
The cover image was created by the transcriber and is placed in the public domain.
Project Gutenberg's Rust, Smut, Mildew, & Mold, by Mordecai Cubitt Cooke This eBook is for the use of anyone anywhere in the United States and most other parts of the world at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org. If you are not located in the United States, you'll have to check the laws of the country where you are located before using this ebook. Title: Rust, Smut, Mildew, & Mold Author: Mordecai Cubitt Cooke Release Date: October 9, 2020 [EBook #63416] Language: English Character set encoding: UTF-8 *** START OF THIS PROJECT GUTENBERG EBOOK RUST, SMUT, MILDEW, & MOLD *** Produced by deaurider, Barry Abrahamsen, and the Online Distributed Proofreading Team at https://www.pgdp.net (This file was produced from images generously made available by The Internet Archive)
The cover image was created by the transcriber and is placed in the public domain.

Plate VII.
W. West imp.
THE BRITISH FUNGI: A Plain and Easy Account of British Fungi: with especial reference to the Esculent and other Economic Species. Illustrated with Coloured Plates of 40 Species. 2nd Edition. Fcap. 8vo., price 6s.
“The author is a thorough mycophagist, well acquainted with the peculiar features by which the most remarkable of the edible kinds of Fungi may be known.”—Gardeners’ Chronicle.
“A very readable volume upon the lowest and least generally understood race of plants. For popular purposes the work could not have been better done.”—Athenæum.
INDEX FUNGORUM BRITANNICORUM: A Complete List of Fungi found in the British Islands to the Present Date. Arranged so as to be applicable either as a Check-List or for Herbarium Labels. Royal 8vo., price 2s. 6d.
THE first edition of this Work having for some time been out of print, and the demands of the public encouraging the publisher to proceed with a new edition, I have added, in a second Appendix, descriptions of all the species discovered in Britain since 1865, so far as they relate to the Orders included in this volume. The success which has attended the sale of this Work, and the number of fresh observers it has brought into the field, has greatly tended to the necessity for a second Appendix. A larger number of observers, over a still more extended area, will, it is hoped, add further to our list; by increasing the number of known species. Hitherto one great cause of the paucity of students of Fungi in this country, especially of the Microscopic forms, has been the want of text-books on the subject, containing descriptions of the species, with figures illustrative of the genera. Although this little volume only partly supplies that want, by including the species found on living plants alone, it has already proved of service; this and its companion volume, “Introduction to British Fungi,” being (with but one exception) the only books on Fungi which have passed into a second edition in this country; a fact which appears to prove that they have succeeded in furnishing a desideratum, and in giving an impetus to the study. It is hoped that similar results will follow the publication of this new edition.
| CHAP. | PAGE | |
| I. | Cluster-Cups | 1 |
| II. | Spermogones | 22 |
| III. | Di-morphism | 33 |
| IV. | Mildew and Brand | 45 |
| V. | Complex Brands | 67 |
| VI. | Smuts | 77 |
| VII. | Complex Smuts | 90 |
| VIII. | Rusts | 95 |
| IX. | Rusts (continued) | 110 |
| X. | White Rusts | 124 |
| XI. | Moulds | 138 |
| XII. | White Mildews or Blights | 162 |
| XIII. | Suggestions | 179 |
| Appendix, Classification, and Descriptions of Fungi contained in this volume | 189 | |
| Appendix II. | 223 | |
| Index | 239 |
IN these latter days, when everyone who possesses a love for the marvellous, or desires a knowledge of some of the minute mysteries of nature, has, or ought to have, a microscope, a want is occasionally felt which we have essayed to supply. This want consists in a guide to some systematic botanical study, in which the microscope can be rendered available, and in which there is ample field for discovery, and ample opportunity for the elucidation of facts only partly revealed. Fungi, especially the more minute epiphyllous species, present just such an opportunity as many an ardent student would gladly take advantage of; one great obstacle to the pursuit being hitherto found in the absence of any hand-book to this section of the British Flora, embracing the emendations, improvements, and additions of the past twenty-seven years (the period at which the fifth volume of the “English Flora” made its appearance). 2It would be incompatible with our object, and beyond our limits, to introduce an entire mycological flora to our readers in these pages; but we hope to communicate such information as will serve to prepare the way still more for such an additional Flora, should it ever be produced, and render the demand still wider and more general for such an extension of our botanical literature. It is true that one work has of late years issued from the press on this subject, but notwithstanding its utility to scientific men as a record of species, it is practically useless to those we address, from the absence of all specific descriptions of microscopic fungi.
Let not the reader imagine, from what we have just stated, that it is our intention to burden him with a dry series of botanical descriptions; as much of this as we deem essential to render the book available to the botanical student, we have preferred to add in the form of an Appendix. Useful as these may be to some, we hope to be enabled to furnish for others something more; and although we at once disclaim any intention of including all the microscopic, or even the epiphytal fungi, in our observations, yet we trust, by a selection of common and typical species for illustration, and by an adherence to certain well-defined groups and sections, to demonstrate that the microscopist will find an eligible field for his observations in this direction, and the botanical student may gain 3some knowledge of their generic and specific distinctions.
It is exceedingly difficult to give a logical definition of what constitutes a fungus. It is no less difficult to furnish a popular description which shall include all and nothing more. If, for example, we particularize the spots and markings on the leaves and stems of herbaceous plants, so commonly met with from early spring till the fall of the last leaf, and even amongst the dead and decaying remains of the vegetation of the year, we may include also such spots and marks as result from insect depredations or diseased tissue. It is not always easy, with a cursory observation under the microscope, to determine whether some appearances are produced by fungi, insects, or organic disease: experience is the safest guide, and until we acquire that we shall occasionally fail.
If we take a stroll away from the busy haunts of men, though only for a short distance,—say, for example (if from London), down to New Cross,—and along the slopes of the railway cutting, we shall be sure to find the plant called the goatsbeard (Tragopogon pratensis) in profusion. In May or June the leaves and unopened involucres of this plant will present a singular appearance, as if sprinkled with gold-dust, or rather, being deficient in lustre, seeming as though some fairy folk had scattered over them a shower of orange-coloured chrome or turmeric powder. Examine this singular 4phenomenon more closely, and the poetry about the pixies all vanishes; for the orange powder will be seen to have issued from the plant itself. A pocket lens, or a Coddington, reveals the secret of the mysterious dust. Hundreds of small orifices like little yellow cups, with a fringe of white teeth around their margins, will be seen thickly scattered over the under surface of the leaves. These cups (called peridia) will appear to have burst through the epidermis of the leaf and elevated themselves above its surface, with the lower portion attached to the substratum beneath. In the interior of these cup-like excrescences, or peridia, a quantity of the orange-coloured spherical dust remains, whilst much of it has been shed and dispersed over the unoccupied portions of the leaves, the stems, and probably on the leaves of the grass or other plants growing in its immediate vicinity. These little cups are fungi, the yellow dust the spores,[1] or ultimate representatives of seed, and the epiphytal plants we have here found we will accept as the type of the group or order to which we wish to direct attention (Plate I. figs. 1-3).

Plate I.
W. West imp.
1. Protospores they should be called, because, in fact, they germinate, and on the threads thus produced the true spores, or fruit, are borne.
Amongst the six families into which fungi are divided, is one in which the spores are the principal feature, as is the aurantiaceous dust in the parasite of the goatsbeard. This family is named 5Coniomycetes, from two Greek words, meaning “dust-fungi.” This group or family includes several smaller groups, termed orders, which are analogous to the natural orders of flowering plants. Without staying to enumerate the characteristics of these orders, we select one in which the spores are enclosed in a distinct peridium, as in our typical plant they are contained within the cups. This order is the Æcidiacei, so called after Æcidium, the largest and most important of the genera included within this order.
The Æcidiacei are always developed on living plants, sometimes on the flowers, fruit, petioles, or stems, but most commonly on the leaves: occasionally on the upper surface, but generally on the inferior. The different species are distributed over a wide area; many are found in Europe and North America, some occur in Asia, Africa, and Australia. When the cryptogamic plants of the world shall have been as widely examined and as well understood as the phanerogamic plants have been, we shall be in a better position to determine the geographical distribution of the different orders of fungi. In the present incomplete state of our knowledge, all such efforts will be unsatisfactory.
But to return to the goatsbeard, and its cluster-cups. The little fungus is called Æcidium tragopogonis, the first being the name of the genus, and the last that of the species. Let us warn the young student against falling into the error of supposing 6because in this, and many other instances, the specific name of the fungus is derived from the plant, or one of the plants, upon which it is found, that therefore the species differs with that of the plant, and that, as a rule, he may anticipate meeting with a distinct species of fungus on every distinct species of plant, or that the parasite which he encounters on the living leaves of any one plant is necessarily specifically distinct from those found on all other plants. One species of Æcidium, for instance, may hitherto have been found only on one species of plant, whereas another Æcidium may have been found on five or six different species of plants. The mycologist will look to the specific differences in the parasite without regard to the identity or distinctness of the plant upon which it is parasitic.
Before the Æcidium breaks through the epidermis, the under surface of the leaves of the goatsbeard will appear to be covered with little elevations or pustules, paler at the apex; these soon become ruptured, and the fungus pushes its head through the opening, at the same time bursting by radiating fissures. The teeth thus formed resemble those of the peristome of some mosses. All around the orifice of the peridium the teeth become recurved, and the orange spores are exposed, crowded together within. At first, and while contained within the peridium, these spores are concatenate or chained together, but 7when dispersed they are scattered singly about the orifice, often mixed with the colourless cells arising from the partial breaking up of the teeth of the peridium.
Let us pause for a moment in our examination of the individual cups, to ascertain their manner of distribution over the leaves. In this instance they are scattered without any apparent order over the under surface, but generally thickest towards the summit of the leaves; occasionally a few are met with on the upper surface. Sometimes two or three touch at the margins, but we have never met with them truly confluent; generally there is a space greater than the width of the cups around each, the stratum or subiculum from whence they arise is scarcely thickened, and there are no spots or indications on the opposite surface. If a leaf be taken fresh and the cuticle stripped off, which it will sometimes do very readily, the orifices through which the Æcidium has burst will appear in irregular holes. If a section be made of one or two of the fungi in situ, they will be seen to spring from beneath the cuticle, the peridium to be simple, and rounded at the base, the spores clustered at the bottom, and the fringe to be a continuation of its cellular substance.
The spores in this species are orange, subglobose, sometimes angular, and indeed very variable both in size and form, though the majority are comparatively large. Each of these bodies is, 8doubtless, capable of reproducing its species, and if we compute 2,000 cluster-cups as occurring on each leaf, and we have found half as many more on an ordinary-sized leaf, and suppose each cup to contain 250,000 spores, which again is below the actual number, then we shall have not less than five hundred millions of reproductive bodies on one leaf of the goatsbeard to furnish a crop of parasites for the plants of the succeeding year. We must reckon by millions, and our figures and faculties fail in appreciating the myriads of spores which compose the orange dust produced upon one infected cluster of plants of Tragopogon. Nor is this all, for our number represents only the actual protospores which are contained within the peridia; each of these on germination may produce not only one but many vegetative spores, which are exceedingly minute, and, individually, may be regarded as embryos of a fresh crop of cluster-cups. And this is not the only enemy of the kind to which this unfortunate plant is subject, for another fungus equally prolific often takes possession of the interior of the involucre wherein the young florets are hid, and converts the whole into a mass of purplish black spores even more minute than those of the Æcidium, and both these parasites will be occasionally found flourishing on the same plant at the same time (Plate V. figs. 92-94).
Naturally enough, our reader will be debating within himself how these spores, which we have 9seen, are shed in such profusion, can enter the tissues of the plants which give subsequent evidence of infection; in fact, how the yellow dust with which the goatsbeard of to-day is covered will inoculate the young plants of next year. If one or two of these spores are sprinkled upon the piece of the cuticle which we have recommended to be removed from the leaf for examination, it will be seen that they are very much larger than the stomata or breathing-pores which stud the cuticle: hence it is clear that they cannot gain admittance there. There remains but one other portal to the interior of the plant—namely, the spongioles, or extremities of the roots. Here another difficulty arises; for the spores are as large as the cells through which they have to pass. This difficulty may be lessened when we remember that what are termed the spores which are discharged from the cups are not the true spores, but bodies from which smaller seed-like vesicles are produced; yet, even then there will be much need of an active imagination to invent hypotheses to cover the innumerable difficulties which would encounter their passage through the vessels of the infected plants. The Rev. M. J. Berkeley proved many years ago that the spores of bunt, for example, may be caused to infect all the plants the seeds of which had been placed in contact with them; but this affection did not necessarily accrue from the absorption of the 10spores, or the ultimate sporidia produced after three or four generations. It is possible that the granular or fluid contents of the spores may be absorbed by the plant, and as a result of this absorption, become inoculated with the virus, which at length breaks out in fungoid growths. Much has been done to elucidate this mystery of inoculation, but much also remains a mystery still. There is no doubt that the inoculation takes place at an early age,[2] probably in the seeds of many plants; in others it may be conveyed with the moisture to the roots; but the spores themselves have certainly not yet been traced traversing the tissues of growing plants.
2. Dr. de Bary has lately shown that in many similar instances the seed-leaves are inoculated. It will be necessary to refer more particularly to his experiments hereafter.
If, instead of going in search of goatsbeard and its attendant fungus, we turn our steps northward and enter one of the Highgate or Hampstead woods, where the pretty little wood-anemone (Anemone nemorosa) flourishes abundantly, and turn up the radical leaves, one by one, and examine their under-surfaces, we shall at length be rewarded by finding one covered with similar cluster-cups to those we have been describing as occurring on the goatsbeard, but far less commonly. Leaf after leaf will be found covered with the brown spots of another fungus called Puccinia anemones, with which nearly every plant will be 11more or less infected in the spring of the year; and at length, if we persevere, the anemone cluster-cup (Æcidium leucospermum) will be our reward (Plate I. figs. 4-6). The specific name will suggest one point of difference between the two fungi, as in this instance the spores are white, and somewhat elliptic. Probably this species is not common, as we have found it but seldom, though often in search of it. A nearly allied species has been found on Anemones in gardens, having but few large teeth about the orifice, though not constantly four, as the name would indicate (Æ. quadrifidum).
A walk through almost any wood, in the spring of the year, will reward the mycologist with another cluster-cup (Æcidium), in which the peridia are scattered over the whole surface of the leaf. This will be found on the wood spurge, giving a sickly yellowish appearance to the leaves, on the under surface of which it is found. By experience one may soon learn to suspect the occurrence of parasites of this nature on leaves, from the peculiar exhausted and unhealthy appearance which they assume as the spores ripen, and which will spare the labour of turning over the leaves when there are no distinct spots on the upper surface. Æ. Euphorbiæ is found on several species of Euphorbium or spurge, but we have always found it most abundantly on the wood spurge in the Kentish woods between Dartford 12and Gravesend. The spores in this species are orange, and externally it bears considerable resemblance to the goatsbeard cluster-cup, but the spores are rather smaller and paler, the teeth are less distinct and persistent, the subiculum is more thickened, and the peridia are more densely crowded.
There is another group of species belonging to the same genus of fungi in which the arrangement of the peridia is different. One of the first of our native wild flowers, in making its appearance after the departure of frost and snow, is the little yellow celandine (Ranunculus ficaria).
And one of the earliest parasitic fungi in spring is an Æcidium which flourishes on its glossy leaves. So common is Æcidium ranunculacearum on this species of Ranunculus, that it can scarcely have escaped the eye of any one who has taken the trouble to examine the plant. It appears in patches on the under surface of the leaves or on their petioles, in the latter case swelling and distorting them. Sometimes these patches are nearly circular, at others of very irregular form, and varying 13in size from less than one-twelfth of an inch to half an inch in diameter. It is found on several species of Ranunculus, as R. acris, bulbosus, and repens, but most commonly on R. ficaria. The leaf is thickened at the spot occupied by the parasite, and generally without indication on the opposite surface. Sometimes one spot, at others several, occur on the same leaf. The peridia are densely crowded together, often arranged in a circinate manner, i.e., like a watch-spring, or the young frond of a fern. The spores are orange, but slightly varying in tint on different species of Ranunculus (Plate II. figs. 12-14). One of the smaller clusters, when collected before the spores are dispersed, or the teeth of the peridium discoloured, mounted dry as an opaque object, makes a very excellent slide for an inch or half-inch objective; and the same may be said of many others of the same genus.
Less common than the foregoing is the species of Æcidium which attacks the violet. The sweetest of flowers as well as the earliest, in despite both of its odour and its humility, becomes a victim to one or more of the ubiquitous race of fungi. Thickened spots at first appear on the leaves; the petioles, or flower stem, or even the calyx, become swollen and distorted; and at length the cluster-cup breaks through. The spots on the leaves upon which the peridia are scattered are yellowish, generally larger than the clusters on the pilewort, and seldom with more than one spot on each leaf. The peridia, or 14cups, are irregularly distributed over the spots, not crowded together as in the last species; and the teeth are large, white, and distinct. The spores are at first orange, but at length become brownish. This species may be found in spring, as late as June, most commonly on the dog-violet, but also on other species of Viola.
It is not a very desirable occupation to search a bed of nettles, and turn over the individual leaves to look for minute fungi. A very pretty Æcidium is nevertheless far from uncommon in such a habitat. Fortunately it occurs very often on the petioles of the leaves and on the stem, distorting them very much; and in such situations flourishing, apparently, more vigorously than when occupying the under surface of the leaves (Plate I. fig. 10). In the latter situation the clusters of peridia are small, seldom exceeding a dozen in a spot, but several spots may be found on the same leaf. On the stem they are clustered around for upwards of an inch in length, and their bright orange colour in such a situation renders them very conspicuous objects. The peridia are always closely packed together upon a thickened base, and offer but slight variations from the forms already enumerated, save that they widen slightly at the mouth, so as to become nearly campanulate. The spores are orange, and very profuse.
During the past summer we noticed, for the first time, a very pretty little species of cluster-cup (Æcidium) on the wood sanicle (Sanicula Europæa) in 15Darenth wood. It was far from uncommon, and we believe it to be specifically distinct from its nearest ally, found on the earthnut leaves, and those of some other umbelliferous plants. The little cups are in small clusters of four or five together, on the under surface and on the petioles; they are small, but the teeth are relatively large, white, and distinct. The spores are of a pallid, yellowish colour, and not so profuse as in the last species. A darker spot on the upper surface of the leaf generally indicates their presence. This species was found many years ago by Carmichael at Appin, and called by him Æcidium saniculæ; but we find no notice of its occurrence since, though it seems to be far from uncommon at Darenth, and probably elsewhere, should the sanicle be common also.
Recently we found the bedstraw cluster-cup (Æcidium galii) on the great hedge bedstraw (Galium mollugo), and as it has not been figured before, we have included it amongst our illustrations (Plate II. figs. 15-17). Though very insignificant when occurring on the small leaves of the yellow bedstraw (Galium verum), it is a prominent object on the above-named species.
We received, for the first time, in July, 1864, from Mr. Gatty, student at Winchester, a portion of a plant of Thesium humifusum (which is by no means common in Britain), covered with beautiful cluster-cups of a species never before recorded as occurring in this country (Plate III. figs. 50, 51) 16named Æcidium Thesii, but which is far from uncommon on the Continent. It occurred in this instance on the Downs, in the vicinity of Winchester.
It is unnecessary here to refer to other allied species of Æcidium, except one to be presently noticed, since we have, at the end of the volume, enumerated and given descriptions of all the species hitherto found in Britain. Suffice it to say that the Buckthorn cluster-cups on the alder buckthorn (Rhamnus frangula), is usually very common in the Highgate and Hornsey woods, and on the common buckthorn (Rhamnus catharticus) in the neighbourhood of Dartford, in Kent. That on the honeysuckle we have found but very rarely. On the gooseberry and red-currant leaves, commonly in some years and rarely in others; whilst a few of those described we have never collected. The species on different composite plants is subject to great variation, and on most may be found in the autumn; one variety only, on the leaves of Lapsana communis, we have met with in the spring.
Very few years ago farmers generally believed that the cluster-cups of the berberry (Berberis vulgaris), were productive of mildew in corn grown near them; this opinion even received the support of Sir J. Banks, but no fungi can be much more distinct than those found on corn crops and this species on the leaves of the berberry. In this instance the cups are much elongated, and cylindrical, 17the clusters vary much in size, and the spots on the upper surface of the leaf are reddish, bright, and distinct. The teeth are white and brittle, and the orange spores copious (Plate I. figs. 7-9).
There are scarcely any of the epiphyllous fungi forming equally handsome or interesting objects for low powers of the microscope, than the genus to which attention has just been directed; and they possess the advantage of being readily found, for that locality must be poor indeed which cannot furnish six species during the year. We have found half of the number of described species within little more than walking distance of the metropolis, within a period of little more than three months, and should be glad to hear of the occurrence of any of the rest.
We have three species of fungi very similar in many respects to the foregoing, but differing in others to such an extent as to justify their association under a different genus and name. The hawthorn is a bush familiar to all who love the “merry month of May,” but it may be that its parasite has been unnoticed by thousands. If, for the future, our readers will bear this subject in their minds when they stand beneath a hawthorn hedge, they may become acquainted with clusters of singular brown pustules on the leaves, petioles, and fruit well worthy of more minute examination (Plate II. fig. 22). They scarcely claim the name of cups, and their lacerated and fringed margins rather 18resemble the pappus crowning the fruits of some composite plants than the cups of Æcidium. The peridia are very long, and split down throughout their length into thread-like filaments of attached cells; these gradually fall away and break up into their component parts till but short portions remain attached to the base of the peridia. These cells are elongated and marked on the surface with waved lines, forming in themselves pretty objects for a high power of the microscope (Plate II. figs. 23, 24). If the teeth of Æcidium resemble the peristome of some mosses, such as Splachnum; the threads of this species of Rœstelia, except in not being twisted, somewhat resemble the peristomes of other mosses of the genus Tortula. The spores in this species are less conspicuous, being of a light brown, and the whole plant, from its modest hue, may be readily passed over without attracting attention unless occurring in abundance.
The leaves of pear-trees afford a second species of this genus sufficiently distinct to commend it to our notice. Sometimes it is very common, at others but few examples are to be met with. The clusters occur on the under surface, and consist of half-a-dozen or less of large peridia, pointed at the apex and swelling in the middle so as to become urn-shaped (Plate II. figs. 20, 21). These vessels or thecæ split into numerous threads or laciniæ, which remain united together at the apex. Like the species already noticed, this is brown and inconspicuous 19except on account of its size, for it is the largest of all that we have had occasion to notice.
The third species occurs on the under surface of the leaves of the mountain-ash. The peridia are clustered on a rusty orange-coloured spot which is visible on the upper surface (Plate II. figs. 18, 19). They are long and cylindrical, with an evident tendency to curvature; the mouth is serrated, but not split up into threads, as in the species found on the hawthorn. There will often be found instead of well-developed peridia, what at one time were regarded as abortive peridia, forming a thickened orange or rust-coloured spot, studded with minute elevated points. These spots are clusters of spermogones, which organs are described in detail in our second chapter. The clusters and spores are of a brighter reddish-brown than in either of the other species. All are remarkably distinct, and perhaps the most curious and interesting of any that we have passed in review. To botanists, the species found on the hawthorn is known as Rœstelia lacerata, that on pear-leaves as Rœstelia cancellata, and the one on the leaves of the mountain-ash as Rœstelia cornuta.
Dr. Withering observed the spore-spots on the leaves of the mountain-ash, but was evidently puzzled to account for them. He writes (in his Arrangement of British Plants), “The spots on the leaves of Sorbus aucuparia consist of minute globules intermixed with wool-like fibres. On 20examining many of them in different states, I at length found a small maggot in some of the younger spots, so that the globules are probably its excrement, and the fibres, the woody fibres of the plant unfit for its food.” We now-a-days smile at such simple and singular conjectures. It affords evidence of the manner in which the speculations of one generation become follies in the next.
Only two species of cluster-cups are described in Withering’s Flora under the genus Lycoperdon: one of these is now called Æcidium compositarum, and is found on various composite plants; the other includes the species found on the wood-anemone and that on the moschatel, and also probably a species of Puccinia on the wood-betony.
To render this chapter more complete, though of less importance to the microscopist, we may allude to the other two genera comprised within this order. Peridermium is the name of one genus which contains two British species found on the leaves and young shoots of coniferous trees. In this genus the peridium bursts irregularly, and does not form cups, or horns, or fringed vessels. The most common species is found on the needle-shaped leaves of the Scotch fir (Plate II. fig. 27), and also on the young twigs, in the latter instance larger and more prominent than in the former. The elongated peridia burst irregularly at their apices without forming teeth (fig. 28).
In the genus Endophyllum, as its name implies, 21the peridium is imbedded within the substance of the succulent leaves. The only species we possess is found rarely upon the common house-leek.
We have derived much pleasure in viewing the astonishment and delight exhibited by friends to whom we have personally communicated specimens of the little fungi we have enumerated for examination under the microscope; and we recommend with confidence this group of parasitic plants, unfortunately so little known, as well worthy of the attention of all who are interested in the minute aspects of nature, and who can recognize the hand—
IN addition to their spore-bearing spots, lichens have for some time been known to possess other organs, termed spermogones, which are probably concerned more or less in the reproductive process. The first intimation of the existence of similar bodies in the entophytal fungi originated with M. Unger in 1833, but it was left to Dr. de Bary and the Messrs. Tulasne, twenty years later, to examine and determine satisfactorily the nature and value of the spermogones of the Uredines. It was at first believed that the smaller pustules—which sometimes precede, and sometimes accompany, the cluster-cups and some other allied fungi—were distinct species developed simultaneously therewith, or members of a new genus, which, under the name of Æcidiolum exanthematum, found a place in the mycologic system.
Without staying to trace the stages through which the elucidation of their true nature proceeded, it will suffice for our purpose to tell what is now known of these secondary organs; to accomplish which we must stand greatly indebted to the independent researches of Messrs. de Bary and Tulasne. It has been demonstrated that both these bodies, namely, the primary organs or cluster-cups, 23and the secondary organs or spermogones, are developed from the same mycelium; but the value of the latter is still undetermined. If they possess any fecundative power, the process has not been traced; or if they are in themselves reproductive, they have not at present been seen to germinate. Their uses, therefore, in the economy of the parasitic plant of which they are now known to form a part is still a mystery, and they remain valueless in the determination of genera and species. Any speculation which might regard them as male organs would be premature, and without support in fact. Hitherto only some species of the genera described in the foregoing chapter, and others belonging to genera not hitherto named, have been ascertained to possess spermogones. Of the former are the Rœsteliæ, some species of Æcidium, as those of Euphorbia, &c., and Peridermium Pini.
These spermogones are of a very simple structure—very delicate, indeed; so much so, that they will scarcely bear preparation for demonstration. De Bary states that they originate from plain, delicate, inarticulate threads, about half the thickness of the mycelium (the root-like branching fibres which form the fundamental stratum of fungoid growths), which are developed in large quantities, and closely packed together. These threads are compacted together so as to form an outer enveloping integument or peridium, which is 24either globular or hemispherical (or in some instances elongated), more or less immersed, and at length opening at the apex (fig. 153) by a regularly formed minute ostiolum. The inner wall of the peridium is covered with a thick forest of simple filaments standing on end. From the summit of these filaments or sterigmata, the spermatia are borne. These are either isolated or associated together in strings or chaplets, are exceedingly minute, of an ovoid or oblong shape, and are produced in such numbers as to fill the cavity of the spermogone. Besides these, a viscid fluid is secreted, in which the spermatia are immersed, and which is expelled with them from the orifice of the peridium. According to the density of this fluid, or the hygrometric state of the atmosphere, it appears sometimes in drops, and sometimes oozing out in threads or cirrhi from the spermogones. To compare minute things with gigantic, as a recent author has observed, it resembles the lava issuing from the crater of a volcano. The colour of this spermatiferous matter is commonly orange, but in some instances brown, though not constantly of the same colour as the spores produced from the same mycelium. This gelatinous substance is dissolved away from the granular bodies which are immersed in it, by adding a little water upon the slide on which the mass is placed for examination. The granules, or spermatia, then exhibit those peculiar movements 25which have been observed in the similar bodies in lichens, and fitly described as “a sort of oscillating motion, as of a body attached at one extremity.” The cause of this motion is at present uncertain, vibratile ciliæ, to which similar movements are referred, being altogether absent; but probably, as De Bary believes, the cause may be found in the influence of exosmose.
The largest spermatia yet examined (those of Peridermium Pini) have a length equal to 1/2500 of an inch, but their width seldom exceeds 1/100000 of an inch, whilst in others their length does not exceed the width of those just named.
Messrs. Tulasne affirm that all these corpuscles, as well as the mucilaginous fluid, evolve an appreciable odour, resembling that of the pollen of the willow. M. Léveillé compares the odour to that of orange flowers, and M. de Bary to that of the evening primrose.
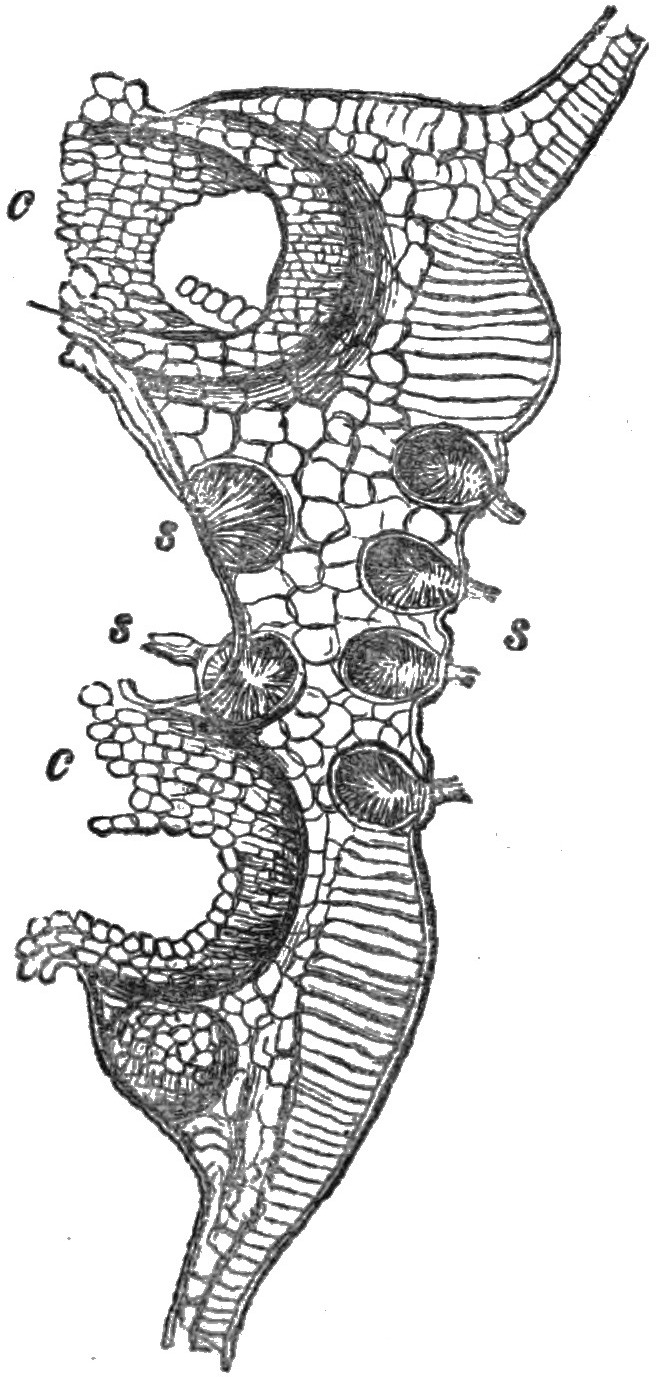
Fig. 1.—Æcidium grossulariæ. c. Cluster-cups. s. Spermogones.
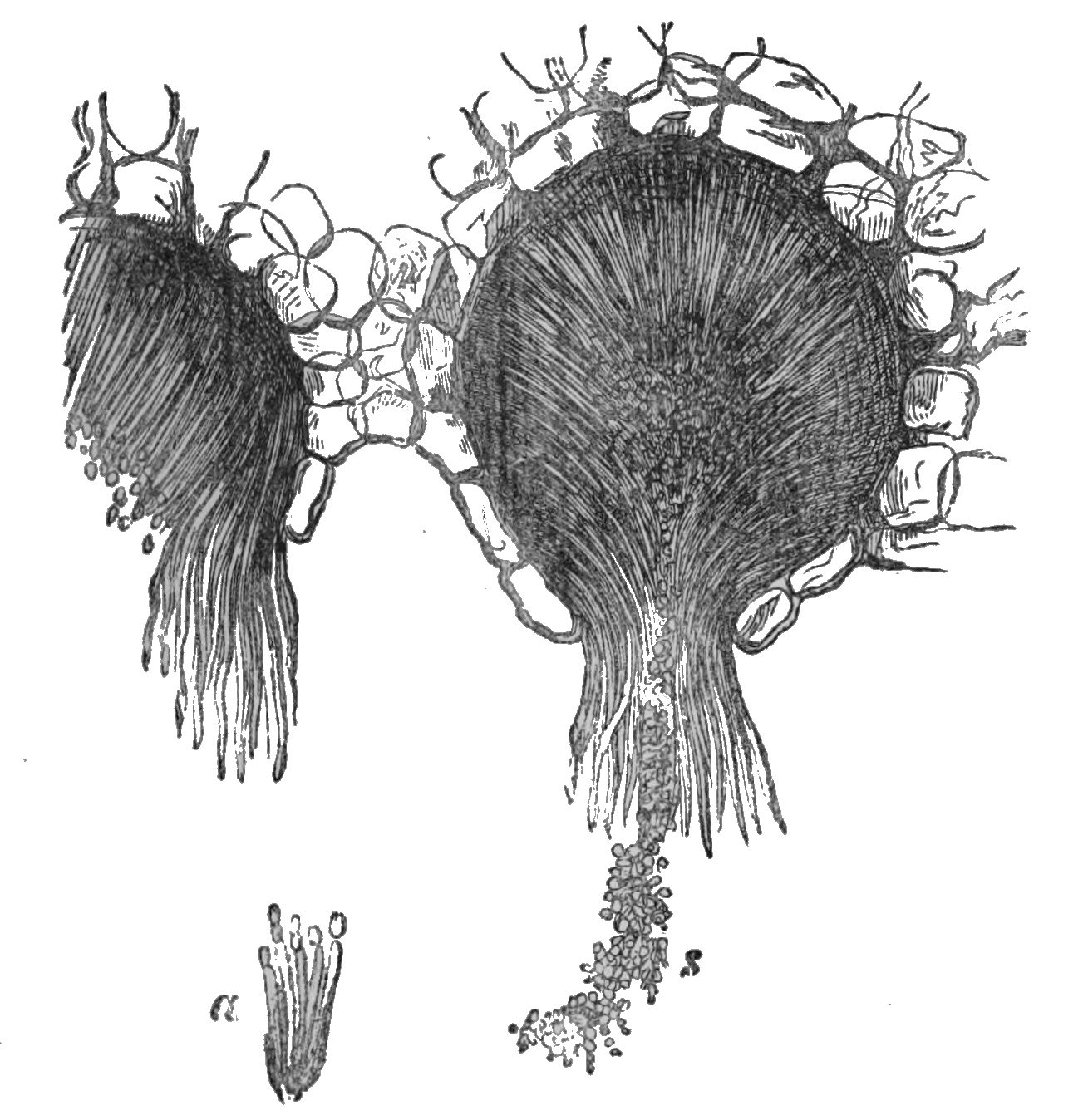
Fig. 2.—Section of ripe spermogones of Æcidium Euphorbiæ. s. Spermatia. a. Sterigmatæ bearing spermatia (De Bary).
The spermogones do not always appear like pustules on the surface of the leaves, for sometimes their presence is only indicated by minute depressed punctures which are scarcely visible; generally, however, they may be recognized by an obtuse, or otherwise a pointed, protuberance that surmounts them. The margin of the orifice is sometimes furnished with short hairs, but is more frequently ornamented with a pencil of long hairs, which are stiff and erect, and of the colour of the enclosed spermatia.
26In many of the species of Æcidium the cups are disposed in a more or less regular circle, the centre of which is occupied by a group of spermogones; at the same time, the corresponding spot on the opposite surface of the leaf will frequently be found also occupied by other spermogones—in some instances in greater number than on the same surface of the leaf on which the cups are seated. This is the case in the Æcidium which is found upon the leaves of the coltsfoot, and that of the honeysuckle.
Very bright orange-coloured spots may be observed in autumn (we have encountered them often in August and September) upon the leaves of pear trees, and which are covered with little tubercles, at first of the same colour, but ultimately becoming brown. These pustules are so many spermogones belonging to Rœstelia cancellata, a kind of cluster-cup found in the same localities. These spots have long since been noticed, and regarded as connected with the Rœstelia, but in what manner has until recently been unknown. The Rev. M. J. Berkeley noticed them in the English Flora in 1836, or at least the granulations on the upper surfaces of the leaves bearing R. cancellata, R. cornuta, and R. lacerata, and called them abortive pseudoperidia. Before this (in 1804) they had been observed by Rebentisch. An examination of one of these spots under a low power of the microscope, and afterwards a section of one or more of the pustules, cut with a sharp razor, and viewed with a higher power, will give an 29idea of the nature of the bodies we are attempting to describe. During the past summer we have noticed very similar orange spots on leaves of the berberry containing spermogones on both surfaces, and these appeared before any cups had been found on that plant. In this instance no cups were produced from the spots on the leaves examined, and which were carefully noticed at intervals until they withered and fell.
In some instances, as in Rœstelia cornuta, which is found on the leaves of the mountain-ash, the cups are produced on the lower, but the spermogones almost exclusively on the upper surface.
The spermogones of Peridermium Pini are white, few in number, and are developed, not only in the spring, but sometimes reappear in the autumn upon the same leaves that produced them at the commencement of the year.
In such instances as those of the Æcidium of the spurge, and also the goatsbeard, in which the cluster-cups are arranged in no appreciable order, the spermogones are scattered amongst them, and even in some instances appear on different leaves. The spermogones are common on the wood spurge in spring, scattered over both surfaces of the leaves before the cluster-cups make their appearance, and gradually these latter are developed amongst them, commencing from the apex of the leaves and proceeding in the order of their development towards the base. In this instance the spermogones are 30bright yellow, as are afterwards the cups and spores of the Æcidium. In most instances the appearance of the spermogones precedes that of the sporiferous organs, but the latter follow sufficiently speedy for perfect development before the decadence of the spermogones takes place.
After the expulsion of the spermatia and the fluid which accompanies them, the whole mass dries up; and where many spermogones have been clustered together in the same spot a brown homogeneous crust is formed upon the epidermis; where they are produced singly, a brownish incrustation is visible about the mouth of each spermogone.
Re-agents applied to the spermogones whilst in full vitality indicate the presence of a considerable amount of a protein substance, which, with sugar and sulphuric acid, produces a deep purple red colour.
From what we have already stated of the method of occurrence of these organs, the following is the only order, apparently, preserved in their development, although no definite rules can at present be affirmed. The spore spots of cluster-cups are generally found upon the under surfaces of the leaves on which they are produced, and the spermogones are most numerous on the upper. When both the cluster-cups and the spermogones appear in the same group on the same surface, the spermogones commonly occupy the centre, and the cups are arranged in a circular manner about them. In other, and fewer instances, both 31organs stand together indiscriminately upon the same surface.
The spermogones are also developed centrifugally, at least so far as at present observed, for when they are produced in a cluster the central one first opens and discharges its contents, and thus the development proceeds outwards from the centre to the circumference. When the spermogones are scattered, as in those of Euphorbia, they are first observed at the apex of the leaf, whence they are developed in succession towards the base. The latter should be sought for on the young plants of the wood spurge in March or April, at which time we have found them abundant at Darenth wood, near Dartford.
It must not be concluded, from the fact that we have not yet adverted to spermogones in connection with other fungi, that they are peculiar to the Æcidiacei. Such is by no means the case. As we have hereafter described other genera and species in which spermogones occur, it would be out of place to enter upon further details here. Let it suffice therefore that we state that they have been found in members of the genera, Aregma, Triphragmium, Puccinia, Lecythea, Trichobasis, and Uredo, but they have been found much more generally in Rœstelia and Æcidium than in any other genus.
As comparatively little is yet known of these bodies, a fair field is open to the enterprising microscopist, with time at his disposal, and a good 32store of perseverance, to win for himself renown in the discovery of fresh facts, and the elucidation of some of the mysteries which yet enshroud these interesting organisms. From the foregoing pages he will learn the direction in which his researches should tend, and he may be assured that every new fact is of importance when carefully ascertained.

