) in einer oder in mehreren Lagen übereinander vorkommen. Die
Amphidisken haben gezackte oder ganze Ränder.
Fig. 39. Spongilla fragilis (Leidy). Nach einem in Alkohol konservierten Stück.
½ natürl. Gr. (Tegelsee, 6. Juli 1888.)
II. Gattung Trochospongilla (Vejdovsky).
Ausgezeichnet durch die glatten (bei einer ausländischen Art) oder
rauhen Skelettnadeln und durch den glatten Rand der Amphidisken, welche
an der Basis einer hohen Luftkammerschicht eingebettet liegen.
Der Schwamm überzieht als weissliche oder strohgelbe Kruste von
geringerer oder grösserer Ausdehnung fremde Körper. Die Skelettnadeln
sind scharf zugespitzt und bis auf die beiden Enden mit sehr starken
Dornen besetzt. Die Spongiolinsubstanz ist stark entwickelt und setzt
dem Zerreissen des Schwammes einen Widerstand entgegen, der grösser als
bei den übrigen Meyeninen ist. Die Amphidisken haben die Form einer
Garnspule. Die Luftkammerschicht besteht aus radiär gestellten Säulen,
deren jede aus übereinander liegenden Zellen gebildet wird. In dieser
Schicht liegen oft Nadeln, welche den Gerüstspikula gleichen, aber von
geringeren Dimensionen sind.
Auch diese Art lebt in stehenden und fliessenden Gewässern Deutschlands
und scheint selten zu sein. Sie wurde bei uns bisher gefunden: Sabor
in Schlesien (Ehrenberg), Spree in Berlin (Lieberkühn,
Weltner), Tegelsee bei Berlin (Weltner), Hellensee bei
Lanke (Weltner).
III. Gattung Ephydatia (Lamouroux).
Die Skelettnadeln sind entweder ganz glatt oder ganz rauh oder es
finden sich beide Sorten in einem Schwamm. Die mit gezackten Rändern
versehenen Amphidisken liegen in ein-, zwei- oder selbst dreifacher
Schicht in der Luftkammerschicht. Die Amphidisken sind entweder alle
von gleicher Länge oder sie sind ungleich lang. Fleischnadeln vorhanden
oder fehlend.
Fig. 40. Trochospongilla erinaceus (Ehrenberg) nach Vejdovsky. ½
natürl. Grösse.
4. Ephydatia Mülleri (Lieberkühn).
Diese Art bildet Krusten mit einer glatten oder unregelmässigen
Oberfläche, an der man oft kurze Zapfen (Fig. 41) oder blattförmige
oder mäandrisch gewundene Fortsätze sieht. Selten scheinen verzweigte
Exemplare vorzukommen. Die Farbe ist weiss, gelb, gelbbraun oder
hellgrün. Die Skelettnadeln sind entweder ganz glatt oder ganz
rauh, oder es finden sich sowohl glatte als rauhe Nadeln in ein und
demselben Schwamme. Die Rauhigkeit[S. 217] der Nadeln ist entweder nur
eine geringe oder sie tritt sehr deutlich als kleine Höcker auf,
die aber stets kleiner sind als bei Trochosp. erinaceus. Die
Nadeln sind gerade oder schwach gekrümmt, scharf zugespitzt und zu
Bündeln vereinigt, deren Kittmasse weniger stark als bei der eben
genannten ist. Die Luftkammerschicht ist in geringerer oder grösserer
Mächtigkeit vorhanden. Die Amphidisken bilden entweder nur eine,
zwei oder selten drei Lagen um die Gemmula. Im letzteren Falle ist
die dritte Lage keine ununterbrochene. Die Amphidisken haben einen
dicken Schaft; die Zähne der Endscheiben sind entweder glatt oder an
ihren Rändern gezähnt. — Ganz charakteristisch für diese Art sind
die Blasenzellen des Weichteiles, welche modifizierte, mit einer sehr
grossen, amylumhaltigen Flüssigkeitsalveole ausgestattete Zellen der
Bindesubstanzschicht sind [Wierzejski[79]].
Diese Art wurde von Lieberkühn nach Exemplaren aus der Spree
aufgestellt. Die Amphidisken der in diesem Flusse lebenden Eph.
Mülleri zeigen einen kurzen Schaft und Endscheiben, welche wenig
aber tief eingezackt sind.
Fig. 41. Ephydatia Mülleri (Lieberkühn). ½ natürl. Grösse, nach einem
getrockneten Exemplar. (Tharandt, Schlossteich.)
In stehenden und fliessenden Gewässern verbreitet in ganz Deutschland.
[S. 218]
5. Ephydatia fluviatilis (Autt.).
Krustenförmige Massen von sehr wechselnder Gestalt bildend, es
kommen ganz flache Krusten vor, daneben andere, die mehr klumpige
Form erreichen. Die Oberfläche ist entweder glatt (Fig. 42) oder mit
seichten Buckeln, oder mit spitzigen kurzen Fortsätzen (Fig. 43), oder
mit rippenförmigen, öfters gewundenen Erhabenheiten versehen. Wenn der
Schwamm Verzweigungen zeigt, so rühren diese von dem Substrat her,
welches derselbe überzogen hat (Fig. 44).
Fig. 42. Ephydatia fluviatilis (Autt.) nach dem Leben. ½ natürl.
Grösse. (6. Juni 1890, Tegelsee.)Fig. 43. Ephydatia fluviatilis (Autt.) nach einem in Alkohol konserv. Stück.
½ natürl. Grösse. (Spree, Berlin: Waisenbrücke, 26. Aug. 1886.)
Die Farbe ist smaragdgrün, hellgrün, hellisabellgelb, schmutzig-weiss
oder weiss. Die glatten, schlanken, allmählich oder plötzlich scharf
zugespitzten, leicht gekrümmten Skelettnadeln sind wie bei[S. 219]Sp.
fragilis nur durch wenig Spongiolinsubstanz mit einander verbunden;
der Schwamm ist daher leicht zerreissbar und brüchig. Öfters finden
sich unter den Nadeln auch kurze, dicke oder auffallend lange und dicke
Formen. Die Gemmulae sind gelblich und haben eine dicke Hülle, die
Luftkammerschicht hat eine äussere Kutikula. Die Amphidisken sind daher
ganz in der Luftkammerschicht eingeschlossen und bilden eine einfache
Lage in derselben. Der Schaft ist dünn, glatt oder bedornt und oft in
der Mitte etwas eingezogen. Er ist doppelt so lang, als der Durchmesser
der Scheiben beträgt. Diese sind am Rande durch ihre zahlreichen, nicht
tiefen Einschnitte ausgezeichnet. Doch finden sich häufig Exemplare,
bei denen die mit langem Schafte ausgestatteten Scheiben nur wenige
tiefe Einschnitte zeigen.
Sehr gemein in stehenden und fliessenden Gewässern in Deutschland.
Auch im Brackwasser, so in der Untertrave bei Travemünde, deren Wasser
0.34% Salzgehalt zeigt (Lenz). Auch in der Dievenow bei Cammin
in Pommern, welche bis zu der genannten Stadt von dem eindringenden
Seewasser durchschnittlich 2–3mal im Monat während eines Jahres schwach
versalzt werden soll (Weltner).
Der Schwamm bildet kleine grüne Polster. Die Skelettnadeln sind gerade
oder leicht gekrümmt und mitunter fein bedornt. Diese Art ist durch
ihre zahlreichen Fleischnadeln von den übrigen Meyeninen unterschieden.
Die Nadeln sind gerade oder ein wenig gebogen und mit dornenähnlichen,
oft am Ende gerundeten Fortsätzen bewehrt. Die Gemmulae tragen eine
grosse Pore, deren[S. 220] Rand nach oben in einen breiten Trichter ausläuft,
selten ist an Stelle des Trichters ein kurzes Rohr entwickelt. Die
Amphidisken sind fast alle gleich lang, die längeren ragen über die
Luftkammerschicht hervor. Ihre gedornten Schäfte sind schlank und
länger als der Durchmesser der Scheiben, welche ziemlich regelmässig
und tief gezackt sind. Die Zacken sind fein gekerbt.
Dieser Schwamm, welcher vielleicht nur eine Übergangsform zu der
folgenden Gattung darstellt, ist bisher nur bei Kvasetice im Bezirk
Deutschbrod (Böhmen), an Euspongilla lacustris sitzend, von
Fr. Petr gefunden.
IV. Gattung Carterius (Potts).
Mit glatten Skelettnadeln und dornigen Fleischspikula. Die Gemmulae
tragen ein langes, gerades Porusrohr, dessen Spitze eine unregelmässig
gelappte Scheibe trägt. In der Luftkammerschicht liegen Amphidisken
von zweierlei Länge, die einen haben eine Länge, welche der Dicke der
Luftkammerschicht gleichkommt, die anderen sind länger und ragen mit
ihrer Scheibe über jene Schicht hinaus.
Fig. 45. Cart. Stepanowi (Dyb.). ½ natürl. Grösse n. d. Leben. (Nach
Petr.)
Der Schwamm ist nur von geringen Dimensionen und bildet (selbständige?)
Verzweigungen. Die Farbe ist blassgrün. Die Skelettnadeln sind glatt,
gerade oder gebogen und scharf zugespitzt. Die zahlreich vorhandenen
Fleischnadeln haben eine gebogene oder gerade Gestalt und sind mit
Dornen besetzt, welche in der Mitte der Nadel am stärksten sind. Das
Porusrohr an den Gemmulae ist gerade oder leicht gebogen und endet in
eine gelappte Scheibe. Die Luftkammerschicht besteht aus zahlreichen
kleinen Zellen. Es sind zweierlei Amphidisken vorhanden; die längeren,
über die Oberfläche der Gemmula hinausragenden betragen etwa ein
Drittel oder die Hälfte aller Amphidisken, welche sämtlich an den
Schäften mit starken Dornen bewehrt sind.
[S. 221]
Gefunden in Europa bisher nur in einem See (Wilikoje) bei Charkow
(Dybowski) und in einem Teich bei Deutschbrod in Böhmen
(Petr).
Anhang.
Wir müssen hier noch einer von Joseph erwähnten Spongilla
stygia gedenken. Diese neue Form wurde in der Grotte Gurk in
Unterkrain von Joseph gefunden und ist völlig durchsichtig.
Die Nadeln sind glatt und an einem Ende keulenförmig verdickt. Nach
Marshall[85] sollen die Gemmulae wirklich fehlen. Es wäre
wünschenswert, diese neue Art genauer zu studieren.
Tabellen zur Bestimmung von Spongilliden sind nach dem jeweiligen
Standpunkte unserer Kenntnis dieser Tiere von Dybowski 1878[86],
Vejdovsky[87], Potts
und Girod[88] gegeben worden.
Wir entwerfen hier einen Schlüssel für die europäischen Formen.
0
Gemmulae mit Amphidisken
1
Gemmulae ohne Amphidisken
5
1
Die Ränder der Amphidisken sind ganzrandig,
Skelettnadeln mit starken Dornen besetzt
Trochospongilla erinaceus.
Die Ränder der Amphidisken sind gezackt,
Skelettnadeln glatt oder rauh
2
2
Fleischnadeln vorhanden; unter den Amphidisken
lassen sich deutlich zwei Sorten, längere und kürzere, unterscheiden
3
Fleischnadeln fehlend; Amphidisken alle ziemlich
gleich lang
4
3
Porus der Gemmula in ein langes Rohr ausgezogen
Carterius Stepanowi.
Porus der Gemmula in einen breiten Trichter
ausgehend oder (selten) in eine kurze Röhre verlängert
Ephydatia bohemica.
4
Amphidisken mit langem Schaft, Endscheiben meist mit
zahlreichen aber nicht sehr tiefen Einschnitten. Luftkammerschicht
mit dünner Kutikula umschlossen. Skelettnadeln glatt. Spongiolinsubstanz
schwach entwickelt
Ephydatia fluviatilis.
Amphidisken mit kurzem Schaft, Endscheiben oft nur
mit wenigen aber tiefen Einschnitten. Luftkammerschicht ohne äussere
Kutikula. Skelettnadeln glatt oder rauh oder beide Sorten vorhanden.
Spongiolin ziemlich reichlich
Ephydatia Mülleri.
5
Gemmulae zu 2–30 in Gruppen in einer dicken, deutlich
zelligen Membran eingeschlossen. Der grösste Teil der Gemmulae bildet an
der Basis des Schwammes eine kontinuierliche einfache Lage, in einer ebenso
gestalteten Membran eingebettet. Fleischnadeln fehlen
Spongilla fragilis.
Gemmulae immer einzeln in den Maschen des
Schwammgerüstes steckend. Fleischnadeln vorhanden
Euspongilla lacustris.
7. Verbreitung.
Wenn diejenigen Spongien, welche man, vielleicht nicht ganz mit Recht,
als eine eigene Familie der Spongillidae anderen Familien
gegenübergestellt hat, auch vorzugsweise Bewohner des süssen Wassers
sind, so haben wir doch schon erfahren, dass auch einzelne Arten im
brackischen Wasser leben können. Wir fügen dem jetzt hinzu, dass auch
in den Gewässern Floridas Spongilliden gefunden worden sind, welche
gelegentlich in versalztem Wasser zu leben gezwungen sind (Potts
1890). Ja, es fehlt auch nicht an solchen Arten, die sowohl das
süsse Wasser als das Meer bewohnen, da die von Pallas im
Baikalsee entdeckte Spongia baicalensis (Pall.) nunmehr auch im
Behringsmeer entdeckt worden ist (Dybowski).
[S. 223]
Alle Süsswasserschwämme leben vorzugsweise auf festen, abgestorbenen
oder doch nicht mehr lebenden Gegenständen; eine Auswahl wird dabei
nicht getroffen. Es kommt ihnen nicht darauf an, einen alten Schuh,
einen Nagel, ein Stück Leder, einen Fetzen Tuch, eine alte Muschel oder
Eierschale zur Unterlage zu benutzen. Seltener wachsen sie auf lebenden
Baumwurzeln oder Konchylien. Schlamm ist ihr ausgesprochener Feind und
nur Potts erwähnt einen Fall, in welchem sich eine Spongillide
auf Schlammboden angesiedelt hatte. Auch in der Spree lebt Eusp.
lacustris im Schlamm; an solchen Exemplaren ist aber stets der im
Schlamme steckende Teil abgestorben.
Die Tiefen, in denen diese Schwämme leben, sind sehr verschieden.
Bei uns werden sie dicht unter der Wasseroberfläche gefunden und sie
steigen bis 4 m hinab. Forel fand im Lac de Joux noch
in 20 m Tiefe eine Spongillide, vermisste die Schwämme aber in
der Tiefenregion des Genfersees (von 15 m an bis zur grössten
Tiefe des Sees von 334 m). Auch bei uns scheinen die Schwämme
in der Tiefe der Seen zu fehlen; Grundproben, in denen Schwammnadeln
gefunden wurden, beweisen nicht das Vorkommen von lebenden Schwämmen
am Boden, da, wie wir in der Einleitung erwähnt haben, absterbende
Spongillidenstücke häufig auf der Wasseroberfläche treiben können.
Dagegen hat Dybowski den Baikalseeschwamm in einer Tiefe
von 100 m gefunden. Dieser Schwamm ist hier in 2–6 m
Tiefe rasenförmig, in 6–25 m baum- oder strauchförmig und in
25–100 m wieder rasenförmig. Fügen wir noch hinzu, dass die im
Bodensee in 12–15 Klaftern Tiefe lebende Spongilla friabilis
von Wartmann (Esper, „Pflanzenthiere“ II. 1794), das
sogenannte „Fischbrot“, wohl kaum ein Schwamm ist, so hätten wir über
die Tiefenverhältnisse das wichtigste erwähnt.
Wir haben schon Gelegenheit gehabt, den Einfluss des Lichtes auf
die Farbe unserer Schwämme zu betonen. Viele und wohl die meisten
sind am Lichte lebend grün. Andere ebenfalls dem Lichte ausgesetzte
zeigen statt des grünen ein braunes Pigment, welches ganz anders
gestaltet ist als jenes. Sowohl die grünen als[S. 224] die braunen Formen
sind an immer beschatteten Lokalitäten gelblich oder gänzlich farblos.
Kräpelin sammelte gelbliche und farblose Exemplare in den Röhren
der Hamburger und de Vries reinweisse Stücke in den Kanälen der
Rotterdamer Wasserleitung.
Die Höhen, in denen Süsswasserschwämme vorkommen, erstrecken sich
nach den bisherigen Beobachtungen bis auf etwa 1900 m. Al.
Brandt erwähnt (Zool. Anzeiger 1879 u. 1880) ihr Vorkommen
in zwei armenischen Alpenseen. In zahlreichen Seen der Schweiz
hat man sie gefunden (Du Plessis-Gouret, Forel,
Imhof). Sie fehlen auch in unseren Gebirgsseen nicht, wie ihr
Vorkommen im Schwarzwald (Retzer) und in den Maaren der Eifel
(Zacharias) beweist, während sie im Kleinen und Grossen Teiche
des Riesengebirges vermisst werden.
Die Art der Verbreitung.
Die Verbreitung kann durch Teilung, Larven und Gemmulae geschehen.
Was es mit der Teilung auf sich hat, ersieht man aus den
Auseinandersetzungen auf S. 200. — Die Verbreitung durch Larven und
Gemmulae wird bei den in Flüssen lebenden Schwämmen vorwiegend auf
passivem Wege geschehen; die Fortpflanzungskörper werden, dem Laufe des
Stromes folgend, flussabwärts schwimmen, bis sie an einem Gegenstande
haften bleiben. Nur in den stillen Buchten können die Larven, wie es
im allgemeinen in den von der Strömung weniger beeinflussten Teichen
und Seen der Fall ist, aktiv zur Verbreitung der Art dienen. Auch
hier wird diese eine langsame sein, da die Larven schwerlich grössere
Reisen in horizontaler Richtung auszuführen imstande sind. Dazu ist
ihre Schwärmzeit zu kurz. Wir haben zwar Larven, welche im Oktober
ausgeschwärmt waren, noch längere Zeit lebend im Aquarium beobachtet;
nach tagelangem Umherirren starben sie aber, ohne sich festzusetzen, am
Boden des Gefässes ab.
Weit wichtiger für die räumliche Verbreitung der Art sind die Gemmulae
und wir sehen nun, dass diese einem doppelten Zweck dienen. Sie
ermöglichen erstens die Fortexistenz der Spezies überhaupt (s. oben S.
201) und zweitens deren Verbreitung.[S. 225] Wenn es auch eine Anzahl Arten
giebt (z. B. Sp. fragilis und Troch. erinaceus bei uns),
bei denen ein Teil der Gemmulae ganz fest an ihre Unterlage gekittet
ist, so fallen doch auch hier die lose im Skelett liegenden aus, wie es
bei den meisten Gemmulae der übrigen Arten der Fall ist. Es wird also
an einem Süsswasserschwamm der eine Teil der Gemmulae den Schwamm an
derselben Stelle wieder erzeugen, an welcher der Mutterschwamm wuchs,
der andere Teil der Gemmulae wird zerstreut. Isoliert man frische,
ausgebildete Gemmulae unserer Arten, so sieht man, dass der eine Teil
an die Oberfläche des Wassers steigt, während der andere untersinkt.
Wir kennen die Ursache dieser Erscheinung noch nicht, obwohl schon
Turpin (1838) schwimmende Gemmulae bekannt waren. — Die Art, in
welcher die sich vom Schwamme loslösenden Gemmulae verbreitet werden
können, hängt vom Bau der Hüllen dieser Körper ab. Sie ist in einer
kurzen Schrift vorzüglich erläutert worden [Marshall[85]], deren
Resultate wir hier wiedergeben.
Bei einer Anzahl tropischer Arten sind die Gemmulae mit einer dicken
Luftkammerschicht ausgerüstet. Zur Zeit der Trockenheit werden solche
Gemmulae leicht durch den Wind hinweggetragen und zerstreut. „Passiv
beweglich mit aërostatischem Apparat — Flugform (der trockenen
Jahreszeit).“
Dann giebt es Gemmulae, welche auf dem Wasser schwimmend zur
Verbreitung dienen. Das ist z. B. bei Eusp. lacustris der Fall.
Die Gemmulae dieser Kategorie tragen Belagsnadeln, mit denen sie sich
hie und da anheften. „Passiv bewegliche Schwimmform mit Ankerapparat
zum Treiben auf der Oberfläche vor dem Winde.“
Wieder andere Gemmulae, z. B. Ephyd. fluviatilis, sind durch
ihren Amphidiskenbelag schwerer und besser geschützt als die der
vorhergehenden Sorte. Sie sinken zu Boden und werden fortgerollt; sie
kommen ihrer Schwere wegen eher zur Ruhe. „Schwimmform mit Hemmapparat
zur langsamen Fortbewegung in fliessendem Wasser.“
Die geologische Verbreitung der Spongilliden scheint sich bis in
den Jura zu erstrecken. Wenigstens weisen einige Nadeln, die[S. 226]
man in jurassischen Süsswasserablagerungen gefunden, darauf hin
(Young). Auch hat Carter im Diluvium von Altmühl
in Bayern Nadeln beschrieben, welche fast ganz den Spikula von
Spongilla Mackayi gleichen.
8. Parasiten und Kommensalen der
Süsswasserschwämme.
Zur Zeit der Gemmulabildung stellen sich zahlreiche niederste Tiere
ein, um an dem im Zerfall begriffenen Weichteil unserer Schwämme
Mahlzeit zu halten. Lieberkühn hat eine Anzahl solcher Wesen
aufgeführt, welche er besonders in Spongilliden zur Winterzeit antraf.
Wir wiederholen ihre Namen nicht, da ihre Reihe nicht erschöpfend ist
und wir selbst durch Hinzufügen einiger anderer die Liste nicht beenden
würden.
Dagegen leben an und in einer frischen, in üppigem Wachstum
befindlichen Spongillide eine Anzahl Organismen, die wir zweckmässig in
zwei Gruppen sondern: Parasiten und Kommensalen.
Der Parasiten sind nur wenige. Dybowski schildert einen
Flohkrebs, den er Gammarus parasiticus nennt, und der auf der
Oberfläche des von uns schon so oft genannten Süsswasserschwammes
des Baikalsees lebt. Dieses Tier ist von grüner Farbe und wird, wenn
man es isoliert hält, gelblich. Es nährt sich also von dem grünen
Pigment (Zoochlorellen) der Spongillide. Auch eine andere Art, einen
Gammarus violaceus Dyb., ebenfalls vom Baikalsee, lernen wir
durch diesen Forscher als einen gelegentlichen Bewohner des Schwammes
kennen, er zeigt sich dann in grüner Färbung, var. virens. —
Ein anderer Parasit ist von Jackson in England beobachtet. Es
ist ein peritriches Infusor, Cyclochaeta spongillae tauft es
der Autor, welches auf der Oberfläche und den oberflächlichen grünen
Partien der Euphyd. fluviatilis lebt. — Ein diesem sehr
nahestehendes Infusor, vielleicht eine Trichodina, lebt nach
Alenitzin in Süsswasserschwämmen (Bütschli, Bronn,
„Klass. u. Ordn. d. Tierreichs“. Bd. I, p. 1808). — Auch bei uns nährt
sich ein Tier vom Süsswasserschwamm. Zur Winterzeit sammelt man[S. 227] öfters
im Tegelsee Exemplare des perennierenden Schwammes, welche grosse
Gänge einer Phryganidenlarve zeigen. Wir haben ein solches Exemplar
mit zwei dieser Larven in Fig. 46 abgebildet. Diese Tiere fressen sich
gewöhnlich vom Rande her in den Schwamm hinein und bohren sich mit dem
Kopfe so tief in das Gewebe, dass man die Gehäuse erst ablösen muss,
um der Tiere selbst sichtbar zu werden. Sie nähren sich, vielleicht
aus Not, von dem grünen Schwamme, denn sie zeigen einen grüngefärbten
Leib. Sie scheinen seit ihrer Erwähnung von Pallas, „Elenchus
Zoophytorum“, 1766, nicht wieder gefunden worden zu sein.
Als einen Übergang von Symbiose zum Parasitismus bezeichnen M. und
A. Weber[75] das Vorkommen einer Fadenalge Trentepohlia
(Chroolepus) spongophila, in Süsswasserschwämmen Sumatras.
Diese grünen Algen leben in farblosen Exemplaren von Ephyd.
fluviatilis und erzeugen besonders um die Oskula herum grüne
Flecke. Ausser dieser Alge erwähnen die genannten Autoren noch eine
Anzahl anderer, welche nur gelegentliche Gäste der Schwämme sind.
Fig. 46. Eph. fluviatilis (Lbkn.) von Phryganiden angefressen. Zwei ders. sieht
man unten links in ihrem tütenförm., gebogenen Gehäuse. (20. Novbr. 1887.
Tegelsee.) ½ nat. Grösse.
Als Kommensalen der Spongilliden sind uns nur folgende bekannt
geworden. Sehr häufig lebt in den Kloakenhöhlen oder in grösseren
Löchern der Schwämme, und wie es scheint besonders in Ephyd.
fluviatilis, die Larve einer Neuroptere: Sisyra (Branchiotoma)
spongillae Westw. Sie wurde in den Süsswasserschwämmen von
Hogg entdeckt und steckt mit dem Kopfe nach aussen gerichtet
im Schwamme. — Weit häufiger als dieses Tier lebt ein anderes
auf der äusseren Haut unserer Schwämme. Man kann einige hunderte
gezüngelter Würmchen, Nais proboscidea, von ein und demselben
Schwammexemplar erhalten. — Einige Male trafen wir auch eine
kleine Cladocere aus der Gruppe der Lynceiden auf der äussern Haut
des Schwammes an. Besonderes Interesse verdient ein unscheinbares,
nur[S. 228] 0.02 mm langes, aber in der Spongillide sehr häufig
anzutreffendes Tierchen. Es ist eine Acinetine: Podophrya fixa,
welche schon Lieberkühn erwähnt. Wir haben kaum einen Schwamm
untersucht, in dessen Kanälen dieses Tier nicht zugegen war.
Am lebenden Schwamm kann man es beobachten, wie es geduldig an ein
und derselben Stelle der Kanalwand festgeheftet sitzt, seine langen
Saugröhrchen nach Beute ausstreckend. Welcher Art ist diese Nahrung?
Nährt sich das Tier von den eingedrungenen, zerfallenen organischen
Bestandteilen oder muss es auf die günstige und, wie wir glauben,
seltene Gelegenheit lauern, ein eingedrungenes Infusor oder ein anderes
Glied aus der Reihe der Protozoen zu erhaschen? Oder ist es gar ein
Parasit des Schwammes? — Wir haben nur selten ein gestieltes Exemplar
dieser Podophrya zu Gesicht bekommen und müssen es unentschieden
lassen, ob wir in ihm die ungestielte Abart der Podophrya fixa
oder die Podophrya libera Perty zu erblicken haben. — Ganz
im Gegensatze hierzu steht das Vorkommen von Milbenlarven (Atax?
Noll), die wohl eher einen Schutz, als Nahrung, in den Kanälen
des Schwammes suchen. Sie scheinen nicht allzuhäufig zu sein, obwohl
sie auch bei Berlin mehrmals gefunden wurden.
9. Das Sammeln, Konservieren und
Untersuchen von Spongilliden.
Bei dem Sammeln von Süsswasserschwämmen kommt es auf den Zweck
an, den man dabei verfolgt. Wünscht man sich nur die Schwämme als
Schaustücke für Sammlungen zu beschaffen, so muss man natürlich ganze,
möglichst unverletzte Exemplare zu erhalten suchen und das ist im
allgemeinen viel schwieriger als das Sammeln von Material, welches
man zu anatomisch-histiologischen oder entwickelungsgeschichtlichen
Untersuchungen braucht, für welche unversehrte Schwammexemplare nicht
nur unnötig, sondern sogar ungeeignet sind.
Bei hohem Wasserstande oder bei tief wachsenden Schwämmen kann man sich
manchmal nur durch Tauchen helfen,[S. 229] um die Stücke in gutem Zustande
an die Oberfläche zu bringen. In anderen Fällen, wenn die Schwämme an
losen Gegenständen, Baumreisern, Brettern, Steinen etc., sitzen, genügt
ein bootshakenähnliches Instrument, an welchem unten ein Beutel
befestigt ist. Wir haben auch am Grunde der Spree mit dem Schleppnetz
manch schönes Stück erhalten, allein man ist dabei zu sehr dem Zufall
anheimgegeben. Die an grossen Balken, Pfeilern etc. angehefteten Stücke
wird man zweckmässig mit dem Kratzer ablösen, falls man sie nicht mit
dem Messer abheben kann. Die so erhaltenen Exemplare sind zwar nie
ganz vollständig erhalten, aber doch noch brauchbar. Übrigens wird
es bei öfteren Besuchen eines an Spongilliden reichen Wassers auch
einmal gelingen, ihnen mit der Hand bequem beizukommen. — Das Sammeln
der Schwämme geschieht zweckmässig zweimal im Jahre. Im Frühling oder
Herbst findet man die Gemmulae in den Schwämmen, letztere sind
in den Sommermonaten am üppigsten entwickelt. Wir möchten auch das
Augenmerk auf die perennierenden Spongilliden richten. Oft haben wir
uns in den Wintermonaten, auf der klaren, dünnen Eisdecke liegend,
an dem Anblick erfreut, den solch ein kleiner Wald abgestorbener
Schilfstengel, welche mit schön grün gefärbten Schwämmen besetzt sind,
am Grunde des Tegelsees bietet.
Die auf die eine oder andere Art gesammelten Exemplare kann man
trocken oder in Spiritus aufbewahren. Die zu trocknenden tötet man
zweckmässig sofort nach dem Entnehmen aus dem Wasser in starkem
Alkohol und trocknet sie erst nach einigen Stunden an der Luft. Man
erhält so ein gutes Präparat, an dem vorzüglich die äussere Haut als
glänzende Membran hervortritt (Möbius). Will man einen Schwamm
in Spiritus aufbewahren, so muss man den Alkohol, in welchem das Stück
abgetötet wurde, nach einigen Stunden mehrmals wechseln. Das Abtöten
geschieht in 96%-Alkohol, da der Schwamm äusserst wasserhaltig ist;
zum Konservieren genügt 75%-Spiritus. Um einen Schwamm, den man zu
konservieren wünscht, zu töten, ist es nötig, schnell zu verfahren.
Man wählt sich in dem von Spongilliden bewohnten Wasser ein gutes
Exemplar aus, löst es unter Wasser von seiner[S. 230] Unterlage oder bringt
es mit derselben bis an die Wasseroberfläche und zieht es nun schnell
aus dem Wasser, schwenkt es behutsam von dem in den Kanälen steckenden
Wasser aus und bringt es schnell in Alkohol. Hat man nur kleinere
Stücke und viel Alkohol zur Verwendung, so braucht man den Schwamm
nicht erst von seinem eingesogenen Wasser zu befreien. Jedenfalls ist
ein längeres Betrachten eines aus dem Wasser gezogenen Schwammes nicht
am Platze; wer den Schwamm im Leben kennen lernen will, muss ihn im
Aquarium beobachten, und wer Bau und Struktur zu erkennen wünscht,
muss sich gutes in Alkohol konserviertes Material beschaffen. Um
ein solches zu anatomischen und histiologischen Untersuchungen des
Weichteiles zu erhalten, kann man nicht die ganzen Schwämme verwenden.
Man braucht hierzu kleine bis 1 cm grosse Stückchen, die man
unter Wasser aus dem Schwamme ausschneidet und sofort in absol. Alkohol
oder in 96%igem, den man aber bald wechseln muss, abtötet (F. E.
Schulze). Noch besser werden die Zellen einer Spongillide fixiert,
indem man die Stückchen in Überosmiumsäure oder Sublimatlösung
bringt, auswäscht und in starkem Alkohol konserviert. Man muss dann
solche Stücke in Böhmerscher Hämatoxylinlösung oder in Boraxkarmin
färben, um die Einzelheiten zu erkennen. Für manche Teile ist eine
Doppelfärbung nötig, z. B. um den Kern der reifen Eier und ihrer
Furchungskugeln vom Dotter unterscheiden zu können. Fiedler
hat hierüber das Nähere angegeben. Derselbe Autor hat zum Abtöten
der Spongillidenstückchen verschiedene Mittel angewandt. Man wird
aus der Arbeit Fiedlers[89] ersehen, dass man zur Erkennung
verschiedener Strukturen verschiedene Wege einschlagen muss. Die so
vorbereiteten Stücke werden mittels des Mikrotoms in dickere und
dünnere Schnitte zerlegt.
Die nach dieser Methode gewonnenen Resultate muss man unbedingt am
lebenden Schwamm kontrollieren. Man wird sich dann überzeugen, was
natürlich ist und was künstlich hervorgebracht war. Um den Schwamm im
Leben anatomisch zu studieren, kann man sich eines jungen, aus einer
Gemmula oder Larve gezogenen bedienen. Oder man kann auch grössere,
bis ½ cm grosse Stücke[S. 231] verwenden, die man auf folgende Weise
erhält. Aus einem ganz frischen Schwamme schneidet man schnell mit
einem scharfen Messer senkrecht zur Oberfläche dünne Scheiben von ½–2
mm Dicke. Diese setzt man in ein kleines mit Wasserpflanzen
(Elodea) besetztes Aquarium auf Objektträger. Nach einigen Tagen haben
solche Stücke Haut und Oskulum neu entwickelt und sind meist an dem
Glase festgewachsen. Man nimmt nur diejenigen aus dem Aquarium, deren
Oskularrohr an der Seite liegt, bedeckt sie mit einem Deckglas und kann
lange Zeit an ihnen Beobachtungen machen. Nur muss man darauf achten,
ob aus dem Oskulum beständig der Wasserstrom austritt, um sicher zu
gehen, dass man einen lebenden Schwamm vor sich hat. Man wird sich
nun von der Formveränderlichkeit aller zelligen Elemente des
Schwammes überzeugen. „Every living part of the sponge that is soft
is subject to polymorphism.“ Carter.
Den Bau des Skelettes erkennt man, indem man von dem im Alkohol
konservierten Material mit der Hand dünne Schnitte macht und diese
in absoluten Alkohol, Terpentin und dann in Kanada-Balsam bringt.
Der zwischen dem Gerüste liegende Weichteil stört zwar, aber man ist
sicher, von dem Skelett nichts verloren zu haben.
Ausserdem muss man sich aber vom Weichteil ganz befreite
Gerüstpräparate in folgender Weise beschaffen. Man nimmt dazu einen
frischen Schwamm, aus dem man in verschiedenen Richtungen dünne
Scheiben schneidet. Diese maceriert man bei Ofenwärme in starkem
Ammoniak (zuerst von F. E. Schulze bei Hornschwämmen angewandt).
Die Lösung des Weichteils geschieht sehr schnell. Man erhält das
Skelett rein, indem man sehr behutsam mit Wasser und dann mit Alkohol
auswäscht. Um die Spongiolinsubstanz sichtbar zu machen, färbt man mit
Eosin oder Karmin. Bei der Maceration fallen immer eine Anzahl Nadeln
aus, man vergleicht deshalb diese Präparate mit denen, an welchen
Gerüst und Weichteil vorhanden sind. Noll hat zum Macerieren Eau
de Javelle angewandt, Girod[88] gebraucht auch Sodalösung. Das
erstere wirkt sehr energisch und zerstört oft mehr als man wünscht.
— Die Herstellung der Präparate von isolierten Nadeln geschieht[S. 232]
durch Kochen eines Stückchens der Spongillide in Salzsäure. Ist alles
zerfallen, so füllt man das Reagensglas mit Wasser voll, schüttelt um
und lässt absetzen. Das Auswaschen muss so lange fortgesetzt werden,
bis das Wasser säurefrei ist. Die Nadeln bettet man in Kanada-Balsam
ein; in Glycerin oder Glyceringelatine treten sie nicht hervor. Bei
dem Auswaschen verliert man viele Belagnadeln der Gemmulae. Die
Fleischnadeln von Eusp. lacustris gehen, wenn man schnell
verfährt, fast ganz verloren. Man muss deshalb auch Zupfpräparate
vom Schwamme untersuchen. Dagegen bietet Eusp. lacustris, und
zwar nur diese Art, die Möglichkeit, ein vollständiges Skelettgerüst
grösserer Teile des Schwammes darzustellen. Da hier die mächtig
entwickelte Spongiolinsubstanz in Kalilauge unlöslich ist, so kann
man grössere Zweige hierin kochen und erhält sehr zierliche weisse
Gerüstbäumchen. Verwendet man zum Kochen eine starke Kalilauge, so
bleibt von dem Skelett nichts als die Spongiolinsubstanz zurück,
weil die Lauge die Nadeln (Kieselsäureanhydrit) auflöst. In solchen
Präparaten treten dann die Nadeln als Lücken in der Kittmasse hervor
(Dybowski).
Die Struktur der Gemmulaschale lehren Dünnschnitte kennen, die man
nicht zu färben braucht. Um den Bau des Keimes zu erforschen, muss man
Dünnschnitte einer Doppelfärbung unterwerfen, sonst lassen sich Kerne
und Dottermaterial kaum von einander unterscheiden. Auch hier muss man
zur Kontrolle das lebende Objekt untersuchen.
Die Entwickelung des jungen Schwammes aus der Larve wird am besten
mit dem Horizontalmikroskop verfolgt. Daneben sind Dauerpräparate
anzufertigen.
Auf die Einzelheiten der hier angegebenen Manipulationen gehen wir
nicht weiter ein. Die wenigen Bemerkungen werden genügen mit dem
Hinweise, dass man das Übrige aus den Arbeiten Lieberkühns
und Carters, ferner bei Potts, Girod[88],
Fiedler und Maas[90] ersehen kann. Und endlich probiert
jeder, kontrolliert die von anderen angegebenen Methoden und findet
selbst die Mittel und Wege, mit und auf denen er zum Ziele kommt.
[S. 233]
Litteratur.
Ein vollständiges Verzeichnis der Arbeiten über Süsswasserschwämme
existiert nicht. Man findet Litteratur bis zum Jahre 1842 bei
G. Johnston, A history of British Sponges and Lithophytes.
Eine Liste der Arbeiten bis 1882 hat W. Dybowski, Studien
über die Süsswasserschwämme des russischen Reiches in Mém. Acad.
Imp. St. Pétersbourg, VIIᵉ série, T. XXX, No. 10 gegeben. Beide
Verzeichnisse sind zwar nicht erschöpfend, aber man kann mit ihnen
weiterarbeiten. Wem die Abhandlung von Dybowski nicht zur Hand ist,
kann sich die Litteratur aus den Verzeichnissen über Spongienwerke
von Bowerbank, A Monograph of the British Spongiadae, Vol. 4.
1882, dann besonders von G. C. J. Vosmaer, Spongien, Bronns
Klassen und Ordnungen des Tierreichs, II. Bd. 1882–87, und von R.
v. Lendenfeld, A Monograph of the Horny Sponges. London 1889,
beschaffen. In dem zuletzt genannten Werke findet man eine vorzügliche
Zusammenstellung aller wichtigen Arbeiten über Spongien. —
Die Litteratur über die grünen Körper der Spongilliden ersieht
man aus K. Brandt, Über die morphologische und physiologische
Bedeutung des Chlorophylls bei Tieren. Mittlg. Zool. Stat. Neapel, Bd.
4. 1883. — Wir fügen diesem Verzeichnis noch hinzu: J. E. Gray,
On the Situation and Rank of Sponges in the Scale of Nature. The Zool.
Journ. Vol. I. 1824.
In folgendem geben wir nur ein Verzeichnis der wichtigeren Werke,
welche nach Carters erster Arbeit (1848) erschienen sind. Die
Zahlen vor den Autornamen beziehen sich auf die Verweise in unserem
Texte.
[Bei einigen Literaturstellen handelt es sich um allgemeine
Nachschlagewerke, denen im Text keine direkten Verweise
gegenüberstehen. Diese werden im Folgenden mit Zahlen gefolgt von
runden Klammern dargestellt. (Anmerkung des Bearbeiters)]
1) J. S. Bowerbank, Further Report on the Vitality of the
Spongiadae. Rep. 27. Meeting Brit. Assoc. Advanc. Sc. 1857, p. 121–125.
London 1858.]
2) H. J. Carter, dessen zahlreiche Arbeiten in den Annals and
Magazine of Natural History, London. Vol. 1, p. 303–311. 1848. —
Vol. 4, p. 81–100. 1849. — Vol. 14, p. 334–335. 1854. — Vol. 17, p.
101–127. 1856. — Vol. 18, p. 39–45.[S. 234] — Vol. 20, p. 21–41. 1857. —
Vol. 3, p. 1–20. 1859. — Vol. 3, p. 331–343. 1859. — Vol. 14, p.
97–111. 1874. — Vol. 16, p. 1–40. 1875. — Vol. 10, p. 362–372. 1882.
— Vol. 12, p. 329–333. 1883. — Vol. 15, p. 18–20. 1885.]
3) W. James Clark, The American Spongilla, a craspedote,
flagellate Infusorian. Americ. Journ. Sc. Vol. 2, p. 426–436. 1871.]
4) W. Dybowski, Studien über die Spongien des russischen
Reiches mit besonderer Berücksichtigung der Spongienfauna des
Baikalsees. Mém. Acad. Impér. Sc. St. Pétersbourg, VIIᵉ série, T. 27,
No. 6, p. 1–71. 1880.]
5) W. Dybowski, Studien über die Süsswasserschwämme des
russischen Reiches. Das., T. 30, No. 10, 26 p. 1882.]
6) W. Ganin, Beiträge zur Kenntniss und Entwickelung der
Spongien. Warschau 1879. 88 p. (Russisch!)]
7) P. Girod, Les Éponges des Eaux douces d’Auvergne. Trav.
Labor. Zool. Girod. Clermont-Ferrand. T. 1. 11 p. 1888.]
8) Al. Goette, Untersuchungen zur Entwickelungsgeschichte von
Spongilla fluviatilis. Hamburg und Leipzig. 64 p. 1886.]
9) Fr. Hilgendorf, Zwei Arbeiten in Sitzungsberichte Ges.
Naturf. Fr. Berlin 1882, p. 26. — 1883, p. 87–90.]
10) G. H. Hinde, On some new Species of Uruguaya, Carter, with
Remarks on the Genus. Ann. Mag. Nat. Hist. Vol. 2, p. 1–12. 1888.]
11) H. Jackson, On a New Peritrichous Infusorian (Cyclochaeta
spongillae). Quart. Journ. Micr. Sc. Vol. 15, No. 5, p. 243–49. 1875.]
[S. 235]
12) N. Lieberkühn, dessen zahlreiche Arbeiten im Archiv für
Anatomie, Physiologie und wissenschaftliche Medizin von Joh. Müller.
1856, p. 1–19, p. 399–414, p. 496–514. — 1857, p. 376 bis 403. —
1859, p. 353–382, p. 515–529. — 1863, p. 717 bis 730. — 1865, p.
732–748. — 1867, p. 74–86.]
13) N. Lieberkühn, Ueber Protozoen. Zeitschr. wiss. Zool. Bd.
8, p. 307–310. 1857.]
14) N. Lieberkühn, Ueber Bewegungserscheinungen der Zellen.
Schrift. Ges. Beförd. Gesammt. Naturw. zu Marburg. Vol. 9, p. 335 bis
385. 1872. (Die Arbeit erschien schon 1870.)]
15) O. Maas, Zur Metamorphose der Spongillalarve. Zool. Anz.
Jahrg. 12, p. 483–487. 1889.]
16) Ed. v. Martens, Ueber einige Ostasiatische
Süsswasserthiere. Arch. f. Nat. Jahrg. 34, Bd. 1, p. 61–64. 1868.]
17) E. Metschnikoff, Spongiologische Studien. Zeitschr. wiss.
Zool. Bd. 32, p. 349–387. 1879.]
18) F. C. Noll, Beiträge zur Naturgeschichte der
Kieselschwämme. Abh. Senckenb. Nat. Ges. Bd. 15. 58 p. 1888.]
19) Fr. Petr, drei Arbeiten in Sitzungsber. kön. böhm. Ges.
Wiss. Prag. Math.-naturw. Klasse. 1885, p. 99–111. — 1886, p. 147–174.
— 1887, p. 203–214.]
[S. 236]
20) F. Vejdovský, drei Arbeiten in Sitzungsber. kön. Böhm. Ges.
Wiss. Nat. Kl. Prag. 1883, p. 328–340. — 1884, p. 167 bis 172. —
1886, p. 175–189.]
21) F. Vejdovský, Einiges über Spongilla glomerata. N. Zool.
Anz. Jahrg. 9, p. 713–15. 1886.]
22) F. Vejdovský, Diagnosis of the European Spongillidae.
Erschien in dem Werk von Potts, No. 30, p. 172–180.]
23) G. C. J. Vosmaer, Spongien. Bronns Kl. u. Ordn. d.
Tierreichs. II. Bd. 1882–87. 498 p.]
24) W. Weltner, Zwei Arbeiten in Sitzungsber. Ges. Naturf.
Freunde Berlin. 1886, p. 152–157. — 1888, p. 18–22.]
25) A. Wierzejski, Le développement des Gemmules des Éponges
d’eau douce d’Europe. Arch. Slav. Biol. T. 1, p. 23 bis 47. 1886.]
26) John T. Young, On the occurence of a Freshwater Sponge in
the Purbeck Limestone (Spongilla purbeckensis). The Geolog. Magaz. Vol.
5, p. 220–221. 1878.]
[72]W. Retzer, Die deutschen Süsswasserschwämme.
Inaug.-Diss. 30 p. Tübingen 1883.
[73]Edw. Potts, Contributions towards a Synopsis of
the American Forms of Freshwater Sponges, with descriptions of those
named by other authors and from all parts of the world. Proceed. Acad.
Nat. Sc. Philadelphia 1887, p. 158–279. (Hauptwerk über Systematik.
Enthält auch die zahlreichen früheren Arbeiten von demselben Autor;
viele interessante Beobachtungen über die Biologie.)
[74]M. Weber, Spongillidae des indischen Archipels.
Zool. Ergebnisse einer Reise in Niederl. Ost-Indien. 1. Heft, p. 30–47.
Leiden 1890.
[75]M. u. A. Weber, Quelques nouveaux cas de
Symbiose. Das. p. 48–72.
[76]E. Ray Lankester, On the Chlorophyll-corpuscles
and amyloid Deposits of Spongilla and Hydra. Quart. Journ. Micr. Sc.
Vol. 22, p. 229–254. 1882.
[77]C. Keller, Ueber Spermabildung bei Spongilla.
Zool. Anz. Jahrg. 1, p. 314–315. 1878.
[78]C. Keller, Ueber den Bau von Reniera semitubulosa
Ow. Zeitschr. wiss. Zool. Bd. 30, p. 563–586. 1878.
[79]A. Wierzejski, Bemerkungen über
Süsswasserschwämme. Zool. Anz. Jahrg. 10, p. 122–126. 1887.
[80]A. Wierzejski, Beitrag zur Kenntnis der
Süsswasserschwämme. Verh. Zool.-bot. Ges. Wien. Jahrg. 1888, Bd. 38, p.
529–536.
[81]R. v. Lendenfeld, Die Süsswasser-Cölenteraten
Australiens. Zool. Jahrb. Vol. 2, p. 87–108. 1887.
[82]W. Marshall, Über einige neue von Herrn
Pechuël-Loesche aus dem Congo gesammelte Kieselschwämme. Jenaische
Zeitschr. für Naturw. Vol. 16, p. 553–577. 1883.
[83]R. v. Lendenfeld, Experimentelle Untersuchungen
über die Physiologie der Spongien. Zeitschr. wiss. Zool. Bd. 58, 297 p.
1889 (Referat im Biol. Centralbl. Bd. 10. 1890).
[84]W. Marshall, Einige vorläufige Bemerkungen über
die Gemmulae der Süsswasserschwämme. Zool. Anz. Jahrg. 6, p. 630 bis
634, p. 648–52. 1883.
