The Project Gutenberg EBook of Australasian Fossils, by Frederick Chapman
This eBook is for the use of anyone anywhere in the United States and most
other parts of the world at no cost and with almost no restrictions
whatsoever. You may copy it, give it away or re-use it under the terms of
the Project Gutenberg License included with this eBook or online at
www.gutenberg.org. If you are not located in the United States, you'll have
to check the laws of the country where you are located before using this ebook.
Title: Australasian Fossils
A Students' Manual of Palaeontology
Author: Frederick Chapman
Contributor: E. W. (Ernest Willington) Skeats
Release Date: March 16, 2019 [EBook #59074]
Language: English
Character set encoding: ISO-8859-1
*** START OF THIS PROJECT GUTENBERG EBOOK AUSTRALASIAN FOSSILS ***
Produced by MFR, Tom Cosmas and the Online Distributed
Proofreading Team at http://www.pgdp.net (This file was
produced from images generously made available by The
Internet Archive)
BY THE SAME AUTHOR.
The Foraminifera
An Introduction to the Study of the Protozoa
by
FREDERICK CHAPMAN, A.L.S., F.R.M.S.
This book has been written with a view of
meeting a demand which has arisen for a concise
account of the Foraminifera, suited to the
requirements of the student of Natural History
and Palaeontology.
With 14 plates and 42 illustrations in the Text.
DEMY 8vo. CLOTH, 10s. 6d.

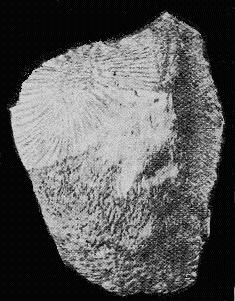
A FOSSIL CRINOID
(Helicocrinus plumosus), about 5/6 nat. size,
in Silurian Mudstone, Brunswick, Victoria.
(Spec. in Nat. Mus., Melbourne).
A Students’ Manual of Palaeontology
Palaeontologist to the National Museum, Melbourne.
Formerly Assistant in the Geological Department of the Royal College of Science, London.
Assoc. Linnean Soc. [Lond.], F.R.M.S., etc.
Author of “The Foraminifera,” “A Monograph of the Silurian Bivalved Mollusca of Victoria,” “New or Little-known Victorian Fossils in the National Museum,” etc.
With an Introduction by
PROFESSOR E. W. SKEATS, D.Sc., F.G.S.
GEORGE ROBERTSON & COMPANY PROPY. LTD.,
Melbourne, Sydney, Adelaide, Brisbane and London.
1914.
| Page | |||
| Preface | 10 | ||
| Introduction by Professor E. W. Skeats, D.Sc., F.G.S. | 13 | ||
PART I.—GENERAL PRINCIPLES. |
|||
| Chap. | I. | —Nature and uses of Fossils | 21 |
| " | II. | —Classification of Fossil Animals and Plants | 34 |
| " | III. | —The Geological Epochs and Time-range of Fossils | 41 |
| " | IV. | —How Fossils are Found, and the Rocks They Form | 51 |
PART II.—SYSTEMATIC PALAEONTOLOGY. |
|||
| Chap. | V. | —Fossil Plants | 82 |
| " | VI. | —Fossil Foraminifera and Radiolaria | 95 |
| " | VII. | —Fossil Sponges, Corals and Graptolites | 107 |
| " | VIII. | —Fossil Starfishes, Sea-lilies and Sea-urchins | 133 |
| " | IX. | —Fossil Worms, Sea-mats and Lamp-shells | 152 |
| " | X. | —Fossil Shell-fish | 174 |
| " | XI. | —Fossil Trilobites, Crustacea and Insects | 220 |
| " | XII. | —Fossil Fishes, Amphibians, Reptiles, Birds and Mammals | 257 |
| Appendix.—Notes on Collecting and Preserving Fossils | 315 | ||
| Index | 321 | ||
| Fig. | Page | |
| 1. | Fossil Shells in clay | 22 |
| 2. | Tracks, probably of Crustaceans | 22 |
| 3. | Structure of Silicified Wood in tangential section: Araucarioxylon Daintreei, Chapm. | 24 |
| 4. | Portrait of William Smith | 26 |
| 5. | Raised Beach: Brighton, England | 28 |
| 6. | Raised Beach: Torquay, Victoria | 28 |
| 7. | Marine Fossils in Volcanic Tuff: Summit of Snowdon | 29 |
| 8. | Kitchen Middens: Torquay, Victoria | 30 |
| 9. | Submerged Forest on the Cheshire Coast | 30 |
| 10. | Pecten murrayanus, Tate. A fossil shell allied to a living species | 32 |
| 11. | Cliff section: Torquay, Victoria | 42 |
| 12. | Diagram of superposition of Strata | 42 |
| 13. | Diagram of the Range-in-time of Australasian Fossils | 50 |
| 14. | Diprotodon skeletons in situ: Lake Callabonna, S. Australia | 51 |
| 15. | Bird remains on sand dunes: King Island, Bass Strait | 52 |
| 16. | Impression of Bird’s feather in Ironstone: Western Victoria | 52 |
| 17. | A Fossil Turtle: Notochelone costata, Owen sp. | 52 |
| 18. | A Ganoid Fish: Pristisomus crassus, A. S. Woodward | 54 |
| 19. | A fossil Insect in amber (Tipula sp.) | 54 |
| 20. | A fossil Crustacean: Thalassina emerii, Bell | 55 |
| 21. | An Ammonite: Desmoceras flindersi, McCoy sp. | 55 |
| 22. | Belemnites: Belemnites diptycha, McCoy | 56 |
| 23. | A Group of Lamp-shells: Magellania flavescens, Lam. sp. | 56 |
| 24. | Zoarium of a living Polyzoan: Retepora sp. | 58 |
| 25. | A fossil Polyzoan: Macropora clarkei, T. Woods sp. | 58 |
| 26. | Fossil Worm-tubes: (?) Serpula | 60 |
| 27. | A living Sea-urchin: Strongylocentrotus erythrogrammus, Val. | 60 |
| 28. | A fossil Sea-urchin: Linthia antiaustrails, Tate | 60 |
| 29. | A fossil Brittle-Star: Ophioderma egertoni, Brod. sp. | 60 |
| 30. | A fossil Crinoid: Taxocrinus simplex, Phillips sp. | 62 |
| 31. | Graptolites on Slate: Tetragraptus fruticosus, J. Hall sp. | 62 |
| 32. | A Stromatoporoid: Actinostroma « 7 » | 63 |
| 33. | Corals in Devonian Marble: Favosites | 64 |
| 34. | Siliceous Skeleton of a living Sponge: (?) Chonelasma | 64 |
| 35. | Spicules of a fossil Sponge: Ecionema newberyi, McCoy sp. | 65 |
| 36. | Nummulites: N. gizehensis, Ehr. var. champollioni, De la Harpe | 65 |
| 37. | Cainozoic Radiolaria | 66 |
| 38. | Radiolaria in Siliceous Limestone | 67 |
| 39. | Travertin Limestone, with leaves of Beech (Fagus) | 67 |
| 40. | Freshwater Limestone with shells (Bulinus) | 68 |
| 41. | Hardened mudstone with Brachiopods (Orthis, etc.) | 69 |
| 42. | Diatomaceous Earth | 72 |
| 43. | Lepidocyclina Limestone | 73 |
| 44. | Coral in Limestone: Favosites grandipora, Eth. fil. | 74 |
| 45. | Crinoidal Limestone | 74 |
| 46. | Turritella Limestone | 75 |
| 47. | Ostracodal Limestone | 75 |
| 48. | Halimeda Limestone | 77 |
| 49. | Tasmanite: a Spore Coal | 77 |
| 50. | Kerosene Shale | 77 |
| 51. | Bone Bed | 77 |
| 52. | Bone Breccia | 79 |
| 53. | Cainozoic Ironstone with Leaves (Banksia) | 80 |
| 54. | Girvanella conferta, Chapm., in Silurian Limestone | 83 |
| 55. | Palaeozoic Plants | 83 |
| 56. | Restoration of Lepidodendron | 84 |
| 57. | Stem of Lepidodendron (Lepidophloios), showing leaf-scars | 84 |
| 58. | Upper Palaeozoic Plants | 85 |
| 59. | Map of Gondwana-Land | 87 |
| 60. | Mesozoic Plants | 88 |
| 61. | Cainozoic Plants | 90 |
| 62. | Eucalyptus leaves from the Deep Leads | 92 |
| 63. | Palaeozoic and Mesozoic Foraminifera | 97 |
| 64. | Lepidocyclina marginata, Mich. sp. Sections of shell showing structure | 99 |
| 65. | Cainozoic Foraminifera | 100 |
| 66. | Fossil Radiolaria | 103 |
| 67. | Palaeozoic Sponges and Archaeocyathinae | 108 |
| 68. | Cainozoic Sponges | 111 |
| 69. | Silurian Corals | 111 |
| 70. | Upper Palaeozoic Corals | 116 |
| 71. | Cainozoic Corals | 118 |
| 72. | Stromatoporoidea and Cladophora | 121 |
| 73. | Lower Ordovician Graptolites « 8 » | 125 |
| 74. | Lower Ordovician Graptolites | 125 |
| 75. | Upper Ordovician and Silurian Graptolites | 127 |
| 76. | Fossil Crinoids | 135 |
| 77. | Fossil Starfishes | 140 |
| 78. | Protaster brisingoides, Gregory, in Silurian Sandstone | 142 |
| 79. | Gregoriura spryi, Chapm., in Silurian Mudstone | 143 |
| 80. | Cainozoic Sea-urchins | 145 |
| 81. | Cainozoic Sea-urchins | 147 |
| 82. | Fossil Worms | 153 |
| 83. | Palaeozoic Polyzoa | 156 |
| 84. | Cainozoic Polyzoa | 157 |
| 85. | Lower Palaeozoic Brachiopods | 159 |
| 86. | Silurian and Devonian Brachiopods | 161 |
| 87. | Carbopermian Brachiopods | 163 |
| 88. | Mesozoic Brachiopods | 165 |
| 89. | Cainozoic Brachiopods | 167 |
| 90. | Lower Palaeozoic Bivalves | 176 |
| 91. | Palaeozoic Bivalves | 179 |
| 92. | Carbopermian Bivalves | 180 |
| 93. | Lower Mesozoic Bivalves | 181 |
| 94. | Cretaceous Bivalves | 183 |
| 95. | Cainozoic Bivalves | 185 |
| 96. | Cainozoic Bivalves | 186 |
| 97. | Fossil Scaphopods and Chitons | 188 |
| 98. | Lower Palaeozoic Gasteropoda | 192 |
| 99. | Silurian Gasteropoda | 194 |
| 100. | Upper Palaeozoic Gasteropoda | 195 |
| 101. | Mesozoic Gasteropoda | 197 |
| 102. | Cainozoic Gasteropoda | 199 |
| 103. | Cainozoic Gasteropoda | 200 |
| 104. | Late Cainozoic and Pleistocene Gasteropoda | 201 |
| 105. | Palaeozoic Cephalopoda | 206 |
| 106. | Mesozoic and Cainozoic Cephalopoda | 208 |
| 107. | Diagram restoration of an Australian Trilobite (Dalmanites) | 224 |
| 108. | Cambrian Trilobites | 226 |
| 109. | Older Silurian Trilobites | 228 |
| 110. | Newer Silurian Trilobites | 230 |
| 111. | Carboniferous Trilobites and a Phyllopod | 232 |
| 112. | Silurian Ostracoda | 236 |
| 113. | Upper Palaeozoic and Mesozoic Ostracoda | 238 |
| 114. | Cainozoic Ostracoda | 239 |
| 115. | Fossil Cirripedes | 242 |
| 116. | Cirripedes. Lepas anatifera, Linn.: living goose barnacle, and L. pritchardi, Hall: Cainozoic | 242 |
| 117. | Ceratiocaris papilio, Salter | 244 |
| 118. | Ordovician Phyllocarids | 245 |
| 119. | Silurian Phyllocarids « 9 » | 245 |
| 120. | Fossil Crabs and Insects | 247 |
| 121. | Silurian Eurypterids | 249 |
| 122. | Thyestes magnificus, Chapm. | 259 |
| 123. | Gyracanthides murrayi, A. S. Woodw. Restoration | 260 |
| 124. | Teeth and Scales of Palaeozoic and Mesozoic Fishes | 260 |
| 125. | Cleithrolepis granulatus, Egerton | 263 |
| 126. | Tooth of Ceratodus avus, A. S. W., and phalangeal of a carnivorous Deinosaur | 264 |
| 127. | Scale of Ceratodus ? avus | 265 |
| 128. | The Queensland Lung-fish: Neoceratodus forsteri, Krefft | 266 |
| 129. | Leptolepis gregarius, A. S. W. | 266 |
| 130. | Cretaceous and Cainozoic Fish-teeth | 268 |
| 131. | Cainozoic Fish remains | 270 |
| 132. | Bothriceps major, A. S. W. | 273 |
| 133. | Ichthyosaurus australis, McCoy | 277 |
| 134. | Fossil Reptiles | 278 |
| 135. | Impression of Bird’s feather, magnified, Cainozoic: Victoria | 281 |
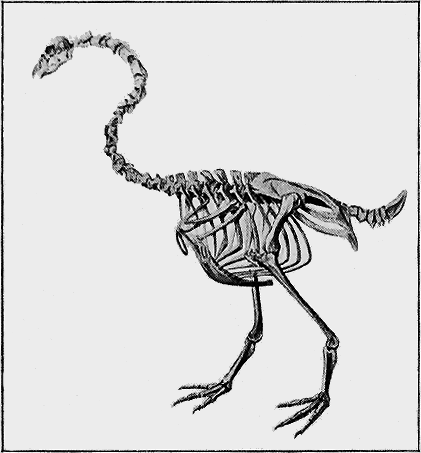
| 136. | Cnemiornis calcitrans, Owen | 284 |
| 137. | Dinornis maximus, Owen. Great Moa | 284 |
| 138. | Pachyornis elephantopus, Owen | 285 |
| 139. | Skeleton of Sarcophilus ursinus, Harris sp. | 288 |
| 140. | Skull of fossil specimen of Sarcophilus ursinus | 288 |
| 141. | Thylacinus major, Owen. Hind part of mandible | 289 |
| 142. | Phascolomys pliocenus, McCoy. Mandible | 290 |
| 143. | Cainozoic Teeth and Otolith | 291 |
| 144. | Skeleton of Diprotodon australis, Owen | 291 |
| 145. | Right hind foot of Diprotodon australis | 292 |
| 146. | Restoration of Diprotodon australis | 292 |
| 147. | Skull and mandible of Thylacoleo carnifex, Owen | 293 |
| 148. | Wynyardia bassiana, Spencer | 294 |
| 149. | Tooth of Scaldicetus macgeei, Chapm. | 297 |
| 150. | Impressions of footprints in dune sand-rock, Warrnambool | 301 |
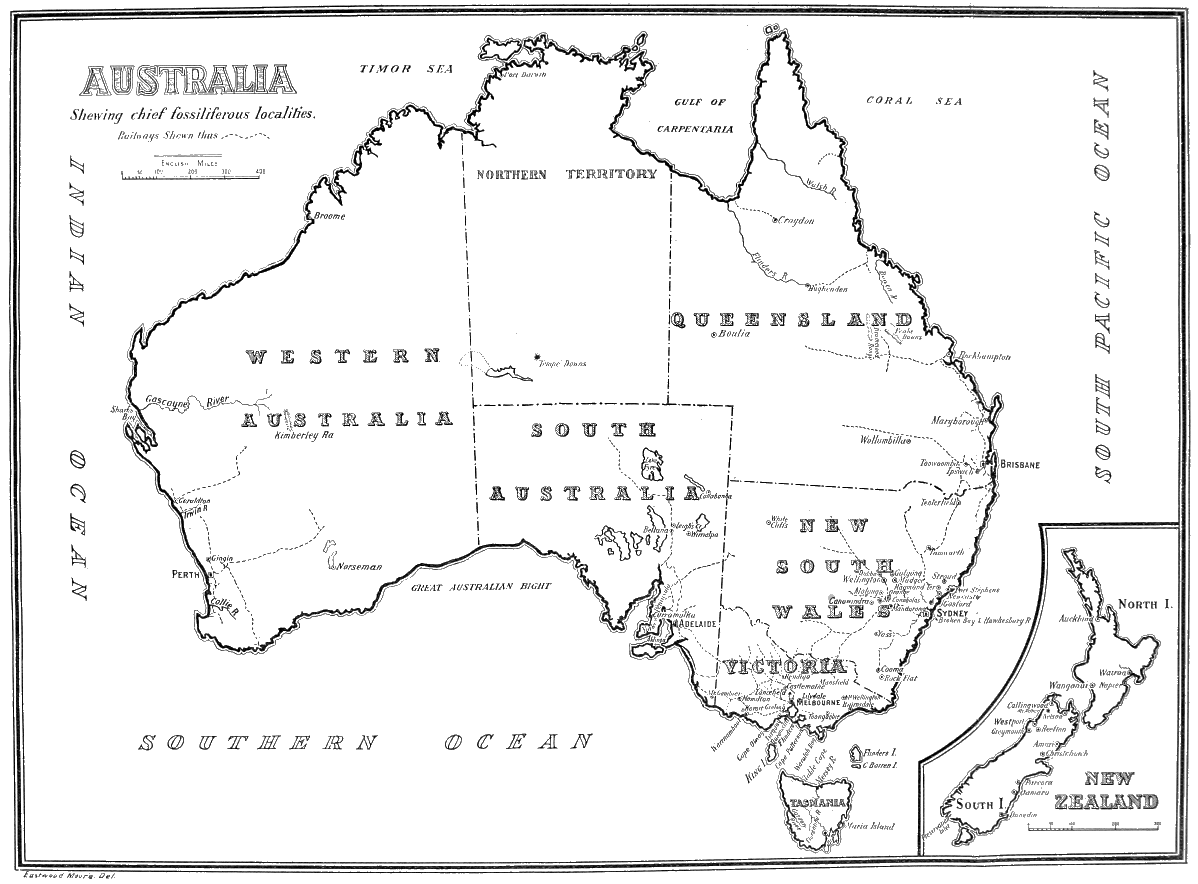
| Map of Australia, showing chief fossiliferous localities | Map |
The more important discoveries of fossils in the southern hemisphere have received, as a rule, very meagre notice in many of the text-books of Geology and Palaeontology published in England, Germany and America, and used by Australasian students. It is thought, therefore, that the time has arrived when an attempt should be made to collect the main facts bearing upon this subject, in order to present them from an Australasian standpoint. With this in view, references to fossils occurring in the northern hemisphere are subordinated, seeing that these may be easily obtained on reference to the accepted text-books in general use.
The present work does not presume to furnish a complete record of Australasian palaeontology, since that would mean the production of a much more extensive and costly volume. Sufficient information is here given, however, to form a groundwork for the student of this section of natural science, and a guide to the collector of these “medals of creation.”
The systematic portion of this book has been arranged primarily from the biological side, since Palaeontology is the “study of ancient life.” Taking each life-group, therefore, from the lowest to the highest types, all the divisions represented by fossils are dealt with in turn, beginning with their occurrence in the oldest rocks and ending with those in the newest strata.
If a commendation of the study of fossils, apart from its scientific utility, were needed, it could be « 11 » pointed out that palaeontology as a branch of geology is, par excellence, an open-air study: and since it requires as handmaids all the sister sciences, is a subject of far-reaching interest. Microscopy and photography are of immense value in certain branches of fossil research, the former in the examination of the minute forms of mollusca, foraminifera and ostracoda, the latter in the exact portraiture of specimens too intricate to copy with the brush, or too evanescent to long retain, when out of their matrix, their clean fresh surfaces. With geology or palaeontology as an objective, a country walk may be a source of much enjoyment to its students, for “in their hand is Nature like an open book”; and the specimens collected on a summer excursion may be closely and profitably studied in the spare time of the winter recess.
The author sincerely trusts that students may share the same pleasure which he has derived from the study of these relics of past life; and that the present attempt to show their relationship both in geological time and biological organisation, may be the means of inducing many to make further advances in this fascinating subject.
In the production of this work several friends and collaborators have materially assisted, their aid considerably increasing its value. It is therefore with grateful thanks that the author acknowledges the help and encouragement given by Professor E. W. Skeats, D.Sc., who has not only been good enough to write the Introductory passages, but who has carefully gone over the MS. and made many helpful « 12 » suggestions. Mr. W. S. Dun, F.G.S., Palaeontologist to the Geological Survey Branch of the Department of Mines, Sydney, has also rendered generous help in giving the benefit of his full acquaintance of the palaeontology of his own State. To the Trustees of the National Museum the author is under special obligations for permission to photograph many unique fossil specimens in the Museum collection, comprising Figs. 3, 16-18, 20-22, 28-31, 35, 39, 40, 45, 46, 51-54, 57, 62, 78, 79, 127, 133, 136, 147 and 148. The author’s thanks are also due to Dr. E. C. Stirling, M.D., M.A., F.R.S., for permission to use Figs. 143, 144 and 145, whilst similar privileges have been accorded by Prof. A. G. Seward, F.R.S., Dr. F. A. Bather, F.R.S., and Mr. C. L. Barrett. Prof. T. W. Edgeworth David, F.R.S., has kindly cleared up some doubtful points of stratigraphy and further increased the author’s indebtedness by the loan of a unique slide of Radiolaria figured on p. 69. Mr. Eastwood Moore, to whom special thanks are due, has greatly added to the pictorial side of this work by his skillful help in preparing many of the illustrations for the press, as well as in the drawing of the several maps. The grouped sets of fossils have been especially drawn for this work by the author. They are either copied from authentic specimens or from previously published drawings; references to the authorities being given in the accompanying legends. Dr. T. S. Hall has kindly read the section on Graptolites and Mammalia. For many helpful suggestions and the careful reading of proofs, thanks are especially owing to Mr. W. E. G. Simons, Mr. R. A. Keble, and to my wife.
Geological Department,
The University, Melbourne.
William Smith, the Father of English Geology, used to apologize for the study of palaeontology by claiming that “the search for a fossil is at least as rational a proceeding as the pursuit of a hare.” Those of us who are accustomed to take the field, armed with a hammer, in the search for “medals of creation” and from time to time have experienced the sporting enjoyment of bringing to light a rare or perfect specimen are quite prepared to support his claim. But the student of fossils needs the help of a text book to guide him to the literature on the subject, to help him with his identifications or to indicate that some of his finds are new and hitherto undescribed. European and American workers have long been provided with excellent books treating generally of fossils, but the illustrations have been quite naturally taken mainly from forms occurring in the Northern Hemisphere. Our own fossil forms both plants and animals are numerous, interesting and in many cases peculiar, but the literature concerning them is so widely scattered in various « 14 » scientific publications that a warm welcome should be given to this book of Mr. Chapman’s, in which the Australian evidence is brought together and summarised by one, whose training, long experience, and personal research qualify him to undertake the task. Especially will teachers and students of Geology and Palaeontology value such an undertaking. Workers in other countries who have only partial access to the Australian literature on the subject should also find this a valuable book of reference.
In the study of fossils we are concerned with the nature, evolution and distribution of the former inhabitants of the earth. The study of Palaeontology may be justified as a means of scientific discipline, for the contributions the subject makes to the increase of natural knowledge and the unfolding of panoramas of ancient life. It also provides perhaps the most positive evidence in the story of evolution. So, too, the student of the present day distribution of animals and plants finds the key to many a problem in zoo-geography in the records of past migrations yielded by the study of fossils in different lands. The stratigraphical geologist is of course principally concerned with two important aspects of the study of fossils.
The masterly generalisation of William Smith that strata can be identified by their fossil contents established by close study of the rocks and fossils of the British Oolites has been confirmed generally by subsequent work. The comparative study of the fossil contents of rocks in widely separated areas has proved to be the most valuable means by which the « 15 » correlation of the rocks can be effected and their identity of age established. In some cases the recognition of a single fossil species in two areas separated, perhaps, by thousands of miles may suffice to demonstrate that the rocks are of the same age. For example, a graptolite such as Phyllograptus typus is found in many parts of the world, but has only a very restricted range in time. It has been found only in rocks of Lower Ordovician age. Its occurrence in Wales and in the rocks of Bendigo practically suffices to establish the identity in age of the rocks in these widely separated areas.
Generally, however, much closer study and a more detailed examination of a large number of the fossils of a rock series are required before the age of the rocks can be surely established and a safe correlation made with distant localities.
The stratigraphical generalisations to be made from the study of fossils however must be qualified by certain considerations. Among these are the fact that our knowledge of the life forms of a given geological period is necessarily incomplete, that the differences in the fossil contents of rocks may depend not only on differences of age but also in the conditions under which the organisms lived and the rocks were accumulated, and that forms of life originating in one area do not spread themselves immediately over the earth but migrate at velocities depending on their mode of life and the presence or absence of barriers to their progress.
Our incomplete knowledge of the forms living in remote geological periods arises partly from the fact « 16 » that some forms had no permanent skeleton and were therefore incapable of preservation, partly to the obliteration of the skeletons of organisms through subsequent earth movements in the rocks or through the solvent action of water. Many land forms, too, probably disintegrated on the surface before deposits were formed over the area. Apart from these causes which determine that a full knowledge of the fossils from ancient rocks in particular, will never be acquired, our knowledge is incomplete by reason either of difficulty of access to certain areas or incomplete search. As a result of later discoveries earlier conclusions based on incomplete evidence as to the age of a rock series, have not infrequently been modified.
The study of the present distribution of animals and plants over the earth is a help in the attempt to decide how far the fossil differences in the sets of rocks are due to differences in the ages of the rocks or to differences in the conditions under which the organisms lived. The present, in this, as in many other geological problems, is the key to the past.
We know, for instance, that differences of climate largely control the geographical distribution of land animals and especially of land plants, and for that reason among others, fossil plants are generally less trustworthy guides to geological age than fossil animals.
In the distribution of marine animals at the present day we find that organisms of simple structure are generally more wide-spread and less susceptible to changes in their environment than are the more complex organisms with specialised structures. Hence we find, for instance, a fossil species of the « 17 » Foraminifera may persist unchanged through several geological periods, while a species of fossil fish has in general not only a short range in time but often a restricted geographical extent. If we consider the marine organisms found at the present day we find a number of free-swimming forms very widely distributed, while a large number are restricted either by reason of climate or of depth. Certain organisms are only to be found between high and low tide levels, others between low tide level and a depth of thirty fathoms, while many quite different forms live in deeper waters. If we confine our attention to shallow-water marine forms we note that certain forms are at the present day restricted to waters of a certain temperature. We find, therefore, a contrast between arctic and tropical faunas, while other types characterize temperate latitudes. Climatic and bathymetrical differences at the present day therefore lead to distinct differences in the distribution of certain organisms, while other forms, less sensitive to these factors, range widely and may be almost universally distributed. Similar conditions obtained in past geological times, and therefore in attempting to correlate the rocks of one area with those of another those fossils which are most wide-spread are often found to be the most valuable.
Attention should also be paid to the conditions under which the deposits accumulated, since it is clear that rocks may be formed at the same time in different areas and yet contain many distinct fossils by reason of climatic or bathymetrical differences. Among living marine organisms we find certain forms restricted to sandy or muddy sea-bottoms and others « 18 » to clear water, and these changes in the conditions of deposition of sediment have played their part in past geological periods in determining differences in the fossil faunas of rocks which were laid down simultaneously. We not infrequently find mudstones passing laterally into limestones, and this lithological change is always accompanied by a more or less notable change in the fossil contents of the two rock types. Such facts emphasize the close connection between stratigraphy and palaeontology, and indicate that the successful tracing out of the geological history of any area is only possible when the evidence of the stratigrapher is reinforced by that provided by the palaeontologist. The fact that species of animals and plants which have been developed in a particular area do not spread all over the world at once but migrate very slowly led Huxley many years ago to put forward his hypothesis of “homotaxis.” He agreed that when the order of succession of rocks and fossils has been made out in one area, this order and succession will be found to be generally similar in other areas. The deposits in two such contrasted areas are homotaxial, that is, show a similarity of order, but, he claimed, are not necessarily synchronous in their formation. In whatever parts of the world Carboniferous, Devonian and Silurian fossils may be found, the rocks with Carboniferous fossils will be found to overlie those with Devonian, and these in their turn rest upon those containing Silurian fossils. And yet Huxley maintained that if, say, Africa was the area in which faunas and floras originated, the migration of a Silurian fauna and flora might take place so slowly « 19 » that by the time it reached Britain the succeeding Devonian forms had developed in Africa, and when it reached North America, Devonian forms had reached Britain and Carboniferous forms had developed in Africa. If this were so a Devonian fauna and flora in Britain may have been contemporaneous with Silurian life in North America and with a Carboniferous fauna and flora in Africa.
This could only be true if the time taken for the migration of faunas and floras was so great as to transcend the boundaries between great geological periods. This does not appear to be the case, and Huxley’s idea in its extreme form has been generally abandoned. At the same time certain anomalies in the range in time of individual genera have been noted, and may possibly be explained on such lines. For instance, among the group of the graptolites, in Britain the genus Bryograptus occurs only in the Upper Cambrian and the genus Leptograptus only in the Upper Ordovician rocks. In Victoria these two genera, together with typical Lower Ordovician forms, may be found near Lancefield preserved on a single slab of shale. In the same way, in a single quarry in Triassic rocks in New South Wales, a number of fossil fish have been found and described, some of which have been compared to Jurassic, others to Permian, and others to Carboniferous forms in the Northern Hemisphere.
Another point which the palaeontologist may occasionally find evidence for is the existence of “biological asylums,” areas which by means of land or other barriers may be for a long period separated from the main stream of evolution. We know that « 20 » the present fauna and flora of Australia is largely of archaic aspect, as it includes a number of types which elsewhere have long ago become extinct or were never developed. This appears to be due to the long isolation of Australia and, as Professor Gregory happily puts it—its “development in a biological backwater.” We have some evidence that similar asylums have existed in past geological periods, with the result that in certain areas where uniform conditions prevailed for a long time or where isolation from competition prevented rapid evolution, some organisms which became extinct in other areas, persisted unchanged in the “asylum” into a younger geological period.
The broad generalizations that rocks may be identified by their fossil contents and that the testimony of the rocks demonstrates the general order of evolution from simple to complex forms, have only been placed on a surer footing by long continued investigations. The modifications produced by conditions of deposit, of climate and of natural barriers to migration, while introducing complexities into the problems of Palaeontology, are every year becoming better known; and when considered in connection with the variations in the characters of the rocks, provide valuable and interesting evidence towards the solution of the ultimate problems of geology and palaeontology, which include the tracing out of the evolution of the history of the earth from the most remote geological period to that point at which the geologist hands over his story to the archaeologist, the historian, and the geographer.
ERNEST W. SKEATS.
GENERAL PRINCIPLES.
NATURE AND USES OF FOSSILS.
Scope of Geology.—
The science of GEOLOGY, of which PALAEONTOLOGY or the study of fossils, forms a part, is concerned with the nature and structure of the earth, the physical forces that have shaped it, and the organic agencies that have helped to build it.
Nature of Fossils.—
The remains of animals and plants that formerly existed in the different periods of the history of the earth are spoken of as fossils. They are found, more or less plentifully, in such common rocks as clays, shales, sandstones, and limestones, all of which are comprised in the great series of Sedimentary Rocks (Fig. 1).
According to the surroundings of the organisms, whether they existed on land, in rivers, lakes, estuaries, or the sea, they are spoken of as belonging to terrestrial, fluviatile, lacustrine, estuarine, or marine deposits.


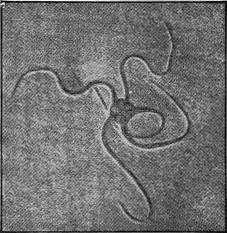
The name fossil, from the Latin ‘fodere’ to dig,—‘fossilis,’ dug out,—is applied to the remains of any animals or plants which have been buried either in sediments laid down in water, in materials gathered together by the wind on land as sand-dunes, in beds of volcanic ash, or in cave earths. But not only remains of organisms are thus called fossils, for the name is also applied to structures only indirectly connected with once living objects, such as rain-prints, ripple-marks, sun-cracks, and tracks or impressions of worms and insects (Fig. 2).
Preservation of Fossils.—
In ordinary terms, fossils are the durable parts of animals and plants which have resisted complete decay by being covered over with the deposits above-named. It is due, then, to the fact that they have been kept from the action of the air, with its destructive bacteria, that we are able to still find these relics of life in the past.
Petrifaction of Fossils.—
When organisms are covered by a tenacious mud, they sometimes undergo no further change. Very often, however, moisture containing mineral matter such as carbonate of lime or silica, percolates through the stratum which contains the fossils, and then they not only have their pores filled with the mineral, but their actual substance may also undergo a molecular change, whereby the original composition of the shell or the hard part is entirely altered. This tends almost invariably to harden the fossils still further, which change of condition is called petrifaction, or the making into stone.

Structure Preserved.—
Petrifaction does not necessarily destroy the structure of a fossil. For example, a piece of wood, which originally consisted of carbon, hydrogen, and nitrogen, may be entirely replaced by flint or silica: and yet the original structure of the wood may be so perfectly preserved that when a thin slice of the petrifaction is examined under a high power of the microscope, the tissues with their component cells are seen and easily recognised (Fig. 3).
Early Observers.—
Remains of animals buried in the rocks were known from the earliest times, and frequent references to these were made by the ancient Greek and Roman philosophers.
Xenophanes.—
Xenophanes, who lived B.C. 535, wrote of shells, « 25 » fishes and seals which had become dried in mud, and were found inland and on the tops of the highest mountains. The presence of these buried shells and bones was ascribed by the ancients to a plastic force latent in the earth itself, while in some cases they were regarded as freaks of nature.
Leonardo da Vinci.—
In the sixteenth and seventeenth centuries Italian observers came to the fore in clearly demonstrating the true nature of fossils. This was no doubt due in part to the fact that the Italian coast affords a rich field of observation in this particular branch of science. The celebrated painter Leonardo da Vinci (early part of the sixteenth century), who carried out some engineering works in connection with canals in the north of Italy, showed that the mud brought down by rivers had penetrated into the interior of shells at a time when they were still at the bottom of the sea near the coast.
Steno.—
In 1669, Steno, a Danish physician residing in Italy, wrote a work on organic petrifactions which are found enclosed in solid rocks, and showed by his dissection of a shark which had been recently captured and by a comparison of its teeth with those found fossil in the cliffs, that they were identical. The same author also pointed out the resemblance between the shells discovered in the Italian strata and those living on the adjacent shores. It was not until the close of the eighteenth century, however, that the study of fossil remains received a decided impetus. It is curious to note that many of these later « 26 » authors maintained the occurrence of a universal flood to account for the presence of fossil shells and bones on the dry land.

Fossils an Index to Age.—
A large part of the credit of showing how fossils are restricted to certain strata, and help to fix the succession and age of the beds, is due to the English geologist and surveyor, William Smith (Fig. 4). “The Father of English Geology,” as he has been called, published two works[1] in the early part of last century, in which he expressed his view of the value of fossils to the geologist and surveyor, and showed that there was a regular law of superposition of one bed upon another, and that strata could be identified at distant localities by their included fossils. Upon « 27 » this foundation the work of later geologists has been firmly established; and students of strata and of fossils work hand in hand.
[1] “Strata identified by Organised Fossils,” 1816-1819; and “Stratigraphical System of Organised Fossils,” 1817.
Stratigraphy.—
That branch of geology which discusses the nature and relations of the various sediments of the earth’s crust, and the form in which they were laid down, is called Stratigraphy. From it we learn that in bygone times many of those places that are now occupied by dry land have been, often more than once, covered by the sea; and thus Tennyson’s lines are forcibly brought to mind—
“There where the long street roars hath been
The stillness of the central sea.”
Elevated Sea-beds.—
A striking illustration in proof of this emergence of the land from the sea is the occurrence of marine shells similar to those now found living in the sea, in sea-cliffs sometimes many hundreds of feet above sea-level. When these upraised beds consist of shingle or sand with shore-loving shells, as limpets and mussels, they are spoken of as Raised Beaches. Elevated beaches are often found maintaining the same level along coast-lines for many miles, like those recorded by Darwin at Chili and Peru, or in the south of England (Fig. 5). They also occur intermittently along the Victorian coast, especially around the indents, where they have survived the wear and tear of tides along the coast line (Fig. 6). They are also a common feature, as a capping, on many coral islands which have undergone elevation.

Sea-beds far from the Present Coast.—
Marine beds of deeper water origin may be found not only close to the coast-line, but frequently on the tops of inland hills some miles from the sea-coast. Their included sea-shells and other organic remains are often found covered by fine sediment forming extensive beds; and they may frequently occur in the position in which they lived and died (Fig. 7). Although it is well known that sea-birds carry shell-fish for some distance inland, yet this would not account for more than a few isolated examples.
Raised Beaches as Distinct from Middens.—
Again, it may be argued that the primitive inhabitants of countries bordering the coast were in the habit of piling up the empty shells of the edible molluscs used by them for food: but these “kitchen middens” are easily distinguished from fossil deposits like shelly beaches, by the absence of stratified layers; and, further, by the shells being confined to edible species, as the Cockle (Cardium), the Blood-cockle (Arca), the Mussel (Mytilus), and the Oyster (Ostrea) (Fig. 8).


Submerged Forests.—
Evidence of change in the coast-line is shown by the occurrence of submerged forest-land, known as “fossil forests,” which consist of the stumps of trees still embedded in the black, loamy soil. Such forests, « 31 » when of comparatively recent age, are found near the existing coast-line, and may sometimes extend for a considerable distance out to sea (Fig. 9).
From the foregoing we learn that:—
1.—Fossils afford data of the various Changes that have taken place in past times in the Relative Positions of Land and Water.
Changes of Climate in the Past.—
At the present day we find special groups of animals (fauna), and plants (flora), restricted to tropical climates; and others, conversely, to the arctic regions. Cycads and tree-ferns, for example, seem to flourish best in warm or sub-tropical countries: yet in past times they were abundant in northern Europe in what are now temperate and arctic regions, as in Yorkshire, Spitzbergen, and Northern Siberia, where indeed at one time they formed the principal flora.
The rein-deer and musk-sheep, now to be found only in the arctic regions, once lived in the South of England, France and Germany. The dwarf willow (Salix polaris) and an arctic moss (Hypnum turgescens), now restricted to the same cold region, occur fossil in the South of England.
In Southern Australia and in New Zealand, the marine shells which lived during the earlier and middle Tertiary times belong to genera and species which are indicative of a warmer climate than that now prevailing; this ancient fauna being like that met with in dredging around the northern coasts of Australia (Fig. 10).

From the above evidence we may say that:—
2.—Fossils teach us that in Former Times the Climate of certain parts of the earth’s surface was Different from that now existing.
Fossils as Guides to Age of Strata.—
In passing from fossil deposits of fairly recent origin to those of older date, we find the proportion of living species gradually diminish, being replaced by forms now extinct. After this the genera themselves are replaced by more ancient types, and if we penetrate still deeper into the series of geological strata, even families and orders of animals and plants give place to others entirely unknown at the present day.
From this we conclude that:—
3.—Fossil Types, or Guide Fossils, are of great value in indicating the Relative Age of Geological Formations.
Gradual Evolution of Life-forms from Lower to Higher Types.—
As a general rule the various types of animals and plants become simpler in organisation as we descend the geological scale. For example, in the oldest rocks the animals are confined to the groups of Foraminifera, Sponges, Corals, Graptolites, Shell-fish and Trilobites, all back-boneless animals: whilst it was not until the Devonian period that the primitive fishes appeared as a well-defined group; and in the next formation, the Carboniferous Series, the first traces of the Batrachians (Frog-like animals) and Reptiles are found. Birds do not appear, so far as their remains are known, until near the close of the Jurassic; whilst Mammals are sparsely represented by Monotremes and Marsupials in the Triassic and Jurassic, becoming more abundant in Cainozoic times, and by the Eutheria (Higher Mammals) from the commencement of the Eocene period.
It is clear from the above and other facts in the geological distribution of animal types that:—
4.—The Geological Record supports in the main the Doctrine of Evolution from Simpler to more Complex types; and fossils throw much light upon the Ancestry of Animals and Plants now found Living.
THE CLASSIFICATION OF FOSSIL ANIMALS AND PLANTS.
An elementary knowledge of the principles underlying the classification of animals and plants is essential to the beginner in the study of fossils.
The Naming of Animals.—
In order to make a clearly understood reference to an animal, or the remains of one, it is as necessary to give it a name as it is in the case of a person or a place. Before the time of Linnaeus (1707-1778), it was the custom to refer, for example, to a shell, in Latin[2] as “the little spiral shell, with cross markings and tubercles, like a ram’s horn;” or to a worm as “the rounded worm with an elevated back.” Improvements in this cumbersome method of naming were made by several of the earlier authors by shortening the description; but no strict rule was established until the tenth edition of Linnaeus’ “Systema Naturae” (1758), when that author instituted his binomial nomenclature by giving each « 35 » form enumerated both a generic and specific name. In plain words, this method takes certain life-forms closely related, but differing in minute particulars, and places them together in a genus or kindred group. Thus the true dogs belong to the genus Canis, but since this group also includes wolves, jackals, and foxes, the various canine animals are respectively designated by a specific name; thus the dog (Canis familiaris), the dingo (C. dingo), the wolf (C. lupus), the jackal (C. aureus), and the fox (C. vulpes). The generic name is placed first. Allied genera are grouped in families, (for example, Canidae), these into orders (ex. Carnivora), the orders into classes (ex. Mammalia), and the classes into phyla or subkingdoms (ex. Vertebrata).
[2] The Latin description was used more commonly than it is at present, as a universal scientific language.
Plants are classified in much the same way, with the exception that families and orders are, by some authors, regarded as of equal value, or even reversed in value; and instead of the term phylum the name series is used.
Classification of the Animal Kingdom.
| NAME OF PHYLUM. | FORMS FOUND FOSSIL | |
| I.— | PROTOZOA | Foraminifera, Radiolaria. |
| II.— | COELENTERATA | Sponges, Corals, Stromatoporoids, Graptolites. |
| III.— | ECHINODERMATA | Crinoids, Starfishes, Brittle-stars, Sea-urchins. |
| IV.— | VERMES | Worms (tube-making and burrowing kinds). |
| V.— | MOLLUSCOIDEA | Polyzoa or Sea-mats, Brachiopods or Lamp-shells. |
| VI.— | MOLLUSCA | Shell-fish: as Bivalves, Tusk-shells, Chitons or Mail-shells, Gasteropods or Snails, Pteropods or Sea-butterflies; Cuttle-fishes. |
| VII.— | ARTHROPODA | Joint-footed animals: as Trilobites, Cyprids, Crabs and Lobsters, Centipedes, Spiders and Insects. « 36 » |
| VIII.— | VERTEBRATA | Fishes, Amphibians, Reptiles, Birds and Mammals. |
Classification of Animal Kingdom.
The first seven groups of the above classification are back-boneless animals or Invertebrata; the eighth division alone comprising the animals with a vertebra or backbone.
Characters of the Several Phyla.—
In the first group are placed those animals which, when living, consist of only one cell, or a series of similar cells, but where the cells were never combined to form tissues having special functions, as in the higher groups.
PROTOZOA.—
The Amoeba of freshwater ponds is an example of such, but owing to its skin or cortex being soft, and its consequent inability to be preserved, it does not concern us here. There are, however, certain marine animals of this simple type of the Protozoa which secrete carbonate of lime to form a chambered shell (Foraminifera); or silica to form a netted and concentrically coated shell held together with radial rods (Radiolaria); and both of these types are found abundantly as fossils. They are mainly microscopic, except in the case of the nummulites and a few other kinds of foraminifera, which are occasionally as large as a crown piece.
COELENTERATA.—
The second group, the Coelenterata, shows a decided advance in organisation, for the body is multicellular, and provided with a body-cavity which serves for circulation and digestion. The important divisions of this group, in which the organisms have hard parts capable of being fossilised, are the limy and flinty Sponges, the Corals, and allied groups, as well as the delicate Graptolites which often cover the surface of the older slates with their serrated, linear forms, resembling pieces of fret-saws.
ECHINODERMATA.—
The third group, Echinodermata, comprises the Sea-lilies (Crinoids), Starfishes and Sea-urchins, besides a few other less important types; and all these mentioned are found living at the present day. Their bodies are arranged in a radial manner, the skin being strengthened by spicules and hardened by limy deposits ultimately forming plates. They have a digestive canal and a circulatory system, and are thus one remove higher than the preceding group.
VERMES.—
The fourth group, Vermes (Worms), are animals with a bilateral or two-sided body, which is sometimes divided into segments, but without jointed appendages. Those which concern the student of fossils are the tube-making worms, the errant or wandering worms which form casts like the lob-worm, and the burrowing kinds whose crypts or dwellings become filled with solid material derived from the surrounding mud.
MOLLUSCOIDEA.—
Group five, the Molluscoidea, contains two types; the Flustras or Sea-mats (Polyzoa) and the Lamp-shells (Brachiopoda). They are at first sight totally unlike; for the first-named are colonies of compound animals, and the second are simple, and enclosed between two valves. They show in common, however, a bilateral symmetry. The mouth is furnished with fine tentacles, or with spirally rolled hair-like or ciliated processes.
MOLLUSCA.—
The sixth group, the Mollusca, includes all shell-fish. They are soft-bodied, bilaterally symmetrical animals, without definite segments. The shells, on account of being formed of carbonate of lime on an organic basis, are often found preserved in fossiliferous strata.
ARTHROPODA.—
The seventh group, the Arthropoda, or joint-footed animals, are distinguished by their segmented, lateral limbs, and by having a body composed of a series of segments or somites. The body and appendages are usually protected by a horny covering, the ‘exoskeleton.’ The group of the Trilobites played an important part in the first era of the formation of the earth’s crust; whilst the other groups were more sparsely represented in earlier geological times, but became more and more predominant until the present day.
VERTEBRATA.—
The great group of the Vertebrata comes last, with its chief characteristic of the backbone structure, which advances in complexity from the Fishes to the Higher Mammals.
A Simplified Classification of the Vegetable Kingdom.
| SERIES. | FORMS FOUND FOSSIL. | |
| I.— | THALLOPHYTA | Sea-weeds: as Corallines and Calcareous Algae. |
| II.— | BRYOPHYTA | Mosses, Liverworts. |
| III.— | PTERIDOPHYTA | Fern-like plants, as Horse-tails, Club-mosses and true Ferns. |
| IV.— | PTERIDOSPERMEAE | Oldest Seed-bearing plants, with fern-like foliage. |
| V.— | GYMNOSPERMEAE | Plants with naked seeds, as Cycads (Fern-palms), Ginkgo (Maiden-hair Tree), and Conifers (Pine trees). |
| VI.— | ANGIOSPERMEAE | Flowering plants, as Grasses, Lilies and all ordinary trees and plants. |
Characters of the Plant Series.
THALLOPHYTA.—
The first series, the Thallophytes, are simple unicellular plants, and occupy the same position in the vegetable kingdom as the Protozoa do in the animal kingdom. Fossil remains of these organisms seem to be fairly well distributed throughout the entire geological series, but, owing to the soft structure of the fronds in most of the types, it is often a matter of doubt whether we are dealing with a true thallophyte or not. Many of the so-called sea-weeds (fucoids) may be only trails or markings left by other organisms, as shell-fish and crustaceans.
BRYOPHYTA.—
The second series, the Bryophytes or moss plants, are represented in the fossil state by a few unimportant examples.
PTERIDOPHYTA.—
The third series, the Pteridophytes, includes the Ferns found from the Devonian up to the present day, Horse-tails and allied forms, like Equisetites, and the Club-mosses and Lepidodendron of the Carboniferous period in various parts of the world.
PTERIDOSPERMEAE.—
The fourth series, the Pteridospermeae, comprises some of the earliest seed-bearing plants, as Alethopteris and Neuropteris. They occur in rocks of Upper Palaeozoic age as far as known.
GYMNOSPERMEAE.
The fifth series, the Gymnospermeae, contains the most important types of plants found fossil, especially those of the primary and secondary rocks: they were more abundant, with the exception of the Coniferae, in the earlier than in the more recent geological periods.
ANGIOSPERMEAE.—
The sixth series, the Angiospermeae, comprises all the Flowering Trees and Plants forming the bulk of the flora now living, and is divided into the kinds having single or double seed-leaves (Monocotyledones the Dicotyledones respectively). This important group came into existence towards the close of the Cretaceous period simultaneously with the higher mammals, and increased in abundance until modern times.
THE GEOLOGICAL EPOCHS: AND THE TIME RANGE OF FOSSILS.
Superposition of Strata.—
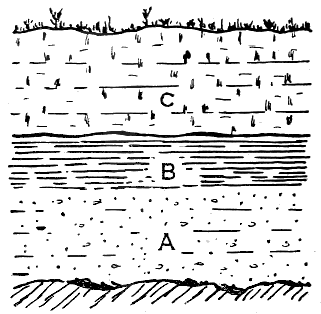
Fossils are chiefly found in rocks which have been formed of sediments laid down in water, such as sandstone, shale and most limestones. These rocks, broadly speaking, have been deposited in a horizontal position, though really slightly inclined from shore to deep-water. One layer has been formed above another, so that the oldest layer is at the bottom, and the newest at the top, of the series (Fig. 11). Let us, for instance, examine a cliff showing three layers: the lower, a sandstone, we will Call A; the intermediate, a shale or clay bed, B; and the uppermost, a limestone or marl, C (Fig. 12). In forming a conclusion about the relative ages of the beds, we shall find that A is always older than B, and B than C, provided no disturbance of the strata has taken place. For instance, the beds once horizontally deposited may have been curved and folded over, or even broken and thrust out of place, within limited areas; but occurrences like these are extremely rare. Moreover, an examination of the surrounding country, or of deep cuttings in the neighbourhood, will tell us if there is any probability of this inversion of strata having taken place.

This law of superposition holds good throughout the mass of sedimentary rocks forming the crust of the earth.
(1). Thus, the position of the strata shows the relative ages of the beds.
Differences in Fossil Faunas.—
Turning once again to our ideal cliff section, if we examine the fossils obtained from bed A, we shall find them differing in the number of kinds or species common to the other beds above and below. Thus, there will be more species alike in beds A and B or in B and C. In other words the faunas of A and B are more nearly related than those of A and C. This is explained by the fact that there is a gradual change in specific forms as we pass through the time series of strata from below upwards; so that the nearer one collecting platform is to another, as a rule, the stronger is the community of species.
Guide Fossils.—
Certain kinds of fossils are typical of particular formations. They are known as guide fossils, and by their occurrence help us to gain some idea of the approximate age of rocks widely separated by ocean and continent. Thus we find fossils typical of the Middle Devonian rocks in Europe, which also occur in parts of Australia, and we therefore conclude that the Australian rocks containing those particular fossils belong to the same formation, and are nearly of the same age.
(2). The included fossils, therefore, give evidence of the age of the beds.
Value of Lithological Evidence.—
The test of age by rock-structure has a more restricted use, but is of value when taken in conjunction with the sequence of the strata and the character of their included fossils.
To explain both the valuable and the uncertain elements of this last method as a determinant of age, we may cite, for instance, the Upper Ordovician slates of Victoria and New South Wales as an example of uniform rock formation; whilst the yellow mudstones and the grey limestones of the Upper Silurian (Yeringian series) of the same states, are instances of diverse lithological structures in strata of similar age. A reference in the latter case to the assemblages of fossils found therein, speedily settles the question.
(3). Hence, the structure and composition of the rocks (lithology), gives only partial evidence in regard to age.
Strata Vertically Arranged.—
The Stratigraphical Series of fossiliferous sediments comprises bedded rocks from all parts of the world, which geologists arrange in a vertical column according to age.
A general computation of such a column for the fossiliferous rocks of Europe gives a thickness of about 14 miles. This is equivalent to a mass of strata lying edgewise from Melbourne to Ringwood. The Australian sediments form a much thicker pile of rocks, for they can hardly fall short of 37 miles, or nearly the distance from Melbourne to Healesville.
This vertical column of strata was formed during three great eras of time. The oldest is called the Primary or Palaeozoic (“ancient life”), in which the animals and plants are of primitive types. This is followed by the Secondary or Mesozoic (“middle life”), in which the animals and plants are intermediate in character between the Palaeozoic and the later, Cainozoic. The third era is the Tertiary or Cainozoic (“recent life”), in which the animals and plants are most nearly allied to living forms. These great periods are further subdivided into epochs, as the Silurian epoch; and these again into stages, as the Yeringian stage.
Vertical Column of Fossiliferous Strata, Australia.
| ERA. | EPOCHS IN EUROPE. | EQUIVALENT STRATA IN AUSTRALIA. |
| CAINOZOIC or TERTIARY (Note 1). | HOLOCENE | Dunes, Beaches, and Shell-beds now forming. |
| PLEISTOCENE | Raised Beaches, River Terraces, Swamp Deposits with Diprotodon, Cave Breccias, Helix Sandstone. | |
| PLIOCENE | Upper.—Estuarine beds of bores in the Murray basin, Marine beds of Limestone Creek, Glenelg River, Vic. (Werrikooian). | |
| Lower.—Kalimnan red sands (terrestrial) and shell marls (marine) of Victoria, Deep Leads (fluviatile) in part, Upper Aldingan of South Australia. | ||
| MIOCENE « 46 » | Deep Leads in part: Leaf-beds of Bacchus Marsh, Dalton and Gunning. Janjukian Series of C. Otway, Spring Creek, and Table Cape. Batesford Limestone. Polyzoal Rock of Mt. Gambier and the Nullarbor Plains. Older Cainozoic of Murray basin, Lower Aldingan Series of S. Australia, Corio Bay and Bairnsdale Series. | |
| OLIGOCENE | Shelly clays and leaf-beds of the Balcombian Series at Mornington; also Shell-marls and clays with Brown Coal, Altona Bay, and lower beds at Muddy Creek, W. Vict. | |
| EOCENE | Probably no representatives. | |
| MESOZOIC or SECONDARY | CRETACEOUS | Upper.—Leaf-beds of Croydon, Q. Desert Sandstone,Q. Radiolarian Rock, N. Territory. Gin-gin Chalk, W.A. |
| Lower.—Rolling Downs Formn., Q. Lake Eyre beds, S.A. | ||
| JURASSIC | Marine.—Geraldton, W.A. | |
| Freshwater.—Carbonaceous sandstone of S. Gippsland, the Wannon, C. Otway and Barrabool Hills. Ipswich Series, Q. Mesozoic of Tasmania, Talbragar beds, N.S.W. | ||
| TRIASSIC | Upper leaf-beds at Bald Hill, Bacchus Marsh, Vict. Hawkesbury Series (Parramatta Shales, Hawkesbury Sandstone, Narrabeen beds), N.S.W. Burrum Beds, Q. | |
| PALAEOZOIC or PRIMARY « 47 » | PERMIAN and CARBONIFEROUS, UPPER | Carbopermian (Note 2), Coal Measures of New South Wales, W. Australia, Queensland (Gympie Series) and Tasmania. Gangamopteris beds of Bacchus Marsh, Vict. Upper Carboniferous of Clarence Town, N.S.W. |
| CARBONIFEROUS, LOWER | Fish and Plant beds, Mansfield, Vict. Grampian sandstone; Avon River sandstone, Vict. (?) Star beds, Queensland. Lepidodendron beds of Kimberley, W.A. (Note 3). | |
| DEVONIAN | Upper.—Sandstones of Iguana Creek, with plant remains. Lepidodendron beds with Lingula, Nyrang Creek, N.S. Wales. | |
| Middle.—Fossiliferous marbles and mudstones of Buchan, Bindi and Tabberabbera, Vict. Rocks of the Murrumbidgee, N.S. Wales, and of Burdekin, Queensland. | ||
| SILURIAN | Upper.—(Yeringian stage).—Lilydale, Loyola, Thomson River, and Waratah Bay, Vict.; Bowning and Yass (in part), N.S. Wales; Queensland. | |
| Lower (Melbournian stage).—Melbourne, Heathcote, Vict.; Bowning and Yass (in part), N.S. Wales. Gordon R. Limestone. | ||
| ORDOVICIAN, UPPER and LOWER | Slates (graptolitic).—Victoria and New South Wales. (?) Gordon River Limestone, Tas., in part (Note 4). Larapintine series of Central Australia. | |
| CAMBRIAN | Mudstones and limestones of Tasmania, South Australia, Victoria and W. Australia. | |
| PRE-CAMBRIAN « 48 » | Fossiliferous rocks doubtful chiefly represented by schistose and other metamorphic rocks. |
1.—The classification of the Cainozoics as employed here is virtually the same as given by McCoy in connection with his work for the Victorian Geological Survey. The writer has obtained further evidence to support these conclusions from special studies in the groups of the cetacea, mollusca and the protozoa. The alternative classification of the cainozoics as given by one or two later authors, introducing the useful local terminology of Hall and Pritchard for the various stages or assises is as follows:—
| TATE AND DENNANT. Stages. |
HALL AND PRITCHARD. Stages. |
||
| Werrikooian | Pleistocene Pliocene |
Werrikooian | Pliocene. |
| Kalimnan | Miocene | Kalimnan | Miocene. |
| Janjukian | (?) Oligocene | Balcombian | Eocene. |
| Balcombian | Eocene | Janjukian and Aldingan in part |
Eocene. |
| Aldingan (lower beds at that loc.) |
Eocene | ||
2.—Or Permo-carboniferous. As the series is held by some authorities to partake of the faunas of both epochs, it is preferable to use the shorter word, which moreover gives the natural sequence. There is, however, strong evidence in favour of using the term Permian for this important series.
3.—Mr. W. S. Dun regards the Lepidodendron beds of W. Australia, New South Wales and Queensland as of Upper Devonian age. There is no doubt, from a broad view of the whole question as to the respective age of these beds in Australia, that the one series is continuous, and probably represents the Upper Devonian and the Lower Carboniferous of the northern hemisphere.
4.—These limestones contain a fauna of brachiopods and corals which, at present, seems to point to the series as intermediate between the older Silurian and the Upper Ordovician.
Vertical Column of Fossiliferous Strata, New Zealand.
| ERA. | EPOCHS IN EUROPE. | EQUIVALENT STRATA IN NEW ZEALAND. | |||
| CAINOZOIC or TERTIARY |
HOLOCENE | River Alluvium. Beach Sands and Gravel. | |||
| PLEISTOCENE | Raised Beaches. Older Gravel Drifts. Moraines. Boulder Clays. |
||||
| PLIOCENE |
|
||||
| MIOCENE | Oamaru series. | ||||
| OLIGOCENE | Waimangaroa series. | ||||
| MESOZOIC or SECONDARY |
CRETACEOUS | Waipara series (of Hutton). | |||
| JURASSIC | Mataura and Putataka series. | ||||
| TRIASSIC | Wairoa, Otapiri and Kaihiku series. | ||||
| PALAEOZOIC or PRIMARY |
PERMIAN | Aorangi (unfossiliferous) series. | |||
| (?)CARBONIFEROUS | Maitai series (with Spirifer and Productus.) (?)Te Anau series (unfossiliferous). |
||||
| SILURIAN | Wangapeka series. | ||||
| ORDOVICIAN | Kakanui series (with Lower Ordovician graptolite facies). | ||||
| CAMBRIAN | Unfossiliferous. Metamorphic schists of the Maniototo series. |
Note 1.—Based for the most part, but with some slight modifications,
on Prof. J. Park’s classification in “Geology of New Zealand,” 1910.

