Project Gutenberg's Anatomy of the Cat, by Jacob Reighard and H. S. Jennings
This eBook is for the use of anyone anywhere in the United States and most
other parts of the world at no cost and with almost no restrictions
whatsoever. You may copy it, give it away or re-use it under the terms of
the Project Gutenberg License included with this eBook or online at
www.gutenberg.org. If you are not located in the United States, you'll have
to check the laws of the country where you are located before using this ebook.
Title: Anatomy of the Cat
Author: Jacob Reighard
H. S. Jennings
Illustrator: Louise Burridge Jennings
Release Date: December 1, 2018 [EBook #58394]
Language: English
Character set encoding: ISO-8859-1
*** START OF THIS PROJECT GUTENBERG EBOOK ANATOMY OF THE CAT ***
Produced by deaurider, Harry Lamé and the Online Distributed
Proofreading Team at http://www.pgdp.net (This file was
produced from images generously made available by The
Internet Archive)
Please see the Transcriber’s Notes at the end of this text.
For some illustrations, larger versions are available by clicking the link in the illustration caption
(Fig. nnn) (not available in all formats).
ANATOMY OF THE CAT
BY
JACOB REIGHARD
Professor of Zoology in the University of Michigan
AND
H. S. JENNINGS
Instructor in Zoology in the University of Michigan
WITH
ONE HUNDRED AND SEVENTY-THREE ORIGINAL FIGURES
DRAWN BY
LOUISE BURRIDGE JENNINGS
NEW YORK
HENRY HOLT AND COMPANY
1901
Copyright, 1901,
BY
HENRY HOLT & CO.
ROBERT DRUMMOND, PRINTER, NEW YORK.
[iii]
PREFACE.
Although the cat has long been in common use for the
practical study of mammalian anatomy, a clear, correct, not
too voluminous account of its structure, such as should be in
the hands of students in the laboratory, has remained a
desideratum. A number of works have been published on the
cat, some of them of much value, yet there is none which
fulfils exactly the conditions mentioned. The books which
have appeared on this subject are the following:
1. Strauss-Durckheim, H. Anatomie descriptive et comparative
du Chat. 2 vols. Paris, 1845.
2. Mivart, St. George. The Cat: an Introduction to the
Study of Back-boned Animals, especially Mammals. New
York, 1881.
3. Wilder, Burt G., and Gage, Simon H. Anatomical
Technology as applied to the Domestic Cat. New York, 1882.
4. Gorham, F. P., and Tower, R. W. A Laboratory
Guide for the Dissection of the Cat. New York, 1895.
5. Jayne, H. Mammalian Anatomy. Vol. I. Philadelphia,
1898.
The first of these works treats only of the muscles and
bones, and is not available for American students. Its excellent
plates (or Williams’s outline reproductions of the same)
should be in every laboratory.
The second book named is written in such general terms
that its descriptions are not readily applicable to the actual
structures found in the dissection of the cat, and experience
has shown that it is not fitted for a laboratory handbook. It
contains, in addition to a general account of the anatomy of
the cat, also a discussion of its embryology, psychology,
palæontology, and classification.
[iv]
The book by Wilder and Gage professedly uses the cat as
a means of illustrating technical methods and a special system
of nomenclature. While of much value in many ways, it does
not undertake to give a complete account of the anatomy of
the animal.
The fourth work is a brief laboratory guide.
The elaborate treatise by Jayne, now in course of publication,
is a monumental work, which will be invaluable for reference,
but is too voluminous to place in the hands of students.
At present only the volume on the bones has been published.
As appears from the above brief characterization, none of
these books gives a complete description of the anatomy of the
cat in moderate volume and without extraneous matter. This
is what the present work aims to do.
In the year 1891-92, Professor Reighard prepared a partial
account of the anatomy of the cat, which has since been in use,
in typewritten form, in University of Michigan classes. It has
been used also at the Universities of Illinois, Nebraska, and
West Virginia, and in Dartmouth College, and has proven so
useful for college work in Mammalian Anatomy that it was
decided to complete it and prepare it for publication. This
has been done by Dr. Jennings.
The figures, which are throughout original, are direct reproductions
of ink drawings, made under the direction of Dr.
Jennings by Mrs. Jennings.
The book is limited to a description of the normal anatomy
of the cat. The direct linear action of each muscle taken alone
has been given in the description of muscles; other matters
belonging to the realm of physiology, as well as all histological
matter, have been excluded. It was felt that the monumental
work of Jayne on the anatomy of the cat, now in course of
publication, forms the best repository for a description of variations
and abnormalities, so that these have been mentioned in
the present volume only when they are so frequent as to be of
much practical importance.
Except where the contrary is stated, the descriptions are
based throughout on our own dissections and observations and
are in no sense a compilation. For this reason we have not[v]
thought it necessary to collect the scattered references to the
anatomy of the cat that may occur in the literature. A
collection of such references may be found in Wilder and
Gage’s Anatomical Technology. In addition to the works
already referred to, we have of course made use of the standard
works on human and veterinary anatomy. Among these
should be mentioned as especially useful the Anatomie des
Hundes by Ellenberger and Baum. Other publications which
have been of service in the preparation of the work are Windle
and Parson’s paper On the Myology of the Terrestrial Carnivora,
in the Proceedings of the Zoological Society of London
for 1897 and 1898, T. B. Stowell’s papers on the nervous
system of the cat in the Proceedings of the American Philosophical
Society (1881, 1886, 1888) and in the Journal of
Comparative Neurology (vol. I.), and F. Clasen’s Die Muskeln
und Nerven des proximalen Abschnitts der vorderen Extremität
der Katze, in Nova Acta der Ksl. Leop.-Carol. Deutschen
Akademie der Naturforscher, Bd. 64.
Nomenclature.—The question of nomenclature has been
one of difficulty. What is desired is a uniform set of
anatomical names,—a system that shall be generally used by
anatomists. At present the greatest diversity prevails as to
the names to be applied to the different structures of the body.
The only set of terms which at the present time seems to have
any chance of general acceptance is that proposed by the
German Anatomical Society at their meeting in Basel in 1895,
and generally designated by the abbreviation BNA. This
system has therefore been adopted, in its main features, for use
in the present work. It seems impossible at the present time,
however, to impose any one set of terms absolutely upon
anatomists of all nations, and we have felt it necessary to use
for certain familiar structures, in place of the BNA terms,
names that have come to have a fixed place in English
anatomy, and may almost be considered component parts of
the English language. The German anatomists have expressly
recognized the fact that this would be to a greater or less
degree necessary among anatomists of different nations, and
have characterized their list as for the present tentative, and[vi]
capable of farther development. The only purpose of a name
is that it shall furnish a key to a common understanding;
where the BNA name does not furnish such a key to English
readers, and where there is a term in established English usage
that does serve this purpose and seems unlikely to be supplanted,
we have used the latter. But we have endeavored to
make the number of these exceptions as small as possible, and
in such cases we have usually cited at the same time the term
proposed by the German society, followed by the abbreviation
BNA. When, on the other hand, we have adopted a BNA
term for which there is also a commonly used English equivalent,
the latter has likewise usually been cited in parenthesis.
In deciding whether or not to use in a given case the BNA
term many difficult cases arose. Will the common English
name innominate bone (os innominatum) be replaced by the
BNA term os coxæ or coxal bone? We have held this to be
highly improbable, and have therefore used the term innominate
bone, merely citing os coxæ (BNA) as a synonym. In the
same way we have used centrum as a designation of a part of a
vertebra, in place of corpus (BNA); premaxillary bone or premaxilla
in place of os incisivum (BNA); malar bone in place
of os zygomaticum (BNA); trapezoid as a name of one of the
bones of the carpus, in place of os multangulum minus (BNA),
etc. In other cases where it has seemed probable that the
BNA term would come into common use, though now unfamiliar,
this and the more common English expression are
both used or used alternatively; such has been the case, for
example, with the Gasserian ganglion or semilunar ganglion
(BNA). In naming the cerebral sulci and gyri the system in
use for man is not well fitted for bringing out the plan of those
in the brain of the cat, so that it was necessary to reject the
BNA names for these structures.
As to the use of the Latin terms and their equivalents in
English form, we have made a practice of employing in the
text sometimes one, sometimes the other; this has the advantage
of giving variety, and of impressing the interchangeability
of the Latin and English forms on the mind of the student.
Where a given structure is called by two equally well-known[vii]
names, we have used both, holding that the student should
become familiar with each and recognize their identity of
meaning.
In general we have maintained the principle that the
primary purpose of such a work as the present is not to illustrate
or defend any particular system of nomenclature, but to
aid in obtaining a knowledge of the structures themselves.
With this end in view, we have used such terms as would in
our judgment best subserve this purpose, making the BNA
system, as the one most likely to prevail, our basis. In applying
the system we have had to keep in mind a number of
sometimes conflicting principles. In some cases the judgment
of other anatomists will doubtless differ from our own; but this
we feel to be inevitable. The matter of an absolutely uniform
nomenclature is not ripe for settlement at the present time.
Some further explanation is needed in regard to the topographical
terms, or terms of direction, used in the present
work. We have adopted the BNA terms in this matter also.
The terms superior, inferior, anterior, and posterior have been
avoided, as these terms do not convey the same meaning in
the case of the cat as they do in man, owing to the difference
in the posture of the body. In place of these terms are used
dorsal and ventral, cranial and caudal. As terms of direction
these, of course, must have an absolutely fixed meaning, signifying
always the same direction without necessary reference
to any given structure. For example, cranial means not
merely toward the cranium, but refers to the direction which is
indicated by movement along a line from the middle of the
body, toward the cranium; after the head or cranium is
reached, the term still continues in force for structures even
beyond the cranium. Thus the tip of the nose is considered
to be craniad of the cranium itself. Lateral signifies away
from the middle plane; medial toward it. Inner and outer or
internal and external are used only with reference to the structure
of separate organs, not with reference to the median plane
of the body.
In describing the limbs the convexity of the joint (the elbow
or knee) is considered as dorsal, the concavity being therefore[viii]
ventral. Medial refers to that side of the limb which in the
normal position is toward the middle of the body; lateral to
the outer side. Terms of direction which are derived only from
the structure of the limb itself are in some cases more convenient
than the usual ones. In the fore limbs the terms radial
(referring to the side on which the radius lies) and ulnar
(referring to the side on which the ulna lies) are used; in the
hind limbs the terms tibial and fibular are used in a similar
manner. Distal means toward the free end of a limb or other
projecting structure; proximal, toward the attached end.
For all these terms an adverbial form ending in -ad has
been employed. Experience has shown this to be very useful
in practice, and while not expressly recommended by the BNA,
it is not condemned. Terms ending in -al are therefore
adjectives; those ending in -ad are adverbs.
In compounding these terms of direction, the hyphen has
been omitted in accordance with the usage recommended by
the Standard Dictionary. Thus dorsoventral is written in
place of dorso-ventral, etc. The student will perhaps be
assisted in understanding these compounds if he notes that
the first component always ends in -o, so that the letter o practically
serves the purpose of a hyphen in determining how the
word is to be divided.
In one particular the BNA nomenclature is not entirely
consistent. While recommending or at least permitting the
use of the general terms dorsal and ventral in place of the
human posterior and anterior, and cranial and caudal in place
of superior and inferior, it retains the words anterior, posterior,
superior, and inferior as parts of the names of definite organs.
For example, we have the muscle serratus anterior in place
of serratus ventralis; serratus posterior inferior in place of
serratus dorsalis caudalis. This is very unfortunate, from a
comparative standpoint, but we have felt it necessary to retain
the BNA terms in order that the structures of the cat may
receive the same names as the corresponding structures of
man.
In the matter of orthography we have endeavored to follow
the best English anatomical usage, as exemplified in Gra[ix]y’s
Human Anatomy,—therefore writing peroneus in place of
peronæus, pyriformis in place of piriformis, etc.
The book is designed for use in the laboratory, to accompany
the dissection and study of the structures themselves.
Anatomy cannot be learned from a book alone, and no one
should attempt to use the present work without at the same
time carefully dissecting the cat. On the other hand, anatomy
can scarcely be learned without descriptions and figures of the
structures laid bare in dissection, so that this or some similar
work should be in the hands of any one attempting to gain a
knowledge of anatomy through the dissection of the cat.
The figures have all been drawn from actual dissections,
and have been carefully selected with a view to furnishing the
most direct assistance to the dissector. It is hoped that no
figures are lacking that are required for giving the students the
necessary points of departure for an intelligent dissection of any
part of the body. The fore limb is illustrated somewhat more
fully than the hind limb, because it was thought that the fore
limb would usually be dissected first; the hind limb will be
easily dissected, with the aid of the figures given, after the
experience gained in dissecting the fore limb.
As the book is designed to accompany the dissection of the
specimen in the laboratory, it was deemed best to give succinct
specific directions for the dissection of the different systems of
organs, together with suggestions as to methods of preserving
and handling the material. These are included in an appendix.
[x]
[xi]
CONTENTS.
| |
|
|
|
PAGE |
| The Skeleton of the Cat |
1 |
| |
I. The Vertebral Column |
1 |
| |
Thoracic Vertebræ |
1 |
| |
Lumbar Vertebræ |
7 |
| |
Sacral Vertebræ: Sacrum |
8 |
| |
Caudal Vertebræ |
11 |
| |
Cervical Vertebræ |
11 |
| |
Ligaments of the Vertebral Column |
16 |
| |
II. The Ribs |
18 |
| |
III. The Sternum |
20 |
| |
IV. The Skull |
21 |
| |
Occipital Bone |
22 |
| |
Interparietal |
25 |
| |
Sphenoid |
25 |
| |
Presphenoid |
29 |
| |
Temporal |
30 |
| |
Parietal |
36 |
| |
Frontal |
37 |
| |
Maxillary |
39 |
| |
Premaxillary |
41 |
| |
Nasal |
42 |
| |
Ethmoid |
42 |
| |
Vomer |
44 |
| |
Palatine |
45 |
| |
Lachrymal |
46 |
| |
Malar |
47 |
| |
Mandible |
47 |
| |
Hyoid |
49 |
| |
The Skull as a Whole |
49 |
| |
Cavities of the Skull |
57 |
| |
Joints and Ligaments of the Skull |
61 |
| |
V. The Thoracic Extremities |
62 |
| |
Scapula |
62 |
| |
Clavicle |
64 |
| |
Humerus |
64 |
| |
Radius |
67 |
| |
Ulna |
68 |
| |
Carpus |
69 |
| |
Bones[xii] of the Hand |
71 |
| |
Joints and Ligaments of the Thoracic Limbs |
73 |
| |
VI. The Pelvic Extremities |
76 |
| |
Innominate Bones |
76 |
| |
Femur |
79 |
| |
Patella |
80 |
| |
Tibia |
80 |
| |
Fibula |
82 |
| |
Tarsus |
82 |
| |
Bones of the Foot |
85 |
| |
Joints and Ligaments of the Pelvic Limbs |
86 |
| The Muscles |
93 |
| |
I. Muscles of the Skin |
93 |
| |
II. Muscles of the Head |
96 |
| |
A. Superficial Muscles |
96 |
| |
B. Deep Muscles |
107 |
| |
a. Muscles of Mastication |
107 |
| |
b. Muscles of Hyoid Bone |
112 |
| |
III. Muscles of the Body |
115 |
| |
1. Muscles of the Back |
115 |
| |
A. Muscles of the Shoulder |
115 |
| |
B. Muscles of the Vertebral Column |
123 |
| |
a. Muscles of the Lumbar and Thoracic Region |
126 |
| |
b. Dorsal Muscles of the Cervical Region |
131 |
| |
C. Muscles of the Tail |
136 |
| |
2. Muscles on the Ventral Side of the Vertebral Column |
138 |
| |
A. Lumbar and Thoracic Regions |
138 |
| |
B. Muscles on the Ventral Side of the Neck |
139 |
| |
3. Muscles of the Thorax |
144 |
| |
A. Breast Muscles (Connecting the Arm and Thorax) |
144 |
| |
B. Muscles of the Wall of the Thorax |
148 |
| |
4. Abdominal Muscles |
153 |
| |
IV. Muscles of the Thoracic Limbs |
156 |
| |
1. Muscles of the Shoulder |
156 |
| |
A. Lateral Surface |
156 |
| |
B. Medial Surface |
161 |
| |
2. Muscles of the Brachium or Upper Arm |
164 |
| |
3. Muscles of the Antibrachium or Forearm |
172 |
| |
Fascia of the Forearm |
172 |
| |
A. Muscles on the Ulnar and Dorsal Side of the Forearm |
173 |
| |
B. Muscles on the Radial and Ventral Side of the Forearm |
179 |
| |
4. Muscles of the Hand |
184 |
| |
A. Between the Tendons |
184 |
| |
B. Muscles of the Thumb |
184 |
| |
C. Between the Metacarpals |
185 |
| |
D. Special Muscles of the Second Digit |
185 |
| |
E. Special Muscles of the Fifth Digit |
185 |
| |
V. Muscles of the Pelvic Limbs |
186 |
| |
1. Muscles of the Hip |
186 |
| |
A. On the Lateral Surface of the Hip[xiii] |
186 |
| |
Fascia of the Thigh |
186 |
| |
B. On the Medial Surface of the Hip |
192 |
| |
2. Muscles of the Thigh |
194 |
| |
3. Muscles of the Lower Leg |
203 |
| |
A. On the Ventral Side |
203 |
| |
B. On the Dorsal and Lateral Surfaces |
209 |
| |
4. Muscles of the Foot |
212 |
| |
A. Muscles on the Dorsum of the Foot |
212 |
| |
B. Muscles on the Sole of the Foot |
212 |
| |
C. Muscles of the Tarsus |
215 |
| The Viscera |
217 |
| |
I. The Body Cavity |
217 |
| |
II. Alimentary Canal |
221 |
| |
1. Mouth |
221 |
| |
Glands of the Mouth |
223 |
| |
Teeth |
224 |
| |
Tongue |
226 |
| |
Muscles of the Tongue |
228 |
| |
Soft Palate |
229 |
| |
Muscles of the Soft Palate |
230 |
| |
2. Pharynx |
231 |
| |
Muscles of the Pharynx |
232 |
| |
3. Œsophagus |
234 |
| |
4. Stomach |
234 |
| |
5. Small Intestine |
236 |
| |
6. Large Intestine |
237 |
| |
7. Liver, Pancreas, and Spleen |
239 |
| |
III. Respiratory Organs |
243 |
| |
1. Nasal Cavity |
243 |
| |
2. Larynx |
246 |
| |
Cartilages of the Larynx |
247 |
| |
Muscles of the Larynx |
249 |
| |
3. Trachea |
251 |
| |
4. Lungs |
252 |
| |
Thyroid Gland |
254 |
| |
Thymus Gland |
254 |
| |
IV. Urogenital System |
255 |
| |
1. Excretory Organs |
255 |
| |
Kidneys |
255 |
| |
Ureter |
256 |
| |
Bladder |
256 |
| |
(Suprarenal Bodies) |
257 |
| |
2. Genital Organs |
257 |
| |
A. Male |
257 |
| |
B. Female |
263 |
| |
Muscles of the Urogenital Organs, Rectum, and Anus |
268 |
| |
a. Muscles Common to the Male and Female |
268 |
| |
b. Muscles Peculiar to the Male |
271 |
| |
c. Muscles Peculiar to the Female[xiv] |
272 |
| The Circulatory System |
274 |
| |
I. The Heart |
274 |
| |
II. The Arteries |
280 |
| |
1. Pulmonary Artery |
280 |
| |
2. Aorta |
281 |
| |
A. Thoracic Aorta and its Branches |
281 |
| |
Common Carotid Artery |
283 |
| |
Subclavian Artery |
290 |
| |
B. Abdominal Aorta and its Branches |
301 |
| |
External Iliac Artery and its Branches |
309 |
| |
III. The Veins |
315 |
| |
1. Veins of the Heart |
315 |
| |
2. Vena Cava Superior and its Branches |
316 |
| |
Veins of the Brain and Spinal Cord |
324 |
| |
3. Vena Cava Inferior and its Branches |
325 |
| |
Portal Vein |
326 |
| |
IV. Lymphatic System |
330 |
| |
1. Lymphatics of the Head |
331 |
| |
2. Lymphatics of the Neck |
332 |
| |
3. Lymphatics of the Thoracic Limbs |
332 |
| |
4. Lymphatics of the Thorax and Abdomen |
333 |
| |
5. Lymphatics of the Pelvic Limbs |
334 |
| The Nervous System |
335 |
| |
I. The Central Nervous System |
336 |
| |
1. Spinal Cord |
336 |
| |
2. The Brain |
339 |
| |
(1) Myelencephalon |
344 |
| |
(2) Metencephalon |
347 |
| |
(3) Mesencephalon |
351 |
| |
(4) Diencephalon |
352 |
| |
(5) Telencephalon |
357 |
| |
II. The Peripheral Nervous System |
369 |
| |
1. Cranial Nerves |
369 |
| |
I. Olfactory Nerve |
369 |
| |
II. Optic Nerve |
369 |
| |
III. Oculomotor Nerve |
369 |
| |
IV. Trochlear Nerve |
370 |
| |
V. Trigeminal Nerve |
370 |
| |
VI. Abducens |
375 |
| |
VII. Facial Nerve |
375 |
| |
VIII. Auditory Nerve |
377 |
| |
IX. Glossopharyngeal Nerve |
378 |
| |
X. Vagus Nerve |
378 |
| |
XI. Accessory Nerve |
382 |
| |
XII. Hypoglossal Nerve |
383 |
| |
2. Spinal Nerves |
383 |
| |
A. Cervical Nerves |
383 |
| |
The Brachial Plexus |
386 |
| |
B. Thoracic Nerves[xv] |
393 |
| |
C. Lumbar Nerves |
394 |
| |
Lumbar Plexus |
395 |
| |
D. Sacral Nerves and Sacral Plexus |
399 |
| |
E. Nerves of the Tail |
404 |
| |
3. Sympathetic System |
404 |
| Sense Organs and Integument |
409 |
| |
I. The Eye |
409 |
| |
II. The Ear |
415 |
| |
III. Olfactory Organ |
426 |
| |
IV. Organ of Taste |
426 |
| |
V. Integument |
427 |
| Appendix: Practical Directions |
429 |
| Index |
473 |
[xvi]
[xvii]
LIST OF ILLUSTRATIONS.
| FIG. |
PAGE |
| 1. |
Skeleton |
2 |
| 2. |
Fourth Thoracic Vertebra |
3 |
| 3. |
Fourth Thoracic Vertebra |
3 |
| 4. |
Thoracic Vertebræ |
5 |
| 5. |
Lumbar Vertebræ |
7 |
| 6. |
Sacrum |
9 |
| 7. |
Sacrum |
9 |
| 8. |
Caudal Vertebra |
11 |
| 9. |
Caudal Vertebra |
11 |
| 10. |
Cervical Vertebræ |
12 |
| 11. |
Sixth Cervical Vertebra |
13 |
| 12. |
Atlas |
13 |
| 13. |
Axis |
15 |
| 14. |
Ligaments of the Odontoid Process |
18 |
| 15. |
Rib |
19 |
| 16. |
Sternum |
20 |
| 17. |
Occipital Bone |
22 |
| 18. |
Occipital Bone |
22 |
| 19. |
Interparietal |
25 |
| 20. |
Sphenoid |
25 |
| 21. |
Presphenoid |
29 |
| 22. |
Temporal |
31 |
| 23. |
Temporal |
31 |
| 24. |
Tympanic Bulla |
33 |
| 25. |
Petrous Bone |
34 |
| 26. |
Frontal |
37 |
| 27. |
Maxillary Bone |
39 |
| 28. |
Maxillary Bone |
39 |
| 29. |
Premaxillary |
41 |
| 30. |
Nasal |
42 |
| 31. |
Ethmoid and Vomer |
43 |
| 32. |
Ethmoid and Vomer |
43 |
| 33. |
Palatine |
45 |
| 34. |
Lachrymal |
46 |
| 35. |
Malar |
46 |
| 36. |
Mandible |
48 |
| 37. |
Mandible |
48 |
| 38. |
Hyoid |
49 |
| 39. |
Skull, Dorsal Surface[xviii] |
50 |
| 40. |
Skull, Side View |
53 |
| 41. |
Skull, Ventral Surface |
55 |
| 42. |
Cavities of Skull |
57 |
| 43. |
Skull, Median Section |
60 |
| 44. |
Scapula |
62 |
| 45. |
Scapula |
62 |
| 46. |
Clavicle |
64 |
| 47. |
Humerus |
65 |
| 48. |
Humerus |
65 |
| 49. |
Radius and Ulna |
68 |
| 50. |
Radius and Ulna |
68 |
| 51. |
Bones of the Hand |
70 |
| 52. |
Ligaments of the Elbow |
74 |
| 53. |
Ligaments of the Elbow |
74 |
| 54. |
Innominate Bone of Kitten |
76 |
| 55. |
Innominate Bone |
77 |
| 56. |
Femur |
79 |
| 57. |
Tibia and Fibula |
81 |
| 58. |
Bones of the Foot |
83 |
| 59. |
Calcaneus |
83 |
| 60. |
Knee-joint |
89 |
| 61. |
Knee-joint |
89 |
| 62. |
Muscles of the Skin |
94 |
| 63. |
Muscles on Dorsal Side of Head |
97 |
| 64. |
Muscles of Face |
102 |
| 65. |
Ventral Muscles of Thorax, Neck, and Head |
109 |
| 66. |
Pterygoid and Palatal Muscles |
112 |
| 67. |
Muscles of Tongue, Hyoid, and Pharynx |
114 |
| 68. |
First Layer of Body Muscles |
117 |
| 69. |
Deep Muscles of the Vertebræ and Ribs |
125 |
| 70. |
Dorsal Muscles of Lumbar and Caudal Regions |
127 |
| 71. |
Deep Muscles of Neck |
135 |
| 72. |
Muscles on the Ventral Surface of the Cervical Vertebræ |
143 |
| 73. |
Second Layer of Body Muscles |
149 |
| 74. |
Diaphragm |
152 |
| 75. |
Lateral Muscles of Arm |
158 |
| 76. |
Origin of Lateral Muscles on Scapula |
160 |
| 77. |
Medial Muscles of Arm |
162 |
| 78. |
Origin of Medial Muscles on Scapula |
163 |
| 79. |
Deep Medial Muscles of Arm |
167 |
| 80. |
Deep Lateral Muscles of Arm |
169 |
| 81. |
Areas of Origin of Muscles on Ventral Surface of Humerus |
171 |
| 82. |
Areas of Origin of Muscles on Medial Side of Humerus |
171 |
| 83. |
Areas of Origin of Muscles on Dorsal Surface of Left Humerus |
171 |
| 84. |
Tendons on Back of Hand |
175 |
| 85. |
Deep Muscles of Forearm |
177 |
| 86. |
Insertions of Muscles on Radius and Ulna |
178 |
| 87. |
Insertions of Muscles on Radius and Ulna |
182 |
| 88. |
Mm. Lumbricales, etc.[xix] |
183 |
| 89. |
Deep Muscles of Palm of Hand |
184 |
| 90. |
Lateral Muscles of the Leg |
192 |
| 91. |
Medial Muscles of the Leg |
197 |
| 92. |
Deep Medial Muscles of Thigh |
200 |
| 93. |
Teeth of the Upper Jaw |
225 |
| 94. |
Teeth of the Lower Jaw |
226 |
| 95. |
Tongue, Epiglottis, etc. |
227 |
| 96. |
Muscles of Tongue, Hyoid, and Pharynx |
229 |
| 97. |
Stomach |
235 |
| 98. |
Colon and Cæcum |
238 |
| 99. |
Ileocolic Valve |
238 |
| 100. |
Liver |
240 |
| 101. |
Liver |
240 |
| 102. |
Pancreas and Spleen |
242 |
| 103. |
Cartilages of Nose |
244 |
| 104. |
Cartilages of Larynx |
247 |
| 105. |
Muscles of Larynx |
250 |
| 106. |
Bronchi |
253 |
| 107. |
Thymus Gland |
254 |
| 108. |
Kidney |
255 |
| 109. |
Section of Kidney |
255 |
| 110. |
Testis |
260 |
| 111. |
Male Genital Organs |
262 |
| 112. |
Female Urogenital Organs |
265 |
| 113. |
Muscles of Urogenital Organs and Anus in Male |
270 |
| 114. |
Muscles of Urogenital Organs of Female |
272 |
| 115. |
Heart |
276 |
| 116. |
Heart |
276 |
| 117. |
Inside of Heart |
278 |
| 118. |
Vessels of Thorax |
282 |
| 119. |
Common Carotid and Internal Jugular |
284 |
| 120. |
Branches of External Carotid |
288 |
| 121. |
Arteries of Brain |
291 |
| 122. |
Vessels and Nerves of the Axilla |
295 |
| 123. |
Vessels and Nerves of the Arm |
299 |
| 124. |
Palmar Arch |
301 |
| 125. |
Cœliac Artery |
302 |
| 126. |
Abdominal Blood-vessels |
305 |
| 127. |
Medial Vessels and Nerves of the Leg |
310 |
| 128. |
Deep Arteries of Foot |
314 |
| 129. |
Thoracic Blood-vessels |
317 |
| 130. |
Superficial Vessels and Nerves of the Forearm |
319 |
| 131. |
Blood-vessels of the Face |
322 |
| 132. |
Portal Vein |
327 |
| 133. |
Spinal Cord, cranial portion |
336 |
| 134. |
Section of Spinal Cord |
337 |
| 135. |
Origin of Spinal Nerves |
337 |
| 136. |
Cauda Equina, etc. |
338 |
| 137. |
Brain, Dorsal View[xx] |
340 |
| 138. |
Brain, Ventral View |
342 |
| 139. |
Diagram of Brain |
343 |
| 140. |
Diagram of Brain |
343 |
| 141. |
Dorsal View of Midbrain and ’Tween-brain |
350 |
| 142. |
Ventral View of Midbrain and ’Tween-brain |
352 |
| 143. |
Longitudinal Section of Brain |
356 |
| 144. |
Lateral View of Brain |
358 |
| 145. |
Diagram of Sulci and Gyri |
359 |
| 146. |
Diagram of Sulci and Gyri |
361 |
| 147. |
Corpus Callosum |
363 |
| 148. |
Fornix, Hippocampus, and Corpus Striatum |
364 |
| 149. |
Transverse Section of Brain |
366 |
| 150. |
Transverse Section of Brain |
366 |
| 151. |
Transverse Section of Brain |
366 |
| 152. |
Transverse Section of Brain |
367 |
| 153. |
Transverse Section of Brain |
367 |
| 154. |
Cranial Nerves |
374 |
| 155. |
Nerves of Face |
376 |
| 156. |
Cranial Nerves in the Neck |
379 |
| 157. |
Sympathetic and Vagus in the Thorax |
381 |
| 158. |
Nerves of the Neck |
384 |
| 159. |
Brachial Plexus |
387 |
| 160. |
Nerves and Vessels of Axilla |
389 |
| 161. |
Nerves and Vessels of Forearm |
391 |
| 162. |
Lumbar and Sacral Nerves |
398 |
| 163. |
Great Sciatic Nerve |
401 |
| 164. |
Sympathetic and Vagus in Abdomen |
407 |
| 165. |
Nictitating Membrane |
410 |
| 166. |
Muscles of Eyeball |
411 |
| 167. |
Diagram of Eye |
413 |
| 168. |
Cartilage of External Ear |
417 |
| 169. |
Muscles of External Ear |
419 |
| 170. |
Tympanic Membrane |
422 |
| 171. |
Malleus and Incus |
423 |
| 172. |
Stapes |
424 |
| 173. |
Membranous Labyrinth |
425 |
[1]
ANATOMY OF THE CAT.
THE SKELETON OF THE CAT.
The skeleton of the cat consists of 230 to 247 bones exclusive
of the sesamoid bones (44) and the chevron bones (8).
These are divided as follows: head 35-40, vertebral column
52-53, ribs 26, sternum 1-8, pelvis 2-8, upper extremities 62,
lower extremities 54-56. The number of bones varies with
the age of the individual, being fewer in the old than in the
young animal, owing to the fact that in an old animal some
bones that were originally separate have united.
I. THE VERTEBRAL COLUMN. COLUMNA VERTEBRALIS.
The vertebral column, spinal column, or back-bone, consists
of a varying number of separate bones, the vertebræ. At its
cranial end are seven vertebræ (cervical, Fig. 1, c) which are
without ribs and support the head; caudad of these are thirteen
rib-bearing vertebræ (thoracic, Fig. 1, m); caudad of these are
seven that are again without ribs (lumbar, Fig. 1, o); these
are followed by three vertebræ (sacral, Fig. 1, x) which are
united into a single bone, the sacrum, which supports the
pelvic arch. Following the sacral vertebræ are twenty-two
or twenty-three small ribless vertebræ which support the tail
(caudal, Fig. 1, y).
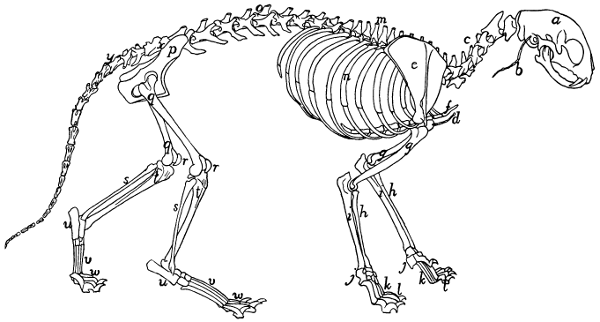
Fig. 1.—Skeleton of Cat.
a, skull; b, hyoid; c, cervical vertebræ; d, clavicle; e,
scapula; f, sternum; g, humerus; h, radius; i, ulna; j, carpus; k, metacarpus;
l, phalanges; m, thoracic vertebræ; n, ribs; o, lumbar vertebræ; p, innominate
bones; q, femur; r, patella; s, fibula; t, tibia; u, tarsus; v,
metatarsus; w, phalanges; x, sacrum; y, caudal vertebræ.
Thoracic Vertebræ. Vertebræ thoracales
(Fig. 4).—The
thoracic vertebræ are most typical, and the fourth one of these
may therefore be first described (Figs. 2 and 3). It forms an
oval ring which has numerous processes and surrounds an
opening which is the vertebral foramen (a). The ventral one-third[2]
[3]
of this ring is much thickened and forms the centrum or
body (corpus) (b) of the vertebra. The centrum is a semicylinder,
the plane face of which bounds the vertebral canal,
while the curved surface is concave longitudinally and is
directed ventrad. The dorsal plane surface of the centrum is
marked by a median longitudinal ridge on either side of which
is an opening (nutrient foramen) for a blood-vessel. The
ends are nearly plane, the caudal being slightly concave; they
are harder and smoother than the other surfaces. They may
be easily separated in a young specimen as thin plates of bone
known as epiphyses.
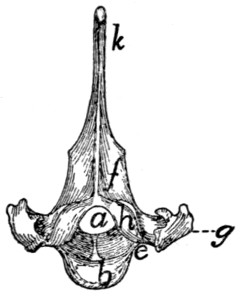
Fig. 2.—Fourth Thoracic Vertebra,
Cranial End.
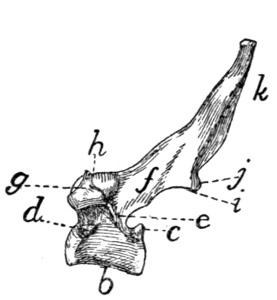
Fig. 3.—Fourth Thoracic Vertebra,
Side View.
a, vertebral foramen; b, centrum; c,
caudal, and d, cranial, costal demifacets;
e, radix or pedicle; f, lamina; g, transverse process; h, cranial articular facet; i,
caudal articular facet; j, caudal articular process; k, spinous process.
At the caudal end of the centrum, at its dorsolateral angle,
is a smooth area on each side continuous with the surface of the
epiphysis and bounded dorsolaterally by a sharp ridge of bone
(c). It is a costal demifacet. In corresponding positions at
the cranial end of the centrum are two demifacets not limited
by bony ridges (d). When the centra of two contiguous
thoracic vertebræ are placed together in the natural position the
cranial costal demifacets of one together with the caudal demifacets
of the other form two costal facets (Fig. 4, e), one on
each side, and each receives the head of a rib.
The dorsal two-thirds of the vertebral ring forms the vertebral
arch which is continued dorsally into the long, bluntly
pointed spinous process (Figs. 2 and 3, k) for attachment of
muscles.
[4]
The vertebral arch (each half of which is sometimes called
a neurapophysis) rises on each side from the cranial two-thirds
of the dorsolateral angle of the centrum, as a thickened portion,
the radix or pedicle (Figs. 2 and 3, e), which forms the
ventral half of the lateral boundary of the vertebral canal.
From the dorsal end of each radix a flat plate of bone, the
lamina (f), extends caudomediad to join its fellow of the
opposite side and form the vertebral arch. Owing to the fact
that the radix rises from only the cranial two-thirds of the
centrum there is left in the caudal border of the vertebral arch
a notch bounded by the radix, the lamina, and the centrum.
There is also a slight excavation of the cranial border of the
radix. When the vertebræ are articulated in the natural position,
these notches form the intervertebral foramina (Fig. 4,
d), for the exit of the spinal nerves.
At the junction of radix and lamina the arch is produced
craniolaterad into a short process, the transverse process (g),
knobbed at the end. On the ventral face of its free end the
transverse process bears a smooth facet, the transverse costal
facet or tubercular facet (Fig. 4, c), for articulation with the
tubercle of a rib.
On the dorsal face of each lamina at its cranial border is a
smooth oval area, the cranial articular facet (superior articular
facet of human anatomy) (Figs. 2 and 3, h). Its long axis is
oblique and it looks dorsolaterad. The slight projections of
the cranial edge of the laminæ on which the facets are situated
are the inconspicuous cranial articular processes (prezygapophyses).
On the ventral surface of each lamina at the caudal border,
near the middle line is a similar area, the caudal articular
facet (inferior articular facet of human anatomy) (i); these
occupy the ventral surfaces of two projections which form the
caudal (inferior) articular processes (postzygapophyses) (j).
These are separated by a median notch. When the vertebræ
are in their natural position the caudal articular facets lie dorsad
of the cranial facets and fit against them. They thus strengthen
the joint between contiguous vertebræ, while permitting slight
rotary motion.
[5]
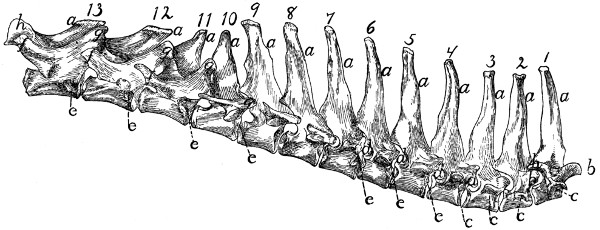
Fig. 4.—Thoracic Vertebræ,
Side View.
a, spinous processes; b, cranial articular processes;
c, transverse costal facets; d, intervertebral
foramina; e, costal facets; f, accessory processes; g, mammillary processes;
h, caudal articular processes.
Differential Characters of the Thoracic Vertebræ (Fig. 4).—Following
the thoracic vertebræ caudad there is to be seen
a gradual increase in the size of the centra brought about by
an increase in their craniocaudal and transverse measurements.
The dorsoventral measurements remain nearly the same.
The costal facets (Fig. 4, e) shift caudad so that on the
eleventh, twelfth, and thirteenth thoracic vertebræ each lies
entirely on the cranial end of its centrum, while the caudal end[6]
of the centrum immediately preceding is not marked by any
part of it. In the eleventh thoracic vertebra each costal facet
is usually still confluent with the smooth cranial end of the
centrum. In the twelfth vertebra the facets are separated by
smooth ridges from the cranial end of the vertebra, while in
the thirteenth vertebra they are separated by rough ridges.
The spinous processes (a) of the first four are of about the
same length. They then decrease in length to the twelfth,
while the twelfth and thirteenth are slightly longer than the
eleventh. The first ten slope more or less caudad, while the
spinous process of the tenth (anticlinal) vertebra is vertical
and those of the eleventh, twelfth, and thirteenth point craniad.
Each of the transverse processes of the seventh thoracic
vertebra shows a tendency to divide into three tubercles; one
of these is directed craniad, the mammillary process (or metapophysis),
one caudad, the accessory process (or anapophysis),
while the third (transverse process proper) looks ventrad and
bears the transverse costal facet. This division becomes
more prominent in the succeeding vertebræ, being most
marked in the ninth and tenth. In the eleventh, twelfth, and
thirteenth vertebræ the mammillary (g) and accessory (f)
processes are very pronounced, while the transverse costal
facet and that part of the transverse process which bears it have
disappeared. The ribs of the eleventh, twelfth, and thirteenth
vertebræ are thus attached to their respective centra by their
heads alone.
The cranial articular processes (b) are prominent on the
first two thoracic vertebræ; back of these they are very small
as far as the eleventh, so that the articular facets seem to be
borne merely upon the dorsal surface of the cranial edge of the
laminæ. In the eleventh, twelfth, and thirteenth the cranial
articular processes are large, bearing the articular facets on
their medial surfaces, while the mammillary processes appear
as tubercles on the lateral surfaces of the articular processes.
The caudal articular processes (h) are prominent in the first
thoracic, then smaller until the tenth is reached; in the tenth,
eleventh, twelfth, and thirteenth they are large and their facets
are borne laterally, so as to face the corresponding cranial[7]
facets. Thus from the tenth to the thirteenth thoracic vertebra
rotary motion is very limited, owing to the interlocking of the
articular processes.
The Lumbar Vertebræ. Vertebræ lumbales
(Fig. 5).—The
last thoracic vertebræ form the transition to the typical
lumbar vertebræ. These are larger than the thoracic vertebræ.
The centra are of the form of the centra of the thoracic vertebræ,
and increase in length to the sixth, but the seventh is about
the length of the first. They increase in breadth to the last.

Fig. 5.—Lumbar Vertebræ.
a, cranial articular processes; b,
mammillary processes; c, caudal articular processes; d, accessory
processes; e, transverse processes; f, spinous processes.
[8]
The cranial articular processes (Fig. 5, a) are prominent
and directed craniodorsad; they have the facets on their
medial surfaces, while their dorsolateral surfaces bear the
mammillary processes (b) as prominent tubercles. The caudal
articular processes (c) are likewise large; their facets look
laterad. When the vertebræ are articulated they are received
between the medially directed cranial processes.
The accessory processes (d) are well developed on the
first vertebra, diminish in size to the fifth or sixth, and are
absent on the seventh and sometimes on the sixth.
The transverse processes (more properly pseudo-transverse
processes) (e) arise from the lateral surface of the centra; are
flat and are directed ventrocraniolaterad. The first is small,
and they increase in length and breadth from the first to the
sixth, those of the last being slightly smaller than in the sixth.
The free ends of the last four are curved craniad.
The spinous processes (f) are flat and directed craniodorsad.
They increase in length to the fifth and then decrease.
The first five are knobbed at the end. In a dorsal view the
spinous process and cranial articular processes of each vertebra
are seen to interlock with the caudal articular processes and
accessory processes of the preceding vertebra in such a way as
to prevent rotary motion, and this arrangement may be traced
craniad as far as the eleventh thoracic vertebra.
Sacral Vertebræ. Vertebræ sacrales
(Figs. 6 and 7).—The
three sacral vertebræ are united in the adult into a single
bone, the os sacrum, or sacrum. In a kitten the three vertebræ
are separate, while in an animal almost mature the first
two are united and the third is still separate. The sacrum lies
between the last lumbar and the first caudal vertebræ and
articulates laterally with the two innominate bones. It is
pyramidal, with the base of the pyramid directed craniad, and
is perforated by a depressed longitudinal canal, the sacral
canal, which is a continuation of the vertebral canal, and by
four large foramina dorsally and four ventrally. It may be
described as having a cranial end or base and a caudal end or
apex, a dorsal, a ventral, and two lateral surfaces.
The base is slightly oblique and presents a smooth transversely[9]
oval articular facet (the cranial end of the centrum of
the first sacral vertebra), for articulation with the centrum of
the last lumbar vertebra. Dorsad of this is the sacral canal,
more depressed than the vertebral arch craniad of it. It supports
a spinous process (Fig. 6, a) which is directed dorsad.
At the junction of its lamina and radix is seen the prominent
cranial articular process (b) with sometimes slight indications
of a mammillary process on its lateral surface. Laterad of the
articular facet is seen the cranial face of the expanded “pseudo-transverse
process” (c) of the first sacral vertebra. The
ventral border of the base is concave ventrad, forming an arc
of about 120 degrees. The apex shows the caudal end of the
last sacral centrum. Dorsad of this are the vertebral arch with
a very short spinous process (a′), and the caudal articular
processes (d). Laterad of the centrum appears the laterally
directed thin transverse process (e).
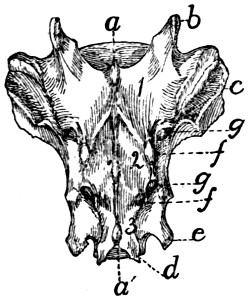
Fig. 6.—Sacrum, Dorsal Surface.
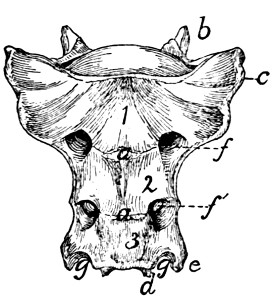
Fig. 7.—Sacrum, Ventral Surface.
Fig. 6.—1, 2, 3, the three sacral
vertebræ. a, a′, spinous processes; b, cranial
articular process of first sacral vertebra; c, expanded transverse process of first
sacral vertebra; d, caudal articular processes of third sacral vertebra; e, transverse
processes of third sacral vertebra; f, tubercles formed by fused articular processes
of the vertebræ; g, dorsal (or posterior) sacral foramina.
Fig. 7.—1, 2, 3, the three sacral
vertebræ. a, the transverse ridges formed by
the union of the centra; b, cranial articular processes of first vertebra; c, transverse
process of first vertebra; d, caudal articular processes of third vertebra; e, transverse
processes of third sacral vertebra; f, f′, ventral (or anterior) sacral foramina;
g, notch which helps to form third ventral sacral foramen.
The ventral or pelvic surface (Fig. 7) is smooth, concave
craniad, convex caudad, and crossed by two transverse ridges
(a) along which are seen the ossified remains of the intervertebral
fibro-cartilages. At the ends of the first ridge is a pair of
nearly circular ventral (or anterior) sacral foramina (f) for[10]
the passage of sacral nerves. At the end of the second ridge
is a pair of ventral sacral foramina (f′), smaller than the first
pair and continued laterocaudad into shallow grooves for the
ventral rami of the sacral nerves. That portion of the bone
lying laterad of a line joining the medial borders of these two
pairs of foramina is known as the lateral mass of the sacrum
and is composed of the fused transverse processes of the sacral
vertebræ. At the caudal margin of the ventral surface there
is a notch between the lateral mass and the centrum (g).
When the caudal vertebræ are articulated, this notch helps to
form a foramen for the third sacral nerve.
The dorsal surface (Fig. 6) is narrower at its cranial end
than is the ventral surface. Its cranial border bears laterally
a pair of cranial articular processes (b) with their medially
directed facets and between them it is concave, so that a large
dorsal opening is left into the vertebral canal between the last
lumbar vertebra and the sacrum. Caudad of the articular
processes are two pairs of tubercles (f). These are the fused
cranial and caudal articular processes of the sacral vertebræ.
Caudad of them are the caudal articular processes of the last
sacral vertebra (d). Craniolaterad of the middle and cranial
tubercles are dorsal (posterior) sacral foramina (g) for the
transmission of the dorsal rami of the sacral nerves. Three
spinous processes (a) appear between these rows of tubercles.
They decrease in height caudad. That part of the surface included
between the spinous process and the tubercles is made
up of the fused laminæ of the sacral vertebræ. That part
between the tubercles and a line joining the lateral margins of
the dorsal (posterior) sacral foramina is formed by the fused
radices of the sacral vertebræ.
The lateral surface may be divided into two parts. Craniad
is a large rough triangular area with equal sides and with one
of its angles directed ventrocraniad. It is the lateral face of
the pseudo-transverse process of the first sacral vertebra (Fig.
6, c). A smooth curved surface (the auricular facet) along its
ventral edge articulates with the ilium, while the dorsal portion
is rough for attachment of ligaments. Caudad is the
narrow longitudinal triangular area of the lateral faces of[11]
the fused transverse processes of the second and third sacral
vertebræ.
Caudal Vertebræ. Vertebræ caudales
(Fig. 1, y, and
Figs. 8 and 9).—The caudal vertebræ (21-23 in number)
decrease gradually in size to the last one. Caudad they
become longer and more slender
and lose the character of vertebræ.
They become finally reduced
to mere centra,—slender
rods of bone knobbed or enlarged
at their two ends (Fig. 8). The
last one is more pointed than the
others and bears at its caudal end
a small separate conical piece,
the rudiment of an additional
vertebra.

Fig. 8.
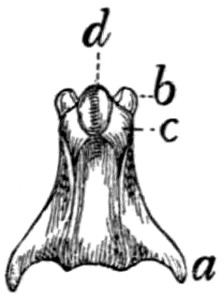
Fig. 9.
Fig. 8.—Caudal Vertebra, from near
the caudal end of the tail.
Fig. 9.—Fourth Caudal Vertebra,
ventral view. a, transverse processes;
b, cranial articular processes; c, hæmal
processes; d, chevron bone.
The parts of a typical vertebra—vertebral arch, transverse
processes, cranial and caudal articular processes—may be
recognized in the vertebræ as far back as the eighth or ninth.
The transverse processes (Fig. 9, a) are directed caudad and
decrease rapidly in length. They are very small on the ninth
vertebra, but may be recognized for a considerable distance
back of this. The spinous process disappears at about the
fourth caudal vertebra, and the vertebral canal becomes
gradually smaller caudad, until on the eighth or ninth vertebra
it becomes merely a groove open dorsad.
Caudad of the third vertebra for a considerable distance,
each centrum bears on each lateral face at its cranial end a
short anterior transverse process, and on its ventral face at its
cranial end a pair of rounded tubercles, hæmal processes (c),
which articulate with a small pyramidal chevron bone (d) so
as to enclose a canal. These structures disappear caudad.
Cervical Vertebræ. Vertebræ cervicales
(Fig. 10).—The
cervical vertebræ number seven. The first two of these are so
peculiar as to require a separate description, so that the last five
may be first considered.
Passing craniad from the fourth thoracic vertebra to the
third cervical there is a gradual transition. The centra of the[12]
cervical vertebræ are broader and thinner than those of the
thoracic vertebræ, while the vertebral arches and vertebral
canal are larger (Fig. 11). The caudal end of each centrum
is concave and looks dorsocaudad when the centrum is held
with its long axis horizontal. The cranial end of the centrum
is convex and looks ventrocraniad when the centrum is horizontal.
These peculiarities are more marked in the third
vertebra than in the seventh. The spinous processes grow
rapidly shorter as we pass craniad; the fifth, sixth, and seventh
are directed dorsocraniad, the third and fourth dorsad.
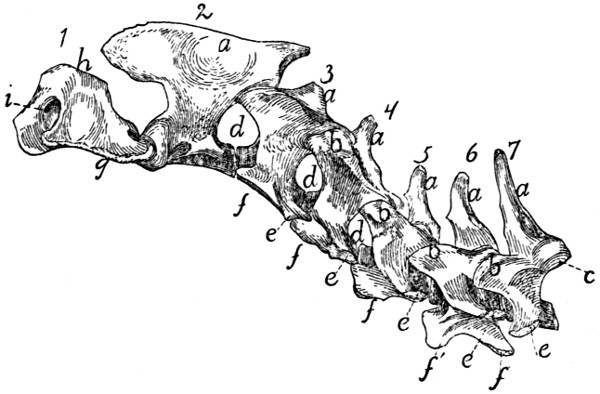
Fig.
10.—Cervical Vertebræ, Side View.
a, spinous processes; b, cranial articular processes; c, caudal articular facet; d,
intervertebral foramina; e, transverse process proper; f, processus costarius; g,
wing of the atlas; h, dorsal arch of the atlas; i, atlantal foramen.
The caudal articular processes are situated at the junction
of the radices and laminæ; their facets (Fig. 10, c) look
ventrocaudolaterad. The cranial articular processes also
become more prominent than is the rule in the thoracic vertebræ;
they are borne at the junction of radix and lamina and
have their facets (Fig. 11, b) directed dorsomediad. The
cranial and caudal articular processes of each side are joined
by a prominent ridge which is most pronounced in the third,
fourth, and fifth vertebræ.
The characteristic feature of the cervical vertebræ is their
transverse process, so called. In each of them it arises by
two roots, one from the centrum and one from the arch.[13]
These two roots, which are broad and thin, converge and unite
so as to enclose a canal or foramen, the foramen transversarium
(Fig. 11, g), for the vertebral artery. Laterad of the
foramen the two parts of the process are, in the third cervical,
almost completely united, the dorsal part being, however, distinguishable
as a tubercle at the caudolateral angle of the thin
plate formed by the process as a whole. This dorsal component
is the transverse process proper (Figs. 10 and 11, e),
while the ventral portion represents a rib, and is hence known
as the processus costarius (f). The expanded plate formed
by the union of these two processes is directed nearly ventrad
and somewhat craniad in the third, fourth, and fifth vertebræ.
The two components of the process gradually separate as we
pass caudad; in the fourth and fifth vertebræ the part which
represents the transverse process proper forms a very prominent
tubercle at the caudolateral angle of the plate formed by the
processus costarius. In the sixth (Fig. 11) the two parts are
almost completely separated; the dorsal part forms (e) a slender
knobbed process, while the processus costarius is divided into
two portions (f and f′) by a broad lateral notch. In the
seventh the ventral part (processus costarius) is usually quite
lacking, though sometimes represented by a slender spicule of
bone. In the former case the foramen transversarium is of
course likewise lacking.

Fig. 11.—Sixth Cervical Vertebra,
Cranial End.
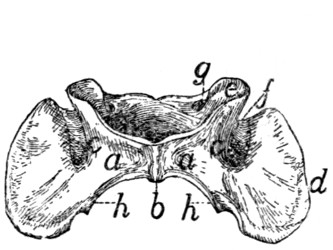
Fig. 12.—Atlas, Ventral View.
Fig. 11.—a, spinous process; b,
cranial articular facet; c, lamina; d, radix or
pedicle; e, transverse process proper; f, f′, processus costarius; g, foramen transversarium;
h, centrum; i, vertebral canal.
Fig. 12.—a, ventral arch; b, tuberculum
anterius; c, lateral masses; d, transverse
processes; e, cranial articular facets; f, groove connecting the foramen transversarium
with the atlantal foramen; g, atlantal foramen; h, caudal articular
facets.
[14]
The Atlas (Fig. 10, 1;
Fig. 12).—The first cervical vertebra
or atlas has somewhat the form of a seal ring. The
centrum is absent; it has united with the second vertebra to
form the odontoid process or dens. Its place is taken in the
atlas by a narrow flat arch of bone, narrower at the ends than
in the middle, the ventral arch (Fig. 12, a) of the atlas. This
connects the lateral, thicker portions of the ring ventrally and
bears on its caudal margin a blunt tubercle (tuberculum
anterius, Fig. 12, b). Laterally the ring is thickened, forming
thus the lateral masses (c) which are continued into the broad
thin transverse processes (Fig. 10, g; Fig. 12, d). Each
lateral mass bears at its cranial end on its medial surface a
concave, pear-shaped facet, cranial (or superior) articular facet,
(Fig. 12, e) for articulation with the condyles of the skull.
These facets look craniomediad. Dorsad of each is a foramen,
the atlantal foramen (Fig. 10, i; Fig. 12, g), which pierces
the dorsal arch at its junction with the lateral mass. Caudal
to the facet, on the medial face of each lateral mass, within the
vertebral canal, is a tubercle. To the two tubercles are
attached the transverse ligament (Fig. 14, b) which holds in
place the odontoid process (dens) of the axis.
That part of the lateral mass which bears the articular facet
projects craniad of the dorsal arch and is separated by a deep
triangular notch from the transverse process. Along the
bottom of this notch runs a groove (Fig. 12, f), convex
craniad, which connects the cranial end of the foramen transversarium
and the atlantal foramen. The vertebral artery
passes along it. The foramen transversarium is circular. It
is bounded laterally by the lateral masses, and dorsally by the
dorsal arch.
The dorsal arch (Fig. 10, h) is two to three times as broad
as the ventral, has a thick convex cranial border with a median
notch, and a thin concave caudal border.
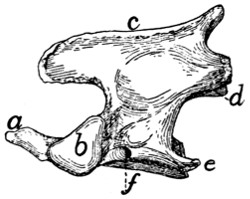
Fig. 13.—Axis or Epistropheus,
Side View.
a, odontoid process or
dens; b, cranial articular
facets; c, spinous process;
d, caudal articular facet;
e, transverse process; f,
foramen transversarium.
The caudal articular facets (Fig. 12, h) are borne by the
caudal ends of the lateral masses. They are slightly concave,
triangular, and look caudomediad, so that their dorsal borders
form with the caudal border of the dorsal arch nearly a semicircle.
The transverse processes are flat and directed laterad.[15]
The attached margin of each is about two-thirds the length of
the thinner free margin. The somewhat thicker caudal end
of the transverse process projects further caudad than any other
part of the vertebra and is separated by a slight notch from the
caudal articular facet. From the bottom of this notch the
foramen transversarium extends craniad and opens at the
middle of the ventral face of the transverse process.
Epistropheus or Axis (Fig. 10, 2;
Fig. 13).—The second
cervical vertebra (epistropheus or axis) is not so wide as the
atlas but is much longer. Craniad the centrum is continued
into a slender conical, toothlike projection,
the dens or odontoid process (Fig.
13, a) which represents the centrum of the
atlas. The dens is smooth below for
articulation with the ventral arch of the
atlas. It is rougher above. Laterad of
the dens the centrum bears a pair of large
cranial articular facets (b) which look
craniolaterad. These have each the form
of a right-angled triangle with rounded
angles, one side of the triangle being
nearly horizontal. Each is separated from
the articular face of the dens by a roughened groove. The
spinous process (c) runs the length of the vertebral arch. It
extends craniad of the vertebral arch nearly as far as the dens,
as a flat rounded projection. Caudad of the vertebral arch it
projects for a short distance as a stout triangular spine. The
caudal articular facets (d) are borne on thickenings of the
caudolateral portions of the arch; they face almost directly
ventrad. The transverse process (e) is slender and triangular
and directed nearly caudad. Its apex reaches no farther than
the caudal or articular face of the centrum. Its base is traversed
by the foramen transversarium (f).
Differential Characters of the Cervical Vertebræ.—It is
possible to identify each of the cervical vertebræ:
The first by the absence of the centrum.
The second by the dens or odontoid process.
The third by the small spinous process and slightly marked[16]
tubercle of the transverse process, and by a median tubercle on
the cranial border of the vertebral arch.
The fourth by the spinous process directed dorsad, and the
short thick tubercle of the transverse process not trifid.
The fifth by the spinous process directed craniad, and the
more slender spine-like tubercle of the transverse process not
trifid.
The sixth by the trifid transverse process.
The seventh by the long spinous process and the slender
simple transverse process, and by the usual absence of the
foramen transversarium.
LIGAMENTS OF THE VERTEBRAL COLUMN.
Fibro-cartilagines intervertebrales.—The separate vertebræ
(except the atlas and axis) are united by the disk-shaped
intervertebral fibro-cartilages, which are situated between the
centra of the vertebræ. Each consists of a central pulpy portion
and a fibrous outer portion, covered by strong intercrossing
tendinous fibers which unite with the periosteum of the
vertebræ.
Ligamentum longitudinale anterius.—On the ventral
face of the centra of the vertebræ, from the atlas to the
sacrum, lies a longitudinal ligament, the anterior longitudinal
ligament. It is very small, almost rudimentary, in the cervical
region: large and strong in the thoracic and lumbar regions.
Ligamentum longitudinale posterius (Fig. 14, a).—A
corresponding ligament (posterior longitudinal ligament) lies
on the dorsal surface of the centra (therefore within the vertebral
canal). It is enlarged between each pair of vertebræ and
closely united to the intervertebral fibro-cartilages.
Ligamentum supraspinale.—Between the tips of the
spinous processes of the thoracic and lumbar vertebræ extend
ligamentous fibers. They are not united to form a distinct
band, and can hardly be distinguished from the numerous
tendinous fibers of the supraspinous muscles. Together they
represent the supraspinous ligament. From the tip of the
spinous process of the first thoracic vertebra to the caudal end
of the spine of the axis extends a slender strand representing[17]
the ligamentum nuchæ or cervical supraspinous ligament.
It is imbedded in the superficial muscles of this region, some
of which take origin from it.
Ligamentous fibers are also present between the spinous
processes of the vertebræ (ligamenta interspinalia): between
the transverse processes (ligamenta intertransversaria), and
between the vertebral arches (ligamenta flava).
Capsulæ articulares.—The joints between the articular
processes are furnished with articular capsules attached about
the edges of the articular surfaces. These are larger and looser
in the cervical region.
Atlanto-occipital Articulation.—The joint between the
atlas and the occipital condyles has a single articular capsule,
which is attached about the borders of the articular surfaces of
the two bones. This capsule is of course widest laterally,
forming indeed two partially separated sacs, which are, however,
continuous by a narrow portion across the ventral middle
line. This capsule communicates with that which covers the
articular surface of the dens, and through this with the capsule
between the atlas and axis. That portion of the capsule which
covers the space between the ventral arch of the atlas and the
occipital bone represents the anterior atlanto-occipital membrane;
it is strengthened by a slender median ligamentous
strand. The posterior atlanto-occipital membrane covers in
the same way the space between the dorsal arch of the atlas and
the dorsal edge of the foramen magnum. In it a number of
different sets of fibers, with regard to direction and to degree
of development, may be distinguished; these have sometimes
been considered separate ligaments.
The lateral ligaments of the atlas begin at the lateral
angle of the cranial margin of the atlas, at about the junction
of its dorsal and ventral arches, and pass cranioventrad to the
jugular processes.
Articulation between the Axis and Atlas.—The articular
capsule is large and loose, being attached to dorsal and ventral
borders of the atlas, about the articular surfaces of the axis,
and to the cranial projection of the spine of the atlas. It also
passes craniad along the ventral side of the dens and communicates[18]
here with the capsule of the atlanto-occipital articulation.
In the dorsal part of the capsule a short
strong ligamentous strand is developed,
connecting the caudal border of the dorsal
arch of the atlas with the tip of the
cranial projection of the spinous process
of the axis.
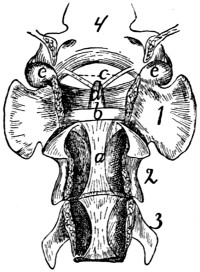
Fig. 14.—Ligaments of
the Odontoid Process
or Dens.
First three cervical vertebræ
and base of the skull,
with dorsal surface removed.
a, ligamentum
longitudinale posterius; b,
transverse ligament of the
atlas; c, ligamenta alaria;
d, odontoid process; e, occipital
condyles; 1, 2, 3,
the first three cervical vertebræ;
4, basal portion of
the occipital bone.
The dens or odontoid process is
held in place by the transverse ligament
(Fig. 14, b) of the atlas, which
passes across the process as it lies within
the vertebral canal of the atlas. The
transverse ligament is attached at its
two ends to the medial surface of the
sides of the atlas at about the region
where the dorsal and ventral arches of
the atlas unite.
From the cranial end of the odontoid
process the two ligamenta alaria (Fig.
14, c) diverge craniolaterad to the rough
ventromedial angle of the condyles of
the occipital bone.
II. RIBS. COSTÆ (Figs. 1 and 15.)
The cat has thirteen pairs of ribs. One of the fifth pair
(Fig. 15) may be taken as typical. It is a curved flattened rod
of bone attached at its dorsal end to the vertebral column,
and at its ventral end to a cartilage (costal cartilage, Fig.
15, f) which serves to unite it to the sternum.
The most convex portion of the bone is known as the
angle (e). Each rib presents a convex lateral and a concave
medial surface, a cranial and a caudal border. The
borders are broad dorsad and narrow ventrad, while the surfaces
are narrow dorsad and broad ventrad. The rib has thus
the appearance of having been twisted.
The rib ends dorsad in a globular head or capitulum (a),
by which it articulates with the costal demifacets of two contiguous
thoracic vertebræ. Between the capitulum and angle[19]
on the lateral surface is an elevated area, the tubercle, marked
by the smooth tubercular facet (c) for articulation with the
transverse process of a vertebra.
The constricted portion between the
head and tubercle is known as the
neck (collum) (d). The angle is
marked by a projecting process (e)
(angular process) on its lateral border,
for attachment of a ligament.
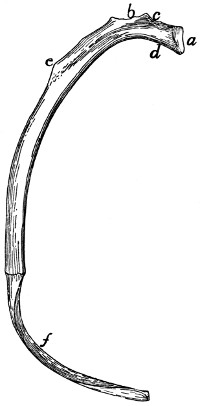
Fig. 15.—Fifth Rib of Left
Side, Cranial View.
a, head; b, tubercle; c, tubercular
facet; d, neck; e, angle,
with angular process; f, cartilage.
The ribs increase in length to
the ninth (the ninth and tenth are
of the same length) and then decrease
to the last. They decrease
in breadth behind the fifth. The
first is nearly in a dorsoventral
plane, while the others have their
dorsal ends inclined slightly craniad.
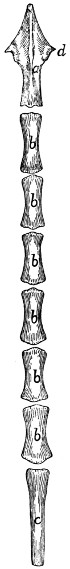
Fig. 16.—Sternum,
Ventral
View.
a, manubrium;
b, the separate
pieces forming
the body; c, bony
part of the xiphoid
process
(the expanded
cartilaginous portion
not being
shown); d, facet
for attachment of
first rib.
The tubercles become less prominent
as we pass caudad and are absent
on the last two or three ribs, which
do not articulate with the transverse
process.
The first nine ribs (true ribs or
costæ veræ) are attached separately
to the sternum by their costal cartilages.
The last four (false ribs or costæ spuriæ) are not
attached separately to the sternum. The costal cartilages of
the tenth, eleventh, and twelfth are united to one another at
their sternal ends. They may be united also to the ninth
costal cartilage or to the sternum by a common cartilage of
insertion, or they may be quite free from the sternum. The
thirteenth costal cartilages are free (floating ribs).
Ligaments of the Ribs.—The articular surfaces between
the head of the rib and the centra, and between the tubercle
and the transverse process of the vertebra, have each an
articular capsule. There are also a number of small ligamentous
bands from the tuberosity and the neck of the rib to
the transverse process of the vertebra.
[20]
III. STERNUM. (Fig. 16.)
The sternum consists of three portions, a
cranial piece or manubrium (a), a caudal piece
or xiphoid process (c), and a middle portion or
body (corpus), which is divided into a number
of segments (b).
To the sternum are united the ventral ends
of the first nine ribs. It thus forms the median
ventral boundary of the thorax. Since the
thorax decreases in dorsoventral measurement
craniad, the long axis of the sternum is inclined
from its caudal end dorsocraniad, and if continued
would strike the vertebral column in the
region of the first cervical vertebra.
The manubrium (a) makes up about one-fifth
the whole length of the sternum and projects
craniad of the first rib. It has the form of a
dagger and presents a dorsal surface and two
lateral surfaces, the latter uniting ventrad to
form a sharp angle. In the middle of the lateral
surface near the dorsal margin is an oval articular
surface (d) borne on a triangular projection.
It looks caudodorsad and is for the first costal
cartilage.
The caudal end articulates with the body by
a synchondrosis and presents a slightly marked
oval facet on each side for the second costal
cartilage.
The body consists of six cylindrical pieces (b)
enlarged at their ends and movably united by
synchondroses. They increase in breadth from
the first, and decrease slightly in length and
thickness. At the caudal end of each near its
ventral border there is a pair of facets looking
caudolaterad. They are for the costal cartilages.
The xiphoid process (c) is a broad thin plate
of cartilage at its caudal end; bony and cylindrical[21]
at its cranial end. It is attached by its base to the
last segment of the body by a considerable cartilaginous interval,
while the opposite end is free and directed caudoventrad.
The cartilage of the ninth rib is attached to the lateral
face of the cartilage between the xiphoid and the body, and
just caudad of this the common cartilage of insertion of the
tenth, eleventh, and twelfth costal cartilages is attached, if
present.
IV. THE SKULL.
The bones of the head consist of the skull proper together
with a number of separate bones forming part of the visceral
skeleton; these are the lower jaw, the hyoid, and the ear-bones.
The skull proper is considered as divided into cranial and
facial portions. The former includes all the bones which take
part in bounding the cranial cavity or cavity of the brain; the
latter includes the bones which support the face.
The cranial portion of the skull includes all that part
enclosing the large cavity which contains the brain. For convenience
this portion may be considered as made up of three
segments, each of which forms a ring surrounding a part of the
cranial cavity. The first or caudal segment or ring consists of
the occipital bone (with the interparietal) surrounding the
foramen magnum. The second segment consists of the
sphenoid ventrad, the parietals laterad and dorsad. Between
the first and second segments are intercalated laterally the
temporal bones containing the auditory organ. The third
segment or ring consists of the presphenoid ventrad, of the
frontals laterad and dorsad. The cranial opening of this ring
is closed by the lamina cribrosa of the ethmoid.
The cranial portion of the skull therefore contains eleven
separate bones: one occipital, one interparietal, two temporals,
one sphenoid, two parietals, one presphenoid, two frontals,
and one ethmoid.
The facial portion of the skull is much smaller than the[22]
cranial, and lies craniad of the latter; it encloses the nasal
cavity. It contains the following thirteen bones: two palatines,
one vomer, two maxillaries, two lachrymals, two premaxillaries,
two nasals, two malar or zygomatic bones. The
two halves of the mandible or lower jaw are frequently included
in the facial portion of the skull, making in all fifteen separate
bones in this part of the skull.
In the following the bones of the skull are first described
separately, then an account is given of the skull as a whole.
Occipital Bone. Os occipitale
(Figs. 17 and 18).—The
occipital bone forms the most caudal portion of the cranium,
entering into the formation of its caudal wall and of its base. It
connects the cranium with the vertebral column and surrounds
a large opening, the foramen magnum (d), by means of which
the cranial cavity communicates with the vertebral canal.
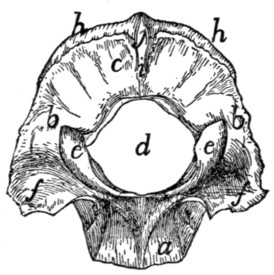
Fig. 17.—Occipital Bone, Caudal
or Outer Surface.
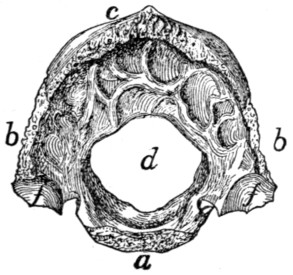
Fig. 18.—Occipital Bone, Inner
Surface.
a, basilar portion; b, lateral
portions; c, squamous portion; d, foramen magnum;
e, occipital condyles; f, jugular processes; g, jugular notch; h, lambdoidal
ridge; i, external occipital crest; j, external occipital protuberance.
In young kittens four portions may be distinguished in this
bone. These are, a basal portion (the basioccipital bone),
two lateral portions (the exoccipital bones), and a dorsal portion
(the supraoccipital bone). These four bones remain
separate through life in many lower vertebrates. In the adult
cat they are completely united into a single bone, but it is
convenient to describe this bone as made up of four parts: a
basal portion (a) (pars basilare), corresponding to the basioccipital,
two lateral portions (b) (partes laterales), corresponding
to the exoccipitals, and a squamous portion (c)
(squama occipitalis) corresponding to the supraoccipital.
[23]
The basilar portion (a) as seen from the dorsal or ventral
surface is oblong and flattened. It is broadest at the junction
of its middle and last thirds, and tapers toward both ends. It
presents a cranial end and a caudal end, a dorsal, a ventral,
and two lateral surfaces. Its caudal end, which helps to form
the ventral boundary of the foramen magnum, is concave from
side to side. The cranial end is transversely elongate, about
five times as broad as high, pointed laterally and roughened
for attachment to the caudal end of the body of the sphenoid,
which has a corresponding form. The joint is a synchondrosis.
The dorsal surface is concave from side to side, forming a
longitudinal groove in which rest the pons and medulla.
The concavity is more pronounced caudad, where the bone is
thinner at its middle.
The ventral surface is marked by three parallel longitudinal
ridges. One of them is median and expands caudad into a
smooth triangular elevated area which extends to the foramen
magnum. The other two ridges run near the lateral edges of
the bone. Between them and the median ridge the surface is
smooth and is depressed caudad. Laterad of each lateral ridge
is a rough triangular surface overlaid in the natural state by the
medial edge of the tympanic bulla.
The lateral surfaces are smooth and sharp and abut against
the petrous portion of the temporal bone. They pass into the
lateral portions of the occipital caudad.
The lateral portions (b) of the occipital arise from the
caudal margin of the basilar portion in the transverse plane; a
short distance laterad of the line of junction they turn dorsad
at an angle of nearly ninety degrees. They form the lateral
boundaries of the foramen magnum and pass dorsally into the
squamous portion.
The external surface of each presents an elongated elevated
spirally curved surface, the occipital condyle (c), for articulation
with the atlas. A small part of each condyle is formed
from the basal portion. The two condyles are separated from
one another ventrally by a narrow notch, and each extends
(laterad) along the border of the foramen magnum to a point
slightly dorsad of the transverse diameter of the latter.
[24]
Laterad of each condyle the bone is elevated into a blunt
triangular projection, the jugular process (f), which covers the
caudal end of the tympanic bulla. Between the jugular process
and the condyle is a deep depression.
The internal surface (Fig. 18) is concave dorsoventrally,
following the outline of the foramen magnum. It is convex
from side to side. It is smooth except at its outer margin,
which is rough for articulation with the mastoid portion of the
temporal bone. The cranial face of the jugular process (f)
shows a rough concavity for the reception of the bulla tympani.
Mediad of the jugular process is a notch (jugular notch) (g)
which when the bones are articulated forms part of the boundary
of the jugular foramen. Mediad of this notch is a foramen
which forms one end of the hypoglossal canal. It passes dorsocaudad
into the cranial cavity and transmits the hypoglossal
nerve. Dorsad of the hypoglossal canal is the cranial opening
of the condyloid canal, which passes caudad and opens just
craniad of the dorsal end of the condyle. It transmits a vein.
The outer border of this portion is rough for articulation with
the petrous and mastoid portions of the temporal.
The squamous portion (c) has the form of a sector of a
circle whose arc is a little more than ninety degrees. The
central angle of the sector is truncated and bounds the foramen
magnum dorsally. The arc of the sector forms the dorsal
margin of the bone, while along the radii it passes into the
lateral portions. Its dorsal portion is thick and porous; its
ventral portion near the foramen magnum is thin and compact.
The external surface (Fig. 17) is marked by a prominent
ridge, the lambdoidal ridge (h), parallel with the dorsal border
and near to it. The narrow portion of the outer surface that
lies dorsad of the ridge forms an angle of about ninety degrees
with the remainder of the surface.
A median crest (i) extends ventrad from the middle of the
lambdoidal ridge toward the foramen magnum; this is the
external occipital crest. At its junction with the lambdoidal
ridge it is elevated into a tubercle, the external occipital
protuberance (j).
[25]
The inner surface (Fig. 18) presents depressions for the
convolutions of the cerebellum.
The dorsal border is thick and rough for articulation with
the parietals and interparietal. The ventral border abuts on
the foramen magnum and is thin and smooth.
Interparietal Bone. Os interparietale
(Fig. 19).—This
is a small triangular bone lying between the parietals, with its
apex directed craniad, and its base in contact with the squamous
portion of the occipital.
Its dorsal surface is arrow-shaped and has its posterior
border notched. It is marked by a median crest (part of the
sagittal crest) which is continued craniad from the middle of
the lambdoidal crest.
The ventral surface is irregularly triangular, smooth, and
concave. The three borders are rough for articulation with
the parietals and occipital.

Fig. 19.—Interparietal
Bone, Outer Surface.
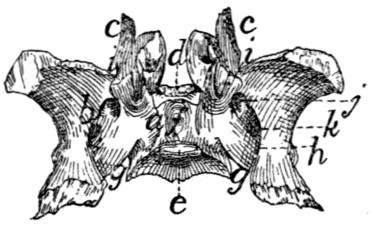
Fig. 20.—Sphenoid Bone, Dorsal or
Internal Surface.
Fig. 19, showing the sagittal crest running craniocaudad across its middle.
Fig. 20.—a, body; b, wings;
c, pterygoid process; d, tuberculum sellæ; e, dorsum
sellæ; f, sella turcica; g, notch which aids in forming the foramen lacerum;
h, longitudinal groove of alisphenoid; i, notch which aids in forming the orbital
fissure; j, foramen rotundum; k, foramen ovale.
Sphenoid Bone. Os sphenoidale
(Fig. 20).—The sphenoid
bone of man is represented in the cat by two entirely
distinct bones,—one cranial, the other caudal. The cranial
portion may be designated as the presphenoid (Fig. 21); the
caudal part will be described as the sphenoid (proper) (Fig.
20).
The sphenoid bone in the kitten is in three parts: a central
portion, the basisphenoid, and two lateral portions, the alisphenoids.
In many lower vertebrates these three bones are
permanently distinct, but in the adult cat they are united to
form the sphenoid bone. To these there is added a fourth
element, separate in many vertebrates as the pterygoid bone.[26]
The sphenoid may thus be described as composed of a central
portion, the body (a) (basisphenoid), and of two thin expanded
wings (b) (alisphenoids, alæ magnæ of the human sphenoid);
each of which has arising from it a thin curved process, the
pterygoid process (c), directed craniad and largely made up of
the pterygoid bone.
The body of the sphenoid (a) lies in the middle line of the
base of the skull. It is wedge-shaped, with the converging
sides of the wedge directed laterad and its apex pointed
craniad.
It has six surfaces, of which the dorsal and a part of the
laterals look into the cranial cavity. The cranial end articulates
with the body of the presphenoid, and the caudal with the
body of the occipital.
The dorsal surface is triangular, with one apex of the triangle
truncated, elevated, and directed craniad. This elevation
is the tuberculum sellæ (d). Just caudad of the middle
the surface presents a rectangular elevation with rounded
angles, the dorsum sellæ (e). The cranial end of the dorsum
sellæ presents at each dorsolateral angle a very small smooth
tubercle which represents one of the posterior clinoid processes
of man. Between this elevation and the elevated cranial end
of this surface there is a deep excavation, the sella turcica (f),
in which in the natural state is lodged the hypophysis. Near
the cranial end of the sella is a small foramen, probably
nutrient. At the caudal end of the body a slight notch (g)
separates it from the wing: this notch forms a part of the
foramen lacerum. Against this notch fits the apex of the
petrous bone, and from it a groove (carotid groove) is continued
mediocraniad to the sella turcica.
The ventral surface (Fig. 41, 3) is triangular, smooth, and
nearly flat; it is marked by a median ridge which is the continuation
craniad of the ridge on the ventral face of the basilar
portion of the occipital.
Its caudal angles are separated from the rest of the bone
by sharp triangular elevations, laterad of which are rough
triangular areas, overlaid when the bones are articulated by a
triangular spine from the tympanic bulla.
[27]
Its lateral surfaces are mostly covered by the wings. They
appear at the sides of the elevated cranial end of the dorsal
surface as triangular areas.
The caudal end is concave, rough, and has the form of the
cranial end of the basilar part of the occipital.
The cranial end is nearly square and rough for articulation
with the body of the presphenoid.
The Wing (alisphenoid; ala magna of the human sphenoid)
(Fig. 20, b).—This is a thin quadrilateral plate of bone attached
by its medial border to nearly the whole of the lateral surface
of the body. Its middle portion lies nearly in the same plane
as the body, but its ends are curved dorsad so that its internal
surface is concave and its external surface is convex. The
curvature is most pronounced near the long lateral border, so
that this border forms nearly a semicircle.
The internal surface supports the occipital lobe of the cerebrum.
It is marked by a rounded groove (h) which is parallel
with the lateral surface of the body. The dorsal margin of the
groove projects mediad in the form of a sharp ridge which is
broadest caudad, where it often reaches nearly to the posterior
clinoid process. The groove passes craniad into three foramina.
The first (cranial) of these, the orbital fissure (i), is
large and lies between the wing, the body, and the pterygoid
process. It is incomplete, but is completed by the presphenoid.
The second foramen is small and rounded; it is the foramen
rotundum (j). The third, foramen ovale (k), is larger and
oval and penetrates the wing through about the middle of its
longitudinal axis. Another minute foramen penetrates the
sphenoid between the wing and the body of the bone, just
laterad of the tuberculum sellæ. This foramen is continuous
craniad with a groove on the dorsal surface of the pterygoid
process; the groove and foramen constitute the pterygoid
canal. It transmits a nerve.
The external surface shows the orbital fissure, the foramen
rotundum and the foramen ovale, bounded ventrally by a sharp
ridge, which is continued onto the pterygoid process. Between
this ridge and the body the surface is longitudinally grooved
for the tuba auditiva or Eustachian tube.
[28]
The semicircular margin of the bone articulates with the
squamous portion of the temporal. At the junction of its
caudal and middle third there is sometimes a toothlike projection
which underlies the root of the zygoma.
The whole of the cranial margin, except the lateral end,
articulates with the wing of the presphenoid. At this end the
angle formed by the junction of lateral and cranial borders is
produced into a flat process, which passes dorsocaudad
between the squamous portion of the temporal and the frontal,
and articulates by the roughened internal surface of its free end
with a similar process from the parietal.
The caudal margin laterad of the groove is bevelled and
roughened at the expense of the dorsal surface and is overlaid
by the ventral end of the tentorium. Mediad of the groove it
projects caudad as a slender point, the lingula of the sphenoid.
This is received into a narrow cleft between the apex of the
petrous bone and the bulla tympani.
The pterygoid process (c) is a nearly square, thin plate of
bone. The medial surface is smooth and concave, the lateral
face is convex and marked by two parallel ridges. The medial
one of these is continued craniad from the bony septum which
separates the orbital fissure from the foramen rotundum, and
the lateral one from the septum which separates the foramen
rotundum from the foramen ovale. A sharp triangular spine
projects laterad from near the caudal end of the lateral ridge.
The two ridges and that part of the lateral surface of the
bone included between them form a part of the sphenoid bone
known as the pterygoid process of the sphenoid bone, in those
cases where the pterygoid is a separate bone.
The remainder of the process is equivalent to the pterygoid
bone of other vertebrates.
Between the caudal margin of this bone and the lateral of
the two ridges, i.e., between the pterygoid bone and the
pterygoid process of the sphenoid, is a long deep fossa, the
internal pterygoid fossa (Fig. 40, s). The laterocaudal
margin of the pterygoid process projects caudad, as a curved
triangular spine, the hamulus or hamular process (Fig. 40, t;
Fig. 43, i).
[29]
The Presphenoid Bone. Os presphenoidale
(Fig. 21).—In
a young cat this bone is in three pieces, a basal portion
(presphenoid) and two wings (orbitosphenoid bones). These
bones remain distinct throughout life in many
lower vertebrates, but in the adult cat they fuse
to form a single bone. We may nevertheless
conveniently describe this bone as made up of
a body (a) (the basisphenoid), and two wings
(b), the orbitosphenoids (the alæ parvæ of the
human sphenoid).
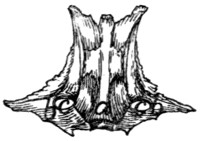
Fig. 21.—Presphenoid
Bone,
Ventral View.
a, body; b, wings;
c, optic foramina.
The body (a) lies in the base of the skull in
the median line, craniad of the basisphenoid. It has the form
of a rectangular prism about twice as long as broad. It is
hollow, and the cavity is divided by a median longitudinal partition
into two cavities (sphenoidal sinuses, Fig. 43, l). The
sphenoidal sinuses are continued craniad into the cavities of
the ethmoid. The body has six surfaces:
The dorsal or internal surface (Fig. 42, n) looks into the
cranial cavity and is continuous with the dorsal surface of the
wings. The caudal end of the body is depressed, and when
united to the basisphenoid aids in forming the cranial wall of
the sella turcica. At each caudolateral angle is a short spine,
the anterior clinoid process. At about one-third the length
of the bone from the caudal end is a transverse groove (chiasmatic
groove, Fig. 42, m) for the optic chiasma. Its ends
lead into two round foramina (the optic foramina, Fig. 42, l;
Fig. 21, c) which pass craniolaterad between the body and
the wings of the presphenoid and transmit the optic nerve and
the ophthalmic artery.
The ventral surface (Fig. 21) is hour-glass-shaped and
marked by a smooth median ridge, continuous with the ridge
on the basisphenoid and overlaid at its cranial end by the
vomer. The caudal end presents a rough triangular area on
each side, for articulation with the pterygoid process of the
sphenoid bone, while the cranial end has similar areas overlaid
by the nasal portion of the palatine bones.
The lateral surface looks towards the orbitotemporal fossa.
It is notched near the caudal end by the ventral border of the[30]
optic foramen (c). Caudad of this foramen the surface presents
an oblique groove which forms in the natural condition the
medial boundary of the orbital fissure.
Craniad of the optic foramen the surface is smooth and
marked near its ventral border by a longitudinal ridge which
forms part of the dorsal boundary of a fossa, the external pterygoid
fossa (Fig. 40, p).
The caudal end presents ventrally a quadrangular rough
surface for articulation with the body of the sphenoid. The
cranial end presents the two sphenoidal sinuses separated by
a median partition.
The median partition articulates by its free border with the
lamina perpendicularis of the ethmoid. At its ventral end is
the abruptly truncate end of the median ridge of the ventral
surface, which is continuous with the ventral cartilaginous portion
of the lamina perpendicularis. The lateral walls of the
sphenoidal sinuses are continued craniad of the dorsal and ventral
walls and of the median partition, and articulate ventrally
with the nasal portion of the palatine bones, and dorsally with
the orbital plate of the frontal. Between them is received the
caudal ends of the labyrinths of the ethmoid in the middle,
while between their dorsal edges is received the caudal end of
the cribriform plate, and between their ventral edges the
expanded end of the vomer.
The wings (b) arise each from nearly the whole of the
dorsolateral angle of the body. They form prominent nearly
horizontal triangular projections over the optic foramina.
The dorsal and ventral surfaces are smooth and continuous
respectively with the dorsal and lateral surfaces of the body.
The dorsal surface looks into the cranial cavity, while the ventral
looks into the orbitotemporal fossa.
Craniad of the apex of the wing its border articulates with
the ventral edge of the orbital portion of the frontal bone.
Caudad of the apex the border articulates with the cranial
border of the wing of the sphenoid.
Temporal Bone. Os temporale
(Figs. 22 and 23).—This
forms a part of the lateral wall of the cranium, filling the gap
between the occipital and the sphenoidal segments. It is made[31]
up of three portions which are distinct in kittens but somewhat
firmly united in adult cats. In lower vertebrates these portions
are distinct bones called the Squamous, the Petrous, and the
Tympanic bones. In the cat they may be described as the
squamous (a), petrous (b), and tympanic (c) portions of the
temporal bone.
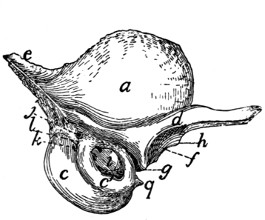
Fig. 22.—Temporal Bone, External
Surface.
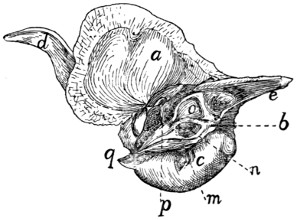
Fig. 23.—Temporal Bone, Internal
Surface.
a, squamous portion; b, petrous portion;
c, c′, tympanic portion (c, entotympanic;
c′, ectotympanic); d, zygomatic process; e, mastoid portion of the petrous;
f, mandibular fossa; g, postmandibular process; h, tuberculum articulare; i, external
auditory meatus; j, stylomastoid foramen; k, pit for tympanohyal bone;
l, mastoid process; m, grooves bounding the jugular foramen; n, internal auditory
meatus; o, appendicular fossa; p, hiatus facialis; q, styliform process of tympanic
bulla.
The squamous portion (a) (squama temporalis) is thin and
oval or has the form of an equilateral triangle with rounded
angles, with a curved process, the zygomatic process (d),
arising from its ventral border. Its outer surface (Fig. 22) is
convex and smooth and gives origin to part of the temporal
muscle. Its inner surface (Fig. 23) is concave and smooth
except near the margins, where it is bevelled and rough. The
roughened border is broader dorsad and caudad. The ventral
margin of the bone is turned mediad at its cranial end so that
the lateral face of the inverted portion looks ventrad. By this
portion of its lateral face the squamous rests upon the tympanic
bulla, and its edge articulates with the tentorium and the wing
of the sphenoid. The remaining (caudal) portion of the ventral
border overlies the mastoid portion (e) of the petrous. By the
remainder of its border the bone articulates with the parietal
dorsad and with the wing of the sphenoid craniad. The[32]
roughened portion of its inner surface overlies the margins of
both these bones.
The zygomatic process (d) is formed by the confluence of
two roots. One of these starts from the ventral end of the
lambdoidal ridge and passes along the ventral margin of the
squama dorsad of the external auditory meatus. The other
arises abruptly from the cranioventral angle of the bone. The
process thus formed is at first broad and passes horizontally
laterad and slightly craniad. It soon grows more slender and
turns gradually craniad, while at the same time it twists so that
the surface which is dorsal at the base becomes medial at the
tip; the posterior root which is continuous with the caudal
border at the base is continuous with the dorsal border at the
apex. On the ventral surface of the base is a transversely
elongated concave articular surface, the mandibular fossa (f),
for the condyloid process of the lower jaw. Caudad of this is
a sharp transverse ridge, the postmandibular or postglenoid
process (g), and craniad of the lateral end of the fossa a slight
tubercle, the tuberculum articulare (h). Near its apex the
zygomatic process is more slender and its ventral border is
bevelled for articulation with the malar or zygomatic bone.
The tympanic (Figs. 22 and 23, c; Fig. 24) is expanded
into a large hollow olive-shaped bone which is known as the
auditory bulla and encloses the tympanic cavity. Its substance
is very compact. Unlike the tympanic of most other
mammals it is developed from two bones, known as the ectotympanic
(Fig. 22, c′) and entotympanic (c). These are
strongly marked in young kittens, and can usually be easily
distinguished in adult cats. The entotympanic (Figs. 22 and
23, c) forms the larger part of the bulla, constituting its ventral
and medial surfaces; it is thin, smooth, and transparent. The
ectotympanic (Fig. 22, c′) surrounds the external auditory
meatus: it is thicker and more opaque than the entotympanic.
The bulla lies ventrad of the squamous, and in an external view
conceals a large part of the petrous.
On its lateral surface it presents near the dorsal border an
irregular oval opening, that of the external auditory meatus
(Fig. 22, i), which leads into the tympanic cavity. Caudad of[33]
the external auditory meatus is a nearly dorsoventral groove,
which, when the bone is articulated, forms a part of the boundary
of the stylomastoid foramen (Fig. 22, j); just ventrad of this
groove is a pit (k) which lodges the tip of the tympanohyal
bone.
Craniad the bone is produced into a short spine, the styliform
process (q), which lies in a horizontal groove in the
ventral surface of the basisphenoid. Laterad of this spine is
a groove for the tuba auditiva or Eustachian tube.
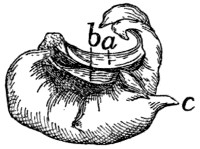
Fig. 24.—Tympanic
Bulla, Isolated,
Medial Surface.
a, inner end of auditory
meatus; b, partition
dividing tympanic cavity;
c, styliform process.
The medial surface (Fig. 24) presents in the middle near
its ventral margin a short triangular spine which lies in the
natural state against the ventral surface of
the basilar portion of the occipital.
Caudad of this spine the surface is
marked by two or three vertical parallel
grooves (Fig. 23, m). They indicate the
portion of the bone which bounds the jugular
foramen, and are possibly impressions of
the ninth, tenth, and eleventh nerves.
The dorsal two-thirds of the medial
surface is lacking in the disarticulated bulla
(Fig. 24), so that the cavity of the bone is
exposed. This opening is in the natural state closed by the
petrous bone. The caudal end is rough where it is overlaid
by the jugular process.
On the inner surface of the tympanic bulla is seen the thickened
margin of the inner end of the auditory meatus (Fig.
24, a). To it is attached the membrana tympana. In the
median dorsal line this margin is notched for the reception of
the incus and head of the malleus. From the lateral wall of
the cavity at the line of junction of the ectotympanic and entotympanic
a thin bony partition (b) rises. It runs almost directly
mediad; is concave dorsally and divides the tympanic cavity
into two chambers.
The Petrous Portion (Fig. 23, b, and Fig. 25).—This consists
of two parts, a very dense part (the petrous portion proper,
Fig. 25), which has the form of a triangular pyramid and
encloses the auditory labyrinth, and a less dense part, the[34]
mastoid portion (Figs. 22 and 23, e), which is flattened and
triangular and is attached by its base to the base of the
pyramid.
The petrous portion may be described as having a base and
three sides, lateral, dorsal, and medial. It completes the
medial wall of the tympanic bulla, so
that it is not possible to see it from the
exterior of a skull except through the
auditory meatus (Fig. 22, i). When the
bones of the skull are articulated its dorsal
surface is covered by the tentorium
and alisphenoid. Its lateral face looks
into the tympanic cavity, while the
medial face looks into the cranial cavity.
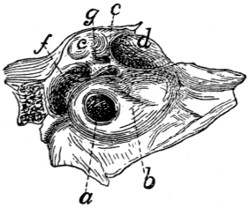
Fig. 25.—Petrous Bone of
Right Side, Lateral
Surface, Enlarged.
a, fenestra cochleæ; b, promontory;
c, fenestra vestibuli;
d, fossa for the tensor
tympani muscle; e, fossa for
incus and malleus; f, fossa
continuous with stylomastoid
foramen; g, foramen leading
to facial canal.
Its lateral face (Fig. 25) (medial
wall of the tympanic cavity) presents
just ventrad of the middle of its base a
large circular foramen, the fenestra
cochleæ (a) (or fenestra rotunda), which
looks caudolaterad; it leads into the
cochlea. The fenestra cochleæ lies at the summit of a nipple-like
elevation, the promontory (b), which is continued toward
the apex of the bone as a gradually diminishing semicylindrical
ridge, due to the presence within it of the bony cochlea.
Dorsad of the fenestra cochleæ is the much smaller fenestra
vestibuli (c) (or ovalis) which leads into the vestibule. It is
occupied in the natural condition by the base of the stapes.
Dorsocraniad of the fenestra vestibuli is a large fossa (d)
which contains the tensor tympani muscle. Dorsocaudad of
this, partly bounded by the squamous portion of the temporal,
is another large fossa (e), the cranial end of which is occupied
by the incus, while its caudal end is occupied by the head of
the malleus.
Nearly caudad of this fossa and separated from it by an
oblique bony septum is a third fossa (f) which is narrow and
curved. It is continuous with a notch in the mastoid portion
of the bone. When the tympanic is articulated the notch is
converted into a foramen (stylomastoid foramen, Fig. 22, j)[35]
for the exit of the seventh nerve. The fossa gives passage to
the seventh nerve and also lodges the stapedius muscle. A
groove may be traced from the stylomastoid foramen to the
caudal border of the fossa for the tensor tympani muscle, where
it passes into a canal (g). The groove and canal are parts of
the canalis facialis or facial canal (aqueductus Fallopii) for
the passage of the seventh nerve through the petrous bone.
The medial surface (Fig. 23, b) of the petrous portion
shows near its middle a fossa, the internal auditory meatus (n).
This is divided by a partition of bone into a dorsal and ventral
part. The dorsal portion is the beginning of the facial canal
(aqueductus Fallopii) by which the seventh nerve passes
through the petrous bone to emerge at the stylomastoid
foramen. The ventral portion shows at its bottom several
small foramina for the auditory nerve.
Dorsocaudad of the internal auditory meatus is a deep
fossa (o) for a small lobe, the so-called appendicular lobe, of
the cerebellum. This may be called the appendicular fossa.
The dorsal surface is triangular and presents near its apex
a foramen—the hiatus facialis (p), the opening of a canal
which joins the canalis facialis and transmits the superficial
petrosal branch of the nerve of the pterygoid canal (Vidian
nerve). That part of the dorsal surface which lies caudad of
the hiatus facialis is known as the tegmen tympani.
The base of the petrous is attached to the mastoid portion
(Fig. 23, e).
(For an account of the structures within the petrous bone
and the tympanic cavity, see the description of the internal
and middle ear.)
The mastoid portion (Figs. 22 and 23, c) is attached by its
base to the pyramidal petrous portion, with which it forms an
angle of about 120 degrees. It appears in the lateral wall of
the skull between the parietal bone and the occipital (Fig.
40, d). The lambdoidal ridge is continued on its outer surface
to the caudal border of the external auditory meatus. Caudad
of the stylomastoid foramen it forms a slight nipple-like
eminence, the mastoid process (Fig. 22, l). Its inner face
looks into the cranial cavity.
[36]
Parietal Bone. Os parietale
(Figs. 39, 40, and 43, 3).—The
parietal bones form the larger part of the lateral and dorsal
boundary of the cranial cavity. Each is a thin rectangular
bone, compact and curved and with a deeply notched shelf of
bone, the tentorium (Fig. 42, e, and Fig. 43, f), projecting
inward from near the caudal margin.
The outer surface is smooth and convex. The highest part
of the convexity, a little caudad of the middle of the bone, is
known as the parietal tubercle or eminence (Fig. 39, d); it
marks the point of beginning ossification. An obscure curved
ridge (Fig. 39, e), running from the caudodorsal angle or a
point craniad of it craniolaterad, indicates the boundary of the
origin of the temporal muscle. Near the ventral border the
surface is roughened and is covered in the natural state by a
part of the squamous portion of the temporal bone.
The inner surface (Fig. 43, 3
and 3′) is smooth and marked
by ridges and grooves for the convolutions of the cerebrum.
Near the medial border is a ridge which, when the bone is
articulated with that of the opposite side, forms a shallow groove
for the superior sagittal sinus. Beginning near the middle of
the ventral margin and passing dorsad is a groove for the
middle meningeal artery. The tentorium (Fig. 43, f) arises
from the inner surface near its caudal margin and projects
mediad as a thin curved or notched shelf of bone which
separates the cerebellar fossa (Fig. 43, I) of the cranium from
the cerebral fossa (Fig. 43, II). When the parietals are
articulated there is left between the tentoria a large foramen
by means of which the two fossæ communicate. The foramen
is bounded laterally and dorsally by the free margins of the
tentoria, while the ventral end of each tentorium articulates
with the alisphenoid, and its dorsal end with the opposite tentorium.
The medial border is straight and is united by suture to the
opposite bone.
The cranial border is bevelled at the expense of the inner
surface and articulates with the frontal. Just ventrad of the
middle of the border projects a sharp spine which fits into a
corresponding notch in the caudal border of the frontal.
[37]
The ventral border is concave, sharp, and bevelled at the
expense of the outer surface, for articulation with the squamous
portion of the temporal, except near the cranial end, where it
articulates with the wing of the sphenoid.
The caudal border is thick and porous medially, but thin
laterally, and bevelled at the expense of the inner surface for
articulation with the interparietal and mastoid portion of the
temporal.
Frontal Bone. Os frontale
(Figs. 39, 40, and 41, 5; Fig.
43, 8; Fig. 26).—The frontal bones meet one another in the
median dorsal line so as to form the roof of the skull between
the parietal and nasal bones. A part extends also ventrad,
forming a large part of the medial wall of the orbit and a part
of the temporal fossa.
The bone may be divided into two portions, a plate forming
the cranial portion of the roof of the skull and a part of the
roof of the nasal cavity, the frontal plate (Fig. 40, 5), and a
part descending into the orbit, the orbital plate (Fig. 40, 5′).
The frontal plate (Fig. 40, 5) is a right-angled triangle with
the hypothenuse lateral. Its dorsal surface is convex and
smooth. The cranial two-thirds
of its lateral border is separated
from the orbital fossa by a ridge,
the supraorbital arch or margin
(Fig. 39, i; Fig. 40, o);
the caudal third passes gradually
into the temporal fossa. At its
cranial angle is a triangular projection,
the frontal spine or
nasal spine (Fig. 26, a), which
fits into a space between the
nasal and maxillary bones.
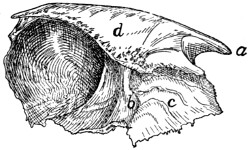
Fig. 26.—Frontal Bone, Medial
Surface.
a, frontal spine; b, transverse ridge;
c, surface applied to the ethmoid; d,
vertical plate of medial border.
The ventral surface is concave and smooth over its caudal
one-half and helps to form the cranial part of the brain-case.
It presents slight ridges and depressions for convolutions of the
cerebrum. At its narrowed middle region the ventral surface
is marked by a thick transverse ridge (Fig. 26, b). Caudally
the ridge descends by a gentle slope to the level of the ventral[38]
surface of the bone. The cranial end of the ridge is pierced
by an oval foramen through which the frontal sinus (Fig. 43,
m, m′), which lies within the ridge, communicates with the
spaces in the ethmoid bone (nasal cavity). Craniad of the
ridge the surface (Fig. 26, c) is rough and, together with the
raised medial border of the bone and the orbital plate, encloses
a rectangular space which in the natural state receives a portion
of the labyrinth of the ethmoid. The ventral surface is marked
at its medial edge by a thin longitudinal ridge which, when the
bones are articulated, is continuous with one of the vertical
lamellæ of the ethmoid.
The medial border forms a vertical plate (d), broadest
craniad and roughened for articulation with its fellow of the
opposite side except at its cranial end, where it articulates with
the border of the nasal bone.
The caudal border is roughened, bevelled at the expense
of the outer surface, and articulated with the parietal bone
except at its ventral end, where it articulates with the alisphenoid.
The lateral border is smooth, and it is here that the orbital
plate is joined to the frontal plate at right angles. Along its
cranial two-thirds this union is marked by a sharp ridge, the
supraorbital margin (Fig. 40, o) or arch. This ridge extends
caudolaterad as a triangular projection, the zygomatic (or
postorbital) process (Fig. 40, n), which is flattened on its
cranioventral face near its extremity and forms part of the
boundary of the orbital fossa. At its cranial end the lateral
border articulates with the nasal and maxillary bones.
The orbital plate (Fig. 40, 5′) arises from the ventral surface
of the lateral border of the frontal plate. It is directed ventrad,
is smooth and concave on its outer surface, and forms the dorsal
portion of the medial wall of the orbital fossa. Near its ventral
border it bears the small ethmoidal foramen, for the artery of
the same name.
On the caudal one-half of its inner surface (Fig. 26) it
assists the caudal part of the dorsal plate in forming the brain-case.
The cranial one-half of its inner surface is marked off
from the remainder of the surface by a sharp irregular ridge[39]
which is for articulation with the cribriform plate of the ethmoid.
Craniad of this the surface is marked by ridges and looks into
the nasal cavity.
The cranial margin is produced dorsally in the form of a
blunt triangular spine. Mediad of this spine the bone articulates
with the lachrymal bone.
The ventral border articulates by its cranial one-third with
the orbital plate of the palatine, and by its caudal two-thirds
with the body and wing of the presphenoid.
Maxillary Bone. Maxilla
(Figs. 27 and 28).—The
maxillary bone forms the cranial and lateral portions of the
roof of the mouth. The bones of opposite sides meet craniad,
but diverge caudad to enclose the palatal plates of the palatine
bones. Each consists of a thick prismatic ventral portion or
body (a) and a thin flat plate, the frontal process (b), extending
dorsad from the cranial part of the bone.
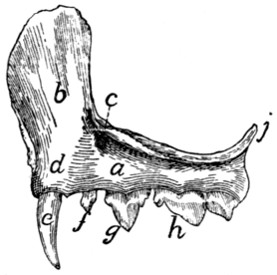
Fig. 27.—Maxillary Bone,
Lateral Surface.
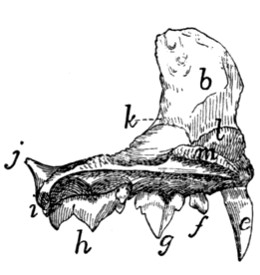
Fig. 28.—Maxillary Bone,
Medial Surface.
a, body; b, frontal process;
c, infraorbital foramen; d, elevation for root of
canine tooth; e, canine tooth; f, first premolar; g, second premolar; h, third premolar;
i, molar tooth; j, zygomatic process; k, beginning of lachrymal canal; l,
ridge to which the ventral nasal concha is attached; m, nasal crest of palatine
process.
The body (a) has the form of a triangular prism whose
broader dorsal face looks into the nasal cavity and orbit, while
the ventral face looks into the mouth, and the lateral face
toward the cheek. From the junction of the dorsal and lateral
surfaces at the cranial end the large flat curved frontal process
(b) passes dorsad, while the teeth are implanted along the
border, alveolar border or process, formed by the junction of
the ventral and lateral surfaces.
[40]
The lateral surface is continuous with the lateral surface of
the frontal process and shows at the base of the frontal process
on its caudal border the large infraorbital foramen (Fig.
27, c), for the vessels and nerves of the same name. Near the
medial end of the surface is a cylindrical elevation (d) for the
root of the canine tooth (e).
The ventral surface is smooth and looks into the roof of the
mouth.
On the dorsal surface caudal and cranial halves may be distinguished.
The caudal one-half enters into the floor of the
orbit. The lateral edge of this portion is divided into two
laminæ, between which is received the end of the malar bone.