[85]W. Marshall, Vorläufige Bemerkungen über die
Fortpflanzungsverhältnisse von Spongilla lacustris. Sitzungsber.
Naturf. Ges. Leipzig. Jahrg. 1884, p. 22–29.
[86]W. Dybowski, dessen Arbeiten in den
Sitzungsberichten der Naturforschenden Gesellschaft zu Dorpat. Vol. 4,
1878, p. 527–534. Vol. 6, 1884, p. 507–515. Vol. 7, 1885–86, p. 44–45,
p. 64–75, p. 137–139, p. 295–298. Ferner im Zoolog. Anzeig. Jahrg. 7,
1884, p. 476–480.
[87]F. Vejdovský, Die Süsswasserschwämme Böhmens.
Abhandl. kön. Akad. Ges. Wiss. Math.-nat. Kl. Prag. Vol. 12. 44 p.
1883.
[88]P. Girod, Les Spongilles. Leur recherche, leur
préparation, leur détermination. Revue scientif. Bourbonnais et du
Centre de la France. T. 2, 11 p. 1889.
[89]K. Fiedler, Über Ei und Spermabildung bei
Spongilla fluviatilis. Zeitschr. wiss. Zool. Bd. 47, p. 85–128. 1888.
[90]O. Maas, Über die Entwicklung des
Süsswasserschwamms. Zeitschr. wiss. Zool. Bd. 50, p. 527–554. 1890.
[S. 237]
Die Strudelwürmer
(Turbellaria).
Von Dr. Otto Zacharias in Plön
(Holstein).
[S. 239]
Zusammen mit den im nächsten Kapitel zu schildernden Rädertieren
(Rotatoria) machen die Turbellarien einen ansehnlichen Bestandteil der
Süsswasserfauna aus, und es dürfte wohl kaum einen Bach, Fluss, Tümpel
oder See geben, worin nicht wenigstens eine oder die andere Art jener
kleinen, abgeplatteten Würmchen zu finden wäre, deren Körperoberfläche
mit einem dichten Wimperbesatze ausgestattet ist. Die einzelnen Cilien
dieser flimmernden Hautbedeckung wirken wie zahllose winzige Ruder
und ermöglichen es den Tierchen, gewandt und schnell durchs Wasser zu
gleiten. Dabei entsteht in unmittelbarer Nähe derselben ein beständiger
Strudel, der durch eingestreute Karminkörnchen für jedes Auge sichtbar
gemacht werden kann. Wegen dieses eigentümlichen Nebenumstandes, der
mit der Ortsbewegung jener Geschöpfe verknüpft ist, nennt man dieselben
„Strudelwürmer“ oder Turbellarien — eine Bezeichnung, die nicht ohne
Weiteres verständlich ist.
Um sich derartige Tiere zu verschaffen, braucht man bloss dem
nächstgelegenen Teiche Wasserpflanzen (besonders Fadenalgen und
Meerlinsen) zu entnehmen und dieselben — mit Wasser von derselben
Lokalität übergossen — in geräumigen Glasschalen ruhig stehen zu
lassen. Schon nach einigen Stunden wird man bei dieser Prozedur die
Wahrnehmung machen, dass zahlreiche Würmchen aus dem Pflanzengewirr
hervorkommen und nun an den Wänden der Glasgefässe langsam
umherkriechen. Von da können[S. 240] sie leicht mit einem Spatel oder mit
Hilfe eines Glasröhrchens weggenommen werden.
Im allgemeinen sind die stehenden Gewässer reicher an Turbellarien
als die fliessenden, und während in manchen Wasseransammlungen nur
einige wenige Arten gefunden werden, giebt es wieder andere, die eine
Fülle von verschiedenen Spezies darbieten. Letzteres ist z. B. der
Fall hinsichtlich des kleineren von den beiden bekannten Hochseen des
Riesengebirges; hier habe ich selbst das Vorkommen von nicht weniger
als 19 Turbellarienspezies festgestellt[91], darunter solche, die zu
den allerseltensten gehören. Und dabei ist jener See (im Volksmunde
„Kleiner Koppenteich“ genannt) nur etwa 255 Ar (= 10 Morgen) gross.
Alle Strudelwürmer des Süsswassers haben ihren Aufenthalt in der
Uferzone oder auf dem Grunde der Seen und Teiche. Im freien,
pflanzenleeren Wasser findet man sie niemals, mit einziger Ausnahme
von Castrada radiata, einer winzigen, glashell durchsichtigen
Form, welche eine bedeutende Schwimmfähigkeit besitzt. Diese Spezies
habe ich mehrfach mit dem feinen Netz aus der Mitte des Müritz-Sees (in
Mecklenburg) gefischt.
Was die Jahreszeit anlangt, welche für den Turbellarienfang am
geeignetsten ist, so hat man die Erfahrung gemacht, dass im Hochsommer
weniger Spezies gefunden werden, als im zeitigen Frühjahr und
besonders kurz nach der Schneeschmelze. Diese Wahrnehmung
bestätigt sich nicht bloss bezüglich der Gewässer des flachen Landes,
sondern auch an den Gebirgsseen. Den Grund für diese Erscheinung hat
man höchst wahrscheinlich mit in dem Umstande zu erblicken, dass
während der heissen Sommermonate der Sauerstoffgehalt der meisten
Wasseransammlungen stark verringert ist, oder vielleicht auch
darin, dass die zu jener Zeit lebhafter vor sich gehende Zersetzung
vegetabilischer Stoffe vielen Arten verderblich wird.
[S. 241]
Allgemeines.
Bei einem Blicke auf die Turbellarienfauna unserer Gewässer
unterscheidet man sogleich zwei Haupttypen, nämlich einesteils
grössere platte Würmer von 1–2 cm Länge, bei denen ein
baumförmig verästelter Darm durch die Haut hindurch wahrnehmbar ist,
und andernteils kleine (nur wie winzige Fadenfragmente aussehende)
Würmchen, deren Verdauungskanal eine einfach gestreckte (stabartige)
oder sackähnliche Gestalt besitzt. Man unterscheidet demgemäss
rhabdocöle und dendrocöle Strudelwürmer. Letztere werden
ihres abgeflachten Körpers halber auch „Planarien“ genannt. Fig. 47
und 48 veranschaulichen in schematischer Weise die den Darmtractus
(d) betreffenden Unterschiede. Damit sind aber bei beiden
Hauptabteilungen auch noch andere Abweichungen im Bau verbunden, die
wir sogleich etwas näher ins Auge fassen wollen.
Fig. 47.Fig. 48.
A. Rhabdocoela.
(Rhabdocoelida, L. v. Graff.)
Wenn man einen zu dieser Gruppe zählenden Strudelwurm seiner
augenfälligsten Beschaffenheit nach skizzieren soll, so kann das etwa
wie folgt geschehen. Ein solches Tier besitzt einen schlauchförmigen
Körper und einen in diesen eingeschlossenen Behälter (Darm) für
Aufnahme und Verdauung der Nahrung. Die Mundöffnung ist je nach den
einzelnen Gattungen bauchständig (Mesostoma) oder terminal (Vortex).
Alle Rhabdocöliden sind afterlos. Zwischen Darm und Leibesschlauch
liegen die Fortpflanzungsorgane, welche einen zwitterartigen Charakter
tragen. Die Begattung ist dieser Einrichtung gemäss stets eine
wechselseitige. In manchen Gattungen (Microstoma, Stenostoma) kommt[S. 242]
aber neben der geschlechtlichen auch eine ungeschlechtliche Vermehrung
vor und letztere bildet mehrfach sogar die Regel. Das zierliche
Meerlinsenkettchen (Catenula lemnae) z. B. pflanzt sich
ganz ausschliesslich auf diese Weise fort.
Zum besseren Verständnis des Geschlechtsapparates der
Rhabdocölen diene die untenstehende Abbildung (Fig. 49), in welcher
die hauptsächlich in Betracht kommenden Teile desselben schematisch
dargestellt sind.
Die zentral liegende Geschlechtsöffnung (pg) führt zunächst
in das Atrium genitale (a), einen erweiterungsfähigen
Hohlraum, der jederseits den Ausführungsgang des Keim- und
Dotterstockes (ks und ds) aufnimmt, während sich
an seinem oberen (resp. vorderen) Ende das männliche Zeugungsorgan
(p) inseriert, welches mit den beiden Hoden (h)
in Verbindung steht. Die reife Keimzelle gelangt nach ihrer Ablösung
vom Keimstocke in jenes Atrium und nimmt hier eine entsprechende
Portion Dottermasse in sich auf. Dann erfolgt die Befruchtung durch ein
anderes hermaphroditisches Individuum der nämlichen Spezies, welches
seinen Penis (p) durch den Porus genitalis bei pg
einführt. Die Umhüllung des nunmehr entwickelungsfähig gewordenen Eies
mit einer schützenden Chitinschale — zu welcher das Material von
besonderen Drüsen oder vom Zellenbelag der Atriumwand geliefert wird —
bildet den Schluss zu diesen Vorgängen.
Fig. 49.
Schema des Geschlechtsapparates.
Die geschilderten Verhältnisse finden wir indessen nur bei den
höchstorganisierten Rhabdocöliden-Familien (bei Mesostomiden,
Vorticiden u. s. w.), während z. B. die einfacher gebauten
Macrostomiden (Fig. 52 S. 256) der gesonderten Dotterstöcke ermangeln
und lediglich Ovarien besitzen. An letzteren zeigt aber
der untere Abschnitt die merkwürdige Eigenschaft, Dotterkörnchen
produzieren zu können, sodass die heranreifenden Keimzellen jenes[S. 243]
Sekret aus ihrer unmittelbaren Umgebung zu beziehen und sich auf Kosten
desselben zu vergrössern im stande sind.
Zwischen diesem primitiven Verhalten und dem Geschlechtsapparate mit
vollständig getrennten Keim- und Dotterstöcken vermittelt der bei
Prorhynchus stagnalis vorliegende Fall, wo sich die einheitliche
Eierstocksdrüse in zwei verschieden funktionierende Abschnitte
differenziert hat, von denen der eine bloss Keimzellen, der andere nur
Dotterflüssigkeit sezerniert. Durch räumliche Sonderung jener beiden
Drüsenhälften, also durch eine allmählich eingetretene Arbeitsteilung
zwischen denselben, haben wir uns die zuerst beschriebene und in Fig.
49 dargestellte Einrichtung hervorgegangen zu denken.
Bei der Mehrzahl der Rhabdocöliden besteht eine Duplizität der
weiblichen Geschlechtsdrüsen; doch giebt es auch einige Gattungen
(Vortex, Mesostoma, Gyrator), bei denen sie nur in einfacher Anzahl
erscheinen. Eine Reduktion der männlichen Drüsen ist seltener zu
konstatieren, doch kommt sie gleichfalls bei Gyrator und noch einigen
anderen Gattungen vor.
Zur genaueren Orientierung über den Bau der Turbellarien ist es
erforderlich, dass wir nach Betrachtung der Generationsorgane nun auch
den ungegliederten, weichen Körper und den Verdauungsapparat derselben
einer näheren Analyse unterziehen. Ersterer besteht, wenn wir bei
unserer Untersuchung von innen nach aussen gehen, aus einem mächtig
entwickelten Hautmuskelschlauche, dessen Zusammensetzung aus
Längs- und Querfasern, welche parallel und unmittelbar an einander
liegen, schon den älteren Forschern bekannt war. Jene Fasern besitzen
bei 0.0005 bis 0.002 mm Breite eine oft sehr bedeutende Länge;
wenigstens lassen sich an Zupfpräparaten nicht selten Fragmente von
0.5 bis 0.9 mm isolieren. Ihrer sonstigen Beschaffenheit nach
sind sie homogen, stark lichtbrechend, glatt und kernlos. Je nach
den einzelnen Arten und Gattungen weist der Hautmuskelschlauch eine
verschiedene Anordnung seiner Elemente auf. Bei manchen Spezies
liegen die Längsfasern zu innerst und werden von einer äusseren
Ringfaserschicht umgeben. Dies ist z. B. der Fall bei Mesostoma
Ehrenbergii.[S. 244] Bei einer nahe verwandten Art (Mesostoma
lingua) befindet sich zwischen beiden Schichten noch eine solche
aus Diagonalfasern, und bei Microstoma lineare ist die
Zusammensetzung des Muskelnetzes gerade umgekehrt, nämlich so, dass die
Längsfasern die äussere, die Ringfasern die innere Schicht bilden. Auf
diesen (seiner Gestalt nach sehr veränderungsfähigen) Muskelschlauch
folgt die sogenannte Basalmembran, ein gallertartiges Häutchen
von feinkörniger Struktur, welches dem Turbellarienkörper Festigkeit
verleiht und zugleich dem darüber liegenden Epithel zur
Unterlage dient.
Letzteres besteht aus einer einfachen Zellenlage, deren einzelne
Elemente durch eine Kittsubstanz mit einander verbunden sind. Zwischen
ganz flachen Epithelzellen und hohen cylindrischen existieren bei den
verschiedenen Spezies alle möglichen Übergänge. Jede solche Zelle
besitzt, je nach ihrer Gestalt, einen scheiben- oder spindelförmigen
Kern und mehrere feine Protoplasmafortsätze (Cilien), die in ihrer
Gesamtheit das charakteristische Wimperkleid darstellen, womit die
Körperoberfläche aller Strudelwürmer bedeckt ist. Bei manchen Spezies
lässt sich über dem Epithel noch eine äusserst feine Kutikula
nachweisen, welche mit zahllosen winzigen Öffnungen für den
Durchtritt der Cilien versehen ist. Mit Hilfe einer einprozentigen
Höllensteinlösung habe ich dieses äusserst zarte Gebilde an frischen
Exemplaren von Macrostoma viride, Microstoma lineare
und Stenostoma leucops deutlich sichtbar gemacht. Man lässt
zu diesem Zwecke einen einzigen Tropfen jener Flüssigkeit unter das
Deckglas laufen und kann dann unterm Mikroskop wahrnehmen, wie sich
jenes glashelle, doppelt contourierte Häutchen auf einzelnen Strecken
oder auch im ganzen Umkreise des Wurmes binnen wenigen Minuten abhebt.
Bei der zuletzt genannten Art geschieht die Loslösung so schnell, dass
zur Herstellung des betreffenden Präparates oft nur einige Sekunden
erforderlich sind.
Ausser den Cilien kommen bei gewissen Gattungen von Rhabdocöliden auch
noch längere, borstenähnliche Epithelfortsätze vor, die vielfach
zitternde oder schwingende Bewegungen ausführen. Bei Macrostoma
viride (Fig. 52) sind diese „Geisselhaare“[S. 245] über den ganzen Körper
verbreitet; die längsten stehen aber am vorderen und hinteren Ende des
Tierchens. Das Gleiche sehen wir bei Macrostoma hystrix und
anderen Vertretern desselben Genus. Auch bei der von E. Sekera
neuerdings beschriebenen Bothrioplana alacris[92] finden wir (am
vorderen Saume des Kopfes) starre Borsten, die — wie es den Anschein
hat — zur Vermittelung von Tastempfindungen dienen.
Als eigentümliche Einlagerungen treten in der Haut bei fast allen
Strudelwürmern des Süsswassers ausserordentlich winzige Stäbchen
(Rhabditen) auf, welche — zu kleinen Paketen oder Bündeln
vereinigt — sich über die gesamte Leibesoberfläche verteilen. Ihre
Entstehung nehmen diese bald nadel-, bald keulenförmigen Gebilde in
besonderen Zellen, die dem zwischen Darm und Hautmuskelschlauch sich
ausspannenden Bindegewebe, welches als Parenchym bezeichnet
wird, angehören. Von hier aus rücken die Stäbchen auf eine noch nicht
hinlänglich festgestellte Weise bis zum Epithel vor und dringen in
die Zellen desselben ein, wo sie dauernd verbleiben. Bei manchen
Rhabdocöliden ist auch das Parenchym selbst mit zahlreichen Rhabditen
durchsetzt, und sie bilden dann im Innern desselben ganze Reihen
oder Strassen, die gewöhnlich im Vorderende der betreffenden Tiere
zusammenlaufen. Das sieht man deutlich bei mehreren Mesostomiden, z. B.
bei M. rostratum und M. viridatum.
Eine mikroskopische Analyse des Parenchyms ergiebt, dass
dasselbe aus Muskelfasern, Bindegewebssträngen und mehrfach
verästelten Zellen besteht, die gleichfalls bindegewebiger Natur
sind. Erst durch die Methode der Schnittserien war es möglich,
die einzelnen Bestandteile des Parenchyms festzustellen und ein
Lückensystem innerhalb desselben nachzuweisen, welches von einer
farblosen, blutartigen Flüssigkeit erfüllt ist. Diese besitzt aber
keine selbständige Zirkulation, sondern wird lediglich durch die
Kontraktionen des Hautmuskelschlauchs gelegentlich in Bewegung versetzt.
Sehr häufig sind die Fasern des Parenchymgewebes mit
Pigmentkörnchen durchsetzt, und es kommt dann das zu stande, was
v. Graff[93]
sehr passend „reticuläre Pigmentierung“ genannt[S. 246]
hat. Hiervon rührt die dunklere Färbung der Rückenfläche bei manchen
Rhabdocölen her, die niemals in den epithelialen Zellen ihren Sitz hat
— obwohl dies bei flüchtiger Ansicht so scheinen könnte.
Im Parenchym ist auch das Gehirnganglion der Turbellarien
gelegen oder, richtiger gesagt, es ist in dieses Gewebe meistenteils
vollständig eingebettet. Seinem feineren Baue nach besteht dasselbe
aus einer Anhäufung von feinkörniger oder zartfaseriger Substanz und
einer Rinde von Ganglienzellen mit runden oder auch ovalen Kernen. Zwei
Längsnervenstämme, die vom Gehirn abgehen und zu beiden Körperseiten
verlaufen, sind bei zahlreichen Rhabdocölen nachgewiesen; bei einigen
Spezies hat man auch mehrfache Kommissuren, welche die Hauptstämme mit
einander verbinden, vorgefunden.
Hinsichtlich des Parenchyms ist noch zu bemerken, dass es bei den
verschiedenen Gattungen in mehr oder minder starker Entwickelung
angetroffen wird. Wir kennen Formen, deren Leibeshöhlung so gut wie
frei von parenchymatösen Muskeln und Bindegewebsbalken ist, während
andere wieder das gerade Gegenteil solchen Verhaltens darbieten.
Was nun schliesslich den Verdauungsapparat der Rhabdocölen
anlangt, so besteht derselbe aus Schlund (Pharynx) und Darm. Letzterer
ist nach dem Parenchym zu entweder durch eine bindegewebige Scheide
abgegrenzt, oder er besitzt eine Muskelausstattung, an welcher wir
— wie beim Hautschlauche — Längs- und Ringfasern unterscheiden
können. Das Darmlumen ist mit einem kontinuierlichen Epithelbelag
ausgestattet, dessen Zellen membranlos sind, und die an ihrer Basis
rundliche, resp. ovale Kerne besitzen. Gewöhnlich ist dieses Epithel
an der unteren (ventralen) Seite des Darmes etwas höher als an der
oberen. Die einzelnen Zellen desselben erscheinen häufig an ihrem
freien Ende keulig verdickt und mit zahlreichen Fetttröpfchen erfüllt.
Was die Verdauung und Assimilation der in den Darm aufgenommenen
Nahrungsobjekte anlangt, so scheinen die grösseren Epithelzellen
kleine organische Partikelchen direkt in sich aufnehmen zu können,[S. 247]
indem sie dieselben nach Art der Rhizopoden (vergl. Kapitel 4) mit
ihrem Protoplasmaleibe umfliessen. Es fände demnach in diesem Falle
eine sogenannte „intracelluläre Verdauung“ statt. Diese Ansicht wird
durch darauf bezügliche Experimente von Isao Ijima (jetzt
Professor an der Universität Tokio) zu fast vollkommener Gewissheit
erhoben. Dr. Ijima fütterte Planarien mit dem Dotter von Hühnereiern
und fand bald darauf die gefressenen Dotterkugeln überall in den
Zellen der Darmverzweigungen seiner Versuchs-Dendrocölen wieder. Auch
entdeckte derselbe Forscher, dass der Darmkanal bei Dendrocoelum
lacteum, welcher so deutlich durch die Haut sichtbar ist, seine
schwärzliche Beschaffenheit von kleinen Schlammteilchen erhält, welche
eingeschlürft und in die Epithelzellen mit aufgenommen worden sind[94].
Was in diesem Bezug für die Planarien experimentell erwiesen ist, darf
ohne Zweifel auch für die Rhabdocöliden als gültig angesehen werden,
zumal Krukenberg auch an mehreren Zoophyten die Aufnahme von
Fremdkörpern durch die Entodermzellen konstatiert hat.
Zum Ergreifen oder Einschlürfen der Nahrung besitzen alle Turbellarien
ein dickmuskulöses, kompliziert gebautes Organ, den Schlund
(Pharynx), welcher entweder die Gestalt eines starkwandigen Rohres oder
diejenige eines zwiebelartigen Bulbus hat. Die übergrosse Mehrzahl
der Rhabdocölen ist mit einem Pharynx bulbosus ausgestattet,
der bei den Mesostomiden eine plattgedrückte kugelige Form zeigt
und sich, von oben her gesehen, wie eine Rosette ausnimmt. Das ist
der sogenannte Pharynx rosulatus. Seine Achse steht stets
senkrecht auf der Längsachse des Körpers. Eine Modifikation desselben
ist der allen Vorticiden zukommende tonnenförmige Schlund (Pharynx
doliiformis). In seinem feinen Bau ist er dem vorigen sehr
ähnlich, aber doch auch leicht wieder von ihm zu unterscheiden, weil
er im ganzen eine stärker entwickelte Muskulatur besitzt. Ausserdem
ist seine Achse der Körperachse immer parallel, oder doch nur wenig
gegen dieselbe geneigt. Mit seiner Spitze ist das Schlundtönnchen in
den weitaus meisten Fällen dem Vorderende des Wurmleibes zugekehrt.
Hierzu kommt noch der Pharynx variabilis, dessen Merkmal eine[S. 248]
grosse Fähigkeit zu Gestaltveränderungen ist, insofern er mannigfache
Windungen und Krümmungen auszuführen vermag und ausserdem weit
hervorgestülpt werden kann. Einen derartigen Schlund finden wir bei dem
merkwürdigen Plagiostoma Lemani, einer Rhabdocöle von marinem
Habitus, die aber im Süsswasser lebt und von Prof. F. A. Forel
zuerst im Genfersee entdeckt wurde. In der Familie der Monotiden,
welche gleichfalls nur einen einzigen Vertreter im salzfreien Wasser
hat[IV], begegnet uns eine Schlundform, die als Pharynx plicatus
bezeichnet wird. Der hauptsächlichste Unterschied zwischen dieser und
den anderen Pharyngealformen besteht darin, dass bei ihr der zwischen
innerer und äusserer Muskelschicht gelegene Raum in offener Verbindung
mit der Leibeshöhle steht, und nicht — wie beim Ph. bulbosus
— gegen letztere abgeschlossen ist. Der Monotidenschlund stellt
demnach eine blosse Ringfalte der Körperhaut dar, die indessen eine
grosse Beweglichkeit zeigt und sich in überraschender Weise rüsselartig
verlängern kann.
[IV] Es ist dies der von mir 1884 in den Koppenteichen des
Riesengebirges aufgefundene Monotus lacustris. Z.
Zuletzt müssen wir auch noch des Wassergefässsystems gedenken,
welches in Gestalt eines zarten Röhrennetzes mit zwei stärkeren
Hauptstämmen den Turbellarienkörper vom vordern bis zum hintern Ende
durchzieht. Die einzelnen Verästelungen desselben zu verfolgen ist mit
mannigfaltigen Schwierigkeiten verknüpft, und daraus erklärt es sich,
dass wir bei manchen Spezies noch sehr wenig über den Gesamtverlauf
dieser Röhrenleitung wissen. Ihrer physiologischen Bedeutung nach
stellt sie höchst wahrscheinlich ein Ausscheidungsorgan dar, welches
verbrauchte Stoffe aufnimmt und ansammelt, um dieselben durch die
Öffnungen, mit denen die Hauptstämme an der Körperoberfläche ausmünden,
fortzuschaffen. Als Ausnahme finden wir bei sämtlichen Arten der
Gattung Stenostoma anstatt zweier nur einen einzigen Hauptstamm,
welcher in der Nähe des hinteren Körperendes sich öffnet.
[S. 249]
B. Dendrocoela.
(Dendrocoelida, L. v. Graff.)
Die vorstehend gegebene Orientierung über die Grundzüge der
Rhabdocöliden-Organisation hat im wesentlichen auch für die grösseren
Turbellarien mit dreizipfeligem und baumförmig verzweigtem Darm
Gültigkeit. Dieselben besitzen gleichfalls einen Hautmuskelschlauch
und eine Basalmembran mit einer darüber befindlichen Lage von
flimmernden Epithelzellen. Letztere sind indessen nicht platt, sondern
hoch cylindrisch gestaltet; in ihrem Innern enthalten sie aber genau
solche Stäbchen, wie wir sie bei den Rhabdocölen antreffen. Ebenso ist
bei unseren Süsswasserdendrocölen (Tricladen) der Raum zwischen der
Hautmuskulatur und dem Darmkanal mit seinen Ausbuchtungen von einem
lockeren Bindegewebe und von zahlreichen (dorsoventral und quer sich
ausspannenden) Parenchymmuskelfasern erfüllt. Dazu kommt noch das
Vorhandensein eines reichmaschigen Wassergefässsystems, der Besitz
eines Gehirnganglions mit davon ausstrahlenden Seitennerven und ein
hermaphroditischer Geschlechtsapparat, der aus Keim- und Dotterstock,
blasigen Hoden, sowie einem zapfenförmigen Begattungsorgan besteht.
Eine ausgebildete Leibeshöhle, wie sie bei vielen Rhabdocöliden
gefunden wird, existiert bei den Tricladen nicht, sodass man es sich
erklären kann, wenn die älteren Zoologen zur Bezeichnung eines solchen
Thatbestandes auf den Ausdruck „parenchymatöse Würmer“ verfielen.
Das Darmepithel hat bei denselben genau die nämliche Beschaffenheit
wie bei den stabdarmigen Turbellarien. Die Zellen desselben sind
langgestreckt, nackt und häufig birnförmig verdickt. Jede besitzt einen
rundlichen Kern, der am basalen Ende liegt. Den gleichen Verhältnissen
begegneten wir bei den Rhabdocöliden. Hinsichtlich des Schlundes
stimmen die Planarien fast ganz genau mit den Monotiden überein, die,
wie bereits erwähnt wurde, einen Pharynx plicatus besitzen. Über
den feineren Bau desselben findet man ausführliche Angaben in L. v.
Graffs Rhabdocöliden-Monographie (S. 87 und 88).
[S. 250]
Wenn die Planarien fressen, so stülpen sie ihren Pharynx oft
bis zu einer erstaunlichen Länge aus dem Munde hervor. Er führt
dabei wurmförmige Bewegungen aus, als wenn er die geeignetsten
Nahrungsobjekte aussuchen wollte. Dabei erweitert sich sein freies Ende
gewöhnlich trompetenartig, sodass auch grössere Beutetiere (Crustaceen,
Insektenlarven z. B.) ergriffen und verschluckt werden können. Trennt
man den hervorgestreckten Tricladenrüssel durch einen Scherenschnitt
an seiner Wurzel ab, so fährt derselbe — wenn er feucht gehalten wird
— oft noch mehrere Stunden lang mit seinen Gestaltveränderungen fort.
Diese Lebenszähigkeit erklärt sich hinlänglich durch die reichliche
Innervation des in Rede stehenden Organs, über welche wir durch
Ijimas Untersuchungen Aufschluss erhalten haben. Etwa zwischen
der äusseren Ringfaserschicht des Pharynx und den Ausführungsgängen
der Speicheldrüsen sind die Nerven zu einem Plexus verbunden, der
gegen das freie Ende hin eine beträchtliche Anschwellung aufweist. An
dem nicht minder zählebigen Rüssel des Süsswasser-Monotus (Monotus
lacustris) ist von M. Braun und mir die Anwesenheit eines
dicken Ringes von Nervenfasern festgestellt worden, der auf Quer- und
Längsschnitten in der Mitte des Pharynx zum Vorschein kommt und einen
deutlichen Zusammenhang mit den ventralen Längsnerven erkennen lässt.
Sinnesnerven, die vom Gehirn aus zu den mehr oder minder
lappenartigen Seitenteilen des Kopfes laufen, sind, wie bei den
Rhabdocölen, so auch bei den Tricladen nachgewiesen. Ebenso besitzt
die Mehrzahl unserer Planarien Sehorgane (Augen), die entweder
zu zweien auf der dorsalen Fläche des Kopfes stehen, oder in grösserer
Anzahl (40–60) den Rand des ganzen vorderen Körperteiles umsäumen
(Polycelis).
Die Planarien produzieren nach stattgehabter (wechselseitiger?)
Befruchtung Eier, die zu 30–40 Stück in ein kugeliges oder elliptisches
Cocon eingeschlossen sind. Letzteres wird mittels eines weisslichen
Sekretes an Wasserpflanzen befestigt. Und zwar geschieht dies schon
sehr früh im Jahre, etwa im Februar oder März. Der Inhalt des Cocons
besteht aus einer Flüssigkeit, in der sich eine[S. 251] grosse Menge von
Dotterzellen (über 10000 sind gezählt worden) befinden, dazwischen ist
aber nur die oben angeführte Anzahl von Eiern sichtbar. Diese letzteren
sind nackte Zellen von geringerer Grösse als die Dotterelemente;
sie haben nur 0.035 bis 0.044 mm im Durchmesser. — Über die
Embryonalentwickelung der Süsswasserdendrocölen besitzen wir ausser Dr.
J. Ijimas Beobachtungen an Dendrocoelum lacteum noch eine
neuere Arbeit des französischen Zoologen Paul Hallez, die sehr
ausführliche Angaben und zahlreiche erläuternde Tafeln enthält[95].
Neben der geschlechtlichen Fortpflanzung kommt bei einigen Planarien
auch eine ungeschlechtliche durch Querteilung vor. Ich habe
diese vielfach in Zweifel gezogene Thatsache 1885 an Planaria
subtentaculata Dugès (aus einem Bache bei Hirschberg i. Schl.) mit
Sicherheit festgestellt und seinerzeit detailliert beschrieben[96]. Der
Hauptsache nach verläuft jener Vorgang folgendermassen. Zuerst zeigt
sich am Beginn des hinteren Leibesdrittels (und zwar immer dicht hinter
dem Eingang zur Rüsseltasche) eine seichte Einschnürung, welche von Tag
zu Tag Fortschritte macht. Während dieser Zeit liegen die Tiere oft
stundenlang ganz still. Nach drei bis vier Tagen bereits kann man mit
der Lupe die ziemlich perfekt gewordene Querteilung konstatieren. Die
Abtrennung des Tochtersprosses von der Mutter erfolgt nunmehr unter
ganz eigentümlichen Umständen. Merkwürdigerweise nämlich löst sich
derselbe erst in seiner mittleren Partie von letzterer ab, während
er zu beiden Seiten damit noch in Verbindung bleibt. Hat sich das
Tochterteilstück definitiv abgetrennt, so bemerkt man am Vorderende
desselben ein kleines, pigmentfreies Zäpfchen: den sich neu bildenden
Kopf. Nach Verlauf von 24 Stunden unterscheidet man schon Augenpunkte
an demselben. Demnächst bildet sich auch eine neue Rüsselhöhle und ein
neuer Pharynx aus, sodass das durch Teilung des Mutter-Individuums
entstandene Tier keinerlei Organisationsmängel zeigt. Diese
Teilungserscheinungen waren aber nur während der warmen Sommermonate
zu beobachten und bemerkenswert ist dabei, dass an den sich so
fortpflanzenden Exemplaren keine Spur von Geschlechtswerkzeugen zu
entdecken[S. 252] war. Der nämliche Vorgang ist unlängst auch an einer anderen
Dendrocöle (Planaria albissima Vejd.) von Dr. E. Sekera
beobachtet worden, sodass die älteren Angaben von Dalyell und
Dugès, welche früher schon über Querteilung bei Planarien
berichtet haben, nunmehr bestätigt sind.
Zu den am meisten in unseren Gewässern verbreiteten Planarien
gehören ausser dem milchweissen Dendrocoelum lacteum Oerst.,
Polycelis nigra O. Fr. Müller, Planaria polychroa
O. Schm. und Pl. lugubris. Eine der grössten einheimischen
Planarien ist das von Dr. W. Weltner im Tegelsee bei Berlin und
später auch in der Spree aufgefundene Dendrocoelum punctatum
Pallas, welches im ausgestreckten Zustande eine Länge von 3–4 cm
erreicht[97]. Alle diese Tiere sind in Grösse, Gestalt und Färbung
ziemlich variabel, und deshalb genügen solche äussere Merkmale bei der
Artbestimmung nicht. Hierzu müssen vielmehr die sehr formbeständigen
Geschlechtsorgane verwendet werden, deren Analyse freilich in manchen
Fällen ebenso zeitraubend wie schwierig ist. Nur auf diesem Wege, der
zuerst von Oscar Schmidt betreten wurde, gelangt man zu einer
sicheren Identifizierung der Spezies.
Bei einer allgemeinen Orientierung, wie sie hier bezweckt
wird, darf schliesslich auch der Hinweis auf das Vorkommen von
Landplanarien nicht fehlen. Und zwar kennen wir derartige Würmer
nicht bloss aus tropischen Ländern, sondern auch als Mitglieder der
einheimischen Fauna. Wir finden dieselben bei einiger Achtsamkeit
unter Holzscheiten, die im Walde lagern, zwischen feuchtem Moos und
an der Unterseite von Hutpilzen. Bis vor kurzem war nur eine einzige
einheimische Art bekannt, nämlich Rhynchodesmus terrestris O.
Fr. Müller. Die grössten Exemplare sind 2–2.5 cm lang und von
schwarzer Färbung; die Rückenseite ist stark gewölbt, die Bauchfläche
hingegen zu einer Kriechsohle ausgebildet. In ihren Bewegungen sind die
Tiere sehr träge, und wie es scheint, lieben sie kühle und schattige
Aufenthaltsorte[98]. Dr. J. v. Kennel fand Exemplare von
Rhynchodesmus in der Umgebung von Würzburg (unter Steinen), Dr. H.
Simroth mehrere in den Wäldern bei Leipzig, und ich welche in der
Nähe von Hirschberg in Schlesien.
[S. 253]
Unterirdisch lebend, d. h. im feuchten Erdreich sich aufhaltend, ist
1887 von Fr. Vejdovsky eine zweite einheimische Landplanarie
entdeckt worden, welche einer ganz neuen Gattung angehört. Sie ist
nur 4–6 mm gross und etwa 0.5 mm breit. Der genannte
Prager Forscher fand sie in einem Erdhaufen bei Bechlin in Böhmen,
den er eines anderen wissenschaftlichen Zweckes wegen durchsuchte.
Das betreffende Tierchen (Microplana humicola) ist vollkommen
durchsichtig, ermangelt der Kopflappen und besitzt nur auf der
Bauchseite eine Cilienbekleidung. Ihrem ungefähren Aussehen nach ist
sie in Fig. 50 wiedergegeben. Das Nähere darüber muss man aus der
darauf bezüglichen Abhandlung Prof. Vejdovskys ersehen[99].
Um endlich noch der Verwandtschaft von Dendrocölen und
Rhabdocölen ein Wort zu widmen, so verdient Erwähnung: dass
wir in dem von Prof. M. Braun (1880) begründeten Genus
Bothrioplana ein interessantes Verbindungsglied zwischen jenen
beiden Unterabteilungen kennen gelernt haben. Es handelt sich dabei
um Tiere, die in der Form des Darmes und im Bau ihres Schlundes eine
fast vollständige Übereinstimmung mit den Dendrocölen bekunden, während
sie durch mehrere andere Merkmale (Mangel der Stäbchenstrassen,
Anordnung der Rhabditen zu Paketen, Besitz von Wimpergrübchen an den
Kopfseiten) an die Rhabdocölen erinnern[100]. Braun fand von
diesen Turbellarien zwei Spezies in Dorpater Brunnenschächten, und
ich habe später (1886) deren noch zwei aus dem Kleinen Koppenteiche
des Riesengebirges gefischt[101]. Ein Habitusbild der einen Art,
die ich B. silesiaca genannt habe, ist in Fig. 51 S. 254
gegeben. Es ist ein augenloses Tierchen von 2.5 mm Länge, an
welchem sofort das verbreiterte Kopfende auffällt. Mit diesem wühlt
es unruhig suchend beständig in dem feinen Mud umher, worin es sich
aufhält. Überhaupt sind diese Bothrioplanen durch äusserst hastige
Bewegungen charakterisiert, die im schroffen Gegensatz zu dem ruhigen
Dahingleiten der gewöhnlichen Planarien stehen. Die[S. 254] Hautfarbe der
kleinen Würmchen ist weiss oder hellgrau. Der Darm schimmert von innen
her bräunlich durch. Die gesamte Körperoberfläche ist mit Cilien
bedeckt und überall sieht man im Epithel Stäbchenpakete, welche aus
je drei Rhabditen bestehen. Gelegentlich treten am Hinterende gewisse
Haftorgane (Klebzellen) hervor, wie sie bei vielen Rhabdocölen
beobachtet werden. Der hintere Körpersaum ist jederseits mit einzelnen
steifen Borsten besetzt, während das vordere Ende frei davon ist und
nur die gewöhnlichen kurzen Cilien trägt. Mit etwas längeren Wimpern
scheinen die beiden Wimpergruben (wg) zu beiden Seiten des
Kopfes ausgestattet zu sein. Der Verdauungskanal hat, wie unsere Figur
zeigt, eine gestreckt-ringförmige Gestalt, und hiervon lässt sich
die dreigabelige Beschaffenheit des Dendrocölen-Darmes ungezwungen
ableiten, wenn wir annehmen, dass die stärkere Entwickelung des
Schlundes bei den Tricladen den Anstoss zu einer Kontinuitätstrennung
in der unteren Ringhälfte gab. Dadurch entstanden naturgemäss die
beiden hinteren Darmschenkel der Planarien, und es wurde Raum zur
Unterbringung des mächtigen Pharynx dieser Würmer geschaffen.
Entschieden unterstützt wird diese zuerst von M. Braun
geäusserte Ansicht durch die Lage des Schlundes unmittelbar hinter
der Gabelungsstelle, und auch dadurch, dass derselbe von hier aus ein
ansehnliches Stück zwischen die beiden in Frage kommenden Darmäste
hineinragt.
Das Gehirn (c) liegt bei Bothrioplana silesiaca in der
halsartigen Einschnürung, mit welcher sich der breitere Kopfteil vom
übrigen Körper absetzt. Die zweite von mir im Kleinen Koppenteich
aufgefundene Spezies (B. Brauni) des nämlichen Genus ähnelt in
der Form ihres Gehirnganglions und im Verlauf der Exkretionsgefässe
den Rhabdocölen noch mehr als die erstbeschriebene Art. Ihr fehlen
auch die Wimpergrübchen, und die Stäbchenpakete enthalten bei ihr eine
grössere Anzahl (4–5)[S. 255] Rhabditen. Jedes einzelne dieser Pakete macht
den Eindruck, als habe es eine leichte schraubenförmige Drehung um
seine Längsachse erfahren. Der Kopfteil ist bei dieser und auch bei der
vorigen Spezies fast vollkommen rhabditenfrei.
Durch diese Tiere wird, wie schon gesagt, die sonst zwischen
Rhabdocölen und Dendrocölen bestehende Kluft überbrückt, und deshalb
haben wir es in denselben mit recht interessanten Übergangsformen
zu thun, die des näheren Studiums wohl wert sind. Hinsichtlich der
Geschlechtswerkzeuge ist bis jetzt festgestellt, dass die Keim- und
Dotterstöcke paarig sind, und dass auf jeder Seite eine Doppelreihe
von Hodenbläschen im mittlern Teile des Körpers vorhanden ist. Diese
und die vorstehend berichteten Befunde rechtfertigen es, dass wir die
Bothrioplanen als niederstes Genus den Tricladen anschliessen, wiewohl
anderseits nicht zu verkennen ist, dass sie in mehrfacher Hinsicht mit
den Rhabdocölen übereinstimmen.
Kurze Beschreibung einiger
Rhabdocöliden-Spezies.
Für den Zweck dieses Kapitels, welcher darin besteht, den Leser in die
Turbellarienfauna des Süsswassers einzuführen, empfiehlt es sich nun,
auf die vorhergegangene allgemeine Orientierung eine Schilderung der
am häufigsten vorkommenden Gattungen folgen zu lassen. Dies kann am
besten durch die Vorführung einzelner Spezies geschehen, und hierbei
werden wir Gelegenheit haben, noch einige Punkte nachzutragen, die wir
bei Beschreibung der generellen Organisationsverhältnisse nur flüchtig
berühren konnten oder ganz übergehen mussten.
Fahndet man in unseren Gewässern auf Turbellarien, so besteht
das Fangergebnis am häufigsten aus Arten, welche den Gattungen
Macrostoma, Microstoma, Stenostoma,
Mesostoma und Vortex angehören. Aus diesen greifen
wir daher je einen Vertreter heraus und unterwerfen ihn einer etwas
eingehenderen Schilderung.
Macrostoma virideE.
v. Ben.
Dies ist ein etwa 2 mm grosses Tierchen, an dem man schon bei
Lupenbesichtigung den grünlich oder gelb gefärbten Darm[S. 256] erkennt.
Bei stärkerer Vergrösserung (Fig. 52) entdecken wir sofort ein
abgerundetes Vorderende und einen spatelartig geformten Schwanzteil,
der reichlich mit Stäbchenpaketen gespickt ist, die zumteil über
die Haut hervorragen. Auch der übrige Körper ist mit solchen
Rhabditenbündeln versehen, aber nicht in dem Masse wie das Kopf- und
Hinterende. Die Stäbchen liegen in sehr verschiedener Anzahl (zu 2–5
Stück) beisammen und haben eine keulenförmige Gestalt. Die Flimmerhaare
der Epithelzellen sind sehr fein und kurz; dazwischen stehen aber
lange Geisselborsten, die sich über die ganze Oberfläche des Tieres
verbreiten. Im Übrigen bietet die Organisation der Macrostomiden
mancherlei primitive Verhältnisse dar. Sie besitzen ein ganz einfaches
Schlundrohr, welches die schlitzförmige Mundöffnung (m) mit
dem Darmsack (d) verbindet. Das Gehirn (g) erscheint
lediglich als eine Bogenkommissur der beiden Längsnervenstämme, und
unterscheidet sich in Form und Masse nicht viel von diesen. Dasselbe
trägt indessen hochentwickelte Sehorgane, die mit einer stark gewölbten
Linse ausgestattet sind. Die Höhlung des Darmkanals ist durchweg
mit Cilien besetzt, deren Länge etwa der Höhe der Epithelzellen
gleichkommt, denen sie aufsitzen. Zu beiden Seiten des Darms liegen
die kompakten kegelförmigen Hoden (h) und dicht dahinter die
paarigen Ovarien (ov). Bei p sehen wir den Penis, der
bei allen Arten der Gattung Macrostoma mit einem chitinösen Ansatze
versehen ist. Bei der in Rede stehenden Spezies ist letzterer S-förmig,
aber so, dass die beiden Krümmungen nicht in einer Ebene liegen. Fig.
53 stellt diese Penis-Armatur bei sehr starker Vergrösserung dar.
Was das Wassergefässsystem anlangt, so kann man sich bei M.
viride die zwei seitlichen Hauptstämme leicht zur Ansicht bringen.
Es scheint, dass sich dieselben oberhalb des Mundes vereinigen und noch
einige kleine Verästelungen nach dem Vorderende hinschicken.
Die Macrostomiden gehören zu den Rhabdocölen mit zwei
Geschlechtsöffnungen, einer männlichen und einer weiblichen. Die
erstere liegt im verbreiterten Hinterteile, die andere in der
ungefähren Höhe der Ovarien; beide natürlich auf der Bauchseite.
Bei Macrostoma viride konnte ich auch einige Beobachtungen
über die Spermatozoen-Entwickelung machen, welche diesen
Vorgang in seinen Hauptzügen klarstellen. Zerzupft oder zerquetscht
man einen reifen Hoden vorsichtig auf dem Objektträger, so hat man in
einem und demselben Präparate gleich alle Stadien der Spermatogenese
beisammen. Ich habe dieselben der Reihe nach in Fig. 54 abgebildet und
mit Buchstaben bezeichnet. Den Ausgangspunkt für die Entwickelung der
Samenkörper bilden die vollkommen kugeligen und ganz durchsichtigen
Hodenzellen oder Spermatogonien (a). Diese zumeist
mehrkernigen Gebilde verwandeln sich durch Einschnürung ihres
Protoplasmaleibes in ebensoviele Spermatocyten oder Samenzellen
(d), die aber zunächst in Zusammenhang mit einander bleiben.
Jede einzelne Spermatocyte lässt zwei kleine Fortsätze aus sich
hervorsprossen, von welchen der eine in der Folge zum Schwanzfaden
des Spermatozoon, der andere zur sogenannten „Nebengeissel“ desselben
wird. Die einkernigen Spermatogonien (b) entwickeln sich genau
auf dieselbe[S. 258] Weise; nur zuweilen kommt es vor (c), dass die
Nebengeissel schon weit hervorgesprosst ist, ehe sich noch irgend eine
Spur von der Bildung des Schwanzfadens zeigt. Bei fortschreitender
Entwickelung lösen sich die Spermatocyten aus ihrem ursprünglichen
Verband (d) und nehmen das in e dargestellte Aussehen
an, indem sich ihr protoplasmatischer Teil in die Länge streckt.
Erst zu allerletzt kommt die bislang im Ruhezustande verbliebene
Kernsubstanz in Bewegung, um in dem immer spindelförmiger werdenden
Spermatozoenkopfe sich gleichmässig zu verteilen. In f sehen wir
dann das völlig fertige Samenkörperchen von Macrostoma viride,
wie es zu vielen hunderten in dem Präparate eines zerdrückten Hodens
vorhanden zu sein pflegt. Der Schwanzteil dieser Körperchen besitzt
eine ausserordentlich lebhafte Schlängelbewegung, wogegen die steife
Nebengeissel nur mit mässiger Geschwindigkeit hin und her pendelt.
Die am meisten verbreitete Spezies der Gattung Macrostoma ist übrigens
nicht diese, sondern M. hystrix Oerst.; in den wesentlichen
Bauverhältnissen stimmen aber beide mit einander überein. Die
letztgenannte Art ist dadurch merkwürdig, dass sie nicht bloss im
süssen, sondern auch im salzigen Wasser vorkommt. Auch lebt sie nicht
bloss in seichten Pfützen und Tümpeln, sondern verträgt ebensogut
den Aufenthalt in beträchtlichen Tiefen. Nach einer Angabe von
Duplessis ist sie sogar noch im Grundschlamme des Genfersees
zu finden[102]. An schwimmenden Holzstückchen und dem Wellenschlag
ausgesetzten Wasserpflanzen vermögen sich die Tierchen mit Hilfe
ihrer „Klebzellen“ festzuhalten, welche am untern Saume des
spatelähnlichen Hinterendes in grosser Anzahl befindlich sind. Solchen
eigentümlichen Haftapparaten begegnen wir auch bei mehreren anderen
Turbellariengattungen.
Microstoma lineareOerst.