HOW FOSSILS ARE FOUND: AND THE ROCKS THEY FORM.
As already noticed, it is the hard parts of buried animals and plants that are generally preserved. We will now consider the groups of organisms, one by one, and note the particular parts of each which we may reasonably expect to find in the fossil state.
MAMMALS.—The bones and teeth: as the Diprotodon remains of Lake Callabonna in South Australia (Fig. 14), of West Melbourne Swamp, Victoria, « 52 » and the Darling Downs, Queensland. Rarely the skin, as in the carcases of the frozen Mammoth of the tundras of Northern Siberia; or the dried remains of the Grypotherium of South American caves.
BIRDS:—Bones: as the Moa bones of New Zealand and the Emu bones of the King Island sand-dunes (Fig. 15). Very rarely the impressions of the feathers of birds are found, as in the ironstone occurring in the Wannon district of Victoria (Fig. 16), and others in fine clays and marls on the continent of Europe and in England. Fossil eggs of sea-birds are occasionally found in coastal sand-dunes of Holocene age.
REPTILES.—Skeletons of fossil turtles (Notochelone) are found in Queensland (Fig. 17). Whole skeletons and the dermal armour (spines and bony plates) of the gigantic, specialised reptiles are found in Europe, North America, and in other parts of the world.
FISHES.—Whole skeletons are sometimes found in sand and clay rocks, as in the Trias of Gosford, New South Wales (Fig. 18), and in the Jurassic of South Gippsland. The ganoid or enamel-scaled fishes are common fossils in the Devonian and Jurassic, notably in Germany, Scotland and Canada: and they also occur in the sandy mudstone of the Lower Carboniferous of Mansfield, Victoria.
INSECTS.—Notwithstanding their fragility, insects are often well preserved as fossils, for the reason that their skin and wings consist of the horny substance called chitin. The Tertiary marls of Europe are very prolific in insect remains (Fig. 19). From « 54 » the Miocene beds of Florissant, Colorado, U.S.A., several hundred species of insects have been described.

 |
Fig 19—A Fossil Insect
(Tipula sp.) in Amber.
Nat. size. Oligocene beds; Baltic Prussia. (F.C. Coll.)
|


CRUSTACEA.—The outer crust, or exoskeleton, of these animals is often hard, being formed of a compound of carbonate and phosphate of lime on an organic, chitinous base. The earliest forms of this group were the trilobites, commencing in Cambrian times, and of which there is a good representative series in Australian rocks. Remains of crabs and lobsters are found in the various Cainozoic deposits in Australia (Fig. 20), and also in the Jurassic in other parts of the world.
MOLLUSCA.—The Cuttle-fish group (Cephalopoda, “head-footed”), is well represented by the Nautilus-like, but straight Orthoceras shells commencing in Ordovician times, and, in later periods, by the beautiful, coiled Ammonites (Fig. 21). The true cuttle-fishes possess an internal bone, the sepiostaire, which one may see at the present day drifted on to the sand at high-water mark on the sea-shore. The rod-like Belemnites are of this nature, and occur abundantly in the Australian Cretaceous rocks of South Australia and Queensland (Fig. 22).
Elephant-tusk shells (Scaphopoda) are frequent in our Tertiary beds: they are also sparingly found in the Cretaceous, and some doubtful remains occur in the Palaeozoic strata of Australia.
The shells of the ordinary mollusca, such as the snails, whelks, mussels, and scallops, are abundant in almost all geological strata from the earliest periods. Their calcareous shells form a covering which, after the decay of the animal within, are from their nature among the most easily preserved of fossil remains. There is hardly an estuary bed, lake-deposit, or sea-bottom, but contains a more or less abundant assemblage of these shell-fish remains, or testacea as they were formerly called (“testa,” a shell or potsherd). We see, therefore, the importance of this group of fossils for purposes of comparison of one fauna with another (antea, Fig. 1).
The chitons or mail-shells, by their jointed nature, consisting of a series of pent-roof-shaped valves united by ligamental tissue, are nearly always represented in the fossil state by separate valves. Fossil examples of this group occur in Australia both in Palaeozoic rocks and, more numerously, in the Cainozoic series.
MOLLUSCOIDEA.—The Brachiopods or Lamp-shells consist generally of two calcareous valves as in the true mollusca (Fig. 23), but are sometimes of horny texture. Like the previous class, they are also easily preserved as fossils. They possess bent, loop-like or spiral arms, called brachia, and by the movement of fine ciliated (hair-like) processes on their outer edges conduct small food particles to the mouth. The brachia are supported by shelly processes, to which are attached, in the Spirifers, delicate spirally coiled ribbons. These internal structures are often beautifully preserved, even though they are so delicate, from the fact that on the death of the animal the commissure or opening round the valves is so tightly closed as to prevent the coarse mud from penetrating while permitting the finer silt, and more rarely mineral matter in solution, to pass, and subsequently to be deposited within the cavity. At the Murray River cliffs in South Australia, a bed of Cainozoic limestone contains many of these brachiopod shells in a unique condition, for the hollow valves have been filled in with a clear crystal of selenite or « 59 » gypsum, through which may be seen the loop or brachial support preserved in its entirety.
The Sea-mats or Polyzoa, represented by Retepora (the Lace-coral) (Fig. 24) and Flustra (the Sea-mat) of the present sea-shore, have a calcareous skeleton, or zoarium, which is easily preserved as a fossil. Polyzoa are very abundant in the Cainozoic beds of Australia, New Zealand, and elsewhere (Fig. 25). In the Mesozoic series, on the other hand, they are not so well represented; but in Europe and North America they play an important part in forming the Cretaceous and some Jurassic strata by the abundance of their remains.
WORMS (VERMES).—The hard, calcareous tubes of Sea-worms, the Polychaeta (“many bristles”) are often found in fossiliferous deposits, and sometimes form large masses, due to their gregarious habits of life; they also occur attached to shells such as oysters (Fig. 26). The burrows of the wandering worms are found in Silurian strata in Australia; and the sedentary forms likewise occur from the Devonian upwards.
ECHINODERMATA.—Sea-urchins (Echinoidea) possess a hard, calcareous, many-plated test or covering and, when living are covered with spines (Fig. 27). Both the tests and spines are found fossil, the former sometimes whole when the sediment has been quietly thrown down upon them; but more frequently, as in the Shepherd’s crown type (Cidaris), are found in disjointed plates, owing to the fact that current action, going on during entombment has caused the plates to separate. The spines are very rarely found attached to the test, more frequently « 60 » being scattered through the marl or sandy clay in which the sea-urchins are buried. The best conditions for the preservation of this group is a marly limestone deposit, in which case the process of fossilisation would be tranquil (Fig. 28).
The true Starfishes (Asteroidea), are either covered with calcareous plates, or the skin is hardened by rough tubercles; and these more lasting portions are preserved in rocks of all ages. The shape of the animal is also often preserved in an exquisite manner in beds of fine mud or clay.
The Brittle-stars (Ophiuroidea) have their body covered with hard, calcareous plates. Their remains are found in rocks as old as the Ordovician in Bohemia but their history in Australia begins with the Silurian period (Fig. 29). From thence onward they are occasionally found in successive strata in various parts of the world.
The bag-like echinoderms (Cystidea) form a rare group, restricted to Palaeozoic strata. The plates of the sack, or theca, and those of the slender arms are calcareous, and are capable of being preserved in the fossil state. A few doubtful remains of this group occur in Australia.
The bud-shaped echinoderms (Blastoidea) also occur chiefly in Devonian and Carboniferous strata. This is also a rare group, and is represented by several forms found only in New South Wales and Queensland.
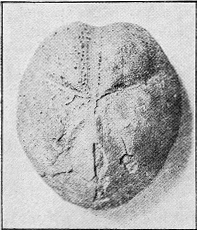
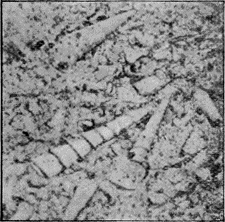
The well known and beautiful fossil forms, the Stone-lilies (Crinoidea) have a very extended geological history, beginning in the Cambrian; whilst a few species are living in the ocean at the present day. The many-jointed skeleton lends itself well to fossilisation, and remains of the crinoids are common in Australia mainly in Palaeozoic strata (Fig. 30). « 62 » In Europe they are found abundantly also in Jurassic strata, especially in the Lias.
 |
Fig. 30.
A Fossil Crinoid (Taxocrinus simplex, Phillips sp.) About 1/2 nat. size. Wenlock Limestone (Silurian), Dudley, England. (Nat. Mus. Coll.)
|


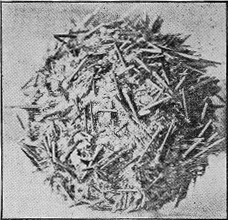
HYDROZOA.—The Graptolites (“stone-writing”) have a chitinous skin (periderm) to the body or hydrosome, which is capable of preservation to a remarkable degree; for their most delicate structures are preserved on the surfaces of the fine black mud deposits which subsequently became hardened into slates. In Australia graptolites occur from the base of the Ordovician to the top of the Silurian (Fig. 31).
Another section of the Hydrozoa is the Stromatoporoidea. These are essentially calcareous, and their structure reminds one of a dense coral. The « 64 » polyps build their tiers of cells (coenosteum) in a regular manner, and seem to have played the same part in the building of ancient reefs in Silurian, Devonian and Carboniferous times as the Millepora at the present day (Fig. 32).
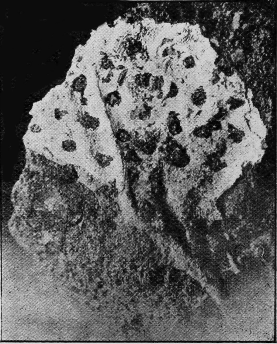
ANTHOZOA.—The true Corals have a stony skeleton, and this is capable of easy preservation as a fossil. There is hardly any fossiliferous stratum of importance which has not its representative corals. In Australia their remains are especially abundant in the Silurian, Devonian (Fig. 33), and Carboniferous formations, and again in the Oligocene and Miocene.
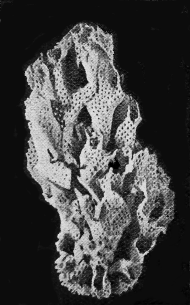
SPONGES.—The framework of the sponge may consist either of flinty, calcareous, or horny material (Fig. 34). The two former kinds are well represented in our Australian rocks, the first appearing in the Lower Ordovician associated with graptolites, and « 65 » again in the Cretaceous and Tertiary rocks (Fig. 35); whilst the calcareous sponges are found in Silurian strata, near Yass, and again in the Cainozoic beds of Flinders, Curlewis and Mornington in Victoria.
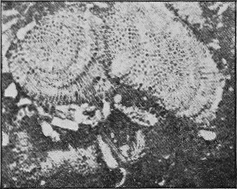
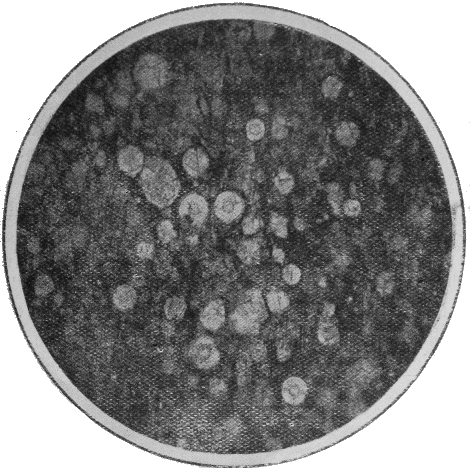
PROTOZOA.—The important and widely-distributed group of the Foraminifera (“hole-bearers”) belonging to the lowest phylum, the Protozoa, generally possess a calcareous shell. The tests range in size from tiny specks of the fiftieth of an inch in diameter, to the giant Nummulite, equalling a five shilling piece in size (Fig. 36). Their varied and beautiful forms are very attractive, but their great interest lies in their multifarious distribution in all kinds of sediments: they are also of importance because certain of the more complex forms indicate « 66 » distinct life zones, being restricted to particular strata occurring in widely-separated areas.
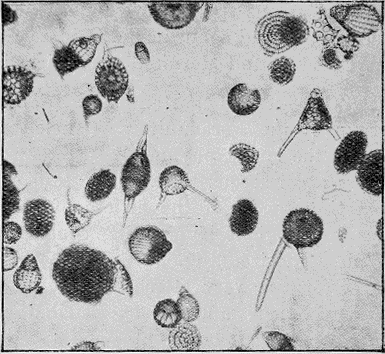
Members of the allied order of the Radiolaria have a flinty shell (Fig. 37); and these organisms are often found building up siliceous rocks such as cherts (Fig. 38).
PLANTS.—The harder portions of plants which are found in the fossil state are,—the wood, the coarser vascular (vessel-bearing) tissue of the leaves, and the harder parts of fruits and seeds.
Fossil wood is of frequent occurrence in Palaeozoic, Mesozoic and Cainozoic strata in Australia, as, for instance, the wood of the trees called Araucarioxylon and Dadoxylon in the Coal measures of New South Wales (see antea, Fig. 3).

Fossil leaves frequently occur in pipe-clay beds, as at Berwick, Victoria, and in travertine from near Hobart, Tasmania (Fig. 39). Fossil fruits are found in abundance in the ancient river gravels at several hundreds of feet below the surface, in the “deep leads” of Haddon, Victoria, and other localities in New South Wales, Queensland and Tasmania.


FOSSILIFEROUS ROCKS.
Section I.—ARGILLACEOUS ROCKS.
Under this head are placed the muds, clays, mudstones, shales and slates. MUDS are usually of a silty nature, that is, containing a variable proportion of sand (quartz) grains. Such are the estuarine muds of Pleistocene and Recent age, containing brackish water foraminifera and ostracoda, and those shells of the mollusca usually found associated with brackish conditions. Lacustrine mud can be distinguished by the included freshwater shells, as Limnaea, Coxiella (brackish), Cyclas and Bulinus, as well as the freshwater ostracoda or cyprids (Fig. 40).
CLAYS are tenacious mud deposits, having the general composition of a hydrous silicate of alumina with some iron. When a clay deposit tends to split into leaves or laminae, either through moderate pressure or by the included fossil remains occupying distinct planes in the rock, they are called SHALES.
Clays and Shales of marine origin are often crowded with the remains of mollusca. The shells are sometimes associated with leaves and other vegetable remains, if forming part of an alternating series of freshwater and marine conditions. An example of this type of sediments is seen in the Mornington beds of the Balcombian series in Victoria.
MUDSTONE is a term applied to a hardened clay deposit derived from the alteration of an impure limestone, and is more often found in the older series of rocks. Mudstones are frequently crowded with fossils, but owing to chemical changes within the rock, the calcareous organisms are as a rule represented by casts and moulds. At times these so faithfully represent the surface and cavities of the organism that they are almost equivalent to a well preserved fossil (Fig. 41).
SLATE.—When shale is subjected to great pressure, a plane of regular splitting called cleavage is induced, which is rarely parallel to the bedding plane or surface spread out on the original sea-floor: the cleavage more often taking place at an appreciable angle to the bedding plane. The graptolitic rocks of Victoria are either shales or slates, according to the absence or development of this cleavage structure in the rock.
Section II.—SILICEOUS ROCKS.
In this group are comprised all granular quartzose sediments, and organic rocks of flinty composition.
SANDSTONES.—Although the base of this type of rock is formed of quartz sand, it often contains fossils. Owing to its porous nature, percolation of water containing dissolved CO2 tends to bring about the solution of the calcareous shells, with the result that only casts of the shells remain.
FLINTS and CHERTS.—These are found in the form of nodules and bands in other strata, principally in limestone. In Europe, flint is usually found in the Chalk formation, whilst chert is found in the Lower Greensands, the Jurassics, the Carboniferous Limestone and in Cambrian rocks. In Australia, flint occurs in the Miocene or Polyzoal-rock formation of Mount Gambier, Cape Liptrap and the Mallee borings. Flint is distinguished from chert by its being black in the mass, often with a white crust, and translucent in thin flakes; chert being more or less granular in texture and sub-opaque in the mass. Both kinds appear to be formed as a pseudomorph or replacement of a portion of the limestone stratum by silica, probably introduced in solution as a soluble alkaline silicate. Both flint and chert often contain fossil shells and other organic remains, such as radiolaria and sponge-spicules, which can be easily seen with a lens in thin flakes struck off by the hammer.
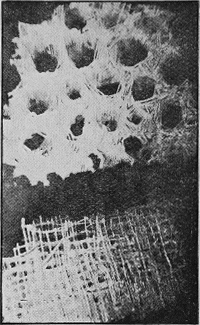
DIATOMITE is essentially composed of the tiny frustules or flinty cases of diatoms (unicellular algae), usually admixed with some spicules of the freshwater sponge, Spongilla. It generally forms a layer at the bottom of a lake bed (Fig. 42).

Section III.—CALCAREOUS ROCKS.
LIMESTONES FORMED BY ORGANISMS.—Organic limestones constitute by far the most important group of fossiliferous rocks. Rocks of this class are composed either wholly of carbonate of lime, or contain other mineral matter also, in varying proportion. Many kinds of limestones owe their origin directly to the agency of animals or plants, which extracted the calcareous matter from the water in « 73 » which they lived in order to build their hard external cases, as for example the sea-urchins; or their internal skeletons, as the stony corals. The accumulated remains of these organisms are generally compacted by a crystalline cement to form a coherent rock.
The chief groups of animals and plants forming such limestone rocks are:—
(a) FORAMINIFERA.—Example. Foraminiferal limestone as the Nummulitic limestone of the Pyramids of Egypt, or the Lepidocyclina limestone of Batesford, near Geelong, Victoria (Fig. 43).
(b) CORALS.—Ex. “Madrepore limestone,” or Devonian marble, with Pachypora. Also the Lilydale limestone, with Favosites, of Silurian age, Victoria (Fig. 44).
(c) STONE-LILIES.—Ex. Crinoidal or Entrochial limestone, Silurian, Toongabbie, Victoria (Fig. 45). Also the Carboniferous or Mountain limestone, Derbyshire, England.
(d) WORM-TUBES.--Ex. Serpulite limestone of Hanover, Germany. Ditrupa limestone of Torquay and Wormbete Creek, Victoria.
(e) POLYZOA.—-Ex. Polyzoal limestone, as the so-called Coralline Crag of Suffolk, England; and the Polyzoal Rock of Mount Gambier, S. Australia.
(f) BRACHIOPODA.—Ex. Brachiopod limestone of Silurian age, Dudley, England. Orthis limestone of Cambrian age, Dolodrook River, N. E. Gippsland.
(g) MOLLUSCA.—Ex. Shell limestone, as the Turritella bed of Table Cape, Tasmania, and of Camperdown, Victoria (Fig. 46), or the Purbeck Marble of Swanage, Dorset, England.
(h) OSTRACODA.—Ex. Cypridiferous limestone, formed of the minute valves of the bivalved ostracoda, as that of Durlston, Dorset, England (Fig. 47).
(i) CADDIS FLY LARVAE.—Ex. Indusial limestone, formed of tubular cases constructed by the larvae of the Caddis fly (Phryganea). Occurs at Durckheim, Rhine District, Germany.
(j) RED SEAWEEDS.—Ex. Nullipore limestone, formed by the stony thallus (frond) of the calcareous sea-weed Lithothamnion, as in the Leithakalk, a common building stone of Vienna.
(k) GREEN SEAWEEDS.—Ex. Halimeda limestone, forming large masses of rock in the late Cainozoic reefs of the New Hebrides (Fig. 48).
(l) (?) BLUE-GREEN SEAWEEDS.—Ex. Girvanella limestone, forming the Peagrit of Jurassic age, of Gloucester, England.
Section IV.—CARBONACEOUS and MISCELLANEOUS ROCKS.
COALS and KEROSENE SHALES (Cannel Coal).—These carbonaceous rocks are formed in much the same way as the deposits in estuaries and lagoon swamps. They result from the sometimes vast aggregation of vegetable material (leaves, wood and fruits), brought down by flooded rivers from the surrounding country, which form a deposit in a swampy or brackish area near the coast, or in an estuary. Layer upon layer is thus formed, alternating with fine mud. The latter effectually seals up the organic layers and renders their change into a carbonaceous deposit more certain.
When shale occurs between the coal-layers it is spoken of as the under-clay, which in most cases is the ancient sub-soil related to the coal-layer immediately above. It is in the shales that the best examples of fossil ferns and other plant-remains are often found. The coal itself is composed of a partially decomposed mass of vegetation which has become hardened and bedded by pressure and gradual drying.
Spore coals are found in thick deposits in some English mines, as at Burnley in Yorkshire. They result from the accumulation of the spores of giant club-mosses which flourished in the coal-period. They « 77 » are generally referred to under the head of Cannel Coals. The “white coal” or Tasmanite of the Mersey Basin in Tasmania is an example of an impure spore coal with a sandy matrix (Fig. 49).
The Kerosene Shale of New South Wales is related to the Torbanite of Scotland and Central France. It « 78 » occurs in lenticular beds between the bituminous coal. It is a very important deposit, commercially speaking, for it yields kerosene oil, and is also used for the manufacture of gas. The rock is composed of myriads of little cell-bodies, referred to as Reinschia, and first supposed to be allied to the freshwater alga, Volvox; but this has lately been questioned, and an alternative view is that they may be the megaspores of club-mosses (Fig. 50).
The coals of Jurassic age in Australia are derived from the remains of coniferous trees and ferns; and some beautiful examples of these plants may often be found in the hardened clay or shale associated with the coal seams.
The Brown Coals of Cainozoic or Tertiary age in Australia are still but little advanced from the early stage, lignite. The leaves found in them are more or less like the present types of the flora. The wood is found to be of the Cypress type (Cupressinoxylon). In New Zealand, however, important deposits of coal of a more bituminous nature occur in the Oligocene of Westport and the Grey River Valley, in the Nelson District.
BONE BEDS.—The bones and excreta of fish and reptiles form considerable deposits in some of the sedimentary formations; especially those partly under the influence of land or swamp conditions. They constitute a kind of conglomerate in which are found bone-fragments and teeth (Fig. 51). These bone-beds are usually rich in phosphates, and are consequently valuable as a source of manure. The Miocene bone-bed with fish teeth at Florida, U.S.A., « 79 » is a notable example. The nodule bed of the Victorian Cainozoics contains an assemblage of bones of cetaceans (whales, etc.).

BONE BRECCIAS.—These are usually formed of the remains of the larger mammals, and consist of a consolidated mass of fragments of bones and teeth embedded in a calcareous matrix. Bone-breccias are of frequent occurrence on the floors of caves which had formerly been the resort of carnivorous animals, and into which they dragged their prey. The surface water percolating through the overlying calcareous strata dissolved a certain amount of lime, and this was re-deposited on the animal remains lying scattered over the cave floor. A deposit so formed constitutes a stalagmite or floor encrustation. As examples of bone-breccias we may refer to the limestone at Limeburners Point, Geelong (Fig. 52); and the stalagmitic deposits of the Buchan Caves.
IRONSTONE.—Rocks formed almost entirely of limonite (hydrated peroxide of iron) are often due to the agency of unicellular plants known as diatoms, which separate the iron from water, and deposit it as hydrous peroxide of iron within their siliceous skeletons. In Norway and Sweden there are large and important deposits of bog iron-ore, which have presumably been formed in the beds of lakes.

Clay ironstone nodules (sphaerosiderite) have generally been formed as accretions around some « 81 » decaying organic body. Many clay ironstone nodules, when broken open, reveal a fossil within, such as a coprolitic body, fern frond, fir-cone, shell or fish.
Oolitic ironstones are composed of minute granules which may have originally been calcareous grains, formed by a primitive plant or alga, but since replaced by iron oxide or carbonate.
The Tertiary ironstone of western Victoria is found to contain leaves, which were washed into lakes and swamps (Fig. 53); and the ferruginous groundmass may have been originally due to the presence of diatoms, though this yet remains to be proved.
FOSSIL PLANTS.
Cambrian Plants.—
The oldest Australian plant-remains belong to the genus Girvanella. This curious little tubular unicellular organism, once thought to be a foraminifer, shows most affinity with the blue-green algae (Cyanophyceae), an important type of plant even now forming calcareous deposits such as the calcareous grains on the shores of the Salt Lake, Utah, and the pea-grit of the Carlsbad hot springs. Girvanella problematica occurs in the Lower Cambrian limestones of South Australia, at Ardrossan and elsewhere.
Silurian Plants.—
Amongst Silurian plants may be mentioned the doubtful sea-weeds known as Bythotrephis. Their branch-like impressions are fairly common in the mudstones of Silurian age found in and around Melbourne. They generally occur in association with shallow-water marine shells and crustacea of that period.
The genus Girvanella before mentioned is also found in the Silurian (Yeringian) of Lilydale and the Tyers River limestone, Victoria (Fig. 54).
Haliserites is a primitive plant of the type of the club-mosses so common in the rocks of the Carboniferous period. This genus is found in some abundance in the Yeringian stage of the Silurian in Gippsland (Fig. 55).
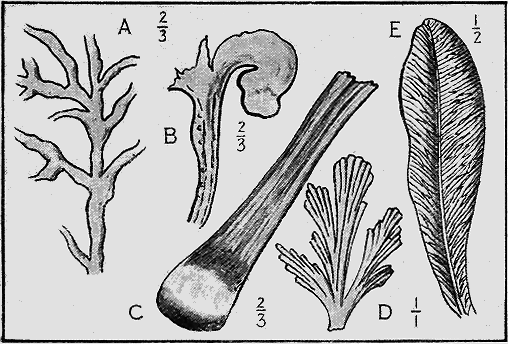
 |
Fig. 56.—Restoration of Lepidodendron elegans. (After Grand’Eury.)
|

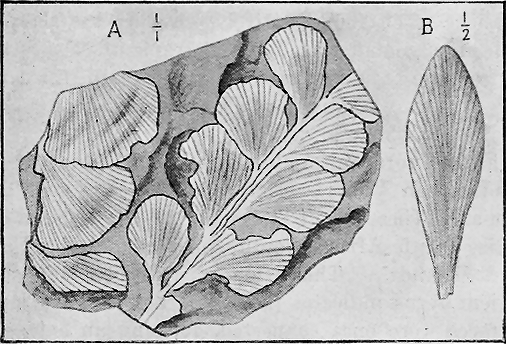
Devonian and Carboniferous Plants.—
Plant-life was not abundant, however, until Upper Devonian and Carboniferous times. In the rocks of these periods we meet with the large strap-shaped leaves of Cordaites and a fern, Sphenopteris, in the first-named series; and the widely distributed Lepidodendron with its handsome lozenge-scarred stems in the later series (Fig. 56). Cordaites has been found in Victoria in the Iguana Creek beds (Upper Devonian), and it also probably occurs at the same horizon at Nungatta, New South Wales. Lepidodendron occurs in the Lower Carboniferous sandstone of Victoria and Queensland (Fig. 57): in New South Wales it is found at Mt. Lambie, Goonoo, Tamworth and Copeland in beds generally regarded as Upper « 86 » Devonian. Both of these plants are typical of Carboniferous (Coal Measure) beds in Europe and North America. The fern Rhacopteris is characteristic of Upper Carboniferous shales and sandstones near Stroud, and other localities in New South Wales as well as in Queensland (Fig. 58). These beds yield a few inferior seams of coal. Girvanella is again seen in the oolitic limestones of Carboniferous age in Queensland and New South Wales.
Carbopermian Plants.—
The higher division of the Australian Carboniferous usually spoken of as the Permo-carboniferous, and here designated the Carbopermian (see Footnote 2, page 48), is typified by a sudden accession of plant forms, chiefly belonging to ferns of the Glossopteris type. The lingulate or tongue-shaped fronds of this genus, with their characteristic reticulate venation, are often found entirely covering the slabs of shale intercalated with the coal seams of New South Wales; and it is also a common fossil in Tasmania and Western Australia. The allied form, Gangamopteris, which is distinguished from Glossopteris by having no definite midrib, is found in beds of the same age in Victoria, New South Wales, and Tasmania. These plant remains are also found in India, South Africa, South America and the Falkland Islands. This wide distribution of such ancient ferns indicates that those now isolated land-surfaces were once connected, forming an extensive continent named by Prof. Suess “Gondwana-Land,” from the Gondwana district in India (Fig. 59).
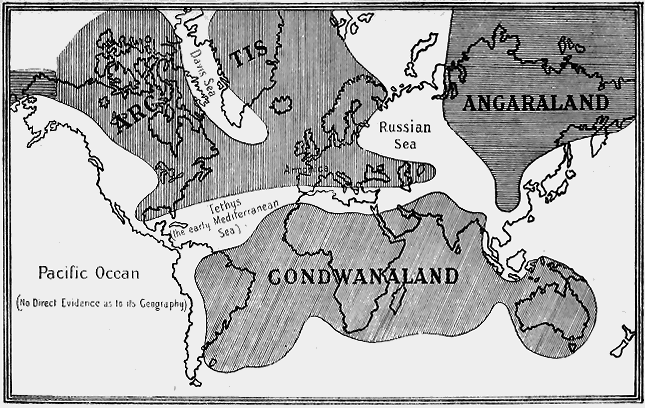 |
||
| E. M. del. | (After J. W. Gregory). | |
| Fig. 59—Map of the World in the Upper Carboniferous Era. | ||
Triassic Plants.—
The widely distributed pinnate fern known as Thinnfeldia is first found in the Trias; in the Narrabeen shales near Manly, and the Hawksbury sandstone at Mount Victoria, New South Wales. It is also a common fossil of the Jurassic of South Gippsland, and other parts of Victoria. The grass-like leaves of Phoenicopsis are frequently met with in Triassic strata, as in the upper series at Bald Hill, Bacchus Marsh, and also in Tasmania. The large Banana-palm-like leaves of Taeniopteris (Macrotaeniopteris) are common to the Triassic and Lower Jurassic beds of India: they are also met with in New Zealand, and in the upper beds at Bald Hill, Bacchus Marsh.
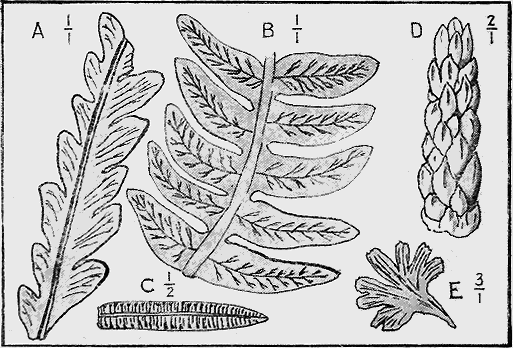
Jurassic Plants.—
The Jurassic flora of Australasia is very prolific in plant forms. These range from liverworts and horse-tails to ferns and conifers. The commonest ferns were Cladophlebis, Sphenopteris, Thinnfeldia and Taeniopteris. The conifers are represented by Araucarites (cone-scales, leaves and fruits), Palissya and Brachyphyllum (Fig. 60). The Ginkgo or Maiden-hair tree, which is still living in China and Japan, and also as a cultivated plant, was extremely abundant in Jurassic times, accompanied by the related genus, Baiera, having more deeply incised leaves; both genera occur in the Jurassic of S. Gippsland, Victoria, and in Queensland. The Jurassic flora of Australasia is in many respects like that of the Yorkshire coast near Scarborough. In New Zealand this flora is represented in the Mataura series, in which there are many forms identical with those of the Australian Jurassic, and even of India.
Cretaceous Plants.—
An upper Cretaceous fern, (?) Didymosorus gleichenioides, is found in the sandstones of the Croydon Gold-field, North Queensland.
Plants of the Cainozoic.—Balcombian Stage.—
The older part of the Cainozoic series in Australasia may be referred to the Oligocene. These are marine beds with occasional, thick seams of lignite, and sometimes of pipe-clay with leaves, the evidence of river influence in the immediate neighbourhood. The fossil wood in the lignite beds appears to be a Cupressinoxylon or Cypress wood. Leaves referable « 90 » to plants living at the present day are also found in certain clays, as at Mornington, containing Eucalyptus precoriacea and a species of Podocarpus.
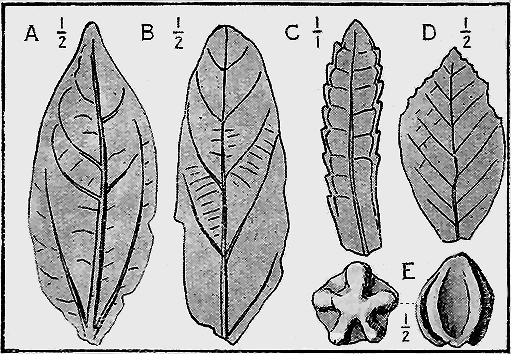
Miocene Leaf-beds.—Janjukian Stage.—
Later Cainozoic deposits, evidently accumulated in lakes, and sometimes ferruginous, may be referred to the Miocene. They are comparable in age with the Janjukian marine beds of Spring Creek and Waurn Ponds in Victoria. These occur far inland and occupy distinct basins, as at the Wannon, Bacchus Marsh (Maddingley), and Pitfield Plains. Leaf-beds of this age occur also on the Otway coast, Victoria, containing the genera Coprosmaephyllum, Persoonia and Phyllocladus. In all probability the Dalton and « 91 » Gunning leaf-beds of New South Wales belong here. Examples of the genera found in beds of this age are Eucalyptus (a species near E. amygdalina), Banksia or Native Honeysuckle, Cinnamomum or Cinnamon, Laurus or Laurel, and Fagus (Notofagus) or Beech (Fig. 61). In the leaf-beds covered by the older basalt on the Dargo High Plains, Gippsland, leaves of the Ginkgo Murrayana occur.
In South Australia several occurrences of leaf beds have been recorded, containing similar species to those found in the Cainozoic of Dalton and Vegetable Creek, New South Wales. For example, Magnolia Brownii occurs at Lake Frome, Bombax Sturtii and Eucalyptus Mitchelli at Elizabeth River, and Apocynophyllum Mackinlayi at Arcoona.
Fruits of the “Deep Leads.”—
The Deep Leads of Victoria, New South Wales and Tasmania probably begin to date from the period just named, for they seem to be contemporaneous with the “Older Gold Drift” of Victoria; a deposit sometimes containing a marine fauna of Janjukian age. This upland river system persisted into Lower Pliocene times, and their buried silts yield many fruits, of types not now found in Australia, such as Platycoila, Penteune and Pleioclinis, along with Cupressus (Spondylostrobus) and Eucalyptus of the existing flora (Fig. 62).
Pleistocene Plants.—
The Pleistocene volcanic tuffs of Mount Gambier have been shown to contain fronds of the living Pteris (Pteridium) aquilina or Bracken fern, and a Banksia in every way comparable with B. marginata, a « 92 » species of the Native Honeysuckle still living in the same district.

The siliceous valves of freshwater diatoms constitute the infusorial earths of Victoria, Queensland, New South Wales and New Zealand. The commonest genera met with are Melosira, Navicula, Cymbella (or Cocconema), Synedra, Tabellaria, Stauroneis and « 93 » Gomphonema. They are, generally speaking, of Pleistocene age, as they are often found filling hollows in the newer basalt flows. In Victoria diatomaceous earths are found at Talbot (See Fig. 42), Sebastopol and Lancefield; in Queensland, at Pine Creek; in New South Wales, at Cooma, Barraba, and the Richmond River; and in New Zealand at Pakaraka, Bay of Islands. In the latter country there is also a marine diatomaceous rock in the Oamaru Series, of Miocene age.
COMMON OR CHARACTERISTIC FOSSILS OF THE FOREGOING CHAPTER.
Girvanella problematica, Nicholson and Etheridge. Cambrian: S. Australia.
Bythotrephis tenuis, J. Hall. Silurian: Victoria.
Haliserites Dechenianus, Göppert sp. Silurian and Devonian: Victoria.
Cordaites australis, McCoy. Upper Devonian: Victoria.
Lepidodendron australe, McCoy. Lower Carboniferous: Victoria and Queensland. Up. Devonian: New South Wales.
Rhacopteris inaequilatera, Göppert sp. Carboniferous: New South Wales.
Glossopteris Browniana, Brongniart. Carbopermian: New South Wales, Queensland, Tasmania and W. Australia.
Gangamopteris spatulata, McCoy. Carbopermian: Victoria, New South Wales and Tasmania.
Thinnfeldia odontopteroides, Morris sp. Triassic: New South Wales. Jurassic: Victoria, Queensland and Tasmania.
Cladophlebis denticulata., Brongn. sp., var. australis, Morris. Jurassic: Queensland, New South Wales, Victoria, Tasmania and New Zealand.
Taeniopteris spatulata, McClelland. Jurassic: Queensland, New South Wales, Victoria, and Tasmania.
(?) Didymosorus gleichenioides, Etheridge fil. Upper Cretaceous: Queensland.
Eucalyptus precoriacea, Deane. Oligocene: Victoria.
Eucalyptus, Banksia, Cinnamomum, Laurus and Fagus. Miocene: Victoria, New South Wales and Tasmania.
Spondylostrobus Smythi, von Mueller. (Fruits and wood). Lower Pliocene: Victoria and Tasmania.
Pteris (Pteridium) aquilina, Linné, and Banksia cf. marginata, Cavanilles. Pleistocene: Victoria and South Australia.
LITERATURE.
Girvanella.—Etheridge, R. jnr. Trans. R. Soc. S. Australia, vol. XIII. 1890, pp. 19, 20. Etheridge, R. and Card, G. Geol. Surv. Queensland, Bull. No. 12, 1900, pp. 26, 27, 32. Chapman, F. Rep. Austr. Assoc. Adv. Sci., Adelaide Meeting (1907), 1908, p. 337.
Devonian Ferns and Cordaites.—McCoy, F. Prod. Pal. Vict. Dec. V., 1876, p. 21. Dun, W. S. Rec. Geol. Surv. New South Wales, vol. V. pt. 3, 1897, p. 117.
Lepidodendron.—McCoy, F. Prod. Pal. Vict., Dec. I. 1874, p. 37. Etheridge, R. jnr. Rec. Geol. Surv, New South Wales, vol. II., pt. 3, 1891, p. 119. Idem, Geol. and Pal. Queensland, 1892, p. 196.
Carboniferous Fungi.—Etheridge, R. jnr. Geol. Surv. W.A., Bull, No. 10, 1903, pp. 25-31.
Carboniferous Ferns.—Dun, W. S. Rec. Geol. Surv. New South Wales, vol. VIII. pt. 2, 1905, pp. 157-161, pls. XXII. and XXIII.
Glossopteris.—Feistmantel, O. Mem. Geol. Surv. New South Wales, Pal. No. 3, 1890. Arber, N. Cat. Foss. Plants, Glossopteris Flora, Brit. Mus., 1905.
Gangamopteris.—McCoy, F. Prod. Pal. Vict., Dec. II. 1875, p. 11.
Jurassic Plants.—McCoy, F. Prod. Pal. Vic., Dec. II. 1875, p. 15. Woods, T. Proc. Linn. Soc. New South Wales, vol. VIII. pt. I. 1883, p. 37. Etheridge, R. jnr. Geol. Pal. Queensland, 1892, p. 314. Dun, W. S. (Taeniopteris), Rep. Austr. Asso. Adv. Sci., Sydney, 1898, pp. 384-400. Seward, A. C. Rec. Geol. Surv. Vic., vol. I. pt. 3, 1904; Chapman, F. Ibid., vol II. pt. 4, 1908; vol. III., pt. 1, 1909. Dun, W. S. Rec. Geol. Surv. New South Wales, vol. VIII. pt. 4, 1909, p. 311.
Older Cainozoic Plants.—McCoy, F. Prod. Pal. Vic., Dec. IV. 1876, p. 31. Ettingshausen, C. von. Mem. Geol. Surv. New South Wales, Pal. No. 2, 1888. Idem, Trans. New Zealand Inst., vol. XXIII. (1890), 1891, p. 237. Deane, H. Rec. Geol. Surv. Vict., vol. I. pt. 1, 1902, pp. 15, 20.
Lower Pliocene Deep Leads.—McCoy, F. Prod. Pal. Vict., Dec. IV. 1876, p. 29. Mueller, F. von. Geol. Surv. Vic., New Veg. Foss., 1874 and 1883.
Pleistocene and other Diatom Earths.—Card, G. W. and Dun, W. S., Rec. Geol. Surv. New South Wales, vol. V. pt. 3, 1897, p. 128.
FOSSIL FORAMINIFERA AND RADIOLARIA.
Protozoans, Their Structure.—
The animals forming the sub-kingdom PROTOZOA (“lowliest animals”), are unicellular (one-celled), as distinguished from all the succeeding higher groups, which are known as the METAZOA (“animals beyond”). The former group, Protozoa, have all their functions performed by means of a simple cell, any additions to the cell-unit merely forming a repetitional or aggregated cell-structure. A familiar example of such occurs in pond-life, in the Amoeba, a form which is not found fossilised on account of the absence of any hard parts or covering capable of preservation. Foraminifera and Radiolaria, however, have such hard parts, and are frequently found fossilised.
Foraminifera: Their Habitats.—
The FORAMINIFERA are a group which, although essentially one-celled, have the protoplasmic body often numerously segmented. The shell or test formed upon, and enclosing the jelly-like sarcode, may consist either of carbonate of lime, cemented sand-grains, or a sub-calcareous or chitinous (horny) covering. The Foraminifera, with very few exceptions, as Mikrogromia, Lieberkuehnia, and some forms of Gromia, are all marine in habit. Some « 96 » genera, however, as Miliolina, Rotalia and Nonionina, affect brackish water conditions.
Since Foraminifera are of so lowly a grade in the animal kingdom, we may naturally expect to find their remains in the oldest known rocks that show any evidence of life. They are, indeed, first seen in rocks of Cambrian age, although they have not yet been detected there in Australian strata.
Cambrian Foraminifera.—
In parts of Siberia and in the Baltic Provinces, both Cambrian and Ordovician rocks contain numerous glauconite casts of Foraminifera, generally of the Globigerina type of shell. In England some Middle Cambrian rocks of Shropshire are filled with tiny exquisitely preserved spiral shells belonging to the genus Spirillina, in which all the characters of the test are seen as clearly as in the specimens picked out of shore-sand at the present day.
Silurian Foraminifera.—
The Silurian rocks in all countries are very poor in foraminiferal shells, only occasional examples being found. In rocks of this age at Lilydale, Victoria, the genus Ammodiscus, with fine sandy, coiled tests, is found in the Cave Hill Limestone.
So far as known, hardly any forms of this group occur in Devonian strata, although some ill-defined shells have been found in the Eifel, Germany.
Carboniferous Foraminifera.—
The Carboniferous rocks in many parts of the world yield an abundant foraminiferal fauna. Such, for instance, are the Saccammina and Endothyra Limestones of the North of England and the North « 97 » of Ireland. The Australian rocks of this age have not afforded any examples of the group, since they are mainly of estuarine or freshwater origin.

Carbopermian Foraminifera.—
In Australia, as at Pokolbin, New South Wales, in the Mersey River district, Tasmania, and in the Irwin River district, Western Australia, the Permian rocks, or “Permo-carboniferous” as they are generally called, often contain beds of impure limestone crowded with the chalky white tests of Nubecularia: other interesting genera occur at the first named locality as Pelosina, Hyperammina, Haplophragmium, Placopsilina, Lituola, Thurammina, Ammodiscus, Stacheia, Monogenerina, Valvulina, Bulimina, « 98 » (?)Pleurostomella, Lagena, Nodosaria, Frondicularia, Geinitzina, Lunucammina, Marginulina, Vaginulina, Anomalina and Truncatulina. The sandy matrix of certain Glossopteris leaf-beds in the Collie Coal measures in W. Australia have yielded some dwarfed examples belonging to the genera Bulimina, Endothyra, Valvulina, Truncatulina and Pulvinulina; whilst in the Irwin River district similar beds contain Nodosaria and Frondicularia (Fig. 63).
Triassic Foraminifera.—
The Triassic and Rhaetic clays of Europe occasionally show traces of foraminiferal shells, probably of estuarine habitat, as do the Wianamatta beds of New South Wales, which also belong to the Triassic epoch. The Australian representatives are placed in the genera Nubecularia, Haplophragmium, Endothyra, Discorbina, Truncatulina, and Pulvinulina. These shells are diminutive even for foraminifera, and their starved condition indicates uncongenial environment.
Jurassic Foraminifera.—
The Jurassic limestones of Western Australia, at Geraldton, contain many species of Foraminifera, principally belonging to the spirally coiled and slipper-shaped Cristellariae. Other genera present are Haplophragmium, Textularia, Bulimina, Flabellina, Marginulina, Vaginulina, Polymorphina, Discorbina, and Truncatulina.
Cretaceous Foraminifera.—
In the Lower Cretaceous rocks known as the Rolling Downs Formation in Queensland, shells of the Foraminifera are found in some abundance at Wollumbilla. They are represented chiefly by Cristellaria and Polymorphina.
Cainozoic Foraminifera.—
The Cainozoic strata in all parts of the world are very rich in Foraminifera, and the genera, and even many species are similar to those now found living. Certain types, however, had a restricted range, and are therefore useful as indicators of age. Such are the Nummulites and the Orbitoides of the Eocene and the Oligocene of Europe, India and the West Indies; and the Lepidocyclinae of the Miocene of Europe, India, Japan and Australia (Fig. 64).
The genus Lepidocyclina is typically represented in the Batesford beds near Geelong, Victoria by L. tournoueri, a fossil of the Burdigalian stage (Middle Miocene) in Europe, as well as by L. marginata. A limestone with large, well-preserved tests of the same genus, and belonging to a slightly lower horizon in the Miocene has lately been discovered in Papua.

Some of the commoner Foraminifera found in the Cainozoic beds of Southern Australia are—Miliolina vulgaris, Textularia gibbosa, Nodosaria affinis, Polymorphina elegantissima, Truncatulina ungeriana and Amphistegina lessonii (Fig. 65). The first-named has a chalky or porcellanous shell; the second a sandy test; the third and fourth glassy or hyaline shells with excessively fine tubules; the fifth a glassy shell « 101 » with numerous surface punctations due to coarser tubules than usual in the shell-walls; whilst the last-named has a smooth, lenticular shell, also hyaline, and occurring in such abundance as often to constitute a foraminiferal rock in itself.
Pleistocene Foraminifera.—
The estuarine deposits of Pleistocene age in southern Australia often contain innumerable shells of Miliolina, Rotalia and Polystomella. One thin seam of sandy clay struck by the bores in the Victorian Mallee consists almost entirely of the shells of the shallow-water and estuarine species, Rotalia beccarii.
Radiolaria: Their Structure.—
The organisms belonging to the order RADIOLARIA are microscopic, and they are all of marine habitat. The body of a radiolarian consists of a central mass of protoplasm enclosed in a membranous capsule, and contains the nuclei, vacuoles, granules and fat globules; whilst outside is a jelly-like portion which throws off pseudopodia or thin radiating threads. The skeleton of Radiolaria is either chitinous or composed of clear, glassy silica, and is often of exquisitely ornamental and regular form.
Habitat.—
These tiny organisms generally live in the open ocean at various depths, and sinking to the bottom, sometimes as deep as 2,000 to 4,000 fathoms, they form an ooze or mud.
Subdivisions.—
Radiolaria are divided into the four legions or orders,—Acantharia, Spumellaria, Nasselaria and Phaeodaria: only the second and third groups are found fossil. The Spumellarians are spherical, ellipsoidal, or disc-shaped, and the Nasselarians conical or helmet-shaped.
Cambrian Radiolaria.—
Certain cherts or hard, siliceous rocks of the palaeozoic era are often crowded with the remains of Radiolaria, giving the rock a spotted appearance. (See antea, Fig. 38). Some of the genera thus found are identical with those living at the present day, whilst others are peculiar to those old sediments. In Australia, remains of their siliceous shells have been found in cherts of Lower Cambrian age near Adelaide. These have been provisionally referred to the genera Carposphaera and Cenellipsis (Fig. 66).
Ordovician Radiolaria.—
Radiolaria have been detected in the Lower Ordovician rocks of Victoria, in beds associated with the Graptolite slates of this series. In New South Wales Radiolarian remains are found in the cherts and slates of Upper Ordovician age at Cooma and Mandurama.
Silurian Radiolaria.—
The Silurian black cherts of the Jenolan Caves in New South Wales contain casts of Radiolaria.
Devonian Radiolaria.—
The Lower Devonian red jaspers of Bingera and Barraba in New South Wales have afforded some casts of Radiolaria, resembling Carposphaera and Cenosphaera.
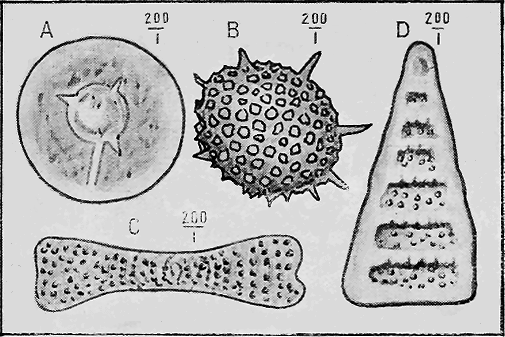
The large number of fifty-three species have been found in the radiolarian rocks of Middle Devonian age at Tamworth in New South Wales (Fig. 66). These have been referred to twenty-nine genera comprising amongst others, Cenosphaera, Xiphosphaera, Staurolonche, Heliosphaera, Acanthosphaera and Spongodiscus.
Cretaceous Radiolaria.—
Although certain silicified rocks in the Jurassic in Europe have furnished a large series of Radiolaria, the Australian marine limestones of this age have not yielded any of their remains up to the present. They have been found, however, in the Lower Cretaceous of Queensland, and in the (?)Upper Cretaceous of Port Darwin, N. Australia. The Radiolaria from the latter locality belong to the sub-orders Prunoidea, « 104 » Discoidea and Cyrtoidea (Fig. 66). The rock which contains these minute fossils is stated to be eaten by the natives for medicinal purposes. As its composition is almost pure silica, its efficacy in such cases must be more imaginary than real.
Cainozoic Radiolaria.—
Cainozoic rocks of Pliocene age, composed entirely of Radiolaria, occur at Barbados in the West Indies. No Cainozoic Radiolaria, however, have been found either in Australia or New Zealand up to the present time.
COMMON OR CHARACTERISTIC FOSSILS OF THE FOREGOING CHAPTER.
FORAMINIFERA.
Nubecularia stephensi, Howchin. Carbopermian: Tasmania and New South Wales.
Frondicularia woodwardi, Howchin. Carbopermian: W. Australia and New South Wales.
Geinitzina triangularis, Chapm. & Howchin. Carbopermian: New South Wales.
Pulvinulina insignis, Chapman. Trias (Wianamatta Series): New South Wales.
Marginulina solida, Terquem. Jurassic: W. Australia.
Flabellina dilatata, Wisniowski. Jurassic: W. Australia.
Vaginulina striata, d’Orbigny. Lower Cretaceous: Queensland.
Truncatulina lobatula, W. and J. sp. Lower Cretaceous: Queensland.
Miliolina vulgaris, d’Orb. sp. Cainozoic: Victoria and S. Australia.
Textularia gibbosa, d’Orb. Cainozoic: Victoria and S. Australia.
Nodosaria affinis, d’Orb. Cainozoic: Victoria and S. Australia.
Polymorphina elegantissima, Parker and Jones. Cainozoic: Victoria, Tasmania, and S. Australia.
Truncatulina ungeriana, d’Orb. sp. Cainozoic: Victoria, King Island, and S. Australia.
Amphistegina lessonii, d’Orb. Cainozoic: Victoria and S. Australia.
Lepidocyclina martini, Schlumberger. Cainozoic (Balcombian and Janjukian): Victoria.
L. tournoueri, Lemoine and Douvillé. Cainozoic (Junjukian): Victoria.
Cycloclypeus pustulosus, Chapman. Cainozoic (Janjukian): Victoria.
Fabularia howchini, Schlumberger. Cainozoic (Kalimnan): Victoria.
Hauerina intermedia, Howchin. Cainozoic (Kalimnan): Victoria.
Rotalia beccarii, Linné sp. Pleistocene: Victoria and S. Australia.
Polystomella striatopunctata, Fichtel and Moll sp. Pleistocene: Victoria and S. Australia.
RADIOLARIA.
(?) Carposphaera sp. Lower Cambrian: South Australia.
(?) Cenellipsis sp. Lower Cambrian: South Australia.
Cenosphaera affinis, Hinde. Devonian: New South Wales.
Staurolonche davidi, Hinde. Devonian: New South Wales.
Amphibrachium truncatum, Hinde. Upper Cretaceous: Northern Territory.
Dictyomitra triangularis, Hinde. Upper Cretaceous: Northern Territory.
LITERATURE.
FORAMINIFERA.
Carbopermian.—Howchin, W. Trans. Roy. Soc. S. Austr., vol. XIX. 1895; pp. 194-198. Chapman, F. and Howchin, W. Mem. Geol. Surv. New South Wales, Pal. No. 14, 1905. Chapman, F. Bull. Geol. Surv. W. Austr., No. 27, 1907, pp. 15-18.
Trias.—Chapman, F. Rec. Geol. Surv. New South Wales, vol. VIII. pt. 4, 1909, pp. 336-339.
Jurassic.—Chapman, F. Proc. Roy. Soc. Vict., vol. XVI. (N.S.), pt. II., 1904, pp. 186-199.
Cretaceous.—Moore, C. Quart. Journ. Geol. Soc., vol. XXVI. 1870, pp. 239 and 242. Howchin, W. Trans. Roy. Soc. S. Austr., vol. VIII. 1886, pp. 79-93. Idem, ibid., vol. XIX., 1895, pp. 198-200. Idem, Bull. Geol. Surv. W. Austr., No. 27, 1907, pp. 38-43.
Cainozoic.—Howchin, W. Trans. Roy. Soc. S. Austr., vol. XII. 1889, pp. 1-20. Idem, ibid., vol. XIV. 1891, pp. 350-356. Jensen, H. I. Proc. Linn. Soc. New South Wales, vol. XXIX. pt. 4, 1905, pp. 829-831. Goddard, E. J. and Jensen, H. I. ibid., vol. XXXII. pt. 2, 1907, pp. 308-318. Chapman, F. Journ. Linn. Soc. Lond. Zool., vol. XXX. 1907, pp. 10-35.
General.—Howchin, W. Rep. Austr. Assoc. Adv. Sci., Adelaide Meeting, 1893, pp. 348-373.
RADIOLARIA.
Lower Cambrian.—David, T. W. E. and Howchin, W. Proc. Linn. Soc. New South Wales, vol. XXI. 1897, p. 571.
Devonian.—David, T. W. E. Proc. Linn. Soc. New South Wales, vol. XXI. 1897, pp. 553-570. Hinde, G. J. Quart. Journ. Geol. Soc., vol. LV. 1890, pp. 38-64.
Upper Cretaceous.—Hinde, G. J. Quart. Journ. Geol. Soc., vol. XLIX. 1893, pp. 221-226.
FOSSIL SPONGES, CORALS AND GRAPTOLITES.
SPONGES.
Characteristics of Sponges.—
The Sponges are sometimes placed by themselves as a separate phylum, the Porifera. With the exception of a few freshwater genera, they are of marine habit and to be found at all depths between low tide (littoral) and deep water (abyssal). Sponges are either fixed or lie loosely on the sea-floor. They possess no organs of locomotion, and have no distinct axis or lateral appendages. They exist by setting up currents in the water whereby the latter is circulated through the system, carrying with it numerous food particles, their tissues being at the same time oxygenated. Their framework, in the siliceous and calcareous sponges, is strengthened by a mineral skeleton, wholly or partially capable of preservation as a fossil.
Cambrian and Ordovician Sponges.—
The oldest rocks in Australia containing the remains of Sponges are the Cambrian limestones of South Australia, at Ardrossan and elsewhere. Some of these sponge-remains are referred to the genus Protospongia, a member of the Hexactinellid group having 6-rayed skeletal elements. When complete, « 108 » the Protospongia has a cup- or funnel-shaped body, composed of large and small modified spicules, which form quadrate areas, often seen in isolated or aggregated patches on the weathered surface of the rock. Protospongia also occurs in the Lower Ordovician slates and shales of Lancefield (P. oblonga), and Bendigo (P. reticulata and P. cruciformis), in Victoria (Fig. 67 A). At St. David’s, in South Wales, the genus is found in rocks of Middle Cambrian age. The South Australian limestones in which Protospongia occurs are usually placed in the Lower Cambrian.