Plate II.
W. West imp.
BEFORE entering further and more fully upon the subject of this volume, it may be advisable to attempt an explanation of a phenomenon of no uncommon occurrence in many groups of Fungi, and which is termed di-morphism.
In the Uredines, or uredo-like fungi, as well as other of the Coniomycetes (in which the spores are the principal feature), the same fungus appears under two or more distinct forms, not necessarily mere differences of age, but so distinct that they have been regarded (and some are so still) as different species belonging to different genera, often far removed from each other, and bearing different names. One plant, for instance, sprinkled over the under surface of a rose-leaf, like turmeric powder, has its mycelium, or root-like threads, penetrating the tissue, whilst bearing above its spherical golden-coloured spores. Its vegetative system is complete, and, apparently, its reproductive also; hence it seems to claim recognition as a perfect plant, and under the name of Uredo Rosæ was so recognized, until microscopical investigation determined otherwise. Thus it has been discovered that certain dark brown spots which appear later 34in the season are produced upon the same mycelium, and are indeed aggregations of more perfect and complex fruits of the same plant. Before this point was satisfactorily decided, the brown spores, which are borne on long stalks, and are themselves septate or divided (apparently or really) by transverse partitions into a complex fruit, received the name of Puccinia Rosæ. At this period, Uredo Rosæ and Puccinia Rosæ, or the yellow fungus and the dark brown fungus, were believed to be distinct and different plants; now, on the contrary, they are believed to be different forms of fruit produced by the same plant; i.e., an instance of di-morphism. Aregma mucronatum, Fr., is the present scientific name of what is regarded as the perfect fungus, whilst the uredo-form either bears the name of Lecythea Rosæ, Lev., or by some mycologists is rejected altogether as a spurious species.
During the summer it is not uncommon to find the leaves of some grasses, of the hop, of roses, and many other plants, covered with a kind of white mould, which appears under the microscope as a multitude of small transparent colourless cellules, generally attached to each other in a moniliform or beaded manner. These moulds were long known under the generic name of Oidium, to which genus the vine disease was also referred. More minute investigation and more careful examination proved that these moulds were not in 35themselves perfect plants, but merely conditions of other fungi of a higher order, little differing it is true in external appearance to the naked eye, but offering material differences in structure under the microscope. Upon the white mould-like threads, spherical bodies are produced in the autumn, containing little sacs or asci filled with spores; and in this condition the plants are arranged under the genus Erysiphe, whilst the species of Oidium which represented their imperfect condition, are excluded from the system. Here, again, we have examples of di-morphism.
In the Journal of the Microscopical Society, Mr. F. Currey has detailed several instances of di-morphism which have fallen within his experience. In one instance he has shown that a small simple spored fungus, termed Cryptosporium Neesii, Ca., is only a state or condition of a fungus with compound fruit, belonging to the Sphœria section of ascigerous fungi, called Valsa suffusa, Fr. Both plants are exactly alike externally, but the perithecium, or flask-like receptacle containing the fructification, in one instance only holds naked spores, and in the other the spores are contained in little elongated vesicular bags or asci, which are packed within the perithecium.
Whilst writing this, one of the most wonderful books in a book-producing age lies beside us; it is the second volume of a work on fungi, by the brothers Tulasne; and this, as well as its predecessor, 36is devoted to this very subject of a multiplicity of form in the fructification of these plants. Illustrated by the most exquisite of engravings which art has ever produced, it also unfolds many a page in the history of these organisms, for which mycologists were not altogether unprepared. In noticing this work, one of our most eminent authors on mycological subjects quotes as an example Dothidia ribis, Fr., one of our most common fungi, which occurs in the form of little black shields on dead twigs of currants and goose-berries. Here we have, he says, naked spores (conidia) growing on the external cells of the stroma; we have naked spores of a second kind (stylospores) produced in distinct cysts (pycnides); we have minute bodies of a third kind (spermatia) produced again in distinct cysts, resembling very closely similar bodies in lichens; and we have a third kind of cysts, containing the usual sporidia in sausage-shaped hyaline sacs (asci). Even here, however, we have not done with marvels; for if the stylospores are placed in water, they produce in the course of twenty-four hours conidia of a second order, exactly analogous to those which arise on the germination of the spores of the rusts and mildews which affect our cereals and other plants.
Further reference is also made to three species of moulds, which M. Tulasne has shown to be only varied forms of the mycelium of a species of 37Sphœria common to various plants; these moulds having been hitherto regarded as fungi perfect in themselves.
In the Uredines, to which much of this volume is devoted, the genera known as Lecythea and Trichobasis are by some mycologists excluded altogether, as containing only species which are mere forms of more highly-developed uredines, such as Puccinia, Aregma, and others. On the other hand, they are retained by those who possess a lingering doubt whether both forms may not be distinct, though developed from the same pustule. As the two forms are distinct in appearance, it will better answer our present purpose to treat them separately, notwithstanding the belief that, in a scientific point of view, the evidence is all in favour of their union.
In fungi of this kind the mycelium, or delicate root-like threads, consists of thin filaments, which are spread through all parts of the plant occupied by the parasite, traversing the intercellular passages, but rarely perforating and entering the cells. This compacted and interwoven mycelium forms a kind of cushion beneath each pustule, on which the fruits of the parasite rest. By the increase of this cushion and the swelling of the fruit, the epidermis which covers them is distended, and ultimately ruptured, so that, when ripened, the spores escape. It must be remembered that the fruit is of from two to four kinds. Small bodies, called spermatia, 38which are derived from the spermogones, and which have not yet been known to germinate; Stylospores, produced either singly, or in bead-like, or moniliform, strings, and which either precede or are associated with the true spores; Spores, sometimes simple, but often complex; and Sporidia, or secondary sporules, which are produced on the germinating threads of the true spores.
The various genera of these endophytes owe their distinctions to the form, or mode of development of their true spores. In one instance these spores are united in pairs, or divided by a septum, so that they are two-celled: these are named Puccinia. In another instance the spores are one-celled, and at first borne upon a stalk or peduncle, from which they are detached in ripening: such are called Trichobasis. It is unnecessary here to indicate all the variations to illustrate the fact that the generic distinctions are based upon the characters of the true spores. How unsatisfactory such a mode will appear, when subjected to experience day by day, a botanist would suspect. In the same pustule, resting upon the same cushion of mycelium, the spores of an Aregma will be found with those of a Lecythea, and those of a Puccinia with Trichobasis. More than this has even been affirmed. The alternation of generations, known to students in the animal world, is here repeated in the vegetable. Dr. de Bary declares that certain data appear to indicate that Æcidium constitutes 39not a genus by itself, but are organs in the development of some other germs and species, possessing its spermogonia, its Æcidium; its Uredo, and its spores, properly speaking; whilst in others the Uredo-form the Puccinia-form, and the Æcidium-form may alternate. It is not our intention to enter deeply upon the discussion of this subject, of so little interest to the beginner, and so out of place in an introduction to the study. That forms and conditions are multifarious, and that an entire revision of the classification is inevitable, are facts which do not require many words to establish. Already it is to be feared that in this brief chapter we have said too much, and must recommend its perusal again, when the names and characters of the genera alluded to have been rendered more familiar.
It could scarcely have been permitted that the student should go far without being cautioned that there is such a thing as di-morphism in microscopic fungi; and the explanation of such a phenomenon must presuppose a certain amount of knowledge which, thus far, the reader could not have acquired. Hence an anomaly, to escape from which an ultimate return to the subject will be necessary.
In a recent account of Dr. de Bary’s experiments,[3] an interesting history is given of the development of a rust-like fungus, which is common 40on many plants of the pea and bean tribe. As it may serve to illustrate some of the preceding, as well as subsequent, remarks on development, an abstract shall close this chapter.
3. De Bary—“Annales des Sciences Naturelles,” ser. 4, vol. xx.
The spores of this species (Uromyces appendiculatus) are oboval cells, terminated by a rounded point, provided with a deep brown, smooth, epispore, or outer coating, and a distinct, but colourless endospore, or inner coating. These enclose a granular matter, which surrounds what has been termed the nucleus, but which appears to be a vacuole. At the top of the epispore is a pore which is characteristic of the genus. The spores are supported on a colourless, or slightly-tinted pedicel of considerable length (Plate VII. fig. 150). By means of this pedicel, the spores are attached to the fostering plant, on which they form pustules or sori of a blackish colour, and variable extent. These spores are ripened towards the end of summer or beginning of autumn. During winter they remain in a state of repose, but in the following spring the faculty of germination developes itself. At this period, when moistened or placed on a humid soil, they germinate at the end of a few days. The spore then emits a curved and obtuse tube, which soon ceasing to elongate itself, gives origin to three or four sporidia, of a reniform or kidney shape. When cultivated on moistened glass, these sporidia also emit a short, thin, slender tube, which produce in turn secondary 41sporidia. Here vegetation ends in the artificial culture above indicated.
When the sporidia are sown upon the epidermis of a favourable plant, the germ-tube being emitted, penetrates the wall of any approximate cellule, swells and increases into a cylindrical tube equal in thickness to the original sporidia, and therefore four or five times the diameter of the germ-tube before it entered the cellule. The contents of the sporidia and external portion of its germ-tube pass into the portion within the cellule, and then these external portions perish, and all evidence of the entry is obliterated, except a very minute point at which the tube remains attached to the inner surface of the wall of the cellule. The enclosed tube soon elongates, divides, and becomes branched. These branches perforate the inner walls of the epidermis, and pass into the intercellular spaces of the parenchyma to become mycelium. This takes place within 24 hours. A few days afterwards the mycelium is spread through the parenchyma. At length the surface of the same spots which had been sown in the first instance with the sporidia, become of a whitish tint, rapidly increasing and intensifying. Three days after, little protuberances appear on the surface of the white spots. These are of an orange colour, and many of them are surmounted by a little drop of mucilaginous fluid. These are spermogones. Their number daily increases, and a little time after appear numerous large 42globular protuberances intermingled with them. These soon rupture the epidermis, and take the orange colour and cylindrical form of cluster-cups (Æcidium). At length the summit of the peridia opens to allow the escape of the stylospores. It is easy to assure oneself that the spermogones and cluster-cups proceed from the mycelium of the sporidia which had been sown. During several days the length and number of the peridia of the Æcidium continue to increase. One month after sowing, brownish or blackish points make their appearance upon the whitish spots, around, or intermingled with the cluster-cups. These increase rapidly in number and magnitude. Examined by the microscope, they present the ordinary fructification of Uromyces, mingled with stylospores. Thus the mycelium of the cluster-cups engenders at the end of its vegetation fruits equal in all points to those from whence they are in the first instance derived.
The stylospores of the cluster-cups possess the irregular, globular form and structure of their congeners. They are filled with orange granular matter, and provided with a colourless, finely-punctated epispore. When these stylospores are sown on the moistened epidermis of a favourable plant, the germ-tube at first creeps along the surface, but as soon as its extremities find a stomate, it enters it and elongates itself in the air-cavity below the orifice, receives the contents of the original 43stylospore and exposed portion of its tube, then separates itself from those parts, which become dispersed. The active part increases and ramifies, and produces a mycelium which spreads through the intercellular passages of the parenchyma. At the end of from six to eight days, the whitish spots appear on the surface of the fostering plant, and indicate that the fructification of the parasite is about to commence. The epidermis is elevated and broken, and little brown pustules appear through the openings. These are the stylospores of Uredo, which are produced in immense quantities, and soon cover the pustules with a deep brown dust. Later, the formation of the stylospores is arrested, and the true germinating spores appear in the same pustules.
The stylospores of Uredo are borne singly at the top of short filaments. On arriving at maturity they detach themselves. They are of a globular form, with a reddish-brown epispore, provided with little pointed prominences, and three pores at equal distances. After maturity they germinate in precisely the same manner as the stylospores of the cluster-cups. They enter only through the stomata of the epidermis. The pulvinules are identical with those which the stylospores of Æcidium originate, and they also produce true spores at the end of their vegetation. No other fruit arises from them. These organs, therefore, always reproduce the same form to which they owe their origin. 44The result of these investigations shows that the bean rust (Uromyces appendiculatus), besides spermogones, possesses four sorts of reproductive organs, which all serve to propagate the species, but that one alone of them produces it in a form always identical, whilst the others present well-marked alternations of generation. Hence it is concluded that there are,
I. Spores which produce in germinating the promycelium, and
II. Sporidia.—These give place to a mycelium, which bears afterwards—
III. Æcidium.—Particular organs which engender stylospores, and which produce—
IV. Uredo, the second form of the stylospores, and later spores (No. I.), which are always associated with Uredo in the same pustule. The spores and stylospores of Uredo come also upon the old mycelium, which has previously produced Æcidium. The Uredo stylospores always produce Uredo, and true spores.
DR. WITHERING’S “Arrangement of British Plants” in 1818 reached its sixth edition. This is less than half a century ago, and yet the whole number of species of Fungi described in that edition was only 564, of which three hundred were included under the old genus Agaricus. Less than eighty of the more minute species of Fungi, but few of which deserve the name of microscopic, were supposed to contain all then known of these wonderful organisms. Since that period, microscopes have become very different instruments, and one result has been the increase of Withering’s 564 species of British Fungi to the 2,479 enumerated in the “Index Fungorum Britannicorum.” By far the greater number of species thus added depend for their specific, and often generic characters, upon microscopical examination. The proportion which the cryptogamic section bears to the phanerogamic in our local Floras before 1818, now almost involuntarily causes a smile. Even such authors as were supposed to pay the greatest possible respect to the lower orders of plants could never present an equal number of pages devoted to them, as to the higher orders. Relhan, for instance, only occupies one-fifth 46of his “Flora Cantabrigiensis,” and Hudson one-fourth of his “Flora Anglica,” with the Cryptogamia. At the present time, it will be seen that, with a liberal allowance for “hair-splitting,” the number of British species of flowering plants scarcely exceeds three-fourths of the number of Fungi alone, not to mention ferns, mosses, algæ and lichens, and yet we have no “Flora” which contains them, and but a minority of our botanists know anything about them. If we need excuse for directing attention to some of the most interesting of these plants, let the above remarks suffice in lieu of formal apology.
“Mildew” is just one of those loose terms which represent no definite idea, or a very different one to different individuals. Talk of mildew to a farmer, and instantly he scampers mentally over his fields of standing corn in search of the brown lines or irregular spots which indicate the unwelcome presence of Puccinia graminis, known to him, and to generations of farmers before him, as “mildew.” Try to convince a Norfolk farmer that anything else is “mildew,” and he will consider you insane for your pains. Speak of mildew in your own domestic circle, and inquire of wives, or daughters, or servants, what it means, and without hesitation another, and even more minute species of fungus, which attacks damp linen, will be indicated as the true mildew, to the exclusion of all others; and with equal claims to antiquity. Go to 47Farnham, or any other hop-growing district, and repeat there your question,—What is mildew?—and there is every probability that you will be told that it is a kind of mould which attacks the hop plant, but which differs as much from both the mildew of the farmer and the laundry-maid as they differ from each other. The vine-grower has his mildew, the gardener his mildewed onions, the stationer his mildewed paper from damp cellars, the plasterer his mildewed walls, and in almost every calling, or sphere in life, wherever a minute fungus commits its ravages upon stock, crop, or chattels, to that individual owner it becomes a bug-bear under the name of “mildew.” Reluctantly this vague term has been employed as a portion of the title to this chapter, but it must be limited in its application to the “mildew of corn,” known to botanists as Puccinia graminis, and not to include the numerous other microscopic Fungi to which the name of mildew is often applied.
The origin of this term and its true application may undoubtedly be traced to mehl-thau, “meal dew.” A singular proof of the ignorance which prevails in regard to all the fungal diseases of corn, may be found in the fact that at least one of our best etymological dictionaries states that the mildew in corn is the same as the ergot of the French. Had the writer ever been a farmer, he would have known the difference; had he ever seen the two, he could scarcely have made such a mistake. It 48is barely possible for him ever to have heard the ergot of grain called by the name of mildew.
How long this disease has been known, is an unsolved problem. About the middle of the last century a tract was published on this subject in Italy, but this was probably not even the first intimation of its fungoid character. Before such conclusion had been arrived at, men may have struggled in the dark, through many generations, to account for a phenomenon with which they were doubtless familiar in its effects. In 1805, Sir Joseph Banks published his “Short Account,” illustrated by engravings from the inimitable drawings of Bauer, whereby many in this country learnt, for the first time, the true nature of mildew.

Plate III.
W. West imp.
With a view to the clearer understanding of these parasites in the phases of their development, let us select one, and we cannot do better than adhere to that of the wheat and other graminaceous plants. A fine day in May or June dawns upon our preparations for a stroll, far enough into the country to find a wheat-field. Even now, with the area of the metropolis constantly widening, and banishing farmers and wheat fields farther and farther from the sound of Bow-bells, a corn field may be reached by a good stiff walk from Charing-Cross, or a six-penny ride at the most, in nearly any direction. Having reached the field, it may be premised that a walk into it of less than twenty yards will be sure to reward you with the fungus we are in quest of. 49Look down at the green leaves, especially the lower ones, and you will soon find one apparently grown rusty. The surface seems to be sprinkled with powdered red ochre, and grown sickly under the operation. Pluck it carefully, and examine it with a pocket lens. Already the structure of a healthy leaf is familiar to you, but in the present instance the cuticle is traversed with numerous longitudinal cracks or fissures, within which, and about their margins, you discern an orange powder, to which the rusty appearance of the leaf is due. Further examination reveals also portions in which the cuticle is distended into yellowish elongated pustules, not yet ruptured, and which is an earlier stage of the same disease. This is the “rust” of the agriculturist, the Trichobasis rubigo-vera of botanists, the first phase of the corn mildew.
To know more of this parasite, we must have recourse to the microscope; having therefore collected a few leaves for this purpose, we return homewards to follow up the investigation. We will not stay to detail the processes of manipulation, since these will not offer any deviation from the ordinary modes of preparation and examination of delicate vegetable tissues.
The vegetative system of the “rust,” and similar fungi, consists of a number of delicate, simple, or branched threads, often intertwining and anastomosing, or uniting one to the other by means of lateral branchlets. These threads, termed the 50mycelium, penetrate the intercellular spaces, and insinuate themselves in a complete network, amongst the cells of which the leaf, or other diseased portion of the plant, is composed. High powers of the microscope, and equally high powers of patience and perseverance, are necessary to make out this part of the structure. We may regard the whole mycelium of one pustule, or spore-spot, as the vegetative system of one fungal plant. At first this mycelium might have originated in a number of individuals, which afterwards became confluent and combined into one for the production of fruit, that is to say, an indefinite number of points in the vicinity of the future mycelium developed threads; and these, in the process of growth, interlaced each other, and ultimately, by means of transverse processes, became united into one vegetative system, in which the individuality of each of the elementary threads became absorbed, and by one combined effort a spore-spot, or cluster of fruit, was produced. In the first instance a number of minute, transparent, colourless cellules are developed from the mycelium: these enlarge, become filled with an orange-coloured endochrome, and appear beneath the cuticle of the leaf as yellowish spots. As a consequence of this increase in bulk, the cuticle becomes distended in the form of a pustule over the yellow cellules, and at length, unable longer to withstand the pressure from beneath, ruptures in irregular, more or less elongated fissures (Plate VII. 51fig. 141), and the yellow bodies, now termed spores (whether correctly so, we do not at present inquire), break from their short pedicels and escape, to the naked eye presenting the appearance of an orange or rust-coloured powder. In this stage the spores are globose, or nearly so, and consist of but one cell Plate VII. figs. 142, 144). It will afford much instructive amusement to examine one of these ruptured pustules as an opaque object under a low power, and afterwards the spores may be viewed with a higher power as a transparent object. The difference in depth of tint, the nearly colourless and smaller immature spores, and the tendency in some of the fully matured ones to elongate, are all facts worthy of notice, as will be seen hereafter.
A month or two later in the season, and we will make another trip to the cornfield. Rusty leaves, and leaf-sheaths, have become even more common than before. A little careful examination, and, here and there, we shall find a leaf or two with decidedly brown pustules intermixed with the rusty ones, or, as we have observed several times during the past autumn, the pustules towards the base of the leaf orange, and those towards the apex reddish-brown. If we remove from the browner spots a little of the powder, by means of a sharp-pointed knife, and place it in a drop of water or alcohol on a glass slide, and after covering with a square of thin glass, submit it to examination under a quarter-inch objective, a different series of forms will be 52observed. There will still be a proportion of subglobose, one-celled, yellow spores; but the majority will be elongated, most with pedicels or stalks, if they have been carefully removed from the leaf, and either decidedly two-celled, or with an evident tendency to become so. The two cells are separated by a partition or dissepiment, which divides the original cell transversely into an upper and lower cell, with an external constriction in the plane of the dissepiment (Plate IV. fig. 59). These bilocular or two-celled spores are those of the “corn mildew” (Puccinia graminis), which may be produced in the same pustules, and from the same mycelium, as the “corn rust,” but which some mycologists consider to be a distinct fungus, others only a modification or stage of the same fungus. After an examination of the different forms in the allied genera to which these chapters are devoted, we shall be able with less of explanation and circumlocution to canvass these two conflicting opinions.
Let us proceed, for the third and last time, to our cornfield, when the corn is nearly or fully ripe, or let us look over any bundle of straw, and we shall find blackish spots, from the size of a pin’s head to an inch in length, mostly on the sheaths of the leaves, often on the culm itself. This is the fully developed mildew, and when once seen is not likely afterwards to be confounded with any other parasite on straw (fig. 57). The drawings of Bauer have already been alluded to. Bauer was botanical 53draughtsman to George III., and his exquisite drawings, both of the germination of wheat and the fungi which infest it, are marvels of artistic skill. A reduced figure from part of one of his drawings is given (Plate IV. fig. 58), exhibiting a tuft of the bilocular spores of Puccinia graminis bursting through a piece of wheat straw. These closely-packed tufts or masses of spores, when examined with a common lens, seem, at first, to resemble the minute sorus of some species of fern; but when seen with higher powers, the apparent resemblance gives place to something very different. The tufts consist of multitudes of stalked bodies, termed spores, which are constricted in the middle and narrowed towards either extremity. The partition, or septum, thrown across the spore at the constriction, separates it into two portions, each of which consists of a cell-wall enclosing an inner vesicle filled with the endochrome (fig. 59) or granular contents, in which a nucleus may often be made out. This species of Puccinia is very common on all the cereals cultivated in this country, and on many of the grasses. A variety found on the reed was at one time considered a distinct species; but the difference does not seem sufficient to warrant a separation. However near some other of the recognized species may seem to approximate in the form of the spores, a very embryo botanist will not fail to observe the distinctive features in the spores of the corn mildew, and speedily recognize 54them amongst a host of others; subject, as they may be, to slight deviations in form, resulting either from external pressure, checks in development, or other accidental circumstances, or the variations of age.
There is no doubt in the minds of agriculturists, botanists, savans, or farm-labourers, that the mildew is very injurious to the corn crop. Different opinions may exist as to how the plants become inoculated, or how infection may be prevented or cured. Some have professed to believe that the spores, such as we have seen produced in clusters on wheat straw, enter by the stomata, or pores, of the growing plant, “and at the bottom of the hollows to which they lead they germinate and push their minute roots into the cellular texture.” Such an explanation, however plausible at first sight, fails on examination, from the fact that the spores are too large to find ingress by such minute openings. It is improbable that the spores enter the growing plant at all. The granular contents of the spores may effect an entrance either through the roots or by the stomata, or the globose bodies produced upon the germination of the spores may be the primary cause of infection. We are not aware that this question has been satisfactorily determined. It is worthy of remembrance by all persons interested in the growth of corn, that the mildew is most common upon plants growing on the site of an old dunghill, or on very rich soil. 55As the same Puccinia is also to be found on numerous grasses, no prudent farmer will permit these to luxuriate around the borders of his fields, lest they should serve to introduce or increase the pest he so much dreads.
The germination of the spores of the corn mildew is a very interesting and instructive process, which may be observed with a very little trouble. If the spores be scraped from the sori of the preceding year (we are not sure that those of the current year will succeed), and kept for a short time in a damp atmosphere under a glass receiver, minute colourless threads will be seen to issue both from the upper and lower divisions of the spores. These will attain a length several times that of the spores from whence they spring. The extremities of these threads ultimately thicken, and two or three septæ are formed across each, dividing it into cells, in which a little orange-coloured endochrome accumulates. From the walls of each of these cells, or joints, a small pedicel, or spicule, is produced outwards, the tip of which gradually swells until a spherical head is formed, into which the orange-coloured fluid passes from the extremities of the threads.[4] A quantity of such threads, bearing at their summits from one to four of these orange-coloured, spherical, secondary fruits, supply 56a beautiful as well as interesting object for the microscope. When matured, these globose bodies, which Tulasne has called sporidia, fall from the threads, and commence germinating on their own account. It is not impossible that the sporidia, in this and allied genera, may themselves produce a third and still more minute fruit, capable of diffusion through the tissues of growing plants, or gaining admission by their stomata. Nothing of the kind, however, has yet been of certainty discovered.
4. Similar in all essential particulars to the germination of Aregma (Plate III. fig. 45).
Forty other species of Puccinia have been recorded as occurring in Great Britain, to all of which many of the foregoing remarks will also apply—viz., such as relate to their two-celled spores being found associated with, and springing from, the same mycelium as certain orange-coloured one-celled spores; and also the main features of the germinating process.

Plate IV.
W. West imp.
A very singular and interesting species is not uncommon on the more delicate grasses, being found chiefly confined to the leaves, and produced in smaller and more rounded, or but slightly elongated, patches (Plate IV. fig. 60). We have met with it plentifully amongst the turf laid down in the grounds of the Crystal Palace at Sydenham, and also on hedge-banks and in pastures. The spores are rather smaller than those of Puccinia graminis, but, like them, much elongated, slightly constricted, and borne on persistent peduncles. The most 57prominent distinction may be found in the apices of the spores, which, in this instance, are not attenuated, but crowned with a series of little spicules, or teeth, whence the specific name of coronata has been derived (Plate IV. fig. 62).
The Labiate family of plants and its ally the Scrophulariaceæ are also subject to the attacks of several kinds of Brand, a name, by the bye, often applied locally to the corn mildew and other similar parasites, and which may have originated in the scorched or burnt appearance which the infected parts generally assume. In the former natural order the different kinds of mint, the ground-ivy, the wood-sage, and the betony, and in the latter, the water figwort and several species of veronica, or speedwell, are peculiarly susceptible; and on most a distinct species of Puccinia is found. To provide against doubt which the less botanical of our readers may possess of the meaning or value of the term Puccinia, which has already occurred two or three times in this chapter, a brief explanation may be necessary, which more scientific readers will excuse.
In botany, as in kindred sciences, acknowledged species have their trivial, or specific name, generally derived from the Latin. In the last species referred to, this was coronata, meaning crowned, in reference to the coronated apex of the fruit. Any indefinite number of species with some features in common are associated together in a group, which is 58termed a genus, and the term prefixed to the specific name of each species constituting that genus is its generic name, also commonly derived from the Latin or Greek. In this instance it is Puccinia, derived from the Greek puka, meaning closely packed, singularly applicable to the manner in which the spores are packed together in the pustules. The common features, or generic distinctions, of this genus, are uniseptate spores borne on a distinct peduncle.
In returning to the species found on Labiate plants, let us suppose ourselves to have strolled towards Hampstead Heath, and south of the road leading from Hampstead to Highgate, near certain conspicuous and well-known arches, built for a purpose not yet attained, are two or three muddy ponds nearly choked up with vegetation. Some fine autumnal afternoon, we must imagine ourselves to have reached the margin of the most northern of these ponds, and amidst a thick growth of reeds, sedges, and other water-loving plants, to have found the water-mint in profusion and luxuriance, with every leaf more or less occupied, on its under surface, with the yellow spores of a species of rust (Trichobasis) mixed with the browner septate spores of the mint brand (Puccinia Menthæ). This is common also on the horse-mint and corn-mint; we have found it on the wild basil and wild thyme, and once only on marjoram. Having collected as many leaves as we desire, and returned to home 59and the microscope, we proceed to examine them in the same manner as we have already examined the mildew, and as a result of such proceeding arrive at the following conclusions:—The pustules are small and round, never elongated as in the corn mildew, and generally confined to the under surface of the leaves (Plate IV. fig. 69). The spores are subglobose, slightly constricted, and the two cells nearly two hemispheres, with their flat surfaces turned towards each other (fig. 70). The form delineated in figure 75 is that of the sorus of many of the epiphytal brands, the centre being occupied by the closely-packed spores, surrounded to a greater or less extent by the remains of the ruptured epidermis.
Although the species of Puccinia (P. glechomatis) found on the leaves of the ground-ivy is said to be very common, we sought it in vain amongst every cluster of that plant met with during last summer and autumn, until, nearly despairing of finding it at all, we at last encountered a plot of ground-ivy covering the ground to the width of two or three yards, and in length eight or ten, nearly every plant being attacked by the brand. This was in the corner of a pasture, and the only time we found infected plants. The fungus, however, may be as common as the plant in other localities. The pustules on the leaves are larger than those of the mints, and also confined to the inferior surface (fig. 73). The spores are elliptic and but slightly 60constricted; the apex is often pointed, though not always so much as in our figure (fig. 74).
Of other species found on allied plants we have not considered it necessary to give figures, or write much. The betony brand (P. Betonicæ, DC.) does not seem to be common enough to be readily found by any one desiring to examine it for himself; and the same may be said of the figwort brand (P. Scrophulariæ, Lib.), the wood-sage brand (P. Scorodoniæ, Lk.), and the speedwell brand (P. Veronicarum, DC.); all of these are, however, characterized by a distinct feature, or features, which have been considered of sufficient importance to constitute a separate species.
We have had occasion to refer incidentally to the brand found on the under surface of the leaves of the wood-anemone (P. Anemones, P.). This is one of the earliest and commonest species. Go wherever the wood-anemone abounds, in any of the woods lying immediately to the north of the metropolis, or any of the woods in Kent, and from March to May it will not be difficult to find attenuated, sickly-looking leaves, with the under surface covered with the pustules of this brand, looking so like the sori of some fern (fig. 65) that it has been, and still is, sometimes considered as such. In Ray’s “Synopsis” (3rd edition, 1724), it is described in company with the maidenhair and wall-rue ferns; a figure is given of it in the same work (t. iii. fig. 1), and it is stated,—“this capillary was gathered by 61the Conjuror of Chalgrave.” When, afterwards, it was better understood, and the spots came to be regarded as true parasitic fungi, it still for a long time continued to bear the name, not even yet quite forgotten, of the Conjuror of Chalgrave’s fern.
An examination of the spores, both collectively in the pustules, and separately under a high power, will not fail to convince any one who has examined only the species we have already alluded to, that this parasite on the anemone (P. Anemones) is a true Puccinia, and a most interesting one. The two cells of the spores are nearly spherical, and the constriction is deeper and more positive than in any of the preceding. Moreover, the surface of the spore is minutely and beautifully echinulate, or covered with erect spines (Plate IV. fig. 66). Some few other of the species found in Britain have echinulate spores, but those are not common like the present. One word of caution to the amateur in search of the Puccinia on the anemone. It will be fruitless looking for it on the large foliaceous bracts of the flower-stalk, since these may be turned up carefully, till the back aches with stooping, ere a solitary pustule will be found; but the true leaves, proceeding from the rhizomes, are certain soon to afford you specimens.
Everybody knows the dandelion, but it is not every one who has noticed the fungi found upon its leaves. These are most commonly of two kinds, or probably the unilocular and bilocular forms of 62the same species: the latter we have found in the month of May, and the former in August and September. The lower leaves of young seedlings have generally rewarded us with the best specimens of the septate-fruited brand (Puccinia variabilis, Grev.). The pustules occur on both sides of the leaf, and are very small and scattered (fig. 82). The spores are singularly variable in form: sometimes both divisions are nearly equal in size; sometimes the upper, and sometimes the lower, division is the smallest; occasionally the septum will be absent altogether; and more rarely, the spores will contain three cells. From the very variable character of the spores (fig. 83), the specific name has been derived.
No species in the entire genus makes so prominent an appearance as the one found on the radical leaves of the spear thistle (Carduus lanceolatus). This latter plant is exceedingly abundant, and so is its parasite (Puccinia syngenesiarum, Lk.). From the month of July till the frosts set in we may be almost certain of finding specimens in any wood. The leaves have a paler roundish spot, from one-twelfth to one-fourth of an inch in diameter, on the upper surface, and a corresponding dark brown raised spot on the under surface, caused by an aggregation of pustules, forming a large compound pustule, often partly covered with the epidermis. The individual pustules are small, but this aggregate mode of growth gives the clusters great prominence, 63and therefore they are not easily overlooked (Plate IV. fig. 63). Although not confined to this species of thistle, we have not yet found this Puccinia on any other plant. The spores are elliptical, rather elongated, constricted, and without spines (fig. 64).
Other species of Puccinia are found on Composite plants, but with none of these is the present fungus likely to be confounded, if regard be had to its peculiar habit. The leaves, for instance, of the common knapweed (Centaurea nigra) are often sprinkled with the small pustules of the centaury brand (Puccinia compositarum, Sch.); these generally occupy the under surface of the lower radical leaves (fig. 67); occasionally a few of the pustules appear on the upper surface. We have not often found this fungus in the neighbourhood of London on the leaves of the knapweed, but, on the other hand, we have encountered it very commonly on those of the saw-wort (Serratula tinctoria). The spores are oval, scarcely constricted, and not attenuated in either direction (fig. 68). Other Composite plants than those above named are liable to attacks from this parasite.
In our school-days we remember to have spent many a stray half-hour digging for “earthnuts,” under which name we, as well as our elders and betters, knew the tubers of Bunium flexuosum. Not then, nor for many years after, did we notice, or regard if we did notice, the distorted radical 64leaves and leaf-stalks, and the blackish-brown spots, which reveal the cause in the presence of a brand, or parasitic fungus, of this genus (Puccinia Umbelliferarum, DC.), which is extremely common on this, as well as some other allied plants. If any spot is searched where this plant grows in any profusion, before the flowering stalks have made their appearance above the surrounding grass, this Puccinia will be readily found by the twisted, contorted, sickly appearance of the infested leaves (fig. 71), the petioles of which are often swollen and gouty in consequence. The sporidia are shortly stalked, and generally very much constricted (fig. 72). The species found on the stems of the hemlock, and also that on Alexanders (Smyrnium Olusatrum), are distinct; the spores of the latter being covered with tubercles or warts (figs. 55, 56). During a botanical ramble through Darenth Wood in April of the year just passed away, in some parts of which the sanicle abounds, we found the bright glossy leaves of this singular and interesting plant freely sprinkled with the pustules of a Puccinia (P. Saniculæ, Grev.), which is not at all uncommon on this, but has not hitherto been found on any other plant. Dr. Greville, of Edinburgh, was the first to describe this, as well as many other of our indigenous minute Fungi. For many years he has toiled earnestly and vigorously at the lower cryptogams, as evidenced by his “Scottish Cryptogamic Flora,” published in 1823; and yet his continual additions 65to the records of science show him to be earnest and vigorous still.
We have by no means exhausted the catalogue of Fungi belonging to this genus found in Britain, nor even those commonly to be met with; but the fear of prolixity, and the desire to introduce a description of other forms into the space still remaining to us, prompt us to dismiss these two-celled brands with but a brief allusion to such as we cannot describe. Box-leaves are the habitat of one species, and those of the periwinkle (Plate VI. fig. 132) of another. One vegetates freely on the leaves of violets through the months of July and August, and another less frequently on the enchanter’s nightshade. Several species of willow-herb (Epilobium) are attacked by one Puccinia (Plate IV. figs. 78, 79), and a single species by another. Plum-tree leaves, bean-leaves, primrose leaves, and the half-dead stems of asparagus, have their separate and distinct species, and others less commonly attack the woodruff, bedstraw (Plate VIII. figs. 172, 173), knotgrass, ragwort, and other plants less common, more local, or, to the generality of the non-botanical, but imperfectly known.
We have found, not uncommonly in the autumn, the scattered pustules of a brand on the stems and leaves of the goat’s-beard, occupying the places which were scarred with the remains of cluster-cups that had flourished on the same spots a month or two previously (Plate IV. fig. 76). The pustules are by no means minute, but elongated and bullate; the 66spores beautifully studded with warts (Plate IV. fig. 77). This species cannot certainly be identical with Puccinia compositarum (Schlecht), P. syngenesiarum (Lk.), or P. tragopogonis (Corda). In none of these do the spores appear to be warted, and the habits of both the latter are different. Its nearest associate appears to be P. centauriæ (Corda), at least in the fruit, and whilst the form and character of these organs are considered of any value in the determination of species, smooth spores cannot be associated, we think, with tuberculate or echinulate spores under the same name.
In the spores of the species to which attention has been more specially directed we have types of the principal forms. In the “corn-mildew” they are elongated, and tapering towards either end; in the “coronated brand” the apex is crowned with spicular processes; in the “wind-flower brand” the entire spores are echinulate; in the “mint brand” they are globose; in the “composite brand” elliptic; in the “earth-nut brand,” nearly cut in two at the septum; and in the “dandelion brand,” so variable in form that no two are precisely alike. On the other hand, all are characterized by a transverse septum dividing each spore into two cells.
FROM the twin-spored genus we pass to another, in which the spores are usually divided into three cells, and which, from this cause, has been named Triphragmium. Only one species has hitherto been found in this country, and that not very commonly, on the leaves of the meadow-sweet, Spiræa ulmaria (Plate III. fig. 47). Externally, it much resembles, in the size and character of the pustules, many of the above-named brands, but when seen under the microscope this similarity disappears. In general outline the spores are nearly globose, and externally papillose. In one species, found on the Continent, but not hitherto in Great Britain, the spores are covered with curious long-hooked spines, by means of which they adhere tenaciously to each other. In germination, the spores of Triphragmium do not offer any noteworthy deviation from those of Puccinia,[5] and the chief interest of our indigenous 68species lies in the three-celled form of its spores (fig. 48), to which occasionally those of Puccinia variabilis approximate, and may be regarded as the link which unites the two genera.
5. Mr. Currey has only seen the tips of the germinating threads swell, and become septate, each of the joints thus formed falling off and germinating without producing spherical sporidia; whilst Tulasne figures globular sporidia, as will be seen in our fig. 49, reduced from the figure by Tulasne.—(Vide Currey, in “Quarterly Journal of Microscopical Science,” 1857, pp. 117, &c.)
The old story of “Eyes and no Eyes” is too often literally true, not only with the children it was written to amuse and instruct, but also with children of a larger growth who scorn such baby tales, and disdain such baby morals. Out of more than a thousand indigenous species of microscopic fungi, of which there is generally some evidence afforded of their presence visible to the naked eye, how few are there of the millions that inhabit our island who can count twenty species that they have ever seen; still fewer that have noticed one hundred. Amongst the twenty species known to the few will probably be included one which appears in autumn in prominent black spots, the size of a large pin’s head, or half a turnip seed, with the flat face downwards, sprinkling the under surface of blackberry leaves (fig. 39); with larger, reddish, purplish, or reddish-brown spots on the upper to indicate the presence of the fungus beneath. Just at the time when blackberries are ripe, these spots are in perfection on the leaves, and their eyes must have been sadly at fault who could ever have gathered their own blackberries without seeing the discoloured leaves. The coloured spots on the face of the leaf are due to the diseased state of the tissues caused by the parasite on the opposite surface. As much of the 69leaf as contains two or three of the black pustules should be removed carefully with a knife or sharp scissors, and submitted to microscopical examination; each will be seen to consist of a dense tuft of blackish, elongated, stalked bodies, clustered as in fig. 44, but much more numerously and closely packed together. These are the spores of the blackberry brand (Aregma bulbosum, Fr.). A few of these spores should be removed on the point of a sharp penknife, placed on a glass slide with a drop of distilled water or alcohol, covered with thin glass, and then viewed with a quarter-inch objective. Each spore has a stalk longer than itself, thickened below, and containing a yellow granular core. The spore itself is much longer than in any of the Pucciniæ, of a dark brown colour, and apparently divided by several transverse partitions into three, four, or more cells, the whole surface being covered with minute warts or prominences (Plate III. fig. 41).
In 1857, Mr. F. Currey investigated the structure of these spores, and the results of these experiments were detailed in the “Quarterly Journal of Microscopical Science.” One conclusion arrived at was, that “the idea of the fruit consisting of sporidia united together and forming a chain, is certainly not in accordance with the true structure. The sporidia are not united to one another in any way, but, although closely packed for want of space, they are in fact free in the interior of what may be called a sporangium or ascus.” To arrive at this 70conclusion, careful examination was necessary, and new modes of manipulation essential. The details of one method employed were to the following effect:—A sufficient number of spores were removed on the point of a lancet, and placed on a slide in a drop of alcohol. Before the spirit was quite evaporated, two or three drops of strong nitric acid were added, and the whole covered with thin glass. The slide was then warmed over a spirit-lamp, the acid not being allowed to boil, but only gradually heated to boiling point. By this means the fruit was found to consist of an outer membrane, nearly transparent, and studded with tubercles; that this membrane enclosed a number of cells which constituted the apparent joints, and which were naturally flattened at either end by mutual pressure. When the outer membrane was dissolved or ruptured, these cells escaped, and became detached from each other. The cells, thus set free, exhibited a brownish or yellow ring around a paler area, in the interior of which an inner cell was visible, sometimes globular, often irregular in shape. The examination of the ring was not entirely satisfactory; it appeared to be sometimes marked with concentric lines having the appearance of wrinkles. The inner cell had granular contents and a central nucleus. When perfectly free they were spherical in form, with a distinct membrane of their own; and colourless, except when acted upon by reagents. The means employed to determine the 71existence of these cells was to soak the spores in muriatic acid; then, upon pressure of the glass cover, the outer membrane and ringed cells were ruptured, and the inner cell escaped (Plate III. fig. 46).
Germination may be induced in these spores by keeping them in a moist atmosphere (fig. 45); but the mode does not differ from that described above as occurring in the “corn mildew.” Mr. Currey writes:—“I know no microscopical object of greater beauty than a number of fruits of Phragmidium in active germination.” By Phragmidium he means the Aregma of this work, of which Phragmidium is a synonyme.
Well may the reader remark on arriving thus far, “Does all this examination and detail refer to the fruit borne in the little blackish spots on bramble leaves, which I have hitherto overlooked?” Ay, and to several similar spots on other plants. Examine carefully the raspberry leaves in your garden, and you will probably find similar, but smaller, pustules also on the under surface (Plate III. fig. 42). We say probably, because none of our British species seem to be equally uncommon with this. During the past year we examined hundreds of plants, and did not find a single pustule. This species was named by Dr. Greville Aregma gracilis, which name it still continues to bear (fig. 43). Such a fate will not await you if you should proceed in the autumn to some chalky district where the burnet is common. Go, for instance, to Greenhithe or Northfleet, on the 72North Kent Railway, in August or September, where the burnet is plentiful, and the leaves will present the appearance of having been peppered beneath, from the number of minute pustules of the burnet brand scattered over the under surface (fig. 30, upper leaflets). Or if you prefer collecting nearer home, visit some neighbouring garden, if your own does not contain many roses, and the leaves will be found equally prolific in an allied species (fig. 36). Should gardens and roses be alike unattainable, any bank or wood will furnish the barren strawberry (Potentilla fragariastrum), and during the latter part of the summer, or in autumn, another species of Aregma will not be uncommon on the under surface of the leaves (fig. 33). All these species will be found accompanied by the orange spores of species of Lecythea, which some mycologists consider to be distinct fungi, and others to be merely forms or conditions of Aregma. These spores are represented in Plate III. figs. 31, 34, 37, and 40. From the magnified figures of the spores of the different species of Aregma (figs. 32, 35, 38, 41, and 43), it will be apparent that they have all certain features in common, i.e., cylindrical spores containing from three to seven cells. This may be called the generic character, common to all the species of the genus Aregma. Again, each species will be observed to possess its own distinct features, which may be termed its specific character. In one, the apex of 73the spores will be obtuse, in another acutely pointed, in another bluntly pointed, &c. In one species the number of cells will usually be four, in another five or six, in another seven or eight. The stem in one species will be slender and equal, in another thickened or bulbous. So that in all there will be some permanent peculiarity for each not shared by the others.
One other form of brand (Xenodochus carbonarius), presenting, it is believed by some, generic differences from all that we have as yet noticed, remains to be briefly alluded to. This form appears to be very uncommon in this country, but, when found, is parasitic on the leaves of the great burnet (Sanguisorba officinalis), a plant of local distribution. The parasite appears to the naked eye in small tufts or pustules resembling those of an Aregma, but, when microscopically examined, the cells of the spores are found to be numerous, indeed, considerably more than in the most complex Aregma (fig. 29). This, however, seems to be the only distinction, for the cells are free in the interior of the investing membrane, and in all points of structure, in so far as it has been examined, identical with Aregma. Whether it is logical to consider a four-celled spore an Aregma, and a seven-celled spore an Aregma, and exclude a ten or twelve-celled spore from the same genus on account of the number of its cells, does not appear to us clearly answerable in the affirmative.
During the course of this and the preceding 74chapter we have passed rapidly through four genera of parasitic fungi so nearly allied, that one is almost led to doubt the validity of the generic distinctions. These may be presented briefly thus:—
| Spores two-celled | Puccinia. |
| Spores three-celled | Triphragmium. |
| Spores four to seven-celled | Aregma. |
| Spores many-celled | Xenodochus. |
It has been seen that the habit, mode of growth, germination, and structure, except in the number of cells, scarcely differ; but it is not our province here to enter upon the discussion of such a subject.
The association of one-celled, orange-coloured spores with the brown two or more celled spores passed in review is another feature worthy of a passing notice, and which opens a field for discussion. It is generally admitted that these two forms are the production of the self-same mycelium or vegetative system, but it is not so generally admitted that they are but two forms or phases of the fruit of the same plant. It is not at all uncommon in the history of mycology to find two forms which were for a long time considered to be distinct plants producing different forms of fruit, and which bore different names, and were located in different genera, at length proved to be only the self-same plant in different conditions, and ending in one name being expunged from the list. Such a fate probably awaits, at no distant date, the orange spores which precede or accompany the species in 75the present genera. Already Tulasne and some others accord them no place in their system.
It may be added, for the benefit of any who wish to pursue the study of this interesting branch of Cryptogamic Botany, that the leaves of the plants containing the parasitic fungi now noticed may be collected and preserved by drying between folds of blotting-paper, or the leaves of a book, and will retain their character, with the exception of colour in the orange forms, so as to be eligible for examination at any period of the year for twenty years to come. Each species, when dry, may be transferred to an old envelope, and labelled outside with the name, date of collection, and locality; and one hundred such envelopes will constitute a miniature herbarium in a very small compass.
ONE of the fungal diseases of corn long and widely known has obtained amongst agriculturists different appellations in different localities. In some it is the “smut,” in others it is respectively “dust-brand,” “bunt-ear,” “black-ball,” and “chimney-sweeper,” all referring, more or less, to the blackish soot-like dust with which the infected and abortive ears are covered. This fungus does not generally excite so much concern amongst farmers as the other affections to which their corn-crops are liable. Perhaps it is not really so extensively injurious, although it entirely destroys every ear of corn upon which it establishes itself. Wheat, barley, oats, rye, and many grasses are subject to its attacks, and farmers have been heard to declare that they like to see a little of it, because its presence proves the general excellence of the whole crop. No one who has passed through a field of standing corn, after its greenness has passed away, but before it is fully ripe, can have failed to notice, here and there, a spare, lean-looking ear, completely blackened with a coating of minute dust (Plate V. fig. 98). If he has been guilty of brushing in amongst the corn, it will still be remembered how 77his hands and clothing became dusted with this powder; and if at the time he should have been clad in sombre black, evidence will have been afforded—in the rusty-looking tint of the powder when sprinkled upon his black continuations—that, however sooty this powder might appear whilst still adhering to the ears of corn, it has an evident brown tint when in contact with one’s clothes. This powder, minute as it is, every granule of it constitutes a spore or protospore capable of germination, and ultimately, after several intermediate stages, of reproducing a fungus like the parent of which it formed a part. During the growth of the plant its virulent contents flow like a poison through the innermost tissues, and at length attack the peduncle or axis of the spikelets of the ear, raising up the essential organs and reducing them to a rudimentary state. Brongniart, who made this species the special subject of observation, states that the fleshy mass which is occupied by the fungus consists entirely of uniform tissue, presenting large, almost quadrilateral cavities, separated by walls, composed of one or two layers of very small cells filled with a compact homogeneous mass of very minute granules, perfectly spherical and equal, slightly adhering to each other, and at first green, afterwards free or simply conglomerate towards the centre of each mass, and of a pale rufous hue; at length the cellular walls disappear, the globules become completely insulated, and the whole mass is changed 78into a heap of powder, consisting of very regular globules, perfectly alike, black, and just like the reproductive bodies of other fungi (Plate V. fig. 99). A scientific botanist of some repute, M. Unger, published a work in Vienna during the year 1823, in which he sought to prove that this, and allied species of fungi, were not fungi at all, but merely broken up cells, or disruptured and altered conditions of certain portions of the diseased plants. The most satisfactory refutation of this theory may be found in the fact that the spores of the smut can be seen to germinate under favourable conditions, and produce fruit, whereas, if they were only the ordinary cells of the plant broken up by disease, fructification would not take place.
The spores in this species are exceedingly minute. It has been ascertained that forty-nine of them would be contained within a space the one-hundred-and-sixty-thousandth part of a square inch; hence one square inch of surface would contain little less than eight millions. These myriads of spores are shed from the ears, and nothing remains but the barren matrix in which they were borne when the farmer proceeds to gather in his crops. At that time he sees no more of the “smut,” all remembrance of it for the time is gone, his only thought is to stack his corn in good condition. But the millions of spores are dispersed, ten millions at least for every ear that has been “smutted,”—and will they not many of them reappear next year, 79and thus year after year, with as much certainty as the grain upon which they are parasitic?
Like many of the parasitic fungi, so destructive in the farm and the garden, this species belongs to the family in which the spores are the distinctive feature. After many botanical changes, the “smut” is at length regarded as a fixed resident in the genus Ustilago; with the specific name of segetum, which latter signifies “standing corn;” it is therefore the Ustilago, or smut of the standing corn. The characters of the genus are, chiefly, that the spores are simple and deeply seated, springing from delicate threads, or in closely-packed cells, ultimately breaking up into a powdery mass. Fifteen members of this genus have been described as British. One of these (U. maydis) attacks the maize or Indian corn grown in this country in a similar manner as the common smut attacks wheat or barley; but as maize is not an established crop with us, a more minute description of this species is unnecessary; the spores are figured in Plate V. fig. 108. Another species (U. hypodytes) makes its appearance at first beneath the sheaths of the leaves surrounding the stems of grasses (fig. 100), and ultimately appears above and around them as a purplish-black dust (fig. 101). The seeds of sedges, the leaves and stems of certain definite species of grass, the flowers of scabious (Plate VI. figs. 123-125), the receptacles of the goatsbeard, the anthers of the bladder campion, and other allied plants, and the 80seeds of the Bistort family, are all liable, more or less, to the attacks of one or other of the residue of the fifteen species of Ustilago already referred to as indigenous to Britain.