Caudad this edge is prolonged into the short dorsally directed
zygomatic process (j). The cranial half of the dorsal surface
looks into the nasal cavity and is separated from the caudal
half by a sharp vertical lamina of bone which runs caudomediad
from the base of the nasal process. To the dorsal edge
of this lamina are articulated the lachrymal bone and a part of
the palatine. At the point where the lamina joins the base of
the nasal process a foramen is seen leading into a canal, the
nasolachrymal canal (k). Craniad of the lamina the surface
is concave. Where it becomes continuous with the inner edge
of the frontal process there is attached to it a thin bone, the
ventral nasal concha (or maxilloturbinal), which is rolled into
an irregular spiral. The nasolachrymal canal opens ventrad
of its cranial end.
The cranial third of this part of the bone projects further
mediad than does the rest of the medial border, forming thus
the broad palatine process. This is rough on its medial edge
for articulation with the premaxillary and the palatine process
of the opposite bone. This medial edge rises also dorsally into
a low ridge, the nasal crest (m), which is roughened for articulation
with the vomer. The caudal two-thirds of the medial
edge articulates with the palatine bone.
The cranial end of the bone articulates with the premaxilla.
The caudal end is smooth.
The frontal process (b) presents on its inner surface, which
looks into the nasal cavity, certain transverse ridges which are[41]
in relation with the ethmoid bone. Its outer surface is smooth.
By its cranial border it articulates with the nasal bone dorsally
and with the premaxillary bone ventrally.
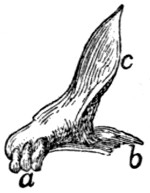
Fig. 29.—Premaxillary
Bone, Obliquely
Craniolateral
Aspect.
a, the three incisor
teeth; b,
palatal portion of
the bone; c, nasal
process.
Its dorsal end articulates medially with the nasal spine of
the frontal bone, and caudally with the orbital plate of the same
bone.
Premaxillary Bone. (Os incisivum BNA.) Premaxilla
(Fig. 29).—The premaxillary bones bear the incisor teeth and
form the cranial portion of the roof of the mouth.
Each consists of an irregular, horizontal palatal portion (b)
and of a perpendicular nasal process (c) which forms part of
the lateral boundary of the nares and enters into
the formation of the lateral wall of the nasal
cavity.
The palatal portion has in its caudal border
a deep notch for the foramen incisivum or anterior
palatine canal, which lies between it and the
maxillary and transmits blood-vessels and nerves.
It articulates with the maxillary bone by this
border.
The medial border is raised into a thin crest
of bone which, besides forming the medial wall of
the foramen incisivum or anterior palatine canal,
articulates by its medial border with the bone of
the opposite side, forming a sort of median trough (sulcus palatinus)
which projects dorsad into the nasal cavity and receives
the ventral border of the nasal septum. The caudal end of this
border articulates laterad with the maxilla, dorsad with the
vomer.
Its craniolateral border bears the incisor teeth (a).
The nasal process (c) presents three surfaces, all elongated
and triangular; one, the medial surface, is smooth and concave
and looks into the nasal cavity. Its dorsal border is rough for
articulation with the nasal bone dorsad, and smooth ventrad
where it aids in forming the nares.
The lateral surface is smooth.
The caudal surface is rough for articulation with the maxillary
bone.
[42]
Nasal Bone. Os nasale
(Fig. 30).—The nasal bones fill
the space between the nasal process of the premaxillary, the
frontal process of the maxillary, and the nasal spine
of the frontal bone (Fig. 39, 7). They thus form
part of the dorsal wall of the nasal cavity near the
middle line.

Fig. 30.—Nasal Bone,
Dorsal
View.
Each may be described as consisting of two
elongated triangular lamellæ, one vertical, the other
horizontal. The vertical lamella is curved slightly
ventrad and has its apex directed craniad. It is
applied by its medial surface against the vertical
lamella of the opposite bone, the two thus forming
a median vertical partition, the nasal crest (Fig. 43, 12), which
extends ventrad into the nasal cavity and, by joining the dorsal
edge of the lamina perpendicularis, helps to form the internasal
septum.
The horizontal lamella is attached to the dorsal margin of
the vertical lamella in such a way that its apex lies opposite
the base of the vertical lamella. It helps to roof in the nasal
cavity, and by its base forms a part of the dorsal boundary of
the narial opening. By its lateral margin it articulates with
the nasal spine of the frontal at its caudal end, with the frontal
process of the maxillary at its middle, and with the nasal process
of the premaxilla at its cranial end. The lateral angle of its
base projects in a curved line which forms the dorsal part of
the lateral boundary of the narial opening.
From the lateral border of the horizontal lamella a bony
plate curves ventrad and mediad, enclosing a narrow fossa which
receives a part of the ethmoid. This is the concha nasalis
superior (nasoturbinal bone).
Ethmoid Bone. Os ethmoidale
(Figs. 31 and 32).—The
ethmoid bone closes in the cranial cavity at its cranial end and
extends forward into the nasal cavity, which it largely fills.
It consists of a median vertical portion, the lamina perpendicularis
(Fig. 43, n; Fig. 42, p), forming a part of the nasal
septum, of two lateral portions made of thin sheets of bone
variously folded and united—the labyrinths (or ethmoturbinals),
which fill the greater part of the nasal cavity; and of[43]
a transverse perforated plate, the cribriform plate (lamina
cribrosa), attached to the caudal end of the lamina perpendicularis
and the labyrinths.
The lamina perpendicularis (Fig. 43, n; Fig. 42, p) is a
flat four-sided bone. By its caudal margin it is continuous with
the cribriform plate; by its ventral margin it is enclosed by the
halves of the vomer; by its dorsal margin it unites with the
crest formed by the vertical portion of the nasal bone craniad
and with the vertical lamina of the medial margin of the frontal
caudad, while its cranial margin is continued into the septal
cartilage of the nose. Its lateral faces are smooth and free.
The lamina cribrosa or cribriform plate (Fig. 42, o) is
elongated heart-shaped, with the apex of the heart ventrad.
Its caudal face is concave and looks into the cranial cavity.
It presents three irregular longitudinal rows of holes, one
median and two lateral, for the passage of the olfactory fibres
from the cranial cavity into the nasal cavity. Its cranial face
is continuous along the medial line with the lamina perpendicularis,
and at the sides with the labyrinths.
The notch in the heart is directed dorsad and receives the
vertical lamina of the medial border of the frontal bone. The
apex of the heart articulates with the cranial end of the dorsal
surface of the presphenoid. Its lateral margins are articulated
with the ethmoidal ridges on the medial surface of the frontal
bone.
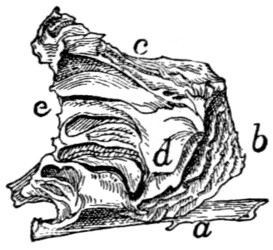
Fig. 31.—Ethmoid and Vomer,
Side View.
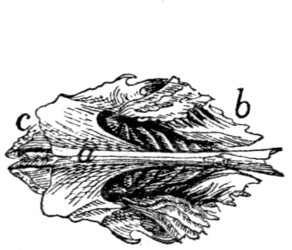
Fig. 32.—Ethmoid and Vomer,
Ventral View.
a, vomer; b, vertical cells of the labyrinth of the ethmoid; c, horizontal cell of
the same; d, part of the ethmoid that forms the lamina papyracea; e, edge of cribriform
plate.
The labyrinths (Figs. 31 and 32) are attached to the cranial
face of the lamina cribrosa, one on each side of the lamina
perpendicularis. Each is made of thin bony plates irregularly[44]
folded so as to enclose spaces, the ethmoid cells. In each
may be distinguished a cranial portion (b), in which the cells
are nearly vertical, and a caudal portion (c), in which the cells
are nearly horizontal.
The medial surfaces are separated by a space from the
lamina perpendicularis. This space is broadest along the
junction of the horizontal and vertical portions of the labyrinth.
There are thus formed two passageways which correspond to
the superior meati of human anatomy.
The lateral surfaces come into contact with the frontal
process of the maxillary and the orbital plate of the frontal
bone. On the lateral surface of each labyrinth there is a thin
irregular lamina of bone lying in a dorsoventral longitudinal
plane and closing in some of the ethmoid cells laterally (d).
A small part of this lamina, situated near the caudoventral
angle of the bone, appears in the orbital fossa on the external
surface of the skull between the presphenoid, palatine, and
frontal bones or between the lachrymal, palatine, and frontal
bones. Sometimes in the entire skull two such pieces may be
seen, one in each of these positions. This corresponds to the
lamina papyracea of human anatomy.
The dorsocaudal angle of each bone is received into the
space between the orbital plate of the frontal and the vertical
lamina of the medial border of the frontal. Its ventrocaudal
angle is received between the cranial extensions of the lateral
walls of the presphenoid, while its ventral surface is overlaid
caudally by the expanded portion of the vomer, to which it is
attached at its caudolateral angles.
Vomer
(Figs. 31 and 32, a).—The vomer consists of two
thin laminæ of bone which ensheath the ventral margin of the
lamina perpendicularis (or the cartilaginous plate which continues
ventrad from this margin) and unite ventrad of it; the
two thus form a trough open dorsad.
Each becomes horizontal near its caudal end and at the
same time expands. The expanded portion lies ventrad of the
labyrinth of the ethmoid, closing in some of its cells: its lateral
angles are united with the labyrinths.
At its caudal end the bone articulates with the body of the[45]
presphenoid, and each half of it is produced caudad near the
middle line into a triangular spine which lies ventrad of the
body of the presphenoid. The horizontal portion of the bone
helps to separate the olfactory and respiratory passages of the
nasal chamber, while its vertical portion contributes to the
formation of the nasal septum.
The ventral margin formed by the junction of the two
halves of the bone is smooth and free caudad, but at its cranial
end is broad and rough for articulation with the palatal
processes of the maxillæ.
Palatine Bone. Os palatinum
(Fig. 33).—The palate
bone or palatine bone consists of two portions, a horizontal or
palatal portion (a) and a perpendicular or nasal portion (b),
uniting at an angle of about forty-five degrees.
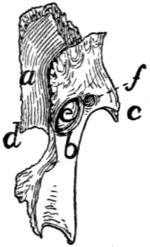
Fig. 33.—Palatine
Bone,
Dorsal View.
a, horizontal
portion; b, perpendicular
portion;
c, maxillary
spine; d, posterior
nasal spine; e,
sphenopalatine
foramen; f, caudal
opening of
posterior palatine
canal.
The horizontal portions (a) of the two bones are received
between the maxillary bones and form the caudal and medial
part of the roof of the mouth. Each is irregularly
quadrilateral in form, with the caudolateral angle
produced caudad into a long process which is
continuous with the perpendicular portion of the
bone. The lateral margin of the horizontal portion
articulates over its cranial half with the
maxillary bone. At about its middle a short thick
maxillary spine (c) projects caudolaterad. The
remainder of the lateral margin is directly continuous
with the perpendicular plate of the bone.
The medial margin is rough for articulation with
the corresponding margin of the opposite palatine;
the caudal angle of this margin projects
caudad as the short posterior nasal spine (d).
The caudal margin forms a free edge which
bounds the choanæ; it passes laterally into the
perpendicular portion.
The ventral surface (Fig. 41, 8) looks into the mouth.
Near the middle of its craniolateral margin are two or more
small foramina (Fig. 41, q) which form the cranial termination
of the posterior palatine canal. The dorsal surface is smooth
and looks into the nasal cavity.
[46]
The perpendicular or nasal portion (Fig. 33, b) of the
palatine is thin and irregularly quadrilateral in form. It is
attached by its cranial two-thirds to the dorsal surface of the
horizontal portion. The outer surface is concave and looks
into the orbital fossa. The inner surface is convex and looks
into the nasal cavity.
The perpendicular portion is marked by two foramina just
craniad of the middle. The larger dorsal oval foramen is the
sphenopalatine foramen (e). The smaller ventral foramen is
the caudal opening of the posterior palatine canal (f). From
this opening the canal passes craniomediad, lying in the substance
of the palatine bone; it opens on the ventral surface of
the horizontal portion at the small openings previously
described (Fig. 41, q).
By its cranial margin it articulates with the lachrymal bone.
By its dorsal margin it articulates craniad with the orbital plate
of the frontal: with the lamina papyracea at its middle, and
with the body of the presphenoid caudad. The caudal half of
the dorsal margin is partially divided into two lamellæ with a
rough surface between them: this rough surface lies against
the ventral surface of the presphenoid. The caudal margin
articulates with the pterygoid portion of the sphenoid.
Lachrymal Bone. Os lachrymale
(Fig. 34; Fig. 39, 10).—The
lachrymal bone is a thin pentagonal scale of bone filling
the interval between the horizontal plate of the palatine, the
maxillary, and the orbital plate of the frontal. Its outer surface
looks into the orbit, its inner surface into the nasal cavity.
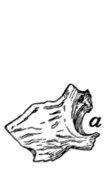
Fig. 34.—Lachrymal Bone of
Left Side, External Surface.
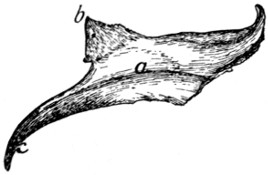
Fig. 35.—Malar Bone of Right Side,
Lateral Surface.
Fig. 34.—a, notch forming the beginning of the lachrymal canal.
Fig. 35.—a, ridge for origin of the masseter muscle; b, frontal process; c, zygomatic
process.
[47]
Near the middle of its cranial border it is notched obliquely
by a foramen (a), the beginning of the nasolachrymal canal.
Malar Bone. Jugal Bone. Os zygomaticum
(Fig. 35).—The
malar or zygomatic bone is a flat curved plate of bone
which forms the lateral wall of the orbit and together with the
zygomatic process of the temporal forms the zygomatic arch.
Its outer surface is smooth and marked by a longitudinal ridge
(a) for attachment of the masseter muscle.
At its caudal end the bone is continued into two processes:
one, the frontal process or orbital process (b), is a triangular
spine of bone directed caudomediad; when the bones are
articulated it lies opposite the zygomatic process of the frontal
to which it is joined by a ligament (orbital ligament). The
other, zygomatic process (c) of the malar bone, extends ventrocaudad
and articulates with a similar process from the temporal
to form the zygomatic arch above mentioned.
Its inner surface is smooth and looks into the orbit, except
that of the zygomatic process, which looks into the temporal
fossa.
Its cranial border is roughened at the expense of both surfaces
and articulates with the maxillary bone. Its other
borders are smooth except the dorsal border of the zygomatic
process, which is roughened for attachment to the zygomatic
process of the temporal.
The Mandible. Mandibula
(Figs. 36 and 37).—The
mandible (or inferior maxillary bone) is composed of two
halves which come together at the cranial end and form the
lower jaw. At its caudal end each half articulates with the
temporal bone at the mandibular fossa, and at its cranial end it
joins the opposite bone, the suture being known as the symphysis
of the jaw (symphysis menti) (Fig. 37, a).
Each half consists of a horizontal portion, the body (b),
bearing teeth on one of its borders (the alveolar border), and
of a vertical portion, the ramus (c).
The body (b) has the form of a flattened cylinder and has
two surfaces and two borders. The lateral surface (Fig. 36)
is smooth and presents near its cranial end a foramen (or
sometimes two), the mental foramen (d), forming the cranial[48]
termination of the mandibular canal. At its caudal end is a
deep fossa continuing on to the ramus, the coronoid fossa, or
masseteric fossa (e).
Fig. 36.—Mandible, Lateral Surface.
Fig. 37.—Mandible, Medial Surface.
a, symphysis; b, body; c,
ramus; d, mental foramina; e, coronoid fossa; f, mandibular
foramen; g, angular process; h, coronoid process; i, condyloid process;
1, 2, 3, the three incisor teeth; 4, the canine tooth; 5, 6, the premolars; 7, the
molar tooth.
The medial surface (Fig. 37) is smooth and has near its
caudal end a foramen, the mandibular foramen (f), which
communicates with the mandibular canal leading lengthwise
through the bone to the mental foramen. The cranial end is
roughened for attachment to the bone of the opposite side.
The ventral border is smooth and rounded; it ends caudally
in a blunt point, the angular process (g). The dorsal
(alveolar) border is slightly curved and bears the sockets
(alveoli) for the teeth. It is continuous with the cranial margin
of the coronoid process.
The ramus is divided into two portions, the coronoid process
(h) and the condyloid process (i). The coronoid process (h)
extends dorsocaudad as a thin plate of bone with smooth surfaces
and borders. Its outer surface is partly occupied by the
coronoid fossa (e). The condyloid process (i) has the form of[49]
a semicylindrical transverse piece of bone attached to the
caudal margin of the coronoid process. It articulates with the
mandibular fossa of the temporal bone.
Hyoid Bone. Os hyoideum
(Fig. 38 and Fig. 104).—The
hyoid bone forms the support for the tongue and gives
origin to muscles passing to the
tongue and larynx. It also
supports the thyroid cartilage
(Fig. 104, 1).
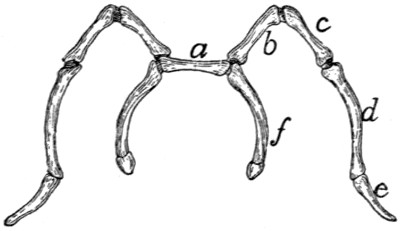
Fig. 38.—Hyoid Bone, Dorsal View.
a, body; b, c, d, e, cranial cornu; f,
caudal cornu; b, ceratohyal; c, epihyal;
d, stylohyal; e, tympanohyal; f, thyrohyal.
It consists of a transverse
bony bar, the body (Fig. 38, a)
and of two cornua or horns
attached to each end of the
body.
The cranial cornu (lesser
cornu of human anatomy) is the longer (Fig. 38, b-e). Each
arises from the cranial face of the body at its lateral end, curves
laterad, and then caudodorsad. It consists of four bony pieces
movably united by cartilage.
The terminal piece is the tympanohyal (e); it is imbedded
in the tympanic bulla just ventrad of the stylomastoid foramen.
It is not therefore seen attached to the cornu after the latter
has been separated from the skull. The other pieces become
successively shorter toward the body, and are called stylohyal
(d), epihyal (c), and ceratohyal (b).
The caudal cornua (f) (greater cornua of human anatomy)
arise from the ends of the body. Each consists of a single
piece of bone, the thyrohyal (f), which passes caudolaterad;
its free end is united to a process of the thyroid cartilage (Fig.
104, 1).
The Skull as a Whole.
—In the following description of the
skull as a whole the mandible, hyoid, and ear-bones are not
included.
The skull forms a bony box which contains the brain and
is produced craniad into the facial portion which encloses the
nasal cavity and forms the framework of the face.
In dorsal view (Fig. 39) the skull presents a smooth convex
surface, broadest caudad, with the two zygomatic arches[50]
(g) curving out some distance laterally. The following bones
are visible in dorsal view: the occipital (1), interparietal (2),
parietals (3), temporals (4),
frontals (5), malar or zygomatic
bones (6), nasals (7), maxillaries
(8), premaxillaries (9), and
lachrymals (10).
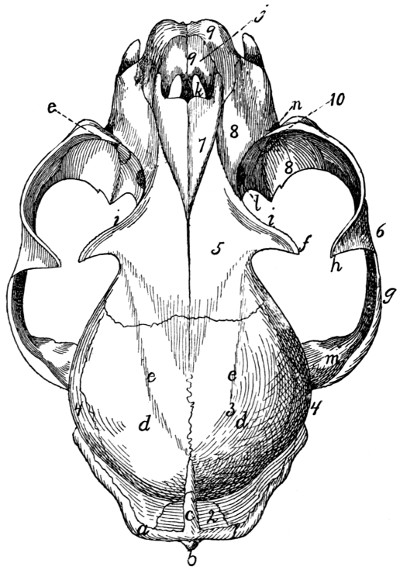
Fig. 39.—Skull, Dorsal Surface.
1, occipital bone; 2, interparietal bone; 3, parietal bones; 4, temporal; 5, frontal;
6, malar; 7, nasal; 8, maxillary; 9, premaxillary; 10, lachrymal, a, lambdoidal
ridge; b, external occipital tubercle; c, sagittal crest; d, parietal eminence; e, line
which forms the dorsal boundary of the temporal fossa; f, zygomatic process of the
frontal; g, zygomatic arch; h, frontal process of the malar; i, supraorbital arch; j,
nares; k, foramen incisivum or anterior palatine foramen; l, sphenopalatine foramen;
m, zygomatic process of the temporal; n, infraorbital foramen; o, opening of lachrymal
duct.
The caudal boundary of the dorsal surface is marked by the
prominent lambdoidal ridge (a) which passes from the middle
cranioventrad along each side to the root of the zygomatic
arch: it is borne by the occipital and temporal bones. From
the middle of the lambdoidal ridge a second ridge, the sagittal[51]
crest (c), passes craniad in the middle line across the interparietal
bone: it varies greatly in extent, reaching in a very old
and muscular cat to the cranial border of the parietals, while
in kittens it does not exist. The most prominent portions of
the skull in this region, just craniad of the middle of the parietal
bones, are known as the parietal tubercles or eminences (d).
A faint curved line (e) runs from the cranial end of the sagittal
crest craniolaterad to the base of the zygomatic process of the
frontal: it marks the dorsal boundary of the origin of the temporal
muscle, and may therefore be considered the dorsal
boundary of the temporal fossa. This fossa extends from its
dorsal boundary as far laterad and caudad as the lambdoidal
ridge (a), and as far craniad as a line connecting the tip of the
zygomatic process of the frontal (f) with the frontal process of
the malar (h). The temporal muscle takes origin from its
surface.
The middle portion of the dorsal surface is formed by the
frontals (5). Each frontal presents laterally a prominent
zygomatic process (f), extending ventrolaterad toward a corresponding
(frontal) process (h) of the malar bone. These two
processes mark the boundary between the orbital fossa (craniad)
and the temporal fossa (caudad). Craniad of the zygomatic
process of the frontal a sharp margin separates the dorsal surface
of the skull from the wall of the orbital fossa: this is the
supraorbital arch or margin (i).
The cranial portion of the dorsal surface is formed by the
maxillary (8), nasal (7),
and premaxillary bones (9). Just
craniad of the nasals, bounded ventrad and craniad by the
premaxillaries, appears the large opening of the nares (j),
leading into the nasal cavity.
The zygomatic arch (g) is formed by the zygomatic process
of the temporal (m) and the malar or zygomatic bone (6).
Each presents near its middle a prominent dorsocaudally
directed process, the frontal process (h) of the malar bone. The
zygomatic arch forms the lateral boundary of the temporal and
orbital fossæ, which are separated by a line connecting the
frontal process of the malar (h) and the zygomatic process of
the frontal (f).
[52]
A portion of the floor of the orbit and the opening of the
lachrymal canal (o) may also be seen in dorsal view; they are
described in connection with the lateral surface.
The caudal surface of the skull is formed largely by the
occipital bone (Fig. 17), surrounding the foramen magnum
(Fig. 17, d). At the sides of the foramen magnum are the
two prominent curved occipital condyles (e) for articulation
with the atlas. Craniolaterad of the condyles, separated from
them by a deep notch, are the jugular processes (f) of the
occipital, closely applied to the caudal ends of the tympanic
bullæ.
Dorsad of the foramen magnum are faint indications of a
median ridge running dorsad, the external occipital crest
(Fig. 17, i); this rises at its junction with the lambdoidal ridge
to form the prominent external occipital tubercle (Fig. 39, b).
The dorsal and dorsolateral boundaries of the posterior surface
are formed by the lambdoidal ridge (Fig. 17, h; Fig. 39, a).
The lateral surface of the skull (Fig. 40) is much more
complicated than the dorsal and posterior surfaces. Caudally
the occipital condyles (a) and external occipital crest (b) are
visible; dorsocaudad the sagittal crest (c).
Extending from the caudal end of the sagittal crest the
lambdoidal ridge (d) is seen passing ventrocraniad to the tympanic
bulla, thence craniad to the root of the zygomatic arch.
In the ventral part of the caudal region the tympanic bulla (e)
is visible with the jugular process (f) of the occipital pressed
close against its caudal end. Just craniad of the jugular process
the mastoid process (g) of the temporal rests against the side
of the bulla. Beneath the cranial edge of this process is the
opening of the stylomastoid foramen (h) for the seventh nerve,
while just ventrad of the foramen is the small pit (i) in the
tympanic bulla for the reception of the tympanohyal bone.
Craniad of the stylomastoid foramen is the large opening of the
external auditory meatus (j), leading into the middle ear.
Immediately dorsocraniad of the external auditory meatus
the zygomatic arch begins as the zygomatic process (k) of the
temporal bone. On the cranial surface of the base of this
process is the deep mandibular fossa (l) for the condyle of the[53]
mandible. This fossa is bounded caudally by the prominent
postmandibular process (m).
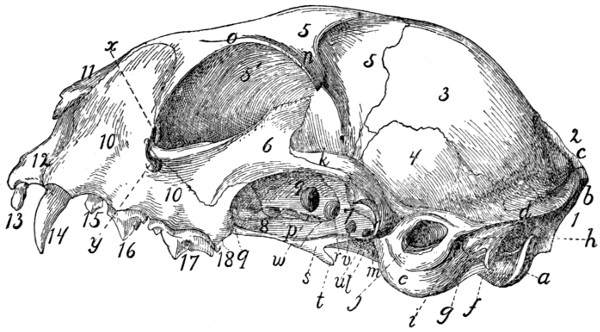
Fig. 40.—Skull, Side View.
1, occipital bone; 2, interparietal; 3, parietal; 4, temporal; 5, 5′, frontal; 6,
malar; 7, sphenoid; 8, palatine; 9, presphenoid; 10, maxillary; 11, nasal; 12, premaxillary;
13, incisor teeth; 14, canine; 15, 16, 17, premolars; 18, molar. a, occipital
condyle; b, external occipital crest; c, sagittal crest; d, lambdoidal ridge; e,
tympanic bulla; f, jugular process; g, mastoid process; h, stylo-mastoid foramen; i,
pit for tympanohyal bone; j, external auditory meatus; k, zygomatic process of temporal
bone; l, mandibular fossa; m, postmandibular process; n, zygomatic process
of the frontal; o, supraorbital margin; p, external pterygoid fossa; q, sphenopalatine
foramen; r, orbital fissure; s, internal pterygoid fossa; t, hamulus; u, foramen
ovale; v, foramen rotundum; w, optic foramen; x, opening of lachrymal canal; y,
infraorbital foramen.
All that portion of the lateral surface of the skull which lies
craniodorsad of the lambdoidal ridge may be divided (excluding
the zygomatic arch) into three main parts, the temporal
fossa, the orbital fossa, and the face. The boundaries of the
temporal fossa have been given. The orbital fossa is bounded
externally by a prominent semicircular ridge formed chiefly by
the zygomatic arch, the zygomatic process of the frontal (n),
and the supraorbital arch (o) of the frontal, which may be
traced to the cranial root of the zygomatic arch. The orbital
fossa may be considered to end caudally and ventrally at the
level of the optic foramen (w); ventrad of it are certain smaller
fossæ. Immediately ventrad is the long external pterygoid
fossa (p), from which arises part of the external pterygoid
muscle. This fossa begins at the sphenopalatine foramen (q)
and extends caudad to the orbital fissure (r); it is separated
by a ridge from the orbital fossa. Caudoventrad of the external[54]
pterygoid fossa and separated from it by a sharp ridge is the
small narrow internal pterygoid fossa (s), which extends ventrad
without interruption on to the surface of the hamulus (t)
and caudad to within two or three millimeters of the tympanic
bulla. From it the internal pterygoid muscle takes origin.
The hamulus (t) projects caudoventrad in this region, forming
a prominent feature in a lateral view.
Four foramina leading into the cranial cavity are visible in
a lateral view of the skull, craniad of the tympanic bulla. The
one nearest the bulla is the foramen ovale (u) for the third
division of the fifth nerve; next craniad of this is the foramen
rotundum (v) for the second division of the fifth nerve. These
two foramina pierce the alisphenoid: just craniad of them,
between the alisphenoid and the orbitosphenoid, is the large
orbital fissure (r) (foramen lacerum anterius), which transmits
the third, fourth, and sixth cranial nerves and the first division
of the fifth. Dorsocraniad of the orbital fissure is the optic
foramen (w), for the optic nerve.
Ventrad of the cranial portion of the orbit is the large
sphenopalatine foramen (q), for the nerves and arteries of the
same name. Just craniad of this is the small caudal opening
of the posterior palatine canal, which passes through the substance
of the palatine bone and opens on its ventral surface
near its cranial margin. Just dorsad of the cranial root of the
zygomatic arch is the opening of the lachrymal canal (x),
while the root of the arch is pierced by the large infraorbital
foramen (y), which transmits the infraorbital nerves and artery
from the orbit.
The teeth (13-18), implanted along the alveolar border of the
maxillary and premaxillary, form a prominent feature in a
lateral view: they are described in the account of the alimentary
canal.
The ventral surface of the skull (Fig. 41) is very complex.
It is separated by the orbits into a caudal and a cranial portion,
united by a narrow median trough-like part. Laterad of this
trough-like part are visible parts of the orbit and the zygomatic
arches, which do not properly belong to the ventral surface and
have already been described.
[55]
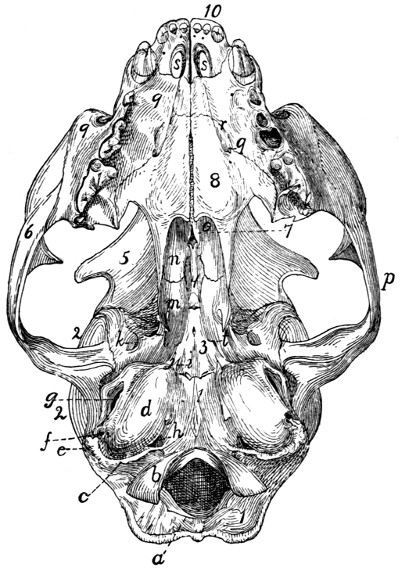
Fig. 41.—Skull, Ventral View.
1, occipital bone; 2, temporal; 3, sphenoid; 4, presphenoid; 5, frontal; 6, malar;
7, vomer; 8, palatine; 9, maxillary; 10, premaxillary. a, foramen magnum; b,
occipital condyles; c, jugular process; d, tympanic bulla; e, mastoid process; f, stylomastoid
foramen; g, external auditory meatus; h, jugular foramen; i, styliform process;
j, groove for Eustachian tube; k, foramen ovale; l, foramen rotundum; m,
pterygoid process of sphenoid; n, perpendicular plate of palatine; o, choanæ or posterior
nares; p, zygomatic arch; q, cranial end of posterior palatine canal; r, palatine
grooves; s, foramina incisiva or anterior palatine foramina; t, opening of pterygoid
canal.
Caudally there appear in the ventral view the foramen
magnum (a), occipital condyles (b), and jugular processes (c).
In front of the jugular processes the two tympanic bullæ (d)
form prominent features, with the mastoid process (e), the
stylomastoid foramen (f), and the external auditory meatus
(g) on their lateral surfaces. All these structures have been
described. The tympanic bullæ (d) are placed with long axes
directed craniomediad, so that they converge toward their
cranial ends. At the caudomedial angle of each bulla is the[56]
large jugular foramen (h), for the ninth, tenth, and eleventh
nerves. Opening into the mediocaudal margin of the jugular
foramen is the smaller hypoglossal foramen, for the twelfth
nerve.
The craniomedial end of the tympanic bulla projects craniad
as the styliform process (i). Just laterad of this process is the
opening (j) into the tympanic bulla by which the tuba auditiva
or Eustachian tube passes into the middle ear. A faint groove
for the tube passes craniomediad from this opening, on the surface
of the sphenoid. Craniolaterad of the opening for the tuba
auditiva is the foramen ovale (k); craniad of this the foramen
rotundum (l) is faintly indicated. On the surface of the
sphenoid just craniad of the styliform process of the bulla tympani
is the minute opening of the pterygoid canal (t). The
orbital fissure and optic foramen are not seen in the ventral
view.
The middle region of the ventral surface is narrow: it is
formed by a trough-like fossa which is bounded laterally by the
pterygoid processes (m) of the sphenoid and the perpendicular
plates of the palatines (n). Ventrad of this lies, in the natural
condition, the soft palate, converting the fossa into the nasal
portion of the pharynx or nasopharynx. Craniad this fossa is
bounded by the free caudal edges of the palatines; beneath
which the fossa communicates with the nasal cavity by the two
choanæ (o). Laterad of this median fossa are visible in the
ventral view parts of the temporal and orbital fossæ, bounded
laterally by the zygomatic arches (p).
The cranial part of the ventral surface is a somewhat triangular
plane area formed by the palatal portions of the palatines
(8), maxillaries (9),
and premaxillaries (10), which together
constitute the hard palate (palatum durum). Laterad and
craniad this area is bounded by the alveolar borders of the
maxillaries and premaxillaries bearing the teeth. The hard
palate is marked near the cranial border of the palatine bones
with two or more foramina which form the cranial termination
of the posterior palatine canal (q). Two faint grooves pass
from these foramina a short distance craniad, gradually converging:
these are known as the palatine grooves (r)[57]
(sulci palatini). Near the cranial end of the hard palate are two
large openings close together near the middle line: these are
the foramina incisiva (or anterior palatine foramina) (s).
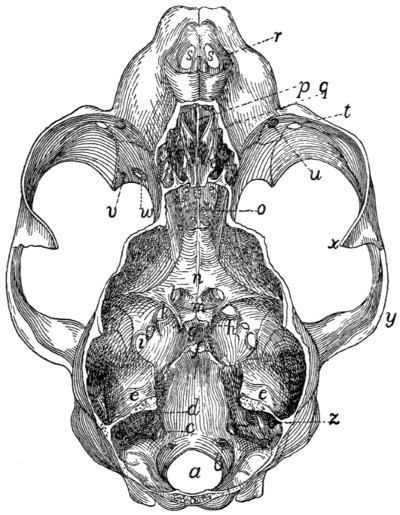
Fig. 42.—Skull, with
Dorsal Surface Removed, showing the Cranial
and Nasal Cavities.
a, foramen magnum; b, caudal end of hypoglossal canal; c, jugular foramen; d,
internal auditory meatus; e, tentorium, forming the cranial boundary of the cerebellar
fossa; f, dorsum sellæ; g, sella turcica; h, anterior clinoid processes; i, foramen
ovale; j, foramen rotundum; k, orbital fissure; l, optic foramen; m, chiasmatic
groove; n, presphenoid bone; o, cribriform plate; p, lamina perpendicularis of ethmoid;
q, labyrinths of ethmoid; r, nares; s, foramina incisiva or anterior palatine
foramina; t, infraorbital foramen; u, opening of the lachrymal canal; v, caudal
opening of posterior palatine canal; w, sphenopalatine foramen; x, frontal process of
the malar; y, zygomatic process of the temporal; z, appendicular fossa, in the petrous
bone.
Cavities of the Skull
(Figs. 42 and 43).—The bones of
the cranial portion of the skull enclose the cranial cavity for
the brain; the facial bones enclose the nasal cavity, for the
olfactory organ.
The cranial cavity is divisible into three principal fossæ:
the cerebellar fossa (Fig. 43, I) caudad,
for the cerebellum;[58]
the cerebral fossa (II) in the middle, for the cerebrum; the
small olfactory fossa (III) at the cranial end for the olfactory
bulb of the brain.
The cerebellar fossa (I) is bounded caudally by the occipital
bone enclosing the foramen magnum (Fig. 42, a). Its ventral
surface is formed by the basilar portion of the occipital and the
petrous portions of the temporals; its lateral surface by the
mastoid portions of the temporals and parts of the parietals and
occipital. Its roof is formed by the parietals and interparietal.
Craniad the cerebellar fossa is partly separated from the cerebral
fossa by the tentorium (Fig. 42, e; Fig. 43, f) formed by
the two parietals: this encloses a quadrangular opening by
which the two fossæ communicate. The caudal, dorsal, and
lateral walls of the cerebellar fossa are deeply marked by fossæ
for the lobes of the cerebellum; the small appendicular fossa
(Fig. 43, e), forming a deep indentation in the petrous bone
near its dorsocaudal end, is particularly noticeable.
The following openings are found in the walls of the cerebellar
fossa. Caudad is the large foramen magnum (Fig. 42, a)
by which the brain-cavity communicates with the vertebral
canal. Near the caudal margin of the foramen magnum, on
its lateral side, just mediad of the dorsal end of the occipital
condyle, is the caudal opening of the condyloid canal (Fig. 43, a)
which passes craniad through the substance of the occipital
bone to open just caudad of the petrous: it transmits a vein.
The condyloid canal varies greatly in size in different specimens.
A few millimeters craniad of the edge of the foramen
magnum on the floor of the fossa is the small opening of the
hypoglossal canal (Figs. 42 and 43, b), for the twelfth nerve.
Just craniad of this, at the caudomedial border of the petrous,
is the large jugular foramen (c). On the petrous itself, near
the middle, is the internal, auditory meatus (d) divided into the
dorsal facial canal for the seventh nerve, and a ventral passage
for the eighth nerve. At the cranial end of the cerebellar
fossa is the large opening bounded by the free edges of the
tentorium.
The cerebral fossa forms much the largest part of the
cranial cavity. It is bounded by the parietals (Fig. 43,
3′),[59]
squamous portions of the temporals (4), frontals (8), the
sphenoid (5), and presphenoid (6). A slight rounded ridge
on its lateral wall at about the position of the suture between
the frontals and parietals separates a smaller cranial portion
sometimes called the anterior fossa, from a larger caudal portion
sometimes known as the middle fossa of the cranial cavity.
The walls of the cerebral cavity are marked with numerous
ridges and shallow furrows for the cerebral convolutions.
The floor of the cerebral cavity is bounded caudad by the
prominent dorsum sellæ (Fig. 42, f; Fig. 43, g), just craniad
of which is the rounded depression known as the sella turcica
(Fig. 42, g; Fig. 43, h), for lodgment of the hypophysis. A
number of foramina pierce the floor of the cavity in this region.
Just ventrad of the cranial tip of the petrous portion of the
temporal is the small foramen lacerum (medius). Craniad
and laterad of this is a row of four foramina: the caudal one is
the foramen ovale (Fig. 42, i); then come in order the foramen
rotundum (j), the orbital fissure (k), and the optic foramen
(Fig. 42, l; Fig. 43, k). The two optic foramina are connected
by the shallow transverse chiasmatic groove (Fig. 42, m), for
the optic chiasma. Another small foramen continues caudad
from a groove on the floor of the orbital fissure; this opens on
the ventral surface of the sphenoid, between the wing and the
body of the bone. The groove and foramen constitute the
pterygoid canal, which transmits a nerve,—the nerve of the
pterygoid canal, or Vidian nerve.
The cranial cavity narrows at its cranial end to form the
small olfactory fossa (Fig. 43, III) which lodges the olfactory
bulbs. This is bounded by the frontals and the lamina cribrosa
(Fig. 42, o) of the ethmoid; caudad it opens directly into the
cerebral fossa. Numerous openings through the lamina
cribrosa for the olfactory fibres connect the olfactory fossa with
the nasal cavity. The roof of the fossa is marked by a prominent
median crest from the united edges of the frontals.
The nasal cavity is almost completely filled by the ethmoid
and vomer and the conchæ nasales. Its roof is formed by the
nasal bones and portions of the frontals; its sides by the
frontals, lachrymals, maxillaries, premaxillaries, and palatine[60]
bones; its floor by the horizontal plates of the palatines,
maxillaries, and premaxillaries.
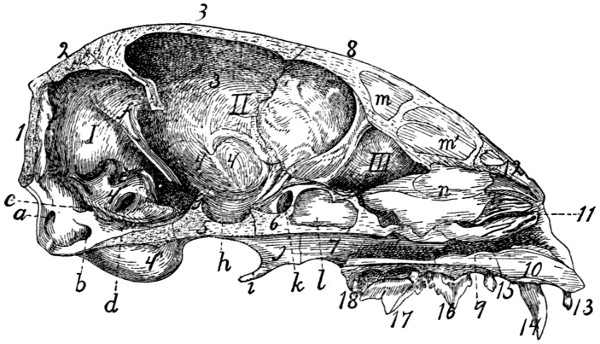
Fig. 43.—Skull, Median
Longitudinal Section, showing the Cavities.
I, cerebellar fossa; II, cerebral fossa; III, olfactory fossa. 1, occipital bone; 2,
interparietal; 3, 3′, parietal; 4, temporal (4, squamous portion; 4′, petrous portion;
4″, tympanic portion); 5, sphenoid; 6, presphenoid; 7, palatine; 8, frontal; 9, maxillary;
10, premaxillary; 11, ethmoid; 12, nasal; 13, incisor teeth; 14, canine;
15, 16, 17, premolars; 18, molar. a, condyloid canal; b, hypoglossal canal; c,
jugular foramen; d, internal auditory meatus; e, appendicular fossa; f, tentorium; g,
dorsum sellæ; h, sella turcica; i, hamular process; j, pterygoid process of sphenoid;
k, optic foramen; l, presphenoid sinus; m, m′, frontal sinus; n, lamina perpendicularis
of the ethmoid (broken at cranial edge).
The nasal cavity opens craniad by the large nares (Fig.
39, j; Fig. 42, r), which are bounded by the premaxillary and
nasal bones. In the natural condition this opening is divided
by a median cartilage which is continuous with the lamina perpendicularis
(Fig. 43, n) of the ethmoid, thus forming a partition
which divides the nasal cavity into two separate halves.
From the floor of the cranial part of the cavity rises a ridge
formed of the nasal crests of the maxillaries and premaxillaries,
and the cranial portion of the vomer. Farther caudad the
vomer spreads out in a horizontal plane and separates from the
floor of the cavity, so that the nasal cavity is thereby divided
by a horizontal partition into dorsal and ventral portions. The
ventral portion is small, forming the inferior meatus of the
nose; it ends caudally at the choanæ (posterior nares, Fig.
41, o) which lead into the nasopharynx. That portion
of the nasal cavity lying dorsad of the vomer is almost completely[61]
filled by the ethmoid and the conchæ nasales, superior
and inferior. It is bounded caudally by the lamina cribrosa of
the ethmoid (Fig. 42, o). The nasal cavity communicates with
the cranial cavity by the foramina for the olfactory fibres in the
lamina cribrosa; with the nasopharynx by the choanæ; with the
exterior of the body by the nares; with the mouth-cavity by
the foramina incisiva or anterior palatine foramina (Fig. 42, s);
with the orbit by the sphenopalatine foramen and the nasolachrymal
canal. It communicates directly also with the
frontal sinuses (Fig. 43, m, m′), the sphenoidal sinuses (Fig.
43, l), and with the cells of the labyrinths of the ethmoid.
JOINTS AND LIGAMENTS OF THE SKULL.
Sutures of the Skull.—The bones of the skull join each
other by means of immovable articulations known as sutures.
These sutures are designated by combining the names of the
bones between which they are situated: as, sphenofrontal suture
(sutura sphenofrontalis), between the sphenoid and frontal;
nasomaxillary suture (sutura nasomaxillaris), between the
nasal and maxillary bones. When a suture joins the two
corresponding bones of opposite sides the prefix inter is used,
as the intermaxillary suture (sutura intermaxillaris) between
the maxillaries. The sutures bounding the parietals have,
however, received special names not derived in this manner.
The suture caudad of the parietals, separating them from the
occipital and interparietal, is known as the lambdoidal suture;
that between the two parietals is the sagittal suture; that
separating the parietals and squamous portions of the temporals
is the squamous suture; that between the parietals and frontals
is the coronal suture. The suture separating the two frontals
also is known as the frontal suture, in place of interfrontal.
Articulations of the Mandible.—In man the two halves of
the mandible are united craniad, so as to form a single bone.
In the cat the two halves are separate, but articulate closely at
the symphysis menti by a thin interarticular cartilage.
The articulation of the mandible at the mandibular fossa of
the temporal is covered with a close articular capsule. The[62]
mandibular fossa is lined with cartilage. A slender ligament
passes from the angular process of the mandible caudad to the
external auditory meatus, being attached to the latter about
8 millimeters from its medial end. This is the stylomandibular
ligament.
V. BONES OF THE THORACIC EXTREMITIES.
Scapula
(Figs. 44 and 45).—The scapula may be described
as a flat triangular bone with one angle rounded. It lies
beneath the muscles on the lateral face of the thorax near its
cranial end. From its lateral surface there projects a flat ridge
(Fig. 44, g), the spine of the scapula. The ventral end of the
ridge is free as a curved process, the acromion process (Fig.
44, j).
I, fossa supraspinata; II,
fossa infraspinata; III, fossa subscapularis; IV, fossa
for teres major. a, vertebral border; b, coracoid border; c, glenoid border; d, glenoid
angle and fossa; e, coracovertebral angle; f, glenovertebral angle; g, spine; h,
tuberosity of the spine; i, metacromion; j, acromion; k, supraglenoidal tubercle;
l, incisura scapulæ; m, coracoid process; n, groove indicating portion of spine; o, o′,
ridges for attachment of muscle-fibres.
The ventral angle of the scapula (d), the glenoid angle[63]
(lateral angle of human anatomy), is much heavier than the
others and bears a concave, pear-shaped articular facet, the
glenoid fossa, for articulation with the humerus. The border
with which this angle is more nearly continuous may be called
the glenoid border (c) (axillary border of human anatomy).
Near the narrower cranial end of the glenoid fossa is a
small curved projection of the bone, the coracoid process (Fig.
45, m). The border upon which it lies is the coracoid border
(b) (superior border of human anatomy). The third border is
turned toward the vertebral column and is the vertebral
border (a).
The angle between the glenoid and vertebral borders is the
glenovertebral angle (f) (inferior angle of human anatomy),
and that between the coracoid and vertebral borders the
coracovertebral angle (e) (medial angle of human anatomy).
The medial or costal surface (Fig. 45) is smooth and nearly
flat. A shallow furrow (n) marks the position of the spine of
the scapula. Between the furrow and the coracoid border are
two oblique parallel ridges (o and o′) for the insertion of muscle-fibres.
Near the glenoid border is a well-marked ridge separating
the subscapular fossa (III), comprising the greater part
of the medial surface of the scapula, from the fossa in which
the teres major muscle has origin (IV). The surface presents
several nutrient foramina usually directed toward the glenoid
angle.
The lateral surface (dorsal surface of human anatomy)
(Fig. 44) is divided by the spine (g) into two portions. The
portion of the scapula craniad of the spine and the cranial
surface of the spine bound the supraspinous fossa (fossa supraspinata)
(I), while the surface caudad of the spine and the
caudal portion of the spine bound the infraspinous fossa (fossa
infraspinata) (II).
The spine (g) begins as a triangular elevated area in the
middle of the vertebral margin and runs toward the glenoid
angle. It rises gradually for about two-fifths of its length and
then the margin becomes broader and the spine remains of the
same height to its glenoid end. There is a rough thickening,
the tuberosity (h) of the spine, situated on its free border about[64]
midway between the tip of the acromion and the vertebral end
of the spine. The spine is inclined toward the glenoid margin
so as to form an angle of about 60 degrees with the caudal half
of the lateral surface.
At the base of the acromion process (j) the margin of the
spine presents a flat triangular projection, the metacromion (i),
directed toward the glenoid border.
The acromion (j) continues in the direction of the spine.
It is thicker than the spine, smooth and rounded on both its
surfaces and both its borders, and its apex is connected by
fibrous tissue to the clavicle.
The coracoid border (b) presents a slight rounded notch,
the incisura scapulæ or suprascapular notch (l), just dorsad of
the glenoid angle, and at its ventral end bears the coracoid
process (m) which is directed ventromediad.
The glenoid angle (d) is the only one requiring special
mention. Between the root of the coracoid process and
the glenoid cavity it presents a tubercle, the supraglenoidal
or bicipital tubercle (k), for the tendon of origin of the biceps
muscle. The glenoid angle is separated by a contracted neck
from the rest of the bone. Between this angle and the inner
margin of the acromion there is left a deep notch, the great
scapular notch.

Fig. 46.—Clavicle.
a, medial end; b, lateral
end.
Clavicle. Clavicula
(Fig. 46).—The clavicle in the cat is
greatly reduced. It is a slender curved rod
of bone imbedded in the muscles of the
shoulder and connected by fibrous tissue to
the apex of the acromion process. The
lateral end (b) is slightly enlarged.
Humerus
(Figs. 47 and 48).—The humerus forms the support
of the upper arm and articulates by its proximal end with
the scapula at the glenoid cavity, and by its distal end with the
radius and ulna, the bones of the lower arm. It is a nearly
cylindrical bone with enlarged ends, and is so curved that its
dorsal and ventral borders are hooked at the opposite ends so
that it has the form of an Italic f.
The proximal end of the bone bears on its dorsomedial
portion a thickening, the head of the humerus (a), which bears[65]
a smooth ovoid articular facet by which the bone articulates
with the glenoid cavity of the scapula. The head is not
separated from the body by a distinct anatomical neck as in the
human humerus.
a, head; b, greater tuberosity; c,
lesser tuberosity; d, bicipital groove; e, pectoral
ridge; f, deltoid ridge; g, rough area for insertion of latissimus dorsi and teres major;
h, nutrient foramen; i, capitulum; j, trochlea; l, coronoid fossa; m, radial fossa; n,
medial epicondyle; o, lateral epicondyle; q, supracondyloid foramen.
Along the lateral border of the proximal end of the shaft is
a high rough ridge semicircular in side view, the great
tuberosity (b). It gives attachment to muscles and is marked
on its dorsal border by a deep depression for the tendon of the
infraspinatus muscle. On the medial margin of the proximal
end closely associated with the head is a smaller elevation, the
lesser tuberosity (c), also for muscular attachment.
Between the greater and lesser tuberosities on the ventral[66]
surface is seen a broad groove, the sulcus intertubercularis or
bicipital groove (d), which passes distad onto the surface of
the shaft. In the natural state it is converted into a canal by
overlying tendons and lodges the tendon of the biceps muscle.
The shaft is nearly cylindrical at its middle, but its dorsoventral
diameter is slightly greater than its mediolateral
diameter. Its proximal end is flattened mediolaterad, while its
distal end is flattened dorsoventrad.
From the ventral margin of the greater tuberosity a ridge,
the pectoral ridge (e), is continued onto the surface of the shaft,
and from the dorsal margin another ridge, the deltoid ridge (f),
passes distad and ventrad so as to meet the pectoral ridge near
the middle of the ventral surface of the bone. On the medial
margin of the bone near the junction of the first and second
fourths is a roughened area (g) for the attachment of the tendons
of the latissimus dorsi and teres major muscles, and on the
same surface near the junction of the second and last thirds is
a nutrient foramen (h).
The distal end of the bone presents a smooth saddle-shaped
articular surface, which, in well-marked bones, is divided, when
seen from the ventral surface, by a slight nearly median ridge
into two unequal portions, lateral and medial (i and j). The
lateral half is rounded and is called the capitulum (i). It is
broader ventrad than dorsad, and is not continued onto the
dorsal surface of the bone. It is for articulation with the
proximal end of the radius.
The medial half of the surface, the trochlea (j), is concave
and passes directly into the capitular surface laterad, but is
limited mediad by a sharp ridge. It is continued onto the
dorsal surface of the bone, where it is limited also laterad by a
ridge. It articulates with the semilunar notch of the ulna.
Proximad of the trochlea the dorsal surface presents a deep
fossa, the olecranon fossa, which receives the olecranon of the
ulna when the arm is straightened. On the ventral surface
(Fig. 47) are two shallower fossæ separated by a longitudinal
ridge. The one over the trochlea receives the coronoid process
of the ulna when the arm is bent, and is called thence the
coronoid fossa (l). The one over the capitulum, the radial[67]
fossa (m), receives a triangular facet on the proximal end of
the radius at the same time. Between the radial and coronoid
fossæ on one side and the olecranon fossa on the other is only
a thin plate of bone. On the medial surface of the distal end
is a considerable roughened elevation, the medial epicondyle
(n) (epitrochlea). It gives origin to flexor muscles and to the
ulnar collateral ligaments of the elbow-joint. Opposite the
medial epicondyle over the capitulum is the lateral epicondyle
(o) for the origin of extensor muscles of the forearm and of the
radial collateral ligaments of the elbow-joint. From the lateral
epicondyle a ridge, the lateral supracondyloid ridge (p), continues
proximad, curving onto the dorsal surface of the bone
and ending about opposite the junction of the deltoid and
pectoral ridges.
Proximad of the medial epicondyle the bone is pierced near
its medial margin by an oblique oval foramen, the supracondyloid
foramen (q).
Radius
(I, Figs. 49 and 50).—In the usual position the
radius lies with its proximal end on the lateral side of the arm,
articulating with the capitulum of the humerus. The proximal
end is thus laterad of the proximal end of the ulna. Its distal
end, however, lies on the medial side of the distal end of the
ulna, so that the radius in the natural position crosses ventrad
of the ulna.
The radius is a curved bone slightly flattened dorsoventrally,
with enlarged ends. It may be described as consisting
of a shaft and of a proximal and a distal end. Its proximal
end presents on the ventral surface a tuberosity, the bicipital
tuberosity (c), for the insertion of the tendon of the biceps
muscle. Proximad of this the bone is contracted to form a
neck (b) which is surmounted by a head (a). The head has
on its proximal surface a depressed oval facet by which it
articulates with the capitulum, and on its ulnar border a long
narrow facet, the articular circumference (d), for articulation
with the radial notch of the ulna; also a triangular facet (e),
which fits into the radial fossa of the humerus.
The shaft is convex dorsad and concave ventrad. The
distal end is somewhat pyramidal. From its medial or radial[68]
side a wedge-shaped process, the styloid process (f), extends
distad. The distal surface of the end together with the lateral
surface of the styloid process form a concave articular cavity
(g) which fits against the scapholunar bone.
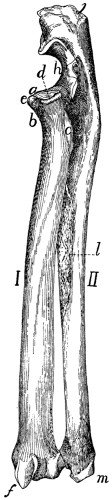
Fig. 49.—Radius
and Ulna, Dorsolateral View.

Fig. 50.—Radius
and Ulna, Ventromedial View.
I, radius; II, ulna. a,
head of radius; b, neck; c, bicipital tuberosity; d,
articular circumference; e, facet for radial fossa of humerus; f, styloid process of
radius; g, facet for articulation of scapholunar bone; h, semilunar (or great sigmoid)
notch of ulna; i, coronoid process; j, olecranon; k, area for insertion of brachialis
and clavobrachial muscles; l, rough area for attachment of interosseous membrane;
m, styloid process of ulna.
The dorsal surface of the distal end is marked by longitudinal
grooves for tendons, and its lateral or ulnar surface bears
a concave facet for articulation with the ulna.
Ulna
(II, Figs. 49 and 50).—The
ulna is a long slender[69]
bone, flattened mediolaterad. It is enlarged at its proximal
end and becomes gradually smaller toward the distal end.
The proximal end is marked ventrally by a deep excavation,
the semilunar notch, or great sigmoid cavity (h). By
the saddle-shaped articular surface of the semilunar notch it
articulates with the trochlea. This articular surface is divided
into two parts by a transverse non-articular area. The distal
boundary of the semilunar notch is a blunt process, the coronoid
process (i), which bears on its lateral surface a concave
facet, the radial notch, for the head of the radius.
The portion of the bone proximad of the semilunar notch is
called the olecranon (j). It fits into the olecranon fossa of the
humerus when the arm is straightened, and is rough at its end
for the insertion of tendons.
The body of the ulna becomes triangular distad. The
distal end is slightly larger than the shaft just proximad of it,
and bears on its radioventral side a hemispherical head for
articulation with the radius. Distad of the head the bone continues
as the flattened styloid process (m), which projects distad
from its dorsolateral side and is smooth on the medial side of
its apex, for articulation with the cuneiform bone of the wrist.
Carpus
(Fig. 51).—The carpus (wrist) consists of seven
bones arranged in two rows, three in the proximal row and
four in the distal row. Beginning on the medial side of the
hand (thumb or radial side), the first bone in the proximal row
is the scapholunar (a) (equal to the scaphoid or navicular and
lunar of the human hand). It articulates with the radius.
The next is the cuneiform (b), articulating with the styloid
process of the ulna, and the next, which is attached to the
cuneiform and projects freely ventrad, is the pisiform (c).
In the distal row the bone on the radial side is the trapezium
(d); the next is the trapezoid (e), the next the os magnum (f),
and the last the unciform (g). The distal row articulates with
the metacarpals or bones of the palm of the hand (1-5).
In the kitten the scapholunar is represented by three bones,
the scaphoid or navicular, on the radial side, the lunare, between
the scaphoid and the cuneiform, and a centrale, which
lies distad of the other two.
[70]
Scapholunar Bone. Os scapholunaris (Fig. 51, a).—The
scapholunar is a quadrangular bone with the ventroradial angle
produced into a blunt process. Its proximal surface is smooth
and articulates with the distal end of the radius. The distal
end is marked by oblique ridges and articulates with the unciform,
os magnum, trapezoid, and trapezium. The ulnar surface
articulates with the cuneiform, and the dorsal surface of the
ventroradial process with the radial sesamoid.
Cuneiform Bone. (Os triquetrum BNA) (Fig. 51, b).—The
cuneiform bone has the form of a flattened pyramid. Its
base articulates with the unciform, its
proximoulnar surface with the pisiform
except at its dorsal margin, where it
articulates with the styloid process of
the ulna. On its proximoradial surface
is a smooth facet for articulation
with the scapholunar.
Pisiform Bone. Os pisiforme (Fig. 51, c).—The pisiform bone is
about twice as long as broad, with
enlarged ends. Its dorsal end articulates
with the cuneiform, and on its
proximal surface, separated from the
above by a smooth ridge, is a smooth
facet for articulation with the styloid
process of the ulna.
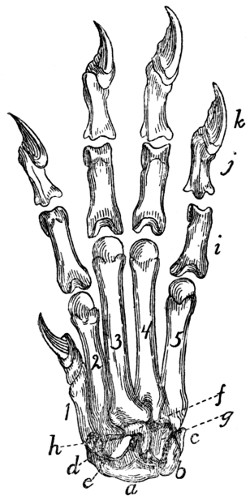
Fig. 51.—Carpus, Metacarpus,
and Phalanges,
Dorsal Surface.
a, scapholunar bone; b,
cuneiform; c, pisiform; d, trapezium;
e, trapezoid; f, os magnum;
g, unciform; h, radial
sesamoid; i, proximal phalanges;
j, second phalanges; k, distal
phalanges; 1, 2, 3, 4, 5, metacarpals
in order from the radial
side.
Unciform Bone. (Os hamatum
BNA) (Fig. 51, g).—The unciform is
a wedge-shaped bone with the apex
of the wedge directed proximad, and
smooth for articulation with the
scapholunar. By a part of its ulnar
surface it articulates with the cuneiform,
and by its radial surface with the
os magnum. Its distal end articulates
with the fourth and fifth metacarpals.
Os magnum. (Os capitatum
BNA) (Fig. 51, f).—The
os magnum may be described as an[71]
oblong plate bearing on its proximal surface a semicircular
ridge which crosses it diagonally. The proximal end of the
bone articulates with the scapholunar. Its distal end articulates
with the third metacarpal except near its ventroulnar angle,
where it articulates with the fourth metacarpal. Its ulnar surface
articulates with the unciform. Its radial border articulates
with the trapezoid, the third metacarpal, and, by two facets,
with the second metacarpal.
Trapezoid. (Os multangulum minus BNA.) (Fig. 51, e).—The
trapezoid is somewhat wedge-shaped, with the apex of the
wedge pointing ventrad. Its proximal side articulates with the
scapholunar, its distal side with the second metacarpal, its ulnar
side with the os magnum, and its radial side with the trapezium.
Trapezium. (Os multangulum majus BNA.) (Fig. 51, d).—The
trapezium has the form of a triangular prism curved into
a semicircle. The convex face looks proximad and articulates
by its ventral half with the scapholunar. The ulnar surface
articulates with the second metacarpal dorsally, and ventrally
with the trapezoid. Its radial surface articulates with the first
metacarpal.
Bones of the Hand or Manus
(Fig. 51, 1-5).—The Metacarpals.
Metacarpus.—The metacarpals are the five bones of
the palm of the hand; they are numbered from one to five,
beginning with the thumb. They are cylindrical elongated bones
with enlarged ends. The distal end is called the head, and the
proximal end the base. Each head bears a hemispherical
articular facet which is marked over its ventral half by a
prominent smooth ridge. The surface dorsad of the ridge
articulates with the proximal head of a phalanx. The ridge
and the surface at its sides are for a pair of sesamoid bones.
The first metacarpal (1) is the shortest. Its head is oblique,
and it articulates by the ulnar half of its proximal surface with
the trapezium (d); by the radial half with the radial sesamoid
(h).
The second metacarpal (2) is marked on the proximal part
of its dorsal surface by an oblique groove passing from the
radial side distad to the ulnar side. The base articulates with
the trapezoid (e). The ulnar surface of the proximal end[72]
articulates with the os magnum (f) and third metacarpal, while
the radial surface articulates with the trapezium (d).
The third metacarpal (3) is the longest, and its base is
rhomboidal with a projecting dorsoradial angle separated by a
groove from the rhomboid surface. The proximal end articulates
with the os magnum (f) and second metacarpal; the
radial surface of the proximal end with the second, and the
ulnar surface with the fourth, metacarpal.
The fourth metacarpal (4) has a base similar in form to that
of the third, and when placed in position with the fifth the two
form a hemispherical facet which articulates with the unciform
(g) and os magnum (f). The fourth metacarpal articulates by
its radial side with the third, and by its ulnar side with the fifth.
The fifth metacarpal (5) articulates by its proximal end
with the unciform (g), and by the radial side of its proximal
end with the fourth metacarpal.
Digits (Fig. 51).—The first digit of the hand is called the
pollex (thumb), the second the index, the third the medius,
the fourth the annularis, the fifth the minimus.
The first digit has two phalanges, each of the others three
phalanges.
Phalanges (Fig. 51, i, j, k).—The phalanges of the proximal
row (i) are elongated, flattened dorsoventrally and curved
so as to be longitudinally convex dorsad. All have thickened
ends. The proximal end is notched, and its proximal surface
looks dorsad and is concave for the head of the metacarpal.
The distal end is pulley-shaped, and the pulley surface extends
farther on to the ventral than on to the dorsal surface, and
serves for articulation with the middle phalanx.
The phalanges of the middle row (j) are like those of the
proximal row, but shorter. The proximal surface is triangular
and marked by a median facetted ridge. The whole surface is
smooth and adapted to the distal end of the phalanx of the first
row. The distal end is transversely elongated, so as to be
cylindrical, and projects more toward the ulnar than toward
the radial side. The distal phalanx (k) articulates with this
cylinder so that when it is fully extended it lies on the ulnar
side of the middle phalanx.
[73]
The distal phalanx (k) has the form of a quadrangular
prism. It is excavated on its proximal surface for articulation
with the middle phalanx. Its distal surface presents dorsad a
deep excavation from the bottom of which arises a compressed
plate of bone having the form of a bird’s beak. The depression
receives the base of a claw, and the beak-like projection supports
the claw.
Sesamoid Bones of the Hand. Ossa sesamoidea.—The hand
contains, in addition to those already described, eleven small
bones that are developed in tendons.
One of these, the radial sesamoid (Fig. 51, h), is closely
applied to the radial end of the scapholunar bone. It is
developed in the tendon of the extensor brevis pollicis muscle.
The other ten occur in pairs as small flattened curved bones
on the ventral side of the joint between each metacarpal and
the phalanx with which it articulates.
JOINTS AND LIGAMENTS OF THE THORACIC LIMBS.
The shoulder-joint is an arthrodial or ball-and-socket joint.
The bones entering into its formation are the scapula and the
humerus.
The capsular ligament or articular capsule is very ample
and allows for extended movement of the humerus. It is
attached to the edge of the glenoid fossa of the scapula and
passing distad covers the head of the humerus and is inserted
at the line of junction of the shaft and the epiphysis which forms
the head of the bone. On the lateral side of the ventral surface
of the humerus the attachment continues distad about two
centimeters along the lateral edge of the bicipital groove. On
the medial side the insertion passes over the proximal end of
the lesser tuberosity. A strong transverse band passes from
the greater tuberosity to the lesser tuberosity and bridges the
bicipital groove, converting it into a canal. The lateral and
medial parts of the capsule are strengthened by thicker bands
of fibres, the more prominent medial one of which passes from
the coracoid process of the scapula to the lesser tuberosity.
To the capsule are closely united parts of the supraspinatus,[74]
infraspinatus, coracobrachialis, and subscapularis muscles. A
synovial membrane lines the capsule within and forms a sheath
around the biceps tendon, so that the latter does not actually
enter the synovial capsule.
The elbow-joint (Figs. 52 and 53) is a ginglymus or
hinge-joint. The bones which enter into it are the humerus,
radius, and ulna.
The capsule of the joint forms a sac, with the following
attachments to the bones: (1) To the humerus it is attached
at the proximal edge of the coronoid and radial fossæ; to the
sides of the capitulum and trochlea distad of the two epicondyles,
and to the distal edge of the olecranon fossa. (2) To
the ulna it is attached at the edges of the radial and semilunar
notches; (3) to the radius around the articular facet, two or
three centimeters distad of the border. Many of the muscles
of this region are closely attached to the capsule.
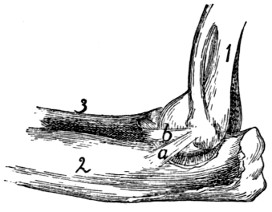
Fig. 52.—Ligaments of Elbow-joint,
Medial Side.
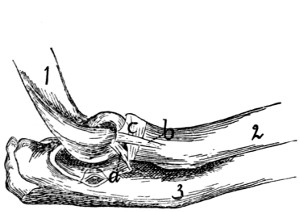
Fig. 53.—Elbow-joint, Lateral
View.
Fig. 52.—1, humerus; 2, ulna; 3, radius. a and b, the two medial collateral
ligaments.
Fig. 53.—1, humerus; 2, radius; 3, ulna. a, dorsal collateral ligament; b, ventral
collateral ligament; c, annular ligament.
Closely connected with the capsule of the joint are the
collateral ligaments. The two medial collateral ligaments
(Fig. 52) arise from the medial epicondyle. One (b) passes
distad and laterad to the interval between the radius and ulna;
here it divides, one branch going to the head of the radius,
while the other is attached to the lateral surface of the ulna at
the edge of the semilunar notch. The second medial ligament
(a) lies dorsad of the first; it passes to the medial surface of
the ulna, at the distal edge of the semilunar notch.
[75]
The two collateral ligaments on the lateral side (Fig. 53)
arise from the lateral epicondyle. The ventral one (b) passes
almost directly distad and is inserted into the lateral surface of
the proximal end of the radius about one centimeter from the
articular surface; its inner surface is partly united to the annular
ligament (c) of the radius. The dorsal one (a) is attached
to the lateral border of the semilunar notch of the ulna.
Articulations of Radius and Ulna.—The proximal radio-ulnar
articulation (Fig. 53) is by a pivot-joint or trochoid.
The two bones are held in place by the annular ligament (Fig.
53, c). This is attached on the lateral side to the dorsal border
of the radial notch of the ulna, passes around the head of the
radius, receiving some ligamentous fibres which come from the
lateral epicondyle, and is attached to the coronoid process of
the ulna. The annular ligament is closely united with the
capsule of the joint.
The radius and ulna are united for about their middle third
by the thin interosseous membrane, which fills the interosseous
space between their adjacent edges.
The Wrist.—At the wrist or carpus there are in reality
three joints, the first between the radius and ulna proximad
and the first row of carpals distad, the second between the two
rows of carpal bones, the third between the distal row of
carpals and the metacarpals. The first two are movable joints;
the third is not. Each of these three joints has a capsule, and
the bones entering into the joints are interconnected by numerous
ligaments. These ligaments are named by combining the
names of the two bones which they interconnect. Ligaments
which interconnect bones of the same row in the carpus are
sometimes distinguished as interosseous ligaments, as contrasted
with intercarpal ligaments, which connect together
bones of different rows. According to their position the ligaments
may also be distinguished as dorsal, ventral, and lateral.
Detailed descriptions and figures of all these ligaments are
given by Strauss-Durckheim.
Metacarpals.—The joint between the carpals and metacarpals
has been described. At the distal end of the metacarpals
the articulations with the phalanges have each a[76]
capsule. The joint is further strengthened by a double lateral
ligament on each side. The two sesamoid bones at each joint
are interconnected by a strong transverse ligament, and each
is connected with the head of the metacarpal and the base of
the first phalanx by a lateral ligament.
Phalanges.—Between the phalanges the joints possess capsules,
and each has a radial and an ulnar lateral ligament.
VI. BONES OF THE PELVIC EXTREMITIES.
Innominate Bones. Ossa innominata. (Os coxæ BNA)
(Figs. 54 and 55).—The two innominate bones articulate
with the sacrum and extend thence caudoventrad and finally
turn mediad and unite in the middle line, forming the symphysis
pubis. They thus form an arch, the pelvic arch, pelvic
girdle or pelvis, which is closed dorsad by
the sacrum.
In the middle of the lateral surface of
each bone is a hemispherical depression, the
acetabulum (Fig. 55, d), which receives the
head of the femur.
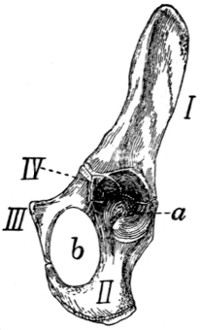
Fig. 54.—Innominate
Bone of Kitten,
Ventrolateral
View.
I, ilium; II, ischium;
III, pubis; IV, acetabular
bone. a, acetabulum;
b, obturator foramen.
In the kitten each innominate bone is
composed of three principal parts united by
sutures (Fig. 54). From the sacrum to the
acetabulum is a single bar, the ilium (I).
Caudad of the acetabulum are two bars.
The dorsal one of these is the ischium (II),
and the ventral one is the pubis (III). The
ischium enters into the formation of the
acetabulum (a), but the pubis does not.
The two bones, however, are in contact at
the ventral edge of the acetabulum. From
this point they diverge, but unite with one another again near
the middle line and thus enclose an oval foramen, the obturator
foramen (b). Wedged between the ilium, ischium, and
pubis at their point of junction and helping to form the acetabulum
is a small irregular bone, the acetabular bone (IV).
In the adult cat these four parts are united into a single bone
which is nevertheless usually described, ignoring the acetabular[77]
piece, as made up of ilium (Fig. 55, I), ischium (II), and
pubis (III).
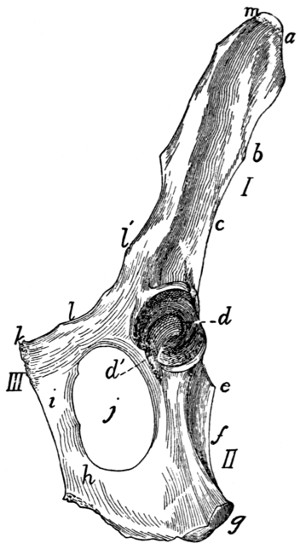
Fig. 55.—Innominate Bone of
Adult Cat, Ventrolateral
View.
I, ilium; II, ischium; III, pubis.
a, crest of the ilium; b, posterior inferior
spine; c, great sciatic notch; d,
acetabulum; d′, incisura acetabuli; e,
spine of the ischium; f, lesser sciatic
notch; g, tuberosity of the ischium;
h, ramus of the ischium; i, ramus of
the pubis; j, obturator foramen; k,
pubic tubercle; l, iliopectineal line;
l′, ilio-pectineal eminence; m, anterior
superior process.
The ilium (I) is somewhat contracted at the middle and
broader at its ends. One end enters into the acetabulum
(Fig. 55, d) and forms about
one-fifth the articular surface.
This end is also the thickest part
of the bone. The lateral surface
of the ilium is concave for the
attachment of muscles. The medial
surface is smooth over its
acetabular half and rough over its
sacral half. The rough portion is
marked at its junction with the
smooth portion by the ear-shaped
auricular impression by which
the bone articulates directly with
the sacrum. The caudal half of
that part of the medial surface
craniad of the auricular impression
gives attachment to the ilio-sacral
ligaments which bind the ilium to
the sacrum. The dorsal border
is straight at its cranial end and
concave and rounded at its caudal
end. Between the two portions
and at the dorsal edge of the
auricular surface is a protuberance
corresponding to the posterior
inferior spine (Fig. 55, b) of the
human ilium. The concavity of
the dorsal border (c) corresponds
to the great sciatic notch of the
human ilium. At its caudal end is the short spine of the
ischium (e), which is not a part of the ilium. The ventral border
of the ilium is broad caudad, becoming narrower craniad. The
lateral margin of the ventral border is continued to a tuberosity
at the edge of the acetabulum; its medial margin is called the[78]
iliopectineal line (l) and extends on the pubis to the symphysis.
An eminence, the iliopectineal eminence (l′), on the iliopectineal
line, lies opposite to the acetabulum at about the junction
of the ilium and pubis. The cranial end of the bone is thickened,
forming the crest (a) of the ilium. At the junction of
the crest with the ventral border is a projection, the anterior
superior process (m) of human anatomy.
The pubis (III) (including the acetabular bone) enters into
the formation of the acetabulum (d) constituting about one-sixth
the circumference, but less than one-sixth its area. It
may be described as a flat, curved bone, contracted at the
middle and expanded at the ends. The dorsal end enters
into the acetabulum; the ventral end unites with the opposite
bone at the symphysis pubis and sends caudad a projection, the
ramus (i) of the pubis, which unites with the ramus of the
opposite bone to form about two-thirds of the entire symphysis.
At the sides of the symphysis a slightly marked angle projects
craniad from each of the pubic bones; these two together constitute
the pubic tubercle (k), for the origin of the rectus
abdominis muscle. The surfaces of the ramus are smooth.
One of its borders is concave and enters into the formation of
the obturator foramen (j). Another of its borders is the iliopectineal
line (l). Its third border is rough for the symphysis.
The ischium (II) has the form of a triangular prism contracted
at the middle. Its cranial end forms nearly two-thirds
of the acetabulum. Its caudal end bears dorsad a rough
thickening, the tuberosity of the ischium (g). From the
caudal half of the ventral border of the bone a sickle-shaped
process, the ramus (h) of the ischium, curves medioventrad
and then craniad and joins the ramus of the pubis. Its medial
border is rough and enters into the symphysis, forming the
caudal one-third. The lateral angle of the bone is rounded.
Its dorsal angle is marked near the cranial end by the spine (e)
of the ischium. The concavity between this spine and the
tuberosity corresponds to the lesser sciatic notch (f) of human
anatomy.
The acetabulum (d) is cup-shaped. The ventral one-sixth
of its border is deficient and a broad groove extends from the[79]
deficiency to the bottom of the cup. The deficiency, incisura
acetabuli, or acetabular notch (d′), is closed naturally by a
ligament, and the groove gives origin to the ligament (ligamentum
teres) which attaches the head of the femur.
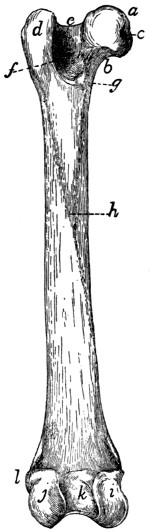
Fig. 56.—Left Femur,
Ventral Side.
a, head; b, neck; c, depression
for round ligament;
d, great trochanter;
e, trochanteric fossa; f,
intertrochanteric line; g,
lesser trochanter; h, linea
aspera; i, medial condyle;
j, lateral condyle; k, intercondyloid
fossa; l, lateral
epicondyle.
Femur
(Fig. 56).—The femur is the
proximal bone of the posterior extremity.
It consists of enlarged proximal and
distal ends connected by a nearly cylindrical
shaft. The proximal end presents
on its medial side a hemispherical head
(a) which fits into the acetabulum. It is
supported by a neck (b) which is contracted
near the head and expanded dorsoventrally
where it joins the remainder of
the bone. The medial surface of the head
presents near its ventral border a depression
(c) for the insertion of the round ligament
of the femur. Ventrad the articular
surface of the head extends as an acute
projection onto the shaft, so that the whole
articular surface appears somewhat pear-shaped.
On the lateral side of the proximal
end opposite the head is a projecting
mass, the great trochanter (d), forming
the end of the shaft. On the medial side
of the great trochanter at its junction with
the neck is a deep fossa, the trochanteric
fossa or digital fossa (e) for the insertion
of muscles. From the ventral surface of
the great trochanter a ridge, the intertrochanteric
line (f), is continued distad,
ending in a pyramidal projection, the
lesser trochanter (g), which serves for
the insertion of muscles. A second ridge
is continued to the lesser trochanter from
the neck. A slight but well-marked
ridge, the spiral ridge or line, runs round two sides of the
neck parallel to the second ridge.
[80]
The shaft is nearly straight and cylindrical. A rough line
is continued along its ventral surface from the lesser trochanter,
and a similar line along its lateral surface from the greater
trochanter; these unite ventrad to form the linea aspera (h).
On its ventral surface is a nutrient foramen, directed proximad.
The shaft gradually widens distad and ends in two condyles
(i and j) which are continuous dorsad but separated ventrad
by a deep notch, the intercondyloid fossa (k). The distal
surface of the shaft and condyles is articular. This articular
surface is larger on the lateral condyle (j). The part of the
articular surface on the end of the shaft (patellar surface) is
for the patella; that part of it on the condyles and separated
by the notch is for the tibia.
On the lateral surface of the lateral condyle is a slight
prominence, the lateral epicondyle (l), and on the medial surface
of the medial condyle is another prominence, the medial
epicondyle; both are for the attachment of ligaments.
Patella
(Fig. 1, r).—The patella is a small flat bone with
a pear-shaped outline, having its apex distad. It lies against
the articular surface at the lower end of the shaft of the femur.
It thus covers the knee-joint. The inner surface is smooth and
convex from side to side, but concave in a proximodistal line.
It fits against the lower end of the femur. Its outer surface is
rough and concave. It is a sesamoid bone inserted in the
tendon of the quadriceps femoris muscle.
Three other sesamoid bones are found in the region of the
knee (see Fig. 61, p. 89). Two are in the tendons of the
gastrocnemius muscle, proximad of the two condyles of the
femur. The third is in the tendon of the popliteus muscle,
just laterad of the lateral condyle of the femur.
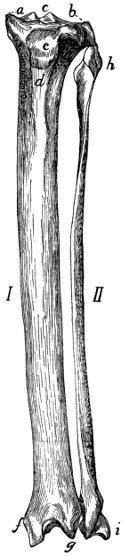
Fig. 57.—Tibia and
Fibula of Left
Leg, Dorsal View.
I, tibia; II, fibula.
a, medial condyle of
the tibia; b, lateral condyle;
c, spine of the
tibia; d, crest of the
tibia; e, tubercle for attachment
of the patellar
ligament; f, medial
malleolus; g, projection
of dorsal surface of the
tibia; h, head of fibula;
i, lateral malleolus.
Tibia
(I, Fig. 57).—The tibia is the longer of the two
bones of the leg between the knee and the ankle, and is the
longest bone of the body. It has a triangular shaft and
enlarged proximal and distal ends.
The proximal end is curved ventrad and projects into two
prominences, the tuberosities, on either side. Each tuberosity
bears on its proximal end an articular facet for the condyles of
the femur; these are known respectively as the lateral and[81]
medial condyles (a and b) of the tibia. The
condyles are oval, convex dorsoventrad, and
concave from side to side. The two condyles
are separated at the middle of their
contiguous margins by a bicuspid projection,
the spine of the tibia (c). They are continuous
dorsad, but separated ventrad by a
deep notch between the tuberosities, the
popliteal notch. On the distal side of the
lateral condyle is an elongated facet for the
proximal end of the fibula.
The shaft is triangular, smallest at about
its middle and enlarged at both ends. It
presents dorsal, medial, and lateral borders,
and medial, lateral, and ventral surfaces.
The lateral surface is concave proximad.
The medial surface is convex. The two
are continuous at the distal end. At their
proximal ends the border separating them
is raised into a prominent ridge, the crest
(d) of the tibia, the proximal end of which
contributes to increase the proximal surface
of the bone, and presents an oblong
tubercle (e) for the insertion of the ligamentum
patellæ (ligament of the quadriceps
femoris muscle). The ventral surface
is concave proximad where it abuts upon the
tuberosities. Its proximal half is crossed by
two rough parallel lines, the distal one of
which crosses in a spiral course from the
lateral to the medial border; near its beginning
is a nutrient foramen.
The distal end extends farther distad on
its medial side. The extension is the medial
malleolus (f). The malleolus presents two
grooves on its medial surface for the tendons
of muscles. On the lateral side of the distal
end is an oblique triangular facet for the distal end of the[82]
fibula. The ventral surface of the distal end presents an oblique
border which passes from the apex of the malleolus
proximolaterad. The dorsal surface extends into a V-shaped
projection (g) between the malleolus and the fibular facet.
The distal end presents an oblique ridge running from the
apex of the V-shaped extension of the dorsal surface to near
the base of the medial malleolus. The ridge and the concavities
on either side of it, the medial one of which is deeper,
fit against the proximal trochlear surface of the astragalus.
Fibula
(II, Fig. 57).—The fibula lies at the lateral side
of the tibia in the shank. It is a slender triangular bone with
enlarged proximal and distal ends.
The proximal end or head (h) is flattened. It bears a facet
on its proximomedial surface for articulation with the tibia,
and is longitudinally grooved on the outer surface.
The shaft has a very sharp medial border. This border is
turned toward the tibia and gives attachment to the interosseous
membrane, which runs between the tibia and fibula.
The distal end is expanded to form the lateral malleolus (i).
This bears a facet on the proximal portion of its medial surface
near its dorsal margin, for the tibia, and distad of this is a
second facet for the astragalus. The ventral and lateral surfaces
are grooved for tendons.
Tarsus
(Fig. 58).—The tarsus consists of seven bones.
The longest of these, lying on the lateral side of the foot and
forming the support of the heel, is the calcaneus or os calcis (a).
It articulates distad with a bone, the cuboid (c), which bears
the fourth and fifth metatarsals. Lying between the calcaneus
and the tibia is the astragalus or talus (b), the distal end of
which articulates with the boat-shaped navicular or scaphoid
(d). The scaphoid bears on its distal surface the three cuneiform
bones, lateral (e), medial (g), and intermediate, bearing
the rudiment of the first metatarsal and the second and third
metatarsals.
Astragalus. (Talus BNA) (Fig. 58, b).—The astragalus
may be divided into body, neck, and head. The body is
marked on its proximal surface by a deep pulley-like groove
for the articulation with the distal end of the tibia, and on its[83]
lateral and medial surfaces by curved
facets for articulation with the malleoli
of the tibia and fibula. This entire
surface for articulation with the bones
of the leg is known as the trochlea.
The lower surface is marked by two
facets separated by a groove; these are
for articulation with corresponding
facets on the calcaneus. Distally the
bone contracts to form the neck and
enlarges at the end, forming the head,
which is smooth on its distal surface for
articulation with the navicular or scaphoid.
Calcaneus (Fig. 58, a, and Fig. 59).—The
calcaneus (os calcis) is the largest
bone of the foot and forms the heel.
It is two or three times as long as broad
and has six surfaces: dorsal, ventral,
medial, lateral, proximal, and distal.
The proximal one-half of the dorsal
surface (Fig. 59) is smooth, while the
distal half is broadened and bears two
facets which are separated by a groove.
These articulate with the corresponding
facets on the astragalus. The medial[84]
facet is borne on a projection of the bone, the sustentaculum
tali (c). Distad of the facets the surface is rough. The ventral
surface is smooth. The proximal end (b) is grooved for the
tendon of Achilles. The lateral surface is smooth and marked
by a grooved tubercle, the peroneal tubercle (d), near the distal
end. The medial surface is marked by part of the articular
facet for the astragalus, and also by the grooved sustentaculum
tali. The distal end (a) articulates with the cuboid.
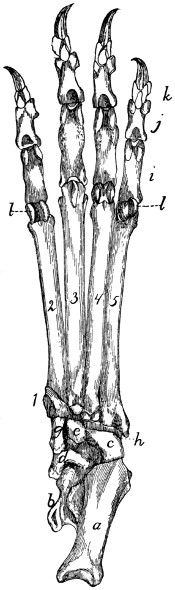
Fig. 58.—Tarsus, Metatarsus,
and Phalanges
of Left Foot, Ventral
View.
a, calcaneus; b, astragalus;
c, cuboid; d, scaphoid; e, lateral
cuneiform; g, medial
cuneiform; h, peroneal groove,
for the tendon of the peroneus
longus muscle; i, proximal
phalanges; j, second row of
phalanges; k, distal phalanges;
l, sesamoid bones. 1, rudimentary
first (medial) metatarsal;
2, 3, 4, 5, the other
metatarsals.
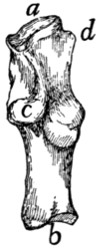
Fig. 59.—Calcaneus of Right
Foot, Dorsal View.
a, distal facet for cuboid; b,
proximal end with groove for
tendon of Achilles; c, sustentaculum
tali; d, peroneal tubercle.
Cuboid. Os cuboideum (Fig. 58, c).—The cuboid has
somewhat the form of a cube and articulates by its proximal
end with the calcaneus (a), and by its distal end with the fourth
and fifth metatarsals. Its medial surface articulates with the
scaphoid (d) and lateral cuneiform (e). The ventral surface is
marked near its distal end by an oblique ridge, distad of which
is a deep groove, the peroneal groove (h), for the tendon of
the peroneus longus muscle.
Scaphoid. (Os naviculare pedis BNA) (Fig. 58, d).—The
scaphoid is a boat-shaped bone. Its proximal surface is
marked by a concave facet for the head of the astragalus (b),
and its distal surface has three facets for the lateral (e), intermediate
and medial (g) cuneiform bones. At the junction of
the ventral with the medial surface is a prominent tubercle.
The lateral surface bears two linear facets for articulation with
the calcaneus (a) and cuboid (c).
Lateral Cuneiform. Ectocuneiform. Os cuneiforme tertium
BNA (Fig. 58, e).—The lateral cuneiform is a wedge-shaped
bone with a hooked process extending from the ventral
sharp angle of the bone. It articulates by its proximal end
with the scaphoid (d), and by its distal end with the third
metatarsal. The medial surface bears near its distal end two
facets for the second metatarsal, and on its proximal end a
facet for the intermediate cuneiform. The caudal surface has
a facet on its proximal end for the cuboid (c).
Intermediate Cuneiform. Mesocuneiform. Os cuneiforme
secundum BNA.—The intermediate cuneiform is small and
wedge-shaped, with the base of the wedge dorsad. It lies
between the lateral cuneiform and the medial cuneiform, articulates
by its proximal end with the middle facet of the scaphoid,[85]
and bears on its distal end the second metatarsal. It is not
visible in ventral view.
Medial Cuneiform. Entocuneiform. Os cuneiforme primum
BNA (Fig. 58, g).—The medial cuneiform lies on the medial
side of the foot. It is a flat triangular bone about twice as long
as broad, and broader at its proximal end than at the distal
end. It bears on its distal end the rudimentary first metatarsal.
The proximal end is oblique and bears a concave facet
for the lateral distal facet of the scaphoid (d). The lateral
surface has a concave facet at its proximal end for the intermediate
cuneiform, while the distal portion is applied against
the medial surface of the second metatarsal.
Bones of the Foot or Pes
(Fig. 58).—Metatarsals. Metatarsus
(Fig. 58, 1-5).—The metatarsals are five in number.
They bear a close resemblance to the metacarpals, but they
may be distinguished by their bases.
The first metacarpal (1) is rudimentary and conical. Its
base has a facet for the distal end of the medial cuneiform (g),
while the outer surface fits into a depression on the inner surface
of the base of the second metatarsal.
The second (2). The proximal surface of the base is triangular,
corresponding to the distal end of the intermediate
cuneiform. The medial surface is marked by two concavities,
one along the proximal border for the distal end of the medial
cuneiform, and one distad of this for the first metatarsal. The
lateral surface bears on the proximal margin an oblique triangular
facet dorsad and a similar facet ventrad, both for the
lateral cuneiform (e). Distad of these facets is a rough ridge.
The third (3). The proximal end of its base is a triangular
facet with the apex directed ventrad and the sides excavated.
It is for the distal end of the lateral cuneiform (e). Its medial
surface presents a depression which receives the ridge of the
second metatarsal. On the lateral surface a short distance
distad of the proximal border is a triangular concave facet, and
near the proximal border ventrad a second concave facet.
Both are for the fourth metatarsal.
The fourth (4). The proximal end is convex, notched
medially and facetted for the cuboid (c). Its medial surface[86]
bears dorsad, a short distance from its proximal end, a smooth
tubercle, and ventrad a small convex facet. Both articulate
with facets on the lateral surface of the third metatarsal. The
lateral surface has a sinuous facet along its dorsal border, and
ventrad of this a depression. There is a second facet along
the ventral border. Both facets are for the fifth metatarsal,
and the depression is for ligaments.
The fifth (5) has its base flattened and expanded so as to
be wedge-shaped, with the apex of the wedge directed proximad.
Its dorsal end extends into a tubercle. It thus presents
only lateral and medial surfaces. The medial surface shows
two tubercles, one distad of the other. The distal tubercle
and the distal half of the proximal tubercle are facetted and fit
into the sinuous facet on the fourth metatarsal. A narrow
facet on the ventral border of the surface articulates with the
facet on the ventral border of the lateral surface of the fourth
metatarsal. The proximal half of the distal tubercle is facetted
for the cuboid (c). The lateral surface is smooth, non-articular,
and obliquely grooved.
Phalanges (Fig. 58, i, j, k).—There are three phalanges
in each of the four digits, and these are almost identical with
those described for the manus.
Sesamoid Bones. Ossa sesamoidea (Fig. 58, l).—The
sesamoid bones are found at the joints between the metatarsals
and phalanges, and are in all respects like those of the manus.
JOINTS AND LIGAMENTS OF THE PELVIC LIMBS.
Ligaments of the Pelvis.—The ilium and sacrum are articulated
at the auricular facet of the ilium and the corresponding
rough surface of the sacrum. The joint is an amphiarthrosis, permitting
very little movement. A capsular ligament surrounds
the articular surface, being attached to the bones about its circumference;
it is short and strong. Craniad of the capsule is a thick
very short ligament, composed of very strong transverse fibres
passing from the rough surface of the sacrum to the corresponding
rough surface of the ilium. This forms the lateral iliosacral
ligament, which is united at its caudal border to the capsule.
A strong, wide ligamentous band passes from the dorsal[87]
border of the ilium to the sides of the sacrum. This is indistinctly
subdivided into several bands, which together represent
the long and short posterior iliosacral ligaments of man.
Symphysis pelvis.—The medial borders of the pubis and
ilium meet in the middle line ventrad of the pelvis and are here
united by cartilage. The joint is strengthened by numerous
small bands which pass across the line of junction from one side
to the other; these occur on both surfaces.
The Hip-joint.—The hip-joint is an enarthrosis, or ball-and-socket
joint in which more than half the spherical head of
the femur is received into the acetabulum. The depth of the
acetabulum is increased by a rim of fibrocartilage about its
margin, forming the labrum glenoidale. This passes across
the acetabular notch, forming the transverse ligament of the
acetabulum; beneath it blood-vessels and nerves pass into the
acetabular cavity.
The capsule of the joint is large and loose. It is attached
about the margin of the acetabulum, and passes over the head
of the femur, to be attached to the bone several millimeters
distad of the head. It thus encloses both the head and the
neck of the femur.
The ligamentum teres, or round ligament, is a very strong,
short ligament which passes from the depression in the head of
the femur to the bottom of the acetabulum.
The Knee-joint (Figs. 60 and 61).—The joint between the
femur and the tibia is very complex. The surfaces of the condyles
of the femur do not correspond to those of the condyles
of the tibia. Between the ends of the two bones are placed
two disks of cartilage, the menisci, or semilunar cartilages
(Fig. 60, c and d; Fig. 61, a and b), of such a form that the
congruity of articular surfaces is restored. Each meniscus has
a proximal surface corresponding to the form of one of the
condyles of the femur, and a distal surface corresponding to a
condyle of the tibia. The menisci are held in position by ligaments.
The knee-joint permits not only backward and forward
movement, but also a small amount of rotary motion.
The joint has two capsules, one on the dorsal (convex)
side, the other on the ventral side. The two communicate[88]
only by a small passageway lying within the joint between the
ends of the bones. The dorsal one is attached to the femur
several millimeters proximad of the patellar surface and some
distance on each side of the latter. The patella is imbedded
in its outer wall, and it is attached to the tibia on the edges of
the articular surface of the latter, from the crest to the tuberosities.
The capsule is also attached laterally and medially to
the sides of the menisci, and is closely united to the patellar
ligaments. Its cavity contains a mass of yellow fat.
The ventral capsule is attached to the borders of the articular
surfaces of the femur and tibia on their ventral sides, to the
menisci, and to the epicondyles of the femur and the tuberosities
of the tibia. Its walls are stronger and its cavity smaller
than those of the dorsal capsule.
The ligaments of the knee-joint (Figs. 60 and 61), aside
from the capsules, may be classified into: (1) those which are
connected with the patella; (2) collateral ligaments (Fig. 60,
i and j), which pass from the epicondyles directly distad along
the sides of the joint to the tibia or fibula; (3) crucial ligaments
(Fig. 60, g and h; Fig. 61, c and d), which cross within the joint
from one side of the femur to the opposite side of the tibia; (4)
ligaments which hold the menisci in place (Fig. 60, e and f;
Fig. 61, e and f).
(1) Ligaments of the Patella.—The patella is imbedded
in the dorsal wall of the dorsal capsule of the joint.
From its distal end a strong tendon or ligament passes distad
to the crest of the tibia. This is known as the ligamentum
patellæ: it may be considered a part of the tendon of M. quadriceps
femoris. On the lateral side the capsule of the joint is
strengthened by the transverse fibres of the tendon of M. plantaris,
which aid in holding the patella in place.
(2) Collateral Ligaments.—Of these there are two.
The ligamentum collaterale fibulare (Fig. 60, j) is attached to
the lateral epicondyle of the femur and passes distad across
the tendon of the plantaris muscle to the head of the fibula.
Dorsad of the fibular ligament and parallel with it passes the
tendon of origin of the extensor longus digitorum. The ligamentum
collaterale tibiale (Fig. 60, i; Fig.
61, g) begins on[89]
the medial epicondyle of the femur and passes distad to the lateral
tuberosity of the tibia; part of it passes one to one and a half
centimeters distad of the tuberosity to be attached to a rough
ridge on the side of the tibia.
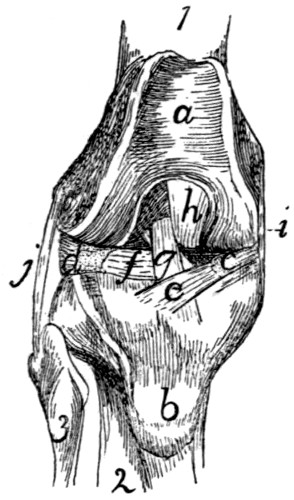
Fig. 60.—Knee-joint, from the
Dorsal or Patellar Side.
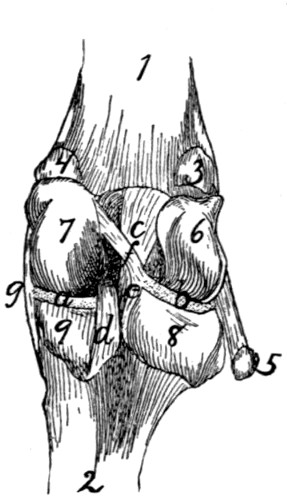
Fig. 61.—Knee-joint, from the
Ventral or Flexor Side.
Fig. 60.—The patella has been removed. 1, femur; 2, tibia; 3, fibula. a,
patellar surface of femur; b, tubercle for attachment of ligamentum patellæ; c, medial
meniscus; d, lateral meniscus; e, f, ligaments of the menisci; g, ligamentum cruciatum
anterius; h, ligamentum cruciatum posterius; i, ligamentum collaterale tibiale;
j, ligamentum collaterale fibulare.
Fig. 61.—The capsule of the joint has been opened. 1, femur; 2, tibia (fibula
not shown); 3, sesamoid bone in lateral head of M. gastrocnemius; 4, sesamoid in
medial head of M. gastrocnemius; 5, tendon of M. popliteus, with sesamoid bone;
6, 7, lateral and medial condyles of the femur, with the intercondyloid notch between
them; 8, 9, lateral and medial condyles of the tibia, with the popliteal notch between
them; a, b, medial and lateral menisci; c, ligamentum cruciatum anterius; d, ligamentum
cruciatum posterius; e, f, ligaments of the lateral meniscus; g, ligamentum
collaterale tibiale.
(3) Crucial Ligaments.—There are two of these also.
The ligamentum cruciatum anterius, or anterior crucial ligament
(Fig. 60, g; Fig. 61, c), is a thick, strong ligament which
begins on the dorsal part of the proximal end of the tibia nearer
the medial side (Fig. 60, g), and passes ventrad and proximad,
between the ends of the two bones forming the joint, into
the intercondyloid fossa of the femur, and becomes attached to
the medial surface of the lateral condyle of the femur (Fig.
61, c). It is composed of two partially separated bands, forming
a slight angle with one another. It is crossed near its
dorsal and ventral ends by two of the ligaments of the menisci.
The ligamentum cruciatum posterius, or posterior crucial ligament[90]
(Fig. 60, h; Fig. 61, d), begins on the tibia at the edge
of the popliteal notch (Fig. 61, d), nearer the medial side, and
passes dorsad and proximad to be attached to the ventral edge
of the patellar surface of the femur, in the intercondyloid fossa
(Fig. 60, h).
(4) Ligaments of the Menisci.—There are five of
these, connecting the menisci with the femur or tibia. One
(Fig. 60, e) passes from the dorsal edge of the medial meniscus
(c) transversely across the anterior crucial ligament (g) to
the proximal end of the tibia nearer the lateral side. A second
(Fig. 60, f) passes from the dorsal margin of the lateral
meniscus (d) transversely beneath the anterior crucial ligament
(g) to the proximal end of the tibia nearer the ventral side and
medial border. A third (Fig. 61, f) passes from the ventral
margin of the lateral meniscus obliquely across the anterior
crucial ligament (c) to the lateral side of the medial condyle of
the femur. The fourth (Fig. 61, e) is small, passing from the
medial angle of the ventral border of the lateral meniscus distad
to the popliteal notch. The fifth passes from the ventral border
of the medial meniscus laterad beneath the posterior crucial
ligament to the proximal end of the tibia, nearer the ventral
and lateral sides.
Articulations between the Tibia and Fibula.—At the
proximal end the fibula is as it were suspended from the distal
side of the overhanging lateral tuberosity of the tibia by strong
ligamentous tissue. The capsule of the joint is formed by an
extension of the capsule of the knee-joint which passes between
the tibia and fibula on the ventral side. Forming the dorsolateral
wall of this extension is a strong, thick ligament which
passes directly from the head of the fibula to the lateral surface
of the lateral tuberosity of the tibia. A second more delicate
ligament passes from the head of the fibula dorsoproximad to
the tubercle laterad of the crest of the tibia, bridging a groove
through which passes the tendon of M. extensor longus digitorum.
The tibia and fibula are connected throughout their length
by an interosseus membrane. This is broad and very thin in
its proximal part, narrower and thicker distad.
[91]
Distad the two malleoli forming the ends of the fibula and
tibia are closely and immovably united. The capsule of the
joint is here an extension proximad of the capsule of the articulation
with the astragalus. On the dorsal side a short broad
band of strong fibres passes from the surface of the tibia
obliquely laterodistad to the border of the fibula; this forms the
anterior ligament of the lateral malleolus. On the ventral
side a very much weaker set of fibres forms the ventral wall of
the articular capsule; it is called the posterior ligament of the
lateral malleolus. Tibia and fibula are also connected on the
dorsal side by the ligamentum transversum cruris, or transverse
ligament of the lower leg (Fig. 91, 5), which spans the
tendons of Mm. extensor longus digitorum and tibialis anterior.
From the middle of the distal margin of this a slender supporting
ligament passes distad and is inserted on the dorsal
surface of the tarsus. The grooves in the two malleoli for the
passage of the tendons are spanned by ligamentous fibres
(retinacula) for holding the tendons in place.
Articulation between the Leg and the Foot.—At the
distal end there is formed between the two malleoli a deep
irregular fossa, into which is received the trochlea of the
astragalus. The joint is covered by a large articular capsule,
which passes also, as above noted, between the tibia and fibula.
In addition to the capsule the following ligaments may be distinguished:
(a) On the lateral side, (1) a short ligament from
the fibula to the astragalus, directed toward the proximal end
of the foot; (2) a ligament from the fibula to the calcaneus,
attached to the latter proximad of the peroneal tubercle;
(3) a stronger ligament from the fibula to the calcaneus, lying
beneath the last-mentioned and directed toward the proximal
end of the bone. (b) On the medial side may be distinguished
(1) a strong ligament from the tibia (medial malleolus) to the
sustentaculum tali, and passing thence onto the scaphoid; (2)
a short ligament from the medial malleolus to the astragalus.
The Tarsus.—The articulations between the separate bones
of the tarsus and between the tarsus and metatarsus have a
considerable number of variously communicating articular capsules.
The separate bones are connected by many ligaments.[92]
The ligaments of the ventral surface (ligamenta plantaria) are
especially well developed. Here may be noticed particularly a
very large calcaneocuboid ligament, and a large calcaneocuneiform
ligament which passes from the sustentaculum tali to the
medial cuneiform. Those on the dorsal surface (ligamenta
dorsalia) are less strong and numerous. Many ligaments,
longitudinal, transverse, and oblique, pass also between the
separate bones (ligamenta interossea). Ligamenta lateralia,
on the lateral and medial borders of the foot, are also distinguishable.
Strauss-Durckheim enumerates ninety-four ligaments
of the tarsus; an account of each of these does not form
part of the plan of the present work.
The ligaments of the metatarsus and phalanges are of the
same general character as in the forelimb. Of these Strauss-Durckheim
enumerates thirty-six; they will not be described
here.
[93]
THE MUSCLES.
I. THE MUSCLES OF THE SKIN. (Fig. 62.)
M. cutaneus maximus (Fig. 62, b).—This is a very large,
thin muscle which covers almost the whole side of the body.
It arises from the outer surface of the latissimus dorsi (Fig. 68,
m) near its ventral end and from the bicipital arch (Fig. 65, t′)
in the axilla; from the linea alba for a considerable distance
(two or three inches) caudad of the base of the xiphoid process,
and from the thorax over a line joining the axilla and the base
of the xiphoid. Sometimes a few fibres take origin from the
fascia which covers the pectoantibrachialis on the ventral side
of the arm.
From their origin the fibres diverge. The cranial ones
curve about the base of the forelimb and are inserted into the
skin at or near the middle line caudad of the first thoracic
vertebra. The most cranial fibres of all are inserted about one
to one and a half centimeters from the middorsal line; thence
the line of insertion approaches the middorsal line to reach it
at about the eighth or ninth thoracic vertebra. The middle
fibres run parallel to the middorsal line in the lumbar and
sacral regions as far as the root of the tail, a small bundle
passing onto the dorsal side of the tail, another onto the ventral
side. The fibres of the caudal portion pass onto the thigh, the
ventral ones running in the fold of skin which stretches from
thigh to abdomen, and are finally lost in the fascia along a line
connecting the knee and the root of the tail. A strong fascia
connects the adjacent borders of the cutaneus and platysma and
lies over the scapular region.
[94]
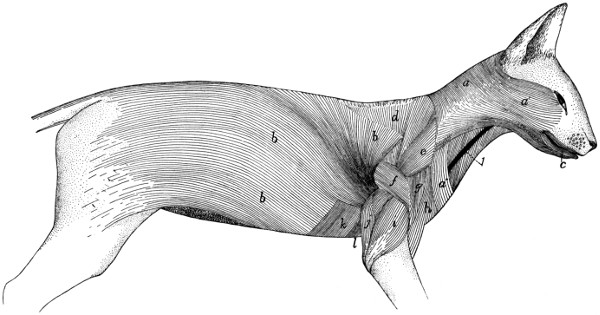
Fig. 62.—Muscles of the Skin.
a, a′, a″, M. platysma; b,
M. cutaneus maximus; c, M. orbicularis oris. Between the platysma and the cutaneus maximus are seen the following
deeper muscles: d, M. acromio-trapezius; e, M. levator scapulæ ventralis;
f, M. spino-deltoideus; g, M. acromio-deltoideus; h, M. clavobrachialis;
i, caput laterale of M. triceps brachii; j, caput longum of M. triceps brachii;
k, M. xiphi-humeralis; l, M. pectoralis minor. 1,
external jugular vein.]
[95]
This muscle lies immediately beneath the integument. It
covers the first layer of body muscles.
Action.—Moves the skin.
M. platysma (Figs. 62 and 64, a,
a′, a″).—This muscle
forms a thin layer of fibres covering the sides of the neck and
face, in close relation with the integument. Several more or
less distinct portions may be distinguished.
(1) Most of the fibres (a) arise from the middorsal line, from
the occiput to the first thoracic vertebra, in a narrow fascia
common to the muscles of the two sides. The most cranial
fibres of this region arise as a small bundle from the external
occipital crest, beneath the levator auris longus (Fig. 63,
g, g′).
From this origin in the middle line the fibres pass craniolaterad.
The most cranial fibres curve about the ventral side
of the ear and pass toward the caudal angle of the eye, where
they unite with fibres of the zygomaticus (Fig. 64, d) or corrugator
supercilii lateralis (Fig. 64, k), or pass to the lower
eyelid. Caudad of these the fibres cover the side of the face
and become lost among the facial muscles, some passing to the
lower eyelid, some to the fibrous pad which supports the
vibrissæ, some to the angle of the mouth, some to the lower
lip. The most ventral fibres meet the fibres of the opposite
muscle just ventrad of the symphysis of the mandible.
The ventral free border of the platysma is separated on the
ventral side of the neck from the border of the opposite muscle
by a wedge-shaped area having its point at the symphysis
menti.
The fibres of this portion of the muscle are interrupted by
an attachment to the skin, along a line passing from the base
of the ear to about the middle of the coracoid border of the
scapula, The dorsal (a) and ventral (a′) portions of the
muscle, separated by this line of attachment, are sometimes
described as separate muscles (the supercervicocutaneus and
cervicofacial, respectively, of Strauss-Durckheim).