Another genus of Sponges, Hyalostelia, whose affinities are not very clear, occurs in the South Australian Cambrian at Curramulka. This type is represented by the long, slightly bent, rod-like « 109 » spicules of the root-tuft, and the skeletal spicules with six rays, one of which is much elongated.
Stephanella maccoyi is a Monactinellid sponge, found in the Lower Ordovician (Bendigo Series) of Bendigo, Victoria.
Silurian Sponges.—
Numerous Sponges of Silurian age are found in the neighbourhood of Yass, New South Wales, which belong to the Lithistid group, having irregular, knotty and branching spicules. These sponges resemble certain fossil fruits, generally like diminutive melons; their peculiar spicular structure, however, is usually visible on the outside of the fossil, especially in weathered specimens. The commonest genus is Carpospongia.
Receptaculites: Silurian to Carboniferous.—
In Upper Silurian, Devonian, and Carboniferous times the curious saucer- or funnel-shaped bodies known as Receptaculites must have been fairly abundant in Australia, judging by their frequent occurrence as fossils. They are found as impressions or moulds and casts in some of the mudstones and limestones of Silurian age in Victoria, as at Loyola and Wombat Creek, in west and north-east Gippsland respectively. In the Devonian limestones of New South Wales they occur at Fernbrook, near Mudgee, at the Goodradigbee River, and at Cavan, near Yass; also in beds of the same age in Victoria, at Bindi, and Buchan (Fig. 67, B.C.). Receptaculites also occur in the Star Beds of Upper Devonian or Lower Carboniferous age in Queensland, at Mount Wyatt. It will thus be seen that this genus has an extensive geological range.
Carbopermian Sponges.—
A Monactinellid Sponge, provisionally referred to Lasiocladia, has been described from the Gympie beds of the Rockhampton District, Queensland. Lasiocladia, as well as the Hexactinellid Sponge Hyalostelia, occurs in the Carbopermian of New South Wales.
Cretaceous Sponges.—
No sponge-remains seem to occur above the Carbopermian in Australia until we reach the Cretaceous rocks. In the Lower Cretaceous series in Queensland a doubtful member of the Hexactinellid group is found, namely, Purisiphonia clarkei. In the Upper Cretaceous of the Darling Downs District pyritized Sponges occur which have been referred to the genus Siphonia, a member of the Lithistid group, well known in the Cretaceous of Europe.
Cainozoic Sponges.—
A white siliceous clay, supposed to be from a “Deep Lead,” in the Norseman district in Western Australia, has proved to consist almost entirely of siliceous sponge-spicules, belonging to the Monactinellid, the Tetractinellid, the Lithistid, and the Hexactinellid groups (Fig. 69 A, B). The reference of the deposit to a “deep lead” or alluvial deposit presents a difficulty, since these sponge-spicules represent moderately deep water marine forms. This deposit resembles in some respects the spicule-bearing rock of Oamaru, New Zealand, which is of Miocene age.


In the Cainozoic beds of southern Australia Sponges with calcareous skeletons are not at all uncommon. The majority of these belong to the Lithonine section of the Calcispongiae, in which the spicules are regular, and not fixed together. Living examples of these sponges, closely related to the fossils, have been dredged from the Japanese Sea. The fossils are found mainly in the Janjukian, at Curlewis, in the Moorabool River limestones, and in the polyzoal rock of Flinders, all in Victoria. They belong to the genera Bactronella, Plectroninia and Tretocalia (Fig. 68, D and E). Some diminutive forms also occur in the older series, the Balcombian, at Mornington, namely, Bactronella parvula. At Boggy Creek, near Sale, in Victoria, a Tetractinellid Sponge, Ecionema newberyi, is found in the Janjukian marls; spicules of this form have also been noted from the clays of the Altona Bay coal-shaft (Fig. 68 C).
The ARCHAEOCYATHINAE: an ancient class of organisms related both to the Sponges and the Corals.
Archaeocyathinae in Cambrian Strata.—
These curious remains have been lately made the subject of detailed research, and it is now concluded that they form a group probably ancestral both to the sponges and the corals. They are calcareous, and generally cup-shaped or conical, often furnished at the pointed base with roots or strands for attachment to the surrounding reef. They have two walls, both the inner and the outer being perforated like sponges. As in the corals, they are divided by transverse septa and these are also perforated. Certain of the genera as « 113 » Protopharetra (Fig. 67 D), Coscinocyathus, and Archaeocyathina, are common to the Cambrian of Sardinia and South Australia, whilst other genera of the class are also found in Siberia, China, Canada and the United States. A species of Protopharetra was recently detected in a pebble derived from the Cambrian limestone in the Antarctic, as far south as 85 deg. An Archaeocyathina limestone has also been found in situ from Shackleton’s farthest south.
CORALS (Class Anthozoa).
Rugose Corals.—
Many of the older types of Corals from the Palaeozoic rocks belong to the Tetracoralla (septa in multiples of four), or Rugosa (i.e., with wrinkled exterior).
Ordovician Corals.—
In Great Britain and North America Rugose Corals are found as early as Ordovician times, represented by Streptelasma, Petraia, etc. In Australia they seem to first make their appearance in the Silurian period.
Silurian Corals.—
In rocks of Silurian age in Australia we find genera like Cyathophyllum (with single cups or compound coralla), Diphyphyllum, Tryplasma and Rhizophyllum, the first-named often being very abundant. The compound corallum of Cyathophyllum approximans presents a very handsome appearance when cut transversely and polished. This coral is found in the Newer Silurian limestone in Victoria; it shows an alliance with C. mitchelli of the Middle « 114 » Devonian of the Murrumbidgee River, New South Wales (Fig. 69 A).
Silurian Hexacoralla.—
It is, however, to the next group, the Hexacoralla, with septa in multiples of six, twelve, and twenty-four, that we turn for the most varied and abundant types of Corals in Silurian times. The genus Favosites (Honey-comb Coral) is extremely abundant in Australian limestones (Fig. 69 B, C), such as those of Lilydale, Walhalla, and Waratah Bay in Victoria, and of Hatton’s Corner and other localities near Yass, in New South Wales. Pleurodictyum is also a familiar type in the Australian Silurian, being one of the commonest corals in the Yeringian stage; although, strange to say, in Germany and N. America, it is typical of Devonian strata (Fig. 69 E). Pleurodictyum had a curious habit of growing, barnacle fashion, on the side of the column of the crinoids or sea-lilies which flourished in those times. Syringopora, with its funnel-shaped tabulae or floor partitions, is typical of many Australian limestones, as those from Lilydale, Victoria, and the Delegate River, New South Wales. Halysites (Chain Coral), with its neat strings of tubular and tabulated corallites joined together by their edges, is another striking Coral of the Silurian period (Fig. 69 F). This and the earlier mentioned Syringopora, is by some authors regarded as belonging to the Alcyonarian Corals (typically with eight tentacles). Halysites is known from the limestones of the Mitta Mitta River, N.E. Gippsland, Victoria; from the Molong and Canobolas districts in New « 115 » South Wales; from the Gordon River limestone in Tasmania; and from Chillagoe in Queensland. Abroad it is a well known type of Coral in the Wenlockian of Gotland in Scandinavia, and Shropshire in England, as well as in the Niagara Limestone of the United States.
Silurian Octocoralla.—
Perhaps the most important of the Octocoralla is Heliolites (“Sunstone”), which is closely allied to the Blue Coral, Heliopora, a frequent constituent of our modern coral reefs. The genus Heliolites has a massive, calcareous corallum, bearing two kinds of pores or tubes, large (autopores) containing complete polyps, and small (siphonopores) containing the coenosarc or flesh of the colony. Both kinds of tubes are closely divided by tabulae, whilst the former are septate. Heliolites is of frequent occurrence in the Silurian limestones of New South Wales and Victoria (Fig. 69 G).
Devonian Corals.—
The Middle Devonian beds of Australia are chiefly limestones, such as the Buchan limestone, Victoria; the Burdekin Series, Queensland; and the Tamworth limestone of New South Wales. These rocks, as a rule, are very fossiliferous, and the chief constituent fossils are the Rugose and Perforate Corals. Campophyllum gregorii is a common form in the Buchan limestone (Fig. 70 A), as well as some large mushroom-shaped Favosites, as F. gothlandica and F. multitabulata. Other genera which may be mentioned as common to the Australian Middle Devonian rocks are, Cyathophyllum, Sanidophyllum and « 116 » Spongophyllum, Heliolites is also found in limestones of this age in New South Wales and Queensland.
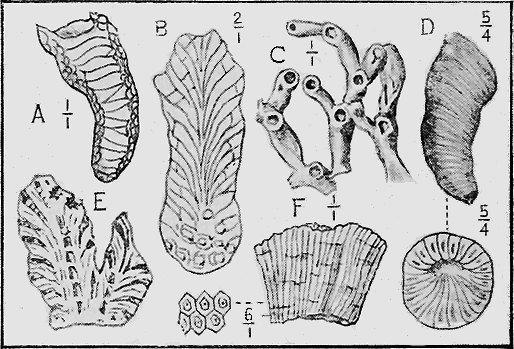
In the Burdekin Series (Middle Devonian) in Queensland we also find Cystiphyllum, Favosites gothlandica, and Pachypora meridionalis (Fig. 70 B), whilst in beds of the same age at Rough Range in Western Australia are found Aulopora repens (Fig. 70 C), and another species of Pachypora, namely, P. tumida.
Carbopermian Corals.—
The only true Carboniferous marine fauna occurring in Australia, appears to be that of the Star Beds in Queensland, but so far no corals have been found. « 117 » The so-called Carboniferous of Western Australia may be regarded as Carbopermian or even of Permian age. The marine Carbopermian beds of New South Wales contain several genera of Corals belonging to the group Rugosa, as Zaphrentis (Fig. 70 D), Lophophyllum, and Campophyllum. Of the Tabulate corals may be mentioned Trachypora wilkinsoni, very typical of the Upper Marine Series (Fig. 70 E) and Cladochonus.
In the Gympie beds of the same system in Queensland occur the following rugose corals, Zaphrentis profunda and a species of Cyathophyllum.
In the Carbopermian of Western Australia the rugose corals are represented by Amplexus, Cyathophyllum, and Plerophyllum, which occur in rocks on the Gascoyne River.
The imperfectly understood group of the Monticuliporoids, by some authors placed with the Polyzoa (Order Trepostomata), are well represented in Australia by the genus Stenopora (Fig. 70 F). The corallum is a massive colony of long tubes set side by side and turned outwards, the polyp moving upwards in growth and cutting off the lower part of the tube by platforms like those in the tabulate corals. Some of the species of Stenopora, like S. tasmaniensis, of New South Wales and Tasmania, are found alike in the Lower and Upper Marine Series. S. australis is confined to the Bowen River Coal-field of Queensland. Stenopora often attains a large size, the corallum reaching over a foot in length.
Neither Jurassic or Cretaceous Corals have been found in Australasia, although elsewhere as in « 118 » Europe and India, the representatives of modern corals are found in some abundance.
Cainozoic Corals.—
In Tertiary times the marine areas of southern Australia were the home of many typical solitary Corals of the group of the Hexacoralla. In the Balcombian beds of Mornington, Victoria, for instance, we have genera such as Flabellum, Placotrochus, Sphenotrochus, Ceratotrochus, Conosmilia, Trematotrochus, Notophyllia and Balanophyllia (Fig. 71).

Corals especially characteristic of the Janjukian Series are Paracyathus tasmanicus, Stephanotrochus tatei, Montlivaltia variformis, Thamnastraea sera and « 119 » Dendrophyllia epithecata. The stony axis of the Sea-pen, Graphularia senescens, a member of the Octocoralla, is also typical of this stage, and are called “square-bones” by the quarrymen at Waurn Ponds, near Geelong, where these fossils occur.
The Kalimnan Corals are not so abundantly represented as in the foregoing stages, but certain species of Flabellum and Trematotrochus, as F. curtum and T. clarkii, are peculiar to those beds. Several of the Janjukian Corals persist into Kalimnan times, some dating as far back as the Balcombian, as Sphenotrochus emarciatus. The Sea-pen, Graphularia senescens is again found at this higher horizon, at Beaumaris; it probably represents a varietal form, the axis being smaller and more slender.
Other examples of the Octocoralla are seen in Mopsea, two species of which are found in the Janjukian at Cape Otway; the deeper beds of the Mallee; and the Mount Gambier Series.
A species of the Astraeidae (Star-corals) of the reef-forming section, Plesiastraea st.vincenti, is found in the Kalimnan of Hallett’s Cove, South Australia.
HYDROZOA.
The few animals of this group met with in fossil faunas are represented by the living Millepora (abundant as a coral reef organism), Hydractinia (parasitic on shells, etc.), and Sertularia (Sea-firs).
Milleporids and Stylasterids.—
Although so abundant at the present time, the genus Millepora does not date back beyond the Pleistocene. The Eocene genus Axopora is supposed « 120 » to belong here, but is not Australian. Of the Stylasterids one example is seen in Deontopora, represented by the branchlets of D. mooraboolensis, from the Janjukian limestone of the Moorabool Valley, near Geelong.
Hydractinia.—
Hydractinia dates from the Upper Cretaceous rocks in England, and in Australia its encrusting polypidom is found attached to shells in the polyzoal limestone of Mount Gambier (Miocene).
Stromatoporoids.
An important group of reef-builders in Palaeozoic times was the organism known as Stromatopora, and its allies. The structures of these hydroid polyps resemble successional and repetitional stages of a form like Hydractinia. As in that genus it always commenced to grow upon a base of attachment such as a shell, increasing by successive layers, until the organic colony often reached an enormous size, and formed great mounds and reefs (see antea, Fig. 32). The stromatoporoid structure was formed by a layer of polyp cells separated by vertical partitions, upon which layer after layer was added until a great vertical thickness was attained. This limestone-making group first appeared in the Silurian, and probably reached its maximum development in Middle Devonian times, when it almost disappeared, except to be represented in Carbopermian strata by a few diminutive forms.
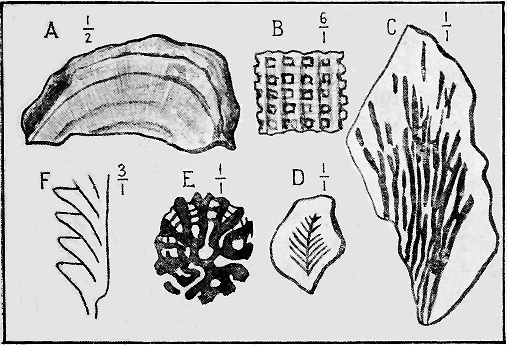
Silurian Stromatoporoids.—
In the Silurian limestones of Victoria (Lilydale, Waratah Bay, Walhalla and Loyola), and New South Wales (near Yass), Stromatoporoids belonging to the genera Clathrodictyon (probably C. regulare), Stromatopora and Idiostroma occur. Stromatoporella has been recorded from the Silurian rocks of the Jenolan Caves, New South Wales.
Devonian Stromatoporids.—
The Middle Devonian strata of Bindi, Victoria, yield large, massive examples of Actinostroma. This genus is distinguished from the closely allied Clathrodictyon by its vertical pillars passing through several laminae in succession. Rocks of the same « 122 » age in Queensland contain Stromatopora, whilst in Western Australia the Rough Range Limestone has been shown to contain Actinostroma clathratum (Fig. 72 A, B) and Stromatoporella eifeliensis.
Cladophora.
Palaeozoic Cladophora.—
Some branching and dendroid forms of Hydrozoa probably related to the modern Calyptoblastea (“covered buds”), such as Sertularia and Campanularia, are included in the Cladophora (“Branch bearers”). They existed from Cambrian to Devonian times, and consist of slender, forking branches sometimes connected by transverse processes or dissepiments, the branches bearing on one or both sides little cups or hydrothecae which evidently contained the polyps, and others of modified form, perhaps for the purpose of reproduction. The outer layer, called the periderm was of chitinous material. They were probably attached to the sea-floor like the Sertularians (Sea-firs).
Dictyonema and Allies.—
Remains of the above group are represented in the Australian rocks by several species of Dictyonema (Fig. 72 E, F) occurring in the Lower Ordovician of Lancefield, and in similar or older shales near Mansfield. Some of these species are of large size, D. grande measuring nearly a foot in width. The genera Callograptus, Ptilograptus (Fig. 72 C, D) and Dendrograptus are also sparsely represented in the Upper Ordovician of Victoria, the two former from San Remo, the latter from Bulla.
Graptolites (Graptolitoidea).—
Value of Graptolites to Stratigraphist.—
The Graptolites were so named by Linnaeus from their resemblances to writing on the slates in which their compressed remains are found. They form a very important group of Palaeozoic fossils in all parts of the world where these rocks occur, and are well represented in Australasia. The species of the various Graptolite genera are often restricted to particular beds, and hence they are of great value as indicators of certain horizons or layers in the black, grey or variously coloured slates and shales of Lower Ordovician to Silurian times. By their aid a stratum or set of strata can be traced across country for long distances, and the typical species can be correlated even with those in the older slates and shales of Great Britain and North America.
Nature of Graptolites.—
The Graptolites were compound animals, consisting of a number of polyps inserted in cups or thecae which budded out in a line from the primary sicula or conical chamber, which chamber was probably attached to floating sea-weed, either by a fine thread (nema), or a disc-like expansion. This budding of the polyp-bearing thecae gives to the polypary or colony the appearance of a fret-saw, with the teeth directed away from the sicula.
The habit of the earlier graptolites was to branch repeatedly, as in Clonograptus, or to show a compound leaf-like structure as in Phyllograptus. Later « 124 » on the many-branched forms had their branches reduced until, as in Didymograptus, there were only two branches. Sometimes the branches opened out to direct the thecae upwards, the better to procure their food supply. In Diplograptus the thecae turned upwards and acquired a support by the formation of a medium rod (virgula), often ending in a disc or float. In Silurian times Monograptus prevailed, a genus having only a single row of thecae supported by a straight or curved virgula. In Retiolites the polypary opened out by means of a net-work of fine strands, rendering it better able to float, at the same time retaining its original strength.
Lower Ordovician Graptolites, Victoria.—
The Lower Ordovician slates and shales of Victoria have been successfully divided into several distinct series by means of the Graptolites. These, commencing at the oldest, are:—
(1) Lancefield Series. Characterised by Bryograptus clarki, B. victoriae, Didymograptus pritchardi, D. taylori and Tetragraptus decipiens. Other forms less restricted are, Clonograptus magnificus (measuring over a yard in breadth), C. flexilis, C. rigidus, Leptograptus antiquus and Tetragraptus approximatus (Fig. 73).
(2) Bendigo Series. Characterised by Tetragraptus fruticosus, T. pendens, Trichograptus fergusoni and Goniograptus thureaui. This series also contains Tetragraptus serra (ranging into Darriwill Series), T. bryonoides, T. quadribrachiatus, T. approximatus (base of the series), Phyllograptus typus, Dichograptus octobrachiatus, Goniograptus macer and many Didymograpti, including D. bifidus (Fig. 74).
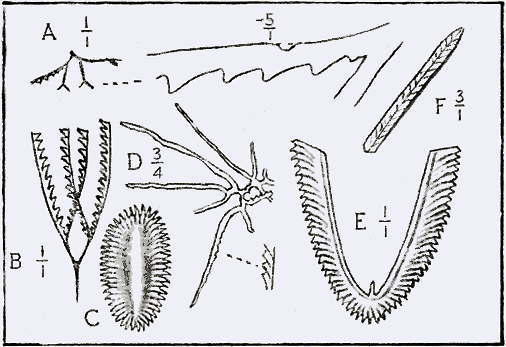
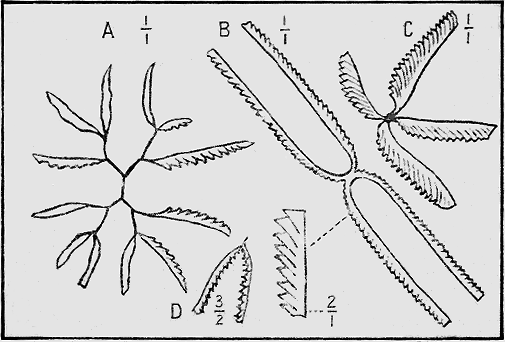
(3) Castlemaine Series. Characterised by Didymograptus bifidus, D. caduceus and Loganograptus logani. Phyllograptus persists from the Bendigo Series. It also contains Tetragraptus serra, T. bryonoides, T. quadribrachiatus, Goniograptus macer and several Didymograpti.
(4) Darriwill Series. Characterised by Trigonograptus wilkinsoni. Also contain Diplograptus, Glossograptus and Lasiograptus, whilst Didymograptus is rare.
Lower Ordovician Graptolites, New Zealand.—
In New Zealand Lower Ordovician Graptolites are found in the Kakanui Series, at Nelson, north-west of South Island. Some of the commoner forms are Didymograptus extensus, D. caduceus, Loganograptus logani, Phyllograptus typus, Tetragraptus similis and T. quadribrachiatus.
Graptolites agreeing closely with those of the Lancefield Series of Victoria occur near Preservation Inlet in the extreme South-west, and have been identified as Clonograptus rigidus, Bryograptus victoriae and Tetragraptus decipiens.
Upper Ordovician Graptolites, Victoria.—
The Upper Ordovician rocks of Victoria, as at Wombat Creek and Mount Wellington in Gippsland, and at Diggers’ Rest near Sunbury, contain the double branched forms like Dicranograptus ramosus, Dicellograptus elegans and D. sextans; the sigmoidal form Stephanograptus gracilis; and the diprionidian « 127 » (biserial) forms as Diplograptus tardus, Climacograptus bicornis, Cryptograptus tricornis, Glossograptus hermani and Lasiograptus margaritatus (Fig. 75).
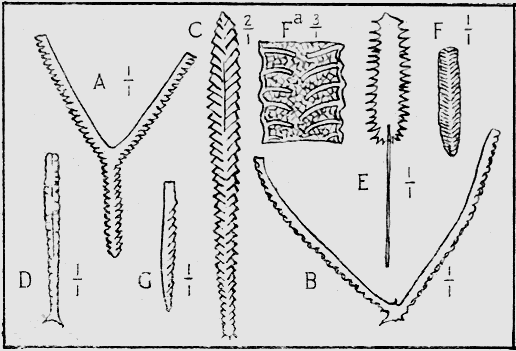
Upper Ordovician Graptolites, New South Wales.—
In New South Wales, at Tallong, the Upper Ordovician Graptolites are well represented by such forms as Dicellograptus elegans, Dicranograptus nicholsoni, Diplograptus carnei, D. foliaceus, Cryptograptus tricornis and Glossograptus quadrimucronatus, etc. Other localities in New South Wales for this Graptolite fauna are Stockyard Creek, Currowang, Tingaringi, Lawson, and Mandurama.
Tasmania.—
From Tasmania a Diplograptus has been recorded, but the particular horizon and locality are uncertain.
Silurian Graptolites, Victoria.—
In the Silurian shales at Keilor, in Victoria, Monograptus is a common genus, and Cyrtograptus and Retiolites australis (Fig. 75 F) also occur. Several species of Monograptus have also been found at South Yarra and Studley Park. At the latter place and Walhalla Monograptus dubius, which is a Wenlock and Ludlow fossil in Britain, has been found in some abundance (Fig. 75 G).
COMMON OR CHARACTERISTIC FOSSILS OF THE FOREGOING CHAPTER.
SPONGES.
Protospongia sp. Cambrian: S. Australia.
Hyalostelia sp. Cambrian: S. Australia.
Protospongia oblonga, Hall. L. Ordovician: Victoria.
Stephanella maccoyi, Hall. L. Ordovician: Victoria.
Carpospongia sp. Silurian: Yass, New South Wales.
Receptaculites fergusoni, Chapman. Silurian: Victoria.
Receptaculites australis, Salter sp. Devonian: Victoria and New South Wales. Carboniferous: Queensland.
(?) Lasiocladia hindei, Eth. fil. Carbopermian: Queensland.
Purisiphonia clarkei, Bowerbank. Lower Cretaceous: Queensland.
Geodia sp. Cainozoic: W. Australia.
Tethya sp. Cainozoic: W. Australia.
Ecionema newberyi, McCoy sp. Cainozoic: Victoria.
Plectroninia halli, Hinde. Cainozoic (Janjukian): Victoria.
Tretocalia pezica, Hinde. Cainozoic (Janjukian): Victoria.
ARCHAEOCYATHINAE.
Protopharetra scoulari, Etheridge, fil. Cambrian: S. Australia.
Coscinocyathus australis, Taylor. Cambrian: S. Australia.
Archaeocyathina ajax, Taylor. Cambrian: S. Australia.
CORALS.
Cyathophyllum approximans, Chapman. Silurian: Victoria.
Tryplasma liliiformis, Etheridge, fil. Silurian: New South Wales.
Favosites grandipora, Etheridge fil. Silurian: Victoria.
Pleurodictyum megastomum, Dun. Silurian: Victoria.
Halysites peristephicus, Etheridge, fil. Silurian: New South Wales.
Heliolites interstincta, Linné sp. Silurian: Victoria.
Campophyllum gregorii, Eth. fil. Middle Devonian: Victoria and Queensland.
Cystiphyllum australasicum, Eth. fil. Middle Devonian: New South Wales and Queensland.
Favosites multitabulata, Eth. fil. Middle Devonian: Victoria and New South Wales.
Pachypora meridionalis, Eth. fil. Middle Devonian: Queensland.
Zaphrentis culleni, Eth. fil. Carboniferous: New South Wales.
Lophophyllum corniculum, de Koninck. Carboniferous: New South Wales.
Zaphrentis profunda, Eth. fil. Carbopermian: Queensland.
Campophyllum columnare, Eth. fil. Carbopermian: New South Wales.
Trachypora wilkinsoni, Eth. fil. Carbopermian: New South Wales.
Stenopora tasmaniensis, Lonsdale. Carbopermian: Tasmania and New South Wales.
Flabellum gambierense, Duncan. Cainozoic: Victoria, S. Australia and Tasmania.
Placotrochus deltoideus, Duncan. Cainozoic: Victoria, S. Australia and Tasmania.
Sphenotrochus emarciatus, Duncan. Cainozoic: Victoria, S. Australia, and Tasmania.
Ceratotrochus exilis, Dennant. Cainozoic: Victoria.
Conosmilia elegans, Duncan. Cainozoic: Victoria.
Balanophyllia armata, Duncan. Cainozoic: Victoria.
Thamnastraea sera, Duncan. Cainozoic: Victoria and Tasmania.
Graphularia senescens, Tate sp. Cainozoic: Victoria and S. Australia.
HYDROZOA.
Clathrodictyon (?) regulare, Rosen sp. Silurian: Victoria.
Actinostroma clathratum, Nicholson. Devonian: W. Australia.
Stromatoporella eifeliensis, Nich. Devonian: W. Australia.
Dictyonema pulchella, T. S. Hall. Lower Ordovician: Victoria.
Ptilograptus sp. L. Ordovician: Victoria.
Callograptus sp. Lower Ordovician: Victoria.
GRAPTOLITES.
Bryograptus victoriae, T. S. Hall. Lower Ordovician (Lancefield Series): Victoria.
Tetragraptus fruticosus, J. Hall. L. Ordovician (Bendigo Series): Victoria.
Didymograptus caduceus, Salter. L. Ordovician (Castlemaine Series): Victoria. Also New Zealand.
Didymograptus bifidus, J. Hall. L. Ordovician (Castlemaine Series): Victoria. Also New Zealand.
Trigonograptus wilkinsoni, T. S. Hall. L. Ordovician (Darriwill Series): Victoria.
Dicranograptus ramosus, J. Hall sp. Upper Ordovician: Victoria.
Monograptus dubius, Suess. Silurian: Victoria.
Retiolites australis, McCoy. Silurian: Victoria.
LITERATURE.
SPONGES.
Cambrian.—Tate, R. Trans. R. Soc. S. Austr., vol. XV. (N.S.), 1892, p. 188.
Ordovician.—Hall, T. S. Proc. R. Soc. Vict., vol. I. pt. I. 1889, pp. 60, 61 (Protospongia). Idem, ibid., vol. XI. (N.S.), pt. II. 1899, pp. 152-155 (Protospongia and Stephanella).
Silurian to Carboniferous.—Salter, J. W. Canad. Org. Rem. Dec. I. 1859, p. 47. Etheridge, R. jnr. and Dun, W. S. Rec. Geol. Surv. New South Wales, vol. VI. 1898, pp. 62-75. Chapman, F. Proc. R. Soc. Vict. vol. XVIII. (N.S.), pt. 1, 1905, pp. 5-15.
Carbopermian.—Etheridge, R. jnr., in Geol. and Pal. Q., 1892, p. 199.
Cretaceous.—Bowerbank, J. S. Proc. Zool. Soc. Lond., 1869, p. 342. Etheridge, R. jnr. in Geol. and Pal. Queensland, 1892, pp. 438, 439 (Purisiphonia).
Cainozoic.—McCoy, F. Prod. Pal. Vict., Dec. V. 1877. Chapman, F. Proc. R. Soc. Vict., vol. XX. (N.S.), pt. 2, 1908, pp. 210-212 (Ecionema). Hinde, G. J. Quart. Journ. Geol. Soc., vol. LVI., 1900, pp. 50-56 (calcisponges). Idem, Bull. Geol. Surv. W. Austr., No. 36, 1910, pp. 7-21 (sponge-spicules).
ARCHAEOCYATHINAE.
Etheridge, R. jnr., Trans. R. Soc. S. Austr., vol. XIII. 1890, pp. 10-22. Taylor, T. G. Mem. Roy. Soc. S. Austr., vol. II., pt. 2, 1910 (a monograph).
CORALS.
Silurian.—Etheridge, R. jnr. Rec. Geol. Surv. New South Wales, vol. II. pt. 1, 1890, pp. 15-21 (Silurian and Devonian). Idem, ibid., vol. II. pt. 4, 1892, pp. 165-174 (Silurian and Devonian). Idem, in Pal. and Geol. Queensland, 1892. Idem, Rec. Austr. Mus., vol. I., No. 10, 1891, pp. 201-205 (Rhizophyllum). Id., ibid., vol. III. No. 2, 1897, pp. 30-33 (Columnaria). Id., Prog. Rep. Geol. Surv. Vict., No. 11, 1899, pp. 30-36. Idem, Mem. Geol. Surv. New South Wales, No. 13, pt. I., 1904 (Halysites). Id., ibid., No. 13, pt. 2, 1907 (Tryplasma). De Koninck, L. G. ibid., Pal. No. 6, 1898. Shearsby, A. J. Geol. Mag., Dec. V., vol. III. 1906, pp. 547-552. Chapman, F. Rec. Geol. Surv. Vict., vol. II. pt. 1, 1907, pp. 67-80.
Devonian.—Etheridge, R. jnr. and Foord, A. H. Ann. Mag. Nat. Hist., ser. V., vol. XIV., 1884, pp. 175-179 (Alveolites and Amplexopora = Litophyllum). Etheridge, R. jnr., in Geol. and Pal. Queensland, 1892. Idem, Proc. Linn. Soc. New South Wales, vol. IX. 1895, pp. 518-539. Id., Rec. Geol. Surv. New South Wales, vol. VI. pt. 3, 1899, pp. 152-182 (Tamworth District). Id., Rec. Austr. Mus., vol. IV. No. 7, 1902, pp. 253-260. De Koninck, L. G. Mem. Geol. Surv. New South Wales, Pal. No. 6, 1898. Chapman, F. Rec. Geol. Surv. Vict., vol. III, pt. 2, 1912, pp. 215-222.
Carbopermian.—Etheridge, R. jnr. Mem. Geol. Surv. New South Wales, Pal. No. 5, 1891. Idem, in Geol. and Pal. Queensland, 1892. Id., Bull. Geol. Surv., W. Austr., No. 10, 1903, pp. 8-10.
Cainozoic.—Duncan, P. M. Quart. Journ. Geol. Soc., vol. XXVI. 1870, pp. 284-318; vol. XXXI. 1875, pp. 673-678; vol. XXXII. 1876, pp. 341-351. Woods, T. Proc. Linn. Soc. New South Wales, vol. XI., 1878, pp. 183-195; ibid., vol. XXX. 1879, pp. 57-61. Idem, Trans. Roy. Soc. S. Austr., vol. I., 1878, pp. 104-119. Dennant, J. Trans. R. Soc. S. Austr., vols. XXIII. (1899) to XXVIII. (1904).
STROMATOPOROIDS.
Hinde, G. J. Geol. Mag., Dec. III. vol. VII, 1890, p. 193.
GRAPTOLITES.
McCoy, F. Prod. Pal. Vict., Decades I. (1874): II. (1875): V. (1877). Hall, T. S. Proc. Roy. Soc. Vict., vol. IV. p. I. 1892, pp. 7, 8 (Dictyonema). Idem, Geol. Mag. Dec. IV. vol. VI. 1899, pp. 438-451; Id., Rep. Austr. Assoc. Adv. Sci., Brisbane, 1909, pp. 318-320. Id., Rec. Geol. Surv. Vict., vol. I. pt. 4, 1906, pp. 266-278. Id., ibid., vol. III. pt. 2, 1912, pp. 188-211. Idem, Rec. Geol. Surv. New South Wales, vol. VII. part 1, 1910, pp. 16, 17. Ibid., pp. 49-59.
FOSSIL SEA-LILIES, STARFISHES, BRITTLE-STARS AND SEA-URCHINS.
Divisions of Echinodermata.—
The sub-kingdom of ECHINODERMATA includes the above groups comprised in the Classes Crinoidea, Asteroidea, Ophiuroidea and Echinoidea. Besides these are the less important classes of the Cystidea or sac-shaped echinoderms (of which no definite remains are recorded from Australian rocks); the Blastoidea or bud-shaped echinoderms (of which four genera are known from Australia); the Edrioasteroidea or sessile starfishes (unknown in Australia); and the Holothuroidea or sea-cucumbers (represented as fossils by the skin spicules and plates, an example of which has been recorded from Australia).
CRINOIDEA, or Sea-lilies.
Crinoidea, their General Structure.—
These often beautiful and graceful animals resemble a starfish mounted on a stalk. They are composed of calcareous joints and plates, and are therefore important as rock-formers. The stalk or column may be either short or long, and is generally rooted, in the adult stage, in the mud of the sea-floor. Fossil Crinoids were sometimes furnished with « 134 » a coiled termination, which could be entwined around such objects as the stems of sea-weeds. The crinoid column is composed of numerous plates, and is round or pentagonal. Upon this is fixed the calyx or cup, with its attached arms, which serve to bring food to the mouth, situated on the upper part of the cup. The arms are grooved, and the water, being charged with food particles (animalcula), pours down these channels into the mouth. The stem elevates the animal above the mud or silt of the sea-floor, thus making it more easy for it to obtain its food supply. The stalks of fossil Crinoids sometimes reached the enormous length of 50 feet. Their calcareous skeleton is built upon a plan having five planes of symmetry; this pentamerism is found throughout the crinoids, the blastoids and the free-moving echinoderma. Crinoids range from moderately shallow- to deep-water, and at the present day are almost restricted to abyssal conditions. The more ancient types usually found their habitats amongst reefs or in comparatively clear water, where there was a marked freedom from sediment, although that was not an essential, as seen by their numerous remains in the Australian mudstones and sandstones.
Cambrian Crinoids.—
The group of the Crinoidea first appears in the Upper Cambrian, and persists to the present time. In North America the genus Dendrocrinus occurs in the Cambrian and Ordovician; and some stem-joints from the Upper Cambrian limestone of the Mount Wellington district, Victoria, may be provisionally referred to this genus.
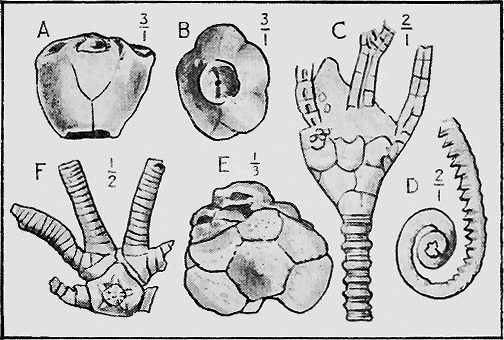
Ordovician Crinoids.—
No undoubted Crinoid remains have been found in the Australian Ordovician; although many genera are found elsewhere in that system, chiefly in N. America, as Reteocrinus, Hybocrinus, Heterocrinus and Dendrocrinus, and in Europe and North America, as Rhodocrinus and Taxocrinus.
Silurian Crinoids.—
The Silurian Crinoidea of Australia are largely represented by the remains of the columns or stalks, which are often found in such abundance as to constitute large masses of sub-crystalline limestone, as that of Toongabbie, Victoria. The columns of the Crinoids do not usually possess sufficient characters « 136 » to enable the forms to be identified. There are, however, more perfect and identifiable remains of several very interesting generic types in the Silurian faunas as follows:—
In New South Wales Pisocrinus is represented with some reservation by (?) P. yassensis, found at Limestone Creek, near Yass (Fig. 76 A, B).
In Victoria, Helicocrinus plumosus and Botryocrinus longibrachiatus occur at Brunswick and Flemington, respectively (Fig. 76). The former is a delicate and handsome species, having a small cup with finely pinnate arms, which are forked once, and with a pentagonal stem coiled at the distal end (see Frontispiece). The genus Botryocrinus is found in rocks of a similar age in North America and England. Hapalocrinus victoriae, a member of the Platycrinidae, has been described from the mudstone of South Yarra, near Melbourne. The species above mentioned are of Melbournian age, belonging to the lower stage of the Silurian system.
Devonian Crinoids.—
In the Middle Devonian of Queensland, fragmentary crinoid stems are found interbedded with the limestone of the Broken River.
Thin slices of the limestone of the same age from Buchan, Victoria, show numerous ossicles and stem-joints of Crinoids.
Similar remains have also been recorded from the Devonian of the Kimberley district and the Gascoyne River in Western Australia.
Carboniferous Crinoids.—
The Carboniferous (Star Beds) of Queensland has yielded remains of Actinocrinus.
The Matai Series of New Zealand, which may be regarded as almost certainly of Carboniferous age, contains remains of a Cyathocrinus, found in the limestone of the Wairoa Gorge.
Carbopermian Crinoids.—
The Carbopermian (Upper Marine Series) of New South Wales yields the interesting Crinoid having a large, globular cup, known as Phialocrinus; the best known species of this genus are P. konincki (Fig. 76 E) and P. princeps. Beds of the same age in New South Wales, also in the Upper Marine Series, contain the aberrant Crinoid with strongly sculptured plates of the calyx in the decorticated condition, Tribrachiocrinus clarkei.
Poteriocrinus and Platycrinus are, with some reservation, recorded from the Gympie Series at Stanwell and the marine beds of the Bowen River Coal-field respectively, both in Queensland.
In Western Australia the Carbopermian rocks of the Gascoyne River are known to contain crinoid stems, tentatively referred to either the Rhodocrinidae or the Actinocrinidae. There is also a species of Platycrinus known from the Gascoyne and Irwin Rivers, and from the Kimberley District.
Triassic Crinoids.—
The Kaihiku Series of Nelson, New Zealand, has yielded some crinoid stems, but the genus has not yet been determined.
Cretaceous Crinoids.—
In the Lower Cretaceous Limestone of Queensland, at Mitchell Downs and Wollumbilla, a typical Crinoid, closely allied to the living Pentacrinus is found, namely, Isocrinus australis (Fig. 76 F).
The Upper Cretaceous opal deposits of White Cliffs in Wilcannia, New South Wales, contain many opalised fossil remains, amongst them being Isocrinus australis, already noticed as occurring in the Lower Cretaceous of Queensland.
Cainozoic Crinoids.—
Pentacrinus stellatus is a species founded on some deeply indented pentagonal stem-joints found in the Oamaru Series (Miocene) at Curiosity Shop, South Canterbury, New Zealand, and also occurring in the Chatham Islands. This species has been identified in the Aire Coastal beds in Victoria, of the same age. Another generic type, Antedon, the beautiful “Feather Star,” is frequently met with in Janjukian strata in Victoria and South Australia, as at Batesford and Mount Gambier, represented by the denuded crown and the ossicles of the arms of a comparatively large species; whilst another and smaller form has been described from beds of the same age from borings in the Victorian Mallee, under the name of A. protomacronema.
BLASTOIDEA—Bud-shaped Echinoderms.
Distribution and Characters of Blastoidea.—
This forms a small class which has a few representatives in the rocks of Australia. Elsewhere they are chiefly of Devonian and Carboniferous ages. In Australia they are confined, so far as known, to sediments of the Carboniferous System. The animal was rooted to the sea-floor and a jointed stem was usually present. The cup or theca, as before noted, is bud-shaped, and consists of basal, radial and deltoid plates, the edges of which are folded inwards into « 139 » the thecal cavity, and thus the internal organs came into contact with the incurrent water. The cup bears five food grooves, bordered by numerous arms or brachioles, which directed the incurrent particles into the thecal cavity.
Carbopermian Blastoids.—
Three genera of blastoids have been recorded from the Gympie Beds, or Carbopermian, of the Rockhampton District of Queensland. They are, Mesoblastus, Granatocrinus and Tricoelocrinus. A similar fossil in beds of like age, and provisionally referred to the genus Metablastus, has been lately recorded from Glenwilliam, Clarence Town, New South Wales.
ASTEROIDEA, or Starfishes.
Characters of True Starfishes.—
These free-moving echinoderms are usually five-sided, though sometimes star-shaped, with numerous arms surrounding a central disc. The mouth is central on the under side of the disc, and the anus above and near the centre (excentric), the latter being covered by a porous plate called the madreporite. The hydraulic system of starfishes consists of tubes extending along the grooved arms and giving off side branches which end in processes called podia and terminating in suckers. The podia pass through pores in the floor plates of the grooves, and communicate within the body with distensions called ampulla. By this means the podia serve as feet, and can be withdrawn by the expulsion of the water in them into the ampulla. The stout flexible covering of the starfish is strengthened by calcareous plates and bars, « 140 » owing to the presence of which they are often preserved as fossils.
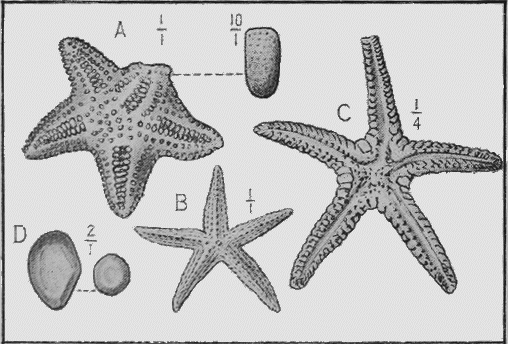
Silurian Starfishes.—
The oldest Australian fossil Starfishes are found in the Silurian. In Victoria they occur in some abundance in the lower, Melbournian, series, but appear to be absent or at all events very scarce in the upper, or Yeringian series. The commonest genus is Palaeaster, of which there are two species, P. smythi (Fig. 77 A) and P. meridionalis, found alike in the sandy and argillaceous strata near Melbourne. Urasterella is another genus found in the Silurian rocks near Melbourne, in which the marginal series of plates seen in Palaeaster are wanting, giving to the starfish a slender, long-armed aspect (Fig. 77 B).
Carbopermian Starfishes.—
In the Lower Marine Series of the Carbopermian of New South Wales a very large species of Palaeaster occurs (P. giganteus), measuring 7 inches from point to point across the disc (Fig. 77 C). Two other species of the same genus occur in this series (P. stutchburii and P. clarkei) the latter also ranging into the Upper Marine Series.
Cainozoic Starfishes.—
No remains of true Starfishes have been recorded from Australia between the Carbopermian and the Tertiary systems. In the Janjukian Series of Victoria the marginal plates of a species of Pentagonaster are typical fossils. They have been recorded from Waurn Ponds, Spring Creek near Torquay, and Batesford (Fig. 77 D). In the Mallee Bores, both marginal and abactinal plates of this genus are found in polyzoal limestone (Miocene). Pentagonaster also occurs in the Lower Muddy Creek beds (Oligocene), and the Upper beds of the same locality (Lower Pliocene). A species of Astropecten has been described from the Waikari River, New Zealand (Oamaru Series).
OPHIUROIDEA, or Brittle-stars.
Characters of Brittle-Stars.—
The Brittle-stars are frequently found at the present day cast up on the fine sandy beaches of the coast. They are easily distinguished from true starfishes by having a definite central disc, to which the arms are attached. The arms are used for locomotion and prehension, and have their grooves covered « 142 » over with plates. The ossicles of the arms are moveable and controlled by muscles which enable them to be used as feet. The lower surface of the disc has a central arrangement of five rhomboidal sets of jaws, formed of modified ossicles, called the mouth frame, whilst the upper surface bears, between one set of arms, the madreporite or covering plate to the water vascular system, as in starfishes.

Silurian Brittle-Stars.—
The Brittle-stars in Australia first appear in the Silurian, but in England and Bohemia date back to the Ordovician. Protaster is the commonest genus, and is represented by P. brisingoides of the Melbournian stage of Silurian strata at Flemington (Fig. 78). It also occurs rarely in the Yeringian beds at Yering, both Victorian localities. A very ornamental form, Gregoriura spryi, occurs in the same « 143 » division of the Silurian at South Yarra. In this fossil the delicate spines attached to the adambulacral ossicles are well preserved and form a marginal fringe to the arm (Fig. 79). Sturtzura is another Silurian genus, found in the Wenlock of England and in the Melbournian of Flemington, Victoria.
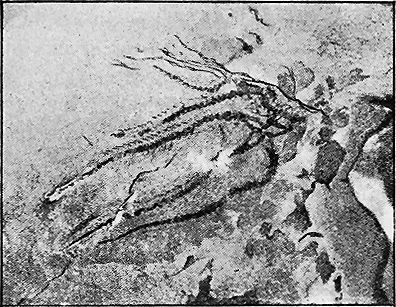
Cainozoic Brittle-Stars.—
From the Victorian Cainozoic beds, in the Lower Pliocene of Grange Burn, Hamilton, a vertebral ossicle of an ophiurian has been obtained, which has been provisionally referred to the genus Sigsbeia.
ECHINOIDEA, or Sea-urchins.
This group is an important one amongst Australian fossils, especially those of Cainozoic age.
Characters of Sea-urchins.—
Echinoids are animals enclosed in a spheroidal box or test composed of numerous calcareous plates, disposed geometrically as in the Starfishes, along five principal lines. The test in the living condition is more or less densely covered with spines. The mouth is on the under surface. The anus is either on the top of the test (dorso-central), or somewhere in the median line between the two lower ambulacra. The ambulacra (“a garden path”) are the rows of perforated plates on the upper (abactinal) surface sometimes extending to the lower surface, through which protrude the podia, which in Starfishes are situated in grooves on the lower surface.
Silurian Palaeechinoids.—
The Palaeechinoids are represented in the Silurian of Australia by occasional plates, as at Bowning, New South Wales, and near Kilmore, Victoria, whilst spines are not uncommon in certain Silurian limestones at Tyer’s River, Gippsland.
Carbopermian Palaeechinoids.—
In the Carbopermian of New South Wales, tests of Archaeocidaris have been recorded, and also a plate of the same genus in the Gympie Beds of Rockhampton, Queensland.
Regular Echinoids.—
The regular Echinoids date from Permian times. They have two vertical rows of plates for each ambulacrum and inter-ambulacrum. The mouth is on the underside, and the anus abactinal (on the upper side) and near the centre.
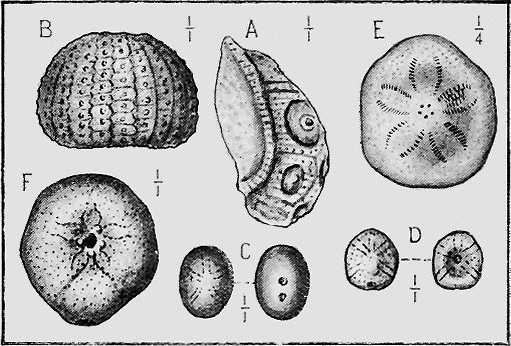
Cainozoic Regular Echinoids.—
In Australasia they make their first appearance in strata of Tertiary age, and some species, as Paradoxechinus novus, range through Balcombian strata to Kalimnan in Victoria, or Oligocene to Lower Pliocene, but are more typically Janjukian. Echinus (Psammechinus) woodsi (Fig. 80 B) is common in Janjukian strata in Victoria and South Australia and occurs sparingly in the Kalimnan. Another common form of the regular Echinoids in Southern Australia is Cidaris australiae (Fig. 80 A), ranging from Janjukian to Kalimnan, occurring more frequently in the older series. In New Zealand a species of Cidaris (C. striata), is known from the « 146 » Oamaru Series at Brighton. An Echinus occurs in the Oamaru Series of Broken River, and two species of that genus in the Wanganui formation of Shakespeare Cliff. Temnechinus macleayana has been recorded from the Cainozoic (Miocene or Pliocene) of Yule Island, Papua.
Irregular Echinoids.—
The irregular Echinoids are not known before the Upper Cretaceous in Australia, and are very common in the Tertiaries. They are distinguished by the anus (periproct) passing backward from the apex, as compared with the regular forms, and by the elongation of the test and the loss of the strong solid spines, which are replaced by thin, slender hair-like spines. The animal is thus better fitted to burrow through the ooze on which it feeds.
Cretaceous Irregular Echinoids.—
An interesting form, Micraster sweeti, is found in the Upper Cretaceous or Desert Sandstone of Maryborough in Queensland, which reminds one of typical European species of this genus.
Cainozoic Irregular Echinoids.—
Amongst the Australian Cainozoic Echinoids of the irregular type the following may be mentioned. The little subglobular test of Fibularia gregata, and Echinocyamus (Scutellina) patella (Fig. 80 C, D) are Janjukian in age. The large Clypeaster, C. gippslandicus (Fig. 80 E), ranges from the Oligocene to Lower Pliocene in Victoria (Balcombian to Kalimnan), and vies in size, especially in the Janjukian, with some large species like those from Malta and Egypt. This genus includes some of the largest known sea-urchins. The biscuit urchin, Arachnoides (Monostychia) « 147 » australis, is commonest in the Janjukian, but ranges from Balcombian to Kalimnan. A common urchin from the polyzoal rock of Mt. Gambier is Echinolampas gambierensis, which is also found in the Lower beds of Muddy Creek. A typical Janjukian fossil is Duncaniaster australiae, formerly thought to belong to the Cretaceous genus Holaster. Although found living, the genus Linthia attained its maximum development both in size and abundance, in Janjukian or Miocene times, as seen in L. gigas (having a length of 71/2 inches) and L. mooraboolensis. Echinoneus dennanti is restricted to the Janjukian. Several species of Eupatagus occur in the Cainozoic or Tertiary beds of South Australia, Victoria and New Zealand; Lovenia forbesi (Fig. 81 C) is common in the Janjukian to Kalimnan, both in Victoria and South Australia. In the latter State also occur the following genera:—Studeria, Cassidulus, Echinolampas, Plesiolampas, Linthia, Schizaster and Brissopsis. In New Zealand the following Cainozoic genera, amongst others of the irregular sea-urchins, may be cited:—Hemipatagus, Brissopsis, Hemiaster, and Schizaster (Fig. 81).

A clypeastroid, Peronella decagonalis has been described from the (?) Lower Pliocene of Papua.
Cainozoic Holothuroidea.—
The HOLOTHUROIDEA (Sea-Cucumbers) are represented in Australian deposits by a unique example of a dermal spicule of wheel-like form, referred to Chiridota, obtained from the Cainozoic (Janjukian) beds of Torquay. This genus is also known from the “calcaire grossier” or Middle Eocene of the Paris Basin, and is found living in all parts of the world.
COMMON OR CHARACTERISTIC FOSSILS OF THE FOREGOING CHAPTER.
CRINOIDS.
(?) Pisocrinus yassensis, Eth. fil. Silurian: New South Wales.
Helicocrinus plumosus, Chapman. Silurian: Victoria.
Botryocrinus longibrachiatus, Chapm. Silurian: Victoria.
Hapalocrinus victoriae, Bather. Silurian: Victoria.
Actinocrinus sp. Carboniferous: Queensland.
Cyathocrinus sp. Carboniferous: New Zealand.
Phialocrinus konincki, Clarke sp. Carbopermian: New South Wales.
Phialocrinus princeps, Eth. fil. Carbopermian: New South Wales.
Tribrachiocrinus clarkei, McCoy. Carbopermian: New South Wales.
(?) Platycrinus sp. Carbopermian: Queensland.
Platycrinus sp. Carbopermian: W. Australia.
Isocrinus australis, Moore sp. Cretaceous: Queensland.
Pentacrinus stellatus, Hutton. Miocene: New Zealand, Chatham Ids. and Victoria.
Antedon protomacronema, Chapman. Miocene: Victoria (deep borings).
BLASTOIDS.
(?) Mesoblastus australis, Eth. fil. Carbopermian: Queensland.
STARFISHES.
Palaeaster smythi, McCoy. Silurian: Victoria.
Palaeaster meridionalis, Eth. fil. Silurian: Victoria.
Urasterella selwyni, McCoy. Silurian: Victoria.
Palaeaster giganteus, Eth. fil. Carbopermian (L. Mar. Ser.): New South Wales.
Palaeaster clarkei, de Koninck. Carbopermian (L. and Up. Mar. Ser.): New South Wales.
Pentagonaster sp. Miocene: Victoria.
Astropecten sp. Miocene: New Zealand.
BRITTLE-STARS.
Protaster brisingoides, Gregory. Silurian: Victoria.
Gregoriura spryi, Chapman. Silurian: Victoria.
Sturtzura leptosomoides, Chapman. Silurian: Victoria.
(?) Sigsbeia sp. Lower Pliocene: Victoria.
ECHINOIDS.
Palaeechinus sp. Silurian: Victoria.
(?) Archaeocidaris selwyni, Eth. fil. Carbopermian: New South Wales.
Micraster sweeti, Eth. fil. Cretaceous: Queensland.
Cidaris (Leiocidaris) australiae, Duncan. Miocene and Lower Pliocene: Victoria and S. Australia.
Cidaris striata, Hutton. Miocene: New Zealand.
Echinus (Psammechinus) woodsi, Laube sp. Miocene and L. Pliocene: Victoria and S. Australia.
Temnechinus macleayana, T. Woods. Cainozoic (? Lower Pliocene): Papua.
Fibularia gregata, Tate. Miocene: Victoria and S. Australia.
Echinocyamus (Scutellina) patella, Tate sp. Oligocene to Miocene: Victoria and S. Australia.
Clypeaster gippslandicus, McCoy. Oligocene to L. Pliocene: Victoria.
Arachnoides (Monostychia) australis, Laube sp. Oligocene to L. Pliocene: Victoria and S. Australia.
Echinoneus dennanti, Hall. Miocene: Victoria.
Duncaniaster australiae, Duncan sp. Miocene: Victoria.
Lovenia forbesi, T. Woods sp. Miocene and L. Pliocene: Victoria and S. Australia.
Hemiaster planedeclivis, Gregory. Miocene: Victoria.
HOLOTHURIAN.
Chiridota sp. Miocene: Victoria.
LITERATURE.
CRINOIDS.
Silurian.—Etheridge, R. jnr. Rec. Austr. Mus., vol. V. No. 5, 1904, pp. 287-292 (Pisocrinus). Bather, F. A. Geol. Mag., Dec. XV. vol. IV. 1897, pp. 337-345 (Hapalocrinus). Chapman, F. Proc. R. Soc. Vict., vol. XV. (N.S.), pt. II. 1903, pp. 107-109 (Helicocrinus and Botryocrinus). Bather, F. A. Ottawa Nat., vol. XX. No. 5, 1906, pp. 97, 98.
Carboniferous and Carbopermian.—De Koninck, L. G. Mem. Geol. Surv. New South Wales, Pal. No. 6, 1898, pp. 121-126. Etheridge, R. jnr., in Geol. and Pal. Queensland, 1892, pp. 207-219. Idem, Mem. Geol. Surv. New South Wales, Pal. No. 5, 1892, pp. 75-119.
Cretaceous.—Moore, C. Quart. Journ. Geol. Soc., vol. XXVI. 1870, p. 243. Etheridge, R. jnr., in Geol. and Pal. Queensland, 1892, p. 439 (Isocrinus).
Cainozoic.—Hutton, F. W. Cat. Tert. Moll. and Ech. of New Zealand, 1873, p. 38.
BLASTOIDS.
Carbopermian.—Etheridge, R. jnr., in Geol. and Pal. Queensland, 1892, pp. 210-213. Taylor, T. G. Proc. Linn. Soc. New South Wales, 1908, pp. 54-59 (? Metablastus).
STARFISHES.
Silurian.—McCoy, F. Prod. Pal. Vict., Dec. I., 1874, pp. 41-43. Etheridge, R. jnr. Rec. Austr. Mus., vol. I., No. 10, 1891, pp. 199, 200.
Carboniferous and Carbopermian.—Etheridge, R. jnr. Mem. Geol. Surv. New South Wales, Pal. No. 5, pt. 2, 1892, pp. 70-75. De Koninck, L. G. Ibid., Pal. No. 6, 1898, p. 127.
Cainozoic.—Hall, T. S. Proc. R. Soc., Vict., vol. XV. (N.S.), pt. I. 1902, pp. 81, 82 (Pentagonaster). Hutton, F. W. Cat. Tert. Moll, and Ech. New Zealand, 1873, p. 38.
BRITTLE-STARS.
Silurian.—Gregory, J. W. Geol. Mag., Dec. III. vol. VI. 1889, pp. 24-27. Chapman, F. Proc. R. Soc. Vict., vol. XIX. (N.S.), pt. II. 1907, pp. 21-27.
Cainozoic.—Hall, T. S. Proc. R. Soc. Vict., vol. XV. (N.S.), pt. I. 1902, p. 82 (cf. Sigsbeia).
ECHINOIDS.
Silurian.—Chapman, F. Rec. Geol. Surv. Vict., vol. II. pt. 1, 1907, pp. 77, 78.
Carbopermian.—Etheridge, R. jnr. Mem. Geol. Surv. New South Wales, Pal. No. 5, pt. 2, 1892, pp. 67-69.
Cretaceous.—Etheridge, R. jnr., in Geol. and Pal. Queensland, 1892, pp. 559, 560.
Cainozoic.—T. Woods. Trans. Adelaide Phil. Soc., 1867. Laube, G. C. Sitz, k. k. Ak. Wiss. Wien, vol. LIX. 1869, pp. 183-198. Hutton, F. W. Cat. Tert. Moll, and Ech. New Zealand, 1873, pp. 38-43. Duncan, P. M. Quart. Journ. Geol. Soc., vol. XXXIII. 1877, pp. 42-73. Tate, R. Quart. Journ. Geol. Soc., vol. XXXIII. 1877, pp. 256-258. Idem, Southern Science Record, 1885, p. 4. Idem, Trans. R. Soc. S. Austr., vol. XIV. pt. 2, 1891, pp. 270-282. McCoy, F. Prod. Pal. Vict., Dec. VI. VII. 1879, 1883. Gregory, J. W. Geol. Mag., Dec. III. vol. VII. 1890, pp. 481-492. Ibid., Dec. III. vol. IX. 1892, pp. 433-437. Cotteau, G. H. Mem. Zool. France, vol. II. No. 4, 1889, p. 228; vol. III. No. 5, 1890, pp. 537-550; vol. IV. No. 5, 1891, pp. 620-633. Bittner, A. Sitz. k.k. Ak. Wiss. Wien, 1892, vol. 101, pp. 331-371. Hall, T. S. Proc. Roy. Soc. Vic., vol. XIX. (N.S.), pt. II. 1906, pp. 48, 53. Chapman, F. Proc. Roy. Soc. Vict., vol. XX. (N.S.), pt. II. 1908, pp. 214-218. Pritchard, G. B. ibid., vol. XXI. (N.S.), pt. I. 1908, pp. 392-400.
HOLOTHURIAN.
Cainozoic.—Hall, T. S. Proc, R. Soc. Vict., vol. X. (N.S.), pt. I. 1902, pp. 82, 83.
FOSSIL WORMS, SEA-MATS and LAMP-SHELLS.
The first-named group, the ringed worms, belong to the phylum Annelida, so-called because of the ring-like structure of their bodies. The two remaining groups, the Polyzoa or Sea-mats and the Brachiopods or Lamp-shells, are comprised in the phylum Molluscoidea, or mollusc-like animals.
WORMS (Annelida).
Annelida and their Fossil Representatives.—
These animals, owing to the scarcity of hard parts within their bodies, play a rather insignificant role as a fossil group. Worms are laterally symmetrical animals, with a dorsal and a ventral surface. They are segmented, the body being formed of numerous rings. Only those of the Class Chaetopoda (“bristle-feet”) are represented by identifiable fossil remains. Fossil worms, moreover, chiefly belong to the Order Polychaeta (“many bristles”). The horny jaws of these worms are sometimes found in the older rocks and are known as conodonts.
Silurian Conodonts.—
Conodonts belonging to three genera are known from Australia. They are all from the Silurian of the Bowning District, near Yass, New South Wales, and are referred to the genera Eunicites, Oenonites and Arabellites.
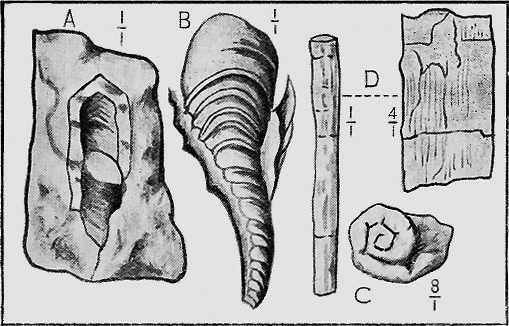
Palaeozoic Errant Worms.—
The wandering Worms (Polychaeta errantia) are also recognised by their impressions, trails, borings and castings. Burrows formed by these worms are seen in Arenicolites, found in the Silurian sandstone of New South Wales, near Yass, and in the Carbopermian (Gympie Series) near Rockhampton, Queensland. The membranous-lined burrows of Trachyderma (T. crassituba), occur in some abundance in the Silurian mudstones in the neighbourhood of Melbourne, « 154 » Victoria (Fig. 82 A). The genus Trachyderma is common also to Great Britain and Burmah, in beds of the same age.
Worm Tracks.—
Some of the curious markings on the Carboniferous sandstone of Mansfield, Victoria, may be due to worm trails and castings, especially since they are associated with sun-cracks and ripple-marks.
Sedentary Worms.—
The sedentary or tube-making Worms (Polychaeta tubicola) are represented by numerous forms. The long conical tube of Cornulites tasmanicus is recorded from the Silurian of Zeehan, Tasmania (Fig. 82 B). Spirorbis occurs in the Middle Devonian of Victoria (Fig. 82 C), and W. Australia, and also in the Carbopermian of W. Australia. Torlessia is found in the Trias or Lower Jurassic of the province of Canterbury, New Zealand (Fig. 82 D). The genus Serpula is widely distributed, occurring in the Carbopermian (Upper Jurassic Series), near East Maitland, New South Wales (S. testatrix), in the Jurassic of W. Australia (S. conformis), in the Lower Cretaceous of Wollumbilla, Queensland (S. intestinalis), and the Darling River, north west of New South Wales, (S. subtrachinus), as well as in Cainozoic deposits in Victoria (S. ouyenensis). Ditrupa is very abundant in some shelly deposits of Janjukian age in Victoria.
MOLLUSCOIDEA.
The Sea-mats (Polyzoa) and the Lamp-shells (Brachiopoda) constitute a natural group, the MOLLUSCOIDEA, which, although unlike in outward « 155 » form, have several physiological structures in common. The respiratory organs lie in front of the mouth, and are in the form of fleshy tentacles or spiral appendages. These animals are more nearly allied to the worms than to the molluscs.
POLYZOA.
Characters of Polyzoa.—
These are almost exclusively marine forms, and are important as fossils. They form colonies (polypary or zoarium), and by their branching, foliaceous or tufty growth resemble sea-weeds. The cells in which the separate zoöids lived have peculiar characters of their own, which serve to distinguish the different genera.
Subdivisions of Polyzoa.—
Polyzoa are divided into the Sub-classes Phylactolaemata, in which the mouth of the zoöid has a lip, and the series of tentacles is horse-shoe shaped; and the Gymnolaemata, in which there is no lip to the mouth, and the tentacles form a complete circle. The first group forms its polypary of soft or horny material, which is not preserved fossil. The latter has a calcareous polypary, and is of much importance as a fossil group. This latter sub-class is further subdivided into the following Orders, viz.:—Trepostomata (“turned mouths”), Cryptostomata (“hidden mouths”), Cyclostomata (“round mouths”), and Cheilostomata (“lip mouths” furnished with a moveable operculum).
Trepostomata (Palaeozoic).—
The Order Trepostomata may include some genera as Monticulipora and Fistulipora, previously referred « 156 » to under the corals. They become extinct after Permian times. Fistulipora occurs in certain Gippsland limestones.