Plate V.
W. West imp.
Although we do not profess to teach practical men how to grow good corn, or how they shall get rid of, or keep clear from, the many foes to which their crops are exposed, yet a suggestion may be offered, based upon the facts obtained in our botanical researches, supported by the analogy of allied circumstances. In this instance the extreme minuteness and profusion of the spores would evidently render all the corn liable to the attachment of, perhaps only two or three, spores to the seed coat. Some ears of corn in nearer proximity to the smutted ears may be covered with spores which yet remain invisible to the naked eye, and when these grains are mixed with others in the heap, the chances are not much in favour of any handful not becoming charged with spores. If the majority of these were not redeemed from destruction by the many changes, shiftings, rubbings, and scrubbings to which the seed corn is liable between the time of its reaping and the period of its sowing, we might expect a very large crop of “smutted” corn. Under ordinary circumstances we can scarcely imagine that the loss arising from infected ears would repay much special labour to prevent it, only that to a large extent the precautions taken to cleanse the seed corn from the spores of one fungus 81will also avail for another, and while cleaning it of the spores of “smut,” those of “bunt” will also be removed. The facts that we rely upon chiefly as indicating the remedy are that the spores are only superficially in contact with the seed corn, and that they are of less specific gravity, causing them to float on the surface of any fluid in which the corn may be immersed. Again, the spores of many species of fungi will not germinate after saturation with certain chemical solutions. One of the most successful and easy of application is a strong solution of Glauber’s salts, in which the seed corn is to be washed, and afterwards, whilst still moist, dusted over with quick-lime. The rationale of this process consists in the setting free of caustic soda by the sulphuric acid of the Glauber’s salt combining with the lime, and converting it into sulphate of lime. The caustic soda is fatal to the germination of the spores of “bunt,” and probably also of “smut;” although, as already intimated, except in cases where these affections of the corn are very prevalent, we shall be informed by the agriculturist that the cost of labour in the prevention will not be compensated in the cure.
Experience has also taught us that many fungi flourish in proportion to the wetness of the season, or dampness of the locality. A wet year is always exceedingly prolific in fungi, and a dry season correspondingly barren, at least in many kinds, whilst others, as the experience of 1864 has convinced us, are 82exceedingly common. In a field or wood the mycologist reaps his richest harvest of mycological specimens in the lowest and dampest spots, in swamps, ditches, and ill-drained nooks. This is a fact worth knowing as much by the farmer as the amateur botanist in search of specimens for his herbarium.
One of the most unmistakable species of “smut” is that which infests the goatsbeard, on which we have already described an Æcidium. Generally about the same time as the cluster-cups make their appearance on the leaves, some of the unopened flower-heads of this plant will be found considerably altered in appearance by the shortening of the segments of the involucre, and at length by the whole inflorescence being invested with a copious purplish-black dust. If, by any means, the lobes of the involucre are any of them separated, the enclosed dust escapes, blackening the fingers and clothing of the collector, as if it were soot (Plate V. fig. 92). A little of this dust submitted to the microscope will be found to consist of myriads of small globose spores, nearly uniform in size and shape; and if a higher power be employed, each of these will appear to have a papillose or minutely granulated surface. The florets, dwarfed in size and contorted, or the remains of them, are embedded in the mass of spores (fig. 93), and if one or two of these are removed and placed under a good one-inch objective, every part will be found covered with adhering spores, to the apparent 83exhaustion of its substance. Of course, the florets are never developed when subjected to the attack of “smut.” The whole plant assumes a faded, sickly appearance, even before the spores are fully ripened. We would recommend our readers, if they collect one of the infected flower-heads, to put it into a box or paper by itself, for if placed in the box with other specimens it will so sprinkle them with its black powder as to render them nearly useless for microscopic examination: everywhere the microscope will detect, where the unaided eye failed to recognize a trace, the ubiquitous spores of Ustilago receptaculorum (Plate V. fig. 94).
In the fenny districts of the eastern counties a species of “smut” called Ustilago typhoides attacks the stems of reeds, forming thick swollen patches of several inches in length (fig. 128), sometimes occupying the whole space between two joints or nodes, and lying beneath the sheath of the leaves. The spores in this species are larger than in the species which attacks the culms of grasses in a similar manner (Plate VI. fig. 129).
There are not many features in the rest of the species of this genus of sufficient interest to the general reader or microscopist to render it advisable to furnish any detailed account of them. We may, however, note that in a species found on the leaves of the common cock’s-foot grass the spores are large, obovate, and rough, with minute granules (figs. 117, 118). This species is known botanically as Ustilago 84salveii, and externally bears considerable resemblance, except in the size and colour of the spores (fig. 119), to another species much more common, and which occurs on the leaves of Poa aquatica and P. fluitans. The last-named species forms long parallel sori, extending often for several inches along the leaves of the aquatic grasses just named, giving them a very singular appearance (Plate V. figs. 105, 106). The spores are small (fig. 107), not being more than one-fourth the length of the last species, and smooth, whilst those are minutely granulated.
An interesting species occurs, very rarely, on the stems of such grasses as Aira cæspitosa and A. aquatica. The sori are in bands at regular distances apart (Plate VI. fig. 120), each band being composed of a number of short parallel sori (fig. 121). The spores are not more than one-third of the size of those in U. longissima.
Sedges are also subject to attack from other species of smut; one of these (U. olivacea) appears to convert the seeds into a fine olive-coloured dust (Plate VI. fig. 126), which gives to the fruit a similar appearance to that presented by corn when attacked by Ustilago segetum. Another species, which also absorbs the seeds, becomes hardened and consolidated more than in any other species, and, though larger than the normal seeds, still retaining their form (figs. 109, 110). This is Ustilago urceolorum, the spores of which are also figured (fig. 111).
The beaksedge (Rhyncospora alba) suffers from 85an allied species which affects it in a similar manner (Plate V. figs. 96, 97), but is not equally common. The spores of Ustilago utriculosa, found on different species of Polygonum, instead of being granulated, are reticulated on the surface (Plate VI. figs. 114, 116). The chief interest attaching to Ustilago antherarum consists in its habitat, for it is developed in the anthers of the flowers of the bladder campion, and other plants of the same natural order. The anthers are much swollen and distorted by this parasite, which is not uncommon, though easily overlooked unless specially sought after (Plate V. figs. 102-104). A list of all the British species will be found at the close of this volume. It will be noted that as in the genus Æcidium the prevailing colour of the spores is orange, so in the genus Ustilago it is black, with a purplish or violaceous tinge.
Four diseases in wheat of fungal origin are known and recognized in the popular language of the farm as “mildew,” “rust,” “smut,” and “bunt.” Sometimes one and sometimes another is most prevalent, and he is an exceedingly fortunate individual who can walk through his fields and find only one of them, especially if that one should be sparingly distributed. It has been our good fortune to dwell much amongst cornfields, and the terror of the word “mildew” to a farmer’s ears is not unfamiliar in our reminiscences of the past, ere we discarded the much-loved country to become a dweller in town. The subject of our present 86remarks inspired no such alarm in the districts of our experience, but in some seasons and localities it is certainly one of the “pests of the farm.” Under the different appellations of “bunt,” “pepper brand,” “bladder brand,” and sometimes “smut,” this infection is very generally known. Externally there is no appearance, except to the practised eye, that anything is wrong. There is no black impalpable dust about the ears as in the true “smut,” no red withered leaves or spotted stem as in the “rust” and “mildew,” and no stunted growth or malformation, evident to the casual glance, by which the insidious foe can be recognized; but stealthily and secretly the work is accomplished, and until the “bunted” grains make their appearance in the sample, the disease may, perchance, be unchallenged.
Externally the “bunted” grain is plumper, and whilst the corn is still green these will be of a brighter green than the rest (Plate V. fig. 84). When broken, the farinaceous interior will be found replaced by a minute black dust of a very fœtid, unpleasant odour, and greasy to the touch (fig. 85). This powder constitutes the spores of the “bunt” mixed with myceloid threads. It may happen that much of the corn in a field is “bunted,” and the discovery not made till the wheat is being ground for flour; then the odour and colour will speedily decide the produce to be unfit for human food. We have not the least doubt that “bunted” corn, when ground 87with flour, is injurious in proportion to its extent, whilst at the same time we can scarcely conceive an intelligent miller grinding up a sample containing any large proportion of “bunted” grains in ignorance of the fact.
If we break open a grain of wheat infested with the “stinking rust” or “bunt,” and then place some of the powder in a drop of water on a glass slide, and submit this to the microscope, first using the half-inch power, then the quarter, or fifth, and finally an eighth or tenth, we shall find that this minute dust consists of myriads of globose brown bodies termed spores, which possess certain reproductive functions. These spores will be found mixed with a number of delicate branched threads, to which they are attached by a short stalk or pedicel, visible with the higher powers (fig. 86). The surface of the spores you will also observe to be beautifully reticulated. These features just described as visible in the “bunt” are the characteristics of the genus to which it belongs (Tilletea), and of which it is the only British species. An allied species infests the Sorghum or durra, a grain but little cultivated in Europe, but found extensively in Africa and Asia, and also apparently found on the Bajra of India.
The interesting experiments of the Rev. M. J. Berkeley on the germination of “bunt” spores have been already alluded to. They were undertaken shortly after the outbreak of the potato 88disease, to ascertain, if possible, the mode by which the minute spores of fungi inoculate growing plants; and although at that time only a bare suspicion of the nature of the bodies resulting from the germination of “bunt” spores was entertained, succeeding examinations in the same direction have brought to light extraordinary facts, and manifested the progress of the successive developments of four generations. The spores of “bunt” are larger than those of the different species of “smut,” and reticulated on the surface (Plate V. fig. 86). When these are made to germinate a kind of stem is protruded (fig. 87), upon which small clusters of elongated thread-like spores of the second generation, or sporidia, are produced (fig. 88). After a time these spores conjugate, or become united by short transverse processes in the same manner as has been observed in some of the lower forms of Algæ (fig. 89). The conjugated spores in the next stage germinate and produce a third kind of fruit, different from either of the preceding, and constituting a third generation (fig. 90). These in turn germinate and produce a fourth order of reproductive organs (fig. 91), so that in the process of growth the “bunt” spores evidently pass through four generations. Hence, as one result, the number of germinating bodies is greatly increased, as well as their power of inflicting injury in a corresponding diminution in size. There are still many points in the history of the growth and 89development through successive generations of the “bunt” spores, but enough is known, on the one hand, to show that this is a true vegetative parasite, and not merely a diseased condition of the tissues of the wheat plant, and on the other that it is perfectly distinct from all the phases of the other and similar parasitic fungi which affect the wheat crop.
SOME of the microscopic fungi are the most unpromising and uninteresting objects to the naked eye which could well be imagined. No one would suppose that the black dust so profusely shed in such genera as Ustilago and Polycystis could be better than as much soot; unless he has learnt by experience not to judge by appearances, but to suspend judgment until examination. The axiom will sooner or later force itself upon all who examine minute objects with the microscope, that all organic nature, whether animal or vegetable, increases in interest in proportion to the magnifying power. Seen by the unaided eye, moulds are all nearly alike, and they seem to be “moulds” and nothing more. “Smuts,” again, sometimes attack one organ, and sometimes another, with very little variation in colour; and “rusts” are all “only rusts” with a paler or more intense rusty tint, until the marvellous combination of lenses, so appropriately named a microscope, unfolds a new world, and exposes its new inhabitants unparalleled in the old world of larger life, in form, habit, development, and mystery.
A very interesting, though small group of fungi, 91allied to the preceding, are included botanically under the genus called Polycystis, in allusion to the many cells of which the spores are composed.[6] In the most recent work on British Fungi, approximating to a Flora—viz., “Berkeley’s Outlines”—only three species are recorded, whilst the most common, at least around London, is omitted in error; for it could scarcely have been unknown as indigenous to this country. This last is the crow-foot smut (Polycystis pompholygodes, Lev.), found on the leaves and petioles of the common creeping buttercup (Ranunculus repens), distorting them very much, and also occurring on the wood-anemone and some other Ranunculaceous plants. The leaves and their footstalks, when attacked, become swollen, as if blistered at first, and ultimately burst in an irregular manner, exposing a mass of blackish soot-like dust (Plate IX. fig. 183), which on examination will be found to consist of the many-celled spores alluded to (Plate IX. fig. 184). Each of these spores appears to have a transparent outer membrane, either enclosing an unequal number, from two or three to five or six, distinct cells, compressed together into a spherical form by the outer integument, or the interior is divided by septa into as many cells. Each of these divisions contains a dark brownish endochrome, or cell-contents. 92As may be anticipated, the spores in all the species associated in this genus are interesting objects for the microscope. The species on the buttercup may be found through the summer and autumn on Ranunculus repens, especially whenever that plant is met with in very damp situations. We have seldom found the plant in any profusion without its attendant fungus.
6. Rabenhorst has proposed Urocystis as the name of this genus, on the ground that Polycystis was priorly applied to a genus of Algæ.
Another species of these many-celled smuts is not uncommon in gardens, on the sweet violet, attacking the footstalks of the leaves more commonly than the leaves themselves, and swelling and contorting them (Plate IX. fig. 185). In general structure the spores are very similar to those of the last species, save that the cells are smaller, and a larger number are collected together (Plate IX. fig. 186). So far as we have yet examined the spores of this and the preceding species, they appear to consist of separate and distinct vesicles (probably spores), contained within a hyaline sac or outer membrane, and not to be a single spore divided into cells by numerous septa.
A species of equal interest (Polycystis Colchici, Tul.) is found on the autumnal crocus, or meadow-saffron (Colchicum autumnale). The spores approach nearer to those of the last than of the prior species.
A fourth species occurs on the leaves of rye (Plate IX. fig. 187), forming elongated parallel blackish lines (Polycystis parallela, B. & Br.). It 93has also been found on the leaves of some grasses, but does not appear to be very common.
Many similar features are possessed by the two members of a genus named Tubercinia, which have been found in this country. One of the species is parasitic upon a plant which we who inhabit southern England never meet with, but which is not uncommon in Scotland, i. e., Trientalis Europæa. The parasite attacks the leaves about the month of September, forming bullate or blistered patches one-eighth to one-sixth of an inch broad, containing a mass of black spores (Plate III. fig. 52). These spores are irregularly globose, large, and opaque, consisting of a number of distinct cells (Plate III. fig. 53). Never having seen other than dried specimens—kindly communicated by Dr. Dickie of Aberdeen, the discoverer of this species—we cannot add much to its history beyond the published description by the Rev. M. J. Berkeley.
The other species occurs on potatoes, and is, during some seasons, common in all parts of Great Britain. The spores are curious, being composed of a number of cells arranged in the form of a hollow sphere, with one or two apertures communicating with the interior (Plate III. fig. 54). They are generally attached by a delicate thread. This species, sometimes confounded with the potato scab, was first described by Mr. Berkeley, about the time of the appearance of the “potato disease,” with which, however, it is in no way connected.
94Thus it will be seen that, inasmuch as we have complex brands in which the number of cells are considerably increased, so have we “complex smuts” in which, instead of one cell, we have many. In the last instance the two genera associated together in this chapter agree. The spores in both are distinctly cellular, but in the last genus far more opaque and consolidated than in the first. Whilst it may be doubted whether the compound spores of Polycystis are anything more than a number of individual spores with a gregarious habit, invested with an outer membrane, such a hypothesis cannot (as far as our individual examinations extend), be made to include Tubercinia. No doubt has yet been thrown on the genuine character of either of these genera. No Uredo or Æcidium, no Trichobasis or Puccinia has been ascertained or suspected to appear as a prior or subsequent form. In their supposed integrity they offer an interesting study, and in their development a good subject for investigation.
UNFORTUNATELY, this group of fungi contains species but too well known for their ravages amongst graminaceous plants, especially the cereals. “Corn rust,” as it is generally called, has a reputation little better than mildew, and it really deserves no better, for it is only another form of that pest of the farm, from the mycelium of which the corn-mildew is at length developed. There are two species very closely allied (doubtless only forms of the same species with different spores) which attack the leaves and culms of growing corn, and, bursting through the cuticle in the manner represented in Plate VII. fig. 141 (magnified), give a peculiar rusty appearance to the plant, as represented in Plate VII. fig. 140. One of these corn-rusts is botanically named Trichobasis rubigo-vera (Lev.), or the “true rust Trichobasis;” the latter, which is the generic name, being a compound of two Greek words (thrix, a hair, and basis, a foundation), on account of the spores being at first furnished at their base with a short, thread-like peduncle, which at length falls away (Plate VII. fig. 142). The other Corn rust is Trichobasis linearis, or “line-like Trichobasis,” because the sori or pustules are linear, 96or lengthened out like a line; the spores nearly double the length of those of the other Corn rust (Plate VII. fig. 144), and not so bright in colour. By intermediate forms these two rusts pass insensibly the one into the other, so that it is sometimes difficult to distinguish them. Both have the spores clustered together in the pustules, at first (Plate VII. fig. 143) attached by their peduncles, but they soon become free, and are scattered like rust-powder over the plant. Adverting to the attack of rust upon the wheat crops to an unusual extent a few years since, the Rev. M. J. Berkeley wrote as follows:—
We have seen rust more prevalent in white wheats, especially in the variety called Russian white, which has red smooth chaff, than we ever remember it. It is, however, confined to particular spots; and while in one field not a single leaf is free, insomuch that a person walking through the wheat is completely painted with the spores, of a fine rust-red; an adjoining field, separated only by a hedge, has not a rusty leaf. So long, however, as the rust is confined to the leaf, it is, we believe, perfectly harmless. The grain swells in spite of it, and the only effect is that the flag dies a little earlier, which is not undesirable when it is too luxuriant. If, however, it gets to any extent upon the chaff, much more if it attacks the seed itself, as is sometimes the case, it is very mischievous.

Plate VI.
W. West imp.
There is, indeed, a notion that rust is merely a form of mildew, and this may be true; but if so, it is a harmless form. The true rust (T. rubigo-vera), if a form of mildew, preserves its own characters to the end, and merely presents a second form of fruit, a circumstance which is exceedingly common amongst the lower fungi. It may exist in company with the mildew, but it also more frequently is developed without a trace of mildew intermixed. As far, then, as its immediate effects upon the plant 97go, it may be treated as a distinct parasite, though advanced botanists may look upon it as offering a case of what they call dualism, which may be easily understood on a comparison with such plants as marigold and orache, which produce seeds of two different forms. Its ultimate effects may, indeed, be injurious, as, if it be really a second form of mildew, it is capable of propagating that pest. The case is quite different with Trichobasis linearis, which is in fact merely the young state of the mildew. When once that supposed species makes its appearance, it is quite certain that there will be mildew to a greater or less extent. While the spores of the one remain unaltered, though intermixed with the true bipartite spores of the mildew, the other exhibits every intermediate state of form and colour.
M. Tulasne, to whom we have already alluded as a high authority on this subject, is of opinion that all the species enumerated in this group or genus, are mere conditions of other species, and therefore excludes it altogether. Another genus (Lecythea), for the same reason, has shared the same fate.
It must not be forgotten that twenty-eight species of rust belonging to this genus are now recorded as occurring in Great Britain. Some of these have yellow or yellowish spores; the rest have the spores of a decidedly brown colour. One of the most attractive of the yellow-spored species is that which appears on the groundsel (T. Senecionis), and a good notion of its external appearance may be gained from our figure (Plate VII. fig. 145). It is not always to be found without searching closely, for in some localities we have found it very sparingly, whilst elsewhere—as, for instance, on Wandsworth 98Common—we have met with it as soon or as late as there were groundsel leaves for it to grow upon. No spot or change in the appearance of the upper surface of the leaves indicates the presence of the parasite beneath; this is, however, often betrayed by the golden-yellow streaks which appear on the stem. Many a time and oft the boys in the semi-rural districts about town have marvelled and questioned each other concerning the reason why, in gathering “groundsel” for the little bird at home, we should be so particular, and pass so many promising plants, plucking here and there a leaf, and seldom collecting one which they consider eligible for the purpose. Collectors of minute fungi must expect to overhear occasionally even hints touching their sanity from those who, without the remotest idea of their mission, think they must be slightly “wrong in the head” to gaze so narrowly and intently, amongst nettles, groundsel, grass, or dry leaves, and only take an occasional fragment of a rotten stick, or two or three sickly leaves, carefully deposit them in their wallet, hat, or pocket, and then “move on.”
It will be unnecessary to repeat what has been stated in a former chapter on spermogones. It will be remembered that these organs are found developed in connection with some of the species of the present genus, as well as with the cluster-cups, or Æcidiacei. They consist externally of small conical elevations, pierced at the apex, which contain 99minute, cellular, linear bodies called spermatia, invested with a kind of mucous substance, that over-flows with them from the orifice of the spermogone, like lava from the crater of a miniature volcano. The rust in company with which they have been found most plentifully, is that which covers the under surface of the leaves of the commonest of all thistles (Cnicus arvensis). The external form of one of these spermogones is figured Plate VII. fig. 153. The rust possesses, when fresh, a peculiar odour, which is said to resemble that of orange-flowers; whence was derived its name of “sweet-smelling rust” (Trichobasis suaveolens, Lev.). M. Tulasne writes:—“With respect to this species, it is, in my opinion, but the first form of a Puccinia, analogous to P. Compositarum, D.C.: the spermogones with which it is mixed being very abundant, it ought to be placed with that Puccinia. I should, perhaps, retain some doubts upon the legitimacy of this relationship, if it had not been my fortune to meet on another species of the same kind (Puccinia Anemones, P.) spermogones perfectly characterized, and in which the colour, habit, and position would not allow me to hesitate an instant in attributing them to the Puccinia.” In this species of rust the whole under surface of the leaf is covered with a purplish-brown dusty coating of the spores, from the numerous pustules which are produced (Plate VII. fig. 151). The plants, when attacked, have a paler and more sickly appearance; the leaves 100have a tendency to fold backwards at their margins, and thenceforth their growth seems to be determined. Sowerby, in his “British Fungi,” says:—“Two or three sorts of flies are occasionally found dead on this plant at the time of the fungus being upon it, which is after wet weather in the summer, or early in autumn; being apparently tempted by its flavour, they over-eat themselves, or else are destroyed by some poison.” This rust has spores resembling, in general characters, those of the yellow-spored series Plate VII. fig. 152).
We have not thought it necessary to give figures of many species, partly on account of the uncertainty existing in many minds whether they ought to be regarded as species, and whether they will long claim a place in the British Flora; and partly on account of the similarity which exists between them, at least so far as they are of interest to the microscopist only.
During the autumn of last year, whilst on a botanical excursion through a portion of Epping Forest, the “great bog” became a centre of some interest. Bogs are generally attractive spots to those who are in search of microscopic organisms. On this occasion the chief objects of interest were the small brown pustules (Plate VIII. fig. 168) with which the upper surface of a large number of the leaves of the pennywort (Hydrocotyle vulgaris) were sprinkled. These pustules were brown, orbicular, regular, and in habit seemed to resemble rather 101those of most of the Pucciniæ than of a Trichobasis. The large, vigorous, and healthy leaves were less affected. Microscopical examination, at first incomplete, led us to the conclusion that it was a species of Uredo, which had been met with in France (Uredo Hydrocotyles, Mont.); but a re-examination, to which we were prompted by Mr. Currey, led to the conclusion that it belonged rather to the present genus; but it can scarcely be associated with any species already described, notwithstanding its apparent affinity with the brown rust found on umbelliferous plants, in which the pustules are invariably developed on the under surface. Under these circumstances, we have called it the Pennywort rust (Trichobasis Hydrocotyles), whilst still doubtful whether it is not the same fungus as that described by Montagne, with whose description it agrees in everything, except what is implied by the generic name. It should not be forgotten, that the work in which Montagne first described this species was published when the genus Uredo embraced the present genus, which was separated from it by Leveille twelve years later; and it is possible that this species was included in a more recent work by the same author, in error and without re-examination, under the old name. An examination of authentic specimens of Montagne’s plant would settle the point; but we know of no published collection which contains it. The characteristic difference between the two genera lies in 102the presence of a peduncle in the early stage of Trichobasis Plate VIII. fig. 169), and its absence in all stages of Uredo. Without wandering further into a subject which has not the merit of being very popular, let us away to some green lane in search of violets, and having found them, take a little of the brown dust from one of the small pustules on the leaves, upon the point of a penknife; place this, with a drop of water, upon a glass slide, and make a record of what we observe.
The field is covered with the myriad spores of a rust of a nearly spherical shape, brownish in colour, and here and there one with a short transparent colourless stalk or pedicel. This is the violet rust (Trichobasis Violarum, B.), very common all through the summer and autumn, generally on the under surface of the leaves of violets, in woods and hedgerows. Should it so happen that the spores when placed under the microscope are found to be two-celled, it will prove that instead of a rust, or Trichobasis, being under examination, a brand, or Puccinia, has been found, which is almost equally common, and which may, without such a test, be easily mistaken for a rust. According to the theory of di-morphism, this is the higher form or complete fruit of the same fungus, which in its simple-celled state is called Trichobasis Violarum.
A similar circumstance may befall the student in examining the rust of labiate plants (Trichobasis Labiatarum, Lev.), which occurs on different species 103of mint, especially the water-mint, about the month of August. We have found a few of the two-celled spores of the Puccinia imbedded in the pustules of this rust almost constantly, whilst the one-celled spores are not uncommon in the pustules of what is regarded as the true mint brand (Puccinia Menthæ, P.).
In spring the young leaves of the periwinkle (Vinca major) will, in some situations, become thickened considerably, and ultimately browned on both faces with the pustules of a rust (Trichobasis Vincæ), which though covered with a conidioid dust are often very tardy in bursting the epidermis. It is almost impossible, after the pustule is ruptured, to find one in which the bilocular spores of Puccinia (fig. 132) are not largely intermingled with the unilocular spores of the “rust” (Plate VI. fig. 131). Later in the season by two or three months, other leaves of the same plants will be found occupied by the smaller and more widely scattered pustules of Puccinia Vincæ, in which the unilocular spores of the “rust” will seldom be found. In this instance the leaves are scarcely thickened, and the colour of the spores is much darker. The under surface of the leaves is commonly alone occupied, and corresponding paler spots on the upper surface indicate the presence of the parasite beneath.
The garden and field bean is liable in some seasons to become quite rust-coloured in consequence of the profusion of spores with which the leaves 104and stems are covered, from the bean rust (Trichobasis Fabæ, Lev.), which in like manner is considered as the simple stage or form of the bean brand (Puccinia Fabæ, Lk.). If the legumes are also examined, a few pustules will sometimes be found on them. Beans thoroughly infected with this rust or brand are seldom of much service in either field or garden.
Beetroot, or mangold wurzel, is another example of garden and field produce which is subject to a similar visitation. This rust often has very red spores when produced on red varieties of beet. During September, 1863, it (Trichobasis Betæ, Lev.) was sent us on the leaves of the wild beet (Beta maritima).
All the species of Polygonum are exceedingly subject to the attacks of the Persicaria rust (Trichobasis Polygonorum, B.), which nearly covers the leaves, till the entire plants seem to be smothered in Scotch snuff. If it were allowable to affirm of any plants that they are martyrs to parasitism, such might be said of the Persicarias and their allies.

Plate VIII.
W. West imp.
Sedges are subject to the attacks of a rust much resembling the corn rust. When it occurs on some species of Carex, the upper surface of the leaf has corresponding pale spots, and the pustules themselves are surrounded by a yellowish margin. This species (Trichobasis caricina) is far from uncommon (Plate VIII. figs. 170, 171). The sedge-like plants belonging to the genus Luzula have also their own 105species of rust (Trichobasis oblongata), the spores of which are deeper in colour than in the sedge rust (Plate VII. figs. 158, 159).
Amongst the remaining species of Trichobasis (the reader must pardon our using the generic name, as we have no equivalent), that found on the leaves of various composite plants is the most common (Trichobasis Cichoracearum, Lev.). It occurs on some thistles, on the saw-wort, dandelion, several species of hawkweed, and similar plants. The pustules are small and more diffused than in the species found on Cnicus arvensis, and they as often appear on the upper as on the lower surfaces of the leaves.
On umbelliferous plants three species are recorded; one with yellow spores (Trichobasis Petroselini, B.); another with a blistered habit, and brown, ovate, or oblong spores (T. Umbellatarum, Lev.); and a third with tawny, obovate, or egg-shaped spores (T. Heraclei, B.), which is found solely on the cow-parsnip. The species of Puccinia corresponding to some of these species of Trichobasis are known, but, in other cases, probability, or speculation if you please, occupies the place of knowledge.
During the month of September, 1864, it was our good fortune to spend a week in revisiting the scenes of our boyhood, and exploring the minute botany of one of the marshy districts of East Norfolk. One day of the seven, memorable to us for 106the discovery of three specimens of a large Boletus (B. cyanescens),[7] not found, to our knowledge, since the days of Sibthorpe, was further enriched by a species of Trichobasis, new to Britain, and apparently uncommon on the Continent. This rust was found on the leaves of the “grass of Parnassus” (Parnassia palustris) on a narrow strip of marsh near Irstead church. It was sought in vain elsewhere. The leaves were scarcely changed in appearance, except by the presence of the pustules. There were no discoloured spots, but the pustules appeared sometimes plentifully, more often scattered, on both surfaces of the leaves: they were small, of a bright brown, with oval spores; the latter were, in their early stages, shortly stalked. We have called this species Trichobasis Parnassiæ. It is possibly the same as published by Westendorp in his “Herbier Cryptogamique Belge” as Uredo Parnassiæ, but we know of no copy which we can consult, and have failed in discovering any other species to which we can refer it. It is certainly a Trichobasis and not an Uredo, according to the present limitation of the latter genus.
7. Two of these specimens were found at the bottom of a hedge-bank, amongst grass, by the side of the road leading from Neatishead-street to Irstead Rectory, and the third in a similar position by the Norwich road, two hundred yards from the turning which leads to Neatishead-street.
Although the evidence against the retention of the species of Lecythea (as the genus is named) 107amongst Fungi as true species, on the ground of di-morphism, is even stronger than against the group just illustrated, we cannot pass them altogether in silence, especially in a popular treatise. Those who are residents in town, and yet possess their little plot of garden-ground, with only two or three pet roses, may have had the misfortune of seeing them smothered with a yellow blight. This golden visitation, unwelcome as it is, may afford a subject for the microscope, and for a small space in this chapter. At first there will not appear to be any important difference between the spores of the yellow series of the last genus and those of the present; but a closer examination will reveal one important distinction, viz., the presence of colourless elongated, abortive spores. The species are not so numerous by half as those of Trichobasis, even when three anomalous forms are included, which species are included by some mycologists in two other genera. One very common rust of this group has already been alluded to (Plate II. fig. 37), and which is known botanically as Lecythea Rosæ. A similar one is found on the bramble, and another on the burnet. All these three species are produced at first on spots which are afterwards more or less occupied by the long, many-celled spores of the dark brown brands called Aregma or Phragmidium, between which and the simple yellow spores of the rust almost every intermediate form may often be found in the same pustule. Thus, from the same 108mycelium as that of the rose rust, the rose brand is afterwards developed; whilst from the nidus of the bramble rust (Plate III. fig. 40) the bramble brand is also at length produced; and the successor to the burnet rust (Plate III. fig. 31) is the burnet brand. Besides these, a rust belonging to the same genus may be found on the leaves of the poplar, the spurge, and the common valerian, and two or three species on willows. It can scarcely have escaped notice, that the goat-willow is almost constantly afflicted with a rust on the under surface of the leaves (Plate VIII. fig. 160). This species will again come under notice as the summer spores of a truly dimorphous species.
One of the rusts separated by some botanists from this genus is found (possibly most commonly) on the leaves of the raspberry; but during the past autumn we have met with it plentifully on the upper surface of the leaves of one or two species of bramble, and have never seen it growing on the raspberry, although in all descriptions of the species that is stated to be its habitat. Even to the naked eye this is so distinct, that no one could well confound it with any other. It appears very late in the autumn, and the spots are scattered at some distance apart from each other (Plate VIII. fig. 162); each spot or pustule forming a ring (Plate VIII. fig. 163 enlarged) encircling a cluster of spermogones which occupy the centre.
Another rejected species (Lecythea Lini, Lev.) 109occurs on the little purging flax (Linum catharticum), forming small pustules on the leaves (Plate VIII. fig. 165); these burst irregularly, and remain surrounded by the remains of the ruptured epidermis (Plate VIII. fig. 166). The yellowish spores are subglobose (Plate VIII. fig. 167), and in the first instance concatenate, or chained together like a necklace, which circumstance has been taken advantage of to place it, with one or two other species, in a separate genus.
We cannot claim for the species brought into notice in the present chapter any attractive features resulting from singularity of form, complexity of structure, or delicate tracery, whereby they might commend themselves to mere “searchers after curiosities,” or be recommended from friend to friend as “sensation” objects for the microscope. They do possess an interest and a value, but such as would not be appreciated by those who seek to pass an idle half-hour by gazing at some new thing.
A QUARTER of a century ago, and all the fungi enumerated in the preceding and in the present and following chapters would have been arranged under three genera, called respectively Æcidium, Puccinia, and Uredo. Under the last-named genus all the species illustrated in the present chapter, beside many others, would have found “a habitation and a name.” There are still a few which bear the old generic name, and, if only out of respect, we shall grant them the first place.
Let the first bright day in May witness the student beside a cluster of plants of Mercurialis perennis, which it will not be difficult to find in many localities, and, on turning up the lower leaves, he will meet with our first illustration of a genuine Uredo, in the form of yellow confluent patches, with a powdery surface (Plate VII. fig. 133). This will be Uredo confluens. By the way, the generic name is in itself suggestive, which it always should be in all instances, but unfortunately is not; it is derived from the Latin word uro, “I burn,” and is peculiarly applicable in instances where the leaves acquire a blistered, burnt, or scorched appearance, occasioned by the 111presence of the fungus. The microscopical features of the spores of this genus are, briefly, a more or less spherical form, without any pedicel or footstalk Plate VII. fig. 134). These spores are at first produced each in a separate cell, but when ripe become free, and are at times with difficulty distinguished from such forms as Trichobasis, unless the pedicels in the early stage of the latter genus are regarded.
A rare species in Britain is the oak-leaf rust (Uredo Quercus), in which the sori or pustules are minute, and at first yellow, but afterwards orange. It occurs on the under surface of the leaves, and was, we believe, first found in this country by Mr. D. Stock, in the neighbourhood of Bungay, nearly five-and-twenty years since. That gentleman informs us that it was not at all uncommon, but always appeared on the young shoots which had sprung up from the trunks or roots of trees that had been cut down. We are uncertain whether it has been found by any one since that time in this country, although it is not uncommon in France.
The leaves of the common sorrel are often sparely sprinkled with the pustules of a rust (Uredo bifrons, Grev.), which derives its specific name from the fact that the pustules, which appear on both surfaces of the leaves, are often opposite to each other Plate VII. fig. 137). These pustules are generally seated on a discoloured spot (Plate VII. fig. 138 enlarged), and are surrounded by the 112remains of the ruptured epidermis. The spores are globose and brown Plate VII. fig. 139). We have not met with any other Uredine on the sorrel leaves, though one having a similar appearance to the naked eye is not uncommon on several species of dock.
The fern rust (Uredo Filicum), which occurs on the under surface of the fronds of two or three species of ferns, we have never met with, and do not think that it can be considered common. It has been found in the West of England, on Cystopteris, and Sowerby collected it, probably not very far from London.
One of the most common is the rust found on the leaves of the enchanter’s nightshade (Circæa lutetiana), sometimes nearly covering the under surface with its tawny snuff-coloured spores Plate VII. fig. 135). The plant on which it is found is rather local, but the rust (Uredo Circææ) seems to abound wherever the plant on which it is parasitic flourishes. Another fungus of a very similar external appearance may be collected, more rarely, from the leaves of the same plant; but in this the spores are two-celled, or divided by a transverse partition. This fact is mentioned to guard against disappointment, should the spores be found to differ from the characters of this section, and to show that the microscope is essential to the study of fungi. The spores of the true Uredo are globose and without any partition Plate VII. fig. 136).