Einzelindividuen von dieser Spezies kommen höchst selten vor;
gewöhnlich findet man nur Ketten, die, wie Fig. 55 veranschaulicht, aus
mehreren ungleichaltrigen Exemplaren bestehen. Dieser Befund findet
seine Erklärung in der Thatsache, dass Microstoma lineare[S. 259] sich
vorwiegend durch Querteilung oder, richtiger gesagt, durch terminale
Knospenbildung fortpflanzt. In nachstehender Figur haben wir eine
Kette von vier Individuen vor uns, deren Entwickelung wie folgt zu
denken ist. Aus dem ursprünglichen Muttertiere (A) ging B
als Tochterspross auf die nämliche Art hervor, wie jetzt die Knospe
b aus ihm selbst, oder wie a aus A. Der Zeit
nach ist die Reihenfolge dabei diese: A, B, b,
a. Das heisst: A erzeugte zunächst B; hierauf
entstand die Knospe b und dann erst die mit a bezeichnete
neue aus dem alten Mutterindividuum A.
Bei genauerer Beobachtung dieser eigentümlichen Fortpflanzungsweise
bemerkt man, dass stets nur das hinterste Drittel des Muttertieres zur
Erzeugung des Tochtersprosses verwendet wird. Mit dem Auftreten zweier
Augenpunkte und der Bildung einer Einschnürung, welche die künftige
Trennungsstelle andeutet, nimmt der Knospungsprozess seinen Anfang.
In der Folge vergrössert sich das Tochterstück allmählich und holt
mit seinem rascheren Wachstum den sich gleichfalls regenerierenden
mütterlichen Teil ein, bis beide in ihren Dimensionen sich fast
vollkommen gleichen. Mit der Neubildung eines Gehirns und Schlundes am
Spross erreicht die Entwickelung des letztern ihren Abschluss, und es
erfolgt seine Lostrennung vom Stammorganismus. Aber bevor er selbst
noch fertig ausgebildet war, wurde in seinem hintern Leibesdrittel
bereits die dritte Generation angelegt, bezüglich deren sich der
nämliche Sprossungsvorgang wiederholt, und so geht diese Vermehrung
immer fort, den ganzen Sommer hindurch.
Fig. 55. Microstoma lineare.
[S. 260]
Erst im Herbst treten andere Verhältnisse ein. Dann finden wir Ketten,
an deren Teilsprossen man die Anwesenheit von Geschlechtsorganen
wahrnimmt, woraus beiläufig hervorgeht, dass geschlechtliche und
ungeschlechtliche Zeugung bei demselben Individuum von Microstoma
lineare und zur nämlichen Zeit stattfinden kann. Jene Ketten sind
aber immer nur eingeschlechtlich, d. h. sie bestehen entweder aus
lauter weiblichen oder aus lauter männlichen Individuen. Die ersteren
besitzen ein einfaches Ovarium, die letzteren paarige, keulenförmige
Hoden und ein Begattungsorgan, dessen chitinöser Teil eine leichte,
schraubenförmige Windung zeigt.
Fig. 56.
Nesselkapsel mit hervorgeschnelltem Faden.
Über den sonstigen Bau von Microstoma lineare können wir
uns kurz fassen. Die flimmernde Epidermis besteht bei dieser
Spezies aus polygonalen Zellen, zwischen denen die birnförmigen
Nesselkapseln liegen. Diese sind über den ganzen Körper
zerstreut und gleichen in ihrem Aussehen fast ganz den gleichnamigen
Gebilden bei den Süsswasserpolypen, nur dass sie ein wenig kleiner
sind, als bei diesen. Sie haben eine Länge von 0.015 mm und
der herausgeschnellte Faden misst etwa 0.12 bis 0.14 mm. Am
Halsteile der Kapsel sitzen vier kleine Widerhaken. Leydig
hat mit gewohntem Scharfblick noch eine zweite Art von Nesselkapseln
bei Microstoma lineare entdeckt, die eine ovale Gestalt haben
und nicht mit Widerhaken versehen sind. Ich hebe diesen Umstand
ausdrücklich hervor, weil wir bei Hydra das gleiche Verhalten
beobachten. Fig. 56 stellt eine grössere Kapsel mit ausgestossenem
Faden dar.
Der Leibesraum ist bei Microstoma lineare von einem
reichen Maschenwerke heller Fasern durchsetzt, zwischen denen
zahlreiche kleine Bindegewebszellen und -kerne eingelagert sind. Das
Gehirn ist in dieses Parenchym eingebettet. Es besteht aus zwei
seitlichen Ganglien und einer Kommissur, die den Schlund umfasst. Zwei
Längsnerven durchziehen den Körper. Das Wassergefässsystem wurde
zuerst von M. Schultze bei dieser Spezies entdeckt, aber nicht
genauer beschrieben. Ich habe dasselbe neuerdings genauer studiert
und gefunden, dass zwei seitliche[S. 261] Hauptstämme vorhanden sind, deren
Verästelungen nach der Mittellinie zu konvergieren und ein ventral
stärker als dorsal entwickeltes Kanalnetz bilden[103]. Am Kopfende der
einzelnen Kettenglieder liegen die Augen in Gestalt rostroter
Pigmenthäufchen. Dicht dahinter befinden sich die Wimpergrübchen
— kleine tiefe Becher mit kreisrunder Öffnung, die innerlich mit
Flimmerepithel ausgekleidet sind. Der Darmtraktus besitzt,
wie man am lebenden Tiere deutlich sieht, ebenfalls wimpernde
Epithelzellen, und der einfache Schlund ist mit zahlreichen
Drüsenzellen (Pharyngealzellen) besetzt, die einen förmlichen Mantel
um denselben bilden. Die Farbe der Tierchen ist ein dunkles
Gelbbraun, sodass sie in dieser Beziehung fast genau mit Hydra
fusca übereinstimmen. Ein Hauptfundort für dieselben sind Tümpel
stehenden Wassers, deren Boden mit zerfallendem Laube oder sonstigen
Pflanzenresten bedeckt ist.
Stenostoma leucopsO. Schm.
Auch bei dieser Gattung haben wir es fast immer nur mit Ketten zu thun,
die aber selten aus mehr als zwei Gliedern bestehen. Ihre Länge beträgt
gewöhnlich 2–3 mm. Dem unbewaffneten Auge erscheinen sie als
kleine weisse Linien.
Wie bei Microstoma lineare, so finden wir auch hier zu beiden
Seiten des Kopfteiles Wimpergruben (Fig. 57 wg S. 262),
die mit einer zierlichen Zellenrosette umgeben sind. Echte Augen sind
nicht vorhanden; vielleicht sind aber die dem Gehirn anhängenden
eigentümlichen Bläschen (bl) als lichtperzipierende Organe zu
deuten.
Das Gehirn hat bei den Stenostomiden eine mächtige Entwickelung,
wie aus den Figuren 57 und 58 hervorgeht. Dasselbe setzt sich
auch schärfer als bei den übrigen Rhabdocölen gegen das umgebende
Bindegewebe ab. Es besteht aus zwei ausserordentlich grossen
mehrlappigen Ganglien, die unmittelbar vor dem Munde gelegen sind. Man
unterscheidet jederseits einen grössern hintern (h) und einen
schmälern Vorderlappen (v). Nach B. Landsberg[104],
der den Bau dieser Gebilde spezieller analysiert[S. 262] hat, überwiegt in
denselben der gangliöse Teil gegen den faserigen, was bei den anderen
Turbellarien nicht der Fall ist.
Darm und Schlund sind bei Stenostoma leucops
(Fig. 57) sowohl wie bei Stenostoma unicolor (Fig. 58)
in allen wesentlichen Stücken so gebaut wie bei Microstoma
lineare, sodass wir auf das dort Gesagte verweisen können. Das
Wassergefässsystem hingegen hat bei den Stenostomiden eine besondere
Gestaltung, insofern es aus einem einzigen Kanal besteht, der am
Hinterende ausmündet und von hier aus (der Mittellinie des Körpers
folgend) bis in den Kopfteil verläuft, um hier in einer Schlinge
umzubiegen und zurückzukehren. Was aus diesem rückläufigen Teile wird,
ist noch nicht genau klargestellt. L. v. Graff vermutet, „dass
derselbe sich in feinere Zweige auflöst“. Ich habe aber von einer
solchen Verästelung auch mit den besten Linsen nichts entdecken können.
Stenostoma unicolor unterscheidet sich von Stenostoma
leucops schon bei Lupenvergrösserung durch den besser markierten
Kopfteil, durch den schlankeren Habitus und durch zahlreiche einzellige
Drüsen im Darmepithel, welche schwärzliche Konkremente enthalten.
Beide Spezies sind sehr häufige Erscheinungen in unseren stehenden
und fliessenden Gewässern; sie sind auch sehr leicht aufzubewahren,
wenn man sie in kleine Glasdosen mit algenhaltigem Wasser bringt. In
derartigen Miniaturaquarien leben die Tierchen viele Monate[S. 263] lang und
pflanzen sich unausgesetzt durch terminale Knospung (vergl. S. 259)
fort.
Fig. 59. Catenula lemnae.
Zu den Stenostomiden stellt v. Graff auch das gewandt
schwimmende Meerlinsenkettchen (Catenula lemnae),
dessen Aussehen in Fig. 59 veranschaulicht ist. Diese Rhabdocöle ist
ein fast ständiger Bewohner kleiner Moortümpel und man trifft sie
in solchen stets massenhaft an. Wie bei den vorhin geschilderten
Formen, so vermehrt sich auch diese durch Querteilung. Am Vorderende
befindet sich eine kopfartige Anschwellung, in welcher das Gehirn
mit dem Hörbläschen (ot) liegt. Der cylindrische Schlund
flimmert auf seiner Innenfläche, ebenso der Darm, welcher übrigens,
falls er leer ist, nur mit Mühe wahrgenommen werden kann. Der Raum
zwischen Darm und Hautmuskelschlauch wird von sehr grossen, dicht
an einander gedrängten Parenchymzellen eingenommen, die in unserer
Figur mit pz bezeichnet sind. Das Wassergefässsystem hat genau
denselben Charakter wie bei Stenostoma leucops und stellt
einen äusserst feinen, vielfach geschlängelten Kanal dar, der auf
der dorsalen Seite des Tieres (subcutan) vom vordern bis zum hintern
Körperende verläuft. Abgesehen von diesem Exkretionskanal aber und
von dem Vorhandensein eines markierten Kopfteils kann ich zwischen
Catenula lemnae und den Stenostomiden auch nicht die geringste
morphologische Verwandtschaft erblicken, so dass ich es nur als einen
Notbehelf ansehen kann, wenn wir das Meerlinsenkettchen einstweilen
mit in die Gattung Stenostoma aufnehmen. Eigentlich steht Catenula
unter den Rhabdocölen völlig isoliert da; wir suchen vergeblich nach
Anknüpfungspunkten für dieses sonderbare Wesen, in welchem Dugès
seinerzeit ein den Bothriocephalen verwandtes Tier zu sehen glaubte.
[S. 264]
Mesostoma viridatumM. Sch.
Die Mesostomiden, d. h. diejenigen Rhabdocölen, welche eine auf der
Mitte der Bauchseite gelegene Mundöffnung und einen rosettenförmigen
Schlund besitzen, stellen eine sehr verschiedenartige Gesellschaft
dar, sodass Prof. M. Braun unlängst mit Recht den Vorschlag
gemacht hat, den Genusnamen Mesostoma ganz über Bord zu werfen,
resp. ihn nur noch für Arten beizubehalten, die ungenügend bekannt
sind. Die Charaktere dieser Gruppe sind von vornherein zu allgemein
gefasst worden und daher erklärt es sich, dass alles, was nach Lage
und Form des Pharynx ihr nicht ganz direkt widersprach, stets bequeme
Unterkunft darin finden konnte. In Fig. 60 sehen wir einen sehr kleinen
Repräsentanten der Gattung Mesostoma, ein hellgrünes Würmchen von etwa
Millimetergrösse, welches fast überall in klaren Gewässern zu finden
ist. Im Gegensatz zur Mehrzahl seiner Gattungsverwandten ist dasselbe
augenlos. Der Schlund liegt bei dieser Art etwa am Anfange des zweiten
Körperdrittels, und ebendaselbst gewahrt man eine Verbindung zwischen
den beiden seitlichen Stämmen des Wassergefässsystems, deren weiterer
Verlauf aber schwer zu verfolgen ist. Gleich hinter dem Pharynx
(ph) liegt die von Cilien umsäumte Geschlechtsöffnung
(gp) und nicht weit davon der birnförmige Penis (p),
der einen chitinösen Ausführungsgang besitzt. Die Leibeshöhle
ist fast ganz erfüllt von Parenchym, und nicht selten findet man
Individuen, welche im Innern 6–8 braunschalige, elliptische Eier
enthalten, so dass zwischen Darm und Leibeswand kaum noch ein freier
Raum übrig bleibt.
Fig. 60. Mesostoma viridatum.
In den Zellen der Epidermis entdeckt man zahlreiche Rhabditen
von 0.018 mm Länge, und bei tieferer Einstellung des Mikroskops
zeigen sich im Parenchym ganze Strassen solcher Stäbchen, die nach dem
Vorderende zu konvergieren.
[S. 265]
Die grüne Färbung des Tierchens rührt von winzigen,
chlorophyllhaltigen Körnchen her, die eine förmliche Schicht unter der
Haut bilden; dieselben sind höchst wahrscheinlich als einzellige Algen
zu betrachten, die in einem symbiotischen Verhältnisse zu ihrem Träger
stehen. Dergleichen Chlorophyllkörner kommen auch bei einigen anderen
Turbellarien und ausserdem bei mehreren Infusorienspezies vor.
In unseren europäischen Seen und Wassertümpeln sind im ganzen etwa
zwanzig Arten von Mesostomiden einheimisch. Eine ähnliche Verbreitung
derselben konstatiert W. A. Silliman für Nordamerika. Die
Gattung Mesostoma weist übrigens neben sehr kleinen (1–5 mm
grossen) Formen auch recht stattliche Vertreter auf. So z. B. das 10–15
mm lange und 4–5 mm breite Mesostoma Ehrenbergii,
welches vollkommen platt gestaltet und glashell durchsichtig ist. Diese
Spezies tritt oft so massenhaft auf, dass die untergetauchten Stengel
der Wasserpflanzen ganz damit bedeckt sind. Hiernach kommt Mesostoma
tetragonum O. Schm., welches 8 bis 10 mm lang wird. Durch
zwei Lamellen, die auf der Rücken- und Bauchseite des Tieres vom
vordern bis zum hintern Ende sich erstrecken und wie Flossen gebraucht
werden, hat dasselbe im Querschnitt eine vierkantige Gestalt. Diese
charakteristische Eigentümlichkeit ist in der Speziesbezeichnung zum
Ausdruck gebracht. Das ungemein häufige Mesostoma lingua O.
Schm. (7–8 mm) gehört auch noch zu den grösseren Erscheinungen,
wogegen die meisten anderen bei uns vorkömmlichen Spezies (z. B.
Mesostoma rostratum Ehrb., M. personatum O. Schm. etc.)
bedeutend kleiner sind und zwischen Algenfäden oft nur mit Mühe
entdeckt werden können.
Vortex truncatusEhrb.
Hier haben wir eine vollständig kosmopolitische Art vor uns, die
von Lappland bis nach Ägypten verbreitet ist, und die man in den
schlammigen Tümpeln der Ebene ebenso häufig antrifft, wie in klaren,
kalten Gebirgsseen. Es sind (Fig. 61 a S. 266) kleine, etwa 1.5
mm lange Tierchen von hell- oder dunkelbrauner Färbung. Sie
besitzen ein abgestutztes Kopfende und ein zugespitzes Schwänzchen.[S. 266]
Die Haut als solche ist vollkommen farblos und mit Stäbchen gespickt,
welche meist zu zweien bei einander liegen. Der Sitz des bräunlichen
Pigments sind die Bindegewebszellen des Parenchyms. Ist der Farbstoff
in grosser Menge vorhanden, so können die Würmchen ein fast schwarzes
Kolorit annehmen. Am Kopfe stehen zwei schwarze halbmondförmige Augen
und der Schlund hat die für die Vorticiden schon eingangs (S. 247)
erwähnte typische Tonnengestalt. Alle Vortex-Arten sind durch ein mehr
oder weniger kompliziertes (chitinöses) Kopulationsorgan ausgezeichnet,
welches aus paarigen Leisten mit Stachelbewehrung besteht. Bei der in
Rede stehenden Spezies (V. truncatus) ist dasselbe ziemlich
einfach gebaut. Es setzt sich aus zwei gabelig auseinanderweichenden,
etwas gekrümmten Ästen zusammen, die auf ihrer konkaven Seite eine
Reihe feiner Zähnchen (Fig. 61 b) tragen. Man bringt sich diese
Hartgebilde am besten zur Ansicht, wenn man mit möglichster Vorsicht
ein Quetschpräparat des ganzen Tieres herstellt.
Fig. 61. Vortex truncatus Ehrb.
Die meisten Vortex-Spezies sind kleine, unscheinbare Würmchen.
Nur V. viridis M. Sch. und V. scoparius O. Schm.
sind Riesen unter ihren Gattungsgenossen, insofern sie oft eine
Länge von 5–6 mm erreichen. Die letztgenannte Art besitzt
ein Kopulationsorgan, welches in seiner Form lebhaft an ein paar
kurzgestielte Stallbesen erinnert, die dicht beisammen liegen. Daher
der sonderbar klingende lateinische Beiname, der aber sehr treffend
gewählt ist. Zu den kleinsten Spezies, die wie V. truncatus
nur 1–1.5 mm gross sind, gehören V. pictus O. Schm.,
V. Hallezii v. Graff, V. sexdentatus v. Graff und V.
cuspidatus O. Schm.
Die vorstehenden kurzen Andeutungen über die speziellere Organisation
einiger häufiger vorkommenden Strudelwürmer sollten lediglich dazu
dienen, den Anfänger mit diesen interessanten,[S. 267] aber noch viel zu wenig
beachteten Tieren bekannt zu machen. Das eingehendere Studium derselben
muss an der Hand von Prof. L. v. Graffs ausgezeichneter
„Monographie der Turbellarien“ (Leipzig 1882, Wilh. Engelmann)
geschehen, in welcher auch ein umfassender Litteraturnachweis zu finden
ist, der noch die neuesten Arbeiten berücksichtigt. Ein Atlas mit
20 lithographischen Tafeln erläutert die bis in das kleinste Detail
gehenden Ausführungen des umfangreichen Textes. — Ein sehr brauchbares
litterarisches Hilfsmittel beim Studium der Turbellarien findet man
auch in einer 1885 erschienenen grösseren Abhandlung von Prof. M.
Braun (Rostock), welche die rhabdocöliden Strudelwürmer Livlands
behandelt und dieselben durch vorzügliche Abbildungen veranschaulicht.
Weiteres ersehe man aus der diesem Kapitel angehängten Übersicht der
Litteratur.
Präparationsmethode.
Es ist hier vielleicht der passendste Ort, einige Worte über
die Art und Weise zu sagen, wie man Turbellarien zum Zwecke der
mikroskopischen Untersuchung gut konservieren kann. Dies gelingt
meinen Erfahrungen zufolge am raschesten und sichersten mit erwärmter
Lösung von Quecksilberchlorid in Wasser. Für grössere Arten nimmt
man diese Flüssigkeit konzentrierter als für kleinere; aber in jedem
Falle erfüllt sie ihren Zweck. Die Tiere werden mit möglichst wenig
Wasser in ein Uhrgläschen gebracht und in dem Augenblicke, wo sie am
schönsten ausgestreckt sind, schnell mit dem Sublimat übergossen. Je
nach der Grösse der Objekte muss letzteres 5–20 Minuten einwirken. Dann
wässert man die getöteten Würmer längere Zeit aus, damit keine Spur
des Quecksilbersalzes in den Geweben zurückbleibt. Zur Aufbewahrung
benutzt man 70prozentigen Alkohol. Die so konservierten Rhabdocölen
und Dendrocölen halten sich Jahre lang und können jederzeit, nach
vorhergegangener Färbung, zur Herstellung von Schnittserien verwendet
werden. —
[S. 268]
Zur Anfertigung von Totalpräparaten, die alles Notwendige erkennen
lassen, hat Prof. M. Braun ein Verfahren angegeben[105],
welches hier mitgeteilt werden soll. Dasselbe eignet sich besonders
gut für kleine Formen. Der genannte Forscher verfährt dabei wie
folgt. Er lässt bei gelindem Deckglasdruck zu dem Wasser, in welchem
sich das zu konservierende Tier befindet, eine Mischung von 3 Teilen
Langscher Flüssigkeit[V] und einem Teil einprozentiger Osmiumsäure
zufliessen, die sehr rasch tötet, zumal wenn man sie etwas erwärmt
anwendet. Sobald das Tier undurchsichtig geworden ist, die Gewebe also
geronnen sind, wird die überschüssige Flüssigkeit mit Löschpapier
abgetupft und 45prozentiger Alkohol unter das Deckglas gebracht.
Derselbe wird im Verlauf einiger Minuten mehrere Male erneuert und dann
durch 70prozentigen ersetzt. Nunmehr kann man das Deckglas vorsichtig
abheben; das Tier bleibt in der Regel an letzterem oder am Objektträger
haften und wird in dieser Lage mit 90prozentigem Alkohol behandelt.
Hierauf giebt man 1–2 Tropfen Karminlösung auf das Objekt, die man etwa
drei Minuten einwirken lässt. Diese Zeit genügt zur Färbung; dann wird
mit destilliertem Wasser abgespült, mit allmählich zu konzentrierendem
Alkohol entwässert, bis man schliesslich zur Aufhellung mit Nelkenöl
oder Kreosot schreiten kann. Sodann erfolgt der Einschluss in
Kanada-Balsam und die Prozedur — welche etwa 20 Minuten in Anspruch
nimmt — ist beendet. Auf diesem Wege erhält man in vieler Beziehung
hübsche Präparate. Die Osmiumsäure bräunt gewisse Teile (Dotterstöcke
und Keimdotterstöcke z. B.) und hebt dieselben stärker hervor, während
die kernreichen Teile des Geschlechtsapparats (Hoden, Ovarien und
sonstige Drüsen) intensiv gefärbt werden. Leider kann man diese Methode
bloss bei solchen Arten verwenden, die sich etwas komprimieren lassen,
ohne zu zerplatzen. In anderen Fällen führt nur die Härtung der Tiere
(s. oben) und die Zerlegung derselben in Schnittserien zum Ziele.
[V] Diese besteht aus 5 Teilen Sublimat, 5 Teilen Eisessig und
100 Teilen Wasser.
[S. 269]
Geographische Verbreitung
der Turbellarien.
Durch zahlreiche faunistische Exkursionen, welche ich in dem Zeitraume
von 1884–1889 in den verschiedensten Teilen Deutschlands ausgeführt
habe[VI], hat sich mir die Überzeugung aufgedrängt, dass die
Verbreitung der Strudelwürmer nicht längs gewisser Linien erfolgt,
aus denen eine Abhängigkeit dieser Tiere von klimatischen Einflüssen
oder von der Bodenbeschaffenheit der bezüglichen Wasseransammlungen
zu erkennen wäre. Die Eier dieser Tiere scheinen vielmehr
durch zufällig sich darbietende Transportgelegenheiten überallhin
verschleppt werden zu können und da, wo es die äusseren Verhältnisse
nur irgend gestatten, die Ansiedelung der Art zu ermöglichen. Die
meisten Turbellarien-Eier sind hartschalig und widerstandsfähig, so
dass sie nicht leicht durch Stoss oder Druck Schaden nehmen können.
Mit solchen Eigenschaften ausgerüstet, können sie also selbst dann
gut erhalten bleiben, wenn das Muttertier, in dessen Leibeshöhle sie
sich befinden, bei der Luftreise am Gefieder eines Wasservogels durch
Austrocknung zu Grunde gehen sollte. Besucht nur der Transporteur
gelegentlich ein anderes Wasserbecken, so gelangen sie wohlbehalten
und lebenskräftig wieder in ihr Element und entwickeln sich dort
ebenso gut wie in dem Weiher oder Tümpel, welchem sie durch[S. 270] Zufall
— samt der mit ihnen trächtigen Mutter — entrissen wurden. Auf
diese Art erklärt sich auch die Launenhaftigkeit des Vorkommens
mancher Turbellarienspezies, die sonst völlig rätselhaft wäre. Es wird
uns von dem nämlichen Gesichtspunkte aus auch die Besiedelung hoch
gelegener Bergseen mit Strudelwürmern begreiflich, da man letztere
doch unmöglich als aktiv dorthin gewandert ansehen kann. Der
Kleine Koppenteich im Riesengebirge, der eine Höhenlage von
1180 m hat, beherbergt — wie schon eingangs mitgeteilt —
14 Arten von Turbellarien[VII], darunter auch eine sehr seltene Form
(Monotus lacustris Zach.), welche zu einer Gattung gehört, die
sonst nur Meeresbewohner umfasst[VIII]. Dieser Süsswasser-Monotus wurde
inzwischen auch im Peipus-See (Russland) und in zahlreichen Seen der
Schweiz aufgefunden. Nichtsdestoweniger bleibt es überraschend, dass
er durch die Gunst des Zufalls auch in jene verlorene Felsenschlucht,
auf deren Grunde der Kleine Koppenteich liegt, verpflanzt werden
konnte. Dies erklärt sich nur aus der Widerstandsfähigkeit seiner
Eikörper, die infolgedessen weit verschleppt werden können, ohne
ihre Entwickelungsfähigkeit einzubüssen. Und in gleicher Weise haben
wir uns die Ansiedelung der übrigen 13 Arten von Strudelwürmern an
jener Lokalität zu erklären, wobei aber nicht ausgeschlossen sein
soll, dass gelegentlich auch einmal eine lebende Turbellarie am
Gefieder eines Vogels hängen bleiben und dadurch in ein anderes (nahe
gelegenes) Wasserbecken übergeführt werden kann. Viele Strudelwürmer
besitzen sogenannte „Klebzellen“ an ihrem Hinterende, mit welchen
sie sich an allerlei feuchten Gegenständen festzuheften vermögen.
Diese eigentümlichen Organe werden ihre[S. 271] guten Dienste wohl auch beim
Transport durch Vögel oder Wasserkäfer leisten, wenn es gilt, einen
Halt gegen das Fortgewehtwerden durch Luftzug während des Fluges jener
Tiere zu gewinnen[106].
1. „Studien über die Fauna des Grossen und Kleinen Teiches im
Riesengebirge.“ Zeitschr. f. wiss. Zoologie. 41. Bd. 1885.
2. „Ergebnisse einer zoolog. Exkursion in das Glatzer-, Iser- und
Riesengebirge.“ Zeitschr. f. wiss. Zoologie. 43. Bd. 1886.
3. „Zur Kenntnis der pelagischen und littoral. Fauna norddeutscher
Seen.“ Zeitschr. f. wiss. Zoologie. 45. Bd. 1887.
4. „Faunistische Studien in westpreuss. Seen.“ Schrift. d. naturf.
Gesellschaft zu Danzig. 1887.
5. „Zur Kenntnis der Fauna des Süssen und Salzigen Sees bei Halle a.
d. S.“ Zeitschr. f. wiss. Zoologie. 46. Bd. 1888.
6. „Über das Ergebnis einer Seen-Untersuchung in der Umgebung von
Frankfurt a. d. O.“ Monatl. Mitteil. aus dem Gesamtgebiete der
Naturw. Nr. 8, 1888/89.
7. „Faunistische Untersuchungen in den Maaren der Eifel.“ Zool.
Anzeiger Nr. 295, 1888.
8. „Zur Kenntnis der Microfauna fliessender Gewässer Deutschlands.“
Biolog. Centralbl. Nr. 24, 1888.
[VII]Mesostoma rostratum Ehrb., Mesost. viridatum
M. Sch., Macrostoma viride van Bened., Stenostoma leucops
O. Schm., Vortex truncatus Ehrb., Vortex Hallezii
v. Graff, Prorhynchus stagnalis M. Sch., Prorhynchus
curvistylus M. Braun, Prorhynchus maximus n. sp., Gyrator
hermaphroditus Ehrb., Bothrioplana silesiaca n. sp.,
Bothriopl. Brauni n. sp., Monotus lacustris Zach. und
Planaria abscissa Ijima.
[VIII] Was das numerische Verhältnis der marinen
Turbellarien-Formen zu denjenigen des Süsswassers anbelangt, so
sei bei dieser Gelegenheit (nach v. Graff) angeführt, dass
160 Meeresbewohnern (darunter 15 Parasiten) 97 Süsswasser- und 1
Landbewohner (der hammerköpfige Prorhynchus) gegenüberstehen. —
Die Familie der Monotiden galt bisher für eine ausschliesslich marine
Gruppe.
Auch in viel höher als 1000 m gelegenen Bergseen finden wir noch
Turbellarien (Rhabdocölen sowohl als Dendrocölen), wie von Prof. Fr.
Zschokke (Basel) neuerdings an einigen Seen des Rhätikons, jener
gewaltigen Grenzkette zwischen Vorarlberg und Graubünden, nachgewiesen
worden ist. Er fand da oben noch Microstoma lineare und eine
Planarien-Spezies[107].
Diese Thatsache (und die Auffindung von Turbellarien in noch anderen
alpinen Seen) unterstützt die Theorie von deren passiver Migration,
sodass wir kein Bedenken tragen dürfen, uns die Verbreitung jener
leicht verletzbaren Würmer in derselben Weise zu denken, wie dies
von den kleinen Krebstieren längst bekannt ist, nämlich durch ihre
widerstandsfähigen Eikörper und die gelegentliche Verschleppung von
erwachsenen Exemplaren, welche am feuchten Gefieder eines wandernden
Wasservogels vor Austrocknung geschützt blieben und so eine Luftreise
nach einem andern Wasserbecken ohne Schaden für Leib und Leben zu
überstehen vermochten.
[S. 272]
Litteratur.
[91] Vergl. O. Zacharias, Zur Kenntnis der niedern
Tierwelt des Riesengebirges nebst vergleichenden Ausblicken. Mit 6
Illustrationen. 1890.
[92]E. Sekera, Prispěvky ku známostem o turbellariéch
sladnovodních. Prag 1888. Mit 4 Tafeln.
[93]L. v. Graff, Monographie der Turbellarien (I.
Rhabdocoelida). 1882. Mit 20 Figurentafeln.
[94]Isao Ijima, Untersuchungen über den Bau und
die Entwickelungsgeschichte der Süsswasser-Dendrocölen (Tricladen).
Zeitschr. f. wiss. Zoologie, 40. Bd. 1884. Mit 4 Tafeln.
[95]Paul Hallez, Embryogénie des Dendrocoeles d’eau
douce. Avec 5 planches. 1887.
[96]O. Zacharias, Über Fortpflanzung durch spontane
Querteilung bei Süsswasserplanarien. Zeitschr. f. wiss. Zoologie, 43.
Bd. 1885.
[97]W. Weltner, Dendrocoelum punctatum Pallas bei
Berlin. Sitzungsber. d. Königl. preuss. Akademie d. Wissenschaften.
1887. Mit 1 Tafel.
[S. 273]
[98]J. v. Kennel, Die in Deutschland gefundenen
Landplanarien. Arbeiten des Zool.-zootom. Instituts zu Würzburg. 5. Bd.
1879.
[99]F. Vejdovský, Note sur une nouvelle planaire
terrestre (Microplana humicola nov. gen., nov. sp.). Revue biologique
du Nord de la France. No. 4, 1890.
[100]M. Braun, Über Dorpater Brunnenplanarien. Mit 1
Tafel. 1881.
[101]O. Zacharias, Zwei neue Vertreter des Genus
Bothrioplana (M. Braun). Zool. Anz. No. 229, 1886.
[102]Du Plessis-Gouret, Essay sur la faune profonde
des lacs de la Suisse. 1885.
[103]O. Zacharias, Das Wassergefässsystem bei
Microstoma lineare. Zool. Anz. No. 196, 1885.
[104]B. Landsberg, Über einheimische Microstomiden.
Programm des Königl. Gymnasiums zu Allenstein. 1887.
[105]M. Braun, Die rhabdocölen Turbellarien Livlands.
Mit 4 Tafeln. 1885.
[106]O. Zacharias, Über Anpassungserscheinungen im
Hinblick auf passive Migration. Biolog. Centralbl. No. 4, 1889.
O. Schmidt, Die rhabdocölen Strudelwürmer des süssen Wassers.
1848. — Ergebnisse der Untersuchung der bei Krakau[S. 274] vorkommenden
Turbellarien. Sitzungsber. Akad. d. Wiss. zu Wien. 25. Bd. 1857. — Die
dendrocölen Strudelwürmer aus den Umgebungen von Graz. Zeitschr. f.
wiss. Zool. 10. Bd. 1858
und
M. Schultze, Beiträge zur Naturgeschichte der Turbellarien. Mit
7 Tafeln. 1861.
[S. 275]
Die Rädertiere
(Rotatoria).
Von Dr. Ludwig H. Plate in Marburg.
[S. 277]
Zu den Lebewesen des süssen Wassers, deren Körperbau wegen ihrer
geringen Grösse nur mit Hilfe des Mikroskopes studiert werden kann,
gehört eine Tiergruppe, die in manchen Organisationsverhältnissen sich
einerseits an die im vorigen Kapitel vom Herausgeber dieses Werkes
geschilderten Turbellarien anlehnt, anderseits einzelne Gattungen
enthält, deren Körper lange, mit Borsten besetzte, an die Gliedmassen
der Arthropoden in morphologischer und physiologischer Hinsicht
auffallend erinnernde Anhänge besitzt. Es ist die kleine, an häufigen
Gattungen nicht allzu reiche Familie der Rädertiere oder „Rotatoria“,
deren Besprechung sich demnach ungezwungen zwischen die von den
Strudelwürmern und den Entomostraken handelnden Schilderungen einfügt.
Die Rädertiere sind vorwiegend Bewohner des süssen Wassers und zwar,
man kann sagen, jeder Form, in der dasselbe in unseren Breitengraden
auftritt; doch enthält auch das Meer eine ganze Anzahl von Gattungen,
deren meiste Arten dem Süsswasser angehören, aber nur in seltenen
Fällen treten die marinen Vertreter in solcher Individuenzahl auf, dass
sie einen charakteristischen Bestandteil der pelagischen Fauna bilden.
Wir werden daher im folgenden vornehmlich auf die Süsswasserformen
eingehen und nur gelegentlich der Meeresbewohner Erwähnung thun. Die
Rotatorien sind, wie angedeutet wurde, keineswegs wählerisch in Bezug
auf ihre Fundplätze; der fliessende Strom und die sprudelnde Bergquelle
gewähren ihnen ebensogut alle Bedingungen zur Erhaltung des[S. 278] Lebens,
wie der stagnierende Sumpf, und die nie austrocknenden Wasserbecken
grösserer Teiche und Seen werden ebenso zahlreich von verschiedenen
Gattungen und Arten bevölkert wie die vergängliche Lache, die sich
irgendwo nach einem heftigen Regenschauer bildet und schon nach wenigen
Tagen der Macht der Sonne weichen muss. Selbst an Orten, die immer
nur ganz vorübergehend mit dem feuchten Elemente in Berührung kommen,
die der Zeitdauer nach viel eher als trocken denn als nass bezeichnet
werden müssen, also unter Existenzbedingungen, wie sie ungünstiger kaum
für einen Wasserorganismus gedacht werden können, selbst dort begegnen
wir dem munteren Völkchen der Rotatorien. Derartige Fundorte sind z. B.
die Rinnen auf den Dächern alter Häuser, sobald sich in ihnen Sand und
Schlamm angesammelt hat; ferner leben viele Rotatorien in dichten Moos-
und Flechtenpolstern sowie in den oberflächlichen Erdlagen zwischen den
Wurzeln von Gräsern und anderen Kräutern. Da die Rotatorien allseitig
von Wasser umspült sein müssen — eine feuchte Atmosphäre genügt nicht
—, um ihre Lebenskräfte bethätigen zu können, so folgt daraus, dass
dieselben an den letztgenannten Fundorten nur nach einem Regenguss oder
nach reichlichem Taufall zum Genusse ihres Daseins kommen. Sobald ihr
Lebenselement wieder abgeflossen oder verdunstet ist, schrumpfen sie
zu einem winzigen Körnchen zusammen, das regungslos daliegt und wie
ein Staubatom vom Winde überall hin geführt werden kann, um bei der
nächsten Anfeuchtung wieder aufzuquellen und, vom Scheintod erweckt,
aufs neue in den Kreis der Lebewesen einzutreten. Viele Rädertiere
— wir können sie als „Erdrotatorien“ den ständigen Wasserbewohnern
gegenüberstellen — sind demnach in wunderbarer Weise an solche
periodisch auftretende Lebensbedingungen angepasst und führen ein
intermittierendes Dasein, auf das ich eingehender weiter unten (s. IV)
zu sprechen kommen werde.
Obwohl nun die meisten Wasserrotatorien ausgezeichnete Schwimmer
sind, die vielfach pfeilschnell sich mit Hilfe ihres „Räderapparates“
zu bewegen vermögen, so giebt es doch nur verhältnismässig wenige
Gattungen, die beständig schwimmen und[S. 279] daher auch dort vorkommen, wo
das Wasser fast frei von Pflanzen und Treibmaterial ist, also in der
Mitte unserer Teiche und Seen. Man pflegt gegenwärtig die im offenen
Meere lebenden Tiere und Pflanzen im Gegensatz zu den die Küste
und den Meeresgrund bewohnenden als „pelagische“ zu bezeichnen und
wendet diesen Ausdruck, übertragend, aber wenig logisch, auch auf die
Süsswasserfauna an. Zu den pelagischen Rädertieren des Süsswassers
gehören vornehmlich die Gattungen Asplanchna, Anuraea,
Triarthra, Polyarthra und Synchaeta, für die
weiter charakteristisch ist, dass sie dort, wo sie vorkommen, meist
in sehr grosser Individuenzahl, zu Tausenden und Abertausenden,
angetroffen werden. Die Mehrzahl der Rotatorien liebt es hingegen,
abwechselnd zu schwimmen und sich vorübergehend mittels besonderer am
Hinterende des Körpers angebrachter „Fussdrüsen“ festzuheften, und
sie halten sich aus diesem Grunde zwischen Wasserpflanzen oder, bei
seichten Gewässern, dicht über dem Boden auf. Bei einigen wenigen
Gattungen endlich ist die Beweglichkeit noch mehr beschränkt. So
treffen wir in der Abteilung der Philodiniden nur Formen an, die,
wenn sie ungestört sind, sich fest irgendwo vor Anker legen und ihren
Räderapparat nur zum Herbeistrudeln von Nahrung benutzen; erst wenn sie
beunruhigt werden, verlassen sie teils spannerraupenartig kriechend,
teils frei schwimmend den Platz. Endlich giebt es noch eine kleine
Familie, die Melicertiden, welche nur in der Jugend frei beweglich
sind, später aber sich dauernd an Wasserpflanzen oder untergetauchten
Gegenständen festheften.
Ehe wir uns nun einer vergleichenden Schilderung der ganzen Gruppe
der Süsswasserrotatorien zuwenden, wollen wir eine bestimmte Art
möglichst eingehend anatomisch und biologisch untersuchen, um so
einen gesicherten Ausgangspunkt zu gewinnen, von dem aus der Leser in
das Studium dieser interessanten kleinen Wesen eindringen und einen
Massstab an die innerhalb derselben vorkommenden Variationen des
Körperbaues und der Lebensweise anlegen kann. Wir wählen als Typus der
Klasse eine der gemeinsten, überall sich findenden Arten, die zuerst
von dem dänischen Zoologen Otto Friedrich Müller 1773 entdeckt[S. 280]
und später von dem grossen Berliner Erforscher der kleinsten Lebewesen,
Christian Gottfried Ehrenberg, im Jahre 1838 unter dem Namen
Hydatina senta, das Krystallfischchen,
beschrieben wurde. Das Tierchen gehört zu den ständigen
Wasserbewohnern, und wir können uns dasselbe ebenso wie seine übrigen
Verwandten mit Hilfe eines sehr engmaschigen seidenen kleinen Netzes
aus sogen. „Müllergaze“ leicht verschaffen. Exemplare von Hydatina
finden sich häufiger in kleinen stehenden Tümpeln als in grösseren
Wasserbecken, was vielleicht nur damit zusammenhängt, dass sie sich
in letzteren auf einen grösseren Raum verteilen, während sie sich
in ersteren nicht selten in enormen Scharen ansammeln, sodass wir
nur für einige Augenblicke das Netz durch das Wasser hin und her zu
ziehen und dann den im Grunde desselben angesammelten Niederschlag in
einem Wasserglase abzuspülen brauchen, um mit einer schwachen Lupe
Tausende von durchsichtigen etwa ½ mm langen Hydatinen sich
rastlos im Wasser umhertummeln zu sehen. Sehr häufig erkennt man von
jedem Tierchen nicht viel mehr als einen grünen zentralen Fleck, der
sich bei Besichtigung mit einem Mikroskope als ein im Magen liegender
Nahrungsballen von kleinen grünen Algen oder Flagellaten erweist. Um
eine volle Einsicht in den Bau der Hydatinen zu gewinnen, bedarf es
eines Mikroskopes, dessen Vergrösserungskraft sich etwa bis zu einer
500fachen steigern lässt. Mit Hilfe eines „Tropfenzählers“ übertragen
wir einige der Tierchen in ein Uhrschälchen, greifen zur Lupe und
bringen eines derselben mittels der Pipette in einem kleinen Tropfen
auf den Objektträger, bedecken ihn mit einem Deckglase und betrachten
nun das Rädertier mit einer schwachen Vergrösserung. Jenachdem sich
noch relativ viel oder wenig Wasser unter dem Deckglase befindet,
schwimmt das Tier rasch oder langsam umher, sodass wir — durch Zusatz
oder durch Absaugen von Wasser — es vollkommen in der Hand haben,
die Geschwindigkeit der Fortbewegung so zu regulieren, dass eine
Beobachtung der natürlichen Körpergestalt möglich ist. Zum[S. 281] Studium der
inneren Organisation ist es jedoch unumgänglich nötig, die lebhaften
Bewegungen der Hydatina ganz oder nahezu zu hemmen, und hierzu stehen
uns drei Mittel zur Verfügung. Einmal lässt sich durch Absaugen eines
Teiles des Wassers ein solcher Druck auf das Objekt ausüben, dass
dasselbe fast unbeweglich daliegt; wird die Körpergestalt hierbei
auch unnatürlich breit, so hat doch diese Methode den grossen Vorzug,
die Lebensäusserungen fast aller Organe der Beobachtung selbst mit
starken Vergrösserungen zugängig zu machen. Zweitens kann man durch
Zusatz eines Tropfens einer dünnen Lösung von Kokain (1 Teil auf 20
Teile Wasser) die Hydatinen in vielen Fällen vorübergehend lähmen;
leider ist dieses Mittel nicht immer zuverlässig und geht auch nur
zu oft Hand in Hand mit Verzerrungen der natürlichen Körpergestalt.
Der letztere Übelstand beeinträchtigt sehr häufig auch die Resultate,
welche mit der dritten Methode, der Abtötung unter dem Deckglase oder
im Uhrschälchen, erzielt werden. Am meisten anzuraten ist die Anwendung
1%iger Überosmiumsäure, da diese die Gewebe vorzüglich fixiert. Um
das Schwarzwerden der Objekte zu verhüten, ist es nötig, dieselben,
nachdem die Säure ½ bis 2 Minuten eingewirkt hat, mit destilliertem
Wasser auszuwaschen. Eine Abtötung durch langsames Erwärmen des Wassers
erhält die Tiere nicht selten schön ausgestreckt, ist aber bei feineren
histologischen Studien nicht verwertbar.
Die Körpergestalt der Hydatina senta (Fig. 62 S. 282)
ist die eines nicht sehr breiten, dafür aber ziemlich hohen Kegels,
etwa eines Zuckerhutes. Der Basis des Kegels entspricht das vordere
Körperende, während die Spitze in zwei kleine dolchförmige Anhänge,
die „Fusszehen“, ausgezogen ist. Die Gestalt des völlig ausgestreckten
Tieres weicht nur darin von der eines mathematischen Kegels ab, dass
sie erstens etwas abgeplattet, gleichsam zusammengedrückt ist, sodass
der Querschnitt keinen Kreis, sondern ein Oval darstellen würde, und
dass sie zweitens hinter dem Vorderende eine ringförmige Einschnürung
besitzt, um sich dafür in der Mitte um so stärker auszubauchen. Es
ergiebt sich daraus eine Gliederung des Körpers in drei undeutlich von
einander[S. 282] geschiedene Regionen, die von vorn nach hinten als Kopf,
Mittelleib und Schwanz (letzterer von der Spitze des Kegels und den
zwei Zehen gebildet) unterschieden werden. Betrachten wir den Kopf
genauer, so sehen wir, dass seine nach vorn gekehrte Fläche nicht
eben ist, sondern dass sie sich zu einer tiefen trichterförmigen
Grube (gr) einsenkt. Der Rand dieses Trichters beschreibt kein
regelmässiges Oval, sondern ein Dreieck, dessen zwei bauchständige
oder „ventrale“ Seiten nach hinten winkelig vorgezogen sind, sodass
sie etwas hinter dem „Dorsal-Rand“ liegen. Denken wir uns demnach
einen Kegel an seiner Basis schräg abgestutzt und die so erhaltene
Fläche trichterförmig eingestülpt, so ergeben sich annähernd gleiche
Verhältnisse wie am Kopfe der Hydatina senta.
Schon eine flüchtige Betrachtung unseres Tierchens zeigt uns, dass
die Kraft, welche dasselbe so[S. 283] munter im Wasser umhertreibt, vom
Kopfende ausgehen muss, denn während der Mittelkörper und der Schwanz
regungslos durch das flüssige Element dahingleiten, bemerken wir am
Kopfe eine lebhafte Strudelung, deren Ursache uns ein starkes Objektiv
erkennen lässt. Der ganze Rand des Kopfes und auch ein grosser Teil
der Trichterwandung ist mit zahllosen Härchen (r) besetzt,
die abwechselnd schnell hakenförmig zusammenknickend nach hinten
schlagen und sich dann wieder langsam ausstrecken; jedes Härchen
stellt gleichsam ein Ruder dar, das um einen festen Punkt herum hin
und her bewegt wird. Bei manchen Rotatorien, namentlich den meisten
Erd- und Moosbewohnern, ruft das Spiel dieser Wimpern oder „Cilien“
den Eindruck eines sich drehenden Rades hervor; indem nämlich
immer nur wenige Ciliengruppen in den Fokus der betreffenden Linse
gleichzeitig hineintreten, und diese dabei den Kopfrand umkreisend
nach einander sichtbar werden, wird auf das Auge ein ähnlicher Reiz
ausgeübt, wie ihn die Speichen eines in Drehung befindlichen Rades
hervorrufen. Aus diesem Grunde hat man der ganzen Abteilung den Namen
„Rädertiere“ gegeben, und bezeichnet man die Summe der zur Bewegung
dienenden Härchen als „Räderapparat“ oder „Räderorgan“.
Bei unserer Hydatina hält es nun gar nicht leicht, den Bau dieses
Wimpertrichters genau festzustellen, so dass in Einzelheiten die
Angaben der verschiedenen Forscher vielfach differieren. Unsere
Abbildung zeigt, wie der Räderapparat im wesentlichen aus zwei
Cilienkränzen, einem äussern und einem innern, besteht. Jener wird aus
einer Reihe sehr zarter und langer Cilien gebildet, zwischen die sich
nur auf der Ventralseite, ungefähr in der Mitte jeder Seitenhälfte,
einzelne derbere Borsten einschieben. Der innere Kranz hingegen ist
vielgestaltiger und zerfällt in drei von einander getrennte Zonen.
An der Rückenwand erhebt er sich zu fünf grösseren Polstern, die mit
starken Borsten besetzt sind und hinter denen eine Doppelschnur von
kleineren Cilien entlang zieht. Auf der Ventralfläche des Kopftrichters
breitet sich jederseits der Medianlinie ein Wimperband aus, das aus
einer Reihe grosser und einer Reihe kleiner Härchen gebildet wird.