Cryptostomata (Palaeozoic).—
In the order Cryptostomata we have the genus Rhombopora with its long, slender branches, which occurs in the Silurian of Victoria and the Carbopermian of Queensland and W. Australia (Fig. 83 C). Of this order a very important Australian genus is Fenestella, the funnel-shaped zoaria of which are found in the Silurian of Victoria and New South Wales, and also in the Carboniferous of the latter State. Fenestella also occurs in the Carbopermian of « 157 » W. Australia and Tasmania (Fig. 83 A). Accompanying the remains of Fenestella in the Carbopermian rocks, and closely related to it, are found the genera Protoretepora and Polypora (Fig. 83 B, D).
Polyzoa have been noticed in Jurassic rocks in W. Australia, but no species have been described.
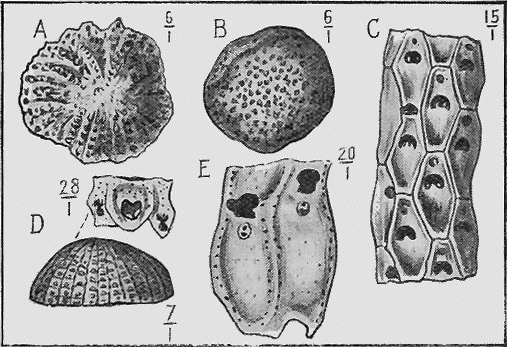
Cheilostomata (Cretaceous).—
Species of the genera (?) Membranipora and (?) Lepralia, belonging to the Cheilostomata, have been described from the Lower Cretaceous of the Darling River, New South Wales, and Wollumbilla, Queensland, respectively.
Cainozoic Polyzoa.—
A very large number of genera of the Polyzoa have been described from the Tertiary strata of South Australia and Victoria. Some of the principal of these are Crisia, Idmonea, Stomatopora, Lichenopora, Hornera, Entalophora and Heteropora of the order Cyclostomata; and Catenicella, Cellaria, Membranipora, Lunulites, Selenaria, Macropora, Tessarodoma, Adeona, Lepralia, Bipora, Smittia, Porina, Cellepora and Retepora of the order Cheilostomata. Many of these genera, and not a few Australian species, are found also in the Cainozoic or Tertiary beds of Orakei Bay, New Zealand (Fig. 84).
BRACHIOPODA (Lamp-shells).
Brachiopods: Their Structure.—
These are marine animals, and are enclosed in a bivalved shell. They differ, however, from true bivalves (Pelecypoda) in having the shell on the back and front of the body, instead of on each side as in the bivalved mollusca. Each valve is equilateral, but the valves differ from one another in that one is larger and generally serves to attach the animal to rocks and other objects of support by a stalk or pedicle. Thus the larger valve is called the pedicle valve and the smaller, on account of its bearing the calcareous supports for the brachia or arms, the brachial valve. Generally speaking, the shell of the valve is penetrated by numerous canals, which give the shell a punctate appearance. Some brachiopod shells, as Atrypa and Rhynchonella, are, however, devoid of these.
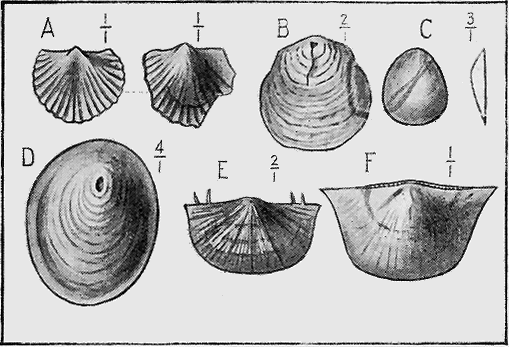
Cambrian Brachiopods.—
Brachiopods are very important fossils in Australasian rocks. They first appear in Cambrian strata, as for example, in the Florentine Valley, in Tasmania, where we find Orthis lenticularis (Fig. 85 A). In Victoria, near Mount Wellington, in the mountainous region of N.E. Gippsland, Orthis platystrophioides is found in a grey limestone. In South Australia the grey Cambrian limestone of Wirrialpa contains the genus Huenella (H. etheridgei). This genus is also found in the Middle and Upper Cambrian of N. America.
Ordovician Brachiopods.—
Coming to Ordovician rocks, the limestones of the Upper Finke Basin in South Australia contain Orthis « 160 » leviensis and O. dichotomalis. The Victorian mudstone at Heathcote may be of Ordovician age or even older; it has afforded a limited fauna of brachiopods and trilobites, amongst the former being various species of Orthis, Chonetes, and Siphonotreta. The latter genus is represented in both the Lower and Upper Ordovician rocks of slaty character in Victoria (Fig. 85 B).
Silurian Brachiopods.—
The Silurian system in Australasia as in Europe, N. America and elsewhere, is very rich in brachiopod life. It is impossible to enumerate even all the genera in a limited work like the present, the most typical only being mentioned.
In New Zealand the palaeozoic fauna is at present imperfectly worked out, but the following genera from the Wangapekian (Silurian) have been identified, viz., Chonetes, Stricklandinia, Orthis, Wilsonia, Atrypa, and Spirifer. The specific identification of these forms with European types is still open to question, but the species are undoubtedly closely allied to some of those from Great Britain and Scandinavia.
The Victorian Silurian Brachiopods are represented by the horny-shelled Lingula, the conical Orbiculoidea, a large species of Siphonotreta, Stropheodonta (with toothed hinge-line), Strophonella, Chonetes (with hollow spines projecting from the ventral valve, one of the species C. melbournensis being characteristic of the Melbournian division of Silurian rocks), Orthis, Pentamerus, Camarotoechia, Rhynchotrema, Wilsonia, Atrypa (represented by the world-wide A. reticularis), Spirifer and Nucleospira (Figs. 85, 86).
New South Wales has a very similar assemblage of genera; whilst Tasmania possesses Camarotoechia, Stropheodonta and Orthis.
Devonian Brachiopods.—
The Devonian limestones and associated strata are fairly rich in Brachiopods. The Victorian rocks of this age at Bindi and Buchan contain genera such as Chonetes (C. australis), Spirifer (S. yassensis and S. howitti) and Athyris.
In New South Wales we again meet with Spirifer yassensis, veritable shell-banks of this species occurring in the neighbourhood of Yass, associated with a species of Chonetes (C. culleni) (Fig. 86 D, E).

In the Upper Devonian of New South Wales abundant remains occur of both Spirifer disjunctus and Camarotoechia pleurodon (var.).
The Upper Devonian Series at Nyrang Creek near Canowindra, New South Wales, contains a Lingula (L. gregaria) associated with the Lepidodendron plant beds of that locality.
Queensland Devonian rocks contain Pentamerus, Atrypa and Spirifer. In Western Australia the Devonian species are Atrypa reticularis, Spirifer cf verneuili, S. musakheylensis and Uncinulus cf. timorensis.
Carboniferous Brachiopods.—
The Carboniferous Brachiopod fauna is represented in New South Wales at Clarence Town and other localities by a species which has an extensive time-range, Leptaena rhomboidalis var. analoga, and the following, a few of which extend upwards into the Carbopermian:—Chonetes papilionacea, Productus semireticulatus, P. punctatus, P. cora, Orthothetes crenistria, Orthis (Rhipidomella) australis, O. (Schizophoria) resupinata, Spirifer striatus, S. bisulcatus, Cyrtina carbonaria and Athyris planosulcatus.
In New Zealand the Matai series, referred to the Jurassic by Hutton, as formerly regarded by Hector, and latterly by Park, as of Carboniferous age, on the ground of a supposed discovery of Spirifer subradiatus (S. glaber) and Productus brachythaerus in the Wairoa Gorge. Although these species may not occur, the genera Spirifer and Productus are present, which, according to Dr. Thomson, are distinctly of pre-Triassic types.
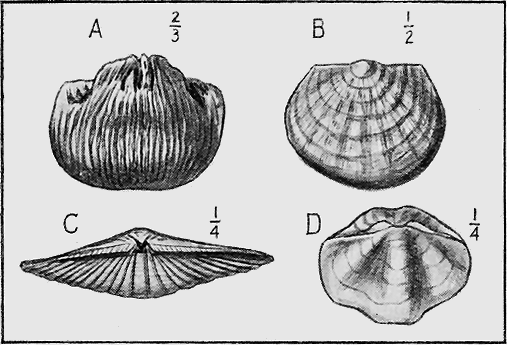
Carbopermian Brachiopods.—
The Brachiopod fauna of Carbopermian age in New South Wales is rich in species of Productus and Spirifer. Amongst the former are P. cora (also found in Western Australia, Queensland and Tasmania), P. brachythaerus (also found in Western Australia and Queensland), (Fig. 87 A), P. semireticulatus (also found in Western Australia, Queensland and the Island of Timor, and a common species in Europe), and P. undatus (also found in Western Australia and Queensland, as well as in Great Britain and Russia). Strophalosia is an allied genus to Productus. It is a common form in beds of the same age in W. Australia, Tasmania, and New South Wales. The best « 164 » known species is S. clarkei (Fig. 87 B). This type of shell is distinguished from Productus in being cemented by the umbo of the ventral valve, which valve is also generally less spinose than the dorsal. When weathered the shells present a peculiar silky or fibrous appearance. The genus Spirifer is represented in W. Australia by such forms as S. vespertilio, S. convolutus, S. hardmani, S. musakheylensis, and S. striatus; whilst S. vespertilio and S. convolutus are common also to New South Wales (Fig. 87 C), and the latter only to Tasmania. S. vespertilio is found in the Gympie beds near Rockhampton, Queensland; and S. tasmaniensis in Queensland (Bowen River Coal-field, Marine Series), New South Wales and Tasmania. Of the smoother, stout forms, referred to the sub-genus Martiniopsis, we may mention S. (M.) subradiatus, which occurs in W. Australia, New South Wales, and Tasmania (Fig. 87 D).
In the Queensland fauna, the Gympie series contains, amongst other Brachiopods Productus cora, Leptaena rhomboidalis var., analoga, Spirifer vespertilio and S. strzeleckii.
Other Carbopermian Brachiopod genera found in Australian faunas are Cleiothyris, Dielasma, Hypothyris, Reticularia, Seminula, Cyrtina, and Syringothyris.
Triassic Brachiopods.—
The Kaihiku Series of New Zealand (Hokonui Hills and Nelson) are probably referable to the Trias. The supposed basal beds contain plants such as Taeniopteris, Cladophlebis, Palissya and Baiera. Above these are marine beds containing Brachiopods belonging to « 165 » Spiriferina, Rhynchonella, Dielasma and Athyris. The succession of these beds presents some palaeontological anomalies still to be explained, for the flora has a decided leaning towards a Jurassic facies.
Next in order of succession the Wairoa Series, in the Hokonui Hills and Nelson, New Zealand, contains Dielasma and Athyris wreyi.
The succeeding series in New Zealand, the Otapiri, or Upper Triassic contains the Brachiopod genera Athyris[3] and Spiriferina, found at Well’s Creek, Nelson.
[3] Referred by Hector to a new sub-genus Clavigera, which name, however, is preoccupied.
Jurassic Brachiopods.—
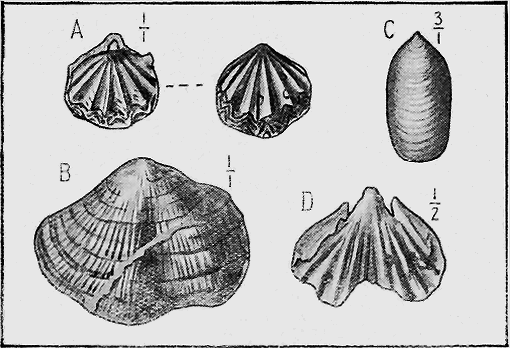
The marine Jurassic beds of W. Australia, as at Shark Bay and Greenough River, contain certain « 166 » Rhynchonellae allied to European species, as R. variabilis (Fig. 88 A), and R. cf. solitaria.
Lower Cretaceous Brachiopods.—
The Lower Cretaceous or Rolling Downs Formation of Queensland has yielded a fair number of Brachiopods, principally from Wollumbilla,—as Terebratella davidsoni (Fig. 88 B), (?) Argiope wollumbillensis, (?) A. punctata, Rhynchonella rustica, R. solitaria, Discina apicalis and Lingula subovalis. From beds of similar age in Central South Australia and the Lake Eyre Basin Lingula subovalis (Fig. 88 C), and Rhynchonella eyrei have been recorded; the latter has been compared with a species (R. walkeri) from the Middle Neocomian of Tealby in Yorkshire.
Upper Cretaceous Brachiopod.—
A solitary species of the Brachiopoda occurs in the Upper Cretaceous of Australia, namely, Rhynchonella croydonensis (Fig. 88 D) of the Desert Sandstone of the Croydon Gold-fields and Mount Angas, Queensland.
Cainozoic Brachiopods.—
The Brachiopoda of the Cainozoic or Tertiary strata of Australia and New Zealand are well represented by the genera Terebratula, Magellania, Terebratulina, Terebratella, Magasella and Acanthothyris. In the Balcombian or Oligocene of southern Australia occur the following:—Terebratula tateana, Magellania corioensis, M. garibaldiana and Magasella compta (Figs. 89 A, D); and most of these range into the next stage, the Janjukian, whilst some extend even to the Kalimnan. Terebratulina suessi, Hutton sp. (= T. scoulari, Tate) ranges through the Balcombian « 167 » and Janjukian, but is most typical of the Janjukian beds in Victoria: it also occurs in the Oamaru Series of New Zealand (= Janjukian). Acanthothyris squamosa (Fig. 89 F) is typical of the Janjukian of southern Australia, and it occurs also in the Pareora beds of the Broken River, New Zealand. The latter are green, sandy, fossiliferous strata immediately succeeding the Oamaru stone of the Hutchinson Quarry beds. A. squamosa is said to be still living south of Kerguelen Island. Magellania insolita is a Victorian species which is also found in the Oamaru Series of New Zealand.

Whilst many of the older Tertiary brachiopods range into the next succeeding stage of the Kalimnan in Victoria, such as Magellania insolita, Terebratulina « 168 » catinuliformis (Fig. 89 E) and Magasella compta, one species, Terebratella pumila, is restricted to the Kalimnan, occurring at the Gippsland Lakes.
The next stage, the Werrikooian, typical in upraised marine beds on the banks of the Glenelg River in western Victoria, contains Magellania flavescens, a species still living (see antea, Fig. 23), and M. insolita, having the extraordinarily wide range of the whole of the Cainozoic stages in southern Australia.
COMMON OR CHARACTERISTIC FOSSILS OF THE FOREGOING CHAPTER.
WORMS.
Eunicites mitchelli, Eth. fil. Silurian: New South Wales.
Oenonites hebes, Eth. fil. Silurian: New South Wales.
Arabellites bowningensis, Eth. fil. Silurian: New South Wales.
Arenicolites sp. Silurian: New South Wales.
Trachyderma crassituba, Chapm. Silurian: Victoria.
Cornulites tasmanicus, Eth. fil. Silurian: Tasmania.
Spirorbis ammonius, M. Edw. var. truncata, Chapm. Mid. Devonian: Victoria.
Spirorbis omphalodes, Goldfuss. Devonian: W. Australia.
Serpula testatrix, Eth. fil. Carbopermian: New South Wales.
Torlessia mackayi, Bather. Lower Mesozoic: New Zealand.
Serpula conformis, Goldfuss. Jurassic: W. Australia.
Serpula intestinalis, Phillips. Lower Cretaceous: Queensland.
Serpula subtrachinus, Eth. fil. Lower Cretaceous: New South Wales.
Serpula ouyenensis, Chapm. Cainozoic: Victoria.
Ditrupa cornea, L. sp. var. wormbetiensis, McCoy. Cainozoic: Victoria.
POLYZOA.
Rhombopora gippslandica, Chapm. Silurian: Victoria.
Fenestella australis, Chapm. Silurian: Victoria.
Protoretepora ampla, Lonsdale. Carbopermian: W. Australia, New South Wales, Queensland, and Tasmania.
Polypora australis, Hinde. Carbopermian: W. Australia.
Rhombopora tenuis, Hinde. Carbopermian: W. Australia.
Rhombopora laxa, Etheridge sp. Carbopermian: Queensland.
Membranipora wilsonensis, Eth. fil. Lower Cretaceous: New South Wales.
(?) Lepralia oolitica, Moore. Lower Cretaceous: Queensland.
Lichenopora australis, MacGillivray. Cainozoic: Victoria.
Heteropora pisiformis, MacGillivray. Cainozoic: Victoria.
Cellaria australis, MacGillivray. Cainozoic: Victoria.
Membranipora macrostoma, Reuss. Cainozoic: Victoria (also living).
Selenaria marginata, T. Woods. Cainozoic: Victoria (also living).
Macropora clarkei, T. Woods sp. Cainozoic: Victoria.
Adeona obliqua, MacGill. Cainozoic: Victoria.
Lepralia burlingtoniensis, Waters. Cainozoic: Victoria.
Bipora philippinensis, Busk sp. Cainozoic: Victoria (also living).
Porina gracilis, M. Edwards sp. Cainozoic: Victoria (also living).
Cellepora fossa, Haswell, sp. Cainozoic: Victoria (also living).
Retepora fissa, MacGill. sp. Cainozoic: Victoria (also living).
BRACHIOPODA.
Orthis lenticularis, Wahlenberg sp. Cambrian: Tasmania.
Orthis platystrophioides, Chapm. Cambrian: Victoria.
Huenella etheridgei, Walcott. Cambrian: S. Australia.
Orthis leviensis, Eth. fil. Ordovician: S. Australia, (?) Victoria.
Siphonotreta discoidalis, Chapm. Ordovician: Victoria.
Siphonotreta maccoyi, Chapm. Ordovician: Victoria.
Lingula yarraensis, Chapm. Silurian: Victoria.
Orbiculoidea selwyni, Chapm. Silurian: Victoria.
Chonetes melbournensis, Chapm. Silurian: Victoria.
Stropheodonta alata, Chapm. Silurian: Victoria.
Orthis elegantula, Dalman. Silurian: Victoria.
Pentamerus australis, McCoy. Silurian: Victoria and New South Wales.
Conchidium knightii, Sow. sp. Silurian: Victoria and New South Wales.
Camarotoechia decemplicata, Sow. sp. Silurian: Victoria.
Rhynchotrema liopleura, McCoy sp. Silurian: Victoria.
Atrypa reticularis, L. sp. Silurian: New South Wales and Victoria. Devonian: New South Wales, W. Australia and Queensland.
Spirifer sulcatus, Hisinger sp. Silurian: Victoria.
Nucleospira australis, McCoy. Silurian: Victoria.
Chonetes australis, McCoy. Mid. Devonian: Victoria.
Chonetes culleni, Dun. Mid. Devonian: New South Wales.
Spirifer yassensis, de Koninck. Mid. Devonian: New South Wales and Victoria.
Spirifer cf. verneuili, de Kon. Mid. Devonian: New South Wales and W. Australia.
Lingula gregaria, Eth. fil. Upper Devonian: New South Wales.
Spirifer disjunctus, Sow. Up. Devonian: New South Wales.
Productus cora, d’Orb. Carboniferous: New South Wales and Queensland.
Orthothetes crenistria, Sow. sp. Carboniferous: New South Wales.
Spirifer striatus, Sow. Carboniferous: New South Wales.
Productus brachythaerus, Sow. Carbopermian: New South Wales, Queensland, W. Australia.
Strophalosia clarkei, Eth. sp. Carbopermian: New South Wales, Tasmania and W. Australia.
Spirifer (Martiniopsis) subradiatus, Sow. Carbopermian: New South Wales, Tasmania and W. Australia.
Spirifer convolutus, Phillips. Carbopermian: New South Wales, Tasmania and W. Australia.
Cleiothyris macleayana, Eth. fil. sp. Carbopermian: W. Australia.
Dielasma elongata, Schlotheim sp. Trias (Kaihiku Series): New Zealand.
Athyris wreyi, Suess sp. Trias (Wairoa Series): New Zealand.
Athyris sp. Trias (Otapiri Series): New Zealand.
Rhynchonella variabilis, Schlotheim sp. Jurassic: W. Australia.
Terebratella davidsoni, Moore. Lower Cretaceous: Queensland.
Rhynchonella solitaria, Moore. Lower Cretaceous: Queensland.
Lingula subovalis, Davidson. Lower Cretaceous: Queensland and S. Australia.
Rhynchonella croydonensis, Eth. fil. Upper Cretaceous: Queensland.
Terebratula tateana, T. Woods. Cainozoic (Balcombian and Janjukian): Victoria and S. Australia.
Magellania corioensis, McCoy, sp. Cainozoic (Balcombian and Janjukian): Victoria and S. Australia.
Magellania garibaldiana, Davidson sp. Cainozoic (Balcombian and Janjukian): Victoria and S. Australia.
Magasella compta, Sow. sp. Cainozoic (Balcombian to Kalimnan): Victoria and S. Australia.
Terebratula suessi, Hutton sp. Cainozoic (Balcombian and Janjukian): Victoria, S. Australia, and New Zealand (Oamaru Series.)
Acanthothyris squamosa, Hutton sp. Cainozoic (Janjukian): Victoria and S. Australia, New Zealand (Oamaru Series) (also living).
Terebratella pumila, Tate. Cainozoic (Kalimnan): Victoria.
Magellania flavescens, Lam. sp. Pleistocene: Victoria (also living).
LITERATURE.
WORMS.
Silurian.—Etheridge, R. jnr. Geol. Mag., Dec. III. vol. VII. 1890, pp. 339, 340. Idem, Proc. Roy. Soc. Tas. (for 1896), 1897, p. 37. Chapman, F. Proc. R. Soc. Vict., vol. XXII. (N.S.), pt. II. 1910, pp. 102-105.
Devonian.—Hinde, G. J. Geol. Mag., Dec. II. vol. VII. 1890, p. 199. Chapman, F. Rec. Geol. Surv. Vict., vol. III. pt. 2, 1912, p. 220.
Carboniferous.—Etheridge, R. jnr. Bull. Geol. Surv. W. Australia, No. 10, 1903, p. 10.
Carbopermian.—Etheridge, R. jnr. Mem. Geol. Surv. New South Wales. Pal. No. 5, 1892, pp. 119-121.
Lower Mesozoic.—Bather, F. A. Geol. Mag., Dec. V. vol. II. 1905, pp. 532-541.
Lower Cretaceous.—Etheridge, R. jnr. Mem. Soc. Geol. Surv. New South Wales, Pal. No. 11. 1902, pp. 12, 13.
Cainozoic.—Chapman, F. Proc. R. Soc. Vict., vol. XXVI. (N.S.) pt. I. 1913, pp. 182-184.
POLYZOA.
Silurian.—Chapman, F. Proc. R. Soc. Vict., vol. XVI. (N.S.), pt. I. 1903, pp. 61-63. Idem, Rec. Geol. Surv. Vic., vol. II., pt. 1, 1907, p. 78.
Carboniferous.—Hinde, G. J. Geol. Mag. Dec. III. vol. VII. 1890, pp. 199-203.
Carbopermian.—De Koninck Mem. Geol. Surv. New South Wales, Pal. No. 6, 1898, pp. 128-140.
Cainozoic.—Stolicka, F. Novara Exped., Geol. Theil., vol. I. pt. 2, pp. 87-158. Waters, A. W. Quart. Journ. Geol. Soc., vol. XXXVII. 1881, pp. 309-347; ibid., vol. XXXVIII. 1882, pp. 257-276 and pp. 502-513; ibid., vol. XXXIX. 1883, pp. 423-443; ibid., vol. XL. 1884, pp. 674-697; ibid., vol. XLI. 1885, pp. 279-310; ibid., vol. XLIII. 1887, pp. 40-72 and 337-350. MacGillivray, P. H. Mon. Tert. Polyzoa Vict., Trans. Roy. Soc. Vict., Vol. IV. 1895. Maplestone, C. M. “Further Descr. Polyzoa Vict.,” Proc. Roy. Soc. Vict., vol. XI. (N.S.), pt. I. 1898, pp. 14-21, et seqq.
BRACHIOPODA.
Cambrian.—Tate, R. Trans. R. Soc. S. Austr., vol. XV. 1892, pp. 185, 186. Etheridge, R. jnr. Rec. Austr. Mus., vol. V. pt. 2, 1904, p. 101. Walcott, C. D. Smiths. Misc. Coll., vol. LIII. 1908, p. 109. Chapman, F. Proc. R. Soc. Vic., vol. XXIII. (N.S.), pt. I. 1911, pp. 310-313.
Ordovician.—Etheridge, R. jnr. Parl. Papers, S. Aust., No. 158, 1891, pp. 13, 14. Tate, R. Rep. Horn Exped., pt. 3, 1896, pp. 110, 111. Chapman, F. Rec. Geol. Surv. Vict., vol. I. pt. 3, 1904, pp. 222-224.
Silurian.—McCoy, F. Prod. Pal. Vic. Dec. V. 1877, pp. 19-29. Eth., R. jnr. Rec. Geol. Surv. New South Wales, vol. 3, pt. 2, 1892, pp. 49-60 (Silurian and Devonian Pentameridae). Idem, Proc. Roy. Soc., Tas., (for 1896), 1897, pp. 38-41. De Koninck, L. G. Mem. Geol. Surv. New South Wales, Pal. No. 6, 1898, pp. 20-29. Dun, W. S. Rec. Geol. Surv. New South Wales, vol. VII. pt. 4, 1904, pp. 318-325 (Silurian to Carboniferous). Ibid., vol. VIII. pt. 3, 1907, pp. 265-269. Chapman, F. Proc. R. Soc. Vict., vol. XVI. (N.S.), pt. 1, 1903, pp. 64-79. Ibid., vol. XXI. (N.S.), pt. 1, 1908, pp. 222, 223. Ibid., vol. XXVI. (N.S.) pt. 1. 1913, pp. 99-113.
Devonian.—McCoy, F. Prod. Pal. Vict., Dec. IV., 1876, pp. 16-18. Foord, A. H. Geol. Mag., Dec. III. vol. VII. 1890, pp. 100-102. Etheridge, R. jnr. Geol. and Pal. Queensland, 1892, pp. 64-68. De Koninck, L. G. Mem. Geol. Surv. New South Wales, Pal., No. 6, 1898, pp. 64-85. Chapman, F. Proc. R. Soc. Vict., vol. XVIII. (N.S.), pt. 1, 1905, pp. 16-19.
Carboniferous.—Etheridge, R. jnr. Rec. Austr. Mus., vol. IV. No. 3, 1901, pp. 119, 120. Idem, Geol. Surv. W. Austr., Bull. No. 10, 1903, pp. 12-23. Dun, W. S. Rec. Geol. Surv. New South Wales, vol. VII., pt. 2, 1902, pp. 72-88 and 91-93.
Carbopermian.—Sowerby, G. B., in Strzelecki’s Phys. Descr. of New South Wales, etc., 1845, pp. 275-285. McCoy, F. Ann. Mag. Nat. Hist., vol. XX. 1847, pp. 231-236. Foord, A. H. Geol. Mag. Dec. III. vol. VII. 1890, pp. 105 and 145-154. Etheridge, R. jnr. Geol. and Pal. Queensland, 1892, pp. 225-264. De Koninck, L. G. Mem. Geol. Surv. New South Wales, Pal., No. 6, 1898, pp. 140-203. Dun, W. S. Rec. Geol. Surv. New South Wales, vol. VIII. pt. 4, 1909, pp. 293-304.
Lower Cretaceous.—Moore, C. Quart. Journ. Geol. Soc., vol. XXVI. 1870, pp. 243-245. Etheridge, R. jnr. Mem. R. Soc. S. Austr., vol. II. pt. 1, 1902, pp. 8, 9.
Upper Cretaceous.—Etheridge, R. jnr. Geol. and Pal. Queensland, 1892, p. 560.
Cainozoic.—McCoy, F. Prod. Pal. Vict., Dec. V. 1877, pp. 11-13. Tate, R. Trans. R. Soc. S. Austr., vol. III. 1880, pp. 140-170. Idem, ibid., vol. XXIII. 1899, pp. 250-259. Hutton, F. W. Trans. N.Z. Inst., vol. XXXVII. 1905, pp. 474-481 (Revn. Tert. Brach.).
FOSSIL SHELL-FISH (MOLLUSCA).
Molluscan Characters.—
The phylum or sub-kingdom Mollusca is a group of soft-bodied animals (mollis, soft), which, although having no external skeleton, usually possess the protective covering of a shell. This shell is secreted from the outer skin or mantle, and is composed of carbonate of lime (calcareous) with a varying proportion of organic material.
Hard Parts.—
Fossil molluscan remains consist practically of the shells, but the calcareous apertural lid (operculum) of some kinds is often preserved, as in Turbo and Hyolithes; or the horny lids of others, as Bithynia of the European Pleistocene “brick earths.” The cuttle-fishes have hard, horny beaks and internal bones, and the latter are frequently found fossil in Australia.
Characters of Pelecypoda.—
The class for first consideration is the important one of the Bivalved Mollusca, the LAMELLIBRANCHIATA (“plate-gills”) or PELECYPODA « 175 » (“hatchet foot”). The shells are double, hinged dorsally and placed on either side of the animal, that is, they are left and right. The height is measured on a vertical line drawn from the beaks or umbones to the ventral margin. The length is the greatest distance between the margins parallel with a line drawn through the mouth and posterior adductor impression. The thickness is measured by a line at right angles to the line of height. The shell being placed mouth forward, the valves are thus left and right. The anterior is usually shorter, excepting in some cases, as in Donax and Nucula.
Hinge Structure.—
In the absence of the animal, the character of the hinge-structure is very important. Some are without teeth (edentulous). The oldest forms have been grouped as the “Palaeoconcha,” and it has been shown that here, although well-developed teeth were absent, the radial ribs of the surface and ventral areas were carried over to the dorsal margin and became a fixed character in the form of crenulations or primitive teeth.
The taxodont type of hinge teeth shows alternating teeth and sockets, as in Nucula.
The schizodont type is seen in the heavy, variable teeth of Trigonia and Schizodus.
The isodont type of hingement is a modification of the taxodont, represented by two ridges originally divergent below the beak, and forming an interlocking series of two pairs of teeth and sockets as in Spondylus; or where the primitive hinge disappears as in Pecten, the divergent ridge-teeth (crura) may only partially develop.
The dysodonts have a feeble hinge-structure derived from the external sculpture impinging on the hinge-line, as in Crenella.
The pantodonta are an ancient palaeozoic group which seems allied to the modern teleodont or long toothed shells, but the laterals may exceed a pair in a single group, as in Allodesma.
The diogenodonta have lateral and cardinal teeth upon a hinge-plate, but never more than two laterals and three cardinals in any one group, as in Crassatellites.
The cyclodonta have extremely arched teeth, which curve out from under the beaks, as in Cardium.
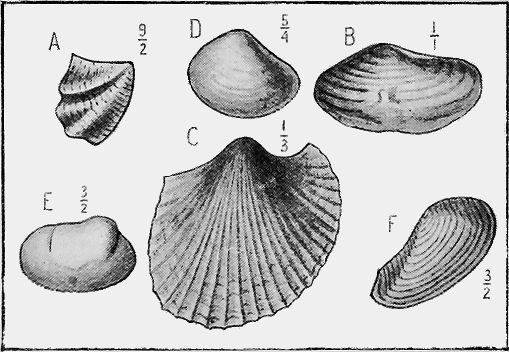
The teleodonts include the more highly developed types of hinge, with attenuated teeth and sockets. Common shells of our coast, and from Cainozoic beds, belonging to this group are Venus, Mactra and Meretrix.
The asthenodonta are boring and burrowing molluscs that have lost the hinge dentition from disuse as Corbula and Pholas.
Cambrian Bivalve.—
The earliest example of a bivalved shell in Australian rocks is Ambonychia macroptera (Fig. 90 A), which occurs in the Cambrian Limestone of Curramulka, S. Australia. It is quite a small form, being less than a quarter of an inch in length.
Ordovician Bivalve.—
In the basal Ordovician mudstone of Heathcote, Victoria, there is a bivalve which in some respects resembles a Modiolopsis (?M. knowsleyensis), but the exact relationship is still doubtful.
Silurian Bivalves.—
The Silurian sandstones, mudstones, slates and limestones of Australia and New Zealand, unlike the older rocks just mentioned, contain a rich assemblage of bivalve fossils. In Victoria the lower division or Melbournian stage contains the following principal genera:—Orthonota, Grammysia, Leptodomus, Edmondia, Cardiola, Ctenodonta, Nuculites, Nucula, Palaeoneilo, Conocardium, Modiolopsis and Paracyclas. The upper division or Yeringian stage contains other species of similar genera to those in the Melbournian, as Grammysia, Palaeoneilo and Conocardium; whilst Panenka, Mytilarca, Sphenotus, « 178 » Actinodesma, Lunulicardium, Actinopteria and Cypricardinia are, so far as known, peculiar to this and a still higher stage. Cardiola is a widely distributed genus, occurring as well in Tasmania; whilst in Europe it is found both in Bohemia and Great Britain. Its time-range in the northern hemisphere is very extensive, being found in beds ranging from Upper Ordovician to Devonian. Actinopteria is found also in New South Wales and New Zealand, and Pterinea and Actinodesma in New South Wales.
The molluscs with a taxodont hinge-line (beset with numerous little teeth and sockets) are quite plentiful in the Australian Silurian; such as Nucula, a form common around Melbourne (N. melbournensis (Fig. 90 D)); Nuculites, which has an internal radial buttress or clavicle separating the anterior muscle-scar from the shell-cavity, and which is found likewise in the Melbourne shales (N. maccoyianus (Fig. 90 E)); Ctenodonta, represented in both the Melbournian and Yeringian stages (C. portlocki); and Palaeoneilo, a handsome, subrostrate generic type with concentric lamellae or striae, commonest in the Melbournian, but occasionally found in the younger stage (P. victoriae Fig. 90 F, Melbournian;—P. raricostae, Yeringian). Conocardium is represented by two species in Victoria (C. bellulum and C. costatum); whilst in New South Wales C. davidis is found at Oakey Creek. In New Zealand Actinopteria and Pterinea occur in the Wangapeka series (Silurian).
Devonian Bivalves.—
The compact limestone and some shales of Middle Devonian age in the N.E. Gippsland area in Victoria, « 179 » contain several as yet undescribed species belonging to the genera Sphenotus, Actinodesma and Paracyclas.
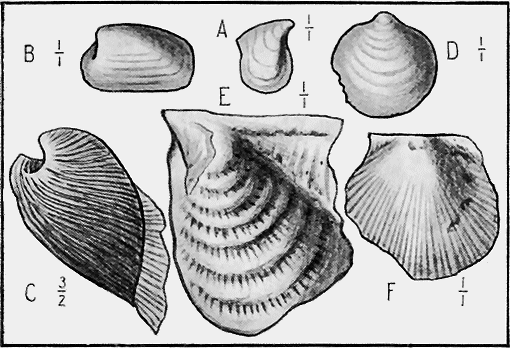
The genera Paracyclas, Aviculopecten and Pterinea have been recorded from New South Wales, chiefly from the Yass district. The derived boulders found in the Upper Cretaceous beds forming the opal-fields at White Cliffs, New South Wales, have been determined as of Devonian age. They contain, amongst other genera, examples of Actinopteria (A. australis), Lyriopecten (L. gracilis) (Fig. 91 F), and Leptodesma (L. inflatum and L. obesum).
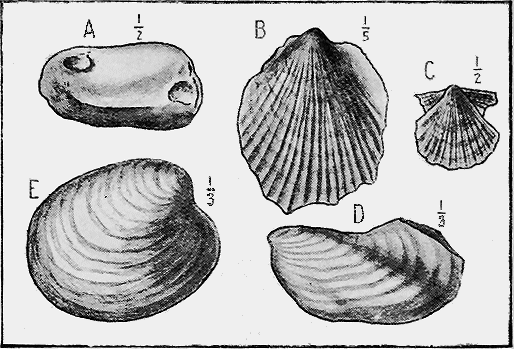
Carbopermian Bivalves.—
One of the most prolific palaeozoic series for bivalved mollusca is the Carbopermian. To select from the numerous genera and species we may mention Stutchburia farleyensis (Fig. 92 A) and Edmondia nobilissima from Farley, New South Wales; and Deltopecten limaeformis (Fig. 92 B), found in the Lower Marine Series at Ravensfield, New South Wales, and in the Upper Marine Series at Burragorang and Pokolbin in the same State, in Queensland at the Mount Britton Gold-field, and in Maria Id., Tasmania. Deltopecten fittoni occurs in both series in New South Wales, and in the Upper Marine Series associated with “Tasmanite shale” in Tasmania. Aviculopecten squamuliferus is a handsome species found alike in Tasmania and New South Wales; whilst A. tenuicollis is common to W. Australia and New South Wales. Other characteristic bivalves of the Carbopermian of New South Wales « 181 » are Chaenomya etheridgei (Fig. 92 D) and Pachydomus globosus (Fig. 92 E). The gigantic Eurydesma cordatum is especially characteristic of the New South Wales Lower Marine Series, and is also found in Tasmania. All three species are found in Queensland.
Triassic Bivalves.—
The Triassic rocks of New South Wales were accumulated under either terrestrial, lacustrine, or brackish (estuarine) conditions. Hence the only bivalved mollusca found are referred to the freshwater genera Unio (U. dunstani) and Unionella (U. bowralensis and U. carnei (Fig. 93 A)). The latter genus differs from Unio in the structure of the adductor muscle-impressions.
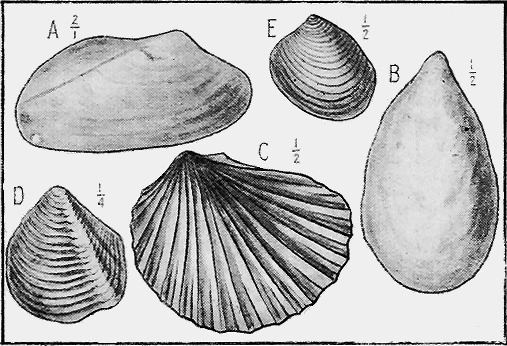
The Queensland Trias (Burrum Formation) contains a solitary species of bivalved mollusca, Corbicula burrumensis. This genus is generally found associated with freshwater or brackish conditions.
In New Zealand marine Triassic beds occur, containing, amongst other genera, a species of Leda. In the succeeding Wairoa Series the interesting fossil, Daonella lommeli occurs. This shell is typical of the Norian (Upper Trias) of the Southern Tyrol. Above the Daonella bed occurs the Trigonia bed, with that genus and Edmondia. In the next younger stage, the Otapiri Series, near Nelson, there are fine-grained sandstones packed full of the remains of Mytilus problematicus (Fig. 93 B) and Monotis salinaria (Fig. 93 C), the latter also a Norian fossil.
Jurassic Bivalves.—
Jurassic bivalved molluscs are plentiful in the W. Australian limestones, as at Greenough River. Amongst others may be mentioned Cucullaea semistriata, Ostrea, Gryphaea, Trigonia moorei (Fig. 93 D), Pecten cinctus, Ctenostreon pectiniforme and Astarte cliftoni (Fig. 93 E). Several of the species found are identical with European Jurassic fossils.
Jurassic strata in Victoria, being of a freshwater and lacustrine nature, yield only species of Unio, as U. dacombei, and U. stirlingi.
The Jurassic beds of S. Australia contain a species of Unio named U. eyrensis. In the same strata which contains this shell, plant remains are found, as Cladophlebis and Thinnfeldia, two well-known types of Jurassic ferns.
Lower Cretaceous Bivalves.—
In Queensland the Lower Cretaceous limestones and marls contain a large assemblage of bivalves, the more important of which are Nucula truncata (Fig. 94 A), Maccoyella reflecta (Fig. 94 B), M. barkleyi, Pecten socialis and Fissilunula clarkei (Fig. 94 C), from Wollumbilla; and Inoceramus pernoides, I. carsoni and Aucella hughendenensis from the Flinders River (the latter also from New South Wales).
In the Lake Eyre District of S. Australia we find Maccoyella barkleyi, which also occurs in Queensland and New South Wales (at White Cliffs), Trigonia cinctuta, Mytilus rugocostatus and Modiola eyrensis. The handsome bivalve, Pleuromya plana occurs near Broome in W. Australia.

Upper Cretaceous Bivalves.—
The Upper Cretaceous or Desert Sandstone at Maryborough, Queensland, has yielded amongst others, the following shells:—(Nucula gigantea, Maccoyella reflecta also found in the Lower Cretaceous of Queensland, New South Wales and S. Australia), and Fissilunula clarkei (also found in the L. Cretaceous of New South Wales, Queensland and S. Australia). Some of these beds, however, which were hitherto believed to belong to the Upper and Lower Series respectively may yet prove to be on one horizon—the Lower Cretaceous. Cyrenopsis opallites (Fig. 94 E) of White Cliffs, New South Wales, appears to be a truly restricted Upper Cretaceous species.
The Cretaceous of New Zealand (Amuri System) contains Trigonia sulcata, Inoceramus sp. and the curious, contorted shell, Conchothyra parasitica (Fig. 94 F) which is related to Pugnellus, a form usually considered as a sub-genus of Strombus.
From Papua an Inoceramus has been recorded from probable Cretaceous beds.
Cainozoic Bivalves.—
In Victoria, South Australia, and the N.W. of Tasmania, as well as in New Zealand, Cainozoic marine beds are well developed, and contain an extensive bivalved molluscan fauna. Of these fossils only a few common and striking examples can here be noticed, on account of the limits of the present work.
The commonest genera are:—Ostrea, Placunanomia, Dimya, Spondylus, Lima, Pecten, Arca, Barbatia, Plagiarca, Cucullaea, Glycimeris, Limopsis, Nucula, Leda, Trigonia, Cardita, Cuna, Crassatellites, Cardium, « 185 » Protocardium, Chama, Meretrix, Venus (Chione), Dosinea, Gari, Mactra, Corbula, Lucina, Tellina, Semele and Myodora.

Persistent Species.—
To mention a few species of persistent range, from Balcombian to Kalimnan, we may cite the following from the Cainozoic of southern Australia:—Dimya dissimilis (Fig. 95 A), Spondylus pseudoradula (Fig. 95 B), Lima (Limatula) jeffreysiana, Pecten polymorphoides (found also in the Oamaru Series, New Zealand) (Fig. 95 C), Amusium zitteli (found also in both the Waimangaroa and Oamaru Series of New Zealand), Barbatia celleporacea, Cucullaea corioensis, Limopsis maccoyi, Nucula tenisoni, Leda vagans (Fig. 95 D), Corbula ephamilla and Myodora tenuilirata.
Balcombian Bivalves.—
On the other hand, many species have a restricted range, and these are invaluable for purposes of stratigraphical correlation. For example, in the Balcombian we have Modiola praerupta (Fig. 95 E), Modiolaria balcombei, Cuna regularis, Cardium cuculloides, Cryptodon mactraeformis, Verticordia pectinata and V. excavata.
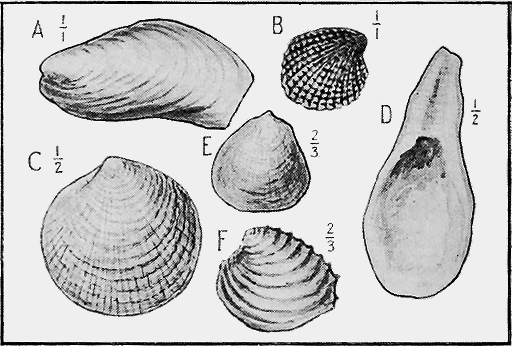
Janjukian Bivalves.—
In the Janjukian Series restricted forms of bivalves are exceptionally numerous, amongst them being:—Dimya sigillata, Plicatula ramulosa, Lima polynema, Pecten praecursor, P. eyrei, P. gambierensis, Pinna cordata, Modiola pueblensis (Fig. 96 A), Arca dissimilis, « 187 » Limopsis multiradiata, L. insolita, Leda leptorhyncha, L. crebrecostata, Cardita maudensis, C. tasmanica (Fig. 96 B), Cuna radiata, Lepton crassum, Cardium pseudomagnum, Venus (Chione) multitaeniata, Solenocurtus legrandi, Lucina planatella (Fig. 96 C), Tellina porrecta and Myodora lamellata.
In Papua a Pecten (P. novaeguineae) has been recorded from the ? Lower Pliocene of Yule Island.
Kalimnan Bivalves.—
The Kalimnan beds contain the following restricted or upward ranging species:—Ostrea arenicola, O. manubriata (Fig. 96 D), Pecten antiaustralis (also in the Werrikooian Series), Perna percrassa, Mytilus hamiltonensis, Glycimeris halli, Limopsis beaumariensis (also Werrikooian) (Fig. 96 E), Leda crassa (also living), Trigonia howitti, Cardita solida, C. calva (also living), Erycina micans, Meretrix paucirugata, Sunetta gibberula, Venus (Chione) subroborata (Fig. 96 F), Donax depressa, Corbula scaphoides (also living), Barnea tiara, Lucina affinis, Tellina albinelloides and Myodora corrugata.
Werrikooian Bivalves.—
The next stage, the Werrikooian (Upper Pliocene), contains a large percentage of living species, as Ostrea angasi, Placunanomia ione (ranging down into Janjukian), Glycimeris radians, Leda crassa (also a common Kalimnan fossil), various species of Venus (Chione), as V. strigosa and V. placida, and Barnea australasiae.
Pleistocene Bivalves.—
The bivalved shells of the Pleistocene are similar to those now found living round the Australian coast, as Pecten asperrimus, Mytilus latus, Leda crassa, Soletellina biradiata and Spisula parva.
Pleistocene shells of bivalved genera occur in the coastal hills of Papua, including the following:—Cultellus, Corbula, Mactra, Tellina, Venus (Chione), Dione, Dosinea, Leda and Arca.
The SCAPHOPODS (“digger foot”) or the “Elephant-tusk shells” are adapted, by their well-developed foot, to burrow into the mud and sand.

Devonian Scaphopods.—
This group of mollusca makes its first appearance in Australasian sediments in the Middle Devonian (Murrumbidgee beds) of New South Wales, represented by Dentalium tenuissimum.
Jurassic Scaphopods.—
In the Jurassic strata of the Mataura Series of New Zealand, Dentalium huttoni (Fig. 97 A) occurs at the Kowhai River and Wilberforce.
Cretaceous Scaphopods.—
Dentalium wollumbillensis occurs in the drab and dark-coloured limestones of the Lower Cretaceous of the Lake Eyre Basin in S. Australia, and the same species is also found in the Lower Cretaceous (Rolling Downs Formation) of Wollumbilla, Queensland.
Cainozoic Scaphopods.—
The Cainozoic beds both of New Zealand and southern Australia yield many species of Dentalium, the commonest and most widely distributed being the longitudinally ribbed D. mantelli (Fig. 97 B), which ranges from the Balcombian to the Werrikooian stages in Australia, and is also typical of the Oamaru Series in New Zealand, where it is accompanied by the ponderous species, D. giganteum, which attained a length of over six inches. Another form common in our Cainozoics is the smooth-shelled D. subfissura; this also has a wide range, namely Balcombian to Kalimnan.
Palaeozoic Chitons.—
The POLYPLACOPHORA or Chitons (“Mail-shells”), first appeared in the Ordovician. In Australia « 190 » Chelodes calceoloides (Fig. 97 C) is found in the Silurian of Derrengullen Creek, Yass, New South Wales; and another species of the genus is found in beds of the same age at Lilydale, Victoria. Between that period and the Cainozoic or Tertiary there is a gap in their history in Australia.
Cainozoic Chitons.—
Ischnochiton granulosus (Fig. 97 D) is a Balcombian species of the modern type of “mail-shell,” occurring not infrequently in the clays of Balcombe’s Bay, Port Phillip, Victoria. Cryptoplax pritchardi (Fig. 97 E) is a curious form belonging to the attenuated, worm-like group of the Cryptoplacidae, until lately unknown in the fossil state; it is found in the Kalimnan Series near Hamilton, Victoria. Several other genera of the chitons are found fossil in the Australian Cainozoics which still live on our coasts, as Lorica, Plaxiphora and Chiton. The first-named genus is represented fossil by Lorica duniana from the Turritella bed (Janjukian) of Table Cape, Tasmania.
Characters of Gasteropoda.—
The GASTEROPODA (“belly-foot”) or univalve shells possess a muscular foot placed beneath the stomach and viscera. In the Heteropoda this foot is modified as a vertical fin, and in the Pteropoda as two wing-like swimming membranes close to the head. The mantle lobe is elevated along the back like a hood, and its surfaces and edges secrete the shell which contains the animal. The shell is typically a cone (example, Patella or Limpet) which is often « 191 » spirally coiled either in a plane (ex. Planorbis), conically turbinoid (ex. Trochus), or turreted (ex. Turritella). The body and shell are attached by muscles, the spiral forms being attached to the columella or axial pillar, and the bowl-shaped forms to the inner surface of the shell.
Gasteropod shells are normally right-handed (dextral), but a few genera as Clausilia, Bulinus and Physa, are left-handed (sinistral). The height or length of the shell is measured from the apex to the lower margin of the mouth. In coiled shells we may regard them as a more or less elongated cone wound round a central pillar, the columella, or around a central tube. A turn or coil of the shell is a whorl, and together, with the exception of the last, form the spire. The line between two adjacent whorls is the suture. When the columella is solid the shell is said to be imperforate, and when a central tube is left by the imperfect fusion of the whorls, it is perforate. The opening of the tubular columella is termed the umbilicus, and this is sometimes contracted by the encroachment of shell matter termed the callus. The aperture is entire when the rim is uninterrupted; and channelled when there is a basal notch, where the siphon which conducts water to the gills is lodged.
As a rule the large heavy gasteropods inhabit shallow water. The following living genera are characteristic of rocky shore-lines; Risella, Buccinum, Purpura and Patella. Genera typical of sandy shores are Nassa, Natica, Cypraea, Turritella and Scala.
Cambrian Gasteropods.—
From the Cambrian of South Australia Prof. Tate described some minute Gasteropods which he referred to the genera Stenotheca (S. rugosa, var. paupera), Ophileta (O. subangulata) (Fig. 98 A), and Platyceras (P. etheridgei). In these beds at Curramulka the following Pteropods were found by the same authority, viz., Salterella planoconvexa, Hyolithes communis (Fig. 98 C) and H. conularioides.
The Cambrian Limestone of the Kimberley District, W. Australia, contains the characteristic Pteropod Salterella hardmani (Fig. 98 B). The shell is a conical tube, straight or slightly curved, and measuring scarcely an inch in length.
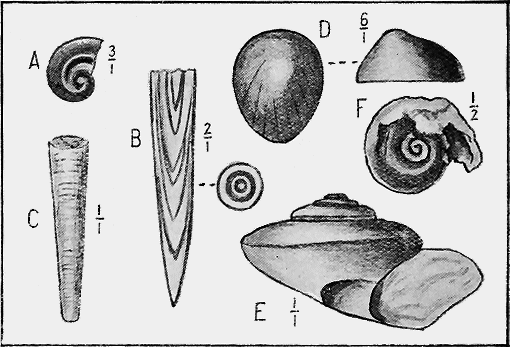
The Upper Cambrian of the Mersey River District in Tasmania has afforded some doubtful examples of the genus Ophileta.
In the Upper Cambrian Limestones of the Dolodrook Valley, near Mt. Wellington, Victoria, a minute limpet shaped Gasteropod occurs, named Scenella tenuistriata (Fig. 98 D).
Ordovician Gasteropods.—
Ordovician limestones with fossil shells occur in the Leigh’s Creek District in South Australia, and also at Tempe Downs and Petermann and Laurie’s Creeks, W. of Alice Springs. The euomphaloid shell Ophileta gilesi was described from Laurie’s Creek, and Eunema larapinta from the Tempe Downs. A pleurotomarid, Raphistoma browni (Fig. 98) occurs near Leigh’s Creek, and at Laurie’s and Petermann Creeks. A Pteropod, Hyolithes leptus, has been described from the Lower Ordovician of Coole Barghurk Creek, near Meredith, Victoria.
Silurian Gasteropods.—
The Silurian Gasteropods are fairly well represented, especially in the upper stage, and are widely distributed throughout the Australian fossiliferous localities. Moreover, some of the species are identical with those found as far off as North America and Europe. In Victoria the shales and sandstones of the lower stage (Melbournian) contain the genera Bellerophon, Cyrtolites and Loxonema. The Pteropoda include Tentaculites, Coleolus, Hyolithes and Conularia (C. sowerbii (Fig. 99 F), a species also found in Great Britain). The Victorian limestones and mudstones of the upper stage (Yeringian) « 194 » are somewhat rich in Gasteropods, such genera occurring as Pleurotomaria, Phanerotrema (with cancellated shell and large slit-band), Murchisonia, Gyrodoma, Bellerophon, Trematonotus (a spiral shell with a large trumpet-shaped mouth and a dorsal row of perforations in place of a slit-band), Euomphalus, Cyclonema, Trochus (Scalaetrochus), Niso (Vetotuba), Loxonema, Platyceras and Capulus. The section Pteropoda contains Tentaculites, Hyolithes and Conularia.