(2) A band of fibres one or two centimeters across (a″)
arises in the fascia of the side of the neck just craniad of the
middle of the coracoid border of the scapula, and passes caudoventrad[96]
toward the manubrium, its fibres crossing the fibres of
the first part of the platysma at right angles. These fibres
become lost in the fascia ventrad of the manubrium, or pass
across the middle line to intermingle with the corresponding
fibres of the opposite side. This portion of the platysma is
sometimes absent.
The platysma is everywhere subcutaneous, except at its
dorsocranial angle, where a small bundle of fibres is covered
by the levator longus auris. It covers the deeper muscles of
the neck and head. Closely attached to its inner surface are
the submentalis and depressor conchæ, whose fibres bridge
over the ventral interval between the borders of the platysmas
of opposite sides.
Action.—Moves the skin of the face and neck.
II. THE MUSCLES OF THE HEAD.
A. Superficial Muscles.
—The most superficial layer
of muscles on the face and head is formed by differentiation of
the fibres of the platysma. The muscles thus formed are not
clearly distinct from each other; in this region sets of fibres
differing in direction and in origin or insertion receive separate
names even though the different sets of fibres are closely interwoven.
In the quadrangle on the dorsal surface of the head enclosed
between the two eyes and the two ears, a thin superficial sheet
of fibres is found, in which a number of different sets may be
distinguished (Fig. 63). These have received the following
names.
M. intermedius scutulorum (Fig. 63, a).—This consists
of a broad thin sheet of transverse fibres between the two
external ears. The fibres are attached at either end to the
scutiform cartilage (1) of the two ears, and pass without interruption
across the middle line. At its cranial edge this muscle
is continuous with the corrugator supercilii medialis (b); at its
lateral edge with the frontoscutularis; at its caudal edge with
the levator auris longus (g).
Relations.—Outer surface with the integument. Inner surface[97]
with the galea aponeurotica (to which the muscle is closely
united), the epicranius muscle (h) and the temporal muscle (n).
Action.—Draws the two ears dorsad, toward the middle
line.
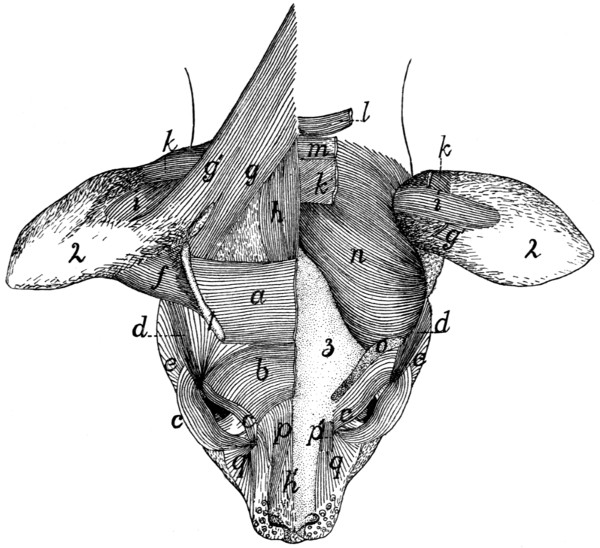
Fig. 63.—Muscles on the Dorsal
Surface of the Head.
On the right side are shown the superficial muscles; on the left side the superficial
muscles have mostly been removed, exposing the deeper muscles and the bone.
a, M. intermedius scutulorum; b, M. corrugator supercilii medialis; c, M. orbicularis
oculi; d, M. corrugator supercilii lateralis; e, cranial end fibres of M. platysma; f,
M. adductor auris superior; g, g′, M. levator auris longus (g, cranial portion; g′,
caudal portion); h, h′, M. epicranius (h, M. occipitalis;
h′ M. frontalis); i, M. transversus
auriculæ; k, M. auricularis superior (cut on the left side); l, M. abductor
auris brevis; m, M. abductor auris longus; n, M. temporalis; o, cut origin of M.
frontoscutularis; p, p′, M. levator labii superioris alæque nasi (p′, the origin from
the maxillary bone); q, angular head of M. quadratus labii superioris. 1, scutiform
cartilage; 2, external ear; 3, bones of the skull.
M. corrugator supercilii medialis (b).—This consists of
a thin sheet of scattered fibres lying craniad of the last and
intermingling with it. The fibres take origin near the middle
line, pass laterad, then curve craniad, converging, to be[98]
inserted into the whole extent of the upper eyelid, especially
near the caudal angle. Here the fibres unite with those of the
orbicularis oculi (c).
This muscle is continuous caudad with the intermedius
scutulorum (a), craniad with the orbicularis oculi (c); laterad
with the corrugator supercilii lateralis (d). Toward the medial
side the fibres lose themselves in a tendinous sheet that joins
the galea aponeurotica.
Relations.—Outer surface with the integument. Inner surface
with the frontoscutularis and the skull.
Action.—Raises the upper eyelid.
M. orbicularis oculi (Fig. 63, c; Fig. 64, s).—This consists
of two thin bands of muscle-fibres which lie one in either
eyelid parallel to its border and unite at the angle of the eye.
Origin by short tendon-fibres from a tubercle on the surface
of the frontal process of the maxillary bone just dorsad of the
orbital end of the lachrymal canal and between the two parts
of the quadratus labii superioris (Fig. 63, p and q).
The muscle splits into two parts which pass into the two
eyelids. At the outer angle of the eye the two bands unite
by the intervention of tendon-fibres between the muscle-fibre
bundles of the two.
Relations.—Outer surface with the integument. Inner surface
with the inner membrane of the eyelid.
Action.—Closes the eye.
M. corrugator supercilii lateralis (Fig. 63, d; Fig. 64, k).—This
consists of a number of scattered fibres which arise from
among the fibres of the frontoscutularis, and from the tendon
lying just craniad of the external opening of the ear, to which
are united also parts of the zygomaticus (Fig. 64, d) and submentalis
(Fig. 64, c). The fibres pass craniad, converging so
as to form a narrow band which is inserted at the caudolateral
angle of the eye, where it unites with the orbicularis oculi
(Fig. 64, s). This muscle is continuous on the medial side
with the corrugator supercilii medialis (Fig. 64, j) and the
frontoauricularis, on the lateral side with the platysma.
Relations.—Outer surface with the integument. Inner surface
with the frontal bone and the frontoscutularis.
[99]
Action.—Pulls the angle of the eye caudad; at the same
time pulls the external ear craniad.
M. frontoauricularis.—A few of the fibres which are
attached along the upper eyelid sometimes pass dorsocaudad,
mingled with fibres of the corrugatores supercilii medialis and
lateralis and the frontoscutularis, to the craniomedial angle of
the auricular cartilage, where they unite with the fibres of the
adductor auris superior. These fibres are sometimes distinguished
as the frontoauricularis muscle.
M. levator auris longus (Fig. 63, g and g′). (Part of the
auricularis posterior of man.)—This lies on the caudal half of
the dorsal surface of the head, forming a laterocaudal continuation
of the intermedius scutulorum (a).
Origin from the middle line of the neck dorsad of the atlas,
and from the sagittal crest for about one centimeter craniad of
the external occipital tubercle. The fibres form a broad thin
sheet which passes craniolaterad as far as the caudal end of the
scutiform cartilage (1). Here the muscle divides; the major
portion is attached to the scutiform cartilage (1), its fibres
intermingling with those of the intermedius scutulorum (a).
The caudal portion of the muscle (g′) passes onto the surface
of the auricle, extending one or two centimeters distad; here
it is inserted on an oblique line which lies directly craniad of
the insertion of the transversus auriculæ (i).
The caudal portion of this muscle (g′) having origin above
the atlas and insertion on the auricle, is sometimes considered
a separate muscle, the supercervicoauricular or cervicoauricular.
The cranial portion (g) with origin on the sagittal crest
and insertion on the scutiform cartilage might be distinguished
as the occipitoscutularis.
The levator auris longus is continuous at its cranial end
medially with M. epicranius (h); laterally with M. intermedius
scutulorum (a).
Relations.—Outer surface with the integument. Inner
surface with the temporal muscle (n), the auricularis superior
(k), the abductor longus auris (m), a narrow strand of the
platysma, and the clavotrapezius.
Action.—Pulls the external ear dorsocaudad.
[100]
The four following muscles lie partly or entirely beneath
those already described; they are doubtless formed as differentiations
of the inner layers of the platysma.
M. auricularis superior (or attollens auris) (Fig. 63, k).—This
muscle forms a band about one centimeter broad lying
just beneath the levator longus (g).
Origin on the sagittal crest for about one centimeter craniad
of the interparietal bone. The muscle passes laterad onto the
surface of the auricle and is inserted into the auricular cartilage
a little caudad of the middle of its convex surface. At its
lateral end the cranial margin is closely united to the under
surface of the levator auris longus (g).
Relations.—Outer surface at the medial end with the epicranius
(h), the abductor auris longus (m), and levator auris
longus (g); at the lateral end with the skin. Inner surface
with the temporal muscle (n) and the auricular cartilage.
Action.—Pulls the external ear dorsad.
M. abductor auris longus (Fig. 63, m; Fig. 64, q). (Part
of the auricularis posterior of man.)
Origin on the sagittal crest dorsad of the interparietal bone,
caudad of that of the auricularis superior (k), which it partly
covers.
The muscle passes laterad as a flat band 8 to 10 millimeters
wide over the caudal surface of the concha of the ear, and is
inserted (Fig. 64, q) on the lateral surface of the eminentia
conchæ, caudad of the antitragus.
Relations.—Outer surface with the levator auris longus (g)
and the integument. Inner surface with the auricularis superior
(k), the abductor auris brevis (l), and the concha.
Action.—Pulls the external ear caudad.
M. abductor auris brevis (Fig. 63, l).
Origin by a tendon from the lambdoidal crest for one or
two centimeters laterad of the middle.
The muscle passes lateroventrad lying beneath the abductor
longus (m) as a flat band 6 to 8 millimeters wide which is
inserted into the medial surface of the most proximal portion
of the concha, just distad of its junction with the cartilaginous
auditory meatus.
[101]
Relations.—Outer surface with the abductor longus (m), a
small strand of the platysma and the concha. Inner surface
with the temporal muscle (n).
Action.—Pulls the concha caudad.
M. epicranius (or occipitofrontalis) (Fig. 63, h and h′).
Origin on the sagittal crest or suture, just craniad of the
origin of the levator longus auris, to the inner surface of which
this muscle is closely united. From the origin the fibres pass
craniad forming a band (h) about 8 or 10 millimeters in width,
the two muscles lying close to one another and partly united
in the middle line. About two centimeters craniad of their
origin the fibres of both muscles end in a tendinous sheet, the
galea aponeurotica, which covers the surface of the skull in the
region between the ears and eyes, and is formed by the inner
surface of the intermedius scutulorum (a) and other muscles of
this region. The galea aponeurotica passes craniad onto the
surface of the nose, where it gives origin again to a thin sheet
of muscle-fibres (h′) which are inserted into the integument
near the cranial ends of the nasal bones.
M. epicranius is thus formed of two muscular portions
(h and h′), connected by a long tendinous sheet. The caudal
portion is frequently distinguished as the occipital muscle
(M. occipitalis, h), the cranial portion, on the nose, as the
frontal muscle (M. frontalis, h′).
Relations.—Outer surface of the occipitalis (h) with the
intermedius scutulorum (a), the levator auris longus (g), and
the integument; inner surface with the auricularis superior (h),
the abductor auris longus, and the bone. Outer surface of
the frontalis (h′) with the integument; inner surface with the
bones of the skull.
Action.—Moves the integument of the dorsal surface of the
head and of the nose.
Ventrad of the external ear the following three differentiated
portions of the platysma may be distinguished.
M. zygomaticus (major) (Fig. 64, d).—A slender band
connecting the angle of the mouth with the scutiform cartilage
of the ear.
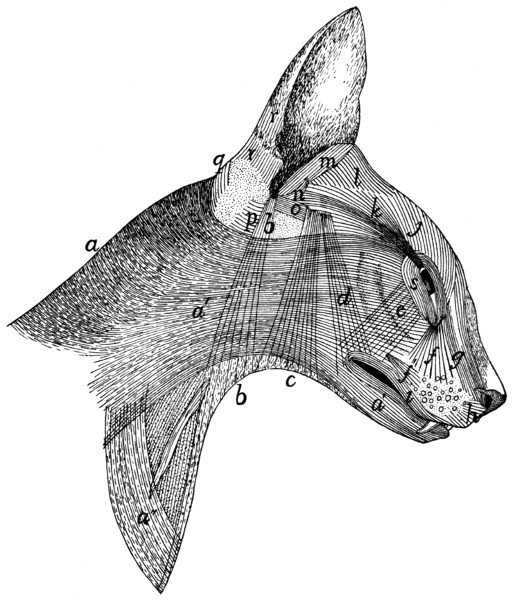
Fig. 64.—Superficial
Muscles on the Lateral Surface of the Head and Neck.
a, a′, a″, M. platysma;
b, M. depressor conchæ; c, M. submentalis; d, M. zygomaticus
(b, c, and d lie beneath the platysma and are seen through it); e, M. zygomaticus
minor; f′, M. caninus; f, g, parts of M. quadratus labii superioris (f,
angular head; g, M. levator labii superioris alæque nasi); h, M. myrtiformis; i, M.
orbicularis oris; j, M. corrugator supercilii medialis; k, M. corrugator supercilii
lateralis; l, M. adductor auris superior; m, M. helicis; n, M. antitragicus; o, M.
adductor auris inferior; p, M. conchæus externus; q, M. abductor auris longus; r,
parts of M. auricularis externus; s, M. orbicularis oculi.
At the angle of the mouth the fibres arise from among those[102]
of the orbicularis oris (i), from both the upper and the lower
lip. They pass thence dorsocaudad across the zygomatic arch
as a band 8 or 10 millimeters wide which is attached to a
tendinous aponeurosis lying just craniad of the external opening
of the ear. To this aponeurosis are attached also, wholly or
partly, the corrugator supercilii lateralis (k) and the submentalis[103]
(c). The aponeurosis is united with the caudoventral angle
of the scutiform cartilage and with the ventral edge of the
cranial margin of the auricular cartilage.
This muscle is united at its cranial end with the orbicularis
oris (i); throughout its length more or less with the platysma
(a, a′); at its caudodorsal end with the corrugator supercilii
lateralis (k) on its medial side, and with the submentalis (c) on
its lateral side.
Relations.—Outer surface with the integument and the
platysma (a′). Inner surface with the masseter, temporal,
frontoscutularis, and adductor auris inferior (o).
Action.—Draws the angle of the mouth dorsocaudad; the
external ear ventrocraniad.
M. submentalis (Fig. 64, c).—A flat band, similar to the
last, and closely connected for some distance to its caudal
border.
It arises as scattered fibres near the ventral middle line at
about the level of the larynx, the fibres of the opposite muscles
crossing the middle line and intermingling. The fibres pass
dorsad, converging so as to form a narrower band, which
unites dorsad with the zygomaticus (d), to be inserted with it
into the tendinous aponeurosis above described.
Relations.—Outer surface with the integument and platysma
(a′). Inner surface with the muscles of the hyoid, the
digastricus, the parotid gland, the masseter, the zygomatic arch
and the temporal muscle.
Action.—Draws the external ear ventrad.
M. depressor conchæ (Fig. 64, b).—A thin band of fibres,
caudad of the last and parallel with it. The muscle arises as
scattered fibres on the ventral surface of the neck, one or
two centimeters craniad of the manubrium, the fibres of the
muscles of opposite sides crossing the middle line and interdigitating.
They pass dorsad, gathering together to form a
narrow band which is inserted into the summit of the antitragus.
Relations.—Outer surface with the skin at the ventral end,
and with the platysma (a′) further dorsad. Inner surface with
the deep muscles of the neck and with the parotid gland.
Action.—Draws the external ear ventrad.
[104]
Lying deeper than the muscles thus far described, but
closely connected with a number of them, are the following:
M. frontoscutularis. Origin (Fig. 63, o) on the frontal bone, along the supraorbital
margin, from the craniomedial angle of the eye caudad
to the zygomatic process of the frontal.
The fibres form a large muscle which passes dorsocaudad
to the scutiform cartilage and is mostly attached along its
ventrolateral border. Some of the outer fibres, however, pass
distad of the scutiform cartilage toward the cartilage of the ear,
thus joining the adductor auris superior (f).
The outer surface of this muscle is closely connected with
fibres of the intermedius scutulorum (a), the corrugatores
supercilii medialis (b) and lateralis (d), and the frontoauricularis,
when this exists. The outer surface is covered near the
origin by the orbicularis oculi (c), farther dorsad by the muscles
just mentioned and the integument. The inner surface is in
relation with the frontal bone and the adductor auris inferior
(Fig. 64, o).
Action.—Pulls the ear craniad.
M. adductor auris inferior. (Fig. 64, o.) (Part of the
auricularis anterior of man.)—A small muscle lying beneath
the frontoscutularis.
Origin on the ligament which connects the zygomatic
process of the frontal with the frontal process of the malar.
The fibres form a thin band about 7 millimeters wide which
passes craniad, closely united to the inner surface of the frontoscutularis.
At the caudal edge of the zygomaticus (d) the
muscle is interrupted by a short tendinous interval; the fibres
then continue to their insertion on the tip of the antitragus.
Relations.—Outer surface with the frontoscutularis, the
zygomaticus (d), and the integument. Inner surface with the
temporal muscle and M. antitragicus (n).
Action.—Pulls the ear craniodorsad.
The remainder of the muscles of the ear, which, with the
exception of the tragicus lateralis, merely interconnect the
cartilages of the external ear or parts of these cartilages, are[105]
described in connection with the account of the Auditory
Organ.
Owing to the presence of the fibrous pad to which the
whiskers are attached, the facial muscles between the eye and
the mouth differ in the cat in some respects from those of
related animals. This fibrous pad interrupts the muscles, frequently
breaking muscles which are elsewhere single into two.
M. zygomaticus minor, or malaris (Fig. 64, e).—This
muscle is not always present; when it exists it forms a very thin
flat band of fibres passing from the ventral side of the eye
toward the angle of the mouth.
Origin among the fibres of the orbicularis oculi (s) in the
lower eyelid. The fibres pass ventrad; the insertion varies.
In some specimens the insertion is among the fibres of the
orbicularis oris (i), at the angle of the mouth, beneath the
zygomaticus (d). In other cases this muscle is said to be
inserted into the pad on which the whiskers rest.
Relations.—Outer surface with the integument and near its
insertion with the zygomaticus major (d). Inner surface with
the malar bone and the masseter muscle.
Action.—Pulls the angle of the mouth (or the whiskers)
dorsad.
M. orbicularis oris (Fig. 64, i).—This forms a very thin
layer of subcutaneous concentric fibres which surround the
mouth, that of the upper lip being thicker than that of the
lower lip. The part of the muscle in the lower lip has its
fibres intermingled with those of the platysma (a′). In the
median line the fibres of the upper lip are interrupted by a
raphe, and caudad of this are intermingled with those of the
caninus.
M. quadratus labii superioris (Fig. 63, p and q; Fig. 64,
f and g).—This is a complex of muscle-fibres attached chiefly
to the fibrous pad on which the whiskers rest. In it two parts
can be distinguished more or less completely.
(1) M. levator labii superioris alæque nasi (Fig. 63, p;
Fig. 64, g) on the sides of the nose. This arises (a) as a continuation
of the frontal portion of the epicranius, and (b) in a[106]
small bundle (Fig. 63, p) from near the dorsal end of the
frontal process of the maxillary bone, in common with the
next. The two slips thus formed unite craniad and are inserted
partly into the integument on the outer side of the wing of the
nose, partly into the skin of the upper lip, lateroventrad of the
nose, and partly into the pad on which the whiskers are located,
in connection with the next.
(2) The angular head (caput angulare), or levator labii
superioris proprius (Fig. 63, q; Fig. 64, f) is much larger
than the last. It takes origin from a small tubercle at the
cranial border of the orbit, close to the origin of the orbicularis
oculi. The fibres diverge from their origin to their insertion
among the whiskers and in the fibrous pad on which the
whiskers rest.
Action.—Erects the whiskers and raises the upper lip.
M. caninus or levator anguli oris (Fig. 64, f′).
Origin in a depression on the lateral surface of the maxillary
bone, just craniad of the last premolar tooth. The fibres
pass craniad, diverging, toward the whiskers, and have their
insertion into the pad of tissue on which the whiskers rest.
Action.—Retracts the whiskers and raises the upper lip.
M. buccinator.—A thin muscle against the mucous membrane
of the upper lip, partly united with the orbicularis oris
and lying beneath M. caninus.
Origin on the outer surface of the superior maxillary bone
in the depression between the infraorbital foramen and the first
premolar tooth. The fibres pass ventrad, diverging, and
mingle with those of the deep part of the orbicularis oris.
Relations.—Outer surface with the caninus and with the
orbicularis oris. Inner surface with the mucous membrane of
the upper lip.
Action.—Raises the upper lip.
M. myrtiformis (probably corresponds to a part of the
human M. nasalis) (Fig. 64, h).—A thin sheet of muscle-fibres
lying craniad of the levator labii superioris alæque nasi and
passing from the whiskers to the nose and upper lip. It is distinguishable
from the alæque nasi by the direction of the
fibres.
[107]
Origin from the pad upon which the whiskers rest. The
fibres diverge and have their
Insertion upon the wing of the nose and into the skin of the
upper lip near the median line.
Action.—Dilator of the nares and elevator of the upper lip.
M. “moustachier” (Str.-D.) (probably corresponds to
one of the Mm. incisivi).—It lies on the upper lip near the
median line.
Origin from the outer surface of the premaxilla near the
suture along the ventral border of the narial opening. The
fibres diverge and pass caudad into the upper lip within the
superficial fibres of the orbicularis oris.
Insertion.—The skin of the upper lip just craniad of the
whiskers. Some of the fibres pass into the pad on which the
whiskers rest.
Action.—It carries the lip craniad.
M. quadratus labii inferioris.—A thin flat band extending
almost the length of the lower lip.
Origin from the alveolar border of the mandible, between
the molar tooth and the canine. The fibres pass dorsad into
the lower lip, where they intermingle with those of the orbicularis
oris. The muscle is perhaps not constant.
Action.—Depressor of the lower lip.
Sometimes a few fibres arising from the cranial portion of
the alveolar border of the mandibula pass mediad, seeming to
join corresponding fibres from the opposite side. Such fibres
would constitute a M. transversus menti.
B. Deep Muscles of the Head.
(a) Muscles of Mastication.
M. digastricus (Fig. 65, b).—The digastric is a thick prismatic
muscle lying mediad of the angle of the lower jaw, connecting
it with the base of the skull.
Origin by fleshy fibres from the outer surface of the jugular
process of the occipital bone, and by a thin tendon from the
tip of the mastoid process and from the ridge between the
mastoid and the jugular processes. The muscle passes craniad,
becoming at the same time broader and more voluminous.
[108]
Insertion.—The ventral border of the mandible craniad of
the caudal border of the molar tooth. The insertion area
extends slightly onto both surfaces of the bone and is triangular,
with its apex craniad.
Relations.—Outer (ventral) surface with the following,
beginning at the caudal end: the rectus capitis lateralis, the
cleidomastoid (h), the submaxillary gland (2), the stylohyoid
(d), a large lymph-gland (3), and the integument. Inner
(dorsal) surface with the jugulohyoid, the tympanic bulla, the
masseter (a), the hyoglossus, and the mylohyoid (c).
Action.—Depressor of the lower jaw.
M. masseter (Figs. 65 and 66, a).—The masseter forms
the projecting mass so prominent in the cat behind and below
the eye. Its outer surface is covered by a strong aponeurosis.
The muscle is divided into three layers which are distinct as
to origin, insertion, and direction of fibres, but which are
otherwise not distinct.
The superficial layer takes
Origin by the superficial aponeurosis from the ridge which
runs lengthwise of the outer surface of the malar bone, and by
fleshy fibres from the surface of the malar bone ventrad of this
ridge. The fibres pass obliquely caudad to their insertion.
Insertion.—The caudal half of the lateral margin of the
ventral border of the mandible, by a sheet of fascia which lies
on the inner surface of the muscle. The superficial fibres curve
ventrad of the mandible and are inserted into a tendinous intersection
between this muscle and the internal pterygoid (Fig.
66, c).
The middle layer takes
Origin from the ventral border of the malar bone, from
opposite the molar tooth to within three millimeters of the
mandibular fossa. The fibres pass ventrad and have their
Insertion into the mandible at the ventral border of the
external coronoid fossa by means of the aponeurosis which
covers the inner surface of the muscle.
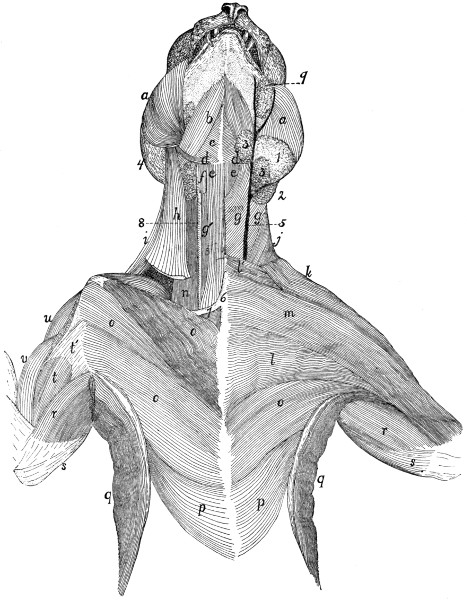
Fig. 65.—Muscles
on the Ventral Surface of the Thorax, Neck and
Head.
The left side shows the first layer of muscles beneath the skin and skin-muscles;
the right side shows the second layer, except in the head region. a, M. masseter;
b, M. digastricus; c, M. mylohyoideus; d, M. stylohyoideus; e, M. sternohyoideus;
f, M. thyreohyoideus; g, M. sternomastoideus; g′, M. sternothyreoideus; h, M.
cleidomastoideus; i, M. levator scapulæ ventralis; j, M. clavotrapezius; k, M. clavobrachialis;
l, M. pectoralis major; m, M. pectoantibrachialis; n, part of M. scalenus;
o, M. pectoralis minor; p, M. xiphihumeralis; q, M. latissimus dorsi; r, M. epitrochlearis;
s, caput longum of M. triceps brachii; t, M. biceps brachii; t′, bicipital
arch; u, M. acromiodeltoideus; v, M. brachialis. 1, parotid gland; 2, submaxillary
gland; 3, lymphatic glands; 4, external ear; 5, external jugular vein; 6, manubrium;
7, first rib; 8, common carotid artery; 9, molar gland.
The deep layer takes
Origin by a strong tendon from the ventral border of the
zygoma just craniad of the mandibular fossa. The fibres pass
cranioventrad, diverge and have their
[109]
[110]
Insertion into the external coronoid fossa.
Relations.—Outer surface with the following, beginning at
the caudal border: the external ear, the parotid gland (Fig.
65, 1), the platysma, the submentalis, the zygomaticus (major),
the zygomaticus minor. Ventral border with a lymph gland
caudad of the angle of the mouth (Fig. 65, 3), and the digastric
muscle (b). Inner surface with the bones.
Action.—A very powerful elevator of the lower jaw.
M. temporalis (Fig. 63, n).—The temporal muscle is the
great mass taking origin from the temporal fossa and having
its insertion on both surfaces and both borders of the coronoid
process of the mandible. It may be divided into two layers,
deep and superficial.
The superficial layer. The temporal fascia stretches over
the temporal fossa, being attached to its borders; that is, to the
sagittal and lambdoidal crests, to the curved ridge which connects
the sagittal crest with the zygomatic process of the frontal
bone, to the caudal border of this process, to the caudal border
of the malar bone, to the caudal border of the ligament connecting
the malar bone and the zygomatic process of the
frontal, and to the dorsal border of the zygomatic process of
the temporal bone and its dorsal root. The craniolateral part
of this fascia is much stronger than the remainder.
The muscle-fibres take origin from the strong craniolateral
part of the inner surface of the temporal fascia, from the groove
on the temporal bone dorsad of the dorsal root of its zygomatic
process, from the whole inner surface of this process and of the
zygomatic process of the malar bone, and sometimes also from
that part of the frontal bone which lies within the temporal
fossa.
Insertion.--The outer surface of the coronoid process of the
mandible dorsad of the coronoid fossa, and both borders of the
coronoid process. The cranial fibres may be inserted craniad
of the coronoid process onto the aponeurosis covering the deep
portion of the muscle. The caudal part of this portion is more
or less distinct and is sometimes described as a separate head.
The deep portion.
Origin by fleshy fibres from the whole surface of the temporal[111]
fossa, except in cases where the cranial part of the fossa
is occupied by the superficial portion. The fibres converge
toward the coronoid process, and the ventral third of the outer
surface is covered by strong fascia.
Insertion.—The whole inner surface of the coronoid process
of the mandible.
Relations.—Outer surface with the external ear, the ear-muscles
(Fig. 63), the epicranius (Fig. 63, h), the corrugatores
supercilii lateralis (d) and medialis (b). Inner surface with
the skull, the pterygoideus externus (Fig. 66, b), and the structures
within the orbit.
Action.—Elevator of the lower jaw.
M. pterygoideus externus (Fig. 66, b).—This lies on the
medial side of the mandible, ventrad of the ventral portion of
the temporal.
Origin.—The external pterygoid fossa (Fig. 40, h), which
lies on the lateral surface of the perpendicular plate of the palatine
bone, and on the surface of the pterygoid process of the
sphenoid bone dorsad of the internal pterygoid fossa. The
fossa extends from the foramen rotundum to the sphenopalatine
foramen. The muscle passes with parallel fibres laterad,
ventrad, and caudad and has its
Insertion by a strong flat tendon into the medial surface of
the mandible near its ventral border between the opening of
the mandibular canal and the base of the angular process.
Relations.—Ventral surface with the masseter (a), the
digastric, and the soft palate with its muscles. Dorsal surface
with the temporal, and near its origin with the muscles of the
eye, and other structures in the orbit.
Action.—Elevator of the lower jaw.
M. pterygoideus internus (Fig. 66, c).—This lies caudad
of the pterygoideus externus, nearly parallel to it, and is connected
with it at its lateral end.
Origin from the whole surface of the internal pterygoid
fossa (Fig. 40, s).
Insertion by fleshy fibres into the ventral surface of the
external pterygoid and its tendon, into the medial surface of
the angular process of the mandible and caudad of the angular[112]
process for about one centimeter into a fascia common to it and
the masseter muscle, and into the stylomandibular ligament.
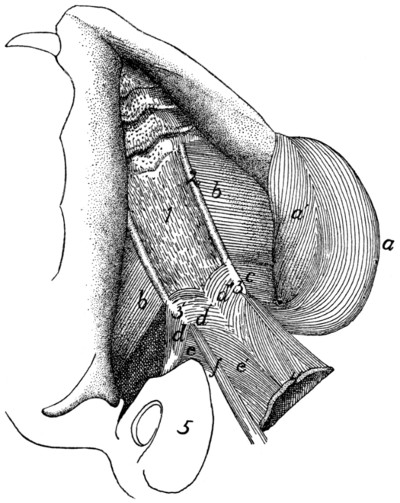
Fig. 66.—Obliquely
Ventral View of the Deep Muscles of the Lower
Jaw and Palate.
The tongue and other structures forming the floor of the mouth have been removed,
showing the roof of the mouth. On the right side the masseter and pterygoideus
internus have been removed, a, a′, M. masseter; b, M. pterygoideus externus;
c, M. pterygoideus internus; d, d′, M. tensor veli palatini; e, e′, M. levator
veli palatini; f, M. constrictor pharyngis superior. 1, aponeurosis of the velum
palatinum; 2, ridges formed by the palatines and pterygoid portions of the sphenoid;
3, hamular processes; 4, free caudal border of the velum palatinum; 5, bulla tympani.
Relations.—Ventral surface with the styloglossus and the
soft palate. Dorsal surface with the bulla tympani (5) and
base of the skull. Cranial surface with the pterygoideus
externus (b), with which it is partly united.
Action.—Assists the pterygoideus externus.
(b) Muscles of the Hyoid Bone.
M. stylohyoideus (Fig. 65, d).—The stylohyoid is a
ribbon-like muscle about three millimeters wide. It lies on[113]
the superficial surface of the digastric, caudad and ventrad of
the angle of the jaw.
Origin by fleshy fibres from the outer surface of the stylohyal
bone near its middle. The muscle curves over the outer surface
of the digastric (b), and ends about one centimeter from
the middle line in a slender flat tendon.
Insertion into the middle of the ventral surface of the body
of the hyoid bone. Some of the fibres of the mylohyoid (c) are
inserted into its tendon.
Relations.—Outer surface with the submaxillary gland (2)
and the submentalis muscle. Inner surface with the digastric (b),
the hyoglossus (Fig. 67, h), and the geniohyoid (Fig. 67, g).
Action.—Raises the hyoid.
M. geniohyoideus (Fig. 67, g).—The geniohyoid is a long
slender muscle which lies next to the median line between the
symphysis menti and the hyoid bone, closely applied to its
fellow of the opposite side and within (dorsad of) the mylohyoid
(Fig. 65, c).
Origin from the ventral half of the inner surface of the
mandibula for about one millimeter from the symphysis.
Insertion by muscle-fibres into the lateral half of the ventral
surface of the body of the hyoid bone.
Relations.—Outer (ventral) surface with the stylohyoid
(Fig. 65, d) and the mylohyoid (Fig. 65, c); medial border with
the muscle of the opposite side. Lateral border with the
hyoglossus (Fig. 67, h) and genioglossus (Fig. 67, f). Inner
(dorsal) surface with the genioglossus.
Action.—Draws the hyoid forward.
M. jugulohyoideus (Fig. 67, b).—The jugulohyoid is a
small flat quadrangular muscle covering the outer part of the
ventral surface of the tympanic bulla.
Origin from the ventral border of the jugular process.
Insertion into the caudal side of the cartilaginous part of the
stylohyoid opposite the origin of the styloglossus.
Relations.—Ventral surface with the digastric. Dorsal surface
with the tympanic bulla (3).
Action.—Draws the hyoid backward and thus gives a firmer
surface of origin for the styloglossus.
[114]
M. mylohyoideus (Fig. 65, c).—The mylohyoid is a large
thin muscle seen after the reflection of the digastric, filling with
its fellow of the opposite side the whole angle between the rami
of the lower jaw. Its fibres are transverse.
Origin from the middle of the medial surface of the body
of the mandible between the caudal opening of the mandibular
canal and the symphysis of the jaw.

Fig. 67.—Muscles
of Tongue, Hyoid Bone, and Pharynx.
a, M. tragicus lateralis; b, M. jugulohyoideus; c, M. pterygoideus externus; d,
partially cut surface of M. pterygoideus internus; e, M. styloglossus; f, M. genioglossus;
g, M. geniohyoideus; h, M. hyoglossus; i, M. glossopharyngeus; j, M.
constrictor pharyngis medius; k, M. constrictor pharyngis inferior; l, M. stylopharyngeus;
m, M. sternohyoideus (cut); n, M. cricothyreoideus; o, M. sternothyreoideus
(cut); p, M. thyreohyoideus. 1, mandible; 1′, angular process of
mandible; 2, stylomandibular ligament; 3, bulla tympani; 4, trachea; 5, œsophagus;
6, thyroid gland; 7, isthmus of the thyroid gland.
Insertion with the opposite muscle into a median raphe
which extends from the symphysis of the jaw to the hyoid bone
and is closely united to the external surface of the geniohyoid.
Some of the fibres gain insertion into the body of the hyoid[115]
bone through their insertion into the tendon of the stylohyoid
(d).
Relations.—Outer surface with the submentalis (Fig. 64, c)
and the digastric (Fig. 65, b). Lateral edge with the masseter
(a), the digastric (b), and the mandibula. Caudal border with
the stylohyoid (d). Inner (dorsal) surface with the geniohyoid
(Fig. 67, g), the hyoglossus (Fig. 67, h), and the genioglossus
(Fig. 67, f).
Action.—Raises the floor of the mouth and thus brings the
hyoid forward.
M. ceratohyoideus.
Origin, the two proximal pieces of the cranial horn of the
hyoid. The fibres pass caudad parallel to one another and
within the middle constrictor of the pharynx.
Insertion into the whole length of the caudal cornu of the
hyoid bone.
Relations.—Outer surface with the middle constrictor of the
pharynx. Inner surface with the pharynx.
Action.—Draws craniad the body of the hyoid by bringing
its cornua together and bending the joint between the two
pieces of the cranial cornua.
The muscles connecting the hyoid and the tongue are
described in connection with the description of the tongue.
III. MUSCLES OF THE BODY.
1. Muscles of the Back.
A. Muscles of the Shoulder
(connecting the forelimb
with the back) (Fig. 68).
M. trapezius.—The trapezius muscle is divided in the cat
into three portions, the spinotrapezius, the acromiotrapezius,
and the clavotrapezius.
M. spinotrapezius, or trapezius inferior (Fig. 68, j).—The
spinotrapezius (caudal part of the human trapezius) is a flat
triangular muscle.
Origin from the tips of the spinous processes of all or
nearly all the thoracic vertebræ and from the intervening
supraspinous ligament. The origin may extend from a point
anywhere between the first and fourth spines to any point
between the eleventh and thirteenth.
[116]
Insertion along an S-shaped line (Fig. 76, k-l) which
crosses the tuberosity of the scapular spine and forms an angle
with the spine of about forty-five degrees. The line extends
on one side onto the fascia covering the supraspinatus muscle,
and on the other side onto the fascia covering the infraspinatus,
but does not reach the borders of the scapula. The cranial
two-thirds of the insertion is by a flat tendon which is broader
craniad.
Relations.—Outer surface with the cutaneus maximus, and
craniad with the acromiotrapezius (h). Inner surface with the
latissimus dorsi (m) caudad and with the rhomboideus, supraspinatus,
and infraspinatus craniad. Caudal border free; cranial
border in contact with the acromiotrapezius (h).
Action.—Draws the scapula dorsocaudad.
M. acromiotrapezius, or trapezius superior (Fig. 68, h).—The
acromiotrapezius (middle part of the human trapezius) is
a flat four-sided muscle just craniad of the spinotrapezius, from
the median dorsal line to the scapular spine.
Origin along the median dorsal line from the spinous process
of the axis to a point anywhere between the spinous processes of
the first and fourth thoracic vertebræ. Its caudal two-thirds is
from a flat tendon which is directly continuous with the muscle
of the opposite side and thus bridges the depression between
the vertebral borders of the scapulæ. This tendon narrows
craniad, and its cranial fourth is attached by a fascia to the
spinous processes of the cervical vertebræ. At the cranial
border of the muscle the fibres sometimes reach the middle
line. The fibres of the muscle run nearly transversely.
Insertion (Fig. 76, h).—Into the outer surface of the metacromion,
the glenoid border of the scapular spine from the
metacromion to the tuberosity, and along a line continued for
about one centimeter from the tuberosity onto the surface of
the spinotrapezius (j) at the junction of its muscular and tendinous
portions.
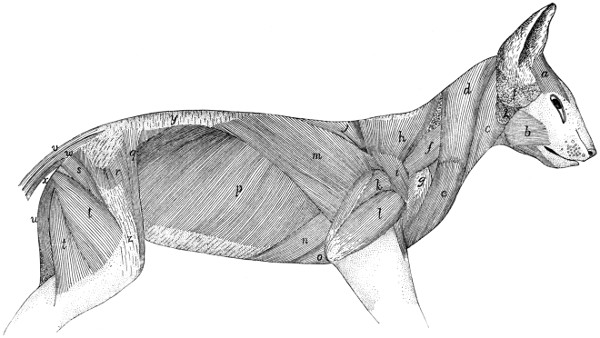
Fig. 68.—First
Layer of Body-muscles, beneath the Skin and Skin-muscles.
a, M. frontoscutularis; b, M. masseter; c, M. sternomastoideus;
d, M. clavotrapezius; e, M. clavobrachialis; f, M. levator scapulæ
ventralis; g, M. acromiodeltoideus; h, M. acromiotrapezius; i, M. spinodeltoideus; j,
M. spinotrapezius; k, caput longum of M. triceps
brachii; l, caput laterale of M. triceps brachii; m, M. latissimus dorsi; n, M. xiphihumeralis;
o, M. pectoralis minor; p, M. obliquus
abdominis externus; q, M. sartorius; r, M. tensor fasciæ latæ; s,
M. caudofemoralis; t, M. biceps femoris; u, M. semitendinosus; v, M.
extensor caudæ lateralis; w, M. abductor caudæ externus; x, M. flexor caudæ
longus; y, lumbodorsal fascia, superficial layer; z, fascia lata.
1, parotid gland; 2, submaxillary gland.
Relations.—Outer surface with the cutaneus maximus, the
integument and platysma. Inner surface with a mass of fat
partially covering the rhomboideus; with the occipitoscapularis,
spinotrapezius, supraspinatus, levator scapulæ, and splenius.[117]
[118]
Near its origin its caudal border touches the outer surface of the
spinotrapezius (j). Its cranial border is overlaid near its
insertion by the levator scapulæ ventralis (f) and is closely
related to the caudal border of the clavotrapezius (d) near its
origin.
Action.—The two muscles hold the scapulæ together.
M. clavotrapezius (Fig. 68, d).—The clavotrapezius
(cranial part of the human trapezius) is a flat muscle passing
from the lambdoidal crest and middorsal line to the clavicle
and covering the side of the neck.
Origin.—The medial half of the lambdoidal crest and the
middorsal line between the crest and the caudal end of the
spine of the axis. The fibres are parallel and pass caudoventrad
so that the muscle covers the ventral surface of the neck
caudally and fills the concavity at the front of the shoulder.
Insertion into the clavicle and into a raphe between the
clavotrapezius and the clavobrachial (e) muscles. This raphe
extends along the clavicle and for some distance laterad of it.
The muscle is partly continuous with the clavobrachial (e).
Relations.—Outer surface with the platysma, and at its
craniomedial angle with the levator auris longus. Inner surface
with the rhomboideus, splenius, occipitoscapularis, levator
scapulæ, cleidomastoid, and supraspinatus. The cranial edge
touches the sternomastoid. The caudal border touches the
acromiotrapezius near the origin.
Action.—Draws the scapula craniodorsad.
This muscle and the clavobrachial (e) are sometimes considered
as forming a single muscle, the cephalohumeral, or
cephalobrachial.
M. occipitoscapularis (or levator scapulæ dorsalis or
rhomboideus capitis) (Fig. 73, a).—The occipitoscapularis
(equivalent to a part of the human rhomboideus) is a slender
flat muscle from the lambdoidal ridge to the coracovertebral
angle of the scapula.
Origin, the medial half of the lambdoidal ridge beneath the
clavotrapezius (Fig. 68, d). The origin does not extend quite
to the middle line. The muscle passes almost directly caudad.
Its caudal end is thicker and narrower than the cranial end and[119]
is wedged between the rhomboideus dorsally and the levator
scapulæ ventrally.
Insertion either into the inner surface of the levator scapulæ
near its dorsocranial border at its insertion or into the scapula
near the coracovertebral angle (Fig. 78, f) or into both. Its
fibres are very closely related to those of the rhomboideus.
Relations.—By its outer surface with the clavotrapezius
(Fig. 68, d) and acromiotrapezius (h). At its insertion it is
often overlaid by either the rhomboideus or the levator scapulæ
or both. By its inner surface with the splenius (Fig. 73, b).
Its caudal third is wedged between the rhomboideus and the
levator scapulæ.
Action.—Draws forward and rotates the scapula.
M. rhomboideus (probably equivalent to the human rhomboideus
major and rhomboideus minor).—A rather thick trapezoid
muscle connecting the vertebral border of the scapula with
the spinous processes of the adjacent vertebræ.
Origin from the caudal two or three fifths of the cervical
supraspinous ligament and from the sides and tips of the first
four thoracic vertebral spines and the interspinous ligaments
caudad of these four spines. It passes lateroventrad, and the
fibres converge somewhat so that the line of insertion is shorter
than the line of origin.
Insertion by a short tendon (1 millimeter long) into the
vertebral border of the scapula, the line of insertion passing
gradually from the inner (Fig. 78, e) to the outer (Fig. 76, i)
surface; and by fleshy fibres into the outer surface of the glenovertebral
angle of the scapula (Fig. 76, i) in close connection
with the origin of the teres major (j). A bundle of fibres at
the cranial border may be inserted into the cranial border of
the levator scapulæ at its insertion.
Relations.—By its outer surface with the spinotrapezius
(Fig. 68, j), acromiotrapezius (h), latissimus dorsi (m), and
clavotrapezius (d); it is usually partially separated from these
muscles by a mass of fat. By its inner surface near its insertion
with the serratus anterior (Fig. 73, i) and levator scapulæ
(Fig. 73, h), and craniad with the splenius (Fig. 73, b), and
the tendon of the serratus posterior superior (Fig. 73, l). At[120]
its lateral end it touches the occipitoscapularis (Fig. 73, a),
serratus anterior (i), levator scapulæ (h), supraspinatus (Fig.
75, a), and infraspinatus (Fig. 75, c), and the teres major (Fig.
75, d).
Action.—Draws the scapula toward the vertebral column.
M. cleidomastoideus (clavicular portion of the human sternocleidomastoid)
(Fig. 65, h).—A narrow flat muscle between
the mastoid process and the clavicle.
Origin from the apex and caudal margin of the mastoid
process of the temporal bone, by tendon and muscle fibres.
The muscle passes caudad, becoming broader and thinner.
Insertion into the lateral four-fifths of the clavicle within
the insertion of the clavotrapezius, and into the raphe which
continues laterad of the clavicle.
Relations.—Outer or lateral surface at the caudal end with
the clavotrapezius (Fig. 65, j); further craniad with the
depressor conchæ (Fig. 64, b), the platysma, and the sternomastoid
(Fig. 65, g). Inner or medial surface with the supraspinatus
(Fig. 75, a) caudad, with the mass of fat in the axilla
at its middle, with the levator claviculæ ventralis (Fig. 65, i)
and obliquus capitis superior (Fig. 71, e) craniad.
Action.—Pulls the clavicle craniad when the head is fixed.
Turns the head and depresses the snout when the clavicle is
fixed.
M. levator scapulæ ventralis, or levator claviculæ (Fig.
68, f; Fig. 65, i; Fig.
72, c, c′, c″) (not usually found in man).
Origin (Fig. 72, c′, c″) in two parts. (1) By fleshy fibres
from the ventral surface of the atlantal transverse process near
its caudal border (c′). (2) By a small tendon from the ventral
surface of the basioccipital opposite the middle of the length of
the bulla tympani (c″) in common with the longus capitis (a).
The two heads unite to form a flat band which passes caudoventrad
and is inserted (Fig. 76, f) by a flat short tendon,
about eight millimeters broad, into the outer surface of the
metacromion along its ventral border, and into the infraspinatus
fossa for one or two millimeters distad of that border.
Relations.—Near the insertion the dorsal border overlaps
the border of the acromiotrapezius (Fig. 68, h), and the ventral[121]
border is attached by strong fascia to the border of the clavotrapezius
(Fig. 68, d). The outer surface is in relation with
the longissimus capitis (Fig. 73, g), clavotrapezius (Fig.
68, d), cleidomastoid (Fig. 73, d), platysma, and the skin.
The inner surface is in relation caudad with the supraspinatus
and with a mass of fat and a lymphatic gland which occupy the
hollow of the shoulder. Craniad it is in relation with the
cleidomastoid (Fig. 73, d), the scalenus (Fig. 73, f), the
longus capitis (Fig. 73, e), and the obliquus superior (Fig.
71, e).
Action.—Pulls the scapula craniad.
M. latissimus dorsi (Fig. 68, m; Fig. 65,
q; Fig. 77, e).—A
large triangular sheet covering rather more than the dorsocranial
half of the abdomen and thorax. It arises from the
middorsal line, from the pelvis to the fifth thoracic spine, and
is inserted into the humerus.
Origin from the tips of the neural spines of the vertebræ
from the fourth or fifth thoracic to about the sixth lumbar.
As far as the tenth or eleventh thoracic vertebra the origin is
directly by muscle-fibres. Caudad of this is a broad triangular
tendon which may be so closely united to the tendons of the
underlying muscles that it cannot be separated. The muscle
passes cranioventrad, the fibres converging to the axilla. In
the axilla a part of the cutaneus maximus and the epitrochlearis
(Fig. 65, r) take origin from the outer surface.
Insertion.—The muscle then ends in a flat tendon, to the
cranial surface of which the fibres of the teres major (Fig.
77, c) are attached. Five to ten millimeters from the humerus
the muscle-fibres of the teres give place to the tendon-fibres,
and the conjoined tendon of the teres and latissimus which is
thus formed (Fig. 79, d′) is inserted into a rough elongated
area (Fig. 82, f) which lies parallel with the pectoral ridge on
the medial surface of the shaft of the humerus (Fig. 48, g).
The teres tendon forms the proximal part of the conjoined
tendon. A part of the tendon of the latissimus may join the
pectoralis minor at its distal or caudal border (Fig. 65), and
thus it may contribute to the formation of both pillars of the
bicipital arch (Fig. 65, t′).
[122]
Relations.—Its outer surface is covered caudad by the
cutaneus maximus, and craniad by the spinotrapezius (Fig.
68, j) and the long head of the triceps (Fig. 68, k). The
cranial border comes into relation with the spinotrapezius (j)
and teres major (Fig. 77, c). The caudal border is in contact
at the ventral end with the xiphihumeralis (Fig. 65, p) and
pectoralis minor (Fig. 65, o). The inner surface of the latissimus
covers near its cranioventral end parts of the xiphihumeralis
(Fig. 68, n), pectoralis minor (Fig. 68, o), teres major
(Fig. 77, c), and serratus anterior (Fig. 73, i). The caudal
border covers a portion of the obliquus externus abdominis
(Fig. 68, p); the dorsal and middle portions cover parts of the
longissimus dorsi (Fig. 69, f″), the spinalis dorsi (Fig. 69, g),
the serratus posterior superior (Fig. 73, l), and serratus posterior
inferior (Fig. 73, n).
Action.—Pulls the arm caudodorsad.
M. serratus anterior and levator scapulæ (Fig. 73, i and h)
in the cat are continuous at both origin and insertion, and it is
barely possible to separate their contiguous borders without
cutting the fibres. The muscle extends from the transverse
processes of the last five cervical vertebræ and from the first
nine or ten ribs to the medial surface of the scapula near its
vertebral margin (Fig. 78, c and d).
M. serratus anterior (or serratus magnus) (Fig. 73, i).
Origin.—From the first nine or ten ribs in as many subdivisions.
The first subdivision arises from the whole or nearly
the whole of the caudal border of the first rib. The next four
or five arise either from the ribs or from the costal cartilages
near or at their junction with the ribs. The last four subdivisions
arise from the ribs at increasing distances from the cartilages.
The fibres converge and the subdivisions join one
another.
Insertion (Fig. 78, c) into a narrow area on the medial surface
of the scapula near the vertebral border. The glenoid
half of the insertion is by a short tendon, the remainder by
muscle-fibres.
Relations.—Outer (lateral) surface with the latissimus dorsi
(Fig. 68, m), teres major (Fig. 77,
c), subscapularis[123] (Fig.
77, a), and near the origin with the rectus abdominis (Fig.
73, k), the scalenus (Fig. 73, f-f‴), and the obliquus externus
(Fig. 68, p). Inner (medial) surface with the serratus posterior
superior (Fig. 73, l) and its tendon, and with the intercostales
externi (Fig. 73, m). At its insertion the muscle touches the
rhomboideus. The cranial edge is united to the caudal edge
of the levator scapulæ (Fig. 73, h).
Action.—Depressor of the scapula.
M. levator scapulæ (Fig. 73, h).
Origin.—From the dorsal tubercles of the transverse processes
of the last five cervical vertebræ and from the ligaments
between the tubercles. A slip sometimes arises from the atlas.
Insertion (Fig. 78, d) into a triangular area on the medial
surface of the scapula near its vertebral border. It is continuous
with the insertion of the serratus anterior.
Relations.—Outer (lateral) surface with a mass of fat separating
it from the clavotrapezius (Fig. 68, d) and cleidomastoid
(Fig. 65, h), and with the subscapularis (Fig. 77, a).
Inner (medial) surface with the splenius (Fig. 73, b), the tendon
of the serratus posterior superior (Fig. 73, l), and the cervical
portion of the longissimus dorsi (Fig. 69, f″). Near the insertion
the muscle touches the occipitoscapularis (Fig. 73, a) and
rhomboideus. Caudal border united with the serratus anterior
(Fig. 73, i).
Action.—Draws the scapula cranioventrad.
B. Muscles of the Back
(interconnecting parts of the
vertebral column) (Fig. 69).—The muscles connected with the
vertebral column form a mass which is less markedly differentiated
into distinct muscles than is the case in the limbs. A
great longitudinal mass of fibres begins in the sacral region and
extends along the vertebral column to the head. This mass
contains fibres running in various directions and attached at one
or both ends to the sacrum, the innominate bones, the spinous,
transverse, and articular processes, and to the arches, of the vertebræ;
to the ribs and to the head, and having in general the
function of moving the vertebræ in various ways, or of moving
parts connected with the vertebræ. In some regions, especially
the cervical, it is possible to distinguish clearly distinct muscles[124]
which have been differentiated from this mass. In the greater
part of its extent, however, it is possible to distinguish only
more or less closely interconnected bundles,—the more clearly
marked of which have received special names.
The great dorsal mass running along each side of the
vertebral column, occupying the space between the spinous
processes and the transverse processes, receives the general
name of M. extensor dorsi communis; the separate muscles
are to be considered as parts or differentiations of this. The
largest part of the mass is in the lumbar region (first portion
of the longissimus dorsi, Fig. 69, f). At the caudal end of
the thorax a lateral mass, parts of which become connected
with the ribs, is separated off from the main part of the
muscle; this extends craniad into the cervical region as
M. iliocostalis (h). The main portion of the longissimus dorsi
(f′) continues craniad on the medial side of the iliocostal into
the neck region. At about the level of the eighth or ninth
thoracic vertebra a strip begins to become separated off on the
medial side of the longissimus; further forward it becomes
clearly distinct, forming the spinalis dorsi (g). The separate
muscles connecting the cervical vertebræ with each other and
with the head are differentiations of the extensor dorsi communis.
A still deeper set of fibres, interconnecting the vertebræ,
forms the multifidus spinæ and a number of other small
muscles.
A description of the different bundles of fibres, under the
names usually given, follows. Certain general principles are
observed in some of the names used. Muscles which interconnect
the spinous processes of the vertebræ receive the name
spinalis. Those interconnecting contiguous spinous processes
are called interspinalis. Muscles attached at one end to
transverse processes, at the other to the spinous processes,
receive the name transversospinalis. Semispinalis has the
same signification as transversospinalis, but is a name usually
applied to subdivisions of the transversospinalis group. The
intertransversarii are muscles interconnecting the transverse
processes.
[125]
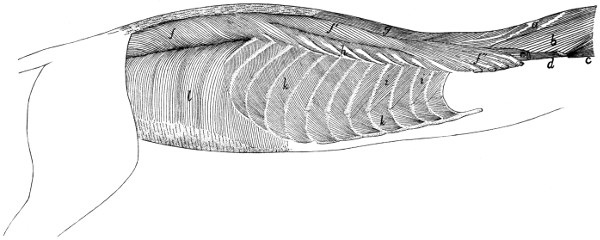
Fig. 69.—Deep
Muscles of Body, Exposed after Removal of the Muscles shown in Fig. 73.
a, M. biventer cervicis; b, M. complexus;
c, M. obliquus capitis superior; d, M. longus atlantis; e,
cut end of M. longissimus capitis; f, f′, f″,
M. longissimus dorsi; g, M. spinalis dorsi; h, M. iliocostalis; i,
Mm. intercostales externi; k, Mm. intercostales interni; l, M. transversus abdominis.
[126]
(a) Muscles of the Lumbar and Thoracic Regions.
—The
muscles of the lumbar and thoracic regions are mostly covered
by a strong fascia, known as the lumbodorsal fascia (Fig.
68, y). This consists of two sheets, the superficial sheet being
applied directly to the outer surface of the inner sheet, or separated
from it by a mass of fat.
The superficial sheet (Fig. 68, y) overlies the lumbar region
and the caudal half of the thoracic region. On the medial side
it is attached to the spinous processes of the vertebræ and is
united closely to the deeper layer. Laterally this fascia is
continuous with the latissimus dorsi (m) and obliquus abdominis
externus (p). Caudad it is attached to the spine of the ilium
and becomes continuous with the fascia covering the gluteus
muscles.
The deeper sheet (Fig. 70, c) is of a tendinous character,
forming the external tendinous layer of the longissimus dorsi,
many of whose fibres take origin from its under surface. It is
described more fully in the account of this muscle.
M. longissimus dorsi (Fig. 69, f, f′,
f″; Fig. 70, a
and b).—A very large muscle, filling most of the region
between the spinous processes and transverse processes of the
lumbar and thoracic vertebræ, and extending into the cervical
region.
The muscle is largest in the lumbar region (Fig. 69, f;
Fig. 70), where it is divided into a narrow medial (Fig. 70, a)
and a thick lateral portion (Fig. 70, b), the latter being again
partially subdivided by the fascia. The two parts unite farther
craniad.
The medial division (Fig. 70, a) consists of muscular
bundles connecting the spinous processes of the vertebræ with
the accessory and mammillary processes of other vertebræ; it
is continuous caudad with the extensor caudæ lateralis (Fig.
70, f). The muscle-fibres take origin in the sacral region by
small round tendons from the spinous processes of the last two
sacral and the caudal vertebræ. They curve cranioventrad,
forming a large belly, and are inserted into the accessory and
mammillary processes of the lumbar vertebræ. In the thoracic
region this division unites with that portion of the lateral division[127]
in which the fibres coming
from the fascia of origin are inserted
into the laminæ and transverse
processes of the thoracic
vertebræ.
The lateral division (Fig.
70, b) is much larger than the
medial one, forming in the
lumbar region a nearly cylindrical
mass.
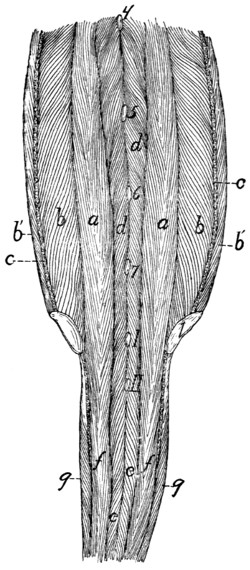
Fig. 70.—Muscles on the Dorsal
Side of the Vertebral Column
in the Lumbar, Sacral, and Caudal
Regions.
Both sheets of the lumbodorsal fascia
have been removed, the deep layer (c)
being cut where it passes into the longissimus
dorsi. 1, crest of ilium; 4-7, tips
of spinous processes of the fourth to
seventh lumbar vertebræ. I, II, tips of
spinous processes of first two sacral vertebræ.
a, b, M. longissimi dorsi (a,
medial portion; b, lateral portion; b′,
portion taking origin from the lumbodorsal
fascia); c, cut edge of deep layer
of lumbodorsal fascia; d, M. multifidus
spinæ; e, M. extensor caudæ medialis;
f, M. extensor caudæ lateralis, g, M.
abductor caudæ externus.
Origin from the crest of the
ilium (Fig. 70, 1) and the medial
surface of the ilium as far caudad
as the auricular impression; also
from the deep layer of the lumbodorsal
fascia (c). This fascia
is connected with the crest of
the ilium and with the tips of
the spinous processes of the
vertebræ in the lumbar and
thoracic regions, and from it a
large proportion of the fibres of
the longissimus take origin. In
the lumbar region it dips into
the muscle as an intermediate
longitudinal sheet (c), partially
dividing it lengthwise into two
parts. Fibres taking origin from
the lateral surface of this intermediate
sheet curve cranioventrad,
and are inserted on the
transverse processes of the lumbar
vertebræ. Fibres taking
origin from the medial surface
of the sheet pass mediocraniad
and are inserted into the accessory
processes and the surfaces
of the vertebral arches. In the thoracic region (Fig. 69,
f′)[128]
most of the fibres are inserted in separate bundles by small
tendons on the transverse processes of the vertebræ, while
some of the medial fibres unite with tendinous strands which
become attached to the laminæ and articular processes of the
vertebræ. At the region of the eighth or ninth thoracic vertebra
the spinalis dorsi (Fig. 69, g) begins to be separated off
on the medial side, the separation becoming complete only
some distance farther craniad; the longissimus dorsi then continues
into the cervical region (f″). Bundles of fibres become
attached in the manner above described to transverse processes
of all the thoracic vertebræ. In the cervical region (f″) the
muscle spreads out and becomes thinner, and bundles of fibres
become attached to the transverse processes of the cervical
vertebræ as far forward as the second. In the more cranial
portion of its extent the muscle receives fibres having origin on
the articular processes and laminæ of the cervical and a few of
the more cranial thoracic vertebræ.
The portion of the muscle which is inserted on the cervical
transverse processes (f″) is sometimes distinguished as the
longissimus cervicis; it is not well separated from the rest of
the muscle in the cat. The longissimus capitis (Fig. 69, c;
Fig. 73, g) is to be considered a differentiated cranial portion
of this muscle.
Relations.—Outer surface with the following, beginning
with the caudal end: the sartorius (Fig. 68, q), the latissimus
dorsi (Fig. 68, m), the lumbodorsal fascia (Fig. 68, y), the
serratus posterior inferior (Fig. 73, n) and superior (Fig.
73, l), and the levator scapulæ (Fig. 73, h). Lateral margin
with the abdominal muscles, the iliocostal (Fig. 69, h), and
the levator scapulæ (Fig. 73, h). Medial side with the multifidus
spinæ (Fig. 70, d), the spinous processes of the thoracic
vertebræ, the spinalis dorsi (Fig. 69, g), the complexus (Fig.
69, b), the biventer cervicis (Fig. 69, a), and the longissimus
capitis (Fig. 69, e).
Action.—Extends the vertebral column.
M. iliocostalis (Fig. 69, h).—This is a muscle about 1½
to 2 centimeters wide, lying on the dorsal portion of the ribs,
laterad of the longissimus dorsi (f′). It is composed of many[129]
partly separated bundles, each with an oblique tendon. The
muscle begins at the last or next to the last rib, where it is
connected with the longissimus by a rather small bundle of
fibres. The rest of the muscle-fibres take origin in bundles
from the lateral surface of the ribs, at about the angles, or from
thin tendons connecting the angles, of the ribs; they pass
obliquely craniad and are inserted, usually by tendons, on the
lateral surface of the third or fourth rib craniad of the one on
which the given bundle has origin. The insertions on any
given rib lie ventrad of the origins on the same rib. In the
cat this muscle is confined to the thoracic region.
Relations.—Outer surface with the serratus posterior inferior
(Fig. 73, n) and superior (Fig. 73, l), and the levator scapulæ
(Fig. 73, h). Inner surface with the dorsal surface of the ribs,
the external intercostals (Fig. 69, i), and craniad with the
levatores costarum. Medial edge in contact with the longissimus
dorsi (Fig. 69, f′).
Action.—Draws the ribs together.
M. spinalis dorsi (Fig. 69, g).—A muscle interconnecting
the spinous processes of vertebræ some distance apart, in the
thoracic and cervical regions. It lies on the medial side of the
longissimus dorsi (f′).
Origin by strong tendons from the tips of the spinous processes
of the tenth to the thirteenth thoracic vertebræ. These
tendons represent the cranial portion of the deep layer of the
lumbodorsal fascia, and give origin also to many fibres of the
longissimus dorsi, so that the two muscles are completely
united at the origin of the spinalis. They become separated at
about the level of the sixth thoracic vertebra, the spinalis forming
a strong separate bundle passing into the neck region close
against the sides of the spinous processes. The insertion is by
fleshy bundles into the sides of the spinous processes of the first
nine or ten thoracic vertebræ and of the cervical vertebræ as
far forward as the second. Some of the fibres of this muscle
pass craniad into the biventer cervicis (Fig. 69, a).
Relations.—Outer surface with the tendons of the serratus
posterior inferior (Fig. 73, n) and serratus posterior superior
(Fig. 73, l), and with the splenius (Fig. 73,
b), biventer[130]
cervicis (Fig. 69, a), and complexus (Fig. 69, b). Lateral
and ventral surface with the longissimus dorsi (f′); medial
surface with the spinous processes and the multifidus spinæ.
Action.—Extensor of the vertebral column.
M. multifidus spinæ.—This consists of bundles of fibres
which have origin on the transverse processes or neighboring
parts, pass craniodorsad across one or more vertebræ, and are
inserted into the spinous processes of vertebræ lying some distance
craniad of the origin. They lie deeper than the muscles
previously described. The muscle is most strongly developed
in the lumbar region (Fig. 70, d), where it forms a thick interwoven
mass in which it is difficult to distinguish separate
bundles. The fibres in this region have origin on the accessory
or mammillary processes and usually pass over more than one
vertebra between origin and insertion; their insertions reach
the dorsal ends of the spinous processes, so that part of the
muscle lies immediately beneath the lumbar fascia. In other
regions the multifidus is covered by other muscles. In the
thoracic region the separate bands are more distinct, and
usually pass in their course over but one intervening vertebra.
In the cervical region the bands are interconnected, forming a
fairly distinct single muscle, which is described separately
below as the semispinalis cervicis (Fig. 71, c). The portion
of this muscle attached to the head (semispinalis capitis) forms
the biventer cervicis (Fig. 69, a) and complexus (b). Caudad
this muscle passes onto the tail as the extensor caudæ medialis
(Fig. 70, c).
Relations.—Outer and lateral surface in the lumbar region
with the longissimus dorsi (Fig. 69, f) and the lumbodorsal
fascia (Fig. 68, y); in the thoracic region with the longissimus
dorsi (Fig. 69, f′) and spinalis dorsi (g). Inner surface with
the arches, articular processes, and spinous processes of the
vertebræ.
Action.—Extends the back when the muscles of both sides
work together. Turns the vertebral column obliquely sideways
when one set acts alone.
The deepest layer of the multifidus forms what is sometimes[131]
distinguished as the Mm. rotatores; no separate layer of this
sort is to be made out in the cat.
Mm. interspinales.—Muscle-bundles passing from the
spinous process of one vertebra to that of the vertebra immediately
craniad or caudad of it. They are best developed in
the lumbar region.
Mm. intertransversarii.—Muscle-fibres interconnecting
the transverse processes. In the lumbar region the intertransversarii
mediales connect the accessory and mammillary
processes; the intertransversarii laterales lying between the
transverse processes. In other regions only one set of the
intertransversarii is to be distinguished.
(b) Dorsal Muscles of the Cervical Region.
—The clavotrapezius
(Fig. 68, d) and occipitoscapularis (Fig. 73, a) have
been described in connection with the muscles of the shoulder.
The remaining muscles of this region may be considered as
differentiations of the general vertebral musculature (M. extensor
dorsi communis).
M. splenius (Fig. 73, b.)—A large sheet of muscle covering
the dorsal part of the side of the neck, beneath the trapezii.
Origin from the whole cervical ligament and from the fascia
covering the deeper muscles along a line which extends from
the first thoracic spinous process caudolaterad for about two
centimeters.
Insertion by a thin tendon into the whole lambdoidal ridge.
Laterad the tendon may be fused with that of the longissimus
capitis (Fig. 73, g).
Relations.—Outer surface with the sternomastoid (Fig.
68, c), occipitoscapularis (Fig. 73, a), clavotrapezius (Fig.
68, d), rhomboideus, tendon of the serratus posterior superior
(Fig. 73, l), and the levator scapulæ (Fig. 73, h). Lateral edge
closely united with the longissimus capitis (Fig. 73, g). Inner
surface with the longissimus capitis, complexus (Fig. 69, b),
and biventer cervicis (Fig. 69, a).
Action.—Lateral flexor of the head. The two together
elevate the head.
M. longissimus capitis (trachelomastoideus) (Fig. 73, g;
Fig. 69, e).—This is a slender muscle lying
close against the[132]
lateral border of the splenius (Fig. 73, b), and sometimes fused
with the splenius at its craniad end. It is a cranial continuation
of the longissimus dorsi (Fig. 69, f″).
Origin by five slips which are attached by strong tendons
to the anterior articular processes of the last four cervical
vertebræ. The tendons are common to this muscle and the
complexus. The five slips unite to form a flat belly which has
its
Insertion by a strong rounded tendon into the mastoid
process of the temporal bone. The tendon may be closely
united to that of the splenius.
Relations.—Lateral surface with the sternomastoid (Fig.
68, c), the levator scapulæ (Fig. 73, h), and longissimus dorsi
(Fig. 69, f″). Medial surface with the splenius (Fig. 73, b)
(to which it is partly united), the complexus (Fig. 69, b), and
the longus atlantis (Fig. 69, d).
Action.—Lateral flexor of the head.
M. biventer cervicis (Fig. 69, a) (medial portion of
M. semispinalis cervicis et capitis).—The biventer cervicis is
a flat muscle which lies beneath the splenius (Fig. 73, b), next
the median line of the neck, with its fibres longitudinal.
Origin in three or four slips from the surface of the three
or four aponeurotic arches which take origin from the tips of
the spinous processes of the last cervical and the first three
thoracic vertebræ and pass laterad and caudad to the anterior
processes of the second, third, fourth, and fifth thoracic vertebræ.
These arches are deviated from their straight course by
four aponeurotic bands which pass from them obliquely caudad
and dorsad to the sides of the spinous processes of the vertebræ
into which the arches are inserted. The arches form the
beginning of the sheet of fascia which passes outside of the
longitudinal supraspinous muscles, and the anchoring bands
form the beginning of a similar sheet which passes within the
longitudinal supraspinous muscles. An additional slip may
take origin from the cervical ligament between the last cervical
and the first thoracic vertebræ. The fibres form a flat band
divided by two or three oblique tendinous intersections. It
adheres closely to the cervical supraspinous ligament.
[133]
Insertion by a strong tendon into the medial part of the
lambdoidal crest.
Relations.—Outer surface with the splenius (Fig. 73, b) and
at the caudal end with the longissimus dorsi (Fig. 69, f″).
Inner surface with the spinalis dorsi (Fig. 69, g), the semispinalis
cervicis (Fig. 71, c), and the rectus capitis posterior
major (Fig. 71, a). Medial edge with the muscle of the opposite
side; lateral edge with the complexus (Fig. 69, b).
Action.—Raises the snout.
M. complexus (Fig. 69, b) (lateral portion of M. semispinalis
cervicis et capitis).—The complexus is closely associated
with the biventer cervicis (a) and lies on its lateral side
beneath the splenius (Fig. 73, b).
Origin by tendinous bands from the anterior articular
processes of the last five or six cervical and the first one, two,
or three thoracic vertebræ. The same bands give origin
externally to the fibres of the longissimus capitis (Fig. 69, e),
and internally to those of deeper muscles of the neck. The
six or more muscular slips continued from these tendons unite
to form a flat band, the
Insertion of which is by a flat tendon into the medial third
of the lambdoidal crest some distance ventrad of the free edge
of the crest. Near the insertion the lateral border of the
muscle is connected by an aponeurotic band to the lateral
border of the transverse process of the atlas.
Relations.—Outer surface at the dorsal border with the
biventer cervicis (Fig. 69, a); at the middle with the splenius
(Fig. 73, b); at the ventral border with the longissimus capitis
(Fig. 69, e) and the longissimus dorsi (Fig. 69, f″). Inner
surface with the cervical portion of the spinalis dorsi, with the
semispinalis cervicis (Fig. 71, c), the obliquus capitis inferior
(Fig. 71, b), the rectus capitis posterior major (Fig. 71, a), and
the obliquus superior (Fig. 71, e).
Action.—Raises the head.
M. spinalis dorsi extends into the cervical region; it has
been described.
M. semispinalis cervicis (Fig. 71, c).—This represents
that portion of the multifidus spinæ which extends into the[134]
cervical region. It is not composed of distinct bundles, like
the multifidus of the thoracic region, but all the fibres are
united into a fairly well-defined muscle.
Origin from the articular processes of the last five cervical
vertebræ.
Insertion into the spinous processes of the cervical vertebræ
up to the second, the largest part of the muscle being inserted
into the caudal end of the spine of the atlas.
Relations.—Outer surface with the spinalis dorsi (Fig.
69, g) and the complexus (Fig. 69, b). Inner surface with
the vertebræ.
Action.—Extends the neck.
M. longissimus dorsi extends into the cervical region,
where it is often distinguished as the longissimus cervicis
(Fig. 69, f″). It has been described.
M. longus atlantis (Fig. 71, f; Fig. 69, d).—This represents
a differentiated portion of the longissimus dorsi.
Origin from the transverse process and the side of the
vertebral arch of the third cervical vertebra.
Insertion into the caudolateral angle of the wing of the
atlas.
Relations.—Dorsal surface with the complexus (Fig. 69, b),
the longissimus capitis (Fig. 69, e), and the obliquus inferior
(Fig. 71, b). Ventral surface with the scalenus (Fig. 71, g),
longissimus dorsi (Fig. 69, f″), and levator scapulæ (Fig.
71, h).
Action.—Extends the neck and turns the head sideways.
M. rectus capitis posterior major (Fig. 71, a).
Origin.—The whole length of the spinous process (or
crest) of the axis, or its caudal part only. It is united by a
raphe to the opposite muscle. The muscle passes craniad and
laterad to its
Insertion into an area ventrad of the medial part of the
lambdoidal crest beneath the insertion of the complexus (Fig.
69, b) and the biventer cervicis (Fig. 69, a).
Relations.—Dorsal surface with the biventer cervicis (Fig.
69, a). Ventral surface with the rectus capitis posterior medius
(Fig. 71, d) and the obliquus capitis inferior.
[135]
Action.—Raises the snout.
M. rectus capitis posterior medius (Fig. 71, d). (This
is in man a part of the rectus capitis posterior major.)—It lies
beneath the rectus capitis posterior major (Fig. 71, a).
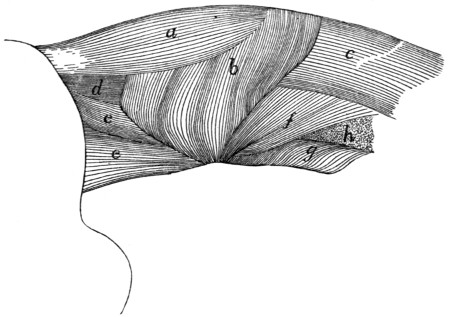
Fig. 71.—Deep
Muscles of the Neck, after the Removal of Mm. Biventer
Cervicis and Complexus.
a, M. rectus capitis posterior major; b, M. obliquus capitis inferior; c, M. semispinalis
cervicis; d, M. rectus capitis posterior medius; e, M. obliquus capitis superior;
f, M. longus atlantis; g, M. scalenus (part of cervical portion); h, cut end of M.
levator scapulæ.
Origin from the cranial end of the axial spine. It forms a
triangular prismatic mass which passes craniad and has its
Insertion into an elongated area on the occipital bone,
ventrad of the median half of the lambdoidal crest.
Relations.—Dorsal surface with the rectus capitis posterior
major (Fig. 71, a). Ventral surface with the atlas and the
rectus capitis posterior minor.
Action.—Assists the rectus capitis posterior major.
M. rectus capitis posterior minor.—A small triangular
muscle beneath the rectus capitis posterior medius (Fig. 71, d).
Origin by the apex of the triangle from the cranial border
of the dorsal arch of the atlas for about two millimeters laterad
of the median line. The muscle passes over the ligament
which connects the atlas to the occiput and adheres to it.
Insertion into an elongated area on the occipital bone,[136]
ventrad of the insertion of the rectus capitis posterior medius
for about five millimeters next to the median line.
Relations.—Dorsal surface with the rectus capitis posterior
medius. Ventral surface with the posterior atlanto-occipital
membrane.
Action.—Assists the rectus capitis posterior major.
M. obliquus inferior (Fig. 71, b).
Origin from the whole lateral surface of the spine of the
axis. The parallel fibres form a thick mass which passes
laterad and craniad.
Insertion into the dorsal surface of the transverse process
of the atlas along a narrow area near its lateral margin.
Relations.—Outer surface with the rectus capitis posterior
major (Fig. 71, a), the complexus (Fig. 69, b), and the
obliquus superior (Fig. 71, e). Inner surface with the axis.
Action.—Rotates the head.
M. obliquus capitis superior (Fig. 71, e; Fig. 72, e).—This
is a triangular muscle passing from the outer border of the
atlas laterad of the recti capitis posteriores to the occiput.
Origin from the lateral border of the transverse process of
the atlas on its ventral margin. The fibres pass craniad and
diverge.
Insertion into the caudal side of the mastoid process of the
temporal bone and into a line parallel with the lambdoidal
ridge and ventrad of it, from the mastoid process to within one
centimeter of the median line.
Relations.—Outer surface with the splenius (Fig. 73, b),
complexus (Fig. 69, b), and longissimus capitis (Fig. 73, g).
Inner surface with the obliquus inferior (Fig. 71, b), the wing
of the atlas, and the rectus lateralis (Fig. 72, d). Ventral
border with the cleidomastoid (Fig. 73, d) and one head of the
levator scapulæ ventralis (Fig. 72, c′).
Action.—Flexes the head laterally.
Mm. interspinales and intertransversarii are found in the
neck region also; they are described with the muscles of the
back.
C. Muscles of the Tail.
—M. extensor caudæ medialis
(Fig. 70, e).—This is a continuation caudad of
the multifidus[137]
spinæ (Fig. 70, d); it lies next to the dorsal median line, the
muscles of right and left side touching one another in the
middle line.
Origin by numerous fleshy bundles from the spinous
processes of the sacral and first caudal vertebræ. The fibres
pass caudad and are inserted by tendons into the articular
processes and the dorsal surface of the caudal vertebræ.
Action.—Extends (raises) the tail.
M. extensor caudæ lateralis (Fig. 70, f).—This is a continuation
caudad of the medial portion of the longissimus dorsi
(Fig. 70, a); it lies just laterad of the extensor caudæ medialis
(Fig. 70, e).
Origin in many fleshy bundles from the articular processes
of the sacral vertebræ, and the transverse processes of the
caudal vertebræ. The fibres curve dorsocaudad and are
inserted by many long slender tendons on the dorsal surfaces
of the caudal vertebræ. The muscle grows continually smaller
as it passes caudad.
Action.—Raises the tail.
M. abductor caudæ (coccygis) externus (Fig. 68, w; Fig.
70, g).—A rounded muscle on the lateral surface of the cranial
part of the tail.
Origin from the medial side of the dorsal border of the
ilium, and from the dorsal surface of the sacrum. The muscle
passes caudad, lying just ventrad of the extensor lateralis, and
is inserted into the transverse processes and lateral surfaces of
the caudal vertebræ, as far back as the eighth or ninth.
Action.—Bends the tail sideways.
M. abductor caudæ internus (or M. coccygeus).—A large
flat muscle, having origin on the spine of the ischium. The
muscle passes dorsomediad, spreading out, and is inserted into
the transverse processes of the second to the fourth caudal
vertebræ.
Relations.—Lateral surface with the caudofemoralis (Fig.
68, s), gluteus maximus, and pyriformis. Medial surface with
the iliocaudalis and the flexor caudæ longus (Fig. 68, x).
M. iliocaudalis (Str.-D.) (Fig. 162,
11′, page 398).—This
represents a portion of the levator ani of man, and in some[138]
specimens is united with that muscle (Fig. 162, 11) in the
cat.
Origin along the ventral half of the medial surface of the
ilium, caudad of the sacrum. The fibres pass caudad and are
inserted by a flat tendon into the ventral surface of the caudal
vertebræ, from the second or third to about the seventh.
Relations.—Lateral surface with the gluteus maximus
and abductor caudæ internus (Fig. 68, w). Medial surface with
the levator ani (Fig. 162, 11), with which this muscle may be
partly united.
Action.—The two muscles together flex the tail.
M. flexor caudæ longus (Fig. 162,
12; Fig. 113, c, page
270).
Origin on the ventral surface of the last lumbar vertebra,
of the sacrum, and of the transverse processes of the caudal
vertebræ. Caudad the muscle forms long, strong tendons
which are inserted on the ventral surface of the tail.
Relations.—Lateral surface with the ilium, the gluteus
maximus, the abductor caudæ internus, the iliocaudalis, and the
skin of the tail. Dorsally it touches in the caudal region the
abductor caudæ externus (Fig. 68, w), medially the flexor caudæ
brevis (Fig. 162, 13).
Action.—Flexes the tail.
M. flexor caudæ brevis (Fig. 162, 13).
Origin on the ventral surface of the caudal vertebræ, from
the first to the seventh or eighth. The principal head comes
from the first vertebra. The bundles thus formed pass caudad
and are inserted each into the ventral surface of a vertebra
some distance caudad of the origin. The muscle extends to
about the tenth caudal vertebra, and its most caudal parts are
closely interconnected with the flexor caudæ longus.
Relations.—Lateral edge with the flexor caudæ longus.
Medial edge in the sacral region with the levator ani (Fig. 162,
11) and the iliocaudalis (Fig. 162,
11′); in the caudal region
with the muscle of the opposite side.
Action.—Flexes the tail.
2. Muscles on the Ventral Side of the Vertebral Column.
A. Lumbar and Thoracic Regions.
—The iliopsoas[139] (Fig.
162, 8) belongs in this group, but since it moves the leg it is
described with the muscles of the pelvic limbs.
M. psoas minor (Fig. 162, 9,
page 398).—This muscle lies
along the ventral surface of the vertebral column, next to the
peritoneum, and extends from the thoracic vertebræ to the
ilium.
Origin usually by five heads, from the caudal border of the
centra of the last two (or one) thoracic and first three (or four)
lumbar vertebræ. These heads also serve as origin for a part
of the iliopsoas, and are closely united at their origin with the
quadratus lumborum. They unite to form an, at first, rather
large flat muscle, which rapidly becomes smaller caudad,
finally forming a slender tendon which is inserted on the iliopectineal
line, just craniad of the acetabulum.
Relations.—Dorsolateral surface with the quadratus lumborum
and iliopsoas (Fig. 162, 8). Ventral surface with the
pleura and diaphragm near the origin; with the peritoneum
caudad.
Action.—Flexes the back in the lumbar region.
M. quadratus lumborum.—A flat muscle lying on the
ventral surface of the transverse processes of the lumbar vertebræ.
Craniad it has origin on the ventral surface of the last
two thoracic vertebræ, and by a few fibres from the last rib.
The muscle passes caudad, lying on the lateral side of the
psoas minor (Fig. 162, 9), and becomes attached to each of the
transverse processes of the lumbar vertebræ. Caudad the
muscle passes into a strong flat tendon which is inserted into
the anterior inferior spine of the ilium.
Relations.—Dorsal surface with the transverse processes
and the intertransverse muscles. Ventral and medial surfaces
with the psoas minor (Fig. 162, 9) and
iliopsoas (Fig. 162, 8).
Action.—Bends the vertebral column sideways.
B. Muscles on the Ventral Side of the Neck
(Fig. 65, page 109).—M.
sternomastoideus (Fig. 65, g; Fig.
68, c).—The sternomastoid (sternal portion of the human
sternocleidomastoid) is a flat band one to three centimeters
wide extending from the cranial end of the manubrium and the
midventral line craniad of it to the lambdoidal ridge.
[140]
The origin extends from the cranial end of the manubrium
along the midventral line as far as the caudal border of the
cricoid cartilage, and is in two parts. The caudal portion
arises from the lateral surface of the manubrium along the
dorsal half of its cranial end, and from the median raphe for
about one centimeter craniad of the manubrium. Its fibres are
parallel and tend to diverge into two layers. The cranial
portion arises by means of fibres which cover the median line
and interdigitate with the fibres of the opposite muscle between
the cricoid cartilage and a point a few millimeters craniad of
the manubrium. Its caudal border is thus overlaid by the
caudal portion. Its fibres converge and join those of the
caudal portion. The muscle passes dorsocraniad and is
Inserted by means of a flat tendon into the lateral half of
the lambdoidal ridge and into a continuation of the ridge onto
the mastoid portion of the temporal bone as far as the mastoid
process. The thickest part of the tendon is inserted into the
mastoid process.
Relations.—Outer surface at the caudal end with the pectoralis
major (Fig. 65, l); in the middle part with the platysma,
the depressor conchæ (Fig. 64, b), and with the external jugular
vein (Fig. 65, 5), which crosses it obliquely; at the cranial
end with the submaxillary (Fig. 65, 2)
and parotid (1) glands.
Inner surface with the sternohyoid (Fig. 65, e), sternothyroid
(g′), internal jugular vein, longus capitis muscle, the cleidomastoid
(h), levator scapulæ ventralis (i), a large lymphatic
gland ventrad of the ear, and the splenius (Fig. 73, b).
Action.—One muscle turns the head and depresses the
snout. Both together depress the snout.
M. sternohyoideus (Fig. 65, e).—A slender muscle on the
midventral line of the neck close to the opposite muscle.
Origin from the cranial border of the first costal cartilage.
The muscle passes craniad closely united near its caudal end
to the sternothyroid (g′).
Insertion (Fig. 67,
m) into the outer half of the ventral surface
of the body of the hyoid bone caudad of the origin of the
geniohyoid (Fig. 67, g).
Relations.—Outer surface with
the sternomastoid[141] (Fig.
65, g). Inner surface with the thyrohyoid (Fig. 65, f; Fig.
67, p), the trachea (Fig. 67, 4), and the larynx.
Action.—Draws the hyoid caudad. Raises the ribs and
sternum when the hyoid is fixed.
M. sternothyreoideus (Fig. 65, g′).—The sternothyroid
lies beneath the sternohyoid and is connected with it at its
caudal end.
Origin from the first costal cartilage beneath the sternohyoid.
Insertion (Fig. 67, o) into the lateral part of the caudal
border of the thyroid cartilage of the larynx.
Relations.—Outer surface with
the sternohyoid (Fig. 65, e)
and laterally with the sternomastoid (Fig. 65, g). Inner surface
with the trachea (Fig. 67, 4), and at the lateral edge with
the thyroid gland (Fig. 67, 6).
Action.—Pulls the larynx caudad.
M. scalenus (Fig. 73, f-f‴).—This is a large and complex
muscle lying on the ventral side of the neck and the
lateral surface of the thorax. It might equally well be classified
with the muscles of the thorax. It is divisible into a considerable
number of interconnected bundles which are sometimes
described as separate muscles.
The largest, middle portion (scalenus medius) (f′) takes
origin by thin tendons from the sixth, seventh, eighth, and
ninth ribs, just dorsad of their junction with the cartilages;
these tendons unite near their origins to form a flat band which
becomes fleshy at about the fifth rib, passes craniad, and unites
just craniad of the first rib with the other parts of the muscle.
The dorsal portion (scalenus posterior) (f″) has origin by a
very slender tendon from about the middle of the outer surface
of the third or fourth rib; it extends craniad as a narrow band
which passes laterad between the heads of the serratus anterior
(i) that are attached to the second and third (or third and
fourth) ribs, and unites with the other heads. The ventral
head (part of scalenus anterior) (f‴) arises by one or two
minute tendons from the cartilages of the second and third ribs;
they are partly united with the transversus costarum (j). This
ventral head passes craniad and unites with the other heads.
[142]
These three divisions unite just craniad of the first rib with
each other and with the cervical portion of the muscle (f)
(part of scalenus anterior). This consists of a number of
small bundles of fibres which arise from the first rib and the
transverse processes of the first thoracic and the last six cervical
vertebræ.
The insertion is onto the transverse processes of all the cervical
vertebræ, including the axis and atlas. According to Strauss-Durckheim
the fibres from any given transverse process are inserted
into the transverse processes of all the vertebræ craniad of it.
The muscle is partly continuous craniad with the longus
capitis (e). The cervical nerves pass out between the bundles
of the cervical portion.
Relations.—Outer (lateral) surface in the thoracic region
with the external oblique (Fig. 68, p), the pectoralis minor
(Fig. 65, o), and in part with the serratus anterior (Fig. 73, i);
in the cervical region with the sternomastoid (Fig. 65, g).
Inner (medial) surface with the serratus anterior (Fig. 73, i),
the levator scapulæ (Fig. 73, h), the longus atlantis, the transverse
processes of the cervical vertebræ, and the longus capitis
(Fig. 73, e).
Action.—Flexes the neck and draws the ribs craniad.
M. longus capitis (or rectus capitis anterior major) (Fig.
72, a; Fig. 73, e).—This is a long muscle lying on the ventral
aspect of the cervical vertebræ. With the muscle of the
opposite side and the underlying longus colli (Fig. 72, g′) it
forms a trough in which lie the œsophagus, pharynx, and
trachea.
Origin by five (or six) heads from the ventral margins of
the transverse processes of the cervical vertebræ from the
second to the sixth inclusive. The heads unite into a common
belly, the lateral border of which is united with the levator
scapulæ ventralis (Fig. 72, c) near its insertion.
Insertion into the body of the sphenoid bone between the
bulla and the middle line. The insertion extends craniad
onto the basisphenoid.
Relations.—Ventral surface with the sternomastoid (Fig.
65, g) and the large lymphatic gland ventrad of the external[143]
ear. Lateral surface with the scalenus (Fig. 72, h) and craniad
with the levator scapulæ ventralis
(Fig. 72, c-c″). Medial
surface with the carotid artery,
the trachea, œsophagus, and
pharynx. Dorsal edge with
the vertebræ, the longus colli
(Fig. 72, g′), and the rectus capitis
anterior minor (Fig. 72, b).
Action.—Lowers the snout.
M. rectus capitis anterior
minor (Fig. 72, b).—This is a
flat band beneath (dorsad of)
the longus capitis (a).
Origin from the ventral surface
of the inferior arch (body)
of the atlas (3) for about five
millimeters laterad of the middle
line.
Insertion into a deep depression
on the basioccipital caudad
of the insertion of the longus
capitis (a) and between the
caudal end of the bulla (1) and
the median line.
Relations.—Ventral surface
with the longus capitis (a).
Dorsal surface with the anterior
(ventral) atlanto-occipital membrane
(2).
Action.—Depresses the
snout.
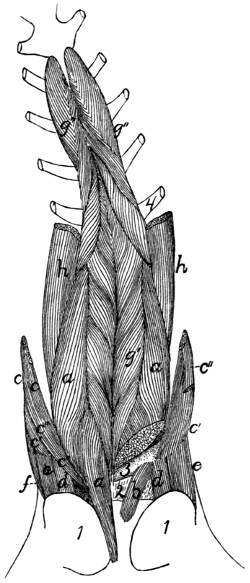
Fig. 72.—Muscles on the Ventral
Surface of the Cervical Vertebræ.
On the left side the longus capitis and
one head of the levator scapulæ ventralis
have been cut, in order to show the rectus
capitis anterior minor. a, M. longus
capitis; b, M. rectus capitis anterior
minor; c, M. levator scapulæ ventralis
(c′, its atlantal head; c″, its occipital
head); d, M. rectus capitis lateralis; e,
M. obliquus capitis superior; f, M. longissimus
capitis; g, M. longus colli (g′, its
cervical portion; g″, its thoracic portion);
h, part of M. scalenus. 1, bulla
tympani; 2, ventral atlanto-occipital
membrane; 3, ventral arch of atlas; 4,
first rib.
M. rectus capitis lateralis (Fig. 72, d).—This lies ventrad
of the obliquus superior (e) on
the ventral face of the transverse
process of the atlas and at first
appears to be a part of the obliquus superior.
[144]
Origin from the median half of the ventral surface of the
transverse process of the atlas. The fibres form a cylindrical
mass which passes craniad and slightly laterad.
Insertion into the fossa laterad of the condyle of the occipital
bone.
Relations.—Ventral surface with the digastric (Fig. 65, b)
and the lymphatic gland ventrad of the ear. Dorsal surface
with the wing of the atlas.
Action.—Flexes the head laterally.
M. longus colli (Fig. 65, g′, g″).—A long, rather slender
muscle lying on the ventral surface of the cervical and first six
thoracic vertebræ. It consists of many separate bundles, and
is divisible into a thoracic and a cervical portion.
The thoracic portion (g″) has origin by six heads from the
ventral surface of the first six thoracic vertebræ. The separate
heads join to form a band which passes craniad and is inserted
for the most part into the processus costarius of the sixth
cervical vertebra; a portion of it joins the cervical division of
the muscle.
The cervical portion (g′) is a continuation craniad of the
thoracic portion. It arises in small bundles from the transverse
processes and sides of the ventral surfaces of the centra of the
cervical vertebræ. These bundles pass craniomediad, so that
those from the muscles of opposite sides meet and are inserted
on the centra of the vertebræ in the middle line, each pair of
bundles forming a V opening caudad. The most cranial
insertion is into the tubercle on the middle of the ventral surface
of the ventral arch of the atlas (3).
Relations.—Dorsal surface with the centra of the vertebræ.
Ventral surface in the thorax with the trachea; in the neck
with the œsophagus. Medial edge with the muscle of the
opposite side. Lateral margin in the cervical region with the
longus capitis (a).
Action.—Bends the neck.
3. Muscles of the Thorax.
A. Breast-muscles (connecting the arm and thorax)
(Fig. 65, page 109).
The muscles connecting the arm with the sternum, corresponding
to the pectoralis muscles of man, fall into a considerable[145]
number of not always clearly separated bundles in the cat.
We shall distinguish by name four such bundles; these might
be, and frequently are, further subdivided.
M. pectoantibrachialis (Fig. 65, m).—This is a small flat
bundle about one or two centimeters wide, forming the most
superficial portion of the pectoralis group.
Origin on the lateral surface of the manubrium. The
muscle passes laterad onto the arm and is inserted by a flat
tendon into the superficial fascia of the dorsal border of the
forearm, near the elbow-joint. The tendon is continuous
craniad with the border of the clavobrachial (k), so that a bundle
of fibres may pass from this muscle to be inserted with the
clavobrachial, or a bundle from the clavobrachial to be inserted
with this. The tendon is continuous caudad with that of the
epitrochlearis (r). The fascia of insertion may be traced to an
attachment into the dorsal border of the ulna. This muscle
usually receives near its insertion a slip, sometimes called its
caudal division, from the deep layer of the pectoralis major.
Relations.—Outer surface with the integument. Inner surface
with both divisions of the pectoralis major (l) on its medial
two-thirds, and with the biceps (t) and bicipital arch (t′) in its
lateral one-third. The cranial border is closely applied to that
of the clavobrachial (k), except at the two ends.
Action.—Draws the arm mediad.
M. pectoralis major, or ectopectoralis (Fig. 65, l, l′).—In
this it is usually possible to distinguish a superficial and a
deep layer.
The superficial (l′) layer is a flat band of fibres one or two
centimeters wide.
Origin from a raphe in the midventral line, along the
cranial half of the manubrium and for five to ten millimeters
craniad of it.
Insertion (Fig. 81, f).—The pectoral ridge of the middle
third of the shaft of the humerus, slightly dorsad of the line of
insertion of the deep portion.
Relations.—Outer surface with the clavobrachial (Fig.
65, k) and integument, the medial end caudally with the
pectoantibrachialis (m), and the distal end with the brachialis[146]
(v). Inner surface with the deep portion (l), the fibres of
which it crosses obliquely.
The deep portion (l) is a flat band of parallel fibres about
three times as broad as the superficial portion.
Origin.—The ventral surface of the manubrium and of the
first three divisions of the sternum, and the median raphe for
about one centimeter craniad of the manubrium. It passes
directly laterad.
Insertion (Fig. 81, e) along a line which begins at the
infraspinatus fossa of the great tuberosity, and runs parallel to
the deltoid ridge until it reaches the pectoral ridge at the junction
of the second and third fourths of the humerus and then
continues in the direction of the pectoral ridge as far as the
junction of the third and fourth fourths of the humerus. Some
of its fibres may pass caudad of the biceps and be inserted with
the epitrochlearis. Into the part of the line which is parallel
to the deltoid ridge the muscle is inserted by a flat tendon;
into the remainder of the line it is usually inserted directly by
muscle-fibres, though at its caudal end its insertion may again
be tendinous.
Relations.—Outer surface with the cutaneus maximus, the
outer layer of the pectoralis major (Fig. 65, l′), the pectoantibrachialis
(m), and the clavobrachial (k). Inner surface with
the pectoralis minor (o), the proximal end of the humerus, the
coracobrachialis (Fig. 77, f), and the supraspinatus (Fig.
77, d) at the insertion of the latter. At the cranial part of its
origin it touches the sternomastoid (Fig. 65, g) and sternohyoid
(Fig. 65, e).
Action.—Draws the arm mediad and turns the foot forward.
M. pectoralis minor, or entopectoralis (Fig. 65, o).—This
is a fan-shaped mass of fibres, flat but thicker than the pectoralis
major.
Origin from the lateral half of the first six divisions of the
body of the sternum and sometimes from the xiphoid process.
The fibres are divisible into several bundles which vary in
extent and distinctness and are sometimes described as separate
muscles. They pass craniolaterad and converge so that the line
of insertion is about one-half as long as that of the origin.
[147]
Insertion (Fig. 81, d) into the humerus along a line which
forms the ventral border of the bicipital groove at the proximal
end of the bone, passes thence onto the pectoral ridge and
continues in the direction of the pectoral ridge nearly as far as
the middle of the length of the humerus. The cranial and
caudal fibres are inserted by means of a thin tendon; the
middle fibres directly. The cranial border of the tendon of
insertion is continued as a thin tendon to the coracoid process;
it is closely united with the insertion of the supraspinatus.
Relations.—By its outer surface with the pectoralis major
(Fig. 65, l) over its cranial half; with the cutaneus maximus,
latissimus dorsi (Fig. 65, q), and integument over its caudal
half. By its inner surface with the xiphihumeralis (Fig. 65, p),
rectus abdominis (Fig. 73, k), and a mass of fat in the axilla.
The tendon of the xiphihumeralis is also connected with its
inner surface near its insertion. Its cranial border is free. Its
caudal border is in relation at its medial end with the xiphihumeralis,
at its lateral end with the tendon of the latissimus
dorsi (Fig. 65, q).
Action.—Draws the arm toward the middle line.
M. xiphihumeralis (Fig. 65, q).—A long, thin, narrow
muscle which may be considered as part of the pectoralis
minor.
Origin a median raphe along the xiphoid process or at an
angle to the median line on the rectus abdominis muscle. It
passes craniad, becoming gradually smaller, and about two
centimeters from its insertion ends in a thin tendon.
Insertion.—It ends in a flat tendon which in passing is connected
by a strong fascia with the tendon of the latissimus
(Fig. 65, q). It passes along the inner surface of the pectoralis
minor (o) to be inserted with its cranial fibres near the ventral
border of the bicipital groove.
Relations.—Outer surface with the cutaneus maximus at
its medial end, with the pectoralis minor (o) at its distal end.
Inner surface with the external oblique (Fig. 68, p), the rectus
abdominis (Fig. 73, k), and a mass of fat in the axilla. Borders
free. Some of the fibres of the cutaneus maximus are attached
to its outer surface.
[148]
Action.—Assists the pectoralis minor.
B. Muscles of the Wall of the Thorax.
—M. serratus
posterior superior (Fig. 73, l).—A thin sheet of muscle
and tendon beneath the serratus anterior (i) on the dorsal part
of the thorax and neck.
Origin by fleshy slips from the outer surfaces of the first
nine ribs just ventrad of their angles. The origin may extend
as far as the tenth or eleventh ribs. The first slip is usually
delicate. The fibres are directed dorsad and craniad and unite
to form a continuous sheet. This ends along a longitudinal
line opposite the vertebral transverse processes in the aponeurosis
which covers the longissimus dorsi and other muscles in this
region. Its insertion is into the median dorsal raphe between
the axial spinous process and the tenth thoracic spinous
process.
Relations.—Outer surface with the serratus anterior (i), the
latissimus dorsi (Fig. 68, m), and the rhomboideus. Inner
surface with the external intercostals (Fig. 73, m), the iliocostal
(Fig. 69, h), the longissimus dorsi (Fig. 69, f), the spinalis
dorsi (Fig. 69, g), and the splenius (Fig. 73, b).
Action.—Draws the ribs craniad.
M. serratus posterior inferior (Fig. 73, n).—The serratus
posterior inferior is a thin muscle lying caudad of the serratus
posterior superior (l) and sometimes overlying the caudal end
of the latter.
Origin by four or five heads from the last four or five ribs.
In some specimens only three heads may be present. The
separate bundles pass dorsocaudad and unite to form a continuous
sheet which ends in a continuation of the aponeurosis of
the serratus posterior superior (l).
Insertion into the lumbar spinous processes and the intervening
interspinous ligaments.
Relations.—Outer surface with the latissimus dorsi (Fig.
68, p), and at the origins with the intercostales externi (Fig.
73, m). Inner surface with the intercostales interni (Fig. 69, k)
at the origin and with the intercostales externi (Fig. 69, i),
iliocostal (Fig. 69, h), longissimus dorsi (Fig. 69, f), and the
caudal end of the spinalis dorsi (Fig. 69, g).
[149]
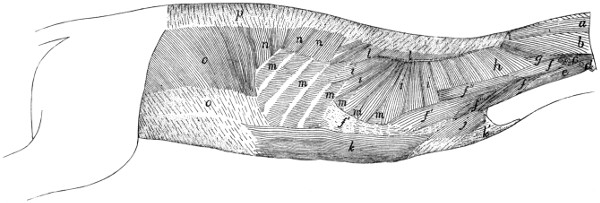
Fig. 73.—Body
Muscles exposed after Removal of the Forelimb and the Muscles shown in Fig. 68.
a, M. occipitoscapularis (cut); b, M. splenius; c,
M. levator scapulæ ventralis (cut); d, M. cleidomastoideus (cut); e, M. longus capitis; f,
M. scalenus (f′, middle division; f″, dorsal division; f‴,
ventral division; f, cervical portion); g, M. longissimus capitis; h, M. levator
scapulæ (cut); i, M. serratus anterior (cut); j, M. transversus costarum; k,
M. rectus abdominis (k′, its tendon); l, M. serratus posterior
superior; m, M. intercostales externi; n, M. serratus posterior inferior; o,
M. obliquus abdominis internus; p, lumbodorsal fascia.
[150]
Action.—Assists the serratus posterior superior.
M. transversus costarum, or sternocostalis externus (Fig. 73,
j).—This is a thin muscle applied to the cranial part
of the side of the thorax, covering the cranial end of the rectus
abdominis (k).
Origin by tendon from the side of the sternum between the
attachments of the third and sixth ribs. The muscle passes
dorsocraniad to its
Insertion on the first rib and the lateral portion of its costal
cartilage.
Relations.—Outer surface with the pectoralis minor (Fig.
65, o). Inner surface with the scalenus (Fig. 73, f‴), the
rectus abdominis (Fig. 73, k), and the intercostales externi
(Fig. 73, m). At its insertion it is united with a portion of the
scalenus (Fig. 73, f‴).
Action.—Draws the sternum forward.
Mm. levatores costarum.—Small muscles having origin
on the transverse processes of the thoracic vertebræ, passing
caudoventrad, and becoming inserted on the angle of the rib
lying immediately caudad of the origin. They are continuous
with the external intercostals.
Relations.—Outer surface with the longissimus dorsi (Fig.
69, f), and craniad with the iliocostal (Fig. 69, h). Inner
surface with the internal intercostals (Fig. 69, k).
Action.—Pull the ribs dorsocraniad.
Mm. intercostales externi (Fig. 73, m; Fig. 69, i).—The
external intercostals are placed in the outer portion of the
intercostal spaces. They are composed of bundles of fibres
attached by their ends to the adjacent borders of the ribs and
having in general the direction of the external oblique muscle,
i.e., they pass from their cranial ends caudoventrad. They
occupy the intercostal spaces between the true ribs and extend
even caudad into the spaces between the false ribs. They are
lacking between the ventral ends of the costal cartilages of the
first six to eight ribs, so that the internal intercostals (Fig.
69, k) are here exposed. The more caudal external intercostals
are more nearly craniocaudal in direction.
Relations.—Outer surface with obliquus abdominis externus[151]
(Fig. 68, p), latissimus dorsi (Fig. 68, m), serratus posterior
inferior (Fig. 73, n) and superior (Fig. 73, l), serratus anterior
(Fig. 73, i), scalenus (Fig. 73,
f), and iliocostal (Fig. 69, h).
Inner surface with the internal intercostals (Fig. 69, k).
Action.—Protractors of the ribs.
Mm. intercostales interni (Fig. 69, k).—The internal
intercostals are similar to the external intercostals, beneath
which they lie. Their fibres pass between the ribs at nearly
right angles to those of the external intercostals and have nearly
the direction of the fibres of the internal oblique. They occupy
all the intercostal spaces from the first to the thirteenth ribs.
Relations.—Outer surface with the external intercostals
(Fig. 69, i), and ventrad with the scalenus (Fig. 73, f), transversus
costarum (Fig. 73, j), and rectus abdominis (Fig.
73, k). Inner surface with the pleura and the transversus
thoracis.
Action.—Retractors of the ribs.
M. transversus thoracis (triangularis sterni; sternocostalis
internus).—This represents a thoracic portion of the transversus
abdominis. It consists of five or six flat muscular bands lying
on the inner surface of the thoracic wall.
Origin on the lateral borders of the dorsal face of the
sternum, opposite the attachments of the cartilages of the third
to the eighth ribs. The six bands thus formed, each about one
centimeter wide, pass laterad and are inserted into the cartilages
of the ribs near their junction with the ribs, and into the fascia
which covers the inner surface of the internal intercostals in this
region.
Relations.—Outer surface with the internal intercostals and
the cartilages of the ribs. Inner surface with the pleura.
Diaphragma (Fig. 74).—The diaphragm consists of a central
so-called semilunar tendon (e) and of muscular fibres which
pass radially from the body wall to the tendon. It forms a
complete oblique partition between the abdominal and thoracic
cavities. The dorsal end is farther caudad than the ventral.