Plate IX.
W. West imp.
113We may also observe that a very interesting species may be found on several species of St. John’s-wort (Hypericum), but especially on the under surface of the leaves of the Tutsan, covering them with its golden-coloured spores (Plate VIII. fig. 174). Another occurs on the leaves of some of the wild garlics (Allium); one is not uncommon on some of the stitchworts; another on saxifrages; another on willow-herbs (Epilobium), and one on the leaves of the cowberry. All of these have yellowish spores. A species with brown spores occurs on sea-lavender (Statice). Space to write on our own part, and patience to read on the part of others, induce us to dismiss all these species with the bare allusion to them and the plants on which they may be found. Some of them may occur in one locality and some in another, and enough has perhaps been said to enable any one to place any that he may find in the proper genus, if not with the scientific name.
From the numerous instances we have met with of persons wholly disinterested in the subject, collecting and making inquiries concerning the bright orange patches next to be referred to, we are led to conclude that most persons have, at least, seen them. These patches of orange-coloured powder burst through the cuticle, and appear on the young branches of wild roses, extending sometimes for two or three inches in length Plate VII. fig. 147), and form one of the most beautiful and 114obtrusive of the dust-like fungi. It is not confined to roses; but the meadow-sweet, on which it is also found, does not grow in localities where its parasite is so liable to meet the gaze of the ordinary wayfarer. The habit of this rust is more or less that of those which are associated with it. The pustules are not small and orbicular as in most instances in other groups (except Polycystis), but are large and irregular, and generally but few together or single. The spores are variously coloured, and have peduncles, or footstalks, of a greater or less length. In the rose rust these spores are profuse, but the peduncle is short (Plate VII. fig. 148).
Plants of the pea and bean tribe are liable to be attacked by one of these rusts, and in this instance the spores are so characteristic that no one could well confound them with any other. Externally it appears as an irregular brownish pustule, breaking through the epidermis and filled with an impalpable powder, not unlike a pinch of “brown rappee” snuff. The spores are ovoid, with a very long peduncle, whence its name (Uromyces appendiculata). It appears on a great variety of plants, but from the peculiarity of the spores Plate VII. fig. 150), is easily recognized. Our figure represents it on the leaf of a vetch (Plate VII. fig. 149). Almost at the commencement of this volume (Chap. III.) we had occasion to refer in detail to some experiments made by De Bary on the spores 115of this species. In the remarks then made occurs a recommendation of reperusal, which would obviate any repetition here.
An allied species, and a beautiful one, is to be found on the stinking iris (Iris fœtidissima), and another on the under surface of the leaves of primroses.
The pilewort (Ranunculus ficaria) we have already seen attacked by one species of microscopic fungi, and we have now to record the occurrence of another. Some plants appear to be destined both alive and dead to become the prey of others. The common nettle, for instance, as if in retribution for the annoyance it often occasions on account of its stinging propensities, has not less than twenty different species of minute fungi, to say nothing of coleopterous and lepidopterous insects, which make a home, sometimes upon its green leaves, and sometimes on its dead stems. We might almost state that it has a flora and a fauna of its own. The pilewort, too, has many foes; but these are fewer in number, and mostly attack the living plant. The cluster-cups have been already noticed; some do not fall within the limits of this volume, but one, which is found in May and June, belongs to the present genus. It appears like a purplish-brown powder bursting through blistered spots on the leaves and footstalks Plate VII. fig. 156). The spores are small, and are, of course, provided with pedicels Plate VII. fig. 157).
116The under surface of the leaves of the white Dutch clover are often sprinkled with black spots, which are nearly round and very numerous. These are so many clusters of fungi belonging to a different section, in which the threads are the important feature. But another parasite is also found on leaves of the same plant, in which the pustules are far less numerous and regular, and are often found on the petiole as well as the leaf, distorting them and twisting them in various directions Plate VII. fig. 154). This is the clover rust (Uromyces apiculata, Lev.), which is a parasite on numerous plants, being found also on the great water-dock and other kinds of dock. The spores are ovoid and brown, with a short peduncle (Plate VII. fig. 155). A very beautiful species occurs on the leaves of the ladies-mantle (Alchemilla), but hitherto we have not been fortunate enough to collect it.
It can scarcely be too great an assumption to suppose that every one is acquainted with the goat-willow (Salix caprea), or that every schoolboy knows the birch (Betula alba). It may be proceeding a step too far to affirm that all who know these trees well enough to distinguish the one from the other, will have observed the under surfaces of the leaves of both sprinkled with a golden dust, during the summer months, and which are the spores of a parasitic fungus. So common is this orange-coloured powder on leaves of the trees 117above-named, that we can hardly believe any one to have had a branch of either in his hand and not observed it, provided any leaves adorned the branch in question. What this parasite is, and what its associates, it is our province to endeavour to explain. Our figure (Plate VIII. fig. 160) represents a leaf of the sallow or goat-willow, with the under surface exhibiting yellow patches, consisting of spores, which are magnified in the next figure (Plate VIII. fig. 161). This exceedingly common rust is termed Lecythea caprearum, Lev., when in the condition figured; but in reality this is only the summer stage, bearing the summer fruit of Melampsora salicina, which latter attains its mature development on the same leaves in the succeeding winter (Plate IX. fig. 191) or early spring. Of course this latter remark applies to the fallen leaves, for at this period all the willows and other deciduous trees are bare. But the leaves, before they fall, give evidence of the parasite at work; and if the collected decaying mass of rubbish at the base of sallow bushes be examined about March, these leaves will be found bearing upon them mature heaps of elongated, wedge-shaped spores, closely packed side by side (Plate IX. fig. 192), and which, whilst still adherent, may often be found in active germination, as represented in an allied species at the bottom of our plate (Plate IX. fig. 197). This phenomenon consists in the production of cylindrical tubes, more or less elongated, from the upper 118extremity (rarely from the base) of the prismatic spores. These tubes are straight or twisted, simple or forked, and each of them becomes divided by transverse septa into four unequal cells towards their apex, from each of which is produced a spicule bearing a sporidium, or spherical secondary fruit, in the same manner as in the genera Puccinia and Aregma. It should be observed, that the winter spores of this rust are borne on the opposite surface of the leaves to the summer spores: for whilst the latter are developed from the under surface, the former are found on the upper. This being also an instance of di-morphism, the summer condition, when spherical spores are produced, should not be regarded as a distinct plant, and the name of Lecythea caprearum does not merit retention in the list of fungi.
There are five species of this interesting group, or genus, found in Great Britain, to the residue of which we may only briefly allude. It has already been stated that, in summer, the yellow spores of a rust are found on the under surface of birch-leaves. These must be sought on the young twigs or suckers, proceeding from the stumps of trees which have been cut down: pale discoloured spots on the upper surface of the leaves indicate the presence of the rust beneath. This is the Uredo betulina of old authors (begging their pardons, for some of them still live), the Lecythea longicapsula of more recent times, and the summer spores of Melampsora betulina, 119according to M. Tulasne and his disciples. When fresh, it is reputed to exhale a faint odour, as of the primrose. During the winter and spring months the wedge-like spores of the second crop are matured on the fallen leaves (Plate IX. figs. 189, 190), and these are capable of a speedy germination, and the production of secondary reproductive bodies, as in the willow rust above alluded to.
Probably, also, the similar rust on the poplar (Plate IX. figs. 195, 196), or on the aspen, may be met with under like conditions; i.e., the summer spores, which are yellow and spherical, on the living leaves, and the brown permanent masses of winter spores on the fallen and decaying leaves. The sole remaining British species is not uncommon on leaves of the common spurge in gardens, and whilst the yellow pulverulent spores occur on the upper, it will not be improbable that black permanent spots will be found on the lower leaves (Plate IX. figs. 193, 194), enclosing closely-packed, rudimentary, elongated or wedge-shaped cellules of the winter spores.
Any one may make himself acquainted with the genus Coleosporium with but little trouble, which the acquisition will more than compensate. A summer stroll into any locality in which the common coltsfoot can be found, will be certain to prove sufficient. Let the spot selected be any station on the North Kent Railway, for those who reside in town, or even a trip to the Crystal Palace and a 120stroll in the grounds, and when the well-known leaves of the coltsfoot are descried, the under surface of the first leaf will doubtless give proof of the presence of the fungus in question, by the orange spores amongst its dense woolly hairs. Sometimes the leaf is almost covered beneath with the bright orange-coloured dust. This is the coltsfoot rust (Coleosporium tussilaginis, Lev., Plate VIII. fig. 180), which may serve as a type of the rest. It may be observed that a species of cluster-cup, or Æcidium, with spores of nearly the same colour, is also to be found on the leaves of the same plant; but in this case the upper surface of the leaf has also corresponding purplish spots, and, what is of still more importance, the spores are seated in small fringed cups. The rust is common till the wintry frosts have set in, and is far more conspicuous than the cluster-cups. A kind of di-morphism prevails in all the species of this genus. Some of the pustules resolve themselves into a kind of powder, whilst others remain entire and solid. Generally there is the largest proportion of globose, dust-like, free spores, produced in the earliest developed fungi, whilst they become more rare towards the close of the season. The permanent spore-spots consist of obovate cellules placed side by side, each of which is divided transversely by three or four septa, and is filled with an orange-red endochrome (Plate VIII. fig. 181); the exterior being enveloped in a kind of mucous layer. The arrangement of spores when packed 121together in the pustule is shown in Plate VIII. fig. 182, from an allied species. When these spores germinate, which they do with great readiness, each division emits a long tube, which generally remains simple and undivided, and from its extremity is produced a reproductive body of an obovate or nearly kidney-shape. These filaments are about 1/120th of an inch in length, of a colourless transparent membrane, along which the orange-red contents of the spores pass into the newly-formed sporidia, or reproductive bodies by which they are terminated. Most of these reniform sporidia disengage themselves from the filaments on which they are produced, and either elongate themselves into a simple and uniform filament, or swell at the extremity as if to reproduce a second spore. If the newly-formed sporidia do not become free, they increase the length of their primitive filament, which by a frequent repetition of the process becomes a tube swelling out at unequal distances.

Plate X.
W. West imp.
The summer spores, or pulverulent spores of the first generation, which are analogous to the Uredospores of Aregma, are also capable of germination, for, if placed in favourable circumstances, they will develop very long filiform processes, which either remain simple or become more or less branched, but always nearly uniform in their diameter. M. Tulasne states that he has observed this germination many times, though we have been less fortunate.
122Of the six species of this genus known to occur in Great Britain, the majority may be commonly met with. That very widely-diffused plant, the wood cow-wheat (Melampyrum pratense), known well to all amateur botanists for its pertinacity in drying black, and presenting anything but an inviting appearance to claim for it a place in the herbarium—also becomes the matrix for the development of a member of this genus; i.e., the cow-wheat rust (Coleosporium Rhinanthacearum, Lev.), and which is found on other allied plants, as the little eyebright (Euphrasia officinalis), &c. (Plate VIII. fig. 176). In colour and habit it resembles the last-named species, and its free, echinulate spores (Plate VIII. fig. 177) form a pretty object for the microscope.
Another equally common species is found inhabiting the leaves of the sow-thistles (Sonchus arvensis and S. oleraceus), and in the autumn may generally be found on either of those plants, presenting the appearance delineated in our plate (Plate VIII. fig. 178). The permanent spores resemble in many points those of the first species, as will be seen from the figure from De Bary’s treatise on this subject (Plate VIII. fig. 179). This is certainly one of the most showy of uredinous fungi, and could not be well overlooked.
The butter-bur rust (Coleosporium petasites, Lev.) and the Campanula rust (Coleosporium Campanulæ, Lev.) are found, the former on the leaves of the 123butter-bur, and the latter on those of the harebell and other Campanulæ, less frequently. We have, however, indicated sufficient, since their great similarity in unprofessional eyes will furnish, in one or two species, all that is desirable for the microscopist.
Unless some similar plan to the following be adopted for examining the species of this genus, it may result in disappointment; for the slight attachment of the joints to each other will otherwise present only a mass of simple echinulate cellules, if a portion be only removed from the leaf on the point of a lancet. This method consists in making a thin vertical section of a pustule in which the spores are contained; by this means the arrangement of the fruit and the mucedinous threads from whence they proceed may be observed. Any person possessed of the cardinal virtues of microscopy—patience and perseverance—will be rewarded in this instance; whilst those who are deficient will lose an object worthy of the virtues they dare not boast. But few instances have occurred in this and the preceding chapters in which the exercise of any great ingenuity or application has been called for; the most juvenile or truest tyro at the microscope may see for himself much of what has been indicated, whilst a few opportunities have occurred for more practised manipulists to prove that they are neither juveniles nor tyros.
ALLUSION has already been made to the important memoir recently published by Dr. de Bary. “White rusts” occupy a conspicuous position in that memoir, and the experiments therein detailed, with the conclusions arrived at, will be largely drawn upon in furnishing the present chapter. Whilst believing that we have fairly represented the views, and faithfully narrated the story of research, if not literally, but denuded of some technicality, yet in such manner as to convey the sense of our author, we claim no originality or merit save for the garb in which it appears, without addition, stricture, or confirmation of our own.
What is the external appearance presented by the “white rust” of cabbages, and allied cruciferous plants, is soon told. During summer and autumn it occupies the surface of the leaves and stems of the shepherd’s-purse (Capsella bursa-pastoris), with elongated narrow white spots like streaks of whitewash (Plate X. fig. 198), and later in the season the leaves of cauliflowers and cabbages become ornamented with similar patches, arranged in a circular manner (Plate X. fig. 199), forming spots as large as a sixpence. Wherever 125these spots appear, the plant is more or less deformed, swollen, or blistered, even before the parasite makes its appearance at the surface. These white pustules have a vegetative system of ramifying threads which traverse the internal portion of the plants on which they are found: these threads constitute what is termed the mycelium. Not only when the plant is deformed and swollen with its undeveloped parasite do we meet with the threads of mycelium in its internal structure, but also in apparently healthy portions of the plant, far removed from the evidently infected spots. These threads are unequal in thickness, much branched, and often with thick gelatinous walls filled with a colourless fluid. They creep insidiously along the intercellular passages, and are provided with certain appendages in the form of straight thread-like tubes, swollen at their tips into globular vesicles (Plate X. fig. 204). These threads do not exceed in length the diameter of the mycelium which bears them. The appendages communicate in their interior with the mycelium, and contain within them the same fluid, which at length becomes more watery, and the terminal vesicles have their walls thickened, so as to resemble, on a casual observation, granules of starch. Dr. de Bary conceives that these appendages serve a similar purpose to the tendrils or suckers of climbing phanerogamic plants; i.e., to fix the mycelium to the cells which are to supply the parasite with nourishment. As 126these appendages are always present, it is easy to discover the mycelium wherever it exists amongst the tissues of an affected plant.
The white pustules already alluded to contain the fruit of the parasite. Bundles of clavate or club-shaped tubes are produced upon the mycelium beneath the epidermis of the infested plant, forming a little tuft or cushion, with each tube producing at its apex reproductive cells, designated “conidia.” These conidia appear to be produced in the following manner:—The tips of the clavate tubes generate them in succession. At first a septum, or partition, divides from the lower portion of the tube a conidium cell; this becomes constricted at the septum and assumes a spherical shape, at length only attached by a short narrow neck. Beneath this again the same process is repeated to form another and another conidium in succession, until a bead-like string of conidia surmount each of the tubes from which they are produced (Plate X. fig. 200). At length the distended epidermis above is no longer able to bear the pressure of the mass of engendered conidia within, and is ruptured irregularly, so that the conidia, easily separating from each other at the narrow neck, make their escape.
As long since as 1807, M. Prevost described the zoospores, or moving spores, of these conidia, and his observations were confirmed by Dr. de Bary three years since, and are now adverted to by him again 127in further confirmation. If the conidia (white spherical bodies ejected from the pustules of the “white rust”) are sown in a drop of water on a glass slide, being careful to immerse them entirely, they will rapidly absorb the water and swell; soon afterwards a large and obtuse papilla, resembling the neck of a bottle, is produced at one of the extremities. At first vacuoles are formed in the contents of each conidium; as these disappear, the whole protoplasm (granular substance filling the conidium) becomes separated by very fine lines of demarcation, into from five to eight polyhedric portions, each with a faintly coloured vacuole in the centre. These portions are so many zoospores. Some minutes after the internal division, the papilla swells and makes itself an opening, through which the zoospores are expelled one by one, without giving any signs of movement of their own. They take a flat disk-like or lenticular form, and group themselves about the opening, whence they have been expelled, in a globular mass. Soon, however, they begin to move, vibratile ciliæ show themselves, and by means of these appendages the entire globule oscillates, the zoospores disengage themselves from each other, the mass is broken up, and each zoospore swims off on its own account (Plate X. fig. 208).
The free zoospores are of the form of a planoconvex lens, obtuse at the edge. Beneath the plane face, out of the centre, and towards that 128point of the margin which during the movement of the zoospore is foremost, is a disk-shaped vacuole, with two ciliæ of unequal length attached to its margin; the shorter cilia is directed forwards, and the longer in the opposite direction, during the evolutions of the zoospores.
The zoospores are produced within from an hour and a half to three hours after the sowing of the conidia in water. They are never absent if the conidia are fresh, or even a month old, but beyond this period their artificial generation is very uncertain. This little experiment is a very simple and interesting one, and may be performed by any one who will take the trouble to follow out these instructions.
From this simple experiment, let us turn for a moment to the plant in its natural condition when affected by the white rust. If, after rain or dew, when the little drops of moisture hang like pearls about the sickly pallid leaves of the shepherd’s-purse, bespattered with the white pustules of the rust, we collect and examine a drop of water from the immediate neighbourhood of one of the pustules, we shall commonly find empty conidia and zoospores in different stages of development.

Plate XI.
W. West imp.
Water alone seems to be essential to them, and for this the conidia may remain unchanged for a month, and literally burst into activity at the first gentle shower, till the whole surface of the plant is swarming with zoospores. We may no longer 129doubt that a true vegetable produces from itself bodies endowed with active motion, resembling low forms of animal life, and yet in themselves not animalcules, as some would suggest, but essentially vegetable, as we shall hereafter demonstrate. To scientific men this is not new, except as regards fungi, for in algæ such bodies have long been recognized.
A second kind of reproductive organs are described by Dr. de Bary; and if future examinations confirm his observations, as they doubtless will, this feature is an important one. It is true that M. Caspary long since detected similar bodies in moulds (allied to that which produces the potato disease), but he only knew them in a limited sense compared with what De Bary has revealed. These fruits are hidden amid the tissues of the plant on which the “white rust” is parasitic, and only betray their presence by the coloration of those tissues. To these bodies it is proposed to give the name of “oogonia” and “antheridia,” on account of their presumed sexuality, the “oogonia” representing the female, and the “antheridia” the male organs.
The oogonia are large spherical or ovoid cells, with a thickish membrane containing a granular protoplasm, or formative fluid. They are produced either terminally or laterally upon the threads of the mycelium, from which they are separated by septa or partitions.
130The antheridia are somewhat blunt-shaped or obovate cellules, considerably smaller than the oogonia, with slightly thickened walls, and containing a finely granular protoplasm. These are produced upon branches of the mycelium which do not bear oogonia. The obtuse extremities of these branches, which are to be developed as antheridia, are applied to the surface of the growing oogonia, to which they adhere, become distended, assume their obovate form, and by the formation of a septum at their base, their contents are isolated from those of the threads of the mycelium, and thus the antheridia are perfected.
When these bodies have attained their full dimensions, the large granules which are contained in the oogonium accumulate at its centre, and form an irregular, somewhat spherical mass, which is called by De Bary a gonosphere. This gonosphere having been formed, a straight tube shoots out from the antheridium which perforates the wall of the oogonium, passes through the fluid which surrounds the gonosphere, elongating itself until it touches that body. From this period a membrane begins to be formed about the gonosphere, which thenceforth maintains a regular spheroidal form. It may be observed that the extremity of the tube which proceeds from the antheridium does not open, and the fecundation, if such it be, is produced solely by contact. After this contact of the two bodies, the gonosphere acquires a new name, 131and is called an “oospore.” The membrane which at first invests this organ is very thin, but by deposits from the surrounding fluid it attains to a greater thickness, and is at length of a yellowish-brown colour, having its surface studded with large obtuse warts (Plate X. fig. 206). One of these warts, larger than the rest, forms a kind of thick sheath around the fecundating tube.
The oospores do not give evidence of any appreciable change for some months. For instance, those collected by De Bary in June did not attain their ulterior development until the commencement of December. The method adopted was as follows:—Parts of the plants containing ripe oospores were preserved in the dried state. When examination was considered desirable, the portion to be employed was immersed in water for a day or two; it was then placed on a humid soil, or mould covered with blotting-paper. The tissues enclosing the oospores were decomposed, and at the end of from four to eight days their germination might be observed when placed in a drop of water. This method again corresponds with the ordinary processes by which the plant naturally decays on exposure to the influences of the atmosphere, and the oospores germinate under the favour of a shower of rain.
If the oospore, after the decay of the tissues, is isolated and placed in a drop of water, the brown investing membrane will be seen to rupture 132irregularly, and its contents (enclosed in a transparent inner membrane) issuing from the orifice. As in the case of the conidia, this body at first contains vacuoles, and is afterwards divided into polyhedric portions; these pass into zoospores, which congregate at the centre into a globular mass (Plate X. fig. 207). They afterwards separate, and for some minutes float about in the vesicle in which they were generated. Ultimately the membrane ruptures, and the zoospores swim about in water just as those produced from the conidia had done. The number contained in each oospore is considerable, and may be estimated at not less than one hundred.
The zoospores, whether produced from conidia or from oospores, appear to be the same. The movements of both in the water last from two to three hours; then they cease, the ciliæ disappear, and the zoospores remain at rest, taking meanwhile a globular form. Afterwards these spores (for having ceased all motion they are no longer zoospores) emit a thin tube from some portion of their surface, such tube attaining a length of from two to ten times that of the spore whence it proceeds. The extremity of these tubes swells and forms a kind of cell, into which the contents of the spore pass through the medium of the tube (Plate X. fig. 209).
Thus far, and thus far only, has Dr. de Bary been enabled to trace the development of the 133zoospores in a drop of water. Another series of experiments was instituted by this mycologist having especial reference to the parasitism of the “white rust.” He made numerous observations to ascertain whether the spores, or the germinating tubes, entered by the roots of growing plants, and satisfied himself that they did not. Plants of garden-cress, mustard, and shepherd’s-purse had their roots immersed in water impregnated with zoospores. After one or two days, though the surfaces of the roots were covered with zoospores that had emitted their germinating tubes in all directions, none had penetrated or showed the least tendency to penetrate the epidermis. Other plants were planted in flowerpots and watered at the roots with water charged with zoospores, and for two days the pots were left standing in the water similarly charged, then the plants were removed, cultivated in the ordinary manner, grew up healthy, and gave no signs of the white rust. Care had been taken that neither stems nor leaves should come in contact with water containing zoospores.
If a drop of water thus charged is placed on the surface of a living leaf of the shepherd’s-purse, for instance, and left at rest for a few hours and examined minutely at the end of that period, they will be found to have germinated. Let the epidermis be removed carefully and placed on a glass slide and submitted to the microscope. Many zoospores will be found to have produced from that 134point of their surface which is nearest to one of the stomata, or pores of the leaf, its slender tube, and to have thrust it through those openings, with the swollen extremity resting in the air-cavity situated beneath the pore. If many days, or even weeks, are allowed to pass, and the leaf is examined again, or another leaf similarly treated, and kept in a living and vigorous condition by remaining attached to the parent plant, still no further change or advance will be observed, the germs will appear fresh, and still in the same condition. Hence it is concluded that plants are not infected through the medium of their leaves.
If the cotyledons, or seed-leaves, are watered with similar impregnated water, a different result has been observed to take place. The germination of the tubes till their entrance at the stomata is the same; but, having entered, the swollen extremity elongates, becomes branched, and takes all the appearance of mycelium such as we at first described. If the infected plant endures through the winter, the mycelium endures with it, to recommence vegetating in the spring.
The experiments which Dr. de Bary performed were all upon plants of the common garden-cress. It will be unnecessary to repeat all the details of these, as given in the memoir recently published on the subject, but it will suffice to give a summary of results. In two series of plants cultivated at different periods from good seeds, one hundred and 135five plants which, had not received the water impregnated with zoospores upon their cotyledons vegetated without any indications of the parasite. Amongst the eighteen plants which were inoculated by watering the cotyledons, four only were not attacked by the parasite, fourteen bore the “white rust.” In six of these it did not extend beyond the cotyledons; in the others it also appeared on the stems and leaves.
From these experiments it may be deduced that plants are not infected by spores of the parasite entering at the roots, or by their leaves, but that inoculation takes place through the medium of the cotyledons, or seed-leaves; that the agents in this inoculation are the zoospores produced either from the conidia or the oospores; that they do not enter the stomata or pores themselves, but thrust out a germinating tube, into the extremity of which the contents of the zoospores pass; that when these tubes have entered the stomata of the cotyledons they branch and ramify, becoming a true mycelium, from which fruitful parasites are developed; that if a plant so infested lives through the winter, the parasite lives with it, to vegetate again in the spring.
The immense number of zoospores capable of being produced from a single infested plant is almost beyond calculation. It is easy for a million of conidia to be developed from such a plant, each producing from five to eight zoospores, besides a 136large number of oospores, each containing a hundred zoospores. It can scarcely be considered marvellous that the white rust should be so common on plants favourable to its development, the marvel being rather that any plant should escape.
Plate XII.
W. West imp.
Until recently it was doubtful whether more than one or two species of Cystopus (white rust) were known. It is now certain that we have three in Great Britain, and three or four others are found elsewhere. Of the British species one is found on many cruciferous plants, as the shepherd’s-purse, garden-cress, mustard, radish, and plants of the cabbage kind. This is the Cystopus candidus. Another occurs on the goat’s-beard, salsify, and scorzonera, which is called Cystopus cubicus. Both have great external resemblances, but both possess specific internal differences. In the Goat’s-beard rust (Plate X. fig. 201) the terminal conidia in the bunches or fascicles of conidia which are produced within the pustules are spheroidal, large, and of a yellow-brown tint, whilst the residue are cylindrical, smaller (Plate X. fig. 202), and more or less compressed. In the crucifer rust the conidia are all equal in the pustules and globose. The oospores in the former of these are subglobose and the warts on their surface are solid; whilst in the latter the oospores are truly globose, and the warts on the surface are hollow (Plate X. fig. 210). The third species is the Sandspurry white rust (Cystopus Lepigoni), which was found on the common sandspurry 137(Spergularia rubra) by Mr. R. G. Keeley, in Swanscombe Marshes (September, 1864). Of the other species it is not improbable that one or two may yet be found in this country. Without attempting to indicate their microscopic differences, it may be serviceable to name the species of phanerogamic plants on which they are likely to be found. The Purslane white rust (Cystopus Portulacæ, D. C.) should be sought on the purslane, which, though of limited cultivation, is exceedingly liable to attack from this parasite, and the Thistle white rust (Cystopus spinulosus, D. By.) may probably be met with on the leaves of the common thistle (Cnicus arvensis) or some of its allies.
Considerable interest is now attached to these parasites, which, as far as we at present know, differ materially in their reproduction from the other dust-like or uredinous fungi with which they have long been associated. Dr. de Bary proposes the union of these with the mould-like fungi of the genus Peronospora, to which the mould infesting the potato belongs, so as to constitute by themselves a group apart from the genera with which both have heretofore been associated. Whether his views will be accepted by mycologists time will speedily prove. Under any circumstances, microscopical and botanical science will reap considerable benefit from his researches.
TWENTY years since, and some of these little pests were altogether unknown, whilst others were only recognized and partly understood by a few scientific men. During the period to which we have alluded more than half the present species contained in the genus Peronospora had never been observed, and amongst these the most devastating of its tribe, the associate and undoubted cause of the potato disease.
Parasitic fungi are far more numerous, both in individuals and species, than most persons are aware, and cultivated plants of all kinds are more or less subject to their ravages. Some are more susceptible than others, of which the corn and grass tribe, or Graminaceæ, as they are termed by botanists, is an example. Not less than thirty species have been recorded upon plants of this natural order, and of these nearly one-half are found upon the living plants. Upon the potato plant, again, no less than ten different kinds of fungi have been described; whilst upon other and more fortunate plants only one or two parasites of this nature establish themselves.

262.—Turnip Mould.
Peronospora parasitica.

263.—Onion Mould.
Peronospora Schleideniana.
It will be sufficient for our present purpose to 139state that one of the six families into which fungi are divided for scientific purposes is called Hyphomycetes, a name compounded of two Greek words signifying “thread” and “mould,” or “fungus,” and is applied to this group because the thread-like filaments of which they are largely composed are the most prominent feature. In this family there are again a number of smaller groups called orders, having an equal value to the natural orders of flowering plants; and one of these orders, called Mucedines, has the fertile threads perfectly distinct from the mycelium or spawn. These threads are sometimes simple and sometimes branched; they may be articulated or without articulations or septa, short or long, erect or creeping, hyaline or whitish, mostly free from colour, and are not coated with a distinct membrane. The spores are generally simple, sometimes solitary, at others in pairs, or strung together like beads for a necklace. Amongst all this variety of arrangement there is order, for these are but features, or partly the features, of the different genera of which the Mucedines are composed. One of the genera is termed Peronospora, and to this the parasitic fungus of the potato, and some others to which we shall have occasion to refer, belong. In this genus the threads are generally branched, but without articulations. The spores, or seed-like bodies, are of two kinds; one kind is borne on the tips of the branches; and the other kind, which is larger and globose, is borne 140upon the creeping mycelium or spawn. All the members of this genus with which we are acquainted are parasitic on living plants, inducing in them speedy decay, but preceding that decay of which they are themselves the cause. Hence we have deemed it the more advantageous course both for writer and reader to associate together the different species of this particular genus of parasitic moulds in one chapter, rather than bring together the different kinds of fungi, belonging perhaps to widely separated genera, but all associated with, or parasitic upon, the same plant. The botanical student will thank us for following this plan, and the general reader will labour under no disadvantage, in this instance at least, from the similarity of the diseases produced in the plants infested.
It has been recently proposed to associate the genera Peronospora and Cystopus together in one group, under the name of Peronosporei; but with the discussion of this subject we have nothing to do in this volume. Having announced this fact, we shall continue to notice them in the several positions heretofore accorded to them.
The mycelium (root-like fibres) in this genus greatly resembles that described for the “white rusts,” though neither so thick, nor is the membrane so gelatinous as in that genus. In some instances the mycelium is confined to the inter-cellulary passages; but in most cases they also perforate the cells of the plant which nourishes 141them. The sucker-like bodies already described in Cystopus are often found produced on the mycelium of Peronospora, but occasionally they appear to be absent, especially in the mould causing the potato disease.
From the mycelium erect threads are produced, upon which one form of fruit, which may be termed “acrospores,” is borne. These filaments are sometimes single, and sometimes in small tufts or fascicles. In some instances they are considerably branched, so as to present a dendroidal or tree-like appearance; in others they are nearly simple, being only surmounted by short spicules; or, in one instance, quite simple, and only surmounted by a single acrospore. The branching habit is by far the most common.
Each ultimate branch in the ramification of the fertile filaments engenders a single acrospore. Its extremity, at first thin and pointed, swells in the form of a globular vesicle, which soon takes the elliptical or ovate shape of the perfected acrospore, and at length separates itself from the branch that supports it.
In all instances the acrospores have a similar structure, but with minute differences in form, &c., which have their importance in the determination of species. In most cases the apex of the acrospore is obtuse, and the entire body has a violet tint, more or less deep; in some it is completely colourless. These acrospores, when placed in favourable 142conditions, will germinate, and, in fact, comport themselves in the manner of true spores.
During the year 1861, Dr. de Bary published an account[8] of the discovery by him of zoospores, similar to those already described in connection with the conidia of the “white rusts,” produced from the acrospores of the mould which originates the potato disease. In the same author’s memoir of 1864, already quoted, the observations there made are confirmed. When the acrospores of the potato mould and the parsnip mould are sown in water upon a glass slide, their contents become divided, and vacuoles are formed, as already described in Cystopus; these parts are expelled through an apical orifice, and, when free, take the form of perfect zoospores, and commence swimming about in the fluid surrounding them.
8. Die gegenwartig herrschende Kartoffelkrankheit, ihre Ursache und ihre Verhütung. Von Dr. A. de Bary. Leipsig: 1861.
These zoospores are oval, or semi-oval, with a structure resembling that of the “white rusts,” save that the two ciliæ, or vibratile hairs, proceed from the same point. The number of zoospores from each acrospore of the potato mould is stated to be from six to sixteen, and in the parsnip mould from six to fourteen. Their ultimate development is the same as has already been described. In the potato mould, the production of the zoospores is much favoured by the exclusion of light.
143Another mode of germination in the acrospores of the potato mould has been observed by the same eminent mycologist. This results when the spores are sown upon a humid body, or on the surface of a drop of water. The acrospore emits from its summit a simple tube, the extremity of which swells into an oval vesicle; into this the contents pass, and it isolates itself by a partition from the germ-tube. Thus it becomes a duplicate of the acrospore from whence it was derived. This secondary body has also the power of producing a tertiary cellule in a similar manner. Both the second and third cellule, when immersed in water, produce zoospores in the ordinary manner, as above described.
Yet another and a third mode of germination is described by the same author, in which the acrospore emits from its apex a germ-tube, which elongates considerably; and into this long and tortuous tube the contents of the acrospore pass, and accumulate at the opposite extremity.
The germ-tubes, produced in the manner last described, when developed on the surface of a favourable plant, perforate the cells of the epidermis, or enter by the stomata. In the case of the potato mould, the germ-tubes enter by the stomata; but in the majority of species the germ-tubes do not enter by the natural pores of the leaves upon which the acrospores are sown, but perforate the cellules, and thus effect admission into the tissues of the 144plant, where they extend, ramify themselves, and become a mycelium. This mycelium originates branched threads, bearing acrospores at the tips of their branchlets, and in many species of Peronospora another kind of reproductive body upon the threads of the mycelium itself. To these bodies we must briefly address ourselves.
This last kind of reproductive organs (not yet positively found in the potato mould) appear to be wholly analogous to the oogonia of the “white rusts” already described, producing oospores in like manner. Dr. de Bary avows with regret that his numerous efforts to observe the germination of these oospores were unsuccessful. Nevertheless, he considers that the perfect resemblance between them and the oospores of the “white rusts” will justify him in concluding that the germination in both is very similar. It will be unnecessary to repeat here the observations already made on the growth and development of oogones and zoospores. What has been advanced respecting these organs in Cystopus will apply also to Peronospora.

264.—Potato Mould.
Peronospora infestans.