In the Silurian of New South Wales the chief Gasteropod genera are Bellerophon (B. jukesi), Euomphalus, Omphalotrochus, and Conularia (C. sowerbii.).
In Tasmania are found Raphistoma, Murchisonia, Bellerophon, Helicotoma, Trochonema and Tentaculites.
Devonian Gasteropods.—
The derived boulders of the White Cliffs opal field have been referred to the Devonian system, but of this there is some doubt, as the Gasteropods noted from these boulders closely resemble those of the Silurian fauna: they are Murchisonia Euomphalus (E. culleni), and Loxonema. The genus Murchisonia has also been recorded from the Baton River, New Zealand (Wangepeka Series) by MacKay.
The Middle Devonian Gasteropod fauna in Victoria, as found in the Buchan and Bindi Limestones, comprises Murchisonia, Trochus, and Platyceras.

In New South Wales the best known genera are Pleurotomaria, Murchisonia, Bellerophon, Euomphalus and Loxonema. The two latter genera have also been obtained at Barker Gorge, Western Australia.
Carboniferous Gasteropods.—
Carboniferous Gasteropoda have been found in New South Wales, belonging to the genera Gosseletina (G. australis) (Fig. 100 A) and Yvania (Y. konincki) (Fig. 100 B), both of which have their countertypes in the Carboniferous of Belgium. Y. konincki is also found in the Carbopermian (Gympie beds) of Rockhampton, Queensland, while Y. levellii is found in the Carbopermian of Western Australia.
Carbopermian Gasteropods.—
The Carbopermian gasteropods of New South Wales are Pleurotomaria (Mourlonia), Keeneia platyschismoides, Murchisonia, Euomphalus, Platyschisma (P. oculum) (Fig. 100 E), Loxonema and Macrocheilus. Examples of the genus Conularia are sometimes found, probably attaining a length, when complete, of 40 centimetres.
In Tasmania we find Conularia tasmanica, a handsome Pteropod, also of large dimensions. Platyschisma, Pleurotomaria (Mourlonia), Bellerophon and Porcellia are amongst the Carbopermian Gasteropods of Queensland.
In Western Australia Pleurotomaria (Mourlonia), Bellerophon, Euomphalus, Euphemus, Platyceras, and Loxonema occur in the Carbopermian.
Jurassic Gasteropods.—
Jurassic gasteropods are found sparingly in the « 197 » limestone of the Geraldton District and other localities in Western Australia. The more important of these are Pleurotomaria (P. greenoughiensis), Turbo (T. australis) (Fig. 101 A) and Rissoina (R. australis) (Fig. 101 B).
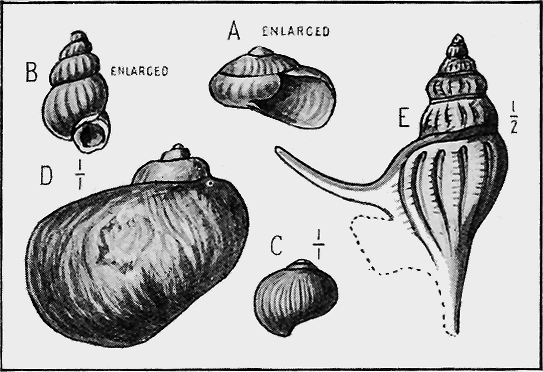
Cretaceous Gasteropods.—
The Queensland gasteropod fauna comprises Cinulia a typical Cretaceous genus, Actaeon and Natica. These occur in the Lower Cretaceous or Rolling Downs Formation. Cinulia is also found in South Australia at Lake Eyre with Natica (N. ornatissima) (Fig. 101 C). Pseudamaura variabilis (Fig. 101 D) is found in New South Wales, Queensland and South Australia; whilst Anchura wilkinsoni occurs in Queensland and South Australia.
In New Zealand the Waipara Greensands (Cretaceous) contain a species of Rostellaria (R. waiparensis) (Fig. 101 E).
Cainozoic Gasteropods.—
Cainozoic Gasteropods are exceedingly abundant in beds of that system in Australasia. The Cainozoic marine fauna in Australia is practically restricted to the States of Victoria, South Australia, and Tasmania; whilst New Zealand has many species in common with Australia.
Genera.—
The commonest genera of the marine Cainozoic or Tertiary deposits are:—Haliotis, Fissurellidea, Emarginula, Subemarginula, Astralium, Liotia, Gibbula, Eulima, Niso, Odostomia, Scala, Solarium, Crepidula, Calyptraea, Natica, Rissoa, Turritella, Siliquaria, Cerithium, Newtoniella, Tylospira, Cypraea, Trivia, Morio, Semicassis, Lotorium, Murex, Typhis, Columbella, Phos, Nassa, Siphonalia, Euthria (Dennantia), Fusus, Columbarium, Fasciolaria, Latirus, Marginella, Mitra, Volutilithes, Voluta, Harpa, Ancilla, Cancellaria, Terebra, Pleurotoma, Drillia, Conus, Bullinella and Vaginella.
Persistent Species.—
Amongst the Cainozoic Gasteropoda of southern Australia which have a persistent range through Balcombian to Kalimnan times, we find:—Niso psila, Crepidula unguiformis (also Werrikooian and Recent), Natica perspectiva, N. hamiltonensis, Turritella murrayana, Cerithium apheles, Cypraea leptorhyncha, Lotorium gibbum, Volutilithes antiscalaris « 199 » (also in Werrikooian), Marginella propinqua, Ancilla pseudaustralis, Conus ligatus and Bullinella exigua.
Balcombian Gasteropods.—
Species restricted to the Balcombian stage include Scala dolicho, Seguenzia radialis, Dissocheilus eburneus, Trivia erugata, Cypraea ampullacea (Fig. 102 A), C. gastroplax, Colubraria leptoskeles, Murex didymus (Fig. 102 B), Eburnopsis aulacoessa (Fig. 102 C), Fasciolaria concinna, Mitra uniplica, Harpa abbreviata, Ancilla lanceolata, Cancellaria calvulata (Fig. 102 D), Buchozia oblongula, Pleurotoma optata, Terebra leptospira and Vaginella eligmostoma (Fig. 102 E), (also found at Gellibrand River).
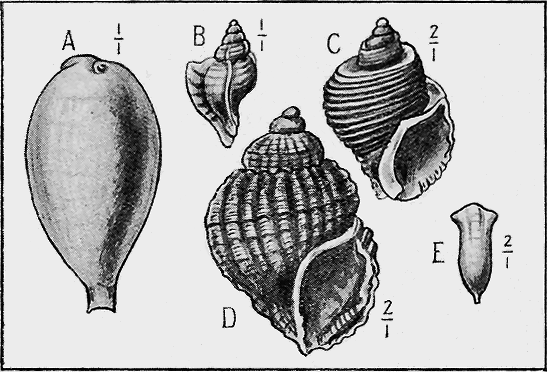
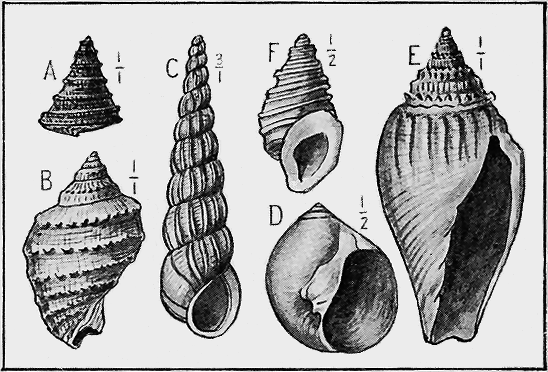
Janjukian Gasteropods.—
Species of Gasteropods restricted to the Janjukian stage include:—Pleurotomaria tertiaria, Haliotis mooraboolensis, Liotia lamellosa, Thalotia alternata, Eutrochus fontinalis (Fig. 103 A), Astralium hudsonianum, Turbo atkinsoni, Odostomia polita, Scala lampra (Fig. 103 C), Natica gibbosa (Fig. 103 D) (also found in the Pareora Series of the Oamaru system and in the Wanganui beds of New Zealand), Calyptraea subtabulata, Turritella aldingae, Cerithiopsis mulderi, Cerithium flemingtonense, Cypraea platyrhyncha, C. consobrina, Morio wilsoni (Fig. 103 B), Lotorium abbotti, Murex otwayensis, Eburnopsis « 201 » tesselatus, Tudicla costata, Latirus semiundulatus, Fusus meredithae, Columbarium spiniferum, Voluta pueblensis, V. heptagonalis, V. macroptera (also recorded from Hall’s Sound, Papua) (Fig. 103 E), Volutilithes anticingulatus (also from Papua), Harpa clathrata, Bela woodsi, Bathytoma paracantha and Volvulella inflatior.
Dolium costatum, allied to the “Fig-Shell” has been noted from the Cainozoic clays (? Lower Pliocene), Yule Island, Papua.

Kalimnan Gasteropods.—
Species of Gasteropods restricted to the Kalimnan Stage, or only passing upwards include:—Bankivia howitti (Fig. 104 A), Liopyrga quadricingulata, Calyptraea corrugata, Natica subvarians, Turritella « 202 » pagodula, Eglisia triplicata (Fig. 104 B), Tylospira clathrata, Cypraea jonesiana, Lotorium ovoideum, Sistrum subreticulatum, Voluta masoni (Fig. 104 C), Ancilla papillata (Fig. 104 D), Cancellaria wannonensis, Drillia wanganuiensis (also in the Petane Series of New Zealand), Terebra catenifera, T. geniculata (Fig. 104 E) and Ringicula tatei.
New Zealand Cainozoic Gasteropods.—
Characteristic Gasteropoda of the Oamaru Series in New Zealand are Pleurotomaria tertiaria (also in the Australian Janjukian), Scala lyrata, Natica darwinii, Turritella cavershamensis, Ancilla hebera (also in the Australian Balcombian and Janjukian) and Pleurotoma hamiltoni. Gasteropods of the Awatere Series in New Zealand are Natica ovata, Struthiolaria sulcata (Fig. 103 F), and Scaphella corrugata (found also in the Oamaru Series). The Putiki beds of the Petane Series in New Zealand contain Trophon expansus, Pisania drewi and Pleurotoma wanganuiensis.
Werrikooian Gasteropods.—
The marine gasteropods of the Werrikooian of southern Australia, as found at Limestone Creek, Glenelg River, Western Victoria, and the Moorabool Viaduct near Geelong, are nearly all living at the present time, with the exception of a few older Cainozoic species. Amongst these latter are Conus ralphi, Pleurotoma murndaliana, Volutilithes antiscalaris and Columbarium craspedotum.
Pleistocene Gasteropoda.—
The Pleistocene land mollusca, and especially the gasteropods of Australia, present some striking « 203 » points of interest, for whilst most of the species are still living, some appear to be extinct. The travertine deposits of Geilston, near Hobart, Tasmania contain Helix geilstonensis and H. stanleyana, the latter still living. The calcareous Helix sandstone of the islands in Bass Strait are largely composed of shells of that genus and generally represent consolidated sand-dunes which have undergone a certain amount of elevation. One of the prevalent species is Helix simsoniana (Fig. 104 F), a handsome keeled form, somewhat related to the living H. launcestonensis. It is found in some abundance in the Kent’s Group and in the adjacent islands.
The large ovoid land-shells, Panda atomata, although still existing, are found associated with extinct marsupials, as Thylacoleo, in the stalagmitic floor of the Buchan Caves, Gippsland.
The Diprotodon-breccias of Queensland have afforded several species of Helix and other land-shells, as well as the brackish-water genus Melania. The Raised Beaches of Queensland, New South Wales, Victoria, and Tasmania all contain species of land and freshwater shells identical with those now found living in the same localities.
The Raised Beaches of New Zealand contain numerous marine shells all having living representatives. Some of these elevated beaches occur as high as 150 feet above sea-level at Taranaki, and at 200 feet near Cape Palliser in Cook Strait.
Many species of Pleistocene Mollusca identical with those now living in Torres Strait, the China Sea and the Philippine Islands are found in Papua. They « 204 » occur in the greenish sandy clay of the hills near the present coast line and comprise the following genera of Gasteropods:—Ranella, Nassa, Mitra, Oliva, Terebra, Conus, Strombus, Bulla and Atys.
Characters of Cephalopoda.—
The highest class of the mollusca is the CEPHALOPODA (“head-feet”). In these shell-fish the extremity of the body or foot is modified, and furnished with eyes, a funnel and tentacles. It has also strong horny beaks or jaws which make it a formidable enemy to the surrounding life in the sea. In the chambered forms of this group the animal partitions off its shell at regular intervals, like the Pearly Nautilus and the Ammonite, inhabiting only the last chamber cavity, but still communicating with the earlier series by a continuous spiral tube (siphuncle). In some forms like the living squid and the extinct Belemnite, the shell is internal and either spoon-shaped, or dart-shaped, that is, subcylindrical and pointed.
Characters of Cephalopod Shells.—Nautiloidea.—
In geological times the nautiloid forms were the first to appear (in the Ordovician), and they were either straight shells, as Orthoceras, or only slightly curved, as Cyrtoceras. Later on they became more closely coiled, and as they were thus less likely to be damaged, they gradually replaced the straight forms.
The Ammonites have the siphuncle close to the outside of the shell, whilst in the Nautilus it is more or less median. The sutures or edges of the septa in Nautilus and its allies are curved or wavy, but not so sharply flexed or foliaceous as in Ammonites. The « 205 » Nautiloidea range from the Ordovician and are still found living.
Ammonoidea.—
The Ammonoidea appear in Devonian times and die out in the Cretaceous. They were very abundant in Jurassic times, especially in Europe.
Belemnoidea.—
The Belemnoidea, ranging from the Trias to Eocene, comprise the extinct Belemnites, the interesting genus Spirulirostra of Miocene times, and the living Spirula.
Sepioidea.—
The Sepioidea or true Cuttle-fishes (“pen-and-ink fish”) range from the Trias to the present day.
Octopoda.—
The Octopoda, with Octopus and Argonauta (the paper “Nautilus”) are present-day modifications. The male of the latter is without a shell, the female only being provided with a delicate boat-shaped shell secreted by the mantle and the two fin-like expansions of the dorsal arms.
Ordovician Cephalopods.—
The Ordovician cephalopods of Australasia are not numerous, and are, so far as known, practically restricted to the limestones of the Larapintine series at Laurie’s Creek and Tempe Downs, in Central South Australia. Amongst them may be mentioned Endoceras warburtoni (Fig. 105 A), (a straight form in which the siphuncle is partially filled with organic deposits); Orthoceras gossei; O. ibiciforme; Trochoceras reticostatum (a coiled form); and Actinoceras tatei (a genus characterised by swollen siphuncular beads between the septa).

Silurian Cephalopods.—
Silurian cephalopods are more generally distributed, and in Victoria constitute an important factor in the molluscan fauna of that system. Orthoceras and Cycloceras are the best known genera, represented by Orthoceras capillosum, found near Kilmore, Victoria; O. lineare (Fig. 105 B), from the Upper Yarra; Cycloceras bullatum, from the Melbournian of Collingwood and Whittlesea; and C. ibex (Fig. 105 C) from South Yarra and Flemington, in both Melbournian shale and sandstone. The latter species occurs also at Rock Flat Greek, New South Wales. Other Victorian species are Kionoceras striatopunctatum, a well-known European fossil with a reticulated « 207 » and beaded ornament, found near Warburton and at McMahon’s Creek, Upper Yarra.
Orthoceras is also recorded from Tasmania and from the Wangapeka beds of Baton River, New Zealand. Cyclolituites, a partially coiled nautilian is recorded from Bowning, near Yass, New South Wales; whilst the closely related Lituites is noted from the Silurian of Tasmania.
Devonian Cephalopods.—
The only genus of cephalopoda at present recorded from the Devonian of Victoria is Phragmoceras (P. subtrigonum) (Fig. 105 D), which occurs in the Middle Devonian Limestone of Buchan, E. Gippsland. From beds of similar age in New South Wales Orthoceras, Cyrtoceras and Goniatites have been noted; whilst the latter genus also occurs near Kimberley, Western Australia. In Queensland Gyroceras philpi is a characteristic shell, found in the Fanning and Reid Gap Limestones of the Burdekin Formation (Middle Devonian).
Carbopermian Cephalopods.—
The Carbopermian rocks of New South Wales have yielded Orthoceras striatum, Cameroceras, Nautilus and Agathiceras micromphalum (Fig. 105 F). In Queensland the Gympie Formation contains Orthoceras, Gyroceras, Nautilus, Agathiceras micromphalum and A. planorbiforme. In Western Australia the Kimberley rocks contain Orthoceras, Glyphioceras sphaericum and Agathiceras micromphalum; whilst the largest known Australian goniatite, Gastrioceras jacksoni (Fig. 105 E) is found in the Irwin River District. Actinoceras hardmani is an interesting « 208 » fossil from the Carbopermian of Lennard River, N.W. Australia. In Tasmania the genera Orthoceras and Goniatites have been recorded from beds of similar age.
Triassic Cephalopods.—
For Triassic cephalopoda we look to New Zealand, where, in the Mount Potts Spiriferina Beds of the Kaihiku Series a species of Orthoceras has been recorded. The Wairoa Series next in succession contains Orthoceras and an Ammonite.

Jurassic Cephalopods.—
The Jurassic of Western Australia yields a rich cephalopod fauna, from which may be selected as « 209 » typical examples the Nautilus, N. perornatus and the following Ammonites: Dorsetensia clarkei; Normanites australis; and Perisphinctes championensis (Fig. 106 A). These all occur in the Greenough River District, and at several other Jurassic localities in Western Australia.
The Jurassic system of New Zealand (Putataka Series) contains Ammonites aucklandicus and Belemnites aucklandicus, both from the upper marine horizon of that series.
Upper Jurassic Ammonites belonging to the genera Macrocephalites (M. cf. calloviensis) and Erymnoceras (E. cf. coronatum) have been recorded from Papua.
Lower Cretaceous Cephalopods.—
Remains of Cephalopoda are fairly abundant in the Lower Cretaceous of Australasia. From amongst them may be selected the following—Nautilus hendersoni (Fig. 106 B) (Q.); Haploceras daintreei (Fig. 106 C) (Q. and N.S.W.); Desmoceras flindersi (Q. and N.S.W.); Schloenbachia inflatus (Q.); Scaphites cruciformis (N. Terr.); Ancyloceras flindersi (Q. and N.S.W.); Crioceras australe (Fig. 106 D) (Q. and S.A.); Belemites australis (Q.); B. oxys (Q., N.S.W., and S.A.); B. sellheimi (Q. and S.A.); B. diptycha, = canhami, Tate, (Q., N.S.W., and S.A.); and B. eremos (Centr. S.A.).
Upper Cretaceous Cephalopods.—
In the Upper Cretaceous (Desert Sandstone) of Queensland there occurs a Belemnite somewhat resembling Belemnites diptycha, but with a very pointed apex.
Cretaceous Cephalopods, New Zealand.—
In New Zealand the Amuri System (Cretaceous) contains fossils which have been referred to the genera Ammonites, Baculites, Hamites, Ancyloceras and Belemnites, but probably these determinations require some further revision. A species of Belemnite has also been noted from probable Cretaceous beds in Papua.
The Cainozoic System in Victoria contains a true Nautilus, N. geelongensis; and Aturia australis (Fig. 106 E), a nautiloid shell having zig-zag suture lines and septal necks enclosing the siphuncle. A. australis is also found in the Oamaru Series of New Zealand; in Victoria it has an extensive vertical range, from Balcombian to Kalimnan (Oligocene to Lower Pliocene). Species of Nautilus are also found in the Janjukian of the Murray River Cliffs; where, in some cases the shell has been infilled with clear gypsum or selenite, through which can be seen the tubular siphuncle in its original position. Spirulirostra curta (Fig. 106 F) is an interesting cuttle-bone of rare occurrence. The genus is represented by two other species only, occurring in the Miocene of Italy and Germany. In Victoria it is occasionally found in the Janjukian marly limestone at Bird Rock near Torquay.
COMMON OR CHARACTERISTIC FOSSILS OF THE FOREGOING CHAPTER.
PELECYPODA.
Ambonychia, macroptera, Tate. Cambrian: S. Australia. (?) Modiolopsis knowsleyensis, Chapm. L. Ordovician: Victoria.
Orthonota australis, Chapm. Silurian (Melbournian): Victoria.
Grammysia cuneiformis, Eth. fil. Silurian (Melbournian): Victoria.
Leptodomus maccoyianus, Chapm. Silurian (Melbournian): Victoria.
Edmondia perobliqua, Chapm. Silurian (Melbournian): Victoria.
Cardiola cornucopiae, Goldfuss sp. Silurian (Melbournian): Victoria.
Panenka gippslandica, McCoy sp. Silurian (Tanjilian): Victoria.
Ctenodonta portlocki, Chapm. Silurian: Victoria.
Nuculites maccoyianus, Chapm. Silurian: Victoria.
Nucula melbournensis, Chapm. Silurian (Melb.): Victoria.
Palaeoneilo victoriae, Chapm. Silurian (Melb.): Victoria.
Pterinea lineata, Goldfuss. Silurian (Yeringian): Victoria.
Lunulicardium antistriatum, Chapm. Silurian (Tanj.): Victoria.
Conocardium costatum, Cressw. sp. Silurian: Victoria.
Conocardium davidis, Dun. Silurian: New South Wales.
Actinopteria boydi, Conrad sp. Silurian (Yer.): Victoria.
Aviculopecten spryi, Chapm. Silurian (Melb.): Victoria.
Modiolopsis complanata, Sowerby sp. Silurian (Melb.): Victoria.
Goniophora australis, Chapm. Silurian (Yer.): Victoria.
Cypricardinia contexta, Barrande. Silurian (Yer.): Victoria.
Paracyclas siluricus, Chapm. Silurian (Melb.): Victoria.
Actinopteria australis, Dun. Devonian: New South Wales.
Lyriopecten gracilis, Dun. Devonian: New South Wales.
Leptodesma inflatum, Dun. Devonian: New South Wales.
Stutchburia farleyensis, Eth. fil. Carbopermian: New South Wales.
Edmondia nobilissima, de Koninck. Carbopermian: New South Wales.
Deltopecten limaeformis, Morris sp. Carbopermian: New South Wales, Queensland and Tasmania.
Aviculopecten squamuliferus, Morris sp. Carbopermian: New South Wales and Tasmania.
Aviculopecten tenuicollis, Dana sp. Carbopermian: New South Wales and W. Australia.
Chaenomya etheridgei, de Koninck sp. Carbopermian: New South Wales and Queensland.
Maeonia elongata, Dana. Carbopermian: New South Wales.
Pachydomus globosus, J. de C. Sow. sp. Carbopermian: New South Wales, Tasmania and Queensland.
Eurydesma cordatum, Morris. Carbopermian: New South Wales and Queensland.
Unio dunstani, Eth. fil. Trias: New South Wales.
Unionella carnei, Eth. fil. Trias: New South Wales.
Corbicula burrumensis, Eth. fil. Trias: Queensland.
Daonella lommeli, Wissm. sp. Trias: New Zealand.
Mytilus problematicus, Zittel. Trias: New Zealand.
Monotis salinaria, Zittel. Trias: New Zealand.
Cucullaea semistriata, Moore. Jurassic: W. Australia.
Trigonia moorei, Lycett. Jurassic: W. Australia.
Ctenostreon pectiniforme, Schlotheim sp. Jurassic: W. Australia.
Astarte cliftoni, Moore. Jurassic: W. Australia.
Unio dacombei, McCoy. Jurassic: Victoria.
Unio eyrensis, Tate. Jurassic: S. Australia.
Nucula truncata, Moore. Lower Cretaceous: Queensland and S. Australia.
Maccoyella reflecta, Moore sp. L. Cretaceous: New South Wales, Queensland (also U. Cretaceous), and S. Australia.
Maccoyella barkleyi, Moore sp. L. Cretaceous: New South Wales, Queensland and S. Australia.
Fissilunula clarkei, Moore sp. L. Cretaceous: New South Wales, Queensland, and S. Australia; also Up. Cret. in Queensland and South Australia.
Inoceramus carsoni, McCoy. Lower Cretaceous: Queensland.
Trigonia cinctuta, Eth. fil. Lower Cretaceous: S. Australia.
Mytilus rugocostatus, Moore. Lower Cretaceous: Queensland and S. Australia.
Cyrenopsis opallites, Eth. fil. Upper Cretaceous: New South Wales.
Conchothyra parasitica, Hutton. Cretaceous: New Zealand.
Dimya dissimilis, Tate. Cainozoic (Balc.-Kal.): Victoria and South Australia.
Spondylus pseudoradula, McCoy. Cainozoic (Balc.-Kal.): Victoria and South Australia.
Pecten polymorphoides, Zittel. Cainozoic (Balc.-Kal.): Victoria and South Australia; also New Zealand.
Cucullaea corioensis, McCoy. Cainozoic (Balc.-Kal.): Victoria and South Australia.
Leda vagans, Tate. Cainozoic (Balc.-Kal.): Victoria and South Australia.
Corbula ephamilla, Tate. Cainozoic (Balc.-Kal.): Victoria and South Australia.
Modiola praerupta, Pritchard. Cainozoic (Balc.): Victoria.
Pecten praecursor, Chapm. Cainozoic (Janjukian): Victoria.
Modiola pueblensis, Pritchard. Cainozoic (Janjukian): Victoria.
Limopsis insolita, Sow. sp. Cainozoic (Janjukian): Victoria and S. Australia. Also Oamaru Ser., N.Z.
Cardita tasmanica, Tate. Cainozoic (Janj.): Tasmania.
Lucina planatella, Tate. Cainozoic (Janj.): Victoria and Tasmania.
Pecten novaeguineae, T. Woods. Cainozoic (?Lower Pliocene), Yule Island, Papua.
Ostrea manubriata, Tate. Cainozoic (Kal.): Victoria.
Glycimeris halli, Pritch. Cainozoic (Kal.): Victoria.
Limopsis beaumariensis, Chapm. Cainozoic (Kalimnan and Werrikooian): Victoria.
Trigonia howitti, McCoy. Cainozoic (Kal.): Victoria.
Meretrix paucirugata, Tate sp. Cainozoic (Kal.): Victoria.
Venus (Chione) subroborata, Tate, sp. Cainozoic (Kal.): Victoria and South Australia.
SCAPHOPODA.
Dentalium tenuissimum, de Koninck. Mid. Devonian: New South Wales.
Dentalium huttoni, Bather. Jurassic: New Zealand.
Dentalium wollumbillensis, Eth. fil. L. Cretaceous: Queensland.
Dentalium, mantelli, Zittel. Cainozoic: Victoria, S. Australia and New Zealand.
POLYPLACOPHORA.
Chelodes calceoloides, Eth. fil. Silurian: New South Wales.
Ischnochiton granulosus, Ashby and Torr sp. Cainozoic (Balc.): Victoria.
Lorica duniana, Hull. Cainozoic (Janjukian): Tasmania.
Cryptoplax pritchardi, Hall. Cainozoic (Kal.): Victoria.
GASTEROPODA.
Ophileta subangulata, Tate. Cambrian: S. Australia.
Platyceras etheridgei, Tate. Cambrian: S. Australia.
Salterella planoconvexa, Tate. Cambrian: S. Australia.
Salterella hardmani, Foord. Cambrian: W. Australia.
Hyolithes communis, Billings. Cambrian: S. Australia.
Scenella tenuistriata, Chapm. Cambrian (Upper): Victoria.
Ophileta gilesi, Tate. Ordovician: S. Australia.
Raphistoma browni, Tate. Ordovician: S. Australia.
Hyolithes leptus, Chapm. Lower Ordovician: Victoria.
Helicotoma johnstoni, Eth. fil. Ordovician: Tasmania.
Coleolus (?) aciculum, J. Hall. Silurian (Melb.): Victoria.
Hyolithes spryi, Chapm. Silurian (Melb.): Victoria.
Conularia ornatissima, Chapm. Silurian (Melb.): Victoria.
Phanerotrema australis, Eth. fil. Silurian (Yer.): Victoria.
Gyrodoma etheridgei, Cressw. sp. Silurian (Yer.): Victoria.
Trematonotus pritchardi, Cressw. Silurian (Yer.): Victoria.
Bellerophon cresswelli, Eth. fil. sp. Silurian (Yer.) Victoria.
Euomphalus northi, Eth. fil. sp. Silurian (Yer.): Victoria.
Cyclonema australis, Eth. fil. Silurian (Yer.): Victoria.
Trochonema montgomerii, Eth. fil. sp. Silurian: Tasmania.
Bellerophon jukesii, de Koninck. Silurian: New South Wales.
Conularia sowerbii, Defrance. Silurian: Victoria and New South Wales.
Euomphalus culleni, Dun. Devonian: New South Wales.
Gosseletina australis, Eth. fil. Carboniferous: New South Wales.
Yvania konincki, Eth. fil. Carboniferous: New South Wales; and Carbopermian: Queensland.
Bellerophon costatus, Sow. Carbopermian: W. Australia.
Mourlonia humilis, de Koninck. Carbopermian: West Australia and New South Wales.
Pleurotomaria (Ptychomphalina) morrisiana, McCoy. Carbopermian: New South Wales.
Keeneia platyschismoides, Eth. fil. Carbopermian (Lower Marine): New South Wales.
Platyschisma oculum, Sow. sp. Carbopermian: New South Wales and Queensland.
Macrocheilus filosus, Sow. Carbopermian: New South Wales.
Loxonema babbindonensis, Eth. fil. Carbopermian: New South Wales.
Conularia tenuistriata, McCoy. Carbopermian: New South Wales and Queensland.
Conularia tasmanica, Carbopermian: Tasmania.
Murchisonia carinata, Etheridge. Carbopermian: Queensland.
Pleurotomaria greenoughiensis, Eth. fil. Jurassic: W. Australia.
Turbo australis, Moore. Jurassic: W. Australia.
Rissoina australis, Moore. Jurassic: W. Australia.
Cinulia hochstetteri, Moore. Cretaceous: Queensland and S. Australia.
Natica ornatissima, Moore. Cretaceous: S. Australia.
Pseudamaura variabilis, Moore sp. Cretaceous: New South Wales, Queensland and S. Australia.
Anchura wilkinsoni, Eth. fil. Cretaceous: Queensland and S. Australia.
Rostellaria waiparensis, Hector. Cretaceous: New Zealand.
Niso psila, T. Woods. Cainozoic (Balc.-Kal.): Victoria and S. Australia.
Crepidula unguiformis, Lam. Cainozoic (Balc.-Recent): Victoria and Tasmania.
Natica hamiltonensis, Tate. Cainozoic (Balc.-Recent): Victoria and South Australia.
Turritella murrayana, Tate. Cainozoic (Balc.-Kal.): Victoria, S. Australia and Tasmania.
Cerithium apheles, T. Woods. Cainozoic (Balc.-Kal.): Victoria.
Volutilithes antiscalaris, McCoy sp. Cainozoic (Balc.-Werrikooian): Victoria.
Ancilla pseudaustralis, Tate sp. Cainozoic (Balc.-Kal.): Victoria, S. Australia and Tasmania.
Cypraea ampullacea, Tate. Cainozoic (Balc.): Victoria.
Murex didyma, Tate. Cainozoic (Balc.): Victoria.
Eburnopsis aulacoessa, Tate. Cainozoic (Balc.): Victoria.
Cancellaria calvulata, Tate. Cainozoic (Balc.): Victoria.
Vaginella eligmostoma, Tate. Cainozoic (Balc.): Victoria.
Eutrochus fontinalis, Pritchard. Cainozoic (Janjukian): Victoria.
Turbo atkinsoni, Pritchard. Cainozoic (Janjukian): Tasmania and Victoria.
Scala lampra, Tate sp. Cainozoic (Janjukian): S. Australia.
Natica gibbosa, Hutton. Cainozoic (Janjukian): Victoria. Also Oamaru and Wanganui Series: New Zealand.
Morio wilsoni, Tate. Cainozoic (Janjukian): Victoria.
Voluta heptagonalis, Tate. Cainozoic (Janjukian): S. Australia.
Volutilithes anticingulatus, McCoy sp. Cainozoic (Janjukian): Victoria and Tasmania. Also Papua.
Bathytoma paracantha, T. Woods sp. Cainozoic (Janj.): Victoria and Tasmania. Also Papua.
Dolium costatum, Deshayes. Cainozoic. (? Lower Pliocene): Yule Island, Papua.
Bankivia howitti, Pritch. Cainozoic (Kal.): Victoria.
Eglisia triplicata, Tate sp. Cainozoic (Kal.): Victoria.
Voluta masoni, Tate. Cainozoic (Kal.): Victoria.
Ancilla papillata, Tate sp. Cainozoic (Kal.): Victoria.
Drillia wanganuiensis, Hutton. Cainozoic (Kal.): Victoria. Also Petane Series: New Zealand.
Terebra geniculata, Tate. Cainozoic (Kal.): Victoria.
Pleurotomaria tertiaria, McCoy. Cainozoic (Kal.): Victoria. Also Oamaru Series: New Zealand.
Scala lyrata, Zittel sp. Cainozoic (Oamaru): New Zealand.
Natica darwinii, Hutton. Cainozoic (Oamaru): New Zealand.
Turritella cavershamensis, Harris. Cainozoic (Oamaru): New Zealand.
Ancilla hebera, Hutton sp. Cainozoic (Oamaru): New Zealand. Also (Balc. and Janj.): Victoria, South Australia and Tasmania.
Pleurotoma hamiltoni, Hutton. Cainozoic (Oamaru): New Zealand.
Natica ovata, Hutton. Cainozoic (Awatere Series): New Zealand.
Struthiolaria sulcata, Hutton. Cainozoic (Awatere Series): New Zealand.
Trophon expansus, Hutton. Cainozoic (Petane Series): New Zealand.
Pisania drewi, Hutton. Cainozoic (Petane Series): New Zealand.
Bankivia fasciata, Menke. Cainozoic (Werrikooian-Recent): Victoria.
Astralium aureum, Jonas sp. Cainozoic (Werrikooian-Recent): Victoria.
Natica subinfundibulum, Tate. Cainozoic (Balc.-Werr.): Victoria and S. Australia.
Nassa pauperata, Lam. Cainozoic (Werr.-Rec.): Victoria.
Helix tasmaniensis, Sow. Cainozoic (Pleistocene): Tasmania.
Helix geilstonensis, Johnston. Cainozoic (Pleistocene): Tasmania.
Panda atomata, Gray sp. Cainozoic (Pleist.-Rec.): Victoria and New South Wales.
CEPHALOPODA.
Endoceras warburtoni, Eth. fil. Ordovician: S. Australia.
Orthoceras gossei, Eth. fil. Ordovician: S. Australia.
Orthoceras ibiciforme, Tate. Ordovician: S. Australia.
Trochoceras reticostatum, Tate. Ordovician: S. Australia.
Actinoceras tatei, Eth. fil. sp. Ordovician: S. Australia.
Orthoceras capillosum, Barrande. Silurian: Victoria.
Orthoceras lineare, Münster sp. Silurian (Yer.): Victoria.
Cycloceras bullatum, Sow. sp. Silurian (Melbournian): Victoria.
Cycloceras ibex, Sow. sp. Silurian (Melbournian): Victoria.
Kionoceras striatopunctatum, Münster sp. Silurian (Tanjilian): Victoria.
Phragmoceras subtrigonum, McCoy. Mid. Devonian: Victoria.
Gyroceras philpi, Eth. fil. Mid. Devonian: Queensland.
Orthoceras striatum, Sow. Carbopermian: New South Wales.
Agathiceras micromphalum, Morris sp. Carbopermian: New South Wales and W. Australia.
Gastrioceras jacksoni, Eth. fil. Carbopermian: W. Australia.
Actinoceras hardmani, Eth. fil. Carbopermian: N.W. Australia.
Nautilus perornatus, Crick. Jurassic: W. Australia.
Dorsetensia clarkei, Crick. Jurassic: W. Australia.
Normanites australis, Crick sp. Jurassic: W. Australia.
Perisphinctes championensis, Crick. Jurassic: W. Australia.
Ammonites aucklandicus, Hector. Jurassic: New Zealand.
Belemnites aucklandicus, Hector. Jurassic: New Zealand.
Nautilus hendersoni, Eth. fil. Lower Cretaceous: Queensland.
Haploceras daintreei, Etheridge sp. Lower Cretaceous: Queensland and New South Wales.
Ancyloceras flindersi, McCoy. Lower Cretaceous: Queensland and New South Wales.
Crioceras australe, Moore. Lower Cretaceous: Queensland and S. Australia.
Scaphites eruciformis, Eth. fil. Lower Cretaceous: Northern Territory.
Belemnites diptycha, McCoy. Lower Cretaceous: Queensland, New South Wales, and S. Australia.
Belemnites eremos, Tate. Lower Cretaceous: S. Australia.
Nautilus geelongensis, Foord. Cainozoic (Janjukian): Victoria.
Aturia australis, McCoy. Cainozoic (Balc.-Kal.): Victoria. Oamaru Series: New Zealand.
Spirulirostra curta, Tate. Cainozoic (Janjukian): Victoria.
LITERATURE.
MOLLUSCA.
Cambrian.—Foord, A. H. Geol. Mag., Dec. III. vol. VII. 1890, pp. 98, 99 (Pteropoda). Tate, R. Trans. R. Soc. S. Austr., vol. XV. 1892, pp. 183-185 (Pelec. and Gastr.), pp. 186, 187 (Pteropoda). Etheridge, R. jnr. Trans. R. Soc. S. Austr., vol. XXIX. 1905, p. 251 (Pteropoda). Chapman, F. Proc. R. Soc. Vict., vol. XXIII. pt. II. 1910, pp. 313, 314 (Gastr.).
Ordovician.—Etheridge, R. jnr. Parl. Papers, Leg. Assemb., S. Austr., No. 158, 1891, pp. 9, 10 (Gastr. and Ceph.). Tate, R. Rep. Horn. Sci. Exped., pt. 3, 1896, pp. 98-110. Chapman, F. Proc. R. Soc. Vic., vol. XV. pt. II. 1903, pp. 119, 120 (Hyolithes).
Silurian.—McCoy, F. Prod. Pal. Vic., Dec. VI. 1879, pp. 23-29. Etheridge, R. jnr. Rec. Austr. Mus., vol. I. No. 3, 1890, pp. 62-67 (Gastr.). Idem, ibid., vol. I. No. 7, 1891, pp. 126-130 (Pelec. and Gastr.). Cresswell, A. W. Proc. R. Soc. Vict., vol. V. 1893, pp. 41-44. Etheridge, R. jun. Rec. Austr. Mus., vol. III. No. 4, 1898, pp. 71-77 (Gastr.). Idem, Rec. Geol. Surv. New South Wales, vol. V. pt. 2, 1898, pp. 67-70 (Chelodes). De Koninck, L. G. Mem. Geo. Surv. New South Wales, Pal. No. 6, 1898, pp. 29-35. Etheridge, R. jnr. Prog. Rep. Geol. Surv. Vict., No. XI. 1899, pp. 34, 35 (Pelec.). Idem, Rec. Austr. Mus., vol. V. No. 2, 1904, pp. 75-77 (Ceph.). Chapman, F. Proc. R. Soc., Vict., vol. XVI. pt. 11. 1904, pp. 336-341 (Pteropoda). Idem, Mem. Nat. Mus. Melbourne, No. 2, 1908 (Pelecypoda).
Devonian.—McCoy, F. Prod. Pal., Vict., Dec. IV. 1876, pp. 18, 19 (Ceph.). Etheridge, R. jnr. Geol. and Pal. Queensland, 1892, p. 69 (Gyroceras). De Koninck, L. G. Mem. Geol. Surv. New South Wales, Pal. No. 6, 1898, pp. 85-105.
Carboniferous.—Etheridge, R. jnr. Rec. Austr. Mus., vol. III. No. 1, 1897, pp. 7-9 (Actinoceras). Idem, Geol. Surv. W.A., Bull. No. 27, 1907, pp. 32-37.
Carbopermian.—Morris, J., in Strzelecki’s Phys. Descr. of New South Wales, etc., 1845, pp. 270-278 and 285-291. Foord, A. H. Geol. Mag., Dec. III. vol. VII. 1890, pp. 103, 104. Etheridge, R. jnr. Geol. and Pal. Queensland, 1892, pp. 264-296. Idem., Proc. Linn. Soc. New South Wales, vol. IX. 1895, pp. 530-537 (Pelec. and Gastr.). De Koninck, L. G. Mem. Geol. Surv. New South Wales, Pal. No. 6, 1898, pp. 203-274. Etheridge, R. jnr. and Dun, W. S. Mem. Geol. Surv. New South Wales, Pal. No. 5, vol. II. pt. I. 1906 (Palaeopecten). Idem, ibid., vol. II., pt. 2, 1910 (Eurydesma).
Trias.—Zittel, K. Novara Exped., vol. I. Abth. II. Geol. Theil., 1864, pp. 26-29. Etheridge, R. jnr. Mem. Geol. Surv. New South Wales, Pal. No. 1, 1888, pp. 8-14.
Jurassic.—Zittel, K. Novara Exped., vol. I., Abth. II. Geol. Theil., 1864, pp. 20-34. Moore, C. Quart. Journ. Geol. Soc., vol. XXVI. pp. 245-260 (Jurassic and Cretaceous Moll.). Etheridge, R. jnr. ibid., vol. XXVIII. 1872, pp. 317-359 (Palaeozoic, Jur. and Cret. Moll.). Crick, G. C. Geol. Mag., Dec. IV. vol. I. 1894, pp. 385 393 and 433-441 (Ceph.). Chapman, F. Proc. R. Soc. Vict., vol. XVI. pt. II. 1904, pp. 327-332. Marshall, P. Trans. New Zealand Inst., vol. XLI. 1909, pp. 143-145 (New Zealand Ceph.). Etheridge, R. jnr. Geol. Surv. W.A. Bull. No. 36, 1910, pp. 30-40.
Cretaceous.—Etheridge, R. jnr. Geol. and Pal. Queensland, 1892, pp. 445-503 and 561-574. Idem, Geol. Surv. Queensland, Bull. No. 13, 1901, pp. 13-35. Idem, Mem. Roy. Soc. S. Aust., vol. II. pt. 1, 1902 (S.A. Moll.). Idem, Mem. Geol. Surv. New South Wales, Pal. No. 11, 1902, pp. 16-49 (New South Wales Moll.).
Cainozoic.—Zittel, K. Novara Exped. Geol. Theil., vol. I. Abth. II. 1864, pp. 34-55 (Pelec. and Gastr. New Zealand). McCoy, F. Prod., Pal. Vict., Dec. I. 1874; Dec. II. 1875; Dec. III. 1876; Dec. V. 1877; Dec. VI. 1879. Woods, J. E. T. Proc. R. Soc. Tas. (1875), 1876, pp. 13-26 (Table Cape Moll.). Idem, Proc. Linn. Soc. New South Wales, vol. III. 1879, pp. 222-240 (Muddy Creek Moll.). Idem, ibid., vol. IV. 1880, pp. 1-24. « 219 » Hutton, F. W. Trans. New Zealand Inst. vol. IX. 1877, pp. 593-598. Ibid., vol. XVII. 1885, pp. 313-332 (New Zealand Pelec. and Gastr.). Idem, Proc. Linn. Soc. New South Wales, vol. I. 2nd ser. (1886), 1887, pp. 205-237, (distr. lists, Pareora and Oamaru). Idem, Macleay, Mem. Vol. Linn. Soc. New South Wales, 1893, pp. 35-92 (Pliocene Moll. New Zealand). Tate, R. Trans. R. Soc. S. Austr., vol. VII. 1886, pp. 96-158, and vol. IX., 1887, pp. 142-189 (Pelec.); ibid., pp. 190-194 (Scaphopoda); ibid., 194-196 (Pteropoda). Idem, ibid., vol. X. 1888, pp. 91-176; vol. XI. 1889, pp. 116-174; vol. XIII. 1890, pp. 185-235; and vol. XVII. 1893, pp. 316-345 (Gastr.). Idem, Journ. R. Soc., New South Wales, vol. XXVII. 1893, pp. 169-191. Idem, ibid., vol. XXXI. 1897, pp. 392-410 (Gastr. and Pelec.). Idem, Trans. Roy. Soc. S. Austr., vol. XXIII. 1899, pp. 260-277 (Revision of Moll.). Pritchard, G. B. Proc. Roy. Soc. Vic., vol. VII. 1895, pp. 225-231 (Pelec.). Idem, ibid., vol. VIII. 1896, pp. 79-141 (Moll. of T. Cape). Idem, ibid., vol. XI. pt. I. 1898, pp. 96-111 (Gastr.). Idem, ibid., vol. XIV. pt. I. 1901, pp. 22-31 (Pelec.). Idem, ibid., vol. XVI. pt. II. 1903, pp. 87-103 (Pelec.). Idem, ibid., vol. XVI. pt. I. 1903, pp. 83-91 (Pleurotomaria). Idem, ibid., vol. XVII. pt. I. 1904, pp. 320-337 (Gastr.) Idem, ibid., vol. XXVI. (N.S.) pt. I. 1913, pp. 192-201 (Volutes). Hall, T. S. Proc. R. Soc. Vict., vol. XVII. pt. II. 1905, pp. 391-393 (Chitons). Ashby, E. and Torr. W. G. Trans. R. Soc. S. Austr., vol. XXV. 1901, pp. 136-144 (Chitons). Thomson, J. A. Trans. New Zealand Inst., Vol. XL. 1908, pp. 102, 103 (N.Z. Moll.). Chapman, F. Proc. R. Soc. Vict. vol. XX. pt. II. 1908, pp. 218-220 (Chiton). Idem, ibid., vol. XXV. pt. I. 1912, pp. 186-192 (Gastr.).
FOSSIL TRILOBITES, CRUSTACEA AND INSECTS.
Arthropods and their Structure.—
The above-named fossil groups are included by zoologists in the sub-kingdom Arthropoda (“joint-footed animals”). The Arthropods possess a body and limbs composed of a number of jointed segments covered externally with a hard, shelly material and separated by a softer, flexible skin. They have no internal skeleton, and therefore the only portion which can be preserved in the fossil state is the harder part of the outer covering. Under exceptional conditions of fossilisation, however, even frail insects such as ants, wasps and dragon-flies are sometimes found more or less wholly preserved and showing their original minute structure.
Subdivisions of Arthropoda.—
The principal representatives of the group of the Arthropods which are found as fossils include the Trilobites; various Crustacea proper, as Crabs, Lobsters, Shrimps, Pod-shrimps and Water-fleas; the Insects; and occasionally Spiders and Scorpions (Arachnida). The King-crabs and Eurypterids (as « 221 » the extinct Pterygotus) form a separate sub-class, the Merostomata, which are placed by some authors in the group of Spiders and Scorpions: their remains date back to the time when the older Palaeozoic strata were deposited.
Crustacea, an Archaic Group.—
A typical division of the Arthropod group, and one which was well represented from the earliest period up to the present day, is the CRUSTACEA. As the name denotes, these animals are generally invested with a strong shelly covering or “crust,” usually of horny or chitinous material, which in some forms is strengthened by deposits of phosphate of lime. Of the horny condition of the shell the groups of the bivalved Crustacea (Ostracoda) and the “water-fleas” (Entomostraca) supply notable instances; whilst the limy-structured shell is seen in the common crab. Some authorities separate the great extinct group of the Trilobites from the rest of the Crustacea; but it will here be convenient, in a preliminary study, to consider them together.
Development of Crustacea.—
The development of the lower forms of the Crustacea is interesting, from the fact that the young usually escapes from the egg in a larval state known as a “nauplius.” In this stage there are no segments to the body, and but a solitary median eye, such as may be seen in the common water-flea known to microscopists as Cyclops. The three pairs of appendages seen in this larval crustacean represent the two pairs of antennae and the jaws or mandibles of the full-grown form.
Among the higher Crustacea, however, there is no larval form; the young escaping from the egg in a more or less highly developed condition resembling the adult. The group of the Crabs, Lobsters and Shrimps (or Decapoda, i.e., having ten ambulatory feet) exhibit a larval stage in which the young form (“zoea”) has a segmented abdomen and seven pairs of appendages.
Trilobites.—
The first group of arthropods here described is that of the TRILOBITES. These were so named on account of the three-lobed form of the body. This particular feature distinguishes them from the Crustacea proper; which includes the Phyllopods (with leaf-like limbs), as the freshwater Estheria, the Ostracoda or Bivalved Water-fleas, the Barnacles or Cirripedia and the Higher Crustacea (Malacostraca), including Shrimps, Crabs, and Lobsters, of which the oldest representatives are the Pod-shrimps (Phyllocarida).
Habits of Trilobites.—
The remains of these primitive but often strikingly ornamented crustacean-like animals, the trilobites, are found in comparative abundance in the limestones, mudstones, and even the sandstones of the older sedimentary rocks of Australasia. They were amongst the most prolific types of animal life existing in the seas of Palaeozoic times, and are especially characteristic of Cambrian, Ordovician and Silurian rocks. Trilobites, as a group, seem to have adapted themselves to almost all conditions of marine life: « 223 » some are found in the hardened black mud of shallow waters, whilst others are to be looked for in the limestones and excessively fine sediments of deeper waters. In all probability certain of these forms crawled over the soft, oozy sea-bed in order to obtain their food, and consequently their remains in the stratified rocks would be restricted to the fine black shales; whilst the freely swimming forms could change their habitat at will, and would be found alike in sandy or clayey deposits. As some indication of their varied habits, the eyes of trilobites differ greatly in size. They are always compound like the eye of the house-fly, though of a semi-lunar shape. In some forms the eyes are very small or even absent, whilst in others they are exceedingly large and prominent. This latter feature probably indicates their frequenting moderately deep water.
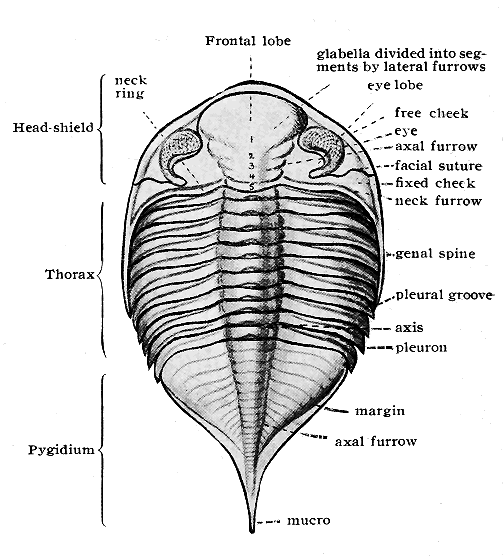
Structure of Trilobites.—
The complete structure and zoological relationship of the trilobites has always been open to some doubt. As regards the former, within recent years exceptionally well-preserved specimens from the Utica Slates and the Cincinnati Limestone of Ohio, rocks of Ordovician age, have been discovered and dissected, whereby our knowledge of the organisation of this group is greatly advanced. These remarkable fossil remains show that the Trilobites bore on their under surface a number of appendages, one pair to each segment, except that of the anal. The front pair is whip-like and served as antennae; the others are branched, the forward portion being a crawling limb, and the hinder, which was fringed with bristles or thin plates, may have served either for swimming or breathing. At the base of the four pairs of appendages attached to the head there was an arrangement for biting the food, from whence it was passed to the mouth. Taking one of the commonest Australasian trilobites, Dalmanites meridianus, for an example of general structure, and looking at the back of the shell or upper surface, we see the trilobate (three-lobed) form well defined (Fig. 107). The central ridge is termed the axis, and on either side of this are arranged the pleural lobes, each well marked transverse division of which, in the central or thoracic region, being a pleuron or rib. The whole body is divided into three more or less distinct portions,—the head-shield or cephalon, the thorax, and the tail-shield or pygidium. The central area of the head-shield is called the glabella or cranidium, against which, on either side, are placed the free cheeks carrying the compound sessile eyes when present. The appendages of the head are pediform or leglike, arranged in five pairs, and biramous or forked, excepting the antennae, which are simple and used as sensory organs. In front of the mouth is the hypostoma or forelip, and behind it is the metastoma or hind-lip. The segments of the head-shield are most closely united, and in all the trilobites are of the same number. Those of the thorax have flexible joints and are variable in number. The segments of the abdomen are fused together and form a caudal shield or pygidium.
The larval stage of the trilobite was a protonauplian form (that is more primitive than the nauplius), the protoaspis; the adult stage, being attained by the addition of segments at the successive moults.
The earliest known trilobites in Australia are some Cambrian species from South Australia, Western Australia, Victoria, and Tasmania.
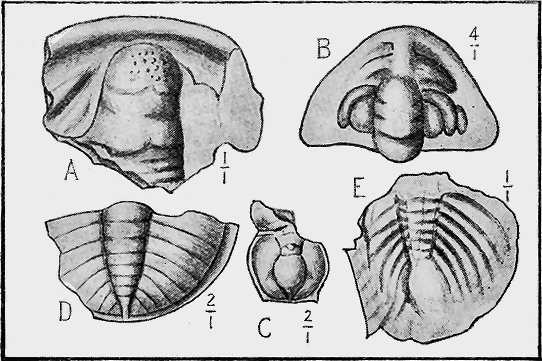
Lower Cambrian Trilobites.—
In the Lower Cambrian Limestone of Yorke Peninsula, South Australia, the following trilobites occur:—a species doubtfully referred to Olenellus (? O. pritchardi); Ptychoparia howchini (Fig. 108 A); P. australis; Dolichometopus tatei (Fig. 108 B); and « 227 » Microdiscus subsagittatus. The Cambrian of the Northern Territory contains Olenellus brownii. In Western Australia Olenellus forresti is found in similar beds.
Upper Cambrian Trilobites.—
The Dolodrook Limestone (Upper Cambrian) of Gippsland, Victoria, contains the remains of the primitive little trilobite Agnostus (A. australiensis, Fig. 108 C); Crepicephalus (C. etheridgei); and Ptychoparia (P. thielei (Fig. 108 D) and P. minima). The Upper Cambrian sandstones of Caroline Creek, Tasmania, contain Dikellocephalus (D. tasmanicus); a species of Asaphus and Ptychoparia (P. stephensi). Beds of the same age in the Florentine Valley, Tasmania, have yielded Dikellocephalus (D. florentinensis, Fig. 108 E).
Ordovician Trilobites.—
Trilobites of Lower Ordovician age or even older, are found in the Knowsley beds near Heathcote in Victoria. They are referred to two genera, Dinesus and Notasaphus. Both forms belong to the ancient family of the Asaphidae. Associated with these trilobites are some doubtful species of sea-weed, spicules of siliceous sponges, traces of threadlike hydrozoa, some fragments of graptolites allied to Bryograptus, and several brachiopods. At the Lyndhurst Gold-fields, near Mandurama, New South Wales, trilobites related to the genus Shumardia have been found associated with brachiopods (lamp-shells), pteropods (sea-butterflies), and graptolites (hydrozoa) of an Upper Ordovician facies.
The limestone beds at Laurie’s Creek and other localities in Central Australia contain remains of Asaphus illarensis, A. howchini and A. lissopelta; whilst in the limestone and quartzite of Middle Valley, Tempe Downs, A. thorntoni also occurs.

Silurian Trilobites.—
Trilobites are well-known fossils in the Australasian Silurian strata. As they occur rather abundantly along with other fossils in rocks of this age they are extremely useful aids in separating the system into the different beds or zones. In Victoria the Silurian is divisible into two sets of beds: an older, or Melbournian stage (the bed-rock of Melbourne) « 229 » and a younger, Yeringian (Lilydale series). Trilobites of Melbournian age are found to belong to the genera Ampyx, Illaenus, Proetus, Cyphaspis, Encrinurus (Cromus) and Homalonotus. The commonest species are Cyphaspis spryi (Fig. 109 B), and Encrinurus (Cromus) spryi from the South Yarra mudstones; and Ampyx parvulus, var. jikaensis (Fig. 109 A), and Homalonotus harrisoni (Fig. 109 C), from the sandstone of Moonee Ponds Creek.
The handsome Dalmanites meridianus and Homalonotus vomer occur at Wandong in what appear to be passage beds between the Melbournian and Yeringian.
The Yeringian of Victoria is far richer in trilobites than the preceding series, and includes the genera Proetus, Cyphaspis, Bronteus, Lichas, Odontopleura, Encrinurus, Calymene, Homalonotus, Cheirurus, and Phacops. The rocks in this division occur as mudstones, limestones, and occasionally sandstones and conglomerates. The mudstones, however, prevail, and these pass insensibly into impure limestones of a blue-black colour, weathering to brown, as at Seville; the change of structure indicating less turbid water. At Lilydale, and on the Thomson River, as well as at Loyola and Waratah Bay, almost pure limestone occurs, which represents clear water conditions, not necessarily deep; there, however, trilobites are scarce, and the prevailing fauna is that of an ancient coral reef. Some described Yeringian species are Lichas australis (Fig. 110 A), Odontopleura jenkinsi (Fig. 110 B) (found also in New South Wales), Encrinurus punctatus (Fig. 110 C), Calymene tuberculosa, « 230 » Bronteus enormis, Phacops sweeti, and P. serratus (Fig. 110 E). In Calymene (“covered up”) the joints of the thorax are facetted at the angles, so that each pleuron could work over that immediately behind; in consequence of this it could roll itself up like a woodlouse or slater, hence the name of the genus. This trilobite also occurs in England, and is there known amongst the quarry men and fossil collectors as the “Dudley Locust.” Perhaps the most characteristic and common trilobite of the Yeringian series in Victoria is Phacops sweeti (Fig. 110 D), formerly identified with Barrande’s P. fecundus, from which it differs in the longer and larger eye with more numerous lenses. It is found in Victoria in the Upper Yarra district near the junction of the Woori Yallock and the Yarra Rivers; north-west of Lilydale; near Seville; at Loyola near Mansfield; and at Fraser’s Creek near Springfield, Kilmore.