The central tendon (e) is thin and irregularly crescent-shaped,
with the convexity ventrad and the horns of the crescent
prolonged as two tendinous bands (e′) which end in two[152]
triangular membranous portions (d) of the diaphragm, one on
each side of the spinal column. It is pierced by an opening
for the vena cava (f).
Fig. 74.—Diaphragm,
Caudal Surface seen Obliquely from the Right.
a, right crus; b, left crus; c,
c′, sternocostal part of diaphragm; d, d′, membranous
portions of the diaphragm; e, central tendon; e′, prolongations of central tendon;
f, opening for posterior vena cava; g, œsophagus; h, aorta; i, M. transversus
abdominis.
The muscular portion is in two parts: (a) The vertebral
portion (a, b) arises by a single tendon from the ventral surface
of the second, third, and fourth lumbar centra. The tendon
diverges into two, the right one (a) of which is much stronger,
and from each of the two arise muscle-fibres. Each mass of
fibres is one of the two crura (sing. crus) of the diaphragm. The
aorta (h) enters the abdomen between the crura dorsally. The
fibres of each crus diverge to be inserted into the central tendon
and the dorsal continuation of its horn. The two sets of fibres
unite ventrad of the opening of the aorta. Between this opening
and the central tendon is another opening for the œsophagus[153]
(g); this lies entirely in that part of the diaphragm which
arises from the right crus (a). The fibres are again united
ventrad of the œsophageal opening.
(b) The sternocostal part (c, c′) takes origin from the
xiphoid process and the last five ribs, by fleshy bundles which
interdigitate with those of the transversus abdominis (i). The
fibres converge to the central tendon (e). Between the crus
of each side and the most dorsal of the costal fibres is the
membranous interval (d) mentioned above.
4. Abdominal Muscles.
M. obliquus abdominis externus
(Fig. 68, p).—A large, thin sheet of muscle covering the whole
abdomen and part of the thorax ventrally.
Origin.—(a) From the last nine or ten ribs by means of as
many tendons, which are interconnected to form arches that
span the slips of the serratus anterior. The muscle-fibres
arise from these tendons and from their intervening arches.
(b) From the lumbodorsal aponeurosis common to it and the
internal oblique. The cranial fibres pass nearly ventrad, the
caudal fibres caudad, and the intervening fibres take an intermediate
course. The fibres end in a thin aponeurosis of insertion
along a curved line which passes at first caudad and then
laterodorsad. The aponeurosis fibres continue in the direction
of the muscle-fibres to the
Insertion into the median raphe ventrad of the sternum from
the insertion of the seventh costal cartilage to the xiphoid
process, into the linea alba from the sternum to the pubic
tubercle, and into the tubercle and the cranial border of the
pubis. Caudad of the xiphoid process the aponeurosis is closely
united to the superficial layer of the internal oblique, where it
forms the outer layer of the sheath of the rectus abdominis.
Laterad of the pubic tubercle the tendon is perforated by the
inguinal canal. In the cat neither the caudal part of the muscle
nor its tendon is attached to the ilium, as it is in man and the
dog, so that no Poupart’s ligament, or inguinal ligament, is
formed.
Relations.—Outer surface with the cutaneus maximus,
the integument, and near the origin with the latissimus dorsi
(Fig. 68, m). Inner surface with the obliquus internus[154] (Fig.
73, o), the rectus abdominis (Fig. 73, k), the intercostales externi
(Fig. 73, m), a small part of the serratus posterior inferior
(Fig. 73, n), and by its dorsal tendon with the longissimus
dorsi (Fig. 69, f).
Action.—Constrictor of the abdomen.
M. obliquus abdominis internus (Fig. 73, o).—A thin
sheet similar to the preceding but of less extent. Its fibres
cross those of the external oblique nearly at right angles and
lie beneath them.
Origin.—(1) Between the fourth and seventh lumbar vertebræ
from the lumbar aponeurosis which is common to it and the
external oblique. The lumbar aponeurosis takes origin from
the lumbar spinous processes and the interspinous ligaments,
and is continuous craniad with the aponeurosis of the serratus
posterior inferior. Laterad it splits into three sheets, two for
the above-named muscles and a third which passes to the
vertebral transverse processes and forms the fascia covering the
supraspinous muscles of the lumbar region. (2) By a similar
aponeurosis from the ventral half of the iliac crest. (3) By
fleshy fibres from the three crural arches. These are three
delicate ligamentous arches which stretch from the crest of the
ilium to the pubic spine. The dorsal one gives exit to the
iliopsoas muscle, the middle to the femoral vessels, and the
ventral to the spermatic cord. In the female the middle and
ventral arches may fuse. The pillar between the dorsal and
middle arches is attached to the iliopectineal eminence.
Insertion.—The fibres pass cranioventrad and end along a
longitudinal line in a thin aponeurosis of insertion, which is
united in the linea alba to those of the external oblique and
transversus. At the caudal end of the aponeurosis all its fibres
pass outside of the rectus abdominis (Fig. 73, k). At its
cranial end the fibres divide into two sheets or laminæ, one of
which passes outside of the rectus and the other inside. The
outer lamina unites with the aponeurosis of the external
oblique, while the inner lamina unites with that of the transversus.
There is thus formed a sheath for the cranial part of
the rectus muscle.
Relations.—Outer surface with the obliquus
externus[155] (Fig.
68, p). Inner surface with the transversus abdominis (Fig.
69, l), and by its ventral tendon with the rectus.
Action.—Compressor of the abdomen.
M. transversus abdominis (Fig. 69, l).—A thin sheet
covering the whole surface of the abdomen and lying beneath
the internal oblique. Its fibres are nearly transverse.
Origin.—(1) By fleshy fibres or by a thin aponeurosis from
the cartilages of all the false and floating ribs, by interdigitation
with the fibres of the diaphragm. (2) From the tips of all the
lumbar transverse processes. (3) From the ventral border of
the ilium. (4) From the dorsal and middle of the three crural
arches, where it may blend partly with the internal oblique.
The muscle is continuous craniad with the transversus thoracis
muscle. Near the lateral border of the rectus abdominis the
muscle ends in a thin aponeurosis of insertion which is continued
(its fibres having the direction of the muscle-fibres) to
the
Insertion in the linea alba.
Relations.—Outer surface with the internal oblique (Fig.
73, o) and the rectus abdominis (Fig. 73, k), dorsad also with
the longissimus dorsi (Fig. 69, f). Inner surface with a thin
fascia covering the peritoneum and ventrocaudad with the
rectus (Fig. 73, k). The dorsal edge of the muscle touches
the iliopsoas and longissimus dorsi; the ventral edge touches
the muscle of the opposite side.
Action.—Constrictor of the abdomen.
M. rectus abdominis (Fig. 73, k).—A rather thick, flat
muscle which lies near the median ventral line separated by
the linea alba from its fellow of the opposite side and stretching
from the pubis to the first costal cartilage. Opposite the first
lumbar vertebra it is approximately four centimeters wide. It
narrows at both ends.
Origin by a strong tendon from the tubercle of the pubis.
The muscle passes craniad at first between the peritoneal fascia
and the transversus aponeurosis, then in the sheath formed by
the internal and external oblique and the transversus aponeurosis.
It emerges from the sheath opposite the xiphoid process
and passes ventrad of the costal cartilages. Between the third[156]
and fourth costal cartilages it ends in a thin tendon which
passes beneath the transversus costarum and is
Inserted into the first costal cartilage near its middle, into
the second costal cartilage near its sternal end, and into the
sternum between the first and fourth cartilages.
Relations.—Outer surface of the cranial part of the muscle
and its tendon with the pectoralis minor (Fig. 68, o) and the
transversus costarum (Fig. 73, j); caudad the muscle is covered
by the outer layer of the rectus sheath. Lateral edge with the
obliquus externus (Fig. 68, p), obliquus internus (Fig. 73, o),
and transversus abdominis (Fig. 69, l). Medial edge with the
muscle of the opposite side. Inner surface with the internal
intercostals (Fig. 69, k) and the rib cartilages; the inner layer
of the rectus sheath and the peritoneum.
Action.—Retracts the ribs and sternum and compresses the
abdomen.
IV. MUSCLES OF THE THORACIC LIMBS.
The muscles connecting the thoracic limbs with the rest of
the body have been described.
1. Muscles of the Shoulder.
A. Muscles on the Lateral Surface of the
Shoulder.
M. deltoideus.—The deltoid muscle of the cat is divided
into two (or three) portions which are together equivalent to
the human deltoid. These are the spinodeltoid, the acromiodeltoid,
and possibly the clavobrachial, which is frequently
called clavodeltoid.
M. spinodeltoideus (Fig. 75, e; Fig. 68, i).—A rather
thick, flat muscle between the scapular spine and the deltoid
ridge of the humerus. It forms a chord of the angle between
the glenoid border of the scapula and the humerus.
Origin by short tendon-fibres from the glenoid border of
somewhat more than the middle third of the spine of the
scapula (Fig. 76, g), and from a tendinous raphe between the
spinotrapezius (Fig. 68, j), acromiotrapezius
(Fig. 68, h), and[157]
infraspinatus (Fig. 75, c). The origin may pass toward the
vertebral border of the scapula onto the infraspinatus muscle
(Fig. 75, c).
Insertion (Fig. 81, h) by a flat tendon upon the deltoid
ridge of the humerus, nearly parallel to that of the pectoralis
major.
Relations.—Outer surface with the integument and at the
insertion with the acromiodeltoideus (Fig. 75, f). Inner surface
with the infraspinatus (c), teres minor (Fig. 80, c), caput
laterale (Fig. 75, h), and caput longum (Fig. 75, g) of the
triceps muscle.
Action.—Flexes the humerus and rotates it outward.
M. acromiodeltoideus (Fig. 75, f; Fig. 68, g).—A flat
muscle which overlies the distal end of the spinodeltoideus
(Fig. 75, c). It connects the acromion with the humerus.
Origin (Fig. 76, d).—From the glenoid border of the
acromion, and sometimes the adjacent metacromion as far as
the tip.
Insertion.—Mostly upon the outer surface of the spinodeltoideus
(Fig. 75, c). The outer fibres are continued to the
bone, especially at the lateral border of the muscle, and are
inserted along a line ventrad of the line of insertion of the
spinodeltoideus, and extending farther distally (Fig. 81, g).
Some of the outer fibres pass into the brachialis (Fig. 80, h).
Relations.—Outer surface with the integument and the
clavobrachial (Fig. 68, e). Inner surface with the infraspinatus
(Fig. 75, c), teres minor (Fig. 80,
c), spinodeltoid (Fig. 75, e),
and caput laterale of the triceps (Fig. 75, h).
Action.—Like that of the spinodeltoid.
M. clavobrachialis (Fig. 65, k;
Fig. 68, e).—A flat, triangular
muscle on the cranial surface of the shoulder, forming
a direct continuation of the clavotrapezius (Fig. 68, d). These
two are frequently described as constituting a single muscle,
the cephalohumeral or cephalobrachial. (The clavobrachial is
frequently given the name clavodeltoid; as its homology with
the human clavodeltoid appears doubtful, it seems well to use
the name clavobrachial, as proposed by Clasen.)
Fig. 75.—Muscles on the
Lateral Surface of
the Arm.
a, M. supraspinatus; b,
part of insertion of M. rhomboideus;
c, M. infraspinatus;
d, M. teres major; e, M. spinodeltoideus;
f, M. acromiodeltoideus;
g, caput longum
of M. triceps brachii; h, caput
laterale of M. triceps brachii;
i, M. brachialis; j, M. biceps;
k, M. brachioradialis; l, M.
extensor carpi radialis longus;
m, M. extensor communis
digitorum (m′, tendon
of same); n, M. extensor lateralis
digitorum; (n′, tendon
of same); o, M. extensor
carpi ulnaris (o′, tendon of
same); p, M. extensor indicis
proprius; q, fifth head of M.
flexor profundus digitorum;
r, M. flexor carpi ulnaris. 1,
vertebral border of the scapula;
2, spine of the scapula;
3, greater tuberosity of the
humerus; 4, olecranon; 5,
transverse ligament of the
wrist.
Origin.—Its superficial fibres are continuations of
the[158]
[159]
clavotrapezius (Fig. 68, d); other fibres have origin from the
clavicle and from a raphe laterad of the clavicle which is common
to this muscle and the clavotrapezius. It passes distad
along the cranial surface of the arm, growing narrower as
it approaches the convexity of the elbow.
Insertion.—Eight to ten millimeters from the ulna it joins
the brachialis muscle (Fig. 79, i) to be inserted with it by a
flat tendon (Fig. 79, i; Fig. 87, c) upon a rough area on the
medial surface of the ulna just distad of the semilunar notch,
and about midway between the dorsal and ventral borders.
Relations.—Outer surface with the integument. Inner
surface with the pectoralis major (Fig. 65, l), biceps (Fig.
77, g), lateral head of the triceps (Fig. 75, h), acromiodeltoid
(Fig. 75, f), and brachialis (Fig. 80, h). Medial border with
the pectoantibrachialis (Fig. 65, m). Lateral border free
except near the insertion, where it is in relation with the
brachialis.
Action.—Flexor of the antibrachium.
M. supraspinatus (Fig. 75, a) occupies the whole of the
supraspinatus fossa of the scapula. It is covered by strong
fascia which stretches from the free edge of the spine to the
coracoid border of the scapula and to the coracoid half of its
vertebral border.
Origin (Fig. 76, b) by fleshy fibres from the whole surface
of the supraspinatus fossa, from the above-mentioned fascia,
and from the subscapularis (Fig. 77, a) craniad of the coracoid
border of the scapula.
Insertion (Fig. 83, a).—It passes over the capsule of the
shoulder-joint, to which it is closely attached, and is inserted
into the free border of the great tuberosity ventrad (or proximad)
of the fossa for the infraspinatus (Fig. 83, c).
Relations.—Outer surface with the spinotrapezius (Fig.
68, j), acromiotrapezius (Fig. 68, h), levator scapulæ ventralis
(Fig. 68, f), clavotrapezius (Fig. 68, d), and cleidomastoid
(Fig. 65, h). Inner surface with the scapula. The distal end
of the coracoid border is closely related to the pectoralis minor
(Fig. 79, f). The glenoid border is related to the origin of
the deltoidei.
[160]
Action.—Extends the humerus after it has been flexed on
the scapula.
M. infraspinatus (Fig. 75, c).—This fills the infraspinatus
fossa, its fibres converging to the insertion on the great tuberosity
of the humerus.
Fig. 76.—Lateral
Surface of the Scapula, with the Areas of Attachment
of Muscles.
a, M. infraspinatus; b, M. supraspinatus; c,
origin of M. biceps; d, M. acromiodeltoideus;
e, M. teres minor; f, M. levator scapulæ ventralis; g, M. spinodeltoideus;
h, M. acromiotrapezius; i, M. rhomboideus; j, M. teres major; k-l, line of insertion
of M. spinotrapezius.
Origin (Fig. 76, a).—By fleshy fibres from the whole infraspinatus
fossa, and by a raphe between it and the teres major
(Fig. 75, d), and sometimes by fibres from the teres minor
(Fig. 80, c), triceps, spinodeltoideus (Fig. 75, e), spinotrapezius
(Fig. 68, j), and subscapularis (Fig. 77, a).
Insertion (Fig. 83, c).—By a flat tendon which passes over
the capsule of the joint, into the ventral half of the infraspinatus
fossa on the great tuberosity of the humerus. (The dorsal half
of the fossa is covered by a synovial bursa.)
Relations.—Outer surface with the spinotrapezius (Fig.
68, j), spinodeltoid (Fig. 68, i), acromiodeltoid (Fig. 68, g),
teres major (Fig. 75, d), and levator scapulæ
ventralis[161] (Fig.
68, f). Inner surface with the scapula. Glenoid border with
the teres minor (Fig. 80, c), the teres major (Fig. 75, d), and
the long head of the triceps (Fig. 75, g).
Action.—Rotates the humerus outward.
M. teres minor (Fig. 80, c).—A small muscle from the
glenoid border of the scapula to the proximal end of the
humerus.
Origin (Fig. 76, e) by a sheet of tendinous fibres from the
glenoid border of the scapula beginning about one-fifth the
length of the border from the glenoid fossa and extending to
its middle. It is often attached to the infraspinatus (Fig.
80, b) and the caput longum of the triceps (Fig. 75, g).
Insertion (Fig. 83, d) by a short tendon into the tubercle
just distad of the infraspinatus fossa on the great tuberosity of
the humerus.
Relations.—Outer surface with the spinodeltoideus (Fig.
75, e), acromiodeltoideus (Fig. 75, f), and the infraspinatus
(Fig. 80, b). Inner surface with the lateral (Fig. 75, h) and long
(Fig. 75, g) heads of the triceps and the capsule of the joint.
Action.—Assists the infraspinatus to rotate the humerus
outward.
B. Muscles on the Medial Surface of the Shoulder.
(Fig. 77.)
M. subscapularis (Fig. 77, a).—A triangular mass from
the subscapular fossa to the lesser tuberosity of the humerus.
Origin (Fig. 78, a).—From the whole subscapular fossa
except along the fusiform area for the attachment of the levator
scapulæ and the serratus anterior near the vertebral border,
and except over a quadrangular area about one centimeter long
near the glenoid angle. The origin is by fleshy fibres directly
from the periosteum except along two or three lines marked
by oblique ridges. To these lines are attached tendinous fibres.
At the glenoid border the area of origin sometimes occupies
part or the whole of the surface of the fossa for the origin of the
teres major, so that the teres arises from the fascia on the surface
of the subscapularis. At the coracoid border some of the
fibres may take origin from the adjacent fascia of the supraspinatus
(Fig. 77, d).
[162]
Insertion (Fig. 82, b).—The fibres converge to the glenoid
border, and the insertion is by
a strong, flat tendon into the
dorsal border of the lesser
tuberosity of the humerus.
Fig. 77.—Muscles on the Medial Side
of the Arm.
M. epitrochlearis, M. clavobrachialis, and the
breast-muscles have been removed. a, M.
subscapularis; b, insertion of M. levator scapulæ
and of M. serratus anterior; c, M. teres major;
d, M. supraspinatus; e, M. latissimus dorsi (e′,
cut edge of that part which becomes united with
the bicipital arch); f, M. coracobrachialis; g, M.
biceps brachii (the capsule of the joint has been
laid open to show its tendon); h, cut insertions
of pectoralis muscles; i, caput longum of M. triceps
brachii; j, long portion of caput mediale
of M. triceps brachii; k, intermediate portion of
caput mediale of M. triceps brachii; l, short portion
of caput mediale of M. triceps brachii; m,
cut end of M. clavobrachialis; n, M. brachioradialis;
o, M. extensor carpi radialis longus; p,
M. extensor carpi radialis brevis; q, M. pronator
teres; r, M. flexor carpi radialis; s, M.
palmaris longus (s′, its tendons); t, M. flexor carpi
ulnaris (t, ulnar head; t′, humeral head); u, third
head of M. flexor profundus digitorum; u′, fifth
head of M. flexor profundus digitorum; u″, one
of the tendons of M. flexor profundus digitorum;
v, M. pronator quadratus; w, M. abductor brevis
pollicis; x, ulnar part of M. flexor sublimis digitorum.
1, transverse ligament of the wrist.
Relations.—Lateral
surface with the scapula
and the capsule of the
shoulder-joint. Medial
surface with the levator
scapulæ (Fig. 73, h), serratus
anterior (Fig. 73, i),
part of the scalenus (Fig.
73, f), the transversus costarum
(Fig. 73, j), and the
coracobrachialis (Fig. 77, f).
Cranial border with the supraspinatus
(Fig. 77, d). Caudal border[163]
with the teres major (Fig. 77, c) and infraspinatus (Fig.
75, c).
Action.—Pulls the humerus inward (mediad).
M. teres major (Fig. 75, d; Fig. 77, c).—A thick muscle,
triangular in cross-section, lying parallel with the glenoid border
of the scapula.
Origin (Fig. 76, j; Fig. 78, b) from the vertebral one-third
of the glenoid border of the scapula, and from fascia covering
the subscapularis (Fig. 77, a)
and the infraspinatus (Fig. 75, c)
over a small area near the glenovertebral
angle of the scapula.
It may touch the insertion of the
rhomboideus.
Insertion (Fig. 82, f) by a
tendon common to it and the
latissimus dorsi (Fig. 79, d′), as
already described.
Relations.—Outer surface
with the caput longum (Fig. 75,
g) and the long portion (Fig.
80, f) of the caput mediale of
the triceps, the latissimus dorsi
(Fig. 77, e), and the cutaneus
maximus. Inner surface with
the serratus anterior (Fig. 73, i),
the scalenus (Fig. 73, f), the
transversus costarum (Fig. 73, j),
and the biceps (Fig. 77, g).
Dorsal border with the subscapularis
(Fig. 77, a) and infraspinatus
(Fig. 75, c).
Action.—Rotates the humerus inward and flexes it in opposition
to the infraspinatus, teres minor, and the deltoidei.
Fig. 78.—Medial Surface of the
Scapula with the Areas of
Attachment of Muscles.
a, M. subscapularis; b, M. teres major;
c, M. serratus anterior; d, M. levator
scapulæ; e, M. rhomboideus; f, M. occipitoscapularis;
g, M. biceps; h, M.
coracobrachialis; i, caput longum of M.
triceps.
[164]
2. Muscles of the Brachium or Upper Arm.
M. clavobrachialis and the pectoralis group (Fig. 65, page
109) lie partly in this region; they have been described.
M. coracobrachialis (Fig. 77, f; Fig. 79, c).—A very
short muscle covering the inner (medial) surface of the capsule
of the shoulder-joint. It extends from the coracoid process
to the proximal end of the humerus.
Origin (Fig. 78, h).—By a round tendon from the tip of
the coracoid process.
Insertion (Fig. 82, e) usually by fleshy fibres on an area
five to eight millimeters long and about half as wide, which lies
on the medial side of the humerus parallel with the ridge which
runs from the dorsal end of the lesser tuberosity to the shaft
and close to it. The proximal end of the area is about one
centimeter from the proximal end of the head of the humerus.
Relations.—Medial surface with the biceps (Fig. 77, g),
pectoralis minor (Fig. 65, o), and teres major (Fig. 77, c).
Lateral surface with the capsule of the shoulder-joint. Dorsal
border with the subscapularis (Fig. 77, a) and the long portion
of the caput mediale of the triceps (Fig. 77, j).
Action.—Adducts the humerus.
The part of the coracobrachialis just described is known as
the short head. A long head is sometimes found. It is a
conical bundle of fibres of varying size, which arises from the
tendon of origin of the short head. It passes distad into a long
and extremely slender tendon, the insertion of which varies
greatly in different individuals. It is commonly on the humerus
in the region of the supracondyloid foramen.
M. epitrochlearis or extensor antibrachii longus (Fig.
65, r).—A thin, flat muscle on the inner or medial side of the
brachium, from the lateral surface of the latissimus dorsi (Fig.
65, q) to the olecranon process of the ulna.
Origin from the lateral or outer surface of the ventral border
of the latissimus dorsi (Fig. 65, q) near the insertion of the
cutaneus maximus. Fibres are often attached to the teres
major and the pectoralis minor.
Insertion by a flat tendon which is closely connected with[165]
that of the pectoantibrachialis (Fig. 65, m) and is continuous
with the general antibrachial fascia, into the caudal border of
the cutaneous (dorsal) surface of the olecranon process of the
ulna.
Relations.—Outer (medial) surface with the integument, the
latissimus dorsi (Fig. 65, q), and the cutaneus maximus. Inner
(lateral) surface with the biceps (Fig. 65, t), caput longum
(Fig. 65, s), and caput mediale (Fig. 77, j, k) of the triceps.
Action.—Extends the antibrachium and tends to supinate
the hand by rotating the ulna.
M. biceps brachii (Fig. 77, g; Fig. 65, t).—A thick, fusiform
muscle lying on the front (ventral) surface of the humerus.
Origin (Fig. 76, c) by a strong, round tendon from the
bicipital tubercle of the glenoid angle of the scapula, at its
coracoid margin. (In man there is a second head—coracoid
head—from the coracoid process.) The tendon passes through
the capsule of the joint and then along the bicipital groove,
which is converted into a canal by a strong ligament.
Insertion by a rounded tendon (Fig. 87, b) on the bicipital
tuberosity of the radius (Fig. 86, k).
Relations.—The tendon of origin passes through the capsule
of the joint, which is covered by a part of the pectoralis minor.
The muscle is then spanned for the middle part of its course
by the bicipital arch (Fig. 65, t′), a description of which is
given below. Farther distad it is covered by the clavobrachial
(Fig. 65, k) and epitrochlearis (Fig. 65, r). At its distal end
(Fig. 79, l′) the muscle passes between the pronator teres on
the radial side and the conjoined tendon of the brachialis and
clavobrachial (Fig. 79, i′) on the other. The biceps lies
throughout most of its length on the ventral surface of the
humerus, and touches along the medial border of its inner surface
the coracobrachialis (Fig. 77, f), the teres major, (Fig.
77, c), the intermediate portion of the caput mediale of the
triceps (Fig. 77, k), and the short portion (Fig. 77, l) of the
same. Its lateral border touches the pectoralis minor (Fig.
65, o) and the brachialis (Fig. 79, i).
Action.—Flexes the forearm, and tends to supinate the
hand.
[166]
The bicipital arch (Fig. 65, t′) is a tendinous arch formed
over the biceps muscle. Its outer or lateral pillar is formed
by the tendon of the pectoralis minor (o), to which may be
added part of the latissimus (q) tendon. Its inner or medial
pillar is formed by the conjoined tendon of the teres major and
latissimus dorsi (Fig. 79, d′), while the xiphihumeralis (Fig.
65, p), cutaneus maximus (Fig. 62, b), and epitrochlearis
(Fig. 65, r) are connected with one or the other pillar or with
the muscles composing them. The caudal portion of the deep
layer of the pectoralis major (Fig. 65, l) may be connected
with the inner pillar.
M. brachialis (Fig. 79, i; Fig. 75, i).—From the lateral
surface of the humerus to the ulna.
Origin (Fig. 83, g, g′) from a long V-shaped line two to
four millimeters wide on the lateral surface of the humerus.
The apex of the V is just distad of the teres minor tubercle (d).
Its dorsal limb (g′) extends along the lateral supracondyloid
ridge to a point opposite the proximal margin of the supracondyloid
foramen; its ventral limb (g) extends in the direction
of the deltoid ridge to the middle of the bone. No
muscle-fibres take origin between the two limbs. The fibres
converge and end in a flat tendon which joins the tendon of
the clavobrachial (Fig. 79, k).
Insertion (Fig. 87, c).—The dorsal portion of the depressed
rough area on the lateral surface of the ulna just distad of the
semilunar notch.
Relations.—Outer surface with the acromiodeltoid (Fig.
75, f), the caput laterale of the triceps (Fig. 75, h), the
brachioradialis (Fig. 75, k), and the proximal end of the extensor
carpi radialis longus (Fig. 75, l). Inner surface with
the humerus. Medial edge with the pectoralis major (Fig.
65, l) and the biceps (Fig. 77, g).
Action.—Flexor of the antibrachium.
M. triceps brachii.—The triceps muscle of the cat, like
that of man, is divisible (as its name indicates) into three main
portions. These are a lateral portion (caput laterale), an
intermediate or long portion (caput longum), and a medial portion
(caput mediale). The first two correspond to the similarly[167]
named heads in man; the medial portion falls into a
number of subdivisions whose homologues are uncertain.
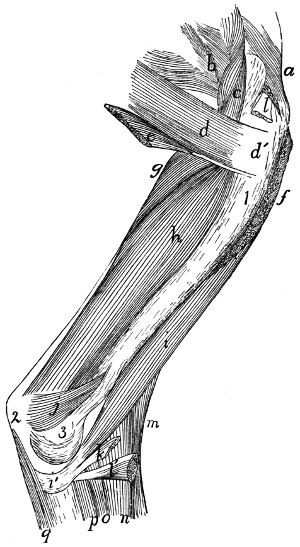
Fig. 79.—Deep
Muscles on the Medial Side of the Humerus.
a, M. supraspinatus; b, M. subscapularis; c, M. coracobrachialis; d, M. teres
major (d′, its tendon); e, part of M. latissimus dorsi joining the teres major; f, cut
end of M. pectoralis minor; g, long part of caput mediale of M. triceps brachii; h, intermediate
part of caput mediale of M. triceps; i, M. brachialis (i′, its tendon); j, short
part of caput mediale of M. triceps; k, cut end of M. clavobrachialis; l, l′, cut ends
of M. biceps; m, M. brachioradialis; n, M. extensor carpi radialis longus; o, M. extensor
carpi radialis brevis; p, fifth head of M. flexor profundus digitorum; q, first
head of M. flexor profundus digitorum. 1, humerus; 2, olecranon; 3, medial epicondyle
of humerus.
Caput laterale (anconeus lateralis) (Fig. 75, h; Fig. 68, l).—The
lateral portion is a flat muscle (most of it subcutaneous)
on the lateral side of the brachium, connecting the proximal
end of the humerus with the olecranon process of the ulna.
[168]
Origin (Fig. 81, h′).—By a flat tendon from the proximal
portion of the deltoid ridge and the distal border of the teres
minor tubercle.
Insertion (Fig. 86, c) by a thin, flat tendon into the lateral
border of the dorsal surface of the ulna between a point opposite
the distal margin of the semilunar notch and the proximal
end of the olecranon.
Relations.—Lateral surface with the integument, and at the
proximal end with the spinodeltoid and acromiodeltoid (Fig.
75, e and f). Medial surface with the brachialis (Fig. 75, i),
the caput longum (g), the caput mediale, the brachioradialis (k),
the anconeus (Fig. 80, l), and the origins of the extensor carpi
radialis longus (Fig. 75, l), the extensor communis digitorum
(m), and the extensor digitorum lateralis (n).
Caput longum (anconeus longus) (Fig. 75, g; Fig. 68, k;
Fig. 77, i).—A prismatic, fusiform mass connecting the scapula
with the olecranon.
Origin (Fig. 78, i) by a thick, flat tendon from a triangular
area one or two centimeters long at the glenoid end of the
scapula on the glenoid border. The base of the area is about
one millimeter from the margin of the glenoid fossa.
Insertion (Fig. 86, b).—The muscle ends in a thick tendon
which passes over the bifurcated ventral end of the olecranon
and is inserted upon the rounded tuberosity which forms the
dorsal angle of the olecranon.
Relations.—Lateral surface with the caput laterale (Fig.
75, h), a portion of the caput mediale (Fig. 77, j and k), the
spinodeltoid (Fig. 75, e), the infraspinatus (Fig. 75, c), and
the integument. Medial surface with the epitrochlearis (Fig.
65, r) and the conjoined portions of the latissimus dorsi and
teres major (Fig. 79, d′). Ventral border with the caput
mediale (Fig. 77, j and k).
Caput mediale.—The medial head consists of three portions
(Fig. 79, g, h, j).
(1) The long portion (anconeus posterior) (Fig. 77, j; Fig.
79, g; Fig. 80, f).
Origin (Fig. 83, e) from a triangular area on the dorsal surface
of the humeral shaft. The base of the area is against the[169]
articular head, and its apex about one-sixth the length of the
humerus from the head. It is between the coracobrachialis
and the lateral head of the triceps.
Fig. 80.—Deep
Muscles on the Lateral Surface of the Humerus.
a, part of M. supraspinatus; b, part of M.
infraspinatus; c, M. teres minor; d, M.
teres major; e, part of M. latissimus dorsi joining M. teres major; f, long part of caput
mediale of M. triceps brachii; g, cut insertion of M. acromiodeltoideus; h, M. brachialis;
i, M. brachioradialis; j, intermediate part of caput mediale of M. triceps brachii;
k, M. extensor carpi radialis longus; l, M. anconeus; m, M. extensor communis digitorum.
1, great tuberosity of humerus; 2, 3, superficial radial and dorsal interosseous
branches, respectively, of the radial nerve.
Insertion (Fig. 86, a).—The muscle ends in a long, slender
tendon which passes through the furrow on the ventral[170]
angle of the olecranon and is inserted into an oblique ridge
which forms the dorsal limit of the furrow. The tendon is
separated from the floor of the furrow by a synovial bursa.
(2) The intermediate portion (anconeus internus) (Fig. 77,
k; Fig. 79, h; Fig. 80, j).
Origin (Fig. 82, g) by fleshy fibres from a triangular area
proximad of the middle of the dorsomedial surface of the
humerus. The length of the area equals about one-fourth the
length of the bone. Its apex points proximad and is almost
continuous with the apex of the area of origin of the long portion.
Insertion (Fig. 87, i) by short tendon-fibres into the medial
border of the ventral and proximal surfaces of the olecranon.
It is connected with the tendon of the long portion.
(3) Short portion (Fig. 79, j; Fig. 77, l).
Origin (Fig. 82, h) from the outer surface of the bony bar
which encloses the supracondyloid foramen of the humerus, as
far as the medial epicondyle.
Insertion (Fig. 87, e) by fleshy fibres into the medial border
of the olecranon distad of the tuberosity for the insertion of the
caput longum.
Relations of the caput mediale.—Lateral surface with the
caput laterale (Fig. 75, h), the humerus, and the anconeus (Fig.
80, l). Dorsal surface with the caput longum (Fig. 77, i),
the epitrochlearis (Fig. 65, r), and the teres major (Fig.
79, d). Ventral surface with the biceps (Fig. 77, g).
Action of the Triceps.—The entire triceps group forms a
powerful extensor of the forearm. The short portion of the
medial head tends also to rotate the arm outward, so as to
supinate the hand.
M. anconeus (Fig. 80, l).—A triangular muscle on the
outside of the elbow-joint from the humerus to the ulna.
Origin (Fig. 83, i).—An irregularly triangular area at the
distal end of the dorsal surface of the humerus. The area of
origin is limited laterally by the sharp lateral supracondyloid
ridge. It sometimes extends onto the lateral epicondyle.
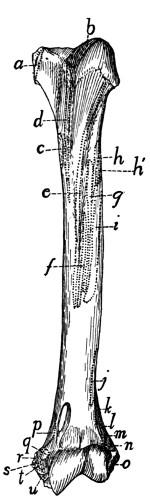
Fig. 81.—Ventral Surface
of Humerus,
with the Areas of
Attachment of Muscles.
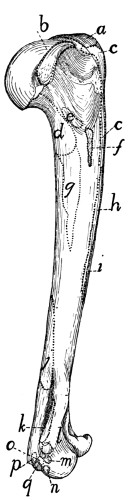
Fig. 82.—Medial Side
of Humerus, with
the Areas of Attachment
of Muscles.
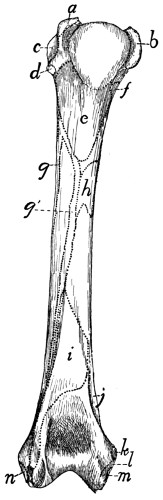
Fig. 83.—Dorsal Surface
of the Left Humerus,
with the Areas of
Attachment of Muscles.
Fig. 81.—a, M. subscapularis; b, M. supraspinatus; c, Mm. teres major and latissimus
dorsi; d, M. pectoralis minor; e, deep layer of M. pectoralis major; f, superficial
layer of M. pectoralis major; g, M. acromiodeltoideus; h, M. spinodeltoideus;
h′, caput laterale of M. triceps; i, M. brachialis; j, second portion of M. brachialis;
k, M. extensor carpi radialis longus; l, M. extensor carpi radialis brevis; m. M. extensor
communis digitorum; n, M. extensor lateralis digitorum; o, M. extensor carpi
ulnaris; p, short portion of caput mediale of M. triceps; q, M. pronator teres; r, M.
palmaris longus; s, third and fourth parts of M. flexor profundus digitorum; t, M.
flexor carpi radialis; u, second head of M. flexor profundus digitorum.
Fig. 82.—a, M. supraspinatus; b, M. subscapularis; c, M. pectoralis minor; d,
long portion of caput mediale of M. triceps; e, M. coracobrachialis; f, Mm. teres
major and latissimus dorsi; g, intermediate portion of caput mediale of M. triceps; h,
deep layer of M. pectoralis major; i, superficial layer of M. pectoralis major; j, M.
anconeus; k, short portion of caput mediale of M. triceps; l, M. pronator teres;
m, M. flexor carpi radialis; n, second head of M. flexor profundus digitorum; o,
third and fourth heads of M. flexor profundus digitorum; p, M. palmaris longus; q,
M. flexor carpi ulnaris.
Fig. 83.—a, M. supraspinatus; b, M. subscapularis; c, M. infraspinatus; d, M.
teres minor; e, long portion of caput mediale of M. triceps; f, M. coracobrachialis;
g, g′, M. brachialis; h, intermediate portion of caput mediale of M. triceps; i, M.
anconeus; j, short portion of medial head of M. triceps; k, M. pronator teres; l,
second and third heads of M. flexor profundus digitorum; m, M. palmaris longus;
n, M. extensor carpi ulnaris.
Insertion (Fig. 86, d).—The lateral surface of the ulna from
the distal margin of the semilunar notch to the proximal end[171]
[172]
of the olecranon. The muscle covers the capsule of the joint
and is closely adherent to it.
Relations.—Outer (dorsal) surface with the caput laterale
(Fig. 75, h) and the intermediate portion (Fig. 80, j) of the
caput mediale. Inner surface with the bones and the capsule
of the joint.
Action.—Keeps the capsule tense and probably rotates the
ulna slightly so as to pronate the hand.
3. Muscles of the Antibrachium or Forearm.
Fascia of the Forearm.
—The muscles of the forearm are
not in contact with the skin, but are covered by the strong
antibrachial fascia. This consists of two layers. The superficial
one is a continuation of the general subcutaneous fascia
of the arm; it covers the forearm as a continuous sheet which
becomes thinner distad, and is finally lost near the wrist. The
deep layer forms a dense, tendinous sheet which is closely
applied to the muscles. It begins partly as a continuation of
the general fascia of the arm, partly from the tendons of the
triceps and epitrochlearis muscles. The sheet is attached to
the dorsal surface of the ulna, between the extensor carpi
ulnaris and the first head of the flexor profundus digitorum.
It surrounds the forearm as a continuous sheath. On the lateral
surface it dips between the extensor muscles, becoming closely
attached to them and forming a partial sheath for each; it is
attached with special firmness to their tendons. On the medial
side of the arm the fascia passes smoothly over the flexors
without dipping between them; it is continuous, however, with
the border of the pronator teres, and distad of this muscle it is
firmly attached to the radius.
At the wrist the fascia becomes attached to the longitudinal
ridges on the dorsal surface of the head of the radius, bridging
thus the intervening grooves and holding the tendons in
place between the ridges. The fascia then continues to form
the dorsal fascia of the hand. On the ventral side of the wrist
the fascia becomes thickened to form a strong transverse
ligament (Fig. 77, 1), which is attached at its radial and ulnar
ends to the carpus, and which holds in place the tendons of the[173]
flexor muscles. Distad of the transverse ligament the fascia
spreads out in the palm of the hand, and becomes united with
the fibrous pad which is situated here. On the fingers the
fascia becomes attached to the phalanges, forming tendinous
sheaths, the so-called vaginal ligaments, for the flexor
muscles. Near the base and the head of the first phalanx
strong, narrow, ring-like bands, the annular ligaments (Fig.
88, 2), are formed in these sheaths; these surround and bind
down the tendons of the flexor muscles.
A. Muscles on the Ulnar and Dorsal Side of the
Forearm (extensors and supinators)
(Fig. 75, p. 158).
M. brachioradialis or supinator longus (Fig. 75, k; Fig.
77, n).—This is a ribbon-like muscle lying on the lateral side
of the brachium and the ventrolateral border of the antibrachium
just beneath the superficial fascia. It connects the
humerus and the radius.
Origin.—By a thin tendon from about the middle fifth of
the dorsal border of the humerus, or sometimes proximad of
the middle. The muscle curves over the outer surface of the
brachialis (Fig. 75, i), to which it is closely applied, and runs
along the lateral border of the antibrachium to its insertion.
Insertion by a short tendon into the ridge which forms the
ventral limit of the groove on the outer surface of the styloid
process of the radius and upon the adjacent ligaments.
Relations.—Outer surface with the integument, and at the
proximal end with the caput laterale of the triceps (Fig. 75, h).
Inner surface with the brachialis (i), the extensor carpi radialis
longus (l) and brevis, and the extensor communis digitorum (m).
Action.—Supinator of the hand.
M. extensor carpi radialis longus (Fig. 75, l; Fig. 77,
o).—This lies along the radial side of the forearm, and connects
the humerus and the second metacarpal.
Origin (Fig. 81, k) from the middle portion of the lateral
supracondyloid ridge between the origin of the brachialis (j)
and the anconeus. About the middle of the forearm the
muscle ends in a slender tendon (Fig. 84, e) which passes
through the groove on the radial side of the dorsal surface of
the distal end of the radius.
[174]
Insertion (Fig. 84, c).—The tendon passes across the carpus
and is inserted onto the dorsal surface of the base of the
second metacarpal on the radial side just distad of the groove
for the radial artery.
Relations.—Outer surface at the proximal end with the
caput laterale of the triceps (Fig. 75, h), farther distad with
the integument and the brachioradialis (k); the distal tendon
with the extensor pollicis brevis (Fig. 84, d). Radial surface
with the brachialis (Fig. 75, i) and the integument. Ulnar
surface with the extensor carpi radialis brevis (Fig. 77, p), and
near the proximal end with the extensor communis digitorum
(Fig. 75, m).
Action.—Extensor of the hand.
M. extensor carpi radialis brevis (Fig. 77, p).—A slender
muscle from the humerus to the third metacarpal.
Origin (Fig. 81, l) from the distal part of the lateral supracondyloid
ridge distad of the extensor longus. Near the
carpus the muscle ends in a slender tendon which passes
through the groove with the extensor longus tendon. The
tendon then diverges from the extensor longus and has its
Insertion (Fig. 84, f) into the radial side of the dorsal surface
of the base of the third metacarpal.
Relations.—Outer surface with the extensor longus (Fig.
77, o), the integument, and distad with the extensor pollicis
brevis (Fig. 84, d). Radial surface with the extensor longus.
Ulnar surface with the extensor communis digitorum (Fig. 75,
m) and the extensor brevis pollicis. Inner surface with the
pronator teres (Fig. 77, q), supinator (Fig. 85, b), and extensor
brevis pollicis (Fig. 85, a).
Action.—Extensor of the hand.
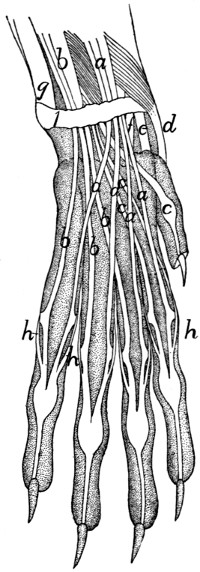
Fig. 84.—Arrangement of
the Tendons on the
Back of the Hand.
a, M. extensor communis
digitorum; b, M. extensor lateralis
digitorum; c, M. extensor
indicis; d, M. extensor
brevis pollicis; e, M. extensor
carpi radialis longus; f, M.
extensor carpi radialis brevis;
g, M. extensor carpi ulnaris;
h, tendons of the interossei.
1, transverse ligament of the
wrist.
M. extensor digitorum communis (Fig. 75, m).—A slender
muscle from the lateral supracondyloid ridge of the humerus
to the phalanges of the second, third, fourth, and fifth digits.
Origin (Fig. 81, m).—The distal surface of the lateral
supracondyloid ridge dorsad of the origin of the extensor carpi
radialis brevis.
At the junction of the middle and distal thirds of the radius the
muscle passes into a large, flat tendon (Fig. 75, m′)
which passes[175]
through the groove on the middle of the dorsal surface of the
distal end of the radius. The groove is converted into a canal
by a strong transverse ligament. The
tendon divides near the groove into four
portions (Fig. 84, a) which diverge and
pass to the four ulnar digits.
Insertions.—Each division of the
tendon as it passes over the dorsal surface
of the first phalanx has its border
connected by strong fascia to the dorsal
surface of the phalanx. It finally passes
through the groove at the distal end of
the first phalanx and is inserted mostly
into the base of the second phalanx; a
portion continues distad to be inserted
into the third phalanx.
Relations.—Outer surface with the
integument and at the origin with the
caput laterale (Fig. 75, h) of the triceps.
Radial surface with the extensor longus
(l) and extensor brevis. Ulnar surface
with the extensor digitorum lateralis (n).
Inner surface with the supinator (Fig.
85, b) and the extensor pollicis brevis
(Fig. 85, a).
Action.—Extensor of the four ulnar
digits.
M. extensor digitorum lateralis (Fig. 75, n). (This corresponds in position
to the extensor digiti quinti proprius
of man.)
Origin (Fig. 81, n) from the lateral
supracondyloid ridge of the humerus
distad of the origin of the extensor
communis.
The muscle passes along the ulnar border and dorsal surface
of the forearm between the extensor communis (Fig. 75, m) and
the extensor carpi ulnaris (o). Near the carpus it ends in a tendon[176]
(n′), which divides immediately into three or four (Fig. 84, b).
These pass through the groove on the ulnar side of the dorsal
surface of the radius and diverge to the digits lying beneath
the extensor communis tendon.
Insertion (Fig. 84, b).—The three tendons on the ulnar
side join the ulnar sides of the tendons of the extensor communis
(a) at their insertions. The radial of the four tendons
(not always present) joins the tendon of the extensor indicis (c)
and may also give a branch to the radial side of the base of the
first phalanx of the third digit.
Relations.—Outer surface with the integument. Radial
border with the extensor communis digitorum (m); ulnar border
with the extensor carpi ulnaris (o). Inner surface with the
supinator (Fig. 85, b) and the extensor brevis pollicis (Fig.
85, a); distad with the extensor indicis (Fig. 85, c).
Action.—Extensor of the four ulnar digits.
M. extensor carpi ulnaris (Fig. 75, o).
Origin.—(1) By a short, broad tendon from the distal portion
of the lateral epicondyle of the humerus (Fig. 81, o) distad
of the origin of the extensor lateralis, and (2) by a smaller
tendon from the ulna at the dorsal tip of the semilunar notch
(Fig. 86, e).
The muscle passes along the ulnar side of the forearm
dorsad of the extensor digitorum lateralis (n). It ends near
the junction of the middle and distal thirds of the forearm in a
large, flat tendon (o′) which is closely adherent to the ligaments
of the wrist and which passes over the distal end of the ulna to
its
Insertion into the tubercle on the ulnar side of the base of
the fifth metacarpal (Fig. 84, g).
Relations.—Outer surface with the integument. Radial
(or ventral) border with the extensor digitorum lateralis (Fig.
75, n); ulnar (or dorsal) border with the flexor profundus digitorum
(q). Inner surface with the radius, the extensor indicis
(Fig. 85, c), and the extensor pollicis brevis (Fig. 85, a).
Action.—Indicated by its name.
M. extensor indicis (proprius) (Fig. 85, c; Fig. 75, p) (includes
extensor pollicis longus also).
[177]
Origin (Fig. 86, f) by short, fleshy fibres from the lateral
surface of the ulna between the semilunar notch and the junction
of the third and fourth (distal) fourths.
The muscle-fibres pass obliquely distad and toward the
radial side and join the tendon (Fig. 85, c′), which runs nearly
the whole length of the radial surface.
Insertion (Fig. 84, c).—The tendon passes through the
groove on the ulnar side of the dorsal surface of the base of the
radius and divides into two. Both of these usually go to the
base of the second phalanx of the second digit, but one may
pass to the pollex, or there may be three divisions going to the
first three digits.
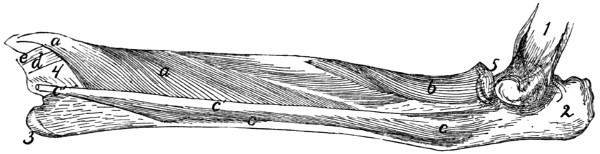
Fig. 85.—Deep
Muscles on the Extensor Surface of the Radius
and Ulna.
a, M. extensor brevis pollicis (a′, its tendon);
b, M. supinator; c, M. extensor indicis
(c′, its tendon); d, tendon of M. extensor carpi radialis brevis; e, tendon of M.
extensor carpi radialis longus. 1, humerus; 2, olecranon; 3, styloid process of
ulna; 4, distal end of radius; 5, head of radius.
Relations.—Outer surface with the integument, the extensor
carpi ulnaris (Fig. 75, o), and the extensor digitorum lateralis
(Fig. 75, n). Dorsal border with the flexor profundus digitorum
(Fig. 75, q). Inner surface with the extensor brevis
pollicis (Fig. 85, a).
Action as indicated by the name.
M. supinator (Fig. 85, b).—A flat muscle wrapped spirally
about the proximal end of the radius.
Origin by a short, strong tendon from the lateral side of the
annular ligament of the radius and by tendinous fibres from the
radial collateral ligament passing from the humerus to the
radius. From the tendon of origin the fibres diverge, passing
toward the radial side and distad.
Insertion (Fig. 86, g).—The dorsal and part of the ventral
surface of the proximal two-fifths of the radius to within[178]
five millimeters of the articular face of the
head.
Relations.—Outer surface with the extensor
carpi radialis brevis, extensor communis
digitorum (Fig. 75, m), and extensor
lateralis digitorum (Fig. 75, n). Radial
border with the pronator teres (Fig. 77, q).
Inner surface with the radius.
Action.—Supinator of the hand.
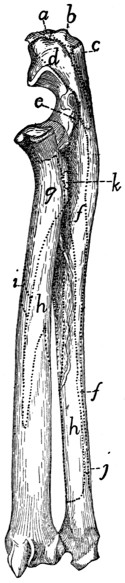
Fig. 86.—Lateral
or Extensor Surface
of Radius and
Ulna, with the
Areas of Attachment
of Muscles.
a, long portion of
caput mediale of M.
triceps; b, caput longum
of M. triceps; c,
caput laterale of M. triceps;
d, M. anconeus;
e, M. extensor carpi
ulnaris; f, M. extensor
indicis; g, M. supinator;
h, M. extensor
brevis pollicis; i, M.
pronator teres; j, M.
flexor profundus digitorum;
k, M. biceps.
M. extensor brevis pollicis or extensor
ossis metacarpi pollicis (Fig. 85, a) (includes
M. abductor longus pollicis of man).
Origin (Fig. 86, h) by fleshy fibres from
the ventral half of the lateral surface of the
shaft of the ulna from the semilunar notch to
the styloid process; from the ulnar half of
the dorsal surface of the radius from the
bicipital tubercle to a short distance distad
of the middle of the bone; and from the
interosseous membrane between these two
areas. The fibres converge to form a strong,
flat tendon (Fig. 84, d) which passes obliquely
over the tendons of the extensor carpi radialis
longus (e) and brevis (f) to its
Insertion into the radial side of the base
of the first metacarpal. The radial sesamoid
bone of the wrist is imbedded in the tendon
at its insertion.
Relations.—Outer surface with the extensor
carpi radialis brevis, the extensor
communis digitorum (Fig. 75, m), extensor
lateralis digitorum (Fig. 75, n), and extensor
indicis (Fig. 85, c). Inner surface with the
radius and ulna, and distad with the tendons
of the two extensores carpi radiales (Fig.
84, e and f).
Action.—Extends and abducts the pollex.
[179]
B. Muscles on the Radial and Ventral Side of
the Forearm (flexors and pronators).
M. pronator teres (Fig. 77, q).
Origin (Fig. 82, l) by a short strong tendon from the
extremity of the medial epicondyle of the humerus.
Insertion (Fig. 86, i) by fleshy fibres and short tendinous
fibres along the medial border of the radius, at its middle.
The radial edge of the muscle is continuous with the strong
deep layer of the antibrachial fascia.
Relations.—Outer surface with the superficial fascia. Radial
border with the tendon of the biceps (Fig. 79, l′), with the
extensor carpi radialis brevis (Fig. 77, p), and the supinator
(Fig. 85, b). Ulnar border with the flexor carpi radialis (Fig.
77, r) and part of the flexor profundus digitorum (Fig. 77, u).
Action.—Pronates the hand by rotating the radius.
M. flexor carpi radialis (Fig. 77, r).—A slender fusiform
muscle.
Origin (Fig. 82, m) from the tip of the medial epicondyle
of the humerus.
Insertion.—The slender tendon passes through a deep
groove between the os magnum and the first metacarpal. The
groove is converted into a canal by the overlying tendons and
muscles. The tendon is finally inserted into the bases of the
second and third metacarpals.
Relations.—Outer surface with the pronator teres (Fig.
77, q), the superficial fascia, and the third head of the flexor
profundus (u). Radial border with the pronator teres (q).
Ulnar border with the third and fourth heads of the flexor profundus.
Inner surface with the ulna proximad and the fifth
head of the flexor profundus (u′) distad.
Action indicated by the name.
M. palmaris longus (Fig. 77, s).—A flat fusiform muscle
beneath the fascia on the medial border of the forearm.
Origin (Fig. 82, p) by a short flat tendon from the distal part
of the medial surface of the medial epicondyle of the humerus.
About one centimeter from the wrist the muscle ends in a
flat tendon which passes through the transverse ligament and
divides on the hand into four or five (or sometimes only three)[180]
tendons (Fig. 77, s′) which diverge to the insertions. The
ulnar portion of the flexor sublimis digitorum (Fig. 77, x) takes
origin from the common tendon before its division.
Insertion.—Each tendon except the first gives off a branch
which spreads out in the trilobed pad in the palm. The middle
two of these may be traced to the integument covering the pad.
The tendon is then inserted (Fig. 88, a) near the base of the
first phalanx onto the outer surface of the perforated portion of
the flexor sublimis tendon (b, b′) at its side, except that to the
thumb, which divides near its distal end into two which are
inserted into the sesamoid bones at the base of the first phalanx.
The tendons of insertion are closely united to the fibrous pulley-ring
at the base of the first phalanx, and each may send a slip
to the base of the phalanx at either side of the ring.
Relations.—Outer surface with the superficial fascia and
one head of the flexor sublimis (Fig. 77, x). Radial border
with the flexor carpi radialis. Ulnar border with the flexor
carpi ulnaris and a part of the flexor profundus digitorum.
Inner surface with the flexor carpi radialis (r), the flexor carpi
ulnaris (t), the flexor profundus digitorum (u), and the radial
head of the flexor sublimis digitorum.
Action.—Flexor of the first phalanx of each of the digits.
M. flexor carpi ulnaris (Fig. 77, t, t′).
Origin.—There are two heads. The first or humeral head
(t′) takes origin in common with the second part of the flexor
profundus from the median surface of the distal end of the
humerus just distad of the medial epicondyle (Fig. 82, q). The
second or ulnar head (t) takes origin by fleshy fibres from the
lateral surface of the olecranon and from the dorsal border of
the ulna from the olecranon to a point distad of the semilunar
notch (Fig. 87, f). The two heads join proximad of the middle
of the forearm. The muscle passes obliquely across the ventral
surface of the forearm to its
Insertion by fleshy and tendinous fibres into the proximal
surface of the pisiform bone.
Relations.—Outer surface with the integument and the
palmaris longus (Fig. 77, s). Inner surface and ulnar border
with the flexor profundus digitorum.
[181]
Action indicated by the name.
M. flexor sublimis digitorum (or perforatus).—This muscle
is in two parts, which are given a common name only because
of the similar structure and insertions of their tendons.
The ulnar part (Fig. 77, x) is a conical muscle taking
origin from the outer surface of the tendon and muscle of the
palmaris longus and from the adjacent ligament. It gives rise
to two or three tendons (Fig. 88, b) which arise from distinct
slips of the muscle; these pass to the ulnar two or three digits.
That passing to the fourth or fifth digit receives an accessory
slip from a small mass of fibres attached to the transverse ligament
on the radial side of the palmaris tendon.
The radial part is a small flat triangular muscle which
takes origin from the outer surface of the tendon formed by the
junction of the tendons of the first and second parts of the flexor
profundus. It divides into two slips, each giving rise to a
tendon. These tendons (Fig. 88, b′) pass to the second and
third digits. That to the third digit may divide into two, one
of which goes to the fourth digit.
Each of the four tendons (Fig. 88, b and b′) is perforated
by a tendon of the flexor profundus (Fig. 88, c) as it passes
through the fibrous pulley-ring at the base of the first phalanx.
It then continues beneath the flexor profundus tendon through
the second pulley-ring (2) and is inserted into the base of the
second phalanx.
Relations.—Outer surface of the ulnar part with the integument;
inner surface with the palmaris longus (Fig. 77, s).
Outer surface of the radial part with the palmaris longus; inner
surface with the flexor profundus.
Action.—Flexor of the second phalanx of digits 2-5.
M. flexor profundus digitorum (or perforans).—This arises
by five heads, the tendons of which join one another at the wrist
to make the strongest and deepest of the flexor tendons.
The first or ulnar head has origin (Fig. 87, g) from the dorsal
half of the medial (radial) surface of the ulna from the proximal
lip of the semilunar notch to within a centimeter of the styloid
process. Its fibres converge to a large flat tendon which forms
the lateral (ulnar) and superficial part of the common tendon.
[182]
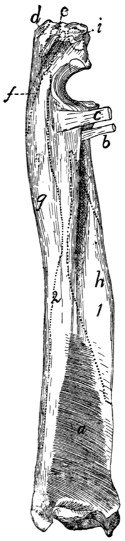
Fig. 87.—Radius and Ulna,
Medial or Flexor Side, with
M. pronator quadratus and the
Areas of Attachment of Other
Muscles.
a, M. pronator quadratus; b,
tendon of M. biceps; c, conjoined
tendon of M. brachialis and M.
clavobrachialis; d, insertion of
caput longum of M. triceps; e,
insertion of short portion of caput
mediale of M. triceps; f, origin
of M. flexor carpi ulnaris; g,
origin of first head of M. flexor
profundus digitorum; h, origin
of fifth head of M. flexor profundus
digitorum; i, intermediate
portion of caput mediale of M.
triceps.
The second head has origin (Fig.
82, n) from the distal end of the
medial epicondyle of the humerus
between the flexor ulnaris and the
third and fourth heads of the profundus.
It may be very closely attached
to these muscles at their origins. Its
tendon joins the radial border of the
tendon of the first head. From this
junction the radial part of the flexor
sublimis arises.
The third head (Fig. 77, u) has
origin (Fig. 82, o) by a strong tendon
from the medial epicondyle of the
humerus, between the palmaris (p)
and flexor radialis (m). Its tendon
forms the radial part of the common
tendon.
The fourth head has origin by a
strong tendon from the medial epicondyle
of the humerus just ventrad of
the origin of the second head and the
flexor ulnaris, to which many of its
fibres are attached. Its tendon forms
the middle and superficial part of the
common tendon.
The fifth or radial head (Fig. 75,
q; Fig. 77, u′) has origin (Fig. 87, h)
from the middle third of the ventral
surface of the shaft of the radius over
an area limited by two oblique bony
ridges; from the adjacent parts of the
interosseous membrane; and from the
ventral part of the medial surface of the
shaft of the ulna between a point about
two centimeters distad of the semilunar
notch and the junction of the
middle and distal thirds of the shaft.[183]
Its very thick tendon forms the middle and deep part of the
common tendon.
The common tendon (Fig. 88, c′)
covers the carpus and metacarpus ventrally.
It divides into five tendons (c)
which pass to the five digits and are inserted
into the bases of the terminal
phalanges. Those of the first four digits
perforate the tendon of the flexor sublimis
(b) at the base of the first phalanx of each
digit. At the same place each passes
through a fibrous pulley-ring attached to
the base of the phalanx. Each then passes
through a second pulley-ring (2) near the
head of the phalanx and is finally inserted
into the base of the terminal phalanx.
Relations.—Outer surface with the
pronator teres (Fig. 77, q), flexor carpi
radialis (Fig. 77, r), palmaris longus (s),
flexor carpi ulnaris (t), integument, and
extensor carpi ulnaris (Fig. 75, o). Inner
surface with the radius, ulna, interosseous
membrane and pronator quadratus (Fig.
87, a).
Action.—Flexor of all the digits.
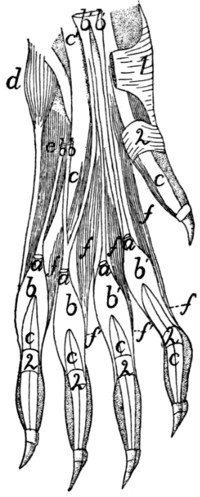
Fig. 88.—Arrangement
of the Tendons of
M. palmaris longus, M.
flexor sublimis digitorum,
and M. flexor profundus
digitorum, with
Mm. lumbricales.
The integument, fibrous
pads, and the palmaris
longus (except the ends of
its tendons) have been removed;
the flexor sublimis
has been cut. a, cut ends
of tendons of M. palmaris
longus; b, tendons of M.
flexor sublimis digitorum,
ulnar division; b′, radial
part of M. flexor sublimis
digitorum; c, tendons of
M. flexor profundus digitorum
(c′, the common tendon);
d, M. abductor digiti
quinti; e, M. flexor brevis
digiti quinti; f, Mm. lumbricales
(f′, their tendons).
1, transverse ligament of
wrist; 2, annular ligaments.
M. pronator quadratus (Fig. 87, a).—A
thick quadrangular muscle whose
fibres run obliquely between the distal
ends of the ulna and radius.
Origin by fleshy fibres from about the
distal half of the ventral (flexor) surface
or border of the ulna and from the interosseous
membrane adjacent to the area.
The fibres pass obliquely distad toward
the radial side to their
Insertion by fleshy fibres into the ventral
(flexor) surface of the radius distad of
the area of origin of the fifth head of the profundus.
[184]
Relations.—Outer surface with the flexor profundus. Inner
surface with the radius, ulna, and interosseous membrane.
Action as indicated by the name.
4. Muscles of the Hand.
A. Between the Tendons.
—Mm. lumbricales (Fig.
88, f, f′).—Four small muscles in the palm of the hand.
Origin by fleshy fibres from the outer (palmar) surface of
the tendon common to the ulnar four parts of the profundus (c′).
Insertion (f′).—The four slips are flat at their origin.
Each becomes cylindrical and curves
about the base of one of the four ulnar
digits and is inserted into the radial side of
the base of the first phalanx close to its
ventral border.
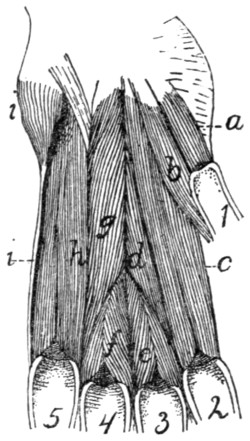
Fig. 89.—Deep Muscles
of the Palm of the
Hand.
a, M. flexor brevis pollicis;
b, M. adductor pollicis;
c, M. interosseus of second
digit (M. flexor brevis digiti
secundi); d, M. adductor
digiti secundi; e, M. interosseus
of third digit; f,
M. interosseus of fourth
digit; g, M. opponens digiti
quinti; h, M. interosseus of
fifth digit (M. flexor brevis
digiti quinti); i, M. abductor
digiti quinti. 1-5,
the digits in order.
Action.—Bend the digits toward the
radial side.
B. Muscles of the Thumb.
—M.
abductor brevis pollicis (Fig. 77, w) (includes
also the opponens pollicis of man).—A
very minute, probably rudimentary
muscle.
Origin from the transverse ligament
(1) which connects the prominent fibrous,
hairless projection that lies over the pisiform
bone with the radial border of the
head of the radius. The muscle passes
radiodistad and ends in a very slender
tendon, the insertion of which is into the
base of the first phalanx of the thumb.
M. flexor brevis pollicis (Fig. 89, a).
Origin by fleshy fibres from the adjacent
borders of the os magnum and scapholunar
bones and from the fascia which
bridges the intervening groove for the flexor carpi radialis.
Insertion by a short tendon into the base of the first
phalanx of the pollex.
Action.—Flexor of the thumb.
[185]
M. adductor pollicis (Fig. 89, b).
Origin by fleshy fibres from the ventral border of the os
magnum. It curves about the base of the first metacarpal on
its ulnar side to its insertion.
Insertion by fleshy fibres into the base of the first phalanx
of the pollex on its ulnar side.
C. Muscles Lying between the Metacarpals.
—Mm.
interossei (Fig. 89).—Small stout muscles lying on the
palmar surfaces of the metacarpals of the second, third, fourth,
and fifth digits.
Origin by fleshy fibres from the ventral or lateral surfaces
of the bases of the metacarpals. Near the distal end of the
metacarpal each divides into two masses which pass onto the
lateral surfaces of the metacarpal, so as to leave the ventral
surface of the distal end exposed.
Insertion partly onto the lateral surfaces of the base of the
first phalanx and its sesamoids, and partly by a slender tendon
(Fig. 84, h) which is continued dorsad to join the extensor
tendon of the digit.
D. Special Muscles of the Second Digit (the
Index).
—M. flexor brevis digiti secundi (or indicis).—This
name may be applied to M. interosseus of the second digit
(Fig. 89, c).
M. abductor digiti secundi.—This name might be applied
to a portion of the interosseus of the second digit which is
sometimes differentiated from the remainder.
Origin from the radial and ventral surfaces of the base of
the second metacarpal, and from the ventral surface of the
trapezium.
Insertion into the radial side of the base of the first phalanx
of the second digit and into its sesamoid.
M. adductor digiti secundi (Fig. 89, d).
Origin from the ventral surface of the os magnum.
Insertion into the ulnar side of the base of the first phalanx
of the second digit.
E. Special Muscles of the Fifth Digit.
—M. abductor
digiti quinti (Fig. 89, i).—A thick, small, conical
bundle.
[186]
Origin from the distal surface of the pisiform bone and from
the transverse ligament on the ulnar side of the pisiform. It
ends in a slender tendon which runs along the ulnar side of the
fifth metacarpal to its
Insertion into the ulnar side of the base of the first phalanx
of the fifth digit.
M. flexor brevis digiti quinti (Fig. 89, h; Fig. 88, e).—This
covers the ventral surface of the fifth metacarpal, and is
really the interosseus of this digit.
Origin by fleshy fibres from the ventral surface of the base
of the fifth metacarpal; from the ventral process of the unciform
bone, and from the transverse ligament proximad of the
fifth metacarpal.
Insertion by fleshy fibres into the ventral border of the
proximal end of the first phalanx of the fifth digit.
M. opponens (adductor) digiti quinti (Fig. 89, g).
Origin by a flat tendon from the ventral surface of the os
magnum on the ulnar side of the adductor pollicis (b). It
passes toward the ulnar side and distad, the fibres diverging to
their
Insertion (1) into nearly the whole of the radial surface of
the fifth metacarpal, (2) into the base of its first phalanx.
V. MUSCLES OF THE PELVIC LIMBS.
1. Muscles of the Hip.
A. On the Lateral Surface of the Hip.
Fascia of the Thigh.
—After the removal of the superficial
fascia with its fat and blood-vessels, there is seen a strong
glistening fascia, the fascia lata (Fig. 68, z, page 117), covering
the vastus lateralis muscle over the dorsal half of the thigh.
Ventrally it dips between the vastus lateralis and the biceps
femoris and, becoming gradually thinner, is lost on the surface
of the former muscle. Passing over the dorsal border of the
thigh, it extends beneath the sartorius and is attached to the
dorsal border of the vastus medialis. Toward its distal end
the fascia is continuous with the tendon of the biceps femoris
and dorsad with the border of the sartorius muscle, while[187]
between these points it is united with the tendon of the vastus
lateralis; it thus gains insertion into the patella. At its proximal
end the fascia receives the insertion of the tensor fasciæ
latæ (Fig. 68, r).
M. tensor fasciæ latæ (Fig. 68, r, page 117;
Fig. 92, a).—The
tensor fasciæ latæ is a thick triangular muscle which
may be recognized by its insertion into the proximal end of the
fascia lata (Fig. 68, z; Fig. 92, a′).
Origin by fleshy fibres as a thick triangular prism from the
outer margin of the ventral border of the ilium craniad of the
auricular impression and from the fascia covering the ventral
border of the gluteus medius (Fig. 90, b) over its cranial half.
The caudal border of the muscle may be continuous with the
cranial border of the gluteus maximus. The muscle spreads
out in a fan-like manner, covering the proximal third of the
dorsal surface of the thigh. The cranial fibres pass farther
distad than the others.
Insertion into the fascia lata (Fig. 68, z) along an irregular
line which begins caudad at the base of the great trochanter
and ends on the cranial side of the thigh at the junction of the
first and second thirds.
Relations.—Outer surface with the cutaneus maximus
(Fig. 62, b, page 94) and craniad with the sartorius (Fig.
68, q). Cranial border with the sartorius (q); caudodorsal
border with the gluteus medius (Fig. 90, b). Inner surface
with the gluteus medius, the rectus femoris (Fig. 92, b), and
the vastus lateralis (Fig. 90, d).
Action indicated by the name. It also assists the quadriceps
femoris in extending the leg.
M. gluteus maximus.—The gluteus maximus is a rather
small quadrangular muscle in the angle between the gluteus
medius (Fig. 90, b) and the caudal vertebræ.
Origin (Fig. 163, 3,
page 401) by fleshy fibres from the tips
of the transverse processes of the last sacral and the first caudal
vertebræ, from the fascia covering the spinous muscles dorsad
of the transverse processes of these vertebræ, and from the
fascia covering the gluteus medius. The muscle passes laterad
and slightly caudad to its
[188]
Insertion by tendon and muscle-fibres into a tubercle on the
caudal side of the great trochanter at the middle of its distal
end, and for a few millimeters into the ridge which continues
distad from this tubercle.
Relations.—Outer surface with the caudofemoralis (Fig.
68, s, page 117), the superficial fascia, and a few fibres of the
cutaneus maximus. Inner surface with the abductor caudæ
internus, the tenuissimus (Fig. 90, g), the great sciatic nerve
(Fig. 163, a), the obturator internus (Fig. 90, e), the gluteus
medius (Fig. 90, b), the gemellus superior, and the pyriformis
(Fig. 163, 7). Cranial border with the gluteus medius and
the tensor fasciæ latæ. Caudal border with the caudofemoralis
(Fig. 68, s).
Action.—Abducts the thigh.
M. gluteus medius (Fig. 90, b).—The gluteus medius is a
very large triangular muscle which connects the ilium and the
sacrum with the great trochanter (1).
Origin by fleshy fibres (1) from the superficial sacral fascia;
(2) from the lateral surface of the fascia which covers the surface
of the supraspinous extensor muscles of the tail; (3) from
the fascia intervening between it and the tensor fasciæ latæ;
(4) by tendon fibres from the dorsal half of the crest of the ilium
and its dorsal border, and the dorsal half of the lateral surface
craniad of the auricular impression; and (5) from the tips of the
transverse processes of the last sacral and the first caudal
vertebræ. The fibres converge to a strong internal tendon.
Insertion into the proximal end of the great trochanter.
Relations.—Outer surface with the gluteus maximus and
tensor fasciæ latæ, and between these with the strong fascia
lying beneath the cutaneus maximus muscle. Inner surface
with the gluteus minimus, the pyriformis, and the gemellus
superior. Cranial border with the tensor fasciæ latæ. Caudal
border with the gluteus maximus.
Action.—Abducts the thigh.
M. pyriformis (Fig. 163, 7).—The pyriformis is a triangular
muscle covered by the gluteus maximus and the gluteus
medius (Fig. 90, b) and overlying the gemellus superior (Fig.
163, 6).
[189]
Origin by fleshy fibres from the tips of the transverse
processes of the last two sacral and the first caudal vertebræ.
The muscle passes laterad through the great sciatic notch to its
Insertion by a flat tendon into an elongated area on the
proximal border of the great trochanter just outside (caudad)
of the insertion of the gemellus superior.
Relations.—Outer surface with the gluteus medius (Fig.
90, b) and the gluteus maximus. Inner surface with the
gemellus superior (Fig. 163, 6), the great sciatic nerve (Fig.
163, a), and the flexor caudæ longus (Fig. 68, x).
Action.—Abductor of the thigh.
M. gemellus superior (Fig. 163, 6).—The gemellus superior
is a triangular muscle lying beneath the pyriformis (Fig.
163, 7). It is broader than the pyriformis, so that it projects
beyond its borders both caudad and craniad. It is closely
united craniad with the gluteus minimus, so that the limits of
the two are definable only with difficulty. Its caudal border
is closely united to the gemellus inferior.
Origin by fleshy fibres from an elongated area on the dorsal
border of the ilium and ischium. The area is narrowed craniad.
It does not quite reach the posterior inferior iliac spine in the
one direction nor the spine of the ischium in the other direction.
The fibres converge to a strong tendon the
Insertion of which is into a triangular area dorsad of the tip
of the great trochanter.
Relations.—Outer surface with the gluteus medius (Fig.
90, b), the pyriformis (Fig. 163, 7), the great sciatic nerve
(Fig. 163, a), and the gluteus maximus. Inner surface with
the ilium, ischium, the capsularis, the caudal edge of the
gluteus minimus (Fig. 163, 5), and the cranial edge of the
obturator internus (Fig. 90, e).
Action.—Rotates the femur and abducts it so as to carry
the foot outward.
M. gluteus minimus (Fig. 163, 5).—The gluteus minimus
is a long triangular muscle beneath the middle of the gluteus
medius (Fig. 90, b) and with its caudal border against or
covering the cranial border of the gemellus superior (Fig.
163, 6), to which it is frequently closely united.
[190]
Origin from the ventral half of the ilium, from near its
cranial end to a point midway between the posterior iliac spine
and the spine of the ischium. The muscle ends in a strong
flat tendon.
Insertion into an oval facet at the base of the dorsal surface
of the great trochanter on its lateral side.
Relations.—Outer surface with the gluteus medius (Fig.
90, b) and at the caudal border with the gemellus superior
(Fig. 163, 6). Inner surface with the capsularis and rectus
femoris (Fig. 92, b).
Action.—Rotates the femur so as to carry the foot out.
M. capsularis (gluteus quartus, or epimeralis).—The capsularis
is a small flat bundle of muscle-fibres which lies obliquely
beneath the gluteus minimus (Fig. 163, 5) and gemellus superior
(6) on the lateral or outer surface of the ilium.
Origin by fleshy fibres from the surface of the ilium over a
triangular area between the origin of the rectus femoris (Fig.
92, b) ventrad, the gemellus superior (Fig. 163, 6) dorsad, and
of the gluteus minimus (Fig. 163, 5) craniad and the acetabulum
caudad. The muscle passes over the smooth surface of
the ilium and the pubis ventrad of the acetabulum and then
over the capsule of the joint.
Insertion by fleshy fibres for about one centimeter in the
middle line on the dorsal surface of the femur distad of the
great trochanter.
Relations.—Outer surface with the gluteus minimus (Fig.
163, 5), the gemellus superior (6),
and the vastus lateralis (Fig.
90, d). Inner surface with the ilium, the rectus femoris (Fig.
92, b), the capsule of the joint, and the vastus medialis (Fig.
92, c).
Action.—Rotates the thigh so as to carry the foot inward,
hence antagonizes the iliopsoas.
M. gemellus inferior.—The gemellus inferior is a flat triangular
muscle situated just caudad of the gemellus superior
and beneath the obturator internus (Fig. 90, e), so that it is
seen on reflecting the latter
Origin from the dorsal one-half of the whole lateral surface
of the ischium between the ischial spine and the ischial tuberosity.[191]
The fibres converge toward its insertion. The inner
surface of the muscle is covered by a strong tendon.
Insertion into the inner surface of the tendon of the
obturator internus (Fig. 90, e) by tendon- and muscle-fibres.
Some of the muscle-fibres are inserted into the capsule of the
joint.
Relations.—Outer surface with the obturator internus (Fig.
90, e). Medial surface with the ischium. Caudal border with
the quadratus femoris (Fig. 90, f). Cranial border with the
gemellus superior.
Action.—Abductor of the thigh. The muscle may be considered
as a separate head of the obturator internus (Fig.
90, e).
M. quadratus femoris (Fig. 90, f).—The quadratus femoris
is a short thick muscle connecting the ischial tuberosity (2)
and the proximal end of the femur. It lies just caudad of the
obturator internus (e) beneath the proximal end of the biceps
femoris (Fig. 68, t).
The origin is by fleshy fibres from a considerable triangular
area on the lateral surface of the ischium near the tuberosity.
The area lies between the origin areas of the gemellus inferior,
biceps, semimembranosus, and the obturator externus.
Insertion into the distal two-thirds of the ventral border of
the great trochanter and about half the adjacent surface of the
lesser trochanter.
Relations.—Outer surface with the tenuissimus (Fig. 90, g),
the biceps (Fig. 68, t), the semitendinosus (Fig. 90, j), and
the great sciatic nerve (Fig. 163, a). Caudal border with the
semimembranosus (Fig. 90, i). Ventral border with the
adductor femoris (Fig. 90, h); dorsal border with the obturator
internus (Fig. 90, e) and gemellus inferior.
Action.—Extensor of the thigh and rotator of the femur so
as to carry the foot inward.
M. obturator externus.—The obturator externus is a flat
triangular muscle beneath the adductor femoris (Fig. 92, g).
Origin by fleshy fibres from the median lip of the obturator
foramen and from both dorsal and ventral surfaces of the rami
of the pubis and ischium adjacent to the lip. Also from the[192]
outer surface of the ramus of the ischium as far as the area for
the quadratus femoris (Fig. 90, f).
The fibres converge to a strong flat
tendon.
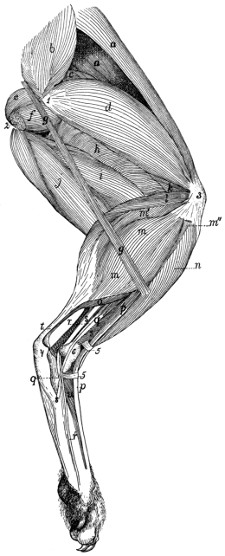
Fig. 90.—Muscles on the Lateral Side of the
Leg, after Removal of the Muscles shown in
Fig. 68 (Biceps, Tensor fasciæ latæ, Caudofemoralis,
and Gluteus maximus).
a, M. sartorius; b, M. gluteus medius; c, M. rectus
femoris; d, M. vastus lateralis; e, M. obturator internus;
f, M. quadratus femoris; g, M. tenuissimus; h, M. adductor
femoris; i, M. semimembranosus; j, M. semitendinosus;
k, M. vastus intermedius; l, M. plantaris;
m, m′, m″, M. gastrocnemius, outer head (m, part from
the external sesamoid bone; m′, from the plantaris; m″,
from the superficial fascia); n, M. tibialis anterior; o, M.
soleus; p, M. extensor longus digitorum; q, M. peroneus
longus; r, M. peroneus tertius (r′, its tendon); s, M.
peroneus brevis; t, tendon of Achilles; u, M. extensor
brevis digitorum. 1, great trochanter of femur; 2,
tuberosity of ischium, with cut origin of M. biceps
femoris; 3, patella; 4, calcaneus; 5, transverse ligaments;
6, lateral malleolus.
Insertion into the proximal portion
of the bottom of the trochanteric
fossa.
Relations.—Ventral surface
with the adductor femoris
(Fig. 92, g). Dorsal
surface with the pubis
and ischium. Caudal
border with the quadratus
femoris (Fig.
90, f).
Action.—Assists
the iliopsoas.
B. Muscles on
the Medial Surface
of the Hip.
—M.
obturator internus
(Fig. 90, e).—The obturator
internus appears as a triangular
muscle caudad of the gemellus
superior.
[193]
Origin by numerous small separate heads from the dorsal
surface of the ramus of the ischium along its symphysis and
following its medial border from the symphysis nearly to the
tuberosity. The fibres form a flat muscle which narrows and
passes through the lesser sciatic notch, turns ventrad and ends
in a strong flat tendon whose
Insertion is into the bottom of the trochanteric fossa of the
femur. The tendon is continued proximad on the inner surface
of the muscle and forms a smooth firm surface by which the
muscle glides over the dorsal border of the ischium. Into the
inner surface of this tendon near its insertion the tendon of the
gemellus inferior is inserted.
Relations.—Medial surface within the pelvis with a mass of
fat separating it from the pelvic organs. Dorsal or outer surface
with the biceps femoris, the tenuissimus, the caudofemoralis,
and the great sciatic nerve. Inner surface with the ischium
and the great sciatic nerve. Caudal (or ventral) border with
the quadratus femoris.
Action.—Abductor of the thigh.
M. iliopsoas (Fig. 162, 8;
Fig. 91, c).—The iliopsoas
(equivalent to the human psoas and iliacus) is a conical muscle
emerging from the abdominal cavity onto the medial surface of
the femur.
Origin.—(a) The portion corresponding to the human psoas
(Fig. 162, 8) arises by ten vertebral heads. The first five of
these come from the five cranial tendons of origin of the psoas
minor (Fig. 162, 9); the sixth from a tendinous expansion
which passes from the tendon of the first head over the ventral
longitudinal muscles to the transverse process of the fifth
lumbar vertebra. The seventh, eighth, ninth, and tenth heads
arise by fleshy fibres from the ventral surfaces of the centra of
the last four lumbar vertebræ.
(b) The portion corresponding to the human iliacus arises
by fleshy fibres from the ventral border of the ilium, from
opposite the auricular impression to the iliopectineal eminence.
The portions of the muscle all converge to form a conical
mass which ends in a strong tendon lying on its outer surface.
[194]
Insertion by tendon and fleshy fibres into the apex of the
lesser trochanter of the femur.
Relations.—Dorsal surface with the quadratus lumborum
(with which this muscle is partly united), the rectus femoris
(Fig. 91, d), and, by the iliac head, with the gluteus minimus.
Ventral and medial surface with the psoas minor (Fig. 162, 9)
and the peritoneum. Lateral edge with the transversus
abdominis (Fig. 162, 4).
Action.—Rotates the thigh so as to carry the foot out; also
flexes the thigh.
2. Muscles of the Thigh.
M. biceps femoris (Fig. 68, t, page 117).—A very large
flat muscle covering about two-thirds of the lateral side of the
thigh.
Origin (Fig. 90, 2).—From the ventral surface of the
tuberosity of the ischium by tendon- and muscle-fibres. The
fibres diverge, and near the knee the mass has spread out, ending
in a fascia. The dorsal border of the muscle and the
common fascia are continuous dorsad with the superficial fascia
of the thigh and with the tendon of the caudofemoralis (Fig.
68, s). Ventrad it is continuous with the superficial fascia of
the shank.
Insertion into rather more than the proximal one-third of
the dorsal border of the tibia along its lateral margin and into
the lateral margin of the patella. In passing over the knee-joint
it is closely united to the underlying ligaments and
tendons.
Relations.—Outer surface with the superficial fascia and
with a few of the most caudal fibres of the cutaneus maximus
(Fig. 62, b, page 94). Cranial (or dorsal) edge with the
caudofemoralis (Fig. 68, s) and the vastus lateralis (Fig. 90, d).
Caudal border with the semitendinosus (Fig. 68, u) and a mass
of fat. Inner surface with the caudofemoralis (Fig. 68, s), the
tenuissimus (Fig. 90, g), the obturator internus (Fig. 90, e),
the quadratus femoris (Fig. 90, f), the semitendinosus (Fig.
68, u), the semimembranosus (Fig. 90, i), the adductor femoris
(Fig. 90, h), the great sciatic nerve
(Fig. 163, a), and distad[195]
with the following muscles of the lower leg: the tibialis anterior
(Fig. 90, n), the extensor longus digitorum (Fig. 90, p), the
peroneus longus (Fig. 90, q), and the lateral head of the
gastrocnemius (Fig. 90, m).
Action.—Abductor of the thigh, and flexor of the shank.
M. tenuissimus or M. abductor cruris (Fig. 90, g).—A
very slender muscle, only three or four millimeters wide.
Origin from the tip of the transverse process of the second
caudal vertebra, in common with the caudofemoralis (Fig.
68, s) or gluteus maximus. It passes obliquely beneath the
biceps femoris (Fig. 68, t) distad and ventrad, to the distal end
of the ventral border of that muscle. Here it becomes continuous
with the ventral border of the biceps, ending in a continuation
of the same fascia into which the biceps is inserted.
Relations.—Outer surface with the caudofemoralis (Fig.
68, s) and the biceps femoris (Fig. 68, t); distad with the
integument. Inner surface with the obturator internus (Fig.
90, e), quadratus femoris (f), semitendinosus (j), adductor
femoris (h), semimembranosus (i), and distad with the muscles
of the lower leg.
M. caudofemoralis (parameralis, Strauss-Durckheim) (Fig.
68, s, page 117).
Origin by a flat tendon from the transverse processes of the
second and third caudal vertebræ. The muscle forms a flat
band which passes distad along the middle of the lateral side
of the thigh over the pelvis and caudad of the great trochanter.
At the middle of the thigh it ends in a very thin tendon. The
tendon passes distad along the medial surface of the biceps
femoris (Fig. 68, t), pierces the fascia lata near the knee, and
passes to its
Insertion into the middle of the lateral border of the patella.
Relations.—Outer surface with the superficial fascia and a
few fibres of the cutaneus maximus; distad with the biceps
femoris (Fig. 68, t). Cranial border with the gluteus maximus;
caudal border with the biceps femoris. Inner surface
with the tenuissimus (Fig. 90, g), the gluteus maximus, the
abductor caudæ internus, and with the obturator internus (Fig.
90, e); distad with the vastus lateralis.
[196]
Action.—Abducts the thigh and helps to extend the shank.
M. semitendinosus (Fig. 90, j).—A long slender muscle
on the ventral (caudal) border of the thigh, between the semimembranosus
(i) and the biceps femoris (Fig. 68, t).
Origin from the apex of the tuberosity of the ischium
beneath the origin of the biceps femoris. The muscle passes
to the medial side of the shank and ends in a thin but strong
tendon (Fig. 92, i′) about five millimeters broad. The tendon
curves proximad and passes beneath the gracilis tendon (Fig.
91, b) to its
Insertion into the crest (dorsal border) of the tibia one or
two centimeters from its proximal end.
Relations.—Lateral surface with the biceps femoris (Fig.
68, t), the integument, a mass of fat, and distad with the gastrocnemius
(Fig. 92, j) and popliteus (Fig. 92, k). Caudal
surface with the integument. Medial surface with the semimembranosus
(Fig. 90, i) and distad with the integument of
the lower leg.
Action.—Flexor of the shank.
M. semimembranosus (Figs. 90 and 91,
i; Fig. 92, h).—A
thick prismatic muscle lying along the ventral (or caudal)
side of the thigh between the semitendinosus (j) and the gracilis
(Fig. 91, b).
Origin by short tendon-fibres from the caudal border of the
tuberosity and the ramus of the ischium. The muscle mass is
divided throughout most of its length into two portions (Fig.
92, h and h′), one of which (h′) lies laterad and caudad of the
other. The two portions pass to the medial side of the knee.
The caudal portion (h′) ends in a strong flat tendon five millimeters
broad, the
Insertion of which is into the medial surface of the femur
on the medial epicondyle, at the middle of its distal border and
into the adjacent medial surface of the tibia behind the lateral
ligament.
The dorsal part has its insertion into the distal one and
one-half to two centimeters of the ridge which is continued
from the medial epicondyle of the femur onto the shaft; and
into the sesamoid bone of the medial epicondyle.
[197]
Relations.—Lateral surface with the semitendinosus (Fig.
90, j), the biceps femoris (Fig. 68, t), and distad with a small
part of the adductor femoris (Fig. 92, g), and with the gastrocnemius
(Fig. 92, j). Medial
surface with the gracilis (Fig.
91, b) and distad with the
sartorius (Fig. 91, a). Cranial
(or dorsal) border at
the origin with the quadratus
femoris (Fig. 90,
f); for the remainder
of its length with the
adductor femoris (Fig.
90, h). Caudal border
with the integument
proximad.
Action.—Extensor
of the thigh.

Fig. 91.—Muscles on the Medial Side of
the Leg.
a, M. sartorius; b, M. gracilis; c, M. iliopsoas; d, M.
rectus femoris; e, M. vastus medialis; f, M. pectineus;
g, M. adductor longus; h, M. adductor femoris; i, M.
semimembranosus; j, M. semitendinosus; k, medial
head of M. gastrocnemius; l, M. plantaris; m, M. flexor
longus hallucis (m′, its tendon); n, M. flexor longus
digitorum; (n′, its tendon); o, M. tibialis posterior; p,
M. tibialis anterior; q, M. interosseus of second digit.
1, patella; 2, calcaneus; 3, medial malleolus; 4, second
metatarsal; 5, transverse ligament of the lower leg.
M. sartorius (Fig. 91,
a; Fig. 68, q).—A large flat
muscle along the medial side
of the thigh near its cranial
(dorsal) border.
Origin from the ventral half of
the crest of the ilium and from the
medial half of its ventral border craniad
of the auricular impression. The caudal
third of the origin is by a thin tendon,
the rest by fleshy fibres.
[198]
Insertion on a long S-shaped line which begins on the
dorsal border of the tibial shaft about two centimeters from its
proximal end, passes thence to the middle of the proximal end
of the medial surface of the tibia, thence across the ligaments
of the knee-joint and the medial surface of the medial epicondyle
to the patella, crosses the patella obliquely to the middle of its
proximal end, and may be continued for some distance proximad
along the middle line on the fascia.
Relations.—Medial surface with the integument and proximad
with the internal oblique muscle (Fig. 68, p). Lateral
(inner) surface with the tensor fasciæ latæ (Fig. 68, r), the
vastus lateralis (Fig. 90, d), the superficial fascia, the vastus
medialis (Fig. 92, c), the rectus femoris (Fig. 92, b), and the
distal end of the semimembranosus (Fig. 92, h′).
Action.—Adducts and rotates the femur and extends the
tibia.
M. gracilis (Fig. 91, b).—The gracilis is a flat, thin, subcutaneous
muscle occupying the ventral half of the medial surface
of the thigh.
Origin by a strong tendon from the caudal three-fourths of
the symphysis of the ischium and pubis. The tendon may
extend caudad of the symphysis in the middle line to the level
of the tuberosity of the ischium. For about its first centimeter
the tendon is common to the two muscles of opposite sides and
gives origin on two sides to the fibres of the adductor femoris.
The common tendon divides into two which pass laterad, each
ending in a convex border from which the muscle-fibres
spring.
Insertion.—It ends in a thin aponeurosis, part of which is
continuous with that of the sartorius. The aponeurosis is finally
inserted into the medial surface of the tibia near its proximal
end, while distally it is continuous with the fascia.
Relations.—Outer (medial) surface with the integument.
Inner (lateral) surface with the semimembranosus (Fig. 91, i)
and the adductor femoris (Fig. 91, h).
Action.—Adducts the leg and draws it caudad.
M. adductor femoris (magnus et brevis) (Fig. 92, g; Fig.
90, h).—The adductor femoris lies between the semimembranosus[199]
(Fig. 92, h) and the femur. Its proximal part is covered
by the semimembranosus, while its distal part lies outside of it.
Origin by muscle-fibres from the rami of the pubis and the
ischium along the whole of the length of the symphysis; from
the ramus of the ischium between the symphysis and the
tuberosity, and from the tendon of origin common to the two
gracilis muscles.
Insertion into nearly the whole of the ventral surface of the
shaft of the femur. The area begins at the base of the great
trochanter opposite the proximal end and the insertion area of
the gluteus maximus. It extends thence distad along the
caudal border of the surface as an area about one millimeter
wide. At the junction of the first and second thirds of the
shaft the area expands and occupies the whole of the ventral
surface of the bone, ending at the intercondyloid fossa.
Relations.—Lateral surface with the biceps (Fig. 68, t), the
great sciatic nerve (Fig. 163, a), the tenuissimus (Fig. 90, g),
and the semimembranosus (Fig. 92, h); dorsolateral border in
contact with the vastus lateralis (Fig. 92, c). Dorsal surface
with the quadratus femoris (Fig. 90, f) and obturator externus
(Fig. 90, e). Caudal surface with the semimembranosus (Fig.
92, h). Medial surface with the gracilis (Fig. 91, b), adductor
longus (Fig. 92, f), pectineus (Fig. 92, e), vastus medialis
(Fig. 92, c), and distad with the medial head of the gastrocnemius
(Fig. 92, j).
Action.—An extensor of the thigh.
M. adductor longus (Fig. 91, g; Fig. 92, f).—A thin
muscle which covers the dorsal half of the medial surface of the
adductor femoris (Fig. 92, g).
Origin by muscle-fibres from the median three-fourths of
the cranial border of the pubis, the line of origin forming a
medial continuation of that of the pectineus (Fig. 92, e).
Insertion by a thin aponeurosis into the external linea
aspera of the femur along the second and third fifths of the
bone.
Relations.—With its medial or cranial surface the adductor
longus forms part of the boundary of a depression among the
muscles at the proximal end of the medial side of the leg.[200]
This depression is called the iliopectineal fossa; it contains the
femoral vein and artery and saphenous nerve imbedded in fat
(Fig. 127). The medial edge of the adductor longus is in
relation with the integument; the lateral edge with the pectineus
(Fig. 92, e). Inner or caudal surface with the adductor
femoris (Fig. 92, g).
Action.—Adductor of the thigh.
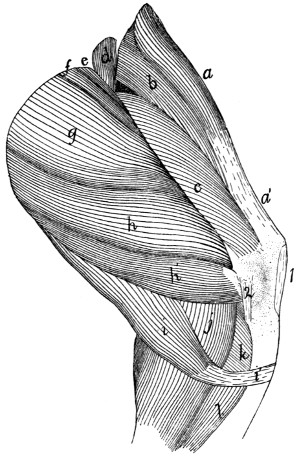
Fig. 92.—Second
Layer of Muscles on the Medial Side of the Thigh.
a, M. tensor fasciæ latæ; a′,
fascia lata; b, M. rectus femoris; c, M. vastus medialis;
d, M. iliopsoas (cut); e, M. pectineus; f, M. adductor longus; g, M. adductor
femoris; h, h′, M. semimembranosus; i, M. semitendinosus (i′, its tendon); j, medial
head of M. gastrocnemius; k, M. popliteus; l, M. flexor longus digitorum. 1, patellar
ligament; 2, ligamentum collaterale tibiale.
M. pectineus (Fig. 92, e).—A flat band of fibres closely
united with the adductor longus (f), of which it appears to be
a lateral continuation.
Origin by fleshy fibres from the lateral one-fourth of the
cranial border of the pubis. The muscle passes over the
smooth outer surface of the pubis between its origin area and[201]
the acetabulum, crosses the iliopsoas (d) obliquely and has its
Insertion by muscle-fibres into an elongated area (five millimeters
in length) on the shaft of the femur just distad of the
lesser trochanter and between the insertion of the adductor
femoris (g) and that of the vastus medialis (c).
Relations.—The cranial edge forms part of the floor of the
iliopectineal fossa. Lateral surface in relation with the iliopsoas
(d) and vastus medialis (c); medial surface with the adductor
longus (f) and adductor femoris (g).
Action.—Adductor of the thigh.
M. quadriceps femoris.—The quadriceps femoris is a very
powerful extensor muscle on the front of the thigh. It consists
of four muscles which unite to form the great extensor of the
shank. It is inserted into the patella and through it, by the
ligamentum patellæ, into the tibia. It may be compared to the
triceps brachii. The parts are:
(1) M. rectus femoris (Fig. 92, b; Fig. 90, c).
Origin by strong tendon from an elongated triangular area
which has its base at the acetabulum and its apex about five
to seven millimeters craniad of the acetabulum, along the ventral
border of the ilium. The area is between that for the
capsularis and that for the iliopsoas. The muscle is flat near its
origin, but soon becomes a triangular prismatic mass which is
united to the dorsal border of the vastus lateralis (Fig. 90, d)
at the junction of the middle and last thirds of the thigh.
Insertion into the oblique area on the outer surface of the
patella near its proximal border in connection with the vastus
lateralis.
Relations.—Outer or lateral surface at the origin with the
gluteus minimus, the edge of the capsularis, the gluteus medius
(Fig. 90, b), and farther distad with the vastus lateralis (Fig.
90, d). Medial surface with the tensor fasciæ latæ (Fig. 92, a),
sartorius (Fig. 90, a), and vastus medialis (Fig. 92, c). Inner
(caudal) surface with the vastus intermedius.
(2) M. vastus lateralis (Fig. 90, d).—A flat triangular
prismatic mass joined distally to the preceding. It covers the
dorsal part of the lateral surface of the thigh.
Origin from a triangular area on the dorsal and lateral surfaces[202]
of the shaft and the great trochanter of the femur. The
base of the area is at the great trochanter, and its apex is at the
junction of the second and third fifths of the shaft on the linea
aspera. The linea aspera forms the ventral boundary of the
triangle, while its dorsal boundary is formed by a line drawn
from its apex to the dorsomedial angle of the great trochanter.
The mass unites with the rectus femoris (c) by its dorsal border
at the junction of the middle and distal thirds of the thigh.
Insertion into the oblique area on the outer surface of the
patella near its lateral border in connection with the rectus
femoris.
(3) M. vastus medialis (Fig. 92, c) lies on the medial side
of the thigh.
Origin by fleshy fibres over a diamond-shaped area on the
shaft of the femur lying between the medial branch of the linea
aspera and the area for the vastus lateralis. Proximad the
area is bounded by the spiral line, and distad it is bounded by
a line parallel to the spiral line and having its proximal end at
about the junction of the first and second thirds of the bone.
The muscle forms a triangular, prismatic mass. It ends in a
thin aponeurosis which is continuous dorsad with the border of
the rectus femoris (Fig. 92, b), and ventrad sometimes with the
tendon of the gracilis (Fig. 91, b).
Insertion into the medial border of the patella and the ligamentum
patellæ. The insertion may extend onto the head of
the tibia.
The adjacent surfaces of the vastus lateralis and the vastus
medialis are connected over their distal thirds by an aponeurosis
which passes transversely beneath the rectus femoris. The
distal end of the aponeurosis is inserted into the proximal
border of the patella. Its proximal end receives the insertion
of some muscle-fibres which form a part of the vastus medialis
as here described. This mass of fibres is described by Strauss-Durckheim
as the crural muscle. Sometimes the transverse
aponeurosis is continuous with only one of the two vasti and
sometimes with neither, so that the insertion of the crural of
Strauss-Durckheim becomes more or less independent.
Relations of the vastus medialis.—Outer (medial) surface[203]
with the sartorius (Fig. 91, a) and the structures in the iliopectineal
fossa. Cranial (or dorsal) surface with the rectus
femoris (Fig. 92, b) and vastus intermedius. Caudal (ventral)
surface with the pectineus (Fig. 92, e), adductor longus (Fig.
92, f), adductor femoris (Fig. 92, g), and semimembranosus
(Fig. 92, h).
(4) M. vastus intermedius.—A flat mass of muscle which
lies beneath the rectus femoris.
Origin from nearly the whole of the dorsal surface of the
shaft of the femur between the areas for the vastus medialis and
vastus lateralis. The area of origin extends distad to an
oblique line the middle of which is about one and one-half
centimeters from the patellar surface, the lateral border of the
area being longer than the medial border.
Insertion by muscle-fibres into the capsule of the joint.
The central fibres are inserted about one centimeter proximad
of the patella, while the lateral mass passes to the level of the
proximal end.
Relations.—Outer surface with the rectus femoris (Fig.
92, b); borders with the vastus medialis (Fig. 92, c) and vastus
lateralis (Fig. 90, d); inner surface with the femur.
Action.—The quadriceps extensor is an extensor of the
shank. The vastus intermedius acting separately is a tensor
of the capsule of the knee-joint.
3. Muscles of the Lower Leg (Crus).
A. On the Ventral Side.
—M. gastrocnemius.—This
is the great muscular mass of the calf. It arises by two heads,
the caput laterale or lateral head (Fig. 90, m), and the caput
mediale or medial head (Fig. 91, k), of the gastrocnemius.
Caput laterale (Fig. 90, m, m′, m″).—The origin of the
lateral head is in four portions:
(1) From the tendon of origin of the plantaris which comes
from the middle of the lateral border of the patella. The tendon
ends in a small conical mass of fibres (m′) which joins the
ventral part of the common head.
(2) By a slender flat tendon, distad of the first, from the[204]
superficial fascia of the shank (m″). The tendon ends in a
conical mass which joins the dorsal border of the common head.
(3) By a strong tendon and by muscle-fibres from the distal
border of the lateral sesamoid bone (Fig. 61, 3) of the femur.
These fibres (m) form the middle and outer part of the common
head.
(4) By muscle-fibres from the middle of the outer surface
of the aponeurosis covering the plantaris (Fig. 90, l) along
rather more than the proximal half of the tibia. The lateral
head thus formed is flat and fusiform in section.
Caput mediale (Fig. 91, k; Fig. 92, j).—The medial head
resembles the lateral head closely.
Origin by a strong tendon from the medial sesamoid bone
of the femur (Fig. 61, 4) and by muscle-fibres from the surface
of the shaft for a short distance proximad of this.
The two heads unite by their contiguous borders at the
junction of the first and second thirds of the tibia. The
common muscle narrows rapidly and ends in a flat tendon
which joins the tendon of the soleus to form the so-called tendon
of Achilles (tendo calcaneus) (Fig. 90, t). This has its
Insertion into the proximal end of the calcaneus (Fig.
90, 4) near its ventral border. This tendon together with that
of the soleus (Fig. 90, o) and the fascia of the shank forms a
tubular sheath for the tendon of the plantaris.
Relations.—(1) Of the lateral head. Lateral (outer) surface
with the biceps femoris (Fig. 68, t) and the superficial fascia.
Medial (inner) surface with the popliteus (Fig. 92, k), the
plantaris (Fig. 90, l) (to which it is closely united), the medial
head of the gastrocnemius (Fig. 91, k), the soleus (Fig. 90, o),
and the peroneus longus (Fig. 90, q). (2) Of the medial head.
Medial (outer) surface with the superficial fascia and the tendons
of the gracilis (Fig. 91, b), semimembranosus (Fig. 92, h′),
and semitendinosus (Fig. 92, i). Lateral (inner) surface at
the origin with the adductor femoris (Fig. 92, g), then with the
plantaris (Fig. 91, l), the popliteus (Fig. 92, k), flexor longus
digitorum (Fig. 91, n), and the lateral head of the gastrocnemius
(Fig. 90, m).
Action.—Extensor of the foot. (It is to be noted that what[205]
is called extension in the foot is analogous to what is called
flexion in the hand, so that the extensors of the foot are most
readily to be compared with the flexors of the hand.)
M. plantaris (Fig. 90, l; Fig. 91, l).—A fusiform muscle
covered distally by the gastrocnemius, but appearing between
the two heads of the gastrocnemius at their proximal ends
(Fig. 90, l).
Origin by a strong tendon from the middle of the lateral
border of the patella and by fleshy fibres from the ventral
border of the lateral sesamoid.
The patellar tendon ends in a flattened head which passes
ventrad over the lateral epicondyle of the femur and then
curves distad; it is closely united with the lateral head of the
gastrocnemius. The muscle ends in a thick tendon which
passes through a sheath formed by the tendons of the gastrocnemius
and soleus, and the fascia of the shank. The tendon
passes over the grooved proximal end of the calcaneus onto its
ventral surface. It is held in place against the groove by two
sheets of aponeurosis, which leave the tendon on either side for
a distance of more than a centimeter and are attached to the
lateral surfaces of the calcaneus at its proximal end. Lateral
motion is thus hindered, while longitudinal motion is permitted.
Between this tendon and the tendon of Achilles is a synovial
bursa which aids the gliding movement of the tendon. In the
sole of the foot opposite the distal end of the calcaneus the
tendon broadens and ends in the flexor brevis digitorum (pedis
perforatus) muscle, which might also be considered as a second
part of the plantaris. In man the plantaris is inserted into the
calcaneus along with the gastrocnemius.
Relations.—The muscle is covered throughout by the two
heads of the gastrocnemius (Fig. 90, m; Fig. 91, k), except
at the proximal end (Fig. 90, l), where it is in contact with the
integument. Inner surface in relation with the popliteus (Fig.
92, k), the soleus (Fig. 90, o), the flexor longus digitorum
(Fig. 91, n), and flexor longus hallucis (Fig. 91, m).
M. soleus (Fig. 90, o).—A flat fusiform muscle lying
beneath the plantaris.
Origin by muscle-fibres from the lateral surface of the head[206]
of the fibula, and by tendon-fibres from the proximal two-fifths
of its ventral border.
Insertion.—The muscle ends in a slender tendon which
joins the lateral border of the gastrocnemius tendon to form the
tendon of Achilles (Fig. 90, t), which forms the sheath of the
plantaris tendon.
Relations.—Outer surface with the gastrocnemius (Fig. 90,
m) and plantaris (Fig. 90, l). Inner surface with the peroneus
longus (Fig. 90, q), peroneus tertius (Fig. 90, r), and flexor
hallucis (Fig. 91, m).
Action.—Assists the gastrocnemius to extend the foot.
M. triceps suræ.—The gastrocnemius and the soleus are
sometimes considered as forming a single muscle, with three
heads, under the name triceps suræ.
M. popliteus (Fig. 92, k).—A triangular muscle passing
from the femur obliquely toward the medial side over the ventral
surface of the proximal end of the tibia.
Origin by a strong tendon from the popliteal groove on the
surface of the lateral epicondyle of the femur. In the tendon
is a sesamoid bone, the popliteal bone (Fig. 61, 5, page 89),
which glides over the ventral part of the lateral articular facet
on the proximal end of the tibia. As the muscle passes over
the joint it is closely attached to the joint capsule. From the
popliteal bone, which with the tendon lies within the capsule
of the knee-joint, the muscle-fibres diverge to their
Insertion into the proximal end of the ventral surface of the
tibial shaft on the medial side of the medial oblique ridge.
The area of insertion extends slightly onto the medial surface
of the bone.
Relations.—Outer surface with the gastrocnemius (Fig.
92, j) and plantaris (Fig. 91, l), and with the tendon of the
semitendinosus (Fig. 92, i). Distal border with the origins
of the soleus and flexor longus digitorum. Inner surface with
the capsule of the knee-joint and the tibia.