265.—Lettuce Mould.
Peronospora gangliformis.
Potato Mould.—Towards the close of the summer of 1845, in the course of a few weeks, every one became aware of the fact that a new disease had appeared which threatened the entire destruction of the potato crop. Until then it seemed to have been almost, although not entirely unknown. It first appeared in the Isle of Wight about the middle of August, and a week afterwards had become 145general in the South of England, and the next week there were but few sound samples of potatoes in the London market. Early in September the disease had commenced its ravages in Ireland, and shortly afterwards it was discovered in Scotland. With the same rapidity it seems to have spread throughout Europe and North America, or at least the western portion of the former and the northern districts of the latter. It must not be imagined, however, that the Isle of Wight was the centre from which this disease spread over such an extended area and with such alarming rapidity. From this spot it doubtless made its first appearance that year amongst our own crops, but there is not the least doubt of its existence both on the continent of Europe and in North America in the previous year, and the farmers of Belgium had noted its appearance in the province of Liège as far back as 1842 and 1843. Other diseases had been observed affecting the potato crop before, and one which was also associated with a parasitic fungus had made its appearance in 1815. It is also exceedingly probable that, in a milder form, the murrain was present with us a year or two before it broke out to such an alarming extent. A correspondent to the Gardeners’ Chronicle, in 1844, notices it in the Isle of Thanet, and another testifies to its occurrence in districts of Ireland for two or three years previous to its general outbreak. The description of the disease in Canada, in 1844, 146contained in a letter addressed to Dr. Bellingham, and quoted by the Rev. M. J. Berkeley,[9] leaves no doubt of its identity:—“During the months of July and August (1844), we had repeated and heavy showers, with oppressive heat, and an atmosphere strongly charged with electricity. Towards the close of the month of August I observed the leaves to be marked with black spots, as if ink had been sprinkled over them. They began to wither, emitting a peculiar, offensive odour; and before a fortnight the field, which had been singularly luxuriant, and almost rank, became arid and dried up, as if by a severe frost. I had the potatoes dug out during the month of September, when about two-thirds were either positively rotten, partially decayed and swarming with worms, or spotted with brownish-coloured patches, resembling flesh that had been frost-bitten. These parts were soft to the touch, and upon the decayed potatoes I observed a whitish substance like mould.”
9. Journal of Horticultural Society of London, vol. i. p. 11.
Although this disease made its first appearance, in the middle of August, 1845, in the Isle of Wight, it had already appeared in Belgium in the same year, a month previously; and although it may have been noticed in other British localities in 1844, it was known in Canada and in St. Helena in the same year to a far greater extent, and in Liège as early as 1842. There are, therefore, good 147grounds for believing that the European centre was Belgium; but if M. Boussingault was correct in stating that “this malady is well known in rainy years at Bogota, where the Indians live almost entirely on potatoes,” then it is not of European but American origin, and is probably derived from districts not far remote from those whence Europe first received the potato itself.
It would occupy too much space to detail the different theories and opinions relative to the causes of this disease to which 1845 and subsequent years gave birth. Suffice it to say, that the lapse of years has silently proved the majority of these to have been fallacious. All such as imputed to peculiar electric conditions, a wet season, or other meteorological influences, the disease which has re-appeared under different conditions and influences, and in seasons remarkable for dryness, are manifestly refuted; whilst its mycological origin has continued to gain adherents, and the gradual accumulation of fresh facts has almost placed it beyond dispute not only that the potato disease is accompanied by, but results from, fungal growth. Unfortunately, this disease has been so prevalent, more or less, during the past eighteen years, that few have been without the opportunity of making themselves acquainted with its external appearance. To this may be added the minute and exact account of its development, as recorded by that excellent mycologist and careful observer, 148the Rev. M. J. Berkeley, in 1846, and to which, even now, nothing of importance can be supplemented or abstracted:—“The leaves began suddenly to assume a paler, and at length a yellowish tint, exhibiting here and there discoloured spots. More or less coinciding with these spots, on the reverse of the leaves, appeared white mealy patches, consisting of a minute mould, proceeding, either singly or in fascicles, from the stomata, and arising from an abundant branched mycelium creeping in every direction through the loose tissue beneath the cuticle. The upper surface rarely, if ever, exhibits the mould, it being almost physically impossible for its delicate threads to penetrate the closely-packed cells which, being arranged side by side, leave scarcely any intercellular passages. The mould, in a few hours from its first piercing the apertures of the stomata, perfects its fruit, and in so doing completely exhausts the matrix, which in consequence withers. No sooner have a number of the leaves been attacked, than the stem itself is subject to change, becoming spotted here and there with dark brown patches, in which the cells are mostly filled with a dark grumous mass, without exhibiting any mucedinous filaments; though, occasionally, I have ascertained their presence. Very rarely fructifying but dwarfed specimens of the mould occur upon it. The stem now rapidly putrefies, the cuticle and its subjacent tissue become pulpy, and separate when touched from the woody 149parts beneath. The whole soon dries up, and in many instances exhibits in the centre the black, irregular fungoid masses which are known under the name of Sclerotium varium, and which are believed to be the mycelium of certain moulds in a high state of condensation.
“If the tubers are now examined, the greater part will often be found smaller than usual, especially if the disease has commenced at an early stage of growth; but in their natural condition, while here and there a tuber, particularly if it has been partially exposed, exhibits traces of disease. The surface is, however, soon marked with livid patches, commencing generally about the eyes, or at the point of connection with the fructifying shoots: these rapidly acquire a spotted appearance, the spots being rather waved, and assuming often a more or less concentric arrangement. Sometimes—especially on the smoother kinds of tuber—two or more regular systems of concentric spots are exhibited on the same tuber. The skin now withers, and is easily separated; the spots become depressed and of a yellowish tinge; and if the tubers be laid in a moist place, in a day or two—sometimes in a few hours—the same mould which destroyed the leaves springs from them, piercing the cuticle from within, yet not scattered, as on the leaves, but forming a conspicuous white tuft. If a section of the diseased tuber be made on the first symptoms of the disease, little brownish or rusty specks are 150found in the cellular tissue, confined, with very rare exceptions, to the space between the cuticle and the sac, if I may so call it, of spiral vessels and their accompanying tissue, which, springing from the subterranean branches, pass into the tuber, making their way to the several buds disposed on the surface. These spots consist at first of a quantity of discoloured cells, mixed more or less with others in a healthy condition. The grains of fecula are for a long time perfectly healthy; the cells themselves, so far from being looser, are more closely bound together than in the more healthy portions. The rusty spots soon exhibit a darker tint, spreading in every direction and becoming confluent; they at length extend beyond the barrier of vascular tissue, and attack the central mass. The tuber, meanwhile, assumes a disagreeable smell, decomposes more or less rapidly, other Fungi establish themselves on the surface, or in the decaying mass, which emits a highly fetid odour, resembling that of decaying agarics; the union of the cells is dissolved, animalcules or mites make their appearance, till at last the whole becomes a loathsome mass of putrescence.”
The form of the mould itself is represented (fig. 264) as exhibited under the microscope, with the nodose swellings of the branches, and spores attached to the tips. These acrospores are filled with a granular mass, from which, as hereafter described, zoospores are produced. The branching 151dendroidal threads of this fungus proceed from a creeping mycelium or spawn of entangled filaments which interpenetrates the matrix, upon which it establishes itself. Upon these threads spherical bodies were long since observed by Dr. Payen, and, under the name of Artotrogus, described by Dr. Montagne as a new species of fungus. Dr. de Bary failed to detect oospores on the mycelium of this mould, although the organs found by Payen in some sort resemble them. De Bary sought in vain, also, for the spherical bodies described by the author above named. It has been calculated that one square line of the under surface of the leaves is capable of producing 3,270 acrospores, each of which yields at least six zoospores, sometimes double that number; thus we have 19,620 reproductive bodies from that small space. The mycelium from the zoospores is capable of penetrating the cellular tissue in twelve hours, and, when established there, it bursts through the stomata of the leaves, and fruit is perfected in from fifteen to eighteen hours. Since the zoospores are perfected and ready to germinate in twenty-four hours from their being placed in water, it becomes almost impossible to calculate the myriads of fungi that may be produced from a single centre. Dr. de Bary has also demonstrated that the brown spots so characteristic of the disease are the result of the action of the spores or zoospores. By placing a quantity of spores in a drop of water 152on the leaves, stems, and tubers under a glass sufficiently air-tight to prevent evaporation, he produced the brown spots, and traced their progress from the earliest stages.
There are a few practical conclusions which may be drawn from these discoveries. In the first place, it is clearly shown by the production of the spots that the fungus is capable of causing the disease, a fact which has been disputed, but now placed beyond doubt. The inference is, that not only is it capable of producing, but is really the cause of the potato murrain. With bodies so minute and active as the zoospores, there can no longer be difficulty in accounting for their penetrating the tissues of the plant. They are most active and productive in wet weather, especially when it is also warm. Moisture appears to be essential, and a dry season the greatest enemy to the spread of the disease. That bodies so minute and subtle should have baffled all efforts to destroy or eradicate, is not now surprising. Whether any method will be found to contend successfully with it, is now more doubtful than ever. A careful reperusal of the old facts by the aid of this new light will tend to the elucidation of much of the mystery in which the subject has been involved. All who have hitherto been sceptical of the mycological source of one of the greatest pests of modern times should study M. de Bary’s pamphlet.

266.—Pea Mould.
Peronospora Viciæ.
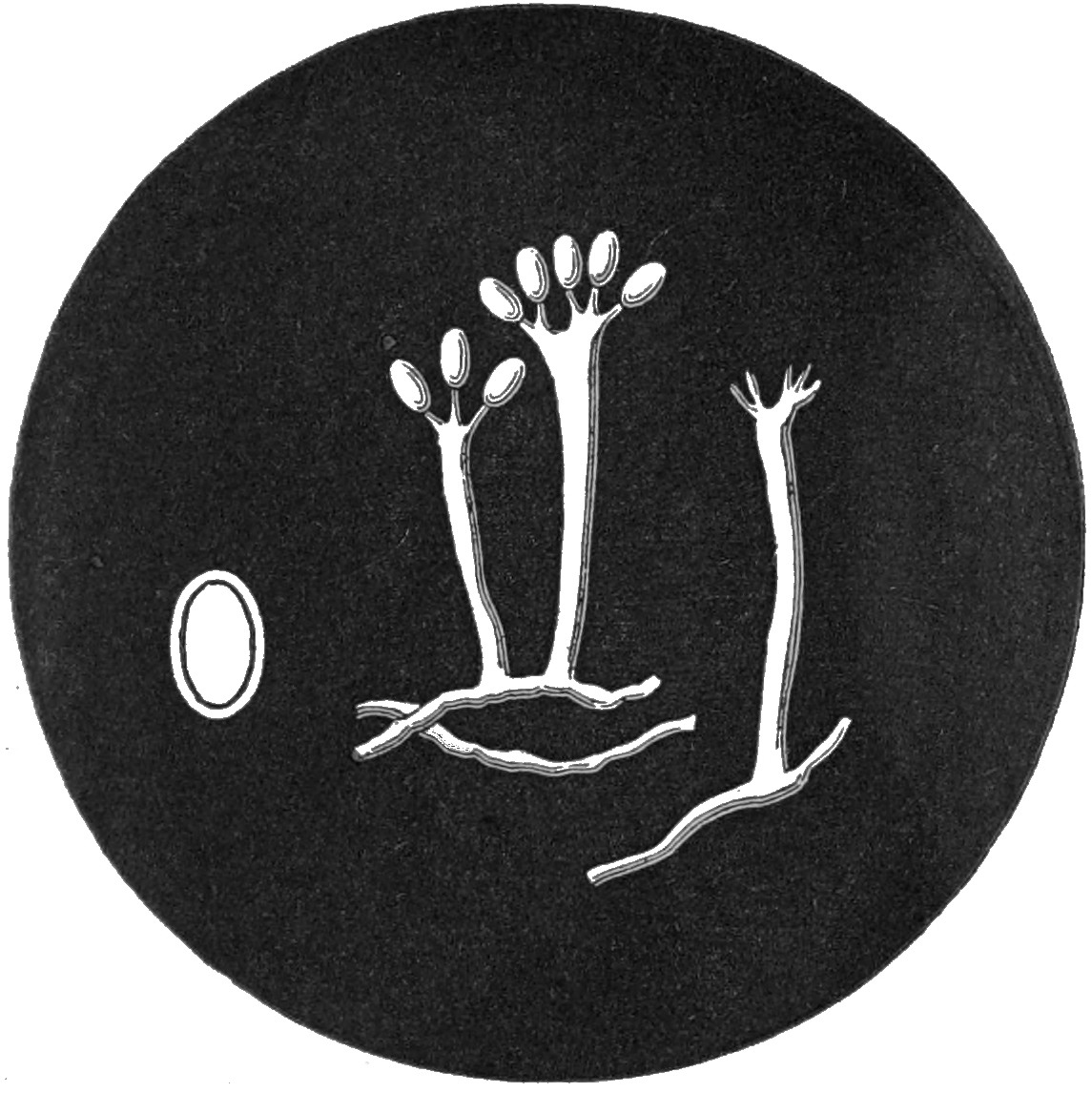
267.—Anemone Mould.
Peronospora pygmæa.
The potato mould has been judiciously named 153Peronospora infestans, or, as it was at first called, Botrytis infestans; but on a revision of the genera Botrytis and Peronospora, it was transferred to the latter genus, in which it remains. Three names were given to it, within a short period of each other, by different mycologists, in ignorance of its having already received a name. The one we have adopted appears to have the priority, at least of publication, and was given by Dr. Montagne. That of Botrytis devastatrix was given by Madame Libert, and Botrytis fallax by M. Desmazières. The principal feature in this species seems to consist in the branches becoming alternately thickened and constricted, so as to resemble a moniliform string or necklace of little bladders or vesicles. The branches are also more erect than in the allied species, and the spores are solitary on the tips or from the sides of the branches, and not in pairs or clusters, and the tips are simple, and not bifid or trifid, as in most of its allies. It need scarcely be remarked, that a high power of the microscope is necessary to make out the distinctive features of the different members of this genus, and that to the naked eye they only appear as a minute whitish mould. As already stated, this little fungus makes its first appearance on the under surface of the leaves, especially the lower ones, of the potato plant, and afterwards attacks the stem, and ultimately the tuber. For examination it is better to select the leaves soon after the fungus makes its appearance.
154Turnip Mould.—Since the advent of the potato murrain a similar disease has been witnessed, though more limited in its extent, amongst Swedish turnips, commencing in little waved irregular lines following the course of the vessels, around which spots are formed by the deposition of dark granules in the same manner as in the potato. In this instance, the leaves apparently are first attacked in a similar manner by a species of mould or Peronospora allied to the one already described, but which has been long known as parasitic upon cruciferous plants, to which the turnip belongs. This species, termed Peronospora parasitica, is white in all stages of its growth. It is much more branched, and the branches are comparatively shorter than in the potato mould, and the tips of the branches are bifid (fig. 262). The acrospores are very large and globose, features also which distinguish this mould from the last. A short time since we were called to witness a bed of splendid cauliflowers, which had, up to that time, been the pride of their cultivator; but, alas! their glory was threatened with speedy annihilation, for in nearly every instance the lower leaves had become more or less covered on their upper surface with yellow spots, and beneath glaucous with the mould we have been describing. The diseased leaves were all immediately removed, but we fear without success, although no positive information has since reached us. The almost unnatural 155vigorous green of the leaves, prior to the appearance of the mould, is not at all an uncommon occurrence: this phenomenon has been noticed in the ears of corn, in which every grain was soon afterwards filled with spores of bunt.
This species was at one time believed to be parasitic on the “white rust,” from which circumstance its specific name was derived. More precise examination proves that it sometimes occurs where no white rust is present, and therefore its parasitism is imaginary. The suckers in this species are large and penetrate the cells, often entirely filling them. This feature is sufficient to distinguish the mycelium of the mould from that of the “rust.” The turnip mould occurs on many cruciferous plants, and especially on the shepherd’s-purse, as well as upon the different varieties of cabbage and its more immediate allies.
Onion Mould.—Another disease, produced by fungi of the same genus, makes its appearance upon young onion plants in the spring. The mould is called Peronospora Schleideniana, and has many features in common with those already described. In this instance the threads are greyish and erect, with alternate branches, not divided by transverse septa, and the spores are obovate, attenuated towards their base (fig. 263). This mould, in some years, is very common and destructive, by preventing the young plants which are attacked from coming to perfection. It is not confined to the 156onion, but appears on other allied species of Allium (to which the onion belongs). The threads form large patches or blotches on the leaves, and sometimes cover them entirely. It very much resembles the turnip mould, from which the form of the spores considerably differs. This is the same species as that described by Caspary, and afterwards by Berkeley, under the name of P. destructor.
Lettuce Mould.—A very similar mould (Peronospora gangliformis) is sometimes very common in spring on the under surface of the leaves of the cultivated lettuce, appearing in definite white mouldy spots. By reference to the figure of a portion of a thread magnified (fig. 265), it will be seen that the peculiar form of the tips of the branchlets evidences the distinctness of this species.
The oospores are small, globose, and of a yellowish-tawny colour. This mould is by no means confined to lettuces, but has also been found on species of ragwort, sow-thistle, nipplewort, endive, and other composite plants; and has from time to time received numerous names, which it is unnecessary to enumerate.
Tare Mould.—The under surface of the leaves of tares, and sometimes also of peas, is liable to attack from an allied species of mould (Peronospora Viciæ). In the spring of 1846 it appeared amongst vetches in some districts to such an extent as at one time to threaten the destruction of the crops; but a succession of dry weather at once abridged 157its power and limited its mischief. Mouldy vetches and mouldy peas are, especially in moist seasons, evils to which the agriculturist knows his crops to be subject; he may not know, however, that this kind of mould (fig. 266) is of so near a kin to that which has acquired such wide-spread fame in connection with the potato. Another species of fungus attacks the garden pea in damp seasons, forming small depressed brownish spots on the leaves and pods; but this is quite distinct from the mould, though probably not less injurious.
The fertile threads are produced in dense clusters, each many times branched, and bearing elliptic acrospores obtuse at their apices, and of a violaceous tint (fig. 266). The oospores are beautifully reticulated and of a yellowish-brown colour (Plate X. fig. 212).
Trefoil and some other allied plants are attacked by another species, characterized by Dr. de Bary as Peronospora trifoliorum, which we have found rather plentifully in some localities on lucern.
The Parsnip Mould (Peronospora nivea, Ung.) is found on many umbelliferous plants; but its attacks upon the parsnip are most to be deplored, because it injures and ultimately destroys an article of human food. The plants infested with this parasite are first attacked in the leaves, but afterwards the roots become spotted and diseased in a similar manner to the potatoes attacked by its congener. The disease has not hitherto been so 158general with the former as the latter; but in some districts it has been far from uncommon.
The fertile threads are collected in bundles, erect, and not so much branched as in many other species. The acrospores are subglobose or ovoid, and papillate at their apices. This species is sometimes called P. umbelliferarum, and sometimes P. macrospora. Generally speaking the average humidity of a season but little affects the production of parasitic fungi. In a dry season, like that of 1864, we found as many species, and these as flourishing and numerous in individuals, as in a proverbially wet year. Such is not the case, however, with the moulds under notice, or such fungi as are reproduced through the medium of zoospores: these are undoubtedly less common in a very dry season; but it must be remembered that a single shower is sufficient for the development of zoospores, and occasional showers or heavy dews will speed them on their course of destruction as readily almost as continuous moisture. The large fungi, on the contrary, become very limited in numbers when the weather is unusually dry.
Spinach Mould.—Spinach is likewise liable to suffer from the establishment of a mould upon the under surface of the leaves: unfortunately this is not unfrequent, and has been known in England certainly for the last fifty or sixty years, since it was figured by Sowerby in his “British Fungi” as many years since. We have lately seen a bed 159of spinach, utterly destroyed by this fungus; whilst on another, not twenty yards apart, not a spotted leaf could be found. This mould is the Peronospora effusa of botanists; it occurs also on some species of goosefoot (Plate X. fig. 215), and probably on knotgrass. To the naked eye it appears in pale purplish-grey patches, which, when examined microscopically, are found to consist of dense bundles of branched threads, bearing ellipsoid acrospores, the membranes of which have a violaceous tint. The oogonia produced upon the mycelium vary considerably in size. The oospores are of the character delineated in our plate (Plate X. fig. 214).
Hitherto all the species of mould to which we have had occasion to refer have been found infesting plants more or less employed as food; but there remain one or two other species to which we must make special reference. One of these affects the most universal of favourites amongst flowers: this is the rose mould. Attention was directed to this mould, and it was described for the first time under the name of Peronospora sparsa, in the columns of the Gardeners’ Chronicle, in 1862. It occurred on a quantity of potted rose-plants in a conservatory. Irregular pale brownish discoloured spots appeared on the upper surface of the leaves; these extended rapidly, and in a short time the leaves withered and shrivelled up, and ultimately the whole plant perished. A delicate greyish mould 160was to be seen by the aid of a lens, scattered over the under surface of the leaves. By the microscope, the branched threads, having the tips furnished with subelliptic spores, were revealed, and an ally of the potato mould found revelling amongst the roses.
During the winter of 1863-4, we found the leaves of several species of dock occupied by a mould which appears to be a very low form of Peronospora. Its presence was indicated by brownish orbicular spots, on which the fertile threads occurred in small bundles. These threads were generally simple, but occasionally forked, bearing rather large elliptical acrospores attached obliquely to the tips of the threads (fig. 269). In consequence of this peculiarity, we have named the species, which does not appear to have been noticed before, Peronospora obliqua. It is clearly very distinct from another species found on dock leaves by Corda.
Of the remaining British species, one (P. Arenariæ) is found on the leaves of the three-veined sandwort (fig. 268); another attacks the red corn-poppy, a third is found on the common nettle, one on the brooklime, another on the wood-anemone (fig. 267), and another on the figwort.
Doubtless all the species in this genus are possessed of the third means of reproduction, by zoospores, as discovered in the potato mould, not only from the acrospores, but also from the oospores.