In New South Wales trilobites are abundant in the Yass district, amongst other localities, where the upper beds, corresponding to the Yeringian of Victoria, are well developed. Dalmanites meridianus is common to the Silurian of New South Wales, Victoria, and Tasmania. In Victoria this handsome species is found in the hard, brown, sandy mudstone of Broadhurst’s and Kilmore Creeks, and, as previously noted, in the hard, blue mudstone of Wandong. At the latter locality specimens may be found in the railway ballast quarry, where they are known to the workmen as “fossil butterflies.” The species also occurs at the famous fossil locality of Hatton’s Corner, Yass; at Bowning; and at Limestone Creek, all in New South Wales. Other trilobites occurring in the Silurian of New South Wales are Odontopleura jenkinsi, O. bowningensis, Cheirurus insignis and Phacops latigenalis (Fig. 109 D).
In the Wangapeka series of New Zealand the calcareous shales and limestones of the upper division contain Calymene blumenbachii, Homalonotus knightii and H. expansus.
Devonian Trilobites.—
Trilobites suddenly became rare in the Australian Devonian. The only known examples of trilobite remains belong to a species of Cheirurus occasionally found in the Middle Devonian limestone of Buchan, « 232 » Victoria; and a species of Proetus in the Devonian of Barker Gorge, Napier Range, West Australia.
Carbopermian Trilobites.—
Trilobites of Carbopermian age are found in New South Wales, Queensland, and Western Australia. All the genera belong to the family Proetidae. The genera Phillipsia (P. seminifera, Fig. 111 A), Griffithides (G. eichwaldi, Fig. 111 B), and Brachymetopus (B. strzelecki, Fig. 111 C) occur in New South Wales. Griffithides eichwaldi is also found in Queensland. Other Queensland species are Phillipsia woodwardi, P. seminifera var. australasica and P. dubia. Phillipsia grandis is found in the Carbopermian of the Gascoyne River, Western Australia.

Phyllopoda in Carboniferous, Triassic and Jurassic.
The PHYLLOPODA, which belong to the Crustacea in the strict sense of the term, comprise the Estheriidae and Cladocera (water-fleas). The former group is represented by Leaia mitchelli, which is found in the Upper Carboniferous or Carbopermian of the Newcastle District, New South Wales. In the still later Hawkesbury series (Triassic) of New South Wales, Estheria coghlani (Fig. 111 D) occurs. This species is a minute form, the carapace measuring from 1.25mm. to 2mm. in the longer diameter of the shell. In the upper part of the Wairoa Series (Triassic) of Nelson, New Zealand, there is found another species of Estheria, identified with a European form E. minuta. Estheria mangaliensis is another form occurring in the Jurassic (Ipswich series) of Queensland. At the present day these little Estheriae sometimes swarm in countless numbers in freshwater lakes or salt marshes.
Ostracoda: Their Structure.—
Passing on to the next group, the bivalved OSTRACODA, we note that these have existed from the earliest geological periods to the present day. They are usually of minute size, commonly about the sixteenth of an inch in length, although some attained a length of nearly one inch (Leperditia). Their bodies are indistinctly segmented, and are enclosed within a horny or calcareous shell. This shell consists of two valves which are joined along the back by a ligament or hinge, the ends and ventral edge remaining quite free. The pairs of appendages present are the antennae (2), mandibles (1), maxillae « 234 » (2), and thoracic feet (2). The only portion found in the fossil state is the bivalved carapace, the two valves being frequently met with still united, especially when these tiny animals have settled down quietly on the sea-bed and have been quickly covered with sediment.
Features of the Ostracod Carapace.—
Since the body parts of the ostracod are wanting in the fossil examples, the generic determination is attended with some difficulty, especially in regard to the smooth or bean-shaped forms. The chief distinctive characters to note are, the contour of the carapace seen in three directions (top, side and end views), the structure of the hinge, and the position and figure of the muscle-spots or points of adhesion of the muscular bands which hold or relax the two valves. The valves in certain genera fit closely upon one another. In others, one overlaps the other, the larger being sometimes the right (as in Leperditia), sometimes the left (as in Leperditella). The hinge-line is often simple or flange-like, or it may consist of a groove and corresponding bar, or there may be a series of teeth and sockets. Lateral eye-tubercles are sometimes seen on the surface of the valve, whilst in the animal there was also a small eye.
Habits of Ostracoda.—
Ostracoda swarmed in many of the streams, lakes and seas of past geological times, and they still exist in vast numbers under similar conditions. Like some other minute forms of life, they played a most important part in building up the rock formations of « 235 » the sedimentary series of the earth’s crust; and by the decomposition of the organism itself they are of real economic value, seeing that in some cases their decay resulted in the subsequent production of oil or kerosene shales and bituminous limestones. The Carboniferous oil shales in the Lothians of Scotland, for example, are crowded with the carapaces of Ostracoda associated with the remains of fishes.
Cambrian Ostracoda.—
Some undescribed forms of the genus Leperditia occur in the hard, sub-crystalline Cambrian Limestone of Curramulka, South Australia.
Silurian Ostracoda.—
In Victoria and New South Wales the oldest rocks from which we have obtained the remains of Ostracoda up to the present, are the uppermost Silurians, in which series they occur both in the limestone and the mudstone. In Victoria their bivalved carapaces are more often found in the limestone; but one genus, Beyrichia, is also met with in abundance in the mudstone. These mudstones, by the way, must have originally contained a large percentage of carbonate of lime, since the casts of the shells of mollusca are often excessively abundant in the rock, and the mudstone is cavernous, resembling an impure, decalcified limestone. These Yeringian mudstones of Victoria seem, therefore, to be the equivalent of the calcareous shales met with in the Wenlock and Gotland Series in Europe; a view entirely in accordance with the character of the remainder of the fauna. One of the commonest of the Silurian ostracods is Beyrichia kloedeni, a form having an extensive distribution in « 236 » Europe. It occurs in the Silurian mudstone of the Upper Yarra District. Other species of the same genus are B. wooriyallockensis (Fig. 112 A), distinguished from the former by differences in the shape of the lobes and its longer valves; also a form with narrow lobes, B. kilmoriensis; and the ornate B. maccoyiana, var. australis. Of the smooth-valved forms, mention may be made of Bythocypris hollii, B. caudalis (Fig. 112 D), and the striking form, Macrocypris flexuosa. Regarding the group of the Primitiae, of which as many as thirteen species and varieties have been described from the Lilydale Limestone, we may mention as common forms P. reticristata (Fig. 112 E) and P. punctata. This genus is distinguished by the bean-shaped or purse-shaped carapace, with its well developed marginal flange and mid-dorsal pit. Other genera which occur in our Silurians and are of great interest on account of their distribution elsewhere. are Isochilina, Aparchites, Xestoleberis, Aechmina, and Argilloecia.
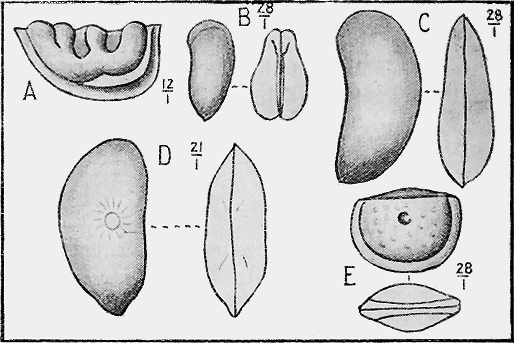
The largest ostracod yet described from Australia, measuring more than a quarter of an inch in length, occurs in the Upper Silurian of Cliftonwood, near Yass, New South Wales. It belongs to the genus Leperditia (L. shearsbii), and is closely related to L. marginata, Keyserling sp.; which occurs in strata of similar age in the Swedish and Russian Baltic area. A limestone at Fifield, New South Wales, probably of Silurian age, contains Primitia, Kloedenia, and Beyrichia.
Devonian Ostracoda.—
The little Primitia cuneus (Fig. 113 A) with a bean-shaped carapace and median pit or depression occurs somewhat frequently in the Middle Devonian Limestone of Buchan, Victoria. Another species, Primitia yassensis, is found in the shaly rock of Narrengullen Greek, New South Wales. It is probable that many other species of the group of the ostracoda remain to be described from Australian Devonian rocks.
Carboniferous Ostracoda.—
In Queensland a conspicuous little ostracod is Beyrichia varicosa from the Star Beds of Corner Creek.
Carbopermian Ostracoda.—
In the Carbopermian of Cessnock, New South Wales, Primitia dunii occurs; and in that of Farley is found Jonesina etheridgei. From both these « 238 » localities Leperditia prominens was also obtained. Another species from New South Wales is Entomis jonesi (Fig. 113 B), described from the Muree Sandstone by de Koninck.
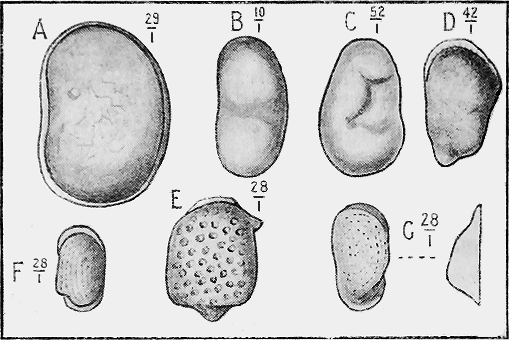
Triassic Ostracoda.—
The Triassic (Wiannamatta Shales) of Grose Vale, New South Wales has afforded a few specimens of ostracoda belonging to Synaphe (S. mesozoica, Fig. 113 C), ? Darwinula, and ? Cytheridea.
Jurassic Ostracoda.—
The marine Jurassic strata of Western Australia at Geraldton, have yielded a small but interesting series of ostracoda, largely of modern generic types. The genera, which were found in a rubbly Trigonia-Limestone, « 239 » are Cythere, Paradoxorhyncha, Loxoconcha, and Cytheropteron.

Cainozoic Ostracoda.—
The fossiliferous clays and calcareous sands of the southern Australian Cainozoic beds often contain abundant remains of ostracoda. The moderately shallow seas in which the fossiliferous clays, such as those of Balcombe’s Bay, were laid down, teemed with these minute bivalved Crustacea. All the forms found in these beds are microscopic. They either belong to living species, or to species closely allied to existing forms. Some of the more prominent of the Balcombian species are Cythere senticosa, a form which is now found living at Tenedos, and C. clavigera « 240 » (Fig. 114 B), with the young form sometimes referred to as C. militaris, a species which may still be dredged alive in Hobson’s Bay. Other genera common in these clays are Bairdia, with its broad, pear-shaped carapace, represented by the still living B. amygdaloides (Fig. 114 A). Cytherella, with its compressed, subquadrate carapace, as seen in C. punctata (Fig. 114 D), a species having an elaborate series of muscle-spots, and which, like the previous species, is found living in Australian seas; and Macrocypris, with its slender, pointed, pear-shaped outline.
Cirripedia: Their Habits and Structure.—
CIRRIPEDIA OR BARNACLES.—These curious modifications of the higher group of Crustacea (Eucrustacea) date back to Ordovician times. They appear to have tried every possible condition of existence; and although they are mostly of shallow water habits, some are found at the great depth of 2,000 fathoms (over two miles). Those which secrete lime or have calcareous shells, attach themselves to stones, pieces of wood, shell-fish, crabs, corals and sea-weeds. Others are found embedded in the thick skin of whales and dolphins, or in cavities which they have bored in corals or shells of molluscs. Some are found parasitic in the stomachs of crabs and lobsters, or within other cirripedes. They begin life, after escaping from the egg, as a free-swimming, unsegmented larva (“nauplius” stage), and before settling down, pass through the free-swimming, segmented “cypris” stage, which represents the pupa condition, and in which state they explore their surroundings in search « 241 » of a suitable resting place for their final change and fixed condition. Just before this occurs, glands are developed in the pupa barnacle, which open into the suckers of the first pair of appendages or antennae. When a suitable place for fixation has been found, these glands pour out a secretion which is not dissolved by water, and thus the barnacle is fixed head downwards to its permanent position. The compound eyes of the “cypris” stage disappear, and henceforth the barnacle is blind. The characteristic plates covering the barnacle are now developed, and the six pairs of swimming feet become the cirri or plumes, with which the barnacle, by incessant waving, procures its food. In short, as remarked by one authority, it is a crustacean “fixed by its head, and kicking the food into its mouth with its legs.”
Cirripedes may be roughly divided into two groups, the Acorn Barnacles and the Goose Barnacles. Although dissimilar in general appearance, they pass through identical stages, and are closely related in most of their essential characters. The latter forms are affixed by a chitinous stalk or peduncle, whilst the acorn barnacles are more or less conical and affixed by the base.
Silurian Cirripedes.—
The stalked barnacles are probably the oldest group, being found as far back as the Ordovician period. In Australia the genus Turrilepas occurs in Silurian rocks, T. mitchelli (Fig. 115 A) being found at Bowning in the Yass District of New South Wales. The isolated plume-like plates of T. yeringiae (Fig. 115 B) are not uncommon in the olive mudstone of the Lilydale District in Victoria.
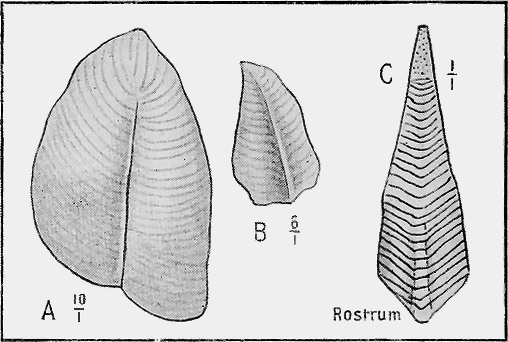

Cainozoic Lepadidae.—
The genus Lepas (the modern goose barnacles) is represented by isolated plates in the Cainozoic (Janjukian) limestones and marls of Waurn Ponds, and Torquay near Geelong: it also occurs in a stratum of about the same age, the nodule bed, at Muddy Creek, near Hamilton, Victoria (L. pritchardi, Fig. 116). In New Zealand the gigantic cirripede, ?Pollicipes aucklandicus (Fig. 115 C), occurs in the Motutapu beds.
Cainozoic Balanidae.—
The Acorn Barnacles are represented in our Cainozoic shell marls and clays by a species of Balanus from the Janjukian of Torquay; whilst two species of the genus occur in the Kalimnan beds at Beaumaris, Port Phillip, in similar beds in the Hamilton District, and at the Gippsland Lakes.
Phyllocarida: Their Structure.—
A large and important group of the higher Crustacea, but confined to the older rocks of Victoria, is the order PHYLLOCARIDA. This seems to form a link between the Entomostraca, including the bivalved Ostracoda and the well-known group of the lobsters, shrimps and crabs. The body of these phyllocarids consists of five segments to the head, eight to the thorax, and from two to eight to the abdomen. The portion usually preserved in this group is the carapace, which covers the head and thorax, and although often in one piece, is sometimes hinged, or « 244 » otherwise articulated along the back. In front of the carapace there is a moveable plate, the rostrum or beak (Fig. 117). There are two pairs of antennae to the head, and the animal is provided with a pair of stalked compound eyes. The thoracic segments are furnished with soft leaf-like legs as in the Phyllopods. The abdomen is formed of ring-like segments, and generally terminates in a sharp tail-piece or telson, often furnished with lateral spines. In many respects the ancient phyllocarids correspond with the living genus Nebalia, which is found inhabiting the shallow waters of the Mediterranean and elsewhere.
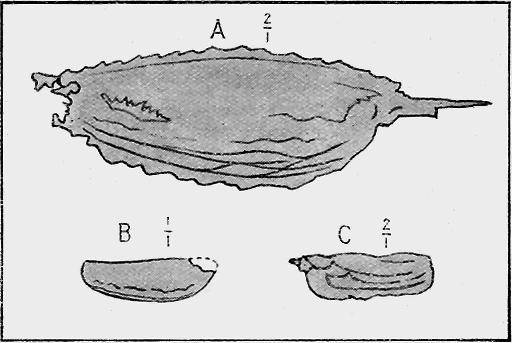

Ordovician Phyllocarids.—
Phyllocarids of the Lower Ordovician slates are referred to the genera Rhinopterocaris, Caryocaris, Saccocaris and Hymenocaris. The first-named is the commonest type; and is found in slates of the Lancefield, Bendigo and Castlemaine Series at the localities named, as well as at Dromana. Rhinopterocaris (Fig. 118 A) is readily distinguished by its long—ovate outline, and this, together with its wrinkled chitinous appearance makes it resemble the wing of a dipterous insect. Caryocaris (Fig. 118 B) is a smaller and narrower form which occurs in the Victorian Lower Ordovician slates, as well as in ice-borne blocks derived from the Ordovician, at Wynyard, in N.W. Tasmania.
Silurian Phyllocarids.—
The chief type of Phyllocarid in the Silurian is Ceratiocaris (Fig. 119). The carapace is typically ovate, straight on one edge, the dorsal, and convexly curved on the other, the ventral. They resemble bean-pods in outline, hence the name “pod-shrimps.” Several species are known from the Victorian shales, mudstones, and sandstones; the forms found in Australia if complete would seldom attain five inches in length, whilst some British species are known to reach the exceptional length of two feet. The long, grooved and jointed telson is not uncommon in the sandstones of Melbourne and Kilmore. Other genera described from Victoria are Aptychopsis and Dithyrocaris.
Lower Cretaceous Crab.—
The earliest example of the DECAPODA in the Australian rocks, so far recorded, is the Lower Cretaceous Prosopon etheridgei (Fig. 120 A) from Queensland, which has affinities with some Jurassic and Neocomian crabs found in Europe. Other crustacean « 247 » remains of less decipherable nature occur in this same deposit.
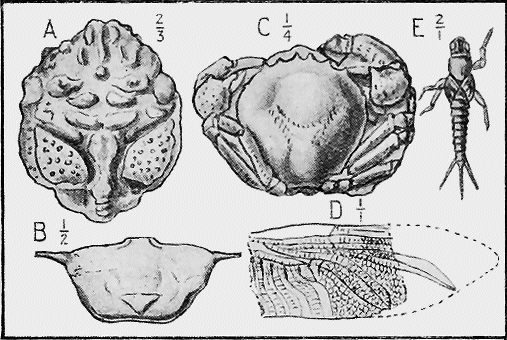
Cainozoic Crabs.—
Of the Cainozoic decapod Crustacea there is a Victorian species of a stalk-eyed crab, Ommatocarcinus corioensis (Fig. 120 B), found in the marls of Curlewis and Port Campbell, and probably of Janjukian age. Various portions of similar Crustacea, consisting of claws and fragmentary carapaces, are found from time to time in the Victorian clays and limestones of Balcombian and Janjukian ages, but they are insufficient for identification. A carapace of one of the Oxystomata (with rounded cephalo-thorax and « 248 » non-salient frontal region) has occurred in the Kalimnan marl of the Beaumaris Cliffs, Port Phillip. It is closely allied to a crab now found in Hobson’s Bay and generally along the Victorian coast.
Remains of a shore-crab (Fam. Cancridae) are found at three localities, in the Oamaru Series, in New Zealand; near Brighton, in Nelson and at Wharekuri in the Waitaki Valley. It has been described under the name of Harpactocarcinus tumidus (Fig. 120 C), a genus of the Cyclometopa or “bow crabs.”
Pleistocene Lobster.—
Numerous remains of a lobster, Thalassina emerii (see antea, Fig. 20), supposed to be of Pleistocene age, occur in nodules found on Queensland and North Australian (Port Darwin) beaches.
Eurypterids in the Silurian.—
The order EURYPTERIDA comprises an extinct group of Crustacea closely allied to the modern King-crab (Limulus). The body was covered with a thin chitinous skeleton, ornamented with regular scale-like markings. This group is represented in Victorian rocks by the remains of Pterygotus (“Sea-scorpions”), animals which often attained a length of six feet. Pterygotus (see Fig. 121 A) had the fore part of the body fused, forming the cephalo-thorax, which was furnished with anterior, marginal facetted eyes and central ocelli or smaller simple ones. To the ventral surface of the body were attached six pairs of appendages. The first pair are modified antennae with pincer-like terminations, used for prehensile « 249 » purposes. Then come four pairs of slender walking feet. The sixth pair of appendages is in the form of powerful swimming feet or paddles, at the bases of which are the comb-like jaws. The abdomen consists of thirteen joints, the last of which, the telson, is spatulate and posteriorly pointed. Fragments of a tolerably large species of Pterygotus occur in the Silurian shales of South Yarra, Melbourne, Victoria. It was probably about 18 inches in length when complete. Of this form, known as P. australis (Fig. 121 B), portions of the chelate (clawed) appendages, and parts of the abdominal segments have been found from time to time, but no complete fossil has yet been discovered.
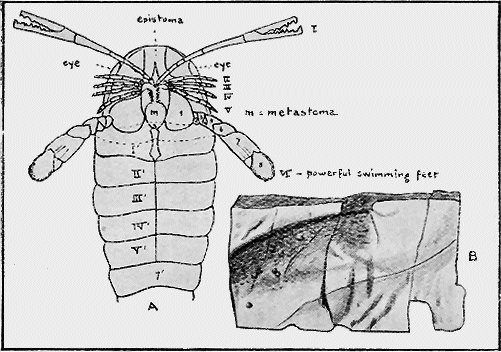
Jurassic Insects.—
Of the group of the INSECTA, the Ipswich Coal measures (Jurassic) of Queensland have yielded an interesting buprestid beetle (Mesostigmodera), whilst beds of the same age in New South Wales contain the remains of a probable Cicada, associated with leaves of the fern Taeniopteris.
Lower Cretaceous Dragon-fly.—
From the Lower Cretaceous of the Flinders River district, Queensland, there has been obtained a fossil dragon-fly, Aeschna flindersensis (Fig. 120 D).
Cainozoic Insects.—
Certain Cainozoic beds of New South Wales, of the age of the Deep-leads of Victoria, and probably equivalent to the Kalimnan terrestrial series, contain a species of Cydnus, a bug-like insect belonging to the order Rhynchota; and there are in the same series a Midge (Chironomus), a Day-fly (Ephemera, Fig. 120 E) and several beetles (? Lagria, Palaeolycus, Cyphon and Oxytelus). The occurrence of these insects of the Deep-leads helps to complete the landscape picture of those far-off Lower Pliocene times, when the old river systems brought down large contributions of vegetable waste from higher lands, in the form of twigs with leaves and fruits; with occasional evidences of the rich and varied fauna of insect life which was especially promoted in the damp and vegetative areas of the lower lands.
COMMON OR CHARACTERISTIC SPECIES OF THE FOREGOING CHAPTER.
TRILOBITES.
Ptychoparia howchini, Eth. fil. Lower Cambrian: South Australia.
Dolichometopus tatei, H. Woodward. Lower Cambrian: South Australia.
Olenellus browni, Eth. fil. Lower Cambrian: Northern Territory.
Agnostus australiensis, Chapm. Upper Cambrian: Victoria.
Ptychoparia thielei, Chapm. Upper Cambrian: Victoria.
Dikellocephalus florentinensis, Eth. fil. Upper Cambrian: Tasmania.
Dinesus ida, Eth. fil. Lower Ordovician: Victoria.
Asaphus illarensis, Eth. fil. Ordovician: Central S. Australia.
Ampyx parvulus, Forbes, var. jikaensis, Chapm. Silurian (Melbournian): Victoria.
Illaenus jutsoni, Chapm. Silurian (Melbournian): Victoria.
Proetus euryceps, McCoy. Silurian: Victoria.
Cyphaspis spryi, Gregory. Silurian (Melbournian): Victoria.
Bronteus enormis, Eth. fil. Silurian (Yeringian): Victoria.
Lichas australis, McCoy. Silurian (Yeringian): Victoria.
Odontopleura jenkinsi, Eth. fil. Silurian: New South Wales. Silurian (Yeringian): Victoria.
Encrinurus punctatus, Brunnich sp. Silurian: New South Wales. Silurian (Yeringian): Victoria.
Encrinurus (Cromus) murchisoni, de Koninck. Silurian: New South Wales.
Encrinurus (Cromus) spryi, Chapm. Silurian (Melbournian): Victoria.
Calymene blumenbachii, Brongn. Silurian (Wangapeka Series): New Zealand.
Homalonotus expansus, Hector. Silurian (Wangapeka Series): New Zealand.
Homalonotus knightii, König. Silurian (Wangapeka Series): New Zealand.
Homalonotus harrisoni, McCoy. Silurian (Melbournian): Victoria.
Homalonotus vomer, Chapm. Silurian: Victoria.
Cheirurus insignis, Beyrich. Silurian: New South Wales.
Phacops sweeti, Eth. fil. and Mitch. Silurian: New South Wales. Silurian (Yeringian): Victoria.
Phacops serratus, Foerste. Silurian (Yeringian): Victoria. Silurian: New South Wales.
Dalmanites meridianus, Eth. fil. and Mitch, sp. Silurian: New South Wales, Victoria and Tasmania.
Cheirurus sp. Middle Devonian: Victoria.
Proetus sp. Devonian: Western Australia.
Phillipsia seminifera, Phillips. Carbopermian: New South Wales.
Phillipsia grandis, Eth. fil. Carbopermian: W. Australia and Queensland.
Griffithides eichwaldi, Waldheim. Carbopermian: New South Wales and Queensland.
Brachymetopus strzelecki, McCoy. Carbopermian: New South Wales.
PHYLLOPODA.
Leaia mitchelli, Eth. fil. Upper Carboniferous: New South Wales.
Estheria coghlani, Cox. Trias: New South Wales.
Estheria minuta, Alberti sp. Trias: New Zealand.
Estheria mangaliensis, Jones. Jurassic: Queensland.
OSTRACODA.
Leperditia sp. Lower Cambrian: S. Australia.
Beyrichia kloedeni, McCoy. Silurian (Yeringian): Victoria.
Beyrichia wooriyallockensis, Chapm. Silurian (Yeringian): Victoria.
Beyrichia maccoyiana, Jones, var. australis, Chapm. Silurian: (Yeringian): Victoria.
Bythocypris hollii, Jones. Silurian (Yeringian): Victoria.
Macrocypris flexuosa, Chapm. Silurian (Yeringian) Victoria.
Primitia reticristata, Jones. Silurian (Yeringian): Victoria.
Leperditia shearsbii, Chapm. Silurian: New South Wales.
Primitia cuneus, Chapm. Middle Devonian: Victoria.
Beyrichia varicosa, T. R. Jones. Carboniferous: Queensland.
Primitia dunii, Chapm. Carbopermian: New South Wales.
Jonesina etheridgei, Chapm. Carbopermian: New South Wales.
Entomis jonesi, de Koninck. Carbopermian: New South Wales.
Synaphe mesozoica, Chapm. sp. Trias: New South Wales.
Cythere lobulata, Chapm. Jurassic: W. Australia.
Paradoxorhyncha foveolata, Chapm. Jurassic: W. Australia.
Loxoconcha jurassica, Chapm. Jurassic: W. Australia.
Cytheropteron australiense, Chapm. Jurassic: W. Australia.
Bairdia amygdaloides, Brady. Cainozoic and living: Victoria.
Cythere senticosa, Baird. Cainozoic. Also living: Victoria.
Cythere clavigera, G. S. Brady. Cainozoic and living: Victoria.
Cytherella punctata, G. S. Brady. Cainozoic and living: Victoria.
Cytherella pulchra, G. S. Brady. Cainozoic and living: Victoria.
CIRRIPEDIA.
Turrilepas mitchelli, Eth. fil. Silurian: New South Wales.
Turrilepas yeringiae, Chapm. Silurian (Yeringian): Victoria.
Lepas pritchardi, Hall. Cainozoic (Janjukian): Victoria.
(?) Pollicipes aucklandicus, Hector sp. Cainozoic (Oamaru Series): New Zealand.
Balanus sp. Cainozoic (Janjukian and Kalimnan): Victoria.
PHYLLOCARIDA.
Rhinopterocaris maccoyi, Eth. fil. sp. Lower Ordovician: Victoria.
Hymenocaris hepburnensis, Chapm. L. Ordovician: Victoria.
Caryocaris marri, Jones and Woodw. L. Ordovician: Victoria and Tasmania.
Caryocaris angusta, Chapm. L. Ordovician: Victoria.
Saccocaris tetragona, Chapm. L. Ordovician: Victoria.
Ceratiocaris cf. murchisoni, Agassiz sp. Silurian: Victoria.
Ceratiocaris pinguis, Chapm. Silurian (Melbournian): Victoria.
Ceratiocaris pritchardi, Chapm. Silurian: Victoria.
Aptychopsis victoriae, Chapm. Silurian (Melbournian): Victoria.
Dithyrocaris praecox, Chapm. Silurian (Melbournian): Victoria.
DECAPODA.
Prosopon etheridgei, H. Woodw. Lower Cretaceous: Queensland.
Ommatocarcinus corioensis, Cresswell sp. Cainozoic (Janjukian): Victoria.
Ebalia sp. Cainozoic (Kalimnan): Victoria.
Harpactocarcinus tumidus, H. Woodw. Cainozoic (Oamaru Series): New Zealand.
Thalassina emerii, Bell. (?) Pleistocene: Queensland and Northern Territory.
EURYPTERIDA.
Pterygotus australis, McCoy. Silurian (Melbournian): Victoria.
INSECTA.
Mesostigmodera typica, Etheridge fil. and Olliff. Jurassic: Queensland.
(?) Cicada lowei, Etheridge fil. and Olliff. Jurassic: New South Wales.
Aeschna flindersensis, H. Woodward. Lower Cretaceous: Queensland.
Chironomus venerabilis, Eth. fil. and Oll. Cainozoic: New South Wales.
Ephemera culleni, Eth. fil. and Oll. Cainozoic: New South Wales.
Palaeolycus problematicum, Eth. fil. and Oll. Cainozoic: New South Wales.
LITERATURE.
TRILOBITES.
McCoy, F. Prod. Pal. Vict., Dec. III. 1876, pp. 13-20, pls. XXII. and XXIII. (Silurian). Hector, J. Trans. N.Z. Inst., vol. IX. 1877, p. 602, pl. XXVII. (Homalonotus). Woodward, H. Geol. Mag., Dec. III. vol. I. 1884, pp. 342-344, pl. XI. (Cambrian). Mitchell, J. Proc. Linn. Soc. New South Wales, vol. II. 1888, pp. 435-440, pl. XI. (Silurian). Foerste, A. F. Bull. Sci. Lab. Denison Univ., vol. III. pt. V. 1888, pp. 122-128, pl. XIII. Etheridge, R. jnr. Proc. Linn. Soc. New South Wales, vol. V. pp. 501-504, pl. XVIII. (Bronteus). Idem, Parl. Papers, Leg. Assemb. S.A., vol. I. No. 23, 1892; ibid., vol. 2, No. 52, 1893 (Asaphus). Id., Geol. Queensland, 1892, pp. 214-216, pls. VII. VIII. and XLIV. (Carboniferous). Id., Proc. R. Soc. Vict., vol. VI. (N.S.), 1894, pp. 189-194, pl. XI. (Bronteus). Id., ibid, vol. VIII. (N.S.), 1896, pp. 56, 57, pl. I. (Dinesus). Id., Rec. Austr. Mus., vol. V. No. 2, 1904, pp. 98-101, pl. X. (Cambrian). Id., Trans. R. Soc. S. Austr., vol. XXII. 1898, pp. 1-3, pl. IV. (Cambrian). Etheridge, R. jnr. and Mitchell, J. Proc. Linn. Soc. New South Wales, vol. VI. 1892, pp. 311-320, pl. XXV.; ibid., vol. VIII. 1894, pp. 169-178, pls. VI. VII.; ibid., vol. X. 1896, pp. 486-511, pls. XXXVIII.-XL.; ibid., vol. XXI. 1897, pp. 694-721, pls. L.-LV. Tate, R. Rep. Horn Exped., 1896, Part 3, Palaeontology, pp. 111, 112, pl. III. De Koninck, L. G. Mem. Geol. Surv. New South Wales, Pal. No. 6, 1898, pp. 36-47 pl. I. (Silurian); pp. 276-281, pl. XXIV. (Carboniferous). Gregory, J. W. Proc. R. Soc. Vict., vol. XIII. (N.S.) pt. II, 1901, pp. 179-182, pl. XXII. (Cyphaspis). Ibid., vol. XV. (N.S.) « 255 » pt. II. 1903, pp. 154-156, pl. XXVI. (Dinesus and Notasaphus.) Chapman, F. Proc. R. Soc. Vict., vol. XXIII. (N.S.), pt. II. 1910, pp. 314-322, pls. LVIII. and LIX. (Cambrian). Ibid., vol. XXIV. (N.S.) pt. II. 1912, pp. 293-300, pls. LXI.-LXIII. (Silurian).
PHYLLOPODA.
Cox, J. C. Proc. Linn. Soc. New South Wales, vol. V., pt. 3, 1881, p. 276 (Estheria). Etheridge, R. jnr. ibid., vol. VII. 1893, pp. 307-310, text fig. (Leaia). Idem, Mem. Geol. Surv. New South Wales, Pal. No. 1, 1888, pp. 6-8, pl. I. (Estheria).
OSTRACODA.
Brady, G. S. in Etheridge, jnr. Geol. Mag., 1876, p. 334 (Cainozoic). De Koninck, L. G. Mem. Geol. Surv. New South Wales, Pal. No. 6, 1898, pp. 33, 36 (Silurian); ibid., pp. 275, 276, pl. XXIV. (Carboniferous). Chapman, F. Proc. R. Soc. Vict., vol. XVI. (N.S.), pt. II. 1904, pp. 199-204, pl. XXIII. (Jurassic). Idem, ibid., vol. XXII. (N.S.), pt. I. 1909, pp. 1-5, pl. I. (Leperditia). Idem, Rec. Geol. Surv. New South Wales, vol. VIII. pt. 4, 1909, pp. 1-3, pl. LIV. (Triassic). Idem, Rec. Geol. Surv. Vict., vol. III. pt. 2, 1912, p. 221, pl. XXXVI. (Primitia). Idem, Proc. R. Soc. Vict., vol. XV. (N.S.), pt. II. 1903, pp. 109-113, pl. XVI. (Beyrichia). Ibid., vol. XVII. (N.S.) pt. I. 1904, pp. 299-312, pls. XIII.-XVII. (Silurian).
CIRRIPEDIA.
Etheridge, R. jnr. Geol. Mag., Dec. III. vol. VII. 1890, pp. 337, 338, pl. XI. (Turrilepas). Hall, T.S. Proc. R. Soc. Vict., vol. XV. (N.S.) pt. I. 1902, pp. 83, 84, pl. XI. (Lepas). Benham, W. B. Geol. Mag., Dec. IV. vol. X. pp. 110-119, pls. IX. X. (? Pollicipes). Chapman, F. Proc. R. Soc. Vict. vol. XXII. (N.S.) pt. II. 1910, pp. 105-197, pls. XXVIII. XXIX. (Turrilepas).
PHYLLOCARIDA.
Etheridge, R. jnr. Rec. Geol. Surv. New South Wales, vol. III. pt. I. 1894, pp. 5-8, pl. IV. (Ordovician). Chapman, F. Proc. R. Soc. Vict. vol. XV. (N.S.), pt. II. 1903, pp. 113-117, pl. XVIII. (Ordovician); ibid., vol. XVII. (N.S.) pt. I. 1904, pp. 312-315, pl. XVII.; ibid., vol. XXII. (N.S.), pt. II. 1910, pp. 107-110, pl. XXVIII. (Silurian). Idem, Rec. Geol. Surv. Vict., vol. III. pt. 2, 1912, pp. 212, 213, pls. XVII. XVIII. (Ordovician).
DECAPODA.
Bell, T. Proc. Geol. Soc. Lond., vol. I. 1845, pp. 93, 94. Text-fig. (Thalassina). Woodward, H. Quart. Journ. Geol. Soc., vol. XXXII. 1876, pp. 51-53, pl. VII. (Harpactocarcinus). Idem., Proc. Linn. Soc. New South Wales, vol. VII. (2), pt. 2, 1892, pp. 301-304 pl. IV. (Prosopon).
Hall, T. S. Proc. R. Soc. Vict., vol. XVII. (N.S.) pt. II. 1905, pp. 356-360, pl. XXIII. (Ommatocarcinus).
EURYPTERIDA.
McCoy, F. Geol. Mag. Dec. IV. vol. VI. 1899, pp. 193, 194, text fig. (Pterygotus).
INSECTA.
Woodward, H. Geol. Mag. Dec. III. vol. I. 1884, pp. 337-339, pl. XI. (Aeschna). Etheridge, R. jnr. and Olliff, A. S. Mem. Geol. Surv. New South Wales, Pal. No. 7, 1890 (Mesozoic and Cainozoic).
FOSSIL FISHES, AMPHIBIANS, REPTILES, BIRDS, AND MAMMALS.
Vertebrates.—
The above-named classes of animals are distinguished from those previously dealt with, by the presence of a vertebral column. The vertebral axis may be either cartilaginous as in some fishes, or bony as in the greater number of animals belonging to this sub-kingdom.
Chordata.—
LINKS BETWEEN THE INVERTEBRATES AND FISHES.—The curious little ascidians or “sea-squirts,” belonging to the group Tunicata, are held by some authorities to be the degenerate descendants of a free-swimming animal having a complete notochord and nerve-tube, structures which are now only seen in the tails of their tadpole-like larvae. The fully developed tunicate is generally sessile and provided with a thick outer coat (tunic) and muscular inner lining. This outer coat in some forms, as Leptoclinum, is strengthened with tiny calcareous spicules, and these are sometimes found in the fossil « 258 » state in Cainozoic clays, as well as in some of the calcareous deep-sea oozes. The little stellate spicules of Leptoclinum are abundant in the Balcombian clays of Mornington, Victoria.
Another primitive form with a notochord is the Lancelet, but this, having no hard parts, is not found in the fossil state.
Primitive Types of Fishes.—
FISHES.—The remains of fishes are naturally more abundant in the fossil condition, owing to their aquatic habits, than those of other vertebrates. The earliest fishes were probably entirely cartilaginous, and some have left only a mere trace or impression on the rocks in which they were embedded. These primitive fishes have no lower jaw, and are without paired limbs. They are sometimes placed in a class by themselves (AGNATHA). The orders of this primitive fish series as represented in Australasia are the Osteostraci (“bony shells”), of which the remains of the Cephalaspis-like head-shield of Thyestes has been found in the Silurian of N.E. Gippsland, Victoria (Fig. 122); and the Antiarchi, with its many-plated cuirass, armoured body-appendages, internal bony tissue, and coarsely tuberculated exterior, as seen in Asterolepis australis, a fossil occasionally found in the Middle Devonian Limestone of Buchan, Gippsland.
True Fishes.—Devonian.—
Of the true fishes (Pisces), the Elasmobranchii (“slit-gills”), a sub-class to which the modern sharks belong, are represented in the Devonian series by the paired spines of a form resembling Climatius, found « 259 » both in Victoria and New South Wales. Remains of Dipnoi (“double-breather” or lung-fishes) occur in the Devonian of Barker Gorge, Western Australia, represented by a new species allied to Coccosteus (“berry-bone” fish); and in a bed of the same age at the Murrumbidgee River, New South Wales by the cranial buckler of Ganorhynchus süssmilchi.


|
Fig. 123. Gyracanthides murrayi, A. S. Woodw. L. Carboniferous. Mansfield, Victoria. (Restoration). About 1/12 nat. size |

Carboniferous Fishes.—
The Lower Carboniferous sandstone of Burnt Creek and other localities near Mansfield, Victoria, contains an abundant fish fauna, associated with stems of Lepidodendron. The slabs of sandstone are often ripple-marked and show signs of tracks and castings of shore-living animals. These deposits were probably laid down in shallow water at the shore margin or in salt lagoons or brackish areas skirting the coast, into which at intervals the remains of the giant lycopods were drifted. The more important of these fish remains are Elasmobranchs, as Gyracanthides murrayi (Fig. 123) and Acanthodes australis; the Dipnoan, Ctenodus breviceps; a Rhizodont or fringe-finned ganoid, Strepsodus decipiens (Fig. 124 A); and a genus related to Palaeoniscus, Elonichthys (E. sweeti, Fig. 124 B, and E. gibbus). The defence spines of Gyracanthides are fairly abundant in the sandstones; whilst on some slabs the large enamelled scales of Strepsodus are equally conspicuous.
From the sandstones of the same age, Lower Carboniferous, in the Grampians of Western Victoria, some small but well-preserved spines belonging to the genus Physonemus have been found associated with a new variety of the well-known European Carboniferous brachiopod, Lingula squamiformis (var. borungensis).
Carbopermian Fishes.—
In the Carbopermian (Gympie Beds) of the Rockhampton District, Queensland, a tooth of a Cochliodont (“snail tooth”) occurs, which has been doubtfully referred to the genus Deltodus (? D. australis). The Cochliodontidae show dentition remarkably like that of the Cestracion or Port Jackson Shark. Another tooth having the same family relationship « 262 » has been referred to Tomodus ? convexus, Agassiz; this is from the Carbopermian of the Port Stephen district of New South Wales. From the Newcastle Coal Measures in New South Wales a Palaeoniscus-like fish, Urosthenes australis has been described.
Carbopermian fish remains are rare in Western Australia. They comprise a wrinkled tooth of Edestus (E. davisii) from the Gascoyne River, belonging to a fish closely related to the Port Jackson shark; and a cochliodont, Poecilodus (P. jonesi, Ag.) from the Kimberley district.
Triassic Fishes.—
Fossil fishes are important and numerous in Australian Triassic beds, especially in New South Wales. At the base of the Hawkesbury or close of the Narrabeen series, the railway ballast quarry near Gosford has yielded an extensive and extremely interesting collection. Near the floor of the quarry there is a band of sandy shale and laminated sandstone 5 feet 9 inches in thickness, and this contains the following genera:—A dipnoan, Gosfordia; and the following ganoids or enamelled scale fishes—Myriolepis, Apateolepis, Dictyopyge, Belonorhynchus, Semionotus, Pristisomus (see antea, Fig. 18), Cleithrolepis (Fig. 125), Pholidophorus and ? Peltopleurus.
Upper Triassic Fishes.—
In the middle of the Wianamatta or Upper Trias Series at St. Peter’s, near Sydney, which contains a fauna described as slightly older in aspect than that of Gosford, having Carbopermian affinities, « 263 » there occur in the hard shale or clay stone the genera Pleuracanthus (a Palaeozoic shark); Sagenodus (a dipnoan related to Ctenodus of the Victorian Carboniferous); and the following ganoids,—Palaeoniscus, Elonichthys, Myriolepis, Elpisopholis, Platysomus and Acentrophorus. From the soft shales were obtained Palaeoniscus, Semionotus, Cleithrolepis and Pholidophorus; an assemblage of genera somewhat comparable with the Gosford fauna.

Lower Mesozoic Fishes.—
From the Lower Mesozoic sandstone (?Triassic) of Tasmania, two species of Acrolepis have been described, viz., A. hamiltoni and A. tasmanicus. The former occurs in the thick bed of sandstone, of nearly « 264 » 1,000 feet, at Knocklofty; the latter species in the sandstone with Vertebraria conformably overlying the Carbopermian at Tinderbox Bay.

Jurassic Fishes.—
The Jurassic beds of Victoria contain three genera. Psilichthys selwyni, a doubtful palaeoniscid was described from Carapook, Co. Dundas; whilst Leptolepis, « 265 » a genus found in the Trias of New South Wales and the Lias and Oolite of Europe, is represented by L. crassicauda from Casterton, associated with the typical Jurassic fern, Taeniopteris. In the Jurassic beds of South Gippsland, at Cape Paterson, an interesting splenial tooth of the mudfish, Ceratodus, was found, named C. avus (Fig. 126). Since then, in a bore-core from Kirrak near the same place a fish scale was discovered (Fig. 127) which, by its shape, size and structure seems to differ in no way from the living lung-fish of Queensland (Fig. 128). It is reasonable to infer that tooth and scale belong to the same species; and in view of the close relationship of the tooth with that of the living mudfish, rather than with that of the Ceratodus found fossil in the Mesozoic of Europe, it may be referred to Neoceratodus, in which genus the living species is now placed.

From the Jurassic beds (Talbragar Series) of New South Wales, an interesting collection of ganoid fishes has been described, comprising Coccolepis australis, Aphnelepis australis, Aetheolepis mirabilis, Archaeomaene tenuis, A. robustus, Leptolepis talbragarensis, L. lowei and L. gregarina (Fig. 129).
Lower Cretaceous Fishes.—
Fish remains are fairly abundant in the Lower Cretaceous of Queensland. They comprise both the sharks and the ganoids. Of the sharks, a specimen, showing seven conjoined vertebrae has been named Lamna daviesii, from the Richmond Downs, Flinders River district; and a tooth referred to Lamna appendiculatus, Agassiz, from Kamileroy, Leichhardt River, N.W. Queensland. The typical Cretaceous genus Corax is represented by a small tooth named C. australis (Fig. 124 C), from the Hamilton River, Queensland, and which closely approaches the tooth of Corax affinis, Agassiz, from the Upper Cretaceous of Europe. Of the ganoid fishes two genera, both members of the family Aspidorhynchidae, have been found in Queensland. Aspidorhynchus sp. and Belonostomus sweeti (Fig. 124 D) have both occurred at Hughenden, Flinders River district. The former genus has a slender body and produced rostrum; in Europe it is more characteristic of Jurassic strata. Belonostomus ranges from the Upper Oolite, Bavaria, « 268 » to the Upper Cretaceous in other parts of the world. Remains of a species of Portheus, one of the predaceous fishes which lived in the Cretaceous period, consisting of a portion of the cranium with the anterior part of the jaws, has been obtained from the Rolling Downs Formation (Lower Cretaceous) near Hughenden, Queensland.
Cretaceous Fishes, New Zealand.—
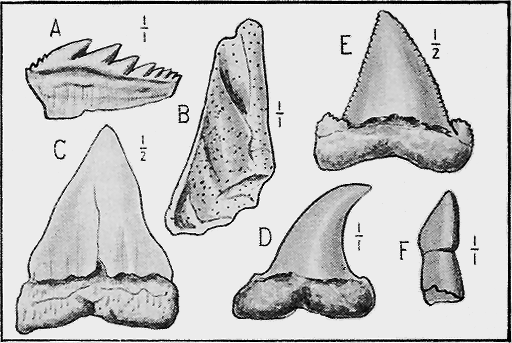
The Cretaceous beds of New Zealand are grouped in ascending order as the Waipara Greensands, the Amuri Limestone and the Weka Pass Stone. In the Waipara beds occur the teeth of Notidanus marginalis « 269 » (Fig. 130 A), and N. dentatus. In the Amuri Limestone N. dentatus is again found, as well as the genus Lamna, represented by L. compressa, Ag. (originally described as L. marginalis, Davis), L. carinata and L. hectori. Two forms of “Elephant fish” are represented by their dental plates, namely Callorhynchus hectori (Fig. 130 B) and Ischyodus thurmanni, Pictet and Campiche (recorded as I. brevirostris, Ag.).
Cainozoic Fishes.—
Fish remains principally consisting of teeth, are common fossils in the Cainozoic beds of southern Australia, particularly in Victoria, and also in New Zealand.
Balcombian Series, Southern Australia.—
The Balcombian beds as seen at Mornington and in the Lower Beds at Muddy Creek, Hamilton, contain the teeth of sharks as Odontaspis contortidens, Lamna crassidens, L. apiculata, Oxyrhina hastalis (rarely), O. minuta, Carcharodon megalodon, and C. robustus.
Janjukian.—
The Janjukian Series (Miocene), represented at Torquay, Waurn Ponds and Table Cape, contains an abundant fish fauna, including amongst sharks, Cestracion cainozoicus, Asteracanthus eocaenicus, Galeocerdo davisi, Carcharoides totuserratus, Odontaspis contortidens, O. incurva, O. cuspidata, Lamna crassidens, L. apiculata (Fig. 130 D), L. compressa, L. bronni, Oxyrhina hastalis (occasional) (Fig. 130 C), O. desori, O. retroflexa, O. minuta, Carcharodon auriculatus (Fig. 130 E), C. megalodon and C. robustus. A species of chimaeroid or Elephant fish « 270 » is represented by a left mandibular tooth named Ischyodus mortoni, from the Table Cape Beds, Tasmania.
The Corio Bay series contains teeth of Acanthias geelongensis, Sphyrna prisca, Odontaspis contortidens, O. attenuata, Oxyrhina minuta, Carcharodon megalodon, amongst sharks; whilst the spine of a Porcupine Fish, Diodon connewarrensis has been obtained from the clays of Lake Connewarre, Victoria.
Kalimnan.—
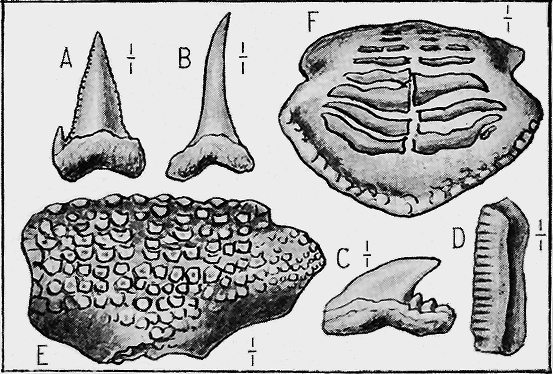
The Kalimnan Series is also prolific in the remains of fishes, the principal localities being Beaumaris and Grange Burn, Hamilton. Amongst the sharks there found are, Notidanus jenningsi (related « 271 » to the Indian Grey Shark), Cestracion cainozoicus (related to the Port Jackson Shark), Asteracanthus eocaenicus, Galeocerdo davisi, G. latidens (Fig. 131 C), G. aduncus, Odontaspis contortidens (Fig. 131 B), O. incurva, O. cuspidata, O. attenuata, Lamna apiculata, L. compressa, Oxyrhina hastalis (abundant), O. desori, O. retroflexa, O. eocaena, O. minuta, Carcharodon auriculatus and C. megalodon. An extinct species of Sting Ray, Myliobatis moorabbinensis (Fig. 131 D), is found at Beaumaris, represented by occasional palatal teeth. Mandibular and palatine teeth of an extinct genus of Elephant Fish, Edaphodon (E. sweeti) are occasionally found at Beaumaris, and at Grange Burn near Hamilton. Two extinct forms of the Wrasse family, the Labridae, are found in Victoria; the pharyngeals of Labrodon confertidens (Fig. 131 E), occurring at Grange Burn, Hamilton, and those of L. depressus, at Beaumaris. The palatal jaws of a Porcupine Fish, Diodon formosus (Fig. 131 F), are frequently met with at the base of the Kalimnan Series, both at Grange Burn and Beaumaris.
Oamaru Series, New Zealand.—
In New Zealand the Oamaru Series, which is comparable in age with the Victorian Janjukian, contains numerous fish remains, chiefly teeth of sharks. These are: Notidanus primigenius, N. marginalis (also occurring in the Waipara Series), Galeocerdo davisi, Odontaspis incurva, O. cuspidata, O. attenuata, Lamna apiculata, L. compressa, Oxyrhina retroflexa, Carcharodon auriculatus, C. megalodon and C. robustus. The teeth of a Sting Ray, Myliobatis plicatilis « 272 » and of a species of Sea-bream, Sargus laticonus, also occur in this series (Fig. 130 F).
Pleistocene.—
A species of fish belonging to the family of the Perches, Ctenolates avus, has been described from freshwater carbonaceous shale of Pleistocene age from Nimbin on the Richmond River, New South Wales.
Amphibians: Their Structure.—
AMPHIBIANS.—This group includes amongst living forms the Frogs, Toads, Newts, and Salamanders. The remains of amphibia are rare in Australasian rocks, and practically limited to the group of the Triassic Labyrinthodonts. The Amphibia are distinguished from Reptiles by certain changes which their young undergo after leaving the egg. In this intermediate stage they breathe by external gills, these being sometimes retained together with the internal lungs in the adult stage. In the older forms of this group the vertebra is of the nature of a notochord, the joints consisting of a thin bony ring with a gelatinous interior. The Labyrinthodontia have a long, lizard-like body, short pectoral limbs as compared with the pelvic, and five-toed feet. The skull is completely roofed over. The teeth are pointed, with a large pulp cavity and wall of infolded or plicated dentine (hence the name labyrinthodont—maze-tooth). The vertebrae are hollow on both sides, sometimes imperfectly ossified, and with a notochordal canal. Ventral aspect with bony thoracic plates. Cranial bones deeply sculptured, and carrying mucus canals.
Carbopermian Labyrinthodonts.—
The genus Bothriceps, probably an Archegosaurian, is represented by two species, B. australis and B. major from New South Wales (Fig. 132). The latter species occurs in the Oil Shale (Carbopermian) of Airly.
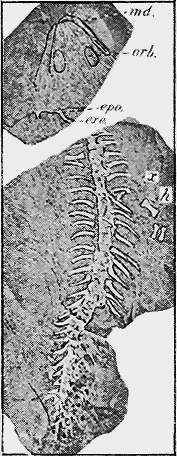
Triassic Labyrinthodonts.—
From the Hawkesbury Series near Gosford, New South Wales, the labyrinthodont, Platyceps wilkinsoni has been described. The skeleton is nearly complete and exposed on the ventral face; the head is « 274 » 27mm. long and 32mm. broad. This specimen is associated with the remains of ganoid fishes, as Palaeoniscus and Cleithrolepis, together with the equisetum-like plant Phyllotheca.
Other, somewhat doubtful remains having similar affinities to the labyrinthodonts are also recorded from the Wianamatta beds (Upper Trias) at Bowral, New South Wales, consisting of a maxilla with teeth and 11 vertebrae with ribs of the left side. Remains of a labyrinthodont, Biloela, supposed to be related to Mastodonsaurus, have been recorded from the Hawkesbury Series of Cockatoo Island, Port Jackson, New South Wales, by W. J. Stephens, and consisting of a pectoral plate compared by that author with M. robustus (now transferred to the genus Capitosaurus).
The only other recorded remains of this group in Australasia are those noted by W. J. Stephens from the Kaihiku Series (Trias) at Nugget Point, Otago; and in the Otapiri Series (Upper Trias) of the Wairoa district, New Zealand.
Reptilia: Their Structure.—
REPTILIA.—The Reptiles are cold-blooded, vertebrated animals, with a scaly skin or armour. Their respiration is essentially by means of lungs, and they are terrestrial or aquatic in habit. The skeleton is completely ossified (bony). Reptiles, although resembling amphibians externally, are more differentiated in structure and of generally larger proportions. They exhibit great diversity of form, especially as regards their extremities. They were even adapted « 275 » for flying, as in the Pterosaurs (“Flying Dragons”) with their membranous wing attached to the anterior limb. The Deinosaurs (“Terrible Reptiles”) were often of great size, exceeding the dimensions of any land mammals, and their limbs were adapted for walking. The marine reptiles, as the Ichthyosauria (“Fish-lizards”) and Sauropterygia (“lizard-finned”) had the limbs transformed into paddles. The neural spines in the vertebra of the Turtles are laterally expanded into a carapace and united with dermal plates. The vertebrae of Reptilia show great variation of form, being biplanate (amphiplatyan), biconcave (amphicoelus), hollow in front (procoelus), or hollow at the back (opisthocoelus). In the case of Reptiles having both pairs of limbs developed, the cervical, dorsal, sacral and caudal regions may be separately distinguished. Amongst the Ophidia (Snakes), Pythonomorpha (“Sea-lizards”) and Ichthyosaurs (“Fish-lizards”) there is no differentiated sacral region. The skull of the Reptiles is nearer that of Birds than Amphibians. The basiocciput (basal bone of the skull at the back) articulates with the atlas (top joint of the backbone) by means of a single condyle (protuberance). All reptiles, with the exception of the Chelonians (Turtles), and a few others, are furnished with teeth: these are formed chiefly of dentine with a layer of enamel.
Dentition.—
Some teeth have solid crowns (pleodont); some grow from persistent pulps (coelodont); socketed teeth (thecodont) are inserted in alveoli; some are fused with the supporting bone along the outer rim or top « 276 » (acrodont); whilst others are developed laterally along the flange-like inner rim of the jaw (pleurodont).
Permian and Triassic Reptiles.—
The history of Reptilia commences in Permian and Triassic times, when they were notably represented by the Theromorphs, Pareiasaurus and Tritylodon in South Africa; the Proterosauria of the European and American Permian and Trias, represented by the lizard-like Palaeohatteria and the dorsally frilled Dimetrodon, with its formidable array of neural spines; also the Rhynchosauria, with their beak-like jaws of the same formations. These two groups constitute the order Rhynchocephalia, which is represented at the present day by the Tuatera of New Zealand.
Triassic Reptile, New Zealand.—
The earliest Australian reptilian record is that of a vertebra of Ichthyosaurus from the Kaihiku Series of Mount Potts, New Zealand (Triassic). This specimen was named I. australis by Hector, but since that species name was preoccupied by McCoy in 1867 it is suggested here that the New Zealand species should be distinguished as I. hectori. The New Zealand occurrence of Ichthyosaurus makes the geological history of the genus very ancient in this part of the world.
Jurassic Reptiles.—
At Cape Paterson, Victoria, in the Jurassic coal-bearing sandstone an extremely interesting discovery was made a few years ago, of the ungual bone (claw) « 277 » of a carnivorous Deinosaur, probably related to Megalosaurus of the European Jurassic and Cretaceous beds (See Fig. 126, 3, 3 A). The presence of an animal like this in Australia points to the former existence of a concomitant terrestrial animal fauna, upon which the deinosaur must have preyed.
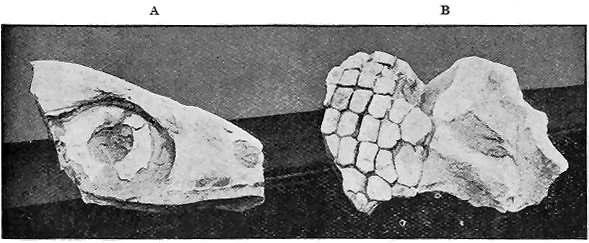
Lower Cretaceous Reptiles.—
The Rolling Downs formation (Lower Cretaceous) of the Thompson and Flinders Rivers in Queensland has yielded remains of a Tortoise, Notochelone costata (see antea, Fig. 17); and the interesting Fish-lizard Ichthyosaurus. Numerous and well preserved remains of I. australis McCoy come from the Flinders River (Fig. 133); whilst I. marathonensis is recorded from Marathon Station, Queensland. The former species is typically represented by a nearly complete skeleton, and was considered by McCoy to « 278 » be one of the largest examples of the genus, since a perfect specimen would probably reach the length of 25 feet. Its teeth resemble those of I. campylodon, Carter, from the English Chalk. Of the Sauropterygia two species of Pliosaurus (P. macrospondylus and P. sutherlandi) have been described from the Lower Cretaceous of the Flinders River; whilst the latter species has also occurred at Pitchery Creek, Central Queensland and at Marathon. P. macrospondylus is distinguished from P. sutherlandi by the roughened edges of the vertebral centra. Another genus of the “lizard-finned” reptiles (Sauropterygia), viz., Cimoliosaurus, occurs in the Upper Cretaceous of White Cliffs, New South Wales (Fig. 134 B, C.)
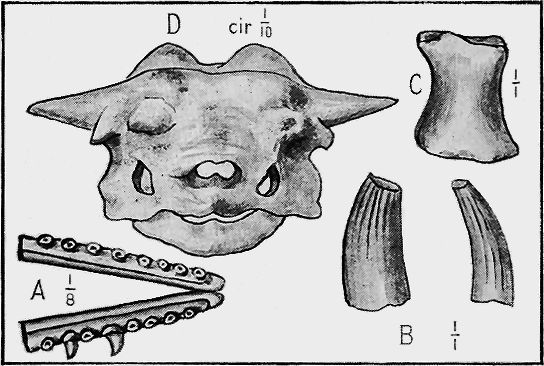
Cretaceous Reptiles, New Zealand.—
The Waipara Series (Cretaceous) of New Zealand contains a fairly large number of reptilian species belonging to several genera among which may be mentioned Plesiosaurus, Polycotylus, and Cimoliosaurus among the Sauropterygia; and Tylosaurus and Taniwhasaurus (Fig. 134 A), marine lizard-like reptiles, belonging to the sub-order Pythonomopha.
Cainozoic and Pleistocene Reptiles.—
The later Cainozoic deposits of Queensland contain remains of Crocodiles referred to Pallymnarchus pollens (from Maryvale Creek) and Crocodilus porosus (from Chinchilla and Arcola, near Brisbane, Queensland). The former species has also occurred at Clunes, whilst Crocodilus porosus is recorded from the Loddon Valley, both in Victoria. Another late Tertiary reptile is the remarkable Horned Turtle, Miolania oweni, which is found in Queensland in Pleistocene deposits (Fig. 134 D), and in the Pliocene (Deep Leads) of Gulgong, New South Wales; whilst a second species of the same genus, M. platyceps, is found in coral sand at Lord Howe Island, 400 miles distant from Australia. This genus has a skull with large bony protuberances, giving it a horned appearance, and the tail is encased in a bony sheath. A species of Miolania is also described from Patagonia. The Cave deposits of Wellington Valley, New South Wales, as well as the fluviatile deposits « 280 » of Queensland, have, yielded the bones of several genera of lizards, including the Giant Lizard (Megalania), which, in its length of 20 feet exceeded that of most living crocodiles.
Birds.—
BIRDS (AVES).—These warm-blooded animals are closely related to Reptiles in many essential particulars; and are generally considered to more nearly approach the Deinosaurs than any other group. The Ratitae (“Raft-breasted” or keel-less birds) and Carinatae (with keeled breast-bones), a sub-class including most modern birds, were probably differentiated before the Cainozoic period.
Jurassic Bird.—
The oldest recorded bird, the remarkable Archaeopteryx, of the Upper Jurassic of Bavaria in Europe, belonging to the Saururae (Reptilian-tailed) is, so far, restricted to the beds of that age.
Miocene Bird, New Zealand.—
The earliest known birds in Australasia occur in the Miocene rocks (Oamaru Series), of New Zealand. In this series, in the Marawhenua Greensands, a Giant Penguin, Palaeeudyptes antarcticus is found at Kakanui near Oamaru, at Curiosity Shop near Christchurch and at Brighton near Nelson, New Zealand: this interesting occurrence shows that these restricted antarctic birds had already become an established type as early as the Miocene.
Victorian Cainozoic Bird.—
The impression of a bird’s feather, probably of a Wader, has lately been described from Western Victoria (see antea Fig. 16 and Fig. 135). This occurs in ironstone, on the surface of which are also impressions of Gum (Eucalyptus) and Native Honeysuckle (Banksia) leaves, of species closely related to those now growing in the same locality. This ironstone is probably of Janjukian age, and may therefore be coincident with the New Zealand occurrence of the Palaeeudyptes in the Oamaru Series.
Pliocene Moa, New Zealand.—
In the Wanganui System (Pliocene) the Putiki Beds have yielded bones of a small Moa (Dinornis), probably the oldest example of the group of great flightless birds which later predominated in New Zealand.
Pleistocene Struthious Birds, Australia.—
Bones of a struthious or Ostrich-like bird, described by Owen under the name of Dromornis australis, a bird as large as the Moa, have been recorded from the Pleistocene of Peak Downs and the Paroo River, Queensland. Indeterminate species of the same genera occur in Phillip Co., New South Wales, and the Mount Gambier Caves, South Australia; whilst Dromaeus patricius is known from King’s Creek, Darling Downs, Queensland.
Genyornis newtoni is an extinct bird allied to the Emeus; it has been found in Pleistocene deposits at Lake Callabonna, South Australia, and other fragmentary remains have been identified by Dr. Stirling and Mr. Zietz from Mount Gambier and Queensland. Regarding the build and habits of Genyornis, those authors remark that “Its legs combine a huge femur nearly as massive, in all but length, as that of Dinornis maximus, and a tibia equalling that of Pachyornis elephantopus with the relatively slender metatarse of Dinornis novae-zealandiae (ingens) and toes which are insignificant beside those of any of the larger moas.”... “In height it may be confidently stated to have been from 6 feet to 6 feet 6 inches, that is if the neck should have been of proportions similar to those of Pachyornis elephantopus.” Those authors also attribute a slow, sluggish habit to the bird, and suggest that herbage rather than roots formed its food. It is very probable that the footprints of birds found in the older dune rock of Warrnambool, Victoria, associated with the doubtful “human footprints” may have been made by Genyornis or a related form.
An extinct Emu, Dromaeus minor, has lately been described from the sub-recent deposits in King Island, Bass Strait.
Pleistocene Carinate Birds, Australia.—
Many genera of carinate birds belonging to living Australian types have been identified by De Vis from the fluviatile deposits on the Darling Downs, Queensland. These include Falcons (Taphaetus and Necrastur); a Pelican (Pelicanus); an Ibis (Palaeopelargus); a Spoonbill (Platalea); Ducks (Anas, Dendrocygna, Biziura and Nyroca); a Darter (Plotus); a Pigeon (Lithophaps); a Ground-pigeon (Progura); a Mound-builder (Chosornis); a Rail (Porphyrio); Moor-hens (Gallinula, Tribonyx and Fulica); and a Stork (Xenorhynchus).
Pleistocene and Holocene Birds, New Zealand.—
In New Zealand numerous remains of birds are found, chiefly in the Pleistocene strata, associated with Moa bones: such are Cnemiornis, the Flightless Pigeon Goose (Fig. 135); Harpagornis, a predatory hawk-like bird larger than any existing eagle; and Aptornis, an extinct Rail. The sand-dunes, peat bogs, swamps, river alluvium, caves and rock shelters of New Zealand often contain numerous remains of the gigantic Moa birds included in the genera Dinornis, Pachyornis and Anomalopteryx, of which perhaps the best known are D. giganteus, D. maximus (Fig. 136), D. robustus, P. elephantopus (Fig. 137), and A. antiqua. Some of the species have become so recently extinct that remains of their skin and feathers have been preserved in fissures in « 284 » the rocks where they were shielded from the influence of air and moisture. The remains of Moa birds are very abundant in some of the localities as at Hamilton in Southland, where, as Hutton estimated, the remains of at least 400 birds were contained within a radius of 25 feet.