Endlich ist auch noch der[S. 284] Grund des Trichters mit zahlreichen
kleinen Cilien besetzt, deren Anordnung sich nicht weiter verfolgen
lässt. Der Räderapparat hat nun nicht allein den Zweck, die Hydatina
unter beständigen Umdrehungen um die Längsachse durch das Wasser zu
treiben, sondern erfüllt auch die viel wesentlichere Aufgabe, die
nötige Nahrung herbeizustrudeln. Unser Krystallfischchen nährt sich,
wie der meist grüne Mageninhalt beweist, vornehmlich von Flagellaten
(Euglenen u. dergl.), kleinen Algen, Diatomeen, Infusorien und
ähnlichen niedrigsten Organismen. Der vom Cilienapparat hervorgerufene
Strudel packt dieselben und schleudert sie auf den Grund des Trichters
(gr) in die daselbst gelegene Mundöffnung, von wo aus sie durch
andere Wimpern in den eigentlichen Verdauungskanal getrieben werden.
Die geschilderten Cilien des Kopftrichters besitzen die Kraft zu ihrer
rastlosen Thätigkeit in sich selbst, sie bewegen sich spontan und
werden nicht etwa durch Muskeln bewegt. Sie müssen naturgemäss fest
in der Haut wurzeln, und daher sehen wir diese am Kopfe etwas
stärker entwickelt als an anderen Körperstellen.
An der glashellen Körperwand der Hydatina lassen sich zwei verschiedene
Lagen unterscheiden. Die äussere (c) ist völlig homogen und
gegen dünne Kalilauge sehr widerstandsfähig, sodass wir ihr eine
hornartige, „chitinige“ Beschaffenheit zuschreiben dürfen. Die innere
(hg) hingegen ist feinkörnig, protoplasmatisch und enthält
von Stelle zu Stelle einen Kern. Besondere Zellgrenzen lassen sich
in dieser Protoplasmaschicht nicht erkennen, sie stellt vielmehr ein
„Zellsyncytium“, d. h. eine durch Verschmelzung der ursprünglich
getrennten Zellen einheitlich und kontinuierlich gewordene Lage dar.
Man bezeichnet sie als „Hypodermis“ und nimmt an, die äussere Schicht,
die „Kutikula“, sei durch Abscheidung von ihr erzeugt. Nur in zwei
Körperregionen schwillt die Hypodermis zu ungewöhnlicher Dicke an.
Dieselben liegen an den beiden Polen des Körpers, nämlich einmal
längs des ganzen Randes des Kopftrichters, woselbst die Hypodermis
zu zahlreichen halbkugeligen Polstern sich verdickt, welche eben den
Cilien des Räderapparates die gewünschte feste Unterlage bieten, und
ferner in den Zehen[S. 285] des Schwanzes, in denen die Hypodermis sich zu
den zwei langen schlauchförmigen und ein gutes Stück nach vorn in die
Leibeshöhle hineinragenden „Fuss- oder Klebdrüsen“ (f)
erweitert. Beobachtet man eine lebende Hydatina, so wird man bald
bemerken, dass diese keineswegs beständig umherschwimmt, sondern dass
sie von Zeit zu Zeit sich an irgend einem untergetauchten Gegenstande
für wenige Minuten festheftet. Das Spiel der Wimpern hört unterdessen
nicht auf, dient dann aber lediglich seiner zweiten (nutritorischen)
Funktion. Die Anheftung erfolgt mittels eines klebrigen Sekretes, das
in Tropfenform von den Fussdrüsen an der Spitze der Zehen durch eine
sehr kleine Öffnung entleert wird und bei Berührung mit dem Wasser
sofort zu einer festen Masse erstarrt. Die Drüsen sind ebenso gebaut
wie die Hypodermis, aus der sie hervorgegangen sind; sie stellen also
ein Zellsyncytium dar ohne zentrales Lumen, d. h. ohne innere Höhle.
Nach vorn läuft jede Drüse in einen feinen bindegewebigen Faden aus,
der sich an der Körperwand befestigt und dadurch das Organ in der
Leibeshöhle suspendiert erhält. Die Ausmündung erfolgt durch ein feines
Röhrchen.
Von den inneren Organen des Krystallfischchens fallen durch ihre
Grösse dem Beobachter vornehmlich diejenigen sofort auf, welche der
Selbsterhaltung und der Fortpflanzung des Tierchens dienen, die
Ernährungs- und die Geschlechtsorgane. Zu den ersteren
kann man, wie wir oben gesehen haben, auch den Cilienbesatz des Kopfes
rechnen. Die von demselben herbeigestrudelten Nahrungsteilchen treten
am Grunde des Kopftrichters in die Mundöffnung über, welche
nicht genau in der Mitte des von den Cilien umstellten Feldes liegt,
sondern stark ventralwärts verschoben ist. Dies hat zur Folge, dass
die dorsale Fläche des Kopftrichters etwas grösser ist, als jede der
beiden ventralen, wie dies auch aus der Betrachtung unserer Abbildung
erhellt. Der Verdauungskanal erstreckt sich unter der Rückenhaut als
ein gerades, der Medianlinie folgendes Rohr nach hinten und gliedert
sich in vier Abschnitte, die vom Munde aus gerechnet nach einander
als Kauapparat oder Mastax, Schlund oder Oesophagus, Mitteldarm[S. 286]
oder Magen und Enddarm oder Rectum bezeichnet werden können. Der
Kauapparat (ma) ist ein relativ grosses, querovales, am
Hinterrande dreilappig ausgezogenes Gebilde, das sich nicht direkt
an die Mundöffnung anschliesst, sondern mit dieser durch ein kurzes
flimmerndes Rohr verbunden ist. Ich sehe in diesem keinen besonderen
Abschnitt des Verdauungskanales, sondern rechne es mit zur Mundöffnung.
Der Mastax wird aus einem äusseren protoplasmatischen Mantel und einem
zentralen chitinigen Kieferapparat zusammengesetzt, dessen eigenartige
Bewegungen dem Beobachter sofort ins Auge fallen. Wie etwa die Backen
eines Nussknackers durch den Druck der Hand gegen einander bewegt und
nach Sprengung der Schale wieder von einander entfernt werden, so
rücken die beiden Seitenteile des Gebisses um einen festen Angelpunkt
abwechselnd gegen und von einander und zermalmen so jedes zwischen
sie geratene Nahrungsteilchen. Der Kieferapparat ist im einzelnen
so kompliziert eingerichtet, dass man seinen Bau nur nach völliger
Isolierung erkennen kann. Bei der Kleinheit des Objektes ist an ein
Zerzupfen des Tieres mittels feiner Nadeln nicht zu denken; wir
opfern daher lieber ein Individuum und bringen einen Tropfen dünner
Kalilauge unter das Deckglas. Dieselbe zerstört, vornehmlich in der
Hitze, sämtliche protoplasmatischen Bestandteile der Hydatina, sodass
nur der chitinige Zahnapparat erhalten bleibt. Jetzt hält es nicht
schwer, sich eine ungefähre Vorstellung von dem zierlichen Gerüste der
Kiefer zu bilden: zwei nach vorn gerichtete Seitenteile werden hinten
durch ein stabförmiges unpaares Gebilde zusammengehalten. Sie tragen
als wichtigsten Bestandteil auf der Innenfläche fünf lanzettförmige,
quergerichtete Zahnleisten, die beim Zusammenklappen des Gebisses
sich zwischen einander schieben und so die Nahrung zerquetschen. Die
den Kieferapparat umhüllende Plasmamasse ist offenbar zum grossen
Teile muskulöser Natur und bewirkt die rhythmischen Bewegungen, wenn
sie auch keine fibrilläre Struktur erkennen lässt. Einzelne Partien
lassen aus ihrem Aussehen auf eine drüsige Funktion schliessen. Auf
der Dorsalfläche und nahe dem Hinterrande des Mastax entspringt der
kurze, sehr enge und schwer sichtbare[S. 287]Schlund. Abgesehen vom
Kauapparat ist er der einzige Abschnitt des Verdauungskanales, welcher
der Flimmercilien entbehrt. Der sich an ihn anschliessende Magen
(sto) ist gross, sackförmig, vorn und hinten etwas verschmälert;
in ihm pflegen bei frisch gefangenen Tieren die Nahrungsbestandteile
so dicht angehäuft zu liegen, dass, wie gesagt, die ganze Hydatina
unter der Lupe nur wie ein wandelnder grüner oder schwärzlicher Fleck
erscheint. Unter dem Mikroskope erweisen sich jene Bestandteile
grösstenteils in beständiger zitternder Bewegung, die durch die langen
Cilien der aus einer Schicht grosser polygonaler Zellen gebildeten
Wandung hervorgerufen wird. Besonders gross sind diese Flimmerhaare am
Übergange des Schlundes in den Magen. Neben dieser Stelle sitzt dem
Magen noch ein Paar drüsiger, birnförmiger Anhänge (dr) an, die
einzigen, welche dem Verdauungskanal überhaupt zukommen. Sie dienen
vermutlich als Leber, indem sie ein die Verdauung unterstützendes
Sekret in den Magen entleeren. Der Enddarm unterscheidet sich
im histologischen Bau nur unwesentlich von dem Magen, von dem eine
ringförmige, muskulöse Einschnürung ihn trennt. Er geht nicht direkt
in den querspaltigen, ungefähr am Anfange des letzten Körperviertels
auf der Rückenseite gelegenen After (a) über, sondern
beide trennt ein kurzer, flimmerloser Zwischenkanal, die sogen.
Kloake. Sie nimmt ausser dem Enddarm die Ausführgänge der
Zeugungs- und Exkretionsorgane auf.
Die Ausscheidung einer harnartigen Flüssigkeit, welche dem Körper
nicht mehr dienliche stickstoffhaltige Stoffe entfernt, besorgen zwei
schmale, dünnwandige Röhren, die Nephridien (ne) —
mit einem unpassenden Namen auch wohl als „Wassergefässe“ bezeichnet
—, welche sich vom Kopfabschnitt der Leibeshöhle an seitlich nach
hinten bis zur Kloake erstrecken; sie verschmelzen hier zu einer
gemeinsamen Blase, einem Harnreservoir (c.v), das in die
Kloake einmündet und seinen Inhalt durch rhythmische Kontraktion
seiner Wände in diese entleert. Bei einem gesunden Tiere folgen
Zusammenziehung und Ausdehnung der Blase einander in regelmässigem
Wechsel, mehrere Male in jeder Minute, sodass man von einem Pulsieren
derselben sprechen kann. Jedes Nierengefäss zeigt[S. 288] aussen eine zarte,
feinkörnige, protoplasmatische Wandung mit hie und da eingestreuten
Kernen und innen einen engen Kanal. An zwei kleinen Stellen, nämlich
vorn im Kopfe und in der Höhe der Magendrüsen, windet sich der Kanal
knotenartig mehrfach hin und her, und hier findet sich dementsprechend
auch eine reichlichere Ansammlung von Protoplasma. Die beiden vorderen
Knäuelpartien stehen ausserdem durch ein enges, bogenförmig unter
der Rückenhaut (genauer gesagt: dem Gehirn) verlaufendes Quergefäss
mit einander in Verbindung. Jeder Nierenschlauch trägt eine Anzahl
eigenartiger sehr kleiner Anhänge (z), die man auch bei den
Turbellarien vorgefunden hat, und die nach der flackernden, unsteten
Bewegung, die man in ihrem Innern stets bemerkt, als „Zitterflammen“
oder „Zitterorgane“ bezeichnet werden. Es sind Ausstülpungen des
Kanallumens, die sich bald mit einer dreieckigen Breitseite, bald
mit einer birnförmigen Schmalseite dem Beobachter präsentieren. Jene
Bewegung wird hervorgerufen durch die Schwingungen einer kleinen
Membran, die dem freien, nach aussen geschlossenen Ende der Ausstülpung
aufsitzt und in ihr Lumen hineinragt. Die ganze Bildung ist wohl
als eine eigentümliche Wimperzelle anzusehen, deren physiologische
Bedeutung freilich noch ganz unklar ist.
Untersucht man eine grosse Anzahl von ausgewachsenen Hydatinen,
so wird man erstaunt darüber sein, bei allen auf der Ventralseite
und in der Mitte des Körpers ein grosses Organ anzutreffen, das
unzweifelhafte Eier enthält und demnach als Geschlechtsorgan des
Weibchens angesehen werden muss. Vergebens aber suchen wir in
diesem Tiere und in anderen Individuen nach irgendwelchen Andeutungen
von einer männlichen Sexualdrüse. Es hat lange gedauert, bis die hier
obwaltenden Verhältnisse klar erkannt worden sind. Bei der Hydatina
sind nämlich fast alle Individuen weiblichen Geschlechtes, und der
Prozentsatz der Männchen ist ein so ausserordentlich niedriger, dass
auf viele Hunderte von ♀ nur ein oder einige wenige ♂ kommen. Die ♂
weichen auch in Grösse und Bau so erheblich von den ♀ ab, dass wir
weiter unten diesen Dimorphismus der Geschlechter besprechen[S. 289] werden.
Da nun die ♂ so sehr viel seltener sind, haben die Hydatinen die
Fähigkeit erworben, sich auch ohne die Männchen „parthenogenetisch“
fortzupflanzen, d. h. die von ihnen erzeugten Eier bedürfen keiner
Befruchtung, sondern entwickeln sich ohne diese „jungfräulich“ zu
neuen Tieren. In dem Ovar bemerkt man zunächst nur eine dichtkörnige
dunkele Protoplasma-Masse (dst), in der acht ovale, von
einem schmalen, hellen Hofe umgebene Gebilde liegen. Wir sehen
hierin acht Kerne, deren Kernkörperchen eine riesige Ausdehnung
gewonnen haben, sodass der eigentliche Kern auf jene Randzone sich
beschränkt. Früher nahm man an, dass diese Kerne sich, von etwas
Protoplasma umgeben, abschnürten und sodann zu Eiern heranwüchsen.
Es hat sich nun neuerdings gezeigt, dass diese Anschauung nicht
richtig ist, sondern dass jene zusammenhängende mit acht grossen
Kernen versehene Protoplasma-Masse nur einen Teil des Ovars darstellt
und zwar denjenigen, der nicht die Eier erzeugt, sondern diese nur
mit Nahrungsdotter versorgt und daher als Dotterstock zu
bezeichnen ist. An seinem Vorderrande erblickt man an Tieren, die mit
Osmium-Säure abgetötet, und deren Kerne hinterher durch irgend eine
der gebräuchlichen Flüssigkeiten, etwa Borax-Karmin, gefärbt worden
sind, noch eine Anzahl sehr kleiner und dicht zusammenliegender Kerne.
Diese stellen den Eierstock (est) dar, denjenigen Teil
der Geschlechtsdrüse, welcher die Keimzellen, die Eier, liefert. Wie
ein Blick auf unsere Abbildung lehrt, werden die Kerne des Eierstockes
nach der rechten Ecke des Vorderrandes (die Bauchseite des Tieres ist
dem Beobachter zugewendet) zu immer kleiner. Sie liegen hier, wie es
scheint, in einem kontinuierlichen Protoplasma-Lager, an das sich nach
links zu deutlich gesonderte kleine Eizellen anschliessen. Je mehr
wir uns der linken Ecke des Ovars nähern, um so grösser werden die
Eizellen, und die grössten schieben sich dabei nach hinten vor und
kommen so neben den linken Seitenrand des Dotterstockes zu liegen,
dem sie sich dicht anschmiegen. In dieser Stellung beginnt nun auch
die ernährende Thätigkeit des Dotterstockes. Letzterer, wie auch die
Eizelle, werden von einer sehr zarten Membran umhüllt, und durch diese
diffundiert die im[S. 290] Dotterstock angesammelte Dottersubstanz hindurch
und gelangt so in die Eizelle hinein. Dieser Übertritt kann natürlich
nur in einer flüssigen Form geschehen und entzieht sich deshalb der
direkten Beobachtung. Aber der Umstand, dass die dem Dotterstocke
angelagerten Eizellen sehr bedeutend an Grösse zunehmen, während die
Plasma-Masse des Dotterstockes abnimmt, lässt keine andere Deutung der
Verhältnisse zu. — Die gesamte Geschlechtsdrüse wird von einer dünnen
Membran sackartig umgeben. Hinter dem Dotterstock umschliesst dieselbe
einen trichterförmig sich nach hinten verengernden Raum, den „Uterus“
(ut), welcher neben der kontraktilen Blase der Nephridien in
die Kloake mündet. Sobald die Eier ihre definitive Grösse erreicht
haben, fallen sie in den Uterus hinein und werden aus diesem durch die
Kloake hindurch einzeln abgesetzt. Die Eier sind von runder oder ovaler
Gestalt und schwanken in der Länge ihrer grössten Achse etwa zwischen
0.10 bis 0.14 mm. Untersucht man eine grössere Anzahl solcher
Eier, so wird man einzelne von besonderer Kleinheit antreffen (0.07 bis
0.11 mm), die im Innern einen sehr auffälligen tiefschwarzen
Fleck aufweisen. Der Breslauer Botaniker Cohn hat zuerst
gezeigt, dass sich aus derartigen Eiern stets Männchen entwickeln. Die
genannten zwei Eisorten sind nun nicht die einzigen, welche unserer
Hydatina zukommen. Da das Tierchen, wie angegeben wurde, mit Vorliebe
sich in ganz seichten Lachen und Gräben aufhält, welche oft schon nach
wenigen Tagen eintrocknen, so würde dasselbe in solchen Fällen stets zu
Grunde gehen, wenn nicht die Natur ein Schutzmittel in der Erzeugung
besonders widerstandsfähiger „Dauereier“ geschaffen hätte; denn die
Hydatina besitzt nicht das von den Erdrotatorien eingangs geschilderte
Eintrocknungsvermögen. Sobald das Tier dem feuchten Elemente entrückt
wird, geht es rettungslos zu Grunde. Jene Dauereier hingegen können
unbeschadet ihrer Lebenskraft eintrocknen. Sie unterscheiden sich von
den gewöhnlichen Eiern äusserlich nur dadurch, dass sie von einer
doppelten Hülle umgeben sind, von denen die äussere sehr derb und
mit einem dichten Haarbesatz versehen ist. Ob auch ein Unterschied
— etwa in der Grösse — zwischen männlichen[S. 291] und weiblichen
Dauereiern existiert, oder ob überhaupt nur letztere vorkommen, was
wahrscheinlicher ist, ist zurzeit noch nicht entschieden. Alle drei
Eisorten vermögen sich nun auf parthenogenetischem Wege zu entwickeln
und zwar die dünnschaligen sehr rasch, ungefähr in 24 Stunden, während
die Entwickelungsdauer der hartschaligen sehr verschieden zu sein
scheint. Jedenfalls können diese wochenlang ohne Veränderung verharren
und bedürfen vielleicht unter Umständen erst einer Trockenperiode.
Früher bezeichnete man die hartschaligen Eier vielfach als
„Wintereier“, indem man annahm, dass sie im Herbste aufträten und die
Art über die rauhe Jahreszeit hinaus erhielten. Cohn stellte
ferner den Satz auf, dass dieselben nur nach erfolgter Begattung
gebildet werden könnten. Beide Ansichten sind nicht richtig. Die
Dauereier werden den ganzen Sommer hindurch gebildet und zwar auch von
Weibchen, die nie mit Männchen in Berührung gekommen sind. Sie dienen
auch nicht als Schutz gegen die Kälte, sondern gegen das Eintrocknen,
daher sie auch bei den Erdrotatorien, die in anderer Weise an das
Versiegen des Wassers angepasst sind, fehlen. Merkwürdigerweise werden
alle drei Eisorten je von besonderen Individuen erzeugt, wenigstens bin
ich auf Grund grösserer Versuchsreihen seinerzeit für die Hydatina
senta zu diesem Schlusse gedrängt worden. Die Mehrzahl aller
Hydatinen legt nur weichschalige weibliche „Sommereier“, und zwar
während ihrer zwei- bis dreiwöchigen Lebensdauer etwa fünfzig Stück;
einige wenige Tiere erzeugen ebenso ausschliesslich die kleinen
weichschaligen Eier, aus denen die Männchen schlüpfen. Das Gleiche gilt
von denjenigen Tieren, welche Dauereier ablegen, nur dass die Zahl
derselben eine viel geringere ist, indem täglich nur 1–5 derselben
abgesetzt werden[IX].
[IX] Nach den neuesten (Comptes rendus, t. CXI. 1890)
Untersuchungen von Maupas, die mir während der Korrektur d. Bog.
bekannt werden, sind die Fortpflanzungsverhältnisse der Hydatina
noch etwas anders. Es finden sich zwei Sorten von ♀: 1) nicht
befruchtungsfähige, die ausnahmslos weibliche parthenogenetische
Eier erzeugen; 2) befruchtungsfähige, die im Falle einer Befruchtung
Dauereier absetzen, bei mangelnder Begattung männliche Sommereier
liefern. Nicht jede Begattung führt zu einer Befruchtung, sondern nur
dann, wenn sie in den ersten acht Stunden nach dem Verlassen des Eies
an einem ♀ vollzogen wird. ♀, die mit der Eierablage begonnen haben,
sind überhaupt nicht mehr einer Befruchtung zugängig, daher man unter
diesen drei verschiedene Arten von Individuen, den drei Eisorten
entsprechend, unterscheiden kann.
[S. 292]
Das Nervensystem der Hydatina besitzt als Zentralorgan ein
über und etwas vor dem Schlundkopf gelegenes Ganglienpaar, das als
„Gehirn“ bezeichnet wird. Betrachtet man dasselbe vom Rücken aus, so
hat es annähernd den Umriss eines Parallelogramms, dessen Längsachse
quergestellt ist. Von der Seite gesehen erscheint es kegelförmig nach
hinten verjüngt. Nach Anwendung von Reagentien lassen sich zahlreiche
in den peripherischen Schichten liegende Kerne erkennen; seine
Struktur ist im einzelnen noch nicht untersucht worden. Von dem Gehirn
ziehen jederseits nach vorn drei Nervenstränge, welche diejenigen
Matrix-Verdickungen versorgen, welche die grossen Borstenbüschel
tragen. Nach hinten setzt sich das Gehirn in einen Nerv fort, der
schräg nach hinten und oben zur Rückenwand zieht und hier ein für die
Rotatorien sehr charakteristisches Sinnesorgan, den „dorsalen Taster“,
bildet. Dicht unter der Haut schwillt nämlich der Nerv zu einem
kleinen, aus nur wenigen bipolaren Ganglien-Zellen gebildeten Ganglion
an, das an eine kreisrunde Öffnung in der Kutikula sich ansetzt. Aus
dieser Öffnung strahlt ein Büschel langer zarter Borsten hervor, welche
offenbar sehr empfindlich sind, den bei Berührung empfundenen Reiz zum
Gehirn weiterleiten und hier zum Bewusstsein kommen lassen. Ein ganz
ähnlich gebautes Sinnesorgan findet sich jederseits ungefähr in der
Mitte des Körpers (lt), doch ist dasselbe wegen seines zarten,
durchsichtigen Baues nicht eben leicht zu finden und bis vor kurzem
von allen Beobachtern übersehen worden. Die „lateralen Taster“ laufen
nach vorn in einen Nerv fort, der sicherlich direkt oder indirekt mit
dem Gehirn in Zusammenhang steht, wenn es auch bis jetzt noch nicht hat
glücken wollen, denselben nachzuweisen.
Auf eine eingehende Darstellung des Muskelsystems der Hydatina
will ich hier verzichten. Es sei nur bemerkt, dass die Muskelfasern
sämtlich glatt sind und unter der Haut liegen, an die sie sich
mit beiden Enden anheften. Der Richtung nach kann man Quer- und
Längsmuskeln unterscheiden. Die ersteren umziehen reifenförmig in
ziemlich gleichen Abständen den Körper und bewirken am lebenden Tier,
wenn sie sich kontrahieren, häufig[S. 293] den Anschein einer segmentalen
Gliederung. Zu den Längsmuskeln rechne ich auch diejenigen Bänder,
welche in etwas schräger Richtung durch die Leibeshöhle ziehen.
Sie bewirken vornehmlich die so häufige und für die Untersuchung
so störende Einstülpung von Kopf und Schwanz in den Mittelkörper.
— Besondere Respirationsorgane fehlen der Hydatina ebensosehr,
wie Blutgefässe. Die Atmung findet offenbar durch die ganze Haut
statt. Die die Leibeshöhle erfüllende Flüssigkeit ist wasserklar,
ohne irgendwelche an Blutkörperchen erinnernde Zellen. Durch die
Wirkung der Muskeln wird sie beständig in der Körperhöhle hin und her
getrieben und kann daher die von den inneren Organen ausgeschiedene
Kohlensäure leicht der Hautfläche zuführen und so den Gasaustausch
vermitteln. Sie dient auch als Antagonist der Längsmuskeln, indem die
eingestülpten Körperpole durch sie unter gleichzeitiger Wirkung der
Ringmuskeln wieder hervorgepresst werden. — Endlich seien hier noch
die zahlreichen zarten Bindegewebsfäden erwähnt, welche sich
zwischen den einzelnen Organen ausspannen und diese in ihrer Lage zu
einander und zur Haut erhalten.
Fig. 63. Hydatina senta ♂. Nach Weber. Seitenansicht.
Schon oben habe ich kurz des eigenartigen
Geschlechts-Dimorphismus gedacht, welcher unserer Hydatina
zukommt. Die früher herrschende Ansicht, derzufolge die Männchen nur
im Herbste auftreten, um die Produktion von Dauereiern hervorzurufen,
ist nicht richtig. Sie finden sich zu allen Jahreszeiten; da sie
aber an Individuenzahl hinter den Weibchen so ausserordentlich
zurückstehen, so wird man ihrer nur dann habhaft werden, wenn
man über grosse Mengen von Hydatinen verfügt. Am zweckmässigsten
ist es in diesem Falle, etwa hundert Weibchen, einzeln oder in
beschränkter Anzahl, in Uhrschälchen zu isolieren und die abgelegten
Eier zu kontrollieren. Bemerkt man unter diesen solche von geringer
Grösse und mit einem schwarzen, wahrscheinlich aus fäkalartigen,
nicht mehr verwertbaren Massen gebildeten Fleck, so hält es nicht
schwer, das zugehörige Muttertier zu ermitteln und sich durch dieses
fortlaufend mit männlichen Individuen versorgen zu lassen. Für die
Männchen ist zweierlei charakteristisch: einmal die geringe
Grösse im Vergleich mit den Weibchen und zweitens[S. 294] die Reduktion
der Organisationshöhe, welche sich vornehmlich in dem fast völligen
Schwunde des Verdauungskanals ausspricht. Es sind äusserst flinke,
kleine Tierchen, die nur ungefähr halb so lang als die Weibchen sind
und blitzschnell im Wasser umherschwimmen. Die Körpergestalt (Fig. 63)
ist ungefähr die gleiche wie bei den Weibchen, nur ist der Kopftrichter
weit flacher und stellt eine seichte Mulde dar, was wohl mit dem
Schwunde der Mundöffnung zusammenhängt. Am Räderapparat erkennt man
den äussern Cilienkranz und die fünf mit stärkeren Cilien besetzten
dorsalen Polster. Das einzige Organ, welches in der Leibeshöhle durch
seine Grösse auffällt, ist der birnförmige Hoden (t),
der durch einen kurzen, mit starker Ringmuskulatur ausgezeichneten
Ausführgang an der der weiblichen Kloakenöffnung entsprechenden Stelle
ausmündet. Er kann bei der Begattung vorgestülpt werden und darf daher
als Penis bezeichnet werden. Im Innern des Hodens bemerkt man bei
geschlechtsreifen Tieren ein siedendes Gewimmel kleiner Samenfäden,
die bei Anwendung sehr starker Objektive einen angeschwollenen Kopf
und einen schwanzartigen Anhangsfaden mit undulierendem Saume erkennen
lassen. An der Wurzel des Ausführganges zeigt die Hodenblase eine
feine Längsstreifung; dieselbe liegt nicht in der Wandung, sondern
wird durch kleine nadelförmige Gebilde hervorgerufen, die bewegungslos
sind und deshalb kaum als Samenfäden gedeutet werden können. Der
Verdauungskanal (sto) hat sich nur in ganz rudimentärer
Form erhalten; er ist bandförmig, ohne Lumen und erstreckt sich von
der Rückenfläche des Hodens bis zur Kopfmulde. Die Berechtigung,
dieses Gebilde als[S. 295] rückgebildeten Darm aufzufassen, ergiebt sich
einmal aus seiner Lage im Körper und dann daraus, dass in ihm die
oben erwähnten schwarzen Körnchen liegen, welche bei den Weibchen
verschiedener anderer Rotatorien im Darm angetroffen werden. Auch das
Exkretionssystem der Männchen ist einfacher als bei den Weibchen; eine
pulsierende, für beide Nephridien gemeinschaftliche Blase fehlt, so
dass dieselben getrennt neben der Penisöffnung ausmünden. Im übrigen
sind die Verhältnisse hier, wie auch im Nerven-, Muskel- und Hautsystem
dieselben wie bei den Weibchen. Da die Männchen nicht imstande sind,
Nahrung aufzunehmen, so ist ihre Lebensdauer eine sehr kurze; ich
habe sie in der feuchten Kammer nur 2–3 Tage zu halten vermocht. Sehr
eigenartige Verhältnisse scheinen bei der Begattung vorzuliegen. Man
trifft nicht zu selten weibliche Tiere an, die in ihrer Leibeshöhle
lebende Samenfäden aufweisen, während man dieselben doch höchstens
im Uterus erwarten sollte. Bringt man nun mehrere (5–6) Männchen mit
einem Weibchen in einem kleinen Wassertropfen zusammen, so nehmen
die Tierchen anfangs, auch bei gegenseitiger Berührung, keine Notiz
von einander. Nach einiger Zeit ändert sich das Bild. Die Männchen
beginnen das Weibchen zu umschwärmen und gelegentlich sieht man ein,
event. auch mehrere Männchen sich dem Weibchen mit vorgestülptem Penis
an irgend einer Körperstelle, aber nicht an der Geschlechtsöffnung,
anheften. Wird nun ein solches Weibchen sofort genauer untersucht, so
finden sich Ballen von Spermatozoen der Innenseite der Haut an eben
jenen Stellen angeheftet, während einige Samenfäden sich schon in der
Leibeshöhle umhertummeln. Es kann daher nicht zweifelhaft sein, dass
es hier zu einer Begattung gekommen ist, wenn es zurzeit auch noch
nicht möglich ist, zu entscheiden, ob diese Samenfäden sich später
in das Ovar einbohren und hier eine Befruchtung herbeiführen; da ich
nach 24 Stunden stets nur abgestorbene Samenkörper in begatteten
Weibchen antraf, so scheint es mir wahrscheinlicher, dass es in solchen
Fällen überhaupt nicht zu einer Befruchtung kommt, der ganze Vorgang
daher eigentlich überflüssig ist, und dass eine solche wohl nur dann
eintritt, wenn es[S. 296] dem Männchen gelingt, die Kloakenöffnung des
Weibchens aufzufinden[X].
[X] Diese Auffassung wird natürlich nach den neuesten Angaben
von Maupas (cf. S. 291) hinfällig.
Nachdem wir im vorstehenden Bau und Biologie eines typischen Rotators
kennen gelernt haben, werden wir verhältnismässig leicht uns ein Bild
der in der ganzen Klasse vorkommenden Organisationsverschiedenheiten
entwerfen können, denn die Rotatorien sind im Vergleich mit anderen
Abteilungen des Tierreiches sehr einförmig zu nennen, und ihre
morphologische Differenzierung schwankt nur innerhalb enggezogener
Grenzen.
II. Vergleichende Schilderung der
Morphologie der Rotatorien[XI].
1. Die Körpergestalt.
Alle Rädertiere sind ungegliedert und bilateral-symmetrisch, d. h. sie
lassen sich nur durch eine in die Längsachse des Tieres gelegte Ebene
in zwei symmetrische Körperhälften — eine linke und eine rechte —
teilen, und dementsprechend kann man an ihnen ein vorderes und ein
hinteres Körperende und eine Bauch- und eine Rückenseite unterscheiden.
Von allen Körperachsen ist diejenige, welche vom vordern zum hintern
Körperpole geht, fast ausnahmslos die längste, und da gleichzeitig die
Breite des Tieres im Verhältnis zur Länge in der Regel sehr gering
ist, so haben die meisten Rotatorien einen gestreckten, wurmartigen
Habitus, dessen Länge zwischen 3 bis 0.05 mm schwankt. Der
Querschnitt durch die Mitte des Körpers zeigt entweder einen runden
bis ovalen Umriss, oder die Dorsoventral-Achse ist nur sehr klein,
und das Tier (z. B. ein Brachionus, Fig. 69 S. 300, oder eine
Polyarthra, Fig. 66 S. 298) erscheint dann stark abgeplattet,
scheibenförmig. Dass alle drei Körperachsen ungefähr gleich lang sind,
und der Körper daher die Gestalt einer Kugel besitzt, kommt nur[S. 297]
bei zwei, auch in der innern Organisation sehr abweichenden Arten
vor, nämlich erstens bei der von Semper auf den Philippinen
entdeckten Trochosphaera aequatorialis, bei der die Stirnfläche
des Kopfes halbkugelförmig aufgeblasen ist, sodass der Ciliensaum
des Räderapparates den Körper in äquatorialer Stellung umzieht, und
zweitens bei Apsilus lentiformis, einer seltenen deutschen
(bei Giessen beobachteten) Art. Annähernder Gleichheit in der Grösse
der Quer- und Längsachsen begegnen wir schon häufiger, z. B. bei den
scheibenförmigen Gattungen Pterodina und Pompholyx. Bei
fast allen Rotatorien ist das vordere Körperende quer abgestutzt,
und die so entstandene „Stirnfläche“ trägt den Räderapparat und, der
Ventralseite genähert, die Mundöffnung. Die Breite des Kopfes ist
bald dieselbe wie in den mittleren Körperregionen, bald wird sie
etwas geringer als diese, so dass sich der Körper nach vorn leicht
verjüngt. Bei besonders mächtiger Entwickelung des Räderapparates (s.
weiter unten) kann der Stirnfläche auch die Maximalbreite des Körpers
zukommen. Hinter der Afteröffnung verjüngt sich der Körper fast aller
Rädertiere und bildet so den zur dauernden oder vorübergehenden
Anheftung dienenden Schwanz (Fig. 62, 63, 64, 65, 69). Dieser Schwanz
kann in sehr verschieden deutlichem Grade sich vom Mittelkörper
absetzen. Bei den in[S. 298]Fig. 62–65 abgebildeten Arten gehen beide ganz
allmählich in einander über. Bei der Asplanchnopus myrmeleo
(Fig. 68) sitzt der kurze in zwei Zehen auslaufende Schwanz nicht am
Hinterende des Rumpfes, sondern fügt sich der Bauchseite ein gutes
Stück weiter nach vorn an. Fig. 69 zeigt uns einen Brachionus,
bei dem der schmale Schwanz fast die halbe Körperlänge erreicht,
von dem viel breitern Mittelkörper sehr scharf abgesetzt ist und
durch besondere Muskeln vorübergehend ganz in diesen zurückgezogen
werden kann. Endlich giebt es eine Anzahl Formen, denen ein echter
Schwanzanhang überhaupt fehlt; so die Gattungen Polyarthra
(Fig. 66), Triarthra, Hertwigia, Asplanchna,
Anuraea (Fig. 67), Sacculus. Bei diesen mangeln mit dem
Schwanze gleichzeitig die Fuss- oder Klebdrüsen, sodass die Tiere
nicht[S. 299] im stande sind, sich festzuheften. — Die Fussdrüsen (f)
sehr vieler Rädertiere münden an der Spitze zweier (selten einer:
Monostyla) sehr verschieden langer dolchförmiger Zehen, die
bald an einander gelegt, bald gespreizt getragen werden. Bei den im
erwachsenen Zustande dauernd festsitzenden Melicertiden, z. B. der
Gattung Floscularia (Fig. 65), sind zwar die Fussdrüsen mächtig
entwickelt, Zehen hingegen fehlen, und der Körper endet mit einer quer
abgestutzten kleinen Fläche. Anderseits giebt es eine Abteilung unter
den Rädertieren, bei denen die Zehen stets in grösserer Zahl als zwei
auftreten. Es sind dies die Philodiniden, wozu der gemeine Rotifer
vulgaris und fast alle Erdrotatorien gehören. Bei diesen läuft
der Schwanz bald in drei (Rotifer, Actinurus), bald in
vier (Philodina[XII]) Zehen aus, die cylindrische fingerförmige
Anhänge darstellen. Vier kurze Zehen kommen auch der an den Beinen
unseres Bachflohkrebses lebenden Callidina parasitica Gigl. zu,
während bei anderen nahen Verwandten noch weitergehende Modifikationen
beobachtet werden. Bei Callidina symbiotica Zel. (Fig. 64)
endigt der Schwanz mit zehn sehr kleinen Zehen, und bei Call.
magna Plate münden die Ausfuhrgänge der Klebdrüsen in eine von
zahlreichen dichtstehenden Kanälen durchbrochene kreisförmige Platte
aus. Alle die genannten Gattungen der Philodiniden besitzen ausserdem
noch zwei „Afterzehen“ oder „Sporen“, welche kurz vor den eigentlichen
Zehen von der Rückenseite ausgehen und in der Gestalt diesen[S. 300] sehr
ähneln (Fig. 64, zh). Bei Call. parasitica fungieren sie
auch als echte Zehen, indem sie Ausführgänge der Klebdrüsen aufnehmen;
bei allen übrigen Arten hingegen enden sie blind geschlossen und sind
daher wohl ihrer ursprünglichen Funktion verlustig gegangen.
[XI] Dieselbe bezieht sich nur auf die Weibchen, da die
Morphologie der Männchen in einem besondern Abschnitte besprochen
werden soll.
[XII] Nach E. F. Weber, während Hudson-Gosse auch
für Philodina nur drei Zehen angeben.
Fig. 64. Callidina symbiotica. Nach Zelinka, etwas geändert.
Ventralansicht.
Fig. 65. Floscularia cornuta. Nach Leydig. Seitenansicht.Fig. 66. Polyarthra platyptera. Nach Leydig. Rückenansicht.Fig. 67. Anuraea aculeata. Nach Hudson-Gosse. Rückenansicht.
Die Fusszehen der Rotatorien können als Körperanhänge
angesehen werden, welche durch Ausstülpungen der Haut entstanden
sind. Ähnliche, aber nicht der Festheftung[S. 301] sondern der Bewegung
dienende Anhänge, die ihren Trägern häufig ein sehr absonderliches
Aussehen verleihen, treffen wir bei einigen Rädertieren an, die
sehr verschiedenen Unterabteilungen des Systems angehören. Ich
erwähne hier nur drei verschiedene Arten derartiger Ausstülpungen.
Bei den Philodiniden entspringt dicht hinter dem Räderapparat ein
langer, aus- und einstülpbarer „Rüssel“ (Fig. 64, rs), der
an seinem verjüngten Vorderende mit beweglichen Cilien besetzt
ist. Der Rüssel ist an seiner Basis fast so breit wie der Körper,
sodass man darüber im Zweifel sein kann, ob man ihn als eine aus der
Rückenfläche hervorgewachsene Neubildung oder nicht vielmehr als
das ursprüngliche Vorderende des Körpers anzusehen hat, in welchem
Falle der Cilienapparat zum grössten Teile auf die Bauchfläche
übergetreten wäre und sich mit der Mundöffnung ein gutes Stück
nach hinten verschoben hätte. Rüssel und Räderapparat sind bei den
Philodiniden nie gleichzeitig in Thätigkeit, sondern in dem Moment,
wo ersterer sich hervorstülpt, wird letzterer eingezogen. Der Rüssel
dient teils als Träger von Sinnesorganen (Tastborsten, Augenflecke),
teils vermittelt er mit den Fusszehen die spannerraupenartige
Bewegungsweise der Philodiniden, wobei seine vordere Wimperplatte
wie eine Saugscheibe der Unterlage angeheftet wird. — Unter den
Arten der Gattung Asplanchna zeichnen sich einige durch den
Besitz von 2–4 kegelförmigen Hautauswüchsen aus, die vom Rücken oder
den Körperseiten entspringen und zumteil eine beträchtliche Höhe
erreichen. Bald treten dieselben nur bei den Männchen auf (so bei
Aspl. Sieboldii Leyd.[XIII] in Vier-, bei Aspl. intermedia
Huds. in Zweizahl), bald sind beide Geschlechter, wenn auch zuweilen
in verschiedener Weise, mit ihnen ausgerüstet (Aspl. amphora
Huds. und Aspl. Ebbesbornii Huds.). Eine besondere Funktion
scheint ihnen nicht zuzukommen. — Ein besonderes Interesse
beanspruchen die bei den Gattungen Polyarthra, Triarthra,
Pedetes und Pedalion[S. 302] vorkommenden Anhänge, da sie
auffallend an die Extremitäten der Entomostraken erinnern. Wie diese
sind sie scharf, zumteil sogar gelenkig, vom Körper abgesetzt, sind
beweglich, tragen nicht selten gefiederte Borsten und dienen der
gleichen Funktion, nämlich zum Rudern. Dennoch sind dieselben nur als
analoge Bildungen anzusehen, denn die zwei wichtigsten Merkmale der
Entomostraken-Gliedmassen, die paarweise und ausschliesslich ventrale
Gruppierung und der Spaltfusscharakter, gehen ihnen völlig ab.
[XIII] Neuerdings hat E. v. Daday die sehr interessante
Beobachtung gemacht, dass bei Aspl. Sieboldii zwei verschiedene
Weibchen vorkommen, erstens männlich-geformte mit vier konischen
Hautanhängen und zweitens solche ohne diese (Math. u. Naturw. Berichte
aus Ungarn. VII. 1889).
Dass nun der Habitus der Rädertiere im einzelnen so vielen kleinen
Veränderungen unterworfen ist, wird vornehmlich durch zwei Organsysteme
bewirkt, nämlich einmal durch die Vielgestaltigkeit des Räderapparates
und dann durch den wechselnden Grad von Festigkeit, welcher der Haut
eigen ist. Wir beginnen daher die vergleichende Betrachtung der
Organsysteme mit diesen.
2. Die Körperhaut.
Wenn ich die Rädertiere oben als ungegliedert bezeichnete, so
scheint die Beschaffenheit der Körperdecke vieler Arten sich hiermit
schwer in Einklang bringen zu lassen. Fanden wir doch schon bei der
Hydatina und ebenso bei Callidina symbiotica (Fig.
64), wie der Körper durch mehrere in ziemlich gleichen Abständen
aufeinanderfolgende Einschnürungen, ähnlich einem Ringel- oder
Bandwurme, in Segmente gegliedert wurde. In der That lässt die Haut
vieler Rotatorien eine Zusammensetzung aus mehreren gleichen Zonen
erkennen, aber da wir bei keinem innern Organe etwas Ähnliches
antreffen, sondern alle nur in Ein- oder Zweizahl vorhanden sind, kann
von einem metameren Körperbaue nicht die Rede sein, sondern höchstens
von einer auf die Haut und die zugehörigen Muskeln beschränkten
Scheinsegmentierung. Diese kommt entweder nur durch die Anordnung
der Ringmuskeln zu stande (Hydatina), oder Hand in Hand mit
dieser geht eine Zusammensetzung der Haut aus abwechselnd derben
und weichen Partien. Die ersteren bilden bei den Philodiniden die
eigentlichen Hautsegmente, die letzteren die je zwei Segmente von
einander sondernden Furchen, welche in Gestalt von Ringfalten sich
unter[S. 303] die Haut des nächsthintern Scheinsegmentes legen. In der
Dicke verhält sich die Kutikula der Falten ganz gleich derjenigen
der Segmente, in die sie ja auch kontinuierlich übergeht. Dass jene
Falten konstante Bildungen sind, lässt sich daher wohl nur aus einer
weicheren Beschaffenheit derselben erklären. Kontrahieren sich die
Längsmuskeln, so schieben sich die schmäleren Scheinsegmente in die
weiteren hinein, ähnlich wie man die Ringe des Tubus eines Fernrohres
in einander stecken kann. — Viel auffallender als bei den Philodiniden
ist der Unterschied zwischen weichhäutigen und derben Partien der
Kutikula bei den sogenannten „gepanzerten“ Rädertieren. Es gehören
hierher zahlreiche Gattungen, von denen manche, wie Brachionus,
Anuraea, Pterodina, Salpina, Colurus,
Metopidia, zu den gemeinsten Vertretern der Klasse zählen.
Der Panzer, d. h. der derbhäutige unelastische Abschnitt der Haut,
umfasst bei diesen Formen nur den Mittelkörper. Die Haut des Kopfes
und des Schwanzes (falls ein solcher überhaupt vorhanden ist) hingegen
bleibt von der gewöhnlichen Konsistenz. Im Panzer nimmt die Kutikula
eine grössere Dicke an und bedeckt sich mit den verschiedenartigsten
Skulpturen, wie Leisten, Rillen, Punkten, Höckern u. dgl., welche
häufig zu sehr zierlichen Zeichnungen zusammentreten. Unsere Abbildung
der Anuraea aculeata (Fig. 67) lässt z. B. die polygonalen
Felder der Rückenfläche des Panzers deutlich erkennen. Sehr häufig
läuft der Panzer dort, wo er in die weiche Haut des Kopfes und des
Schwanzes übergeht oder, obwohl seltener, an anderen Stellen in
zahnartige Fortsätze aus. So verdankt die eben erwähnte Anuraea
ihren Speziesnamen aculeata dem Umstande, dass der Panzer am
Vorderrande sechs, am Hinterrande zwei ansehnliche Dornen trägt. Bei
den verschiedenen Brachionus-Arten differiert vornehmlich die Zahl,
Grösse und Anordnung der Zähne am Vorderrande der Schale; ausser
diesen begegnen wir vielfach zwei kleinen Zähnen an der Basis des
Schwanzes (Fig. 69). Bei einigen Rädertieren erreichen die Stacheln
des Panzers eine solche Länge, dass ganz abenteuerliche Formen
hierdurch entstehen, Anuraea longispina Kellic. z. B. trägt vorn
drei, hinten einen spiessartigen Fortsatz von der Länge des[S. 304] ganzen
Panzers. Bei Stephanops tripus Lord und Steph. Leidigii
Zacharias sitzt der Rückenfläche ein nach hinten gerichteter und
bogenförmig gekrümmter Stachel auf, welcher bei ersterer etwas kürzer,
bei letzterer länger als das ganze Tier ist. — Der Panzer bewirkt
in einzelnen Fällen eine Abweichung von der typischen Lagerung der
inneren Organe. Ist er z. B. sehr flach gebaut (Pterodina,
Metopidia), so kommen die Geschlechtsorgane im entwickelten
Zustande teilweise neben den Darm, anstatt unter ihn, zu liegen. Ferner
hat sich die Öffnung der Kloake von der Dorsalseite auf die Bauchfläche
verschoben bei den Gattungen Pterodina, Anuraea und
Dinocharis, welch’ letztere auch dadurch bemerkenswert ist, dass
bei ihr der Hals panzerartig erhärtet ist, während sonst der Kopf stets
weichhäutig bleibt.
3. Der Räderapparat.
Von allen Organsystemen zeigt keines eine so grosse Vielgestaltigkeit
und einen so verschiedenen Ausbildungsgrad als dasjenige, welches
die Lokomotion und die Nahrungsaufnahme vermittelt, das Räderorgan.
Dennoch zeigt eine vergleichende Betrachtung, dass den meisten
Rotatorien eine gemeinsame Grundform in der Anordnung der Cilien
zukommt, die freilich nicht immer mit gleicher Deutlichkeit zu Tage
tritt. Dieselben stehen nämlich in zwei Kreisen, von denen der eine,
innere in vielen Fällen vor, der andere, äussere hinter der Mundöffnung
gelegen ist; der letztere setzt sich in der Regel in die Mundöffnung
selbst fort. In sehr prägnanter Weise treffen wir einen derartigen
doppelten Wimpersaum, einen praeoralen und einen postoralen, bei den
Gattungen Rotifer, Philodina, Callidina (Fig.