268.—Sandwort Mould.
Peronospora Arenariæ.
269.—Dock Mould.
Peronospora obliqua.
161The fearful rapidity with which this method enables them to multiply themselves may account for their widely spreading and devastating power. No other genus of fungi can parallel this in the number of species injurious to the field or the garden, or in which the injuries inflicted are so great and irremediable.
N.B.—Since the foregoing chapter was in type, the Rev. M. J. Berkeley informs us that both Mr. Broome and himself have examined the mould on dock-leaves, to which we have given the name of Peronospora obliqua, and have come to the conclusion that it is truly a member of that genus, and not hitherto described; but they are also of opinion that it is the same mould as one described by Dr. Montagne as Ascomyces Rumicis. We concur with them in thinking it deficient in the important characteristics of Ascomyces, and therefore retain its proposed name of P. obliqua.
NOTWITHSTANDING the inconvenience to ourselves of calling very different fungi by the same common name of “mildew,” the popular mind does not recognize the inconvenience, since it scarcely troubles itself to inquire whether they are not all the same thing. In obedience to this custom, we again write of “mildew,” or “blight,” as it is called in some districts, but of a very different kind to that which is so detrimental to growing crops of corn. In the present instance it is our intention to illustrate a group of fungi which are exceedingly common, and which differ greatly in appearance and structure from any to which we have had occasion to allude. To obtain a general knowledge of these forms let our reader proceed at once to a clump of rank grass; if it is his fortune to dwell in the country, the walk of a few yards will suffice. Let him examine this clump more carefully, perhaps, than he has been accustomed to do, and we venture to predict that he will find some of the leaves covered with what appears to be a dirty white mould, or mildew (Plate XI. fig. 235). One of these leaves should be collected as carefully and conveyed to the microscope 163as speedily as possible, taking care not to touch, or brush it against any other object so as to disturb the arrangement of the delicate little threads upon its surface. If a small portion, say about an inch, is cut from this leaf with a sharp pair of scissors, and laid upon a slide, or pinned down upon a strip of sheet cork, so as to keep it flat, and then submitted to examination under the microscope, with an inch power, a beautiful forest of crystalline vegetation will be observed. If the examiner on this occasion should not possess a binocular microscope we are sorry for him, because in that case he will not see all that is to be seen under the greatest advantages. If we ever truly enjoy looking through such an instrument, it is on an occasion like this, when a low power is all that is needed, and the object is required to be seen in relief. It is scarcely possible to convey an adequate idea of the beauty of such a scene as the microscope reveals upon this fragment of grass-leaf. Little bundles of delicate threads, clear and crystalline, are seated upon a slender branching mycelium. These threads, sometimes erect, sometimes drooping, flexuous, or prostrate, are composed of numerous roundish or spherical cells attached to each other in a moniliform or bead-like manner (fig. 236). These easily separate from each other. Let a portion of the threads be removed from the leaf on the point of a lancet and laid upon a glass slide, with a thin cover over them. Submit this object to a quarter-inch 164power, as a drop of water is let fall at the edge of the cover and insinuates itself, by capillary attraction, between the two plates of glass. So soon as it touches the moniliform threads, the disunion commences, and almost before they are enveloped in the fluid, two spherules will scarce remain attached to each other. This delicate little mould on the grass leaf at one time bore the name of Oidium monilioides. It is now regarded only as a condition of another minute fungus, to which attention will shortly be directed.
The vine disease, so fearfully destructive on the Continent, and not altogether unknown in this country, is another of these incomplete fungi. From an individual who at the time of its first discovery in the south of England took considerable interest in the subject, it was called Oidium Tuckeri, which name it continued to bear, both here and abroad, until, with many others, probably nearly all of the same genus, it was found to be only a barren state of what is called by mycologists an Erysiphe. The real discoverer of this mildew was undoubtedly the Rev. M. J. Berkeley, who has successfully devoted a long life to the study of these minute organisms, through evil and through good report, and when that study was beset with more difficulties, and received less encouragement than at present. If, towards the autumn, we should again collect some whitened, mouldy, or mildewed grass-leaves, similar in appearance to those mentioned 165above, and carefully look at them with a pocket lens, little black points, almost as small as a pin-point, or more resembling the full stop with which, this sentence closes, will be found scattered over the white threads. The aid of the microscope must be again sought to make out the structure of the little black dots. Closely nestling upon the mycelium, the little points will prove to be spherical brownish, conceptacles, surrounded with transparent floccose appendages. Many other species are far more beautiful than that of the grass-leaf, as will be seen by reference to our plate. The variation consists chiefly in the form of the appendages which spring from the conceptacle and surround it in a radiating (as in figs. 219, 222, 225, and 230), or in a more or less confused and entangled manner (as in figs. 216, 240, 245, and 251). The surface of the conceptacle is minutely reticulated, and its base is attached to the mycelium. When first formed, these globose conceptacles are almost colourless; they afterwards acquire a yellow colour, and are ultimately of a deep brown. The appendages are seldom at all coloured. Within the conceptacle are contained from one to several transparent obovoid sacs, or spore-cases, called sporangia, enclosing a definite number of spores (figs. 218, 224, 228, &c.), which vary in different species. In the hazel mildew, for instance, there are two spores in each sporangium; in the willow mildew four; in the maple mildew 166eight; in the grass mildew, and some others, numerous. The tips of the appendages are variable, and often elegant (figs. 227, 231, 233, 234, and 247), sometimes simple and at others symmetrically branched. All the species occur on the still living and green parts of plants, especially the leaves, and are therefore truly parasitic. A pocket lens will show whether any conceptacles are present on any suspicious leaf which may be collected, but high powers of the microscope are essential for their complete examination. It is during autumn, when vegetation begins to languish, that we shall be most successful in searching for specimens. They will then be found almost everywhere, and the white mycelium forms an object too conspicuous for them to be readily overlooked. Botanically, nearly all the species were at one period included in one genus, under the name of Erysiphe, a name derived from the Greek, and signifying “mildew;” at the present time they are distributed through several genera, the chief distinctions of which are based upon the form of the appendages. Though personally disposed to question the generic value of such distinctions, it would be imprudent to adopt any other names here than those to be found in recent English works on fungi.
The first species in our enumeration is found on cultivated roses. What a deplorable picture does a favourite rose-bush present when attacked by this mildew! The leaves blistered, puckered, and contorted; 167their petioles and the peduncles and calyces of the flowers swollen, distorted, and grey with mould; and the whole plant looking so diseased and leprous that it needs no mycologist to tell that the rose is mildewed. The conceptacle in this species is minute, and contains but one sporangium, which is one of the characters of the genus in which it is now included, and a more justifiable distinction than the ramifications of the appendages. The mycelium is rather profuse, and the threads or appendages which spring from the conceptacle are simple and floccose (fig. 216). The sporangium contains eight ovate spores. This species (Sphærotheca pannosa, Lev.), in its oidioid or conidiiferous form, was for some time known under the name of Oidium leucoconium.
An allied species constitutes the hop-mildew, a visitation with which some of our Kentish friends are too familiar. This is not a prejudiced species in the choice of its habitation, since it is found on many other plants, where it flourishes with equal vigour. The meadow-sweet, burnet, scabious, teasle, dandelion, and other composite plants, plantain, and plants of the cucumber family, all suffer more or less from its roving disposition. The mycelium of whitish threads is even more conspicuous than in the last species, but the conceptacles are often not to be found at all. These are also very minute and most common on the under surface of the leaves. The appendages, or fulcra, are simple, 168floccose (fig. 217), and coloured. The sporangia are found singly in each conceptacle, and each, sporangium contains eight spores.
An autumnal stroll amongst hazel-bushes, when the nuts are ripe, will lead, if the nuts are not a greater attraction, to the discovery of whitish, patches on the under surface of the leaves, caused by the mycelium of the hazel mildew (Phyllactinia guttata, Lev.). These patches are less distinct and conspicuous than in many other species, but the little blackish dots of the conceptacles may be distinguished by sharp eyes without the use of the lens. Though possessing a decided preference for the hazel, this species is also found on the green leaves of the hawthorn, ash, elm, birch, sallow, beech, oak, and hornbeam. The conceptacles are larger than in the two preceding species, and somewhat depressed above. The appendages are few (fig. 219), radiating, rigid, and acicular, or like needles. Each conceptacle contains eight or more sporangia, and each sporangium has from two to four spores (fig. 220). This species being very common, its conceptacles large, and produced copiously, and its appendages distinct, it will prove a good type with which the student of these fungi may commence his examinations. This is the only representative which we possess of the genus established by M. Leveille for such of the Erysiphei as have the conceptacle depressed, and the appendages rigid and simple; by which 169features it is distinguished from genuine species of Erysiphe.
Two species, also common, having many features agreeing with each other, are found on the leaves of the maple and the willow. The willow blight (Uncinula adunca, Lev.) is found irrespectively on various species of poplar and willow (fig. 221). In size and external appearances, to the unaided eye, it seems scarcely to differ from the preceding, but more minute examination will show that in the appendages there is an appreciable difference. Still rigid, but no longer aciculate, the tips bent or curved like a little hook, or curled upon themselves (fig. 223), radiating and numerous (fig. 222), and at length tending upwards. Many sporangia are contained within each conceptacle, each of which is furnished with four spores. The amateur must not be disappointed, if, on examining mature conceptacles with a view to the discovery of the sporangia, he finds only free spores. The investing membrane is very delicate, and disappears generally as the spores are matured.
The “blight” or “mildew” which occurs on the common hedge-maple, as well as on sycamore leaves, is exceedingly conspicuous when occurring on the former plant. The whole bush often presents a hoary appearance as if sprinkled with powdered chalk. In the spring, the under surface of the leaves of the same plant are liable to become hoary from another cause. The whiteness occurs 170in patches, has often a pinkish or violaceous tint, and glistens like hoar-frost. This affection of the leaves was, at one time, believed to be produced by a fungus which was called Erineum acerinum, but now it is regarded as a diseased state of the tissues. In the maple mildew, both surfaces of the leaves are alike affected, and the little, dark, point-like conceptacles will be found studded over both. It is not uncommon to meet with very white leaves, caused by the mycelium, but which bear no fruit. The appendages in this species are shorter than in the last (fig. 225), and the tips are bifid (fig. 226), or divided into two short branches, each of which is bifid, and uncinate or hook-shaped (fig. 227). The conceptacles contain not less than eight sporangia, each of which encloses eight spores.
Amongst the parasites that prey upon the much abused berberry (which has been charged in turn with producing the mildew in corn), is one which causes the green leaves to assume a chalky appearance (fig. 229), though less conspicuously than in the maple blight. This parasite is the berberry mildew (Microsphæria berberidis, Lev.). In such localities as the writer has met with the berberry suffering from mildew, he has invariably found a larger proportion of leaves with the barren mycelium than of those on which the conceptacles were developed. Perhaps in other localities this may not be the case. The appendages, as will be seen on reference to our plate, differ materially from any of 171those to which we have referred; indeed, this genus (or sub-genus) has the most elaborate and beautiful forms in these appendages of any of the Erysiphei. A figure is given of the tip of a fulcrum from a continental species (M. Ehrenbergii, Lev.), not yet found in this country (fig. 233). In the berberry blight the appendages are straight at the base, but afterwards become forked, each fork being again forked, and these yet again branched in a similar manner (fig. 230); so that a complex dichotomous tip is formed to each of the appendages (fig. 231). Each conceptacle contains about six sporangia, and each sporangium contains from six to eight spores (fig. 232).
The common gooseberry is also liable to a visitation from an allied species, in many respects closely similar, but differing in having the tips of the appendages more branched, and the extremities of the ultimate branchlets are not entire and attenuated, as in the berberry mildew; but divided into two toothlike processes. The conceptacles in this species contain from four to eight sporangia, each of which has four or five spores.
In England, the leaves of the guelder-rose, and in France (perhaps also in this country) those of the alder, nourish a parasite belonging to this division. This “blight” possesses so much in common with others to which allusion has been made, that it will scarcely be necessary to describe it in detail. A figure of the tip of one of the appendages of the 172variety found on the alder is given in the Plate XI. fig. 234.
We have found another species which had not been before noticed in this country (M. Hedwigii, Lev.), on the leaves of the mealy guelder-rose in the vicinity of Darenth Wood, near Dartford, in Kent. The mealy character of the leaves of this plant, and the minute size of the conceptacles of the parasite, render it difficult to find; indeed, it could not be noticed unless it were sought for, as we sought it, lens in hand. It only occurs on the under surface of the leaves: the mycelium is very web-like and fugacious, the conceptacles minute, globose, and scattered (fig. 243). Four sporangia, each containing but four spores (fig. 244), are enclosed in each conceptacle, which is surrounded by a few appendages (about six) thrice dichotomous, and thickened at the tips of the ultimate branches, which are incurved (fig. 247).
The species of true Erysiphe are distinguished botanically from the foregoing by the floccose character of the appendages, in which feature they accord with the species found on the rose and the hop, but from which they differ in the conceptacles containing numerous sporangia instead of only one, as in those species.
One of the most common and conspicuous of these is found on the leaves and leaf-like stipules of the garden pea. Every leaf in a crop will sometimes suffer, and the gardener, to his great mortification, 173finds that the mildew is more prolific than his peas. The leaves become sickly and yellow as the mycelium of the fungus spreads over them, when they present a peculiar appearance, as if growing beside a chalky road in dry dusty weather, and had become covered with comminuted chalk. Soon the conceptacles appear, profusely scattered over the white threads, like grains of gunpowder (fig. 237), and after a brief struggle for existence the pea and its parasite die together. In this species (Erysiphe Martii, Lev.), the appendages are nearly transparent, short, and much interwoven with the mycelium (fig. 238), the globose sporangia containing from four to eight spores (fig. 239). It is not confined to peas, although that habitat has been here given for it, because it is so common upon them. Beans, melilot, St. John’s-wort, some umbelliferous plants, and the meadow-sweet, have all been found affected.
The species found on grasses, especially the cocksfoot, has been already alluded to. The conceptacles contain from twenty to twenty-four ovate sporangia, each enclosing eight spores. The appendages and mycelium are much interwoven.
Another of these “white mildews,” not only on account of its frequency of occurrence on certain plants, but also from the numerous species of phanerogamous plants on which it is found (fig. 240), may be truly designated “common” (Erysiphe communis, Lk.); many kinds of crow-foot, especially Ranunculus acris, are subject to its 174parasitism. It is found also on other plants of the same natural order, on the rest-harrow, trefoils, enchanter’s nightshade, bindweed, and knotgrass. There are from four to eight sporangia in each conceptacle, containing from four to eight spores (fig. 241). In this species, more especially, M. Tulasne found curious sucker-like processes developed on the threads of the mycelium (fig. 242): their office may probably be only that of attachment.
Of the other species found in Britain an enumeration will suffice, since they contain no feature of interest to the microscopist; and all the members of this section are far less beautiful than those in other genera (especially Microsphæria).
The leaves of the dogwood or cornel (figs. 245, 246) are the home of one species (E. tortilis, Lk.), and the burdock of another (E. Montagnei, Lev.). Both of these, in addition to the above, have sporangia which contain more than two spores. There are also two species in which only two spores are contained in each sporidium. One of these (E. Linkii, Lev.) is found on both surfaces of the leaves of the mugwort (figs. 248, 249); the other (E. lamprocarpa, Lev.) occurs on salsafy, scorzonera, weasel-snout, and plantain (figs. 250, 251).
These complete the Erysiphei; but there are allied species of too much interest not to be noticed in connection with them. Three very singular fungi are found on damp straw and paper; two on the former and one on the latter. Of the species 175occurring on straw, the most common one is figured, natural size, in our plate (fig. 257); but from this no idea can be formed of its structure, which in some points resembles an Erysiphe. The conceptacles are thin and brittle, and are clothed externally with long dark-coloured branched hairs (figs. 258, 259). The conceptacle contains long narrow sporangia, each enclosing dark, almost black, lemon-shaped sporidia. For low powers this is a very interesting object. The minute structure affords no feature of popular interest. This fungus (which bears the name of Chætomium elatum) is common on old straw, thatch, reeds, matting, &c., resembling small brown tufts of hair, visible to the naked eye.
Paper much exposed to damp will occasionally develop a similar “bristle-mould,” surrounded by a yellowish spot (Chætomium chartarum, Ehrb.); but it is not so common as the last. In habit and structure it is very similar (figs. 252, 253).
In 1838, the Rev. M. J. Berkeley announced the discovery by him of a singular production, for which he was unable to find a fitting location in any genus then established, and for which he accordingly characterized a new one, under the name of Ascotricha. This new species of paper mildew was found by him on some printed paper in a box. It somewhat resembles the other species above alluded to, at a casual glance; but more minute examination will reveal its differences. The author 176to whom we are indebted for this species thus describes its development. At first it appears as a minute branched mould interspersed with globose brownish conidia. As it advances in growth, globose black peridia become visible amongst the threads, clothed with and supported by alternately branched obscurely-jointed filaments, the branches of which generally form an acute angle with the stem (fig. 254). The ramification of these is very peculiar, the stem and main shaft of each sub-division being almost constantly shortened and surmounted by the branches given off near its apex; this, again, is often abbreviated and another branchlet given off, which again surpasses it; and occasionally the same circumstance takes place a third time. The apices are clavate and colourless; the rest of the filaments, when viewed by transmitted light, brown, even, and pellucid: a few globose conidia are usually attached to them (fig. 255). The conceptacle is thin, black to the naked eye, of an olive-brown under the microscope, filled with a mass of linear extremely transparent asci (fig. 256), each containing a single row of broadly elliptic chocolate sporidia. These have a paler border; sometimes the colour entirely vanishes, either from age or abortion, and there is only a minute globose nucleus or more probably a vesicle of air, in the centre; occasionally they become so transparent that the globular bodies alone are visible. After the conceptacles burst, several are frequently collected 177together into an irregular linear body, which consists principally of the conglomerated sporidia.
One other very common and troublesome little fungus (Eurotium herbariorum) will for the present close our examples. This is found creeping over dried plants preserved in herbaria, on decaying fruit, preserves, and various other substances, sometimes animal as well as vegetable, but chiefly the latter. To the naked eye it appears as a myriad of little yellow spherical bodies, of the size of very small pins’ heads, resting upon fine cobweb-like threads (fig. 260). When magnified, the surface of the conceptacles is seen to be reticulated (fig. 261). In the interior the sporidia are borne, contained also, as in the former instance, in asci. It has been considered probable, but as yet not fully proved, that this mildew is a compound fruited (ascigerous) condition of an equally common mould (Aspergillus).
Dr. Shortt, of Chingleput, in a recent report on the growth and production of Indian Cotton, remarks that the plants are subject to the attacks of a kind of mildew. He writes:—“They appear in the form of rounded fibres or thallus, shooting up in the air, having the lamina of the leaf as a base, and feeling villous to the touch. The small fibrillæ that form the nap appear shooting up as sharp projections when seen by the naked eye; under the microscope they are found to consist of pointed tubes, interspersed here 178and there with minute granular cells. It first attacks either the upper surface of the petioles, or the margins of the leaf, gradually extending over the lamina, and matting together the whole leaf into a greyish-white, felty mass. At first it attacks the young shoots and tender leaves, preventing them from expanding. The extension of the parasite deprives the plant of its juices, and eventually either destroys or renders it sterile. The spores seem to be derived from the atmosphere, and finding the plant in a state fit to receive them, from either the results of excessive cultivation, or from the effects of heat and want of moisture rendering it unhealthy, and thus favouring the reception of the spores of the fungi. Another variety speckles the leaves with whitish dots. These remain separate, but the lamina is covered with them, and in time the leaf changes colour, becomes yellowish, and eventually dies away. This is evidently the disease called Bunt, or some variety of it, as it is seated beneath the epidermis, and eventually the spores escape. Under the microscope they seem to consist of small dark cells or spots attached to a thread-like mycelium.”
The writer seems certainly to have made a mistake in its affinities, and on the faith of the above quotation we should be more disposed to regard it as an Erysiphe. It is to be hoped that specimens of the affected leaves will be forwarded to this country for examination.
IF, in offering a few practical suggestions, we either repeat ourselves, or communicate common-place hints, those who may know already all we shall essay to tell them will please to pardon and pass on.
All the information essential under this head relates to collecting, examining, and preserving microscopic fungi.
Collecting does not differ, except in the objects themselves, from any other botanical collecting. Those who attempt it must be prepared to sacrifice their kid gloves and patent-leather boots, to put on waterproofs and perseverance, and come home sometimes disappointed. The requisites for good work are but few, and easily supplied. A strong knife, a pocket lens, and a box or leather bag, will be all that is really essential. But where shall we go—and when? Hedge-banks, the sides of ditches, borders of woods, anywhere, if the plants are to be found on which the fungi are parasitic. We flatter ourselves on being rather successful in collecting, and our favourite localities have always been the dampest places in woods, railway-banks, and waste places. It is a great mistake to endeavour 180to go over a large tract of ground. We have spent a whole day in a little chalkpit, which had fallen into disuse, and grown wild. Fifty yards into a wood is as much, as we attempt, when alone; and a spot six yards square has afforded us occupation for hours. It is better to examine a small space thoroughly than to scamper on, mile after mile, and find nothing.
When? is as much to be noted as where! All the year round we shall be sure of finding something of interest. As soon as the last patch of winter’s snow has melted from the ground, and green leaves begin to unfold themselves, the search may begin. Cluster-cups (Æcidium) will be the earliest forms encountered. On the leaves of Lapsana communis, and the pilewort, these will be found before the majority of plants have burst their buds. Henceforth, other forms will gradually appear, until May or June. One or two species of Puccinia will be seen in April or May, but from that period until autumn, species of Trichobasis will be common. In June and July the smuts belonging to the genus Ustilago are most plentiful, and from August to October Puccinia and its allied genera will have the ascendant; so that from March to October there is continually succeeding each other some species of parasitic fungus belonging to the Uredines. From October to March need be no more a season of repose from the search of these minute plants than from March to October. 181So long as the ground is not covered with snow there will always be something to find on dead leaves, rotten sticks, &c., when there is not a green leaf to be seen. But these belong to a section to which we have studiously avoided all reference in the foregoing pages.
General instructions will not always apply; but in most instances, the lowest and earliest leaves, in which vitality appears to decline, will be most likely to suffer from the attacks of fungi. This rule must not be too stringently applied; the species of Æcidium, for instance, will generally be found on vigorous green leaves.
Having found a plant infected with some rust or brand, and by means of a pocket lens assured yourself that it is such, although the power is insufficient to tell what it is, collect as many leaves as you are likely to require; place them flat one upon the other, to prevent their curling up at the edges, should the weather be hot, and yourself far from home, and lay them in your box; or if you should take in preference an old book with stiff covers, place them separately between the leaves of your book, and they will be in still better condition, if you desire to preserve them. Arrived at home with the results of your trip, proceed at once to lay them between folds of blotting-paper, submit them to a gentle pressure, and change the papers daily until your leaves are dry, not forgetting to keep a scrap of paper with each collection, stating 182date and locality, to which, after microscopical examination, the name may be added. When thoroughly dry, your leaves may be preserved for reference in old envelopes, with the particulars endorsed on the outside. Fungi on leaves will generally be examined to the greatest advantage in the fresh state, but if too much pressure is not employed in the drying, it will not be difficult even in that condition to make out their characteristic features. Care must be taken, by changing their position, that moulds of other kinds do not establish themselves upon the specimens in drying, or that when dried they do not fall a prey to Eurotium herbariorum.
If it is intended to add these leaves to your herbarium, or to form a special herbarium for them, they should be mounted on white paper, first by affixing one or two leaves by means of thin glue to a paper about four inches square, on which the name, date, and locality may be written, and attaching several of these species-papers to a larger or genus-paper, or by devoting each larger paper to a species, adding in future other varieties, and enclosing all the species-papers of the same genus within a folded sheet, on which the name of the genus is written.
We have adopted, for our own herbarium, the “foolscap” size. A sheet of paper receives within its fold the specimens of a single species; these are affixed to the right-hand page, when the sheet 183is open, and a small envelope is attached by its face to the same page at the bottom, in which loose specimens are kept for minute and special examination, or as duplicates. When the sheet is folded, the specific name is written at the right-hand lower corner, or, what is better, a strip containing that name and its number is cut from a copy of the “Index Fungorum,” kept for the purpose, and gummed in its place. The remainder of this page, which is of white cartridge paper, is occupied with memoranda referring to the species enclosed, sketches of the spores, synonyms, references to descriptions, &c. All the species-papers of each genus are placed together within a sheet of brown paper, half an inch larger in each direction, with the name of the genus written at the left-hand corner. A piece of millboard, the size of the covers when folded, separates each order.
When a leaf, or other portion of a plant, is to be examined under the microscope, with the view of determining the genus and species of its parasite, it may be fastened with small pins to a piece of sheet cork, two or three inches square, and about one-eighth of an inch in thickness, such as used for lining entomological cabinets, and so placed under a lens that it may easily be brought into focus, and both hands left at liberty; or a dissecting microscope may be used for the purpose. From one of the pustules the spores may be removed on the sharp point of a penknife, and placed in a drop 184of water on a glass slide. A thin glass cover is placed over the drop of water, and the slide is submitted to examination. For further satisfaction it will often be found necessary to make carefully a thin section of a pustule, and place this under the microscope, a more troublesome but also much more satisfactory method. Reference to the Appendix will soon determine the name and position of the fungus, provided it belongs to the section to which this volume is devoted.
If it is thought desirable to mount the spores as permanent objects, there is no obstacle to such a proceeding. The spores of the different species of Aregma, of Triphragmium, and many of the Pucciniæ, will be worth the trouble. We have tried several media, and only adopted Glycerine or Balsam; either of these, especially the former, if the greater difficulty of securely closing can be overcome, will answer the purpose.
It should be remarked that in the examination of moulds, such as those of the genus Peronospora, included in this work, if any fluid be added, the acrospores are immediately disconnected from the threads, and float in the medium; so that if their mode of attachment or arrangement would be studied, that must be achieved without the addition of any fluid whatever.
The best methods of observing germination, the production of zoospores, &c., have been detailed already.
185Thus do we arrive at the close of the task which we had set ourselves to perform. This fragment of a history of microscopic fungi goes forth to plead for students, and prepare the path for somewhat more complete. Is it not a shame that more than two thousand species of plants (never mind how minute, how insignificant) should be known to exist, and constitute a flora, in a nation amongst the foremost in civilization, and yet be without a complete record? It is nevertheless true that hundreds of minute organisms, exquisite in form, marvellous in structure, mysterious in development, injurious to some, linked with the existence of all, are known to flourish in Britain without a history or description, in the language of, or produced in, the country they inhabit. It is also true that the descriptions, by which they should be known, of hundreds of the rest lie buried in a floating literature whence the youthful and ardent student needs, not only youth and ardour, but leisure and perseverance unlimited to unearth them. This, however, by the way; we may be too great dotards on our native land, and foolish in our desire to see her in advance, and not in the rear in scientific attainments, pursuits, and productions of other and, perhaps, less favoured nations.
Already we fear that some of our readers will have thought our story of rust, brand, and mildew interminable, and looked anxiously for the close. 186On the other hand, we would flatter ourselves into the belief that some have made a new acquaintance with these minute and mysterious forms, and would fain know more. The groups of fungi which we have endeavoured to illustrate have, with but few exceptions, one feature in common, i.e., that they are parasitic on living plants. These constitute but a small portion of the microscopical species found in this country. To the elegant forms of mould which inhabit decaying vegetable substances no allusion has been made. These constitute a fairy flora of forests and gardens with features as varied, and fruits as multiform, as those of the trees and flowers of the earth. The numerous, and often marvellous, phases of low life developed upon dead leaves and rotten sticks, would in description occupy a far greater space than we have devoted to our subject. Yet, for all these, we have not spared so much as a passing word. The treasures still left unopened are far richer than even those we have revealed. The gates of another world have been thrown open, but we have scarcely passed the threshold. A minutely and elaborately illuminated page of the book of Nature has been turned, and we have only perused a single line. We might traverse the primeval forests of the new world, and explore the unknown regions of the old, and not encounter so much to excite our admiration, or cause our wonder, as lies about our feet at home; marvels 187which we tread beneath our feet, or kick from our path, because they appear to be only rotten sticks, withered grass, and decaying leaves. All this may appear as the dream of an enthusiast, or the ravings of one on whom the moon has shone too often. When Columbus spoke of a new world beyond the seas, which he longed to seek and explore, some believed him duped, and others called him mad. We write of no chimerical El Dorado, we speak of no undiscovered world, and yet we seem to allude to wonders still unknown, because so few have had the courage to venture upon the journey for themselves.
In sober earnestness, however, let us commend this pursuit to all who possess a microscope and leisure to use it. It may be for a time the “pursuit of knowledge under difficulties,” on account of the condition of our literature on this special subject; but many workers will produce more readers, and good books will come when there are more to purchase and appreciate. It is not improbable that in more cases than have come under our own observation, microscopists wearied of diatoms and allied forms, or deeming themselves in possession of all that is novel or interesting in this direction, are seeking for a new field of labour, and a new subject to kindle up a new enthusiasm. To these we have advised, and to any more such we continue to advise, that fungi should have a fair trial. If variety is desired, here they will 188have at least 2,000 species for a knowledge of which the microscope is essential. If they thirst for discovery, let them be assured that here also the earnest worker is sure to meet with such a reward. Or if they would acquaint themselves with the manifestations of Divine power as developed in the most minute of created things, let them follow such observers as Tulasne and De Bary, and seek the “why and the wherefore” of the phenomena of mycetal life.
If there should still be any hesitation whether there is in this pursuit sufficient of the element of variety, for those who do not desire to pursue the subject into its deepest scientific recesses, to render it available for them, let them go to a good public library, such as that of the British Museum, and inquire for the large illustrated work by Corda, entitled “Icones Fungorum,” or the more recent volumes by Tulasne (Selecta Fungorum Carpologia), and examine the figures of microscopic fungi in either of those works, and decide for themselves.
| Peridium elongated— | |
| separating in threads | Rœstelia. |
| rupturing irregularly | Peridermium. |
| Peridium abbreviated, or semi-immersed | Æcidium. |
| Peridium immersed | Endophyllum. |
Peridium elongated, at length opening by lateral fissures, or a terminal lacerated mouth. Spermogonia on the opposite surface, on the same or on different leaves.
Rœstelia cancellata, Reb. Pear-leaf Rœstelia; spots yellow, then red, prominent; peridia split to the base into laciniæ, which remain united at the apex.—On Pear leaves. Not very common. Autumn. (Plate II. figs. 20, 21.)
Rœstelia cornuta, Tul. Horn-like Rœstelia; spots rusty-brown; peridia cylindrical, slightly curved, yellowish-brown; spores greyish, at length brown.—On the under surface of the leaves of Mountain-Ash. Not common. August. (Plate II. figs. 18, 19.)
Rœstelia lacerata, Tul. Lacerated Rœstelia; peridia clustered in tufts, brown, elongated, splitting to the base in segments; spores light brown.—On the under surface of the leaves, and on the petioles and fruit of the Hawthorn. Not uncommon. May to July. (Plate II. figs. 22, 26.)
Peridium elongated, at length bursting irregularly. Spermogonia scattered, conspicuous.
Peridermium Pini, Chev.; peridia oblong, scattered, large; spores orange, abundant; spermogonia vernal or autumnal, or both; spermatia large, white.—On leaves and young branches of Scotch Fir. Common in Scotland, occasional in England. Summer. (Plate II. figs. 27, 28.)
Peridermium elatinum, Lk.; simple, immersed; peridia elliptic, pallid; sporidia orange.—On Silver Fir, altering both foliage and ramification. Not common.
Peridium seldom elongated, opening by a terminal mouth, surrounded by a fringe of recurved teeth, or when short bursting irregularly. Spores disposed in chains. Spermogonia on the same or the opposite surface, clustered or scattered, central or intermixed.
Sect. I.—Peridia scattered (not collected in tufts or clusters).
Æcidium leucospermum, DC. White-spored Cluster-Cups; spots yellowish; peridia scattered, often covering the whole under surface; spores white, ovate.—On both sides of the leaves of the wood-anemone. Common. June. (Plate I. figs. 4-6.)
Æcidium quadrifidum, DC. Four-lobed Cluster-Cups; spots brownish; peridia scattered, occupying almost the entire under surface; spores brown, subglobose.—On the under side of leaves of Anemone in gardens. (Lobes at the mouth of the peridium not constantly four.) April to May.
Æcidium albescens, Grev. Moschatel Cluster-Cups; leaf blistered, whitish, scattered; peridia white, split into a few large teeth; spores yellowish-white.—On leaves and petioles of Adoxa moschatellina. Not uncommon. April.
Æcidium Epilobii, DC. Willow-herb Cluster-Cups; spots obliterated; peridia scattered, at length oval, wider above; spores orange, at length brown.—On the under side of leaves of Epilobium montanum, E. hirsutum, and E. palustre, rarely on the upper. Common. June to August.
Æcidium Thesii, Desv. Bastard-toadflax Cluster-Cups; spots obliterated; peridia scattered or biseriate; short, cylindrical, margin irregularly toothed, erect; spores yellowish, then dingy.—On Thesium humifusum. Downs near Winchester. July, 1864. Chipstead, Surrey, Oct. 14, 1864. (Plate III. figs. 50, 51.)
191Æcidium Soldanellæ, Hornsch. Soldanella Cluster-Cups; spots obliterated; peridia solitary, scattered over the inferior surface; spores orange.—On the under surface of the leaves of Soldanella alpina.
Æcidium Tragopogonis, Pers. Goatsbeard Cluster-Cups; spots obliterated; peridia scattered, torn, wider above; spores orange, at length black.—On stems, leaves, and involucres of common Goatsbeard. Very common. May to June. (Plate I. figs. 1-3.)
Æcidium Euphorbiæ, Pers. Spurge Cluster-Cups; spots obliterated, leaf thickened; peridia scattered or crowded, distinct; spores orange.—On the under surface of the leaves of Spurge. Common. May to June.
Æcidium Berberidis, Pers. Berberry Cluster-Cups; spots roundish, bright red; subiculum thickened; peridia in subrotund or oval patches, often elongated; spores orange.—On leaves, peduncles, and fruit of the common Berberry. Common. May to July. (Plate I. figs. 7-9.)
Æcidium Thalictri, Grev. Meadow-Rue Cluster-Cups; collected in roundish clusters; peridia oblong; spores bright orange.—On Thalictrum alpinum. Not uncommon in Scotland.
Æcidium crassum, Pers. Buckthorn Cluster-Cups; spots yellow-brown, subiculum thickened; peridia crowded into a roundish heap, at first globose, yellow, at length open; spores orange.—On Rhamnus catharticus and R. frangula. Common.
Æcidium Periclymeni, DC. Honeysuckle Cluster-Cups; spots variegated, yellow and brown, subiculum thickened; peridia sometimes elongated, in roundish or effused heaps; spores orange.—On the under surface of Honeysuckle leaves. Not common. June to August.
Æcidium Calthæ. Grev. Marsh-Marigold Cluster-Cups; aggregate; peridia somewhat campanulate, with numerous minute marginal teeth; spores bright orange, subglobose or oval.—On leaves and petioles of Caltha palustris. Margin of peridia pale and brittle. Rare. Spring.
Æcidium Ranunculacearum, DC. Crowfoot Cluster-Cups; spots obliterated, subiculum thickened; peridia in irregular heaps, densely crowded together; spores orange.—On leaves of various Ranunculaceæ; very common on R. Ficaria, not uncommon on R. repens, more rarely on R. acris and R. bulbosus. Spring. (Plate II. figs. 12-14.)
192Æcidium Galii, Pers. Bedstraw Cluster-Cups; spots linear or oblong, obscurely brown; peridia scattered, rarely aggregate, dentate, whitish; spores white.—On the leaves of Galium verum and G. mollugo. (Plate II. figs. 15-17.)
I am doubtful whether the Æcidium on Galium mollugo is really this species.—(M. C. C.)
Æcidium Bunii, DC. Pig-nut Cluster-Cups; spots obliterated, subiculum thickened; peridia in irregular subrotund or oval heaps; spores orange.—On Bunium bulbocastanum and Pimpinella saxifraga. Spring.
Æcidium Valerianacearum, Dub. Valerian Cluster-Cups; hypogenous, rarely cauline, spots on a thickened subcircular or oblong base; peridia scattered, more or less crowded, cup-shaped, tawny, margin erect, denticulate; spores dirty-yellow.—On Valeriana officinalis and V. dioica. North Britain.
Æcidium Asperifolii, Pers. Borage Cluster-Cups; clusters subrotund, on a slightly thickened subiculum; peridia scattered; spores yellowish-white.—On leaves of various Boragineæ. Summer. The parts of the leaves on which it occurs are rendered concave on one side and convex on the other.
Æcidium Grossulariæ, DC. Gooseberry Cluster-Cups; spots yellow, bright red on the opposite side, with a yellow border; peridia crowded in roundish heaps, at length brown, and surrounded with a brown area; spores orange.—On leaves and fruit of Gooseberry and Currant. Common. May to June.
Æcidium Urticæ, DC. Nettle Cluster-Cups; spots obliterated, subiculum thickened; peridia disposed in elongated or subrotund heaps, at first subglobose, then gaping; spores orange.—On leaves and stems of Nettles, distorting them very much. Common. June. (Plate I. figs. 10, 11.)
Æcidium Behenis, DC. Bladder-Campion Cluster-Cups; spots yellow, brown on opposite side; peridia somewhat circinating, in subrotund heaps; spores brown.—On Silene inflata. Not common. Some of the peridia are short and open, others larger and closed.
Æcidium Orobi, DC. Bitter-Vetch Cluster-Cups; spots yellow, effused; peridia scattered and disposed in small heaps; spores at length white.—On stems and leaves of Orobus tuberosus. Scotland.
Æcidium Compositarum, Mart. Composite Cluster-Cups; spots purplish, subrotund, confluent above; peridia crowded, in orbicular patches, or circinating, on the under surface; spores orange, oval.
193Var. a. Taraxaci, Grev.; clusters small, scattered.—On leaves of the Dandelion. June to July.
Var. b. Prenanthis, Pers.; spots circular or irregular, purplish; subiculum incrassated.—On leaves of Hawkweed (Hieracium paludosum). Summer.
Var. c. Tussilaginis, Pers.; clusters round, on a thickened base; peridia circinating.—On the under surface of leaves of Coltsfoot and Butter-bur. Common. Autumn.
Var. d. Jacobæa, Grev.; pustular, soon becoming agglomerated, numerous, depressed; peridia splitting into short, brittle, yellowish-white teeth.—On leaves of Senecio Jacobæa and Sonchus arvensis. June to August.
Var. e. Lapsani, Purt.; spots purplish, irregular, confluent, on both sides of the leaves; peridia amphigenous, in irregular patches or scattered, not prominent, teeth numerous, minute, reflexed; spores yellow, oval.—On both surfaces of the leaves of Lapsana communis. April. Not uncommon.
Æcidium Saniculæ, Carm. Sanicle Cluster-Cups; spots purplish, slightly incrassated, small, scattered, roundish; peridia in small circulate clusters, hypogenous, and on the petioles, at first hemispherical, at length open, margin with from 4 to 6 spreading lobes; spores yellowish, elliptical.—On the under surface of the leaves and on the petioles of Sanicula Europæa. Not uncommon. May and June.
Æcidium Violæ, Schum. Violet Cluster-Cups; spots yellowish; peridia in irregular heaps, seriate and scattered; spores orange, at length brown.—On leaves, petioles, and sepals of Violets. Common. May and June.
Æcidium Poterii, Cooke. Burnet Cluster-Cups; spots obliterated, clusters subrotund or elongated; peridia hypogenous and on the petioles, circinating or scattered, immersed, margin irregularly fringed with numerous minute teeth, soon falling away; spores yellowish, oval.—On the under surface of the leaflets and on the petioles of Poterium Sanguisorba. Rare. May and June. Dartford Brent, Kent.
Æcidium Geranii, DC. Cranesbill Cluster-Cups; spots yellow and purple; peridia in circinating clusters; spores yellow, at length brown.—On the under surface of leaves of Geranium pratense and G. dissectum. Not common.
Æcidium Menthæ, DC. Mint Cluster-Cups; spots obliterated; subiculum thickened; peridia scattered, emersed, or aggregate immersed; spores orange, elliptic.—On various Mints. Common.
Æcidium Scrophulariæ, DC. Figwort Cluster-Cups; spots yellowish; peridia in roundish circinate clusters (rarely scattered) on the under surface; spores whitish, becoming tawny.—On 194the leaves of Scrophularia aquatica. Thame and Sydenham, Oxon.
Æcidium Pedicularis, Lobosch. Red-rattle Cluster-Cups; spots obliterated, subiculum thickened; peridia thickly and irregularly clustered, subimmersed; spores dirty, pallid orange.—On petioles, leaves, and stems of Pedicularis palustris. Not common.