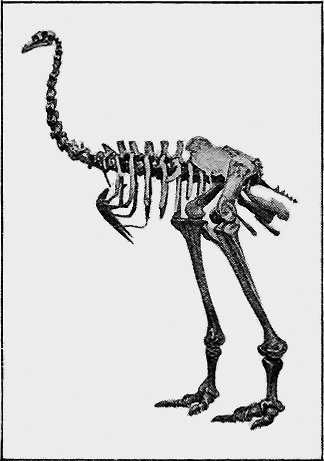
Mammalia: Early Types.—
MAMMALIA.—The history of those warm-blooded animals, the mammals, commences in the early part of the Mesozoic period. It was then that the skull began to assume the characters seen in the modern quadrupeds, « 286 » and their well-formed limb-bones, and fusion of the three bones on each side of the pelvic arch to form the innominate bone, also show relationship to the later types. The earliest ancestral mammalian forms seem to be related to the theromorphic reptiles, predominant in the Permian and Trias. The mammals first to make their appearance were probably related to those of the Monotreme and Marsupial orders. More nearly related to the former is the group of mammals of the Mesozoic period, the Multituberculata.
Multituberculata.—
This group comprises the Triassic Tritylodon (South Africa and Germany); the Upper Jurassic Bolodon (England and United States); the Upper Jurassic to Lower Cainozoic Plagiaulax (England, United States and France); and the Lower Eocene Polymastodon (New Mexico). The molar teeth are ridged longitudinally, and carry numerous tubercles, hence the name of the group, and resemble the deciduous teeth of the Duck-billed Platypus (Ornithorhynchus).
Monotremata.—
The Monotremata are represented at the present day in Australia and New Guinea by the Echidna or Spiny Anteater, and by the Ornithorhynchus or Duck-billed Platypus of Eastern Australia and Tasmania. These egg-laying mammals show relationship towards the reptiles both in structure and in methods of reproduction.
A Pliocene species of Ornithorhynchus (O. maximus) has been recorded from the Deep-leads of Gulgong, « 287 » New South Wales, and the same beds have yielded the remains of Echidna (Proechidna) robusta. Remains of another species, Echidna, (P.) oweni, have been described from the Pleistocene Cave-breccias of the Wellington Valley Caves, New South Wales; and Ornithorhynchus agilis is found in deposits of similar age in Queensland.
Marsupials.—
The Marsupials or pouched mammals belong to the sub-class Metatheria. They are divided into Diprotodontia and Polyprotodontia, accordingly as they possess a single pair of incisor teeth in the lower jaw, or many front teeth, hence the names of the two sub-orders. A later classification of the Marsupials is that of their division into syndactyla and diadactyla.
The diadactyla have the second and third toes separate, and are represented by the family Dasyuridae or Native Cats. These are polyprotodont. They are the most archaic of the marsupial group. Remains of Dasyurus, both of extinct and still living species are found in Pleistocene Cave-breccias in Victoria and New South Wales. The Tasmanian Devil (Sarcophilus ursinus) (Fig. 138, 139) and the Tasmanian Wolf (Thylacinus cynocephalus), still living in Tasmania, have left numerous remains on the mainland, in Victoria and New South Wales. Of the latter genus an extinct species is T. major from the Pleistocene of Queensland (Fig. 140).
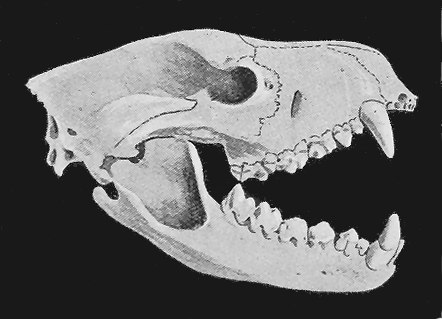
The syndactyla have the second and third toes enclosed in a common skin. The Peramelidae and the Notoryctidae are polyprotodont. The remainder are « 289 » all diprotodont. The Peramelidae or Bandicoot family are represented in Pleistocene Cave-breccias in New South Wales by the genera Peragale and Perameles.
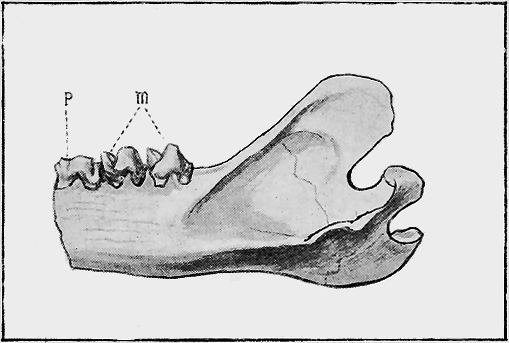
Pleistocene Diprotodonts.—
Pleistocene remains of the diprotodont forms of this syndactylous group are Phascolomys (the Wombat), perhaps ranging as low as Upper Pliocene (P. pliocenus) (Fig. 141); Phascolonus (P. gigas) (Fig. 142 A)[4], a large Wombat from Queensland and New South Wales and South Australia; the Giant Kangaroos, as Macropus titan (Queensland, New South Wales, Victoria and South Australia), Procoptodon goliah (Queensland, New South Wales and Victoria), Sthenurus atlas (New South Wales, Queensland, Victoria and South Australia), Palorchestes azael (Victoria, New South Wales and Queensland); also the great Diprotodon, the largest known marsupial, as large as, and rather taller than, a rhinoceros, found in almost every part of Australia, with an allied form referred to Nototherium occurring also in Tasmania (Figs. 143, 144, 145). Nototherium (Queensland, South Australia and Victoria), was a smaller animal than Diprotodon, with a shorter and broader skull and similar dentition. Remains of the extinct “Marsupial Lion,” Thylacoleo carnifex, an animal allied to the phalangers, have been found in Cave-deposits in New South Wales, Queensland, Victoria and Western Australia. Incised bones of other animals, which are believed to have been gnawed by Thylacoleo, have been found associated with its remains. Thylacoleo possessed a peculiar dentition, the first pair of incisors in the upper jaw being very large and trenchant, whilst the canine and two anterior premolars are small and functionless: the lower jaw has also a pair of large first incisors, behind which are two small premolars, and an enormous chisel-edged last premolar biting against a similar tooth in the upper jaw (Fig. 146).
[4] This genus was described by Owen in 1872 as a sub-genus of Phascolomys founded on some cheek-teeth; and subsequently, in 1884, the same author described some incisors under the name of Sceparnodon ramsayi, which are now known to belong to the same animal that bore the cheek-teeth.

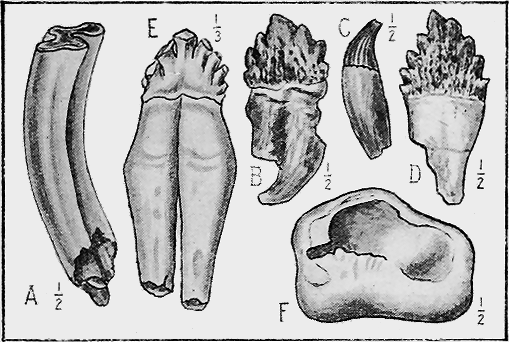

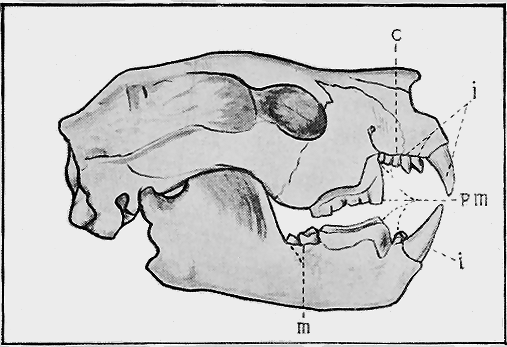
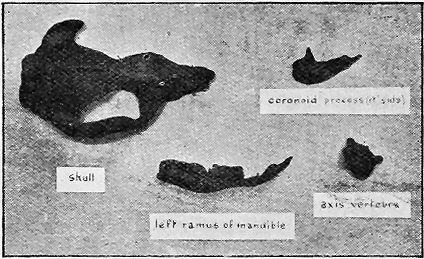
Oldest Known Marsupial.
The oldest marsupial found in Australia is probably Wynyardia bassiana (Fig. 147), whose remains occurred in the Turritella-bed at Table Cape, which is either of Miocene or Lower Pliocene age. This stratum occurs above the well-known Crassatellites-bed (Miocene) of that locality. So far as can be gathered from its incomplete dentition, Wynyardia represents an annectant form between the Diprotodonts and the Polyprotodonts.
Pleistocene Genera, also Living.—
Besides the genera above enumerated, many other marsupials of well-known living species are represented by fossil remains in Cave-deposits and on “sand-blows” in most of the Australian States. The genera thus represented in the Pleistocene deposits of Australia are Bettongia (Prehensile Rat-Kangaroo); Dasyurus (Native Cat); Hypsiprymnus (Rat-Kangaroo); Macropus (Kangaroo); Perameles (Bandicoot); Petaurus (Flying Phalanger); Phalanger (Cuscus); Phascolomys (Wombat); Sarcophilus (Tasmanian Devil); Thylacinus (Tasmanian Wolf).
Cetacea.—
The order Cetacea includes Whales, Dolphins and Porpoises. The earliest known forms belong to the sub-order Archaeoceti, and whilst absent from Australian deposits, are found in the Eocene of Europe, Northern Africa and North America.
Odontoceti: Toothed Whales.—
Remains of Cetacea are first met with in Australian rocks in the Oligocene (Balcombian) of Victoria. At Muddy Creek near Hamilton fragments of ribs and other bones of cetacea, not yet determined, occur in the tenacious blue clays of the lower part of the Clifton Bank section. In Australia and New Zealand the oldest determinable remains of this order belong to the Odontoceti, members of which range from Miocene to Pliocene. Teeth of the toothed whales like Squalodon of the Miocene of France and Bavaria have been found in New Zealand (Kekenodon); in South Australia (Metasqualodon); and in Victoria (Parasqualodon). In Victoria the « 296 » teeth of Squalodontidae occur in the Janjukian beds of Cape Otway, Waurn Ponds and Torquay, represented by molars and anterior teeth of Parasqualodon wilkinsoni (Fig. 142 B, C). The same species also occurs at Table Cape, Tasmania, in beds of similar age. Teeth of Metasqualodon harwoodi (Fig. 142 D) occasionally occur in the white polyzoal rock of the Mount Gambier district, South Australia. The gigantic toothed whale, Kekenodon onamata (Fig. 142 E) occurs in the Marawhenua Greensands (Oamaru Series) at Waitaki Valley, Waihao, Ngapara, Waikouaiti and Milburn in New Zealand. The molar teeth of this striking species, with their serrated crowns, measure nearly five inches in length.
Ear-bones of Whales.—
The tympanic bones of whales are not uncommon in the Janjukian beds of Waurn Ponds, near Geelong, Victoria; and they are occasionally found in the basement bed of the Kalimnan at Beaumaris, Port Phillip. In the absence of any distinctive generic characters they have been referred to the quasi-genus Cetotolithes (Fig. 142 F). McCoy has expressed the opinion that they may perhaps be referable to the ziphioid or beaked whales, for undoubted remains of that group, as teeth of Ziphius geelongensis, occur in these same beds; as well as portions of their rostrate crania, in the Kalimnan basement beds at Grange Burn, near Hamilton. The large curved and flattened teeth of Ziphius (Dolichodon) geelongensis are occasionally found, more or less fragmentary, in the polyzoal rock of Waurn Ponds.