64), Pterodina, Melicerta, Lacinularia und
einigen anderen an. Hier ist auch eine Arbeitsteilung in der Funktion
der beiden Cilienkränze eingetreten. Der praeorale ist mit besonders
starken Cilien versehen und dient zur Fortbewegung des Tieres, während
die schwächeren Flimmern des postoralen Kranzes die Nahrungsteilchen in
den Mund strudeln. Die eigenartige Radbewegung, welche das Spiel der
praeoralen Wimpern dem Auge vortäuscht, ist, wie schon[S. 305] erwähnt wurde,
die Veranlassung zur Benennung der hierher gehörigen Organismen als
Rädertiere gewesen. Unsere Abbildung der Callidina symbiotica
(Fig. 64) zeigt, wie die Cilien dieses vordern Kranzes auf zwei durch
eine mediane Furche von einander getrennten Polstern, Ausstülpungen
der Stirnfläche, randständig angebracht und in der Mitte der Bauch-
und Rückenseite durch eine nackte Stelle unterbrochen sind. Man könnte
daher auch von zwei unvollständigen Cilienkreisen reden, wenn die
Raddrehung derselben nicht in gleichem Sinne dem Auge sich darböte. Der
hintere, postorale Kranz besteht nicht aus einer einfachen Cilienreihe,
sondern stellt einen Flimmerstreifen dar, welcher jederseits aus
der ventralwärts stark lippenförmig vorspringenden Mundöffnung
hervortritt und um jene Stirnpolster herum sich dem Rücken zuwendet.
Eine Verschmelzung in der dorsalen Mediane findet aber auch bei diesem
Kranze nicht statt. — Die Umbildung der beiden für die genannten
Gattungen charakteristischen primitiven Wimpersäume erfolgte bei den
übrigen Rädertieren nach zwei Richtungen hin, jenachdem der postorale,
äussere oder der praeorale, innere Cilienkranz der mächtigere wurde
und besondere Differenzierungen einging. Der erstere Fall ist nur
selten, nämlich bei den festsitzenden Gattungen Floscularia
(Fig. 65) und Stephanoceros, eingetreten. Bei beiden hat sich
die Stirnfläche kelchförmig eingesenkt und trägt im Grunde die von
kleinen Cilien des inneren Kranzes umstellte Mundöffnung. Der Kelchrand
trägt die sehr langen [bei einzelnen Arten (Floscularia mira
Huds.) geradezu enorm grossen] Cilien des äusseren Kranzes und ist in
fünf Anhänge ausgezogen, auf die sich die Wimpern auch fortsetzen. Bei
Floscularia sind es kurze, stumpfkegelförmige Zipfel, bei dem
schönen Stephanoceros Eichhornii hingegen lange, zungenförmige
Fortsätze. — Bei der weitaus grössten Zahl der Rotatorien erleidet
hingegen der innere Cilienkranz mannigfache Umgestaltungen: aus der
ursprünglich einfachen Cilienreihe werden mehrere; einzelne erheben
sich auf besonderen Polstern und nehmen dann die Gestalt derber Griffel
an; andere verlieren ihre Beweglichkeit und werden zu starren langen
Tastborsten; die ursprüngliche ringförmige Anordnung wird verwischt,
indem der[S. 306] Zusammenhang der Cilien durch nackte Partien unterbrochen
wird; es treten sehr feine Cilien sekundär an ursprünglich nackten
Stellen der Stirnfläche auf, und so fort. Auf alle diese zahlreichen
Modifikationen hier näher einzugehen, ist unmöglich, und seien daher
nur noch zwei Abweichungen erwähnt. Bei Besprechung der Körpergestalt
der Philodiniden wurde schon hervorgehoben, dass die Spitze des
dorsalen „Rüssels“ mit einer kleinen Flimmerplatte besetzt ist, also
mit einer Art sekundären Räderorgans (vgl. Fig. 64). Wie dieselbe zu
deuten ist, ob als Bildung sui generis oder als abgespaltener Teil
des ursprünglich einheitlichen Cilienapparates, lässt sich schwer
entscheiden. Letzteres scheint die grössere Wahrscheinlichkeit für sich
zu haben, indem der Dorsalrand der Wimperscheibe sich in verschiedenen
Gattungen in einen zungenförmigen Anhang auszieht, der dann entweder
(Rhinops vitrea) noch vom äusseren Wimpersaum umzogen wird
oder nur einige Tastborsten trägt (Adineta vaga) oder endlich
ganz nackt geworden ist (Metopidia, Stephanops).
Abspaltungen seitlicher Partien des Räderapparates werden auch sonst
beobachtet, so bei der Gattung Synchaeta, die jederseits ein
„Wimperohr“ am Kopfe trägt. — Die zweite hier zu erwähnende Abweichung
betrifft Formen, deren Cilienapparat eine ventrale und vorn am Körper
gelegene Scheibe darstellt (Notommata tardigrada Leyd.,
Adineta vaga Dav., Diglena forcipata Ehr., Digl.
giraffa Gosse), die dicht mit kleinen Wimpern besetzt ist. Von
einer ursprünglich ringförmigen Anordnung der Cilien haben sich keine
Andeutungen erhalten, und die ganze Bildung erinnert ausserordentlich
an die Gastrotrichen, eine kleine Gruppe mikroskopischer
Süsswassertierchen, die ungefähr die Gestalt der Rotatorien haben, aber
auf der ganzen Bauchseite bewimpert sind. — Der Räderapparat fehlt
nur bei wenigen Gattungen, die auch sonst in ihrer Organisation sehr
abweichen, völlig: so bei dem an der Unterseite von Nymphaeablättern
festgehefteten Apsilus lentiformis Metschn., bei dem
Regenwurmparasiten Balatro clavus Clap. und bei dem marinen, auf
Nebalia schmarotzenden Paraseison nudus Plate. Bei manchen der
kleineren Arten scheint er stark reduziert zu sein, ohne dass jedoch
unsere Instrumente zurzeit[S. 307] für ein eingehendes Studium derselben
ausreichten. — Der Cilienapparat kann bei allen Rotatorien mit
Ausnahme der Gattung Adineta vorübergehend in den Mittelkörper
eingestülpt werden. In diesem Zustande ruhen die Cilien, beginnen aber
mit dem Moment der Ausstülpung, offenbar unwillkürlich, wieder ihre
Thätigkeit. Soll bei den Philodiniden der Räderapparat benutzt werden,
so muss der „Rüssel“ eingestülpt im Körper liegen. Anderseits wird
jener sofort eingezogen, wenn das Tier beginnt sich spannerraupenartig
unter abwechselnder Festheftung und Loslösung der Rüsselspitze und der
Zehen fortzubewegen. Man erblickt daher nur ausnahmsweise die beiden
Cilienregionen gleichzeitig in Thätigkeit.
4. Die Muskulatur
der Rädertiere ist noch zu wenig erforscht, um eine vergleichende
Schilderung zu ermöglichen. Bei vielen, vielleicht allen Rotatorien
giebt es in der Mitte des Rumpfes eine Querlinie, von der die grossen
nach vorn und nach hinten laufenden Längsmuskeln ausstrahlen. Neben den
glatten Muskeln, welche weitaus die Mehrzahl bilden, kommen bei einigen
Arten auch quergestreifte vor; solche sind z. B. die Kopfretraktoren
von Pterodina und Polyarthra (Fig. 66, mu). Die
Ringmuskeln verteilen sich gewöhnlich in ziemlich gleichen Abständen
auf den ganzen Körper; nur bei Asplanchna myrmeleo liegen sie
am Halse dicht nebeneinander, hier auf schmalem Querringe einen echten
Hautmuskelschlauch bildend. Bei den Philodiniden ist jeder Ringmuskel
aus einer bestimmten Zahl kleiner Segmente zusammengesetzt.
5. Das Nervensystem
besteht überall aus einem grossen, dorsal vom Schlundkopfe gelegenen
Zentralorgan, dem Gehirn, dessen beide Ganglien breit mit einander
verwachsen sind. Die Gestalt ist bei verschiedenen Arten wechselnd:
rundlich, parallelogrammförmig, bei den Philodiniden dreieckig. Bei
letzteren ist eine Zusammensetzung aus peripherischen Ganglienzellen
und zentraler Fasermasse nachgewiesen, und daher eine solche auch für
die übrigen Arten sehr wahrscheinlich.[S. 308] Vom Gehirn strahlen Nerven
aus einmal an die zwischen den lokomotorischen Cilien oder auf der
Wimperscheibe stehenden Tastborsten, die „Stirntaster“ (t,
Fig. 66, 69), dann nach hinten und oben an den (resp. die) dorsalen
Taster und endlich nach hinten in den Mittelkörper. Wie die Zahl der
Stirntaster, so schwankt auch die Zahl der sie versorgenden Nerven,
über die übrigens nur für wenige Arten Untersuchungen vorliegen. Bei
den Philodiniden laufen die Cerebralnerven nach vorn teils in den
Rüssel hinein, dessen Spitze mit Tastbüscheln versehen ist, teils
versorgen sie das Räderorgan und den Mund. Die nach hinten in den Rumpf
tretenden Nerven scheinen bei den einzelnen Arten der Philodiniden
sich ziemlich verschieden zu verhalten. Bei Callidina magna
fand ich nur einen starken Nerv jederseits, von dem Seitenzweige
an die Haut ausstrahlten. Zelinka hingegen beschreibt von
Call. symbiotica einen ventralen und einen seitlichen Nerv und
bei Discopus synaptae sogar noch einen dritten, dorsalen, die
aber bei dieser Art alle drei nicht direkt, sondern durch besondere
Ganglienzellen mit dem Gehirn zusammenhängen. — Die Stirntaster
der Rädertiere treten bald als einzelne Borsten, bald als Büschel
von solchen auf, während die Rücken- und Seitentaster stets nur in
letzterer Form angetroffen werden. Bei den meisten Gattungen ist
der Rückentaster unpaar, wird aber sehr häufig von einem doppelten,
erst an der Basis des Tasters verschmelzenden Nerv versorgt, was
vielleicht auf eine ursprüngliche Duplicität des Organs hinweist. Die
Arten mit paarigem Rückentaster (Asplanchna, Hertwigia,
Apsilus) würden dann, obwohl im sonstigen Bau wenig typisch,
hierin ein primitives Verhalten bewahrt haben. Bei einigen Arten
erhebt sich der dorsale Taster in auffälligem Masse über die
Rückenfläche, der er gewöhnlich direkt aufsitzt. So treffen wir ihn
bei Lacinularia in Gestalt einer kleinen Doppelpapille, bei
Brachionus (Fig. 69, dt) und Anuraea als kurzen
Kegel, endlich bei den Philodiniden als stabförmigen retraktilen
Tentakel an. In der Regel sitzt der dorsale Taster im Nacken und rückt
nur selten, z. B. bei Asplanchna, weiter nach hinten, sodass
man ihn zunächst, da er paarig ist, mit den Seitentastern verwechseln
kann. Die Dorsal- und[S. 309] Lateraltaster sind für die Rädertiere in hohem
Masse charakteristisch, da sie fast konstant angetroffen werden.
Ersterer fehlt meines Wissens nur bei Conochilus, letztere nur
den Philodiniden und den marinen Seisoniden. Über den Zusammenhang
des zum Lateraltaster gehörigen Nervs mit dem Gehirn liegen zurzeit
noch keine sicheren Mitteilungen vor, da derselbe wahrscheinlich
durch einen im Kopfe liegenden Nervenplexus bewirkt wird. In ihrer
Lage am Körper schwanken die Seitentaster insofern, als sie bald
weiter nach vorn, bald mehr nach hinten die Kutikula durchbrechen.
Die meisten liegen ungefähr am Beginn des hinteren Körperdrittels.
Bei Polyarthra finden sie sich dagegen fast am Hinterrande des
Körpers, bei Asplanchna und Hydatina in der Mitte, und
bei Lacinularia und Melicerta — bei letzterer sind
sie, ebenso wie bei Copeus spicatus, zu zwei langen Tentakeln
ausgezogen — ungefähr in Schlundhöhe. In allen diesen Gattungen stehen
sie hinter dem dorsalen Taster, und nur bei Pterodina liegen
alle drei Sinnesorgane auf einer Querlinie.
Von anderweitigen Sinnesorganen kommen nur Augenflecke den
Rädertieren häufig zu. Sie sind meist unpaar und sitzen als rötlicher
oder schwärzlicher Pigmentfleck der Unterseite des Gehirns an.
In anderen Fällen sind sie paarig und gehören dann dem Rande der
Wimperscheibe an. Ihr Bau ist ausserordentlich einfach, häufig nur
ein Pigmentfleck, dem bei einzelnen Arten noch ein lichtbrechender
weisslicher Körper, die sogenannte Linse, sich hinzugesellt.
6. Der Verdauungskanal
zeigt eigentlich bei allen Rotatorien dieselben Verhältnisse, und
nur einem Abschnitte desselben, dem Kauapparat, kommt eine grössere
morphologische Variabilität zu. Die Mundöffnung liegt nur bei
Floscularia und Stephanoceros genau am Vorderende
der Längsachse des Körpers, sonst ist sie stets ventralwärts
verschoben und kann in einzelnen Fällen (Adineta vaga)
sogar ein beträchtliches Stück nach hinten verlagert sein. Ein
besonderes Mundrohr, wie wir es für Hydatina kennen
lernten, braucht nicht immer vorhanden zu sein, sondern nicht selten
(Eosphora, Diglena) schliesst[S. 310] sich der Mastax direkt
an die Eingangsöffnung an und kann aus dieser zum Ergreifen der Beute
etwas hervorgestossen werden. Anderseits erreicht das Mundrohr bei
Floscularia und Stephanoceros eine ungewöhnliche Länge
und erweitert sich vor dem Zahnapparat zu einem besonderen kropfartigen
Abschnitt. Vielleicht ist der grosse sackartige Raum, in dem bei
Asplanchna die Kiefer liegen, ebenfalls als eine Erweiterung des
Mundrohres anzusehen. — Die Struktur des Kauapparates ist für
die Systematik von hoher Bedeutung. Nach dem Vorgange von Gosse
kann man an demselben meist folgende Zusammensetzung beobachten: an
einen mittleren, nach hinten gerichteten unpaaren Abschnitt, den
„Incus“, heften sich die vorn und seitlich gelegenen paarigen „Mallei“,
von denen jeder aus zwei gelenkig mit einander verbundenen Stücken
besteht. Das eine von diesen, der Uncus, liegt medianwärts und stellt
den eigentlichen Kauapparat dar; er kann in der verschiedensten Gestalt
auftreten: als Träger quergestellter Leisten, als spitzer oder gesägter
Zahn u. dgl. Das andere Stück sitzt dem Uncus aussen an und dient als
Stützbalken für denselben; es wird als Manubrium bezeichnet. Bei nicht
wenigen Rädertieren scheint übrigens der Kauapparat weit einfacher
gebaut zu sein. Bei den Asplanchnen z. B. (Fig. 68) lässt sich nur der
mediane Incus und der paarige gebogene Zahn des Uncus unterscheiden;
ein Manubrium fehlt hier vollständig. Noch reduzierter tritt uns der
Mastax der Philodiniden entgegen; er besteht im wesentlichen aus zwei
halbkreisförmigen, mit derben Querleisten in wechselnder Zahl bedeckten
Chitinplatten, die nur längs des Durchmessers eng aneinanderschliessen,
und sich um diesen als Gelenkachse gegen einander bewegen. Die den
Zahnapparat umhüllende Fleischmasse ist teils muskulöser Natur, teils
scheint sie als Speicheldrüsen zu fungieren. — Der Schlund
ist meist kurz, nur bei Synchaeta und Asplanchna (Fig.
68, oe) von ansehnlicher Länge. Flimmerzellen werden in ihm
sicherlich nicht immer angetroffen, denn gerade die genannten Gattungen
schieben die aufgenommene Nahrung durch eine Art peristaltischer
Bewegung nach hinten. — Im Mitteldarm oder Magen sind Cilien
hingegen stets vorhanden. Mit Ausnahme der Philodiniden wird die
Wand[S. 311] desselben stets aus grossen polygonalen Zellen gebildet,
die in einer Schicht liegen. Bei jener Abteilung hingegen ist die
Magenwand ungewöhnlich dick und besteht aus einer kontinuierlichen
Protoplasmamasse mit zahlreichen eingestreuten kleinen Kernen. In den
Magenzellen der Rotatorien tritt nicht selten ein braunes Pigment auf,
das aber immer nur vorübergehend sich zeigt und bei Nahrungsmangel
sofort verschwindet. Dass dasselbe als Leber funktioniert, ist
daher sehr unwahrscheinlich. Näher liegt es, den zwei konstant am
Vorderende des Magens einmündenden Drüsen eine Förderung der Verdauung
zuzuschreiben. Ihre Gestalt ist sehr verschieden, bald kurz birnförmig,
bald gestreckt und in mehrere Lappen ausgezogen. Vielleicht haben die
bei Triphylus (Diglena) lacustris Ehr. jederseits vorhandenen
drei schlauchförmigen Ausstülpungen eine ähnliche Bedeutung. Mit
Ausnahme der Gattungen Asplanchna und Paraseison,
bei denen der Magen blind geschlossen endet, kommt ein flimmernder
Enddarm allen Rädertieren zu. Die sich an ihn anschliessende
Kloake entbehrt der Cilien mit Ausnahme einer Art (Rhinops
vitrea).
7. Die Exkretionsorgane.
Die Nephridien der Rotatorien zeigen bei allen Arten ein sehr
gleichförmiges Verhalten. Man kann an ihnen in der Regel zwei
zartwandige enge Längskanäle, die mit einer wechselnden Anzahl von
Geisselzellen besetzt sind, und eine gemeinsame kontraktile Blase,
welche in die Kloake ausmündet, unterscheiden. Im Gegensatze zu den
ganz ähnlich gebauten Nierenorganen der Turbellarien, Trematoden und
Cestoden bilden die Nephridien der Rädertiere keine Seitenzweige
und keine netzartigen Anastomosen untereinander, sondern jedes
„Wassergefäss“ zieht als ein leicht hin und her geschlängelter Kanal
von der kontraktilen Blase neben dem Darme nach vorn. In der Regel
reichen die Nephridien bis in die Kopfregion herein, wo sie blind
endigen und mittels zarter Bindegewebsfäden in ihrer Lage erhalten
werden. Nur bei wenigen Arten, z. B. der Gattung Synchaeta, sind
sie weit kürzer und reichen kaum über die Mitte des Körpers hinaus. Bei
vier verschiedenen[S. 312] Rädertieren stehen ausnahmsweise beide Nephridien
im Kopfe durch einen Querkanal mit einander in Verbindung; es sind
dies zumteil im System sehr weit getrennte Arten, nämlich Hydatina
senta, Lacinularia socialis, eine Floscularia-Spezies
und Apsilus lentiformis, sodass diesem Quergefäss vermutlich
noch eine weitere Verbreitung zukommt. Jedes Wassergefäss pflegt an
zwei oder drei Stellen sich knäuelförmig zu verschlingen, wobei die
die Wandung der Schlingen bildenden Zellen zu einer kontinuierlichen
Protoplasmamasse verschmelzen. In den so entstehenden Anschwellungen
gelingt es noch am leichtesten die Kerne der Wandzellen zu erblicken,
welche in den unverschlungenen Partien der Nephridien wegen ihrer
Kleinheit und Zerstreutheit schwer zu erkennen sind. Zellgrenzen haben
sich zwischen den Wandzellen bis jetzt nicht nachweisen lassen. Die
eigenartigen Geisselzellen der Nephridien, wegen ihrer flackernden
Bewegungen auch „Zitterorgane“ oder „Zitterflammen“ genannt, sitzen in
kleinen zartwandigen Ausstülpungen entweder direkt oder mittels eines
kleinen Stieles dem Nierenschlauche an. Jede Nephridie besitzt in der
Regel fünf bis zehn derselben — die Zahl ist für ein und dieselbe Art
konstant —, die sich in mehr oder weniger gleichmässigen Abständen
auf das ganze Organ verteilen. Bei den Asplanchnen wird die Anzahl
derselben meist eine viel grössere, und es tritt dann eine Spaltung
jeder Nephridie in zwei vorn und hinten zusammenhängende Kanäle ein,
von denen der eine dicht mit Zitterorganen besetzt ist (Fig. 68). Der
feinere Bau dieser Geisselzellen ist bis in die jüngste Zeit Gegenstand
lebhaftester Controverse gewesen, die sich vornehmlich darauf bezog,
ob dieselben am freien Ende offen oder geschlossen seien. Nach unserer
Ansicht ist unzweifelhaft das Letztere der Fall. — In die pulsierende
Blase münden die Nephridien mittels einer scharf umschriebenen runden
Öffnung ein. Die Kontraktilität wird durch ein Netzwerk feiner Muskeln,
die in der Wandung sitzen, hervorgerufen. Bei einigen Rotatorien
(Lacinularia, Tubicolaria, Pterodina) fehlt das
gemeinsame Harnreservoir, und die Nephridien münden direkt in die
Kloake. Endlich findet sich noch eine auffallende Modifikation bei
Conochilus volvox und den[S. 313] meisten Philodiniden. Hier hat sich
ein Teil der Kloake zur Harnblase umgewandelt und pulsiert ebenso
regelmässig wie die gewöhnlichen, von der Kloake scharf abgesetzten
Reservoire.
8. Die Klebdrüsen.
Von diesen sind in der Regel, den zwei Zehen entsprechend, nur zwei
vorhanden. Mit der Zahl der Zehen wächst aber auch häufig diejenige der
Drüsenschläuche; so finden wir z. B. bei der sechszehigen Callidina
parasitica und bei der am Schwanzende mit zehn kleinen Zäpfchen
besetzten Call. symbiotica vier Klebdrüsen. Das Sekret der
Fussdrüsen kann vielfach zu dünnen, elastischen Fäden ausgezogen
werden, mit denen sich das Tierchen vorübergehend vor Anker legt.
Eine ganz isoliert dastehende Umbildung haben die Klebdrüsen bei
Monocerca und Diurella erfahren, bei denen sie zu einer
mit kontraktiler Wandung versehenen Blase geworden sind.
9. Der Keimdotterstock und die Eibildung.
Das weibliche Geschlechtsorgan zeigt bei allen Rädertieren, mit
Ausnahme der marinen, auch in anderer Hinsicht sehr abweichenden
Seisoniden, eine Zusammensetzung aus zwei verschieden funktionierenden
Abschnitten. Der eine derselben ist so gross, dass er fast
ausschliesslich die Masse der Sexualdrüse ausmacht. Er liefert nur
die Nährsubstanzen, welche das heranwachsende Ei nötig hat, und wird
daher als Dotterstock bezeichnet. Der zweite, sehr viel kleinere
Abschnitt ist der Erzeuger der Eizellen und führt dementsprechend den
Namen Eierstock. Über Bau und Physiologie dieser beiden Teile des
weiblichen Geschlechtsapparates ist zurzeit kaum mehr bekannt, als
von Hydatina senta geschildert wurde. Es ist daher hier nur
Folgendes nachzutragen. Die Mehrzahl aller Rädertiere hat ein unpaares
Geschlechtsorgan, also nur einen Keimstock und einen Dotterstock. Man
kann sie als „Monogononten“ den Philodiniden gegenüberstellen, da diese
„digonont“ sind, d. h. ein rechtes und ein linkes Ovar, also zwei
Keimstöcke,[S. 314] zwei Dotterstöcke und zwei Ovidukte, besitzen. Keimstock
und Dotterstock haben nun bei jeder Spezies eine konstante Lage zu
einander, infolgedessen die Eier auch immer an derselben Seite des
dotterbildenden Abschnittes zur Entwickelung gelangen. Die Stellung
beider ist aber bei verschiedenen Arten eine verschiedene, und zwar
findet sich der Keimstock bald dem Vorder-, bald dem Hinter-, bald
endlich dem Seitenrande des Dotterstockes angelagert. Im ersten Falle
liegt der Keimstock meist sehr unsymmetrisch, nämlich der linken Ecke
des Dotterstockes angeschmiegt, wenn man von unten auf denselben
blickt. So situiert kommt er z. B. bei den Gattungen Hydatina,
Euchlanis, Brachionus, Triarthra u. a. vor.
Der zweite Typus wird durch Polyarthra, Conochilus,
Asplanchna u. a. vertreten. Unsere Abbildung 68 zeigt, wie
der Dotterstock der Asplanchna myrmeleo nicht die gewöhnliche
Sackform aufweist, sondern sich zu einem langen, hufeisenförmig
gekrümmten Bande gestreckt hat, dem in der Mitte der Hinterseite
der kleine rundliche Eierstock anliegt. Hier finden wir auch nicht
die typische Achtzahl der Kerne, sondern eine weit grössere. Dem
seitlichen und medianen Rande des Dotterstockes ist der Keimstock
bei den Philodiniden angelagert (Fig. 64), ein Verhalten, das auch
wahrscheinlich noch anderen Gattungen zukommen dürfte. Sehr interessant
werden viele Philodiniden, z. B. der gemeine Rotifer vulgaris,
dadurch, dass ihnen ein Ovidukt abgeht. An den Keimdotterstock
setzt sich zwar hinten und vorn ein Faden an, der aber lediglich
als Aufhängeapparat dient. Die Folge ist, dass die Eier in die
Körperhöhle fallen und hier ihre Entwickelung durchmachen. Diese geht
sehr rasch vor sich, und daher findet man selten einen Rotifer
vulgaris, der nicht in seiner Leibeshöhle ein oder zwei munter sich
umherbewegende und völlig ausgebildete Junge neben einigen Embryonen
beherbergte. Da eine besondere Geschlechtsöffnung fehlt, kann die
Geburt nur auf eine etwas gewaltsame Weise geschehen, und in der That
schieben sich die Tierchen mit dem Kopfe voran durch die Wand der
Kloake und dann durch den After ins Freie. Die der Mutter dadurch
zugefügte Risswunde scheint bald wieder zu heilen.
[S. 315]
Die zwei Eisorten, welche wir bei Hydatina kennen lernten, die
hartschaligen Dauereier und die gewöhnlichen, weichhäutigen scheinen
allen Monogononten zuzukommen. Bei den Erdrotatorien hingegen sind
Dauereier noch nicht beobachtet worden, da sie an die Austrocknung
in anderer Weise angepasst sind. Die derbe Schale der Dauereier ist
häufig mit sehr zierlichen Skulpturen: Punkten, Stacheln, Haaren
u. dergl. besetzt. Sie entwickeln sich bald schon nach einigen Wochen,
bald erst nach Monaten. Sie werden meines Wissens stets abgesetzt,
während die gewöhnlichen Eier bei vielen Gattungen (Brachionus,
Anuraea) dem Rücken angeheftet und so herumgetragen werden.
III. Die männlichen
Rotatorien.
Dieselben Verhältnisse, welche die charakteristischen Unterschiede
der männlichen Hydatinen von den weiblichen bilden, kehren bei allen
übrigen Rädertieren wieder. Die Männchen bieten, soweit sie überhaupt
bekannt sind, was nur für eine relativ kleine Anzahl von Gattungen
(etwa 30) zutrifft, in ihrer Organisation viel einfachere Verhältnisse
dar als die zugehörigen Weibchen, und ist dies so zu erklären, dass sie
einerseits überhaupt auf niedrigerer phyletischer Entwickelungsstufe
als die Weibchen stehen geblieben, anderseits auch in Folge der
untergeordneten Rolle, die sie seit dem Auftreten der Parthenogenese
im Geschlechtsleben spielen, rückgebildet sind. Ersteres macht die
grosse Gleichförmigkeit, welche die Mehrzahl der Männchen in Gestalt
und Organisation aufweist, verständlich, während auf letzteres die
geringe Körpergrösse, das Fehlen einer Mundöffnung und die Rückbildung
des Darmkanales und des Räderapparates zurückzuführen ist. Bei den
verschiedenen Gattungen ist der Geschlechtsdimorphismus in verschieden
starkem Grade ausgeprägt. Bei den marinen Seisoniden haben sich die
ursprünglichen Verhältnisse erhalten: Männchen und Weibchen stehen auf
gleicher Organisationshöhe und sind auch annähernd gleich häufig. Bei
den Euchlaniden besitzen die Männchen noch den Panzer in derselben
Gestalt wie die Weibchen, aber sie sind etwas kleiner als diese, und
der Darm ist auf einen unregelmässigen Zellstrang[S. 316] reduziert. In
allen übrigen Gattungen haben die Männchen eine weiche Haut, auch
wenn die Weibchen gepanzert sind, und einen sehr viel kleineren,
walzenförmigen Körper, der sich nach hinten verjüngt und, wie bei den
Weibchen, häufig mit zwei kleinen Zehen endet. Sehr auffallend ist das
♂ der Asplanchna Sieboldii dadurch, dass es vier kegelförmige
Körperanhänge besitzt, die der einen Form[XIV] von ♀ abgehen. Der
Räderapparat erinnert manchmal (Hydatina) noch an denjenigen
der Weibchen, meist aber ist er stark rückgebildet. Ein Stirntrichter
ist nur bei der männlichen Hydatina angedeutet, während sonst
die Wimperscheibe stark halbkugelig vorspringt und keine Spur der
ursprünglichen Mundöffnung erkennen lässt. Die im rudimentären Darm
vieler Männchen vorkommenden schwarzen Körnerhaufen — wahrscheinlich
bestehen sie aus Kalk — treten bei ganz jungen Weibchen einzelner
Gattungen (Brachionus) ebenfalls im Enddarm auf und beweisen die
Richtigkeit der Deutung jenes Zellstranges als eines rückgebildeten
Verdauungskanales. Die primitivere Organisation der Männchen spricht
sich im Nervensystem darin aus, dass die Tastbüschel nie auf besonderen
Hügeln oder Tentakeln stehen und im Exkretionsapparat in dem Fehlen
der kontraktilen Blase bei manchen Gattungen (Hydatina,
Brachionus), deren Weibchen eine solche besitzen; doch giebt es
auch Männchen mit einer Harnblase (Asplanchna, Apsilus).
Ein dorsaler einstülpbarer Penis ist nur bei einzelnen Gattungen
(Hydatina, Brachionus) vorhanden, bei anderen scheint
das hintere, verjüngte Körperende als solcher zu funktionieren
(Conochilus, Polyarthra, Anuraea). Über die
Begattung sind unsere Kenntnisse noch äusserst mangelhaft. Dem
bei Hydatina über sie Gesagten sei hier hinzugefügt, dass
Weber neuerdings bei Diglena catellina[XV] beobachtet
haben will, wie der Penis direkt in die weibliche Kloake geführt
wurde. Merkwürdigerweise kennt man von den gemeinsten Arten unter den
Philodiniden, z. B. von Rotifer vulgaris, die Männchen[S. 317] immer
noch nicht, und fast scheint es, als ob dieselben hier ganz fehlten,
und die Fortpflanzung ausschliesslich parthenogenetisch erfolgte.
[XV] Diese Beobachtung ist vom Herausgeber dieses Werkes bei
derselben Art bestätigt worden.
IV. Einige Bemerkungen über
die Biologie der Rädertiere.
Planmässige und allseitige Studien zur Biologie der Rotatorien
fehlen fast ganz, sodass ich an dieser Stelle nur auf einige
wenige Verhältnisse eingehen kann. — Über die Lebensdauer der
Wasserrotatorien ist nur das bekannt, was S. 295 über Hydatina
senta gesagt wurde. Bei den Erdrotatorien wird sich dieselbe
überhaupt schwerlich ermitteln lassen, da sie je nach Länge und
Zahl der Trockenperioden sehr verschieden ausfallen wird. Aus den
an Hydatina gemachten Beobachtungen scheint hervorzugehen, dass
die Lebenskraft der Weibchen erlischt, wenn der Keimstock sich
erschöpft hat. Die Männchen hingegen führen nur ein ganz ephemeres,
zwei- bis dreitägiges Dasein. — Als Nahrung dienen den Rädertieren
die verschiedenartigsten Mikroorganismen: Bakterien, Flagellaten,
Diatomeen, Infusorien, kleine Algen u. dergl. Einige Arten haben
eine schmarotzende Lebensweise angenommen und sich dadurch an
besondere Ernährungsbedingungen gewöhnt. So lebt Proales
(Notommata) Werneckii Ehr. im erwachsenen Zustande in den Kolben
von Vaucheria und nährt sich von dem Protoplasma dieser Alge.
Notommata parasita Ehr. und Hertwigia volvocicola
Plate leben in Volvoxkolonien. Die genannten Arten sind aber auch
im stande, sich von ihrem Wirte zu entfernen und sind daher nur
als zeitweilige Schmarotzer anzusehen. Albertia vermiculus
Duj. hingegen ist zum echten Entoparasiten geworden, der dauernd
in der Leibeshöhle des Regenwurmes und im Darme verschiedener
Nacktschnecken angetroffen wird. Eine Anzahl Rädertiere werden
konstant auf gewissen Tieren oder Pflanzen angetroffen, weil bestimmte
Lebensgewohnheiten derselben ihnen einen leichten Nahrungserwerb
sichern. Sie sind für den Wirt völlig indifferent, schaden ihm weder,
noch nützen sie ihm. Man hat ein derartiges Zusammenleben zweier
verschiedener Organismen als Raumparasitismus[S. 318] bezeichnet, eine
nicht gerade glückliche Benennung, da man von Parasiten nur dann
reden kann, wenn der eine Organismus durch den andern benachteiligt
wird. Zutreffender ist der Ausdruck „Raumsymbiose“, wenn die von
dem einen Lebewesen geschaffenen räumlichen Verhältnisse dem andern
zu gute kommen. Solche Raumsymbionten sind unter den Rädertieren
die Seisoniden, welche an den Extremitäten der marinen Krebsgattung
Nebalia leben, die Callidina parasitica, welche in
derselben Weise dem Bachflohkrebse sich anheftet, der auf der Haut
von Synapten lebende Discopus synaptae Zel. und endlich
Callidina symbiotica Zel. und Call. Leitgebii Zel.,
welche konstant auf gewissen Lebermoosen der Gattungen Radula,
Lejeunia, Frullania angetroffen werden und hier in
kleinen, von bestimmten Blattteilen gebildeten Höhlen, in denen die
Feuchtigkeit sich relativ lange erhält, leben. — Alle Raumsymbionten
leben in der Regel in einer grössern Individuenanzahl zusammen.
Das Gleiche gilt von vielen festsitzenden Rädertieren, z. B. der
Lacinularia socialis, der Melicerta ringens; indem
die jungen Tierchen sich neben den alten niederlassen, entsteht
eine Art Kolonie. Echte Kolonien werden nur von einem Rotator, dem
Conochilus volvox, gebildet. Sämtliche Individuen einer solchen
stossen im Zentrum einer Gallertkugel zusammen und sind selbst radiär
gerichtet. Der Ähnlichkeit, welche diese beständig rotierenden Kugeln
mit einem Volvox globator haben, verdanken die Tierchen ihren
Speziesnamen. — Gallertumhüllungen von Röhrenform werden auch von
anderen festsitzenden Arten abgeschieden, z. B. von dem schönen
Stephanoceros Eichhornii. Melicerta ringens kann sogar
den Anspruch erheben, ihr Gehäuse mit Hilfe eines Kunsttriebes
zu bauen. Sie weiss die herbeigestrudelten Partikelchen in einer
becherförmigen Vertiefung des Kopfes zu runden Ballen zu formen und
diese mittels einer Gallertausscheidung zu einer sehr regelmässigen
Wohnröhre zusammenzuheften. — Alle festsitzenden Rädertiere machen
natürlich in der Jugend ein freibewegliches Stadium durch. Zwei Arten
der Lokomotion werden bei den Rädertieren beobachtet. Weitaus die
meisten schwimmen mit Hilfe ihres Räderapparates unter beständigen
Drehungen um die Längsachse herum, bald[S. 319] schneller, bald langsamer.
Besonders rasch bewegen sich so alle Männchen durch das Wasser. Die
wenigen Formen mit ventraler Wimperscheibe (Adineta vaga,
Notommata tardigrada) kriechen ohne Rotation über die Unterlage.
Ein spannerraupenartiges Kriechen ist ausser dem Schwimmvermögen den
Philodiniden eigen, wobei sie sich abwechselnd mit den Fusszehen und
der Rüsselspitze anheften. — Unter allen biologischen Erscheinungen
der Rädertiere hat keines seit den Zeiten eines Leeuwenhoek die
Aufmerksamkeit der Naturforscher mehr gefesselt als die wunderbare
Lebenszähigkeit und die schier unverwüstliche Lebenskraft, welche
einigen derselben, nämlich den Erdrotatorien, innewohnt. Die Fähigkeit,
nach einer Periode völliger Austrocknung auf Wasserzusatz wieder
aufzuleben, kommt nur den zwischen Erde und Moos lebenden Philodiniden
zu. Alle echten Wasserbewohner und auch schon die ständig im Wasser
lebenden Philodiniden[XVI] gehen hingegen beim Verdunsten des Wassers
rettungslos zu Grunde. Diesen Tieren ist eben in den Dauereiern
ein besonderes Anpassungsmittel zur Erhaltung der Art gegeben.
Sehr merkwürdig ist es, dass sich viele Erdrotatorien so sehr an
ein intermittierendes Leben gewöhnt zu haben scheinen, dass ihnen
zeitweilige Trockenperioden geradezu zum Bedürfnis geworden sind.
Thut man nämlich philodinidenhaltiges Moos in ein Wassergefäss, so
beobachtet man bei vielen Arten nach einigen Tagen den Eintritt des
Todes. Obwohl Wasserorganismen und obwohl nur im flüssigen Elemente
ihrer Lebenskräfte sich erfreuend, sterben die Tiere dennoch bei
längerem Aufenthalte im Wasser ab, weil ihnen ein solcher in ihren
natürlichen Existenzbedingungen nie zu Teil wird. Die Lebenszähigkeit
der Erdrotatorien erweist sich nun nicht bloss beim Verdunsten des
Wassers, sondern auch gegenüber ungewöhnlich hohen oder niedrigen
Temperaturen, denn beiden muss die auf schwarzem Basaltfelsen zwischen
Flechten lebende, der glühenden Augustsonne nicht minder als der Kälte
der Dezembernacht blossgestellte Callidine gewachsen sein, soll die Art
nicht zu Grunde gehen. So verträgt die Callidina symbiotica,[S. 320]
wie Zelinka gezeigt hat, im eingetrockneten Zustand eine Kälte von
− 20° C. und eine Hitze von + 70°, und mit anderen Philodiniden hat man
ähnliche Erfahrungen gemacht.
[XVI] Einige Wasser-Philodiniden sollen eine Gallertcyste
ausscheiden und in dieser austrocknen können, eine Angabe, die jedoch
noch der Bestätigung bedarf.
V. Überblick über das System der
Rotatorien.
Sieht man das System der Tiere als Ausdruck ihrer phyletischen
Entwickelung, ihrer Stammesgeschichte, an, so lassen sich die
Rädertiere nur mit manchen Bedenken in einem der Endzweige des
natürlichen Stammbaumes unterbringen. Ihre Organisation ist nämlich so
eigenartig, dass sie sich an keine Tiergruppe bei ausschliesslicher
Berücksichtigung der erwachsenen Individuen derselben anschliessen
lassen. Am ungezwungensten reihen sie sich noch auf Grund ihres
Cilienapparates und ihrer Nephridien an die Turbellarien an.
Verständlich wird ihr Verhältnis zu den Evertebraten erst dann,
wenn man in ihnen eine sehr alte, primitive Tiergruppe sieht, deren
Ahnen den Ausgangspunkt für die Phylogenie einer ganzen Anzahl
anderer Tierklassen gebildet haben. Zu dieser Ansicht drängt der
Umstand, dass bei sehr vielen Anneliden, Turbellarien, Mollusken
und Bryozoen im Laufe der Ontogenie Larven auftreten, die eine
ausgesprochene Ähnlichkeit mit Rädertieren aufweisen, Larven, die man
unbedingt für Rotatorien halten würde, wenn sie in diesem Zustande
geschlechtsreif würden und einen Kauapparat besässen. Man nimmt daher
zurzeit an, dass jene Tierklassen und die Rädertiere phyletisch in
einer rotatorienartigen Stammform wurzeln, welche als „Trochophora“
bezeichnet wird.
Die Rotatorien zerfallen, wenn wir von den marinen Seisoniden absehen,
in zwei natürliche Gruppen:
1. Digononta seu Philodinida: Geschlechtsorgane paarig, jedes
mit oder ohne Ovidukt. Mitteldarm von einem Zellsyncytium gebildet.
Stets ohne laterale Taster. Am Rücken ein grosser, einstülpbarer
Rüssel, welcher die spannerraupenartige Bewegung vermittelt. Teils
Erd-, teils echte Wasserbewohner. Gattungen: Rotifer,
Philodina, Actinurus, Callidina. — Adineta.
2. Monogononta: Ovar unpaar, stets mit Ovidukt. Mitteldarm aus
grossen Zellen gebildet. Mit lateralen und dorsalen Tastern.[S. 321] Ohne
Rückenrüssel. Schwimmend oder festsitzend. Nur ständige Wasserbewohner.
— Hierher gehört die Mehrzahl aller Rädertiere, deren weitere
systematische Einteilung am besten aus Hudson-Gosses Monographie zu
ersehen ist. Wir erwähnen nur die Hauptfamilien:
a) Melicertida seu Rhizota. Im Alter festsitzend. Ohne Zehen.
Hinteres Körperende nicht einziehbar. Gattungen: Floscularia,
Stephanoceros, Melicerta, Lacinularia,
Limnias, Oecistes, Conochilus.
b) Illoricata. Haut weich. Gattungen: Asplanchna,
Synchaeta, Hydatina, Notommata.
c) Loricata. Mit Panzer. Gattungen: Rattulus,
Dinocharis, Salpina, Euchlanis, Lepadella,
Colurus, Pterodina, Brachionus, Anuraea.
Alle Abbildungen sind bei starker Vergrösserung gezeichnet. Die
Buchstaben bedeuten:
a
After
c
Kutikula
c.v
Kontrakt. Blase
dst
Dotterstock
dt
dorsaler Taster
dr
Magendrüse
est
Eierstock
f
Fussdrüse
g
Gehirn
hg
Hypodermis
kl
Kloake
l
lateraler Taster
ma
Kauapparat
mu
Muskel
ne
Nephridie
o
Augenfleck
oe
Oesophagus
ov
Ei
re
Enddarm
r
Räderapparat
sto
Magen
t
Tastborste
te
Hoden
ut
Uterus
z
Zitterflamme (Geisselzelle).
[S. 322]
Litteratur.
1) F. Cohn, Die Fortpflanzung der Rädertiere. Zeitschr. f.
wiss. Zool. VII, 1856, p. 431–486.
2) E. v. Daday, Morphologisch-physiol. Beiträge zur Kenntnis
der Hexarthra polyptera. Budapest 1886.
3) C. Eckstein, Die Rotatorien der Umgegend von Giessen.
Zeitschr. f. wiss. Zool. XXXIX, 1884, p. 343–443.
4) G. Chr. Ehrenberg, Die Infusionstierchen als vollkommene
Organismen. Leipzig 1838.
5) H. Grenacher, Einige Beobachtungen über Rädertiere.
Zeitschr. f. wiss. Zool. XIX, 1869, p. 483–497.
6) C. T. Hudson u. P. H. Gosse, The Rotifera or
Wheel-Animalcules. 2 Bde. London 1886 und 1 Supplementband 1889.
Grundlegend für die Systematik.
7) F. Leydig, Über den Bau u. die systematische Stellung d.
Räd. Zeitschr. f. wiss. Zool. VI, 1854, p. 1–120.
8) L. Plate, Beiträge z. Naturgesch. d. Rot. Jenaische
Zeitschr. f. Nat. XIX, 1885, p. 1–120.
9) L. Plate, Untersuch. einiger an den Kiemenblättern des
Gammarus pulex lebenden Ektoparasiten. Zeitschr. f. wiss. Zool. XLIII,
p. 229–235.
10) L. Plate, Über die Rotatorienfauna des bottnischen
Meerbusens etc. ibid. XLIX, p. 1–42.
11) G. Tessin, Über Eibildung und Entwickelung der Rotatorien.
ibid. XLIV, 1886.
12) E. F. Weber, Notes sur quelques Rotateurs des Environs de
Genève. Arch. de Biologie. VIII, 1888.
13) C. Zelinka, Studien über Rädertiere. I. Über die Symbiose
und Anatomie von Rot. aus dem Genus Callidina. Zeitschr. f. wiss. Zool.
XLIV.
14) C. Zelinka, II. Der Raumparasitismus u. die Anatomie von
Discopus Synaptae. ibid. XLVII.
[S. 323]
Die Krebsfauna
unserer Gewässer.
Von Dr. J. Vosseler in Tübingen.
[S. 325]
Die Krebse (Crustacea) werden zu dem grossen Tierkreise der
Gliederfüssler (Arthropoden) gerechnet, welche eigentlich besser
mit dem Namen Kerftiere bezeichnet werden, denn im Grunde genommen
besitzen die meisten höheren Wirbeltiere ebenfalls gegliederte Beine
und müssten somit auch unter den Begriff „Arthropoden“ eingereiht
werden. Im grossen Ganzen kann man die Kerftiere in zwei Klassen
trennen: in solche, welche durch Kiemen atmen (Branchiata
Krebse), und solche, welche durch Tracheen (fein verzweigte Röhrchen,
welche die Luft im ganzen Körper herumleiten) atmen (Tracheata).
Zwischen beiden Klassen finden sich, wie beinahe überall im Tierreiche,
Übergänge. Zu der zweiten Klasse, welche ihrer Organisation wegen
als die höher entwickelte angesehen werden muss, zählt man die
Tausendfüsse (Myriopoda oder Vielfüsser), die Spinnentiere
(Arachnoidea oder Achtfüsser) und die Insekten (Hexapoda
oder Sechsfüsser).
Die Krustaceen (Branchiata) selbst können in zwei
Abteilungen untergebracht werden, welche jedoch nicht vollkommen
wissenschaftlich gegen einander abgegrenzt sind. Dennoch ist es für
die Übersichtlichkeit unseres Stoffes von Vorteil, wenn die von
verschiedenen Lehrbüchern bis jetzt durchgeführte systematische
Trennung beibehalten wird.
Hiernach werden die kleineren Krebse, welche eine einfache
Organisation besitzen und wenigstens teilweise in der Entwickelung
Übereinstimmung zeigen, als Entomostraken den grösseren
Krebsen mit vollkommeneren Einrichtungen, den Malakostraken,[S. 326]
gegenübergestellt. Die Vertreter beider Abteilungen lassen sich leicht
mit blossem Auge unterscheiden und von einander trennen und innerhalb
dieser zwei Gruppen machen sich weitere Verschiedenheiten bei genauer
Beobachtung bemerklich, welche charakteristisch genug sind, um auch für
den Nichtkundigen hier angeführt zu werden.
Streifen wir mit einem kleinen Netz (wie es etwa zum Schmetterlingsfang
benutzt wird) die Wassergewächse ab und durchfahren damit auch in
der Nähe der Uferpflanzen das Wasser, so erbeuten wir eine ganze
Menge kleiner Krustaceen, die, in ein Glas mit Wasser gebracht, ein
unendliches Gewimmel darstellen. Auf einen derartigen Fang wollen
wir jetzt unsern prüfenden Blick werfen. Wir beginnen mit den
grösseren Insassen des durchsichtigen Behälters und fassen ein Tier
ins Auge, welches sich immer am Boden und zwischen den miterhaltenen
Pflanzenteilen zu verstecken sucht. Wird es gestört, so schwimmt es
mit gekrümmtem, seitwärts zusammengerolltem Körper eine kurze Strecke
geradeaus, fast immer auf der einen Körperseite liegend und auch in
der Ruhelage stets die Beine schwingend. Seine Länge beträgt einen
Centimeter und mehr, wenn es ausgewachsen ist. Der Umstand, dass mehr
als drei Beinpaare vorhanden sind, lässt auf einen Krebs schliessen.