Æcidium Primulæ, DC. Primrose Cluster-Cups; spots obliterated; peridia solitary, scattered, and crowded, hypogenous; spores whitish-yellow.—On the under surface of leaves of Primroses. Not common. May.
Æcidium rubellum, Pers. Dock Cluster-Cups; spots purple; peridia circinating, centre free; spores yellowish-white.—On leaves of Dock and Sorrel. Not uncommon in moist localities. May and June.
Æcidium Ari, Berk. Wake-robin Cluster-Cups; spots round, confluent; peridia circinating, not crowded, central ones abortive.—On leaves of Arum maculatum. Not common. June and July.
Æcidium Dracontii, Schwein. Arum Cluster-Cups; spots pallid, extensively scattered over the leaves, sometimes nearly covering them; peridia large, scattered, abundant, disposed without order on the spots; spores orange.—On Arum triphyllum, in gardens. Melbury, 1863 (Rev. M. J. B.) A North American species.
Æcidium Allii, Grev. Garlic Cluster-Cups; spots pale; peridia circinating, not contiguous; spores yellowish.—On leaves of broad-leaved Garlic (Allium ursinum). June and July.
Peridium enclosed within the substance of the leaf, bursting irregularly.
Endophyllum Sempervivi, Lév.; peridia immersed, elliptic or roundish; spores ochraceous, becoming brownish.—On leaves of Houseleek.
| Spores multiseptate— | |
| moniliform | Xenodochus. |
| cylindrical | Aregma. |
| biseptate | Triphragmium. |
| uniseptate | Puccinia. |
| Stroma tremelloid and expanded | Gymnosporangium. |
| clavate or club-shaped | Podisoma. |
Spores multiseptate, moniliform, breaking up into many distinct articulations.—Berk. Outl., pp. 328.
Xenodochus carbonarius, Schl. Burnet Chain-Brand; scattered, in small tufts, hypogenous; spores curved or straight, composed of from 5 to 15 articulations; obtuse at one extremity, slightly attenuate at the other.—On Burnet. Very rare. (Plate III. fig. 29.)
Spores cylindrical, multiseptate, scarcely moniliform, borne on a long peduncle.—Berk. Outl., p. 329.
Aregma mucronatum, Fr. Rose Brand; hypogenous, scattered over the leaves in minute tufts; spores 5- to 7-septate, terminal joint mucronate; peduncles incrassated below, fusiform.—On leaves of various Roses. Autumn. Frequent. (Plate III. fig. 38.)
Aregma acuminatum, Fr. Burnet Brand; hypogenous, scattered in minute tufts; spores multiseptate, terminal joint acuminate; peduncles equal.—On Burnet leaves. July. Common. (Plate III. fig. 32.)
Aregma bulbosum, Fr. Bramble Brand; hypogenous, with a dull red stain on the upper surface; spores in large tufts, 4-septate, terminal joint apiculate; peduncles incrassated, and bulbous at the base.—On Bramble leaves. Autumn. Very common. (Plate III. fig. 41.)
Aregma gracile, Berk. Raspberry Brand; hypogenous, scattered, in small tufts; spores 7- to 9-septate, the terminal joint apiculate; peduncles slender.—On Raspberry leaves. Autumn. (Plate III. fig. 43.)
196Aregma obtusatum, Fr. Strawberry Brand; hypogenous, scattered, in minute tufts; spores multiseptate, terminal joint obtuse; peduncles equal.—On leaves of barren Strawberry. Autumn. Common. (Plate III. fig. 35.)
Spores trilocular, septa mostly vertical and horizontal.—Berk. Outl., p. 332.
Triphragmium Ulmariæ, Lk. Meadow-sweet Brand; spots obliterated; sori at first subrotund, covered with the epidermis; at length, when the cuticle has vanished, effuse; spores brown, subturbinate, divided by a vertical dissepiment, shortly pedicellate.—On leaves of Meadow-sweet, Spiræa ulmaria. Autumn. (Plate III. fig. 48.)
Spores uniseptate, supported on a distinct peduncle.—Berk. Outl., p. 329.
Puccinia graminis, Pers. Corn Mildew; spots pale, diffuse; sori linear, confluent, amphigenous; spores at length black, clavate, very slightly constricted.—On the leaves and culms of corn and grass. Autumn. Very common, and injurious to corn. (Plate IV. figs. 57-59.)
Var. β. Arundinis, Grev. Sori broad; spores more constricted and with longer peduncles.—On the leaves and sheaths of reeds. Very common.
Puccinia striola, Lk. Sedge Mildew; spots pallid; sori linear, crowded, distinct, subconvex; spores at length black, oblong and slightly constricted, or obovate, and not constricted.—On Sedges, Rushes, &c. Autumn. Common.
Puccinia coronata, Cd. Coronated Mildew; spots pallid; sori linear, short, crowded, not confluent, surrounded by the ruptured epidermis; spores shortly pedicellate, pallid; the apex surrounded by obtuse radiating teeth.—On various Grasses. Autumn. Not uncommon. (Plate IV. figs. 60-62.)
Puccinia truncata, B. and Br. Iris Brand; spots obliterated; sori oblong, brown, surrounded by the scarious epidermis; spores obovate-oblong, even, attenuated below, upper cell abruptly truncate.—On Iris fœtidissima. Autumn.
Puccinia Asparagi, DC. Asparagus Brand; spots none; sori reddish-brown, generally on the stems, scattered and crowded, surrounded by the ruptured epidermis; spores oblong, obtuse, constricted; peduncle white, long, thread-like.—On dead stems of Asparagus. Autumn.
197Puccinia Polygonorum, Lk. Polygonum Brand; spots yellowish; sori minute, crowded into orbicular patches; spores brown-black, obovate-oblong, frequently constricted, with the upper joint globose.—On the under surface of the leaves of various species of Polygonum. Autumn. Common.
Puccinia vaginalium, Lk. Knotgrass Brand; spots none; sori hypogenous, subrotund or oblong, at first surrounded by the ruptured epidermis; spores brown, obtusely ovate; pedicels long, filiform.—On leaves and stems of Knotgrass, Polygonum aviculare. Autumn.
Puccinia Thesii, Chaill. Bastard-toadflax Brand; cauline and amphigenous; sori blackish-brown, small, roundish, or oblong, convex, scattered or aggregate, surrounded by the ruptured epidermis; spores ovate, obtuse, scarcely constricted; pedicels elongated.—On Thesium humifusum, in company with Æcidium Thesii. Chipstead, Surrey, Oct. 14, 1864. (A. Grugeon.)
Puccinia Primulæ, Grev. Primrose Brand; hypogenous, deep brown, solitary, scattered, or concentric and subconfluent spores obovate-oblong, slightly constricted.—On Primrose leaves. Not common.
Puccinia Veronicarum, DC. Veronica Brand; spots yellowish; sori subglobose, aggregate, or circinating, central one large; spores brown, obovate-oblong, more or less constricted.—On the under surface of the leaves of several species of Veronica. Not common.
Puccinia Glechomatis, DC. Ground-Ivy Brand; spots brownish; sori subrotund, scattered, hypogenous; spots brown, rather short, subelliptic, scarcely at all constricted.—On leaves of Ground-Ivy, Glechoma hederacea. September and October. Not uncommon. (Plate IV. figs. 73, 74.)
Puccinia Menthæ, Pers. Mint Brand; spots obliterated; sori varying in size, hypogenous, subrotund, scattered; spores at length black, subglobose, or angular; peduncles short.—On leaves of Mint, Wild Thyme, Marjoram, &c. Autumn. Common. (Plate IV. figs. 69, 70.)
Puccinia Scorodoniæ, Lk. Wood-sage Brand; spots obliterated; sori small, confluent in subrotund patches, hypogenous; spores cinnamon; peduncles very long.—On leaves of Wood-sage. Darenth Wood. Not common.
Puccinia Scrophulariæ, Lib. Figwort Brand; spots pallid; sori subrotund, oblong, or linear and confluent, surrounded by the ruptured epidermis; spores very much crowded, subglobose or oval, brownish, plicate, but not truly septate; pedicels elongated.—On Scrophularia aquatica. Penzance. Very rare.
198Puccinia Betonicæ, DC. Betony Brand; spots obliterated; sori hypogenous, subrotund, aggregate, surrounded by the ruptured epidermis; spores very pale-brown, short, obovate, elliptic; peduncles short.—On Stachys Betonica. Rare.
Puccinia Vincæ, Berk. Periwinkle Brand; spots yellowish; sori hypogenous, scattered, subrotund, surrounded by the ruptured epidermis; spores brown, oblong, slightly constricted, lower cell rather attenuated; peduncle very short.—On leaves of Vinca major. (Plate VI. fig. 132.) Autumn. Not uncommon.
Puccinia Campanulæ, Carm. Campanula Brand; spots apparently none; sori large, irregular, crowded, for a long time covered with the epidermis, at length surrounded by it; spores oblong-ovate, or slightly constricted; peduncles very short.—On Campanulæ. Not common.
Puccinia clandestina, Carm. Scabious Brand; spots yellowish; sori very minute, distinct, but collected together in great numbers, dark brown; epidermis evanescent; spores oblong, very slightly constricted, pedunculate.—On Scabiosa succisa. Not common.
Puccinia Compositarum, Sch. Compositæ Brand; spots obliterated or whitish; sori small, subrotund, generally hypogenous, encircled with the ruptured epidermis; spores brown, oval, scarcely constricted.—On the leaves of Centaureæ, &c. Autumn. Common. (Plate IV. figs. 67, 68.)
Puccinia Syngenesiarum, Lk. Thistle Brand; spots obliterated; sori minute, collected in oval blackish-brown, raised spots, covered with the epidermis; spores brown; peduncles very short.—On the leaves of Thistles. Autumn. Common. (Plate IV. figs. 63, 64.)
Puccinia glomerata, Grev. Ragwort Brand; spots pale; sori roundish, depressed, often confluent; spores oblong, very variable; peduncles short.—On leaves of Ragwort, Senecio Jacobæa. Not common.
Puccinia variabilis, Grev. Variable Brand; sori amphigenous, minute, roundish, surrounded by the ruptured epidermis, nearly black; spores variable, obtuse, cells often subdivided; peduncle very short.—On leaves of Dandelion. Summer and Autumn. Not uncommon. (Plate IV. figs. 82, 83.)
Puccinia Valantiæ, Pers. Crosswort Brand; spots obliterated; sori small, subrotund, scattered or crowded; spores pale-brown, obovate, attenuated below, the upper segment globular, easily separating.—On Galium cruciatum. Autumn. Not common.
199Puccinia Galiorum, Lk. Bedstraw Brand; spots obliterated; sori hypogenous, partly covered with and surrounded by the ruptured epidermis, scattered; spores brown.—On the leaves of several species of Galium and Asperula odorata. Not uncommon. (Plate VIII. figs. 172, 173.)
Puccinia Umbelliferarum, DC. Pig-nut Brand; spots obliterated; sori small and scattered, subrotund, surrounded by the remains of the ruptured epidermis; spores brown, broadly elliptic, much constricted; peduncle short.—On various Umbelliferæ. Common. (Plate IV. figs. 71, 72.)
Puccinia Ægopodii, Lk. Goutweed Brand; spots brown; sori minute, subrotund and elongated, surrounded by the ruptured epidermis, often circinating, and forming roundish patches; spores brown; peduncles very short.—On Ægopodium podagraria.
Puccinia Saniculæ, Grev. Sanicle Brand; orbicular, variable in size, blackish-brown, scattered, rather confluent; spores very obtuse; peduncles somewhat elongated.—On Sanicula Europæa. Not uncommon. Summer and Autumn.
Puccinia bullaria, Lk. Hemlock Brand; spots obliterated; sori subrotund or oblong, covered with the epidermis; spores brown, peduncle short, nearly obsolete.—On dry stems of Conium maculatum and other Umbelliferæ. August and Sept.
Puccinia Smyrnii, Cd. Alexander’s Brand; spots obliterated; sori hypogenous, large, solitary, scattered, brown; spores ovoid, obtuse, verrucose, slightly constricted, minutely pedicellate.—On Smyrnium olusatrum. (Plate III. figs. 55, 56.)
Puccinia Anemones, Pers. Anemone Brand; spots obliterated; sori subrotund, surrounded by the ruptured epidermis, scattered, aggregate, and confluent; spores brown, very much constricted, consisting of two nearly globose portions, echinulate; peduncles very short.—On various species of Anemone. Very common on Anemone nemorosa. April and May. (Plate IV. figs. 64, 65.)
Puccinia Calthæ, Lk. Marsh-Marigold Brand; spots brownish; sori small, subrotund, convex, surrounded by the ruptured epidermis, scattered; spores obovate, attenuated below, slightly constricted; peduncle very short.—On leaves of Marsh Marigold. Spring. Rare.
Puccinia Noli-tangeris, Corda. Balsam Brand; spots irregular, confluent, brownish; sori gregarious, reddish-brown; spores oblong, obtuse or attenuated, distinctly apiculate, brown; peduncles rudimentary. On leaves of Impatiens noli-tangere. Gathered at Albury, Surrey, by the Rev. L. Jenyns.—(M. J. B.)
200Puccinia Violarum, Lk. Violet Brand; spots yellowish; sori hypogenous, small, crowded, covered with the epidermis, then surrounded by it; spores brown, elliptic or broadly elliptic, slightly constricted.—On Violets. Autumn. Common.
Puccinia Lychnidearum, Lk. Lychnis Brand; spots yellowish; sori subrotund or oblong, unequal, scattered, rarely confluent; spores white, at length brown, elongated, oblong, slightly constricted.—On leaves and stems of Lychnidæ, Sagina procumbens. Autumn. Common.
Puccinia umbilici, Guep. Penny-wort Brand; seated on pallid spots; sori round, convex, compact, at length confluent in large orbicular patches; spores subglobose, not constricted, shortly pedicellate.—On Cotyledon umbilicus. Penzance, Guernsey, Corwen, N. Wales. W. Pamplin. (Plate IV. figs. 80, 81.)
Puccinia Rhodiolæ, B. and Br. Stonecrop Brand; spots orbicular, brown; sori minute, crowded; spores shortly pedicellate, articulations depressed, sometimes spuriously subdivided.—On leaves of Sedum Rhodiola. Summer. Not common.
Puccinia Saxifragarum, Schl. Moschatel Brand; spots obliterated; sori subrotund, scattered, crowded and confluent, when young surrounded by the epidermis; spores red-brown, rather short, oblong, slightly constricted.—On both surfaces of the leaves of Adoxa moschatellina. Not uncommon. Summer.
Puccinia Chrysosplenii, Grey. Golden-saxifrage Brand; sori of various sizes, few together and confluent, pale brown; spores long, somewhat waved, much attenuated at either extremity; peduncle elongated.—On the under surface of the leaves of Chrysosplenium oppositifolium. Rare.
Puccinia Epilobii, DC. Willow-herb Brand; spots pale; sori hypogenous, subrotund, crowded; epidermis evanescent; spores effuse, cinnamon, broadly elliptic, strongly constricted; peduncles very short.—On the leaves of Epilobium palustre. Not common.
Puccinia pulverulenta, Grev. Great Willow-herb Brand; sori hypogenous, dark brown, scattered or subconfluent, open concentric; spores minute, obovate, slightly constricted, lower cell rather attenuated; peduncle not very short.—On the leaves of Epilobium montanum and E. hirsutum. August and September. Common. (Plate IV. figs. 78, 79.)
Puccinia Circææ, Pers. Nightshade Brand; spots obliterated; sori minute, semiglobose, crowded into subrotund patches, occasionally confluent, generally covered with the epidermis; spores brown, oblong, often acute, sometimes obtuse; peduncles long, thick.—On the leaves of Circæa lutetiana and C. alpina. Autumn. Not common.
201Puccinia Prunorum, Lk. Plum-tree Brand; spots obliterated; sori hypogenous, subrotund scattered, epidermis obliterated; spores brown, peduncles very short.—On the leaves of Plum-trees. September and October. Very common.
Puccinia Fabæ, Lk. Bean Brand; spots none; sori subrotund, or elongated, surrounded by the ruptured epidermis; spores at length black, ovato-globose; peduncle slender.—On Beans. August and September. Not uncommon.
Puccinia Buxi, DC. Box Brand; spots none; sori subrotund, convex, scattered; spores brown, oblong, rather strongly constricted, lower cell slightly attenuated; peduncle very long.—On both surfaces of Box-leaves. April and May. Not uncommon.
Peduncles extremely long, agglutinated by gelatine into a tremelloid expanded mass. Spores uniseptate.
Gymnosporangium Juniperi, Lk.; forming a soft gelatinous, irregular, orange mass; spores ovate or subelliptic, filled with subglobose granules.—On living twigs of Juniperus communis.
Peduncles extremely long, agglutinated by gelatine into a common stem, spreading out above into a clavariæform mass; spores mostly uniseptate.—Berk. Outl., p. 331.
Podisoma Juniperi, Fr.; orange, clavariæform, somewhat branched; stroma simple; spores very long, lanceolate, filled with elliptic granules.—On living branches of Juniperus communis. Not common.
Podisoma foliicolum, B.; epiphyllous, brown-black masses, subglobose, subelliptic, or irregular, consisting of radiating, crowded, very slender, agglutinated filaments, each bearing an elliptic or clavate, very obtuse spore, 3- to 5-septate.—On living leaves of Juniperus communis. Rare.
Podisoma Sabinæ, Fr.; red-brown, tuberculiform and clavate, simple; stroma obliterated; spores obovate, uniseptate.—On living branches of Juniperus Sabinæ. April. Not uncommon.
Spores spherical, reticulated, proceeding from delicate branched threads.
Tilletia caries, Tul. Bunt; included within the germen; spores spherical, rather large, black.—On wheat, filling the grains with dark-coloured spores, fetid when crushed. Autumn. Very common. (Plate V. figs. 84-91.)
Plant deeply seated. Spores simple, springing from delicate threads, or in closely-packed cells, ultimately breaking up into a powdery mass.—Berk. Outl., p. 335.
Ustilago segetum, Ditm. Corn Smut; produced on the receptacle and rachis; epidermis soon ruptured; spores loose, minute, globose, black.—On the ears of corn and grasses. Autumn. Very common. (Plate V. figs. 98, 99.)
203Ustilago urceolorum, Tul. Sedge Smut; produced on the glumes and utricles; epidermis soon bursting; spores in a compact mass, afterwards breaking up, globose, rather large, granulated.—Surrounding the seed of various Carices; as, Carex prœcox, stellulata, recurva, and pseudo-cyperus. Autumn. Not uncommon. (Plate VI. figs. 109-111.)
Ustilago longissima, Tul. Elongated Smut; produced on the leaves in linear, long, parallel, dirty-olive patches; epidermis bursting longitudinally; spores globose, breaking up into minute granules, olive-black.—On leaves of Poa aquatica and fluitans. Summer. Common. (Plate V. figs. 105-107.)
Ustilago olivacea, Tul. Olive Smut; infesting the enlarged receptacle; epidermis soon bursting; spores olive-green, powdery, minute, mixed with filaments.—On Carex riparia. Not common. (Plate VI. figs. 126, 127.)
Ustilago hypodytes, Fr. Grass-culm Smut; produced on the culms beneath the sheaths, afterwards exposed; spores minute, subglobose, brownish-black.—On the culms of various Grasses. Summer. Sometimes not uncommon. (Plate V. figs. 100, 101.)
Ustilago Maydis, Corda. Maize Smut; produced on the stems, germens, &c.; epidermis at length bursting; spores spherical, minute, brownish-black, surface covered with echinulate warts.—On stems, &c., of Zea mays. (Plate V. fig. 108.)
Ustilago Montagnei, Tul. Beaksedge Smut; produced on the seeds; epidermis bursting; spores slightly angular, small, dark-coloured, intermixed sparingly with fragile filaments.—On seeds of Rhyncospora alba. Not common. (Plate V. figs. 96, 97.)
Ustilago typhoides, B. and Br. Reed Smut; prodded on the stems of reeds, forming thick bullate patches several inches long, occupying whole internodes, covered by their sheath; spores globose, rather large.—On stems of Arundo phragmitis. Autumn. Not uncommon. (Plate VI. figs. 128, 129.)
Ustilago Salveii, B. and Br. Cocksfoot Smut; produced on the leaves, forming elongated parallel sori on the upper surface; spores obovate, rather large, rough with minute granules.—On leaves of Dactylis glomerata and other Grasses. Not common. (Plate VI. figs. 117-119.)
Ustilago grammica, B. and Br. Banded Smut; forming little transverse bands consisting of short parallel black lines, 1/12 of an inch or more in length; spores globose, very minute.—On stems of Aira aquatica and A. cæspitosa. Uncommon. (Plate VI. figs. 120-122.)
204Ustilago vinosa, Tul. Oxyria Smut; produced on the swollen receptacles; spores roundish, very small, and minutely papillose, separately pellucid, in clusters, violaceous.—On the receptacles of Oxyria reniformis. Uncommon.
Ustilago utriculosa, Tul. Utricle Smut; produced in the germen and perigonium; epidermis soon ruptured; spores effuse, minute, globose, purple-black.—On Polygonum hydropiper and other Polygona. Autumn. Not uncommon. (Plate VI. figs. 112-116.)
Ustilago flosculorum, Fr. Floret Smut; produced within the florets; spores minute, purplish-brown.—On the florets of Scabiosa arvensis. Not common. (Plate VI. figs. 123-125.)
Ustilago receptaculorum, Fr. Goatsbeard Smut; produced within the receptacles; spores ovate, minute, reticulated, violet-brown, nearly black, very profuse, filling the receptacle.—On the receptacles of Goatsbeard. June, July. Common. (Plate V. figs. 92-95.)
Ustilago antherarum, Fr. Anther Smut; produced on the anthers and germens; spores subglobose, effuse, violet.—On the anthers of Silene inflata, &c. (Plate V. figs. 102-104.)
Stroma composed of little irregular cells forming a lentiform disk, whose surface is covered with many layers of cells, each of which encloses a spore; spores simple, always without any appendage.—Berk. Outl., p. 331.
Uredo Potentillarum, DC. Potentilla Uredo; spots yellowish; sori subrotund and oval, bullate, aggregate, open, confluent; spores subglobose, subcoherent, orange.—On various Rosaceæ. Common.
Uredo Saxifragarum, DC. Saxifrage Uredo; spots pallid; sori subrotund and oval, raised, scattered and aggregate on the under surface; epidermis ruptured, persistent; spores subglobose, yellow.—On various Saxifrages.
Uredo Filicum, Desm. Fern Uredo; spots yellowish; sori subrotund, bullate, scattered and aggregate on the under surface; epidermis at length bursting; spores subglobose, yellow.—On Ferns (Cystopteris, &c.) Not common.
Uredo pustulata, P. Willow-herb Uredo; spots yellowish; sori subrotund, minute, closed, scattered and confluent on both surfaces; spores globose, yellow.—On Epilobium palustre.
205Uredo Hypericorum, DC. St. John’s-Wort Uredo; spots yellowish; sori subrotund, small, bullate, distinct, scattered on the under surface; epidermis at length bursting; spores subglobose, cohering, orange.—On various Hyperica. August. Not uncommon. (Plate VIII. figs. 174, 175.)
Uredo Caryophyllacearum, Johnst. Stitchwort Uredo; spots yellowish; sori subglobose, scattered and aggregate, minute, generally on the under surface; epidermis closed; spores oval, at length yellow.—On various Caryophyllaceæ. Autumn.
Uredo Quercus, Brond. Oak-leaf Uredo; on the under surface; sori yellow, then orange, minute, ovate, and orbicular, slightly prominent, scattered, solitary or agglomerated into minute patches, surrounded by the ruptured epidermis; spores subglobose, pellucid, not cohering.—On Oak-leaves. September. Not common.
Uredo porphyrogenita, Kze. Bird-cherry Uredo; spots purplish; sori subrotund, small, aggregate, hypogenous; epidermis at length ruptured; spores coherent, subglobose.—On leaves of Prunus Padus. Scotland, 1863. (Rev. M. J. Berkeley.)
Uredo Vacciniorum, P. Bilberry Uredo; spots yellow-brown; sori subrotund, minute, aggregate, and scattered, on the under surface of the leaves; epidermis seldom ruptured; spores ovoid, yellowish.—On Vaccinium Myrtillus and V. vitis-idæa. Scotland.
Uredo confluens, DC. Mercury Uredo; on the under surface, depressed, yellow, oblong, concentric, at length confluent; spores nearly oval.—On Mercurialis perennis and M. annua. May, June. Common. (Plate VII. figs. 133, 134.)
Uredo Alliorum, DC. Garlic Uredo; spots obliterated; sori linear, oblong, and oval, on both surfaces; spores ovoid or subglobose, yellow or whitish.—On various species of Allium. Common.
Uredo Statices, Desm. Sea-lavender Uredo; sori few and scattered, orbicular or oval; spores sessile, globose, brown.—On various species of Statice.
Uredo Circææ, A. & S. Nightshade Uredo; sori minute, crowded, subrotund, slightly confluent, on the under surface; spores irregular, ovate, small, ochraceous.—On leaves of Circæa lutetiana. June, July. Common. (Plate VII. figs. 135, 136.)
Uredo bifrons, Grev. Twin-faced Uredo. On both surfaces of the leaves, often opposite, scattered, round, light brown, girt with the remains of the epidermis; spores globose.—On Sorrel leaves (Rumex acetosa and R. acetosella). July, September. Common. (Plate VII. figs. 137-139)
Stroma surrounded or sprinkled with elongated abortive spores. Spores free, invested with their mother-cell, or concatenate.—Berk. Outl., p. 334.
Lecythea Ruborum, Lév. Bramble Rust. Spots pale, brown or purple on the opposite side, sometimes depressed above; sori subrotund, aggregate; epidermis soon bursting; spores globose or subglobose, echinulate, bright ochraceous-yellow.—On the under surface of Bramble leaves. July and August. Very common. (Plate II. fig. 40.)
Lecythea Rosæ, Lév. Rose Rust; spots yellow, small, scattered; spores sub-oval, sometimes minutely pedicellate, orange.—On Rose-leaves. July, September. Extremely common. (Plate II. fig. 37.)
Lecythea Poterii, Lév. Burnet Rust; spots obliterated, rufous on the opposite side; sori subrotund, scattered, minute on the under surface; epidermis bursting; spores subglobose, often pedicellate, intense orange; barren spores pale, cylindrical, and slightly curved.—On Poterium Sanguisorba. Summer. Common. (Plate III. fig. 31.)
Lecythea Populina, Lév. Poplar Rust; hypogenous; sori yellow, roundish or oblong, surrounded by the ruptured epidermis; spores copious, elongated or ovate.—On Poplar and Birch leaves. Summer. Common.
Lecythea Euphorbiæ, Lév. Spurge Rust; spots obliterated; sori subrotund, small, scattered, surrounded by the ruptured epidermis; barren spores pyriform or subglobose, pedicellate; fertile spores subglobose, orange.—On the under surface of the leaves of various species of Euphorbia, especially E. helioscopia and E. peplus. August. Very common.
Lecythea epitea, Lév. Tawny Willow Rust; sori roundish, scattered, at first tawny, at length growing pale, surrounded by the ruptured epidermis; spores subrotund and pyriform.—On the under surface of Willow-leaves.
Lecythea mixta, Lév. Orange Willow Rust; spots yellow; sori subrotund, aggregate, confluent, effuse, permanently surrounded by the ruptured epidermis; spores oblong and pyriform, orange.—On both surfaces of the leaves of Willows.
207Lecythea Saliceti, Lév. Common Willow Rust; spots yellowish; sori subrotund, solitary, or in circles, surrounded by the ruptured epidermis; barren spores subglobose and pedicellate or pyriform; fertile spores subglobose, orange.—On the under surface of Willow-leaves. Autumn. Common.
Lecythea Baryi, Berk. De Bary’s Rust; sori few; cystidia with an abrupt globose head; spores subglobose.—On leaves of Brachypodium pennatum. Very rare.
Lecythea Valerianæ, Berk. Valerian Rust; spots yellowish; sori subrotund, small, confluent, sometimes circinating; epidermis at length bursting; spores reddish-brown, subglobose or clavate, shortly pedicellate.—On Valeriana officinalis. August.
Lecythea gyrosa, Berk. Ringed Rust; spots obliterated; sori minute, confluent, and forming a small distinct ring; epidermis bursting; spores globose and elongato-pyriform, yellow or pale.—On the upper surface of Raspberry and Bramble leaves, forming a more or less perfect ring with the centre unoccupied. September. (Plate VIII. figs. 162-164.)
Lecythea Capræarum, Berk. Sallow Rust; spots obliterated, yellow on the opposite side; sori varied in form, here and there confluent, surrounded by the ruptured epidermis; barren spores subglobose and pyriform, pedicellate; fertile spores subglobose, dirty yellow.—On the under surface of the leaves of Sallows. June to August. Very common. (Plate VIII. figs. 160, 161.)
Lecythea Lini, Berk. Flax Rust; spots yellowish; sori subrotund, scattered, surrounded by the ruptured epidermis; spores globose or pyriform, sometimes pedicellate.—On Linum catharticum. July. Not uncommon. (Plate VIII. figs. 165-167.)
Spores free; attached at first to a short peduncle, which at length falls away.—Berk. Outl., pp. 332.
Trichobasis rubigo-vera, Lév. Round Corn Rust; spots yellow, heaps oval, scattered, generally on the upper surface; epidermis at length bursting longitudinally; spores subglobose, reddish-brown, easily dispersed.—On Grasses and Corn. Throughout the Summer. Very common. (Plate VII. figs. 140-142.)
208Trichobasis linearis, Lév. Long Corn Rust; spots yellow-brown. Sori elliptic, then elongated and linear; epidermis bursting; spores oblong or globose, yellow.—On leaves and sheaths of Corn and Grasses. Summer. Common. Plate VII. figs. 143, 144.)
Trichobasis Glumarum, Lév. Glume Rust; sori minute, round, scarcely convex, subgregarious, often confluent. Spores globose or subovoid, orange, not pedicellate; epispore smooth.—On the glumes of Cereals. August.
Trichobasis Symphyti, Lév. Comfrey Rust; Sori minute, very numerous, scattered, roundish, then confluent; epidermis ruptured, scarcely conspicuous around the margin; spores subglobose, pallid orange.—On Comfrey.
Trichobasis Pyrolæ, B. Wintergreen Rust; spots yellowish-brown on the opposite side; sori globose, minute, scattered or aggregate, on the under surface; epidermis generally closed; spores subglobose, yellow.—On Pyrola rotundifolia, &c.
Trichobasis Petroselini, B. Parsley Rust; spots yellowish; sori subrotund and oval, confluent, on both surfaces; epidermis at length ruptured; spores globose or subglobose, occasionally obsoletely pedicellate, pale yellow.—On various Umbelliferæ.
Trichobasis Senecionis, B. Groundsel Rust; spots obliterated; sori solitary or regularly crowded, subrotund and oval, on the under surface, surrounded by the ruptured epidermis; spores subglobose, orange.—On various species of Groundsel. July. Very common. Plate VII. figs. 145, 146.)
Trichobasis Caricina, B. Sedge Rust; spots red; sori oval, minute, scattered, surrounded by the ruptured epidermis; spores subglobose, reddish, then brown.—On Carex pendula, C. pseudo-cyperus, &c. June to August. Common. (Plate VIII. figs. 170, 171.)
Trichobasis oblongata, B. Luzula Rust; spots oblong, often confluent, yellow-brown; sori elliptic, on both surfaces; epidermis closed; spores brown, obtuse at either extremity.—On Luzulæ. May to July. (Plate VII. figs. 158, 159.)
Trichobasis Scillarum, B. Hyacinth Rust; spots oblong or subrotund, crowded into patches; epidermis bursting longitudinally; spores rubiginous, obovate, shortly pedunculate.—On the Wild Hyacinth, &c.
Trichobasis Cichoracearum, Lév. Hawkweed Rust; on both sides of the leaf, dark, fuscous, minute, round, scattered: spores globose, rarely minutely pedicellate.—On Thistles and Hawkweed. July to September. Common.
209Trichobasis Artemisiæ, B. Mugwort Rust; spots obliterated, brownish on the opposite side; sori subglobose and oval, minute, scattered, on both surfaces; epidermis soon ruptured; spores subglobose, brownish.—On Artemisia vulgaris, &c. Not common.
Trichobasis Labiatarum, Lév. Mint Rust; spots yellowish and brown; sori subrotund, scattered, subaggregate, on the under surface; epidermis ruptured; spores subglobose, brown.—On various Labiatæ. August to September. Common.
Trichobasis Lychnidearum, Lév. Chickweed Rust; spots pallid yellowish; sori subrotund plane, scattered on the under surface, cinnamon, at length brownish; epidermis ruptured; spores globoso-ovoid, sessile, or shortly pedicellate.—On Caryophyllaceæ. Summer and Autumn.
Trichobasis Umbellatarum, Lév. Hemlock Rust; spots yellowish; sori subrotund and ovate, scattered, on the under surface, surrounded by the ruptured epidermis; spores ovate, oval, and oblong in the same heap, brown.—On Conium maculatum, Apium graveolens, and other Umbelliferæ. August and September.
Trichobasis Heraclei, B. Hogweed Rust; on the under surface, scattered, sometimes subconfluent, roundish, light brown, girt by the remains of the epidermis; spores obovate, with a very short peduncle.—On Heracleum spondylium. June and July.
Trichobasis Hydrocotyles, Cooke. Flukewort Rust; without definite spots; sori chiefly on the upper, sometimes on the under surface, scattered, variable, roundish, erumpent, surrounded by the ruptured epidermis; spores subglobose, at length brown; epispore rough with minute tubercles.—On Hydrocotyle vulgaris. Epping, July to September, 1863 and 1864. (Plate VIII. figs. 168, 169.)
Trichobasis Betæ, Lév. Beet-leaf Rust; spots yellow; heaps subrotund and oval, scattered and concentric, on the upper surface; epidermis at length bursting; spores subglobose, shortly pedicellate, brown.—On the leaves of Beet. August and September. Common.
Trichobasis Fabæ, Lév. Bean Rust; spots obliterated; sori subrotund and oval; bullate, scattered and aggregate, surrounded by the ruptured epidermis; spores ovoid, brown.—On Beans. August and September. Common.
Trichobasis Galii, Lév. Bedstraw Rust; spots yellowish; sori subrotund, aggregate, closed; spores globose, reddish.—On Galium verum, saxatile, &c. July and August.
210Trichobasis suaveolens, Lév. Thistle Rust; spots obliterated, yellow on the opposite side; sori subrotund, nearly plane, scattered, at length confluent, on the under surface, surrounded by the ruptured epidermis; spores globose, brown.—On leaves of Thistles, frequently covering the whole under surface. Summer. Common. (Plate VII. figs. 151-153.)
Trichobasis Polygonorum, B. Knotgrass Rust; spots red-yellow, widely effused; sori subrotund, scattered, sometimes forming a ring, epidermis at length bursting; spores somewhat obovate, brown.—On Polygonum aviculare, amphibium, and other species. July to September. Common.
Trichobasis Vincæ, B. Periwinkle Rust; spots yellowish; sori small, subrotund, and oval, on the under surface, surrounded by the ruptured epidermis; spores oval, rather ovoid, brown.—On leaves of Vinca major. May and June. (Plate VI. fig. 130, 131.)
Trichobasis Geranii, B. Geranium Rust; spots yellowish; sori subrotund, nearly plane, scattered or confluent; spores subglobose, brown.—On various species of Geranium.
Trichobasis Violarum, B. Violet Rust; spots yellowish; sori subrotund, scattered, generally on the under surface; epidermis ruptured, persistent; spores subglobose, brown.—On leaves and petioles of Violets. July. Not uncommon.
Trichobasis Parnassiæ, Cooke. Grass of Parnassus Rust; on both surfaces of the leaves; sori at first bullate, at length rupturing the epidermis, scattered, often confluent; spores globose or nearly so, rather large, tawny brown.—On Parnassia palustris. Irstead, Norfolk. September, 1864.
Trichobasis Epilobii, Berk. Willow-herb Rust; spots yellowish; sori subrotund, scattered, surrounded by the ruptured epidermis, often on the under surface; spores subglobose, brown.—On Epilobium montanum, &c. June and July.
Trichobasis Rhamni, Cooke. Buckthorn Rust; seated on definite yellowish spots; sori occurring only on the under surface of the leaves, scattered or collected in clusters, very minute, roundish, or oblong; spores at length rusty brown, subglobose.—On Rhamnus catharticus. Selsdon, Surrey. September 16, 1864. (A. Grugeon.)
Spores unilocular, attached permanently to a decided peduncle of greater or less length.—Berk. Outl., p. 333.
211Uromyces Alliorum, DC. Garlic Rust; spots obliterated, sori linear, oblong, or oval, amphigenous; spores subglobose, yellow.—On species of Allium.
Uromyces Ulmariæ, Lév. Golden Rust; broad, pulverulent, on the under surface of leaves, nerves, and petioles; spores numerous, subglobose, bright reddish-orange.—On Spiræa Ulmariæ, Roses, &c. Common. (Plate VII. figs. 147, 148.)
Uromyces appendiculata, Lév. Long-stemmed Rust; spots yellowish-brown, sori subrotund and oval, confluent, nearly plane, on the under surface. Epidermis bursting, spores ovoid, brown, with a long peduncle.—On Leguminosæ and other plants. August and September. (Plate VII. figs. 149, 150.)
Uromyces apiculosa., Lév. Short-stemmed Rust: spots yellow or brown, sori subrotund, scattered, surrounded by the ruptured epidermis; spores ovoid, brown, shortly pedunculate.—On dock and various other plants. August and September. Common. (Plate VII. figs. 154, 155.)
Uromyces Limonii, Lév. Sea-Lavender Rust; epiphyllous, sori bullate, scattered or disposed in rings; spores ovate.—On Statice limonia.
Uromyces Ficariæ, Lév. Pilewort Rust; spots yellowish, sori scattered, aggregate, confluent, and expanded; epidermis ruptured; spores ovoid, brown.—On Ranunculus Ficaria. May and June. Common. (Plate VII. figs. 156, 157.)
Uromyces Primulæ, Lév. Primrose Rust; spots yellowish, sori subrotund and oval, aggregate, on the under surface; epidermis at length bursting; spores ovoid, brown.—On Primroses.
Uromyces intrusa, Lév. Lady’s-mantle Rust; on the under surface, scattered, or partially aggregate, reddish-brown, rounded, somewhat prominent, minute, very unequal; spores roundish or oval.—On Alchemilla vulgaris.
Uromyces Iridis, Lév. Iris Rust; on the under, rarely on the upper, surface; spots yellow, sori small, pale red-brown, oblong and linear, scattered or aggregate, scarcely convex; epidermis bullate, rarely bursting longitudinally; spores globose or broadly elliptic, pale brown, pellucid.—On Iris fœtidissima. August to September. Not uncommon.
Spores irregular, consisting of several cells.—Berk. Outl., p. 334.
Polycystis Colchici, Tul. Meadow-saffron Smut; sori elongated, bursting irregularly; spores smooth or slightly papillose.—On leaves of Meadow-Saffron. Not common.
212Polycystis Violæ, B. and Br. Violet Smut; sori scattered, elongated, on both surfaces of the leaves and petioles; spores more or less globose, consisting of several cells surrounded by a common irregular crust.—On leaves and petioles of Violets. August. Common. (Plate IX. figs. 185, 186.)
Polycystis pompholygodes, Lév. Buttercup Smut; sori varied in form, bullate, epidermis inflated, at first entire, then bursting irregularly, its remains surrounding the clusters; spores copious, subglobose, black, opaque or pellucid.—On Ranunculus repens and other Ranunculaceæ. Summer. Very common. (Plate IX. figs. 183, 184.)
Polycystis parallela, B. and Br. Rye Smut; sori very long, linear, epidermis bursting longitudinally, spores globose, with several projecting nodules, dark brown.—On culms and sheaths of Rye and on the leaves of Carices. (Plate IX. figs. 187, 188.)
Plant deeply seated; spores multicellular, subglobose, or conchiform. Berk. Outl., p. 335.
Tuburcinia scabies, B. Potato Smut; spores globose composed of minute cells forming together a hollow globe with one or more lacunæ, generally attached laterally by a slender thread, olive. On Potatoes. Common. (Plate III. fig. 54.)
Tuburcinia Trientalis, B. and Br. Trientalis Smut; sori two lines broad, bullate, containing a black mass of rather irregular depressed subglobose spores, which are very opaque and distinctly cellular. Hyphasma white, branched, creeping, delicate.—On leaves of Trientalis Europæa. August and September. Scotland. (Plate III. figs. 52, 53.)
Spores cylindrical, septate, some separating at the joints, some of a different nature, persistent.—Berk. Outl., p. 333.
Coleosporium Tussilaginis, Lév. Coltsfoot Rust; on the under surface, prominent, crowded, generally forming circles, becoming very confluent; spores numerous, sub-ovate, orange-yellow.—On Tussilago Farfara. Summer. Common. (Plate VIII. figs. 180, 181.)
Coleosporium pingue, Lév. Tawny Rose Rust; spots obliterated, sori effuse, on the nerves and petioles of the leaves; spores ovoid, yellowish-brown.—On Roses, &c.
213Coleosporium Petasitis, Lév. Butterbur Rust; on the under surface, minute, depressed, spreading, somewhat aggregate, subconfluent, irregular in form; spores oval, orange, or orange-red.—On Tussilago Petasites. Autumn. Common.
Coleosporium Campanulæ, Lév. Campanula Rust; spots obliterated, brown on the opposite side, sori irregular, confluent, plane, on the under surface; spores subglobose, cohering, yellow, at length pale. On leaves of various Campanulæ. September and October.
Coleosporium Sonchi-arvensis, Lév. Sow-thistle Rust; on the under surface, depressed, irregular in form, scattered, partially confluent; spores ovate, reddish orange.—On Sonchus oleraceus and arvensis. Summer. Common. (Plate VIII. figs. 178, 179.)
Coleosporium Rhinanthacearum, Lév. Cow-wheat Rust; spots none or subferruginous; sori irregular, confluent, on the under, rarely on both surfaces; spores subglobose, compact, golden-yellow.—On Euphrasia officinalis, Bartsia odontites, Melampyrum arvense, &c. August and September. Common. (Plate VIII. figs. 170, 177.)
Spores of two orders, crowded into a dense compact mass, with or without a covering, wedge-shaped. Berk. Outl., p. 333.
Melampsora Euphorbiæ, Cast. Hypophyllous; sori of summer spores golden-yellow, scattered, distinct, sometimes cauline; spores small, subglobose; sori of perfect spores becoming black, small, roundish, spores prismatic, membrane thickened above, dark brown.—On leaves and stems of Euphorbia helioscopia, E. exigua, and other species of spurge. Common. (Plate IX. figs. 193, 194.)
Melampsora Populina, Lév. Hypophyllous, epiphyllous, or amphigenous, at first yellow or orange; summer spores obovate-oblong, attenuated or truncate, echinulate; paraphyses obovate, capitate or claviform, abundant in fully-ripened sori; sori of perfect spores at first tawny yellow, becoming black during the winter, swelling in the spring, and becoming of a cinnamon-colour, hypophyllous, roundish, or oblong; spores prism-shaped, 5-6 together, yellowish, smooth.—On leaves of Populus nigra. Common. Spores perfected in February. (Plate IX. figs. 195, 196.)
214Melampsora Tremulæ, Tul. Hypophyllous; sori of summer spores punctiform, prominent, or papillæform, numerous; spores tawny-yellow, elliptical or obovate; paraphyses slender; sori of perfect spores scattered, at length blackish; spores elongated, compressed, attenuated downwards, yellowish.—On leaves of Populus tremula. Common. Spores perfected during the winter.
Melampsora Betulina, Desm. Hypophyllous; sori of summer spores bright yellow or orange, oblong, cylindrical, or obovate, truncate at the base, echinulate; paraphyses encircling or intermixed, obovate, smooth, hyaline; sori of perfect spores confluent, of an obscure brown in the winter, becoming of a bright orange when mature; spores elongated, attenuated below, polygonal, ochraceous.—On leaves of Betula alba. Common. Spores perfected in January and February. (Plate IX. figs. 189, 190.)
Melampsora Salicina, Lév. Epiphyllous or hypophyllous; sori, or heaps of summer spores, scattered, pale orange, bright orange, or cinereous; spores ovato-globose, paraphyses capitate, rarely obovate; sori of perfect spores epiphyllous, scattered or aggregate, at first yellowish-tawny, then brownish, at length nearly black, bullate; spores oblong, closely packed, and laterally compressed.—On Salix viminalis and S. capræa. Common. Spores perfected in February. (Plate IX. figs. 191, 192.)
Receptacle consisting of thick branched threads; conidia concatenate, at length separating; oospores deeply seated on the mycelium.
Cystopus candidus, Lév. Crucifer White Rust; conidia equal, globose; membrane equal, ochraceous; oospores subglobose, epispore yellowish-brown, with irregular obtuse warts; warts solid.—On Shepherd’s-purse, Cabbages, and other Cruciferæ. Summer. Very common. (Plate X. figs. 198-200, 205-207.)
Cystopus cubicus, Str. Goatsbeard White Rust; conidia unequal; terminal cell sterile, larger than the rest; membrane thickened, ochraceous, rarely yellowish; fertile cells shortly cylindrical; membrane hyaline; oospores globose; epispore brown, verrucose; warts hollow, round, or irregular.—On goatsbeard, salsify, and scorzonera. Summer and autumn. Common. (Plate X. figs. 201, 202, 210.)
Cystopus Lepigoni, De By. Sandwort White Rust; conidia unequal; terminal cell sterile, globose; membrane thickened; fertile cells subglobose or cylindrical; membrane hyaline; oospores globose; epispore brown, tubercles minute, irregular, very convex, often resembling spines.—On Spergutaria rubra. Swanscombe Marshes, September, 1864. (R. G. Keeley.)
Parasitic threads mostly inarticulate. Spores of two kinds:—1. Acrospores on the tips of the branchlets; 2. Oospores large, globose, on the creeping mycelium.
Peronospora infestans, Mont. Potato Mould; threads of mycelium slender, always destitute of suckers; fertile threads thin, gradually attenuated upwards, with one to five branches, one or more inflated vesicles near the apices of the branches; branches either simple or with short branchlets; acrospores ellipsoid or ovoid; apex furnished with a prominent papilla.—On leaves, stems, and tubers of the Potato, causing the potato-murrain. Very common since 1845. (Plate XIV. fig. 264.)
Peronospora nivea, Ung. (P. macrospora, B.). Parsnip Mould; threads of mycelium stout, often torulose; suckers numerous, vesicular, obovate; fertile threads fasciculate, dwarfish, tapering or subulate, or once or twice shortly bifurcate, rarely trifurcate, with one to four horizontal branches near the summit, once, twice, or three times bifurcate; acrospores subglobose or ovoid, with an obtuse papilla at the apex.—On various Umbelliferæ. Common and variable.
Peronospora pygmæa, Ung. (P. curta, Berk.). Anemone Mould; threads of mycelium thickened, often constricted and varicose; suckers minute, obovate, or pear-shaped; fertile threads fasciculate (2-5 or more), simple above or divided at the apex into 2-4 short simple branches, or shortly twice dichotomous, or all simple, obtuse, surmounted by 2-4 short spicules; acrospores ovoid or ellipsoid, variable in size; apices broadly and obtusely papillate.—On Wood-anemone. Not uncommon. (Plate XV. fig. 267.)
Peronospora gangliformis, Berk. Lettuce Mould; threads of the mycelium stout, now and then torulose; suckers vesicular, obovate or clavate; fertile threads 2-6 times dichotomous, sometimes trichotomous; stems and primary branches slender, dilated or inflated above; the ultimate ramuli inflated at the apex into a turbinate or subglobose vesicle bearing from 2-8 subulate processes or spicules; acrospores minute, subglobose; apices with broad depressed papillæ, produced on the spicular processes, On Lettuces and other Compositæ. Frequent. (Plate XIV. fig. 265.)
216Peronospora parasitica, Pers. Cabbage Mould; threads of the mycelium thickened and much branched; suckers numerous, branched; branches clavate, obtuse; fertile threads thick, soft, flexile, equal or unequal, 5-8 times dichotomous, rarely trichotomous; branches always repeatedly trifurcate; acrospores broadly elliptical, very obtuse at the apex, white.—On Cabbages, Shepherd’s-purse, and other Cruciferæ, sometimes in company with Cystopus candidus. Summer and autumn. Common. (Plate XIII. fig. 262.)
Peronospora Viciæ, Berk. Pea Mould; fertile threads densely cæspitose, erect, equal, rarely unequal, 6-7-8 times dichotomous; ultimate ramuli shortly subulate, acute; acrospores ellipsoid, very obtuse at the apex, obtuse or slightly acute at the base; membrane with a violaceous tint.—On Tares, Peas, &c. Frequent. (Plate XV. fig. 266; Plate X. fig. 212.)
Peronospora Arenariæ, Berk. Sandwort Mould; fertile threads slender, 6-7 times equally, rarely unequally, dichotomous; branches spreading; ultimate ramuli slender, acute, subulate, nearly erect; acrospores broadly elliptical, oftentimes very obtuse, small; membrane scarcely violaceous.—On Arenaria serpyllifolia and A. trinervis.—June. (Plate XVI. fig. 268; Plate X. fig. 211.)
Peronospora effusa, Grev. Spinach Mould; fertile threads fasciculate, short, thick, 2-6, rarely 7 times dichotomous above; acrospores broadly ellipsoid, sometimes very obtuse; membrane with a violaceous tint. On Spinach, Goosefoot, and some other allied plants. Spring and autumn. Not uncommon. (Plate X. figs. 214, 215.)
Peronospora Urticæ, Casp. Nettle Mould; fertile threads small, loosely 4-6 times dichotomous; branches flexuose, ultimate ramuli subulate, arcuate, often deflexed; acrospores large, broadly ovoid or subglobose, distinctly pedicellate; apices very obtuse; membrane violaceous.—On leaves of the common Nettle.
Peronospora trifoliorum, De By. Clover Mould; fertile threads cæspitose, equally or unequally 6-7 times dichotomous, rarely trichotomous; ultimate branches subulate, acute, slightly curved; acrospores ellipsoid, very obtuse; membrane with a slightly violaceous tint; oospores large; epispore brown.—On Lucern (Medicago sativa). Highgate and Hampstead, 1864.
217Peronospora grisea, Ung. Veronica Mould; fertile threads erect, fasciculate, grey, 5-7 times regularly dichotomous; branches gradually attenuated; primary oblique erect; others spreading, flexuose; ultimate mostly unequal, slightly arcuate; acrospores ellipsoid or ovoid, obtuse; membrane pale and dirty violet.—On leaves of Veronica beccabunga. May, 1846. (Plate X. fig. 213.)
Peronospora arborescens, Berk. Poppy Mould; fertile threads slender, erect, 7-10 times dichotomous above; branches more or less flexuose, squarrose, spreading, gradually attenuated; ultimate ramuli shortly subulate, more or less arcuate; acrospores very small, subglobose; membrane scarcely violaceous.—On the Corn Poppy. June. Common.
Peronospora Schleideniana, De By. (P. Destructor, B.) Onion Mould; fertile threads robust, erect, not septate, branched alternately; ultimate ramuli forked and uncinate or divaricate; acrospores seated on the tips of the ultimate ramuli, obovoid or nearly pear-shaped, attenuated at the base; membrane of a dirty violet-colour.—On the leaves of various species of Allium. Often plentiful. (Plate XIII. fig. 263.)