Kalimnan-Scaldicetus.—
From the Kalimnan Series (Lower Pliocene) of Beaumaris, Port Phillip, there was described a short time since, a remarkably well preserved specimen of Scaldicetus tooth belonging to a new form, S. macgeei (Fig. 148). Another species of the genus, with teeth of a slender form, has been found in the same geological series, at Grange Burn, near Hamilton. In only one other locality besides Australia does the genus « 298 »occur, viz., at Antwerp, Belgium, in Crag deposits of Lower Pliocene age.
Sirenia.—
The order Sirenia (Manatees and Dugongs) is represented in the Australian Pleistocene by Chronozoön australe. The remains consist of the parietal and upper part of the occipital bones of the skull, and were discovered in the fluviatile deposits on the Darling Downs, Queensland. This fossil skull, according to De Vis, had a shallower temporal fossa and feebler masticating muscles, as well as a less highly developed brain than the existing Dugong.
Carnivora.—
The order Carnivora is represented in Australia by the Native Dog or Dingo (Canis dingo). It is by no means a settled question whether the Dingo can boast of very great antiquity. The evidence of its remains having been found under volcanic tuff beds in Victoria is not very convincing, for the original record does not indicate the precise position where the bones were found. The fact of the remains of the Dingo having been found in Cave deposits often associated with extinct marsupials, goes a good way to prove its antiquity. McCoy was strongly inclined to the view of its Pleistocene age, and points out that it shows cranial characters intermediate between the Dogs of South America and the Old World. Fossil remains of the Dingo, associated with Pleistocene mammalian forms have been recorded from the Wellington, Valley Caves, New South Wales; from the Mount Macedon Cave, near « 299 » Gisborne; and in the neighbourhood of Warrnambool, Western Victoria.
Pinnipedia.—
Of the fin-footed Carnivores or Seals and Walruses, the earliest Australasian record is that of the remains of a small seal in the Okehu shell-beds near Wanganui, found in association with the bones of a small Moa-bird (Dinornis).
Newer Pliocene Seal.—
This seal was referred by Hector to Arctocephalus cinereus, a species synonymous, however, with the widely distributed living Seal, Otaria forsteri, Lesson, of the Southern Ocean. Another and larger species of eared seal allied to the living Fur Seal, Otaria forsteri, occurs in Victoria.
Pleistocene Seal.—
This fossil was named Arctocephalus williamsi by McCoy, and was found in Pleistocene deposits at Queenscliff, Port Phillip, at 5 feet below the surface, in marl and sand stone overlain with limestone. Although referred at the time of description to the Pliocene, it has since been proved that at this locality there is a considerable thickness of practically sub-recent material which is more accurately classed with the Pleistocene. Similar remains of eared seals are not uncommon in the Pleistocene deposits of the Otway Coast.
Subrecent Human Remains.
On turning to the occurrence of “human fossils” in Australia we find the geological evidence for any great antiquity of man on this continent to be very « 300 » scanty and inconclusive. This does not, however, imply that man’s existence in Australia will not eventually be proved to date back far beyond the period of the “kitchen middens” of modern aspect, such as are now exposed on the slopes behind the sea-beaches, and on the inland camping grounds. Almost all the records of Australian human remains that have been found in other than ordinary burial places, have proved to be of comparatively recent date. For example, the partially lime-encrusted body found in the cave in the Mosquito Plains, north of Penola, South Australia, recorded by Tenison Woods, is that of an aborigine who, in the early days of settlement, crawled into the cave in a wounded condition. Other occurrences of human remains in caves, but of fairly recent date are, a child’s skull found in a small cave at Bungonia, Co. Argyle, New South Wales, recorded by Etheridge; and the non-petrified limb-bones found in a cave at Wellington, New South Wales, recorded by Krefft, which were probably washed in from the surface in recent times. As regards the former, in Western Australia, as observed by Froggatt, the natives at the present time seek shelter in caves, where these occur, instead of building mia-mias.
A more interesting, because probably much older, occurrence of human remains has been described by Etheridge and Trickett from one of the Jenolan Caves (Skeleton Cave); and those authors conclude from "The great lapse of time that must have accrued to enable the changes already outlined to have taken place since the introduction of the « 301 » remains into the Skeleton Cave," that these remains are ancient.
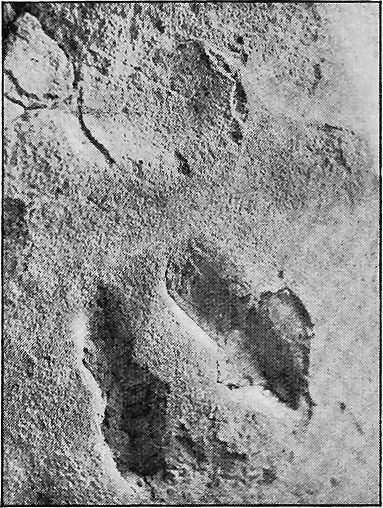
Curious footprints supposed to resemble impressions of human feet with accompanying impress as if made by natives seated, have been long known from the older sand-dune rock of Warrnambool. They were found at Kellas’ Quarry, on the Port Fairy Road in 1890 and at a depth of 54 feet. In November, 1912, a further discovery of similar footprints « 302 » were found at Messrs. Steere Bros.' Quarry, Warrnambool, at a depth of 10 feet, as a block of stone was being removed for building purposes. These footprints are even more obscure than those previously found, and it would be unsafe to affirm their human origin, although they are suggestive of such. Their antiquity is certainly great, since the lavas and tuffs of the Tower Hill district are found overlying this old dune-rock. Other footprints associated with these resemble those of the Dingo and a gigantic bird, possibly like Genyornis.
Probable Origin of Aborigines.—
Ethnology appears to throw more light upon the subject than does geology. Australia has in the past been peopled by two distinct types of man. (1), the ancestors of the Tasmanians, now alas, extinct, who according to some authorities came by way of Australia from Papua through the Malay Peninsula, passing over to Tasmania from the mainland before the separation caused by the subsidence of the Bass Strait area; and who were represented by a negroid or woolly-haired type: (2), the present aboriginals of Australia, showing affinities with the Dravidians of Southern India, a primitive race from whose original stock the white Caucasian races of Europe were derived. By intermarriage with a negroid race like the Melanesian, it is supposed that the black Caucasian gave rise to the present Australian mixed aboriginal type, with negroid features, but possessing the long black hair and keener intellect of the “melanochroi,” as the dark Eurasian stock was termed by Huxley.
Aboriginal Implements.—
The stone implements fashioned by the Tasmanian aboriginals were roughly chipped and of primitive type, of such forms as used at the present day by the Bushmen of South Africa, and representing the eoliths and palaeoliths of early man in the south of England. The implements of the Australian aboriginals on the other hand include besides these both flakes and worked and polished tools, such as were produced by the Neolithic men of Europe, as contrasted with the typically rough palaeolithic tools of the Tasmanian, who never grooved his axes for hafting as did the Australian aboriginal. According to some authorities the Tasmanians represent palaeolithic or even eolithic man in the character of their implements; whilst the Australian resembles the Middle or Mousterian stage of early man in certain of their ethnological characters and in the forms of their implements, although a marked exception is seen in their manufacture of polished adzes, of the neolithic period and in the use of bone implements such as were used in Europe in Upper Palaeolithic times. So far no human remains or handiwork in the form of chipped implements have been found in other than superficial deposits, either in Tasmania or Australia. The incised bone-fragment found near Ballarat, in a bed of silt beneath a sheet of basalt which flowed from Mount Buninyong, is believed by some to be evidence of man’s handiwork in the early Pleistocene, though by others thought to have been cut by the teeth of the “marsupial lion” (Thylacoleo). A stone axe of basalt, grooved for the purpose of « 304 » mounting in a handle, was found in gravel at Ballarat at a depth of 22 inches from the surface. This, however, is no proof of man’s antiquity, for superficial deposits of much greater depth are easily accumulated within a short period. Another implement was found at Maryborough in Queensland in gravels at a depth of 4 feet from the surface, but not below the basalt of the main lead. In this case it is believed that the implement may have fallen into a natural hollow or wombat-burrow. A bone pointer, such as used by native medicine men, was some years ago found buried in the Miocene marls of Waurn Ponds near Geelong. Its presence in so old a rock is easily explained from the fact that in the aboriginal ceremonies the pointer was buried after the incantations. Seeing the difficulties in the way of discovering reliable occurrences of man’s handiwork in isolated examples amongst the older superficial deposits of silt and gravels, the ancient sand-dunes of Victoria, which date back at least to Upper Pliocene, should afford favourable conditions for the preservation of any really ancient kitchen middens, did such exist. Moreover, these deposits would have been less liable to disturbance when once they were covered, than the inland deposits, for the former are now consolidated into a tolerably hard stone.
Antiquity of Man in Australia.—
A strong argument in favour of a considerable antiquity for man in Australia is the fact that the dialects are many, and marriage and tribal customs more complex and intricate than would be found « 305 » in a comparatively recent primitive race. In any case, it is quite possible, if not probable, that man was in southern Australia before the termination of the last phase of volcanic activity, since the tuff beds of Koroit, for example, are quite modern and were laid down on a modern sea-beach strewn with shells identical in species and condition with those now found thrown up in the vicinity at high tide. This view is quite compatible with the occurrence of dingo remains (assuming this animal was introduced by man) in cave deposits in Australia, associated with extinct forms of marsupials.
COMMON OR CHARACTERISTIC FOSSILS OF THE FOREGOING CHAPTER.
FISHES.
Thyestes magnificus, Chapman. Silurian: Victoria.
Asterolepis australis, McCoy. Middle Devonian: Victoria.
Ganorhynchus süssmilchi, Etheridge fil. Devonian: New South Wales.
Gyracanthides murrayi, A. S. Woodward. Lower Carboniferous: Victoria.
Acanthodes australis, A. S. Woodward. Lower Carboniferous: Victoria.
Ctenodus breviceps, A. S. Woodward. Lower Carboniferous: Victoria.
Strepsodus decipiens, A. S. Woodward. Lower Carboniferous: Victoria.
Elonichthys sweeti, A. S. Woodward. Lower Carboniferous: Victoria.
Physonemus micracanthus, Chapman. Lower Carboniferous: Victoria.
(?) Deltodus australis, Eth. fil. Carbopermian: Queensland.
Tomodus (?) convexus, Agassiz. Carbopermian: New South Wales.
Edestus davisii, H. Woodward. Carbopermian: W. Australia.
Peocilodus jonesi, Agassiz. Carbopermian: W. Australia.
Gosfordia truncata, A. S. Woodw. Triassic: New South Wales.
Myriolepis clarkei, Egerton. Triassic: New South Wales.
Apateolepis australis, A. S. Woodw. Triassic: New South Wales.
Dictyopyge robusta, A. S. Woodw. Triassic: New South Wales.
Belonorhynchus gigas, A. S. Woodw. Triassic: New South Wales.
Semionotus australis, A. S. Woodw. Triassic: New South Wales.
Pristisomus latus, A. S. Woodw. Triassic: New South Wales.
Cleithrolepis granulatus, Egerton. Triassic: New South Wales.
Pholidophorus gregarius, A. S. Woodw. Triassic: New South Wales.
Pleuracanthus parvidens, A. S. Woodw. Upper Trias: New South Wales.
Sagenodus laticeps, A. S. Woodw. Upper Trias: New South Wales.
Palaeoniscus crassus, A. S. Woodw. Upper Trias: New South Wales.
Elonichthys armatus, A. S. Woodw. Upper Trias: New South Wales.
Elpisopholis dunstani, A. S. Woodw. Upper Trias: New South Wales.
Pholidophorus australis, A. S. Woodw. Upper Trias: New South Wales.
Psilichthys selwyni, Hall. Jurassic: Victoria.
Leptolepis crassicauda, Hall. Jurassic: Victoria.
Ceratodus avus, A. S. Woodw. Jurassic: Victoria.
Coccolepis australis, A. S. Woodw. Jurassic: New South Wales.
Aphnelepis australis, A. S. Woodw. Jurassic: New South Wales.
Aetheolepis mirabilis, A. S. Woodw. Jurassic: New South Wales.
Archaeomaene tenuis, A. S. Woodw. Jurassic: New South Wales.
Leptolepis talbragarensis, A. S. Woodw. Jurassic: New South Wales.
Lamna daviesii, Eth. fil. Lower Cretaceous: Queensland.
Lamna appendiculatus, Agassiz. Lower Cretaceous: Queensland.
Corax australis, Chapm. Lower Cretaceous: Queensland.
Aspidorhynchus sp. Lower Cretaceous: Queensland.
Belonostomus sweeti, Eth. fil. and A. S. Woodw. Lower Cretaceous: Queensland.
Portheus australis, A. S. Woodw. Lower Cretaceous: Queensland.
Cladocyclus sweeti, A. S. Woodw. Lower Cretaceous: Queensland.
Notidanus marginalis, Davis. Cretaceous: New Zealand.
Lamna compressa, Agassiz. Cretaceous: New Zealand.
Callorhynchus hectori, Newton. Cretaceous: New Zealand.
Ischyodus thurmanni, Pictet and Campiche. Cretaceous: New Zealand.
Odontaspis contortidens, Agassiz. Cainozoic (Bal. and Janj.): Victoria.
Lamna apiculata, Ag. sp. Cainozoic (Bal. and Janj.): Victoria. Also Cainozoic (Oamaru Series): New Zealand.
Carcharodon megalodon, Agassiz. Cainozoic (Bal. Janj. and Kal.): Victoria. Also Cainozoic (Oamaru Series): New Zealand.
Cestracion cainozoicus, Chapm. and Pritch. Cainozoic (Janj. and Kal.): Victoria.
Asteracanthus eocaenicus, Tate sp. Cainozoic (Janj. and Kal.): Victoria.
Galeocerdo davisi, Chapm. and Pritch. Cainozoic (Janj.): Victoria. Also Cretaceous (Waipara Series) and Cainozoic (Oamaru Series): New Zealand.
Carcharoides totuserratus, Ameghino. Cainozoic (Janj.): Victoria.
Odontaspis incurva, Davis sp. Cainozoic (Janj. and Kal.): Victoria. Also Cainozoic (Oamaru Series): New Zealand.
Oxyrhina retroflexa, Agassiz. Cainozoic (Janj.): Victoria. Also Cainozoic (Oamaru Series): New Zealand.
Carcharodon auriculatus, Blainville sp. Cainozoic (Janj. and Kal.): Victoria.
Acanthias geelongensis, Chapm. and Pritch. Cainozoic (Janj.): Victoria.
Ischyodus mortoni, Chapm. and Pritch. Cainozoic (Janj.): Tasmania.
Notidanus jenningsi, Chapm. and Pritch. Cainozoic (Kal.): Victoria.
Galeocerdo aduncus, Agassiz. Cainozoic (Kal.): Victoria.
Oxyrhina hastalis, Agassiz. Cainozoic (rare in Balc. and Janj., abundant in Kal.): Victoria.
Myliobatis moorabbinensis, Chapm. and Pritch. Cainozoic (Kal.): Victoria.
Edaphodon sweeti, Chapm. and Pritch. Cainozoic (Kal.): Victoria.
Labrodon confertidens, Chap. and Pritch. Cainozoic (Kal.): Victoria.
Diodon formosus, Chapm. and Pritch. Cainozoic (Kal.): Victoria.
Notidanus marginalis, Davis. Cretaceous (Waipara Series); and Cainozoic (Oamaru Series): New Zealand.
Myliobatis plicatilis, Davis. Cainozoic (Oamaru Series): New Zealand.
Sargus laticonus, Davis. Cainozoic (Oamaru Series): New Zealand.
Ctenolates avus, A. S. Woodw. Pleistocene: New South Wales.
Neoceratodus forsteri, Krefft, sp. Pleistocene: New South Wales.
AMPHIBIA.
Bothriceps australis, Huxley. Carbopermian: New South Wales.
Bothriceps major, A. S. Woodw. Carbopermian: New South Wales.
Platyceps wilkinsoni, Stephens. Triassic: New South Wales.
REPTILIA.
Ichthyosaurus hectori, Ch. (nom. mut.). Triassic: New Zealand.
(?) Megalosaurus sp. Jurassic: Victoria.
Notochelone costata, Owen sp. Lower Cretaceous: Queensland.
Ichthyosaurus australis, McCoy. Lower Cretaceous: Queensland.
Ichthyosaurus marathonensis, Eth. fil. Lower Cretaceous: Queensland.
Cimoliosaurus leucoscopelus, Eth. fil. Upper Cretaceous: New South Wales.
Plesiosaurus australis, Owen. Cretaceous: New Zealand.
Polycotylus tenuis, Hector. Cretaceous: New Zealand.
Cimoliosaurus haastii, Hector sp. Cretaceous: New Zealand.
Tylosaurus haumuriensis, Hector sp. Cretaceous: New Zealand.
Taniwhasaurus oweni, Hector. Cretaceous: New Zealand.
Pallymnarchus pollens, De Vis. Pleistocene: Queensland and Victoria.
Crocodilus porosus, Schneider. Pleistocene: Queensland and Victoria.
Miolania oweni, A. S. Woodw. Pliocene (Deep-leads): New South Wales. Pleistocene: Queensland.
Miolania platyceps, Owen. Pleistocene: Lord Howe Island.
Megalania prisca, Owen. Pleistocene: Queensland.
BIRDS.
Palaeeudyptes antarcticus, Huxley. Cainozoic (Oamaru Series): New Zealand.
Dinornis sp. Cainozoic (Petane Series): New Zealand.
Pelecanus proavis, De Vis. Pleistocene: Queensland.
Platalea subtenuis, De Vis. Pleistocene: Queensland.
Anas elapsa, De Vis. Pleistocene: Queensland.
Gallinula strenuipes, De Vis. Pleistocene: Queensland.
Fulica prior, De Vis. Pleistocene: Queensland.
Dromornis australis, Owen. Pleistocene: Queensland and New South Wales.
Dromaeus patricius, De Vis. Pleistocene. Queensland.
Dromaeus minor, Spencer. Pleistocene: King Island.
Genyornis newtoni, Stirling and Zietz. Pleistocene: S. Australia.
Cnemiornis calcitrans, Owen. Pleistocene: New Zealand.
Harpagornis moorei, von Haast. Pleistocene: New Zealand.
Aptornis otidiformis, Owen sp. Pleistocene: New Zealand.
Dinornis giganteus, Owen. Pleistocene and Holocene: N. Id., New Zealand.
Pachyornis elephantopus, Owen sp. Pleistocene and Holocene: S. Id., New Zealand.
Anomalopteryx antiqua, Hutton. Pleistocene: S. Id., New Zealand.
MAMMALIA.
Ornithorhynchus maximus, Dun. Cainozoic (Kalimnan or L. Pliocene): New South Wales.
Echidna (Proechidna) robusta, Dun. Cainozoic (Kalimnan): New South Wales.
Ornithorhynchus agilis, De Vis. Pleistocene: New South Wales.
Echidna (Proechidna) oweni, Krefft. Pleistocene: New South Wales.
Wynyardia bassiana, Spencer. Cainozoic (Kalimnan): Tasmania.
Dasyurus maculatus, Kerr sp. Pleistocene: Victoria and New South Wales. Living: Queensland, New South Wales, Victoria and Tasmania.
Phascolomys pliocenus, McCoy. Cainozoic (Werrikooian): Victoria.
Sarcophilus ursinus, Harris sp. Pleistocene: Victoria and New South Wales. Living: Tasmania.
Thylacinus cynocephalus, Harris sp. Pleistocene: Victoria and New South Wales. Living: Tasmania.
Thylacinus spelaeus, Owen. Pleistocene: Queensland and New South Wales.
Thylacinus major, Owen. Pleistocene: Queensland.
Peragale lagotis, Reid sp. Pleistocene: New South Wales. Living: S. Australia and W. Australia.
Perameles gunni, Gray. Pleistocene: Victoria. Living: Queensland and Victoria.
Phascolomys parvus, Owen. Pleistocene: Queensland.
Phascolonus gigas, Owen. Pleistocene: Queensland, New South Wales and S. Australia.
Macropus titan, Owen. Pleistocene: Queensland, Victoria, New South Wales and S. Australia.
Macropus anak, Owen. Pleistocene: Queensland, S. Australia and New South Wales.
Procoptodon goliah, Owen sp. Pleistocene: Queensland, New South Wales and Victoria.
Sthenurus atlas, Owen sp. Pleistocene: Queensland, New South Wales, Victoria, and South Australia.
Sthenurus occidentalis, Glauert. Pleistocene: W. Australia.
Palorchestes azael, Owen. Pleistocene: Queensland, New South Wales and Victoria.
Diprotodon australis, Owen. Pleistocene: Queensland, New South Wales, Victoria and S. Australia.
Nototherium mitchelli, Owen. Pleistocene: Queensland, S. Australia and Victoria.
Thylacoleo carnifex, Owen. Pleistocene: Queensland, New South Wales, Victoria and W. Australia.
Parasqualodon wilkinsoni, McCoy sp. Cainozoic (Janjukian): Victoria and Tasmania.
Metasqualodon harwoodi, Sanger sp. Cainozoic (Janjukian): S. Australia.
Kekenodon onamata, Hector. Cainozoic (Oamaru Series): New Zealand.
Cetotolithes nelsoni, McCoy. Cainozoic (Janjukian): Victoria.
Ziphius (Dolichodon) geelongensis, McCoy. Cainozoic (Janjukian): Victoria.
Scaldicetus macgeei, Chapm. Cainozoic (Kalimnan): Victoria.
Chronozoön australis, De Vis. Pleistocene: Queensland.
Canis dingo, Blumenbach. Late Pleistocene or Holocene: Victoria.
Otaria forsteri, Lesson. Pliocene (Petane Series): N. Id., New Zealand.
Arctocephalus williamsi, McCoy. Pleistocene: Victoria.
LITERATURE.
FISHES.
Silurian.—Chapman, F. Proc. R. Soc. Vict., vol. XVIII. (N.S.), pt. II. 1906, pp. 93-100, pls. VII. and VIII. (Thyestes).
Devonian.—McCoy, F. Prod. Pal. Vict., Dec. IV. 1876, pp. 19, 20, pl. XXXV. figs. 7, 7a, 7b (Asterolepis). Etheridge, R. jnr. Rec. Austr. Mus., vol. VI. pp. 129-132, pl. XXVIII. (Ganorhynchus).
Carboniferous and Carbopermian.—Woodward, H. Geol. Mag., Dec. III. vol. III. 1886, pp. 1-7, pl. I. (Edestus). Etheridge, R. jnr. Geol. and Pal. Queensland, 1892, p. 296, pl. XXXIX. fig. 1 (Deltodus). De Koninck, L. G. Mem. Geol. Surv. New South Wales, Pal. No. 6, 1898, p. 281, pl. XXIV., fig. 11 (Tomodus). Woodward, A. S. Mem. Nat. Mus. Melbourne, No. 1. 1906 (Mansfield Series).
Triassic.—Johnston, R. M. and Morton, A. Proc. R. Soc. Tasmania (1889), 1890, pp. 102-104; ibid. (1890), 1891, pp. 152-154 (Acrolepis). Woodward, A. S. Mem. Geol. Surv. New South Wales, Pal. No. 4, 1890 (Gosford). Ibid. No. 10, 1908 (St. Peters).
Jurassic.—Woodward, A. S. Mem. Geol. Surv. New South Wales, Pal. No. 9, 1895. Id., Ann. Mag. Nat. Hist., Ser. VII. Vol. XVIII. 1906, pp. 1-3, pl. I. (Ceratodus). Hall, T. S. Proc. R. Soc. Vict. vol. XII. (N.S.) pt. II. 1900, pp. 147-151, pl. XIV. Chapman, F. Rec. Geol. Surv. Vict. vol. III. pt. 2, 1912, pp. 234-235, pl. XXXIX. (Ceratodus).
Cretaceous.—Etheridge, R. jnr. Proc. Linn. Soc. New South Wales, vol. III. ser. 2, 1889, pp. 156-161, pl. IV. Idem, Geol. and Pal. Queensland, 1892, pp. 503-504. Davis, J. W. Trans. R. Dubl. Soc. vol. IV. ser. 2. 1888, pp. 1-48, pls. I.-VII. (Cretaceous and Cainozoic of New Zealand). Etheridge, R. jnr. and Woodward, A. S. Trans. R. Soc. Vict., vol. II. pt. II. 1892, pp. 1-7, pl. I. (Belonostomus). Woodward, A. S. Ann. Mag. Nat. Hist., ser. 6, vol. XIX. « 312 » 1894, pp. 444-447, pl. X. (Portheus and Cladocyclus). Chapman, F. Proc. R. Soc. Vict., vol. XXI. (N.S.), pt. II. 1909, pp. 452, 453 (Corax).
Cainozoic.—McCoy, F. Prod. Pal. Vict., Dec. II. 1875, pp. 8-10, pl. XI. (Carcharodon). Chapman, F. and Pritchard, G. B. Proc. R. Soc. Vict., vol. XVII. (N.S.), pt. I. 1904, pp. 267-297, pls. V.-VIII. Idem, ibid, vol. XX. (N.S.), pt. I. 1907, pp. 59-75, pls. V.-VIII. See also Davis, J. W. (Cretaceous).
Pleistocene.—Etheridge, R. jnr. Geol. and Pal. Queensland, 1892, p. 646 (Neoceratodus). Woodward, A. S. Rec. Geol. Surv. New South Wales, vol. VII. pt. 2, 1902, pp. 88-91, pl. XXIV. (Ctenolates).
AMPHIBIA.
Huxley, T. H. Quart. Journ. Geol. Soc., vol. XV. 1859, pp. 647-649, pl. XXII. figs. 1, 2 (Bothriceps). Stephens, W. J. Proc. Linn. Soc. New South Wales, ser. 2. vol. I. 1886, pp. 931-940. Ibid., 1887, pp. 1175-1182, pl. XXII. Ibid., vol. II. 1887, pp. 156-158. Woodward, A. S. Rec. Geol. Surv. New South Wales, vol. VIII. pt. 4, 1909, pp. 317-319, pl. LI. (Bothriceps).
REPTILIA.
Jurassic and Cretaceous.—Hector, J. Trans. N.Z. Inst., vol. VI. 1874, pp. 333-358.
Cretaceous.—McCoy, F. Proc. R. Soc. Vic., vol. VIII. pt. I. 1868, p. 42 (Plesiosaurus). Ibid., vol. IX. pt. II. 1869, p. 77 (Ichthyosaurus). Owen, R. Geol. Mag., Dec. I. vol. VII. 1870, pp. 49-53, pl. III. (Plesiosaurus). Id., Quart. Journ. Geol. Soc. vol. XXXVIII. 1882, pp. 178-183 (“Notochelys” = Notochelone). Etheridge, R. jnr. Proc. Linn. Soc. New South Wales, ser. 2, vol. III. 1889, pp. 405-413, pls. VII. and VIII. (Ichthyosaurus). Id., Geol. and Pal. Queensland, 1892, pp. 505-510. Hutton, F. W. Trans. N.Z. Inst. vol. XXVI. 1894, pp. 354-358, 1 pl. (Cimoliosaurus).
Pleistocene.—Etheridge, R. jnr. Rec. Geol. Surv. New South Wales, vol. I. pt. 3, 1889, pp. 149-152 (Miolania). Id., Geol. and Pal. Queensland, 1892, pp. 647-653.
AVES.
Miocene.—Huxley, T. H. Quart. Journ. Geol. Soc. vol. XV. 1859, pp. 670-677. Also Hector, J. Trans. N.Z. Inst. vol. IV. 1872, pp. 341-346, 1 pl. (Palaeeudyptes). Chapman, F. Proc. R. Soc. Vict. (N.S.) pt. I. 1910, pp. 21-26, pls. IV. and V.
Pleistocene and Holocene.—Von Haast, J. Trans. N.Z. Inst., vol. IV., 1872, pp. 192-196; and vol. VI. 1874, pp. 62-75 (Harpagornis). Owen, R. Memoirs on the Extinct Wingless Birds of New Zealand, London, 1879, 2 vols. De Vis, C. W. Proc. R. Soc. Queensland, vol. VI. pt. I. 1889, pp. 6-8. Id., Proc. Linn. Soc. New South Wales, vol. III. ser. 2, 1888, pp. 1277-1292, pls. XXXIII.-XXXVI. (Carinatae). Etheridge, R. jnr. Rec. Geol. Surv. New South Wales, vol. I. pt. 2, 1889, pp. 126-136, pls. XI.-XIII. (Dromornis). Id., Geol. and Pal. Queensland, 1892, pp. 653-663. Hutton, F. W. Trans. N.Z. Inst., vol. XXIV. 1892, pp. 93-172 (Moas). Id., ibid., vol. XXV. 1893, pp. 14-16, 1 pl. (Anomalopteryx). Id., ibid., vol. XXIX. 1897, pp. 441-557, figs. (Moas). Id., ibid., vol. XXXVIII. 1906, pp. 66 and 67 (Emeus crassus). Hamilton, A. Ibid, vol. XXVI. 1894, pp. 227-257 (Bibliography of Moas). Ibid, vol. XXX. 1898, pp. 445 and 446 (Euryapteryx). Stirling, E. C. and Zietz, A. H. C. Mem. R. Soc. S. Austr., vol. I. pt. II. 1900, pp. 41-80, pls. XIX.-XXIV. (Genyornis). Spencer, W. B. Vict. Nat. vol. XXIII. 1906, pp. 139 and 140; also Spencer, W. B. and Kershaw, J. A. Mem. Nat. Mus. Melbourne No. 3, 1910, pp. 5-35, pls. I.-VII. (Dromaeus minor).
MAMMALS.
Huxley, T. H. Quart. Journ. Geol. Soc., vol. XV. 1859, pp. 676-677 (Phocaenopsis). McCoy, F. Prod. Pal. Vict., Dec. I. 1874, pp. 21, 22, pls. III.-V. (Phascolomys). Ibid, Dec. II. 1875, pp. 7-8, pl. XI. and Dec. VI. 1879, pp. 20 and 21, pl. LV. (Squalodon). Ibid, Dec. III. 1876, pp. 7-12, pl. XXI. (Thylacoleo). Ibid, Dec. IV. 1876, pp. 7-11, pl. XXXI.-XXXIII. (Diprotodon). Ibid, Dec. V. 1877, pp. 7-9, pl. XLI. and XLII. (Arctocephalus). Ibid, Dec. VI. 1879, pp. 5-7, pl. LI. (Macropus): pp. 9-11, pl. LI.-LIII. (Procoptodon): pp. 13-17, pl. LIV. (Cetotolithes); pp. 19 and 20, pl. LV. (Physetodon). Ibid, Dec. VII. 1882, pp. 7-10, pl. LX. (Canis dingo): pp. 11-13, pl. LXXII. and LXII. (Sarcophilus): pp. 23-26, pl. LIX. (Ziphius). Owen, R. Extinct Mammals of Australia, London 1877, 2 vols. Hector, J. Trans. N.Z. Inst., vol. XIII. 1881, pp. 434-436, 1 pl. (Kekenodon). Lydekker, R. Cat. Foss. Mammalia, Brit. Mus. part V. 1887. Id., Handbook to the Marsupialia, and Monotremata. Allen’s Nat. Library, 1894, pt. III. pp. 249-286. De Vis, C. W. Proc. Linn. Soc. New South Wales, vol. VIII. pt. 3, 1883, p. 395 (Sirenian). Id., ibid, vol. X. 1895, pp. 75-133, pls. XIV.-XVIII. (Macropodidae). Id., Proc. R. Soc. « 314 » Vict., vol. XII. (N.S.), pt. I, 1899, pp. 107-11 (Marsupials). Etheridge, R. jnr. Geol. and Pal. Queensland, 1892, pp. 663-683 (Pleistocene Mammals). Dun, W. S. Rec. Geol. Surv. New South Wales, vol. III. pt. 4, 1893, pp. 120-124, pl. XVI. (Palorchestes). Ibid., vol. IV. pt. 3, 1895, pp. 118-126, pls. XI. and XII. (Monotremes). Stirling, E. C. and Zietz, A. H. C. Mem. Roy. Soc. S. Australia, vol. I. pt. I. 1899 (Descr. of Diprotodon, Manus and Pes.). Spencer, W. B. Proc. Zool. Soc. 1900, pp. 776-794, pls. XLIX. and L. (Wynyardia). Hall, T. S. Proc. R. Soc. Vict. vol. XXIII. (N.S.), pt. II. 1911, pp. 257-265, pl. XXXVI. (Rev. of Squalodontidae). Spencer, W. B. and Walcott, R. H. Proc. R. Soc. Vict., vol. XXIV. (N.S.), pt. I. 1912, pp. 92-123, pls. XXXVI.-XXIX. (Thylacoleo). Chapman, F. Rec. Geol. Surv. Vict., vol. III. pt. 2, 1912, pp. 236-238, pl. XL. (Scaldicetus). Woods, J. E. T. Geol. Observations in S. Australia, 1862, pp. 329 and 330 (Human Remains): also Krefft, G. Australian Vertebrata, Recent and Fossil, 1867, p. 91; Etheridge, R. jnr. Rec. Geol. Surv. New South Wales, vol. III. pt. 4, 1893, pp. 128-132; Etheridge, R. jnr. and Trickett, O. Ibid., vol. VII. pt. 4, 1904, pp. 325-328.
The tools and other paraphernalia necessary for fossil collecting are fortunately within the reach of all. The principal of these is a geological hammer, preferably with a pick at one end of the head and the opposite end square-faced. The pick end is useful for digging out fossils from soft clays, or for extracting a block of fossils entire. The square end is employed for breaking up the slabs or masses containing fossils. To get good results, much will of course depend upon one’s skill in striking the right face of a block. If bedding planes are present on the lump from which we wish to extract our fossils, it will be well to strike at right angles to these layers in order to split them asunder, thus exposing a shell-layer corresponding to the original surface of the ancient sea-bed upon which the organisms accumulated. In some cases the splitting of fossiliferous rocks may be best carried out with the pick end, provided it be not too sharply curved. The hammer should be faced with steel, for many fossiliferous rocks, especially compact limestones, are apt to severely try the temper of an ill-made tool.
A chisel, of chilled steel, should accompany the hammer, since this is often of the greatest use in working out large fossils, more particularly those that are buried in a cliff or quarry face. The process of extracting difficult specimens should never be hurried, for one often gets surprisingly good results with a little extra care.
A strong pocket knife may be used in trimming specimens and partially cleaning shells that can be safely manipulated on the spot, but the final cleaning should be left until the return home. The knife is also useful for cleaning slates and shales, since the chisel-edge is frequently a trifle too thick for this kind of work.
For the more delicate fossils, means for careful packing should be provided; chip-boxes and cotton-wool being indispensable for the smaller specimens. A ready method of packing the fossils obtained from the friable, sandy tertiary deposits is to store them in tins, the contents of which can be firmly secured from rattling by filling up with sand. This sand, however, should be taken from the same bed in which the fossils occur, so as to get no admixture of the smaller shells from another formation or deposit; for although we may not wish to examine the finer material ourselves, it will yield in many cases a rich harvest to our microscopical friends, such residues containing microzoa, as shells of foraminifera, polyzoa and carapaces of the ostracoda. The residues referred to may be obtained from many of our marls and rubbly limestones by the simple process of washing in water, and repeatedly pouring off the finest « 317 » clayey mud, until only a sandy deposit remains, which can then be dried and sorted over by the aid of a lens or low power microscope.
Hints on Fossil Collecting.—
As regards the places most suitable for collecting fossils, the Cainozoic beds are perhaps, the most accessible to a beginner, especially in Victoria. For instance, the cliff exposures at Beaumaris, Port Phillip, will afford a plentiful supply of the little heart-shaped sea-urchin, Lovenia, and an occasional Trigonia and Limopsis, as well as many other fossils of the great group of the shell-fish or mollusca. The richest bed containing the sharks’ teeth at the above locality is almost perpetually covered with a bed of shingle, but can be reached by digging at the cliff-base. Isolated specimens, however, although rather the worse for wear, may often be picked up amongst the shingle, having been washed up from the foreshore by the tide. An enticing band of large bivalve shells (Dosinea), can be seen halfway up the cliffs, near the baths at this locality, but are somewhat disappointing, for when obtained they crumble to pieces in the hand, since their shells are composed of the changeable form of carbonate of lime called aragonite, which has decomposed in place in the bed, after the shells were covered up by the deposit.
Good collections of shells of the Balcombian series may be easily made at Balcombe’s Bay and Grice’s Creek, Port Phillip. They can there be dug out of the grey-blue clay with a knife, and afterwards cleaned at leisure by means of a soft tooth brush dipped in water. In the cement stone at the same place « 318 » there are numerous shells of pteropods or “sea-butterflies” (Vaginella), and specimens of the stone may be obtained, showing myriads of the porcelain-like shells, and also their internal casts in the hard greenish coloured matrix.
The ferruginous or ironstone beds seen in the Flemington Railway cutting, Melbourne, is an old marine shell-bank, resting on basalt. The shells have all been dissolved away, and only their casts and moulds remain. These impressions are, however, so faithfully moulded that the ornamentation of each shell can often be reproduced on a squeeze taken with a piece of modelling wax or plasticine. Such fossil remains are easily collected by carefully breaking up the blocks of ironstone with a hammer.
Quarries in the older limestones and mudstones in Victoria, New South Wales and other States, are often good hunting grounds for fossils. The quarry at Cave Hill, Lilydale, for example, will be found very profitable, for the limestone is full of corals and molluscan shells; whilst the friable or rubbly portion is worth breaking down for the smaller fossils. The bed-rock (Silurian) of Melbourne is in places very fossiliferous; the sandstones of Moonee Ponds Creek generally affording a fair number of brachiopods, and occasionally corals. The mudstones of South Yarra, Studley Park, Yan Yean, and other places on the same geological horizon, contain a rich fauna, to be obtained only by the assiduous collector who will search over and break up a large number of blocks. Practice in this work makes a good collector; although of course one must know something about « 319 » the objects looked for, since many apparently obscure fossil remains of great interest might easily be passed over for lack of knowledge as to what should be expected to occur at each particular locality.
Many other good collecting grounds might here be alluded to, but we have purposely cited only a few near Melbourne, since a selection from other parts of Australasia may easily be made from the localities mentioned in connection with the various groups of fossils dealt with in the systematic portion of this work.
Preservation of Fossils.—
Many of the Cainozoic fossils from the shelly sands and clays are extremely delicate, owing in some cases to their being imperfectly preserved, seeing that they frequently contain in their shell-structure layers of the unstable form of carbonate of lime called aragonite. Fossils containing aragonite are:—Calcareous Sponges; Corals; Bivalved shells, except Oysters, Pectens, and the outer layer of Spondylus, Pinna, and Mytilus; Gasteropods (with a few exceptions); and Cephalopods. In some of these, however, a transformation of the aragonite into calcite enables the fossil to be permanently preserved. The delicate fossils referred to should be dipped in weak glue or gelatine and left to dry; after which their final cleaning can be done with the aid of a little warm water and a soft brush.
Certain of the clays and mudstones, both of Cainozoic and Jurassic ages which show remains of plants, such as leaves and fern fronds, are often best treated with a thin « 320 » surface layer of paper varnish, before they lose the natural moisture of the rock; for when they become perfectly dry the thin carbonaceous film representing the original leaf-substance peels off, and the fossil is consequently destroyed. A method of treatment for Cainozoic leaves, by dipping them in warm vaseline and brushing off the superfluous material, has been described by Mr. H. Deane.
Storing Fossils for Reference.—
Fossils specimens are generally best displayed in cardboard trays; or if thin wooden paper-covered tablets are used, say of about 3-16in. thickness and cut to proportionate sizes, the fossils should be held in place by pins for easy removal, unless more than one example can be shown together, exhibiting all aspects, when they can be secured to the tablet by a touch of seccotine. The smaller shells may be displayed in glass topped boxes, which in turn may be stuck down to tablets or placed in trays.
Aboriginal implements, 303
Aborigines, probable origin of, 302
Acanthias, 270
Acanthodes, 261
Acanthosphaera, 103
Acanthothyris, 166, 167
Acentrophorus, 263
Acrolepis, 263
Actaeon, 197
Actinoceras, 205, 207
Actinocrinus, 136
Actinodesma, 178, 179
Actinopteria, 178, 179
Actinostroma, 121, 122
Adeona, 158
Aechmina, 237
Aeschna, 250
Aetheolepis, 267
Agathiceras, 207
AGNATHA, 258
Agnostus, 227
Allodesma, 176
Ambonychia, 177
Ammodiscus, 96, 97
Ammonites, 204, 209, 210
AMMONOIDEA, 205
Amoeba, 36, 95
AMPHIBIA, structure of, 272
Amphistegina, 100
Amplexus, 117
Ampyx, 229
Amusium, 185
Anas, 283
Anchura, 197
Ancilla, 198, 199, 202
Ancyloceras, 209, 210
ANGIOSPERMEAE, characters of, 40
ANNELIDA, 152
Anomalina, 98
Anomalopteryx, 283
Antedon, 138
ANTHOZOA, 64, 113
Antiquity of man in Australia, 304
Aparchites, 237
Apateolepis, 262
Aphnelepis, 267
Apocynophyllum, 91
Aptornis, 283
Aptychopsis, 246
Arabellites, 153
Arachnoides, 146
Araucarioxylon, 68
Araucarites, 89
Arca, 184, 186, 188
Archaeocidaris, 144
Archaeocyathina, 113
ARCHAEOCYATHINAE, 112
Archaeomaene, 267
Archaeopteryx, 280
Arctocephalus, 299
Arenicolites, 153
Argillaceous rocks, 69
Argilloecia, 237
?Argiope, 166
Argonauta, 205
ARTHROPODA, structure and subdivisions of, 38, 220
Asaphus, 227, 228
Aspidorhynchus, 267
Astarte, 182
Asteracanthus, 269, 271
ASTEROIDEA, 139
Asterolepis, 258
Astralium, 198, 200
Astropecten, 141
Athyris, 161, 162, 165
Atrypa, 158, 160, 162
Aturia, 210
« 322 »
Atys, 204
Aucella, 183
Aulopora, 116
Australian fossiliferous strata, 45-48.
AVES, 280
Aviculopecten, 179, 180
Axopora, 119
Bactronella, 112
Baculites, 210
Baiera, 89, 164
Bairdia, 240
Balanophyllia, 118
Balanus, 243
Balcombian bivalves, 186
” gasteropods, 199
Bandicoot, 289, 295
Bankivia, 201
Banksia, 91, 281
Barbatia, 184, 185
Barnacles, 240
Barnea, 187
Bathytoma, 201
Bela, 201
Belemnites, 205, 209, 210
BELEMNOIDEA, 205
Bellerophon, 193, 194, 195, 196
Belonorhynchus, 262
Belonostomus, 267
Bettongia, 295
Beyrichia, 235, 236, 237
Biloela, 274
Bipora, 158
Birds, fossil, 53, 280
Biziura, 283
BLASTOIDEA, distribution and characters of, 61, 138
Blue-green Algae, 76, 82
Bog iron-ore, 80
Bolodon, 286
Bombax, 91
Bone-beds, 78
Bone-breccias, 79
Bothriceps, 273
Botryocrinus, 136
BRACHIOPODA, structure of, 57, 158
Brachiopod limestone, 74
Brachymetopus, 232
Brachyphyllum, 89
Bracken fern, 91
Brissopsis, 148
Brittle-stars, characters of, 61, 141
Bronteus, 229, 230
Bryograptus, 124, 126, 227
BRYOPHYTA, characters of, 39
Buccinum, 191
Buchozia, 199
Bulimina, 97, 98
Bulinus, 69, 191
Bulla, 204
Bullinella, 198, 199
Bythocypris, 236
Bythotrephis, 82
Cainozoic Balanidae, 243
” bird, Victoria, 281
” bivalves, 184
” brachiopods, 166
” brittle-stars, 143
” chitons, 190
” corals, 118
” crabs, 247
” echinoids, irregular, 146
” echinoids, regular, 145
” fishes, 269
” Foraminifera, 99
” gasteropods, 198
” gasteropods, New Zealand, 202
” Holothuroidea, 148
” insects, 250
” Lepadidae, 243
” Ostracoda, 239
” and Pleistocene reptiles, 279
” plants, 89
” Polyzoa, 158
« 323 »
Cainozoic Radiolaria, 104
” scaphopods, 189
” sponges, 110
” starfishes, 141
” strata, 45, 46
Calcareous rocks, 72
” sponges, 112
Callograptus, 122
Callorhynchus, 269
Calymene, 229, 230, 231
CALYPTOBLASTEA, 122
Calyptraea, 198, 200, 201
Camarotoechia, 160, 161, 162
Cambrian bivalves, 177
” brachiopods, 159
” crinoids, 134
” Foraminifera, 96
” gasteropods, 192
” Ostracoda, 235
” plants, 82
” Radiolaria, 102
” sponges, 107
Cameroceras, 207
Campanularia, 122
Campophyllum, 115, 117
Cancellaria, 198, 199, 202
Canis, 298
Cannel coal, 76
Capitosaurus, 274
Capulus, 194
Carbonaceous rocks, 76
Carboniferous brachiopods, 162
” crinoids, 136
” fishes, 259
” Foraminifera, 96
” gasteropods, 196
” Ostracoda, 237
” plants, 85
Carbopermian bivalves, 179
” blastoids, 139
” brachiopods, 163
” cephalopods, 207
” corals, 116
” crinoids, 137
” fishes, 261
” Foraminifera, 97
” gasteropods, 196
” labyrinthodonts, 273
” Ostracoda, 237
” palaeechinoids, 144
” Phyllopoda, 233
” plants, 86
” sponges, 110
” starfishes, 141
” trilobites, 232
Carcharodon, 269, 270, 271
Carcharoides, 269
Cardiola, 177, 178
Cardita, 184, 187
Cardium, 176, 184, 186, 187
CARNIVORA, 298
Carposphaera, 102
Carpospongia, 109
Caryocaris, 244, 246
Cassidulus, 148
Catenicella, 158
Cellaria, 158
Cellepora, 158
Cenellipsis, 102
Cenosphaera, 102, 103
CEPHALOPODA, characters of, 204
Ceratiocaris, 246
Ceratodus, 265, 267
Ceratotrochus, 118
Cerithiopsis, 200
Cerithium, 198, 200
Cestracion, 261, 269, 271
CETACEA, 295
Cetotolithes, 296
Chaenomya, 181
CHAETOPODA, 152
Chama, 185
Changes of climate in the past, 31
CHEILOSTOMATA, 155, 157
Cheirurus, 229, 231
Chelodes, 190
Cherts, 71
Chione, 185, 187, 188
Chiridota, 148
« 324 »
Chironomus, 250
Chiton, 190
Chonetes, 160, 161, 162
CHORDATA, 257
Chosornis, 283
Chronozoön, 298
Cicada, 250
Cidaris, 145
Cimoliosaurus, 279
Cinnamomum, 91
Cinulia, 197
CIRRIPEDIA, habits and structure of, 240
Cladochonus, 117
Cladophlebis, 89, 164, 182
CLADOPHORA, 122
Classification of animals, 35
Clathrodictyon, 121
Clausilia, 191
Clavigera, 165
Clays, 69
Cleiothyris, 164
Cleithrolepis, 262, 263, 274
Climacograptus, 127
Climatius, 258
Clonograptus, 123, 124, 126
Clypeaster, 146
Cnemiornis, 283
Coals, 76
Coccolepis, 267
Cocconema, 92
Coccosteus, 259
COELENTERATA, characters of, 37
Coleolus, 193
Collecting fossils, 317
Colubraria, 199
Columbarium, 198, 201, 202
Columbella, 198
Conchothyra, 184
Conocardium, 177, 178
Conodonts, 153
Conosmilia, 118
Conularia, 193, 194, 196
Conus, 198, 199, 202, 204
Coprosmaephyllum, 90
Coral limestone, 73
Corals, 64, 113
Corax, 267
Corbicula, 182
Corbula, 177, 185, 187, 188
Cordaites, 85
Cornulites, 154
Coscinocyathus, 113
Coxiella, 69
Crassatellites, 176, 184
Crenella, 176
Crepicephalus, 227
Crepidula, 198
Cretaceous (Lower and Upper) cephalopods, 209
” cephalopods, New Zealand, 210
” Cheilostomata, 157
” crinoids, 137
” echinoids (irregular), 146
” (Lower) fishes, 267
” fishes, New Zealand, 268
” Foraminifera, 98
” gasteropods, 197
” plants, 89
” Radiolaria, 103
” (Lower) reptiles, 277
” reptiles, New Zealand, 279
” scaphopods, 189
” sponges, 110
Crinoidal limestone, 74
CRINOIDEA, occurrence and structure of, 61, 133
Crioceras, 209
Crisia, 158
Cristellaria, 98
Crocodilus, 279
Cromus, 229
Crustacea, an archaic group, 221
” development of, 221
” fossil, 54
Cryptodon, 186
« 325 »
Cryptograptus, 127
Cryptoplax, 190
Cryptostomata, 155, 156
Ctenodonta, 177, 178
Ctenodus, 261, 263
Ctenolates, 272
Ctenostreon, 182
Cucullaea, 182, 184, 185
Cultellus, 188
Cuna, 184, 186, 187
Cupressinoxylon, 78, 89
Cupressus, 91
Cuscus, 295
Cuttle-fishes, 205
CYANOPHYCEAE, 82
Cyathocrinus, 137
Cyathophyllum, 113, 115, 117
Cyclas, 69
Cycloceras, 206
Cyclolituites, 207
Cyclometopa, 248
Cyclonema, 194
CYCLOSTOMATA, 155
Cydnus, 250
Cymbella, 92
Cyphaspis, 229
Cyphon, 250
Cypraea, 191, 198, 199, 200, 202
Cypricardinia, 178
Cyprid limestone, 75
Cyrenopsis, 184
Cyrtoceras, 204, 207
Cyrtograptus, 128
Cyrtina, 162, 164
Cyrtolites, 193
Cystideans, 61
Cystiphyllum, 116
Cythere, 239, 240
Cytherella, 240
?Cytheridea, 238
Cytheropteron, 239
Dadoxylon, 68
Dalmanites, 224, 225, 229, 231
Daonella, 182
Darter, 283
?Darwinula, 238
Dasyurus, 287, 295
DECAPODA, 246
Deep Leads, fruits of, 91
” insects from, 250
Deltodus, 261
Deltopecten, 180
Dendrocrinus, 134, 135
Dendrocygna, 283
Dendrograptus, 122
Dendrophyllia, 119
Dennantia, 198
Dentalium, 189
Dentition of Reptiles, 275
Deontopora, 120
Desmoceras, 209
Devonian bivalves, 178
” brachiopods, 161
” cephalopods, 207
” corals, 115
” crinoids, 136
” fishes, 258
” gasteropods, 195
” Ostracoda, 237
” plants, 85
” Radiolaria, 102
” scaphopods, 189
” stromatoporoids, 121
” trilobites, 231
DIADACTYLA, 287
Diatomite, 72
Diatoms, 92
Dicellograptus, 126, 127
Dichograptus, 126
Dicranograptus, 126, 127
Dictyonema, and allies, 122
Dictyopyge, 262
Didymograptus, 124, 126
?Didymosorus, 89
Dielasma, 164, 165
Dikellocephalus, 227
Dimetrodon, 276
Dimya, 184, 185, 186
Dinesus, 227
Dingo, 298, 305
Dinornis, 281, 282, 283, 299
Diodon, 270, 271
« 326 »
Dione, 188
Diphyphyllum, 113
Diplograptus, 124, 126, 127, 128
Diprotodon, 51, 290, 293
Diprotodon-breccias, 203
DIPROTODONTIA, 287
Discina, 166
Discorbina, 98
Dissocheilus, 199
Dithyrocaris, 246
Ditrupa, 154
Ditrupa limestone, 74
Dolichodon, 296
Dolichometopus, 226
Dolium, 201
Donax, 175, 187
Dorsetensia, 209
Dosinea, 185, 188
Drillia, 198, 202
Dromaeus, 282, 283
Dromornis, 282
Duck, 283
Duncaniaster, 147
Ear-bones of whales, 296
Early observers, 24
Eburnopsis, 199, 200
Echidna, 286, 287
Echinocyamus, 146
ECHINODERMATA, characters of, 37, 59
” divisions of, 133
ECHINOIDEA, 143
Echinolampas, 147, 148
Echinoneus, 147
Echinus, 145
Ecionema, 112
Edaphodon, 271
Edestus, 262
Edmondia, 177, 180, 182
Eglisia, 202
Elephant-fish, 269, 271
Elephant-tusk shells, 188
Elevated sea-beds, 27
Elonichthys, 261, 263
Elpisopholis, 263
Emarginula, 198
Emu, 283
Encrinurus, 229
Endoceras, 205
Endothyra, 96, 98
Entalophora, 158
Entomis, 238
Ephemera, 250
Equisetites, 40
Errant worms, 153
Erycina, 187
Erymnoceras, 209
Estheria, 233
Eucalyptus, 90, 91, 281
Eulima, 198
Eunema, 193
Eunicites, 153
Euomphalus, 194, 195, 196
Eupatagus, 147
Euphemus, 196
Eurydesma, 181
EURYPTERIDA, 248
Euthria, 198
Eutrochus, 200
Evolution of life-forms, 33
Fagus (Notofagus), 91
Falcon, 283
Fasciolaria, 198, 199
Favosites, 73, 114, 115, 116
Feather-star, 138
Fenestella, 156, 157
Fibularia, 146
Fishes, fossil, 53
” primitive types, 258
” true, 258
Fish-lizards, 275, 276, 277, 278
Fissilunula, 183, 184
Fissurellidea, 198
Fistulipora, 155, 156
Flabellina, 98
Flabellum, 118, 119
Flightless pigeon goose, 283
Flints, 71
Flying phalanger, 295
Foraminifera, characters of, 36, 95
” fossil, 65
« 327 »
Foraminiferal limestone, 73
Fossil faunas, differences in, 43
Fossiliferous strata, Australia, 45-48
” strata, New Zealand 49
Fossil, origin of name, 23
Fossils an index to age of strata, 26, 32
” nature of, 21
” petrifaction of, 23
” preservation of, 23
” structure preserved in, 24
Fossil wood, 24, 66, 68
Frondicularia, 97, 98
Fruits of the deep leads, 91
Fulica, 283
Fusus, 198, 201
Galeocerdo, 269, 271
Gallinula, 283
Gangamopteris, 86
Ganorhynchus, 259
Gari, 185
GASTEROPODA, characters of, 190
Gastrioceras, 207
Geinitzina, 98
Genyornis, 282, 302
Geological epochs, 45-49
Geology, scope of, 21
Giant kangaroo, 289
” lizard, 280
” penguin, 280
Gibbula, 198
Ginkgo, 89, 91
Girvanella, 76, 82, 86
Glauconite casts of foraminifera, 96
Glossograptus, 126, 127
Glossopteris, 86
Glycimeris, 184, 187
Glyphioceras, 207
Gomphonema, 93
Gondwana-Land, 87
Goniatites, 207, 208
Goniograptus, 124, 126
Gosfordia, 262
Gosseletina, 196
Grammysia, 177
Granatocrinus, 139
Graphularia, 118, 119
Graptolites, Bendigo series, 124
” Lancefield series, 124
” nature of, 63, 123
” Tasmania, 128
GRAPTOLITOIDEA, 123
Gregoriura, 142
Griffithides, 232
Gromia, 95
Ground pigeon, 283
Gryphaea, 182
Grypotherium, 53
Guide fossils, 43
GYMNOSPERMEAE, characters of, 40
Gyracanthides, 261
Gyroceras, 207
Gyrodoma, 194
Halimeda limestone, 75
Haliotis, 198, 200
Haliserites, 83
Halysites, 114
Hamites, 210
Hapalocrinus, 136
Haploceras, 209
Haplophragmium, 97, 98
Harpa, 198, 199, 201
Harpactocarcinus, 248
Harpagornis, 283
Hawk, 283
Helicocrinus, 136
Helicotoma, 195
Heliolites, 115, 116
Heliopora, 115
Heliosphaera, 103
Helix, 203
Hemiaster, 148
Hemipatagus, 148
Heterocrinus, 135
« 328 »
HETEROPODA, 190
Heteropora, 158
Hexactinellid sponge, 107, 110
Hinge-structure, in bivalves, 175
Holaster, 147
HOLOTHUROIDEA, 148
Homalonotus, 229, 231
Hornera, 158
Huenella, 159
Human remains, sub-recent, 299
Hyalostelia, 108, 110
Hybocrinus, 135
Hydractinia, 119, 120
HYDROZOA, 63, 119
Hymenocaris, 244
Hyperammina, 97
Hyolithes, 192, 193, 194
Hypothyris, 164
Hypsiprymnus, 295
Ibis, 283
Ichthyosaurus, 276, 277, 278
Idiostroma, 121
Idmonea, 158
Illaenus, 229
Indusial limestone, 75
Inoceramus, 183, 184
Insects, 53, 250
Ironstone, 80
Irregular echinoids, 146
Ischnochiton, 190
Ischyodus, 269, 270
Isochilina, 237
Isocrinus, 137, 138
Janjukian bivalves, 186
” gasteropods, 200
Jonesina, 237
Jurassic bird, 280
” bivalves, 182
” brachiopods, 165
” cephalopods, 208
” fishes, 264
” Foraminifera, 98
” gasteropods, 196
” insects, 250
” Ostracoda, 238
” Phyllopoda, 233
” plants, 89
” reptiles, 276
” scaphopods, 189
Kalimnan bivalves, 187
” gasteropods, 201
Kangaroo, 295
Keeneia, 196
Kekenodon, 295, 296
Kerosene shale, 77
Kionoceras, 206
Kloedenia, 237
Labrodon, 271
LABYRINTHODONTIA, 272
Lagena, 98
?Lagria, 250
Lamna, 267, 269, 271
Lamp-shells, 57, 158
Lasiocladia, 110
Lasiograptus, 126, 127
Latirus, 198, 201
Laurus, 91
Leaia, 233
Leda, 182, 184, 185, 187, 188
Leonardo da Vinci, 25
Lepas, 243
Leperditella, 234
Leperditia, 233, 234, 235, 237, 238
Lepidocyclina, 99, 100
” limestone, 73
Lepidodendron, 40, 85, 261
” beds, 162
Lepralia, 157, 158
Leptaena, 162, 164
Leptoclinum, 257, 258
Leptodesma, 179
Leptodomus, 177
Leptograptus, 124
Leptolepis, 264, 265, 267
Lepton, 187
Lichas, 229
Lichenopora, 158
« 329 »
Lieberkuehnia, 95
Lima, 184, 185, 186
Limatula, 185
Limestones formed by organisms, 72
Limnaea, 69
Limopsis, 184, 185, 187
Limulus, 248
Lingula, 160, 162, 166, 261
Linthia, 147, 148
Liopyrga, 201
Liotia, 198, 200
Lithistid sponges, 109, 110
Lithological evidence, value of, 44
Lithophaps, 283
Lithothamnion, 75
Lituites, 207
Lituola, 97
Loganograptus, 126
Lophophyllum, 117
Lorica, 190
Lotorium, 198, 200, 202
Lovenia, 147
Lower Cambrian trilobites, 226
” Cretaceous bivalves, 183
” ” brachiopods, 166
” ” cephalopods, 209
” ” crab, 246
” ” dragon-fly, 250
” ” fishes, 267
” ” reptiles, 277
” Mesozoic fishes, 263
” Ordovician graptolites, New Zealand, 126
” Ordovician graptolites, Victoria, 124
Loxoconcha, 239
Loxonema, 193, 194, 195, 196
Lucina, 185, 187
Lung-fish, 265
Lunucammina, 98
Lunulicardium, 178
Lunulites, 158
Lyriopecten, 179
Maccoyella, 183, 184
Macrocephalites, 209
Macrocheilus, 196
Macrocypris, 236, 240
Macropora, 158
Macropus, 289, 295
Macrotaeniopteris, 88
Mactra, 177, 185, 188
Madrepore limestone, 73
Magasella, 166, 168
Magellania, 166, 167, 168
Magnolia, 91
Maiden-hair tree, 89
Mail-shells, 189
MAMMALIA, early types, 285
Mammals, fossil, 51
Manatees and dugongs, 298
Marginella, 198, 199
Marginulina, 98
Marsupial lion, 293
Marsupial, oldest known Australian, 294
Marsupials, 287
” Pleistocene and living, 295
Martiniopsis, 164
Mastodonsaurus, 274
Material for fossil collecting, 315
Megalania, 280
Megalosaurus, 277
Melania, 203
Melosira, 92
Membranipora, 157, 158
Meretrix, 177, 185, 187
Mesoblastus, 139
Mesostigmodera, 250
Mesozoic strata, 46
Metablastus, 139
Metasqualodon, 295, 296
METAZOA, 95
Micraster, 146
Microdiscus, 227
Mikrogromia, 95
Millepora, 119
Milleporids, 119
Miliolina, 96, 100, 101
« 330 »
Miocene bird, New Zealand, 280
” leaf-beds, 90
Miolania, 279
Mitra, 198, 199, 204
Moa-birds, 281-285, 299
Modiola, 183, 186
Modiolaria, 186
Modiolopsis, 177
MOLLUSCA, characters of, 38, 56, 174
MOLLUSCOIDEA, characters of, 38, 57, 154
Monactinellid sponges, 109, 110
Monogenerina, 97
Monograptus, 124, 128
Monostychia, 146
Monotis, 182
MONOTREMATA, 286
Monticulipora, 155
Monticuliporoids, 117
Montlivaltia, 118
Moor-hen, 283
Mopsea, 119
Morio, 198, 200
Mound-builders, 283
Mourlonia, 196
Mud-fish, 265, 267
Muds, 69
Mudstone, 70
MULTITUBERCULATA, 286
Murchisonia, 104, 195, 196
Murex, 198, 199, 200
Myodora, 185, 187
Myriolepis, 262, 263
Mytilarca, 177
Mytilus, 182, 183, 187, 188
Naming of animals, 34
Nassa, 191, 198, 204
Natica, 191, 197, 198, 200, 201
Native cat, 287, 295
” dog, 298
” honeysuckle, 91, 92
NAUTILOIDEA, 204
Nautilus, 204, 207, 209, 210
Navicula, 92
Nebalia, 244
Necrastur, 283
Neoceratodus, 267
Newer Pliocene seal, 299
Newtoniella, 198
New Zealand fossiliferous strata, 49
Niso, 194, 198
Nodosaria, 98, 100
Nonionina, 96
Normanites, 209
Notasaphus, 227
Notidanus, 268, 269, 270, 271
Notochelone, 53, 277
Notophyllia, 118
Nototherium, 293
Nubecularia, 97, 98
Nucleospira, 160
Nucula, 175, 177, 178, 183, 184, 185
Nuculites, 177, 178
Nullipore limestone, 75
Nummulites, 65, 99
Nummulitic limestone, 73
Nyroca, 283
OCTOPODA, 205
Octopus, 205
Odontaspis, 269, 270, 271
ODONTOCETI, 295
Odontopleura, 229, 231
Odostomia, 198, 200
Oenonites, 153
Olenellus, 226, 227
Oliva, 204
Ommatocarcinus, 247
Omphalotrochus, 194
Oolitic ironstone, 81
Ophileta, 192, 193
OPHIUROIDEA, 141
Orbiculoidea, 160
Orbitoides, 99
Ordovician bivalve, 177
” brachiopods, 159
« 331 »
” cephalopods, 205
” corals, 113
” crinoids, 135
” gasteropods, 193
” Phyllocarida, 244
” Radiolaria, 102
” sponges, 108
” trilobites, 227
Ornithorhynchus, 286, 287
Orthis, 159, 160, 161, 162
” limestone, 74
Orthoceras, 204, 205, 206, 207, 208
Orthonota, 177
Orthothetes, 162
OSTRACODA, features of carapace, 234
” habits of, 234
” structure of, 233
Ostrea, 182, 184, 187
Otaria, 299
Oxyrhina, 269, 270, 271
OXYSTOMATA, 247
Oxytelus, 250
Pachydomus, 181
Pachyornis, 282, 283
Pachypora, 73, 116
Palaeaster, 140, 141
Palaeeudyptes, 280, 281
Palaeohatteria, 276
Palaeolycus, 250
Palaeoneilo, 177, 178
Palaeoniscus, 261, 263, 274
Palaeopelargus, 283
Palaeozoic chitons, 189
” Cladophora, 122
” Cryptostomata, 156
” errant worms, 153
” strata, 47
” Trepostomata, 155
Palissya, 89, 164
Pallymnarchus, 279
Palorchestes, 290
Panda, 203
Panenka, 177
Paracyainus, 118
Paracyclas, 177, 179
Paradoxechinus, 145
Paradoxorhyncha, 239
Parasqualodon, 295, 296
Pareiasaurus, 276
Patella, 190, 191
Pecten, 175, 182, 183, 184, 185, 186, 187, 188
PELECYPODA, characters of, 174
” hinge structure of, 175
Pelican, 283
Pelicanus, 283
Pelosina, 97
?Peltopleurus, 262
Pentacrinus, 137, 138
Pentagonaster, 141
Pentamerus, 160, 162
Penteune, 91
Peragale, 289
Perameles, 289, 295
Perisphinctes, 209
Permian and Triassic reptiles, 276
Perna, 187
Peronella, 148
Persoonia, 90
Petaurus, 295
Petraia, 113
Phacops, 229, 230, 231
Phalanger, 295
Phanerotrema, 194
Phascolomys, 289, 295
Phascolonus, 289
Phialocrinus, 137
Phillipsia, 232
Phoenicopsis, 88
Pholas, 177
Pholidophorus, 262, 263
Phos, 198
Phragmoceras, 207
Phryganea, 75
PHYLACTOLAEMATA, 155
PHYLLOCARIDA, structure of, 243
Phyllocladus, 90
Phyllograptus, 123, 126
« 332 »
PHYLLOPODA, 233
Phyllotheca, 274
Physa, 191
Physonemus, 261
Pigeon, 283
Pinna, 186
PINNIPEDIA, 299
Pisania, 202
?Pisocrinus, 136
Placopsilina, 97
Placotrochus, 118
Placunanomia, 184, 187
Plagiarca, 184
Plagiaulax, 286
Planorbis, 191
Plants, fossil, 66
Plant series, characters of, 39
Platalea, 283
Platyceps, 273
Platyceras, 192, 194, 195, 196
Platycoila, 91
Platycrinus, 137
Platyschisma, 196
Platysomus, 263
Plaxiphora, 190
Plectroninia, 112
Pleioclinis, 91
Pleistocene birds, New Zealand, 283
” bivalves, 188
” carinate birds, 283
” diprotodonts, 289
” fish, 272
” Foraminifera, 101
” gasteropods, 202
” lobster, 248
” plants, 91
” seal, 299
Plerophyllum, 117
Plesiastraea, 119
Plesiolampas, 148
Plesiosaurus, 279
Pleuracanthus, 263
Pleurodictyum, 114
Pleuromya, 183
?Pleurostomella, 98
Pleurotoma, 198, 199, 202
Pleurotomaria, 194, 196, 197, 200, 202
Plicatula, 186
Pliocene moa, New Zealand, 281
Pliosaurus, 278
Plotus, 283
Podocarpus, 90
Poecilodus, 262
?Pollicipes, 243
POLYCHAETA, 152, 154
Polycotylus, 279
Polymastodon, 286
Polymorphina, 98, 100
POLYPLACOPHORA, 189
Polypora, 157
POLYPROTODONTIA, 287
Polystomella, 101
POLYZOA, characters of, 59, 155
” subdivisons of, 155
Polyzoal limestone, 74
Porcellia, 196
Porcupine fish, 270, 271
Porina, 158
Porphyrio, 283
Portheus, 268
Poteriocrinus, 137
Prehensile Rat-kangaroo, 295
Preservation of fossils, 319
Primitia, 236, 237
Pristisomus, 262
Procoptodon, 290
Productus, 162, 163, 164
Proechidna, 287
Proetus, 229, 232
Progura, 283
Prosopon, 246
Protaster, 142
Protocardium, 185
Protopharetra, 113
Protoretepora, 157
Protospongia, 107,108
PROTOZOA, characters of, 36, 65, 95
« 333 »
Psammechinus, 145
Pseudamaura, 197
Psilichthys, 264
PTERIDOPHYTA, characters of, 40
PTERIDOSPERMEAE, characters of, 40
Pterinea, 178, 179
Pteris (Pteridium), 91
PTEROPODA, 190, 192, 193, 194
Pterygotus, 248, 249
Ptilograptus, 122
Ptychoparia, 226, 227
Pugnellus, 184
Pulvinulina, 98
Purbeck marble, 74
Purisiphonia, 110
Purpura, 191
RADIOLARIA, characters of, 36, 66
” habitat of, 101
” structure of, 101
” subdivisions, 102
Rail, 283
Raised beaches as distinct from middens, 29
Ranella, 204
Range-in-time of fossils, 50
Raphistoma, 193, 195
Rat-kangaroo, 295
Receptaculites, 109
Regular echinoids, 144
Reinschia, 78
Reptiles, fossil, 53
” dentition of, 275
” structure of, 274
Reteocrinus, 135
Retepora, 158
Reticularia, 164
Retiolites, 124, 128
Rhacopteris, 86
Rhinopterocaris, 244, 246
Rhipidomella, 162
Rhizophyllum, 113
Rhodocrinus, 135
Rhombopora, 156
Rhynchonella, 158, 165, 166
RHYNCHOTA, 250
Rhynchotrema, 160
Ringicula, 202
Risella, 191
Rissoa, 198
Rissoina, 197
Rostellaria, 198
Rotalia, 96, 101
Rugose corals, 113
Saccammina, 96
Saccocaris, 244
Sagenodus, 263
Salterella, 192
Sandstones, 71
Sanidophyllum, 115
Sarcophilus, 287, 295
Sargus, 272
Scala, 191, 198, 199, 200, 202
Scalaetrochus, 194
Scaldicetus, 297
Scaphella, 202
Scaphites, 209
SCAPHOPODA, 188
Scenella, 193
Sceparnodon, 289
Schizaster, 148
Schizodus, 175
Schizophoria, 162
Schloenbachia, 209
Scutellina, 146
Sea-beds far from the present coast, 29
Sea-bream, 272
” -cucumbers, 148
” -firs, 119, 122
” -mats, 154, 155
” -pen, 119
” -urchins, 59, 143
” characters of, 144
Sedentary worms, 154
Seguenzia, 199
Selenaria, 158
Semele, 185
Semicassis, 198
Seminula, 164
« 334 »
Semionotus, 262, 263
SEPIOIDEA, 205
Serpula, 154
Serpulite limestone, 74
Sertularia, 119, 122
Shales, 69
Sharks, 267, 269, 270, 271
Shell-limestone, 74
Shumardia, 227
Sigsbeia, 143
Siliceous rocks, 71
Silicified wood, 24
Siliquaria, 198
Silurian bivalves, 177
” brachiopods, 160
” brittle-stars, 142
” cephalopods, 206
” cirripedes, 241
” conodonts, 153
” corals, 113
” crinoids, 135
” Foraminifera, 96
” gasteropods, 193
” graptolites, Victoria, 128
” Hexacoralla, 114
” Octocoralla, 115
” Ostracoda, 235
” palaeechinoids, 144
” Phyllocarida, 246
” plants, 82
” Radiolaria, 102
” sponges, 109
” starfishes, 140
” stromatoporoids, 121
” trilobites, 228
Siphonalia, 198
Siphonia, 110
Siphonotreta, 160
SIRENIA, 298
Sistrum, 202
Slate, 70
Smith, William, 26
Smittia, 158
Solarium, 198
Solenocurtus, 187
Soletellina, 188
Sphaerosiderite, 80
Sphenopteris, 85, 89
Sphenotrochus, 118, 119
Sphenotus, 177, 179
Sphyrna, 270
Spirifer, 160, 161, 162, 163, 164
Spiriferina, 165
” -beds, 208
Spirillina, 96
Spirorbis, 154
Spirula, 205
Spirulirostra, 205, 210
Spisula, 188
Spondylostrobus, 91
Spondylus, 175, 184, 185
SPONGES, characteristics of, 64, 107
Spongilla, 72
Spongodiscus, 103
Spongophyllum, 116
Spoonbill, 283
Spore coal, 76
Squalodon, 295
Stacheia, 97
Star-corals, 119
Starfishes, characters of, 61, 139
Staurolonche, 103
Stauroneis, 92
Steno, 25
Stenopora, 117
Stenotheca, 192
Stephanella, 109
Stephanograptus, 126
Stephanotrochus, 118
Sthenurus, 290
Sting-ray, 271
Stomatopora, 158
Storing fossils, 320
Stork, 283
Strata, superposition of, 41
” vertically arranged, 44
Stratigraphical series, general thickness, 44
Stratigraphy, 27
Strepsodus, 261
Streptelasma, 113
« 335 »
Stricklandinia, 160
Stromatopora, 120, 121
Stromatoporella, 121, 122
STROMATOPOROIDS, 63, 120
Strombus, 184, 204
Strophalosia, 163
Stropheodonta, 160, 161
Strophonella, 160
Struthiolaria, 202
Studeria, 148
Sturtzura, 143
Stutchburia, 180
STYLASTERIDS, 119
Subemarginula, 198
Submerged forests, 30
Sunetta, 187
Superposition of strata, 41
Synaphe, 238
SYNDACTYLA, 288
Synedra, 92
Syringopora, 114
Syringothyris, 164
Tabellaria, 92
Taeniopteris, 88, 89, 164, 250, 265
Taniwhasaurus, 279
Taphaetus, 283
Tasmanian devil, 287, 295
” wolf, 287, 295
Tasmanite, 77
Taxocrinus, 135
Tellina, 185, 187, 188
Temnechinus, 146
Tentaculites, 193, 194, 195
Terebra, 198, 199, 202, 204
Terebratella, 166, 168
Terebratula, 166
Terebratulina, 166, 167
Tertiary ironstone, 81
Tessarodoma, 158
TETRACORALLA, 113
Tetractinellid sponge, 110, 112
Tetragraptus, 124, 126
Textularia, 98, 100
Thalassina, 248
THALLOPHYTA, characters of, 39
Thalotia, 200
Thamnastraea, 118
Thinnfeldia, 88, 89, 182
Thurammina, 97
Thyestes, 258
Thylacinus, 287, 295
Thylacoleo, 293, 303
Time-range of fossils, 50
Tomodus, 262
Toothed whales, 295
Torbanite, 77
Torlessia, 154
Trachyderma, 153, 154
Trachypora, 117
Trematonotus, 194
Trematotrochus, 118, 119
TREPOSTOMATA, 155
Tretocalia, 112
Triassic bivalves, 181
” brachiopods, 164
” cephalopods, 208
” crinoids, 137
” fishes, 262
” Foraminifera, 98
” labyrinthodonts, 273
” Ostracoda, 238
” Phyllopoda, 233
” plants, 88
” reptiles, New Zealand, 276
Tribonyx, 283
Tribrachiocrinus, 137
Trichograptus, 124
Tricoelocrinus, 139
Trigonia, 175, 182, 183, 184, 187
Trigonograptus, 126
TRILOBITES, habits of, 222
” structure of, 223
Tritylodon, 276, 286
Trivia, 198, 199
Trochoceras, 205
Trochonema, 195
Trochus, 191, 194, 195
Trophon, 202
« 336 »
Truncatulina, 98, 100
Tryplasma, 113
Tuatera, 276
Tudicla, 201
TUNICATA, 257
Turbo, 197, 200
Turrilepas, 241, 243
Turritella, 191, 198, 200, 201, 202
Turritella -limestone, 74
Tylosaurus, 279
Tylospira, 198, 202
Typhis, 198
Uncinulus, 162
Unio, 181, 182
Unionella, 181
Upper Cambrian trilobites, 227
” Cretaceous bivalves, 184
” Cretaceous brachiopod, 166
” Cretaceous cephalopod, 166
” Triassic fishes, 262
” Ordovician graptolites, New South Wales, 127
” Ordovician graptolites, Victoria, 126
Urasterella, 140
Urosthenes, 262
Vaginella, 198, 199
Vaginulina, 98
Valvulina, 97, 98
Venus, 177, 185, 187, 188
VERMES, characters of, 37
Vertebraria, 264
VERTEBRATA, characters of, 38, 257
Verticordia, 186
Vetotuba, 194
Voluta, 198, 201, 202
Volutilithes, 198, 201, 202
Volvox, 78
Volvulella, 201
Warrnambool footprints, 301
Werrikooian bivalves, 187
” gasteropods, 202
Whales, 295
White coal, 77
Wilsonia, 160
Wombat, 289, 295
Worms, fossil, 59, 152
Worm-tracks, 154
Wrasse family, 271
Wynyardia, 294
Xenophanes, 24
Xenorhynchus, 283
Xestoleberis, 237
Xiphosphaera, 103
Yvania, 196
Zaphrentis, 117
Ziphius, 296
Appended letters indicate the State or Country:—
N.S.W., New South Wales; N.T., Northern Territory; N.Z.,
New Zealand; Q., Queensland; S.A., South Australia; T.,
Tasmania; V., Victoria; W.A., Western Australia.
Adelaide, S.A., 102
Aire Coast, V., 138
Airly, N.S.W., 273
Alice Springs, S.A., 193
Altona Bay, V., 112
Arcola, Q., 279
Arcoona, S.A., 91
Ardrossan, S.A., 82, 107
Bacchus Marsh, V., 88, 90
Balcombe’s Bay, V., 190, 239, 317
Bald Hill, V., 88
Barker Gorge, W.A., 196, 232, 259
Barraba, N.S.W., 93, 102
Batesford, V., 73, 100, 138, 141
Baton River, N.Z., 195, 207
Bay of Islands, N.Z., 93
Beaumaris, V., 119, 243, 248, 270, 271, 296, 297, 317
Bendigo, V., 108, 109, 124, 246
Berwick, V., 68
Bindi, V., 109, 121, 161, 195
Bingera, N.S.W., 102
Boggy Creek, V., 112
Bowen River, Q., 117, 137, 164
Bowning, N.S.W., 144, 153, 207, 231, 241
Bowral, N.S.W., 274
Brighton, N.Z., 146, 248, 280
Broadhurst’s Creek, V., 231
Broken River, N.Z., 146, 167
Broken River, Q., 136
Broome, W.A., 183
Brunswick, V., 136
Buchan, V., 79, 109, 115, 136, 161, 195, 203, 207, 231, 237, 258
Bulla, V., 122
Bungonia, N.S.W., 300
Burdekin, Q., 115, 116
Burnt Creek, V., 259
Burragorang, N.S.W., 180
Camperdown, V., 74
Canobolas district, N.S.W., 114
Canowindra, N.S.W., 162
Canterbury, N.Z., 154
Cape Liptrap, V., 71
Cape Otway, V., 119, 296
Cape Palliser, N.Z., 203
Cape Paterson, V., 265, 276
Carapook, V., 264
Caroline Creek, T., 227
Casterton, V., 265
Castlemaine, V., 126, 246
Cavan, N.S.W., 109
Cessnock, N.S.W., 237
Chatham Ids., 138
Chillagoe, Q., 115
Chinchilla, Q., 279
Clarence Town, N.S.W., 139, 162
Cliftonwood, N.S.W., 237
Clunes, V., 279
Cockatoo Id., N.S.W., 274
Collie, W.A., 98
« 338 »
Collingwood, V., 206
Coole Barghurk Creek, V., 193
Cooma, N.S.W., 93, 102
Copeland, N.S.W., 85
Corio Bay, V., 270
Corner Creek, Q., 237
Croydon, Q., 89, 166
Curiosity Shop, N.Z., 138, 280
Curlewis, V., 112, 247
Curramulka, S.A., 108, 177, 192, 235
Currowang, N.S.W., 127
Dalton, N.S.W., 90, 91
Dargo High Plains, V., 91
Darling Downs, Q., 53, 110, 282, 283, 298
Darling River, N.S.W., 154, 157
Darriwill, V., 126
Delegate River, N.S.W., 114
Derrengullen Creek, N.S.W., 190
Diggers’ Rest, V., 126
Dolodrook River, V., 193, 227
Dromana, V., 246
Dundas Co., V., 264
East Maitland, N.S.W., 154
Elizabeth River, S.A., 91
Fanning River, Q., 207
Farley, N.S.W., 180, 237
Fernbrook, N.S.W., 109
Fifield, N.S.W., 237
Flemington, V., 136, 142, 143, 206, 318
Flinders, V., 65, 112
Flinders River, Q., 183, 250, 267, 277, 278
Florentine Valley, T., 159, 227
Fraser’s Creek, V., 231
Gascoyne River, W.A., 117, 136, 137, 232, 262
Geelong, V., 100, 119, 120, 243
Geilston, T., 203
Gellibrand River, V., 199
Geraldton, W.A., 98, 197, 238
Gippsland Lakes, V., 168, 243
Gisborne, V., 299
Glenelg River, V., 168
Glenwilliam, N.S.W., 139
Goodradigbee River, N.S.W., 109
Goonoo, N.S.W., 85
Gordon River, T., 115
Gosford, N.S.W., 53, 262, 263, 273
Grampians, V., 261
Grange Burn, Hamilton, V., 143, 270, 271, 296, 297
Greenough River, W.A., 165, 182, 209
Grey River, N.Z., 78
Grice’s Creek, V., 317
Grose Vale, N.S.W., 238
Gulgong, N.S.W., 279, 286
Gunning, N.S.W., 91
Haddon, V., 68
Hallett’s Cove, S.A., 119
Hall’s Sound, Papua, 201
Hamilton, N.Z., 285
Hamilton, V., 190, 243, 270, 271, 295, 296, 297
Hamilton River, Q., 267
Hatton’s Corner, N.S.W., 114, 231
Heathcote, V., 160, 177, 227
Hobart, T., 68, 203
Hokonui Hills, N.Z., 164, 165
Hughenden, Q., 267, 268
Iguana Creek, V., 85
Irwin River, W.A., 97, 98, 137, 207
Island of Timor, 163
« 339 »
Jenolan Caves, N.S.W., 102, 121, 300
Kakanui, N.Z., 280
Kamileroy, Q., 267
Keilor, V., 128
Kent’s Group, T., 203
Kilmore, V., 144, 206, 231, 246
Kilmore Creek, V., 231
Kimberley, W.A., 136, 137, 192, 207, 262
King Island, T., 53, 104, 283
King’s Creek, Q., 282
Kirrak, V., 265
Knocklofty, T., 264
Knowsley, V., 227
Koroit, V., 305
Kowhai River, N.Z., 189
Lake Callabonna, S.A., 51, 282
Lake Connewarre, V., 270
Lake Eyre, S.A., 166, 183, 189, 197
Lake Frome, S.A., 91
Lancefield, V., 93, 108, 122, 124, 246
Laurie’s Creek, S.A., 193, 205, 228
Lawson, N.S.W., 127
Leichhardt River, Q., 267
Leigh’s Creek, S.A., 193
Lennard River, W.A., 208
Lilydale, V., 73, 82, 96, 114, 121, 190, 229, 231, 236, 243, 318
Limeburners Point, V., 79
Limestone Creek, Glenelg River, V., 202
Limestone Creek, Yass, N.S.W.; 136, 231
Loddon Valley, V., 279
Lord Howe Id., 279
Loyola, V., 109, 121, 229, 231
Lyndhurst, N.S.W., 227
Maddingley, V., 90
Mallee, V., 71, 101, 119, 138, 141
Mandurama, N.S.W., 102, 127, 227
Manly, N.S.W., 88
Mansfield, V., 53, 122, 154, 231, 259
Marathon Station, Q., 277
Maria Id., T., 180
Maryborough, Q., 146, 184, 304
Maryvale Creek, Q., 279
McMahon’s Creek, V., 207
Melbourne, V., 82, 136, 140, 153, 178, 246
Mersey River, T., 77, 97, 193
Milburn, N.Z., 296
Mitchell Downs, Q., 137
Mitta Mitta River, V., 114
Molong, N.S.W., 114
Moonee Ponds Creek, V., 229, 318
Moorabool River, V., 112, 120, 202
Mornington, V., 65, 70, 90, 112, 118, 258, 269
Mosquito Plains, S.A., 300
Mount Angas, Q., 166
” Buninyong, V., 303
” Gambier, S.A., 71, 91, 119, 120, 138, 147, 282, 296
” Lambie, N.S.W., 85
” Macedon Cave, 298
” Potts, N.Z., 276
” Victoria, N.S.W., 88
” Wellington, V., 126, 134, 159, 193
” Wyatt, Q., 109
Muddy Creek, Hamilton, V., 141, 147, 243, 269, 295
Mudgee, N.S.W., 109
Muree, Raymond Terrace, N.S.W., 238
Murray River Cliffs, S.A., 58, 210
« 340 »
Murrumbidgee River, N.S.W., 114, 189, 259
Napier Range, W.A., 232
Narrengullen Creek, N.S.W., 237
Nelson, N.Z., 78, 126, 164, 165, 182, 233, 248
Newcastle, N.S.W., 233
Ngapara, N.Z., 296
Nimbin, Richmond River, N.S.W., 272
Norseman district, W.A., 110
Nugget Point, Otago, N.Z., 274
Nungatta, N.S.W., 85
Nyrang Creek, N.S.W., 162
Oakey Creek, N.S.W., 178
Oamaru, N.Z., 110, 280
Orakei Bay, N.Z., 158
Otway Coast, V., 90
Pakaraka, N.Z., 93
Papua, 100, 146, 148, 184, 187, 188, 201, 203, 209, 210
Paroo River, Q., 282
Peak Downs, Q., 282
Penola, S.A., 300
Petermann Creek, S.A., 193
Phillip Co., N.S.W., 282
Pine Creek, Q., 93
Pitfield Plains, V., 90
Pitchery Creek, Q., 278
Pokolbin, N.S.W., 97, 180
Port Campbell, V., 247
Port Darwin, N.T., 103, 248
Port Stephen, N.S.W., 262
Preservation Inlet, N.Z., 126
Ravensfield, N.S.W., 180
Reid Gap, Q., 207
Richmond Downs, Q., 267
Richmond River, N.S.W., 93
Rock Flat Creek, N.S.W., 206
Rockhampton, Q., 110, 139, 144, 153, 164, 196, 261
Rough Range, W.A., 116, 122
Sale, V., 112
San Remo, V., 122
Sebastopol, V., 93
Seville, V., 229, 231
Shakespeare Cliff, N.Z., 146
Southland, N.Z., 285
South Yarra, V., 128, 136, 143, 206, 229, 249, 318
Spring Creek, Torquay, V., 141
St. Peter’s, Sydney, N.S.W., 262
Stanwell, Q., 137
Stockyard Creek, N.S.W., 127
Stroud, N.S.W., 86
Studley Park, V., 128, 318
Sunbury, V., 126
Table Cape, T., 74, 190, 269, 270, 294, 296
Talbot, V., 93
Talbragar, 267
Tallong, N.S.W., 127
Tamworth, N.S.W., 85, 103, 115
Taranaki, N.Z., 203
Tempe Downs, S.A., 193, 205, 228
Thompson River, Q., 277
Thomson River, V., 229
Tinderbox Bay, T., 264
Tingaringi, N.S.W., 127
Toongabbie, V., 74, 135
Torquay, V., 74, 141, 148, 243, 269, 296
Tyer’s River, V., 82, 144
Upper Finke Basin, S.A., 159
« 341 »
Upper Yarra, V., 206, 207, 231, 236
Vegetable Creek, N.S.W., 91
Waihao, N.Z., 296
Waikari River, N.Z., 141
Waikouaiti, N.Z., 296
Wairoa, N.Z., 274
Wairoa Gorge, N.Z., 137, 162
Waitaki Valley, N.Z., 296
Walhalla, V., 114, 121, 128
Wandong, V., 229, 231
Wanganui, N.Z., 299
Wannon River district, V., 53, 90
Waratah Bay, V., 114, 121, 229
Warburton, V., 207
Warrnambool, V., 282, 299, 301, 302
Waurn Ponds, V., 90, 119, 141, 243, 269, 296
Wellington Valley, N.S.W., 287, 298, 300
Well’s Creek, N.Z., 165
West Melbourne Swamp, V., 51
Westport, N.Z., 78
Wharekuri, N.Z., 248
White Cliffs, N.S.W., 138, 179, 183, 184, 195, 279
Whittlesea, V., 206
Wilberforce, N.Z., 189
Wilcannia, N.S.W., 138
Wirrialpa, S.A., 159
Wollumbilla, Q., 98, 137, 154, 157, 166, 183, 189
Wombat Creek, V., 109, 126
Woori Yallock Creek, V., 231
Wormbete Creek, V., 74
Wynyard, T., 246
Yan Yean, V., 318
Yass, N.S.W., 65, 109, 114, 121, 153, 161, 179, 190, 207, 231, 237, 241
Yering, V., 142
Yorke Peninsula, S.A., 226
Yule Id., Papua, 146, 187, 201
Zeehan, T., 154
CORRIGENDA.
Page 65, for head-line “Protozoa” read “How Fossils are Found.”
Page 147, for head-line “Characteristic Fossils” read “Sea-urchins.”
Page 273, for head-line “Reptiles” read “Amphibians.”
[Transcriber Note: These changes were not utilized here as they only apply to the titles at the top of the printed pages.]
ERRATUM—Page 47.
In 1st column—for “Mesozoic or Secondary (continued).”
Read “Palaeozoic or Primary” and omit divisional line.
[Transcriber Note: These changes were applied to the text.]
Transcriber Note
Images were moved so paragraphs were not split. Minor typographical errors were corrected. Hyphenation was standardized to the most prevalent form utilized. As the æ ligature was only used five times and “ae” was used more than 50 times, the ligature was converted to "ae".
End of Project Gutenberg's Australasian Fossils, by Frederick Chapman
*** END OF THIS PROJECT GUTENBERG EBOOK AUSTRALASIAN FOSSILS ***
***** This file should be named 59074-h.htm or 59074-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/5/9/0/7/59074/
Produced by MFR, Tom Cosmas and the Online Distributed
Proofreading Team at http://www.pgdp.net (This file was
produced from images generously made available by The
Internet Archive)
Updated editions will replace the previous one--the old editions will
be renamed.
Creating the works from print editions not protected by U.S. copyright
law means that no one owns a United States copyright in these works,
so the Foundation (and you!) can copy and distribute it in the United
States without permission and without paying copyright
royalties. Special rules, set forth in the General Terms of Use part
of this license, apply to copying and distributing Project
Gutenberg-tm electronic works to protect the PROJECT GUTENBERG-tm
concept and trademark. Project Gutenberg is a registered trademark,
and may not be used if you charge for the eBooks, unless you receive
specific permission. If you do not charge anything for copies of this
eBook, complying with the rules is very easy. You may use this eBook
for nearly any purpose such as creation of derivative works, reports,
performances and research. They may be modified and printed and given
away--you may do practically ANYTHING in the United States with eBooks
not protected by U.S. copyright law. Redistribution is subject to the
trademark license, especially commercial redistribution.
START: FULL LICENSE
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full
Project Gutenberg-tm License available with this file or online at
www.gutenberg.org/license.
Section 1. General Terms of Use and Redistributing Project
Gutenberg-tm electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or
destroy all copies of Project Gutenberg-tm electronic works in your
possession. If you paid a fee for obtaining a copy of or access to a
Project Gutenberg-tm electronic work and you do not agree to be bound
by the terms of this agreement, you may obtain a refund from the
person or entity to whom you paid the fee as set forth in paragraph
1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this
agreement and help preserve free future access to Project Gutenberg-tm
electronic works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the
Foundation" or PGLAF), owns a compilation copyright in the collection
of Project Gutenberg-tm electronic works. Nearly all the individual
works in the collection are in the public domain in the United
States. If an individual work is unprotected by copyright law in the
United States and you are located in the United States, we do not
claim a right to prevent you from copying, distributing, performing,
displaying or creating derivative works based on the work as long as
all references to Project Gutenberg are removed. Of course, we hope
that you will support the Project Gutenberg-tm mission of promoting
free access to electronic works by freely sharing Project Gutenberg-tm
works in compliance with the terms of this agreement for keeping the
Project Gutenberg-tm name associated with the work. You can easily
comply with the terms of this agreement by keeping this work in the
same format with its attached full Project Gutenberg-tm License when
you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are
in a constant state of change. If you are outside the United States,
check the laws of your country in addition to the terms of this
agreement before downloading, copying, displaying, performing,
distributing or creating derivative works based on this work or any
other Project Gutenberg-tm work. The Foundation makes no
representations concerning the copyright status of any work in any
country outside the United States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other
immediate access to, the full Project Gutenberg-tm License must appear
prominently whenever any copy of a Project Gutenberg-tm work (any work
on which the phrase "Project Gutenberg" appears, or with which the
phrase "Project Gutenberg" is associated) is accessed, displayed,
performed, viewed, copied or distributed:
This eBook is for the use of anyone anywhere in the United States and
most other parts of the world at no cost and with almost no
restrictions whatsoever. You may copy it, give it away or re-use it
under the terms of the Project Gutenberg License included with this
eBook or online at www.gutenberg.org. If you are not located in the
United States, you'll have to check the laws of the country where you
are located before using this ebook.
1.E.2. If an individual Project Gutenberg-tm electronic work is
derived from texts not protected by U.S. copyright law (does not
contain a notice indicating that it is posted with permission of the
copyright holder), the work can be copied and distributed to anyone in
the United States without paying any fees or charges. If you are
redistributing or providing access to a work with the phrase "Project
Gutenberg" associated with or appearing on the work, you must comply
either with the requirements of paragraphs 1.E.1 through 1.E.7 or
obtain permission for the use of the work and the Project Gutenberg-tm
trademark as set forth in paragraphs 1.E.8 or 1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any
additional terms imposed by the copyright holder. Additional terms
will be linked to the Project Gutenberg-tm License for all works
posted with the permission of the copyright holder found at the
beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including
any word processing or hypertext form. However, if you provide access
to or distribute copies of a Project Gutenberg-tm work in a format
other than "Plain Vanilla ASCII" or other format used in the official
version posted on the official Project Gutenberg-tm web site
(www.gutenberg.org), you must, at no additional cost, fee or expense
to the user, provide a copy, a means of exporting a copy, or a means
of obtaining a copy upon request, of the work in its original "Plain
Vanilla ASCII" or other form. Any alternate format must include the
full Project Gutenberg-tm License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works
provided that
* You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is owed
to the owner of the Project Gutenberg-tm trademark, but he has
agreed to donate royalties under this paragraph to the Project
Gutenberg Literary Archive Foundation. Royalty payments must be paid
within 60 days following each date on which you prepare (or are
legally required to prepare) your periodic tax returns. Royalty
payments should be clearly marked as such and sent to the Project
Gutenberg Literary Archive Foundation at the address specified in
Section 4, "Information about donations to the Project Gutenberg
Literary Archive Foundation."
* You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or destroy all
copies of the works possessed in a physical medium and discontinue
all use of and all access to other copies of Project Gutenberg-tm
works.
* You provide, in accordance with paragraph 1.F.3, a full refund of
any money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days of
receipt of the work.
* You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project
Gutenberg-tm electronic work or group of works on different terms than
are set forth in this agreement, you must obtain permission in writing
from both the Project Gutenberg Literary Archive Foundation and The
Project Gutenberg Trademark LLC, the owner of the Project Gutenberg-tm
trademark. Contact the Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
works not protected by U.S. copyright law in creating the Project
Gutenberg-tm collection. Despite these efforts, Project Gutenberg-tm
electronic works, and the medium on which they may be stored, may
contain "Defects," such as, but not limited to, incomplete, inaccurate
or corrupt data, transcription errors, a copyright or other
intellectual property infringement, a defective or damaged disk or
other medium, a computer virus, or computer codes that damage or
cannot be read by your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium
with your written explanation. The person or entity that provided you
with the defective work may elect to provide a replacement copy in
lieu of a refund. If you received the work electronically, the person
or entity providing it to you may choose to give you a second
opportunity to receive the work electronically in lieu of a refund. If
the second copy is also defective, you may demand a refund in writing
without further opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS', WITH NO
OTHER WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT
LIMITED TO WARRANTIES OF MERCHANTABILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of
damages. If any disclaimer or limitation set forth in this agreement
violates the law of the state applicable to this agreement, the
agreement shall be interpreted to make the maximum disclaimer or
limitation permitted by the applicable state law. The invalidity or
unenforceability of any provision of this agreement shall not void the
remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in
accordance with this agreement, and any volunteers associated with the
production, promotion and distribution of Project Gutenberg-tm
electronic works, harmless from all liability, costs and expenses,
including legal fees, that arise directly or indirectly from any of
the following which you do or cause to occur: (a) distribution of this
or any Project Gutenberg-tm work, (b) alteration, modification, or
additions or deletions to any Project Gutenberg-tm work, and (c) any
Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of
computers including obsolete, old, middle-aged and new computers. It
exists because of the efforts of hundreds of volunteers and donations
from people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future
generations. To learn more about the Project Gutenberg Literary
Archive Foundation and how your efforts and donations can help, see
Sections 3 and 4 and the Foundation information page at
www.gutenberg.org
Section 3. Information about the Project Gutenberg Literary Archive Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg Literary
Archive Foundation are tax deductible to the full extent permitted by
U.S. federal laws and your state's laws.
The Foundation's principal office is in Fairbanks, Alaska, with the
mailing address: PO Box 750175, Fairbanks, AK 99775, but its
volunteers and employees are scattered throughout numerous
locations. Its business office is located at 809 North 1500 West, Salt
Lake City, UT 84116, (801) 596-1887. Email contact links and up to
date contact information can be found at the Foundation's web site and
official page at www.gutenberg.org/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To SEND
DONATIONS or determine the status of compliance for any particular
state visit www.gutenberg.org/donate
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations. To
donate, please visit: www.gutenberg.org/donate
Section 5. General Information About Project Gutenberg-tm electronic works.
Professor Michael S. Hart was the originator of the Project
Gutenberg-tm concept of a library of electronic works that could be
freely shared with anyone. For forty years, he produced and
distributed Project Gutenberg-tm eBooks with only a loose network of
volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as not protected by copyright in
the U.S. unless a copyright notice is included. Thus, we do not
necessarily keep eBooks in compliance with any particular paper
edition.
Most people start at our Web site which has the main PG search
facility: www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.