Die genannten wenigen Merkmale, zu denen noch zwei Paar fadenförmige
Fühler von weniger als Körperlänge zu rechnen sind, genügen, um einen
Angehörigen aus der Abteilung der höheren Krebse zu kennzeichnen
und wir haben in dem beobachteten Tiere einen Flohkrebs oder
Gammarus der Ordnung der Amphipoden vor uns. Fast noch
leichter kenntlich ist ein Verwandter desselben, welcher offenbar ein
ganz schlechter Schwimmer ist. Unter seinem wie bei einer Schildkröte
verbreiterten, an den Seiten überstehenden Rückendach sind ebenfalls
mehr als drei Beinpaare zu sehen und am Kopfe, wie vorhin, zwei Paar
ungleich langer Fühler. Unbehelligt krabbelt der plumpe Geselle langsam
und schwerfällig an der neuen Umgebung herum, immer mit den Fühlern
tastend und prüfend. Berührt man ihn jedoch mit einem Stäbchen, so
zeigt er, dass auch ihm rasche[S. 327] Bewegungen möglich sind, wenn auch
in geringerem Masse und weniger eleganter Ausführung, als seinem
Vorgänger. Die ganze Erscheinung dieses zweiten Krebses erinnert an
die bekannten Kellerasseln, aus deren Sippe er auch in der That einen
Spross vorstellt, welcher das kühle Wasserleben noch nicht mit dem auf
dem Lande vertauscht hat. Wir nennen ihn Wasserassel (Asellus)
und rechnen ihn ebenfalls zu den höheren Krebsen und zwar zur
Ordnung der Isopoden.
Diese beiden eben nach ihren gröbsten Umrissen und Gewohnheiten
erkannten, zu den Malakostraken gehörigen Formen werden wir
später betrachten und wollen nun die kleinen Insassen des Glases,
soweit es nicht Junge der genannten Arten sind, nach ihrem Äussern mit
blossem Auge unterscheiden lernen. Es bleibt uns, da wir Insekten und
Würmer nicht beachten, nur noch das Gewimmel kleiner und kleinster
Wesen, welche oft kaum mehr mit dem blossen Auge erkennbar sind, die
Entomostraken, übrig. Trotzdem genügt auch hier ein wenig Geduld und
sorgsames Zuschauen, um sowohl in den Körperformen als in der Art der
Bewegung noch deutliche Unterschiede wahrzunehmen. Während die einen
durch ununterbrochenes Hüpfen voranzukommen oder sich wenigstens in der
Höhe zu halten suchen, zappeln die anderen scheinbar ziellos im Glase
herum. Diese wie jene setzen sich von Zeit zu Zeit, wie um auszuruhen
von der anstrengenden Bewegung, am Glase oder an Pflanzenteilen fest,
oder lassen sich auf den Grund niedersinken. Unsere Wissbegierde treibt
uns immer weiter, und da sich allmählich das Auge daran gewöhnt hat,
Unterschiede zu entdecken, so wird es nicht schwer fallen, unter den
hüpfenden Tierchen abermals zweierlei zu unterscheiden. Es heisst nun
allerdings etwas genau zusehen, denn wir sind nahezu auf dem Punkte
der Forschung angelangt, wo die Leistungsfähigkeit des unbewaffneten
menschlichen Auges der Wissbegierde des Forschers Grenzen setzt.
Eine langsamere gleichmässige Bewegung, welche eigentlich das Tier,
falls es nicht gestört wird, weniger vorwärtsbringt, als vielmehr
stets in einer gewissen Höhe über dem Boden erhält, des weiteren ein
schwarzer Punkt, welcher am Kopfteil sitzt und das[S. 328] Auge darstellt,
kennzeichnet die Wasserflöhe oder Daphniden, welche zu der Ordnung
der Blattfüsser (Phyllopoden) zählen. Der Leib selbst,
vom Kopfe nicht besonders scharf sich abhebend, ist mehr oder
weniger eiförmig. Über den Umrissen ragt hinten oftmals ein Stachel,
am Kopf zwei wie Hörner emporstehende Fühler, welche die Bewegung
verursachen, hervor. Viel mehr würden in Rücksicht auf ihre Bewegung
die länglichen, vorn dicken, hinten schlanken Hüpfer den Beinamen
„Floh“ verdienen, welche recht häufig mit einem grossen grünlichen
bis weissen Säckchen an jeder Körperseite (den Eiersäcken) und zwei
(manchmal recht langen) Hörnchen, welche wagrecht abstehend getragen
werden, in eleganten weiten Sätzen im Wasser herumschnellen. Wir nennen
sie Hüpferlinge oder Widderchen und reihen sie in die grosse Ordnung
der Ruderfüsser (Copepoden) ein. Dem hüpfenden Teil der
kleinen Tierwelt in unserem Glase hätten wir nun auch ein Plätzchen
im System angewiesen. Wir wenden uns jetzt zu den am schwersten
erkennbaren Formen und verfolgen mit einiger Ausdauer das nächste beste
der unruhig umherzappelnden Geschöpfe. Es wird hier nötig, um leicht
mögliche Verwechslungen auszuschliessen, eine nicht zu den Krebsen
gehörige Gruppe von Tieren in ihrem Habitus zu schildern. Es mag somit
das kleine Wesen einen annähernd kugelrunden Körper, an dem sich
vier nicht versteck-, sondern höchstens anlegbare Beinpaare erkennen
lassen, besitzen. Vielleicht gelingt es sogar, eine schön gefärbte
Zeichnung auf dem Rücken nachzuweisen. Diese Merkmale lassen auf eine
Wassermilbe schliessen, welche wir nicht weiter beachten. Ganz ähnlich
in ihren Bewegungen verhält sich eine Familie der Entomostraken, deren
Angehörige nach aussen von einer zweiklappigen Schale umschlossen
werden, zwischen denen das Körperchen ganz versteckt liegt. Diese
Schale ist der einer Muschel ausserordentlich ähnlich und ihr verdanken
die Insassen den Namen „Muschelkrebse“ (Ostrakoden).
Oft wird man überrascht, wie aus dem Spalt, der die beiden Klappen
trennt, plötzlich eine Anzahl Gliedmassen hervortreten und durch eine
rasche Thätigkeit das Tierchen in etwas unsicheren Linien von Stelle
zu Stelle bewegen. Während der[S. 329] Ruhepausen werden die Beine eingezogen
oder zum Krabbeln am Boden oder an Pflanzenstengeln benutzt. Glaubt das
Tier sich gefährdet, so zieht es ebenfalls sofort seine Beine ein und
schliesst die Schalen. Es liegt dann die nierenförmige Schale, wie ein
unbelebtes Ding, vor uns.
Wenn wir nun noch einer Form gedenken, in welcher die Krebstiere des
süssen Wassers den höchsten Grad der Entwickelung erreicht haben
und für welche, da sie der ganzen Tierklasse den Namen gegeben
hat, ein weiteres Charakteristikum wohl nicht nötig ist, — ich
meine den Flusskrebs —, so kennen wir nun Vertreter aus allen
Abteilungen und Ordnungen der unsere Gewässer bewohnenden Krustaceen.
Selbstverständlich genügt nicht immer das Absuchen einer Fundstelle
allein, um alle, so wie es geschildert wurde, auf einmal vor Augen zu
bekommen; doch wird es nirgends an stehenden und fliessenden Wassern
fehlen, welche fast zu jeder Jahreszeit die typischen Formen zu sammeln
gestatten. Zum Schluss dieser systematischen Betrachtungen lasse ich
eine kurze Übersicht des Wesentlichsten folgen. Wir haben in der
zu den Kerftieren gehörigen Klasse der Krustaceen zwei Abteilungen
unterscheiden gelernt:
Entomostraca oder niedere Krebse,
Malacostraca oder höhere Krebse.
Zu der ersten Abteilung rechnen wir:
Ruderfüsser oder Spaltfüsser
(Copepoden), Hüpferlinge,
Kiemenfüsser
(Branchiopoden), Wasserflöhe,
Muschelkrebse (Ostrakoden).
Zu der zweiten:
Flohkrebse (Amphipoden),
Wasserasseln (Isopoden),
Scherenkrebse
(Dekapoden), Flusskrebs.
[S. 330]
Wir wenden uns zuerst den Gliedern der ersten Abteilung, den
Entomostraken,
zu, da sie in ihrem Bau einfacher angelegt und leichter zu übersehen
sind, als die „höheren Krebse“.
Es fällt schwer, nur einige gemeinsame Kennzeichen für alle hierher
gehörigen Tiere nachzuweisen; denn nicht einmal immer kann man ohne
weiteres einen Kopf- und einen Schwanzteil unterscheiden. Am besten
wird die Abteilung der Entomostraken dadurch charakterisiert, dass
sie ausserordentlich reich an verschiedenen Krebsformen ist, deren
Gliedmassen sowohl in der Zahl als auch Gestaltung ebenso wie die
Segmente des Körpers sehr variieren. Alle Entomostraken besitzen,
wie auch die höheren Krebse, zwei Paar Fühler. Ferner herrscht
unter den nachher zu besprechenden Ordnungen mit Ausnahme nur einer
Unterordnung annähernde Übereinstimmung in der Körpergestalt während
der ersten Entwickelungsstadien nach dem Verlassen des Eies. Nicht alle
Entomostraken-Familien sind im süssen Wasser vertreten, eine derselben,
die der Rankenfüsser (Cirripedien), findet sich nur im Meere vor. Die
Zahl der Gattungen und Arten, welche einer Familie angehören, bewegt
sich in weiten Grenzen. Von Entomostraken sind gegenwärtig weit über
2000 Arten bekannt. Von dieser Summe zählt etwa die Hälfte zu den
Copepoden, ¼ zu den Ostrakoden, ⅐ zu den Phyllopoden und nur ⅛
zu den im Meere lebenden Cirripedien. Die Mehrzahl der Arten auch der
übrigen Familien ausser den Cirripedien lebt ebenfalls im Meere. Aus
der umfangreichen Familie der Copepoden lebt nur der zehnte
Teil im süssen Wasser. Wohl die bedeutendste Rolle von allen niederen
Krebsen spielen in unseren Gewässern die
Copepoden oder Ruderfüsser.
In dieser zahlreichen Familie unterscheiden wir drei Gruppen, von
denen zwei allerdings für das praktische Leben nur wenig Bedeutung
haben, allein nichtsdestoweniger ausserordentlich interessant
sind wegen der enormen Umbildung des Körpers. Die[S. 331] Tiere, welche
diesen Gruppen angehören, leben nämlich parasitisch und haben sich
dieser Lebensweise so angepasst, dass sie als Ruderfüsser kaum mehr
zu erkennen sind und lange falsch beurteilt wurden. Ehe wir uns
diese Schmarotzerkrebse, welche ihrer saugenden Mundteile wegen
Siphonostomata genannt werden, näher betrachten, wollen wir die
freilebenden Ruderfüsser, welche beissende Mundteile besitzen und als
Gnathostomata bezeichnet werden, kennen lernen. Schmarotzende
und freilebende Ruderfüsser werden zusammen als echte Copepoden
oder Eucopepoden den sogenannten Karpfenläusen
(Branchiura oder Kiemenschwänzen) gegenübergestellt.
Freilebende
Ruderfüsser (Gnathostomata).
Die erste Nachricht über das Vorkommen von Ruderfüssern (auch
Spaltfüsser genannt) verdanken wir Leeuwenhoek; sie stammt
aus dem Jahre 1690. Allein erst von der Mitte unseres Jahrhunderts
an wurden sie eigentlich genauer studiert, besonders von
Fischer[108]
in Petersburg und C. Claus[109][110][111].
Die Bedeutung des dem Griechischen entstammenden wissenschaftlichen
Namens ist bereits angegeben worden. Die Tiere, welche unter diese
Familie zu rechnen sind, besitzen eine nur geringe Körpergrösse,
gewöhnlich ½–3 mm, höchst selten misst eine Art 5 mm
oder gar, wie einige im Meer lebende Formen, 9 mm. Der Körper
kann bei den meisten etwa mit einer halbierten Birne verglichen
werden. Die Wölbung würde dem Rücken, die Schnittfläche der Bauchseite
entsprechen. Der grössere, dickere Teil stellt dann den Vorderleib,
der dünnere den Hinterleib dar. Der Vorderleib ist aus fünf Teilen
zusammengesetzt, welche an und für sich starr, dennoch gegen einander
beweglich sind durch dünnhäutige Einschnitte, wie sie auch dem
Hinterleib und den Gliedmassen die Beweglichkeit verleihen. Das grösste
Stück des Vorderleibes ist oft allein so lang wie die übrigen zusammen.
In ihm ist Kopf und Brust vereinigt, weshalb es Kopfbruststück oder
Cephalothorax genannt wird. Da bei den Kerftieren gewöhnlich jeder
Körperring der ersten Anlage nach mit einem Paar Gliedmassen versehen
ist, so lässt sich aus[S. 332] der am Kopfbruststück der Copepoden vorhandenen
Summe derselben schliessen, dass mehrere Ringe zur Bildung dieses
verschmolzen sind. Die nächsten vier, immer noch zum Vorderleib
zählenden Ringe nehmen allmählich an Grösse ab und tragen Beinpaare.
Der auf den letzten folgende Hinterleib zählt fünf Ringe. Diese sind
beim Männchen getrennt erhalten, während beim Weibchen der erste mit
dem zweiten zu einem einzigen Segment (Ring) verschmilzt. Im hintersten
Segmente mündet der Darm nach aussen. Eine Art Steuerruder bildet den
Abschluss des Körpers. Dieses wird, da es aus zwei nebeneinander dem
letzten Hinterleibsringe aufsitzenden Teilen besteht, welche etwa mit
den Zinken einer Gabel verglichen werden können, „Schwanzgabel“ oder
„Furca“ genannt. Beide Zinken tragen am Ende lange Borsten. Länge und
Breite der Körpersegmente sind je nach der Art des Tieres manchem
Wechsel unterworfen. Während bei einigen Familien der Hinterleib so
lang ist wie der Vorderleib, ist bei anderen dieser bedeutend länger
als jener.
Die Copepoden sind, wie die meisten Krebstiere, reichlich mit
Gliedmassen versehen und zwar im ganzen mit etwa elf Paaren, welche
dreierlei Verrichtungen zu erfüllen haben. Zum Tasten und Schwimmen
dienen zwei Fühlerpaare, der Ernährung vier Paar Mundwerkzeuge, auf
welche fünf Beinpaare gleichzeitig mit den Fühlern das Schwimmen
vermittelnd folgen. Von den letzteren ist gewöhnlich das hinterste
verkümmert oder zu einem Greiforgan bei den Männchen umgewandelt.
Wie der Körper und die übrigen Gliedmassen, sind auch die Fühler aus
hintereinander liegenden Segmenten oder Ringen zusammengesetzt. Die
Zahl dieser Ringe ist eine je nach der Art, dem Geschlecht und Alter
verschiedene, sie kann sogar innerhalb einer Art variieren.
Von den drei bei uns vorkommenden Familien von Ruderfüssern haben
die Harpactiden die geringste Zahl von Gliedern, an den Fühlern
nämlich höchstens acht; die Cyclopiden besitzen 6–18, die Calaniden
gar 25. Die Antennen aller Copepoden sind reichlich mit Borsten
und kleinen Sinnesorganen, welche später besprochen werden sollen,
versehen. Wie schon gesagt,[S. 333] dienen diese Fühler wesentlich mit zur
Bewegung. Seltsame Umbildungen erfahren sie bei den Männchen. Während
die weibliche Antenne gewöhnlich fadenförmig ist und gegen das Ende
zu gleichmässig an Dicke abnimmt, wobei die verschiedene Länge der
einzelnen Glieder keine Rolle spielt, ist die des Männchens zu einem
ganz komplizierten Greiforgan umgestaltet. Diese Funktionsvermehrung
trifft bei Harpactiden und Cyclopiden die beiden Antennen, bei den
Calaniden nur die der rechten Seite. Bei der letzteren Familie ist der
fünfte Fuss auf der entsprechenden Seite ebenfalls zu einem Greiforgan
verwandelt. Alle diese geschlechtlich differenzierten Antennen
sind zweimal geknickt (Fig. 70, am) und können nach vorn
einwärtsgeschlagen werden, so dass eine Art Schlinge entsteht, mit der
die Weibchen gefangen und während der Begattung festgehalten werden.
Die zweiten Antennen sind meistens bedeutend kürzer und setzen sich
bei Cyclopiden und Harpactiden aus vier Gliedern zusammen. Bei den
Calaniden sind sie in zwei Äste gespalten, deren einer vier, der andere
sieben Glieder besitzt.
Der Mund ist von einer als Oberlippe bezeichneten bezahnten Platte
überdeckt. An seinen Seiten sitzen zwei Paar Kiefern und ebenso viele
Kieferfüsse, welche in stetiger Bewegung sind und viele Borsten und
Stacheln tragen.
Fig. 70. Cyclops tenuicornis Claus (Weibchen, von oben gesehen). a′ Erste
Antenne — a″ Zweite Antenne — am Antenne des Männchens —
au Auge — m Längsmuskel — M Magen — D Darm —
E Eiersäckchen — Ee Eileiter, mit Eiern gefüllt.
Von den nun folgenden vier Schwimmfusspaaren ist das erste noch am
Kopfbruststück angeheftet und gewöhnlich das kleinste.[S. 334] Alle sitzen
noch am Vorderleib. Sie bestehen aus einem breiten Grundgliede, auf
welchem zwei Äste mit je zwei bis drei Gliedern entspringen; sie werden
als Spaltbeine bezeichnet. Wie die Mundwerkzeuge sind auch besonders
die Endglieder der Beine reichlich mit Borsten und Dornen ausgestattet.
Es wäre nun noch eines fünften, rudimentär gewordenen Fusspaares zu
gedenken, welches dem kleinsten (fünften) Vorderleibssegment aufsitzt,
oft nur durch Borsten angedeutet, oft 1–2gliederig ist oder gar noch
aus einem Grundgliede und zwei kleinen Ästen (Calaniden) besteht. So
unscheinbar es ist, so gross ist seine Bedeutung für die Unterscheidung
der Arten. Wie oben erwähnt wurde, ist der rechte Fuss des rudimentären
Paares bei den Calaniden zu einem Greiforgan umgewandelt und trägt
einen sehr starken langen Dorn am Ende. Bei den Harpactiden erleidet
das dritte Schwimmfusspaar eine entsprechende Umwandlung zum gleichen
Zweck.
Es mögen hier einige Bemerkungen über den Bau der Körperbedeckung
eingefügt werden, welche das Tier gegen die Umgebung abgrenzt und ihm
Schutz verleiht. Bei allen Kerftieren besteht die Haut aus zwei Lagen,
von denen die eine die andere erzeugt. Die äusserste Lage besteht aus
einem „Chitin“ genannten Stoff und ist von sehr wechselnder Dicke.
Während manche Käfer vom Sammler kaum mit der Nadel durchbohrt werden
können, hat die Stubenfliege nur eine dünne Chitindecke und kann daher
leicht zerdrückt werden. Von dieser Chitinhaut sind alle die
weicheren inneren Organe schützend umschlossen; an ihr setzen sich auch
die Muskeln und Sehnen an. Da nun der äussersten Körperschicht die
Aufgabe zufällt, den Körper zu stützen und Ansatzstellen für alle die
Muskeln abzugeben, welche die Gliedmassen etc. bewegen, so kann man
bei den Kerftieren und also auch bei den Krebsen von einem äusseren
Skelett reden. Dieses wird von einer darunter liegenden ganz weichen
Schicht, der Matrix oder Hypodermis, deren Elemente kleine, vielseitige
platte Zellen darstellen, abgesondert. Doch geschieht die Absonderung
nicht immerfort gleichmässig, sondern sie findet nur nach bestimmten
Zwischenpausen bei noch nicht ausgewachsenen Tieren statt.[S. 335] Jeder neuen
Absonderung geht ein Abwerfen der alten Chitinhaut vorauf.
Ähnlich ist auch die Haut, welche den Anfang und das Ende des
Darmrohres auskleidet, zusammengesetzt und wird ebenfalls bei der
Häutung abgestossen.
Da wir nun das Äussere der Copepoden einigermassen kennen, wenden wir
uns zu dem Bau und der Zusammensetzung der inneren Organe und Systeme.
Dieselben sind, trotz ihrer Einfachheit, ihrer Struktur und Funktion
nach doch oft recht schwierig zu erkennen.
Gleich das Nervensystem, von dem ja alle Lebensäusserungen,
willkürliche wie unwillkürliche, ausgehen, ist bei der Kleinheit der
Tiere, zu der sich noch andere Hindernisse gesellen, so schwer zu
entdecken, dass bei mancher Art noch nicht einmal die äussere Form
mit voller Sicherheit festgestellt ist. Von vornherein lässt sich
ja annehmen, dass es sich vom Bau nahe verwandter Arten, von deren
zentralem Nervensystem man genaue Kenntnis hat, nicht allzuweit
entfernen werde. Gewöhnlich lagert sich der Hauptteil, den man als
Hirn bezeichnen kann, wie bei allen Kerftieren, über dem Schlund und
zwar in Form von Ganglien, d. h. Anschwellungen von Nervensträngen,
in denen die Elemente des Nervensystems, Fasern und Zellen, bei
einander liegen. Solche Ganglien sind, der bilateralen Symmetrie des
Körpers entsprechend, gewöhnlich paarweise angelegt. Die Paare selbst
treten ursprünglich in jedem Segment in der Einzahl auf, können aber
sowohl seitlich verwachsen, als auch, wenn Segmente mit einander
verschmelzen, zusammenrücken, so dass mehrere hintereinander gelegene
Paare durch Verwachsung nur noch ein einziges darstellen. Um wieder
auf das „obere Schlundganglion“ (so lautet die wissenschaftliche
Bezeichnung des Hirns) zurückzukommen, so ist es meistens auch durch
seine Grösse vor den anderen ausgezeichnet. Von ihm gehen rechts
und links an den Seiten des Schlundes Nerven (Kommissuren) hin,
welche in ein unter dem Schlunde liegendes Ganglienpaar eintreten.
Auf diese Weise entsteht eine Art Ring, welcher sich eng um das
Rohr des Schlundes herumlegt und „Schlundring“ genannt wird. Dieses
„untere Schlundganglion“ ist mit den weiter nach hinten[S. 336] liegenden
segmentalen Ganglien durch zwei Nervenstränge, wie jene unter sich,
verbunden. Das Bild dieses „Bauchstranges“ gleicht einer Strickleiter.
Wie schon der Name besagt, liegt dieser Bauchstrang, welcher unserem
Rückenmark entspricht, an der Bauchseite unter dem Darm. Ob überall
bei unseren Copepoden auch im Hinterleib solche durch Nerven mit
einander verbundene Ganglien zu finden sind, ist nicht ganz sicher
festgestellt. Bei den Cyclopiden fehlen sie dort, was natürlich nicht
ausschliesst, dass derselbe dennoch reichlich mit Nerven versorgt ist.
Von diesem Zentralnervensystem, welches, wie wir gesehen haben, aus
einem Schlundring und einer Bauchganglienkette sich zusammensetzt,
gehen nun gröbere und feinere sich reichlich verzweigende Nervenäste
zu allen Teilen und Organen des Körpers und vor allem zu den
Sinnesorganen.
Obgleich diese im Vergleich mit anderen Kerftieren nur schwach
entwickelt sind, scheinen die Copepoden dennoch für die meisten
Eindrücke recht empfänglich zu sein. Am leichtesten sichtbar und keiner
Art fehlend ist das Auge. Es ist bei grösseren Arten, namentlich
bei den fast farblosen pelagischen Tieren, ohne weitere Hilfsmittel
durch seine gewöhnlich rot bis rotbraune Färbung kenntlich. Häufig
glänzt es sehr schön.
Ausnahmslos müssen sich die Copepoden mit einem einzigen Auge
behelfen und dieser Umstand hat der Gattung Cyclops mit einer
Anspielung auf den einäugigen Schmiedeknecht Vulcans im Ätna zu ihrem
Namen verholfen. Dieses eine Auge ist so primitiv gebaut, dass wir
ruhig annehmen können, es werde seinem Besitzer zu keiner besonders
vollkommenen Erkenntnis der Umgebung verhelfen und nicht viel mehr
als die Unterscheidung von hell und dunkel ermöglichen. Es sitzt bei
allen Arten mitten auf der Stirn zwischen den beiden Fühlern und ist
bloss bei den Calaniden ein wenig beweglich, sonst aber fest. Mit
einem Vergrösserungsglas sieht man dem schon erwähnten Pigmentfleck
zwei kleine glashelle Linsen aufsitzen, welche meistens den Glanz
verursachen. Denkt man sich nun hierzu einen Nerv, der vom Hirn zum
Auge zieht und unter demselben eine kleine gangliöse Anschwellung
zeigt, so[S. 337] kennen wir die ganze Einrichtung, wie sie einfacher kaum
gedacht werden kann. Die nächstniedere Entwickelungsstufe des Auges
kommt schon sehr früh in der Tierreihe vor bei wirbellosen Tieren und
stellt einen mit Nerven versehenen Pigmentfleck dar, ist also um das
optische Hilfsmittel der beiden Linsen ärmer. Die Linsen selbst sind
aus der Chitinschicht der Haut gebildet und werden bei jeder Häutung
durch neue ersetzt.
Wenn wir somit dem Formen- und Farbensinn der kleinen Hüpferlinge
keine besondere Hochachtung zollen dürfen, so kann man doch aus
manchen Beobachtungen schliessen, dass nicht alle Sinne gleich
schlecht entwickelt sind. Namentlich scheinen der Geruch und
Geschmack gut ausgebildet zu sein. Nur fehlen uns vorderhand die
Mittel, genau zu unterscheiden, welche Organe dem einen, welche dem
anderen Sinne dienen. Es ist ja nicht unmöglich, dass bei den im Wasser
lebenden Tieren ein und dieselbe Vorrichtung beiderlei Perceptionen
ermöglicht, da nur ein Medium vorhanden ist, um die erregenden Stoffe
zu übermitteln — das Wasser. Ob nun die Stoffe gasförmig oder fest
sind, bleibt sich gleich. Da nicht wohl anzunehmen ist, dass unlösliche
Stoffe einen Eindruck auf andere als Seh- und Tastorgane machen, so
ist von diesen abzusehen. Lösliche Stoffe jedoch können nach unseren
Anschauungen von einem Sinnesorgan wahrgenommen werden. Wie bei
den höheren Wirbeltieren ist der Geruchs-Geschmackssinn auf bestimmte
Körperteile beschränkt und zwar wohl ausnahmslos auf die Antennen.
Am ersten Paare derselben sitzen neben einer bedeutenden Anzahl von
Borsten verschiedener Grösse und Form je nach Art, Geschlecht und
Alter wechselnde Mengen äusserst zarter Gebilde auf, welche kaum
einem anderen Zweck dienen dürften, als die nächste Umgebung auf ihre
chemische Beschaffenheit zu prüfen. Es sind dies die bei den Cyclopiden
und Harpactiden von Leydig beschriebenen sogenannten „blassen
Kolben und Cylinder“, welchen bei unserem Calaniden Diaptomus (s. S.
338) feine und lange verborgen gebliebene Gebilde entsprechen, die
zuerst von Imhof[112] beschrieben und von mir[113]
abgebildet wurden. Besonders auffallend ist die Verschiedenheit der Form
dieser[S. 338] Sinnesorgane bei den Cyclopiden, und wohl wert, mit einigen
Worten beschrieben zu werden. Vor allem ist hervorzuheben, dass die
männlichen Antennen stets reichlich mit solchen Kolben und Cylindern
versehen sind, während sie bei den Weibchen entweder gar nicht oder
nur in der Einzahl angetroffen werden. Der letztere Fall ist um so
interessanter, als die Lebensäusserungen der Tiere keineswegs die
Vermutung aufkommen lassen, als fehle diesen der Geruchssinn überhaupt;
man kann somit schliessen, dass auch noch auf andere, bis jetzt noch
nicht ermittelte Weise geschmeckt bezw. gerochen wird. Diejenigen,
welche ein Sinnesorgan besitzen, tragen dasselbe am zwölften Gliede der
ersten Antenne in Form eines auf einem Stiele sitzenden Kölbchens. Ganz
ähnlich in der Form, nur grösser und zahlreicher sind die Kölbchen, mit
welchen die Männchen dieser Weibchen an den Klammerantennen ausgerüstet
sind. Die übrigen Männchen dagegen schmecken mit den „blassen
Cylindern“, welche morphologisch den eben geschilderten Kölbchen
entsprechen. Diese Cylinder sind im letzten Drittel fein behaart und
kommen auch bei unserem Harpactiden Canthocamptus vor. An den langen
Fühlern unseres gemeinsten Calaniden Diaptomus treten beim Weibchen wie
Männchen ganz kleine, äusserst zarte und darum kaum sichtbare Organe
auf, welche etwa die Form einer Lanzenspitze haben. Einen deutlichen
Übergang von den Borsten zu dem Schmeckorgan weisen die an der Antenne
der Heterocope, eines nahen Verwandten des Diaptomus, auf. Viele
derselben entfernen sich mit Ausnahme einer blasigen Auftreibung am
Grunde kaum vom Bau einer Borste.
Es mag hier erwähnt werden, dass alle die genannten Sinnesorgane, so
verschieden ihre Form und Grösse ist, ganz den so zahlreich an allen
Gliedmassen der Copepoden vorkommenden Borsten, welche an den Antennen
sicher mit die Tastempfindung vermitteln, entsprechen und aus diesen
sich entwickelt haben. Nicht allein die Anordnung der „blassen Kolben
und Cylinder“ auf den Gliedern der Antenne dient uns als Beweis dafür,
sondern auch der Umstand, dass, wie beim ebenerwähnten Falle, nicht
selten solche Sinnesorgane angetroffen werden, welche ein Mittelding[S. 339]
zwischen Borste und Kolben (oder Cylinder) darstellen. Es ist dies
ein Beispiel, dass eigentlich Geschmack und Geruch nur verfeinerte
Tastempfindungen sind. Weitere Bemerkungen über die Thätigkeit der
vermutlichen Organe des Geschmacks- und Geruchssinnes werden in dem
Abschnitt über die Lebensweise der kleinen Kruster folgen und wir
wenden uns zur Besprechung des Gehörs, sofern man überhaupt
von einem solchen reden kann. Es ist nämlich bis jetzt nicht gelungen
an Süsswasser-Copepoden auch nur in Spuren ein Organ nachzuweisen,
welches zur Aufnahme von Schallwellen im Wasser spezifiziert wäre.
Wollen wir nicht annehmen, dass die oben geschilderten Sinnesorgane
der Antennen — zu denen ja schliesslich noch alle die feineren
Borsten, namentlich die mit feinen Härchen versehenen zu rechnen sind
— neben den angegebenen Funktionen noch die des Hörens übernehmen, so
können wir nur vermuten, dass Schallwellen, die ja im Wasser ohnedies
viel besser geleitet werden als in der Luft, vom ganzen Körper des
Tieres empfunden werden und somit ebenso wirken, wie ein Reiz auf den
Tastsinn. Da nicht einzusehen ist, von welchem Vorteil eine auch nur
annähernd genaue Unterscheidung der Töne für das Wohlbefinden eines so
niedrig organisierten Wesens sein soll, so wird man wohl begreifen,
dass die geschilderte Art der Schallempfindung ihren Zweck ausreichend
erfüllt. Auch wir empfinden ja den Schall viel öfter als wir es ahnen
zumteil mittels des Tastsinnes. Jeder kann sich davon überzeugen,
wenn er während des Vortrages eines kräftigen Gesangsstückes oder
einer schmetternden Kapelle die Fingerspitzen auf den Deckel eines
frei gehaltenen Buches oder einer kleinen Zigarrenkiste legt. Dass
aber die Copepoden einen gut entwickelten Tastsinn haben, wird
jeder Freund derselben, welcher etwa mit einer Pipette die gewandteren
Schwimmer, wie Diaptomus und Cyclops, aus einem grösseren Gefäss
herausfangen will, zu seinem Ärger erfahren. Mit einer oft geradezu
bewundernswerten Geschicklichkeit wissen die Tiere dem gefahrbringenden
Instrumente, dessen Nähe sie offenbar nicht sehen, sondern lange,
ehe es den Körper berührt, fühlen, auszuweichen, und je öfter und
sorgfältiger man ihnen nahe[S. 340] zu kommen sucht, desto sicherer entfliehen
sie — einmal scheu gemacht — durch einen schnellen Sprung.
Von den Eingeweiden ist am leichtesten der Darmtraktus,
besonders am lebenden Tiere — allerdings so wenig wie das Nervensystem
mit blossem Auge — zu verfolgen. Wir teilen denselben in einen vordern
(Schlund-)Teil, Mittel-(Magen) und Enddarm. Der erste beginnt mit dem
Munde. Von der Mundöffnung an steigt der Schlund beinahe senkrecht in
die Höhe, biegt ein wenig nach vorn der Stirn zu um, erleidet dort eine
Knickung und geht allmählich in eine Erweiterung des Darmrohres, den
Mitteldarm oder Magen, über, welcher sich annähernd durch den ganzen
Vorderleib erstreckt. Mit dem Beginn des Hinterleibs fängt der wieder
dünnere Enddarm an und mündet an der Oberseite des letzten Segmentes
vor der Furca nach aussen. Der Darmtraktus geht somit wie ein gerader
Schlauch mitten durch den Körper. Am Enddarm besonders kann man leicht
eine eigentümliche Erscheinung beobachten, welche dem Darm aller Tiere,
den Menschen nicht ausgenommen, eigen ist. Häufig treten nämlich an
demselben Einschnürungen auf, welche weiter wandern und, da sie rasch
auf einander folgen, wie Wellen sich fortzupflanzen scheinen. Es sind
dies die sogenannten „peristaltischen Bewegungen“, welche, durch die
in der Darmwand liegenden Muskeln erzeugt, der Weiterbeförderung des
Inhalts dienen.
Vergebens würden wir nach Leber und Niere, zwei so
hochwichtigen Organen im tierischen Körper, suchen, wollten wir sie
als gesonderte Teile der Eingeweide vermuten. Nach mühsamen Versuchen
erst ist es gelungen nachzuweisen, dass Zellen, auf dem Magenstück
liegend, die Verrichtung der erwähnten Drüsen übernommen haben. Etwa
innerhalb des Kopfbruststückes ist der Darm von drüsigen Zellen
umgeben, welche mit gelb gefärbten Fettkügelchen erfüllt und ihrem Bau
nach als Leberzellen zu deuten sind. Weiter nach hinten, dem Ende des
Magens zu, enthalten ähnlich geformte Zellen die Stoffe, welche bei
den höheren Tieren von den Nieren abgesondert werden. Durch Einwirkung
chemischer Reagentien lässt sich ihre Zusammensetzung erkennen.[S. 341] Diese
Nierenzellen geben ihren Inhalt am Anfang des Enddarms an diesen zur
Weiterbeförderung nach aussen ab.
Selbst unser Regenwurm ist in dieser Beziehung, so niedrig er sonst
steht, besser eingerichtet, als der Copepode, indem er wenigstens
gesonderte Organe, welche die Funktion der Niere verrichten, besitzt.
Allein nicht nur in diesem einen Falle können wir am Copepoden die
Einfachheit der Mittel bewundern, mit denen die Natur so unendlich
verschiedenes zweckmässig einzurichten weiss. Gleich die Art und
Weise, wie das Blut im Körper der kleinen Geschöpfe zu allen Organen
hingeführt wird, ist bei den Cyclopiden und Harpactiden so originell
und zugleich so primitiv, dass wir kaum begreifen können, wie dadurch
die Thätigkeit eines komplizierten Apparates, wie es das Herz anderer
Tiere ist, vollkommen ersetzt werden kann.
Die beiden eben erwähnten Familien besitzen nämlich nicht einmal die
Andeutung eines Herzens und bei diesen Tieren übernimmt nun
wunderbarerweise der Darm einen Teil der Funktion eines solchen,
ist also in dreifacher Weise beschäftigt. Dadurch, dass er sich in
bestimmter Richtung (vorwiegend in der Vertikalebene auf und ab)
bewegt, wird das Blut wenigstens ordentlich untereinandergeschüttelt,
wenn auch nicht in vorgeschriebenen Bahnen oder Adern herumgeleitet.
Die Lage und der Verlauf des Darmes wurde vorhin geschildert. An der
erwähnten Umbiegung, wo der Schlund in den Magen übergeht, setzen sich
an der Oberseite feine Muskeln an, welche nach vorn und oben gegen
die Stirne zu verlaufen. Diese bewegen bei ihrer Zusammenziehung den
Darm in dieser Richtung. Sehr ausgiebig und regelmässig ist diese
Bewegung nicht. Eine zweite Art von „Darmpulsationen“ vollzieht sich
durch die Thätigkeit anderer Muskeln, welche sich einerseits am Darm
beim Übergang des Magens in den Enddarm ansetzen, anderseits an der
innern Bauchwand des Hinterleibes. Durch diese Muskulatur wird der
Darm gewaltsam nach abwärts und zugleich nach hinten gerissen und
alles Blut, das sich unter ihm befand, nach oben an den Seiten des
Darmes vorbei unter die[S. 342] Rückenhaut getrieben. Gleich darauf kehrt
alles in seine vorige Lage zurück. Die Zahl dieser Bewegungen in einer
bestimmten Zeit ist manchem Wechsel unterworfen und scheint zumteil
willkürlich ausgeführt zu werden. Die Gliedmassen sorgen ausserdem
durch ihre fast immerwährende Bewegung jedenfalls mit dafür, dass das
Blut keinen Augenblick ruhig im Körper verweilt.
Eine viel vollkommenere Blutzirkulation findet bei den Calaniden
statt, da diese ein Herz besitzen. Dieses ist bei unserm Diaptomus
in einer ausserordentlich lebhaften Bewegung und schlägt etwa 150mal
in der Minute, so rasch, dass es beinahe zu vibrieren scheint. Durch
seine energische Thätigkeit werden die in seiner Umgebung liegenden
Organe ebenfalls in einem beständigen Zittern erhalten. Beobachtet
man einen Diaptomus längere Zeit unter dem Mikroskop von einem
dünnen Gläschen bedeckt, so verbraucht er allmählich den in dem
umgebenden Wassertropfen enthaltenen Sauerstoff und dann schlägt das
Herz des absterbenden Tieres immer langsamer, so dass es sich leicht
genau besehen lässt. Man wird dann finden, dass es einem kleinen
Balle gleicht, welcher in der Mitte des Rückens und zwar im dritten
Vorderleibssegment direkt unter der Haut liegt. Feine blasse Fädchen
ziehen von ihm an die umgebenden Teile (Haut, Darm) und halten es in
seiner Lage fest. Es ist mit einigen Öffnungen versehen. Am sichersten
ist eine hintere und vordere, weniger bestimmt solche an den Seiten
beobachtet worden. Blutgefässe fehlen auch hier. Wie bei den zwei
anderen Familien ist auch bei den Calaniden die ganze Körperhöhle mit
Blut erfüllt, so dass die Organe sozusagen darin schwimmen. Zieht
das Herz sich zusammen, so strömt das Blut gegen den Kopf zu aus der
vordern Öffnung, welche demnach als „arterielle“ angesehen werden kann.
Die bei der Kontraktion sich schliessende hintere oder „venöse“ Öffnung
lässt bei der Ausdehnung Blut einströmen, welches von seiner Wanderung
durch den Körper und die Gliedmassen zurückkehrt.
Das Blut selbst ist farblos oder schwach gelblich. Feste
Bestandteile, etwa den Blutkörperchen anderer Tiere entsprechend,[S. 343]
lassen sich nur sehr schwer darin entdecken und wurden früher ganz
vermisst.
Gesonderte Verrichtungen, welche zur Atmung dienen könnten,
fehlen allen echten Copepoden vollständig. Man muss demnach annehmen,
dass der im Wasser gelöste Sauerstoff, der auch diesen kleinen Wesen
unentbehrlich ist, einfach durch die Haut aufgenommen und auf demselben
Wege die Kohlensäure abgeschieden wird. Man wird diese scheinbare
Unvollkommenheit verstehen, wenn man bedenkt, wie leicht sich ein
Gasaustausch durch eine Haut, welche in allen Teilen dünner ist als die
Kiemenhaut grösserer Krebse, vollziehen kann. Es wirkt somit die ganze
Körperoberfläche als Atemorgan, sozusagen als Kieme. Das Bedürfnis
nach Sauerstoff entspricht der Grösse der Tiere und der Lebhaftigkeit
derselben.
Alle Bewegungen im und am Körper werden durch hochentwickelte
Muskeln ausgeführt. Vor allem sind zwei Stränge derselben,
je einer links und rechts von der Mittellinie des Körpers, stark
ausgeprägt und leicht zu sehen. Diese vermitteln die Verschiebung der
Körpersegmente gegen einander. An den Seiten des Körpers und am Bauche
setzen sich die Muskeln der Gliedmassen an. Von der eigentümlichen
Thätigkeit der Darm- und Herzmuskulatur war schon früher die Rede.
Es bliebe uns jetzt noch übrig, da ja nur die wichtigeren Organe
hier Raum finden können, die Fortpflanzungsorgane und daran
anknüpfend die Entwickelung der Copepoden zu besprechen.
Schon bei der Beschreibung der Gliedmassen wurde erwähnt, dass alle
Copepoden getrennten Geschlechts seien, und die äusseren Merkmale,
welche beide Geschlechter trennen, hervorgehoben.
Die Fortpflanzungsorgane nehmen bei den reifen Tieren einen
grossen Teil der Leibeshöhle ein und schimmern namentlich bei den
Cyclopiden durch die dünne Körperwand durch. Da der Inhalt derselben
oft genug eine bläuliche bis grüne Färbung besitzt, erscheinen die
Tiere in dem entsprechenden Tone gefärbt. Die[S. 344] Männchen sind sofort
an ihrem schlankeren Körper und den umgeformten Antennen zu erkennen.
Junge Tiere sind nicht immer leicht zu unterscheiden. Die Weibchen sind
gewöhnlich grösser als die Männchen derselben Art und der Hinterleib
derselben besitzt ein Segment mehr als bei jenen, da das erste und
zweite nicht mit einander verwachsen sind. Bei beiden Geschlechtern
liegen die Fortpflanzungsorgane über dem Darm und unter der Rückenhaut.
Wo ein Herz, wie bei den Calaniden, vorhanden ist, befindet sich dieses
über den Geschlechtsorganen. Sind letztere bei den Weibchen stark
entwickelt und mit Eiern erfüllt, so erstrecken sie sich etwas an den
Seiten des Darmes nach dem Bauche zu abwärts. Die Keime werden in einer
sackähnlichen Drüse erzeugt. Von dieser Drüse treten sie nach rechts
und links in die Eileiter über, welche bei grossen Weibchen durch ihre
eigentümliche Form (Fig. 70 Ee) und den gefärbten Inhalt leicht
sichtbar werden. Die Eileiter reichen weit nach vorn und hinten bis zum
ersten Segment des Hinterleibes, wo sie nach aussen münden. Nach den
Seiten senden dieselben ebenfalls mit Eiern erfüllte Lappen aus. In den
Eileitern werden die Keime (Eier) mit Dotter versehen und erhalten auch
eine zarte Hülle. Die Eier geniessen bis zum Auskriechen der Jungen die
mütterliche Fürsorge und werden nicht einfach in das Wasser entleert.
Vor dem Verlassen des mütterlichen Körpers werden sie nämlich aus einer
Drüse mit einem Klebstoff umhüllt, welcher im Wasser erhärtet und die
Eier unter einander sowohl, als auch mit den Seiten des mütterlichen
Körpers am ersten Ringe des Hinterleibes verkittet. Solche Säckchen
tragen die Cyclopiden zwei, deren jedes etwa mit zwanzig Eiern erfüllt
ist, die Harpactiden und Calaniden dagegen nur eines. Bei ersteren
sind sie oval und links und rechts am Hinterleib, bei letzteren mehr
rundlich und an der Unterseite befestigt. Diese Säckchen sind bei den
Copepoden, die der pelagischen Fauna angehören, stets kleiner als bei
den Uferformen.
Die männlichen Organe sind bei den Cyclopiden mit Ausnahme der
Keimdrüse ebenfalls paarig angelegt, bei den Calaniden und Harpactiden
dagegen nicht. Die Geschlechtsprodukte werden in sogenannten
Spermatophoren an das Weibchen angeheftet. Den[S. 345] Cyclopiden sind solche
Gebilde von rundlicher Form eigen, bei den Calaniden und Harpactiden
stellen sie jedoch langgezogene Flaschen von sehr komplizierter
Einrichtung dar. Ihre Entstehung und Funktion wurde von A.
Gruber vorzüglich geschildert.
Die Befruchtung der Eier findet am Ende der Eileiter während des
Ablegens statt.
In den Säckchen sind, wie im Körper, die Eier von grünlicher bis
bläulicher, selten gelber Farbe. Eine dünne Haut umschliesst den
körnigen fetthaltigen Inhalt. Bald nach der Bildung der Eiersäckchen
beginnt die im Ei schlummernde geheimnisvolle Kraft in der Umbildung
des Dotters zu einem neuen Wesen ihre Wirkung zu entfalten. So
interessant die hierbei sich abspielenden Vorgänge sind, so können sie
hier nicht weiter geschildert werden. C. Claus[110] hat uns
schon vor langen Jahren mit denselben bekannt gemacht. Nach einer,
je nach der Jahreszeit, zwei bis zehn Tage dauernden embryonalen
Entwicklung entschlüpft dem Ei der junge Copepode, welcher zunächst
mit seinen Eltern so wenig Ähnlichkeit hat, dass man ihn lange für ein
ganz besonderes Tier hielt und ihn „Nauplius“ benannte. Alle jungen
Copepoden und auch die meisten übrigen niederen Krebse haben diese
Entwickelungsform des „Nauplius-Stadiums“ gemeinsam, so verschieden
im erwachsenen Zustande die Tiere auch aussehen mögen. In diesem
Jugendkleide stellt sich das kleine Wesen als rundlicher Körper mit nur
drei Gliedmassenpaaren dar, welche an der Bauchseite entspringen und
in der Form ebenfalls wesentlich von denen der Alten abweichen. Die
weitere Entwickelung vom Nauplius zum vollkommenen Tiere geht sozusagen
ruckweise vor sich. Die kleinen Kruster wachsen so wenig wie andere
Kerftiere etwa ähnlich den höheren Tieren allmählich heran, wobei der
schon in seiner Gestaltung fertige Körper eigentlich nur an Grösse
zunimmt, sondern in Pausen. Von Zeit zu Zeit wird, wie bereits früher
beschrieben wurde, den Jungen die Haut zu enge, deren Beschaffenheit
eine Dehnung nur bis zu einem gewissen Grade zulässt. Sie, d. h. die
unbelebte äussere Schicht derselben, wird deshalb abgeworfen und durch
eine neue ersetzt. Mit jedem Wechsel der Haut treten am Körper mehr
Segmente und Gliedmassen,[S. 346] an diesen aber, so weit sie nicht schon
früher vorhanden waren, neue Bestandteile auf. Das Auge, den Darm und
— wo ein solches vorkommt — auch das Herz bringen die Copepoden, wie
die erste Anlage der Fortpflanzungsorgane, mit auf die Welt. Mit dem
Verlassen des Eies entzieht sich der Nauplius der mütterlichen Obhut
und geht von nun an selbständig seinen Weg durchs Leben.