Action.—Rotates the thigh so as to turn the toes inward.
The three following deep muscles on the ventral surface of
the shank are covered by a deep fascia (the deep crural fascia)[207]
which separates them from the overlying muscles. This fascia
stretches from the dorsal border of the tibia about the ventral
side of the shank to the medial border of the tibia. It sends
a slip beneath the border of the fibula.
M. flexor longus digitorum (Fig. 91, m and n).—Owing
to the reduction of the first digit in the cat the tendon of the
flexor longus hallucis (m) has become united to that of the
flexor longus digitorum (n), so that the two might be considered
separate heads of a single muscle. The head corresponding
to the flexor longus hallucis (m) is much larger than that corresponding
to the flexor longus digitorum (n). The two heads
will be described separately.
(1) M. flexor longus hallucis (Fig. 91, m).—This lies
against the ventral surface of the tibia and fibula beneath the
popliteus (Fig. 92, k) and soleus (Fig. 90, o).
Origin by fleshy fibres, (1) from the sheet of deep crural
fascia which dips between it and the tibialis posterior (Fig.
91, o) and the flexor longus digitorum (n). (2) From the ventral
surface of the tibia distad of the lateral oblique line to within
one to three centimeters of the distal end of the shaft. (3) From
the medial surface of the shaft and head of the fibula and from
the whole interosseous ligament. On the fibular side the origin
may extend onto the tendon of the popliteus and the fascia of
the peroneus longus. The fibres form a flat fusiform mass
ending one centimeter from the heel in a strong flat tendon
(m′) which passes over the groove on the distal surface of the
astragalus and over the groove on the sustentaculum tali. The
two grooves are converted into a continuous canal by strong
transverse ligaments, and the canal is lined by an extension of
the synovial bursa of the ankle-joint. Emerging from the
canal the tendon broadens and receives on its medial side the
insertion of the tendon of the flexor longus digitorum (n). The
tendon gives origin on its outer surface to the lumbricales and
to the common plantar ligament. The tendon continues to
broaden until it reaches the middle of the length of the metacarpals;
here it divides into four tendons which pass to the
terminal phalanges of the digits.
Insertion.—The relations of the four tendons and their[208]
insertions are identical with those of the flexor profundus digitorum
of the hand.
Relations.—Outer surface with the soleus (Fig. 90, o) and
plantaris (Fig. 91, l). Lateral border with the soleus, the
peroneus tertius (Fig. 90, r), and peroneus brevis (Fig. 90, s).
Medial border with the tibialis posterior (Fig. 91, o) and flexor
longus digitorum (Fig. 91, n).
Action.—Flexor of the phalanges.
(2) M. flexor longus digitorum (Fig. 91, n).
Origin by muscle- and tendon-fibres from the ventral surface
of the tibia over its proximal half between the oblique
ridges; by fleshy fibres from an aponeurosis between it and the
tibialis posterior (Fig. 91, o), and by tendon from the medial
surface of the head of the fibula. At the junction of the middle
and distal thirds of the bone the muscle ends in a slender tendon
which passes through the ventral groove on the medial
surface of the tibia and, curving onto the sole of the foot,
becomes attached to the medial border of the common tendon
described under the last.
Relations.—Outer surface with the medial head of the gastrocnemius
(Fig. 91, k) and with the superficial fascia. Inner
surface with the tibialis posterior (Fig. 91, o). Medial border
at the proximal end with the popliteus (Fig. 92, k), lateral
border with the tibialis posterior (Fig. 91, o), and flexor longus
hallucis (Fig. 91, m).
Action.—Flexor of the phalanges.
M. tibialis posterior (Fig. 91, o).—A slender flat fusiform
muscle beneath the flexor longus digitorum and between it and
the flexor longus hallucis.
Origin by fleshy fibres from nearly the whole medial surface
of the head of the fibula, from the inner surface of the aponeurosis
between it and the flexor longus digitorum (n), and by
a few fleshy fibres from the ventral surface of the tibia between
the oblique ridges. Some fibres may also arise from the outer
surface of the aponeurosis covering the flexor longus hallucis
(m). The muscle ends at about the middle of the tibia in a
slender flat tendon which passes parallel to the tendon of the
flexor longus digitorum (n) through the dorsal groove on the[209]
medial surface of the distal end of the tibia. This groove is
converted into a canal by a transverse ligament. Beyond the
canal the tendon turns onto the plantar surface of the foot and
passes through a groove on the ventral surface of the scaphoid
bone. It then divides.
Insertion into the outer tuberosity on the surface of the
scaphoid, and onto the proximal end of the ventral surface of
the medial cuneiform.
Relations.—Outer and medial surface with the flexor longus
digitorum (n). Lateral surface with the flexor longus hallucis (m).
Action.—Extensor of the foot.
B. Muscles on the Dorsal and Lateral Surfaces
of the Lower Leg.
—M. peroneus longus (Fig. 90, q).—A
slender, fusiform muscle lying superficially on the lateral side
of the leg, dorsad of the soleus.
Origin by tendon-fibres from the lateral surface of the head
of the fibula and from the proximal half of the lateral surface
of its shaft.
At the junction of the middle and distal thirds of the lower
leg the muscle ends in a slender tendon which passes through
the groove on the lateral surface of the lateral malleolus. This
groove is converted into a canal by a transverse ligament.
The tendon passes through the groove on the peroneal tubercle
of the calcaneus, then turns onto the sole of the foot and passes
through the peroneal groove on the cuboid bone. It then turns
mediad and passes through the groove between the ventral
processes of the lateral cuneiform and the metatarsal until it
reaches the first metatarsal. The entire groove is converted
into a canal by the overlying ligaments.
Insertion into (1) the base of the fifth metatarsal, and (2)
the outer side of the base of the first metatarsal, and (3) by
slender branches into the bases of the other metatarsals.
Relations.—Outer (lateral) surface with the superficial fascia
and the tendon of the biceps femoris. Inner surface with the
peroneus tertius (r) and peroneus brevis (s), and with the
extensor longus digitorum (p). At the proximal end the
dorsal border touches the tibialis anterior (n).
Action.—Flexor of the foot.
[210]
M. peroneus tertius (Fig. 90, r).—A slender fusiform
muscle beneath the peroneus longus (q).
Origin by fleshy fibres from about the second quarter of the
lateral surface of the fibula. The muscle ends in a slender
tendon (r′), which passes with that of the peroneus brevis (s)
through the groove on the ventral border of the lateral malleolus.
The groove is converted into a canal by a transverse
ligament. Emerging from it, the tendon turns and passes along
the outer margin of the foot. In passing the sesamoid at the
base of the first phalanx of the fifth digit it is united to it by a
band which passes from the sesamoid to the tendon.
The insertion is finally into the lateral border of the extensor
tendon of the fifth digit as it passes from the first to the second
phalanx.
Relations.—Outer surface with the peroneus longus (q), the
soleus (o), and the superficial fascia. Ventral border with the
soleus (o) and flexor longus hallucis (Fig. 91, m). Inner surface
with the peroneus brevis (Fig. 90, s).
Action.—Extensor and abductor of the fifth digit and flexor
of the foot.
M. peroneus brevis (Fig. 90, s) lies beneath the other
peronei.
Origin by fleshy fibres from the distal half of the surface of
the fibula, which it embraces. At the lateral malleolus the
muscle ends in a thick tendon which passes through the canal
on the ventral surface of the malleolus. In the canal it is
enlarged and surrounded by a synovial bursa. It then turns
onto the foot and passes over the dorsal surface of the calcaneus
on its lateral side and has its
Insertion into the tubercle on the lateral side of the base of
the fifth metatarsal.
Relations.—Outer surface with the peronei longus (q) and
tertius (r). Inner surface with the bone.
Action.—Extensor of the foot.
M. extensor digitorum longus (Fig. 90, p).—A fusiform
muscle which lies beneath the tibialis anterior (n), against the
interosseous membrane, and between the tibialis anterior and
the peronei.
[211]
Origin by a thin flat tendon from the lateral surface of the
lateral epicondyle of the femur just dorsad of the origin of the
popliteus. The tendon becomes narrower and thicker and
passes through the capsule of the knee-joint and over a slight
groove on the tibia just dorsad of the head of the fibula. The
belly of the muscle extends the entire length of the tibia, passing
beneath the transverse ligament (5) along with the tendon
of the tibialis anterior (n). At the ankle it runs through a
fibrous loop which is attached to the dorsal surface of the
calcaneus (the annular ligament of the calcaneus); it then turns
and passes onto the dorsum of the foot. The muscle ends in
four slender tendons, the two middle of which begin at the
transverse ligament, while the others begin at the fibrous loop.
The tendons diverge to the four digits. Each tendon is connected
with the fibrous sheath surrounding the base of the first
phalanx, and each has a synovial bursa beneath it at this point.
Near the distal end of the first phalanx the tendon is united on
the lateral side to the conjoined tendon of the extensor brevis
digitorum and an interosseus muscle, and on the medial side
to the tendon of the interosseus. The lateral side of the most
lateral tendon is not thus united, but is joined on the outer side
by the tendon of the peroneus tertius.
Insertion.—The tendon thus formed on the dorsum of each
phalanx by the junction of the two or three tendons above
mentioned is firmly attached to the base of the second phalanx
at its proximal end in the middle of its dorsal surface. In
crossing the joint between the first and second phalanges the
tendon forms a fibrous pad which protects the dorsum of the
joint and glides over it. It then continues to be inserted into
the base of the terminal phalanx.
Relations.—Outer surface with the tibialis anterior (n) and
the superficial fascia. Inner surface with a part of the tibialis
anterior, with the bones, the interosseous membrane, and the
peroneus brevis (s).
Action.—Extensor of the phalanges.
M. tibialis anterior (Fig. 90, n).—This is the superficial
muscle covering the lateral side of the tibia.
Origin by fleshy fibres from the proximal one-sixth of the[212]
lateral surface of the shaft of the tibia, from the proximal third
of the medial border of the shaft and head of the fibula, and
from the intervening interosseous ligament. The triangular
body of the muscle covers the lateral surface of the tibia and
the outer surface of the extensor longus digitorum (p). Near
the malleolus it ends in a strong tendon which passes beneath
the transverse ligament along with the tendon of the extensor
longus (p). The tendon crosses the dorsal surface of the foot
obliquely toward its medial side.
Insertion into the outer surface of the first metatarsal.
Relations.—Outer surface with the superficial fascia and the
tendon of the biceps femoris. Inner surface with the extensor
longus (p), the tibia, and the interosseous membrane.
Action.—Flexor of the foot.
4. Muscles of the Foot.
A. Muscles on the Dorsum of the Foot.
—M. extensor
brevis digitorum (Fig. 90, u).—A broad thin muscle
covering the tarsus and part of the metatarsus on the lateral
side of the dorsum of the foot. It may be divided into three
slips.
Origin from the distal border of nearly the whole calcaneal
annular ligament (5), and from the proximal end of the dorsal
surface of the three lateral metatarsals. At the middle of the
metatarsals the muscle ends in three flat tendons which pass
into the three interspaces between the four tendons of the extensor
longus. Each tendon is divided into two branches.
Insertion.—The lateral branch of each tendon is inserted
into the cartilaginous plate which lies in the metatarsophalangeal
articulation of the digit on the outer side. The medial
branch joins the lateral side of the extensor longus tendon on
the dorsum of the first phalanx.
Action.—Extensor of the digits.
B. Muscles of the Sole of the Foot.
—M. flexor
brevis digitorum (pedis perforatus).—This is the direct continuation
of the tendon of the plantaris. It lies immediately
beneath the superficial plantar fascia. The flat belly of the
muscle has its medial border attached by an oblique tendinous[213]
band to the medial surface of the scaphoid and the medial
cuneiform. It divides into four slips which diverge to the four
toes, each ending in a flat tendon. The slips decrease in size
from the lateral to the medial side. The three lateral ones
overlap one another proximally.
Insertion.—Each tendon expands at the distal end of the
metatarsal and is wrapped about the tendon of the deep flexor
which perforates it. The two halves unite beneath the perforating
tendon and pass together with the perforating tendon
through fibrous rings, the annular ligaments. One of these is
attached to the sesamoids at the base of the phalanx. The
other is attached to the head of the first phalanx. The tendons
are finally inserted into the bases of the second phalanges.
The two annular ligaments are connected by fascia so as
to form a continuous canal for the tendons. This canal is lined
by a synovial membrane. Covering this canal is a more superficial
layer of fascia. Before passing into the proximal annular
ligament the two middle tendons unite each with the tendon
of the corresponding lumbrical muscle. Each also gives off
a branch, the lateral one on its lateral side and the medial one
on its medial side. These branches, like those of the two
middle ones, are inserted into the common phalangeal fascia.
Action.—Flexor of the second phalanges of the digits.
M. quadratus plantæ (called also the plantar head of the
flexor longus digitorum) is a thin flat muscle which takes
origin from the dorsal part of the lateral surface of the calcaneus
and the cuboid. Its fibres converge to a flat tendon
which passes transversely across the flexor longus digitorum
and beneath the flexor brevis digitorum and calcaneometatarsal.
Insertion.—Into the medial part of the outer surface of the
tendon of the flexor longus digitorum.
Action.—It holds the flexor longus tendon in place.
Mm. lumbricales.—These are six.
Origin.—The three larger ones have origin from the outer
surface of the expanded portion of the flexor longus digitorum
on its distal half. Each ends in a slender tendon.
Insertions.—The tendons unite with the divisions of the[214]
tendon of the flexor brevis which pass to the three lateral
digits, at their entrance to the first annular ligament. The
one to the fifth digit is sometimes absent.
The other three have origin from the tendon of the flexor
longus digitorum at the point where it divides. Each occupies
one of the three intervals between the four divisions, and its
fibres originate from the proximal ends of the two tendons
which bound its interval.
Insertion by a very slender tendon into the medial side of
the first phalanges of the third, fourth, and fifth digits near
their bases.
Action.—Move the third, fourth, and fifth digits toward the
medial side.
Mm. interossei.—The interosseus of the second digit forms
five portions, which cover the ventral surface of the digit.
Origin.—(1) By two tendons from the ventral process of
the lateral cuneiform. The muscle divides into four parts, three
of which pass to the medial side of the digit and act as
abductors. They are called the long, short, and middle
abductors (abductor longus, brevis, and intermedius digiti
secundi).
Insertions.—The short head into the medial sesamoid of
the metatarsophalangeal joint, the middle into the medial side
of the base of the first phalanx, and the long into the extensor
communis tendon near the distal end of the first phalanx.
The names long, short, and middle refer to the lengths of the
tendons as determined by their points of insertion.
The fourth portion which comes from the lateral cuneiform
has its insertion into the lateral side of the extensor tendon
near the distal end of the first phalanx. It is therefore an
adductor longus digiti secundi. In some cases there is an
adductor brevis from this same head.
(2) The fifth part of the interosseus of the second digit has
origin from the middle of the ligament covering the peroneal
canal, along with the middle adductor of the fifth digit. The
two muscles diverge. Insertion into the outer side of the base
of the first phalanx of the second digit. It is an adductor
medius digiti secundi.
[215]
The interossei of the third and fourth digits are alike.
Origin from the ventral surface of the base of the metatarsal.
The muscles cover the ventral surface of the shaft. At
the head each divides and passes onto the sides of the
metatarsal.
Insertion.—Each half of the muscle shows a tendency to
divide into two parts. One of these, the more superficial and
ventral, ends in a tendon which joins the extensor tendon near
the distal end of the first phalanx. The other part is inserted
by short tendon- and muscle-fibres into the side of the base of
the first phalanx; one of the parts is therefore a middle, and the
other a long, adductor or abductor, the adductores and abductores
digiti tertii and quarti.
The fifth digit has five short muscles.
M. abductor medius digiti quinti.
Origin from the ventral surface of the calcaneus and from
the fifth metatarsal.
Insertion by a slender tendon into the lateral side of the
base of the first phalanx of the fifth digit.
M. adductor medius digiti quinti.
Origin with the adductor medius digiti secundi.
Insertion on the inner side of the base of the phalanx of the
fifth digit.
M. opponens digiti quinti.
Origin by a flat tendon from the middle of the ligament
which covers the peroneal canal. It passes obliquely outward
parallel to the last.
Insertion on the inner side of the shaft of the fifth metatarsal.
The two remaining muscles of the fifth digit take origin
from the outer part of the ligament covering the peroneal canal.
They cover the ventral surface of the shaft.
Insertion.—The lateral one is inserted into the lateral sesamoid
and is therefore an abductor brevis. The medial one is
inserted into the extensor tendon and is therefore an adductor
longus.
C. Muscles of the Tarsus.
—M. calcaneometatarsalis
(part of M. adductor minimi digiti?).—A weak muscle made up[216]
largely of tendon-fibres, some of which pass directly from origin
to insertion. It may be regarded as a ligament.
Origin, the lateral and ventral surface of the calcaneus near
the proximal end. It passes distad and slightly laterad outside
of the quadratus plantæ.
Insertion, the lateral side of the base of the fifth metatarsal,
and the adjacent surfaces of the cuboid and calcaneus.
Action.—Probably causes slight motion of the cuboid on
the head of the calcaneus.
M. scaphocuneiformis.—A small muscle lying on the sole
of the foot in the depression between the lateral cuneiform,
medial cuneiform, and the lateral tubercle of the scaphoid. It
is hidden by the overlying ligaments.
Origin, the lateral tubercle of the scaphoid bone. It passes
distad and laterad.
Insertion, the lateral surface of the medial cuneiform.
Action.—Rotates the medial cuneiform on the scaphoid and
would thus act as an opponens of the great toe if the great toe
were present.
[217]
THE VISCERA.
I. THE BODY CAVITY.
The greater part of the viscera are situated in the body
cavity or cœlom. This is divided by the diaphragm into two
parts, the thoracic cavity and the abdominal cavity. Each
is lined by a serous membrane, in which the part covering the
outer wall of the cavity is distinguished as the parietal layer
from the part covering the viscera, which is known as the
visceral layer.
The thoracic cavity is bounded by the thoracic vertebræ,
the ribs, the sternum, and the diaphragm. The cranial opening
of the cavity is filled by the trachea and œsophagus as they
enter from the neck region. The thoracic cavity is lined by
two thin layers of tissue, the outer one of which is the fascia
endothoracica, while the inner is the pleura. The fascia
endothoracica is a sheet of connective tissue which lines the
entire inner surface of the thoracic cavity, descending from the
dorsal median line to the heart, and passing into the fibrous
layer of the pericardium. The pleura is a thin membrane
covering the fascia endothoracica and corresponding to the
peritoneum of the abdominal cavity. It forms two sacs, the
pleuræ, lining respectively the right and left halves of the
thoracic cavity. Each of these two sacs is closed, the viscera
being suspended within them by folds of the membrane, so that
the cavity is everywhere separated from the viscera by a sheet
of the pleura. That portion of the pleura which lines the
thoracic wall is known as the parietal layer; it may be divided
into that covering the ribs (costal pleura), and that covering
the diaphragm. That portion which covers the viscera is the
visceral layer, or, since it covers chiefly the lungs, it may be[218]
distinguished as the pulmonary pleura. The medial walls of
the two pleural sacs come in contact in the median plane,
forming a median vertical partition passing lengthwise of the
thoracic cavity. This partition is known as the mediastinal
septum. The space between the two layers which make up
the mediastinal septum is known as the mediastinum, or
mediastinal cavity; it contains numerous organs of the thorax.
Three parts are usually distinguished in this cavity: a ventral
mediastinal cavity, containing chiefly blood-vessels and the
thymus gland; a middle mediastinal cavity, enclosing the heart
and the anterior and posterior venæ cavæ; and a dorsal mediastinal
cavity, containing the trachea, the œsophagus, and the
aorta.
The abdominal cavity lies caudad of the diaphragm; in it
are sometimes distinguished the abdominal cavity proper,
extending as far caudad as the cranial edge of the pubis, and
the pelvic cavity, lying caudad of this, in the region surrounded
by the innominate bones and the sacrum. The two cavities
are not distinctly marked off, so that it is convenient to consider
the abdominal cavity as undivided. Both parts are lined
by the peritoneum.
The peritoneum is a thin transparent sheet of connective
tissue supporting on its surface a layer of flattened epithelial
cells, the peritoneal epithelium. It forms a sac which lines the
entire abdominal cavity. This sac is closed in the male; in
the female, however, it communicates with the exterior through
the uterine (or Fallopian) tubes and uteri. All the organs of
the abdominal cavity are outside the sac. In the course of their
development these organs have encroached on the peritoneal
sac. Each has grown against the outer wall of the sac to a
greater or less extent and has forced a part of this wall ahead
of it into the cavity. In some cases the encroachment has
gone so far that the organ in question lies apparently within
the peritoneal cavity, suspended from the wall of the sac by a
fold of that wall. The wall may thus be divided into three
portions. One of these, the parietal layer, lines the wall of
the body cavity. The second (the mesentery in case of the
alimentary canal, or a ligament in the case of another organ)[219]
suspends the organ from the body wall. Between the layers
of each mesentery or ligament blood-vessels may pass to the
organs. The third portion or visceral layer covers the organ
in question, forming its serous covering.
The reflections of the peritoneum to form the mesenteries
and ligaments may be thus described:
Caudally the peritoneum covers the whole surface of the
bladder and is reflected from its ventral wall to the linea alba
as the suspensory ligament of the bladder. Farther craniad
the peritoneum suspends the rectum and colon from the middorsal
line, forming the mesorectum and mesocolon. The
mesocolon continues craniad to the level of the caudal end of
the right kidney and is broadest at its cranial end. At this
end the mesocolon passes into the mesentery proper, which
suspends the small intestine and is very broad and much folded.
Its dorsal border is attached to the median line opposite the
caudal end of the kidneys and is very short compared to its
very long ventral or intestinal border. Toward the cranial end
the mesentery of the jejunum passes gradually into the very
much shorter duodenal mesentery. This is drawn out at the
caudal end of the duodenum into a fold, the duodenorenal ligament
which attaches the duodenum to the kidney.
The mesogastrium or peritoneal fold for the stomach passes
from that part of the median dorsal line lying between the
kidneys and the diaphragm, to the greater curvature of the
stomach. It does not pass directly to the stomach, but passes
first ventrad of the small intestine as far as the pelvis. Thence
it turns craniad to reach the greater curvature of the stomach.
The fold thus formed is called the great omentum. It forms
the dorsal and ventral walls of a sac, the omental sac, the
cavity of which is called the lesser peritoneal cavity. The
descending limb of the fold forms the dorsal wall of the sac,
and its ascending limb forms the ventral wall. Each of these
walls is double like a mesentery, so that the great omentum
consists of four sheets of peritoneum. Between the two sheets
forming the descending limb lies the left half of the pancreas,
which passes thence to the right into the duodenal mesentery.
A transverse fold passes from the descending limb of the[220]
omentum along the cranial border of the pancreas to the
duodenal mesentery. Farther to the right the descending limb
of the omentum, which is here shorter, encloses the spleen and
holds it in position parallel to the greater curvature of the
stomach and about one centimeter from it. This part of the
great omentum is sometimes called the gastrosplenic or
gastrolienal omentum.
The great omental sac communicates with the peritoneal
sac by an opening, the foramen epiploicum or foramen of
Winslow. This opening lies caudad and dextrad of the
caudate lobe of the liver. This lobe itself lies within the sac.
Along the ventral border of the foramen epiploicum pass the
common bile-duct from the liver and the portal vein to the
liver.
The size of the great omental sac is increased by the
lesser omentum. This is a double sheet of peritoneum which
stretches horizontally from the liver to the duodenum and the
lesser curvature of the stomach. It covers ventrad the caudate
lobe of the liver. Its right border is at the foramen epiploicum.
The part of it which stretches to the duodenum is called the
duodenohepatic ligament and contains the bile-ducts and the
portal vein. The part that stretches to the stomach is called
the gastrohepatic ligament. The peritoneum covers the caudal
and cranial surfaces of the liver and is reflected to the adjacent
parts to form the ligaments of the liver.
The suspensory ligament of the liver passes from the
caudal surface of the diaphragm and the median ventral line
for about one or two centimeters caudad of the diaphragm, to
the liver, and extends between its two halves. The ligamentum
teres or round ligament is the thickened free caudal border of
the suspensory ligament. It is the remains of the fœtal umbilical
vein. From the dorsal border of the liver the peritoneum
which covers its cranial surface turns ventrad onto the caudal
surface of the diaphragm, while that which covers its caudal
surface turns dorsad onto the caudal surface of the diaphragm
to reach the dorsal body wall. Between these two sheets a
small linear part of the surface of the liver is closely applied to
the diaphragm without intervening peritoneum. The two[221]
sheets which bound this area constitute the coronary ligament
of the liver. This ligament is broader where it passes from the
left lateral lobe to the diaphragm and is called the triangular
ligament. (There is perhaps a corresponding right triangular
ligament, from the cranial division of the right lateral lobe.)
The caudal division of the right lateral lobe is held to the
kidney of that side by the hepatorenal ligament.
II. THE ALIMENTARY CANAL. APPARATUS DIGESTORIUS.
The alimentary canal may be divided into mouth, pharynx,
œsophagus, stomach, small intestine, and large intestine.
With these are associated certain accessory structures,—the
salivary glands, the liver, and the pancreas. The spleen,
though not belonging to the digestive system, is usually
described in connection with it. The respiratory organs are
almost throughout in close relation with the organs of the
digestive system.
1. The Mouth. Cavum oris.
—The mouth cavity extends
from the lips to the pharynx. It is narrower toward the lips,
broadens caudad as far back as the last teeth, then becomes
narrowed to form the isthmus faucium, by which it communicates
with the pharynx. The mouth cavity is divisible into
the vestibule of the mouth (vestibulum oris), which comprises
that part outside the jaws proper, bounded externally by the
lips and cheeks, and the mouth cavity proper (cavum oris
proprium), which lies within the teeth. That portion of the
vestibule which is bounded by the cheeks is sometimes farther
distinguished as the buccal cavity. The entire mouth cavity
(except the teeth) is lined by the mucous membrane or mucosa.
The lips (labia oris) are thick folds of skin bounding the
entrance to the mouth cavity. The outer surface is covered
with hair; the inner surface is covered with the mucous membrane.
The upper lip is marked in the median line by a deep
external groove which extends upward to the septum of the
nose. Along the inner surface of this groove the lip is closely
united to the jaw by a thick fold, the frenulum of the upper
lip. For some distance on each side of the frenulum the inner[222]
surface of the lip bears numerous large papillæ. The lower
lip is also united to the jaw by a frenulum in the median line;
it is again united to the jaw just caudad of the canine tooth, in
the space between the latter and the first premolar. Caudad
the two lips pass into each other (forming the commissura
labiorum), and unite with the cheek. The muscles of the lips
have been described (page 105).
The cheeks (buccæ) in the cat are comparatively thin and
small, extending from the lips caudad to the ramus of the
mandible. The outer surface is covered with hair; the inner
surface is smooth and somewhat folded. The buccal cavity is
rather small. On the inner surface of the cheek open the duct
of the parotid (Steno’s duct), the ducts of the molar gland, and
that of the infraorbital gland.
The roof of the mouth cavity is formed by the hard and
soft palates. The hard palate (palatum durum) forms the
cranial part of the roof; it is supported by the palatal plates of
the maxillary and palatine bones. The mucosa of the hard
palate is elevated to form seven or eight curved transverse
ridges, which are concave caudad. Between the ridges are
rows of papillæ. In front of the most cranial ridge is a papilla
in the middle line, and at each side of the papilla is the opening
of a small duct (the incisive duct or Stenson’s duct),
which leads dorsad through the incisive foramen to the
vomeronasal organ (or organ of Jacobson), which lies on the
floor of the nasal cavity. Caudad of the hard palate the roof
of the mouth is formed by the soft palate or velum palatinum,
described below.
The floor of the mouth cavity is formed chiefly by the
tongue, which extends as far caudad as the isthmus faucium.
Ventrad of the free edge of the tongue the mucosa forms a
prominent median vertical fold which unites the tongue with
the floor of the mouth beneath it; this fold is the frenulum
linguæ. On each side of the median line at the cranial border
of the floor of the mouth is a prominent papilla, at the apex of
which open the ducts of the submaxillary and sublingual
glands, the former on the lateral side of the apex, the latter
on the medial side.
[223]
The sides of the mouth cavity are formed by the teeth and
the gums, covering the alveolar borders of the mandible,
maxillaries, and premaxillaries.
The mouth cavity presents further for examination the
glands, the teeth, the tongue, and the soft palate.
The Glands of the Mouth (Glandulæ oris).
—There
are five pairs of salivary glands which open into the
mouth cavity.
1. The parotid gland (glandula parotis) (Fig. 65,
1; Fig.
131, 10) is flattened, rather finely lobulated, and lies ventrad of
the external auditory meatus and beneath the dermal muscles.
Its cranial border follows the caudal border of the masseter
muscle and overlies it somewhat; its caudal border is about
three centimeters caudad of the border of the masseter. Its
borders are unevenly lobed. The parotid duct (ductus parotideus:
frequently called Stenon’s or Steno’s duct) is formed
by the union of several smaller ducts near the ventral end of
the cranial border of the gland. It passes craniad imbedded
in the fascia covering the masseter. At the cranial border of
the masseter it turns inward and lies close against the mucous
membrane of the mouth, so that from the inside of the mouth
it appears as a white ridge on the mucosa. It opens on the
inside of the cheek opposite the most prominent cusp of the last
premolar tooth. Along the course of the parotid duct in some
cases one or more small accessory parotid glands are found.
2. The submaxillary gland (glandula submaxillaris)
(Fig. 65, 2, page 109,
and Fig. 131, 11) is approximately kidney-shaped.
Its surface is nearly smooth, the lobulations not
being apparent externally. It lies ventrad of the parotid, at
the caudal edge of the masseter muscle, just caudad of the
angular process of the mandible. The posterior facial vein
(Fig. 131, b) crosses its outer surface, and its cranioventral
border is hidden by two lymphatic glands (Fig. 131, 12) lying
at the sides of the anterior facial vein. The submaxillary
duct (ductus submaxillaris, frequently called Wharton’s duct)
leaves the inner surface of the gland and passes beneath the
digastric and mylohyoid muscles and against the outer surface
of the styloglossus. From the point where the styloglossus[224]
passes into the tongue the duct continues craniad close against
the oral mucosa and parallel to the mandibula. It is accompanied
by the duct of the sublingual, which lies at first dorsad
of it and then mediad. It opens at the apex of the prominent
papilla which lies at the side of the middle line at the cranial
end of the floor of the mouth.
3. The sublingual gland is elongated and conical in form,
with its base against the submaxillary, of which it appears to
be a continuation. It stretches along the submaxillary duct
for about one and one-half centimeters, lying between the
masseter and digastric muscles. The sublingual duct leaves
its ventral side, passes close to the submaxillary duct, at first
dorsad and then mediad of it, and opens on the medial side of
the apex of the same papilla with the submaxillary duct.
4. The molar gland (glandula molaris) (Fig. 65, 9) lies
between the orbicularis oris and the mucosa of the lower lip.
It stretches from the cranial border of the masseter to a point
between the first premolar and the canine. It is flat, broad
caudad, and ends in a point craniad. It has several ducts
which pass straight through the cheek and open on the mucous
surface of the mouth.
5. The infraorbital (or orbital) gland lies in the lateral
part of the orbit on its ventral floor. It is ovoid and about one
and one-half centimeters long and one-third as thick. Its
ventral end rests against the mucosa of the mouth just caudad
of the molar tooth. Its duct leaves the ventral end and opens
into the mouth at a point about three millimeters caudad of the
molar tooth.
The Teeth. Dentes.
—The adult cat has thirty teeth,
fourteen in the lower jaw and sixteen in the upper jaw. There
are twelve incisors, four canines, ten premolars, and four
molars. The tooth formula for the cat is then
i3 —
33 — 3,
c1 —
11 — 1,
pm3 —
32 — 2,
m1 —
11 — 1.
The teeth are implanted in the alveolar borders of the
premaxillaries, maxillaries, and mandible. In each tooth can
be distinguished the root, imbedded in the socket of the bone,[225]
the crown, which projects above the gums, and a narrow neck
connecting the two. The root is composed of one or more
separate fangs; the crown bears one or more points or cusps.
At the cranial end of each jaw are six incisor teeth (Figs.
93 and 94, a). These are imbedded in the alveolar borders of
the premaxillaries and the mandible.
The incisor teeth are
small, with a crown bearing a
sharp edge which is notched so
as to form three minute cusps.
The root of each has a single
fang. The lateral incisors are
the largest in each jaw, and
those of the upper jaw are larger
than those of the lower.
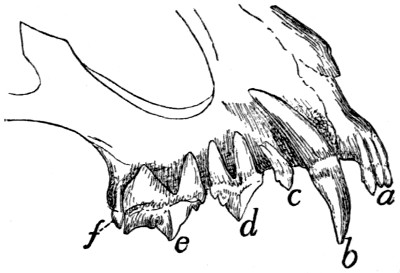
Fig. 93.—Upper Jaw, with Roots
of the Teeth Laid Bare.
a, incisors; b, canine; c, first premolar;
d, second premolar; e, third premolar;
f, molar.
Caudad of the incisors, and
in the upper jaw separated from
them by a slight interval, are the canines (b), two in each jaw.
These are long, strong, pointed teeth, deeply imbedded in the
mandible and maxillaries, their large sockets causing a rounded
swelling on the external surface of the bones. Each has a
single fang and a single cusp. When the mouth is closed the
upper canines lie laterocaudad of the lower ones.
Caudad of the canines there is in each jaw a considerable
interval free from teeth: this is called the diastema. Caudad
of the diastema are the premolar teeth, three pairs (c, d, e) in
the upper jaw and two pairs (c, d) in the lower jaw. These
teeth are compressed sideways, and those of the lower jaw fit
inside of those of the upper jaw. In the upper jaw (Fig. 93)
the first premolar (c) is small and usually has but a single cusp
and a single fang, though occasionally there is a small supplementary
cusp and fang. The second premolar is larger (d);
it has a large central cusp, with a single smaller cranial cusp
and two small caudal cusps, making four in all. This tooth
has two fangs. The third premolar (e) is the largest tooth in
the jaws; it has three large cusps in longitudinal series and a
small cusp lying on the medial side of the first one in the row.
Its root has three fangs. The molar tooth (f) of the upper[226]
jaw is small and lies caudomediad of the last premolar. It has
two small cusps and two fangs.
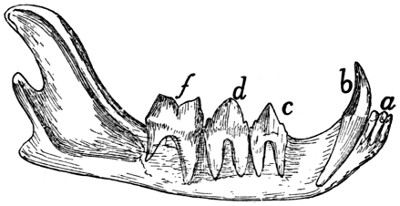
Fig. 94.—Mandible,
with Roots of the Teeth Laid Bare.
a, incisors; b, canine; c, first premolar; d, second
premolar; f, molar.
In the lower jaw (Fig. 94) the two premolars (c and d) are
similar, the caudal one being a little larger. Each has four
cusps; a single large one, a small one craniad of this, and two
small ones caudad of it. Each has two fangs. The single
molar (f) is the largest tooth of the lower jaw; it has two large
cusps and two fangs.
The Deciduous Teeth.—At birth the cat has no teeth.
There appears later a set of twenty-six teeth: twelve incisors,
four canines, and ten molars (six in the upper jaw and four in
the lower). These teeth are later replaced by the permanent
ones above described. The deciduous teeth of the cat are fully
described by Jayne (“Mammalian Anatomy,” vol. 1. p. 319),
where also an account is given of the order of appearance of
the teeth.
The Tongue. Lingua
(Fig. 95).—The tongue is a
muscular organ covered with mucous membrane; in life it is
very mobile. It is an elongated organ, flat above, broadest in
the middle, and very slightly narrowed at each end. It
extends from the incisor teeth to the isthmus faucium and
nearly fills the mouth cavity. The caudal third of the tongue
forms the floor of the mouth cavity, so that the tongue has here
no ventral surface, but is directly continuous with underlying
organs. It is in this region that the extrinsic muscles of the
tongue (except the genioglossus) enter it. The cranial two-thirds
of the tongue is partly or entirely free from the floor of
the mouth, the cranial one-third being completely free and
movable. In about the middle third the ventral surface of the[227]
tongue is held to the floor of the mouth by the fold known as
the frenulum linguæ. The frenulum contains parts of the two
genioglossus muscles, which enter the tongue through it. The
ventral surface and lateral borders of the tongue are smooth,
soft, and free from papillæ. The
dorsal surface is raised into papillæ
of various kinds, and has a slight
median longitudinal furrow. The
caudal part of the dorsal surface is
softer, redder, and marked with
papillæ of a different kind from
those of the rest of the tongue.
From the caudal end a small
median vertical fold, the frenulum
(or plica) glossoepiglottica passes
from the dorsal surface of the
tongue to the cranial surface of the
epiglottis.

Fig. 95.—Tongue, Epiglottis,
and Opening of Larynx.
a, filiform papillæ; b, fungiform
papillæ; b′, very large papillæ at the
sides of the tongue; c, circumvallate
papillæ; d, tonsils; e, epiglottis; f,
plica aryepiglottica; g, arytenoid cartilages
(covered with mucosa); h,
glottis; i, false vocal cords; j, true
vocal cords.
The papillæ of the tongue are
of three kinds. 1. The very numerous
filiform papillæ (a) (papillæ
filiformes); many of them
are horny and tooth-like, with
points turned caudad. These are
most numerous at the middle of
the free end of the tongue. 2. The
fungiform papillæ (b) (papillæ
fungiformes) are found scattered
over the surface of the middle of
the tongue caudad of the large
filiform papillæ. They are enlarged
at their free ends. There
is a prominent row of very large
ones (b′) opposite the circumvallate
papillæ at the borders of the
tongue. 3. The circumvallate papillæ (c) (papillæ vallatæ)
are blunt and each is surrounded by a trench which is bounded
in turn by a raised wall. They are in two rows of two or three[228]
each, which converge near the base of the tongue so as to form
a V with the apex directed caudad.
Muscles of the Tongue
(Fig. 96).—M. genioglossus
(f) passes from the symphysis of the lower jaw into the tongue
and lies beneath (dorsad of) the geniohyoid (g).
Origin from the medial surface of the mandible near the
symphysis and dorsad of the origin of the geniohyoid.
Insertion.—The fibres pass dorsad, diverging in a fan-like
manner and forming a flat vertical plate closely applied to the
muscle of the opposite side. This plate extends along the
caudal three-fourths of the tongue, i.e. as far as it is attached.
The cranial fibres arch craniad to the tip of the tongue, the
caudal fibres arch caudad to the root of the tongue. Dorsad
the muscle is confounded with the muscle of the opposite side.
Action.—Draws the root of the tongue forward and the tip
backward.
M. hyoglossus (h).—From the body of the hyoid bone to
the tongue.
Origin.—(1) From the ventral surface of the body of the
hyoid laterad of the geniohyoid (g), and (2) by a second head
from the ceratohyal.
Insertion.—Both heads penetrate into the tongue between
the styloglossus (e) and the genioglossus (f). The fibres
intermingle with those of the styloglossus (e) and thus help to
form the lateral parts of the tongue. They finally end in the
integument on the dorsum of the tongue at the sides.
Action.—Retracts the tongue and depresses it.
M. styloglossus (e).—From the stylohyal bone to the
tongue.
Origin from the mastoid process of the temporal bone, from
the stylomandibular ligament (2) (which connects the border
of the external auditory meatus with the angular process of the
mandible) and from the proximal cartilaginous portion of the
cranial cornu of the hyoid bone. The fibres pass mediad,
diverging between those of the digastric and hyoglossus (h)
into the lateral part of the tongue.
Insertion.—The fibres pass toward the tip of the tongue,
where the mass finally ends in a point, the superficial ones[229]
gaining insertion into the integument at the sides of the
tongue.
Relations.—Outer surface
with the digastric (Fig. 65, b)
and mylohyoid (Fig. 65, c). Inner surface with the pterygoideus
internus (Fig. 96, d), the cranial cornu of the hyoid,
and the tympanic bulla.
Action.—Retracts the tongue and raises it.
Fig. 96.—Muscles
of Tongue, Hyoid Bone, and Pharynx.
a, M. tragicus lateralis; b, M. jugulohyoideus; c, M. pterygoideus externus; d,
partially cut surface of M. pterygoideus internus; e, M. styloglossus; f, M. genioglossus;
g, M. geniohyoideus; h, M. hyoglossus; i, M. glossopharyngeus; j, M. constrictor
pharyngis medius; k, M. constrictor pharyngis inferior; l, M. stylopharyngeus;
m, M. sternohyoideus (cut); n, M. cricothyreoideus; o, M. sternothyreoideus (cut);
p, M. thyreohyoideus. 1, mandible; 1′, angular process of mandible; 2, stylomandibular
ligament; 3, bulla tympani; 4, trachea; 5, œsophagus; 6, thyroid gland;
7, isthmus of the thyroid gland.
The intrinsic muscles of the tongue (those entirely within
it) are attached to its integument at both their ends. There
are three sets of fibres: a longitudinal, a transverse, and a vertical
one. These are seen most readily in cross-sections.
The Soft Palate. Velum Palatinum
(Fig. 66, [230]
page 112).—The soft palate is the free curtain-like structure which
forms the caudal part of the roof of the mouth. It is attached
to the caudal border of the palatal plates and the ventral border
of the perpendicular plates, of the palatine, and to the pterygoid
processes and hamuli of the sphenoid, and extends some distance
caudad of the hamuli. It thus forms a rather long and
narrow curtain separating the caudal part of the nasal cavity
from the mouth. Caudad it ends in a free arched border (Fig.
66, 4) which is at about the level of the epiglottis, and may lie
against the cranial or the caudal surface of the latter. The
narrowed passage bounded by the margin of the velum palatinum
dorsad and the tongue ventrad is the isthmus faucium.
From the sides of the velum a short distance from the caudal
border a fold of mucosa passes ventrad to the side of the
tongue; a short distance caudad of this a similar fold passes to
the floor of the pharynx. These folds form the cranial and
caudal pillars of the fauces. Between these folds is a shallow
pocket, from the bottom of which there arises a prominent projection
or swelling which is one of the two tonsils (Fig. 95, d).
Each tonsil is a reddish, lobulated gland, lymphoid in the
adult, nearly a centimeter in length, and about one-third as
long as broad, with its long axis craniocaudad.
The velum palatinum consists of two layers of mucous
membrane, oral and nasal, with intervening muscular and connective
tissue. The muscles of the soft palate in the cat are
as follows:
M. tensor veli palatini (Fig. 66, d, d′, page 112).
Origin from the ventral surface of the body of the sphenoid
between the foramen ovale and the groove for the Eustachian
tube. The muscle ends in a flat tendon which passes over the
hamular process (3) of the pterygoid bone.
Insertion by spreading out in the soft palate into an
aponeurosis which joins the aponeurosis of the opposite muscle
and lies between the mucous membrane of the mouth and that
of the nose.
Action.—Stretches the palate.
M. levator veli palatini (Fig. 66, e, e′).—A flat triangular
muscle which lies within the tensor.
[231]
Origin from the surface of the body of the sphenoid mediad
of the groove for the Eustachian tube, from the styliform
process of the bulla tympani, and in part from the Eustachian
tube. The muscle passes caudad, and its fibres then diverge
into the velum palatinum.
Insertion into the velum palatinum, some of the fibres
meeting in the middle line.
Action indicated by the name.
A number of other muscles have been described in the soft
palate of the cat; they are, however, poorly developed and not
easily distinguished. For an account of these, see Stowell,
Proceedings of the Am. Soc. of Microscopists, 1889.
2. The Pharynx.
—At the caudal end of the mouth cavity
the passage for the food and that for the air cross; at the
cranial end the food-passage (mouth) is ventral, the respiratory
passage (nasal cavity) dorsal. Farther caudad the food-passage
(œsophagus) is dorsal, while the respiratory passage (larynx
and trachea) is ventral. In the region of crossing there is
therefore for a certain distance a common passageway for food
and air, and this is known as the pharynx. It extends from the
isthmus faucium, at the free caudal margin of the soft palate, to
the beginning of the œsophagus, at the dorsal or caudal margin
of the opening of the larynx. The dorsal wall of the pharynx
is separated from the base of the skull and the centra of the
cervical vertebræ only by intervening muscles (longus capitis,
levator scapulæ ventralis, and longus colli, Fig. 72, page
143). Its lateral and ventral walls are supported by the hyoid
bone and the cartilages of the larynx.
Craniad the pharynx continues, usually, without break into
the cavity lying dorsad of the soft palate. But at the time of
swallowing the free edge of the soft palate is pushed dorsad
against the dorsal wall of the pharynx, while the caudal part
of the pharynx is drawn craniad, so as to form a cavity continuous
with that of the mouth. In this way the cavity above
the soft palate is completely separated at the time of swallowing
from the rest of the pharynx. This separated portion is
known as the nasopharynx: it is strictly a portion of the
respiratory passage, as the food does not pass into it. The[232]
nasopharynx is continuous craniad by the choanæ with the
nasal cavity; it forms a horizontal tube between and ventrad
of the perpendicular plates of the palatine bones, and has the
same craniocaudal extent as the soft palate. Its dorsal wall
lies against the basis cranii and the longus capitis muscles; its
lateral walls against the pterygoid muscles and the perpendicular
plates of the palatine bones; its ventral wall is the soft
palate. At the middle of its length, at the junction of its
dorsal and lateral wall, are two longitudinal slits about three
millimeters long. These are the medial openings of the
Eustachian tubes, by which the nasopharynx communicates
with the tympanic cavity.
The pharynx proper, situated caudad of the nasopharynx,
is smaller than the latter. It is bounded craniad by the
epiglottis and the margin of the soft palate, and is continuous
between the two, by the isthmus faucium, with the mouth
cavity. Its floor is formed by the cranial end of the larynx.
At its caudal end it passes dorsally into the œsophagus, while
ventrally it communicates with the larynx. Its walls are
muscular.
Muscles of the Pharynx
(Fig. 96).—M. glossopharyngeus
(i).
Origin.—Some fibres on the ventral and lateral part of the
genioglossus (f) leave that muscle near its caudal end. They
form a thin band of diverging fibres which pass outside of the
cranial horn of the hyoid. A similar sheet of fibres leaves the
midventral part of the styloglossus (e). The two sheets unite
and the united muscle crosses the hyoid, turns dorsad, and has
its
Insertion into the median dorsal raphe of the pharynx.
Action.—Constrictor of the pharynx.
M. constrictor pharyngis inferior (k).—A thin sheet of
muscle covering the sides of the pharynx at its caudal end.
Origin from the lateral surfaces of the thyroid and the
cricoid cartilages. The fibres pass dorsad and craniad, the
cranial ones covering the fibres of the middle constrictor (j).
Insertion.—The median longitudinal raphe on the dorsum
of the pharynx. The caudal fibres are transverse and continuous[233]
with the circular fibres of the œsophagus. The cranial
fibres may pass as far as the base of the sphenoid.
Action.—Constrictor of the pharynx.
M. constrictor pharyngis medius (j).—A thin sheet which
covers the middle part of the lateral surface of the pharynx.
Origin.—The ventral two pieces of the cranial horn and
the whole of the caudal horn of the hyoid. The fibres diverge,
passing dorsad.
Insertion into the median dorsal raphe of the pharynx.
The cranial fibres are inserted into the base of the sphenoid
bone. The muscle covers part of the stylopharyngeus (l) and
the superior constrictor (Fig. 66, f, page 112) and is partly
covered by the glossopharyngeus (Fig. 96, i).
Action.—Constrictor of the pharynx.
M. stylopharyngeus (l).
Origin from the tip of the mastoid process of the temporal
bone and from the inner surface of the cartilaginous piece
between the tympanohyal and the stylohyal bones. The
parallel fibres form a flat band which passes ventrocaudad over
the outer surface of the constrictor superior.
Insertion.—The ventral fibres pass beneath the middle constrictor
(j) at its cranial border and, continuing toward the
middle line of the pharynx, gradually lose themselves among
the fibres of the superior constrictor. The dorsal fibres pass
onto the outer surface of the middle constrictor and are lost
among its fibres.
Action.—Constrictor of the pharynx.
M. constrictor pharyngis superior or pterygopharyngeus
(Fig. 66, f, page 112).—A flat, triangular sheet beneath the
constrictor medius.
Origin.—The tip of the hamular process of the pterygoid
bone. The muscle passes caudad, the fibres diverging, and
dips beneath the cranial border of the constrictor medius.
Insertion into the median dorsal raphe of the pharynx.
The dorsal fibres are inserted into the base of the sphenoid.
The ventral fibres pass lengthwise of the pharynx, closely
connected with those of the stylopharyngeus (Fig. 96, l), and
finally reach the level of the larynx.
[234]
Action.—Constrictor of the pharynx.
3. The Œsophagus.
—The œsophagus is a straight tube,
dorsoventrally flat when empty, which extends from the
pharynx to the stomach. It has a uniform diameter when
moderately dilated of about one centimeter. It lies dorsad of
the trachea and against the longus colli muscles (Fig. 72, g′)
covering the centra of the cervical vertebræ, until it reaches
the caudal end of the thyroid gland (Fig. 96, 6); then it passes
to the left and lies laterodorsad of the trachea until it reaches
the bifurcation of the trachea. It there returns to the median
line, passes gradually distad, separated from the vertebræ by
the aorta, and finally pierces the diaphragm about two centimeters
from the dorsal body wall, and enters the stomach. Its
attachment to the diaphragm is loose enough to permit of longitudinal
motion. In passing through the thoracic cavity it lies
in the posterior mediastinum ventrad of the aorta. Its wall
consists of a muscular coat, a submucosa, and a mucosa, and
its inner surface presents many longitudinal folds. It has no
serous covering, its side walls being merely in contact with the
halves of the mediastinal septum.
4. The Stomach. Ventriculus
(Fig. 97).—The stomach
is the widest part of the alimentary canal. It is a pear-shaped
sac, the long axis of which is curved nearly into a semicircle.
The broad end of the sac lies to the left and dorsad; here the
stomach communicates with the œsophagus (a). The narrowed
end extends to the right and lies more ventrad than the other
end; it passes here into the duodenum (g). That portion of
the stomach which communicates with the œsophagus is known
as the cardiac end (b); the opposite is the pyloric end. Owing
to the curved form of the stomach above mentioned it is possible
to distinguish a concave and a convex side. The concave
side is directed craniad and dextrad; it is called the lesser
curvature of the stomach (c). The longer convex border is
directed caudad and to the left; it is called the greater curvature
(d). The greater curvature extends to the left, next to
the œsophagus, into a prominent convexity known as the
fundus (e) of the stomach.
The stomach lies at the cranial end of the abdominal[235]
cavity, mostly to the left of the middle line. Its cardiac end
is in contact by its dorsal surface with the dorsal, nearly horizontal,
portion of the diaphragm. On its ventral side the
cardiac end does not touch the diaphragm, so that a small part
of the œsophagus passes here for a short distance into the
abdominal cavity, to join the stomach. The communication
of œsophagus and stomach is by a simple conical increase in
size of the former. The pyloric end of the stomach extends
to the right of the middle line, becoming constantly smaller;
at its junction with the duodenum there is a constriction which
marks the position of the pyloric valve (f). This valve is
formed by a ring-like thickening of the circular muscle-fibres
of the alimentary canal, forming a sphincter muscle at the
junction of the stomach and duodenum and causing a projection
of the mucosa into the lumen of the canal. The ventral surface
of the stomach lies against the liver except when the stomach
is much distended with food, when the ventral surface comes
to lie against the ventral abdominal wall.
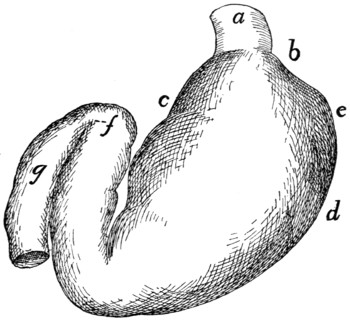
Fig. 97.—Stomach, Ventral View.
a, œsophagus; b, cardiac end of the stomach; c, lesser curvature; d, greater
curvature; e, fundus; f, pyloric valve; g, part of duodenum.
The stomach is supported by the great omentum and the
gastrohepatic ligament. It is connected with the duodenum
by the gastroduodenal ligament; with the spleen by the gastrolienal
ligament.
[236]
The inner surface of the stomach presents longitudinal folds
at its pyloric end and along the greater curvature as far as the
fundus. The prominence of these depends on the degree of
distension. Its walls are composed of an external peritoneal
layer, an internal mucous layer, and an intervening muscular
layer. This may be seen with the unaided eye in sections of
the wall.
5. The Small Intestine. Intestinum tenue.
—The small
intestine lies in numerous coils which take up the greater part
of the space in the abdominal cavity. It has a length about
three times that of the body of the cat. It is usually considered
as divided into three parts, duodenum, jejunum, and ileum;
these divisions are, however, not clearly marked off. The
small intestine is suspended by the mesentery already
described.
The duodenum is that part of the small intestine which follows
the stomach. At the pylorus (Fig. 97, f) the alimentary
canal makes a rather sharp turn so that the first part of the
duodenum forms an angle with the pyloric portion of the
stomach, and extends caudad and slightly toward the right,
soon becoming directed almost entirely caudad and lying along
the right side. About eight or ten centimeters caudad of the
pylorus it makes a U-shaped bend, extending thus craniosinistrad
for four or five centimeters. Here it passes without definite
limit into the jejunum, the duodenum being considered to end
at the next turn caudad. The entire duodenum is about fourteen
to sixteen centimeters in length. Between the two limbs
of the U-shaped bend formed by the duodenum, the duodenal
half of the pancreas is enclosed (Fig. 102, a).
The walls of the duodenum are composed of the serous
(peritoneal) investment, a muscular coat which is made up of
an outer thin, longitudinal layer of fibres and an inner thick,
circular layer, a submucous coat, and inside this a mucous coat.
The mucosa is thrown up into numerous delicate finger-like
villi which give to it a velvety appearance. On the dorsal
wall of the duodenum, about three centimeters distad of the
pylorus, the mucosa presents a slight papilla, at the apex of
which is seen the oval opening of the ampulla of Vater. This[237]
is an ovoid space in the wall of the duodenum. The space
is encroached upon by numerous folds of the walls. The
common bile-duct and the pancreatic duct open into it, the
former extending from the bottom of the ampulla nearly to its
mouth, and the latter extending from the bottom about half-way
to the mouth. Two centimeters caudoventrad of the opening
of the ampulla of Vater is the opening of the accessory pancreatic
duct. It can usually be demonstrated only by passing
a bristle into the duodenum through an opening in the duct.
The jejunum is the part of the small intestine following the
duodenum. It is not separated from the part of the small
intestine following it by any sharp line. In man it constitutes
two-fifths of the small intestine exclusive of the duodenum, and
is characterized by its emptiness after death and by the absence
from it of Peyer’s agminated glands (Peyer’s patches).
The ileum is the portion of the small intestine between the
jejunum and colon. It lies suspended by its mesentery in
numerous folds in the caudal part of the abdominal cavity,
separated from the ventral abdominal wall only by the great
omentum. It is of nearly uniform diameter, but its caudal portion
is thinner-walled than its cranial portion. Its walls have
a microscopic structure like that of the duodenum and jejunum.
On its inner surface and on the inner surface of the jejunum are
seen close-set villi, but these become rather sparser toward the
caudal end of the ileum and disappear about one centimeter
from the opening into the colon. Among the villi of the
caudal end of the ileum are numerous rounded elongations free
from villi. These are the solitary follicles or solitary glands
(lymphatic) of the intestine. These glands when aggregated
together form the agminated glands or patches of Peyer. The
ileum passes at the caudal end into the colon, the opening
being guarded by the ileocolic valve (Fig. 99). This is formed
by a marked projection of the mucosa (f) and transverse
muscle layer (e) of the ileum into the colon. Its surface is
free from villi.
6. The Large Intestine. Intestinum crassum.
—The
large intestine is divided into colon and rectum. The colon
or first part of the large intestine lies against the dorsal body[238]
wall and is separated from the ventral body wall by the folds
of the ileum. It has a diameter about three times that of the
ileum. The opening of the ileum into it is on its side between
one and two centimeters from its cranial end (Fig. 98). The
blind pouch thus formed by the cranial end of the colon is the
cæcum (Fig. 98, c; Fig. 99, a). The cæcum ends in a slight
conical projection which may be considered as the rudiment of
a vermiform appendix. The colon lies at first on the right side
and passes at first craniad; then transversely to the left, then
caudad, lying nearly in the middle line and next to the dorsal
abdominal wall. The colon may thus be distinguished according
to its direction into ascending, transverse, and descending
colon. At its caudal end the colon passes without sharp limit
into the rectum.
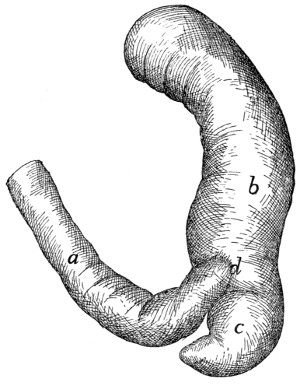
Fig. 98.—Junction of Small and
Large Intestine.
Fig. 99.—Section of the Ileocolic
Valve.
Fig. 98.—a, ileum; b, ascending colon;
c, cæcum; d, position of ileocolic valve.
Fig. 99.—a, cæcum; b, colon;
c, ileum; d, longitudinal muscle layer; e, transverse
muscle layer; f, mucosa; g, ileocolic valve (opened, as when material is passing
into the colon).
At the bottom of the cæcum on its inner surface is seen
a collection of solitary glands forming one of the agminated
glands of Peyer, or Peyer’s patches. The mucous membrane
is without villi. It presents a few considerable elevations,
probably solitary glands.
[239]
The rectum is the terminal portion of the large intestine
lying in the median line close to the dorsal body wall, from
which it is suspended by the short mesorectum. Its structure
is like that of the colon. It opens externally at the anus.
The entire large intestine has a length about one-half that of
the animal. At each side of the anus are two large secreting
sacs, the anal sacs or glands, each about a centimeter in
diameter. These open into the anus one or two millimeters
from its caudal boundary.
Muscles of the Rectum and Anus.—Owing to the close
interrelation of the muscles of the rectum and anus with those
of the urogenital organs, all these muscles will be described
together at the end of the description of the urogenital system.
7. The Liver, Pancreas, and Spleen.
—The Liver. Hepar.—The
liver (Figs. 100 and 101) is a large red-brown organ
occupying the cranial part of the abdominal cavity. It is
closely applied to the caudal surface of the diaphragm and
extends thence ventrad of the stomach so as to conceal all but
its pyloric end. Owing to the position of the stomach the
larger mass of the liver is on the right side and it extends
somewhat further caudad on this side.
The liver is divided by the dorsoventral suspensory ligament
into the right and left lobes, and each half is again divided
into lobes. On the left is a small left median (b) and a larger
left lateral lobe (a). The left lateral (a) extends caudad with
a thin edge which covers the greater part of the ventral surface
of the stomach. On the right there is a large right median
(or cystic) lobe (c, c′). Its cranial surface is dome-shaped
and fitted against the right two-thirds of the caudal surface of
the diaphragm. Its ventral edge is thin, its dorsal edge thick,
and its caudal surface marked by a deep dorsoventral cleft in
which lies the gall-bladder (Fig. 101, f). Dorsad and caudad
of the cystic lobe is the right lateral lobe (d, d′), which is
deeply cleft. Its elongated caudal division (d′) extends in a
point to the caudal end of the right kidney and is adapted to
the medial half of its ventral surface. Its smaller and more
compact cranial division (d) ends ventrally in a thin edge. It
lies between the caudal division (d′) and the cystic lobe (c),
and[240]
[241]
its dorsal surface is adapted to the suprarenal body. The
caudate or Spigelian lobe (Fig. 101, e) is an elongated, triangular,
pyramidal lobe. It lies in the omental sac and partly
closes the foramen epiploicum (foramen of Winslow). At its
base it is connected with the caudal division of the right lateral
lobe (d′).
Fig. 100.—Liver, Cranial Surface.
a, left lateral lobe; b, left median lobe; c,
right median lobe; d, d′, right lateral
lobe; e, gall-bladder; f, opening of posterior vena cava, with the smaller openings of
the hepatic veins.
Fig. 101.—Liver
turned Craniad, showing Dorsocaudal Surface.
a, left lateral lobe; b, left median lobe; c,
c′, right median (or cystic) lobe; d, d′,
cranial and caudal divisions of the right lateral lobe; e, caudate lobe; f, gall-bladder;
g, cystic duct; h, hepatic ducts; i, common bile-duct; j, portal vein; k, part of
duodenum.
The Gall-bladder (Fig. 101, f) is pear-shaped and lies in
a cleft on the caudal (or dorsal) surface of the right median
lobe (c, c′) of the liver. Its larger end is directed caudad (or
ventrad) and is free. By one surface it is in contact with the
liver and not covered by peritoneum, while the other surface
is covered by peritoneum. The peritoneum in passing from
the larger free end to the liver forms one or two ligament-like
folds. By its smaller end the gall-bladder is continuous with
the cystic duct (g). This duct is about three centimeters long
and has a sinuous course. At its distal end it is joined by two
(or more) hepatic ducts (h), bringing the bile from the lobes
of the liver. The relation of these to the cystic duct varies.
They may open into it by a common trunk or separately. Of
these hepatic ducts one is made up by the junction of smaller
hepatic ducts from the left half of the liver and the left half of
the cystic lobe, while the other is similarly formed by smaller
ducts from the right half of the cystic lobe, from both divisions
of the right lateral lobe and from the caudate lobe. The duct
formed by the junction of the hepatic and cystic ducts is the
common bile-duct (ductus communis choledochus) (i). It
passes in the free right border of the gastroduodenal omentum
to the duodenum (k) and opens into it by way of the ampulla
of Vater, in common with the pancreatic duct, at a point on
the dorsal surface of the duodenum and about three centimeters
from the pylorus.
Pancreas.—The pancreas (Fig. 102, a) is a flattened,
closely lobulated gland of irregular outline, about twelve centimeters
long, varying in width from one to two centimeters.
It is bent nearly at right angles at about its middle. One of
the halves (a′) into which it is divided by its bend lies in the
descending limb of the great omentum, and is near the greater
curvature of the stomach (d) and parallel to it. The free end[242]
of this half is in contact with the spleen (e). The other half
(a) lies in the duodenal omentum between the limits of the
duodenal U (c) and reaches to the bottom of the U. The
pancreas has two ducts. The larger pancreatic duct (b)
(sometimes known as the duct of Wirsung) collects the pancreatic
fluid from both halves of the gland, the ductlets from
each half uniting to make two larger ducts, which then unite
near the angle of the gland to make the pancreatic duct. This
is short and broad and opens into the ampulla of Vater together
with the common bile-duct. The accessory pancreatic duct
(duct of Santorini) opens into the duodenum about two centimeters
caudoventrad of the ampulla of Vater. It is formed by
the union of branches which anastomose with those of the
pancreatic duct. It is apparently sometimes lacking.
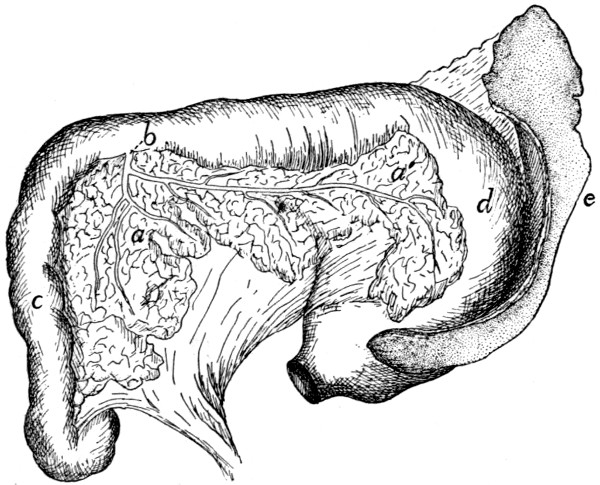
Fig. 102.—Pancreas and Spleen.
The œsophagus has been cut and the stomach turned caudad, so that the dorsal
surface of the stomach and the ventral surface of the duodenum are seen. a, pancreas
(a, duodenal portion; a′, gastric portion); b, pancreatic duct; c, duodenum; d,
stomach; e, spleen.
Spleen. Lien.—The spleen (Fig. 102, e) is a deep red,
flattened, elongated gland belonging to the lymphatic system.
One of its ends, the left, lies against the free end of the gastric
half of the pancreas and is broader than the other end. The
spleen is curved and is suspended in the descending limb of the
great omentum so that it follows the greater curvature of the
stomach (d).
[243]
III. RESPIRATORY ORGANS. APPARATUS RESPIRATORIUS.
The organs of respiration consist of the nasal cavity, the
nasopharynx, the pharynx (also a food-passage), the larynx
(also the organ of the voice), the trachea, the bronchi, and the
lungs. With them are usually described also the thyroid and
thymus glands.
1. The Nasal Cavity. Cavum nasi.
—The osseous framework
of the nasal cavity has already been described (page 59),
and in connection with this description the boundaries of the
cavity and its connections with other cavities have been given.
It consists essentially of a large cavity bounded by the facial
bones and divided by a longitudinal partition into two lateral
halves. The two cavities thus formed are nearly filled by (1)
the labyrinths of the ethmoid (ethmoturbinals), (2) the superior
nasal conchæ or nasoturbinals, projecting into the dorsal part
from the ventral surface of the nasal bones, and (3) the inferior
nasal conchæ, or maxilloturbinals, projecting into the ventral
portion from the medial surfaces of the maxillaries.
There remain to be considered, in addition to the bones,
the cartilaginous framework of certain parts of the nose, and
the mucous membrane. The lamina perpendicularis is continued
by cartilage, especially craniad, in such a way as to
make a complete septum separating the two cavities. This
septum extends from the septum of the external nose caudad
to the lamina cribrosa, and from the internasal suture ventrad
to the vomer and the suture of the premaxillaries. All parts
of the nasal cavity are lined by mucous membrane. This is
continuous at the nares with the integument, while at the
choanæ it passes into the mucosa of the pharynx. It covers
the conchæ nasales and the labyrinths of the ethmoid, passing
into the cellules of the latter. Owing to the crowding together
of the conchæ nasales and the labyrinths the nose is almost
completely filled, only three narrow passageways being distinguishable.
The ventral one of these, known as the ventral or
inferior meatus of the nose, lies ventrad of the inferior nasal
concha, next to the nasal septum. It passes caudad beneath[244]
the horizontal plate formed by the vomer and ethmoid, and
opens caudad at the choanæ into the nasopharynx. The
dorsal or superior meatus of the nose lies just ventrad of the
superior nasal concha, next to the median septum; it leads to
the frontal sinus, the lamina cribrosa, and the caudal parts of
the ethmoid. The middle meatus of the nose has almost disappeared
in the cat; it is simply the narrow space between the
superior and inferior meati. It is practically filled by the
ethmoid, into the cells of which it leads. The mucosa is continued
from the nasal cavity into the sinuses of the frontals and
presphenoid.
At the sides of the nasal septum, near the ventral edge,
and about one to one and a half centimeters caudad of the
nares, there is on each side a small curved cartilaginous tube,
about one centimeter or less in length. This, the vomeronasal
organ, or organ of Jacobson, lies against the septum, between
it and the mucosa. It begins at the incisive canal or anterior
palatine foramen, in the roof of the mouth, curves thence
caudodorsad close against the side of the nasal septum, and
ends blindly in the nasal cavity.
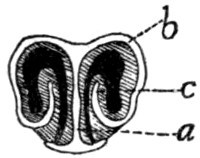
Fig. 103.—Cross-section
of the
Cartilages of
the External
Nose.
a, cartilage of the
median septum; b,
“wings”; c, ridge
formed by internal
portion of wing.
The nares or cranial openings of the nasal cavity are supported
by a number of cartilages which form the framework of
the snout or external nose. The cartilaginous continuation of
the lamina perpendicularis extends some distance craniad of
the tips of the nasal and premaxillary bones, forming the
septum of the external nose (Fig. 103, a).
From the dorsal edge of this projecting cartilaginous
septum, two thin cartilaginous wings
(b) extend laterad, forming the dorsal wall of
the narial opening. Each then turns ventrad
to form the lateral wall of the opening, but
does not form the ventral floor; instead it
curves mediad and finally dorsad, thus extending
from the floor of the opening as a
prominent ridge (c) within the nares. The
cartilaginous wing is thus rolled into a sort of
spiral, ending with a free edge within the narial opening.
From the ventral edge of the median cartilaginous septum there[245]
are likewise lateral extensions, which form part of the ventral
boundary of the nares; these do not quite reach the ventral
parts of the dorsal wings, however, so that a small part of the
narial opening is not bounded by cartilage. A section of the
narial cartilages is shown in Fig. 103. The wings from the
dorsal edge of the septum do not extend quite to the cranial
tip of the nose, so that a notch is formed on the ventrolateral
side of the nares; a section in this region would therefore differ
from that figured.
The framework of the external nose thus formed is covered
externally by thick hairless skin, containing many glands.
From the ventral end of the internarial septum a groove passes
ventrad, partly dividing the upper lip.
The inner surfaces of the cartilages are covered by the
mucosa, which forms a number of ridges. The narial opening
is almost completely divided by the prominent ridge which is
supported by the free edge (c) of the cartilaginous wing above
described. This free edge is covered by a thick layer of
mucosa, and the entire ridge so formed is continuous caudad
with the inferior nasal concha. On the medial side of the
partial partition formed by this ridge and near the ventral side
there begins a slight distance caudad of the outer opening
another ridge, supported by the ventrally incurved portion of
the cartilaginous ring. This soon becomes a thick swelling;
beneath it opens the lachrymal canal, and that part of the
passageway that lies ventromediad of it is the beginning of the
inferior meatus of the nose. About one or two centimeters
caudad of the external opening a third ridge projects from the
dorsolateral wall of the cavity toward the large ridge first
described; dorsomediad of it is a narrow passage which is the
entrance to the superior meatus of the nose.
The olfactory mucosa, or that part to which the olfactory
nerve is distributed, and which therefore acts as the sensory
surface, is confined to the dorsocaudal parts of the nasal
cavity, in the region occupied by the cells of the ethmoid.
The air penetrates to this region probably only by a definite
act of snuffing, the inferior meatus serving as the usual passageway
of air to the lungs.
[246]
The nasopharynx and pharynx are considered under the
alimentary canal (page 231).
2. The Larynx.
The larynx is the enlarged upper end of the air-passage
which leads from the pharynx to the lungs. It is a box composed
of pieces of cartilage connected by ligaments and moved
by muscles, and it is lined by mucous membrane. At the root
of the tongue about one centimeter caudad of the body of the
hyoid bone is the triangular leaf-like epiglottis (Fig. 95, e,
page 227). When food is taken this closes the opening into
the larynx, and the food passes over it into the œsophagus.
It is so curved that its apex is directed craniad. A fold of
mucous membrane, the plica glossoepiglottica or frenulum of
the epiglottis, extends from the middle of its cranial surface
to the root of the tongue, and on each side of this fold is a
depression.
From each side of the base of the epiglottis a fold, plica
aryepiglottica (Fig. 95, f), extends caudad to the base of the
arytenoid cartilage (g). Dorsad of this fold and separated
from it by a depression is a ridge which marks the position of
the caudal hyoid cornu. The plicæ aryepiglotticæ (f) and the
epiglottis (e) form the boundaries of the aditus laryngis, or
opening into the larynx.
The cavity of the larynx is divided into three portions.
The upper one of these is the vestibule of the larynx. It
is bounded caudad by two folds of mucosa (i) that stretch
from the caudal surface of the epiglottis near its base to the
tips of the arytenoid cartilages. These folds are the false
vocal cords (i). Their vibration is said to produce purring.
Caudad of the false vocal cords two folds of the mucosa
stretch from the apices of the arytenoid cartilages to the thyroid
cartilages, near the base of the epiglottis. These folds are
nearer the median plane than the false vocal cords. They
are the true vocal cords (j). The middle portion of the
laryngeal cavity is that between the true and the false vocal
cords. It is produced laterally into a very small pouch or
pocket on each side, the ventriculus. The narrow slit between
the true vocal cords is the glottis (h). It can be narrowed[247]
and widened by the action of muscles. The vocal cords, which
bound it, are set vibrating by currents of air transmitted from the
lungs, and the voice-sounds are thus produced. The caudal
portion (inferior portion) of the laryngeal cavity is that between
the glottis and the first tracheal cartilage. It is narrowed near
the glottis.
Cartilages of the Larynx
(Fig. 104).—There are
three unpaired cartilages, the thyroid (1), cricoid (3), and
epiglottic (2), and two paired cartilages, the arytenoids (4).
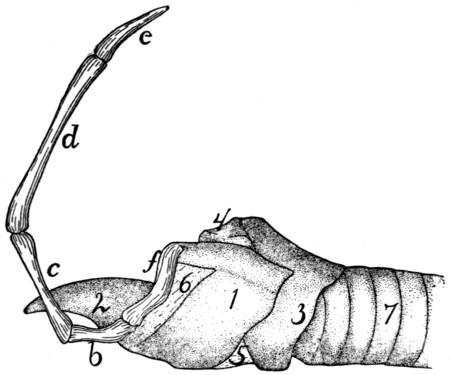
Fig. 104.—Cartilages
of Larynx, with Side View of Hyoid Bone.
b, ceratohyal; c, epihyal; d, stylohyal; e,
tympanohyal; f, thyrohyal. 1, thyroid
cartilage; 2, epiglottis; 3, cricoid cartilage; 4, arytenoid cartilage; 5, cricothyroid
ligament; 6, thyrohyoid ligament; 7, trachea.
The thyroid cartilage (cartilago thyreoidea) (1) has nearly
the form of a visor of a cap, but is relatively broader at its ends
than a cap visor. It forms about two-thirds the circumference
of a circle, and is so situated that it embraces the other cartilages
ventrally and laterally. Its caudal and cranial borders
are oblique to its caudocranial axis and are directed dorsocaudad.
To the middle of the cranial border is attached the
epiglottic cartilage (2), and the whole cranial margin is connected
by membrane (6) to the body and caudal cornua (f)
of the hyoid bone. The dorsal border projects craniad into a
considerable cornu which is attached to the free end of the
caudal hyoid cornu (f). The border also projects caudad into
a process which articulates with a facet on the lateral surface
of the cricoid cartilage (3). In the middle of the
dorsal surface[248]
is a longitudinal ridge for attachment of the vocal cords and
origin of the thyroarytenoid muscles (Fig. 105, d), and at the
caudal end of this ridge the caudal border presents a considerable
rounded notch.
The cricoid cartilage (cartilago cricoidea) (3) has the form
of a seal ring with its broader part dorsad. The broad dorsal
part of the ring is partly embraced by the wings of the thyroid
cartilage (1). Its caudal border is undulating and nearly at
right angles to its caudocranial axis. It is connected by
membrane with the first tracheal ring. The cranial border is
oblique to the long axis and lies in a plane which passes from
the ventral side craniodorsad. Near the median line on each
side it articulates by an oblique facet with one of the two
arytenoid cartilages (4). Near the midventral line the cricothyroid
ligament (5) is attached and it stretches thence to the
midcaudal notch of the thyroid cartilage (1).
The outer surface presents at the middle of each side a
facet for articulation with the thyroid, and is marked in the
median dorsal line by a ridge for the posterior cricoarytenoid
muscle (Fig. 105, b).
The arytenoid cartilages (cartilago arytenoidea) (Fig.
104, 4) are triangular pyramids with base and sides nearly
equilateral triangles. One side articulates with an oblique facet
on the cranial border of the cricoid near the mid-dorsal line.
Near the opposite apex is attached the vocal cord. At the
lateral angle of the base is attached on its dorsal side the posterior
cricoarytenoid muscle (Fig. 105, b), and on its ventral
side the thyroarytenoid (Fig. 105, d) and lateral cricoarytenoid
(Fig. 105, c) muscles. The movements produced by
these muscles carry the arytenoid ends of the vocal cords toward
or from the median plane and thus open or close the glottis.
The epiglottic cartilage (Fig. 104, 2) is flexible (fibro-cartilage)
and of a cordate form. It is so curved that its caudal
surface is convex dorsoventrally and concave from side to side.
It supports the epiglottis. Its cranial surface presents a slight
median ridge for attachment of muscles. By its base it is
attached to the midventral part of the cranial border of the
thyroid cartilage (1). Its position varies so
that it either stands[249]
erect with its apex directed craniad to allow the passage of air
to the lungs, or, as in the act of swallowing, it is turned caudad
over the aditus laryngis so as to allow food to pass over it and
into the œsophagus.
The vocal cords are two fibrous elastic bands. Each is
attached at one end to the apex of the arytenoid cartilage, and
at the other end to the median ridge on the dorsal surface of
the thyroid. Each supports a projecting fold of mucous membrane,
the vibration of which causes the voice.
Muscles of the Larynx.
—1. Muscles moving the entire
larynx.
A. Elevators. M. thyreohyoideus (Fig. 96, p, page 229).—A
flat band on the lateral side of the larynx.
Origin on the lateral part of the caudal border of the
thyroid cartilage.
Insertion on the medial two-thirds of the caudal border of
the caudal cornu of the hyoid.
Action.—Raises the larynx.
The stylohyoid (Fig. 65, d, page 109) and the median and
inferior constrictors (Fig. 96, j and k) of the pharynx, already
described, have the same action.
B. Depressors.—The sternothyroid (Fig. 65, g′), already
described (p. 141).
2. Muscles which move the parts of the larynx one upon
another.
A. Muscles on the Outer Surface of the Larynx.
M. cricothyreoideus (Fig. 96, n).—A broad flat band
which with its fellow covers the ventral surface of the cricoid
cartilage and the cricothyroid ligament.
Origin.—The lateral half of the ventral surface of the cricoid
cartilage. The muscles diverge so as to leave a part of the
cricothyroid ligament between them.
Insertion.—The ventral part of the caudal border of the
thyroid cartilage laterad of the median ventral notch.
M. cricoarytenoideus posterior (Fig. 105, b).—The two
muscles cover the dorsal surface of the larynx.
Origin.—From the dorsal part of the caudal border of the
cricoid cartilage (3) and from its median dorsal crest. The[250]
fibres pass craniolaterad, converging. The lateral fibres are
nearly longitudinal in direction.
Insertion.—The dorsal border of the caudal end of the
arytenoid cartilage (4).
Action.—Moves the arytenoid on its oblique articulation
with the thyroid. The apex of the arytenoid is thus carried
laterad, and the vocal cords are separated so as to widen the
glottis.
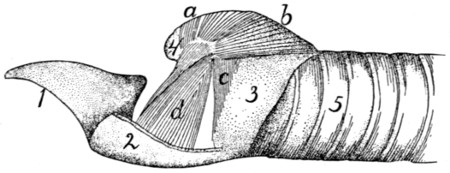
Fig. 105.—Muscles
of the Larynx, as Exposed by Removal of most of
the Left Half of the Thyroid Cartilage.
1, epiglottis; 2, portion of the thyroid cartilage (cut); 3, cricoid cartilage; 4,
arytenoid cartilage; 5, trachea. a, M. arytenoideus transversus; b, M. cricoarytenoideus
posterior; c, M. cricoarytenoideus lateralis; d, M. thyreoarytenoideus.
M. arytenoideus transversus (Fig. 105, a).—A small
unpaired muscle running transversely between the caudal ends
of the arytenoid cartilages, just beneath the mucous membrane
and parallel to the cranial border of the cricoid cartilage.
M. glossoepiglotticus.—A small longitudinal muscle, lying
by the side of its fellow in the frenulum of the epiglottis.
Origin.—The median fibrous septum of the tongue.
Insertion.—The dorsal (cranial) surface of the epiglottic
cartilage in the median line near its attached border.
Action.—Draws the epiglottis craniad.
M. hyoepiglotticus is a small bundle of parallel fibres lying
in the frenulum of the epiglottis craniad and dorsad of the body
of the hyoid bone.
Origin.—The lateral end of the cranial surface of the body
of the hyoid. The two muscles pass craniodorsad, converging,
and unite with the preceding.
Insertion with the preceding. (The lateral portion of the
muscle may continue craniad to the tongue as a second part of
the hyoglossus.)
Action.—Like the preceding.
[251]
B. Muscles on the Inner Surface of the Cartilages of the
Larynx.
M. thyreoarytenoideus (Fig. 105, d).—The thyroarytenoid
is a triangular, flat muscle of considerable size. It lies within
the wing of the thyroid cartilage (2), and its fibres are nearly
dorsoventral in direction.
Origin.—The median longitudinal crest on the dorsal surface
of the thyroid cartilage (2).
Insertion.—The fibres converge to the insertion into the
cranial lip of the laterocaudal angle of the arytenoid cartilage
(4).
Action.—Turns the arytenoid on its oblique articulation so
as to close the glottis.
M. cricoarytenoideus lateralis (Fig. 105, c).—Triangular,
a little smaller than the preceding, caudad of which it is
situated, so that it also is covered by the wing of the thyroid.
Origin.—The lateral part of the cranial border of the
cricoid cartilage (3). The fibres converge, passing dorsad.
Insertion.—The caudal lip of the laterocaudal angle of the
arytenoid cartilage (4).
Action.—Similar to the preceding, so that it closes the
glottis.
3. The Trachea
(Fig. 105, 5;
Fig. 106, a).—The trachea
is that part of the air-passage which extends from the larynx
to the bronchi (Fig. 106). It is a straight tube composed of a
lining mucosa with ciliated epithelium, and a connective-tissue
covering which encloses supporting cartilages. Each tracheal
cartilage is incomplete dorsally where it lies against the
œsophagus, the gap between the two free ends of each ring
being filled with muscular and connective tissue. As a result
of this the diameter of the trachea is not fixed, but can be
increased and diminished. The first ring is broader than the
others. Where the œsophagus leaves the median line, the
dorsal surface of the trachea lies against the longus colli muscles
(Fig. 72, g″). Its ventral surface is against the sternohyoid
(Fig. 65, e) and sternothyroid (Fig. 65, g′) muscles.
Its lateral surfaces are partly covered by the thyroid gland
(Fig. 96, 6), and are in close
relation with the carotid artery[252]
(Fig. 119, a, page 284), the vagus and sympathetic nerves
(Fig. 156, i), and the internal jugular vein (Fig. 119, b).
In the thoracic cavity the great vessels coming from the
heart lie against the ventral surface of the trachea (Fig.
129, 2). At about the level of the sixth rib the trachea
divides into the two main bronchi (Fig. 106). Each bronchus
is supported by incomplete rings of cartilage like those of
the trachea and has otherwise in general the structure of the
trachea. In the lungs the bronchi become divided into many
branches (Fig. 106), in the manner described in the account
of the lungs.
4. The Lungs. Pulmones.
—Immediately after division
of the trachea the two bronchi enter the lungs (Fig. 106).
These are two large, much-lobed organs, which fill the greater
part of the thoracic cavity. The main lobes of the lungs are
completely separated from each other, except in so far as they
are connected by the bronchi and connective tissue; the main
lobes may also be partly subdivided into secondary lobes that
are not thus completely separated. The two lungs are completely
separated from each other, except at the radix, where
they are united by the bronchi; they lie in the right and left
halves of the thoracic cavity, with the mediastinal septum
between them. The bronchi on entering the lungs divide in
the following manner. Each divides at first into two main
branches. The cranial branch on the right side is known as
the eparterial bronchus (b), because it lies craniad of the pulmonary
artery. All the others are hyparterial; i.e., they lie
caudad of the pulmonary artery. The right cranial bronchus
does not further subdivide into large bronchi, but gives off
numerous small branches. The right caudal bronchus divides
into three main branches. There are thus four main branches
of the right bronchus. The left cranial bronchus divides into
two main branches; the left caudal bronchus continues caudad
as a main trunk giving off small branchlets. Of the left
bronchus there are thus but three main subdivisions. The
main subdivisions of the bronchi on the two sides correspond,
as will be seen, with the lobulation of the lungs.
The right lung (Fig. 106,1-4) is slightly larger than the[253]
left (1′-3′).
It divides into three smaller proximal lobes (1-3),
and one large distal one (4). The most cranial one of the
proximal lobes (1) is sometimes partly subdivided. The third
one of the proximal lobes (3) lies mediad of the others; it is
partly subdivided and one-half projects into a pocket in the
mediastinum, so that it comes to lie across the middle line,
extending a short distance onto the left side. This lobe is
frequently called the mediastinal lobe (3). The caudal lobe
(4) of the right lung is large and flat, containing about half the
substance of the lung.
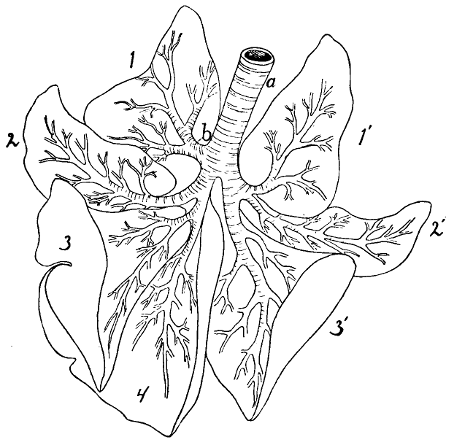
Fig. 106.—Ramifications of the Bronchi, with Outlines of the Lobes
of the Lungs, Ventral View.
1-4, lobes of the right lung; 1′-3′, lobes of the left lung. a, trachea; b, eparterial
bronchus.
The left lung is divided into three main lobes (1′-3′); the
two cranial ones (1′-2′) are, however, partly united at the base,
so that they may be considered subdivisions of but a single
lobe; thus the left lung has but two distinctly separated lobes.
Each lung is attached to the aorta, vertebral column, and
diaphragm by a fold of pleura, the pulmonary ligament. This[254]
is broadest at the caudal lobe of each lung. Each pulmonary
ligament is double, being formed of two sheets of the pleura.
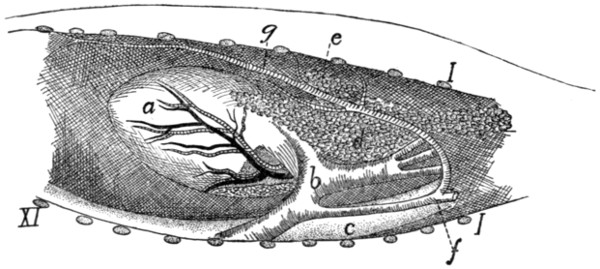
Fig. 107.—Position
of Thymus Gland, from Left Side.
a, heart; b, aorta; c, œsophagus; d,
thymus gland; e, lymphatic gland; f, left
subclavian artery; g, internal mammary artery. I, cut ends of first rib; XI, eleventh
rib.
The Thyroid Gland. Glandula thyreoidea
(Fig. 96, 6).—The
thyroid gland consists of two lateral lobes (6) and a
median lobe or isthmus (7). Each lateral lobe (6) is an
elongated, flattened, lobulated mass with round ends. It is
about two centimeters long and about one-fourth as broad. It
lies at the side of the trachea (4), dorsad of the lateral margin
of the sternohyoid muscle. Its cranial end is at the level of
the caudal border of the cricoid cartilage. The isthmus (7)
is a delicate band two millimeters wide which connects the
caudal ends of the two lateral lobes. It passes ventrad of the
trachea and in close contact with it. The thyroid has no duct.
The Thymus Gland. Glandula thymus
(Fig. 107, d).—The
thymus gland is best developed in young kittens; in the
adult cat it has partly or almost completely degenerated. It is
an elongated, flattened organ, of a pinkish-gray color, which lies
in the mediastinal cavity, between the two lungs and against
the sternum. It extends caudad as far as the heart (a), overlying
the pericardium at its posterior end. At its cranial end
it projects, when well developed, a short distance (about one
centimeter) outside of the thoracic cavity into the neck region.
The caudal end is forked, and the left lobe thus formed is
usually larger than the right. The cranial end may also show
indication of a division into two lobes, but this is frequently not
the case.
[255]
IV. THE UROGENITAL SYSTEM. APPARATUS UROGENITALIS.
1. The Excretory Organs.
Kidney. Ren
(Figs. 108 and 109).
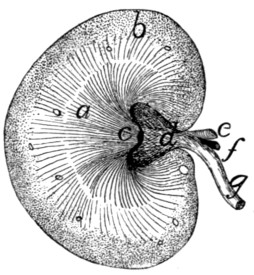
Fig. 109.—Median Longitudinal
Section of Kidney.
Fig. 108.—a, renal artery; b, renal vein; c, ureter.
Fig. 109.—a, medullary portion; b,
cortical portion; c, papilla; d, pelvis; e, renal
artery; f, renal vein; g, ureter.
The kidneys of the cat are compact (i.e., not lobulated) and
have the usual kidney or bean form. They lie in the abdominal
cavity, one on either side of the vertebral column, against
the dorsal body wall, in the region between the third and fifth
lumbar vertebræ. The right kidney is one or two centimeters
farther craniad than the left, and the long axes of the two converge
craniad a little. Each is covered by peritoneum on
its ventral surface only (i.e., it is retroperitoneal). At the
border of the kidney, where the peritoneum passes from it to
the body wall, there is an accumulation of fat, which is most
abundant at the cranial end of the kidney. Within the peritoneal
investment the kidney is enclosed in a special loose
fibrous covering, the capsule or tunica fibrosa, which is continuous
with the fibrous coat of the ureter and pelvis. In the
middle of the median border of each kidney is a notch, the
hilus. It gives exit to the ureter (Fig. 108, c) and renal veins
(b), and entrance to the renal artery (a). On the ventral surface
of the kidney within the capsule are seen grooves radiating
from the hilus. They contain blood-vessels. If the substance
of the kidney is sliced away parallel to the ventral surface for
some distance (Fig. 109), there is exposed a cavity, the sinus,[256]
which lies near the medial border and the opening of which is
the hilus. It contains the pelvis (d) (the expanded beginning
of the duct of the kidney), and also renal vessels (e and f) with
their branches. These structures are enclosed in fat, which fills
the remainder of the sinus. Upon opening the pelvis the
kidney substance is seen to project into it in the form of a cone,
the papilla (c), the apex of which is directed mediad. On the
apex of the papilla are the numerous openings of the uriniferous
collecting-tubes, some of them opening at the bottom of an
apical depression of the papilla.
In a section made parallel to the ventral surface and in the
median plane, the substance of the kidney is seen to consist of
a peripheral darker and more granular cortical portion (Fig.
109, b), and of a central, lighter, less granular medullary
portion (a). Both portions are marked by lines which converge
to the apex of the papilla (c).
The Ureter
(Fig. 108, c; Figs. 111
and 112, b).—The
duct of the kidney begins as the pelvis (Fig. 109, d), a conical
sac the base of which encloses the base of the papilla.
From the apex of the papilla the urine passes into the pelvis.
The outer wall of the pelvis is continuous with the capsule of
the kidney. At the hilus the pelvis narrows to form the ureter
(Fig. 109, g). The ureter passes caudad in a fold of peritoneum
which contains fat. Near its caudal end it passes
dorsad of the vas deferens (Fig. 111, c), turns ventrocraniad,
and pierces the dorsal wall of the bladder (Fig. 111, a)
obliquely near the neck. On the inside of the bladder the
openings of the ureters appear as pores about five millimeters
apart, and each is surrounded by a white, ring-like elevation
of the surface.
The Bladder. Vesica urinaria.
—The bladder (Fig.
111, a) is pear-shaped. It lies in the abdominal cavity between
its ventral wall and the rectum and a short distance craniad of
the pubic symphysis. Caudad it is continued into a rather
long, narrow neck (f) which passes dorsad of the symphysis
to the pelvic cavity.
The bladder is covered by peritoneum and is held in place
by its neck and by three folds of the peritoneum. One of these[257]
passes from its ventral wall to the linea alba and is the suspensory
ligament. Two others pass one from each side of the
bladder to the dorsal body wall at the sides of the rectum.
They are the lateral ligaments of the bladder. They form the
walls of a partly isolated peritoneal pocket into which the
rectum passes; this pocket opens craniad into the peritoneal
cavity. The wall of the bladder is composed of an internal
epithelium, a layer of plain muscle-fibre bundles which cross
one another in various directions, and the external peritoneal
layer.
Suprarenal Bodies. Glandulæ suprarenales.
—The
suprarenal bodies are two ovoid bodies about a centimeter
in the longest diameter, lying craniomediad of the kidneys, but
usually not touching them. In a fresh condition they are of a
pinkish or yellow color. They are usually imbedded in fat and
are covered by peritoneum on their ventral surface. They have
no duct and are of uncertain function.
2. The Genital Organs.
A. The Male Genital Organs.
External Genital
Organs.—The external genital organs are the scrotum and
penis.
The scrotal sac or scrotum is a pouch of integument which
lies ventrad of the anus in the median line against the ischiatic
symphysis. It is marked by a median groove which indicates
the position of an internal septum dividing its cavity into lateral
halves, within each of which is one of the testes.
The penis (Fig. 111, l; Fig. 113,
6; see also page 262) lies
ventrad of the scrotal sac. It projects caudad. It is covered
by the integument, which projects at its end as a free fold, the
prepuce. Within the prepuce is the projecting glans penis
(Fig. 113, 7). It is conical and bears on the ventral side of its
free end the opening of the urethra, the common urinogenital
opening. On the side on which the urethra opens the glans is
connected to the prepuce by a fold of integument, the frenulum.
The surface of the glans is covered with sharp, recurved,
horny papillæ.
[258]
The Scrotum and Testes, and the Ducts of the Testes.—The
scrotum contains the two testes, one in each of its compartments.
Each testis lies in a diverticulum of the abdominal
cavity, which is lined by an extension of the peritoneum.
The testis has the same relation to this peritoneal diverticulum
that the intestine has to the abdominal cavity; i.e., it does not
lie within the cavity of the diverticulum, but is suspended
apparently within it by means of a fold of its wall which acts
as a mesentery. The peritoneal diverticulum is called the
tunica vaginalis propria and consists thus of a parietal layer
and a visceral layer.
The tunica vaginalis propria consists of a slender proximal
part through which the blood-vessels pass to the testis and the
vas deferens from it, and of an expanded distal part in which
lies the testis. Only the distal part lies within the scrotum.
The blood-vessels and vas deferens are suspended in the narrow
part of the tunica vaginalis propria by means of a mesenterial
fold similar to that which suspends the testis, and continuous
with it. This fold and the blood-vessels and vas deferens contained
within it form the spermatic cord (Fig. 111, d) which
passes from the abdomen to the scrotal sac in the narrow part
of the tunica vaginalis propria. In the formation of the human
tunica vaginalis the various layers of the body wall are carried
out by it and form the tunics, or coats of the testis. The one
of these coats next the tunica vaginalis propria (which is
reckoned as one of the coats) is the tunica vaginalis communis
(or fascia propria) and is formed by the transversalis fascia.
Outside of the tunica vaginalis communis is the cremaster
muscle, an incomplete layer formed from the fibres of the
internal oblique muscle. Next is the cremasteric (or intercolumnar)
fascia from the aponeurosis of the external oblique
muscle, and outside of this is the integument forming the
scrotum. In this integument there is a layer of smooth muscle
which is sometimes described as the tunica dartos. In the cat
the tunica dartos and the cremaster muscle are wanting. The
cremaster is replaced by the elevator scroti muscle. The coats
of the testis are thus five, as follows:
1. The scrotum (the integument).
[259]
2. Cremasteric fascia (subcutaneous fascia).
3. The levator scroti muscle (subcutaneous muscle-layer)
(Fig. 113, j).
4. Tunica vaginalis communis (transversalis fascia).
5. Tunica vaginalis propria (peritoneum).
The tunica vaginalis communis is inseparably united with
the parietal layer of the tunica vaginalis propria. Where the
spermatic cord passes from the abdominal wall to the scrotum
it is covered by integument and cremasteric fascia on its ventral
surface only, but is entirely surrounded by the tunica vaginalis
propria and tunica vaginalis communis. The canal by which
the spermatic cord passes through the body wall is known as
the inguinal canal. The end by which it opens into the
abdominal cavity is the internal inguinal ring, and the opposite
end is called the external inguinal ring.
The internal inguinal ring is merely the point of connection
between the proximal tubular portion of the tunica
vaginalis propria and the abdominal cavity. It is circular, and
is situated close against the lateral side of the lateral ligament
of the bladder at its middle.
The external inguinal ring is an oval opening in the aponeurosis
of insertion of the external oblique muscle. The aponeurosis
of this muscle ends caudally in a free border along the
cranial edge of the pubis, from the ilium to the pubic tubercle.
The external ring is just craniad of the end of this aponeurosis.
The inguinal canal between these rings lies along the
lateral border of the rectus muscle. It is one to one and a
half centimeters long. Its medial wall rests on the rectus
muscle; its dorsal wall on the fat contained within the lateral
ligament of the bladder. The lateral and ventral walls lie on
the transversus muscle proximally, on the internal oblique near
the distal end. The wall itself is composed of tunica vaginalis
propria and communis. As it passes within the caudal border
of the internal oblique muscle it receives some aponeurotic
fibres from it. A thin aponeurosis is also continued from the
border of the external ring onto the tunica vaginalis propria.
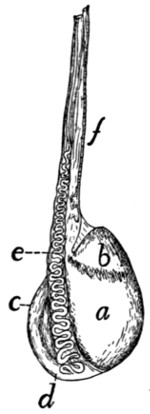
Fig. 110.—Testis.
a, testis; b, caput
epididymis; c, epididymis;
d, cauda
epididymis; e, vas
deferens; f, spermatic
cord.
The testes (Fig. 110) are the organs which produce the
spermatozoa. They lie one in each compartment of the scrotal[260]
sac, enveloped in the membranes or tunics described when
treating of the scrotum. Each is attached to the dorsal wall
of its peritoneal pouch by a mesenteric fold. The testis is surrounded
by the visceral layer of the tunica vaginalis propria,
and within this, by a dense fibrous covering, the tunica albuginea,
which sends septa into its interior. Within the fibrous
covering it is made up of numerous coils of seminiferous tubules
which are readily seen by the naked eye.
The epididymis (Fig. 110, c) is the beginning of the
efferent duct of the testis. It appears as a flat band with a broad
rounded end which lies on the medial surface of
the testis at its cranial end (b). From this
point it passes about the cranial end of the testis
from its medial to its lateral surface, forming
thus a semicircle with the convexity ventrad.
Thence it passes as a narrower band (c) along
the dorsal side of the testis, laterad of the suspending
mesentery, to the caudal end of the
testis (d). At the caudal end of the testis it
is enlarged, passes from its lateral to its medial
surface, and turns at the same time craniad to
become continuous with the vas deferens (e).
The enlarged cranial end is the caput epididymis
(b); the enlarged caudal end the cauda
epididymis (d). The whole epididymis is
encased in a tough fibrous covering similar to
that of the testis. The fibrous covering (albuginea)
of the testis and that of the epididymis
are connected by fibrous tissue. Within the
fibrous covering the head of the epididymis is made up of tubules
which pass from the testis into its end: these are the vasa
efferentia testis. The vasa efferentia unite within the caput
into a single vessel which passes in a very tortuous course to
the cauda. Its numerous windings form the cauda, from the
end of which it passes craniad as the vas deferens (e).
The vas deferens (Fig. 110, e; Fig. 111, c) is a slender
tube much convoluted at its beginning. It passes from the
cauda epididymis (d) along the mesenterial fold of the testis[261]
on its medial side, to the spermatic cord (Fig. 111, d). It
leaves the spermatic cord at the internal inguinal ring and,
curving over the ureter (Fig. 111, b), bends craniad and
approaches the vas deferens of the opposite side dorsad of the
neck of the bladder (Fig. 111, f). The two vasa deferentia
pass caudad together as far as the cranial border of the pubis.
There they enter the prostate gland (g), pierce the dorsal wall
of the neck of the bladder, and open close together on the inner
surface. The two openings are separated by a slight elevation,
the veru montanum.
The Urethra (Fig. 111, h).—The urethra is the common
urinogenital duct which is formed by the union of the neck of
the bladder (f) and the vasa deferentia (c) dorsad of the cranial
border of the pubis. It extends thence to the end of the penis.
It is divided into three portions.
1. The prostate portion is the commencement of the
urethra; it is surrounded by the prostate gland (g).
2. The membranous portion (h) extends from the prostate
portion to a point between the crura of the penis. This portion
is surrounded by the thick compressor urethræ muscle, so
that its wall appears much thicker than it really is.
3. The spongy portion (pars cavernosa) extends along the
ventral side of the penis to its end in the groove between the
corpora cavernosa penis. At its beginning is an enlargement
formed by the bulbocavernosus muscle. This is known as the
bulbus urethræ. The walls of the spongy portion are thick
and vascular and form the corpus cavernosum urethræ or
corpus spongiosum. At its end the corpus cavernosum urethræ
is greatly enlarged and forms the glans penis (m).
Glands of the Urethra.—1. The prostate (Fig. 111, g) is a
bilobed gland lying on the dorsal wall of the urethra and surrounding
the ends of the vasa deferentia (c). It opens into the
urethra at its beginning by numerous small ducts visible to the
naked eye on the inner surface of the urethra.
2. The bulbourethral or Cowper’s glands (Fig. 111, i).—There
are two bulbourethral or Cowper’s glands, one on either
side of the bulbus of the urethra between the ischiocavernosus
and bulbocavernosus muscles. Each has a covering of muscle-fibres[262]
derived from the neighboring bulbocavernosus (Fig.
113, l). Each is said to open by a single duct into the urethra
at the root of the penis.
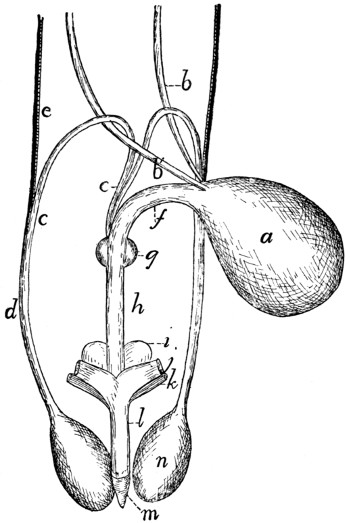
Fig. 111.—Male Genital Organs.
a, bladder; b, b′, ureters; c,
vasa deferentia; d, spermatic cord; e, spermatic artery
and vein; f, neck of bladder; g, prostate gland; h, urethra; i, bulbourethral (or
Cowper’s) gland; j, corpus cavernosum penis, cut from ischium; k, ischiocavernosus
muscle (cut); l, penis; m, glans penis; n, testis.
The penis (Fig. 111, l;
Fig. 113, 6) is a cylindrical organ
with the apex directed backward. It is covered by integument
which projects at its free end in the form of a fold, the prepuce
(Fig. 113), which ensheaths the glans (Fig. 113, 7) of the
penis. Beneath the integument is a layer of strong fibrous
subcutaneous fascia continuous with that of the surrounding
parts. On the dorsum of the penis a thin band of fibrous tissue,
the ligamentum suspensorium penis, is continued from the[263]
middle line beneath the pelvic symphysis. This band divides
distally, and its halves ensheath the glans and thus form a support
of the penis.
The penis is formed by three bodies, the two corpora
cavernosa penis and the single corpus cavernosum urethræ
(corpus spongiosum).
The corpora cavernosa penis (Fig. 111, j). Each is a
cylindrical sheath of dense fibrous tissue within which are
trabeculæ separating blood-sinuses. Each corpus cavernosum
is attached by the one pointed end to the caudal border of the
ramus of the ischium near the symphysis. From their attachments
the corpora cavernosa approach one another, forming
the crura of the penis. They become closely united by their
medial surfaces and pass thus to the free end of the penis where
their somewhat pointed distal ends are imbedded in the
glans (m). A groove is left between the corpora cavernosa on
the dorsum of the penis, and there is a second groove on the
ventral side. In the latter groove lies the urethra.
The corpus cavernosum urethræ is the spongy portion of
the urethra which lies in the groove on the ventral surface of
the penis, between the corpora cavernosa penis. It becomes
greatly enlarged at the distal end of the penis, forming the
glans (m). A small bone (os penis) is imbedded in the distal
end of the penis.
B. Female Genital Organs
(Fig. 112).—The urinal
organs of the female are like those of the male. The neck (k)
of the bladder is, however, much longer, extending almost to
the border of the ischiatic ramus. The urethra is consequently
short.
The genital organs consist of the ovaries (c) (the organs
which produce the eggs), the oviducts or uterine tubes (e)
(Fallopian tubes), which receive the eggs from the ovaries; a
bifid uterus (f and i) in which the ova undergo their development,
and a vagina (m) which leads from the uterus to its
junction with the neck of the bladder.
The neck of the bladder (k) unites with the vagina to form
the urogenital sinus (n) which is very short, leads to the
external opening, and is comparable to the male urethra.[264]
Ventrad of the external orifice of the urogenital sinus is the
clitoris, a rudimentary structure homologous with the penis of
the male.
The Ovaries (c).—The ovaries lie in the abdominal cavity
in the same longitudinal line with the kidneys and a short distance
caudad of them. Each is an ovoid body about one centimeter
long and one-third to one-half as broad. On its surface
are numerous whitish projecting vesicles, the larger of which
show clear centres. They are the Graafian follicles (best seen
in section), which contain the eggs. There may be present
one or more elevations of the size of the largest Graafian follicles,
but of a bright red or brown color. They are the corpora
lutea (sing. corpus luteum),—Graafian follicles from which the
eggs have been discharged.
The ovary is held in position by the broad ligament of the
uterus, a fold of the peritoneum, which passes here from the
uterine tube to the adjacent body wall. The ovary lies in a
sort of a pocket formed by the broad ligament. In the natural
position the pocket opens ventrolaterad. The ovary is further
held in position by the ligament of the ovary (ligamentum
ovarii), a short thick cord which passes from the ventral face
of the ovary at its uterine end to the adjacent ventral surface
of the uterus.
The Uterine Tubes (e).—The uterine (or Fallopian) tubes
or oviducts are the tubes which convey the ova from the ovary
(c) to the uterus (f). Each begins with an expanded trumpet-shaped
opening, the ostium tubæ abdominale (d). Its walls
are thin, and the mucosa of its inner surface is thrown up into
undulating, radiating ridges.