Peronospora violacea, Berk. Scabious Mould; fertile threads branched; acrospores sub-elliptical, violet-coloured. All that is known of this species is contained in the following note from the Rev. M. J. Berkeley:—“It grew on the petals of the common scabious. I have not found it again, and have either lost or mislaid my specimens. You may describe it as læte violacea; floccis ramosis; sporis subellipticis, violaceis. It is probably the Farinaria on Scabious of Sowerby.” (M. J. B.)
Peronospora sordida, Berk. Figwort Mould; forming broad, irregular, dirty, pallid spots on the under surface of the leaves; fertile threads loosely dichotomous above; tips forked, unequal; acrospores obovate, apiculate.—On leaves of Scrophularia. Jedburgh.
Peronospora sparsa, Berk. Rose Mould; fertile threads scattered, by no means torulose, ultimate branches scarcely uncinate, dichotomous, pallid grey; acrospores sub-elliptical.—On the under side of rose-leaves in conservatories.
Peronospora obliqua, Cooke. Dock Mould; threads of the mycelium slender; fertile threads fasciculate, erect, simple, rarely bifurcate, attenuated upwards; acrospores large, ellipsoid, attached obliquely near the base.—On the under surface of dock leaves. Winter and spring. Probably not uncommon. Brownish circular spots on the leaves indicate the presence of this mould, which is so minute that it might otherwise be overlooked. (Plate XVI. fig. 269.)
Mycelium arachnoid; perithecia globose, containing a single globose sporangium; appendages numerous, floccose.—Berk. Outl., p. 404.
Sphærotheca pannosa, Lév. Rose Blight; mycelium thickened, woolly, felted, persistent; conceptacles minute, globose, scattered; appendages floccose, white; sporangium many-spored.—On the branches, calyces, petioles, and leaves of Roses. Very common. (Plate XI. figs. 217, 218.)
Sphærotheca Castagnei, Lév. Hop Blight; on both surfaces; mycelium effuse, web-like, commonly evanescent; conceptacles minute, scattered, globose; appendages numerous, short, flexuose above; sporangium many-spored.—On the leaves of the Hop, Meadow-sweet, and various other plants. Common. (Plate XI. fig. 216.)
Perithecia hemispherical, at length depressed, seated on a persistent or evanescent membranaceo-granular receptacle; appendages straight, rigid, acicular, at length bent back.—Berk. Outl., p. 404.
Phyllactinia guttata, Lév. Hazel Blight; amphigenous; mycelium web-like, often evanescent; conceptacles large, scattered, hemispherical, at length depressed; appendages hyaline, rigid, simple; sporangia 4-20, containing 2-4 spores.—On the leaves of Hawthorn, Hazel, Ash, Elm, Alder, Beech, Birch, Oak, Hornbeam, and various other plants. Common. (Plate XI. figs. 219, 220.)
Mycelium floccose; perithecia globose; appendages rigid, simple, bifid or dichotomous, uncinate, at length bent upwards.—Berk. Outl., p. 404.
219Uncinula adunca, Lév. Willow Blight; mycelium variable; conceptacles scattered or gregarious, minute; appendages simple; sporangia 8-12, sub-pyriform, containing 4 spores.—On the leaves of Willows, Poplars, Birch, &c. Not uncommon. (Plate XI. figs. 221-224.)
Uncinula bicornis, Lév. Maple Blight; amphigenous; mycelium web-like, effuse, evanescent, or like a membrane and persistent; conceptacles large, hemispherical, at length depressed; appendages simple, bifid or dichotomous, uncinate; sporangia 8, sub-pyriform, containing 8 spores.—On the leaves of Maples. Common. (Plate XI. figs. 225-228.)
Mycelium arachnoid; appendages straight, dichotomous; branchlets swelling at the tip, or filiform.—Berk. Outl., p. 404.
Microsphæria Hedwigii, Lév. Mealy Guelder-rose Blight; hypophyllous; mycelium web-like, evanescent; conceptacles minute, globose, scattered; appendages few, very little longer than the diameter of the conceptacles; sporangia 4, ovate, containing 4 spores.—On leaves of mealy Guelder-rose. Near Greenhithe, Kent.
Microsphæria penicillata, Lév. Guelder-rose Blight; amphigenous; mycelium web-like, effuse, evanescent; conceptacles scattered, minute, globose; appendages 8-12, equal to the diameter of the conceptacle; sporangia 4, ovate, rostrate, containing 8 spores.—On leaves of Guelder-rose and Alder. Probably not uncommon. (Plate XI. fig. 234.)
Microsphæria Mougeotii, Lév. Tea-tree Blight; amphigenous; mycelium web-like, oftentimes persistent; conceptacles minute, scattered or gregarious, globose, at length depressed; appendages loosely dichotomous; sporangia 12-16, on a short pedicel, 2-spored.—On leaves of Lycium barbarum. Near Dartford, Kent. October, 1864.
Microsphæria Berberidis, Lév. Berberry Blight; amphigenous; mycelium web-like, oftentimes persistent; conceptacles scattered or gregarious, globose, minute; appendages few (5-10); branchlets long, divaricate, obtuse at their apices; sporangia 6, ovate, containing 6-8 spores.—On leaves of the common Berberry. Autumn. Frequent. (Plate XI. figs. 229-232.)
220Microsphæria grossulariæ, Lév. Gooseberry Blight; amphigenous; mycelium web-like, fugacious or persistent; conceptacles scattered or gregarious, globose, minute; appendages 10-15, vaguely dichotomous, ultimate branchlets bidentate; sporangia 4-8, ovate, containing 4-5 spores.—On Gooseberry-leaves. Autumn. Frequent.
Mycelium arachnoid; appendages floccose, simple or irregularly branched.—Berk. Outl., p. 404.
Erysiphe Linkii, Lév. Mugwort Blight; amphigenous; mycelium web-like; fugacious or persistent; conceptacles minute, globose, scattered, emersed; appendages white, interwoven with the mycelium; sporangia 8-20, pyriform, with elongated pedicels.—On leaves of Mugwort. Autumn. Frequent. (Plate XII. figs. 248, 249.)
Erysiphe lamprocarpa, Lév. Composite Blight; amphigenous; mycelium web-like, fugacious or persistent; conceptacles minute, globose, scattered, or gregarious; appendages coloured, interwoven with the mycelium; sporangia 8-16, shortly pedicellate.—On leaves of Salsify, Scorzonera, Plantain, &c. Autumn. Not uncommon. (Plate XII. figs. 250, 251.)
Erysiphe graminis, D.C. Grass Blight; amphigenous or epiphyllous; mycelium effuse, floccose, persistent; conceptacles large, gregarious or disseminated, hemispherical, at length depressed and semi-immersed; appendages simple or interwoven with the mycelium; sporangia 20-24, ovate, pedicellate, with 8 spores.—On leaves of Grasses. Autumn. Frequent. (Plate XI. figs. 235, 236.)
Erysiphe Martii, Lk. Pea Blight; amphigenous; mycelium web-like, very often evanescent, globose; appendages short, interwoven with the mycelium; sporangia 4-8, globose, pedicellate, with 4-8 spores.—On leaves of Peas, Beans, Umbelliferæ, and other plants. Autumn. Very common. (Plate XI. figs. 237-239.)
Erysiphe Montagnei, Lév. Burdock Blight; amphigenous or hypophyllous; mycelium web-like, evanescent; conceptacles minute, globose, gregarious or scattered; appendages distinct from the mycelium; sporangia 8, ovate, rostrate, with 2-3 spores.—On leaves of Burdock. Not uncommon.
221Erysiphe tortilis, Lk. Cornel Blight; hypophyllous; mycelium web-like, effuse, evanescent; conceptacles minute, globose; appendages ten times as long, free from the mycelium, flexuose; sporangia 4, ovate, rostrate, with 4 spores.—On leaves of the common Dogwood. Autumn. Frequent. (Plate XII. figs. 245, 246.)
Erysiphe communis, Schl. Buttercup Blight; hypophyllous; mycelium effuse, web-like, evanescent or persistent; conceptacles minute, globose, scattered or gregarious; appendages short; sporangia 4-8, ovate, rostrate, with 4-8 spores.—On leaves of various Ranunculaceæ, Leguminosæ, and other plants. Autumn. Very common. (Plate XI. figs. 240-242.)
Perithecium thin, brittle, mouthless; sporangia linear, containing dark lemon-shaped spores. Berk. Outl., p. 405.
Chætomium elatum, Kze. Straw-bristle Mould; perithecium sub-ovate, base radiato-fibrose, hairs of the vertex very long, interwoven, branched; spores broadly elliptic, apiculate at either end.—On mouldering straw, reeds, matting, &c. Very common. (Plate XII. figs. 257-259.)
Chætomium chartarum, Ehb. Paper-bristle Mould; perithecium subglobose, black, surrounded by a bright yellow spot; spores subglobose.—On paper. Stibbington, Hants. Rare. (Plate XII. figs. 252, 253.)
Chætomium glabrum, B. This species has never been described. It was recorded, by name only, in Berkeley’s Outlines, and, the specimens being mislaid, that gentleman is unable to describe it completely and correctly. “It grew abundantly on straw, and differed from Chætomium elatum in being perfectly free from hairs.”—(M. J. B.)—On damp straw.
Perithecium thin, free, mouthless, seated on loose, branched, conidiiferous threads; sporangia linear, containing dark elliptic spores.—Berk. Outl., p. 405.
Ascotricha chartarum, B. Paper Mildew; perithecium thin, olive-brown, seated on radiating flocci; sporangia linear, numerous; spores broadly elliptic, chocolate-coloured.—On white printed paper in a deal candle-box. King’s Cliffe. (Plate XII. figs. 254-256.)
Perithecia reticulated, vesicular, coloured, attached to mucedinous threads; sporangia delicate.—Berk. Outl., p. 405.
Eurotium herbariorum, Lk. Herbarium Mould; perithecium spherical, sub-depressed, yellow, seated upon radiating expanded, branched, intricate flocci.—On plants in herbaria, and various decaying substances. Very common. (Plate XII. figs. 260, 261.)
The following species, belonging to those sections of Microscopic Fungi which are included in this volume, have been found in Britain since the publication of the preceding Appendix:—
Peridermium columnare, Alb. and Schw. Simple, slender, naked, cylindrical, elongated, lacerated at the apex, white; sporidia orange.—On Picea. Watcombe, near Torquay. September, 1867. (E. Parfitt.)
Æcidium Orchidearum, Fiedl. Orchis Cluster-cup; spots large, pallid, orbicular or elongated; peridia circinating, semi-immersed; spores golden-yellow.—On Orchis latifolia. Dilham, Norfolk. June, 1866.
Puccinia discoidearum, Link. Southernwood Brand; spots obliterated; sori subrotund, minute, surrounded by the ruptured epidermis; sporidia brown, oblong or ovoid, somewhat rhomboidal, with both cells attenuated, and triangular; peduncles elongated.—On Artemisia maritima. Swanscombe Marshes. 1865.
Puccinia Virgaureæ, Lib. Golden-rod Brand; spots orbicular, pallid, then yellowish; sori blackish-brown, minute, punctiform, shining, clustered, nearly stellate, convex; sporidia oblong, subconstricted, yellowish-brown above, attenuated and yellowish-white below; peduncles short.—On leaves of Solidago virgaurea. Shere, Surrey. July and August, 1865. (Dr. Capron.)
Puccinia difformis, Kunze. Goosegrass Brand; spots yellowish; sori variable, compact, often in rings; epidermis bullate; sporidia obovate, obtuse, on short pedicels, brown.—On leaves of Galium aparine. July, 1866. Shere and Darenth 224Usually occurring on or near spots previously occupied by Æcidium.
Puccinia Apii, Corda. Celery Brand; sori large, confluent, red-brown, powdery; spores oblong, constricted, brown; epispore smooth, thick; pedicels short, attenuated.—On leaves of Celery. Autumn, 1866, 1867.
Puccinia Asari, Kunze. Asarabacca Brand; spots obliterated; sori small, subglobose, crowded or circinating, ultimately confluent, surrounded by the ruptured epidermis, usually forming a large, roundish, convex, pulverulent mass; sporidia brown, elliptic, constricted.—On leaves of Asarum Europæum. Bettws-y-coed, N. Wales. July, 1866.
Puccinia Mœhringiæ, Fuckel. It is very doubtful whether this deserves to rank as a species distinct from P. Lychnidearum, Lk. I think not. The sporidia are rather shorter than in the last-named species, and the pedicels are very long; otherwise it does not appear to differ from the typical form.—On Mœhringia trinervis. Near Worcester. June, 1867. (Dr. Holl.)
Puccinia fallens, Cooke. Vetch Brand; sori few and small, scattered, intermixed with pustules of Trichobasis; sporidia obovate, on rather long pedicels, of a tawny colour, and slightly constricted at the septum; epispore smooth.—On leaves of Vicia sepium. Liverpool. Autumn, 1865. (R. McLeod.)
Uredo Orchidis, Mart. Orchis Uredo; amphigenous; spots reddish-brown; sori subrotund, arranged in circles, often confluent; sporidia subglobose, golden-yellow.—On Listera ovata and Orchis latifolia. Crosby, near Liverpool. May, June, 1865. (R. G. McLeod.)
Uredo Empetri, D.C. Crowberry Uredo; hypogenous; spots obliterated; sori oval, scattered: the epidermis at first convex, afterwards ruptured and concave; sporidia ovoid or subglobose, bright yellow.—On Empetrum nigrum. Near Llanderfel, N. Wales. May, 1865.
Uredo Euonymi, Mart. Spindle Uredo; spots yellowish; sori roundish, circinating, often confluent; epidermis erumpent; sporidia ovoid and slightly coherent, tawny-yellow.—On leaves of Euonymus Europæus. Darenth Wood, Kent. August, 1864. Rare.
Uredo Tropæoli, Desmz. Nasturtium Uredo: hypogenous; spots pale-yellow; sori minute, roundish, scattered or 225confluent; sporidia ovoid or subglobose, orange.—On leaves of Tropæolum aduncum. Shere. October, 1865. (Dr. Capron.)
Trichobasis fallens, Cooke. Clover Rust; spots obliterated; sori amphigenous, numerous, scattered, subrotund, brown, surrounded by the remains of the ruptured epidermis; spores sub-ovate; pedicels short, hyaline, evanescent; epispore verrucose. Uredo fallens, Desmz.—On leaves of Clover, &c. September. Not uncommon.
Uromyces graminum, Cooke. Cocksfoot Rust; epiphyllous; sori oblong or confluent and linear, convex, black and shining, so as easily to be confounded on casual observation with Dothidea graminis, at length bursting longitudinally; sporidia subglobose or ovate, tawny, with hyaline pedicels of variable length.—On leaves of Dactylis glomerata. Shere. October, 1865. (Dr. Capron.)
Uromyces concentrica, Lév. Described at p. 208 as Trichobasis Scillarum, B.; but it appears to be a good Uromyces, and has been placed in that genus by Lévielle.
Uromyces sparsa, Lév. Sandwort Rust; spots pallid; sori subrotund and oval, amphigenous and cauline; epidermis erumpent; sporidia ovoid, brownish; peduncles thickened, short.—On Spergularia rubra. Swanscombe Marshes. June, 1865.
Uromyces Polygoni, Fuckel. Knotgrass Rust; cauline; sori elongated and confluent, convex, surrounded by the remains of the ruptured epidermis; sporidia subglobose or globose, smooth, yellowish-brown; pedicels very long, thickened, hyaline, persistent. Capitularia Polygoni, Rabh.—On the stems of Polygonum aviculare. October. Common.
Cystopus spinulosus, De Bary. Thistle (White) Rust; conidia in time much elongated; sori erumpent on both surfaces of the leaves, white; oospores globose; epispore brown; tubercles minute, solid, very prominent, often acute and spinulose.—On Thistles. Bungay, Suffolk. September, 1865.
Peronospora candida, Fuckel. Primrose Mould; densely cæspitose, white; threads erect, dichotomously branched; ultimate branchlets short, spreading; sporidia ovate.—On leaves of Primrose. Near Corwen, N. Wales. July, 1866.
Mycelium effuse, web-like, evanescent; conceptacles sphærical, containing one subglobose 8-spored sporangium; spores ovate; appendages few, dichotomous, thickened at their extremities, hyaline.
Podosphæra Kunzei, Lév. Plum Blight; amphigenous; conceptacles minute, scattered, globose; appendages three times the length of the diameter of the conceptacles.—On leaves of Prunus domestica. Shere, Surrey. Sept., 1865. (Dr. Capron.)
Podosphæra clandestina, Lév. Hawthorn Blight; amphigenous; conceptacles minute, globose, scattered; appendages (8-10) equal in length to the diameter of the conceptacles; branches short and rounded at their extremities.—On leaves of the Hawthorn. Upper Holloway. October, 1864; Shere, Surrey, September, 1865. (E. C.)
Uncinula Wallrothii, Lév. Sloe Blight; amphigenous; mycelium web-like, evanescent; conceptacles minute, scattered; sporanges 12-16, pear-shaped, 6-spored; appendages numerous, twice the length of the diameter of the conceptacles.—On leaves of the Sloe, Prunus spinosa. Shere, Surrey. October, 1865. (Dr. Capron.)
Microsphæria comata, Lév. Spindle Blight; hypophyllous; mycelium web-like, fugacious; conceptacles scattered, minute, globose; sporanges 8, ovate, with a beak-like termination at their base, 4-spored; appendages few, six times the length of the diameter of the conceptacles.—On Euonymus Europæus. Shere, Surrey. August, 1865. (Dr. Capron.)
Erysiphe horridula, Lév. Borage Blight; amphigenous; mycelium web-like, sometimes persistent; conceptacles minute, globose, scattered or clustered; sporanges 20-24, oblong-ovate, attenuated downwards, containing 3-4 spores; appendages short, flexuose, and bent upwards.—On leaves of Lycopsis arvensis. Shere. October, 1865. (Dr. Capron.)
Chætomium murorum, Corda. Wall Bristle-mould; gregarious, glaucous, then blackish; perithecium globose, brown; hairs circinate, erect, septate, pulverulent; spores oblong, yellowish. On plaster. Rare. (M. J. B.)
[The following species have been added to the British Flora since the publication of the Appendix.]
Æcidium Parnassiæ, Grav. Parnassia Cluster-cups; hypophyllous; spots pallid; peridia in subrotund patches, irregularly disposed, tawny-yellow, between urceolate and concave; the margin thick and nearly entire; spores pallid. Duby. Bot. Gall. ii. p. 904.—On leaves of Parnassia palustris, near Glasgow. (Dr. Greville.) The original specimen is in the Edinburgh Herbarium.
Æcidium Statices, Desm. Sea-Lavender Cluster-cups; hypophyllous; spots subrotund, or confluent and irregular, purple; peridia in subrotund circinating clusters, sometimes irregularly disposed on the nerves and petioles; urceolate; margin lacerated, white; spores orange. Desm. Exs. No. 132. Cooke, Exs. No. 444.—On leaves and petioles of Statices. June, July. Fleetwood, 1859 (Rev. A. Bloxam). Walney Island, Lancashire, 1871 (C. Bailey). Near Basingstoke, 1871 (R. S. Hill). Near Chichester, 1872 (F. V. Paxton).
Chætomium funicolum, Cooke. Twine Bristle-mould; perithecia scattered, sub-ovate, black; hairs of the vertex very long, dichotomous or simple, erect, slender, acute, black; sporidia lemon-shaped, dingy brown.—On twine suspended in a vessel containing water at the British Museum. (W. Carruthers, F.R.S.) March, 1872. This species is most closely allied to C. elatum, but much smaller and neater. It is wholly black, and without the fibrous base of C. elatum. The hairs are more delicate, not having half the diameter, and the sporidia are scarcely more than half as long or broad. A species of Polyactis afterwards made its appearance on the same mycelium on some portion of the twine left behind with a few immature perithecia.
| Plate | fig. | |
| I. | 1. | Goatsbeard with its cluster-cups (Æcidium Tragopogonis). |
| 〃 | 2. | Fragment of same, slightly magnified. |
| 〃 | 3. | Section of cluster-cup, further magnified. |
| 〃 | 4. | Leaf of wood-anemone with its cluster-cups (Æcidium leucospermum). |
| 〃 | 5. | Portion of same, slightly magnified. |
| 〃 | 6. | Anemone cluster-cups, further magnified. |
| 〃 | 7. | Leaf of Berberry with cluster-cups (Æcidium Berberidis). |
| 〃 | 8. | Cluster of cups from the Berberry, as seen with a lens. |
| 〃 | 9. | Cluster-cups of Berberry, magnified. |
| 〃 | 10. | Nettle-stem distorted by growth of cluster-cups (Æcidium Urticæ). |
| 〃 | 11. | Nettle cluster-cups, magnified. |
| II. | 12. | Leaf of Pilewort with groups of cluster-cups (Æcidium Ranunculacearum). |
| 〃 | 13. | Group of cluster-cups from Pilewort, as seen with a lens. |
| 〃 | 14. | Section of Pilewort cluster-cups, magnified. |
| 〃 | 15. | Bedstraw cluster-cups (Æcidium Galii) on the Great Hedge Bedstraw. |
| 〃 | 16. | Tip of leaflet with cluster of cups, viewed through a lens. |
| 〃 | 17. | Bedstraw cluster-cup from same, magnified. |
| 〃 | 18. | Leaflets of Mountain-ash with horn-shaped cluster-cups (Rœstelia cornuta). |
| 〃 | 19. | Group of Mountain-ash cluster-cups, magnified. |
| 〃 | 20. | Pear-leaf with its cluster-cups (Rœstelia cancellata). |
| 〃 | 21. | Cluster-cup from the same, magnified. |
| 〃 | 22. | Leaves and fruit of Hawthorn with lacerated cluster-cups (Rœstelia lacerata). |
| 〃 | 23. | Elongated cells forming the walls of the cluster-cups, × 250. |
| 〃 | 24. | Single cell, more highly magnified, with its parallel striæ. |
| 230〃 | 25. | Cluster-cups from fruit of Hawthorn, magnified. |
| 〃 | 26. | Section of same, further magnified. |
| 〃 | 27. | Fir-leaves bearing cluster-cups (Peridermium Pini). |
| 〃 | 28. | Isolated cup of same, magnified. |
| III. | 29. | Many-jointed fruit of Burnet chain-brand (Xenodochus carbonarius), magnified. |
| 〃 | 30. | Leaflets of Burnet with rust on the lower, and brand (Aregma acuminatum) on the upper leaflets. |
| 〃 | 31. | Spores of Burnet rust (Lecythea Poterii) × 230. |
| 〃 | 32. | Fruit of Burnet brand (Aregma acuminatum). |
| 〃 | 33. | Leaf of Barren Strawberry with rust and brand intermixed. |
| 〃 | 34. | Spores of Barren Strawberry rust × 230. |
| 〃 | 35. | Fruit of Strawberry brand (Aregma obtusatum) × 300. |
| 〃 | 36. | Leaflet of Rose with its rust and brand intermixed. |
| 〃 | 37. | Spores of Rose rust (Lecythea Rosæ) × 230. |
| 〃 | 38. | Fruit of Rose brand (Aregma mucronatum) × 300. |
| 〃 | 39. | Leaflet of Bramble, with its rust and brand intermixed. |
| 〃 | 40. | Spores of Bramble rust (Lecythea Ruborum) × 230. |
| 〃 | 41. | Fruit of Bramble brand (Aregma bulbosum) × 300. |
| 〃 | 42. | Leaflet of Raspberry with its brand. |
| 〃 | 43. | Fruit of Raspberry brand (Aregma gracile) × 230. |
| 〃 | 44. | Cluster of fruits of Rose brand (Aregma mucronatum) × 230. |
| 〃 | 45. | Fruit of Bramble brand (Aregma bulbosum) in active germination, with sporidia at the tips of the threads × 300. |
| 〃 | 46. | Fruit of Bramble brand (Aregma bulbosum) ruptured, with inner cell escaping × 250 (F. Currey). |
| 〃 | 47. | Portion of leaflet of Meadow-sweet with its brand. |
| 〃 | 48. | Spores of Meadow-sweet brand (Triphragmium ulmariæ) × 300. |
| 〃 | 49. | Spore of Meadow-sweet brand (Triphragmium ulmariæ) in germination, with sporidia borne on the germ-tubes × 300 (Tulasne). |
| 〃 | 50. | Sprig of Thesium humifusum bearing Bastard-toadflax cluster-cups (Æcidium Thesii). |
| 〃 | 51. | Portion of leaf of same, with cluster-cups, enlarged. |
| 〃 | 52. | Leaflet of Trientalis Europæa with its smut (Tuburcinia Trientalis). |
| 〃 | 53. | Spores of the same × 320. |
| 〃 | 54. | Spores of Potato smut (Tuburcinia scabies) × 320. (Berkeley.) |
| 〃 | 55. | Leaflet of Alexanders (Smyrnium olusatrum) with its brand. |
| 〃 | 56. | Spores of Alexanders brand (Puccinia Smyrnii) × 320. |
| 231IV. | 57. | Wheat-straw attacked by mildew (Puccinia graminis). |
| 〃 | 58. | Cluster of spores of corn-mildew, magnified (Bauer). |
| 〃 | 59. | Single spore of corn-mildew (Puccinia graminis) × 300. |
| 〃 | 60. | Portion of blade of grass with coronated mildew (Puccinia coronata). |
| 〃 | 61. | Portion of same enlarged, showing the pustules, or sori. |
| 〃 | 62. | Spore of coronated mildew (Puccinia coronata) × highly. |
| 〃 | 63. | Portion of leaf of Spear-thistle with its brand (Puccinia syngenesiarum). |
| 〃 | 64. | Spores of Thistle brand (Puccinia syngenesiarum) × 320. |
| 〃 | 65. | Leaf of Wood-anemone with brand (Puccinia anemones). |
| 〃 | 66. | Spore of Anemone brand (Puccinia anemones) × 320. |
| 〃 | 67. | Leaf of Centaurea nigra with brand. |
| 〃 | 68. | Spores of Composite-brand (Puccinia compositarum) × 320. |
| 〃 | 69. | Leaf of Mint with Mint brand (Puccinia menthæ). |
| 〃 | 70. | Spore of Mint brand (Puccinia menthæ) × 350. |
| 〃 | 71. | Portion of leaf of Earth-nut with brand (Puccinia umbelliferarum). |
| 〃 | 72. | Spore of Earth-nut brand (Puccinia umbelliferarum) × 320. |
| 〃 | 73. | Leaf of Ground-ivy with its brand (Puccinia Glechomatis). |
| 〃 | 74. | Spore of Ground-ivy brand (Puccinia Glechomatis) × 320. |
| 〃 | 75. | Pustule of brand surrounded by the ruptured epidermis, magnified. |
| 〃 | 76. | Portion of leaf and stem of Goatsbeard with brand. |
| 〃 | 77. | Spores of the same × 320. |
| 〃 | 78. | Portion of leaf of Willow-herb with its brand (Puccinia pulverulenta). |
| 〃 | 79. | Spores of Willow-herb brand (Puccinia pulverulenta) × 320. |
| 〃 | 80. | Leaf of Pennywort with brand (Puccinia umbilici). |
| 〃 | 81. | Spores of Pennywort-brand (Puccinia umbilici) × 320. |
| 〃 | 82. | Portion of leaf of Dandelion with its brand. |
| 〃 | 83. | Spores of Variable-brand (Puccinia variabilis) × 320. |
| V. | 84. | Grain of Wheat infected with Fœtid smut (Tilletia caries). |
| 〃 | 85. | Longitudinal section of the same. |
| 〃 | 86. | Spores of Fœtid smut (Tilletia caries) mixed with delicate branching threads × 320. |
| 232〃 | 87. | Spores of Fœtid smut in germination × highly. |
| 〃 | 88. | Sporidia of the first order borne on the germinating tubes of the fœtid smut (Tilletia caries) × highly. |
| 〃 | 89. | Sporidia of the first order, showing their transverse connection, × highly. |
| 〃 | 90. | Sporidia of the first order, producing sporidia of the second order, × highly. |
| 〃 | 91. | Sporidium of the second order in active germination. |
| 〃 | 92. | Deformed flower-head of Goatsbeard infested with smut (Ustilago receptaculorum). |
| 〃 | 93. | Floret removed, sprinkled with spores of the smut, enlarged. |
| 〃 | 94. | Spores of Goatsbeard smut (Ustilago receptaculorum) × 460. |
| 〃 | 95. | The same in active germination × 460 (Tulasne). |
| 〃 | 96. | Beaksedge (Rhyncospora alba) with its smut (Ustilago Montagnei). |
| 〃 | 97. | Spores of Ustilago Montagnei × 460. |
| 〃 | 98. | Ear of Barley infected with smut (Ustilago segetum). |
| 〃 | 99. | Spores of Corn-smut (Ustilago segetum) × 460. |
| 〃 | 100. | Grass with its smut (Ustilago hypodytes). |
| 〃 | 101. | Spores of Grass-smut (Ustilago hypodytes). |
| 〃 | 102. | Flowers of Bladder-campion with anther smut (Ustilago antherarum). |
| 〃 | 103. | Anther distorted by smut (Ustilago antherarum). |
| 〃 | 104. | Spores of Anther smut (Ustilago antherarum) × 460. |
| 〃 | 105. | Portion of leaf of water grass with Elongated smut (Ustilago longissima). |
| 〃 | 106. | Pustule of same, enlarged. |
| 〃 | 107. | Spores of Elongated smut (Ustilago longissima) × very highly. |
| 〃 | 108. | Spores of Maize smut (Ustilago maydis) × 460. |
| VI. | 109. | Sedge (Carex recurva) with Sedge smut (Ustilago urceolorum). |
| 〃 | 110. | Single fruit covered with the smut. |
| 〃 | 111. | Spores of Sedge smut (Ustilago urceolorum) × 460. |
| 〃 | 112. | Flower of Polygonum persicaria distorted by Utricle smut (Ustilago utriculosa). |
| 〃 | 113. | Section of the same (Tulasne). |
| 〃 | 114. | Inflorescence of Polygonum hydropiper with Utricle smut (Ustilago utriculosa). |
| 〃 | 115. | Section of infected flower of Polygonum hydropiper. |
| 〃 | 116. | Spores of Utricle smut (Ustilago utriculosa) × 460. |
| 〃 | 117. | Leaf of Cocksfoot, grass with smut (Ustilago Salveii). |
| 〃 | 118. | Portion of leaf, showing pustules, × slightly. |
| 〃 | 119. | Spores of Cocksfoot smut (Ustilago Salveii) × 320. |
| 〃 | 120. | Grass stem with Banded smut (Ustilago grammica). |
| 233〃 | 121. | Portion of same, showing arrangement of the pustules, × slightly. |
| 〃 | 122. | Spores of banded smut (Ustilago grammica) × 320. |
| 〃 | 123. | Inflorescence of Scabious with Floret smut (Ustilago flosculorum). |
| 〃 | 124. | Floret occupied by the smut. |
| 〃 | 125. | Spores of Floret smut (Ustilago flosculorum) × 320. |
| 〃 | 126. | Sedge (Carex riparia) attacked by Olive smut (Ustilago olivacea). |
| 〃 | 127. | Spores of Olive smut (Ustilago olivacea) × 460. |
| 〃 | 128. | Reed stem with smut (Ustilago typhoides). |
| 〃 | 129. | Spores of Reed smut (Ustilago typhoides) × 320. |
| 〃 | 130. | Leaf of Periwinkle with its rust (Trichobasis vincæ). |
| 〃 | 131. | Spores of Periwinkle rust (Trichobasis vincæ) × 320. |
| 〃 | 132. | Spores of Periwinkle brand (Puccinia vincæ) × 320. |
| VII. | 133. | Leaf of Mercury with its rust (Uredo confluens). |
| 〃 | 134. | Spores of Mercury rust (Uredo confluens) × 320. |
| 〃 | 135. | Leaf of Enchanter’s Nightshade with its rust. |
| 〃 | 136. | Spores of Nightshade rust (Uredo Circææ) × 320. |
| 〃 | 137. | Leaf of Sorrel with Twin-faced rust (Uredo bifrons). |
| 〃 | 138. | Pustule of Twin-faced rust seated on a coloured spot × slightly. |
| 〃 | 139. | Spores of the same rust × 320. |
| 〃 | 140. | Leaf of Wheat with Corn rust (Trichobasis rubigo-vera). |
| 〃 | 141. | Pustules of the same rust × slightly. |
| 〃 | 142. | Spores of Round corn rust × 320. |
| 〃 | 143. | Tuft of spores of Elongated corn rust (Trichobasis linearis) magnified. |
| 〃 | 144. | Spores of Elongated corn rust (Trichobasis linearis) × 320. |
| 〃 | 145. | Leaf of Groundsel with its rust (Trichobasis Senecionis). |
| 〃 | 146. | Spores of Groundsel rust (Trichobasis Senecionis) × 320. |
| 〃 | 147. | Wild Rose with Golden rust (Uromyces Ulmariæ). |
| 〃 | 148. | Spores of Golden rust × 320. |
| 〃 | 149. | Leaf of Vetch with Long-stemmed rust (Uromyces appendiculata). |
| 〃 | 150. | Spores of the same × 460. |
| 〃 | 151. | Leaf of Common thistle with Sweet-smelling rust (Trichobasis suaveolens). |
| 〃 | 152. | Spores of Sweet rust (Trichobasis suaveolens) × 320. |
| 〃 | 153. | Spermogone of Sweet rust, from common thistle, × highly (De Bary). |
| 〃 | 154. | Clover leaf with Short-stemmed rust (Uromyces apiculosa). |
| 〃 | 155. | Spores of Short-stemmed rust × 320. |
| 〃 | 156. | Leaf of Pilewort with its rust (Uromyces Ficariæ). |
| 〃 | 157. | Spores of Pilewort rust (Uromyces Ficariæ) × 320. |
| 234〃 | 158. | Portion of leaf of Luzula with Oblong rust (Trichobasis oblongata). |
| 〃 | 159. | Spores of Oblong rust (Trichobasis oblongata) × 320. |
| VIII. | 160. | Leaf of Sallow with its rust (Lecythea capræarum). |
| 〃 | 161. | Spores of Sallow rust (Lecythea capræarum) × 320. |
| 〃 | 162. | Bramble leaf with Ringed rust (Lecythea gyrosa). |
| 〃 | 163. | Pustule of Ringed rust (Lecythea gyrosa) × slightly. |
| 〃 | 164. | Spores of Ringed rust (Lecythea gyrosa) × 320. |
| 〃 | 165. | Purging Flax with its rust (Lecythea Lini). |
| 〃 | 166. | Pustule of the same × slightly. |
| 〃 | 167. | Spores of Flax rust (Lecythea Lini) × 320. |
| 〃 | 168. | Leaf of Flukewort with its rust (Trichobasis hydrocotyles). |
| 〃 | 169. | Spores of Trichobasis hydrocotyles × 320. |
| 〃 | 170. | Portion of leaf of Sedge with its rust (Trichobasis caricina). |
| 〃 | 171. | Spores of Sedge rust (Trichobasis caricina) × 320. |
| 〃 | 172. | Whorl of leaves of Hedge Bedstraw with Bedstraw brand (Puccinia Galii). |
| 〃 | 173. | Spores of Bedstraw brand (Puccinia Galii) × 320. |
| 〃 | 174. | Leaf of Tutsan with St. John’s-wort rust (Uredo hypericorum). |
| 〃 | 175. | Spores of St John’s-wort rust (Uredo hypericorum) × 320. |
| 〃 | 176. | Leaves of Cow-wheat with its rust (Coleosporium rhinanthacearum). |
| 〃 | 177. | Spores of Cow-wheat rust (Coleosporium rhinanthacearum) × 320. |
| 〃 | 178. | Portion of leaf of Sow-thistle with its rust (Coleosporium sonchi-arvensis). |
| 〃 | 179. | Spores of Sow-thistle rust × highly (De Bary). |
| 〃 | 180. | Portion of Coltsfoot leaf with its rust (Coleosporium Tussilaginis). |
| 〃 | 181. | Spores of Coltsfoot rust × highly. |
| 〃 | 182. | Group of spores of Coltsfoot rust in situ × highly (Tulasne). |
| IX. | 183. | Leaf of Buttercup with Buttercup smut (Polycystis pompholygodes). |
| 〃 | 184. | Fruit of Buttercup smut (Polycystis pompholygodes) × 460. |
| 〃 | 185. | Violet leaf distorted by Violet smut (Polycystis Violæ). |
| 〃 | 186. | Fruit of Violet smut (Polycystis Violæ) × 460. |
| 〃 | 187. | Portion of Rye-leaf with Rye smut (Polycystis parallela). |
| 〃 | 188. | Fruit of Rye smut (Polycystis parallela) ×460. |
| 〃 | 189. | Dead Birch leaf with Birch wedge-rust (Melampsora betulina). |
| 〃 | 190. | Winter spores of (Melampsora betulina) × 460. |
| 235〃 | 191. | Portion of Sallow leaf with Willow wedge-rust (Melampsora salicina). |
| 〃 | 192. | Winter spores of Melampsora salicina × 460. |
| 〃 | 193. | Leaves of Spurge with Spurge wedge-rust (Melampsora Euphorbiæ). |
| 〃 | 194. | Winter spores of Melampsora Euphorbiæ × highly. |
| 〃 | 195. | Portion of Poplar-leaf with Poplar wedge-rust (Melampsora populina). |
| 〃 | 196. | Winter spores of Melampsora populina × 460. |
| 〃 | 197. | Winter spores of Melampsora betulina in active germination—a sporidia × 466 (Tulasne). |
| X. | 198. | Fruit of Shepherd’s-Purse with White rust (Cystopus candidus). |
| 〃 | 199. | Portion of Cabbage-leaf with White rust (Cystopus candidus). |
| 〃 | 200. | Conidia of White rust (Cystopus candidus) × 360. |
| 〃 | 201. | Portion of Goatsbeard-leaf with White rust (Cystopus cubicus). |
| 〃 | 202. | Conidia of Goatsbeard white rust (Cystopus cubicus) × 360. |
| 〃 | 203. | Portion of Mycelium producing the first of a chain of conidia × 400. |
| 〃 | 204. | Mycelium of White rust with sucker-like processes × 360. |
| 〃 | 205. | Mycelium of White rust with nascent oogonia × 360. |
| 〃 | 206. | Oogonium of Crucifer white rust (Cystopus candidus) × 400 (De Bary). |
| 〃 | 207. | The same, further advanced, ruptured, with zoospores, × 400. |
| 〃 | 208. | Free zoospores of White rust, with their ciliæ, × 400 (De Bary). |
| 〃 | 209. | Resting zoospores in germination × 400 (De Bary). |
| 〃 | 210. | Oogonium of Goatsbeard white rust (Cystopus cubicus) × 400 (De Bary). |
| 〃 | 211. | Oogonium of Sandwort mould (Peronospora arenariæ) × 400 (De Bary). |
| 〃 | 212. | Oogonium of Pea mould (Peronospora viciæ) × 400 (De Bary). |
| 〃 | 213. | Oogonium of Veronica mould (Peronospora grisea) × 400 (De Bary). |
| 〃 | 214. | Oogonium of Spinach mould (Peronospora effusa) × 400 (De Bary). |
| 〃 | 215. | Leaf of Goosefoot with mould (Peronospora effusa). |
| XI. | 216. | Conceptacle of Hop mildew (Sphærotheca castagnei) × 80. |
| 〃 | 217. | Conceptacle of Rose blight (Sphærotheca pannosa) × 80. |
| 〃 | 218. | Sporangium of Sphærotheca pannosa × highly. |
| 236〃 | 219. | Conceptacle of Hazel blight (Phyllactinia guttata) × 80. |
| 〃 | 220. | Sporangium of Phyllactinia guttata × highly. |
| 〃 | 221. | Portion of leaf of Willow with Willow blight (Uncinula adunca). |
| 〃 | 222. | Conceptacle of Willow blight (Uncinula adunca) × 80. |
| 〃 | 223. | Tip of one of the appendages × highly. |
| 〃 | 224. | Sporangium of Willow blight (Uncinula adunca) × highly. |
| 〃 | 225. | Conceptacle of Maple blight (Uncinula bicornis) × 80. |
| 〃 | 226. | Tip of one of the appendages of Uncinula bicornis × highly. |
| 〃 | 227. | Furcate tip of one of the appendages of the Maple blight (Uncinula bicornis) × highly. |
| 〃 | 228. | Sporangium of Uncinula bicornis × highly. |
| 〃 | 229. | Leaf of Berberry with Berberry blight (Microsphæria Berberidis). |
| 〃 | 230. | Conceptacle of Berberry blight × 80. |
| 〃 | 231. | Tip of one of the appendages of Berberry blight (Microsphæria Berberidis) × highly. |
| 〃 | 232. | Sporangium of Berberry blight × highly. |
| 〃 | 233. | Tip of appendage of a continental species of blight (Microsphæria Ehrenbergii) × highly (Léveillé). |
| 〃 | 234. | Tip of appendage of Alder blight (Microsphæria penicillata) × highly. |
| 〃 | 235. | Portion of Grass leaf with blight (Oidium monilioides). |
| 〃 | 236. | Tuft of conidia of Oidium monilioides × 120. |
| 〃 | 237. | Leaflet of Garden Pea with Pea blight (Erysiphe Martii). |
| 〃 | 238. | Conceptacle of Pea blight (Erysiphe Martii) × 80. |
| 〃 | 239. | Sporangium of Erysiphe Martii × highly. |
| 〃 | 240. | Conceptacle of Buttercup blight (Erysiphe communis) × 80. |
| 〃 | 241. | Sporangium of Erysiphe communis × highly. |
| 〃 | 242. | Sucker from the mycelium of Erysiphe communis, magnified. |
| XII. | 243. | Conceptacle of Mealy Guelder-rose blight (Microsphæria Hedwigii) × 80. |
| 〃 | 244. | Sporangium of same, containing spores, × highly. |
| 〃 | 245. | Conceptacle of Cornel blight (Erysiphe tortilis) × 80. |
| 〃 | 246. | Sporangium of same, containing spores, × highly. |
| 〃 | 247. | Tip of appendage of Microsphæria Hedwigii × highly. |
| 〃 | 248. | Sporangium of Mugwort blight (Erysiphe Linkii), containing spores, × highly. |
| 〃 | 249. | Conceptacle of Mugwort blight (Erysiphe Linkii) × 80. |
| 〃 | 250. | Sporangium and spores of Plantain blight (Erysiphe lamprocarpa) × highly. |
| 237〃 | 251. | Conceptacle of Plantain blight (Erysiphe lamprocarpa) × 80. |
| 〃 | 252. | Conceptacle of Paper bristle-mould (Chætomium chartarum) magnified. |
| 〃 | 253. | Sporidium of the same, further magnified. |
| 〃 | 254. | Conceptacle of Paper mildew (Ascotricha chartarum) magnified (Berkeley). |
| 〃 | 255. | Portion of thread of same with conidia, further magnified (Berkeley). |
| 〃 | 256. | Sporangium of same, containing spores (Berkeley). |
| 〃 | 257. | Piece of straw with Bristle mould (Chætomium elatum). |
| 〃 | 258. | Conceptacle of same, slightly magnified. |
| 〃 | 259. | Section of same, further magnified. |
| 〃 | 260. | Dead leaf over-run with Herbarium mildew (Eurotium herbariorum). |
| 〃 | 261. | Conceptacles of the same, seated on their mycelium, magnified. |
| XIII. | 262. | Fertile thread of Turnip mould (Peronospora parasitica). |
| 〃 | 263. | Fertile thread of Onion mould (Peronospora Schleideniana). |
| XIV. | 264. | Fertile thread of Potato mould (Peronospora infestans). |
| 〃 | 265. | Fertile thread of Lettuce mould (Peronospora gangliformis). |
| XV. | 266. | Fertile thread of Pea mould (Peronospora Viciæ). |
| 〃 | 267. | Fertile thread of Anemone mould (Peronospora curta). |
| XVI. | 268. | Fertile thread of Sandwort mould (Peronospora arenariæ). |
| 〃 | 269. | Fertile thread of Dock mould (Peronospora obliqua) × 320. |
PAGE
End of the Project Gutenberg EBook of Rust, Smut, Mildew, & Mold, by
Mordecai Cubitt Cooke
*** END OF THIS PROJECT GUTENBERG EBOOK RUST, SMUT, MILDEW, & MOLD ***
***** This file should be named 63416-h.htm or 63416-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/6/3/4/1/63416/
Produced by deaurider, Barry Abrahamsen, and the Online
Distributed Proofreading Team at https://www.pgdp.net (This
file was produced from images generously made available
by The Internet Archive)
Updated editions will replace the previous one--the old editions will
be renamed.
Creating the works from print editions not protected by U.S. copyright
law means that no one owns a United States copyright in these works,
so the Foundation (and you!) can copy and distribute it in the United
States without permission and without paying copyright
royalties. Special rules, set forth in the General Terms of Use part
of this license, apply to copying and distributing Project
Gutenberg-tm electronic works to protect the PROJECT GUTENBERG-tm
concept and trademark. Project Gutenberg is a registered trademark,
and may not be used if you charge for the eBooks, unless you receive
specific permission. If you do not charge anything for copies of this
eBook, complying with the rules is very easy. You may use this eBook
for nearly any purpose such as creation of derivative works, reports,
performances and research. They may be modified and printed and given
away--you may do practically ANYTHING in the United States with eBooks
not protected by U.S. copyright law. Redistribution is subject to the
trademark license, especially commercial redistribution.
START: FULL LICENSE
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full
Project Gutenberg-tm License available with this file or online at
www.gutenberg.org/license.
Section 1. General Terms of Use and Redistributing Project
Gutenberg-tm electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or
destroy all copies of Project Gutenberg-tm electronic works in your
possession. If you paid a fee for obtaining a copy of or access to a
Project Gutenberg-tm electronic work and you do not agree to be bound
by the terms of this agreement, you may obtain a refund from the
person or entity to whom you paid the fee as set forth in paragraph
1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this
agreement and help preserve free future access to Project Gutenberg-tm
electronic works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the
Foundation" or PGLAF), owns a compilation copyright in the collection
of Project Gutenberg-tm electronic works. Nearly all the individual
works in the collection are in the public domain in the United
States. If an individual work is unprotected by copyright law in the
United States and you are located in the United States, we do not
claim a right to prevent you from copying, distributing, performing,
displaying or creating derivative works based on the work as long as
all references to Project Gutenberg are removed. Of course, we hope
that you will support the Project Gutenberg-tm mission of promoting
free access to electronic works by freely sharing Project Gutenberg-tm
works in compliance with the terms of this agreement for keeping the
Project Gutenberg-tm name associated with the work. You can easily
comply with the terms of this agreement by keeping this work in the
same format with its attached full Project Gutenberg-tm License when
you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are
in a constant state of change. If you are outside the United States,
check the laws of your country in addition to the terms of this
agreement before downloading, copying, displaying, performing,
distributing or creating derivative works based on this work or any
other Project Gutenberg-tm work. The Foundation makes no
representations concerning the copyright status of any work in any
country outside the United States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other
immediate access to, the full Project Gutenberg-tm License must appear
prominently whenever any copy of a Project Gutenberg-tm work (any work
on which the phrase "Project Gutenberg" appears, or with which the
phrase "Project Gutenberg" is associated) is accessed, displayed,
performed, viewed, copied or distributed:
This eBook is for the use of anyone anywhere in the United States and
most other parts of the world at no cost and with almost no
restrictions whatsoever. You may copy it, give it away or re-use it
under the terms of the Project Gutenberg License included with this
eBook or online at www.gutenberg.org. If you are not located in the
United States, you'll have to check the laws of the country where you
are located before using this ebook.
1.E.2. If an individual Project Gutenberg-tm electronic work is
derived from texts not protected by U.S. copyright law (does not
contain a notice indicating that it is posted with permission of the
copyright holder), the work can be copied and distributed to anyone in
the United States without paying any fees or charges. If you are
redistributing or providing access to a work with the phrase "Project
Gutenberg" associated with or appearing on the work, you must comply
either with the requirements of paragraphs 1.E.1 through 1.E.7 or
obtain permission for the use of the work and the Project Gutenberg-tm
trademark as set forth in paragraphs 1.E.8 or 1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any
additional terms imposed by the copyright holder. Additional terms
will be linked to the Project Gutenberg-tm License for all works
posted with the permission of the copyright holder found at the
beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including
any word processing or hypertext form. However, if you provide access
to or distribute copies of a Project Gutenberg-tm work in a format
other than "Plain Vanilla ASCII" or other format used in the official
version posted on the official Project Gutenberg-tm web site
(www.gutenberg.org), you must, at no additional cost, fee or expense
to the user, provide a copy, a means of exporting a copy, or a means
of obtaining a copy upon request, of the work in its original "Plain
Vanilla ASCII" or other form. Any alternate format must include the
full Project Gutenberg-tm License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works
provided that
* You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is owed
to the owner of the Project Gutenberg-tm trademark, but he has
agreed to donate royalties under this paragraph to the Project
Gutenberg Literary Archive Foundation. Royalty payments must be paid
within 60 days following each date on which you prepare (or are
legally required to prepare) your periodic tax returns. Royalty
payments should be clearly marked as such and sent to the Project
Gutenberg Literary Archive Foundation at the address specified in
Section 4, "Information about donations to the Project Gutenberg
Literary Archive Foundation."
* You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or destroy all
copies of the works possessed in a physical medium and discontinue
all use of and all access to other copies of Project Gutenberg-tm
works.
* You provide, in accordance with paragraph 1.F.3, a full refund of
any money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days of
receipt of the work.
* You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project
Gutenberg-tm electronic work or group of works on different terms than
are set forth in this agreement, you must obtain permission in writing
from both the Project Gutenberg Literary Archive Foundation and The
Project Gutenberg Trademark LLC, the owner of the Project Gutenberg-tm
trademark. Contact the Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
works not protected by U.S. copyright law in creating the Project
Gutenberg-tm collection. Despite these efforts, Project Gutenberg-tm
electronic works, and the medium on which they may be stored, may
contain "Defects," such as, but not limited to, incomplete, inaccurate
or corrupt data, transcription errors, a copyright or other
intellectual property infringement, a defective or damaged disk or
other medium, a computer virus, or computer codes that damage or
cannot be read by your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium
with your written explanation. The person or entity that provided you
with the defective work may elect to provide a replacement copy in
lieu of a refund. If you received the work electronically, the person
or entity providing it to you may choose to give you a second
opportunity to receive the work electronically in lieu of a refund. If
the second copy is also defective, you may demand a refund in writing
without further opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS', WITH NO
OTHER WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT
LIMITED TO WARRANTIES OF MERCHANTABILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of
damages. If any disclaimer or limitation set forth in this agreement
violates the law of the state applicable to this agreement, the
agreement shall be interpreted to make the maximum disclaimer or
limitation permitted by the applicable state law. The invalidity or
unenforceability of any provision of this agreement shall not void the
remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in
accordance with this agreement, and any volunteers associated with the
production, promotion and distribution of Project Gutenberg-tm
electronic works, harmless from all liability, costs and expenses,
including legal fees, that arise directly or indirectly from any of
the following which you do or cause to occur: (a) distribution of this
or any Project Gutenberg-tm work, (b) alteration, modification, or
additions or deletions to any Project Gutenberg-tm work, and (c) any
Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of
computers including obsolete, old, middle-aged and new computers. It
exists because of the efforts of hundreds of volunteers and donations
from people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future
generations. To learn more about the Project Gutenberg Literary
Archive Foundation and how your efforts and donations can help, see
Sections 3 and 4 and the Foundation information page at
www.gutenberg.org
Section 3. Information about the Project Gutenberg Literary Archive Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg Literary
Archive Foundation are tax deductible to the full extent permitted by
U.S. federal laws and your state's laws.
The Foundation's principal office is in Fairbanks, Alaska, with the
mailing address: PO Box 750175, Fairbanks, AK 99775, but its
volunteers and employees are scattered throughout numerous
locations. Its business office is located at 809 North 1500 West, Salt
Lake City, UT 84116, (801) 596-1887. Email contact links and up to
date contact information can be found at the Foundation's web site and
official page at www.gutenberg.org/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
[email protected]
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To SEND
DONATIONS or determine the status of compliance for any particular
state visit www.gutenberg.org/donate
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations. To
donate, please visit: www.gutenberg.org/donate
Section 5. General Information About Project Gutenberg-tm electronic works.
Professor Michael S. Hart was the originator of the Project
Gutenberg-tm concept of a library of electronic works that could be
freely shared with anyone. For forty years, he produced and
distributed Project Gutenberg-tm eBooks with only a loose network of
volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as not protected by copyright in
the U.S. unless a copyright notice is included. Thus, we do not
necessarily keep eBooks in compliance with any particular paper
edition.
Most people start at our Web site which has the main PG search
facility: www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.