Es ist dieses Nauplius-Stadium für die Systematik der niederen
Krebse um so wichtiger, als manche erwachsene Formen selbst für das
geübte Auge so wenig Ähnlichkeit mit den nächsten Verwandten haben,
dass man ohne Kenntnis der Entwickelung sie lange mit ganz anderen
Tieren zusammenstellte. Es sind solche weitgehenden Veränderungen der
äussern Gestalt namentlich bei den Rankenfüssern und den parasitischen
Copepoden beobachtet und auf eine Anpassung an die Lebensweise
zurückzuführen. Einige Rankenfüsser setzen sich fest, werden mit
einer harten Kalkschale umschlossen und scheinen in dieser Form alle
Anschauungen über den Bau eines Krebses über den Haufen werfen zu
wollen. In der That wird jeder, welcher zum ersten Mal eine sogenannte
Entenmuschel oder die Seepocken zu Gesicht bekommt, Mühe haben, sie als
Krebse zu erkennen. Von den parasitischen Copepoden werden wir
später noch einiges erfahren.
Nach einer bestimmten Zahl von Häutungen hat das Tier seine endgültige
Grösse und Form erreicht und ist fortpflanzungsfähig geworden. Die
Zeit, die es vom Verlassen des Eies an zu seiner weiteren Entwickelung
nötig hat, ist je nach den äusseren Einflüssen verschieden. Im Sommer
genügen nach Jurines[114] Beobachtungen zwei bis drei Wochen,
während in der kälteren Jahreszeit unter Umständen ebensoviele Monate
nötig sind, bis das Tier fertig ist. Die erreichte Grösse ist bei
Individuen einer Art meist annähernd gleich. Nur ausnahmsweise findet
man solche, welche das normale Mass um vieles (fast ⅓) übersteigen,
ohne dass eine besondere Ursache für ein so auffallendes Wachstum sich
nachweisen liesse.
So interessant das Studium der einzelnen Familien, Gattungen und
Arten, welche unsere süssen Wasser bewohnen, ist, so muss[S. 347] dennoch
auf eine Aufzählung aller bis jetzt beschriebenen Formen verzichtet
werden. In dem angefügten Litteraturverzeichnis sind jedoch für solche,
welche sich eingehender mit den interessanten Tieren abgeben wollen,
die wichtigsten Werke namhaft gemacht, in denen die Beschreibung
und Klassifikation der Copepoden behandelt wird. Wir wenden uns zu
der am niedersten stehenden Familie der Eucopepoden mit kauenden
Mundwerkzeugen, den
Cyclopiden.
Der ganze Körper (Fig. 70) setzt sich (ohne die Furca) aus zehn Ringen
zusammen, von denen fünf auf den Vorderleib, ebensoviele (beim Weibchen
verwachsen die zwei ersten) auf den Hinterleib kommen. Der Vorderleib
ist meistens beträchtlich breiter als der Hinterleib und eiförmig. Die
ersten Antennen setzen sich aus 8–17 (selten 18) Gliedern zusammen und
sind beim Männchen beide zu Greiforganen umgewandelt. Das Auge ist in
der Mitte der Stirne gelegen und hat zwei Linsen. Das Weibchen trägt
seine Eier in zwei gewöhnlich etwas abstehenden Säckchen an beiden
Seiten des Hinterleibs.
Am gemeinsten von allen den Arten, deren Antennen 17gliederige Fühler
besitzen, ist der Cyclops viridis Fischer. Die Fühler dieser
grossen Art sind kaum länger als das erste Vorderleibssegment und
gedrungen; sie tragen am zwölften Gliede ein feines Sinneskölbchen.
Fast gleich häufig trifft man den kleinen Cyclops agilis
Koch an, welchem von Claus, da er am Aussenrande der Furca
eine kleine Säge trägt, der Name serrulatus beigelegt wurde.
Seine Fühler sind schlank und reichen bis zum vierten Segment des
Vorderleibs. Sie besitzen zwölf Glieder, ermangeln aber eines
Sinneskölbchens. Die Eiersäckchen werden sehr abstehend getragen. Von
grossen Cyclopiden mit langen Antennen (viel länger als das erste
Vorderleibssegment) sind etwa noch Cycl. tenuicornis Claus und
signatus Koch zu erwähnen. Diese sehen sich im allgemeinen
ähnlich, allein während C. tenuicornis wie C. viridis
ein Sinneskölbchen an den ersten Antennen trägt, fehlt dieses dem
C. signatus stets. Die Länge von diesen beiden[S. 348] Arten und von
viridis beträgt etwa 3.5 mm. Ein naher Verwandter des
C. agilis interessiert uns durch sein beschränktes Vorkommen. Er
wurde in den Maaren der Eifel von Dr. Otto Zacharias gefunden
und erhielt bei der Beschreibung[115] den Namen C. maarensis[XVII].
Es giebt noch eine bedeutende Anzahl von meist kleinen Arten mit
17gliederigen Antennen, welche kaum länger als das Kopfbruststück sind.
Alle sind aber sehr schwer zu bestimmen, da die Merkmale nur mit Mühe
aufgefunden werden können. Viel seltener trifft man Cyclopiden an,
deren Antennen weniger als zwölf Glieder besitzen. Einer derselben,
C. canthocarpoides, kommt auch im Meere vor.
[XVII] Es ist diese Art nach neueren Untersuchungen vielleicht
mit C. macrurus Sars identisch.
Fig. 71. Canthocamptus minutus Müller, nach Brady (Weibchen von der
Seite).
Die Familie der
Harpactiden
zählt im süssen Wasser zu der am wenigsten bedeutenden. Wie
Cyclops die einzige Gattung unter den Cyclopiden darstellt, so
sind auch die Harpactiden nur durch eine Gattung, Canthocamptus
(Fig. 71), vertreten. Diese Form ist in mehreren Arten sehr weit
verbreitet. Sie wird besonders im ersten Frühjahr in Gesellschaft
von Cyclops in allen unseren kleinen Weihern gefunden, viel
seltener oder gar nicht im Sommer. Der Körper von Canthocamptus
weist dieselbe Gliederung auf, wie die von Cyclops, jedoch ist
Vorder- und Hinterleib[S. 349] an Breite nur wenig verschieden und beide
sind nicht sehr scharf von einander abgesetzt. Die ersten Antennen
sind kürzer als das Kopfbruststück und beim Männchen zu Greiforganen
umgestaltet. Bei letzterem erleidet das dritte Fusspaar eine demselben
Zwecke dienende Formveränderung. Am Hinterleib der Weibchen wird nur
ein Eiersäckchen angetroffen, neben welchem häufig die langen
flaschenförmigen Spermatophoren hängen. Canthocamptus ist ein
schlechter Schwimmer und dreht sich bei seinen Bewegungen um seine
eigene Achse schraubenförmig durch das Wasser. Die grösste bei uns
vorkommende Art — Canth. minutus Müller oder staphylinus
Jurine — misst etwas über einen Millimeter; die meisten anderen sind
noch kleiner. Viel wichtiger und grösser ist die dritte Familie der
Süsswassercopepoden, die der
Calaniden,
obgleich auch sie nur mit wenigen Gattungen bei uns vertreten ist.
Diese sind nicht so allgemein verbreitet, wie die beiden vorhergehenden
Familien, sie kommen aber meist in grossen Massen vor, wenn sie einmal
in einem Wasser eingebürgert sind. Der Vorderleib der Calaniden ist
länger als der Hinterleib und bildet ein langgezogenes, nach hinten
abgestutztes Oval. Der ganze Körper ist annähernd cylindrisch.
Die meist 25gliederigen Antennen sind etwa so lang als der ganze
Körper. Die rechte ist beim Männchen wie auch der rechte Fuss des
fünften Beinpaares zu einem Greiforgan umgestaltet. Das Weibchen
erzeugt nur ein Eiersäckchen, welches wie bei Canthocamptus
an der Unterseite des Hinterleibs klebt. Die am Ende der Furca
aufsitzenden Borsten haben ziemlich gleiche Länge und sind fächerförmig
ausgebreitet. Die hiehergehörigen Gattungen und Arten wurden in einer
sehr ausführlichen Arbeit von Jules de Guerne und Jules
Richard[116] zusammengestellt. Hiernach werden neun bis zehn
Gattungen mit über 70 Arten unterschieden. Am meisten Bedeutung hat
das Genus Diaptomus, von welchem allein an die 60 Vertreter
beschrieben wurden. Gewöhnlich findet man bei uns drei Arten an,
d. h. nicht alle drei beisammen. Die grösste Art ist Diaptomus
castor Jurine,[S. 350] welcher über 3½ mm lang wird und
meistens kleinere Gewässer bewohnt. Diesem sehr ähnlich, jedoch
kleiner, ist D. coeruleus Fischer. Eine fast in allen grossen
Wasserbecken auftretende und als Fischnahrung sehr wichtige Art ist
der pelagisch lebende zierliche D. gracilis, welcher fast keine
Färbung hat und ganz wasserhell ist. Dem letzteren gleicht eine durch
Lilljeborg entdeckte und früher nur im Norden gefundene Art,
Diaptomus graciloides (Fig. 72), welche in einem der schon bei
Cyclops maarensis erwähnten Maare der Eifel, in dem Gemünder
Maar und zwar nur in diesem ebenfalls durch Zacharias gefunden,
sonst aber bis jetzt nirgends in Deutschland angetroffen wurde. Die
Gattung Heterocope bewohnt auch nur grosse Wasserbecken und
zeichnet sich durch bedeutende Grösse und kräftigen Bau aus. Von den
selteneren und wie es scheint mehr dem Norden angehörigen Gattungen
wäre noch als in Deutschlands Nordwesten vorkommend Eurytemora
Poppe zu erwähnen.
Fig. 72. Diaptomus graciloides Lilljeborg. a′ Erste Antenne — a″
Zweite Antenne — E Eiersäckchen.
Alle Calaniden sind vorzügliche Schwimmer. Sie lieben es, häufig mit
dem Rücken nach abwärts gekehrt sich im Wasser zu[S. 351] bewegen. Nicht
allein mit Füssen und Antennen vollziehen sie letzteres, vielmehr
erzeugt auch die rasche Thätigkeit der Mundwerkzeuge, welche zum Zweck
der Nahrungszufuhr ausgeübt wird, ein etwas langsames Fortgleiten.
Bei Diaptomus wurde beobachtet, dass er nachts in einen lethargischen
Zustand, eine Art Schlaf, verfalle.
Die Nahrung aller freilebenden Copepoden besteht in kleinen
Teilchen tierischer und pflanzlicher Substanz, wie sie sich auf dem
Grunde der Gewässer, an Pflanzenstengeln und so weiter vorfindet. Vor
allem scheinen Urtiere und kleine Algen aufgenommen zu werden. Da stets
beides zusammen vorkommt, ist es schwer zu entscheiden, ob die Tiere
in der That Allesfresser sind oder ob der eine oder andere Bestandteil
etwa zufällig in den Darm gelangt. Nach einigen Beobachtungen fressen
die Mütter ihre eigenen Kinder, und dieses kannibalische Betragen würde
die Copepoden als Räuber oder reine Fleischfresser kennzeichnen. Es ist
sehr unterhaltend, den Tieren bei der Suche nach Nahrung zuzusehen.
Wenn der sie beherbergende Behälter etliche Pflanzen enthält, weiden
sie mit Vorliebe den daran sitzenden Detritus unter pickenden
Bewegungen regelrecht ab. Ist dann der Magen mit einem genügenden
Vorrat von Speisen versehen, so sucht das Tier wieder die Gesellschaft
der Genossen auf. Diese ist allen Arten, wie es sich schon aus der Art
des Vorkommens in grossen Scharen ergiebt, geradezu Bedürfnis. Mit
munteren Sprüngen haschen und jagen sich Alte wie Junge gegenseitig
und scheinen dies nur der Unterhaltung wegen zu thun. Mitten durch
dieses lebhafte lautlose Gewimmel stürzen dann oft die von den kleinen
lebhaften Männchen verfolgten Weibchen in wilden Sätzen. Eine Zeitlang
geht die Jagd durch das Gewirre der Wasserpflanzen, dann wieder über
freiere Stellen, bis das Weibchen durch eine geschickte Wendung dem
Verfolger sich entzieht oder, müde geworden, auf Gnade und Ungnade sich
ergiebt.
Schon früher wurde angedeutet, dass wir nur ausnahmsweise in einer wenn
auch noch so kleinen Wasseransammlung, falls sie nur[S. 352] schon einige
Zeit bestand, vergebens nach Copepoden fahnden. In Grönland so gut
wie unter den Tropen sind namentlich die stagnierenden oder langsam
fliessenden Gewässer oft in staunenerregender Weise damit bevölkert
und von dem enormen Reichtum des Meeres an diesen Tieren können wir
uns kaum eine Vorstellung machen. Ich selbst habe einen Fall erlebt,
wo ein kleiner Weiher, dessen auffallende Armut an Pflanzenwuchs
keine reiche Ausbeute versprach, durch die Masse einer einzigen
Copepoden-Art (Diaptomus coeruleus) geradezu gelb gefärbt war.
Mehrfach werden ähnliche Beobachtungen erwähnt, und manche Nachricht,
die für die auffallende Färbung eines Gewässers keinen Grund angiebt,
dürfte auf das massenhafte Vorkommen von Ruderfüssern (vielleicht im
Verein mit Daphnien) zurückzuführen sein. Ganze Wolken der ersteren
färben zu gewissen Zeiten weite Strecken des Meeres und die Fischer
bezeichnen diese Erscheinung als „Rotäsung“ und kennen sie als Vorboten
reichlichen Fanges.
So wenig als in horizontaler Richtung scheinen auch in vertikaler
dem Vorkommen der Ruderfüsser Grenzen gezogen zu sein, sofern
überhaupt Wasser vorhanden ist. Auf dem Kamme des Riesengebirges fand
Zacharias[117], in 2500 m
Meereshöhe Imhof[118]
(im Val di Brutto) noch Copepoden vor, während sie anderseits in
unseren grossen Binnenseen und im Meere die grössten bis jetzt
erforschten Tiefen noch beleben und einen wesentlichen Bestandteil der
in ewigem Dunkel lebenden Fauna ausmachen. Die reichste Abwechslung
in der Beschaffenheit ihrer Wohnorte ertragen entschieden die
Süsswasser-Copepoden. Die Temperatur des Wassers scheint nur insofern
von Einfluss auf die kleinen Körper zu sein, als im Winter unter
der Eisdecke die Vermehrung langsamer vor sich geht. Versuche haben
bewiesen, dass selbst durch längeres vollständiges Einfrieren die
Lebenskraft nicht erlischt. Wenn wir gerade im Winter und im Frühjahr,
nachdem die Macht der wiederkehrenden Sonne die Eisdecke weggetaut hat,
am meisten Ruderfüsser in unseren Tümpeln und Weihern antreffen, so ist
dies mehr dem Umstande zuzuschreiben, dass die Mehrzahl ihrer Feinde
entweder im vollständigen Winterschlaf liegt oder doch in einem solch[S. 353]
lethargischen Zustande sich befindet, dass das Bedürfnis nach Nahrung
nur ein geringes ist. Bei der allgemeinen Verbreitung kann es uns nicht
Wunder nehmen, wenn unter den Mitgliedern der Höhlenfaunen ebenfalls
Copepoden erwähnt werden. Zu den seltneren Fällen gehört es, dass ein
Angehöriger des süssen Wassers im Mineral-, Brack- oder gar Meerwasser
angetroffen wird. Überraschend ist dies bei der Anspruchslosigkeit
unserer Tiere und deren Widerstandsfähigkeit gegen äussere Einflüsse
nicht. Diese ebengenannten Eigenschaften im Verein mit der grossen
Vermehrung und raschen Entwickelung sind die einzigen Mittel, welche
den Ruderfüssern zu einem erfolgreichen Kampfe ums Dasein zur Verfügung
stehen. Schon innerhalb kleiner Wasserbecken sind die meisten Arten
manchmal gezwungen, von einer weitern Eigenschaft, welche für
gewöhnlich nicht in die Augen springt, Gebrauch zu machen: ich meine
die Anpassungsfähigkeit.
Wie im Meere, so kann man auch in unseren Seen dreierlei Regionen
nach den physikalischen und organischen Verhältnissen unterscheiden.
Jede derselben ist durch spezifische Tier- und Pflanzenformen von
den anderen verschieden und dies tritt gerade bei den niederen
Krebstieren am deutlichsten zu Tage. Wollen diese waffenlosen
Wesen in der Konkurrenz mit anderen Tierformen nicht unterliegen,
so müssen sie sich, ob sie nun in der Uferregion oder pelagisch
oder gar in der Tiefenregion leben, den jeweiligen Verhältnissen
anpassen. Es lässt sich schon zum voraus aus der Beschaffenheit der
Lokalität entnehmen, dass z. B. die reich mit Pflanzen bewachsene
Uferzone mit seichtem, warmem Wasser den Tieren bedeutend günstigere
Lebensbedingungen gewähren wird, als die scheinbar leblose Mitte des
Sees mit ihren verborgenen Tiefen. Dort kann ein Wesen auf engem Raum
seine Nahrung zusammenlesen und es hat nicht viel zu sagen, wenn
seine Schwimmwerkzeuge nicht erster Güte sind. Die reiche Ernährung
ermöglicht ein rasches Wachstum und reichliche Fortpflanzung. Die
kleinen Kruster müssten sich ins Unendliche vermehren und somit die
oben erwähnte Erscheinung, dass das Wasser durch ihre Massen sich
färbt, viel öfter verursachen, wenn nicht auch hier ein gewichtiges[S. 354]
Regulativ entgegenwirken würde. In dem so reich belebten seichten
Wasser haben nämlich die grössten Feinde der kleinen Krebse ihren
Wohnsitz aufgeschlagen und verschlingen unzählbare Summen derselben.
In einem aus verschiedenen Stoffen zusammengeklebten Häuschen
lauert die Larve der Köcherfliege, an den Pflanzenstengeln die der
Eintagsfliegen, im Schlamm halb versteckt die Libellenlarve mit ihrem
heimtückischen Fangapparat auf lebende Beute. Muntere junge Fischchen
schnappen spielend ganze Massen von Entomostraken weg und diesem
Vernichtungskriege schliesst sich auch der verschmitzt lauernde,
langsam die Beute erschleichende junge Triton an. Auf Schritt und
Tritt lauert Tod und Verderben, und wenn nicht die gütige Mutter Natur
immer reichliche Nachkommenschaft an der Stelle der im ungleichen
Kampfe Gefallenen eintreten liesse, so wären die Copepoden samt ihren
Verwandten schon längst aus der Liste der Lebewesen gestrichen. In der
Uferregion also kann der Ruderfüsser sich nur dann erhalten,
wenn er sich rasch und reichlich vermehrt. Mit Waffen seiner Feinde
sich zu erwehren erlaubt ihm seine natürliche Ausrüstung nicht.
Ganz anders gestaltet sich das Leben und Treiben unserer Tiere in der
sogenannten pelagischen Zone der Seen bei einer Wassertiefe
von mindestens 15–20 m und einer oft mehrere hundert Meter
betragenden Entfernung vom Ufer. In dem beinahe immer klaren Wasser
fehlt, abgesehen von einigen winzigen Algen und Diatomeen, jeder
Pflanzenwuchs. Von dem von Urtieren reich belebten Detritus der
Uferregion führen Wind und Strömung nur noch Spuren hinaus und
diese bilden, auf weite Strecken verteilt, eine kärgliche Nahrung.
Temperaturschwankungen und Wellenschlag werden in der Tiefe,
welche die Tiere gewöhnlich, namentlich während des Tages, bewohnen,
kaum mehr fühlbar; im übrigen sind jedoch die Verhältnisse für einen
vom Ufer her in diese Region verschlagenen Copepoden so ungünstig als
möglich, zumal auch jede Deckung fehlt, welche das Tier vor seinen
Feinden schützen könnte. Wenn wir trotzdem eben in der pelagischen
Region eine reiche Fauna von Entomostraken antreffen, so ist dies
einzig und allein der weitgehenden Anpassung, welche sich Generationen[S. 355]
hindurch während eines nicht allzukurz zu bemessenden Zeitraumes im Bau
der Kruster vollzog, zuzuschreiben. Als die auffallendste Eigenschaft,
welche alle pelagisch lebenden Krebstiere — auch die des Meeres —
auf dem Wege der natürlichen Zuchtwahl erworben haben, bewundern wir
deren ausserordentliche Durchsichtigkeit und Farblosigkeit. Hierdurch
werden auch die Copepoden dem Auge ihrer Feinde entzogen, denen
sie bei dem Mangel an Verstecken und der den Verfolgern gegenüber
geringen Schnelligkeit auf der Flucht kaum zu entrinnen vermöchten.
Haut, Muskulatur und Nervensystem sind gewöhnlich wasserhell; nur der
Darm (durch seinen Inhalt) und die Geschlechtsorgane zeigen hie und
da noch Färbung. In zweiter Linie macht sich die Anpassung bei der
Fortpflanzung bemerklich; denn alle Copepoden tragen selten mehr als
vier Eier auf einmal in den Eiersäckchen mit sich herum. Der mühsame
Erwerb der im Wasser sehr verteilten Nahrung wirkt einer reichen
Fruchtbarkeit entgegen. Ausserdem sind die Tiere mit kleinen Eiersäcken
beim Aufsuchen des Futters weniger in ihren Bewegungen gehemmt, als
die mit grossen derartigen Anhängseln, und somit diesen beim Kampfe
ums Dasein überlegen. Den höheren Ansprüchen an die Beweglichkeit der
pelagischen Copepoden passten sich die Schwimmwerkzeuge (Fühler und
Beine) ebenfalls in vortrefflicher Weise an, indem sie entweder länger,
oder wenigstens kräftiger entwickelt sind, als bei den Uferformen.
Eigentümlich sind die periodischen Wanderungen der pelagischen Krebse,
indem sie bei Nacht an die Oberfläche steigen, bei Tag dagegen die
Tiefe aufsuchen.
Die als Anpassungserscheinungen zu deutenden Eigenschaften, welche
die Bewohner der pelagischen Region kennzeichnen, können auch den
der Tiefenregion angehörigen von Nutzen sein. Nicht im Meere
allein, dessen tiefste Abgründe bis vor verhältnismässig kurzer Zeit
für unbelebt gehalten wurden, haben sich die Krebstiere und vor allem
die Entomostraken unter einem ganz enormen Wasserdruck eingewöhnt, auch
auf dem Grunde unserer grössten Seen bilden sie einen der wichtigsten
Bestandteile des dort noch herrschenden Lebens. Es sind dies meist
ganz bestimmte Arten,[S. 356] welche nie an der Oberfläche gefunden werden
und deren Verbreitung in allen grösseren Wasserbecken Deutschlands,
Frankreichs und Schwedens darum so schwer zu erklären ist, weil keines
der später zu erwähnenden Transportmittel bei Tieren, die immer in der
Tiefe leben, eine Rolle spielen kann.
Es gelang bis jetzt nie, alle bei uns einheimischen Copepoden in ein
und derselben Wasseransammlung, welchen Umfang diese auch haben mag,
beisammen zu finden. Oft beherrscht eine Art einzig und allein in
unzählbarer Menge irgend einen Weiher, noch öfter aber trifft man
(selbst in unscheinbaren Tümpeln) zwei bis drei Arten an und es ist
eine Seltenheit, wenn z. B. sieben Vertreter einer einzigen Gattung
beisammen leben. Es befremdet diese Unregelmässigkeit des Vorkommens um
so mehr, wenn man erfährt, dass oft ganz dicht nebeneinanderliegende
Weiher eine ganz verschieden zusammengesetzte Copepoden-Fauna aufweisen
können, selbst wenn sie durch kleine Wasseradern mit einander verbunden
sind. Bei seinen Untersuchungen über die Tierwelt der Eifelmaare
fand Zacharias einen Diaptomus (graciloides
Lilljeborg), welcher nur in dem Gemündener Maar (Eifel) vorfindlich
war, und bis jetzt nur in Schweden und der russischen Halbinsel Kola
angetroffen wurde. Wie eine so hochnordische Form so ganz unvermittelt
mitten in Deutschland auftreten kann, bildet noch heute ein Rätsel,
dessen Lösung aber für die Wissenschaft in hohem Masse wichtig und
interessant ist. Auch der schon erwähnte Cyclops maarensis[XVIII]
(ebenfalls aus der Eifel) giebt uns manches zu denken. Wenn nicht
nachgewiesen werden kann, dass er auch in anderen Seen vorkommt,
so müssen wir annehmen, es sei entweder eine Form, welche durch
allmähliche Anpassung an die Lokalität entstanden ist und, selbst wenn
sie verschleppt wird, in anderen Wassern nicht fortkommt, oder am Ende
eine Art, welche überall ausgestorben nur hier sich erhalten hat. Ich
unterlasse es, weitere Gesichtspunkte aufzustellen; diese wenigen mögen
genügen, zu zeigen, wie vieles im Gebiete der Krustaceenkunde, selbst
in den verhältnismässig engen Grenzen der[S. 357] Süsswasserfauna, noch zu
erforschen ist und von welchen Gesichtspunkten man dabei auszugehen hat.
Unsere Kenntnisse von der Art und Weise, wie die Copepoden und
andere niedere Tiere von einem Wasser in das andere, sofern beide
gänzlich abgeschlossen sind, gelangen, sind noch sehr mangelhaft
und wir können, da nur wenige genaue Beobachtungen bis jetzt
angestellt wurden, uns vorderhand fast nur in Vermutungen ergehen.
Man könnte daran denken, dass der Wind, welcher bei der Verbreitung
der Pflanzen eine so wichtige Rolle spielt, auch die Übertragung
der kleinen Copepoden vermittle. Allein der Körper derselben zeigt
so wenig wie die Eier irgend eine Einrichtung, welche einen solchen
Transport wahrscheinlich machte. Im Gegenteil, die ganzen Tiere und
deren Eier sind gegen Austrocknung ausserordentlich empfindlich und
sterben schon bei geringem Wasserverlust ab, und dass Wasserteilchen
mit eingeschlossenen Copepoden oder deren Keimen stundenweit durch
die Macht des Windes fortgerissen werden sollten, ist nicht wohl
anzunehmen. Viel wahrscheinlicher und durch die Untersuchungen der
unten genannten Forscher zumteil bewiesen ist eine andere Art der
„passiven Wanderung“ der niederen Süsswassertiere, welche durch
Schwimmvögel und Wasserinsekten bewerkstelligt wird. Jules de
Guerne hat nämlich aus am Gefieder und den Beinen von Wasservögeln
hängenden Schlammbrocken eine ganze Mikrofauna zu erziehen vermocht.
Nicht nur ganze Tiere, sondern in besonders reicher Anzahl deren
manchmal durch eine dicke Schale der Vertrocknung widerstehende Keime
waren, an den betreffenden Teilen klebend, auf dem Wege verschleppt zu
werden. Kleine Wasseransammlungen, welche für Vögel unzugänglich sind,
werden nach W. Migulas Untersuchungen durch die Vermittlung der
unscheinbaren Wasserkäfer mit verschiedenen Pflanzen- und Tierarten
belebt.
An eine aktive Wanderung selbst über kleine Strecken darf, falls
es nicht im Wasser geschehen kann, bei der Konstitution und Lebensweise
der Copepoden nicht gedacht werden.
Am meisten Schwierigkeit bietet die Eigenartigkeit und gleichmässige
Zusammensetzung der sogenannten Tiefenfauna in den[S. 358] Süsswasserseen
für eine Erklärung dar. Wir finden zwar unter den Copepoden kaum
ausschliessliche Angehörige der Tiefenfauna, allein zugleich mit
Rücksicht auf die später zu behandelnden Daphniden mögen des
Zusammenhangs wegen einige Betrachtungen über die Verbreitung lebender
Wesen, welche in etwa 20–100 und mehr Metern Tiefe ihr Dasein fristen,
hier Platz finden.
Da unsere meisten Binnenseen nicht durch Wasseradern mit einander in
Verbindung stehen, so können die Tiere nicht auf direktem Wege von
dem einen zum andern gelangen, um so weniger, da sie die Tiefe nie
verlassen. Hierdurch ist auch der Transport durch Vögel ausgeschlossen.
Man nahm deshalb an, dass alle diese Seen, was auch zweifellos bei
vielen der Fall war, in früheren Zeiten vom Meer überdeckt gewesen
seien, beim Zurückgehen desselben übrig blieben und mit ihm eine Anzahl
von Tieren, welche der allmählichen Aussüssung dieser sogenannten
„Reliktenseen“ zu widerstehen vermochten. So plausibel diese Hypothese
ist, so giebt sie doch keinen Grund dafür an, warum unter den vielen
tausenden immer nur ganz wenige und überall beinahe dieselben Formen
eine solche Veränderung des Wassers ertrugen. Viel wahrscheinlicher
und neuerdings durch Zacharias[119] vertreten ist eine andere
Ansicht. Hiernach wanderten die in Frage stehenden Arten ganz
allmählich aus dem Meere in das süsse Wasser ein und zwar zu einer
Zeit, wo der grössere Wasserreichtum der Erde für eine solche Wanderung
niederer Tiere eine günstigere Verbindung darbot. Auch heute noch
kann vielfach ein Aufsteigen mariner Formen in unsere Flüsse und Seen
beobachtet werden. Eine plötzliche Überführung eines Mitgliedes der
Süsswasserfauna ins Meer, oder umgekehrt, wird nur von den Parasiten
unter den Copepoden, zu denen wir uns jetzt wenden wollen, ertragen,
während die meisten übrigen Krebse diesem raschen Wechsel erliegen.
Es wurde schon früher erwähnt, dass auf Grund ihrer Entwickelung die
parasitisch lebenden Copepoden zu den Eucopepoden gehören, ursprünglich
ein freies Leben führen und eine cyclopsähnliche Form besitzen. Mit
der Änderung der Lebensweise, welche[S. 359] gewöhnlich schon früh mit der
Auffindung eines entsprechenden Wirtes beginnt, bleiben einzelne
Körperteile in der Entwickelung zurück oder bilden sich, wie namentlich
manche Gliedmassen, zu so seltsamen Formen um, dass die Identität
derselben mit denen des freilebenden Tieres nur schwierig festzustellen
ist. Manchmal geht die Segmentierung des Leibes verloren oder es
verkümmert der Hinterleib. Da die Beine zum Schwimmen bei vielen
Arten nicht mehr benutzt werden, fehlen sie manchmal ganz oder sind
nur in Andeutungen vorhanden. Die Mundteile werden der Lebensweise so
angepasst, dass der Besitzer sich saugend von den Körpersäften seines
Wirtes ernähren kann. Ein Teil derselben, wie auch die zweiten Antennen
bilden gewöhnlich Klammerorgane, welche den Schmarotzer auf dem Körper
des heimgesuchten Tieres befestigen.
Die ersten Antennen sind meistens noch vorhanden, aber gewöhnlich sehr
klein, in etlichen Fällen fehlt eine Gliederung. Dass die zweiten
Antennen in der Regel als Haftorgan funktionieren, wurde oben gesagt.
Die Ober- und Unterlippe ist langgezogen. Jede stellt eine Halbrinne
dar und beide bilden sich aneinanderlegend eine kurze Röhre, in welcher
das erste, zu feinen stilettförmigen Gebilden umgewandelte Kieferpaar
liegt. Das zweite Kieferpaar kann ganz verkümmert sein. Von der
merkwürdigen Umwandlung eines Kieferfusspaares zu einem Haftapparat
kann Figur 73 einen Begriff geben, welche ein Tier darstellt, bei dem
die Kieferfüsse beider Seiten, K, an den Enden verwachsen.
Wollte man alle die Umbildungen, welche der Körper der parasitischen
Copepoden erleidet, aufzählen, so müsste man jede Art für sich
beschreiben. Das Angeführte mag jedoch in Verbindung mit der Abbildung
genügen, um wenigstens eine Vorstellung zu geben von dem, was Anpassung
an parasitische Lebensweise heisst.
Fig. 73. Achtheres percarum von Nordmann. Weibchen von oben mit Eiersäckchen.
K Kieferfüsse verwachsen — D Darm — E Eiersäckchen.
[S. 360]
Weit weniger auffallend, allerdings auch nicht so sehr bekannt, sind
die Veränderungen, welche die inneren Organe erleiden.
Das Nervensystem ist im allgemeinen noch so erhalten, wie wir es
früher kennen lernten, allein die Sinnesorgane sind sehr spärlich
vorhanden. Vor allem fehlt meistens das Auge ganz. Hand in Hand mit
der Verkümmerung der Antennen geht eine solche der daran befindlichen
Sinnesorgane und Borsten. Den meisten Schmarotzern scheint ein Herz
ganz abzugehen. Die Muskulatur entspricht der geringen Beweglichkeit
der Tiere und ist vor allem in den Haftapparaten gut ausgebildet. Am
besten kommen bei den Parasiten der Darm und die Geschlechtsorgane
weg, denn beide zusammen füllen den ganzen Körper beinahe allein aus.
Letztere schwellen zur Zeit der Reife und Befruchtung der Eier oft
ungeheuer an und bedingen die wunderbarsten Gestaltveränderungen.
Das ganze Tier scheint nur noch der Ernährung und Fortpflanzung zu
dienen und bildet so den direkten Gegensatz zu den immer nur wenige
Eier produzierenden pelagischen freilebenden Copepoden. Nicht selten
erreicht oder übertrifft der Inhalt der Eiersäckchen den Umfang des
Körpers an Grösse.
Während so die geschlechtsreifen Weibchen oft eine für einen Copepoden
ganz ansehnliche Grösse erreichen (bis 1.5 cm), bleiben bei
manchen Arten die Männchen zwergartig klein und wurden früher gar nicht
gefunden oder nicht richtig erkannt. Sie halten sich in diesem Falle am
Weibchen auf.
Die schmarotzenden Copepoden bilden, wenn man die im Meere vorkommenden
Formen berücksichtigt, etwa die Hälfte aller bekannten Arten. Ihre
Bedeutung im süssen Wasser ist eine geringe, da sie trotz der enormen
Vermehrung doch selten ihren Wirt (zumeist Fische) am Leben bedrohen.
Manche Arten der Siphonostomata leben nur auf der Haut der von
ihnen bewohnten Fische von den schleimigen Absonderungen derselben,
andere saugen mittels ihres Rüssels das Blut der Wirte und nur wenige
bohren sich geradezu in das Fleisch derselben ein.[S. 361] Besonders häufig
trifft man Parasiten auf den Kiemen an, wo sie immer reichliche
Nahrung, sei es Blut oder Schleim, und zugleich Schutz vor dem
Abgestreiftwerden finden.
Nicht bei allen Arten schmarotzen beide Geschlechter gleichzeitig. Die
Männchen der auf den Kiemen unserer Fische so häufig anzutreffenden
Gattung Ergasilus leben gewöhnlich frei und die Weibchen selbst
zeigen an ihrer Körperform noch viele Ähnlichkeit mit einem Cyclops.
Die ersten Fühler sind gegliedert, ein Auge ist noch vorhanden. Die
beiden Eiersäckchen sind sehr gross und langgestreckt. Die Mundteile
sind, obwohl zum Stechen eingerichtet, von keiner Saugröhre umgeben.
Von den Parasiten, welche bei massenhaftem Auftreten dem Fischzüchter
unter Umständen Schaden verursachen können, ist die Gattung
Lernaeocera bemerkenswert. Auf unseren Fischen leben mehrere
Arten derselben, welche sich oft tief in das Fleisch einbohren, wie
beim Karpfen, oder hässliche Anschwellungen (am Unterkiefer des
Hechtes) verursachen. Bis zur Begattung leben Männchen wie Weibchen
frei; nach derselben sucht dieses einen passenden Wirt und nun beginnen
sich am Körper des Copepoden eine Reihe von Umänderungen zu vollziehen,
nach deren Abschluss das Tier viel eher einem Wurm, denn einem Krebs
ähnlich sieht. Vor allem geht die Segmentierung des Körpers beinahe
ganz verloren, der Hinterleib verkümmert, die Beinpaare, vier an der
Zahl, rücken weit auseinander und sind ausserordentlich klein. Die
ersten Kiefer sind von einer, wie oben beschrieben, gebildeten Röhre
umgeben.
Ein recht häufiger Bewohner der Kiemen unseres wohlschmeckenden
Flussbarsches ist der in Fig. 73 abgebildete Achtheres percarum
von Nordmann. Die grossen nach vorn gebogenen Kieferfüsse, welche an
der Spitze verwachsen sind, geben dem Tiere ein Aussehen, als wollte es
die Hände ringen, und machen es sehr leicht kenntlich.
Wir nehmen hiermit von den echten Ruderfüssern Abschied und wenden uns
zu der zweiten Gruppe der Copepoden, den Karpfenläusen.
[S. 362]
Kiemenschwänze (Branchiura).
Schon bei den parasitischen Eucopepoden mussten wir unsere
Vorstellungen über Bau und Gliederung des normalen Copepoden-Körpers
in vielfacher Hinsicht ändern. Auch bei den ebenfalls der Hauptsache
nach parasitisch lebenden Karpfenläusen treffen wir Gestalten an,
welche nur nach genauer Untersuchung den Ruderfüsser verraten. Die
Organisation der wenigen hierher zu rechnenden Formen ist trotz der
Lebensweise eine sehr hohe und den Verhältnissen vorzüglich angepasst.
Der ganze Körper des bis 6 mm langen Tieres ist nahezu eirund
und ganz plattgedrückt. Der verkümmerte Hinterleib ist noch durch
zwei kleine Läppchen (Schwanzflosse) angedeutet. Dem Umstand, dass
diese zumteil als Kiemen funktionieren, verdanken die Tiere den
Namen „Kiemenschwänze“. Die Mundteile sind zum Saugen eingerichtet.
Die Nahrungsaufnahme wird mittels eines Stachels und einer Röhre
vollzogen. Mit den beiden Kieferfusspaaren hält sich das Tier auf den
von ihm bewohnten Fischen fest. Das erste ist zu diesem Zwecke zu zwei
grossen runden Saugnäpfen umgestaltet, das zweite mit scharfen Klauen
zum Festhaken versehen. Die beiden Antennenpaare sind sehr klein und
unscheinbar. Da die Karpfenläuse, namentlich während der Begattung,
frei umherschwimmen, sind ihre vier Paare von Schwimmbeinen noch ganz
gut ausgebildet. Zum ersten Mal unter den Copepoden treffen wir bei den
Branchiuren zwei komplizierte, weit auseinanderstehende Augen an. Das
Nervensystem ist hochentwickelt. Das Herz entsendet gegen den Kopf zu
das Blut durch eine Aorta; von dem Vorkommen gesonderter Vorrichtungen
zum Atmen war schon die Rede. Auch am Darm zeigt sich eine höhere
Entwickelungsstufe daran, dass die Leberzellen in grossen verästelten
Schläuchen enthalten sind. Die Weibchen hegen keine solche Sorgfalt für
die sich entwickelnden Jungen, dass sie die Eier mit sich herumtragen
würden, sondern entleeren diese einfach in das Wasser oder heften sie
an Gegenständen an. Am häufigsten trifft man die gemeine Karpfenlaus,
Argulus foliaceus[121], an.
[S. 363]
Die Karpfenläuse bewohnen nicht ausschliesslich, wie etwa der Name
vermuten lassen möchte, unsere Karpfenarten, sondern kommen beinahe
auf allen im süssen Wasser, einige sogar auf im Meere lebenden Fischen
vor. Mit ziemlicher Gewandtheit bewegen sie sich auf der schlüpfrigen,
schleimigen Haut der letzteren herum und sind trotz ihrer Grösse nicht
leicht wahrzunehmen, da ihre grünliche Farbe sehr gut mit der der
Fische übereinstimmt und ausserdem die platte Gestalt den eigenartigen
Krebsen gestattet, sich so anzuschmiegen, dass ihre Anwesenheit kaum
auffällt.
Schaden und
Nutzen der Entomostraken.
Da es nun einmal im Wesen unserer realistischen Zeitströmung liegt,
dass ein Gegenstand erst dann unser volles Interesse erregt, wenn wir
erfahren, ob er dem Menschen nützt oder schadet, so mag, ehe wir zu
den übrigen Entomostraken-Familien übergehen, zum Schlusse noch die
Bedeutung der Copepoden und zugleich der sonstigen niederen Krebse für
die übrigen Bewohner des süssen Wassers und die Beziehungen jener zum
Menschen kurz angeführt werden.
Es ist aus dem früher Gesagten zu entnehmen, dass die Parasiten nach
ihrer Lebensweise weder den anderen Süsswassertieren noch dem Menschen
Nutzen gewähren können. Sie schädigen vielmehr die Fische an Gesundheit
und Leben und damit indirekt den Menschen. Dieser immerhin unbedeutende
Schaden wird aber in reichlichem Masse durch die freilebenden
Ruderfüsser und die Entomostraken überhaupt aufgewogen. Diese sind
es, welche das ganze unendlich reiche Material der kleinsten Lebewesen,
sowohl Pflanzen wie Tiere, für die höhere Tierwelt aufschliessen und
nutzbar machen und zwar unter Darangabe ihres Lebens. Unschätzbare
Massen kostbarer Nährstoffe gingen, wenigstens für den Menschen,
verloren, wenn nicht die Entomostraken in unermüdlicher Thätigkeit alle
die kleinsten Urtiere und einzelligen Algen, welche dem Auge unsichtbar
am Boden der Gewässer oder in diesen selbst suspendiert sich vorfinden,
zu sammeln und in ihrem Körper aufzuspeichern vermöchten.
[S. 364]
Es würde zuweit führen, wollte man alle die Tiere, welche ganz oder
teilweise von Entomostraken leben, hier anführen. Der Anwesenheit der
Copepoden vor allem verdanken wir die wohlschmeckenden Blaufelchen
und Saiblinge unserer grossen Seen. Diese und andere nicht weniger
schätzbare Fische leben so ausschliesslich von Entomostraken, dass
sie von der Tafel verschwinden müssten, sobald diese einmal nicht
mehr in genügender Weise sich vermehren würden. Jährlich werden an
verschiedenen Meeresküsten die schon erwähnten Scharen von Copepoden
beobachtet, welche auf weite Strecken hin das Meer rot färben und darum
als „Rotäsung“ (Maidre) bezeichnet werden. Von dieser Erscheinung hängt
der tausende von Menschen ernährende und beschäftigende Heringsfang
ab. Sardellen, Sardinen, Makrelen, ja selbst das riesigste aller
Säugetiere, der Walfisch, leben oft ausschliesslich von den kleinsten
Krebstieren. Die Walfischjäger wissen, dass stets die Jagd in der
Nähe der roten Copepoden-Wolken gute Ausbeute liefert. Es mag, um zu
den Süsswasser-Entomostraken zurückzukehren, noch eines Umstandes
gedacht werden, welcher allein schon genügte, die Unentbehrlichkeit der
niederen Krebse für die Fauna der Seen und indirekt für den Menschen
darzuthun.
Die jungen Fische nämlich, d. h. die sogenannte „Fischbrut“, findet
kaum eine passendere Nahrung in der ersten Zeit nach dem Verlassen
des Eies als Entomostraken. Ob der Fisch später, wie der Barsch und
der Hecht, reiner Räuber, oder, wie der Karpfen, fast ausschliesslich
Pflanzenfresser wird, bleibt sich gleich, in der ersten Jugend frisst
er nur Entomostraken. Wer in rationeller Weise Fischzucht betreiben
will, kann auf einem ganz einfachen Wege sich dieses billigste und
zugleich beste Futter für künstlich erbrütete Fische beschaffen:
indem er nämlich geeignete Teiche trockenlegt, den Grund düngt, wie
der Landmann sein Feld, und nun im Frühjahr zur Laichzeit der Fische
den Teich wieder bespannt, d. h. mit Wasser füllt. Er hat damit alle
Bedingungen erfüllt, welche in kurzer Zeit eine geradezu ungeheuerliche
Vermehrung der wenigen Entomostrakenkeime, welche im Schlamme sich
noch vorfanden oder von aussen zugebracht wurden, ermöglichen.[S. 365] Auf
die grossen Vorteile einer solchen naturgemässen Fütterung weiter
einzugehen, ist hier nicht der Ort. Betont mag jedoch werden, dass
die eben geschilderte Methode einen wesentlichen Faktor für die
gedeihliche Entwickelung unserer Fischereiverhältnisse darstellt.
Man kann geradezu behaupten, dass eine erfolgreiche Aufzucht junger
Fische, sei es in den Räumen einer Fischzuchtanstalt oder im freien
Naturleben, an das Vorkommen von Entomostraken geknüpft ist. In noch
viel ausgedehnterem Masse werden solche Methoden Anwendung finden, wenn
erst die Wissenschaft auf dem so wichtigen Gebiet der Krustaceenkunde
dem praktischen Leben durch ein genaues Studium der Formen und der
Lebensweise der Tiere vorgearbeitet hat. Von den vielen Lücken, welche
hier noch auszufüllen sind, haben wir im Vorstehenden einige wenige
angedeutet. „Gleichmässige Würdigung aller Teile des Naturstudiums ist
aber vorzüglich ein Bedürfnis der gegenwärtigen Zeit, wo der materielle
Reichtum und wachsende Wohlstand der Nationen in einer sorgfältigeren
Benutzung von Naturprodukten und Naturkräften gegründet ist“, sagt
Humboldt in der Einleitung zu seinem „Kosmos“ und heute mehr als je
darf in Rücksicht auf das eben Mitgeteilte diese Mahnung den Männern
der Wissenschaft ins Gedächtnis gerufen werden.
Blattfüsser (Phyllopoden).
Diese Gruppe verdankt den Namen, wie die vorhergehende, der
eigentümlichen Form und Funktion der Beine, deren einer Ast blattförmig
verbreitert ist und teilweise als eine Art Kieme der Atmung dient.
Die Blattfüsser sind fast ebenso häufig wie die Ruderfüsser und nicht
selten ebenso zahlreich in einem Wasser vertreten wie diese. Nur im
Winter treffen wir sie nicht immer an. Trotzdem dass durch Parasitismus
umgeänderte Arten fehlen, umfasst diese Ordnung der niederen Krebse
dennoch eine bedeutende Anzahl recht verschieden gestalteter Wesen. Für
die Beobachtung des lebenden Tieres stellen die meisten Arten wegen
ihres seitlich komprimierten Körpers ein noch viel anziehenderes Objekt
dar als die Angehörigen der zuerst behandelten Ordnung. Nach der Zahl[S. 366]
der Beine und einigen anderen Merkmalen trennt man die Blattfüsser in
zwei Abteilungen. Die einen mit vier bis sechs Beinpaaren bezeichnet
man als
Wasserflöhe
oder Cladocera.
Der Körper dieser wegen der Art ihrer Bewegung „Wasserflöhe“, wegen
der verzweigten Fühler (Hörner) „Cladocera“ genannten Entomostraken
ist nicht in deutliche Segmente geteilt. Nicht einmal immer ist der
Kopf durch einen kleinen Einschnitt vom Rumpfe abgesetzt. Letzterer
ist oft von einer Schale umschlossen. In dieser Schale, deren Form
die äusseren Umrisse des hinteren Körperteiles bedingt, liegt der
Leib frei beweglich. Die beiden Hälften der Schale sind am Rücken mit
einander verwachsen, auf der Oberfläche häufig mit feinen Skulpturen
versehen und gewöhnlich so durchsichtig, dass man die inneren Organe
beobachten kann. Nicht immer ist der Leib ganz in der Schale verborgen,
sondern oft nur zu einem ganz geringen Teil davon überdeckt. In wenigen
Fällen fehlt jede Andeutung einer Schale. Am Kopfe stehen wie zwei
Hörnchen die hinteren zweiästigen Fühler empor, welche zum Schwimmen
dienen. Das erste Paar ist zu kurzen Stummeln rückgebildet, welche
als Sinnesorgan mit sehr feinen perzipierenden Apparaten ausgestattet
sind und beim Männchen zugleich als Greiforgan funktionieren. Von
Mundteilen besitzen die Wasserflöhe jederseits einen Ober- und einen
Unterkiefer. Die Beinpaare sind wie bei den Copepoden zweiästig. Der
äussere Ast ist blattförmig verbreitert und vertritt die Stelle von
Kiemen. Die Zahl der Beinpaare schwankt zwischen vier und sechs. Die
das Zentralnervensystem darstellende Ganglienkette hat, da die Knoten
sehr nahe zusammenrücken, nur eine geringe Länge. Auffallend gross und
schön ist das einen bedeutenden Teil des Kopfes ausfüllende Auge. Es
ist in einem beständigen Zittern begriffen und besteht eigentlich aus
zwei verwachsenen Teilen. Ohne besondere Mühe vermag man die daran
sich ansetzenden Muskeln und Nerven zu sehen. Die lichtbrechenden
Körper (Linsen) glänzen sehr schön und heben sich scharf vom Pigment
der inneren Teile[S. 367] des Auges ab. Bei den meisten Arten verläuft der
Darm gerade durch den Körper und nur in wenigen Fällen bildet er eine
Schlinge. Noch im Kopfteil entspringen an seiner Oberseite zwei gegen
die Augen vorspringende kurze Schläuche, in welchen die Leberzellen
untergebracht sind.