The ostium (d) lies on the lateral side of the ovary (c) at
its cranial end, and the trumpet partly clasps the ovary. From
the ostium the tube (e) turns craniad, then mediad, and then
caudad, so as to describe a curve about the cranial end of the
ovary. It then extends caudad on the mediodorsal aspect of
the ovary to its junction with the uterine cornu (f). It is
sinuous throughout its course, and the first two-thirds (the
vestibulum) is of considerably greater diameter than the last
third. Throughout the last two-thirds of its course it lies in
the free border of the broad ligament. Its mucosa is thrown
into irregular folds, mostly longitudinal, and is lined by ciliated
epithelium. From the foregoing description it is seen that the
ova must pass through the body cavity in order to reach the
ostium tubæ.
[265]
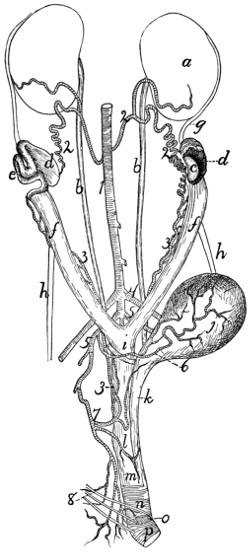
Fig. 112.—Female
Urogenital Organs, Obliquely Ventral View.
a, kidney; b, ureter; c, ovary; d,
ostium tubæ abdominale; e, uterine (Fallopian)
tube; f, cornua of the uterus; g, cranial edge of broad ligament; h, round ligament
of the uterus; i, body of the uterus; j, bladder; k, neck of the bladder; l, position
of cervix uteri; m, vagina; n, urogenital sinus or vestibule, with M. urethralis; o,
corpus cavernosum clitoridis, with M. ischiocavernosus (cut); p, fibres of M. constrictor
vestibuli. 1, aorta; 2, internal spermatic arteries; 3, uterine arteries; 4,
external iliac arteries; 5, hypogastric arteries; 6, umbilical artery; 7, inferior hemorrhoidal
artery; 8, branches of N. pudendus.
[266]
The Uterus (f, i).—The uterus consists of a median portion
or body (i) which is unpaired, and of two horns or cornua
(f) which extend from the body to the uterine tubes. The
body (i) of the uterus is a tube about four centimeters long
which lies in the abdominal cavity, ventrad of the rectum and
between it and the bladder. Its caudal end is at the level of
the cranial border of the pubis. The cranial end of its cavity
is divided by a median dorsoventral partition into lateral halves,
while the cavity of its caudal portion is unpaired. The mucosa
is thrown up into large longitudinal folds. At its caudal end
the uterus projects into the vagina (m), so that when the vagina
is opened the end of the uterus is seen projecting into it as a
prominent papilla. The portion of the uterus thus enclosed by
the vagina is the cervix uteri or neck of the uterus. Its free
end is directed ventrocaudad, and a prominent ridge is continued
from its caudodorsal side along the mid-dorsal wall of the
vagina. The uterine cavity communicates with the vaginal
cavity by a V-shaped opening, the os uteri, which looks ventrocaudad
and has its apex directed cranioventrad. Each horn
(f) of the uterus passes craniolaterad in a nearly straight course
from the body (i) to the uterine tube (e). It narrows rapidly
and becomes continuous with the tube. The mucosa is thrown
into longitudinal folds.
Ligaments of the Uterus.—The Broad Ligament.—The
uterus is held in place principally by the broad ligaments.
These are two folds of the peritoneum, each of which is attached
to the whole length of one of the cornua and the adjacent part
of the uterine tube and to the corresponding lateral surface of
the body of the uterus. Each ligament ends craniad in a concave
free border. Its attached border forms a curved line
which begins laterad of the kidney and extends thence to the
lateral ligament of the bladder. From the lateral ligament of
the bladder the broad ligament extends caudad into the rectovesical[267]
pouch of the peritoneum, which lies between the rectum
and the bladder. It holds the body of the uterus to the lateral
wall of this pouch, and together with the opposite ligament
and the body of the uterus forms thus a transverse partition,
which divides the rectovesical pouch into dorsal and ventral
portions.
The round ligament (h) is a fibrous band which extends
from a point of the body wall, which corresponds exactly to
the internal inguinal ring of the male, to the cornu of the
uterus about two centimeters from the cranial end. It is
attached to the broad ligament by an intervening fold of
peritoneum.
The Vagina (m).—The vagina extends from the os uteri
(at l) dorsad of the symphysis of the pelvis to a point a short
distance craniad of the caudal border of the ischiatic symphysis.
At this point it joins the neck of the bladder (k) to form the
vestibulum or urogenital sinus (n), which is homologous with
the urethra of the male.
Urogenital Sinus (n).—The urogenital sinus extends from
the caudal end of the vagina (m) to the external opening,
which is situated ventrad of the anal opening. It is about a
centimeter long and nearly as wide, and is marked off from
the vagina by a circular fold of mucosa, while its inner surface
presents longitudinal folds. On its ventral wall at its cranial
end is the opening of the neck of the bladder, which is
enclosed by a ring-like elevation of the mucosa, most prominent
at the sides. The external entrance to the urogenital sinus
forms the vulva.
The Clitoris.—The clitoris is a minute organ homologous
with the penis and lying on the ventral floor of the urogenital
sinus. Its distal end lies at the entrance of the urogenital
sinus on its ventral border. In adult specimens the prepuce
of the clitoris appears as a slight elevation of the integument
surrounding a central vascular structure which appears red in
the fresh organ. From the clitoris there are two small corpora
cavernosa clitoridis (o) passing craniad and then diverging
to be attached to the ischiatic rami. The ischiatic portion of[268]
each is covered by a muscle (ischiocavernosus). The clitoris
is said to contain a bone.
Mammary Glands.—The mammary glands secrete the
milk, and lie on the ventral surface of the body beneath the
integument. The separate glands are closely gathered into
two chief masses, one on each side the ventral middle line.
Each of these extends from about the region of the fourth rib
to the caudal end of the abdomen, ending over the pubic
symphysis. On each side the glands are gathered into five
groups, each of which is furnished with a nipple. The nipple
is a projection of the integument, having near its distal end
numerous fine openings for the ducts of the glands. The first
two nipples are on the thorax, the other three on the abdomen,
the most caudal ones being about two or three centimeters in
front of the cranial edge of the pubis.
Rudimentary mammary glands and nipples are present in
the male.
Muscles of the Urogenital Organs, Rectum, and
Anus
(Figs. 113 and 114).—The muscles connected with the
caudal openings of the alimentary canal and of the urogenital
organs are closely interrelated, a single muscle sometimes
acting on parts of both systems. For this reason all these
muscles are described together.
The region lying between the anus and the external opening
of the urogenital organs is known as the perineum. The
perineum is formed chiefly by muscles and fascia.
a. Muscles common to the Male and Female.
—M. sphincter
ani externus (Fig. 113, i; Fig. 114, a).—This muscle is confounded
with the levator scroti (Fig. 113, j) or the levator
vulvæ (Fig. 114, b). The two take origin in common from the
integument on the dorsum of the root of the tail dorsad of the
fifth caudal vertebra. There the fibres from the opposite sides
are intermingled. The common muscle passes ventrad about
the tail as a flat band close to the integument. Dorsad of the
anus the inner fibres of the muscles of the opposite sides are
united. They then separate and surround the anus as a band
five millimeters wide situated beneath the integument. Ventrad
of the anus the fibres are again intermingled. Some fibres[269]
on each side then continue to the scrotum as the levator scroti
(Fig. 113, j) or to the vulva as the levator vulvæ (Fig. 114, b).
Fibres also pass onto the anal pouch and unite with the sphincter
ani internus, forming the constrictors of the anal pouch
(Strauss-Durckheim).
M. sphincter ani internus (Fig. 113, h; Fig. 114, c).—The
sphincter ani internus is a broad and thick band of striated
muscle-fibres which surrounds the rectum at the anus. Dorsad
the band is about two centimeters broad, while ventrad it is
less than one. In the ventral median line some of the fibres
pass craniad to help in forming the bulbocavernosus muscle.
The muscle surrounds the anal sac.
(The muscle here described under this name is that described
under the same name in the cat by Strauss-Durckheim and
Mivart; it corresponds, however, to a part of the sphincter ani
externus of the dog, as described by Ellenberger and Baum.)
M. levator ani (or pubiocaudalis) (Fig. 162, 11).—This
muscle lies in the pelvic cavity. Each muscle forms a nearly
vertical sheet, and between the two are the rectum and the
urethra.
Origin from the symphysis of the pelvis.
Insertion into the midventral line of the centra of the third,
fourth, and fifth caudal vertebræ, close to the muscle of the
opposite side. This muscle is frequently continuous with the
iliocaudalis (Fig. 162, 11′).
Action.—Bends the tail and compresses the rectum.
M. ischiocavernosus (Fig. 113, m; Fig. 114, e).—A small,
flat, spindle-shaped muscle which lies upon the crus of the penis
or clitoris. Each has
Origin from the caudal border of the ramus of the ischium,
about one centimeter from the median line.
Insertion, in the male, into the whole outer surface of the
crus penis, or bulb of the corpus cavernosum penis. In the
female the muscle is smaller than in the male, and the insertion
is into the ventral surface of the urogenital sinus, at the base
of the clitoris.
M. transversus perinei (Fig. 114, i).—A small bundle of
fibres which arises from the medial surface of the ischium, just[270]
dorsad of the origin of the ischiocavernosus, and passes mediad
to join the sphincter ani internus (c).
M. caudoanalis (S.-D.) (Fig. 113, f; Fig. 114, g).—A
slender, flat bundle of fibres having origin on the middle line of
the ventral surface of the second and third caudal vertebræ.
It passes caudoventrad, lying between the levator ani and the
caudorectal (Fig. 113, e; Fig. 114, j), and unites with the
ventral portion of the sphincter ani internus (Fig. 113, h).
Action.—Draws the anus craniodorsad.
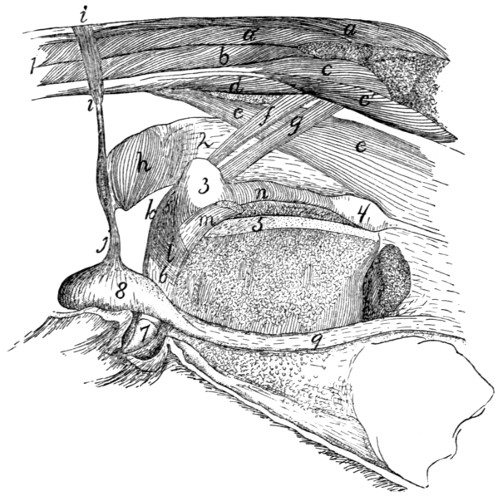
Fig. 113.—Muscles
of the Anus, Urogenital Organs, and Tail in the
Male (Slightly Schematic).
One side of the pelvis has been removed, a, a′, M. extensor caudæ lateralis; b,
M. abductor caudæ externus; c, c′, M. flexor caudæ longus; d, M. flexor caudæ
brevis; e, M. caudorectalis; f, M. caudoanalis; g, M. caudocavernosus; h, M.
sphincter ani internus; i, M. sphincter ani externus; j, M. levator scroti; k, M. rectocavernosus;
l, M. bulbocavernosus; m, M. ischiocavernosus (cut); n, M. compressor
urethræ membranaceæ. 1, tail; 2, rectum; 3, bulbourethral or Cowper’s gland; 4,
prostate gland; 5, symphysis pubis; 6, penis; 7, glans penis; 8, testis; 9, spermatic
cord.
M. caudorectalis (Fig. 113, e; Fig. 114, j).
Origin from the ventral surface of the sixth and seventh
caudal vertebræ. A small, at first unpaired band two or three
millimeters wide is formed, which passes cranioventrad, soon[271]
dividing into two lateral halves. These spread out over the
sides of the rectum, forming a broad sheet of fibres which pass
into the walls of the rectum, among the transverse fibres of the
latter. This muscle is covered by the caudoanalis (Fig.
113, f), caudocavernosus (g) (or caudovaginalis, Fig. 114, h),
iliocaudalis, and levator ani.
b. Muscles Peculiar to the Male
(Fig. 113).—M. levator
scroti (j).—This is a band of fibres which passes ventrad in the
median line from the sphincter ani externus (i) onto the
scrotum. Here it spreads out beneath the skin, forming
especially a well-marked bundle in the median furrow between
the two testes.
M. rectocavernosus, or retractor penis (k).—A small
bundle of fibres which arises in two parts from the ventral surface
of the sphincter ani internus (h). The two parts unite into
a single bundle which passes caudad on the middle line of the
ventral surface of the penis; it is inserted into the corpus cavernosum
just proximad of the glans. The muscle is covered
only by integument, and overlies the ischiocavernosus (m).
M. caudocavernosus (S.-D.) (g).—A slender bundle just
craniad of the caudoanalis (f).
Origin on the median ventral line of the first two caudal
vertebræ. The muscle passes caudoventrad, lying between the
levator ani and the caudorectal (e). It divides into two bands,
one of which is inserted into the base of the corpus cavernosum,
while the other extends farther caudad and is inserted at the
distal extremity of the corpus cavernosum.
Action.—Flexes the penis (bends it backward).
M. bulbocavernosus (accelerator urinæ) (l).—The two
muscles cover the ventral surface of the penis.
Origin of each from a median raphe, which passes from the
bulbous portion of the urethra toward the anus. The fibres
pass toward the distal end of the penis and have their
Insertion into the distal half of the lateral surface of the
corpus cavernosum penis.
M. compressor urethræ membranaceæ (n).—A thick layer
of striated muscle-fibres which surrounds the urethra between
Cowper’s gland (3) and the prostate
(4). The fibres have a[272]
circular course, and the cranial ones are attached to the crura
of the penis. The other fibres have no fixed attachment.
c. Muscles Peculiar to the Female
(Fig. 114).—M. levator
vulvæ (Strauss-Durckheim), or constrictor cunni (b).—This is
homologous with the levator scroti of the male. It consists of
a band of fibres which pass ventrad from the external sphincter
ani (a) and surround the vulva (3), lying immediately beneath
the integument.
M. constrictor vestibuli, or rectovaginalis (Strauss-Durckheim)
(d).
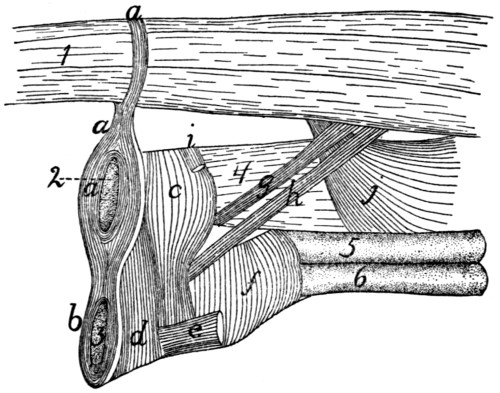
Fig. 114.—Muscles
of the Anus and Urogenital Organs in the Female.
a, M. sphincter ani externus; b, M. levator vulvæ; c, M. sphincter ani internus;
d, M. constrictor vestibuli; e, M. ischiocavernosus (cut); f, M. urethralis; g, M.
caudoanalis; h, M. caudovaginalis; i, M. transversus perinei; j, M. caudorectalis.
1, the tail; 2, anus; 3, vulva; 4, rectum; 5, vagina; 6, neck of the bladder.
Origin from the sides of the sphincter ani internus (c). The
muscle forms a distinct bundle two or three millimeters wide,
which passes ventrocaudad and is inserted into the ventral surface
of the urogenital sinus, caudad of the insertion of the
ischiocavernosus (e).
M. caudovaginalis (Strauss-Durckheim) (h).—A slender
band just craniad of the caudoanalis (g), and corresponding to
the caudocavernosus of the male.
Origin from the median line of the ventral surface of the
first two caudal vertebræ. The muscle passes caudoventrad,
lying between the levator ani and the caudorectal (j), and is
inserted into the ventral side of the urogenital sinus, at the base
of the clitoris.
[273]
M. urethralis (f).—This consists of fibres surrounding the
cranial part of the urogenital sinus and the caudal parts of the
vagina and neck of the bladder.
Origin partly on the caudal part of the symphysis of the
ischium, partly from the ventral surface of the urogenital sinus,
where the fibres are attached to the corpora cavernosa clitoridis.
The fibres pass dorsad over the surface of the sinus, and over
the surface of the union of the vagina and neck of the bladder,
to be inserted into the sides of the vagina and the dorsal surface
of the urogenital sinus.
[274]
THE CIRCULATORY SYSTEM.
I. THE HEART. COR.
The heart lies in the mediastinum, enclosed in the pericardial
sac, and projects rather more toward the left than toward
the right. It is an ovoid or pear-shaped organ, with its long
axis directed approximately craniocaudad. Its caudal end or
apex is, however, directed slightly ventrad and to the left,
while the larger cranial end or base faces slightly dorsad as
well as craniad.
Laterally and dorsally the heart is largely covered by the
lungs. The ventral side and a considerable portion of the
lateral surface are, however, not thus covered, so that they lie
against the thoracic wall. The heart extends from about the
fourth or the fifth to the eighth rib, and its apex touches the
diaphragm.
The cavity of the heart is divided by a longitudinal dorsoventral
septum into lateral halves—a right and a left side.
Each side is again divided by a transverse dorsoventral septum
(auriculoventricular) into two chambers, one of which, the
auricle, lies at the base; the other, the ventricle, lies at the
apex of the heart. There are thus right and left auricles and
right and left ventricles. An external groove, partly filled
with fat, separates the auricular portion of the heart from the
ventricular part; this groove is known as the sulcus coronarius.
Each auricle or ventricle has a single set of blood-vessels
either leaving it or entering it. Thus the blood enters the
right auricle by the inferior and superior venæ cavæ (Fig.
116, d and e) (præcava and postcava). It passes thence into
the right ventricle, and from the right ventricle to the lungs by[275]
a single pulmonary artery (Fig. 115, f). It returns to the
left auricle by the numerous pulmonary veins (Fig. 116, g,
h, i), and passes thence to the left ventricle. From the left
ventricle it passes to the body by a single aorta (Fig. 115, g).
When the heart is viewed from the ventral surface (Fig.
115), a considerable blood-vessel, the coronary artery (q),
running from the base to a point a little to the right of the
apex, indicates the position of the septum (ventricular septum),
between the right (a) and left (b) ventricles. The apex thus
belongs to the left ventricle. From the cranial end of the
right ventricle the pulmonary artery (f) is seen passing
obliquely craniad and toward the left. Beneath the pulmonary
artery the aorta (g) appears rising from the middle of the base
of the heart and passing directly craniad. At the base appears
a part of the auricular appendage of the left auricle (d), and at
the right a part of the right auricular appendage (c). The
former (d) is larger and bent at right angles, so that its free end
is directed toward the apex.
In the dorsal view (Fig. 116) the position of the ventricular
septum is not indicated. On the surface of the left ventricle a
short distance from the ventricular septum and nearly parallel
to it are seen branches of the coronary artery and the coronary
vein passing toward the apex. Craniad of the left ventricle is
seen the left auricle (b) with the pulmonary veins (g, h, i)
opening into it. Craniad of the right ventricle (a) is the right
auricle (c) with the venæ cavæ (d and e) opening into it near
the middle. Coming from beneath the auricles are seen the
pulmonary artery (j) and the aorta (f).
The chambers of the heart.
1. The right auricle (atrium dextrum) (Figs. 115 and
116, c). Externally the right end of the right auricle projects
so as to form its auricular appendage (Fig. 115, c) which lies
at the right of the base of the aorta (Fig. 115, g). The
superior vena cava (Fig. 116, e) is seen entering near the
cranial end of the auricular septum opposite the base of the
aorta. The inferior vena cava (Fig. 116, d) enters the auricle
near the coronary sinus and close to the auricular septum.
The netted appearance which the wall, especially that of the[276]
appendage, presents externally is due to the muscular thickenings
of the wall (musculi pectinati).
Fig. 115.—a, right ventricle;
b, left ventricle; c, right auricular appendage; d,
left auricular appendage; e, conus arteriosus; f, pulmonary artery; g, aortic arch;
h, thoracic aorta; i, vena cava superior; j, innominate artery; k, left subclavian
artery; l, left common carotid; m, right common carotid; n, right subclavian; o,
azygos vein; p, the two innominate veins; q, coronary artery.
Fig. 116.—a, right ventricle; b,
left auricle; c, right auricle; d, vena cava inferior;
e, vena cava superior; f, aorta; g, h, i, groups of pulmonary veins (g, dorsal
group; h, sinistral group; i, dextral group); j, pulmonary artery (division into two);
k, innominate artery; l, left subclavian; m, left common carotid; n, right common
carotid; o, right subclavian; p, azygos vein.
The cavity (including that of the auricular appendage) is
somewhat egg-shaped, with its long axis transverse. The[277]
musculi pectinati, which branch and unite into a network, are
most abundant on its dorsal wall at the right. The axes of
the two venæ cavæ if produced into the cavity of the auricle
would meet one another at its centre and nearly at right
angles. Caudad of the opening of the inferior vena cava is
seen the slit-like opening of the coronary sinus, guarded
craniad by the semilunar valve of the coronary sinus (or valve
of Thebesius). The coronary sinus receives blood from
coronary veins, which collect it from the walls of the heart.
In the auricular septum ventrad of the opening of the
inferior vena cava is seen a faintly marked smooth oval depression,
the fossa ovalis. When the auricular septum is examined
by transmitted light it is seen to be thinner over the fossa
ovalis. There is an opening, foramen ovale, at this point in
fœtal life, so that the blood of the inferior vena cava then
passes directly from the right auricle to the left auricle. Caudad
the cavity of the right auricle communicates with that of
the ventricle by the large oval auriculoventricular opening,
which is guarded by the tricuspid valves (Fig. 117).
2. The right ventricle (ventriculus dexter) (Figs. 115 and
116, a) does not reach quite to the apex of the heart, so that it
makes up less than one-half of its ventricular portion. It
makes a half-spiral turn about the left ventricle, from its lateral
side at the apex toward its ventral side at the base, where it
ends in the pulmonary artery (Fig. 115, e). Its wall is very
thin as compared with that of the left ventricle. Its cavity
(Fig. 117) is nearly flat on its medial side, convex on its lateral
side. It communicates with the auricle craniodorsad. Cranioventrad
the cavity is narrower and turns around toward the
ventral side of the heart and opens into the pulmonary artery.
That portion of the ventricle between the auriculoventricular
opening and the pulmonary artery is the conus arteriosus (Fig.
115, e; Fig. 117, f). Internally the wall presents many
muscular trabeculæ (Fig. 117, a) of various sizes. These are
more numerous over the ventricular wall than on the septum.
They do not occur on the conus (f). Surrounding the auriculoventricular
opening is the tricuspid valve (Fig. 117, d,
d′, d″). It consists of three flaps. One of these
(d′) is septal[278]
(i.e., it lies against the septum), while of the two others which
do not lie against the septum, one is dorsal (d″) and the other
ventral. Each flap is thin, semicircular, and membranous, and
is attached to the border of the auriculoventricular opening by
the diameter of the semicircle. The free border of each is
attached to the wall of the heart by numerous delicate tendinous
bands, the chordæ tendineæ (c), some of which are
attached to the lower face of each valve. The chordæ tendineæ
of the septal valve (or most of them) are attached at their
opposite ends to the septum directly, while those of the dorsal
and ventral flaps are attached to the ends of three or more
band-like muscles, columnæ (or trabeculæ) carneæ (b), which
are fixed by their opposite ends to the ventricular wall.
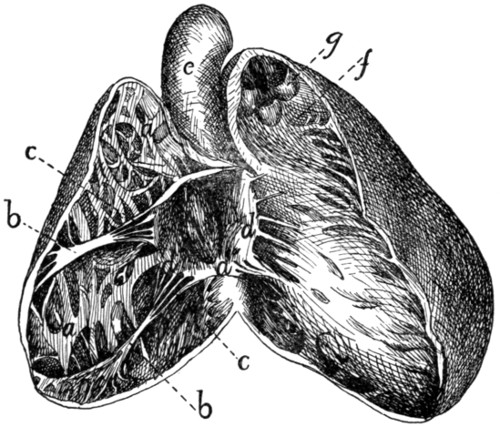
Fig. 117.—Heart,
with Right Ventricle laid open to show the Tricuspid
Valve.
a, trabeculæ; b, columnæ
carneæ; c, chordæ tendineæ; d, d′, d″, the three flaps
of the tricuspid valve; e, aorta; f, conus arteriosus, laid open; g, semilunar valves
of the pulmonary artery.
Between the conus arteriosus (f) and the pulmonary artery
are three pocket-like semilunar valves (Fig. 117, g), one ventral,
one dextral, and one sinistral. Between each valve and
the wall of the pulmonary artery there is an enlargement of the
cavity of the artery, one of the pulmonary sinuses (or sinuses
of Valsalva). Beyond the sinuses the pulmonary artery divides
into right and left branches (Fig. 116, j).
[279]
3. The left auricle (atrium sinistrum) (Fig. 115, d; Fig.
116, b). The auricular appendage (Fig. 115, d) is bent at
right angles and its apex turned caudad. The cavity is thus
irregular. Muscular trabeculæ occur only in the auricular
appendage. When the auricular septum is examined by transmitted
light the position of the fossa ovalis is indicated at about
the middle of the septum as seen from the left side. Ventrad
of it is a fold of the septal wall.
The pulmonary veins enter the dorsal wall of the auricle
in three groups (Fig. 116, g, h, i). Each group opens into a
more or less pronounced sinus or extension of the auricular
cavity. The sinuses may be called, on account of their position,
dorsal (g), sinistral (i), and dextral (h).
4. The left ventricle (Fig. 115, b) occupies rather more
than the left half of the base of the heart. Its walls are two
or three times as thick as those of the right ventricle. Internally
its walls present a few muscular bands, comparable to
the trabeculæ of the left ventricle. There are two very large
columnæ carneæ, one dorsad and one ventrad. At the cranial
end it communicates near the lateral wall with the left auricle
by the auriculoventricular opening, and near the septum with
the aorta. The bicuspid valve (or mitral valve), which guards
the auriculoventricular opening, consists of two flaps, one septal
and one lateral. Their chordæ tendineæ, which come from their
free borders as well as from their outer surfaces, are attached
to the columnæ carneæ, chordæ passing from both valves to
each columna carnea.
The opening into the aorta is guarded by three aortic
semilunar valves, each of which partly conceals an aortic
sinus (or sinus of Valsalva). One valve is dorsal, one sinistral,
and one dextral. In the dextral sinus is the opening of
one of the coronary arteries, which carry blood to the walls of
the heart.
Pericardium.
The pericardium is a sac enclosing the heart. It lies in
the middle mediastinum, and the two halves of the mediastinal
septum with their fat may be dissected away from it. Its wall
is composed of two layers, an external firm fibrous layer and[280]
an internal layer of flattened epithelial cells similar to the
peritoneal epithelium (serous layer). The fibrous layer forms
a sac which repeats roughly the form of the heart. This sac
is attached to the aorta at the point of origin of the subclavian
artery, to the pulmonary artery at its bifurcation, and to the
venæ cavæ and pulmonary veins near their entrance into the
heart. At these points it is continuous with the fibrous coats
of the vessels named, and from them it is reflected over the
heart, forming a complete sac enclosing it but not attached to
it anywhere. The heart lies within this sac. The serous layer
lines the fibrous sac and gives to the surface of the heart and
fibrous layer a smooth glistening appearance. It is reflected
over the heart. The relation of the heart to it is much the
same as the relation of the intestine to the peritoneal sac.
The serous layer consists therefore of two portions, parietal,
lining the sac, and visceral, covering the heart. The parietal
and visceral portions are continuous along a line which runs
approximately parallel to the auriculoventricular groove and
encloses all the great blood-vessels. Within this line the heart
lies against the fibrous layer of pericardium and is not covered
by the serous layer. The serous layer is easily dissected free
from the heart-wall, but its parietal portion is closely adherent
to the fibrous layer.
II. THE ARTERIES. ARTERIÆ.
1. A. pulmonalis, the Pulmonary Artery
(Fig. 115, f).
The pulmonary artery passes craniodorsad and slightly to
the left from the cranial end of the conus arteriosus. One to
one and a half centimeters from the conus it divides into right
and left branches (Fig. 116, j). Just before the division the
dorsal surface of the pulmonary artery is connected by the short
ligamentum arteriosum, or ligamentum Botalli, with the aorta.
This is the remnant of a canal which in fœtal life forms a free
communication between the pulmonary artery and the aorta;
this canal is known as the ductus Botalli. The ligament is
almost or quite obliterated in the adult cat.
The left branch of the pulmonary artery passes to the left
lung, crossing ventrad of the thoracic aorta. It then divides[281]
into branches which pass to the lobes of the left lung. The
point of division of the left branch of the pulmonary artery lies
craniad of all the lobes of the lung, so that the lobes of the left
lung are said to be all hyparterial, i.e., below (or caudad
of) the artery.
The right branch passes under (dorsocaudad of) the aortic
arch, and reaches the right lung at about the junction of the
cranial lobe with the remainder of the lung. The cranial lobe
of the right lung is therefore said to be eparterial, since it is
craniad of the pulmonary artery; the other lobes are hyparterial.
The right branch divides at its entrance to the lung
and is distributed to its lobes.
2. Aorta
(Fig. 115, g; Fig. 118, a).
The aorta is the single great vessel which conveys blood
from the left ventricle. It makes a sharp semicircular curve
dorsad (Fig. 118, a′) and to the left, passes caudad at the left
side of the vertebral column, and passes between the crura of
the diaphragm to reach the abdominal cavity. It is divisible
into thoracic aorta (Fig. 118) and abdominal aorta (Fig. 126).
A. Thoracic Aorta
(Fig. 118).—The first portion of the
thoracic aorta, curved as above described, is the aortic
arch (a′). It lies in the thoracic cavity opposite the interval
between the third and fourth or fourth and fifth ribs. It is
separated from the vertebral column on the right by the
superior vena cava (q), and on the left by the œsophagus (4).
At its beginning it lies a little to the right of the median plane;
but it passes at once to the left side of the vertebral column.
Branches of the Thoracic Aorta.
1. Aa. coronariæ.—The coronary arteries are two. They
arise from the aortic sinuses. The left one (Fig. 118, p) leaves
the aorta on the left side, passes dorsad of the pulmonary artery,
and divides into two branches, one of which follows the auriculoventricular
groove (sulcus coronarius) to the dorsal side of the
heart and sends branches to the adjacent heart-walls, while the
other runs onto the ventricles, following approximately the
ventral border of the ventricular septum. The right coronary
artery passes in the auriculoventricular groove toward the right
and dorsad, and supplies the adjacent walls of the heart.
[282]
2. A. anonyma (Fig. 115, j;
Fig. 118, b).—The innominate
artery passes craniad from the convexity of the aortic arch (a′).
It gives rise first to a small mediastinal artery (m) which
passes ventrad into the mediastinum, then to the left common
carotid (Fig. 115, l), then to the right common carotid (Fig.
115, m). Sometimes the carotids are given off from a common
trunk. Beyond the right common carotid the innominate is
continued as the right subclavian (Fig. 115, n).
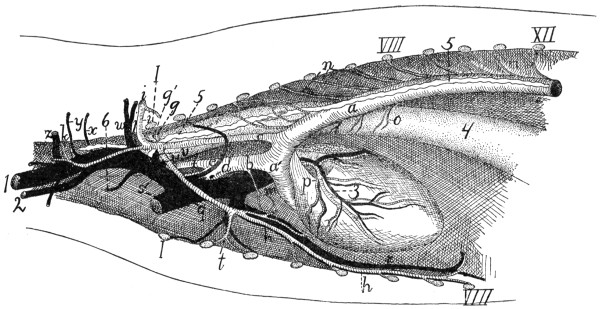
Fig. 118.—Vessels
of the Thorax, Viewed from the Left Side.
a, aorta (a′, aortic arch); b,
innominate artery; c, left subclavian; d, right subclavian; e, right common
carotid; f, left common carotid; g, costocervical axis; g′,
vertebral artery; h, internal mammary artery; i,
axillary artery; j, thyrocervical axis; k, A. transversa scapulæ;
m, mediastinal artery; n, intercostal arteries;
o, œsophageal arteries; p, left coronary artery; q,
superior vena cava; r, internal mammary vein; s, the two
innominate veins; t, sternal artery; u, costocervical vein;
v, vertebral vein; w, axillary vein; x, vein to clavotrapezius
and lateral ends of the pectoral muscles; y, vein accompanying second branch of A. transversa
scapulæ; z, thyrocervical vein. 1, external jugular vein; 2, internal jugular vein; 3, vena cordis magna; 4,
œsophagus; 5, thoracic duct; 6, one of the lymphatic ducts from the neck. I, first rib (cut); VIII, eighth
rib (cut); XII, twelfth rib (cut).
[283]
3. A. subclavia sinistra (Fig. 115, k; Fig. 118, c).—The
left subclavian passes craniad from the convexity of the aortic
arch. It curves about the first rib (Fig. 118, I) to enter the
armpit, where it becomes the axillary artery (i).
4. Aa. intercostales (n).—The intercostal arteries are given
off from the dorsal side of the aorta. Each passes to an intercostal
space and divides into three branches. One of these
passes ventrad along the caudal margin of the cranial one of
the two ribs between which it extends. Another goes to
the deep muscles of the back, and the third enters the spinal
canal through the intervertebral foramen. The arteries for the
first and second (and sometimes the third) intercostal space
usually arise from the subclavian (costocervical axis, Fig.
118, g).
5. Aa. bronchiales.—The bronchial arteries are two, and
arise either from the aorta opposite the fourth intercostal space
or from the fourth intercostal arteries. They accompany the
bronchi to the lungs.
6. Aa. œsophageæ (Fig. 118, o).—The œsophageal arteries
are small branches of varying origin passing to the œsophagus.
7. Aa. lumbales.—The lumbar arteries correspond to the
intercostals, but pass off between the lumbar vertebræ. The
obliquity of the diaphragm throws the origin of one or two
pairs of them within the thorax.
A. carotis communis. The Common Carotid Artery.
(Fig. 115, l and m; Fig. 119, a).
The two common carotid arteries arise from the innominate
artery in the manner already described. Each
passes craniad along the side of the trachea. In the thorax
(Fig. 118, f) the common carotid lies mediad of the subclavian
artery (Fig. 118, c) and dorsad of the superior vena
cava (Fig. 118, q). In the neck (Fig. 119) the artery lies,
accompanied by the vagus and sympathetic nerves and the
internal jugular vein (b), in the space between the longus
capitis muscle (7) and the trachea (14); it is covered ventrally
by the sternomastoid and sternothyroid (2) muscles, lying close
to the lateral border of the latter. Near its origin the common
carotid may give rise to the small inferior thyroid artery. It[284]
then passes to about the level of the larynx without giving off
branches; here it gives off the superior thyroid (c) on the
ventral side and one or more muscular branches (e) on the
dorsal side. One or two centimeters further craniad it gives
off on the dorsal side the internal carotid (g) and the occipital
artery (f). The main artery now takes the name external
carotid (m).
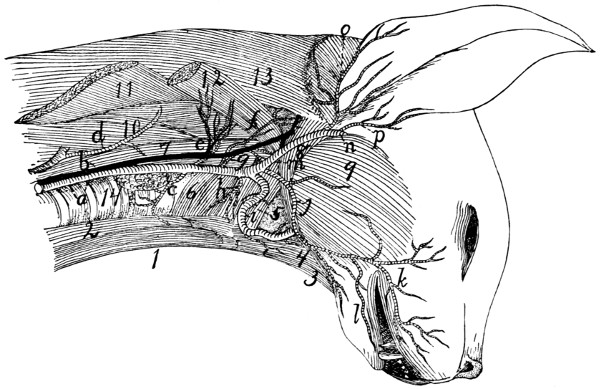
Fig. 119.—Common
Carotid Artery and Internal Jugular Vein.
a, common carotid artery; b, internal jugular vein; c, superior thyroid artery; d,
cervicalis ascendens artery; e, large muscular branches; f, occipital artery; g, internal
carotid; h, branch to larynx; i, lingual artery; j, external maxillary; k, superior
labial; l, inferior labial; m, external carotid; n, internal maxillary; o, posterior
auricular; p, superficial temporal. 1, M. sternohyoideus; 2, M. sternothyreoideus;
3, M. geniohyoideus; 4. M genioglossus; 5, M. constrictor pharyngis medius; 6, M.
constrictor pharyngis inferior; 7, M. longus capitis; 8, M. digastricus; 9, M. masseter;
10, M. scalenus; 11, M. levator scapulæ (cut); 12, M. levator scapulæ ventralis
(cut); 13, M. splenius; 14, trachea.
Branches of the common carotid:
1. A. thyreoidea ima.—The inferior thyroid is a small
artery which arises either from the common carotid near its
origin, or from the innominate before the origin of the carotid.
It passes craniad on the trachea as far as the thyroid gland,
giving branches to the trachea and œsophagus.
2. A. thyreoidea superior (c).—The superior thyroid leaves
the carotid opposite the thyroid cartilage and passes mediad
and caudad, sending branches to the thyroid gland, and the[285]
sternothyroid (2) and sternohyoid (1) muscles. A small
branch, the superior laryngeal, passes to the larynx, and
supplies those muscles of the larynx which are not enclosed
by the cartilages.
3. Rami musculares (e).—One or two branches, usually of
considerable size, leave the common carotid at about the same
level as the superior thyroid and pass to the muscles on the
dorsal side of the neck, the main trunk of the artery passing
between the longus capitis (7) and scalenus muscles (10).
4. A. occipitalis (f).—The occipital artery arises from the
common carotid at about the same point as the internal carotid.
It immediately sends a large branch dorsad, passing between
M. longus capitis (7) and the vertebral column, to the deep
muscles of the neck. The occipital then crosses the outer
surface of the digastric muscle (8) to the back of the skull, and
runs along the lambdoidal crest just beneath the splenius
muscle. It sends a number of branches to the muscles of the
back of the neck; and one of its branches may unite with the
vertebral artery as it lies in the groove on the atlas, or with a
branch of the vertebral.
5. A. carotis interna (g).—The internal carotid artery is
one of the terminal branches of the common carotid. It is very
small. It is given off near or in common with the occipital
artery, passes toward the cranial end of the tympanic bulla,
enters the bulla with the Eustachian tube, and passes into the
skull at the foramen lacerum. Its course is much convoluted
before entering the foramen. Within the skull (Fig. 121, g)
it joins the posterior cerebral artery (Fig. 121, f) at the side
of the hypophysis.
6. A. carotis externa (Fig. 119, m).—After giving off the
internal carotid the continuation of the common carotid artery
receives the name external carotid (m). It passes craniad and
laterad between the digastric (8) and styloglossus muscles,
where it gives off cranioventrad the lingual artery (i) and a
number of small muscular branches; also sometimes the small
laryngeal artery. At the dorsolateral border of the digastric
(8) it gives off the external maxillary artery (j), and about one
centimeter farther craniad the posterior auricular (o). It now[286]
turns mediad, lying against the cartilaginous auditory meatus,
on its cranial side, and gives off the superficial temporal (p).
The artery then continues mediad, taking the name internal
maxillary (n)—so that the internal maxillary artery is to be
considered the terminal branch of the external carotid.
Branches of the external carotid artery (Fig. 119):
a. A. lingualis (Fig. 119, i; Fig. 120, d).—The lingual
artery leaves the external carotid near its beginning and passes
craniomediad along the ventral border of the digastric muscle
(Fig. 120, 9), accompanied by the hypoglossal nerve. It gives
off numerous small branches to the hyoid and pharyngeal
muscles, then passes dorsad of the hyoglossus muscle (Fig.
120, 6), where it gives off a branch which passes transversely
across the middle line to communicate with the artery of the
other side. Beneath the hyoglossus the artery turns craniad
and passes into the tongue. Here it runs along the medial
border of the styloglossus to the tip of the tongue, giving off
numerous branches into the substance of this organ.
b. Rami musculares.—Muscular branches pass to the
digastric and to the hyoid muscles. A small branch (Fig. 119,
h), which may arise either from the external carotid or from
the common carotid near the beginning of the external carotid,
passes to the larynx and supplies the thyroarytenoid and lateral
cricoarytenoid muscles of the larynx.
c. A. maxillaris externa (Fig. 119, j).—The external
maxillary artery leaves the external carotid (m) opposite the
angle of the jaw and at about the dorsal border of the digastric
muscle (8). It passes craniad, lying at first beneath the
digastric muscle and sending a branch to the submaxillary
gland. Opposite the caudal border of the mylohyoid muscle
it gives off the submental artery, turns dorsad, emerges from
beneath the digastric, and passes along the cranial border of the
masseter onto the face. Here it divides into superior (k) and
inferior (l) labial branches, which pass along the upper and
lower lips, respectively, giving off numerous branches.
The submental artery passes to the symphysis menti
between the digastric and mylohyoid muscles, giving off on its
course collateral branches to the muscles of this region.
[287]
d. A. auricularis posterior (Fig. 119, o).—The posterior
auricular leaves the external carotid (m) opposite the middle of
the bulla tympani and beneath the submaxillary gland. It
passes about the base of the ear on its caudal and dorsal sides
outside of the deep muscles of the occipital region, but beneath
the auricular muscles, and sends several branches to the muscles
of the external ear, passing onto the caudal surface of the
concha. It sends also a large branch mediad to the muscles
of the occiput, especially to the temporal muscle, within which
it ramifies. A large branch (anterior auricular, Fig. 131, u)
passes from the caudal side of the concha craniodorsad, and
appears on the cranial side of the external ear, running along
the cranial margin of the auditory opening.
e. A. temporalis superficialis (Fig. 119,
p; Fig. 120, h).—The
superficial temporal artery arises from the external
carotid as the latter lies between the cartilaginous auditory
meatus and the caudal border of the masseter muscle. It
passes dorsad and gives off soon after its origin a muscular
branch to the masseter, and an auricular branch which passes
distad along the concha auris and ramifies over its cranial
surface. The superficial temporal itself passes onto the surface
of the temporal muscle (Fig. 120, 11), to which it gives
numerous branches. It extends to the caudal angle of the eye
(Fig. 131, s), where it divides. One branch passes into the
lower eyelid; the larger branch passes along the dorsal side of
the eye, sending a branch into the orbit and small branches
onto the dorsal surface of the nose.
f. A. maxillaris interna (Fig. 119, n; Fig. 120, i).—The
internal maxillary artery is the continuation of the external
carotid. It turns caudad at the caudal end of the mandible,
then passes craniad, lying dorsad of the pterygoid muscles
(Fig. 120, 10), and against the medial surface of the mandible.
It gives off the inferior alveolar artery (Fig. 120, j), then the
middle meningeal (k), and then continuing mediad divides into
three or four branches. The branches redivide, and the twigs
form a complicated plexus, the carotid plexus (Fig. 120, l),
which surrounds the maxillary division of the fifth nerve near
its exit from the foramen rotundum. One of the larger[288]
branches of the plexus enters the skull through the orbital
fissure, lying beside the hypophysis; it divides in the manner
described below.
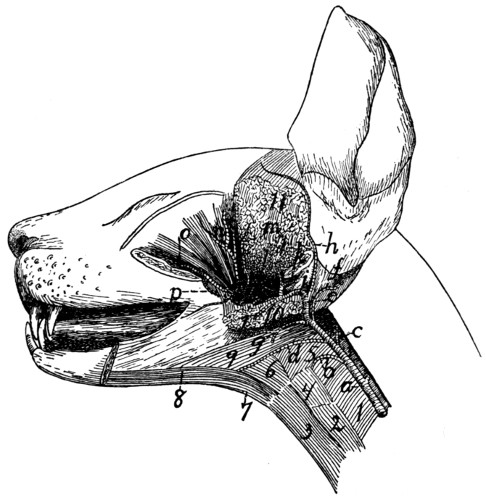
Fig. 120.—Branches
of External Carotid Artery.
a, common carotid; b, branch to larynx; c,
internal carotid; d, lingual; e, external
carotid; f, posterior auricular; g, external maxillary; h, superficial temporal;
i, internal maxillary; j, inferior alveolar; k, middle meningeal; l, carotid plexus; m,
branch to temporal muscle; n, ophthalmic; o, infraorbital; p, lesser palatine. 1, M.
constrictor pharyngis inferior; 2, M. sternothyreoideus; 3, M. sternohyoideus; 4, M.
thyreohyoideus; 5, M. constrictor pharyngis medius; 6, M. hyoglossus; 7, M. geniohyoideus;
8, M. genioglossus; 9, M. digastricus; 10, Mm. pterygoidei externus and
internus (cut); 11, M. temporalis (cut).
Beyond the carotid plexus a main trunk which may be considered
the continuation of the internal maxillary passes craniad,
lying on the dorsal surface of the external pterygoid muscle;
nearly opposite the molar tooth it divides into the infraorbital
(Fig. 120, o) and the sphenopalatine arteries.
Branches of the internal maxillary artery and of the carotid
plexus:
1. A. alveolaris inferior (Fig. 120, j).—The inferior alveolar
(or inferior dental) artery leaves the inferior maxillary just[289]
opposite the condyloid process of the mandible. It enters the
mandibular canal by the mandibular foramen, along with the
nerve of the same name, and traverses the canal, furnishing
branches to the lower teeth. It emerges at the mental foramen,
and its terminal branches are distributed to the chin, but a
branch is continued in the bone beyond the mental foramen
and supplies the incisor and canine teeth of the lower jaw.
2. A. meningea media (Fig. 120, k).—The middle meningeal
is a large vessel which leaves the internal maxillary at
about the same level as the inferior alveolar. It passes into
the foramen ovale and ramifies in the dura mater. Its branches
leave distinct impressions on the inner surface of the bones of
the skull.
3. A large branch (Fig. 121, h) from the plexus passes
into the cranial cavity through the orbital fissure and lies
within the skull at the side of the hypophysis. It gives off
the following branches:
a. A posterior communicating branch, very short, which
extends caudad and joins the internal carotid artery (Fig.
121, g).
b. A. cerebri media (Fig. 121, i).—The middle cerebral
artery passes dorsad on the side of the cerebral hemisphere
along the fissure of Sylvius and divides into numerous branches
which are distributed to the surface of the cerebrum.
c. A. cerebri anterior (Fig. 121, j) Passes dorsad between
the cerebral hemispheres. Just craniad of the optic chiasma
the two anterior cerebral arteries are united by a small communicating
branch, thus completing the circulus arteriosus or
circle of Willis (Fig. 121), surrounding the hypophysis (see
page 292).
4. From the carotid plexus several branches pass, arising
either separately or in common, to the masseter, temporal, and
pterygoid muscles.
5. A. ophthalmica (Fig. 120, n).—The ophthalmic artery
passes from the carotid plexus to the structures in the orbit.
It gives off numerous branches which supply the muscles of the
eyeball, and other structures of this region. It sends an
ethmoidal branch into the nasal cavity through the ethmoidal[290]
foramen in the orbital plate of the frontal bone, then continues
distad to emerge from the orbit on the medial side of the eye;
here it anastomoses with branches of the superficial temporal.
6. A. palatina minor (Fig. 120, p).—The lesser palatine
artery leaves the internal maxillary distad of the carotid plexus,
near the caudal border of the maxillary bone. It passes
ventrocaudad into the soft palate.
7. A. sphenopalatina.—The sphenopalatine is one of the
terminal branches of the internal maxillary; it passes mediad
through the sphenopalatine foramen into the nasal cavity, and
divides into numerous branches which supply the mucous
membrane of the nose. It gives off just before it enters the
sphenopalatine foramen the descending palatine (A. palatina
descendens), which passes into the posterior palatine canal and
emerges on the surface of the hard palate, where it ramifies.
8. A. infraorbitalis (Fig. 120, o).—The infraorbital artery
is a direct continuation craniad of the internal maxillary. It
sends off numerous small branches to the teeth of the upper
jaw, and a rather large branch which passes to the lower eyelid.
It then enters the infraorbital foramen, at the same time
dividing usually into two or three branches; these emerge from
the foramen and supply the parts of the nose and upper lip
adjacent to the foramen (Fig. 131, r).
A. subclavia. The Subclavian Artery.
The left subclavian (Fig. 118, c) arises from the convexity
of the aortic arch just distad of the origin of the innominate
artery, and about two or three centimeters from the heart. It
passes craniad and slightly to the left, and turns into the left
arm just craniad of the first rib.
The right subclavian (Fig. 115, n) is a direct continuation
of the innominate, the artery receiving the name subclavian
after the right common carotid is given off, usually at about the
level of the second or third intercostal space.
The subclavian has the following branches: the vertebral
artery (Fig. 118, g′), the internal mammary (Fig. 118, h),
the costocervical axis (Fig. 118, g), the thyrocervical axis
(Fig. 118, j). Beyond the last-named branch it continues into
the arm as the axillary artery (Fig. 118, i; Fig. 122, g).
[291]
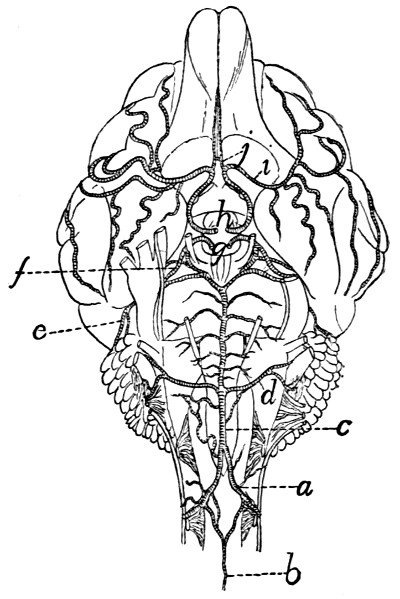
Fig. 121.—Arteries of the Ventral
Surface of the Brain.
a, A. vertebralis; b, A. spinalis anterior;
c, A. basilaris; d, A. cerebelli
inferior posterior; e, A. cerebelli anterior;
f, A. cerebri posterior; g, cut
ends of the two internal carotid arteries;
h, cut ends of branches from the carotid
plexus (g and h are placed within
the circulus arteriosus or circle of
Willis); i, A. cerebri media; j, A.
cerebri anterior.
a. A. vertebralis (Fig. 118, g′).—The vertebral artery
arises from the dorsal surface of the subclavian opposite the
first rib. It passes craniad and dorsad at the side of the
thoracic portion of the longus colli muscle, and enters the
foramen transversarium of the sixth cervical vertebra. It
passes thence craniad through the foramina transversaria (which
together form the vertebrarterial canal) and gives off at the
intervertebral foramina branches
to the muscles of the neck and
branches which pass across the
ventral surface of the spinal
cord to join A. spinalis anterior.
Craniad of the foramen transversarium
of the atlas the vertebral
artery turns dorsad in the groove
on the lateral surface of the atlas.
Here it gives off a large branch
which passes laterodorsad to the
muscles of the neck and may
anastomose with a branch of the
occipital artery. The vertebral
artery then passes into the vertebral
canal through the atlantal
foramen. It passes to the ventral
side of the spinal cord and unites
at about the level of the foramen
magnum with the vertebral artery
of the opposite side (Fig. 121, a)
to form the basilar artery (A.
basilaris) (Fig. 121, c), which
passes craniad along the ventral
middle line of the brain. Just before their union the two vertebral
arteries (a) give off each a branch which passes caudomediad.
These two branches soon unite in the middle line,
forming the anterior spinal artery (A. spinalis anterior) (b),
which passes caudad the entire length of the spinal cord, lying
on its ventral middle line and receiving many communicating
branches from the vertebral, intercostal, and lumbar arteries.
[292]
A. basilaris (c).—This arises by the union of the two
vertebral arteries in the manner just described. It passes
craniad along the ventral middle line of the medulla and pons,
giving numerous small branches to these structures. A large
branch, A. cerebelli inferior posterior (d), passes on each side
to the caudal surface of the cerebellum, on which it ramifies.
At the cranial margin of the pons the basilar artery divides.
From each division a very large branch passes on each side
across the pedunculi cerebri to the cranial part of the cerebellum;
this is A. cerebelli anterior (e). Just craniad of this,
arising from nearly the same point, the smaller A. cerebri
posterior (f) passes laterad to the caudal part of the cerebrum;
it is joined by the internal carotid artery (g). The small
continuations of the two halves of A. basilaris then pass
craniad at the side of the hypophysis to join the posterior communicating
branches from the carotid plexus. As other
branches of the carotid plexus unite across the middle line
craniad of the optic chiasma, an arterial circle is formed on the
base of the brain, surrounding the hypophysis and the optic
chiasma. This is known as the circulus arteriosus, or circle
of Willis (see page 289).
b. A. mammaria interna (Fig. 118, h).—The internal mammary
artery rises from the ventral surface of the subclavian
opposite the first rib, and passes in the mediastinum ventrocaudad
to reach the sternum opposite the third intercostal
space. It sends two or three small branches craniad to the
midventral part of the thoracic wall and then extends caudad
at the side of the sternum. It sends off lateral branches to the
ventral thoracic wall, branches to the mediastinum and pericardium,
a branch to the diaphragm which anastomoses with
the phrenic; and finally it passes out of the thoracic cavity
caudad of the last costal cartilage, extends caudad in a zigzag
course at the lateral border of the rectus muscle and anastomoses
with the inferior epigastric artery.
c. Truncus costocervicalis (Fig. 118, g).—The costocervical
axis arises from the subclavian opposite the first rib,
passes craniodorsad, and divides almost at once into two
branches. The smaller one of these, the superior intercostal[293]
(A. intercostalis suprema), passes caudad and, dividing, supplies
the first and second intercostal spaces, and then passes to
the deep muscles of the back. The other branch divides almost
immediately into two. One of these, A. transversa colli,
passes laterad in front of the first rib and enters the serratus
anterior muscle. In this it passes dorsad, giving off branches
to this muscle and to the levator scapulæ, till it reaches
M. rhomboideus, which it likewise supplies. The other branch
(A. cervicalis profunda), which seems to form a continuation
of the main artery, passes directly dorsad and leaves the
thoracic cavity between the heads of the first and second ribs.
Here it passes into the deep muscles of the neck; it can be
traced in the substance of the complexus muscle as far forward
as the atlas.
d. Truncus thyrocervicalis (Fig. 118, j).—The thyrocervical
axis (or thyroid axis) arises from the subclavian
beneath the first rib, a short distance distad of the origin of the
costocervical axis. It passes laterocraniad and dorsad, lying
on the mediocranial side of the brachial plexus. A short distance
from its origin it gives off a branch, varying much in
size, the cervicalis ascendens (Fig. 119, d), which passes
craniad on the ventral side of the neck, supplying the sternomastoid,
sternohyoid, the cervical portion of the scalenus, and
sometimes other muscles of the neck region. Other branches
pass from the thyroid axis to the inner surface of the clavotrapezius
(Fig. 122, a) and to the ventral end of the
pectoral muscles. At the level of the cranial border of the
scapula the artery takes the name A. transversa scapulæ, or
suprascapularis. This divides into three main branches. The
first branch, sometimes large, sometimes small, passes to the
lymphatic gland in the hollow of the shoulder, and to the
adjacent muscles; it supplies the clavotrapezius, acromiotrapezius,
levator scapulæ ventralis, splenius, occipitoscapularis,
and rhomboideus. A second branch passes between the subscapularis
and supraspinatus muscles, dividing into various
branches which supply the muscles named. The third branch
pierces the supraspinatus muscle, just craniad of the acromion
process, and ramifies in that muscle.
[294]
e. A. axillaris (Fig. 122, g).—The axillary artery is the
continuation of the subclavian laterad of the first rib. It lies
caudad of the brachial plexus and parallel to it. It sends off
the following branches:
1. A. thoracica anterior (Fig. 122, h).—The anterior
thoracic is a slender artery which leaves the ventral side of the
axillary opposite the first rib and passes caudomediad, to supply
the medial ends of the pectoral muscles (g).
2. A. thoracica longa (p).—The long thoracic artery is
larger than the preceding, leaves the axillary a short distance
laterad of it, and passes caudad to the middle portions of the
pectoral muscles (8) and continues to the inner surface of the
latissimus dorsi.
A short distance beyond the long thoracic the axillary
divides into two. The more cranial one of these is the subscapular
(l); the other is the brachial (g′).
3. A. subscapularis (l).—The subscapular artery passes
laterad and gives off a short distance from its origin the
A. thoracicodorsalis (t) and A. circumflexa humeri posterior
(see below); it may also give rise to the circumflexa anterior
humeri (l′), and to the profunda brachii (u); these two
branches, however, rise more frequently from the brachial
artery (g′) and are described in connection with it. The subscapular
artery then passes through the triangular interval
between the scapular end of the long head of the triceps, the
latissimus dorsi, and the glenoid border of the scapula. Within
this interval it sends muscular branches to the long head of the
triceps, the subscapularis, and the latissimus dorsi. That to
the latter muscle is very large. At the border of the scapula,
opposite the tuberosity of the spine, it turns craniad, passes
over the lateral surface of the infraspinatus muscle, supplying
it; crosses the spine, and sends branches into the supraspinatus
fossa in both directions parallel to the scapular spine. These
supply the supraspinatus, acromiotrapezius, and spinotrapezius,
and anastomose with the branches of the transversa scapulæ.
As the subscapularis turns craniad onto the surface of the infraspinatus
it sends dorsad a small branch, the circumflexa
scapulæ, which passes in the infraspinatus fossa close to its[295]
[296]
glenoid border as far as the glenovertebral angle, supplying
the infraspinatus and latissimus dorsi by lateral branches.
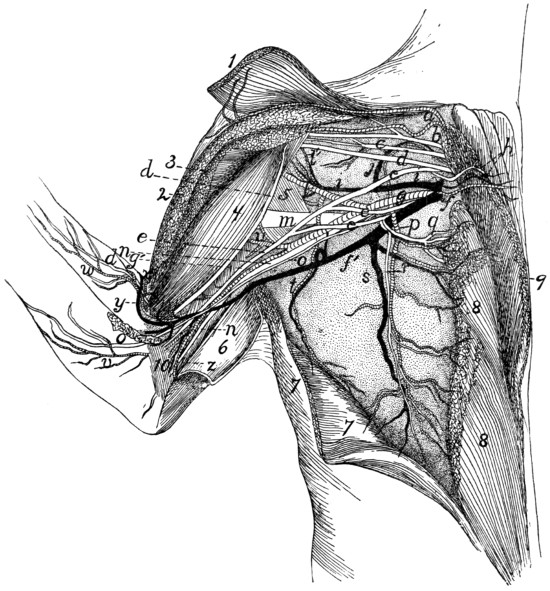
Fig. 122.—Blood-vessels
and the more Ventral Nerves of the Axilla,
Ventral View.
The pectoral and clavobrachial muscles have been cut and laid aside, their ends
being shown; only a part of the nerves are exhibited. 1, M. clavobrachialis; 2, cut
end of M. pectoralis major; 3, cut end of M. pectoralis minor; 4, M. biceps; 5, M.
teres major; 6, M. epitrochlearis, partly cut and turned back; 7, M. latissimus dorsi,
partly cut; 8, M. pectoralis minor; 9, M. pectoralis major; 10, short portion of caput
mediale of M. triceps brachii. a, branches of the thyrocervical axis to clavobrachial
and clavotrapezius muscles; b, suprascapular nerve; c, first subscapular nerve; d,
musculocutaneous nerve; e, median nerve; f, V. axillaris; f′, V. brachialis; g, A.
axillaris; g′, A. brachialis; h, first anterior thoracic nerve, accompanied by the anterior
thoracic artery and vein; i, V. subscapularis; j, large muscular branch (to
subscapular muscle) of the subscapular vein and brachial artery; l, A. subscapularis;
l′, A. circumflexa humeri anterior; m, radial nerve; n, ulnar nerve; o, medial
cutaneous nerve; p, A. thoracica longa; q, r, second anterior thoracic nerve; s, V.
longa thoracica; t, A. and V. thoracicodorsalis; u, A. profunda brachii; v, branch
of brachial artery accompanying medial cutaneous nerve; w, branch of A. collateralis
radialis superior; x, A. collateralis radialis superior; y, V. mediana cubiti; z, A. collateralis
ulnaris.
A. thoracicodorsalis (t).—This arises from the subscapular
a short distance from its origin and passes across the teres
major to the latissimus dorsi (7), giving branches to both these
muscles and to the epitrochlearis.
A. circumflexa humeri posterior.—The posterior circumflex
artery arises from the A. subscapularis close to the origin
of the latter. It passes between the subscapularis muscle and
the teres major, close to the border of the biceps, then between
the lateral and long heads of the triceps. It gives off a branch
to the inner surface of the spinodeltoid and acromiodeltoid,
then passes distad to supply the lateral and long heads of the
triceps.
4. A. brachialis (g′).—The brachial artery is the continuation
of the axillary (g) into the arm beyond the origin of
the subscapular (l). It passes along that side of the biceps
which lies next to the humerus. It passes thus, accompanied
by the brachial vein (f′) and median and ulnar nerves, through
the bicipital arch and afterwards between the biceps (4) and
the intermediate division of the medial head of the triceps to
the supracondyloid foramen of the humerus, through which
it passes with the median nerve to reach the concavity of the
elbow. It gives off the following branches:
a. A. circumflexa humeri anterior (l′).—The anterior circumflex
artery usually leaves the brachial near its origin (but
may arise from the subscapular (l) or one of its branches); it
passes to the biceps (4) near the origin of the latter and sends
a branch proximad to the head of the humerus.
b. A. profunda brachii or superior profunda (u).—This
arises from the first part of the brachial or it may come off from
one of the branches of the axillary (e.g., the subscapularis).
It passes along with the radial nerve onto the dorsal side of the
humerus and supplies the triceps muscle. It also sends
branches to the epitrochlearis (6) and latissimus dorsi (7).
c. Rami musculares.—Muscular branches are given off
near the supracondyloid foramen to the biceps (4), epitrochlearis
(6), and brachialis muscles. A nutrient artery leaves[297]
the brachial proximad of the supracondyloid foramen, either
separately or in common with the muscular branches, and
passes into the nutrient foramen at the junction of the middle
and distal thirds of the shaft of the humerus, supplying the
bone.
d. A. collateralis ulnaris (superior) (or A. anastomotica
magna) (z).—This leaves the brachial artery just proximad of
the supracondyloid foramen and passes to the convexity of the
elbow, supplying the structures about the olecranon.
e. A. collateralis radialis superior (Fig. 130).—This rises
from the axillary artery just proximad of the supracondyloid
foramen (Fig. 122, x), in company with the vena mediana cubiti
(y), passes across the surface of the biceps (4), beneath the
pectoantibrachialis, into the concavity of the elbow. Here it
gives branches to the pectoantibrachialis, clavobrachialis, and
extensor muscles of the forearm. It then passes onto the ventroradial
border of the forearm (Fig. 130) and runs along this
border, in company with the vena cephalica (Fig. 130, c) and
the superficial radial nerve (g), to the wrist, sending off
branches to the integument. At the wrist it turns onto the
dorsum of the hand, passing in a gentle curve to the ulnar side
and distad, and giving off a branch for the space between each
pair of metacarpal bones. These branches (Fig. 130, e)
(Aa. digitales dorsales) pass distad and anastomose with
branches coming from the palm.
f. A. radialis (Fig. 123).—Distad of the convexity of the
elbow the brachial artery takes the name radial artery. It
passes from the supracondyloid foramen on the medial side of
the biceps tendon (1) and beneath the pronator teres muscle
(5), giving off small branches, as far as the middle of the forearm,
where it gives off the ulnar artery (k). It then passes
from beneath the pronator teres (5) and lies on the surface of
the fifth part of the flexor profundus (8), covered only by the
fascia and integument. Near the wrist it sends off a branch
(m) toward the ulnar side of the arm, which passes into the
palm on the surface of the tendon of the flexor profundus,
giving twigs to the digits and to the pad in the palm. A
branch from this may join the ulnar, and the common trunk[298]
thus formed passes into the hand and gives origin to the
branches described under the ulnar artery. The presence and
size of this branch of the radial varies with the size of the ulnar
artery, it being smaller or absent as the ulnar is larger. It is
shown at m, Fig. 123. The radial artery (g) now turns
gradually dorsad and passes beneath the tendon of the extensor
brevis pollicis onto the dorsum of the hand. It passes over the
oblique groove on the dorsal surface of the base of the second
metacarpal beneath the tendon of the extensor carpi radialis
longus and passes between the bases of the second and third
metacarpals into the palm of the hand. Here it passes to the
ulnar side and anastomoses with the ulnar artery to form the
palmar arch.
Branches of the Radial Artery.
1. A. radialis recurrens (i).—The radial recurrent arises
in the concavity of the elbow and sends a branch proximad to
the structures in the concavity; it then continues to the radial
side of the forearm at the elbow, supplying adjacent parts of
the brachialis (2), the extensor carpi radialis, and the extensor
communis digitorum.
2. A. ulnaris recurrens (j).—The ulnar recurrent arises
on the medial side of the tendon of the biceps and supplies
structures in the concavity of the elbow on the ulnar side, also
the pronator teres (5) and proximal ends of the flexor carpi
radialis and flexor profundis digitorum.
3. Rami musculares.—Muscular branches are given off
along the course of the artery, to adjacent muscles. A nutrient
artery to the radius leaves the radial artery about two centimeters
distad of the supracondyloid foramen.
4. Aa interosseæ (anterior (l) and posterior) are given off
usually separately between the origin of the ulnar artery and
the biceps tendon. Sometimes they arise as a common trunk
which soon divides. The posterior interosseous artery passes
distad, supplying the flexor muscles of the forearm. It also
sends a branch to the pronator teres. The anterior interosseous
(l) may arise from the ulnar artery. It passes dorsad to
the interosseous membrane, on which it runs distad; sends a
nutrient branch to the ulna, and then passes to the wrist in the[299]
[300]
substance of the pronator quadratus, which it supplies. It
sends a branch to the dorsal surface of the wrist; this ramifies
on the carpus, forming a network of small arteries.
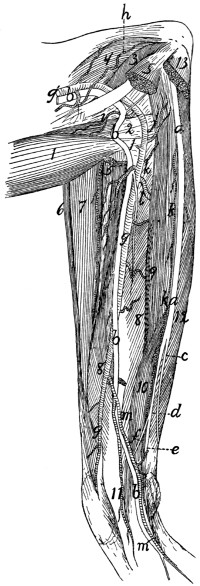
Fig. 123.—Nerves
and Arteries of the Forearm, seen from the
Flexor Side.
Mm. palmaris longus, flexor carpi radialis, flexor carpi ulnaris, and the third and
fourth heads of flexor profundus digitorum have been removed; also part of M. pronator
teres. 1, biceps; 2, conjoined tendon of brachialis and clavobrachialis; 3, short
portion of caput mediale of triceps brachii; 4, intermediate portion of caput mediale
of triceps brachii; 5, cut ends of pronator teres; 6, brachioradialis; 7, extensor carpi
radialis longus and brevis; 8, fifth head of flexor profundus digitorum; 9, second
head of flexor profundus digitorum; 10, radial part of flexor sublimis digitorum; 11,
common tendon of flexor profundus digitorum; 12, first head of flexor profundus
digitorum; 13, cut origin of flexor carpi ulnaris. a, ulnar nerve; b, median nerve;
c, dorsal cutaneous branch of the ulnar nerve; d, palmar branch of ulnar nerve; e,
deep palmar branch; f, superficial palmar branch; g′, A. brachialis; g, A. radialis;
h, A. collateralis ulnaris superior; i, A. radialis recurrens; j, A. ulnaris recurrens;
k, A. ulnaris; l, A. interossea anterior; m, large branch of A. radialis, joining A.
ulnaris.
5. A. ulnaris (k).—The ulnar artery passes beneath the
second, third, and fourth parts of the flexor profundus digitorum
(but outside of the origin of the fifth part), to the inner surface
of the flexor carpi ulnaris. It supplies the flexor carpi ulnaris,
the flexor profundus and palmaris longus, and passes on the
inner surface of the flexor carpi ulnaris to the wrist. Near the
wrist it sends a branch onto the side of the forearm, and
another to its midventral part, and ends in a small branch
to the wrist on the radial side of the pisiform bone. This
branch anastomoses with the radial to form the palmar arch,
described below.
6. The palmar arch (Fig. 124) is formed by the termination
of the radial artery (a) in the palm and its junction with
the end of the ulnar (f). The radial artery reaches the palm
between the bases of the second and third metacarpals and
passes thence toward the ulnar side and distad, piercing the
interosseus muscle of the third digit and lying on the outer
surface of the interossei of the third and fourth digits beneath
the adductors of the second and fifth digits. A small communicating
branch from the radial passes to it between the first
and second metacarpals.
Branches of the palmar arch:
A. princeps pollicis et indicis (c) leaves the palmar arch
near its radial end and sends a branch onto the ulnar side of
the thumb and one onto the radial side of the index.
The palmar interosseæ (d) are three in number. They
leave the palmar arch (b) and pass distad and dorsad in the
intervals between the four ulnar digits. The radial one passes
along the ulnar side of the first digit, the ulnar one along the
radial side of the fifth digit. The middle one divides and supplies
the contiguous sides of the third and fourth digits. Each
of these interosseous arteries sends off muscular branches (e) to
the short muscles in the palm, and branches into the fibrous
pad which occupies the palm of the hand. The palmar arch[301]
also sends branches onto the dorsum of the hand at the sides
of the fourth metacarpal. Branches pass from these proximad
to the wrist.
B. Aorta abdominalis. The Abdominal Aorta.
—The
abdominal aorta emerges into the abdomen from between
the crura of the diaphragm, at about the
level of the second lumbar vertebra. It
passes caudad along the dorsal middle
line, lying to the left of the inferior vena
cava. It gives off parietal branches to
the body wall, and visceral branches to
the viscera, and ventrad of the first sacral
vertebra it gives off two large branches on
each side, the external iliac (Fig. 126, k)
and the hypogastric (Fig. 126, l)—a very
small median vessel, the sacralis media
(Fig. 126, o), continuing the course of the
aorta and passing into the tail.
The aorta gives off the following
branches: A. cœliaca; A. mesenterica
superior; Aa. adrenolumbales; Aa. renales;
A. mesenterica inferior; Aa. iliolumbales;
Aa. lumbales (seven pairs);
Aa. iliacæ externæ; Aa. hypogastricæ.
A. A. cœliaca (Fig. 125).—The cœliac
artery is a large branch which is given off
from the aorta one centimeter or less
caudad of the opening in the diaphragm.
It passes directly ventrad about three or
four centimeters, then divides, usually at once, into three
branches. The most cranial of these is the hepatic (d), the
next is the gastrica sinistra (e), while the third and largest,
seeming to form a continuation of the cœliac, is the splenic (f)
(A. lienalis). The cœliac artery may give rise also, before its
division, to the two phrenic arteries (which, however, usually
arise from the adrenolumbales), and either before or at the point
of division to one or two small Aa. ventriculi dorsales, which,
however, frequently arise from the gastrica sinistra.
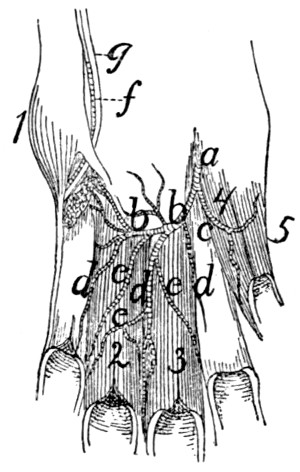
Fig. 124.
The Palmar Arch and
its Branches in the
Palm of the Hand.
The interosseous muscles
have been removed,
except those of the third
and fourth digits. 1, M.
abductor digiti quinti; 2,
3, Mm. interossei of third
and fourth digits; 4, M.
adductor pollicis; 5, M.
flexor brevis pollicis. a,
termination of radial artery;
b, palmar arch; c, A. princeps
pollicis et indicis; d,
Aa. interosseæ; e, muscular
branches of the same;
f, A. ulnaris; g, deep palmar
branch of ulnar nerve.
[302]
Branches of the cœliac artery:
1. A. hepatica (d).—The hepatic artery passes cranioventrad,
pierces the descending limb of the great omentum and
passes craniad to the liver, lying, together with the portal vein
and common bile-duct, in the ventral boundary of the foramen
epiploicum (foramen of Winslow) and contained, together with
the last-named vessels, in a fibrous sheath called the capsule
of Glisson. Just before entering the sheath it gives off the
gastroduodenalis (g). At its termination the hepatic artery
divides, sending branches to the lobes of the liver and a cystic
artery to the gall-bladder.
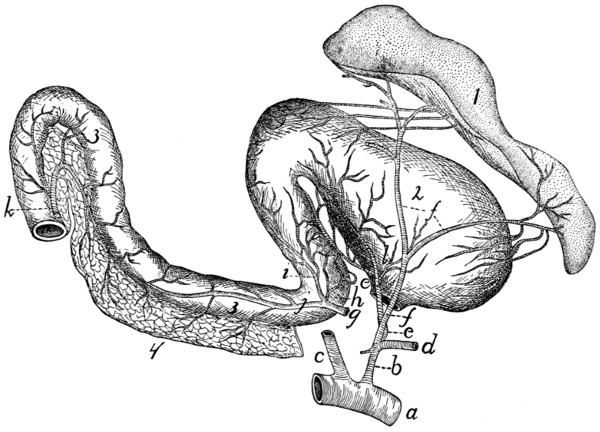
Fig. 125.—The
Cœliac Artery and its Branches.
a, abdominal aorta; b, A. cœliaca; c,
A. mesenterica superior (cut); d, A. hepatica
(cut); e, A. gastrica sinistra; f, A. lienalis; g, A. gastroduodenalis (cut from
A. hepatica); h, A. pylorica; i, A. gastroepiploica dextra; j, A. pancreaticoduodenalis
superior; k, A. pancreaticoduodenalis inferior (cut from inferior mesenteric);
l, A. ventriculi dorsalis. 1, spleen; 2, stomach; 3, duodenum; 4, pancreas.
a. A. gastroduodenalis (g).—This arises from the hepatic
near the pylorus and passes caudad, dividing one or two centimeters
from its origin into three branches, A. pylorica (h),
A. pancreaticoduodenalis superior (j), and A. gastroepiploica
dextra (i). The pylorica (h) (which may arise directly from
the hepatic) passes to the pylorus, thence along the lesser curvature[303]
of the stomach, dividing into numerous branches and
anastomosing with the gastrica sinistra. The pancreaticoduodenalis
superior (j) passes to the duodenum, supplying it and
the duodenal part of the pancreas and anastomosing with the
pancreaticoduodenalis inferior (k). The gastroepiploica dextra
(i) passes along the greater curvature of the stomach from the
pyloric end and supplies the walls of the stomach, sending
branches also to the ascending limb of the great omentum.
These branches anastomose with the terminal branches of the
splenic artery.
2. A. gastrica sinistra (e).—This arises from the cœliac
artery and passes to the lesser curvature of the stomach,
extending along this to the right. It gives off many branches
to the walls of the stomach, and anastomoses with A. pylorica.
A. ventriculi dorsalis (l).—One or two small arteries
which arise either from the gastrica sinistra or the cœliac
artery, and pass toward the dorsal part of the greater curvature
of the stomach near its cardiac end.
3. A. lienalis (f).—The splenic artery is the largest of the
branches of the cœliac axis, of which it appears to be the direct
continuation. It divides into two large branches, one to the
cranial end, the other to the caudal end, of the spleen (1).
From the latter a large branch passes to the pancreas and
descending limb of the great omentum.
B. A. mesenterica superior (Fig. 126, d).—The superior
mesenteric artery is larger than the cœliac. It supplies the
blood to the small intestines and caudal portion of the pancreas
and to the ascending and transverse colon. It arises from the
ventral side of the abdominal aorta about one centimeter
caudad of the cœliac axis and passes caudoventrad, forming a
curve with the convexity dextrad. It gives off the pancreaticoduodenalis
inferior, colica media, colica dextra, ileocolica,
and numerous branches to the small intestine.
1. A. pancreaticoduodenalis inferior (Fig. 125, k).—This
passes to the caudal end of the pancreas and a part of the
duodenum, anastomosing with the pancreaticoduodenalis
superior.
2. A. colica media is a large branch to the transverse and[304]
descending portions of the large intestine. It divides and sends
branches in both directions which anastomose with the colica
dextra and with branches from the inferior mesenteric arteries.
3. A. colica dextra.—A small branch which passes to the
ascending and transverse colon, anastomosing with the colica
media and ileocolica. It is sometimes absent.
4. A. ileocolica.—This passes to the cæcum and supplies
that structure, sending branches to the ileocolic valve and the
caudal end of the ileum, and anastomosing with the colica
dextra and the intestinal branches of the superior mesenteric.
5. The superior mesenteric now divides into about sixteen
terminal branches which pass to the small intestine. In many
cases these branches unite near the intestine, forming arches,
and from these arches numerous short branches pass to the
intestine.
3. A. adrenolumbalis (or lumboabdominalis) (Fig. 126, e).—This
rises from the aorta, one on each side, about two
centimeters caudad of the superior mesenteric. Each passes
laterad onto the dorsal body wall, supplying the muscles of this
region. A large branch passes caudad along the surface of the
muscles dorsad of the kidney and anastomoses with the iliolumbar
artery. From the adrenolumbalis rises usually:
A. phrenica (f).—The phrenic artery rises either from the
cœliac (c) or the adrenolumbalis (e) and passes to the diaphragm,
near its dorsal border. In the diaphragm it passes
ventrad, lying at the medial margin of the costal portion of the
diaphragm, as far as its sternal portion, where it unites with the
artery of the opposite side. The two thus form an arch, from
which radiating arteries pass off to supply the diaphragm.
4. A. renalis (g).—The two renal arteries arise from the
sides of the aorta, usually at about the same point, so that the
left passes caudolaterad and the right craniolaterad (owing to
the position of the kidneys). The artery usually divides just
before entering the kidney (2). It passes dorsad of the vein
(t). The renal artery sometimes sends a branch to the suprarenal
body (1). It also occasionally gives origin to the spermatic
artery (h).
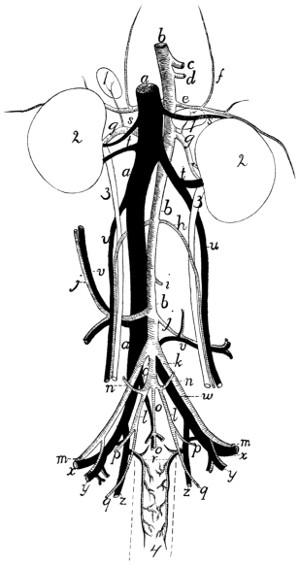
Fig. 126.—Abdominal
Blood-vessels, Ventral View.
a, vena cava inferior; b, abdominal aorta; c,
A. cœliaca (cut); d, A. mesenterica
superior (cut); e, A. adrenolumbalis; f, A. phrenica; g, A. renalis; h, A. spermatica
interna; i, A. mesenterica inferior (cut); j, A. iliolumbalis; k, A. iliaca
externa; l, A. hypogastrica; m, beginning of A. femoralis; n, A. umbilicalis; o, A.
and V. sacralis media (o, the artery; o′, the vein); p, A. glutea superior; q, A.
glutea inferior; r, A. and V. hemorrhoidalis media; s, V. adrenolumbalis; t, V.
renalis; u, V. spermatica interna; v, V. iliolumbalis; w, V. iliaca communis; x, V.
femoralis; y, V. and A. profunda femoris; z, V. hypogastrica. 1, suprarenal body;
2, kidney; 3, ureter; 4, outline (broken) of rectum.
5. A. spermatica interna (h).—This rises from the aorta[305]
[306]
on each side at about the level of the caudal ends of the kidneys,
and passes laterad (or it may arise from the renal). In male
specimens the artery turns caudad, along with the corresponding
vein (u), and passes to the inguinal canal. Thence it
passes along with the vas deferens and the vein (forming
all together the spermatic cord) to the testes and the other
structures in the scrotum.
In females (Fig. 112, 2, page 265) the artery is larger and
passes more nearly directly laterad, being much convoluted.
It passes to the ovary, being now called the ovarian artery.
It sends branches into the ovary (c) and to the cranial end of
the uterus (f); the latter anastomosing with branches of the
uterine artery (3).
6. A. mesenterica inferior (Fig. 126, i).—The inferior mesenteric
artery has its origin from the aorta at about the level of
the last lumbar vertebra. It passes toward the large intestine,
and near it divides into two branches, the colica sinistra,
which passes craniad along the descending colon, anastomosing
with the colica media, and the superior hemorrhoidal,
which passes caudad along the descending colon and rectum
and anastomoses with the middle hemorrhoidal (page 308).
7. A. iliolumbalis (Fig. 126, j).—The iliolumbar arteries
arise from the aorta about two centimeters caudad of the inferior
mesenteric and pass laterad over the ventral surface of the psoas
minor and iliopsoas muscles. Each divides into two main
branches, one passing caudad and the other craniad; these
supply the muscles of this region. The cranial branch anastomoses
with branches of the adrenolumbalis (e). The caudal
branch sends an artery through the abdominal wall to appear
at the cranial edge of the thigh; it passes onto the lateral surface
of M. sartorius and extends here some distance distad.
8. A. lumbalis.—There are usually seven pairs of lumbar
arteries, passing almost directly dorsad from the dorsal surface
of the aorta; the first pair arising just craniad of the diaphragm,
the last at the same level as the origin of the external iliac.
The two arteries of a pair usually arise from the aorta one
behind the other, or they may arise by a common trunk.
They correspond to the intercostal arteries. They pass[307]
between the centra of the vertebræ and the muscles of the
dorsal region, and send one main branch laterad, the other
dorsad; both supply the dorsal muscles. From the dorsal
branch small arteries pass into the vertebral canal to join the
anterior spinal artery.
9. A. iliaca externa (Fig. 126, k).—The external iliac
artery passes obliquely caudad from the aorta, lying ventrad of
the common iliac vein and against the medial surface of the
psoas minor muscle. It passes onto the ventral surface of this
and the iliopsoas and at the same time reaches the tendon of the
abdominal muscles; through a small opening in this tendon
lying ventrocaudad of that for the iliopsoas, it leaves the
abdominal cavity. On its emergence from the abdominal
cavity onto the medial surface of the thigh it receives the name
A. femoralis (m). The branches of the external iliac artery
are described below (page 309).
10. A. hypogastrica (or iliaca interna) (Fig. 126, l).—The
hypogastric or internal iliac arteries are given off from the aorta
usually within a centimeter caudad of the external iliac. Each
passes caudolaterad, lying on the medial side of the common
iliac vein, and divides into branches which supply the structures
within the pelvis and the muscles about the pelvic wall.
There is much variation in the origin and relations of the
branches of this artery. The following seems to be a very
common arrangement: A very short distance from the origin
the umbilical artery (n) is given off. The hypogastric artery
then extends two or three centimeters and gives off A.
glutea superior (p), which passes at once to the pelvic
wall. A little farther caudad the hypogastric divides into the
middle hemorrhoidal (r), to the rectum, and the glutea inferior
(q), which passes out of the pelvis.
Branches of the hypogastric artery:
a. A. umbilicalis (Fig. 126, n).—This small artery arises
from the hypogastric about one centimeter from the beginning
of the latter, and passes ventrad to the bladder. Here it divides
into two branches—the superior vesical to the sides of the
bladder, the inferior vesical to the neck of the bladder and the
urethra.
[308]
b. A. glutea superior (Fig. 126, p).—This passes dorsolaterad
from the hypogastric and reaches the pelvic wall against
the medial surface of the ilium. Here it divides into two
branches. One passes between the iliopsoas muscle and the
ventral border of the ilium to the medial surface of the gluteus
maximus; it gives branches to the gluteus muscles, the pyriformis,
and the rectus femoris. The other branch passes dorsad
of the ilium, sending a branch to the medial surface of the
gluteus medius, and a small branch which joins the lateral
sacral artery and enters thus the sacral canal.
c. A. hemorrhoidalis media (Fig. 126, r).—The middle
hemorrhoidal passes ventrad from the hypogastric, onto the
lateral surface of the rectum. It passes caudad near the ventral
side of the rectum as far as the anus. A short distance from the
origin it gives off a branch which passes almost directly ventrad
toward the beginning of the urethra. In the male this branch
is small, sending twigs to the urethra and the prostate. In
the female it is much larger, forming A. uterina (Fig. 112, 3,
page 265). The uterine artery turns craniad onto the uterus,
which it supplies, passing to the cranial end of the uterine
cornu and anastomosing with the ovarian artery.
The middle hemorrhoidal gives off in the female branches
to the vagina (Fig. 112, m) and neck of the bladder (k), and
a large branch to the urogenital sinus (n); it then supplies the
anal glands and other structures about the anus, and anastomoses
with the terminal branches of the hemorrhoidalis
superior. In the male branchlets are given to the urethra, to
the bulbourethral (or Cowper’s) gland, and to the penis. The
A. dorsalis penis is a small branch which passes along the
median dorsal groove of the penis to the glans. The middle
hemorrhoidal then supplies the structures about the anus and
anastomoses with the superior hemorrhoidal, as in the female.
d. A. glutea inferior (Fig. 126, q).—This is the terminal
portion of the hypogastric. It passes along with the great
sciatic nerve to the great sciatic notch, and reaches the medial
surface of M. pyriformis. It divides into branches which supply
the gluteus and pyriformis muscles, while a small branch
accompanies the great sciatic nerve. A small branch also[309]
passes onto the lateral surface of the tail, extending caudad
along its side.
11. A. sacralis media (Fig. 126, o).—This is the continuation
of the aorta into the sacral and caudal regions. It extends
to near the end of the tail, lying in the ventral middle line
against the sacral and caudal vertebræ. In the caudal region
it passes through the hæmal arches, covered by the chevron
bones. Between the vertebræ it gives off side branches comparable
to the intercostal and lumbar arteries of the aorta.
Branches of the sacralis media:
a. A. sacralis lateralis.—Behind the first sacral vertebra
a large branch, the lateral sacral, is given off on each side.
This enters the first anterior sacral foramen, gives a branch to
the structures in the sacral canal, and sends a dorsal branch
out through the posterior sacral foramen to the muscles on the
dorsal side of the sacrum.
Similar but smaller branches are given off between the succeeding
vertebræ, sacral and caudal.
The External Iliac and its Branches.
—The origin of the
external iliac (Fig. 126, k) is described above (page 307).
It gives off the following branches:
1. A. profunda femoris (Fig. 126, y).—This is given off just
before the external iliac leaves the abdominal cavity. It passes
caudad, and about one centimeter from its origin it gives off
three branches, sometimes all separately, sometimes two in
common. One of these passes mediad in the lateral ligament
of the bladder and ramifies on the lateral surface of that organ.
A second passes through the abdominal wall and into the fat
on the medial surface of the thigh; it sends a branch caudad
toward the external genital organs, while its main trunk passes
distad in the subcutaneous fat almost to the knee. From one
of these two a small branchlet passes in the male to the spermatic
cord and accompanies this to the testis; it represents
thus the external spermatic artery. The third branch,
A. epigastrica inferior, passes directly to the ventral surface of
the rectus abdominis muscle, on which it passes craniad, sending
branches to the muscles of the abdominal walls. It anastomoses
with terminal branches of the internal mammary artery.
[310]
The profunda femoris itself passes between the iliopsoas
and the pectineus muscles, then
to the medial surface of the
adductor femoris. It divides
into branches which supply
the pectineus, adductor
longus, adductor femoris,
caudofemoralis, quadratus
femoris, semimembranosus,
and semitendinosus.
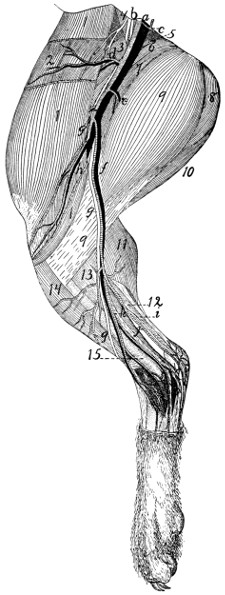
Fig. 127.—Superficial Arteries, Veins, and Nerves on
the Medial Side of the Leg.
a, A. and V. femoralis; b, N. femoralis; c, A. profunda
femoris; d, A. and V. circumflexa femoris lateralis; e, ramus
muscularis; f, A. saphena and V. saphena magna; g, N.
saphenus; h, A. and V. articularis genu suprema; i, N. tibialis;
j, plantar branch of A. saphena; k, dorsal branch of A.
saphena. 1, M. sartorius; 2, M. tensor fasciæ latæ; 3, M.
rectus femoris; 4, M. iliopsoas; 5, M. pectineus; 6, M. adductor
longus; 7, M. adductor femoris; 8, M. semimembranosus;
9, M. gracilis; 10, M. semitendinosus; 11, medial head
of M. gastrocnemius; 12, M. plantaris; 13, M. flexor longus
digitorum; 14, M. tibialis anterior; 15, tibia.
2. A. femoralis (Fig.
127).—This is the continuation
of the external
iliac onto the medial surface
of the thigh. The
artery lies in a triangular
depression between the
borders of the sartorius (1)
and gracilis (9) muscles. The
floor of the depression is formed
by the adductor longus (6), pectineus
(5), vastus internus, and
rectus femoris (3) muscles. This
triangular depression is known as the iliopectineal
fossa, or Scarpa’s triangle; it contains
also the femoral vein (a) and saphenous
nerve (g), which pass along with the artery.[311]
The artery extends distad, and at about the middle of the length
of the thigh it passes, along with the vein and nerve, into a
groove between the vastus medialis and the adductor femoris.
This groove is converted into a canal by the overlying aponeurosis;
it is known as Hunter’s canal or the canalis adductorius.
At the distal end of the adductor femoris the femoral
artery ceases to be superficial and passes between the vastus
medialis and the semimembranosus to the popliteal space; it
now receives the name A. poplitea.
Branches of the femoral artery:
a. A. circumflexa femoris lateralis (Fig. 127, d).—The
lateral circumflex arises from the femoral about one centimeter
from its emergence from the abdominal cavity. It passes
craniad and laterad and divides into two main portions. One
part passes between the rectus femoris and vastus medialis,
gives branches to these muscles, and sends an ascending
branch to the structures about the hip-joint, and a descending
branch into the vastus medialis muscle. The remainder
of the lateral circumflex passes along the inner (lateral) surface
of the sartorius (1) to the cranial border of the leg, giving
branches to the sartorius (1) and tensor fasciæ latæ (2).
b. A large muscular branch (e) passes caudad (or ventrad)
and distad across the distal end of the adductor femoris (7)
and between the gracilis (9) and semimembranosus, supplying
these muscles. Other small muscular branches may be given
off from both sides of the femoral.
c. A. articularis genu suprema (h).—The superior articular
artery arises from the femoral, either in common with the
saphenous artery or separately, a little proximad of the point
where the femoral ceases to be superficial. It passes toward
the knee, between the vastus medialis and semimembranosus,
and covered by the sartorius (1), and ramifies over the medial
surface of the knee-joint and in the structures just proximad of
the joint.
d. A. saphena (f).—The saphenous artery frequently takes
origin in common with the superior articular (h), but may arise
separately at about the same level. It passes distad across the
gracilis (9), accompanied by the saphenous
nerve (g) and vein,[312]
sends several branches dorsad (toward the knee), and at about
the middle of the lower leg it divides into two main branches,
a dorsal branch (k), passing to the dorsum of the foot, and a
plantar branch (j), to the sole of the foot. The dorsal branch
is accompanied by the main saphenous nerve (g) and vein. It
sends one or two branches to the medial side of the ankle-joint,
passes then onto the dorsum of the foot, and divides into
four main branches. The medial one passes along the medial
side of the medial digit; the other three pass to the intervals
between the digits. Each divides into two branches which
supply the contiguous sides of the two digits between which
the interval lies. The dorsal branch thus supplies arteries to
the sides of all the digits except to the lateral side of the lateral
digit. This is supplied by A. suralis.
The plantar branch (j) is larger than the dorsal. It passes
distad on the medial surface of the flexor longus hallucis,
accompanied by the tibial nerve, and across the space between
the tendon of Achilles and the flexor. It gives superficial and
deep branches to the structures about the ankle-joint, and
sends inward a branch from the lateral side (Fig. 128, d)
which joins the termination of A. tibialis anterior to form the
plantar arch. It then passes distad along the plantar surface
of the foot, nearer its medial border. Beneath the pad in the
sole of the foot it divides into three branches which supply the
interosseous spaces between the three digits.
e. A. poplitea.—The main trunk of the femoral artery
passes between the vastus medialis and the semimembranosus
and then through the distal portion of the adductor femoris to
reach the popliteal space. This is the space ventrad (or
caudad) of the knee, between the biceps femoris on one side
and the semimembranosus on the other. Here the artery
receives the name A. poplitea, or popliteal artery. The popliteal
artery gives off a number of large branches as it passes
through the popliteal space, passes between the condyles of
the femur and underneath the popliteal muscle, and finally
turns dorsolaterad between the tibia and fibula, just distad of
the head of the fibula. It now receives the name A. tibialis[313]
anterior and passes distad along the dorsal (anterior) border
of the fibula.
Branches of the popliteal artery:
1. A. suralis.—This is a large branch which passes distad
from the caudal side of the popliteal artery. It sends branches
to the biceps and to the fat in the popliteal space, and passes
onto the ventral border of the lateral head of the gastrocnemius.
It sends branches to both heads of the gastrocnemius and to
the popliteus, passes distad onto the lateral surface of the tendon
of Achilles, and may be traced to the proximal portion of
the dorsolateral side of the foot, where it sends many branches
to the integument about the ankle-joint. It then passes along
the lateral side of the foot and supplies the artery on the lateral
side of the fifth digit.
2. Aa. genu posteriores.—Several small arteries which
pass to the knee-joint.
3. Rami musculares.—Numerous small branches to the
muscles about the popliteal space.
4. A. tibialis posterior.—This is the largest branch of the
popliteal artery, which it leaves just before the latter passes
beneath the popliteal muscle. The tibialis posterior passes at
first mediad, then turns distad, passes across the popliteus
muscle onto the surface of the flexor longus hallucis, and
ramifies in the substance of this muscle and its tendon. It
gives branches also to the gastrocnemius and soleus. (It does
not pass into the foot as does the corresponding artery in
man.)
5. A. tibialis anterior.—The anterior tibial artery is the
continuation of the popliteal after its passage through the interosseous
membrane between tibia and fibula to the dorsal side of
the lower leg. Here it passes distad, lying against the interosseous
membrane, between the extensor longus digitorum and
the tibialis anterior muscle, or partly imbedded in the substance
of the latter. It passes, lying beneath the tendons of these
muscles, through the transverse ligament proximad of the
malleoli, thus reaching the dorsum of the foot. It extends distad
across the tarsus to the space between the second and third
metatarsals, and through this space to the plantar side of the[314]
foot (Fig. 128). Here it receives one or two small branches
(a and d) from the plantar branch of A. saphena (forming
with them the plantar arch
(c)), and passing distad and
slightly laterad, divides into
three main portions (e). These
(Aa. digitales plantares) pass
to the three intervals between
the digits. The medial one
passes along the lateral side
of the second digit; the next
divides, sending a branch to
the lateral side of the third
digit and the medial side of
the fourth; the lateral one
passes along the medial border
of the fifth digit. The Aa.
digitales plantares send numerous
branches to the interosseous
muscles.
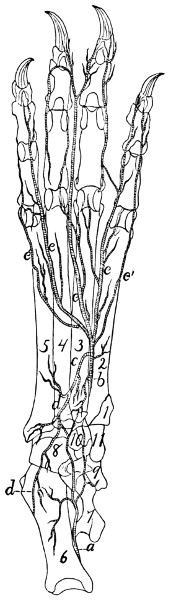
Fig. 128.—Deep Arteries on the
Plantar Surface of the Foot.
a, medial twig from plantar branch of
A. saphena; b, A. tibialis anterior (or
plantaris profunda); c, plantar arch; d,
lateral twig from plantar branch of A.
saphena; e, terminal branches from A.
tibialis anterior.
Branches of the tibialis
anterior:
a. A. tibialis recurrens.—Immediately
after passing
through the interosseous membrane
the anterior tibial sends
a small artery proximad to the
knee-joint.
b. Rami musculares.—Numerous
short branches
throughout the course of the
artery, to the muscles about it.
c. Ramus superficialis.—A
larger branch leaves the
lateral surface of the artery
about one-third the distance
from the knee to the ankle and passes along with the superficial
peroneal nerve, at first beneath M. peroneus longus, then[315]
superficially. It passes onto the dorsal surface of the foot and
becomes connected with terminal branches of A. saphena.
d. A. tarsea medialis.—This arises from the tibialis anterior
just distad of the transverse ligament through which
it passes at the ankle, passes over the medial surface of
the astragalus, and is distributed to the ligaments about the
ankle-joint. It anastomoses with the plantar branch of A.
saphena.
e. A. metatarsea.—The metatarsal artery passes laterad
from the tibialis anterior at the distal boundary of the tarsus,
on the dorsal surface of the foot. It sends branches distad in
the intervals between the third and fourth and the fourth and
fifth metatarsals.
The distal branches (Fig. 128) of the tibialis anterior have
been described in the general description of the artery.
THE VEINS. VENÆ.
Venæ pulmonales. The Pulmonary Veins.
The pulmonary veins follow in the lungs the course of the
bronchi. They enter the left auricle in three groups (Fig.
116, page 276). The first of these (i) comes from the anterior
and middle lobes of the right side, the second (h) from the
corresponding lobes of the left side, and the third (g) from the
terminal lobes of both sides. Each group is composed of two
or three veins and opens into a sinus or extension of the auricle.
The sinuses are from their position dextral (i), sinistral (h),
and dorsal (g).
The Veins of the Body.
The veins of the body may be classified into three main
groups: (1) the veins of the heart; (2) the superior vena cava
and its branches; (3) the inferior vena cava and its branches.
The portal system of veins will be considered with the inferior
vena cava.
1. Veins of the Heart.
The veins of the heart consist of one large vein, the vena
cordis magna, and a number of small veins.
[316]
Vena cordis magna (Fig. 118, 3).—This arises as a number
of scattered branches on the surface of both ventricles. These
unite to form two main trunks. One of these, on the ventral
side of the heart, marks externally the septum between the
two ventricles. It passes beneath the left auricular appendage,
then turns to the left and passes along the sulcus coronarius
to the dorsal side of the heart. Here it unites with the second
main trunk which ramifies over the surface of the left ventricle.
Arriving at the base of the right auricle it enters
this by the coronary sinus, which lies just beneath the opening
of the inferior vena cava.
In addition to the vena cordis magna and its branches
there are a number of small veins, especially in the wall of the
right ventricle. These enter the right auricle by a number of
small separate openings along the sulcus coronarius.
2. Vena cava superior (Fig. 129, b).
The superior vena cava is the great vein returning the
blood from the head, fore-limb, and cranial part of the trunk.
It extends from the level of the first rib on the right side of the
vertebral column to the right auricle. Its caudal end lies
dorsad of the aortic arch. Opposite the right rib it is formed
by the junction of the two innominate veins (m).
Branches of the superior vena cava:
1. V. azygos (d).—The first branch of the superior vena
cava is the azygos vein, which enters the vena cava on the
right side a centimeter or less craniad of the root of the right
lung. It is formed in the abdominal cavity by the confluence
of two or three small veins, which collect the blood from the
muscles of the dorsal wall of the abdomen. The small median
trunk thus formed enters the thoracic cavity between the crura
of the diaphragm and lies on the ventral surface of the centra
of the thoracic vertebræ, slightly to the right of the middle
line. The azygos receives the intercostal veins (n), which
correspond to the intercostal arteries and have the same course
and distribution. In the caudal part of the thoracic cavity the
intercostals enter the azygos separately, but the intercostals of[317]
[318]
the cranial intercostal spaces usually unite two or three
together and enter the azygos by one or more common trunks.
The azygos also receives bronchial and œsophageal veins,
corresponding to the similarly named arteries.
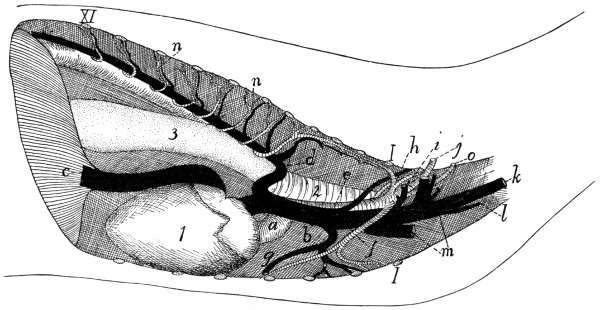
Fig. 129.—Thoracic
Blood-vessels, from the Right Side.
a, aorta; b, vena cava superior; c,
vena cava inferior; d, V. azygos; e, A. subclavia dextra; f, A. mammaria
interna; g, V. mammaria interna; h, costocervical axis; i,
united trunk of vertebral and costocervical veins; j, A.
and V. axillaris; k, V. jugularis externa; l, V. jugularis interna; m,
the two innominate veins; n, intercostal arteries
and veins; o, thyrocervical axis. 1, heart; 2, trachea; 3, œsophagus;
I-XI, cut ends of the first eleven ribs.
2. V. mammaria interna (g).—The internal mammary
veins follow the corresponding arteries. The two veins unite
to form a common trunk which, after receiving a branch from
the cranial part of the sternum, enters the vena cava opposite
the third rib.
3. Vv. anonymæ (Fig. 129, m; Fig. 115, p).—The innominate
veins unite opposite the first intercostal space to form
the vena cava superior. Each extends from the cranial end of
the vena cava superior craniolaterad to a point a short distance
craniad of the first rib and is there formed by the union of the
external jugular (Fig. 129, k) and subclavian (j) veins. The
innominate vein has the following branches:
A and B. Vv. vertebralis and costocervicalis (i).—The
vertebral and costocervical veins unite to form a single trunk
which enters the innominate about opposite the first rib.
These two veins follow the corresponding arteries. (Sometimes
the vertebral and costocervical veins enter the innominate
separately, and in other cases one or more of the components
of the costocervical veins (e.g., the transversa colli) may enter
the axillary vein. In other cases the entire common trunk of
the vertebralis and costocervicalis may enter the superior vena
cava directly.)
C. V. subclavia.—The subclavian vein enters the innominate
at about the level of the first rib. Outside of the thorax
it is called the axillary vein (j); this comes from the arm.
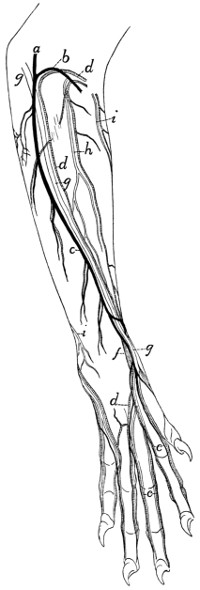
Fig. 130.
Superficial
Arteries,
Veins, and
Nerves on
the Extensor
Side of the
Arm and Dorsum
of the
Hand.
a, V. cephalica;
b, V. mediana cubiti;
c, V. cephalica
antibrachii; d,
A. collateralis radialis
superior; e,
Aa. and Vv. digitales
dorsales; f,
A. radialis; g, superficial
radial
nerve; h, N. musculocutaneus;
i, N.
cutaneus medialis.
Veins of the Arm.—The blood is returned from the arm
by two systems of veins,—a deep and a superficial system.
The deep system is composed of veins which correspond to
the branches of the axillary artery, follow in general the same
course, and receive the same names (Fig. 122). They unite to
form the axillary vein. The brachial vein does not pass with
the brachial artery through the supracondyloid foramen.
The superficial system comprises the vena cephalica and
its branches.
[319]
V. cephalica (Fig. 130).—Several superficial branches
lying in the palm of the hand unite to form a common trunk
which curves about the radial side of
the wrist and here unites with a similarly
formed trunk from the dorsum of
the hand. The vein formed by this
union, vena cephalica (antibrachii) (c),
passes to the elbow accompanying the
radial nerve (g) and the collateral radial
artery (d), and lying on the extensor
carpi radialis longus muscle just beneath
the integument. It receives small lateral
superficial branches. At the elbow it
divides into two: these are the vena
cephalica (humeri) (a) and the vena
mediana cubiti (b). The latter passes
beneath the pectoantibrachialis muscle,
across the biceps, and unites with the
brachial vein in the concavity of the
elbow.
The vena cephalica (humeri) passes
at the elbow onto the upper arm, following
the lateral border of the
clavobrachial muscle. It thus
reaches the acromiodeltoid muscle,
where it divides into two branches.
One passes inward beneath the
acromiodeltoid, and joins the
vena circumflexa posterior (a
branch of the axillary). The
second continues over the
outer surface of the shoulder,
and finally joins the vena
transversa scapulæ (a branch
of the external jugular).
D. V. jugularis externa
(Fig. 131; Fig. 65, 5, page
109).—The external jugular[320]
vein comes from the head and face. It is formed by the junction
of the anterior (Fig. 131, c) and posterior (b) facial veins
near the ventral border of the sternomastoid muscle. It passes
thence obliquely across the sternomastoid to the triangular
interval between it and the clavotrapezius. In the interval
it receives a small branch which accompanies the branch of
the transversa scapulæ artery to the clavobrachialis muscle,
receives next the common trunk of the vena cephalica and
transversa scapulæ, and opposite the cranial end of the
manubrium receives the internal jugular vein (Fig. 129, l).
It then unites with the subclavian to form the innominate
vein. Near the point where it receives the internal jugular the
external jugular vein receives the thoracic duct (Fig. 118, 5).
Branches of the external jugular:
a. V. transversa scapulæ.—This follows the course of the
artery of the same name. One of its branches receives one of
the two terminal divisions of the vena cephalica (q. v.). In
many cases veins which correspond to branches of the transversa
scapulæ artery enter the external jugular separately (as
shown in Fig. 118, x and y).
b. V. cervicalis ascendens.—A small branch which follows
the corresponding branch of the thyrocervical axis.
c. V. jugularis interna (Fig. 118,
2; Fig. 119, page 284).—The
internal jugular vein varies much in size and in the
place where it joins the external jugular. The point of junction
is usually at about the level of the first rib, but may be much
farther craniad. It arises on the ventral side of the basal portion
of the occipital by the junction of a number of veins which
are described below. The internal jugular passes caudad in
the neck region at the side of the trachea, in company with the
common carotid artery and the vagus and sympathetic nerves.
In this region it receives branches which accompany the
branches of the common carotid artery.
The veins which by their union form the internal jugular
vein are the following:
(1) One or two veins from the venous sinuses of the brain,
leaving the cranial cavity by the jugular foramen. These are
described in the account of the veins of the brain.
[321]
(2) A large communicating branch from the posterior facial.
(3) The vena occipitalis, a large branch coming from the
vertebral column (which it leaves by the atlantal foramen) and
from the back of the head.
d. V. facialis anterior (Fig. 131, c).—The anterior facial
vein collects the blood from the face, the tongue, and adjacent
parts; it joins the posterior facial (b) caudad of the angle of
the jaw to form the external jugular vein.
The anterior facial begins over the frontal bone caudad of
the orbit, where it is known as the frontal vein (c″). This
runs along the dorsal border of the orbit, into which it dips.
It sends a communicating branch into the orbit, and receives
small veins (superior palpebral) from the upper eyelid. It
then turns ventrad, passing along the cranial angle of the eye
between the levator labii superioris proprius (5) and the orbicularis
oculi (8) muscles, being called in this region the angular
vein (c′). This receives branches (external nasal veins, k)
from the side of the nose. The vein crosses the malar bone
obliquely (now receiving the name anterior facial, c) and follows
the cranioventral border of the masseter muscle (9). As
it passes the infraorbital foramen it receives through the
foramen a small vein from within the orbit. It receives also
the inferior palpebral (j) from the lower eyelid, and the
superior labial vein (i) from the upper lip. Just caudad of the
angle of the mouth it receives from beneath the cranioventral
margin of the masseter the V. facialis profunda (h) described
below. Still farther caudad it receives the inferior labial vein
(g) from the lower lip, and a small branch from the masseter
muscle. Caudad of the middle of the cranioventral border of
the masseter it receives from beneath the lymphatic gland of
this region the large submental vein (f). Next the anterior
facial vein receives a large communicating branch (V. transversa,
d) which passes transversely across the ventral surface
of the throat and connects the two anterior facial veins of the
opposite sides. From the middle of this communicating branch
an unpaired trunk passes caudad in the middle line and divides
into the two laryngeal veins, which pass to the larynx; a small
branch is continued from the unpaired trunk craniad into the[322]
tongue. The communicating branch receives also near its
junction with the anterior facial a rather large superficial branch
(e) which comes from the sides of the lower jaw and passes
across the lymphatic gland (12) to join the communicating
branch.
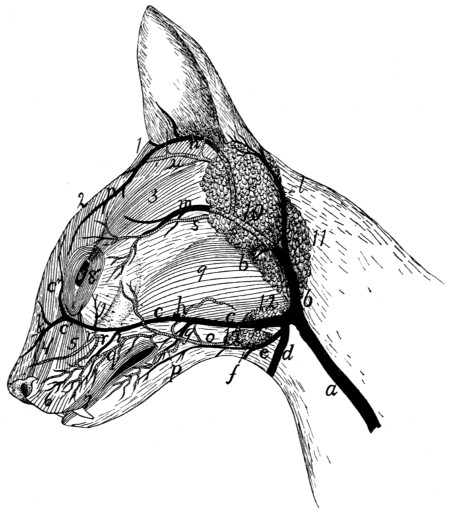
Fig. 131.—Superficial
Veins and Arteries of the Face.
a, external jugular vein; b, posterior
facial vein; c, anterior facial vein (c′, angular
vein; c″, frontal vein); d, communicating branch with opposite anterior facial; e,
superficial branch from lower jaw; f, submental vein; g, inferior labial vein; h, deep
facial vein; i, superior labial vein; j, inferior palpebral vein; k, external nasal veins;
l, posterior auricular vein; m, superficial temporal vein; n, anterior auricular vein;
o, external maxillary artery; p, inferior labial artery; q, superior labial artery; r,
infraorbital arteries; s, superficial temporal artery; u, anterior auricular branch of
posterior auricular artery. 1, M. adductor auris superior; 2, M. corrugator supercilii
medialis; 3, M. corrugator supercilii lateralis; 4, M. levator labii superioris alæque
nasi; 5, M. levator labii superioris proprius; 6, M. myrtiformis; 7, M. orbicularis
oris; 8, M. orbicularis oculi; 9, M. masseter; 10, parotid gland; 11, submaxillary
gland; 12, lymphatic glands.
The following larger branches of the anterior facial are not
described above:
[323]
V. facialis profunda (Fig. 131, h).—The deep facial vein
enters the anterior facial from beneath the masseter, just caudad
of the angle of the mouth. It collects branches from the soft
and hard palates and the teeth, these branches passing along
with the corresponding arteries for a distance, then uniting and
passing ventrolaterad beneath the zygomatic arch to join the
anterior facial. The deep facial vein also receives branches
which come from the venous plexus that lies in the same region
as the arterial carotid plexus.
V. submentalis (Fig. 131, f).—This enters the anterior
facial vein at the ventral border of the masseter muscle and
near the cranial end of the two superficial lymph-glands (12)
in this region. The submental vein is formed at the dorsal
border of the digastric muscle by two branches emerging from
between the digastric and the mandible. The more cranial
of these, V. lingualis, collects branches from the tongue,
from its middle to the tip; it passes through the middle of the
lateral border of the mylohyoid muscle to join the other branch.
The second branch emerges from between the mandible and
the pterygoid muscles. It comes from near the caudal end of
the mandible, where it becomes continuous with a communicating
branch from the posterior facial vein, and receives a
branch which comes from the mandibular canal; also small
branches from the adjacent parts.
e. V. facialis posterior (Fig. 131, b).—The posterior facial
vein arises from the region of the internal maxillary artery, its
terminal branches following the branches of the artery. It
collects blood from the pterygoid, masseter (9), and temporal
muscles, and forms a plexus which is interwoven with the
carotid (arterial) plexus in the region of the orbital fissure. It
is connected with the submental vein by a communicating
branch. From the posterior facial a branch extends ventrad
close against the outer surface of the tympanic bulla and then
turns caudad, receives a branch from the pharynx, then continues
to join the internal jugular. The posterior facial itself
passes laterad along the caudal border of the masseter (9) and
the ventral border of the parotid gland (10), then becomes
superficial and turns ventrad over the outer surface of the submaxillary[324]
gland (11) to join the anterior facial (c) caudad of the
angle of the jaw.
As it passes ventrad of the parotid gland (10), just before
becoming superficial, the posterior facial receives the superficial
temporal (m). This at first follows the superficial temporal
artery (s), then enters the substance of the parotid gland,
joining the posterior facial near the ventral border of the latter.
The superficial temporal receives the anterior auricular (n), a
large vein passing along the cranial border of the auricular
opening and arising dorsad of the eye.
The posterior facial receives also, at the point where it
passes from beneath the parotid gland, the posterior auricular
or great auricular vein (l). This collects blood from the back
of the head and the external ear, its branches following those
of the artery of the same name. It passes along the caudal
border of the parotid gland to join the posterior facial.
Veins of the Brain and Spinal Cord.
—The veins of
the brain form a large number of small vessels which pour their
blood into larger veins lying in folds of the dura mater; these
are known as the venous sinuses of the dura mater. These
sinuses communicate with the venous plexus about the orbital
fissure, coming from the posterior facial vein; with the internal
jugular through the jugular foramen, and with the vertebral
veins, in the vertebral canal. The chief sinuses of the dura
mater on the dorsal side of the brain are as follows:
1. Sinus sagittalis superior.—This lies in the dorsal part
of the falx cerebri, between the two hemispheres of the cerebrum.
It receives veins from the dorsal and middle parts of
the cerebrum, and passes caudad to the tentorium. Here it
enters the sinus transversus. It receives the vena cerebri
magna, a large vein coming from the interior of the brain and
passing directly dorsad at the caudal end of the corpus callosum
to enter the sinus sagittalis.
2. Sinus transversus.—This lies in a canal in the dorsal
border of the tentorium. It receives numerous small veins
from the cerebellum, roof of the skull, etc. One or two centimeters
on each side of the middle line the sinus transversus
passes out of the canal onto the caudal surface of the tentorium,[325]
thence proceeds caudoventrad obliquely over the surface of the
cerebellum, unites with veins from the ventral side of the
brain, leaves the skull by the jugular foramen, forming thus
the inferior cerebral vein, and joins the internal jugular vein.
On the ventral side of the brain are the following sinuses of
the dura mater:
3. Sinus cavernosus.—A short broad venous sinus, one
on each side of the hypophysis, on the body of the sphenoid.
It receives veins from the side and ventral surface of the brain.
The two sinuses are connected by communicating branches
craniad and caudad of the hypophysis. From them branches
pass out through the orbital fissure to join the plexus formed
by the branches of the posterior facial vein.
4. Sinus petrosus inferior.—This arises from the sinus
cavernosus and passes caudolaterad in the groove between the
edge of the petrous bone and the basilar portion of the occipital.
Reaching the jugular foramen it divides; part joins the
termination of the sinus transversus to form the inferior cerebral
vein, which passes through the jugular foramen to join the
internal jugular vein. The other portion of the sinus petrosus
inferior passes through the condyloid canal of the occipital
bone, communicates by a strong transverse branch across the
surface of the basioccipital with the vein of the opposite side,
and enters the spinal canal through the foramen magnum.
Here it joins the sinus of the vertebral column.
5. Sinus columnæ vertebralis.—On the ventral surface of
the vertebral canal, beneath the periosteum, are two wide
venous sinuses, one on each side of the middle line. These
sinuses extend the entire length of the spinal cord. At the
atlantal foramen each sends a strong branch to the internal
jugular vein. Farther caudad they send branches to the vertebral,
intercostal, and lumbar veins, and communicate with each
other by numerous transverse branches. They receive many
small veins from the spinal cord.
3. Vena cava inferior (Fig. 129,
c; Fig. 126, a).
The inferior vena cava (Fig. 126, a) is formed at about the
level of the last lumbar vertebra by the union of the two common[326]
iliac veins (Fig. 126, w). It passes craniad near the dorsal
median line, lying at first dorsad of the aorta (b), then to the
right, then ventrad. It enters the substance of the liver in the
dorsal part of the caudate lobe of the latter, passes through the
liver, and then through the diaphragm near the ventrolateral
edge of the central tendon. It passes then craniad in the
thoracic cavity (Fig. 129, c) ventrad of the caudal lobe of the
right lung, and enters the right auricle.
It receives the following branches (Fig. 126): lumbar, iliolumbalis
(v), spermatica interna (u), renal (t), adrenolumbalis
(s), phrenic, and hepatic veins. All but the last two
named accompany the arteries of the same name.
V. phrenica.—The phrenic veins gather the blood from
the diaphragm and empty into the vena cava as it penetrates
the diaphragm.
V. hepatica.—The hepatic veins vary in number. They
gather the blood from the liver (sent in by the portal vein and
hepatic arteries) and enter the vena cava just caudad of the
diaphragm.
Vena Portæ. The Portal Vein
(Fig. 131).—The
portal vein is the large vein carrying the blood from the
abdominal viscera to the liver. Within the liver the portal
vein breaks up into capillaries; these collect to form the
hepatic veins, which enter the vena cava inferior. The portal
vein (a) is formed near the pyloric end of the stomach (1) by
the union of the superior mesenteric (b) and gastrosplenic (c)
veins and passes thence along the ventral border of the foramen
epiploicum (foramen of Winslow) to the liver (2), where it
divides, going to the lobes of the liver. On its way to the
liver it may receive the pancreaticoduodenalis (d), gastroepiploica
(e), and coronaria ventriculi (f). These may unite with
the portal separately, or any two or all three may unite to form
a single trunk before entering the portal vein.
1. V. coronaria ventriculi (f) gathers the blood from the
lesser curvature of the stomach and anastomoses with the
gastrosplenic veins. It usually empties into the portal vein
near the pylorus.
2. V. pancreaticoduodenalis (d) receives the blood from[327]
the pancreas and first part of the duodenum, and empties into
the vena portæ near to or with the preceding.
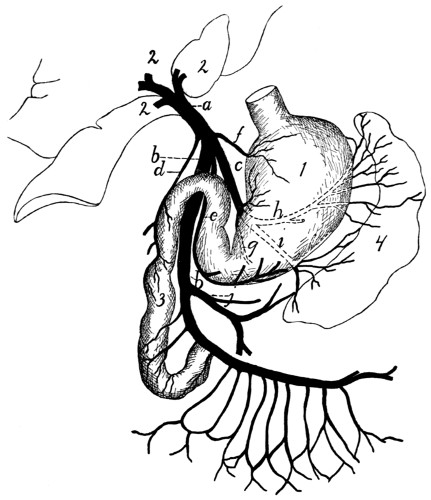
Fig. 132.—Portal Vein.
a, portal vein; b, superior mesenteric; c, gastrosplenic; d, pancreaticoduodenalis;
e, gastroepiploica; f, coronaria ventriculi; g, h, i, branches of gastrosplenic; j, inferior
mesenteric. 1, stomach; 2, liver; 3, duodenum; 4, spleen.
3. V. gastroepiploica (e) comes from the greater curvature
of the stomach and ascending limb of the great omentum. It
empties into the vena portæ ventrodextrad of the opening of
the coronaria ventriculi, or sometimes in common with it.
4. V. gastrolienalis (c).—The gastrosplenic is one of the
veins which unite to make up the portal vein. It is formed by
three branches: one (g) from the horizontal or gastric portion
of the pancreas, one (h) from the stomach and gastric end of
the spleen, the third and largest (i) from the caudal end of the
spleen and descending limb of the great omentum. The three[328]
branches unite and the common trunk joins the superior mesenteric
vein (b) to form the portal vein as above described.
5. V. mesenterica superior (b).—The superior mesenteric
vein receives the blood from the small and large intestines.
It is formed by numerous branches which follow and have the
same name as the corresponding arteries. It receives also the
inferior mesenteric (j) from the large intestine. The numerous
branches unite, and the main trunk thus formed joins the
gastrosplenic (c), and the two united become the portal
vein (a).
Vena iliaca communis. Common Iliac Vein (Fig.
126, w; Fig. 127, page 310).
The common iliac veins (Fig. 126, w) are the two large
vessels which unite in the sacral region to form the inferior
vena cava (a). Each is a large vessel, usually four or five
centimeters in length, which is formed by the junction of the
large external iliac vein with the smaller hypogastric vein
(z). The left common iliac usually receives the V. sacralis
media (o); other lateral branches are as a rule not received by
the common iliac veins.
There is much variation as to the formation of the common
iliac veins and their union to form the vena cava. On this subject
the paper by McClure, in the American Naturalist, vol.
XXXIV. pp. 135-198 (March, 1900), may be consulted with
profit. The more typical variations may be classified as follows:
(1) The two common iliac veins may be longer than
usual, so that the vena cava is formed farther craniad than
usual. (2) There may be in the abdominal region two separate
veins (venæ cardinales) representing the inferior vena
cava, these uniting in the region of the kidneys. (3) The
vena sacralis media may enter into the left common iliac (the
usual condition); or the right common iliac (as in Fig. 126);
or it may fork, one branch passing to the left, the other to the
right common iliac.
Branches of the common iliac veins:
1. V. sacralis media (Fig. 126, o).—This follows the
course of the corresponding artery and usually enters the left
common iliac.
[329]
2. V. hypogastrica (or iliaca interna) (z).—The hypogastric
or internal iliac vein joins the medial side of the
external iliac to form the common iliac (w). Its branches
follow the branches of the corresponding artery except that
the vein from the bladder (umbilical vein) joins the V. hemorrhoidalis
media. Also the vena glutea inferior receives a
large superficial communicating branch (Fig. 163, k′) from the
vena saphena parva. This branch leaves the saphena parva
(Fig. 163, k) at the popliteal space, and passes over the lateral
surface of the biceps muscle to its proximal end. Here it
passes between the biceps and caudofemoralis to join the vena
glutea inferior.
3. V. iliaca externa (Fig. 127, page 310).—This vein follows
the corresponding artery, collecting the blood from the
posterior extremity. Its branches are the same and have in
general the same distribution as those of the artery. The vein
has, however, certain branches in addition to those of the
artery, resulting in a somewhat different general arrangement
of vessels; it will be necessary therefore to give an account of
the system of veins from the foot and lower leg.
The blood is collected from the foot into two sets of veins
which may be distinguished as the deep and the superficial
sets. The veins of the deep set are those which accompany
the branches of the popliteal and anterior tibial arteries. Those
of the superficial set are larger and carry most of the blood.
On the dorsum of the foot blood is collected from the third,
fourth, and fifth digits by two or three superficial metatarsal
veins, which unite opposite the proximal end of the metatarsus
into a single vein. This receives a similar branch coming from
the dorsum of the second digit, and has numerous branches in
the ankle region which form here a network, communicating
with the vena saphena parva (Fig. 163, k), and with branches
from the plantar side of the foot. From this network two or
three main veins pass proximad on the medial side of the leg,
along with the branches of the arteria saphena, and unite with
a similar vein from the plantar side of the foot to form the vena
saphena magna (Fig. 127, k, page 310), which accompanies
the arteria saphena to join the femoral vein (a).
[330]
On the plantar side of the foot small veins pass from the
digits to a superficial venous plantar arch, which lies just
proximad of the fibrous pad on the sole of the foot. At its
medial side this arch extends proximad and dorsad to join that
branch of the vena saphena magna which comes from the
dorsum of the second digit. On the lateral side it extends
proximad along the lateral margin of the foot, sends a communicating
branch dorsad to the branches of the V. saphena
magna, and continues along the lateral border of the foot to
the ankle. It forms the beginning of the V. saphena parva
(Fig. 163, k, page 401). This receives branches from the
lateral surface of the ankle, which anastomose with those of
the saphena magna. The saphena parva then passes proximad
along the lateral surface of the leg, close to its ventral border,
to the popliteal space. At the distal end of the popliteal
space it divides into two branches. One passes inward
through the fat of the popliteal space, receiving branches from
the lymphatic gland situated here, and joins the popliteal vein.
The other (k′) remains superficial and continues proximad over
the lateral surface of the biceps to the proximal end of that
muscle. Here it passes between the biceps and caudofemoralis
to join the inferior gluteal vein.
IV. THE LYMPHATIC SYSTEM. SYSTEMA LYMPHATICUM.
The lymphatic system of the cat has not been worked out
in detail, so that only the main features of the system are given
in the following account.
The lymphatic system consists of a number of vessels, the
lymphatics, containing a colorless fluid called lymph, and of
lymphatic glands, connected with the lymphatic vessels.
The lymphatic vessels are found throughout the body as
slender tubes, frequently united into networks, and containing
many valves. They take origin from the spaces in the connective
tissue, so that they are at first without definite walls.
The fluid in the connective-tissue spaces gradually flows
together into tubes with definite walls, and these tubes, the
lymphatic vessels, finally join the venous system. In their[331]
passage through the lymphatic glands the vessels break up into
fine channels which again unite on leaving the gland. In the
gland lymph-cells are added to the fluid, and probably other
changes are produced.
Most or all of the lymphatic vessels of the body finally unite
into two trunks before entering the veins. The largest trunk
is the thoracic duct (Fig. 118,
5, page 282), which collects the
lymph from the entire caudal half of the body, and from the
left forelimb and the left side of the thorax, head, and neck.
It passes along the dorsosinistral side of the thoracic aorta,
lying against its surface, and enters the left external jugular
vein at its junction with the subclavian. The second main
trunk is the right lymphatic duct; this collects the lymph
from the right side of the thorax, the right forelimb, and the
right side of the neck and head; it enters the right external
jugular.
1. Lymphatics of the Head.
—A number of lymphatic
vessels arise on the sides of the face, especially in the upper
and lower lips. They form a superficial network of vessels,
lying over the ventral half of the masseter muscle and an area
ventrad of it. These lymphatic vessels all enter two large
lymphatic glands (Fig. 131, 12,
page 322) lying at the ventrocaudal
angle of the masseter muscle, covering the union of the
anterior and posterior facial veins. Into these glands pass also
a number of lymph vessels from the back of the head. From
these two lymphatic glands two or three small lymphatic
vessels pass caudad, lying on the surface of the external jugular
vein. Near the point of the shoulder, some distance craniad
of the junction of the external and internal jugular veins, these
vessels enter another small lymphatic gland lying on the dorsal
surface of the external jugular vein. Thence one or two
vessels continue caudad, still on the surface of the external
jugular, and finally unite with the deep lymphatics of the head
and neck and those of the arm to enter the caudal end of the
external jugular vein, usually (on the left side) after junction
with the thoracic duct, from the thorax.
The deep lymphatics of the head come from the internal parts
of the head,—tongue, pharynx, etc., and enter a large lymphatic[332]
gland situated close to the caudoventral surface of the tympanic
bulla, at the side of the pharynx. Thence a large lymphatic
trunk passes caudad at the side of the trachea, along with the
common carotid artery and internal jugular vein, unites with
the superficial vessels from the head, above described, joins
(on the left side) the thoracic duct, and enters the external
jugular vein.
2. Lymphatics of the Neck.
—Two or three lymph-glands
are found in the fatty mass in the neck beneath the clavotrapezius
and levator scapulæ ventralis muscles. These receive
lymphatics from the neck and side of the thorax. The vessels
which arise from them join those from the head and arm to
enter the external jugular vein.
3. Lymphatics of the Thoracic Limbs.
—The lymphatics
of the forelimbs begin as a number of vessels on the dorsum
and palm of the hand. These all pass toward the radial side,
and at about the base of the first metacarpal the vessels from
both sides of the hand unite to form two or three main trunks.
These pass towards the elbow, accompanying the vena cephalica
(Fig. 130). There are usually two of these main trunks,
one lying on each side of the vena cephalica. They accompany
this vein over the shoulder, following that portion which finally
joins the external jugular. In the hollow of the shoulder they
enter the cervical lymph-glands, one or two lymph-glands
imbedded in the fat lying beneath the clavotrapezius and
levator scapulæ ventralis in this region. From the cervical
lymph-glands a single trunk passes mediocaudad, unites with
the lymphatics from the head, and usually (on the left side)
with the cranial end of the thoracic duct, and enters the
external jugular vein at its junction with the subclavian. The
common trunk from the head and arm may enter the jugular
separately, but close to the termination of the thoracic duct.
On the right side the common trunk is joined by one or more
small vessels coming from the lymphatic glands of the thorax
and forming the right lymphatic duct.
There is said to be also a deep system of lymphatics in the
arm, in addition to the superficial system above described; this
is said to accompany the branches of the brachial vein. If this[333]
system is present in the cat, it is much less easily demonstrated
than the superficial system.
4. Lymphatics of the Thorax and Abdomen.
—In the
thorax are a number of lymphatic glands which are apparently
not constant in number and position. One is commonly found
dorsad of the bifurcation of the superior vena cava, one ventrad
of the mammary vein, several small ones about the bifurcation
of the trachea, one or more in the region of the aortic arch,
and a number of small ones scattered in the mediastinum.
The vessels from these glands finally join either the right lymphatic
duct or the thoracic duct; the details have not been
worked out and are probably variable.
In the abdominal cavity a number of mesenteric glands, of
considerable size, are found in the mesentery and in the mesocolon.
In the mesocolon these are usually separate, forming
a chain of glands following the colon about one and a half
centimeters from it. In the mesentery the glands are mostly
united into a very large one, formerly known as the pancreas
aselli; this is the largest lymph-gland in the body. It is a
curved structure, four or five centimeters in length, lying in
the central region of the mesentery.
Into the mesenteric glands pass lymphatic vessels from the
viscera of the abdomen. From the mesenteric glands one or
two large lymphatic vessels pass craniodorsad, at first with the
portal vein, then dorsad of it. This vessel reaches the dorsal
side of the aorta near the cranial end of the kidney, where it
enters a large fusiform vessel, the receptaculum chyli, which
stretches from the cranial end of the kidney craniad between
the crura of the diaphragm into the thorax. The receptaculum
receives other lymphatics from the various organs of the
abdominal cavity as well as vessels coming from the pelvic
region, and from the iliac glands. These lie beside the iliac
artery, receive lymphatics from the hind limbs, and send lymphatics
craniad to the receptaculum chyli. The latter forms
the beginning of the thoracic duct.
The thoracic duct (Fig. 118, 5, page 282) enters the
thoracic cavity between the crura of the diaphragm as a continuation
of the receptaculum chyli. It lies on the dorsal side[334]
of the aorta (a), somewhat to the left. It passes along the
aorta, gradually coming to lie more on its left side, and finally
leaves that vessel near the aortic arch. The duct continues
thence straight craniad along the left side of the œsophagus till
it reaches the external jugular vein at its junction with the subclavian.
In this region the thoracic duct usually unites with
the lymphatics coming from the head and arm, then enters the
external jugular vein, or the innominate at the point of union
of external jugular and subclavian.
In its passage through the thorax the duct frequently
divides into two or three divisions which pass along parallel to
each other for a distance, then reunite. Such division into two
or three parts is especially frequent just before the union with
the jugular vein. The numerous valves in the thoracic duct
give it a beaded appearance.
5. Lymphatics of the Pelvic Limbs.
—A number of lymphatic
vessels are formed on the dorsum and the sole of the
foot. These pass to the lateral side, forming a network on the
lateral and dorsal surface of the ankle-joint, and finally unite
into two main trunks which accompany the vena saphena parva.
In the popliteal space these enter the popliteal gland which
lies imbedded in the fat of that region. From the popliteal
gland one or two trunks accompany the superficial division of
the vena saphena parva across the lateral surface of the biceps
muscle, and finally thus reach the pelvic cavity. Here they
join the lymphatics of the pelvic region, which all pour their
lymph at last into the thoracic duct. Inguinal glands are
either very small or not present in the cat.
The pelvic limbs have perhaps a deep system of lymphatics,
accompanying the deep veins; if so, they are not easily
demonstrable.
[335]

Fig. 133.—Cranial Portion
of the Spinal Cord, with
the Cervical Enlargement.
Dorsal View.
1-8, the eight cervical
nerves. I-II, first two thoracic
nerves. a, dorsal roots (ventral
roots not shown); b, spinal
ganglia; c, dorsal rami; d, ventral
rami; e, posterior median
sulcus; f, posterior lateral sulcus.
NERVOUS SYSTEM.
I. THE CENTRAL NERVOUS SYSTEM.
1. The Spinal Cord. Medulla spinalis
(Figs. 133-136).—The
spinal cord is that portion of the nervous system which
occupies the vertebral canal; it is continuous craniad with the
brain. It has the form of a somewhat flattened cylinder and
extends from the foramen magnum into the caudal region. It
diminishes in diameter after entering the sacral region.
The cord has a cervical and a lumbar enlargement, the
former (Fig. 133) marking the origin of the nerves which pass
to the fore limb, and the latter (Fig. 136) the origin of those
which pass to the hind limb.
The cervical enlargement (Fig. 133) lies in that part of the
vertebral canal bounded by the fourth to seventh cervical and
first thoracic vertebræ. Caudad of the first thoracic vertebra
the cord continues of nearly uniform diameter to the lumbar
enlargement (Fig. 136) which stretches from the third to the
seventh lumbar vertebræ (inclusive). Caudad of the seventh
lumbar vertebra it diminishes uniformly in diameter and ends
in a slender cord, the filum terminale (Fig. 136, C), which
may be traced into the caudal region.
The surface of the cord is marked by a number of longitudinal
grooves or sulci, and fissures. The most prominent
of these is the anterior median fissure (Fig. 134, c), along
the ventral median line; into this the pia mater dips. The
posterior median sulcus (Fig. 133, e; Fig. 134, a) is a shallow
furrow along the dorsal median line. The anterior fissure and
the posterior sulcus thus divide the cord into lateral halves.[336]
Each half is subdivided by the anterior and posterior lateral
sulci.
The posterior lateral sulcus (Fig.
133, f; Fig. 134, b) lies at the side of
the posterior median sulcus. It is
broad and shallow and has the posterior
roots of the spinal nerves emerging
from its bottom.
The anterior lateral sulcus exists
only after the forcible pulling out of
the anterior (ventral) roots of the
spinal nerves. It then marks the line
along which they originate from the
cord.
In cross-section (Fig. 134) the
substance of the cord appears to the
naked eye as composed of a darker
central “gray matter” (g) and of an
outer “white matter” (f). The
gray matter has in section the form of
the letter H. The cord is really
tubular, having a central canal (h)
which appears in section in the middle
of the cross-bar of the H, while the
two ends of each vertical bar extend
toward the anterior and posterior
lateral fissures.
The white matter is divided into
funiculi (or columns) by the fissures
and sulci, so that there is in each half
of the cord an anterior, a posterior,
and a lateral white funiculus.
In the cervical region (Fig. 134)
there appears between the posterior
lateral and posterior median sulci an
intermediate sulcus which divides the
posterior funiculus in this region into two. The median
slenderer of these is the fasciculus gracilis (d) or column of[337]
Goll. The lateral and thicker is the fasciculus cuneatus (e)
or column of Burdach.
The membranes of the spinal cord:
The dura mater is a tough fibrous membrane directly continuous
with the dura mater of the brain. It
differs from the dura mater of the brain in
two particulars:
1. At the foramen magnum it splits into
two layers, one of which lines the bony
vertebral canal and forms its periosteum,
while the other covers the cord.
2. It is separated from the cord by a considerable
space. The dura mater is continuous
with the fibrous sheaths of the spinal
nerves at their points of exit. Along the
sides of the cord it is connected to the pia
mater by a delicate strand of connective tissue
probably equivalent to the “ligamentum
denticulatum” of man.
The arachnoid is a delicate cellular membrane
lying beneath the dura mater, between it and the pia
mater. It forms a continuous investment for the cord, is not
vascular, and is said not to dip into the fissures of the cord.

Fig. 134.—Cross-section
of the
Spinal Cord in
the Cervical Region.
a, posterior median
sulcus; b, posterior
lateral sulcus; c, anterior
median fissure;
d, fasciculus gracilis;
e, fasciculus cuneatus;
f, white matter; g,
gray matter; h, central
canal.
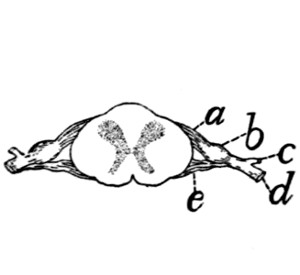
Fig. 135.—Section of
Spinal Cord, showing
the Origin of a Pair
of Spinal Nerves.
a, dorsal root; b, spinal
ganglion; c, dorsal ramus;
d, ventral ramus; e, ventral
root.
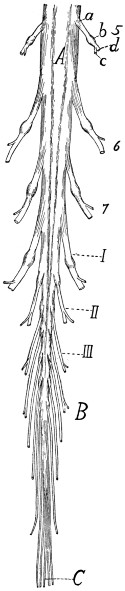
Fig. 136.—Caudal Portion
of the Spinal Cord.
A, lumbar enlargement; B,
cauda equina; C, filum terminale;
5-7, fifth to seventh lumbar
nerves. I-III, the three
sacral nerves. a, dorsal roots;
b, spinal ganglia; c, dorsal
rami; d, ventral rami.
The pia mater invests the cord closely and contains some
blood-vessels. It is a delicate membrane which dips into the
fissures and sulci of the cord and is connected
to it by numerous strands of connective
tissue that pass from it into the
substance of the cord. The nerves pierce
it.
Spinal Nerves.—From the spinal cord
arise the spinal nerves. Of these there
are about thirty-eight pairs in the cat.
Eight are cervical, thirteen thoracic,
seven lumbar, three sacral, and seven or eight caudal. Those
leaving the cervical (Fig. 133,
5-8 and I) and lumbar (Fig.
136) enlargements are larger than the others. The first
cervical nerve leaves the vertebral canal through the atlantal[338]
foramen, the second leaves between the
arches of the atlas and axis, while all
the others leave the vertebral canal by
way of the intervertebral foramina.
Each nerve arises from the cord by a
dorsal and a ventral root. The dorsal
root is chiefly sensory, the ventral motor
in character. The dorsal root (radix
posterior) (Figs. 133, 135, and 136, a)
begins as a number (twelve or more)
of separate nerve-bundles which emerge
from the posterior lateral groove. These
roots lie nearly in a single plane and
pass laterad, converging to penetrate a
ganglion, the spinal ganglion (b) (or
ganglion of the posterior root). All
the spinal ganglia except the first
and second are situated in the intervertebral
foramina or within the vertebral
canal. The first and second are
situated among the muscles surrounding
the place of exit of the nerves.
The ventral root (radix anterior)
(Fig. 135, e) arises as a larger number of
small fibre-bundles which do not lie in
a single plane, so that in a transverse
section several rootlets may appear in a
single section. The rootlets converge
to form a single mass which joins the
dorsal root just as it emerges from the
spinal ganglion. The nerve formed by
the junction of the ventral and dorsal
roots is one of the spinal nerves.
The direction in which the nerves
leave the cord varies. In the cervical
region (Fig. 133) and cranial part of
the lumbar region it is nearly laterad;
at the cervical and lumbar enlargements[339]
it is laterocaudad. The nerves in the sacral and caudal regions
pass almost directly caudad to reach the intervertebral foramina
and form thus a brush which surrounds the filum terminale
(Fig. 136, C) and is called the cauda equina (Fig. 136, B).
Each spinal nerve immediately after leaving the intervertebral
foramen divides into two branches, a dorsal or posterior
branch or ramus (c), and a ventral or anterior ramus (d).
The dorsal ramus is in each case small (except in the first and
second cervical nerves), and is distributed to the longitudinal
muscles and integument of the back.
The ventral rami are larger and each is connected a short
distance beyond its origin with the sympathetic system by a
short ramus communicans or communicating branch. Each
is then distributed to the integument and muscles of the ventral
part of the body, including the limbs.
The ventral rami which pass to the limbs are much larger
than the others. The ventral rami are further distinguished
from the dorsal by the fact that they frequently unite with one
another to form plexuses.
The peripheral distribution of the spinal nerves is described
later.
2. The Brain. Encephalon.
—The brain is that portion of
the central nervous system that is included within the cranial
cavity. It is a direct continuation of the spinal cord, and
presents many of the same essential characters as the latter,
with great modifications in details.
The structure of the brain can best be understood if it be
considered as a modified continuation of the spinal cord, and
the relation of the parts to the essential parts of the cord noted.
The spinal cord is a nearly straight tube, with a central cavity
and thick walls. The brain is likewise tubular, with the
cavities enlarged or subdivided in places, with the walls greatly
thickened, and with a number of bends and constrictions in the
tube. The relation of the structure of the brain to that of the
spinal cord is most easily perceived by an examination of the
brain of some lower vertebrate, as the frog or shark, and such
an examination should be made before proceeding to the study
of the more complicated brain of the cat.
[340]
In the following account of the brain of the cat all parts
will be described as far as possible in relation with the tubular
structure of the brain. The brain will be considered as a
hollow structure, having central cavities, and the solid portions
will be brought into relation as parts of the roof, sides, or floor
of the cavities. The cavities of the brain, forming a direct
continuation of the central canal of the spinal cord, are known
as ventricles (ventriculi).
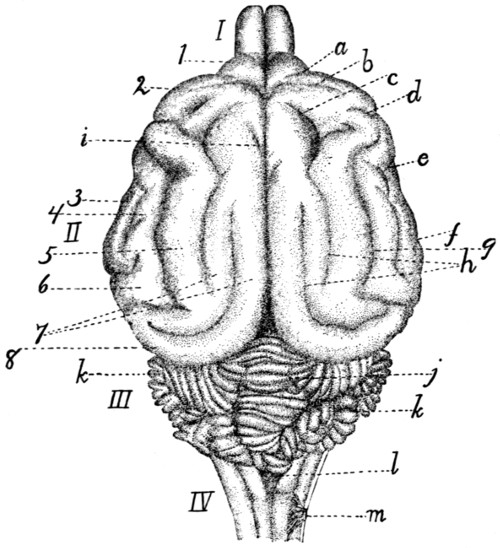
Fig. 137.—Dorsal Surface of the Brain.
I, olfactory bulbs; II, cerebral hemispheres;
III, cerebellum; IV, medulla. a,
præsylvian sulcus; b, cruciate sulcus; c, sulcus ansatus; d, coronal sulcus; e, anterior
sulcus; f, posterior sulcus; g, suprasylvian sulcus; h, separate parts of the lateral
sulcus; i, end of splenial sulcus; j, vermis of cerebellum; k, hemispheres of cerebellum;
l, roof of fourth ventricle; m, first cervical nerve. 1, orbital lobe (or gyrus);
2, sigmoid gyrus; 3, anterior sylvian gyrus; 4, ectosylvian gyrus; 5, suprasylvian
gyrus; 6, posterior sylvian gyrus; 7, parts of marginal gyrus; 8, gyrus compositus
posterior.
In a general view of the cat’s brain from the dorsal side
(Fig. 137) four subdivisions are discernible. At the caudal
end is a small stalk-like portion (IV) which is clearly a slightly
modified continuation of the spinal cord; this is the myelencephalon
or medulla oblongata. Just craniad of this, rising[341]
high above it, is an irregularly lobulated rounded portion, the
cerebellum (III). This is produced as a thickening of the wall
of the original tube; it forms the dorsal part of the metencephalon.
These two portions of the brain have a common
cavity, lying within the medulla oblongata and ventrad of the
cerebellum, and formed by a widening of the cavity of the
spinal cord; it is known as the fourth ventricle. Craniad of
the cerebellum, separated from it by a deep transverse fissure,
are two very large portions, the hemispheres of the cerebrum
(II), separated from each other by a deep longitudinal fissure;
they constitute together the telencephalon. The hemispheres
are produced by a sort of forking of the original central tube;—by
outgrowths on the sides of the tube, the central cavity
extending into the outgrowths. The two outgrowths extend
dorsad, as well as craniad and caudad from the original place
of origin, forming the larger part of the brain; the extensions
of the central cavity which they contain are known as the
lateral ventricles. Finally, in front of the two hemispheres
are seen the two small olfactory bulbs (I) which are mere
extensions of the two hemispheres and contains cavities which
are extensions of the lateral ventricles.
In a ventral view of the entire brain (Fig. 138) certain
parts are visible which in the dorsal view are hidden by the
large cerebral hemispheres. At the caudal end is seen, as
before, the medulla oblongata, or myelencephalon, and craniad
of this, on the ventral side of the cerebellum, a broad transverse
tract, the pons (i); this, like the cerebellum, forms part
of the metencephalon. All this part of the brain formed by
the myelencephalon and metencephalon lies caudad of
(behind) the rest of the brain and is marked off from it by a
great fissure; it therefore receives as a whole the name hindbrain
or rhombencephalon.
Just craniad of the pons are seen two short diverging arm-like
bands of fibres (g), enclosing a small triangular space
between them. These two arms with the space between them
are all that is visible of a division of the brain which in dorsal
view is completely covered by the backward projection of the
cerebral hemispheres. This is the midbrain or mesencephalon.[342]
In this portion of the brain the continuation of the central
cavity is a narrow canal which receives the name cerebral
aqueduct (aqueductus cerebri) or aqueduct of Sylvius.
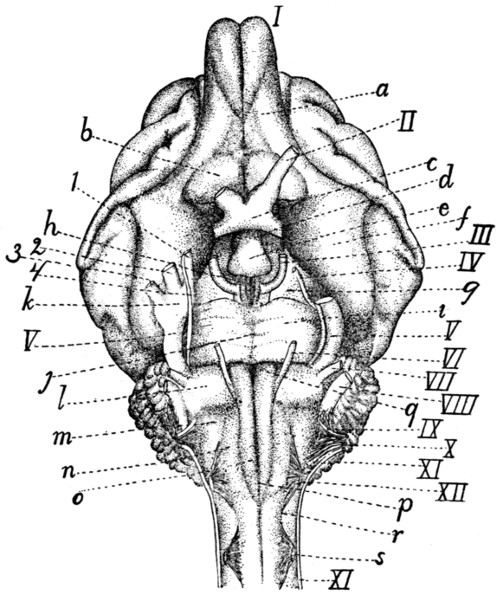
Fig. 138.—Ventral Surface of the Brain.
I-XII, the twelve cranial nerves in order;
I, olfactory bulb; II, N. opticus; III,
N. oculomotorius; IV, N. trochlearis; V, N. trigeminus (1, mandibular nerve; 2,
maxillary nerve; 3, ophthalmic nerve; 4, sensory root); VI, N. abducens; VII, N.
facialis; VIII, N. acusticus; IX, N. glossopharyngeus; X, N. vagus; XI, N. accessorius;
XII, N. hypoglossus. a, tractus olfactorius; b, substantia perforata anterior;
c, optic chiasma; d, infundibulum; e, hypophysis; f, lobus pyriformis, or tractus
postrhinalis; g, pedunculi cerebri; h, substantia perforata posterior; i, pons; j, sulcus
basilaris of pons; k, semilunar (or Gasserian) ganglion; l, trapezium; m, area ovalis;
n, area elliptica; o, pyramidal tracts; p, anterior or ventral fissure; q, foramen
cæcum; r, anterior lateral sulcus; s, first cervical nerve.
Just craniad of the midbrain is a small rather irregular area,
bounded craniad by the band-like optic chiasma (c), from
which arise the optic nerves (II), and showing caudad of this
band a rounded irregular projection (e). This is a fourth division
of the brain which is likewise covered dorsad by the
hemispheres; it is the ’tween-brain or diencephalon. Its
cavity is known as the third ventricle. Just craniad of the[343]
’tween-brain, in the median line, is a deep fissure, showing that
here in reality is the termination of the original tube, the
further extension craniad being due to the pushing forward of
the lateral outgrowths, or cerebral hemispheres, which extend
in ventral view a considerable distance further craniad. The
hemispheres lie also caudad, laterad, and dorsad of the midbrain
and ’tween-brain, so that these parts are almost enveloped
by the hemispheres. The hemispheres and ’tween-brain are
sometimes taken together as forming a single portion, the forebrain
or prosencephalon.
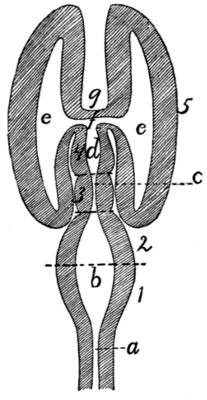
Fig. 139.—Diagram of Brain
from Above.
Fig. 140.—Diagram of Brain from
the Side.
1, myelencephalon; 2, metencephalon; 3, mesencephalon; 4, diencephalon; 5,
telencephalon. a, canal of the spinal cord; b, fourth ventricle; c, cerebral aqueduct
(aqueduct of Sylvius); d, third ventricle; e, lateral ventricle; f, interventricular foramen
(or foramen of Monroe); g, lamina terminalis; h, olfactory bulb.
There are thus altogether five principal subdivisions of the
brain, each enclosing a portion of the central cavity. These
are the myelencephalon, the metencephalon (these two enclosing
the fourth ventricle), the mesencephalon (enclosing the
cerebral aqueduct), the diencephalon
(enclosing the third ventricle), and
the two cerebral hemispheres, constituting
together the telencephalon,
and enclosing the two lateral ventricles.
A plan of the brain, considered
as a tube enclosing cavities,
is given in Figs. 139 and 140. Fig.
139 shows the plan considered as
viewed from the dorsal side, Fig. 140 in a lateral view. These
figures are of course pure diagrams, showing only what may
be called the plan of structure, and omitting all details. In
the actual conditions many important modifications even of the
main features of the plan are met with.
[344]
In the following description the parts of the brain will be
taken up in this order:
- A. Rhombencephalon (primitive hindbrain).
- 1. Myelencephalon.
- 2. Metencephalon.
- B. Mesencephalon (primitive midbrain).
- 3. Mesencephalon.
- C. Prosencephalon (primitive forebrain).
- 4. Diencephalon.
- 5. Telencephalon.
A. Rhombencephalon.—
1. Myelencephalon.
—The myelencephalon
or medulla oblongata is transitional between the
spinal cord and the brain. It has in general the structural
peculiarities of the cord, but these undergo in it a gradual transformation
into the arrangements characteristic of the brain.
It has the form of a flattened and truncated cone, which
widens craniad and is limited at the cranial end by the pons
(Fig. 138, i) ventrally and laterally; by the cerebellum (Fig.
137, III) dorsally. The cranial portion of its dorsal surface
is overhung by the cerebellum. The points of origin of the
roots of the first pair of cervical nerves (Fig. 138, s) may be
taken as indicating the boundary between the spinal cord and
the medulla oblongata; there is no other external marking to
show the limits of the two.
The central canal of that part of the medulla which is
overhung by the cerebellum is greatly widened (Fig. 141, h).
The cavity thus formed is flattened and triangular, with the
apex of the triangle caudad, and is the caudal part of the
fourth ventricle (Fig. 141, h). The roof is very thin (Fig.
143, n) and is intimately connected with the pia mater, so that
in removing the latter the thin roof of the fourth ventricle is
removed with it and the cavity of the fourth ventricle is left
exposed. The thin roof of the fourth ventricle is known as the
velum medullare posterius (Fig. 143, n).
The ventral (anterior) fissure (Fig. 138, p) of the cord
passes onto the medulla oblongata and ends at the pons (i),
its end being slightly deeper and forming what is known as the[345]
foramen cæcum (q). The dorsal (posterior) sulcus (Fig. 141,
a) is well marked; it ends at the caudal boundary of the fourth
ventricle (h).
The anterior lateral sulcus (Fig. 138, r) may be traced
craniad from the origin of the ventral roots of the first cervical
nerves (s), along the lateral border of the area elliptica (n),
then curving mediad to the lateral border of the pyramis (o),
and finally reaching the pons (i). Its position is marked by
the origin of the roots of the hypoglossal nerve (XII).
The posterior lateral sulcus (Fig. 141, b), marked on the
spinal cord by the origin of the dorsal nerve-roots, curves
laterad at the sides of the fourth ventricle (h) owing to the
increasing width of the latter, and ends at an elevated area of
oblique fibres, the area ovalis (f).
The columns or funiculi bounded by the longitudinal
fissures present the following peculiarities:
The anterior white funiculus of the cord is replaced in the
medulla by the pyramidal tracts (pyramides) (Fig. 138, o).
The pyramidal tracts are formed by fibres which emerge from
beneath the pons and pass caudad to disappear just craniad of
the level of the first cervical nerve (s). The pyramidal tracts
are bounded medially by the anterior median fissure (p), but
laterally each is separated from the anterior lateral sulcus (r)
over its caudal part by an elongated area elliptica (n), the
human homologue of which is uncertain. It perhaps represents
the oliva.
Laterad of the cranial portion of the pyramids is an irregular
area known as the trapezium (l) which abuts caudad on
the area elliptica (n) and the area ovalis (m). The area ovalis
(Fig. 138, m; Fig. 141, f) (or zonula Arnoldi) is abroad band
of oblique fibres which passes from the lateral side of the area
elliptica craniodorsad to disappear under the cerebellum.
The posterior white funiculus was divided in the cervical
region into two, the fasciculus gracilis (column of Goll) and
the fasciculus cuneatus (column of Burdach). The fasciculus
gracilis (Fig. 141, c) extends to the caudal end of the fourth
ventricle (h) and ends there in an expansion, the clava (d),
which forms the posterior boundary of the fourth ventricle.[346]
The fasciculus cuneatus (e) passes laterad on account of the
width of the fourth ventricle and appears to end at the area
ovalis (f), but it may be seen passing beneath the area ovalis,
emerging at its cranial border and turning dorsad to enter the
cerebellum. It forms the side walls of a part of the shallow
fourth ventricle.
The lateral funiculi are divided by longitudinal furrows into
three divisions. The dorsal one of these is the fasciculus
cuneatus lateralis or column of Rolando (Fig. 141, g). It
accompanies the fasciculus cuneatus into the cerebellum.
The entire mass formed by the fasciculus gracilis, the
fasciculus cuneatus medialis, and fasciculus cuneatus lateralis is
known as the corpus restiforme, and since its fibres pass into
the cerebellum it is sometimes known as the pedunculus cerebelli
(or crus cerebelli ad medullam).
The following cranial nerves arise from the medulla oblongata
(Fig. 138).
The twelfth nerve (XII) (N. hypoglossus) arises by ten or
fifteen rootlets from the ventral surface of the medulla oblongata
in the anterior lateral fissure (r) opposite the caudal portion
of the area elliptica (n).
The eleventh cranial nerve (XI) (N. accessorius) arises by
numerous rootlets from the lateral surface of the medulla
oblongata and of the spinal cord as far caudad as the sixth or
seventh cervical nerve. These rootlets join to form a nerve
which enters the cranium through the foramen magnum and is
closely associated at its point of exit with the glossopharyngeal
(IX) and vagus (X) nerves. The line of origin on the medulla
passes between the dorsal and ventral roots of the cervical
nerves and is along the middle of the lateral white funiculi.
The tenth cranial nerve (X) (N. vagus) arises by about
eighteen very delicate rootlets from the surface of the area
ovalis. The rootlets are divided into a dorsal and a ventral
series. The dorsal series (about twelve) arise in the groove
which separates the fasciculus cuneatus medialis from the
fasciculus cuneatus lateralis. The ventral series arise somewhat
ventrad of this groove. These rootlets are to be distinguished
from those of the ninth nerve by their smaller size.
[347]
The ninth cranial nerve (IX) (N. glossopharyngeus) arises
from the area ovalis from a line craniad of the dorsal line of
origin of the vagus roots (X) and between these and those of
the auditory (VIII). It arises by a number of rootlets which
are larger than those of the vagus (X), with which this nerve
is closely associated.
The eighth cranial nerve (VIII) (N. acusticus) appears at
the lateral end of the trapezium (l). It arises from an elevation
(Fig. 141, i) which is continued dorsomediad along the cranial
border of the area ovalis.
The seventh cranial nerve (VII) (N. facialis) leaves the
lateral border of the trapezium (l) near its cranial edge,
between the fifth and eighth nerves. It is much smaller than
the eighth nerve.
The sixth cranial nerve (VI) (N. abducens) arises by about
six bundles from the groove between the pyramids and the
trapezii and passes craniad.
2. Metencephalon.
—The metencephalon includes the pons
and the cerebellum.
The pons (Fig. 138, i) is a mass of transverse fibres which
forms the ventral and cranial part of the primitive hindbrain.
It is a modification of the latter brought about by the development
of the cerebellum, and the degree of its development is in
direct ratio to that of the cerebellar and cerebral hemispheres.
The pons forms a projecting mass of fibres which is marked by
a median longitudinal groove, the sulcus basilaris (j), which
indicates the course of the basilar artery (Fig. 121, c). Laterad
the fibres of the pons converge somewhat and turning dorsad
disappear in the cerebellum, forming the brachia pontis (Fig.
141, l).
The fifth cranial nerve (Fig. 138, V) (N. trigeminus) arises
by two roots from the caudal border of the pons, near the
lateral end. The ventral root (4) is small; the dorsal one is
much larger and soon forms the large semilunar ganglion (k)
from which three branches (1, 2, and 3) diverge. The ventral
root (4) joins one of these branches (1).
The cerebellum (Fig. 137, III) is formed by an increase in
size of the cranial portion of the primitive hindbrain. This[348]
increase has affected principally the surface of the roof, so that
as the cerebellum has grown it has been thrown into many
folds, the exact form of which varies in different specimens.
The cerebellum has at the same time increased in size and has
thus extended laterad as well as caudad and craniad. It thus
touches the cerebrum in front (separated from it by the tentorium)
and aids it in concealing the midbrain and ’tween-brain
in dorsal view, while caudad in the same view it conceals the
greater part of the medulla. The connections of the cerebellum
with adjacent parts of the brain are also overhung and concealed.
The whole surface of the cerebellum is thrown up into
numerous folds or gyri, separated from one another by deep
fissures or sulci, which appear at first to render the surface
wholly irregular. The entire mass is, however, divisible into
a central portion, which from its resemblance to a segmented
worm is called the vermis (j) (its cranial part is the superior
vermis, and its caudal part the inferior vermis), and into lateral
portions, the hemispheres (k). The vermis (j) occupies a
median longitudinal position, and its gyri and sulci are in the
main transverse. It is not directly connected with adjacent
parts, and its ventral surface extends farther caudad and craniad
than that of the hemispheres. The ventral part of the superior
vermis is fitted against the posterior corpora quadrigemina.
The hemispheres (k) may again be subdivided into groups
of gyri which have received special names. One of these, the
so-called appendicular lobe, fits into the appendicular fossa of
the petrous bone.
The cerebellum is connected to the adjacent parts by three
tracts of fibres, sometimes known as crura cerebelli. The tract
connecting it with the medulla oblongata is the corpus restiforme;
that connecting it with the pons is the brachium
pontis (Fig. 141, l); these have been described. A third
tract passes craniad to the corpora quadrigemina (Fig. 141, p
and q); this is the brachium conjunctivum (Fig. 141, k).
The cerebellum is composed of white and gray matter, the
latter on the surface (Fig. 143, III). The folds of its surface
present thus a contrivance for increasing the amount of gray[349]
matter. The white matter forms a central mass from which
tracts extend into the folds. The whole mass of white matter
has thus in section (more particularly in a longitudinal section
of the vermis) the appearance of a tree, whence the name
arbor vitæ (Fig. 143).
The fourth ventricle (Fig. 141, h; Fig. 143, m) is the
cavity of the original hindbrain. It begins caudad at the clava
(Fig. 141, d) as a widening and continuation of the central
canal of the spinal cord and extends craniad, becoming wider
and passing ventrad of the cerebellum. It becomes narrower
craniad and ends at the posterior corpora quadrigemina (p),
where it becomes continuous with the aqueductus cerebri (Fig.
141, o; Fig. 143, j). The cavity is shallow and is encroached
upon dorsally by the vermis of the cerebellum (Fig. 143).
The floor of the cavity is known as the fossa rhomboidea
(Fig. 141, h). It is formed by the continuation of the gray
matter which surrounds the central canal of the cord. It is
marked by a median longitudinal groove. At its widest part
are seen two considerable tracts of white fibres (striæ medullares)
which pass from near the median line laterad and extend
into the auditory (eighth) nerves. The floor caudad and
craniad of these striæ is marked by a number of elevations and
depressions. Similarly situated elevations in the human brain
differ from one another slightly in color, are made up of gray
matter, and are the centres of origin of most of the cranial
nerves. Their homologues in the cat appear not to have been
determined.
The side walls of the fourth ventricle (Fig. 141) are formed
by the following in order, beginning caudad: the clava (d),
the corpus restiforme, brachium pontis (l), brachium conjunctivum
(k), and caudal corpora quadrigemina (p) (colliculi
inferiores).
The roof (Fig. 143) is formed caudad by a thin layer of
non-nervous matter which is closely associated with the pia
mater. This thin layer is known as the velum medullare
posterius (n). It connects the dorsal surface of the medulla
with the caudal border of the cerebellum. The pia mater
covering this portion of the roof is vascular and is folded in[350]
toward the floor of the ventricle, forming the choroid plexus
of the fourth ventricle. In the middle the roof of the fourth
ventricle is the cerebellum, while craniad the roof is the velum
medullare anterius (Fig. 143, l). This is a thin layer just
craniad of the cerebellum, connecting it with the corpora quadrigemina,
and attached laterally to the brachia conjunctiva.
Here the fourth ventricle narrows craniad and becomes continuous
with the slender aqueductus cerebri (Fig. 143, j)
(aqueduct of Sylvius). The narrowed portion of the brain is
frequently known as the isthmus rhombencephali.
Fig. 141.—Dorsal
Surface of Myelencephalon, Mesencephalon, and
Diencephalon.
The cerebellum and the greater part of the cerebral hemispheres have been removed.
a, posterior sulcus of cord; b, posterior lateral sulcus; c, fasciculus gracilis;
d, clava; e, fasciculus cuneatus; f, area ovalis; g, fasciculus cuneatus lateralis; h,
fossa rhomboidea or floor of fourth ventricle; i, projection formed by origin of auditory
nerve; j, facial nerve; k, cut end of brachium conjunctivum; l, cut end of brachium
pontis (l′, brachium pontis); m, velum medullare anterius; n, fourth nerve;
o, depression marking caudal end of aqueductus cerebri (aqueduct of Sylvius); p,
caudal corpora quadrigemina (colliculi posteriores); q, cranial corpora quadrigemina
(colliculi anteriores); r, brachium quadrigeminum inferius; s, corpus geniculatum
mediale; t, thalamus; u, striæ medullares; v, third ventricle; w, pulvinar; x, corpus
geniculatum laterale; y, corpus striatum; z, outlines of olfactory bulbs. 1, boundary
between hemispheres and ’tween-brain; 2, pineal body.
The fourth cranial nerve (Fig. 141, n; Fig. 138, IV)
(N. trochlearis) arises from the brain at the craniolateral angle
of the velum medullare anterius.
[351]
B. Mesencephalon.—
3. Mesencephalon.
—The mesencephalon
or midbrain includes the corpora quadrigemina (Fig.
141) and the pedunculi cerebri (Fig. 142). In the primary
midbrain there is a pronounced thickening of the walls accompanied
by a reduction of the central canal. The midbrain does
not thus become very large and is concealed in the dorsal view
by the cerebellum and cerebrum, though its floor appears in
the ventral view just craniad of the pons (Fig. 138, g). Its
narrow canal is the aqueductus cerebri (aqueduct of Sylvius)
(Fig. 143, j). Its roof forms the corpora quadrigemina, and
its floor the pedunculi cerebri.
In a dorsal view (Fig. 141) the roof is seen to be marked
by two pairs of elevations, the corpora quadrigemina (p and q).
The cranial pair (q) (known as the colliculi superiores) are circular
in outline, surrounded on all sides except the cranial one
by a deep groove. From the cranial side a tract of fibres
(brachium quadrigeminum superius, or arm of the cranial corpus)
extends craniad and disappears beneath the thalamus (t).
Between the anterior or cranial corpora quadrigemina lies the
pineal body or epiphysis (corpus pineale) (Fig. 143, y; Fig.
141, 2), a portion of the roof of the ’tween-brain. The caudal
corpora (Fig. 141, p) are larger than the cranial ones, and
ovoid in shape with the long axis vertical. They are united
in the median line, and the velum medullare anterius (m)
stretches between their caudal borders. The brachium of the
caudal corpus quadrigeminum (brachium quadrigeminum inferius)
(r) extends craniad and disappears beneath a considerable
elevation, the corpus geniculatum mediale (s).
Crossing this brachium is seen a small tract of fibres which
extends ventrad, crosses the pedunculus cerebri, and reaches
the medial border of the latter. It is the tractus transversus
peduncularis (Fig. 142, b). Ventrad of the caudal corpus
quadrigeminum is seen a triangular area of oblique fibres which
corresponds in position to the human lemniscus.
The pedunculi cerebri (peduncles of the cerebrum) form
the ventral part of the midbrain. They appear in a ventral
view of the entire brain (Fig. 138) as two broad tracts of fibres
(g) emerging from beneath the pons and diverging from one[352]
another as they pass craniad, finally disappearing beneath the
cerebral hemispheres. Each is made up of many fibre-bundles,
which are apparent in surface view (Fig.
142). The peduncles are separated by
a small triangular space, which is marked
by a median longitudinal sulcus. In this
space, just caudad of the mammillary
bodies, is a small area through which a
number of blood-vessels pass into the
brain. This is known as the posterior
perforated area (or substance) (Fig.
142, j). The cerebral peduncles (a) are
crossed by the tractus transversus peduncularis
(b) (see above).
The third cranial nerve (Fig. 138,
III) (N. oculomotorius) leaves the brain
at the medial border of the cerebral
peduncle (g), just caudad of the tractus
transversus peduncularis.
The aqueductus cerebri (Fig. 143, j;
Fig. 153, d) (or aqueduct of Sylvius) is
the continuation craniad of the fourth
ventricle. It is a narrow passage, one
or two millimeters in diameter, lying
dorsad of the pedunculi cerebri and ventrad of the corpora
quadrigemina.
Fig. 142.—Ventral Surface
of the Midbrain
and ’Tween-brain,
with the Pons.
a, pedunculi cerebri; b,
tractus transversus peduncularis;
c, corpus geniculatum
mediale; d, optic tract; e,
optic chiasma; f, optic nerve;
g, mammillary bodies; h,
tuber cinereum; i, opening
for infundibulum (which has
been removed); j, posterior
perforated area; k, pons.
III, V, VII, VIII, third,
fifth, seventh, and eighth
cranial nerves.
C. Prosencephalon.—The prosencephalon or primitive
forebrain includes the diencephalon or ’tween-brain and the
telencephalon or cerebral hemispheres.
4. Diencephalon.
—The diencephalon or ’tween-brain includes
the thalami and the other parts bounding the third
ventricle. The diencephalon is seen in entire brains only in
ventral view (Fig. 138, c, d, e, etc.)
The diencephalon may be considered as forming almost or
quite the most cranial portion of the median nervous tube,—the
cranial wall of the third ventricle (the lamina terminalis)
(Fig. 143, d), ending in the median line in the deep fissure
between the hemispheres of the cerebrum. Parts of the brain[353]
which extend farther craniad than this are lateral portions, due
to the forward growth of the lateral hemispheres. The two
hemispheres of the cerebrum may be considered as lateral outgrowths
of the central ’tween-brain; these outgrowths have
extended dorsad, laterad, craniad, and caudad, so as to cover
almost completely the ’tween-brain.
In early stages the cerebral hemispheres are projections from
the cranial end of the ’tween-brain, so that the plane of junction
was nearly transverse, the cranial end of the ’tween-brain
joining the caudal end of the hemispheres. With the increasing
size and backward growth of the latter, the attachment to
the ’tween-brain has been shifted from a cranial to nearly a
lateral position, and at the same time the originally lateral surface
of the ’tween-brain has become nearly caudal. This is
shown in Fig. 141, the deep fissure at 1 marking the line of
attachment between the ’tween-brain and the hemispheres.
The dorsoventral plane of junction of ’tween-brain and hemispheres
is (as Fig. 141 shows) not wholly lateral, but oblique,
passing from its cranial end near the middle line caudolaterad.
A second peculiarity of the ’tween-brain lies in the thinness
of its roof. The roof is exceedingly thin and is so intimately
connected with the pia mater that they cannot be removed
separately. The ventral thick floor of the ’tween-brain is
directly continuous with the similar floor of the cerebrum; but
where the roof of the ’tween-brain joins the roof of the cerebrum
along the oblique plane already indicated, the roof is very thin
and is intimately connected to the pia mater, and is at the
same time folded into the lateral ventricles to form, together
with the pia mater, the choroid plexuses of the lateral ventricles
(Fig. 148, e). When the pia mater is removed the thin roof of
the brain along the line of junction of the ’tween-brain and the
cerebrum is brought away and there appears to be a direct
communication between lateral ventricles and the exterior.
We may now take up the parts of the diencephalon in detail.
a. Thalamus (Fig. 141, t).—The two thalami are large
oblique rounded ridges, forming the sides of the ’tween-brain.
They lie just craniad of the cranial corpora quadrigemina (q),
but separated from them by a broad groove, and are completely[354]
covered by the caudally projecting part of the cerebral hemispheres.
The medial larger end of each thalamus is near the
middle line; thence the thalamus extends caudolaterad, and
rises at its lateral extremity into a sharply rounded projection,
the corpus geniculatum laterale (x). From this projection the
thalamus is continued ventrad and then craniomediad as a large
band of fibres, the optic tract (tractus opticus) (Fig. 142, d),
which extends to the optic chiasma (e), where it passes into the
optic nerves (f). On the caudal border of the thalamus, near
the median line, is a very faint projection, the pulvinar (Fig.
141, w); this lies just craniad of the lateral border of the cranial
corpus quadrigeminum (q). Just ventrad of the corpus geniculatum
laterale (x) is the prominent rounded corpus geniculatum
mediale (s); this is connected by a prominent ridge, the
brachium quadrigeminum inferius (r), with the caudal corpus
quadrigeminum (p). In a similar manner the brachium
quadrigeminum superius passes from the cranial corpus quadrigeminum
(q) into the thalamus itself.
Between the two thalami there exists a groove, the sulcus
hypothalamicus. Over this groove lies the roof of the third
ventricle, forming the choroid plexus of the third ventricle
(Fig. 143, w). The medial surface of the two thalami are flat
and extend directly ventrad, forming part of the lateral
boundary of the third ventricle (Fig. 143, h). The two
medial surfaces meet over a considerable area across the narrow
cavity of the third ventricle, and unite, forming the massa
intermedia (Fig. 143, f) or intermediate mass of the thalamus
(“middle commissure”). This connection of the thalami of
the two sides is thus not a primitive one, forming no part of
the roof or floor of the central cavity, but is a secondary connection
due to a growing together of a part of the two sides of
the ventricle across its cavity. Along the dorsal edge of the
medial border of each thalamus passes a distinct white strand,
the stria medullaris (Fig. 141, u); the two striæ meet in an
arch caudad, lying beneath the pineal body (2).
The thalamus is separated craniolaterad by a groove (1)
from the corpus striatum (y), on the floor of the cerebral
hemisphere.
[355]
b. The roof of the third ventricle is thin and united with
the pia mater, as already stated. The pia mater bears many
blood-vessels, and the two are folded into the groove between
the optic thalami, forming the lamina chorioidea epithelialis,
or choroid plexus of the third ventricle (Fig. 143, w). The
roof is attached to the dorsomedial borders of the thalami and
becomes continuous with the choroid plexus of the lateral ventricles
(Fig. 148, e) at the craniolateral borders of the thalami.
The pineal body (corpus pineale) or epiphysis (Fig. 141,
2; Fig. 143, y) is a small conical body which is formed as an
outgrowth of the caudal part of the roof of the third ventricle.
It lies on the roof of the brain between the two cranial corpora
quadrigemina (Fig. 141, q). It is hollow, containing an extension
of the third ventricle. From its craniolateral angles two
white strands, the habenulæ, extend into the striæ medullares
(Fig. 141, u) of the thalami.
Just ventrad of the pineal body is a transverse band of
white fibres, lying in the caudal part of the roof of the third
ventricle. This interconnects the two thalami, and forms the
posterior commissure (Fig. 143, z′) (commissura posterior).
From this commissure a thin sheet of tissue extends to the
pineal body.
c. The floor of the third ventricle appears in a ventral view
of the brain (Fig. 138, Fig. 142) as a somewhat diamond-shaped
space craniad of the pedunculi cerebri (Fig. 142, a) and
bounded along its cranial margin by the optic tracts (Fig.
142, d). The optic tracts come from the thalami, as already
described; they converge and unite to form the optic chiasma
(Fig. 142, e), from which the optic nerves (f) diverge.
Immediately caudad of the optic chiasma lies a considerable
rounded gray elevation, the tuber cinereum (Fig. 142, h).
This bears on its ventral surface in the natural condition the
infundibulum (Fig. 138, d) with the hypophysis (Fig. 138, e);
in cases where the two latter structures have been removed
(Fig. 142) the tuber cinereum (h) bears a small longitudinal
opening (i) for attachment of the infundibulum. The infundibulum
(Fig. 138, d) is a hollow extension of the floor of the
third ventricle, and is attached to the middle of the ventral[356]
surface of the tuber cinereum. It bears at its ventral end the
hypophysis (pituitary body) (Fig. 138, e), a vascular non-nervous
body of unknown function. The hypophysis is lodged in
the sella turcica of the sphenoid bone.
At its caudal border the tuber cinereum bears two white
elevations, the mammillary bodies (corpora mammillaria)
(Fig. 142, g).
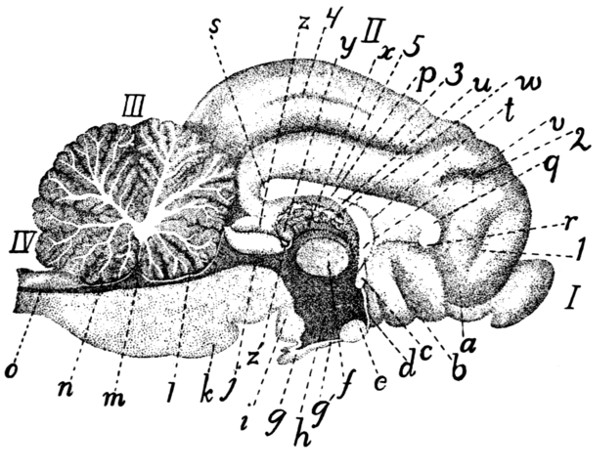
Fig. 143.—Longitudinal
Median Section of Brain.
I, olfactory bulb; II, cerebrum; III, section of cerebellum, showing “arbor
vitæ”; IV, medulla oblongata. a, tractus olfactorius; b, substantia perforata anterior;
c, anterior commissure; d, lamina terminalis; e, optic chiasma; f, massa intermedia
of thalamus; g′, infundibulum; g, hypophysis; h, third ventricle; i, mammillary
body; j, aqueductus cerebri (or aqueduct of Sylvius); k, pons; l, velum medullare
anterius; m, fourth ventricle; n, velum medullare posterius; o, continuation of
canal of spinal cord; p, corpus callosum (body); q, genu of corpus callosum; r, rostrum;
s, splenium; t, septum pellucidum; u, fornix; v, pillars of fornix; w, choroid
plexus of third ventricle; x, stria medullaris; y, corpus pineale; z, cranial corpus
quadrigeminum; z′, posterior commissure. 1, sulcus falcialis; 2, sulcus cruciatus;
3, sulcus splenialis; 4, sulcus marginalis; 5, sulcus supracallosalis (or callosalis).
The third ventricle (Fig. 143, h; Fig. 141, v) is a very
narrow slit-like space, of considerable extent dorsoventrally,
but less than a millimeter in width; it lies between the medial
ends of the thalami (Fig. 141), and extends ventrad into the
tuber cinereum, and thence into the infundibulum (Fig. 143,
g′). Dorsad it extends into the corpus pineale (Fig. 143, y).
The third ventricle communicates caudally near its dorsal
border with the aqueductus cerebri (j); craniolaterally with
the lateral ventricles, through the interventricular foramen[357]
(foramen of Monroe). Its cranial boundary forms in the middle
line a thin plate, the lamina terminalis (d), which is, morphologically,
the cranial termination of the cerebrospinal axis; it
lies at the bottom of the deep fissure between the cerebral
hemispheres. At the dorsal border of the lamina terminalis is
a strong transverse band of fibres connecting the two sides of
the brain; this is the anterior commissure (c). The cavity of
the third ventricle is much encroached upon by the meeting
and secondary union of the two thalami across the middle line,
forming the massa intermedia (f).
The boundaries of the third ventricle, in order, are as follows,
beginning dorsocraniad of the communication with the
aqueductus cerebri: the posterior commissure (Fig. 143, z′),
the pineal body (y), the choroid plexus of the third ventricle
(w), the columns of the fornix (v), the anterior commissure
(c), the lamina terminalis (d), the tuber cinereum with the
infundibulum (g′), the substantia perforata posterior, and the
midbrain. The lateral boundaries are formed by the thalami
(Fig. 141, t).
5. Telencephalon.
—The telencephalon includes the two
cerebral hemispheres. The name cerebrum is also applied to
this portion of the brain; frequently, however, the name cerebrum
is used as signifying the entire mass of the brain craniad
of the rhombencephalon,—therefore including mesencephalon,
diencephalon, and telencephalon. The term will here be used
as synonymous with telencephalon.
The cerebral hemispheres arise as two lateral outgrowths
from the cranial end of the primitive forebrain. They have
undergone great increase in size in the course of evolution, so
as to form the larger part of the brain; at the same time
important modifications of structure have taken place. In
the original condition the medial faces of the two hemispheres
are not connected, the two being separate outgrowths of the
’tween-brain, and connected only through the latter. But
secondary connections have been formed across the fissure
between the two hemispheres, resulting in the production of
the corpus callosum (Fig. 143, p;
Fig. 147; Figs. 149-152, a),
a broad transverse band of white fibres connecting the two[358]
hemispheres dorsad of the ’tween-brain. Ventrad of the corpus
callosum another secondary union has resulted in the production
of the fornix (Fig. 143, u).
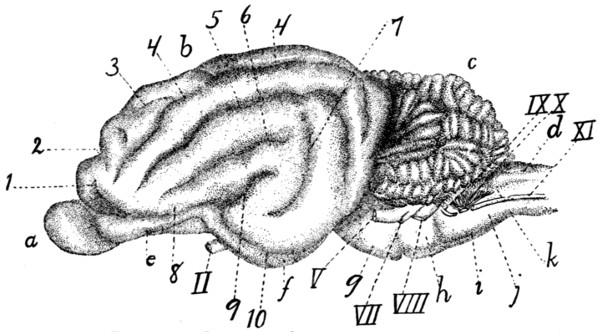
Fig. 144.—Lateral Surface of the Brain.
a, olfactory bulb; b, cerebral hemisphere; c, cerebellum; d, medulla oblongata;
e, tractus olfactorius; f, lobus pyriformis or tractus postrhinalis; g, pons; h, trapezium;
i, pyramid; j, area elliptica; k, area ovalis. II, N. opticus; V, N. trigeminus;
VII, N. facialis; VIII, N. acusticus; IX, N. glossopharyngeus; X, N. vagus;
XI, N. accessorius. 1. sulcus præsylvius; 2, sulcus cruciatus; 3, sulcus ansatus;
4, sulcus lateralis; 5, sulcus suprasylvius; 6, sulcus anterior; 7, sulcus posterior; 8,
sulcus rhinalis; 9. fissura Sylvii; 10. sulcus rhinalis posterior.
External Features.—With increase in size the mass of the
cerebrum shows externally a tendency to divide into three
lobes, one craniad, the frontal (Fig. 145, A); one caudoventrad,
the temporal (B); and one caudodorsad, the occipital
(C). The two latter are not distinctly marked off from one
another. The limit between the temporal and frontal lobes is
marked by a short deep fissure, the lateral fissure (fissura
cerebri lateralis), or fissure of Sylvius (Fig. 144, 9; Fig.
145, a). Each lobe is thrown up into elevations or gyri,
which are separated by grooves or sulci; these are described
below. The homology of the cerebral gyri and sulci of the
cat with those of man is in most cases uncertain.
The sulci and gyri of the cerebral hemispheres may be
described briefly in their main features as follows: On the
lateral surface of the hemisphere (Fig. 145) the lateral fissure
(fissura cerebri lateralis), or fissure of Sylvius, separating
temporal and frontal lobes, forms the most convenient point
of departure for an understanding of the fissures. The fissure[359]
of Sylvius is short but deep, and is formed early during
development. In man it covers an area, the insula, at its
bottom, which may be seen by separating the sides of the
fissure. In the cat the insula is rudimentary and can be
demonstrated only with difficulty.
The lateral surface of the hemisphere is marked by fissures
which form three concentric arches (b-c, d, and e), irregular
and incomplete, about the fissure of Sylvius. These arches all
open ventrad. The first arch (b-c) (that next to the fissure of
Sylvius) lacks the central part, the keystone, so that a complete
arch is not formed, but only the two sides of the arch, as
two separate sulci (b and c). The one of these craniad of the
fissure of Sylvius is the sulcus anterior (b); the other is the
sulcus posterior (c). In the dog this arch is usually complete
and the sulcus forming it is frequently known as the ectosylvian
sulcus (sulcus ectosylvius).
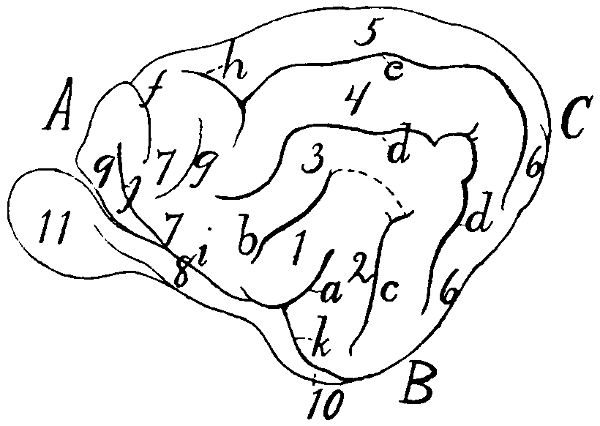
Fig. 145.—Diagram of the Sulci and Gyri on the Lateral Surface
of the Hemisphere.
A, frontal lobe; B, temporal lobe; C, occipital lobe. a, lateral fissure or fissure
of Sylvius; b, sulcus anterior; c, sulcus posterior (the broken line connecting the
ends of a and b serves to show how the first arch is completed to form the ectosylvian
sulcus, as in the dog); d, sulcus suprasylvius; e, sulcus lateralis; f, sulcus cruciatus;
g, sulcus coronalis; h, sulcus ansatus; i, sulcus rhinalis; j, sulcus præsylvius; k,
sulcus rhinalis posterior. 1, anterior Sylvian gyrus; 2, posterior Sylvian gyrus; 3,
gyrus ectosylvius; 4, gyrus suprasylvius; 5, gyrus marginalis; 6, gyrus compositus
posterior; 7, sigmoid gyrus; 8, olfactory tract; 9, orbital gyrus; 10, lobus pyriformis;
11, olfactory bulb.
The gyri included between the fissure of Sylvius on the one
hand and the anterior and posterior sulci on the other are
known as the anterior (1) and posterior (2) Sylvian gyri.
The second arch is formed by the suprasylvian sulcus
(sulcus suprasylvius) (d). The gyrus between the anterior
and posterior sulci ventrad and the suprasylvian sulcus dorsad[360]
is the ectosylvian gyrus (3) (because dorsad of the ectosylvian
sulcus).
The third arched sulcus, next to the medial margin of the
hemisphere, is variable: it forms the lateral sulcus (e). The
gyrus lying between the suprasylvian and lateral sulci is the
suprasylvian gyrus (4) (because dorsad of the suprasylvian
sulcus). The lateral sulcus is sometimes broken into two (as
in Fig. 137, h), the two ends overlapping so as to leave a
small gyrus between the two ends.
The gyrus dorsomediad of the lateral sulcus, extending
onto the medial surface of the hemisphere as far as the splenial
sulcus, is the marginal gyrus (5).
The four sets of gyri included between these three sets of
arched sulci and the fissure of Sylvius unite caudad and craniad
to form single gyri. The caudal one is the gyrus compositus
posterior (6). The cranial one (less regular than the caudal
one) is the sigmoid gyrus (7). The latter curves about the
cruciate sulcus (f) (described below).
In the cranial part of the lateral surface of the hemisphere
are certain sulci and gyri which do not belong to the system
above described. The cruciate sulcus (f) is a short transverse
sulcus passing from the lateral surface of the hemisphere onto
its medial surface, where it extends caudad about a centimeter.
Curving around the lateral end of this, separated from it by
part of the sigmoid gyrus, is the short coronal sulcus (g).
Connected usually with the end of the lateral sulcus (e) and
running nearly parallel with the cruciate sulcus is the short
sulcus ansatus (h).
At its ventral end the fissure of Sylvius (a) joins a longitudinal
groove passing craniad and caudad from the point of
junction. That part which passes craniad is the sulcus
rhinalis (i); it forms the dorsolateral boundary of the olfactory
tract (8). Passing dorsocraniad from the sulcus rhinalis is a
sulcus which separates a very small cranial lobe from the rest
of the hemisphere; this is the supraorbital or præsylvian
sulcus (j). The portion of the hemisphere craniad of this
sulcus is the orbital gyrus (9).
The sulcus rhinalis (i) is continuous caudad with the[361]
sulcus rhinalis posterior (or sulcus postrhinalis) (k). This
extends caudad onto that portion of the hemisphere which faces
the cerebellum. It forms the lateral boundary of a large elongated
oval lobe lying at the side of the ventral floor of the
midbrain and ’tween-brain. This lobe has been called tractus
postrhinalis or lobus pyriformis (Fig. 145,
10; Fig. 138, f).
In addition to the sulci and gyri above mentioned, any
given specimen will usually show a number of small inconstant
sulci and gyri in various regions; these inconstant structures
will not be here described.
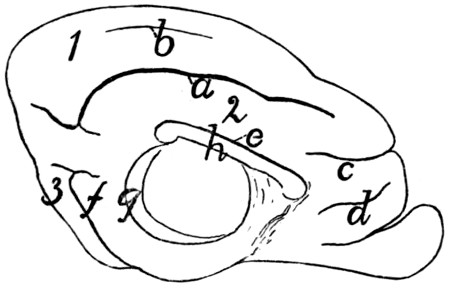
Fig. 146.—Diagram of the Sulci and Gyri on the Medial Surface of
the Hemisphere.
a, sulcus splenialis; b, sulcus marginalis;
c, sulcus cruciatus; d, sulcus falcialis;
e, sulcus supracallosalis; f, sulcus rhinalis posterior; g, hippocampal sulcus; h,
corpus callosum. 1, gyrus marginalis; 2, gyrus fornicatus; 3, gyrus compositus
posterior.
On the medial surface of the hemisphere (Fig. 146), the
following arrangement of sulci and gyri seems to be typical.
Some distance from the dorsal margin a long sulcus runs
parallel with the margin; this is the sulcus splenialis (a).
The marginal gyrus (1) is dorsad of the splenial sulcus, passing
onto the lateral surface of the hemisphere, where it is
bounded by the lateral sulcus. The splenial sulcus extends
onto the caudal surface of the hemisphere. A shallow sulcus
marginalis (b) occurs frequently between the sulcus splenialis
and the dorsal margin of the hemisphere. Cranioventrad or
the cranial end of the splenial sulcus is that portion of the
sulcus cruciatus (c) that lies on the medial surface of the
hemisphere. Ventrad of this is a short shallow furrow which
has been called the sulcus falcialis (d).
Immediately dorsad of the corpus callosum (h), separating
its dorsal surface from a part of the hemisphere, is the[362]
supracallosal or callosal sulcus (e). Between the supracallosal
(e) and splenial (a) sulci is the gyrus fornicatus (2).
On the caudal surface of the hemisphere (that facing the
cerebellum) appear the ends of the sulcus splenialis (a) and the
sulcus rhinalis posterior (f). Hidden in the natural condition
by the thalami and optic tract is the broad hippocampal sulcus
(g), marking externally the course of the hippocampus.
A further extension of the surface of the cerebrum has
taken place in connection with the sense of smell. A mass of
gray matter, the olfactory bulb (Fig. 144, a), reckoned as a
part of the cerebrum, is separated from the latter and lies
against the cribriform plate of the ethmoid bone. From it the
olfactory fibres pass through the perforations of the plate to the
olfactory mucous membrane. The olfactory bulb contains a
cavity, a part of the lateral ventricle.
The bulb lies against the ventral surface of the frontal lobe
and projects craniad of it. It is connected to the cerebrum by
a tract of fibres, the olfactory tract (Fig. 138, a), which is
divisible into two roots, medial and lateral. The medial root
comes from the medial surface of the frontal lobe, where it is
continuous with a tract extending to the cranial end of the
corpus callosum. The lateral root is traceable from an elevated
gyrus-like portion of the cerebrum which lies at the side of the
infundibulum and is known as the lobus pyriformis or tractus
postrhinalis (Fig. 138, f). The lateral root is divisible into a
medial white strand and a lateral gray strand.
That part of the brain comprising the olfactory bulb and
the parts intimately related to it are frequently included under
the term rhinencephalon.
In the triangular area between the two olfactory tracts and
craniad of the optic chiasma appears a mass of gray matter,
subdivided by a longitudinal fissure. This possesses numerous
openings through which blood-vessels pass to the brain substance,
and is thence known as the anterior perforated substance
(substantia perforata anterior) (Fig. 138, b).
Internal Structures of the Cerebrum.—The cavity of each
of the cerebral hemispheres is known as a lateral ventricle.
The two lateral ventricles constitute the first and second of the[363]
ventricles of the brain, whence the application of the names
third and fourth ventricles to the cavities of the ’tween-brain
and hindbrain. The lateral ventricles do not grow at the same
rate as the walls of the hemispheres, so that they remain comparatively
small. The cavity of each ventricle is further
reduced in size by the development on its floor of a large
ridge-like thickening, the corpus striatum (Fig. 148, f).
The dorsal wall or roof of the lateral ventricle joins the thin
roof of the third ventricle on each side along an oblique curved
line (Fig. 141, 1) which follows the cranial or lateral border of
the thalamus (Fig. 141, t). Along this line the thin roof of
the brain is folded in together with the pia mater to form the
choroid plexus of the lateral ventricles (lamina chorioidea
epithelialis) (Fig. 148, e). When this is pulled out there is
left a fissure, the “great transverse fissure of the cerebrum”
(Fig. 141, 1). Just dorsad of the groove
between the thalami there runs a tract of
white fibres known as the fornix (Fig.
148, a). The two halves of the fornix
separate at the cranial ends of the thalami
and pass ventrad, forming thus the
pillars of the fornix (Fig. 148, b; Fig.
143, v). Dorsad of the cranial end of the
fornix the corpus callosum (Fig. 143, p)
passes from one hemisphere to the other.
Caudad of the pillars of the fornix, the
lateral ventricles communicate with the
third ventricle by way of the interventricular
foramen (foramen of Monroe).
The parts of the cerebrum may now
be taken up in detail.
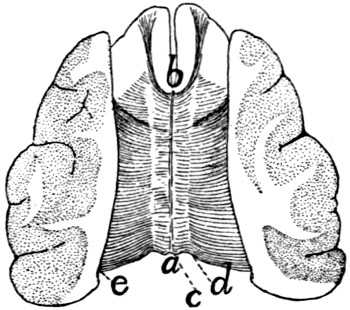
Fig. 147.—The Corpus
Callosum.
The dorsal portion of the
hemispheres has been dissected
away, then sliced off,
showing the plate of transverse
fibres forming the
corpus callosum. a, splenium;
b, genu; c, line marking
the medial edge of the hemispheres;
d, line marking the
lateral boundary of the supracallosal
sulcus; laterad of
this line the corpus callosum
lies in the substance of the
hemispheres, which have
been dissected away; e, line
marking medial limit of cut
surface.
The corpus callosum (Fig. 147; Fig.
143, p; Figs. 149-152, a) is a broad
transverse band of fibres forming a
secondary connection between the medial
walls of the two hemispheres, dorsad of the roof of the third
ventricle. Its outer surface (Fig. 147) is exposed at the
bottom of the fissure which separates the hemispheres. On[364]
each side it passes laterad, forming the roof of the lateral
ventricle. Its cranial part lies dorsad of the corpus striatum,
and its caudal part dorsad of the thalamus. Laterally its fibres
radiate into the substance of the hemispheres. At its cranial
end the corpus callosum bends ventrad and then caudad (Fig.
143). The part which turns to pass ventrad is the genu (Fig.
143, q) or knee, while the part which projects caudad is the
rostrum (r). The caudal border of the corpus callosum is also
thickened and turned ventrad and is called the splenium (Fig.
143, s); it lies dorsad of the cranial corpora quadrigemina (z).
The caudal half of the ventral surface of the corpus callosum
is united with the fornix (Fig. 143, u).
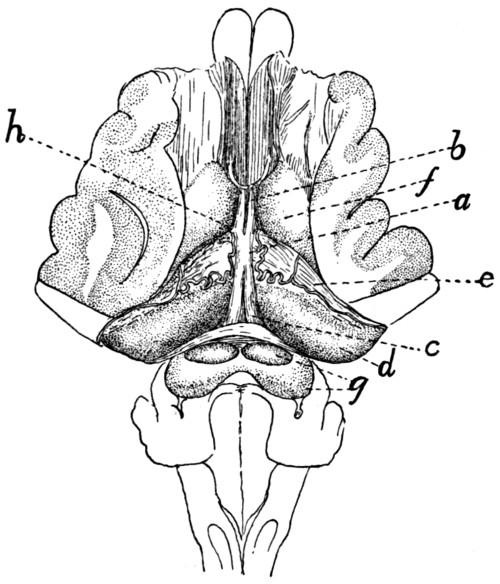
Fig. 148.—Fornix, Hippocampus, and Corpus Striatum.
The dorsal portion of the hemispheres has been dissected away and the corpus
callosum removed. a, fornix; b, columns or pillars of the fornix; c, crura of the
fornix; d, hippocampus; e, choroid plexus of the lateral ventricles overlying the fimbria
(the choroid plexus shows an artery); f, corpus striatum; g, corpora quadrigemina;
h, position of the interventricular foramen (foramen of Monroe).
The fornix (Fig. 148, a; Fig. 143,
u, v; Figs. 150-152, b)
consists of an arched tract of longitudinal fibres near the medial
border of each hemisphere, ventrad of the corpus callosum.
Each tract begins in the mammillary bodies, and passes dorsad,
the two converging until they run side by side, forming a
cylindrical dorsoventral bundle known as the columns or pillars[365]
of the fornix (Fig. 143, v; Fig. 150, e), which cross the
anterior commissure (Fig. 143, c; Fig. 150, f) caudad of the
latter. Caudad of the pillars of the fornix, between these and
the thalamus, lies on each side the interventricular foramen or
foramen of Monroe, a small opening which connects the lateral
ventricles with the third ventricle. Dorsad of the foramen the
fornix turns caudad, the two fibre-tracts of each hemisphere
lying side by side and closely connected (Fig. 148, a), forming
thus another secondary union between the medial surfaces of
the two hemispheres. This portion of the fornix is the corpus
or body (Fig. 148, a); it lies dorsad of the roof of the third
ventricle and passes to the splenium (Fig. 143, s) of the corpus
callosum, and its dorsal surface unites with the ventral surface
of the latter (Fig. 143). Caudad the two halves of the fornix
diverge, forming the crura of the fornix (Fig. 148, c); these
and the body are continuous laterally with the hippocampus
(Fig. 148, d) and the fimbria (Fig. 148, beneath e).
The anterior commissure (Fig. 143, c; Fig. 150, f) is a
transverse band of white fibres which stretches from one hemisphere
to the other about half way between the interventricular
foramen or foramen of Monroe and the floor of the third ventricle,
and just craniad of the pillars of the fornix. This tract
of fibres is developed in the original wall of the third ventricle,
so that it does not form a secondary connection between the
halves of the cerebrum, as do the fornix and corpus callosum.
It lies dorsad of the lamina terminalis (Fig. 143, d) and is
continuous with it.
The septum pellucidum (Fig. 143, t) is a vertical partition
which separates the lateral ventricles and fills the interval
between the corpus callosum dorsad and the fornix ventrad.
It is triangular and translucent. It is formed from the medial
walls of the two hemispheres and therefore is made up of two
laminæ which embrace between them a space which originally
was a part of the fissure separating the hemispheres. This
space has been called the fifth ventricle.
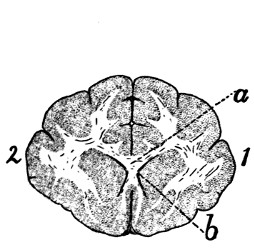
Fig. 149.—Caudal Surface of
Transverse Section of Brain
through the Genu of the
Corpus Callosum.
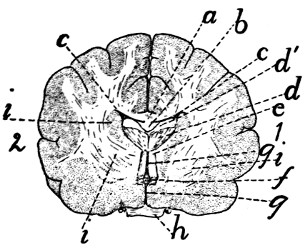
Fig. 150.—Caudal Surface of
Transverse Section of Brain
through the Optic Chiasma
and the Region of the Interventricular
Foramen (Foramen
of Monroe).
Fig. 149.—Gray matter dotted; white matter with a few lines. 1, right hemisphere;
2, left hemisphere. a, corpus callosum, section through genu and rostrum;
b, cavity of lateral ventricles.
Fig. 150.—1, right hemisphere; 2, left hemisphere. a, corpus callosum; b, fornix
(cut); c, cavity of lateral ventricles; d, fimbria (d′, cut surface; d, caudal uncut
surface of the part that curves ventrad); e, pillars of fornix; f, anterior commissure;
g, parts of third ventricle; h, optic chiasma; i, parts of corpus striatum.
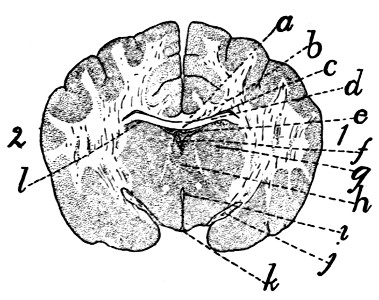
Fig. 151.—Caudal Surface of Transverse Section of the Brain through
the Tuber cinereum and Infundibulum, Craniad of the Hypophysis.
a, corpus callosum; b, fornix (continuous at the sides with the hippocampus); c,
cavity of lateral ventricles; d, hippocampus; e, choroid plexus of the third ventricle
(roof of the diencephalon); f, dorsal part of cavity of the third ventricle; g, section
of thalamus; h, massa intermedia of the thalami, dividing the third ventricle into a
dorsal (f) and a ventral (i) portion; i, ventral part of third ventricle; j, part of optic
tracts; k, thin wall of infundibulum, with part of cavity of third ventricle; l, line
separating roof of thalamus (diencephalon) from floor of hemispheres (telencephalon).
The hippocampus (Fig. 148, d; Fig. 152, d) is an elongated
rounded elevation of the floor of the lateral ventricle.
It is continuous mediad with the fornix (Fig. 148, a) and[366]
extends thence along the inferior horn of the lateral ventricle
to its end in the temporal lobe. It is somewhat narrower at
its lateral end. It is thus curved into a semicircle in conformity
with the inferior horn of the ventricle. Its dorsal surface is
convex and looks into the lateral ventricle; its ventral surface
is concave and rests upon the thalamus and the optic tract.[367]
Along the craniolateral edge of the hippocampus is a broad,
clearly marked fibre-tract, the fimbria; this runs parallel to
the choroid plexus of the lateral ventricle and beneath it, and
is continuous at its medial end with the fornix.
The corpus striatum (Fig. 148, f;
Fig. 141, y; Fig. 150,
i) is a fusiform or ovoid elevation of the floor of the lateral ventricle.
One of its narrow ends lies about opposite the middle
of the thalamus (Fig. 141, t) and it extends thence obliquely
mediad nearly parallel with the thalamus. In the groove
between it and the thalamus is the choroid plexus of the lateral
ventricles (Fig. 148, e). Internally the corpus striatum is
made up of several layers of different texture (Fig. 150, i),
whence its name.
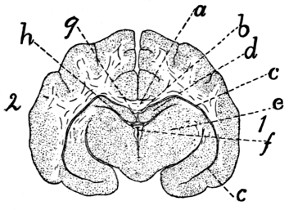
Fig. 152.—Caudal Surface of
Transverse Section of the
Brain through the Pineal
Body and Thalami.
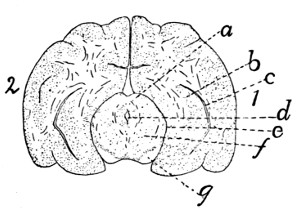
Fig. 153.—Caudal Surface of
Transverse Section of the Brain
through the Cranial Pair of
Corpora Quadrigemina.
Fig. 152.—a, corpus
callosum; b, fornix; c, inferior horn of lateral ventricles;
d, hippocampus; e, thalamus; f, third ventricle; g, pineal body; h, space between
the floor of the hemisphere (hippocampus) and roof of the ’tween-brain (thalamus).
Fig. 153.—a, cranial corpora quadrigemina; b, hippocampus; c, part of inferior
horn of lateral ventricle; d, aqueductus cerebri (aqueduct of Sylvius); e, space
between the outer surface of the midbrain and the lower surface of the hemisphere;
f, cross-section of midbrain; g, pedunculi cerebri.
The choroid plexus of the lateral ventricles (lamina
chorioidea epithelialis) (Fig. 148, e).—The line of junction of
the roof of the third ventricle (i.e., the choroid plexus of the
third ventricle) and the cerebral hemispheres is along the
lateral (or cranial) border of the thalamus (Fig. 141, t).
Along this line (Fig. 141, 1) the brain-wall remains very thin
and becomes closely associated with the pia mater, so that on
the removal of the pia there is left the “transverse fissure”
which leads into the lateral ventricle. In an early stage, before
the cerebrum has grown far caudad, the fissure is exposed in[368]
dorsal view, but after the hemispheres have covered the
thalami the fissure appears to be in the lower face of the hemisphere.
Through this fissure the pia mater extends upward
into the lateral ventricles as a fold separated from the ventricle
by the thin brain-wall. This fold is vascular and is known as
the choroid plexus of the lateral ventricle (Fig. 148, e). It is
a fringe-like fold lying on the fimbria and extending from the
foramen of Monroe (at h) nearly to the end of the inferior horn
of the lateral ventricle.
The lateral ventricles are the extensions into the hemispheres
of the originally unpaired cavity of the forebrain.
Each consists of a portion parallel to the basis cranii and lying
at the side of the septum pellucidum between the corpus callosum
dorsad and the corpus striatum, hippocampus, and fornix
ventrad, and of two horns, an anterior and an inferior horn.
The inferior horn (Fig. 152, c) is a narrow cleft, crescent-shaped
in cross-section, which follows the dorsal surface of the
hippocampus (Fig. 148, d; Fig. 152, d) along the temporal
lobe to its end almost against the tuber cinereum. It thus
passes first caudad and then ventrad and finally mediad. The
choroid plexus and hippocampus project into it from its floor.
The anterior horn extends ventrad and then slightly caudad
in the frontal lobe, following the corpus striatum (Fig. 148, f),
on which it lies.
The interventricular foramen (or foramen of Monroe).—If
the choroid plexus of the lateral ventricle (Fig. 148, e) is
followed to its medial end, it is found to pass through a foramen
(at h) in which it becomes continuous with the opposite plexus
or roof of the third ventricle. This foramen leads from one
lateral ventricle to the other and is connected ventrally by a
median opening with the third ventricle. It is thus Y-shaped
or T-shaped and is the interventricular foramen (or foramen of
Monroe). The foramen lies opposite the middle of the corpus
striatum and caudad of the pillars of the fornix.
Membranes of the Brain.—The membranes of the brain
are the dura mater, the arachnoid, and the pia mater.
The dura mater is a strong fibrous membrane lining the
cranial cavity and covering the brain. It is strongly attached[369]
to the projections of the base of the skull and to the tentorium.
It dips between the cerebral hemispheres and olfactory bulbs,
forming the falx cerebri. It likewise dips between the cerebrum
and the cerebellum, covering both surfaces of the bony
tentorium. The dura mater does not pass into the fissures or
sulci of the surface of the brain. It becomes continuous with
the sheath of the cranial nerves as they leave the skull. It is
continuous with the dura mater of the spinal cord.
The arachnoid and pia mater are essentially similar to the
same structures on the spinal cord. The pia mater dips into
the fissures and sulci of the brain.
II. THE PERIPHERAL NERVOUS SYSTEM.
1. Cranial Nerves. Nervi cerebrales.—
I. N. olfactorius.
—The
fasciculi of the olfactory nerves arise from the
olfactory bulb (Fig. 144, a) and pass through the foramina of
the cribriform plate, upon which the bulb lies, to be distributed
to the olfactory mucous membrane of the nose.
II. N. opticus.
—The second nerve or optic (Fig. 138,
II) arises from the optic chiasma (Fig. 138, c), passes through
the optic foramen, and extends in an S-shaped curve to the
eyeball. Its course is craniodorsad. It pierces the sclerotic
and choroid coats of the eye and spreads out into the retina.
III. N. oculomotorius.
—The third or oculomotor nerve
arises (Fig. 138, III) from the pedunculus cerebri and passes
into the orbit through the orbital fissure. It passes between
the lateral and superior recti, sends a large branch to the
superior rectus, supplies the medial rectus and the retractor
oculi, passes laterad of the optic nerve and supplies the inferior
rectus, and sends a long branch to the inferior oblique (Fig.
166, f). At the point where the branch is given off to the
inferior oblique muscle is the triangular reddish ciliary (or
ophthalmic) ganglion, about two millimeters in diameter. It
is directly attached to the inferior oblique branch of the third
nerve and receives a delicate sensory filament from the
ophthalmic nerve. From the ciliary ganglion proceed two[370]
short ciliary nerves which pass along each side of the optic
nerve to the eyeball. These nerves penetrate the sclerotic at
the sides of the eyeball to be distributed to the ball.
IV. N. trochlearis.
—The fourth nerve, the trochlearis
(or patheticus), arises from the lateral border of the velum
medullare anterius, as already described (Fig. 141, n). It
passes craniad, emerges through the orbital fissure in company
with the oculomotor, abducens, and ophthalmic, passes dorsad
of the superior rectus muscle, and reaches (Fig. 154, l) the
caudal border of the belly of the superior oblique muscle near
its middle. It supplies only the superior oblique.
V. N. trigeminus.
—The fifth nerve, the trigeminus (or
trifacial) arises (Fig. 138, V) by two roots, a large sensory and
a small motor root (4), as already described (p. 347). One
of these, the motor (4), is smaller and more ventral; the other,
the sensory root, is larger and dorsal. The dorsal root soon
enlarges to form a large ganglion, the semilunar (or Gasserian)
ganglion (Fig. 138, k), from which three branches diverge.
One branch is joined by the ventral root (4), which passes over
the ventral surface of the semilunar ganglion; and the nerve
thus formed is the mandibular division (1) of the fifth nerve.
Of the other two branches from the ganglion, the middle and
longest is the maxillary nerve (2), and the smallest is the
ophthalmic (3). The mandibular nerve is thus mixed, motor
and sensory, while the others are sensory.
1. N. ophthalmicus.—The ophthalmic or first division of
the fifth nerve arises from the semilunar (or Gasserian) ganglion.
It passes out of the cranial cavity and into the orbit
by way of the orbital fissure, in company with the third, fourth,
and sixth nerves and with the extension of the carotid
(arterial) plexus.
It passes between the superior and medial recti along with
the third nerve, crosses dorsad of the optic nerve, and divides
into infratrochlear and ethmoidal branches. In the orbital
fissure it gives off the frontal nerve, and while crossing the
optic it gives off the long ciliary nerve.
a. N. frontalis.—The frontal nerve passes along the lateral
border of the superior oblique muscle and then laterad of the[371]
pulley to near the middle of the supraorbital crest of the frontal
bone. Here it passes out of the orbit and is distributed to the
integument of the upper eyelid and the adjacent region at the
side of the nose.
b. N. infratrochlearis.—The infratrochlear nerve passes
between the superior rectus and the superior oblique in the first
part of its course. It then passes ventrad of the superior
oblique and ventrad of the pulley to be distributed to the integument
of the upper eyelid near the inner angle.
c. N. ethmoidalis.—The ethmoidal nerve passes along
with the ethmoidal artery through the ethmoidal foramen (or
foramina) in the orbital plate of the frontal bone. It is finally
distributed to the mucosa of the nose and to the cartilage and
integument of the snout.
d. N. ciliaris longus.—The long ciliary nerve arises from
the ophthalmic and passes along the optic nerve to be distributed
to the eyeball. It divides into several branches before
penetrating the sclerotic.
One or two small communicating branches to the ciliary
ganglion are given off at about the same point as the long
ciliary nerve. (For a description of this ganglion see the
account of the oculomotor nerve, page 369.)
2. N. maxillaris.—The maxillary nerve, the second division
of N. trigeminus, rises from the semilunar (Gasserian)
ganglion and leaves the skull by the foramen rotundum. It
is the sensory nerve of the palate, upper teeth and upper lip,
and of part of the forehead and cheek.
On leaving the foramen the maxillary nerve divides into
three branches, the two infraorbital nerves (Fig. 154, f) and
the sphenopalatine (Fig. 154, g). Two smaller branches are
likewise given off either within or just outside of the foramen,
the lachrymal nerve (Fig. 154, j) and the zygomatic (subcutaneus
malæ) (Fig. 154, i).
a. N. lachrymalis (Fig. 154, j; Fig. 155, m).—The
lachrymal nerve passes along the periorbita to the lachrymal
gland (Fig. 154, 11), to which it gives branches. It then continues
caudad of the zygomatic process of the temporal to the
integument; here it turns caudad and is distributed to the integument[372]
in the region between the eye and the external ear
(Fig. 155, m). It anastomoses with the zygomatic branch of
the seventh nerve (Fig. 155, h).
b. N. zygomaticus (subcutaneus malæ) (Fig. 154, i).—This
arises with the preceding and follows it for some distance.
It passes through a foramen in the frontal process of the malar
bone and is distributed to the lower eyelid and adjacent
integument.
c. Nn. infraorbitales (Fig. 154, f; Fig. 155, l).—The
infraorbital nerves are two of nearly equal size. They pass
through the orbit ventrad of the eyeball to the infraorbital
canal. On their course each divides once or twice and each
divides again in the infraorbital foramen, so that about eight
branches emerge from the infraorbital foramen and diverge to
the integument and whiskers of the upper lip and to the side
and wing of the nose (Fig. 155, l). In the infraorbital canal,
and before reaching it, branches are given to the molar teeth,
and a branch continues in the bone to the canine, incisor, and
premolar teeth.
d. N. sphenopalatinus (Fig. 154, g).—The sphenopalatine
nerve turns mediad from the infraorbitals, directing its
course toward the sphenopalatine foramen. Before reaching
this it gives off the greater palatine nerve (N. palatinus
major), which enters the posterior palatine canal and passes
to the hard palate. The sphenopalatine then usually divides
into two branches which pass along side by side to enter the
sphenopalatine ganglion. This is a large elongated triangular
ganglion lying on the dorsal surface of the external pterygoid
muscle, just laterad of the sphenopalatine foramen.
The following nerves are connected with the sphenopalatine
ganglion:
a. N. palatinus minor.—This leaves the craniolateral
angle of the ganglion and passes to the soft palate.
b. N. nasalis posterior.—The posterior nasal nerve enters
the nasal cavity by the sphenopalatine foramen and is distributed
to the mucosa of the ventral and middle parts of the
nasal cavity.
c. N. canalis pterygoidii (Vidian Nerve).—This is a large[373]
nerve which leaves the caudal angle of the sphenopalatine
ganglion and passes caudad. It enters the orbital fissure, lying
in a groove on its ventral wall (the cranial end of this groove
is sometimes converted into a canal). The groove ends
caudally in a foramen which pierces the sphenoid bone between
the wing and the body and lies just mediad of the foramen
rotundum. The groove and foramen constitute the pterygoid
canal (from which the nerve is named). After emerging from
the pterygoid canal onto the ventral surface of the basisphenoid
the nerve enters the tympanic bulla along with the Eustachian
tube, lying on the medial side of the latter. Just after entering,
on reaching the internal carotid artery, it divides into two.