Das Herz, welches keiner Art fehlt, liegt hinter dem Kopf und Rumpf
trennenden Einschnitt und pulsiert ausserordentlich rasch. Die Richtung
des Blutstromes lässt sich sehr schön an den in der Flüssigkeit
schwimmenden Körperchen verfolgen. Das Herz selbst ist ähnlich
gebaut wie das schon beschriebene von Diaptomus. Hinter dem Herzen
befindet sich zwischen Leib und Schale ein Hohlraum, welcher bei den
Weibchen nicht selten mit grossen Eiern in verschiedenen Stadien der
Entwickelung erfüllt ist. Die Eier werden in einer unter und neben dem
Darm liegenden Drüse erzeugt. Ganz eigentümliche Vorgänge spielen sich
bei der Vermehrung der Tiere ab. Die Wasserflöhe erzeugen nämlich zwei
Arten von Eiern, welche in Form, Grösse, Farbe etc. verschieden sind.
Während der günstigeren Jahreszeit bilden die reifen Weibchen kleine
dünnhäutige Eier, welche sich sehr rasch entwickeln und aus denen nur
Weibchen hervorgehen. So kann selbst, wenn in dem Wasser Männchen
vorkommen, eine ungeschlechtliche Vermehrung längere Zeit hindurch
fortgesetzt werden, allein nur dann, wenn die Lebensbedingungen für
die Tiere recht günstige sind, d. h. wenn genügend Wasser, Nahrung
und Wärme zu Gebote stehen. Beginnt das Wasser in dem von Daphniden
bewohnten Tümpel zu verdunsten oder droht mit Eintritt der kälteren
Jahreszeit den Tieren Gefahr, so werden sogenannte Winter- oder
Dauereier gebildet, welche nur nach vorhergegangener Begattung zu
stande kommen. Diese zeichnen sich durch bedeutendere Grösse und
dickere Haut, vor allem aber dadurch aus, dass sie, um äusseren
Einflüssen besser Widerstand leisten zu können, mit einer recht derben
Hülle, in welcher meistens zwei Eier zugleich eingeschlossen sind,
umgeben werden. Diese Hülle, den sogenannten „Sattel“ (Ephippium),
erhalten die befruchteten Eier erst nach dem Eintritt in den Brutraum.
Den durch das Ephippium geschützten Eiern[S. 368] fällt die wichtige Aufgabe
zu, die Art vor dem Aussterben in austrocknenden Tümpeln zu bewahren
und eine recht ausgiebige Verbreitung zu bewirken. Diese erfolgt
namentlich in Seen, die von wandernden Wasservögeln besucht sind, darum
sehr leicht, weil die Ephippien an der Oberfläche des Wassers schwimmen
und unschwer an anderen Gegenständen haften. Da die Wintereier ebenso
gut der Wärme und Trockenheit als der Kälte widerstehen, können sie
leicht mit dem zu Staub zerfallenden Schlamm austrocknender Gewässer
durch den Wind verweht werden, wie der Blütenstaub der Pflanzen.
Fig. 74. Daphnia (Simocephalus) sima (Liévin). a′ Erste (verkümmerte)
Antenne — a″ Zweite Antenne — Au Auge — B Äusserer Ast
der Beine — Hi Hirn — L Leberschlauch — D Darm — H
Herz — Brtr. Brutraum — SE Sommerei im Brutraum — Est.
Eierstock.
Über die Befruchtungserscheinungen am Eie hat Weismann[122] eine
Reihe gründlicher Untersuchungen angestellt. Die Wasserflöhe verlassen
das Ei in einer Gestalt, welche der der erwachsenen Tiere schon sehr
ähnlich ist, machen also kein freilebendes Cyclopsstadium durch.
Fig. 75. Leptodora hyalina Lilljeborg. A′ Erste Antenne - a″
Zweite Antenne — Au Auge — Hi Hirn — H Herz — S
Schale — E Eier — Est Eierstock — D Darm.Fig. 76. Bythotrephes longimanus Leydig. a′ Erste Antenne — a″
Zweite Antenne — Au Auge — Brtr Brutraum — E Ei — D
Darm — Sch Schwanzstachel, nur zumteil gezeichnet.
Von den vier Familien, in welche die Wasserflöhe eingeteilt werden
und welche von Leydig[123] eingehend in einer Monographie
behandelt wurden, ist die der Daphniden am zahlreichsten vertreten.[S. 369]
Oft färben Angehörige der Gattung Daphnia (Fig. 74) frisch
entstandene Regenwassertümpel durch ihre Massen rot. Die seltsamsten
und interessantesten Formen umfasst die Familie der grossäugigen
Polyphemiden, von denen Fig. 75 und 76 zwei Arten darstellen, welche
in vielen unseren grossen Seen, gewöhnlich[S. 370] in der Tiefe, angetroffen
werden. Der von Leydig zuerst im Magen von Blaufelchen
entdeckte Bythotrephes longimanus zeichnet sich durch seinen
geradezu abenteuerlich langen Schwanzstachel, welcher sich zum Körper
des Tieres wie 10 : 2 verhält, und die verlängerten Beine des ersten
Paares aus. Der Brutraum ist wie bei der wegen ihrer Durchsichtigkeit
so benannten Leptodora hyalina nur klein. Letztere übt,
durch ihre Farblosigkeit begünstigt, ihrem Opfer unsichtbar sich
nahend, ihr räuberisches Handwerk gegen die übrigen Mitglieder der
Entomostrakenfauna aus und entgeht zugleich der Verfolgung ihrer
Feinde. Die Leptodora wird etwa einen Centimeter lang.
Viel grössere Tiere bilden die zweite Unterordnung der Blattfüsser,
die sogenannten Kiemenfüsser (Branchiopoden). Während bei den
Wasserflöhen höchstens sechs Beinpaare vorhanden sind, treffen wir
bei den Kiemenfüssern von 10–40 Paare an. Der Leib ist manchmal, wie
bei den Daphnien, von einer Schale umschlossen und meistens deutlich
gegliedert. Die Ruderantennen sind bei manchen Arten verkümmert.
Die Tiere vermehren sich oft durch viele Generationen hindurch
parthenogenetisch und lange Zeit hielt man einige Arten, da die nur
ganz selten auftretenden Männchen nicht beachtet worden waren, für
Zwitter. Gewöhnlich kommen die zwei grössten Kiemenfüsser gleichzeitig
in kleinen Wasseransammlungen, welche zeitweise vertrocknen, vor. Der
grösste, Apus, verzehrt häufig den kleineren, Branchipus,
weshalb man beim Fange dieser im südlichen Deutschland nur ganz
selten, dann aber in grossen Mengen vorkommenden Krebse darauf
bedacht sein muss, beide in besonderen Behältern aufzubewahren. In
der Gefangenschaft dauern die Branchiopoden leider nur kurze Zeit
aus, bilden aber während des Lebens durch die Seltsamkeit der Form
und Bewegung, welche einem gleichmässigen Kriechen ähnelt, sowie
die teilweise prächtige Färbung ganz anziehende Objekte für die
Beobachtung. Die kleinen Eier sind bei Apus rot, bei Branchipus schön
himmelblau gefärbt und entwickeln sich unter günstigen Umständen in
dem Gefäss, in dem die Tiere gehalten werden. Der Entwickelung muss,
wie es scheint, eine vollkommene[S. 371] Austrocknung und ein Durchfrieren
vorangehen, denn jahrzehntelang sucht man in einem Tümpel vergebens
nach diesen schönen Krustern, trotzdem dass sie schon einmal darin
beobachtet wurden. Erst wenn derselbe einmal längere Zeit trocken lag
treten sie wieder darin auf. Der Leib des 6–7 cm lang werdenden
Apus ist mit einer schildförmigen Schale bedeckt, auf welcher in der
Kopfgegend die Augen unbeweglich sitzen. Am Ende des Hinterleibes
stehen zwei lange Schwanzborsten. Die vielen verbreiterten Beine mit
den Kiemenanhängen sind in beständiger Bewegung. Drei lange Fäden,
welche rechts und links die Seiten des Kopfes überragen, gehören dem
ersten Beinpaare an.
Dem nur 1½ cm langen schlanken Branchipus fehlt die Schale
gänzlich. Die zweiten Fühler des Männchens sind zu Greifhaken
umgeformt. Die Augen sitzen auf Stielen und können bewegt werden. Der
Vorderleib besteht aus elf Segmenten mit ebenso vielen Beinpaaren, der
der Gliedmassen entbehrende Hinterleib aus neun Segmenten. Eine mit
Branchipus verwandte Art, Artemia, lebt nur in sehr salzhaltigem
Wasser und wird, wenn sie allmählich in süsseres übergeführt wird, nach
einer Anzahl von Generationen dem Branchipus ähnlich.
Alle Branchiopoden kommen nur in Binnengewässern, nie im Meere vor. Die
das Ei verlassenden Jungen besitzen die Naupliusgestalt.
Sowohl nach der Zahl der Arten als der Individuen ist in der
Entomostrakenfauna am geringsten die Ordnung der
Muschelkrebse (Ostracoda).
Fig. 77. Cypris fasciata.a′ u. a″ Antennen — Au Auge —
Ab Abdomen — L Leberschlauch — Est Eierstock — m
Muskeln — M Mandibel — Sch Schale.
Schon früher wurde die Art der Bewegung dieser kleinen Krebse
geschildert und erwähnt, dass der Name von der eigentümlichen
muschelähnlichen Schale herrührt, in welche der ganze Körper des Tieres
eingehüllt ist. Die beiden Hälften der hie und da Kalk enthaltenden
Schale sind am Rücken durch eine elastische Haut verbunden, welche den
Schliessmuskeln entgegenwirkt und bei einer Ausdehnung der letzteren
ein Öffnen der Schale verursacht. Der Körper ist nur sehr schwer in
seinen Umrissen erkennbar und undeutlich gegliedert. Von Gliedmassen
treffen wir[S. 372] wieder zwei Paar Fühler an, deren hinteres beinförmig
ist. Es folgen dann nach rückwärts ein Paar Oberkiefer und zwei Paar
Unterkiefer. Da nur zwei eigentliche Beinpaare vorkommen, versieht
ausser den hinteren Fühlern beim Männchen der Taster des zweiten
Unterkieferpaares die Funktion eines solchen und zugleich eines
Greiforganes. Das Ende des Hinterleibes ist wie bei den Wasserflöhen
nach vorn geschlagen. Der verwischten Segmentierung des Körpers und
der zusammengedrängten Gestalt entspricht ein verkürztes Nervensystem.
Es ist nur ein aus zwei verschmolzenen Augen entstandenes Sehorgan
vorhanden, von anderen Sinnesorganen treffen wir wieder blasse
Anhängsel an den Antennen. Das Herz fehlt. Der Darm ist nicht so
einfach wie bei den beiden früher beschriebenen Ordnungen, sondern er
besteht aus mehreren in Form und Bau scharf von einander abgesetzten
Abteilungen. Die Eier werden in zwei langen, nach hinten dünner
werdenden Schläuchen erzeugt. Die Männchen, welche bei manchen Arten
sehr selten sind, besitzen sehr komplizierte Fortpflanzungsorgane. Die
meisten Arten der bei uns vorkommenden Muschelkrebse vermehren sich
vorwiegend auf parthenogenetischem Wege. Die Jungen sind beim Verlassen
des Eies schon von der Schale umgeben. Am häufigsten finden wir in
unseren Tümpeln und Seen die Gattung Cypris (Fig. 77) an. Die
Lebensweise dieser kleinen, höchstens bis 3 mm lang werdenden
Entomostraken gleicht sehr derjenigen der früheren Ordnungen. Munter
zappelnd schwimmen sie zwischen den Wasserpflanzen umher, immer nach
kurz andauernder Bewegung wieder einen Augenblick mit den Fühlern sich
an festen Gegenständen anheftend, um auszuruhen. Die Nahrung besteht in
pflanzlichen und tierischen Stoffen. — Zum Schluss wenden wir uns zu
den
[S. 373]
Höheren Krebsen oder Malakostraken.
Von den zahlreichen Gattungen der Ordnung der
Asseln (Isopoden),
welche zumeist im Meere leben, gehört eine unserer Süsswasserfauna an
— die gemeine Wasserassel, Asellus aquaticus L. (Fig. 78).
Wie man aus der Abbildung ersieht, ist der Körper sehr vollkommen
gegliedert, nur wenige Segmente des Hinterleibs sind verwachsen.
Der nach hinten etwas breiter werdende Leib ist von oben nach unten
zusammengedrückt und steht an den Seiten über. Der mit dem folgenden
Brustringe nicht verwachsene Kopf trägt ein kurzes erstes und ein sehr
langes zweites Fühlerpaar. Von Mundwerkzeugen sind jederseits ein
Oberkiefer, zwei Unterkiefer und ein Kieferfuss vorhanden. Auf diese
folgen sieben Paare von Brustbeinen, deren gleichartige Beschaffenheit
Veranlassung gab, die Ordnung „Isopoden“ (Gleichfüsser) zu benennen.
Der verkürzte Hinterleib trägt sechs zweiästige Beinpaare. Der
Innenast der fünf ersten ist zu einem Kiemenplättchen umgewandelt, das
sechste steht über das Hinterleibsende vor. An den vorderen Fusspaaren
sitzen beim Weibchen Platten, welche sich an der Bauchseite anlegen,
übereinandergreifen und so einen ausdehnbaren Raum zwischen den
Wurzeln der Füsse herstellen, in welchem die Eier gehegt werden. Der
Darm verläuft ohne Windungen zu machen gerade durch den Körper. Schon
äusserlich lassen sich drei Abschnitte an ihm erkennen. Auf eine kurze
Speiseröhre folgt ein fast bei allen höheren Krebsen[S. 374] vorkommender
„Kaumagen“, deshalb so genannt, weil an der Innenseite dieses
Darmstückes zwei Platten mit rauher Oberfläche durch besondere Muskeln
gegeneinander reiben und auf diese Weise eine weitere Zerkleinerung der
Nahrung vollbringen. Schon ziemlich weit vorn im Vorderleib beginnt der
eigentliche Darm, an dessen Anfangsteil zwei Paar langer Leberschläuche
einmünden. Das Herz stellt einen langen, unter dem Rücken liegenden
Schlauch dar, an dessen Seiten das Blut durch paarige Öffnungen
eintritt, um bei einer Kontraktion nach vorn durch eine Aorta wieder in
die Leibeshöhle getrieben zu werden. Die Geschlechtsorgane sind paarig
angelegt und münden beim Weibchen am fünften, beim Männchen am letzten
Brustring nach aussen. Das Nervensystem des Vorderleibes bildet einen
Strang, welcher in jedem Segment zu Ganglien anschwillt; im Hinterleib
sind alle diese Ganglien zusammengerückt und bilden einen grossen
Nervenknoten. Das Sehorgan ist durch zwei Gruppen von Punktaugen von
ähnlichem Bau wie bei Cyclops, welche an den Seiten des Kopfes liegen,
gebildet. An den Antennen sitzen grosse blasse Riechkolben.
Fig. 78. Asellus aquaticus L. a′ u. a″ Antennen — Au Auge —
B Brustbeine — H Hinterleibsbeine — K Kieferfüsse.
Die Asseln ernähren sich von pflanzlichen und tierischen Stoffen. Sie
leben gern zwischen modernden Blättern und grünen Algen. Die Eier und
die ausgeschlüpften Jungen werden in dem durch die oben geschilderten
Brutplatten erzeugten Raume umhergetragen. Die Männchen sind gewöhnlich
grösser als die Weibchen.
An feuchten Orten leben auf dem Lande unter Steinen, in Kellern
u. s. w. verschiedene Verwandte unserer Wasserassel, von denen die
Mauerassel (Oniscus), die Körnerassel (Porcellio) und die
Rollassel (Armadillidium) sehr häufig angetroffen werden. Eine
blinde Wasserassel bewohnt tiefe Seen, Brunnen oder die Gewässer von
Höhlen. Sehr nahe mit den Isopoden verwandt sind die
Flohkrebse (Amphipoden).
Der Körper dieser besonders in klaren Gebirgsbächen und Quellen
ungemein häufigen Tiere ist seitlich zusammengedrückt und wohl
gegliedert. Kopf und erstes Brustsegment sind mit einander verwachsen.
Die beiden Fühlerpaare haben annähernd gleiche[S. 375] Länge; am ersten
entspringt ein kleiner Nebenast. Mundwerkzeuge und Beinpaare sind in
gleicher Anzahl wie bei den Asseln vorhanden. Die Endglieder der ersten
zwei Brustbeinpaare sind zu kleinen Greifhänden umgestaltet. An der
Innenseite der meisten Brustbeine sitzen als Kiemen funktionierende
blasige Gebilde, beim Weibchen ausserdem Brutplatten. Die Beine des
Hinterleibes sind in zwei gleichartige Äste gespalten. Die drei
ersten Paare der Hinterleibsbeine unterscheiden sich in der Form von
den folgenden und dienen zum Schwimmen und zur Erzeugung stetigen
Wasserwechsels an den Kiemen, weshalb sie auch in der Ruhe immerwährend
schwingen. Während der Darm dem der Wasserassel sehr ähnlich und
ebenfalls mit zwei Paaren von Leberschläuchen versehen ist, lässt sich
am Herzen insofern eine höhere Entwickelungsstufe erkennen, als es
weiter vorn im Körper seinen Platz hat und sowohl gegen den Kopf zu als
auch nach hinten Gefässe entsendet. Die Gliederung des Nervensystems
entspricht der Segmentierung des Körpers. Die beiden rundlichen
Augen sind ähnlich zusammengesetzt wie bei den höheren Kerfen. Die
Antennen tragen blasse Riechkolben. Die Geschlechtsorgane mit ihren
Ausmündungen verhalten sich wie bei den Isopoden. Die Eier und Jungen
werden ebenfalls zwischen den Brustbeinen von den Brutplatten bedeckt
getragen. Die Art, wie die bei uns gemeinste Gattung Gammarus
(Fig. 79) sich im Wasser bewegt, wurde eingangs geschildert. Man findet
diesen Flohkrebs am sichersten unter Steinen, in deren Nähe faules
Laub sich vorfindet. Oft schimmert ein kleiner roter Körper durch
die Leibeswand. Es ist dies ein Echinorhynchus, ein häufiger
Parasit. Eine blinde Abart des Gammarus lebt, ebenso wie die
blinde Assel, in tiefen Brunnen oder Höhlen.
Fig. 79. Gammarus pulex L. a′ Erste Antenne — a″ Zweite Antenne —
Au Auge — H Hinterleibsbeine — N Nebenast.
[S. 376]
Die Ordnung der
Zehnfüsser (Decapoden)
ist bei uns nur durch den Flusskrebs vertreten, dessen Geschlecht
durch die schon seit einer Reihe von Jahren wütende Krebspest so
sehr dezimiert wurde, dass manche Flussläufe von diesen in Form und
Gebahren gleich originellen Scherenträgern ganz entvölkert sind. Der
Kopf unseres Flusskrebses ist mit den Brustringen und diese unter
sich zu dem Kopfbruststück verwachsen, welches an der Stirn in einen
Stachel ausläuft. Einige vertiefte Linien auf dem Cephalothorax deuten
noch die einst vorhanden gewesene Trennung in einzelne Segmente an.
Der gegliederte Hinterleib dient als Ruder zum Schwimmen und ist beim
Weibchen breiter als beim Männchen. Die ersten kurzen zweiästigen
Antennen werden von den zweiten weit überragt. Ausser einem Paar
Ober- und zwei Paaren Unterkiefern dienen als Mundwerkzeuge noch
drei Paar Kieferfüsse. Auf diese folgen die grossen Brustbeine,
deren Anzahl zu der Bezeichnung „Zehnfüsser“ Veranlassung gab. Die
ersten drei Beinpaare tragen am Ende Scheren, welche am ersten sehr
stark entwickelt sind und eine kräftige Waffe gegen Feinde bilden,
deren Wirkung manchem Leser bekannt sein dürfte. Die sechs Beinpaare
des Hinterleibes sind zweiästig: Das erste bildet beim Männchen ein
Begattungsglied, das letzte ist bei beiden Geschlechtern verbreitert
und giebt mit dem hintersten Körpersegment zusammen die sogenannte
Schwanzflosse ab. An den Hinterleibsbeinen tragen die Weibchen die
Eier. Die Ganglien des Bauchstranges sind in der Brustgegend nahe
zusammengerückt. Die zwei grossen Augen sitzen auf beweglichen Stielen.
Zum ersten Mal unter den Krebstieren treffen wir ein deutliches
Hörorgan an, welches im ersten Glied der kleinen Antennen liegt. Es
stellt ein kleines nach aussen sich öffnendes Bläschen dar, welches
im Innern mit feinen Härchen ausgekleidet ist und vom Krebs selbst
mit Sandkörnchen erfüllt wird. Diese dienen als Hörsteine und müssen
nach jeder Häutung, da die ganze Auskleidung des Bläschens entfernt
wird, erneuert werden; so lange dies nicht geschehen ist, ist[S. 377] der
Krebs taub. Mehrere Male wirft das Tier seinen Hautpanzer, welcher
ihm bei reichlicher Nahrung bald zu enge wird, ab und erscheint
dann, ehe genügend Kalk in der jungen Haut abgelagert ist, weich und
gewöhnlich rötlich gefärbt als sogenannter „Butterkrebs“. Da dieser
äusserst hilf- und wehrlos ist, kann es ihm passieren, dass er von
seinen eigenen Kameraden gefressen wird, wenn er in der Wahl eines
guten Versteckes nicht vorsichtig genug war. Im hintern Teile des
Cephalothorax liegt das (von oben gesehen) etwa fünfeckige Herz,
von dem aus ein reichverzweigtes Gefässsystem das Blut nach allen
Körperteilen leitet. Zum Atmen dienen verästelte Kiemen, welche am
Grunde der Kieferfüsse und Brustbeine entspringen und unter den nach
den Seiten heruntergebogenen Rändern des Cephalothorax verborgen sind.
Der Darm macht keine Windungen, ist aber am Anfang zu einem mit starken
Chitinzähnen ausgerüsteten Kaumagen erweitert, an dessen vorderer
Wand der zur Verstärkung der Haut nötige Kalk in Form von sogenannten
„Krebsaugen“ vorrätig gehalten wird. Die gelbliche, aus kleinen
Schläuchen bestehende Leber erfüllt einen grossen Teil des Vorderleibs.
Die eigentlich paarigen Geschlechtsschläuche verwachsen auf eine kurze
Strecke mit einander und münden beim Weibchen am Grunde des dritten,
beim Männchen am fünften Beinpaare nach aussen. Wie schon gesagt
entwickeln sich die Jungen unter dem Schutz des mütterlichen Schwanzes
und verlassen das Ei in einer dem erwachsenen Tiere sehr ähnlichen
Gestalt. Eiertragende Weibchen findet man bis in den Juni hinein, und
so lange sollten die Tiere eigentlich geschont werden und nicht nur
während der Monate, deren Name ein R enthält. Im September und in der
ersten Hälfte des Oktober ist das Fleisch der Krebse viel fester und
wohlschmeckender als im Mai, wo recht häufig die Tiere sich vom Winter
her noch nicht gehörig erholt haben.
Der Krebs kommt in allen unseren Gewässern, selbst in ganz kleinen
Bergbächen vor. Am liebsten hält er sich unter Steinen und in
selbstgegrabenen Löchern am Ufer während des Tages versteckt und
geht erst mit Einbruch der Dämmerung auf die Suche nach Nahrung.
Diese besteht hauptsächlich aus Aas und wenigen lebenden[S. 378] wirbellosen
Tieren. Dass er Fische und Frösche fange, mag sich zufällig hie und da
beobachten lassen; ich selbst war nie in der Lage, dies bestätigen zu
können. Trotzdem dass ich lange in grossen Aquarien kleine und grosse
Fische mit Krebsen zusammen hielt, konnte nie eine Verminderung der
Zahl der ersteren wahrgenommen werden. Der Krebs ist auch mit seinen
Scheren viel zu langsam, um rasch bewegliche Tiere erhaschen zu können.
Auf weite Entfernung wittert der Krebs im Wasser liegendes Aas und
seine Vorliebe für dieses wird beim Fange benutzt. Bekannt ist an ihm
das Vermögen, verlorengegangene Gliedmassen zu ersetzen. Wird der Krebs
an der einen oder andern Schere erhascht, so sucht er unter Darangabe
dieser oder unter Umständen beider zugleich zu entfliehen. Schon bei
der nächsten Häutung beginnt der Verlust sich zu ersetzen. Die neue
Schere erreicht aber nie wieder die Grösse der ersten. Auch im Kampfe
unter einander, welcher sehr ausdauernd und erbittert geführt wird,
verlieren die Krebse nicht selten ihre Hauptwaffe. Um eine ordentliche
Grösse zu erreichen und fortpflanzungsfähig zu werden, braucht der
Krebs 4–5 Jahre. In raschfliessenden Waldbächen bleibt er gewöhnlich
kleiner, als in Seen und Flüssen. Nützlich werden die höheren Krebse
dadurch, dass sie durch die Vertilgung von allerhand faulenden Stoffen
eine Art Reinlichkeitspolizei in unseren Gewässern bilden. Ausserdem
wird der Flusskrebs — allerdings nicht von jedermann — als Speise
hochgeschätzt.
Wir beschliessen hiermit unsere Betrachtungen über die Krebstiere.
Der Zweck derselben ist erreicht, wenn sie auch in weiteren Kreisen
Kenntnisse über den Bau und die Lebensweise der wesentlichsten bei uns
vorkommenden Krustaceenformen verbreiten.
[S. 379]
Litteratur.
[Bei einigen Literaturstellen handelt es sich um allgemeine
Nachschlagewerke, denen im Text keine direkten Verweise
gegenüberstehen. Diese werden im Folgenden mit Zahlen gefolgt von
runden Klammern dargestellt. (Anmerkung des Bearbeiters)]
1) W. Baird, Natural History of the British Entomostraca.
London 1850.]
2) C. Claus, Über Bau und Entwicklung parasitischer Krustaceen.
Marburg 1855.]
3) S. Fischer, Über das Genus Cypris und dessen bei Petersburg
und Reval vorkommende Arten. Petersburg 1851.]
4) A. Friç, Die Krustenthiere Böhmens. Prag 1871.]
5) T. H. Huxley, The crayfish. London 1889.]
6) W. Lilljeborg, De crustaceis ex ordinibus tribus: Cladocera,
Ostracoda et Copepoda in Scania occurrentibus. Lund 1843.]
7) J. Lubbock, On some freshwater Entomostraca. Transact. Linn.
Soc. Vol. 24.]
8) W. Migula, Biolog. Centralblatt Nr. 17. 1888.]
9) O. Fr. Müller, Entomostraca seu Insecta testacea. Francfurt
1792.]
10) F. Plateau, Recherches sur les crustacées d’eau douce de
Belgique. Mém. couronnée et des savants étrang. 1867.]
11) S. A. Poppe, Notizen zur Fauna der Süsswasserbecken des
nordwestl. Deutschlands mit besonderer Berücksichtigung der Krustaceen.
Abhandl. naturwiss. Verein Bremen Bd. X. 1889.]
12) J. Richard, Entomostracés nouveaux ou peu connus. Bull. d.
l. soc. zoolog. de France. Tome XIII. 1888.]
13) G. O. Sars, Norges Ferskvandskrebsdyr. Cladocera.
Christiania 1865.]
14) E. G. Zaddach, De apodis cancriformis anatomia et historia
evolutionis. Bonn 1841.]
[108]S. Fischer, Beiträge zur Kenntniss der in der
Umgegend von Petersburg sich findenden Cyklopiden. Bull. d. l. soc.
impér. des Naturalistes de Moscou. Bd. 24 (1851). Bd. 26 (1853).
[109]G. S. Brady, Monograph of the free and
semiparasitic Copepoda of the British Islands. London 1880.
[110]C. Claus, Die freilebenden Copepoden. Leipzig
1863.
[111]C. Claus, Copepodenfauna von Nizza. Marburg und
Leipzig 1866.
[113] a) O. Schmeil, Beiträge zur Kenntnis der
Süsswasser-Copepoden Deutschlands. Inauguraldissertation. Halle 1891.
b) J. Vosseler, Die freilebenden Copepoden Württembergs und
angrenzender Gegenden. Jahreshefte d. Ver. f. vaterländische Naturkunde
in Württemberg. 1886.
[120]A. v. Nordmann, Mikrograph. Beiträge H. 6. Berlin
1832.
[121]F. Leydig, Über Argulus foliaceus. Zeitschr. f.
wissensch. Zoologie 1850.
[122]Aug. Weismann, Beiträge zur Naturgeschichte
der Daphnoiden. Leipzig 1876–79.
[S. 380]
[123]F. Leydig,
Naturgeschichte der Daphniden. Tübingen 1860.
Druck von J. J. Weber in Leipzig.
Webers Naturwissenschaftliche
Bibliothek.
Unter obigem Titel erscheint in unterzeichnetem Verlage eine Reihe
von naturwissenschaftlichen Werken, wovon zurzeit drei Bände
vorliegen.
Jeder Band wird ein in sich abgeschlossenes Ganzes bilden und von
einer Autorität auf dem Gebiete, von welchem er handelt, in klarer,
leichtfasslicher Form, aber doch unter vollständiger Wahrung des
wissenschaftlichen Standpunktes, verfasst werden. Soweit es der Inhalt
erfordert, werden Abbildungen, welche den Text ergänzen und zum
bessern Verständnis desselben dienen, beigegeben werden.
In der Reihe selbst werden Originalarbeiten deutscher Gelehrten und
Forscher mit Übersetzungen von Werken hervorragender ausländischer
Autoren abwechseln.
In Vorbereitung sind folgende Bände:
E. Gerland: Geschichte der Physik.
E. L. Trouessart: Die geographische Verbreitung der Tiere.
W. Marshall: Der Bau der Vögel.
H. Gadeau de Kerville: Leuchtende Pflanzen und Tiere.
W. Marshall: Das Leben der Vögel.
C. Chun: Das Tierleben auf der Oberfläche des Meeres.
Die Bände erscheinen in Zwischenräumen von mehreren Monaten und sind
einzeln käuflich.
Verlagsbuchhandlung von J. J. Weber in Leipzig.
Webers Naturwissenschaftliche
Bibliothek.
Dritter Band:
Die
Sinne und Sinnesorgane
der
niederen Tiere.
Von
E. Jourdan.
Aus dem Französischen übersetzt
von
William Marshall.
Mit 48 in den Text gedruckten Abbildungen.
Preis in Original-Leinenband 3 Mark.
Inhalt:
Erstes Hauptstück.
Kurze Übersicht über den allgemeinen Bau der Organismen.






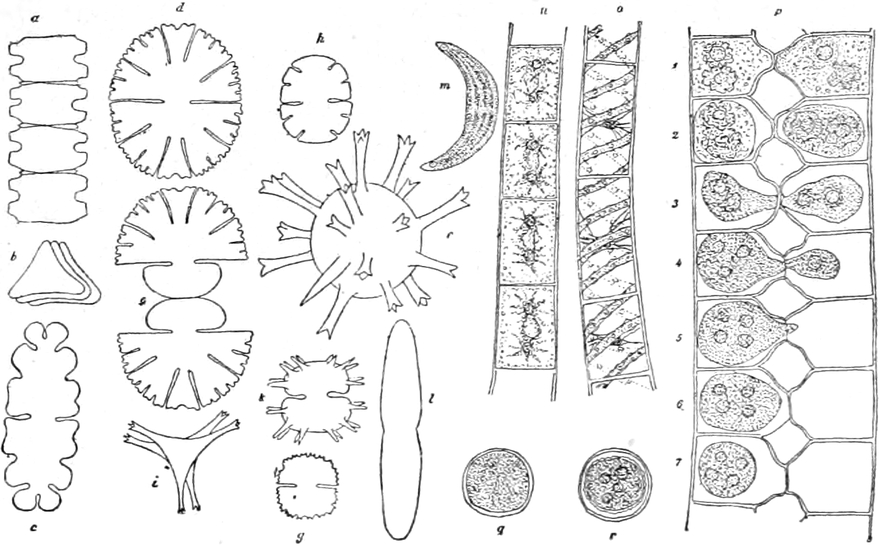









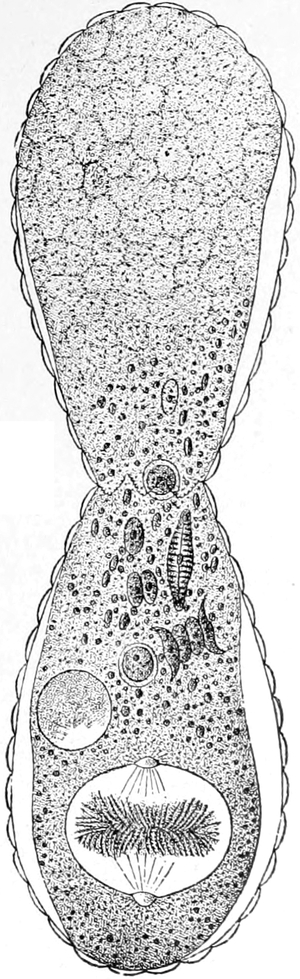












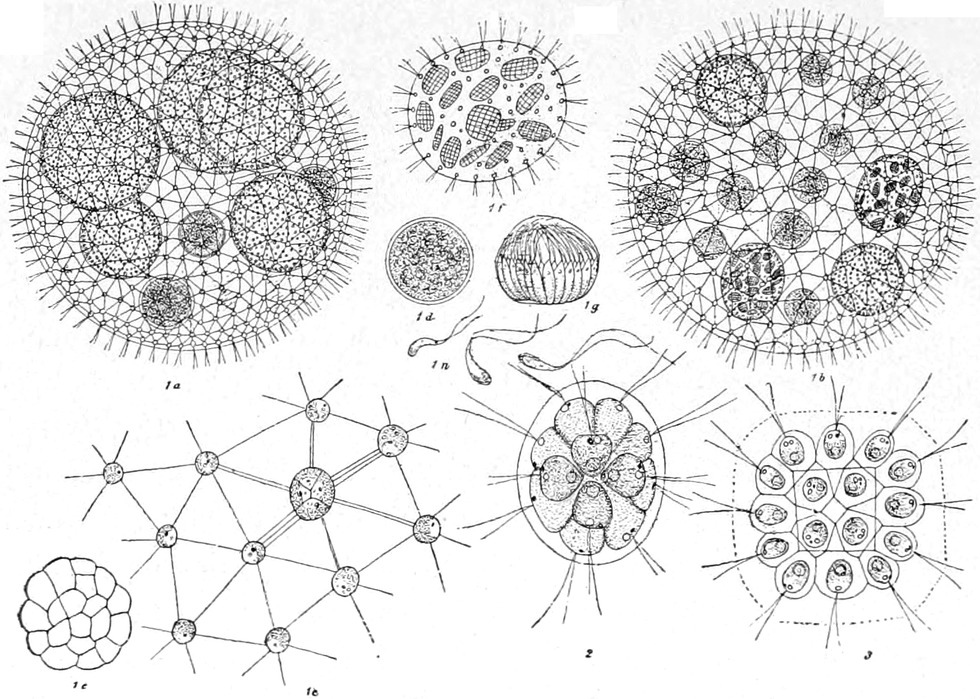


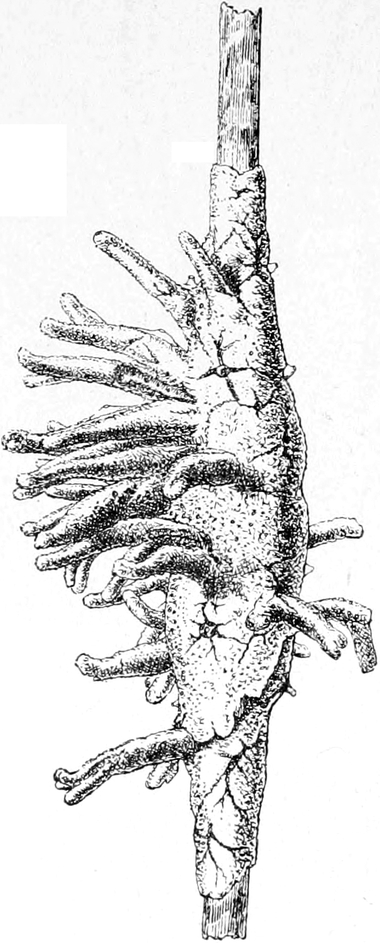
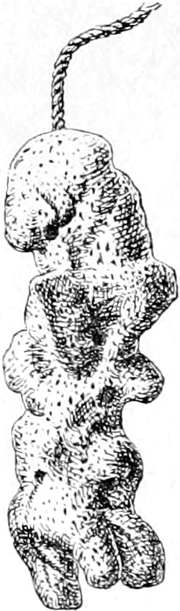


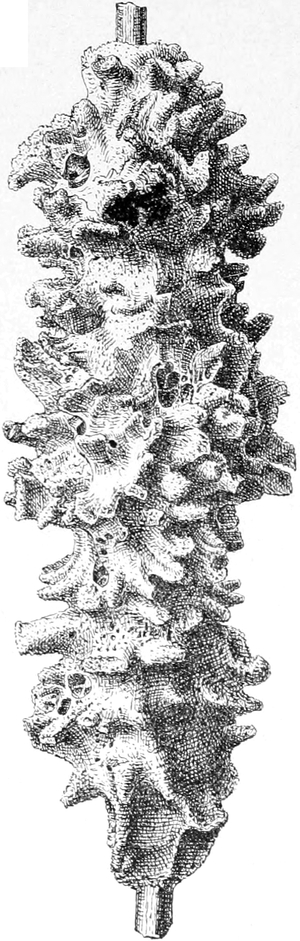









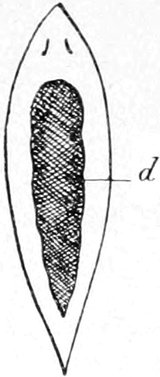



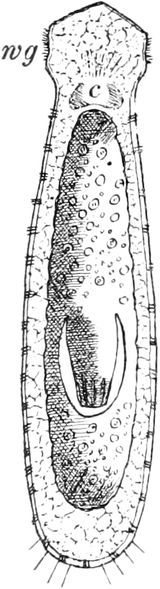





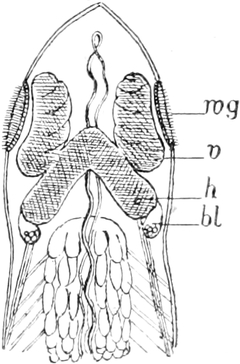






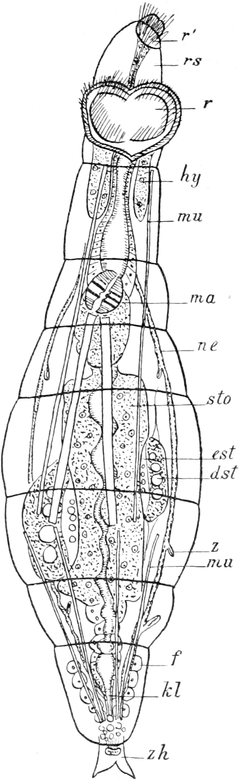





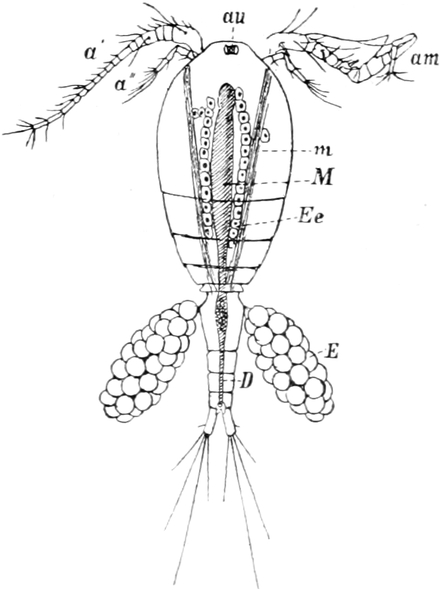
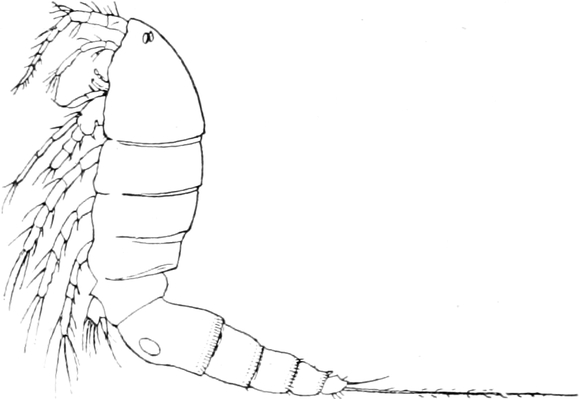


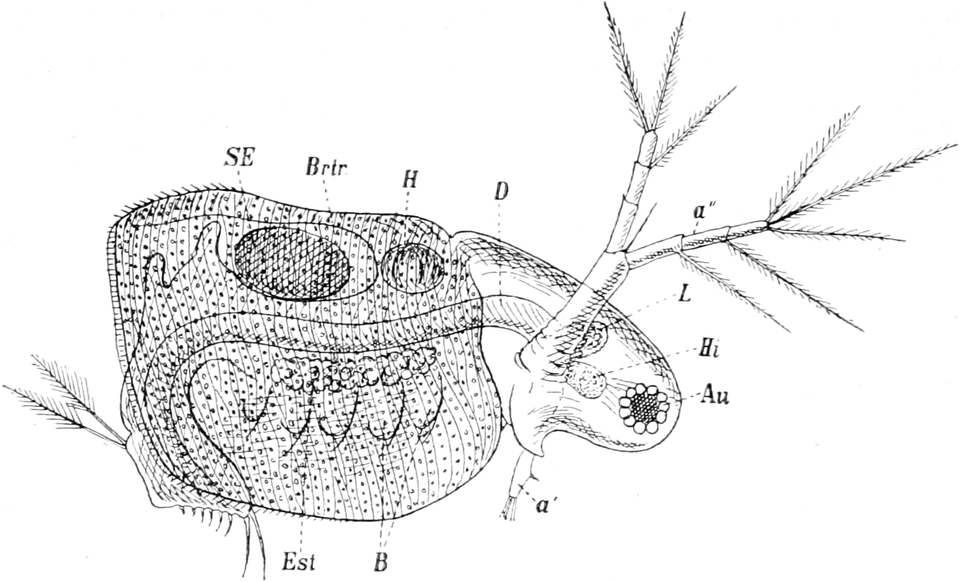



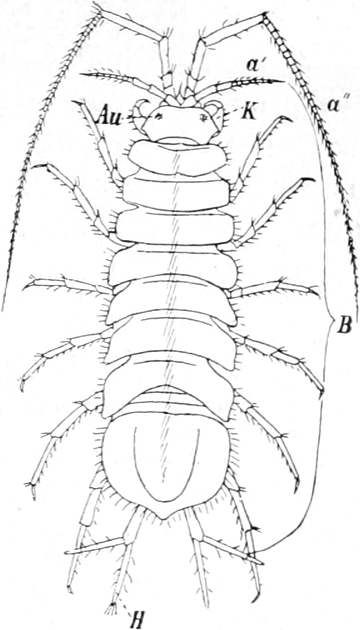




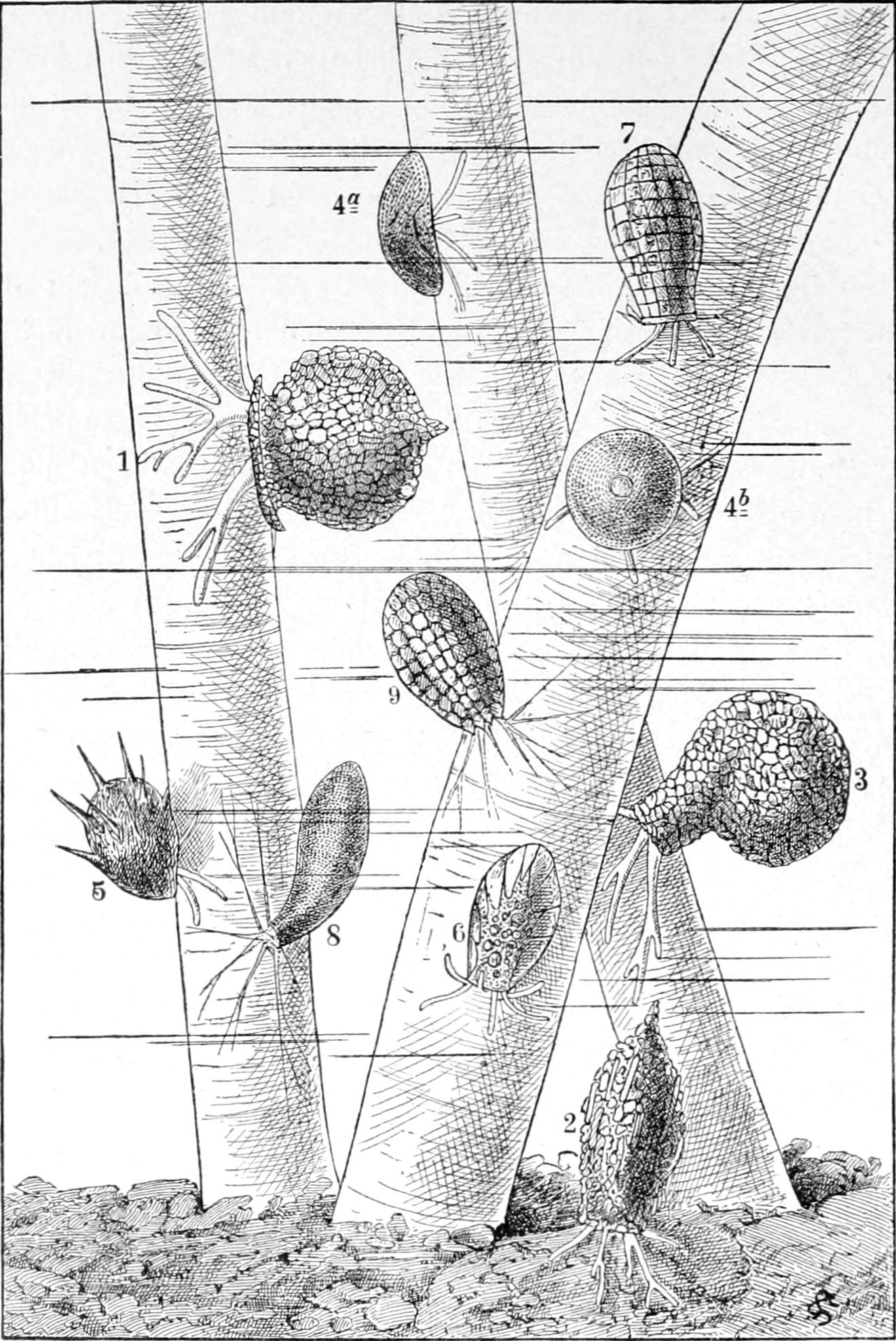{kind=link}