One of these, N. petrosus superficialis major (p. 375), passes
into the hiatus facialis of the petrous bone and joins the facial
nerve. The other, N. petrosus profundus, accompanies the
internal carotid artery caudad, turning therefore out of the
bulla and passing caudad along its medial side; it finally joins
the superior cervical ganglion of the sympathetic system.
3. N. mandibularis.—The third division of the fifth, the
mandibular nerve, takes origin by a strong root from the
semilunar (or Gasserian) ganglion, and receives also after
separating from the ganglion the smaller ventral root (portio
minor) of the fifth nerve. The root from the ganglion is sensory;
the smaller root is motor, so that the mandibular nerve
is both sensory and motor. The nerve passes through the
foramen ovale, and sends off at once the following branches:
a. N. auriculotemporalis (Figs. 154 and 155, n).—This
passes dorsad between the cartilaginous auditory meatus and
the zygomatic process of the temporal bone (Fig. 154, n),
emerges at the caudal border of the masseter muscle (Fig.
155, n), and divides into two chief branches. One, the auricular
branch, passes along the cranial side of the external ear
and is distributed to its integument. The other, the temporal
branch, passes along the zygomatic arch, gives branches to the
temporal muscle and to the skin, follows the zygomatic arch
almost to the angle of the mouth, and anastomoses with the
superior buccal branch of the facial (Fig. 155, k).
[374]
b. N. temporalis profundus.—One or more large branches
passing mediad of the zygomatic arch to the temporal muscle.
c. N. massetericus.—The masseteric nerve passes dorsocraniad
to the masseter muscle.
d. N. pterygoideus.—One or more pterygoid branches
pass to the pterygoid muscles. A small twig from the nerve
to the internal pterygoid passes into the tympanic cavity and
supplies the tensor tympani muscle.
e. N. buccinatorius (Fig. 154, e).—This passes craniad
along the dorsolateral surface of the pterygoid muscles (Fig.
154, 8) to the angle of the mouth. Here it divides into
branches to the masseter muscle, the mucosa of the mouth,
and to the lips.
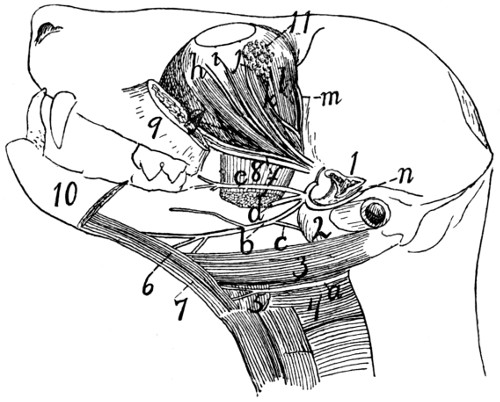
Fig. 154.—Dissection
showing a Number of the Cranial Nerves.
The mandible and zygomatic arch have been cut and removed; the temporal and
internal pterygoid muscles are also removed. a, N. hypoglossus; b, N. lingualis;
c, N. chorda tympani; d, N. alveolaris inferior (cut); e, N. buccinatorius; f, Nn.
infraorbitales; g, N. sphenopalatinus; h, branch of N. oculomotorius; i, N. zygomaticus;
j, N. lachrymalis; k, N. abducens; l, N. trochlearis; m, N. frontalis; n, N.
auriculotemporalis. 1, cut end of zygomatic process of temporal; 2, tympanic bulla;
3, M. digastricus; 4, M. constrictor pharyngis medius; 5, M. hyoglossus; 6, M.
genioglossus; 7, M. geniohyoideus; 8, M. pterygoideus externus (cut); 9, cut
cranial end of zygomatic arch; 10, mandible (cut); 11, lachrymal gland.
After giving off the above branches the mandibular nerve
passes three or four millimeters laterad along the root of the
zygomatic process of the temporal bone, and divides into two
large branches, the inferior alveolar (or dental) nerve (d) and
the lingual nerve (b).
[375]
f. N. alveolaris inferior (Fig. 154, d).—This passes
toward the mandibular foramen, but before reaching it gives
off a branch to the mylohyoid and digastric muscles. It then
enters the foramen, passes through the mandibular canal, and
gives numerous inferior dental branches to the teeth. At the
mental foramen (or foramina) it passes out of the canal and
divides into several mental nerves, to the chin, lower lip, and
mucosa of the lower jaw.
g. N. lingualis (Fig. 154, b).—The lingual nerve passes
between the internal and external pterygoid muscles, and two
or three millimeters from its origin it receives the chorda tympani
nerve (c), a communicating branch from the facial (see
page 376). It passes onto the ventral surface of the external
pterygoid, thence onto the side of the tongue, beneath the
mylohyoid. On the side of the tongue it divides into branches
which enter the tongue and are distributed to its mucous membrane.
One branch passes to the mucosa of the pharynx.
Just before entering the tongue a branch is given off to the
sublingual and submaxillary glands.
VI. N. abducens.
—The sixth nerve, the abducens, arises
from the medulla, as already described (Fig. 138, VI, and page
347). It passes into the orbit through the orbital fissure, then
extends obliquely cranioventrad along the medial surface of
the lateral rectus muscle (Fig. 154, k). At about the middle
of the length of the muscle it divides into two or three branches
which enter at once into the lateral rectus and supply it.
VII. N. facialis
(Fig. 155).—The seventh or facial nerve
arises from the trapezium at the caudal margin of the pons,
craniad of the origin of the eighth nerve, as described in the
account of the brain (Fig. 138, VII). It passes into the
internal auditory meatus, traverses the facial canal through the
petrous bone, and emerges at the stylomastoid foramen.
Within the facial canal the nerve bears an enlargement,
the ganglion geniculi. It gives off within the canal a branch
to the stapedius muscle, and the superficial petrosal and
chorda tympani nerves.
The superficial petrosal (N. petrosus superficialis major)
nerve passes into the hiatus facialis, through the canal which[376]
forms its continuation, and joins the sphenopalatine ganglion
(p. 373).
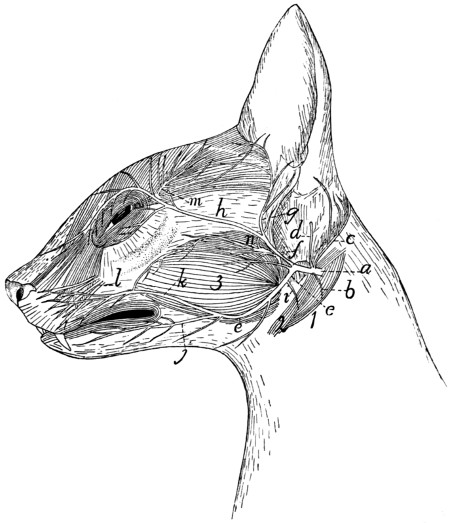
Fig. 155.—Superficial Nerves of the Face.
a, N. facialis; b, branch to digastric; c, N. auricularis posterior; d, branch to
inside of ear; e, ventral ramus; f, dorsal ramus; g, temporal nerve; h, zygomatic
nerve; i, nerve to stylohyoid muscle; j, inferior buccal nerve; k, superior buccal
nerve; l, infraorbital branches of fifth nerve; m, lachrymal branches of fifth nerve;
n, auriculotemporal branches of fifth nerve. 1, M. digastricus; 2, M. stylohyoideus;
3, M. masseter.
The chorda tympani is given off two or three millimeters
before the emergence of the facial at the stylomastoid foramen.
It passes into the tympanic cavity, extends across it between
the malleus and incus, passing close against the tensor tympani
muscle, and leaves it (by a small cleft, the canal of Huguier)
between the bulla and the squamous portion of the temporal.
It then passes craniad (Fig. 154, c),
ventrad of the root of the[377]
zygomatic process of the temporal, and joins the lingual nerve
(Fig. 154, b) (branch of the third division of the fifth nerve).
On emerging from the stylomastoid foramen the facial
nerve (Fig. 155, a) gives off at once a small branch (b) to the
digastric muscle (1), and a larger branch, N. auricularis
posterior (c), which passes dorsocaudad about the base of the
ear, supplying some of its muscles. Another small branch (d)
pierces the cartilaginous external ear and is distributed to its
inner surface. The facial nerve then curves craniad about the
proximal part of the cartilaginous external ear, and divides five
or six millimeters from its emergence into two main branches,
which may be designated as the dorsal (f) and ventral (e)
rami. A third small branch may also rise from the point of
union of the two; it is usually, however, a twig from the dorsal
ramus, and passes to M. submentalis.
The dorsal ramus (f) sends two or three small branches
to the cheek (including the one just mentioned), and divides
into the temporal (g) and the zygomatic (h) branches. The
former (g) passes along the cranial margin of the external ear,
supplying the superficial muscles, and extends into the temporal
region, where it lies deeper than the terminal branches
of the lachrymal nerve (m) (from the fifth cranial). The
zygomatic branch (h) passes across the malar bone to the
caudal angle of the eye, sends branches into both eyelids,
anastomoses with twigs from the lachrymal branch (m) of the
fifth nerve, and passes along the medial side of the eye to the
lateral surface of the nose, where it ramifies.
The ventral ramus (e) gives off a minute branch (i) to the
stylohyoid muscle (2), then proceeds toward the angle of the
mouth and divides into superior (k) and inferior (j) buccal
branches, the former to the muscles of the upper lip and the
contiguous regions, the latter to those of the lower lip and
chin.
The seventh nerve thus supplies most of the muscles of the
head except those of mastication, and of these it supplies the
digastric.
VIII. N. acusticus.
—The eighth or auditory nerve takes
origin (Fig. 138, VIII) from the floor of the fourth ventricle,[378]
as before described (page 347). It passes into the internal
auditory meatus and is distributed to the internal ear within
the petrous bone.
IX. N. glossopharyngeus
(Fig. 156, a).—The ninth or
glossopharyngeal nerve takes origin (Fig. 138, IX) from the
side of the medulla, as described in the account of the brain.
Close to its origin its trunk bears a small enlargement, the
ganglion superius. It passes along with the tenth and
eleventh nerves through the jugular foramen. At its exit it
has a ganglionic enlargement, the ganglion petrosum; this is
connected by fine fibres with the ganglion nodosum (Fig. 156,
d) of the vagus. The glossopharyngeal (Fig. 156, a) then
passes craniad over the surface of the tympanic bulla (15) and
mediad of the digastric muscle. It continues mediad of the
carotid artery, and as it approaches the cranial cornu of the
hyoid divides into two portions, one of which passes to the
muscles and mucosa of the pharynx, and the other to the
tongue, where it is the special nerve of taste.
X. N. vagus.
—The vagus nerve arises from the side of
the medulla in the manner described under the Brain (page
346 and Fig. 138, X). It passes through the jugular foramen
along with the glossopharyngeal and accessory nerves.
Cervical Portion of the Vagus (Fig. 156, d, d′).—In the
foramen, or just before entering it, it presents a ganglionic
enlargement, the ganglion jugulare (or “ganglion of the
root”), and a short distance beyond the foramen it forms a
second ganglion, the ganglion nodosum (d) (or “ganglion of
the trunk”), which lies dorsocaudad of the superior cervical
sympathetic ganglion (e). The ganglia of the vagus and sympathetic
are closely bound together by connective tissue, and
that portion of the vagus craniad of the ganglion nodosum is
interconnected by a network of nervous fibres with the sympathetic
(e), hypoglossal (b), and accessory (c) nerves. From
the ganglion nodosum the vagus (d′) passes caudad, closely
bound up with the sympathetic; the two lie at the side of the
common carotid artery. Just before entering the thorax the
two separate, the vagus being the larger and lying ventrad of[379]
the sympathetic. (In rare cases the two are distinctly separated
throughout their length.)
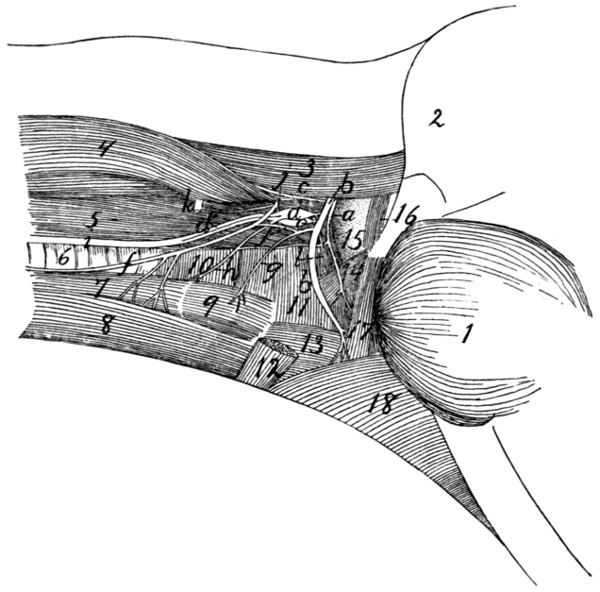
Fig. 156.—Glossopharyngeal,
Hypoglossal, Vagus, Sympathetic, and
First Cervical Nerves in the Neck.
a, N. glossopharyngeus; b, N. hypoglossus; c, N. accessorius; d, ganglion nodosum
of vagus nerve; d′, N. vagus; e, ganglion cervicale superius of sympathetic;
f, ramus descendens of N. hypoglossus (united with first cervical); g, branch to M.
thyrohyoid from ramus descendens of N. hypoglossus; h, N. laryngeus superior of
N. vagus; i, united vagus and sympathetic nerves; j, first cervical nerve; k, second
cervical nerve; l, N. pharyngeus from vagus. 1, M. masseter; 2, outline of external
ear; 3, M. splenius; 4, M. levator scapulæ ventralis; 5, M. longus capitis; 6, trachea;
7, M. sternothyreoideus; 8, M. sternohyoideus; 9, M. thyreohyoideus; 10, M. constrictor
pharyngis inferior; 11, M. constrictor pharyngis medius; 12, cut end of M.
stylohyoideus; 13, M. hyoglossus; 14, M. stylopharyngeus; 15, bulla tympani; 16,
M. jugulohyoideus; 17, M. styloglossus; 18, M. mylohyoideus.
Branches of the vagus in the cervical region (Fig. 156).
a. N. auricularis.—This leaves the ganglion jugulare,
passes into the petrous bone to the facial canal, leaves the skull
with the facial nerve by the stylomastoid foramen, and is distributed
to the external ear.
b. N. pharyngeus (l).—This leaves the vagus craniad of
the ganglion nodosum, passes ventrad, sends a small communicating
branch to N. laryngeus superior (h), and is distributed[380]
to the pharyngeal muscles and cranial part of the
œsophagus.
c. N. laryngeus superior (h).—This arises from the ganglion
nodosum, passes ventrad, crossing the medial surface of
the carotid artery, and reaches the larynx at about the level
of the caudal end of M. thyreohyoideus. It passes into the
larynx and supplies its mucosa.
Thoracic Portion of the Vagus (Fig. 157).—As the vagus
(b) and sympathetic (c) separate to enter the thorax, the vagus
(b) lies more ventrad. The right vagus now lies along the
lateral surface of the trachea, the left one (Fig. 157) along that
of the œsophagus. At about the level of the first rib or further
craniad there arise, apparently from the vagus (but really from
the sympathetic), two slender nerves which pass caudad parallel
and close to the vagus, lying slightly dorsad of it. These are
the cardiac nerves (i); they arise from the sympathetic farther
craniad in the neck region, accompany the vagus, and seem
to branch from it. In cases where vagus and sympathetic are
separate throughout their courses, these nerves arise clearly
from the sympathetic. The cardiac nerves pass to the heart
and aid in forming a network of nerves known as the cardiac
plexus (k).
The vagus usually receives also, at about the level of the
first rib, one or two communicating branches from the middle
cervical ganglion (d) of the sympathetic. It then passes to
the roots of the lungs, crossing the lateral surface of the aortic
arch on the left side, and the medial surface of the azygos vein
on the right side. As it crosses the aortic arch the left vagus
gives off N. laryngeus inferior (j). This curves around
the caudal side of the aortic arch to the lateral surface of the
trachea (o), then extends on the lateral and ventral surface of
the trachea craniad into the neck region. In the neck it passes
to the dorsolateral side of the trachea and reaches the larynx.
It passes into the larynx between the cricoid and thyroid cartilages,
and supplies the muscles of the larynx. It anastomoses
with the superior laryngeal.
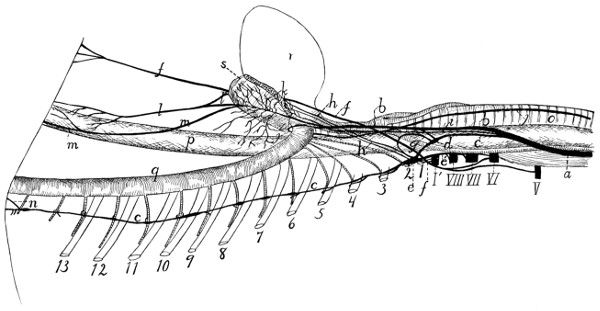
Fig. 157.—Sympathetic,
Vagus, and Phrenic Nerves in the Neck and Thorax.
1-13, the thirteen ribs (cut); V-VIII, the
fifth to eighth cervical nerves (cut); I′, first thoracic nerve (cut). a, combined trunk of vagus
and sympathetic; b, vagus; c, sympathetic; d, middle cervical
ganglion of sympathetic; e, inferior cervical ganglion; e′, communicating
branch to sixth, seventh, and eighth cervical nerves; f, phrenic nerve;
g, loop of sympathetic about subclavian artery, between the middle
and inferior cervical ganglia; h, cardiac branches from the inferior cervical
ganglion; i, cardiac branch passing from sympathetic along
with vagus to heart; j, inferior laryngeal nerve; k, pulmonary and
cardiac plexus; l, ventral œsophageal branch of vagus; l′, similar branch
from the opposite side; m, dorsal œsophageal branch of vagus; n,
great splanchnic nerve; o, trachea; p, œsophagus; q, aorta; r, heart;
s, root of lung.
The right inferior laryngeal nerve is given off from the
right vagus much further craniad than the left; it curves around[381]
[382]
the right subclavian artery just caudad of the origin of the
internal mammary artery, then passes to the trachea, and has
a course and distribution like that of the left side.
On reaching the root of the lungs the vagus divides into
numerous branches which form the pulmonary plexus (k) over
the roots of the lungs, and extends onto the basis of the heart
as the cardiac plexus. From these plexuses numerous
branches pass to the heart, lungs, pulmonary artery, pericardium,
etc. The cardiac branches from the sympathetic, mentioned
above, are also connected with the cardiac plexus.
Caudad of the pulmonary plexus the vagus is continued
on each side as two trunks, a dorsal (m) and a ventral (l)
which pass along the œsophagus (p). The ventral branches
of right and left vagi (l and l′) unite a short distance caudad
of the root of the lung, and the single trunk extends into the
abdominal cavity on the ventral surface of the œsophagus.
The dorsal branches of right and left vagi unite farther back,
near the diaphragm, and the single trunk thus formed (m)
enters the abdominal cavity on the dorsal surface of the
œsophagus. Both divisions give fine nerves to the œsophagus.
Abdominal Portion of the Vagus (Fig. 164, page 407).—After
passing through the diaphragm the ventral division (Fig.
164, l) reaches the lesser curvature of the stomach (1), over
which it ramifies, some branches being traceable almost to the
pylorus. The network of branches thus formed is the anterior
gastric plexus. Some twigs from the ventral division pass
transversely across the cranial end of the stomach and join the
plexus formed by the dorsal division.
The dorsal division (m) of the vagus reaches the greater
curvature of the stomach, where it ramifies, forming the posterior
gastric plexus. Branches from this anastomose with
the anterior gastric plexus and with the cœliac plexus of the
sympathetic (e).
XI. N. accessorius.
—The accessory (or spinal accessory)
nerve arises (Fig. 138, XI) by numerous rootlets from the
lateral surface of the medulla and of the spinal cord as far
caudad as the fifth to seventh cervical nerve. These spinal
rootlets join to form a nerve which enters the cranium through[383]
the foramen magnum. After receiving the rootlets from the
medulla, it leaves the cranial cavity along with the vagus and
glossopharyngeal by the jugular foramen. Just outside the
foramen (Fig. 156, c) it becomes involved in a plexus of fine
branches which interconnect it with the vagus, sympathetic, and
hypoglossal. It then turns laterad and caudad (Fig. 158, 1)
and pierces the cleidomastoid muscle (d), to which it gives
small branches. It then divides: one branch enters the
sternomastoid muscle; the other passes caudad along the
dorsal border of the levator scapulæ ventralis, sends branches
to the clavotrapezius, and may be traced to the acromiotrapezius
and spinotrapezius, which it supplies.
XII. N. hypoglossus
(Fig. 156, b).—The twelfth or
hypoglossal nerve arises from the ventral side of the medulla
(Fig. 138, XII), as previously described. It emerges from
the skull through the hypoglossal canal. At first it passes
ventrad, then gradually turns craniad, following thus a curved
course and passing successively laterad of the vagus and sympathetic
nerves (Fig. 156, d and e), the common carotid artery,
and the cranial cornu of the hyoid bone. A short distance
from its origin the nerve gives off a ramus descendens (Fig.
156, f), which receives a communicating branch from the first
cervical nerve (j), and divides into two branches, one (g) to
the thyrohyoid muscle (9), the other to the sternohyoid (8) and
sternothyroid muscles (7). The main nerve passes beneath the
mylohyoid muscle into the tongue, at first following the lingual
artery, then laterad of it, then crossing it again. It sends
branches to all the muscles of the tongue, and may be traced
to its tip.
The hypoglossal nerve is involved in the plexus just outside
of the jugular foramen, with which the vagus, sympathetic, and
accessory nerves are also connected.
2. Spinal Nerves.
—The origin and general features of the
spinal nerves have been described in connection with the
account of the spinal cord (page 337). A description of their
peripheral distribution will now be given.
A. Cervical Nerves.
—There are eight pairs of cervical
nerves. The first leaves the vertebral canal through the[384]
atlantal foramen. The second passes out between the arches
of the atlas and axis, not through a special intervertebral foramen,
so that its ganglion lies among the muscles of the back
of the neck. The others emerge through the intervertebral
foramina, the eighth one from between the last cervical and
first thoracic vertebræ.
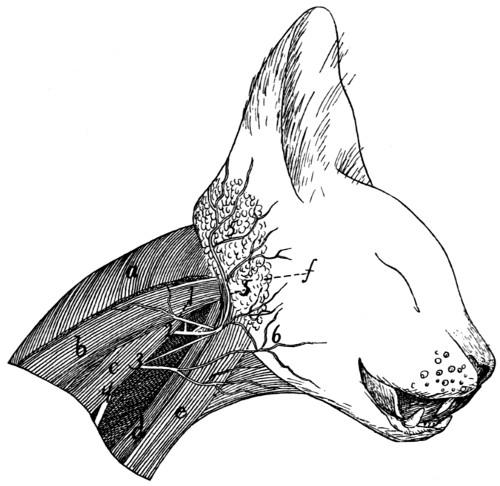
Fig. 158.—Superficial Nerves of the Neck.
M. clavotrapezius has been partly removed. 1, N. accessorius; 2-4, ventral
rami of second to fourth cervical nerves; 5, N. auricularis magnus; 6, N. cutaneus
colli. a, M. clavotrapezius (cut); b, M. splenius; c, M. levator scapulæ ventralis;
d, M. cleidomastoideus; e, M. sternomastoideus; f, parotid gland.
Dorsal Rami (Rami posteriores).—The dorsal ramus of the
first nerve (N. suboccipitalis) supplies the short dorsal muscles
which move the head and connect the atlas and skull. In
the second nerve the dorsal ramus is much larger, forming
N. occipitalis major. It sends small branches to the muscles
about its origin, then turns craniad on the surface of the obliquus
superior muscle, passes through the biventer cervicis and
splenius, joins a small branch from the third nerve, and reaches
the dorsal surface of the back of the head. It passes craniad,
lying beneath the levator auris longus, emerges from between[385]
the two divisions of this muscle, and supplies the skin and
cutaneous muscles between the two external ears. The dorsal
rami of the other cervical nerves supply muscles and integument
on the back of the neck.
Ventral Rami (rami anteriores).—These pass ventrad
between the transverse processes of the vertebræ, except in the
case of the first nerve (Fig. 156, j). This passes ventrad from
the atlantal foramen along the groove for the vertebral artery,
through the notch in the wing of the atlas, and across the lateral
surface of the longus capitis muscle (5). Here it sends a
branch caudad to join the second cervical (k), then crosses the
vagus (d′) and sympathetic and the carotid artery, giving off
communicating branches to the vagus and sympathetic, and
uniting with a branch of the descending ramus of the hypoglossal
(f). At the lateral surface of the larynx it turns
caudad, following the lateral border of the sternothyroid muscle
(7), and is distributed to the sternohyoid (8) and sternothyroid
(7).
The ventral ramus of the second nerve (Fig. 158, 2)
receives a branch from that of the first nerve, passes laterad
between the levator scapulæ ventralis (c) and the cleidomastoid
(d), receives a communicating branch from the third cervical
(3), sends a branch to N. accessorius (1) and numerous small
nerves into the sternomastoid (e) and cleidomastoid (d), then
turns craniad and divides into N. auricularis magnus (5) and
N. cutaneus colli (6).
N. auricularis magnus (5), the great auricular nerve,
passes dorsocraniad across the lateral surface of the sternomastoid
(e) to the lateral and caudal surface of the external ear and
parotid gland (f), where it ramifies. The cutaneus colli (6)
is the smaller, ventral, division of the second nerve; it may
receive also an accession from the third. It passes to the
integument over the ventral part of M. masseter and ventrad
of that muscle.
The third nerve (Fig. 158, 3) communicates with the second
and supplies the levator scapulæ ventralis (c), cleidomastoid
(d), sternomastoid (e), longus capitis, and other muscles of this
region and aids in forming the cutaneus colli (6).
The fourth[386]
(4) and fifth are distributed to the muscles and integument of
the sides of the neck. A branch of the fourth supplies the integument
in the hollow of the shoulder, and one from the fifth
follows the vena cephalica and supplies the integument over
the shoulder. The fifth by sending a branch to aid in forming
the phrenic nerve (Fig. 157, f) may be considered to enter
partly into the brachial plexus.
Owing to the intercommunicating branches between the
ventral roots of the first five cervical nerves, these are sometimes
considered as forming a loose plexus which receives the
name cervical plexus.
The sixth, seventh, and eighth cervical nerves (with a part
of the fifth) become interconnected with each other and with
the first thoracic to form the brachial plexus.
The Brachial Plexus
(Fig. 159).—The brachial plexus is
formed by the ventral rami of the fifth, sixth, seventh, and
eighth cervical nerves and the first thoracic. Of the fifth cervical
only a small part enters into the plexus, forming part of
the phrenic nerve. The formation of the plexus is due to the
union of the different nerves by means of strong connecting
branches or ansæ. The plexus lies in the axilla, along with
the axillary artery and vein; all its component nerves pass
laterad in front of the first rib. Its branches supply the arm
and shoulder.
The precise arrangement of the different strands is
somewhat variable. The plexus is commonly made up in
approximately the following manner (Fig. 159). From the
fifth cervical nerve (V) a small branch joins a similar one
from the sixth to form the phrenic nerve (a); the remainder of
the fifth does not enter into the plexus. From the sixth cervical
(VI) arise parts of the phrenic nerve (a), the suprascapular
(b), the cranial one of the three subscapular nerves (c), the
axillary (d), and the musculocutaneous (f). The sixth also
gives off close to its origin a nerve (b′) which passes to the
inner surface of the levator scapulæ and ramifies over the surface,
supplying this muscle and extending to the rhomboideus,
which it also innervates. The seventh cervical (VII) is the
largest nerve entering into the plexus; it furnishes parts of one[387]
or both of the anterior thoracic nerves (k and n), of the posterior
thoracic (m), the three subscapular nerves (c, e, and i),
the axillary (d), musculocutaneous (f), radial (h), and median
(g) nerves. The eighth cervical (VIII) supplies parts of one
of the anterior thoracic nerves (k), the caudal one of the subscapular
nerves (i), the radial (h), median (g), and ulnar (j)
nerves. The first thoracic (1) curves craniad, so as to leave
the thorax on the cranial side of the first rib; it supplies the
median cutaneous nerve (l) and parts of the radial (h), ulnar
(j), and median (g) nerves; sometimes also a part of one (k)
of the anterior thoracic nerves.
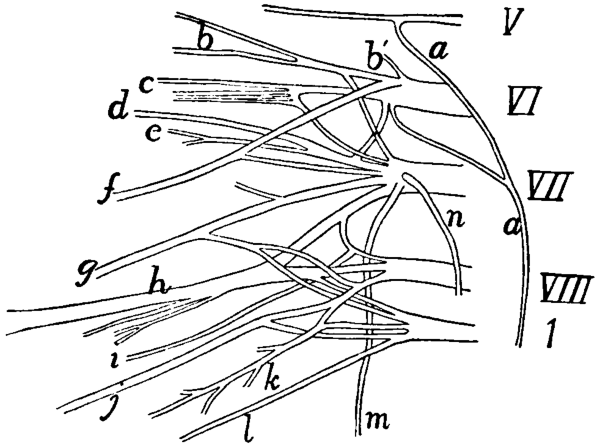
Fig. 159.—Diagram of the Right Brachial Plexus.
V, VI, VII, VIII, the fifth to eighth cervical nerves. 1, the first thoracic
nerve. a, phrenic nerve; b, suprascapular; b′, nerve to serratus anterior and levator
scapulæ muscles; c, first or cranial subscapular nerve; d, axillary nerve; e, second
subscapular; f, musculocutaneous; g, median; h, radial; i, third subscapular; j, ulnar;
k, second anterior thoracic; l, medial cutaneous; m, posterior thoracic; n, first
anterior thoracic.
The following are the nerves which arise from the brachial
plexus:
A. Nerves of the Shoulder and Breast (Fig. 160).
1. Nn. thoracici anteriores.—There are two anterior
thoracic nerves. One (Fig. 160, h; Fig. 159, n) arises from the
seventh cervical and passes to the pectoral muscles (9) along
with the anterior thoracic artery. The other (Fig. 159, k; Fig.
160, q and r) arises usually by two roots, one from the eighth cervical,
one from the first thoracic; it passes along with the long
thoracic artery to be distributed to the pectoralis muscles (Fig.
160, 8) and sometimes also to the latissimus dorsi (7).
[388]
2. N. thoracicus posterior.—The posterior thoracic (or
long thoracic) nerve (external respiratory) (Fig. 159, m) arises
from the seventh cervical nerve near its beginning. It passes
within the scalenus muscle and extends caudad on the outer
surface of the serratus anterior muscle, which it supplies.
3. N. suprascapularis (Figs. 159 and 160, b).—The suprascapular
nerve arises from the sixth (and sometimes the seventh)
cervical. It passes laterad and gives off a branch which passes
over the shoulder-joint and penetrates the clavodeltoid muscle
to be distributed to the integument on the ventral surface of
the upper arm. It then follows the transversa scapulæ artery
into the supraspinatus fossa and accompanies the artery to be
distributed to the supraspinatus and infraspinatus muscles.
4. Nn. subscapulares.—The subscapular nerves are three.
The cranial one (Figs. 159 and 160, c) arises from the sixth and
seventh cervical and supplies the subscapular muscle. The
middle one (Fig. 159, e) arises from the seventh; it supplies
principally the teres major. The caudal one (Fig. 159, i) arises
from the seventh and eighth cervical and supplies the latissimus
dorsi. (The points of origin from the plexus vary.)
5. N. axillaris (or circumflexus) (Fig. 159, d).—The axillary
nerve arises from the sixth and seventh cervical nerves.
It passes toward the shoulder-joint, then follows the posterior
circumflex artery ventrad of the long head of the triceps to be
distributed to the spinodeltoid and acromiodeltoid. A branch
of it continues to the clavobrachial, which it supplies.
B. The Phrenic Nerve (Fig. 157, f, page 381).
6. N. phrenicus.—The phrenic nerve (internal respiratory)
is formed by the junction of two slender branches, one from
the fifth and one from the sixth cervical nerves (Fig. 159, a).
(It is said to receive sometimes a branch from the fourth.) It
passes caudad into the thorax on the ventral surface of the
subclavian artery and then extends to the diaphragm lying
at the side of the inferior and superior venæ cavæ. It is the
motor nerve of the diaphragm.
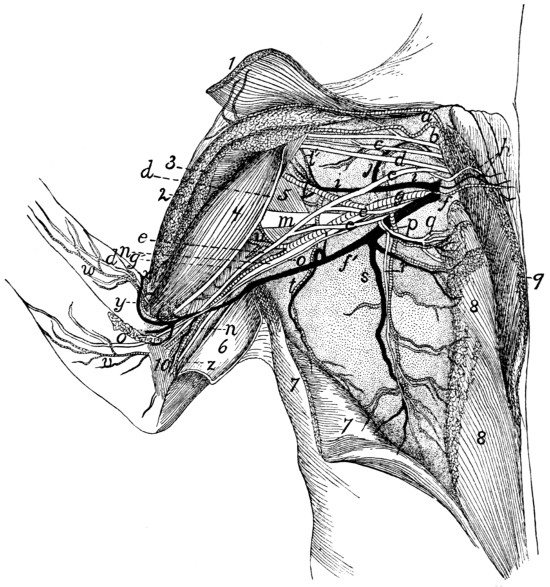
Fig. 160.—Blood-vessels
and the more Ventral Nerves of the Axilla,
Ventral View.
The pectoral and clavobrachial muscles have been cut and laid aside, their ends
being shown; only a part of the nerves are exhibited. 1, M. clavobrachialis; 2, cut
end of M. pectoralis major; 3, cut end of M. pectoralis minor; 4, M. biceps; 5, M.
teres major; 6, M. epitrochlearis, partly cut and turned back; 7, M. latissimus dorsi,
partly cut; 8, M. pectoralis minor; 9, M. pectoralis major; 10, short portion of
caput mediale of M. triceps brachii. a, branches of the thyrocervical axis to clavobrachial
and clavotrapezius muscles; b, suprascapular nerve; c, first subscapular
nerve; d, musculocutaneous nerve; e, median nerve; f, V. axillaris; g, A. axillaris;
h, first anterior thoracic nerve, accompanied by the anterior thoracic artery and vein;
i, V. subscapularis; j, large muscular branch (to subscapular muscle) of the subscapular
vein and brachial artery; l, A. subscapularis; l′, A. circumflexa humeri
anterior; m, radial nerve; n, ulnar nerve; o, medial cutaneous nerve; p, A. thoracica
longa; q, r, second anterior thoracic nerve; s, V. thoracica longa; t, A. and
V. thoracicodorsalis; u, A. profunda brachii; v, branch of brachial artery accompanying
medial cutaneous nerve; w, branch of A. collateralis radialis superior; x, A.
collateralis radialis superior; y, V. mediana cubiti; z, A. collateralis ulnaris.
C. Nerves of the Arm.
7. N. musculocutaneus (Fig. 160, d).—The musculocutaneous
nerve or external cutaneous arises from the ventral[389]
[390]
surface of the sixth and seventh nerves (Fig. 159, f). It passes
toward the shoulder-joint and supplies the biceps (Fig. 160, 4)
and coracobrachialis muscles. It then passes distad, resting on
the dorsal border of the biceps (4), and supplies the brachialis
muscle. Thence it passes dorsad of the biceps to the lateral
side, and reaches the integument by passing between the clavobrachialis
and pectoantibrachialis muscles near the elbow; it is
then distributed to the skin on the radial side of the ventral surface
of the forearm, as far as the wrist (Fig. 130, h, page 319).
8. N. cutaneus medialis (Fig. 160, o).—The medial (or
internal) cutaneous nerve rises from the first thoracic. It
passes distad along the medial side of the biceps, and at the
junction of the second and third thirds of the upper arm reaches
the integument by passing between the epitrochlearis and
pectoantibrachialis muscles. It then curves spirally about the
dorsal border of the forearm and is distributed to the integument
of the forearm on the ulnar side, extending nearly to the
wrist.
9. N. medianus (Fig. 160, e).—The median nerve is
formed by the junction of three branches, one each from the
seventh and eighth cervical with the brachial artery (g) passing
between them, and one from the first thoracic nerve. It
accompanies the brachial artery (g) lying on its medial side,
and passes with it through the supracondyloid foramen; here
it is connected by a branch to the musculocutaneous nerve.
It passes into the forearm (Fig. 161, b), lying at first beneath
the pronator teres (5). Here it gives branches to the pronator
teres and flexor muscles (flexor carpi radialis, palmaris longus,
five heads of the flexor profundus digitorum, but not to the
flexor carpi ulnaris). Continuing along the forearm, lying on the
flexor carpi radialis, it sends a posterior interosseous nerve to
the pronator quadratus muscle and branches to the deep and
superficial divisions of the flexor sublimis digitorum. It thus
supplies the flexor and pronator muscles (except the flexor
carpi ulnaris). It then passes into the palm beneath the transverse
ligament and gives rise to three principal branches. The
first of these supplies the integument of the thumb, sending a
branch on either side of it; and onto the radial side of the[391]
[392]
second digit. The second passes along the contiguous sides
of the second and third digits and supplies their integument.
The third passes along the contiguous sides of the third and
fourth digits to their integument. Each sends twigs into the
trilobed pad in the palm, so that there are thus seven terminal
branches in the palm. From these, branches (probably) pass
to the three radial Mm. lumbricales.
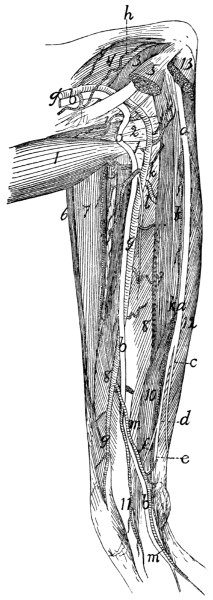
Fig. 161.—Nerves
and Arteries of the Forearm, seen from the
Flexor Side.
Mm. palmaris longus, flexor carpi radialis, flexor carpi ulnaris, and the third
and fourth heads of M. flexor profundus digitorum have been removed; also part of
the pronator teres. 1, M. biceps; 2, conjoined tendon of M. brachialis and M.
clavobrachialis; 3, short portion of caput mediale of M. triceps; 4, intermediate
portion of caput mediale of M. triceps brachii; 5, cut ends of M. pronator teres;
6, M. brachioradialis; 7, M. extensor carpi radialis longus and brevis; 8, fifth head
of M. flexor profundus digitorum; 9, second head of M. flexor profundus digitorum;
10, radial part of M. flexor sublimis digitorum; 11, common tendon of M. flexor
profundus digitorum; 12, first head of M. flexor profundus digitorum; 13, cut origin
of M. flexor carpi ulnaris. a, ulnar nerve; b, median nerve; c, dorsal cutaneous
branch of the ulnar nerve; d, palmar branch of ulnar nerve; e, deep palmar
branch; f, superficial palmar branch; g′, A. brachialis; g, A. radialis; h, A. collateralis
ulnaris superior; i, A. radialis recurrens; j, A. ulnaris recurrens; k, A. ulnaris;
l, A. interossea anterior; m, large branch of A. radialis, joining A. ulnaris.
10. N. radialis.—The radial (or musculospiral) nerve is
formed by the junction of branches from the seventh and eighth
cervical and first thoracic nerves. It immediately sends
branches to the epitrochlearis, long head of the triceps, and
long and intermediate portions of the medial head of the triceps.
It then passes between the long and intermediate portions of the
medial head of the triceps, following the profunda brachii artery,
and curves about the humerus to its cranial side. It supplies
there the lateral head of the triceps and the anconeus, and,
lying on the brachialis muscle, divides into a superficial and a
deep branch. The superficial branch is the superficial radial
nerve (Fig. 130, g); the deep one forms the dorsal (or posterior)
interosseous nerve. The former is sensory and the latter
motor.
The superficial radial nerve (Fig. 130, g, page 319)
becomes cutaneous at the junction of the second and third thirds
of the upper arm, where it emerges from between the lateral
head of the triceps and the brachialis muscle. It then follows
the course of the vena cephalica (c) to the wrist and follows the
dorsal tributary of the same vein onto the dorsum of the hand.
It supplies the integument of the distal part of the ventral surface
of the upper arm and that along the forearm. On the
hand it is distributed to the dorsal surface in the same manner
as the median nerve on the ventral surface, i.e. to the integument
of the thumb on both sides and to that of the medial side
of the second digit; to that of the contiguous sides of the
second and third digits and of the contiguous sides of the third
and fourth digits. There are thus seven terminal branches,
one for each of these regions.
The dorsal interosseous nerve follows the brachialis muscle
and passes onto the forearm between it and the extensor carpi[393]
radialis longus. It supplies the supinator and extensor
muscles of the forearm (brachioradialis, supinator, extensor
carpi radialis longus, extensor carpi radialis brevis, extensor
communis digitorum, extensor digitorum lateralis, extensor
indicis, extensor brevis pollicis). The dorsal interosseous
and radial nerves thus supply all the extensor muscles of
the forearm and upper arm and the supinators (modified
extensors).
11. N. ulnaris (Fig. 160, n).—The ulnar nerve arises from
the eighth cervical and first thoracic and accompanies the
brachial artery (g) and median nerve (e) through the upper
arm. It does not pass through the supracondyloid foramen,
but curves over the dorsal end of the medial epicondyle of the
humerus within the short division of the medial head of the
triceps, and reaches the dorsal border of the forearm. In the
forearm (Fig. 161, a) it lies beneath the flexor carpi ulnaris.
It supplies muscular branches to the flexor carpi ulnaris and
the first or ulnar head of the flexor profundus digitorum. Near
the middle of the forearm it divides into two branches, dorsal
cutaneous (c) and palmar (d).
The dorsal cutaneous branch (c) curves about the ulnar
side of the wrist to the dorsum of the hand and divides into
twigs which supply the dorsal part of the ulnar side of the fifth
digit and of the contiguous sides of the fourth and fifth. The
palmar branch (d) sends a twig to the integument on the flexor
surface of the arm near the wrist, and then divides into deep
palmar (e) and superficial palmar (f) branches.
The superficial palmar branch (f) passes into the palm
and supplies the ventral portion of the integument on the ulnar
side of the fifth digit, and on the contiguous sides of the fifth
and fourth.
The deep palmar branch (e) passes into the palm, beneath
the ligament of the pisiform bone. Just distad of the pisiform
bone it curves toward the radial side and breaks up into a
number of small branches which are distributed to the short
muscles in the palm.
B. Thoracic Nerves.
Dorsal Rami.—The dorsal rami of the thoracic nerves are[394]
small and supply the muscles and integument of the back.
Each gives off a branch directly dorsad to the spinal muscles,
and a lateral branch which reaches the integument some distance
from the middle line.
Ventral Rami.—The ventral ramus of the first thoracic
nerve enters into the brachial plexus, as already described.
The ventral rami of the other thoracic nerves form the intercostal
nerves. Each of these passes ventrolaterad, lying close
to the caudal border of a rib, in company with the intercostal
artery. Branches are given to the intercostal muscles, and at
about the middle of the length of the rib a large lateral branch
is given off, which passes to the more superficial muscles of the
thoracic wall (serrati posteriores, obliquus externus, etc.), its
main branches running dorsad and ventrad. The main intercostal
nerve extends ventrad to the transversus costarum and
rectus abdominis, supplying these muscles.
C. Lumbar Nerves.
—There are seven lumbar nerves,
one passing from the vertebral canal caudad of each lumbar
vertebra.
Dorsal Rami.—The dorsal rami are similar to those of the
thoracic region, sending one branch dorsad to the muscles of
the vertebral column, another dorsolaterad to reach the integument
at about the lateral border of the longissimus dorsi. The
dorsal rami are somewhat smaller caudad.
Ventral Rami.—The last four lumbar nerves are interconnected
to form the lumbar or lumbosacral plexus. The first
three are distinct, and will therefore be described separately.
The first three lumbar nerves are directed strongly caudad
(as well as ventrad), so that on leaving the intervertebral foramen
they pass ventrad of the transverse process of the vertebra
immediately succeeding. Each communicates with the sympathetic
system and gives off near its origin branches to the
muscles on the ventral side of the vertebræ,—the first to the
crus of the diaphragm, the second and third (Fig. 162, II and
III) to the quadratus lumborum and psoas muscles. Each
divides three to five centimeters from its origin into a lateral
and a medial branch, the first having a more cranial, the latter
a more caudal course. The first three nerves of the cat are[395]
represented in man by the iliohypogastric and the ilioinguinal
nerves. There seems no good ground for applying these
names to two of the three in the cat, in preference to the third,
so that we shall speak of these nerves in the cat as simply the
first, second, and third lumbar nerves.
1. The first lumbar nerve arises from the intervertebral foramen
caudad of the first lumbar vertebra. Its lateral division
passes between the transversus and obliquus internus muscles,
then between the obliquus internus and externus. At about
the middle of the abdomen it pierces the obliquus externus and
is distributed to the integument of the middle of the ventral
surface of the abdomen. The medial division passes between
the obliquus internus and transversus, crosses the lateral division
of the second (lying mediad of it), and extends to the
rectus abdominis, which it supplies.
2. The second lumbar nerve (Fig. 162, a) divides, like the
others, into two branches. The lateral branch pierces the
muscles of the abdominal wall at the lateral border of the
longissimus dorsi and passes, lying just beneath the integument,
ventrocaudad nearly to the pubis. It supplies the
integument of the caudal half of the abdomen and of the fold
between the thigh and abdomen. The medial branch passes
at first almost directly caudad, lying on the medial surface of
the transversus abdominis; it curves gradually ventrad, lying
in the substance of the transversus, and reaches the rectus
abdominis three or four centimeters craniad of the pubis.
3. The third lumbar nerve (b) is not united with the fourth,
as in many animals, but remains distinct. It divides one or
two centimeters from its origin. The lateral branch passes
caudad to the outer surface of the abdominal muscles, and
extends, lying just beneath the integument, to that portion of
the abdominal wall which is partly covered by the thigh. The
medial branch passes caudad on the lateral surface of the
iliopsoas muscle, follows the iliolumbar artery for a short distance,
and gives branches to the caudal part of the transversus
and rectus abdominis muscles.
The Lumbar Plexus
(Fig. 162).—The fourth, fifth, sixth,
and seventh lumbar nerves are interconnected by short[396]
branches, forming thus the lumbar plexus. They are also
connected with the sacral plexus, so that the two are often
considered together as the lumbosacral plexus (Fig. 162).
In some other animals the first three lumbar nerves form also
a part of the plexus; but they are not connected with it in the
cat.
The fourth lumbar nerve forms the genitofemoral (c) and
part of the lateral cutaneous (d) nerves, and is connected by a
short strand with the fifth. The fifth aids in forming the
lateral cutaneous (d) and femoral (f) nerves, and is connected
by a short branch with the sixth. The sixth lumbar is large;
it forms the major part of the femoral (f) and a large part of
the obturator nerve (g), and sends a large connecting branch
caudad to join the seventh and thus pass into the sacral plexus.
The seventh passes caudad to join the sacral plexus and aid in
forming the great sciatic nerve (h); by its connection with the
sixth it aids in forming also the obturator nerve (g).
4. N. genitofemoralis (or N. lumboinguinalis) (c, c′, c″).—This
is a direct continuation of the fourth lumbar nerve. Its
medial branch (c″) passes along the medial surface of the
iliopsoas (8) and the psoas minor (9) to the external iliac
artery, accompanies this, lying on the ventral surface, from its
origin to the point where it gives off the profunda femoris, then
accompanies the latter artery and passes onto that branch of it
that spreads out under the integument of the ventral pelvic
region. It crosses the spermatic cord and ramifies in the skin
of the proximal part of the medial side of the thigh. The
lateral branch of the genitofemoralis (c′) pierces the psoas
minor (9), appearing on its ventral surface opposite the fifth
lumbar vertebra. It passes caudad on the ventral surface of
this muscle, crosses the iliolumbar artery, then turns caudolaterad,
passes through the abdominal wall, and is distributed
to the craniomedial surface of the thigh and to the adjacent
abdominal wall.
This nerve is variable in origin and in distribution. Its
lateral branch is sometimes lacking.
5. N. cutaneus femoris lateralis (d).—The lateral cutaneous
nerve arises from the connecting strand between the fourth[397]
and fifth nerves, most of its fibres coming from the fifth. It
passes caudoventrad between the iliopsoas and psoas minor and
accompanies the iliolumbar artery across the ventral surface of
the iliopsoas. It pierces the abdominal wall in company with
the artery and passes onto the cranial border of the thigh. Its
terminal branches are distributed to the integument along with
the branches of the iliolumbar artery,—supplying the lateral
surface of the femur and extending as far as the knee.
6. N. femoralis (f).—The femoral is a large nerve which
rises by strong roots from the fifth and sixth lumbar nerves.
It gives branches to the iliopsoas (8) and passes through that
muscle to its ventral surface. It leaves the abdominal cavity
lying on the ventral surface of the iliopsoas, and at the point
where it pierces the abdominal wall it divides into three (or
four) branches. One (or two) of these pass to the sartorius
muscle. Another passes between the rectus femoris and vastus
medialis muscles, and divides into numerous branches which
supply these muscles and the vastus intermedius.
The third branch of the femoral is N. saphenus or the long
saphenous nerve (Fig. 127, g, page 310). This passes distad
along with the femoral artery and vein, giving a few twigs to
the integument. Where the long saphenous artery (f) separates
from the femoral artery the nerve follows the former and
passes with it along the medial side of the lower leg. At the
knee and in the lower leg it gives off a number of cutaneous
branches, and finally divides just distad of the middle of the
lower leg into two main branches. Both of these pass to the
concavity of the ankle-joint and divide into fine branches, which
form a sort of plexus on the ankle and dorsal surface of the
foot; branches from this can be traced almost to the toes.
7. N. obturatorius (Fig. 162, g).—The obturator nerve
arises from the connecting band between the sixth and seventh
nerves. It lies at first laterad, then dorsad, of the common
iliac vein, and passes almost directly caudad to the cranial
border of M. obturator internus (15). Here it turns sharply
laterad about the pubis, and passes through the obturator
foramen. It gives twigs to the obturator externus muscle, and[398]
[399]
divides into several branches, which innervate the adductor
femoris, adductor longus, pectineus, and gracilis.
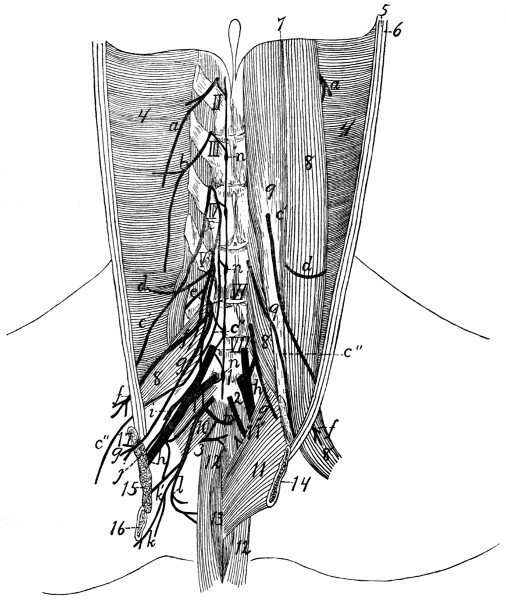
Fig. 162.—The
Lumbar and Sacral Nerves, as seen in a View of the
Dorsal Wall of the Abdominal Cavity, after Removal of the
Viscera.
On the right side the iliopsoas, psoas minor, and quadratus lumborum have been
removed. The symphysis pubis has been split and the two parts divaricated, to
show the pelvic cavity; on the right side parts of the innominate bones and the
levator ani and coccygeus muscles have been removed. II-VII, second to seventh
lumbar nerves; 1-3, first to third sacral nerves. a, second lumbar nerve; b, lateral
and medial branches of third lumbar nerve; c, N. genitofemoralis (c′, its lateral
branch; c″, its medial branch); d, N. cutaneus femoralis lateralis; e, muscular
branch of N. femoralis; f, N. femoralis; g, N. obturatorius; h, N. ischiadicus or
great sciatic nerve; i, N. gluteus superior; j, N. gluteus inferior; k, N. cutaneus
femoris posterior; l, N. pudendus; m, N. hemorrhoidalis inferior; n, part of the
sympathetic (incompletely shown, merely to indicate connections with the spinal
nerves); 4, M. transversus abdominis; 5, 6, cut edges of M. obliquus internus and
externus; 7, dorsal border of the diaphragm; 8, M. iliopsoas (mostly removed on the
right side); 9, M. psoas minor; 10, M. pyriformis; 11, M. levator ani; 11′, M. iliocaudalis;
12, M. flexor caudæ longus; 13, M. flexor caudæ brevis; 14, edge of pubic
symphysis; 15, cut end of M. obturator internus; 16, cut ramus of ischium; 17, cut
ramus of pubis.
D. Sacral Nerves and Sacral Plexus
(Fig. 162).—A
large band passes caudad from the sixth and seventh lumbar
nerves to connect with the sacral nerves; this constitutes the
so-called lumbosacral cord. The sacral nerves are three in
number. Their dorsal rami pass dorsad out of the two posterior
sacral foramina and the foramen caudad of the sacrum, to
the muscles and skin dorsad of the sacrum. The ventral rami
pass from the two anterior sacral foramina and from the foramen
between the last sacral and first caudal vertebræ. The
first is the largest; it joins the lumbosacral cord to form the
great sciatic nerve (N. ischiadicus) (h) and the superior (i) and
inferior (j) gluteal, and sends a branch caudad to connect with
the second sacral. The second and third sacral nerves are
small; they unite with the connecting branch from the first to
form a network from which arises the pudendus (l), cutaneus
femoris posterior (k), hemorrhoidalis inferior (m), and a small
branch to the great sciatic (h).
The nerves arising from the sacral plexus are the following:
1. N. gluteus superior (Fig. 162, i; Fig. 163, j).—The
superior gluteal nerve arises from the lumbosacral cord and the
first sacral nerve; it passes dorsad in the notch between sacrum
and ilium, caudolaterad of the first anterior sacral foramen,
curves over the dorsal border of the ilium between the gluteus
medius and pyriformis muscles (Fig. 163, 7), passes between
the gemellus superior (6) and gluteus minimus (5) and ventrad
of the latter, and reaches the medial surface of the tensor fasciæ
latæ, which it innervates. In its course it sends branches to
the gluteus medius, gluteus minimus (5), and gemellus
superior.
2. N. gluteus inferior (Fig. 162, j; Fig. 163, i).—A small
nerve from the lumbosacral cord and the first sacral nerve.
It passes dorsocaudad, lying on the dorsal surface of the great
sciatic nerve (Fig. 162, h). After leaving the pelvis by the
great sciatic notch it divides beneath the pyriformis (Fig.
163, 7) into two branches; one goes to M. caudofemoralis (4),
the other to M. gluteus maximus (3).
[400]
3. N. ischiadicus (Fig. 162, h; Fig. 163, a).—The great
sciatic nerve, the largest nerve in the body, arises from the
lumbosacral cord (formed chiefly by the sixth and seventh
lumbar nerves), the first sacral nerve and a small branch from
the second sacral. It passes caudodorsad and leaves the pelvis
by passing across the great sciatic notch, between M. pyriformis
and M. gemellus superior. It passes across the tendon
of M. obturator internus (Fig. 163, 9) near the insertion, and
here sends caudad a large muscular branch (b) beneath the
biceps. This branch innervates the biceps, semitendinosus (17)
and semimembranosus (16). Other twigs in this region pass to
the quadratus femoris (10) and tenuissimus. The great sciatic
nerve (a) now passes across the quadratus femoris (10), adductor
femoris (15), and semimembranosus (16), lying beneath the
biceps femoris. It thus enters the popliteal space and approaches
the popliteal artery and vein. Here it gives off one
or two small muscular branches to the distal part of the
biceps, and sends a slender branch, N. suralis (c), along the
medial surface of the biceps to the lateral surface of the lateral
head of M. gastrocnemius (20). The sural nerve (c) becomes
cutaneous at the distal (ventral) border of the biceps, and
divides three or four centimeters proximad of the ankle into
two branches. One passes over the tendon of Achilles (27) to
the proximal end of the calcaneus and ramifies in this region.
The other passes onto the lateral surface of the foot, and supplies
the integument in this region over the tarsus and part of
the metatarsus.
The great sciatic nerve now divides in the popliteal space
into the peroneus communis (d) and the tibialis (e).
Fig. 163.—Dissection of
Leg, Lateral View, to
show Distribution of the
Great Sciatic Nerve.
The biceps, caudofemoralis,
gluteus maximus, gluteus medius,
tensor vaginæ femoris,
and part of the peroneus
longus have been removed.
a, N. ischiadicus
or great sciatic
nerve; b, muscular
branch; c, N. suralis;
d, N. peroneus communis;
e, N. tibialis;
f, N. peroneus profundus;
g, N. peroneus
superficialis
and its terminal
branches; h, branches
of N. cutaneus
femoris posterior; i,
N. gluteus inferior; j,
N. gluteus superior; k,
vena saphena parva;
k′, communicating
branch to V. glutea inferior.
1, M. sartorius; 2,
cut origin of M. gluteus medius;
3, cut origin of M. gluteus
maximus; 4, cut origin of
M. caudofemoralis; 5, M. gluteus
minimus; 6, M. gemellus superior;
7, M. pyriformis; 8, M. flexor caudæ
longus; 9, M. obturator internus; 10,
M. quadratus femoris; 11, tuberosity of the
ischium; 12, great trochanter of femur; 13,
M. rectus femoris; 14, M. vastus lateralis;
15, M. adductor femoris; 16, M. semimembranosus;
17, M. semitendinosus (the slender
ribbon-like muscle crossing this is M.
tenuissimus); 18, M. vastus intermedius; 19,
M. plantaris; 20, M. gastrocnemius; 21, M.
peroneus longus (cut); 22, M. tibialis anterior;
23, M. extensor longus digitorum;
24, transverse ligament; 25, M. peroneus
brevis; 26, M. soleus; 27, tendon of Achilles;
28, M. extensor brevis digitorum; 29,
tendon of M. peroneus tertius.
a. N. peroneus communis (d).—This is the more lateral
of the two divisions of the great sciatic nerve. It passes along
the medial surface of the biceps to the lateral surface of the
lateral head of M. gastrocnemius (20), where it extends to a
point just distad of the head of the fibula. Here it passes
beneath that part of the gastrocnemius which has origin on the
fascia of the shank, passes between the soleus and peroneus
longus (21), then between the peroneus longus and peroneus
tertius. It gives off a number of small muscular
branches,[401]
[402]
then divides two or three centimeters distad of the head of the
fibula into two main branches, N. peroneus superficialis (g)
and N. peroneus profundus (f).
(1) N. peroneus superficialis (g).—The superficial peroneal
nerve passes distad between the peroneus longus and peroneus
tertius. Near the ankle it becomes superficial, passing along
the concavity of the ankle across the transverse ligament which
binds down the tendons of the extensor longus (23) and tibialis
anterior (22), and thus reaches the dorsal side of the foot. Two
or three minute twigs are given off to the ankle, then the nerve
divides into four divisions. These pass distad to the toes, subdividing
so as to send a nerve to each side of each toe.
(2) N. peroneus profundus (f).—The deep peroneal nerve
passes between the tibialis anterior and extensor longus digitorum
muscles, gives branches to these muscles, and passes
distad with the tibialis anterior artery, lying on the inner surface
of the tibialis anterior muscle. It passes onto the dorsum
of the foot, lying on the inner surface of the tendon of the
muscle. On the tarsus it divides into two branches. The
lateral branch passes into the extensor brevis digitorum,
while the medial one extends in the dorsal groove between
metatarsals four and five to the toes; here it divides into two
nerves which supply the contiguous sides of digits four and
five.
b. N. tibialis (e).—The tibial nerve passes distad parallel
with the peroneal nerve, but mediad of it. It passes between
the lateral and medial heads of the gastrocnemius, giving
off muscular branches to these, and to the plantaris and
soleus. It then passes between the plantaris and the medial
head of the gastrocnemius, and reaches the space between the
plantaris and flexor longus hallucis, where it passes distad. It
gives off, in the region just described, muscular branches to the
flexor longus digitorum, flexor longus hallucis, and tibialis
posterior. Below the middle of the lower leg the nerve
becomes superficial (Fig. 127, i, page 310), lying on the ventral
surface of the flexor longus hallucis. It passes in the
depression between the heel and the medial malleolus onto the
plantar surface of the tarsus, sends a small branch to the[403]
plantar surface of the heel, and divides into two branches, the
medial and the lateral plantar nerves.
(1) N. plantaris medialis.—The medial plantar nerve
passes along the medial border of the flexor brevis digitorum
and divides into two branches. One passes to the medial
(second) digit, supplying both sides; the other supplies the
third digit in the same way. Both send branches to the fibrous
pad on the sole of the foot.
(2) N. plantaris lateralis.—The lateral plantar nerve
passes laterad across the tendon of the flexor longus digitorum
to its lateral edge. At about the proximal end of the metatarsi
it divides into two branches. The lateral branch passes distad
to the ventral surface of the fifth digit. The medial branch
passes beneath the tendon of the flexor longus digitorum and
transversely across the interossei; it divides into numerous
tendons which innervate the interossei and other short muscles
of the sole of the foot.
4. N. pudendus (Fig. 162, l; Fig. 112, 8,
page 265).—This
arises (Fig. 162, l) from the sacral plexus caudad of the
great sciatic nerve. It has two roots, one from the second
sacral, one from the third, and it may also receive a small
accession from the great sciatic nerve (h) three or four centimeters
caudad of the origin of the latter. It passes at first
dorsolaterad, laterad of the coccygeus muscle, then curves
mediad toward the anus. It divides into two branches: one
passes to the base of the penis, sends twigs into the compressor
urethræ muscle, and passes onto the dorsal surface of the penis.
This branch is N. dorsalis penis; it extends with the artery of
the same name to the glans. The second branch of N. pudendus
(N. hemorrhoidalis medius) passes to the anus and innervates
the muscles and other structures about the caudal end of
the rectum. In the female, branches of N. pudendus supply
the urogenital sinus and adjacent structures (Fig. 112, 8).
5. N. cutaneus femoris posterior (Fig. 162, k; Fig.
163, h).—This arises from the sacral plexus, its roots coming
chiefly from the second and third sacral nerves, and passes
caudodorsad, at first in close connection with N. pudendus.
It then accompanies the posterior gluteal artery and vein, sends[404]
branches (perineal nerves) into the fat at the sides of the anus,
and branches onto the lateral surface of the biceps muscle. One
of the latter follows the communicating vein (Fig. 163, k′) from
V. saphena parva, and may be traced as far distad as the
popliteal space.
6. N. hemorrhoidalis inferior (or posterior) (Fig. 162, m).—This
arises by two roots, from the second and third sacral
nerves, and passes with the inferior hemorrhoidal artery ventrad
across the lateral surface of the rectum to the urethra. Here
it divides into two parts: one passes craniad to the bladder, the
other caudad onto the ventrolateral surface of the rectum.
A small nerve passes from the sacral plexus, especially
from the third sacral nerve, into M. levator ani (Fig. 162, 11).
Another small nerve from the same region passes to M. coccygeus,
and a third to the proximal end of M. tenuissimus.
E. Nerves of the Tail. Coccygeal Nerves.
—From
the intervertebral foramina of the first seven or eight caudal
vertebræ spinal nerves of the usual type are given off. The
dorsal rami innervate the dorsal muscles of the tail. The
ventral rami are interconnected with each other and with the
last sacral nerve by a longitudinal cord; they innervate the
muscles and integument of the ventral side of the tail.
3. The Sympathetic Nervous System. Systema nervorum
sympathicum.
The sympathetic system consists essentially of a chain of
ganglia on each side of the ventral surface of the vertebral
column, interconnected by longitudinal nerve-cords, and
stretching from the base of the skull to the tail. The ganglia
are connected to the spinal nerves by communicating branches,
and numerous branches pass from them to the abdominal and
thoracic viscera, and to the walls of the lymph- and blood-vessels,
forming complicated plexuses.
Cervical portion (Fig. 156, page 379).—The sympathetic
system begins just caudad of the tympanic bulla as the superior
cervical ganglion (G. cervicale superius) (Fig. 156, e). This
is a large ganglion closely applied to the ganglion nodosum
(d) of the vagus, and lying on its ventrocranial side. From[405]
the superior cervical ganglion the sympathetic trunk passes
caudad usually closely bound up with the vagus (d′), so that
the two can scarcely be distinguished. They lie (i) at the side
of the trachea (6), close against the lateral surface of the carotid
artery. A few centimeters before reaching the first rib the two
separate (Fig. 157, page 381); the sympathetic (c) is here the
smaller and more dorsal of the two. Just craniad of the first
rib the sympathetic usually forms a small swelling, the middle
cervical ganglion (Fig. 157, d), beyond which it divides into
two portions. The larger dorsal division passes dorsocaudad
and joins a large ganglion, the inferior cervical (e), which
lies at the head of the first rib, on the lateral surface of the
longus colli muscle. The ventral branch (g′) passes caudad,
turns laterad about the medial and caudal surface of the
subclavian artery, and likewise reaches the inferior cervical
ganglion.
In some cases the middle cervical ganglion (d) is lacking.
Branches of the Sympathetic in the Cervical Region.—From
the cranial end of the superior cervical ganglion several
nerves pass craniad. Some of these become connected immediately
with the cranial nerves emerging from the jugular
foramen, forming a plexus in this region. Others pass with
the internal carotid artery craniad, forming a plexus about that
artery; from the plexus twigs arise which pass to the fifth and
sixth cranial nerves; doubtless also to the seventh and eighth.
A specially large branch, the deep petrosal nerve (see page
373), passes from the superior cervical ganglion along with the
internal carotid artery to the pterygoid canal, thence through
this to the sphenopalatine ganglion. Before separating from
the vagus the sympathetic gives off one or two cardiac nerves
(Fig. 157, i). These are closely bound up with the vagus (b), so
that they appear usually to have origin from the vagus at about
the level of the first rib. These pass with the vagus to the
heart and take part in the formation of the cardiac plexus (k).
At the middle cervical ganglion (d) usually a communicating
branch is given to the vagus. The branches of the inferior
cervical ganglion (e) fall in the thoracic region, and are
described in that connection.
[406]
Thoracic Portion of the Sympathetic (Fig. 157, page 381).—The
inferior cervical ganglion (e) lies entirely within the
thoracic cavity. It has the following branches:
1. One or two small cardiac branches (h), which pass to
the heart (r).
2. A large communicating branch (e′), which passes
craniad from the cranial end of the ganglion (e) along the
lateral surface of the longus colli muscle, and gives communicating
branches to the sixth, seventh, and eighth cervical
nerves (VI, VII, and VIII).
3. One or two small communicating branches to the first
thoracic nerve (I′).
4. A communicating branch to the second thoracic nerve.
The arrangement of these communicating branches is
somewhat variable; they may arise in various combinations, as
at first united, or as more or less completely separated nerves.
The sympathetic trunk (c) continues caudad from the caudal
end of the inferior cervical ganglion, lying at first on the lateral
surface of the longus colli, then on the lateral surface of the
centra of the vertebræ. For each vertebra there is a slight
enlargement, forming one of the thoracic ganglia of the sympathetic,
and from each ganglion one or two communicating
branches are given off to the corresponding spinal nerve. Just
caudad of the last rib (but while still in the thoracic cavity)
the sympathetic gives off the large N. splanchnicus major or
great splanchnic nerve (n), which passes ventrocaudad and
pierces the diaphragm. The sympathetic itself then passes
through the diaphragm laterad of the crus.
Abdominal Portion of the Sympathetic (Fig. 164).—The
great splanchnic nerve (b) passes through the diaphragm (2)
laterad of the crus, extends two or three centimeters caudoventrad,
and joins a large ganglion lying just caudad of the cœliac
artery (4), almost on the lateral surface of the superior mesenteric
(5). This is the cœliac (or semilunar) ganglion (d).
From it nerves branch in all directions, forming the cœliac
plexus (c), a part of the large solar plexus. Less than a
centimeter ventrocaudad of the cœliac ganglion, on the caudal
surface of the superior mesenteric artery (5), is the somewhat[407]
smaller superior mesenteric ganglion (f); the two are connected
by two strong nerve-trunks.
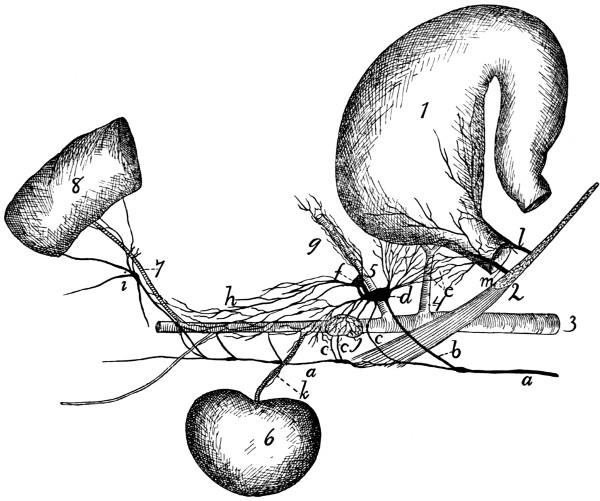
Fig. 164.—Sympathetic
and Vagus in the Abdomen (somewhat Schematic).
1, stomach; 2, crus and cut edge of diaphragm; 3, aorta; 4, cœliac artery; 5,
superior mesenteric artery; 6, kidney; 7, inferior mesenteric artery; 8, large intestine.
a, main trunk of sympathetic; b, great splanchnic nerve; c, lesser splanchnic
nerves; d, cœliac (or semilunar) ganglion; e, cœliac plexus; f, superior mesenteric
ganglion; g, superior mesenteric plexus, following the artery; h, aortic plexus; i,
inferior mesenteric ganglion and plexus; j, suprarenal plexus on suprarenal body; k,
renal plexus following renal artery; l, ventral œsophageal branch of vagus, forming
anterior gastric plexus; m, dorsal œsophageal branch of vagus, forming posterior
gastric plexus.
The cœliac ganglion (d) receives from the sympathetic (a),
in addition to the great splanchnic nerve, two (or three) lesser
splanchnic nerves (c), which arise from the main trunk of the
sympathetic, the first just as it reaches the abdominal cavity,
the second one or two centimeters caudad of the diaphragm.
From the cœliac and superior mesenteric ganglia a network
of nerves passes in all directions to the abdominal viscera.[408]
These nerves follow especially the arteries, forming plexuses
over their surfaces, and passing with them to the organs which
they supply. The cœliac plexus anastomoses with the posterior
gastric plexus of the vagus. Parts of the cœliac plexus
following the branches of the cœliac artery may be distinguished
as hepatic plexus, splenic plexus, etc. A dense
plexus is formed on the surface of the suprarenal body (suprarenal
plexus, j), and a renal plexus (k) follows the renal artery
to the kidneys (6). The superior mesenteric plexus (g) forms
a dense network over the superior mesenteric artery (5) and
follows it to the intestine. A plexus passes caudad on the
aorta (aortic plexus, h). This is a continuation of the cœliac
and superior mesenteric plexuses; it receives a number of
branches from the main sympathetic trunk, lying dorsad of the
aorta. The aortic plexus contains a number of small ganglia.
From the aortic plexus the smaller inferior mesenteric
plexus (i) follows the inferior mesenteric artery (7) to the large
intestine; in this lies a ganglion of considerable size, the
inferior mesenteric ganglion.
In the pelvic region small plexuses are formed in a similar
manner, from branches given off by the main sympathetic
trunk, together with branches from the plexuses of the
abdominal cavity. These plexuses supply the various organs
in the pelvis.
The main sympathetic trunk (a) passes into the abdomen
at the side of the crus of the diaphragm, lying dorsad of the
great splanchnic nerve (b). In the abdomen it lies nearer the
middle line than in the thorax, so that the two trunks of right
and left side are separated by but a few millimeters as they lie
on the ventral surface of the centra of the lumbar vertebræ.
There is a ganglion for each vertebra (Fig. 162, n), and from
each ganglion are given off one or more communicating
branches to the spinal nerves, and usually a branch which
passes ventrad to join one of the plexuses among the viscera.
The first three branches of this kind (Fig. 164, c) are larger
and pass to the cœliac and superior mesenteric ganglia. The
sympathetic may be traced into the pelvis, where it becomes
smaller, and is lost as the caudal region is reached.
[409]
SENSE-ORGANS AND INTEGUMENT.
I. THE ORGAN OF SIGHT. ORGANON VISUS.
1. The Orbit.—The bony framework of the orbital fossa,
in which the organ of sight is situated, has already been
described (page 53). The orbit is not protected by bone on
all sides, being open caudad and ventrad, and partly laterad.
The structures within the orbit are further protected and separated
from other structures by a very thin, tough, transparent
membranous sac, the periorbita, which surrounds them almost
completely and nearly fills the orbit. The periorbita forms a
conical sac with the apex of the cone directed caudoventrad
and attached about the optic foramen and orbital fissure. The
base of the cone is attached about the margin of the orbital
fossa,—to the supraorbital margin of the frontal, to the
maxillary and malar bones, and to the orbital ligament, connecting
the frontal process of the malar with the zygomatic
process of the frontal. The periorbita is partly separated from
the walls of the orbital fossa by masses of fat. It comes in
contact, aside from the bones, with the temporal and pterygoid
muscles, and with nerves and blood-vessels in the orbit.
The sac thus formed encloses the eyeball with its muscles
and glands (Fig. 166), as well as a mass of fat in which these
are imbedded. The periorbita and its contents are freely
movable with relation to surrounding structures.
2. The Eyelids. Palpebræ.—The eyelids are two thin
folds which protect the eye. Externally they are covered with
hair; internally by the thin membranous conjunctiva. The
eyelids contain the Meibomian or tarsal glands, arranged in
short rows passing perpendicularly inward from the edge of the
lids; these may sometimes be seen with the naked eye as[410]
broad yellowish lines. They secrete a substance which prevents
the adhesion of the edges of the two lids. On the edge
of each eyelid, three to four millimeters from the medial angle
of the eye, is one of the openings of the lachrymal canals.
At the medial angle of the eye is the large nictitating
membrane (membrana nictitans), or “third eyelid,” corresponding
to the plica semilunaris of man.
In the cat this is large and may cover the
whole surface of the eye. It is supported
(Fig. 165) by a broad central strip of cartilage
(a), passing from its edge to its inner
angle, and the inner end of this cartilage is
surrounded, on the medial (concave) side of
the membrane, by the large lobulated Harderian
glands (b).
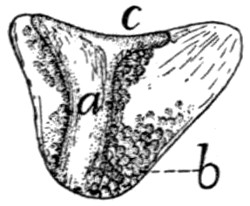
Fig. 165.—Inner
Surface of Membrana
nictitans,
showing the supporting
cartilage and Harderian
glands. a, supporting
cartilage; b,
Harderian glands; c,
outer edge.
The conjunctiva is the thin membrane
covering the inner surface of the lids, the
outer surface of the eyeball, and both surfaces of the nictitating
membrane.
The muscles of the eyelids are M. orbicularis oculi, already
described (page 98), and M. levator palpebræ superioris,
described below.
3. Lachrymal Apparatus.—The lachrymal gland is a
large reddish gland, not lobulated externally, which lies on
the surface of the eyeball just beneath the lateral angle of the
eye. The gland is situated immediately craniad of, and in
contact with, the zygomatic process of the frontal bone (Fig.
154, 11, page 374). It is flat and about one centimeter in
diameter. The ducts which pass from the gland to the eye
are not visible to the naked eye. The fluid secreted by the
gland collects at the medial angle of the eye and passes into
the two openings of the lachrymal canals, one of which is
found on the pigmented edge of each lid, three or four millimeters
from the medial angle of the eye. The canals passing
from these openings soon unite to form the nasolachrymal
duct, which passes through the lachrymal bone, along the
medial surface of the maxillary, to open into the nasal cavity
ventrad of the ventral concha of the nose.
[411]
4. Muscles of the Orbit (Fig. 166).—Within the orbit are
eleven muscles, ten connected with the eyeball, and one,
M. levator palpebræ superioris, with the upper eyelid.
Of the ten muscles of the eyeball, eight are straight
muscles, passing from an origin about the inner end of the
orbit directly distad; the other two have an oblique direction.
Four of the straight muscles are larger and are known as the
Mm. recti (b, c, d); these are distinguished according to their
position as lateral (c), medial, superior (d), and inferior (b).
The four smaller straight muscles constitute together M. retractor
oculi (e). The two oblique muscles are known as
Mm. obliqui and are distinguished according to their position
as inferior (a) and superior.
Fig. 166.—Muscles
of the Eyeball, Lateral View.
a, M. obliquus inferior; b, M. rectus inferior; c, M. rectus lateralis; d, M. rectus
superior; e, parts of M. retractor oculi; f, the oculomotor nerve.
Mm. recti (b, c, d).—The four recti muscles arise from the
bone about the optic foramen and pass toward the eyeball.
They are inserted by thin, flat tendons along a line which
separates the darker caudal part of the sclerotic from the white
zone of the sclerotic which borders the cornea,—the line of
insertion of the four tendons forming thus a circle about the
eyeball. The rectus superior (d) is partly covered by M.
levator palpebræ superioris. The tendon of the rectus inferior
(b) is crossed by M. obliquus inferior (a). The recti muscles
are all innervated by the oculomotor (third) nerve (f), except
the lateral (c), which is supplied by the abducens (sixth) nerve.
M. retractor oculi (e).—The retractor oculi arises about the
optic foramen and divides into four heads which lie nearer the[412]
eyeball than the recti and are therefore partly covered by the
latter. They alternate with the recti and are inserted into
the eyeball at about its equator, except the inferior division,
which is inserted on a line with the recti. They are supplied
by the oculomotor nerve.
M. obliquus inferior (a).—The inferior oblique arises from
the maxillary bone just laterad of the lachrymal bone and
curves over the ventral side of the eyeball along the lines of
insertion of the recti, crossing the tendon of the rectus inferior
(b) to the ventral edge of the tendon of the lateral rectus (c),
where it is inserted. It rotates the eyeball and is supplied by
the third nerve (f).
M. obliquus superior.—The superior oblique takes origin
from the cranial border of the optic foramen, passes dorsocraniad,
and ends in a small rounded tendon which passes
through a fibrous pulley-ring. This ring is situated near the
rim of the orbit, one or two centimeters craniad of the zygomatic
process of the frontal bone, and is attached to the wall of the
orbit by two fibrous bands. After passing through the pulley-ring
the tendon turns and passes laterocaudad; it becomes
much expanded, passes caudad of the superior rectus, and is
inserted into the eyeball along the caudal margin of the insertion
of the superior rectus tendon. The superior oblique is
supplied by the trochlear (fourth) nerve.
M. levator palpebræ superioris.—This is a small thin
muscle which takes origin on the wall of the optic foramen
close to the rectus superior, passes over the outer surface of the
rectus superior and beneath the lachrymal gland, and forms a
thin tendon which is inserted into the margin of the upper
eyelid. It is innervated by the oculomotor.
5. The Eyeball.—The eyeball is approximately spherical
in form, but the cranial surface has a slightly greater curvature
than the caudal portion, so that the eye appears slightly pointed
at the cranial (free) surface. The large cylindrical optic nerve
(Fig. 167, a) enters the eyeball at nearly the centre of the
caudal half of the eye. On the outer surface is seen the transparent
cornea (c), covering the free surface of the eye, and the
opaque sclerotic (b), covering the remainder of the ball.[413]
Through the cornea can be seen the yellow curtain-like iris,
with an opening in its centre, the pupil.
Fig. 167.—Diagram of a
Section of the Eye.
a, optic nerve; b, sclerotic
(b′, thickened “white zone”);
c, cornea; d, choroid; e, iris;
f, ciliary body; g, retina; h,
crystalline lens; i, zonula ciliaris;
j, anterior chamber of
eye; k, vitreous humor.
Coats of the Eye (Fig. 167).—The outer coat of the eyeball
is formed by the sclerotic (or sclera) (b) and the cornea (c).
The sclerotic is the whitish opaque portion covering the caudal
three-fourths of the eye. It forms a tough sac serving as a
protection for the structures within. The cornea (c) is the circular
transparent portion of the outer coat, covering the free
surface of the eye; it is a direct continuation of the sclerotic.
Just before passing into the cornea the sclerotic is much thickened,
forming a broad white zone (b′) about the eyeball near
its equator. To this zone, at its caudal margin, are attached
the rectus muscles of the eyeball. The sclerotic has many
pigment-cells on its inner surface, giving it a brownish appearance
and forming the lamina fusca.
The cornea (c) is transparent and more strongly curved
than the sclerotic. It is thicker than
most of the sclerotic coat, but not so
thick as the white zone of the latter.
Within the sclerotic is the second
coat of the eye, formed by the choroid
(d) and the iris (e). The choroid (d)
is a vascular layer, and contains much
pigment, giving it a dark color. It
lines the inner surface of the sclerotic,
but does not cover the inner surface of
the cornea. At the white zone of the
sclerotic it is thrown into a large number
(about seventy) of meridional folds,
the plicæ ciliares or ciliary folds, which
together constitute the so-called ciliary
body (corpus ciliare) (f). This forms
a circular zone on the inner surface of the white zone of the
sclerotic, and is continuous with the iris (e). The choroid coat
is furnished in certain parts of its area with a layer of cells
containing crystals, which give it a brilliant color, forming the
tapetum. The color of the tapetum is a metallic yellowish[414]
blue or green; it is this layer which causes the cat’s eye to
“shine” in the dark.
The iris (e) is a continuation of the choroid. It projects
inward from the white zone of the sclerotic, forming a circular
curtain lying some distance behind the cornea, and perforated
by an opening, the pupil. The iris is usually yellow in color
on its outer surface, darkly pigmented within. The pupil
varies normally in size and form according to the amount of
light to which the eye is subjected. In a cat killed with
chloroform the pupil is very large and circular; in the living
animal it is elliptical with the long axis dorsoventral, or when
much contracted it is a mere dorsoventral slit.
The inner coat of the eye is formed by the retina (g).
This is the part of the eye which is sensitive to light; it is
formed by an expansion of the optic nerve (a). In a preserved
eye it is usually soft and opaque, and may be seen to line the
caudal half of the inner surface, extending apparently to the
ciliary body. Here it seems to end as a free margin, the ora
serrata; it really becomes thin and passes onto the surface of
the ciliary body, forming the ciliary portion of the retina,
and onto the inner surface of the iris, where it forms the uvea.
Near the centre of the retina the entrance of the optic nerve is
marked by a small round spot, the blind spot.
The Crystalline Lens (h).—The lens is a biconvex transparent
body situated within the eye a little in front of the ciliary
body and just behind the iris. It is more strongly convex in
front than behind. The lens is surrounded by a thin transparent
sac, the capsule of the lens (capsula lentis). The
capsule of the lens is attached all around the equator of the
lens by fibrous bands to the ciliary body, this attachment
forming the zonula ciliaris (or zonula Zinni) (i), by which
the lens is suspended.
Chambers of the Eye.—The lens (h) and zonula ciliaris (i)
form a partition dividing the eye into two parts. The cavity
in front of the lens contains a fluid, the aqueous humor.
This cavity is partly subdivided by the iris into the anterior
and posterior chambers of the eye.
The part of the eye lying behind the lens is much larger[415]
than the part in front of the lens. It is lined chiefly by the
retina, its front boundary being the lens. It contains a jelly-like
substance, the vitreous humor (or vitreous body, corpus
vitreum).
II. THE EAR. ORGANON AUDITUS.
The ear of the cat, like that of man, is composed of three
parts, the external, middle, and internal ear.
The external ear is limited internally by the tympanic
membrane.
The middle ear extends from the tympanic membrane to
the fenestra cochleæ and fenestra vestibuli. It contains the
bones of the ear, and is connected with the pharynx by the
Eustachian tube.
The internal ear is wholly contained within the petrous
bone, and on the surface of the membrane lining its cavities
are distributed the branches of the auditory (eighth) nerve.
The External Ear.
The external ear consists of an expanded portion, the
auricle (auricula) or pinna, and of a canal leading from the
auricle to the tympanic membrane, known as the external
auditory meatus. The concave surface of the auricle is
directed craniolaterad, and its edges form the dorsal and lateral
boundaries of the auditory opening. From the bottom of the
cavity which it partly encloses, the external auditory meatus
passes medioventrad.
The auricle (Fig. 168) is composed of a thin curved plate
of cartilage (cartilago auriculæ), to which numerous muscles
are attached and which is covered on both sides by integument
intimately attached to the cartilage. Distad the edges of the
cartilage form a free border (b); proximad the plate is rolled
together in such a way that its edges (g and h) overlap on the
craniomedial side, thus enclosing a tube or funnel (e) (concha)
which forms the enlarged outer portion of the auditory passage.
Within the cavity of the auricle, on its medial surface,
somewhat below the level of the lowest portion of the margin
of the auditory opening is seen a very prominent pedunculated[416]
process (d). This is really part of a transverse ridge,
extending craniocaudad on the medial wall of the auricular
cavity. This ridge is the antihelix (d); it divides the auricula
into a distal and a proximal portion. The portion distad of
the antihelix is the scapha (a); its surface is smooth except for
two slight longitudinal ridges extending distad from near the
two ends of the antihelix. That portion of the auricle that lies
proximad of the antihelix forms a deep irregular cavity known
as the concha (e). A deep furrow, corresponding to the antihelix,
separates scapha from concha externally. The concha
extends caudad as a deep, rounded pocket; externally this
pocket forms a prominent convexity, the eminentia conchæ
(j), on the caudal surface of the proximal portion of the
external ear. Farther proximad the concha narrows like a
funnel and becomes much compressed laterally, so that a section
of its cavity forms a narrow craniocaudal slit.
The inner surface of the concha is marked by a number of
very prominent ridges and projections. These are usually due
to folds in the cartilage, so that external furrows and depressions
correspond to the internal projections. The edges of these
depressions serve for attachment of muscles (Fig. 169), by contraction
of which the form of the concha is changed,—the folds
of the cartilage thus serving as regions of greater flexibility.
Two or three centimeters proximad of its external opening
the cartilage of the concha ends in a free edge which receives
within itself the distal end of the cartilaginous external auditory
meatus (e′). The two are united together by fibrous tissue.
The cartilaginous auditory meatus (e′) extends as a nearly
cylindrical tube mediad and slightly craniad to the tympanic
membrane in the opening of the bulla tympani. This tube is
lined with integument which continues over the tympanic
membrane. The subcutaneous tissue of the tube contains the
ceruminous glands, which secrete the ear-wax and open on the
surface of the skin.
To understand the relations of the muscles of the external
ear it is necessary to consider a little more fully the structure
of the auricular cartilage (Fig. 168), especially as compared
with the human ear. The ear of the cat differs from the human[417]
ear in the fact that the edges of the auricular cartilages are
rolled together proximad so as to overlap. There is thus no
gap between the tragus (g) and antitragus (h), as in man, but
these two structures partly overlap.
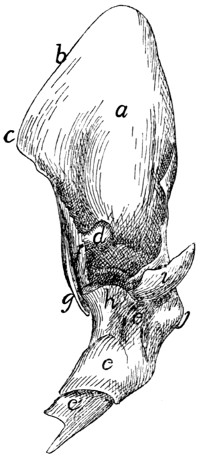
Fig. 168.—Cartilage of the
External Ear, Craniomedial
View.
a, scapha or pinna; b, helix;
c, spine of the helix; d, antihelix;
e, concha; e′, cartilaginous auditory
meatus; f, crista helicis; g,
tragus; h, antitragus; i, processus
uncinatus; j, eminentia conchæ.
The free edge (b) of the auricular cartilage corresponds
partly to the helix of man. At its
mediocranial angle the proximal part
of the auricle is composed of muscle
and integument only, so that the
cartilage when isolated presents here
a deep notch distad of which is
a prominent projecting angle, the
spine (c) of the helix, which serves
for attachment of the adductor auris
superior muscle (Fig. 169, 1). Proximad
of the spine, separated from it
by the deep notch, the cranial edge
(g) of the auricular cartilage approaches
the caudal edge (h), finally
overlapping it, so that the cavity of
the auricle is now completely surrounded.
This overlapping portion
of the cartilage is on the medial side
of the cavity, and in the natural state
is covered by muscles and integument,
so as not to be apparent.
That portion of the cranial edge
of the cartilage that overlaps the caudal
edge forms a cartilaginous plate
which projects proximad as a blunt
point; this plate is the tragus (g).
Along the inner edge of the tragus is
a ridge which forms a continuation of the helix; this is the
crista helicis (f). The portion of the caudal edge of the cartilage
that is partly overlapped by the tragus is an irregular flat
plate known as the antitragus (h). At its caudolateral margin
the antitragus extends distad as a thin, pointed, cartilaginous
spine, the processus uncinatus (i). This supports a sheet of[418]
integument which bears two longitudinal ridges projecting into
the cavity of the ear, with a broad groove between them.
The Scutiform Cartilage.—Closely connected with the
external ear, but not forming a part of it, is the scutiform cartilage.
This lies in the temporal fossa, between the integument
and the temporal muscle, just behind the orbit and
beneath the craniomedial portion of the auricle (Fig. 63, 1,
page 97). It is a narrow cartilage about two centimeters
long, with its long axis craniocaudal. For the greater part of
its length it forms a slender rod, from which a thin cartilaginous
sheet extends a short distance laterad; at its caudal end
the rod forks, the two arms extending nearly at right angles
to the main portion. The scutiform cartilage serves for the
origin or insertion of a number of the muscles of the ear.
Muscles of the External Ear (Fig. 169).—(a) The muscles
connecting the cartilages of the external ear with other parts of
the head have been described (page 96), excepting the tragicus
lateralis (4). These muscles are the following: M. intermedius
scutulorum (Fig. 63, a), M. frontoauricularis, M. levator auris
longus (including the cervicoauricular) (Fig. 63, g, g′),
M. auricularis superior (Fig. 63, k), M. abductor auris longus
(Fig. 63, m), M. abductor auris brevis (Fig. 63, l), M. zygomaticus
(Fig. 64, d), M. submentalis (Fig. 64, c), M. depressor
conchæ (Fig. 64, b), M. frontoscutularis, M. adductor auris
inferior (Fig. 63, f), and M. tragicus lateralis
(Fig. 169, 4).
M. tragicus lateralis (Fig. 169, 4).—A band about seven
millimeters wide, lying beneath the mass of fat about the ear
and running ventrad lengthwise of the concha and close
against it.
Origin (Fig. 96, a, page 229) on the caudal end of the
mandibula, in the cavity found between the condyloid process
and the angular process. The muscle passes dorsad and is
inserted (Fig. 169, 4) on the caudal margin of the tragus and
in the depression on the concha just caudad of the tragus.
Relations.—This muscle lies between the temporal muscle
and the concha.
Action.—Pulls the ear ventrad and probably rotates it
outward.
[419]
(b) The following three muscles connect the scutiform
cartilage with other parts of the external ear.
M. rotator auris or scutuloauricularis inferior.—A flat
craniocaudal band of fibres lying between the auricle and the
temporal muscle.
Origin on the scutiform cartilage,
just caudad of the insertion of the
frontoscutularis, of which this muscle
seems to be a continuation. The
muscle passes caudad as a band seven
or eight millimeters wide, curving
about the medial surface of the auricle,
and is inserted on the caudomedial
surface of the eminentia
conchæ.
Relations.—Lateral surface with
the auricular cartilage; medial surface
with the temporal muscle.
Action.—Rotates the external ear
mediad and caudad.
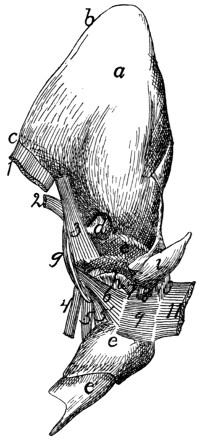
Fig. 169.—Cartilage of the
External Ear, with Muscles.
a-i, as in Fig. 168. 1, M.
adductor auris superior (cut); 2,
M. adductor auris medius (cut);
3, M. helicis; 4, M. tragicus lateralis
(cut); 5, 5′, M. tragicus medialis;
6, M. antitragicus; 7, M.
adductor auris inferior (cut); 8,
M. depressor conchæ (cut); 9, M.
conchæus externus; 10, part of
M. auricularis externus; 11, M.
abductor auris longus (cut).
M. adductor auris superior (Fig.
169, 1; Fig. 63, f). (Part of the
auricularis anterior of man.)
Origin on the dorsal surface of
the scutiform cartilage for its entire
length. The fibres form a continuation
of those of the frontoscutularis;
they pass dorsocaudad and are inserted
into the spina helicis or craniomedial
margin of the auricular cartilage.
This muscle lies in the fold
of skin that connects the craniomedial
angle of the auricular cartilage with
the head, so that it is covered on both
sides by integument.
Action.—Draws the auricle craniad.
M. adductor auris medius (Fig. 169, 2).
(Part of the[420]
auricularis anterior of man.)—A very thin band of fibres eight
to ten millimeters broad.
Origin on the middle two-thirds of the caudoventral edge
of the scutiform cartilage. The fibres pass nearly ventrad
and are inserted along the medial or caudal surface of the
tragus.
Relations.—Outer surface with the auricular cartilage.
Inner surface with the temporal muscle or with the pad of fat
separating this from the auricle.
Action.—Pulls the concha dorsocraniad.
(c) The remainder of the muscles of the ear interconnect
parts of the auricular cartilage.
M. transversus auriculæ (Fig. 63, i).—A band about six
to eight millimeters wide, on the medial convex surface of the
auricle, between the distal ends of M. levator auris longus (g′)
and M. auricularis superior (k).
Origin on the medial surface of the concha, just proximad
of the furrow which corresponds to the antihelix. The muscle
bridges this furrow, passes distad about two centimeters, and is
inserted on the auricular cartilage on a line which forms a
caudal continuation of the line of insertion of the levator auris
longus (g′).
Relations.—Outer surface with the levator auris longus
(g, g′), the auricularis superior (k), and the integument; inner
surface with the auricular cartilage.
Action.—Flexes the scapha mediad on the concha, thus
enlarging the external opening of the concha.
M. auricularis externus (Fig. 169, 10;
Fig. 64, r).—This
consists of small scattered bands of interrupted fibres on the
convex surface of the auricle, caudad of the transversus
auriculæ. The largest of these bands runs parallel with the
transversus auriculæ about five or six millimeters caudad of the
latter. It has origin on the eminentia conchæ distad of the
insertion of the abductor auris longus. Thence the fibres run
distad five or six millimeters and are interrupted by an attachment
to the cartilage. Distad the fibres begin again, and the
muscle runs thence distad for about one centimeter, the distal
end being attached to the cartilage.
[421]
Caudad of this, especially of its proximal portion, are one
or two small bands of fibres usually running parallel with it,
but unconnected with it and with each other. The extent and
distribution of these bands varies, and the direction of fibres is
also variable.
Relations.—Outer surface with the integument; inner surface
with the auricular cartilage.
Action.—Flexes the auricular cartilage.
M. helicis (Fig. 169, 3;
Fig. 64, m).—A muscle about
two centimeters in length and five or six millimeters in width,
on the inner surface of the ear, along its cranial margin.
Origin on the medial surface of the concha, just caudad of
the proximal end of the tragus. The fibres pass distad to be
inserted on the auricular cartilage at the caudal angle of the
notch which lies beneath the cranial edge of the auricle, along
with the caudal fibres of the adductor auris superior (1).
Relations.—Lateral surface with the integument; medial
surface with the tragus.
Action.—Draws proximad the cranial margin of the auricle.
M. antitragicus (Fig. 169, 6;
Fig. 64, n).—A small muscle
about one centimeter in length and three millimeters in thickness,
attached at one end to the caudal border of the antitragus.
The fibres pass mediodistad and are inserted on the
tragus, in common with the fibres of the tragicus medialis
(Fig. 169, 5, 5′).
Relations.—Outer surface with the adductor auris inferior
and the integument; inner surface with the cartilage.
Action.—Constricts the external auditory opening.
M. tragicus medialis (Fig. 169, 5, 5′).—A small, flat muscle,
in two parts, on the medial surface of the concha.
Origin on the ventral end of the tragus. The cranial
fibres form a thick nearly cylindrical bundle which extends
proximad about one centimeter, and is inserted on the cranial
surface of the concha just proximad of a deep fold in the cartilage.
The caudal fibres form a thin sheet which spreads on
the medial surface of the concha and is inserted on an obliquely
proximodistal line on the medial surface of the concha, less
than one centimeter from the free distal edge.
[422]
These two bundles are distinct except at their origin, and
are sometimes considered as separate muscles.
Relations.—Outer surface with the tragicus lateralis (4) and
the fat about the concha. Inner surface with the concha.
Action.—Flexes the concha.
Strauss-Durckheim describes under the name cornetoconchæus
a small muscle bridging the deep groove caused by the
folding of the medial surface of the concha, its proximal end
being close to the insertion of the abductor auris brevis (page
100). This muscle was absent in all of the cases examined for
its presence.
M. conchæus externus (Fig. 169, 9).—A quadrilateral
muscle on the lateral surface of the concha. The cranial end
of this muscle is attached to the concha a short distance distad
of the antitragus along a dorsoventral line six to eight millimeters
in length; the fibres pass thence caudodorsad a distance
of about one centimeter, where they are again attached to the
concha. The muscle thus bridges a shallow depression on the
lateral surface of the concha.
Action.—Constricts the concha.
The Middle Ear.
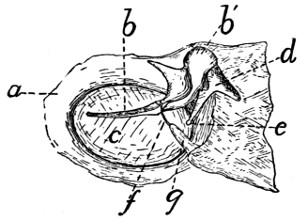
Fig. 170.—Tympanic Membrane,
Viewed from the Inner Side.
The outer wall of the bulla has
been removed, carrying with it the
tympanic membrane with the malleus
and incus; these are now
viewed from the side that was
turned toward the cavity. a, bony
ring forming inner end of external
auditory meatus; b, b′, malleus; c,
tympanic membrane; d, incus; e,
lenticular process; f, chorda tympani
nerve; g, cartilaginous support
for chorda tympani nerve.
The middle ear is enclosed in a
cavity within the tympanic bulla.
Its outer boundary, as well as the
inner boundary of the external ear,
is formed by the tympanic membrane
(Fig. 170, c.) This is a thin,
semitransparent membrane attached
to the bony ring (a) surrounding the
inner end of the external auditory
meatus. It is oval in outline. Across
the dorsal third of its inner surface
runs the handle of the malleus (b),
to which the membrane is firmly
attached and which by pulling on
the membrane gives it the form of a cone instead of a flat surface.
The apex of the cone is directed mediad.
The middle ear, or cavum tympani, is an ellipsoidal cavity[423]
situated in the tympanic bulla. On removing the medial wall
of the bulla, a bony plate is seen separating its cavity into two
chambers. This plate of bone extends in from the lateral wall
of the bulla, forming a complete partition except at the dorsocaudal
part, where a notch in the bone forms a free communication
between the two chambers. The lateral chamber, or
tympanum proper, contains the bones of the ear and is limited
externally by the tympanic membrane. At its cranial end is
seen the opening of the tuba auditiva, or Eustachian tube.

Fig. 171.—Malleus
and Incus,
United in the
Natural Position.
1, malleus; 2, incus.
a, handle of
malleus; b, neck; c,
head; d, process for
attachment of tendon
of M. tensor
tympani; e, wing-like
plate and process
of neck; f, g,
the two processes of
the incus.
The tuba auditiva, or Eustachian tube, is a cartilaginous tube
about one and a half to two centimeters long, passing from the
nasopharynx to the middle ear and placing these two cavities
in communication. The pharyngeal opening is at about the
middle of the length of the nasopharynx, near the dorsal border
of the lateral wall; it is a narrow slit two or three millimeters
long. Thence the tube passes caudolaterad on the ventral
surface of the sphenoid bone, and enters the tympanic cavity
through the opening just laterad of the styliform process of the
tympanic bulla. The tube has a thick, cartilaginous medial
wall, while the lateral wall is thin and formed
of connective tissue. The lumen of the tube
is a curved slit in cross-section.
Bones of the Ear (ossicula auditus).—There
are three small bones in the middle ear, the
malleus (hammer) (Fig. 171, 1), incus (anvil)
(Fig. 171, 2), and stapes (stirrup) (Fig. 172).
The malleus (Fig. 171, 1) consists of a
slender handle (a) with a small neck (b) attached
at an angle to the caudal end of the
handle. At the end of the neck is a rounded
head (c) which articulates with the incus (2),
and together with one part of the incus lies in
a small fossa dorsad of the fenestra vestibuli.
From the medial aspect of the neck extends a
short process of bone (d) to which is attached
the tendon of the tensor tympani muscle. One
side of the neck is produced into a thin wing-like plate, bearing
on its edge a pointed process (e).
[424]
The incus (Fig. 171, 2) bears much resemblance to a two-fanged
molar tooth. It lies in the fossa with the head of the
malleus. Its head or crown is directed craniad and has a
concave surface which articulates with the head of the malleus.
One of the fangs (f) extends caudad in the fossa above mentioned
and is held in position by a ligament. The other fang
(g) extends ventromediad and articulates with the head of the
stapes. This fang terminates in a minute rounded tubercle
known as the lenticular process; this in early stages of development
is a separate bone, the os lenticulare.
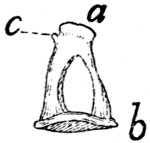
Fig. 172.
Stapes.
a, head; b,
base; c, process
for attachment
of the stapedius
muscle.
The stapes (Fig. 172) has the form of a stirrup, and is the
last in the chain of bones. The small end is termed the head
(a) and the broader part the base (b). The head
of the stapes articulates with one of the processes
of the incus. The base of the stapes fits into the
fenestra vestibuli of the petrous bone and completely
closes it. The stapes is held in position
by the stapedius muscle, which is attached to a
minute process (c) near the head.
Thus the three bones of the middle ear form a
chain which connects the medial surface of the
tympanic membrane with the fenestra vestibuli. As the
fenestra vestibuli forms a portion of the wall of the vestibule,
the vibrations of the tympanic membrane are transmitted
through the chain of bones to the fluid within the vestibule.
Muscles of the Middle Ear.
M. tensor tympani.—The tensor tympani muscle is a short
conical muscle having origin in a small fossa (Fig. 25, d, p.
34) in the petrous bone dorsocraniad of the fenestra vestibuli.
Its very short tendon passes laterad and is inserted into a projection
(Fig. 171, d) on the neck of the malleus.
M. stapedius.—The stapedius muscle is a short muscle
taking origin from a fossa (Fig. 25, f) in the lateral surface of
the petrous bone caudad of that for the incus. It is inserted
into the head of the stapes.
The Internal Ear (Fig. 173).
The internal ear or labyrinth is completely enclosed in the
petrous bone. In the dry bone the internal ear communicates[425]
with the middle ear by the fenestra cochleæ and the fenestra
vestibuli, which have been described in the account of the
petrous bone (page 33). In the recent state the fenestra
vestibuli is closed by the base of the stapes, while the fenestra
cochleæ is covered by a delicate membrane,—so that the internal
ear forms a closed cavity. It is possible to distinguish
a bony labyrinth and a membranous labyrinth (Fig. 173), the
former being the cavity in the petrous bone, while the latter is
the real auditory organ, situated within the cavity. The membranous
labyrinth repeats in general the form of the bony labyrinth,
so that it will not be necessary to distinguish the two in
our account of the main features of the structure. The auditory
nerve is distributed to the membranous labyrinth. For an
account of the minute structure of this organ reference should
be made to a text-book of general histology or physiology.
Fig. 173.—Membranous
Labyrinth.
a, vestibule; b, cochlea;
c, external semicircular
canal; d, superior semicircular
canal; e, posterior
semicircular canal; f, scala
tympani; g, scala vestibuli;
h, fenestra cochleæ; i,
fenestra vestibuli.
The labyrinth (Fig. 173) consists of a cavity, the vestibule
(a), and connected with this a coiled
tube, the cochlea (b), and the three semicircular
canals (c, d, and e).
The cochlea (b) is situated in the promontory,—the
rounded elevation of the
petrous bone extending craniad from the
fenestra cochleæ. It consists of a bony
tube coiled spirally about a central column
known as the modiolus. The coil has
nearly the form of a snail’s shell, and is
somewhat more lengthened in the cat than
in man. A plate of bone, the lamina
spiralis, incompletely divides the cavity of
the canal lengthwise into two parts, known as scalæ (f and g).
One of these opens at the fenestra cochleæ (h) into the tympanic
cavity (or would so open but for the thin membrane covering
the fenestra); this is known as the scala tympani (f). The
other scala is connected with the vestibule (a) and is known
as the scala vestibuli (g).
The vestibule (a) is a pyramidal cavity situated mediad of
the caudal end of the cochlea. It is connected with the scala
vestibuli of the cochlea, and with the three semicircular canals.[426]
The fenestra vestibuli (i) is an opening in its bony wall, filled
in the recent state by the stapes. On the medial surface of the
vestibule are a number of minute orifices through which the
auditory nerve enters.
The semicircular canals (c, d, and e) are three curved
tubes, each forming about two-thirds of a circle, and opening
at both ends into the vestibule. They are imbedded in the
petrous bone. The planes of the three tubes intersect nearly
at right angles; from their position in man they have been
named external (c), superior (d), and posterior (e). The
external one (c) is nearly horizontal in position, and surrounds
the curved fossa (Fig. 25, f) for the stapedius muscle, caudad
of the fenestra vestibuli. The superior (Fig. 173, d) and posterior
(e) are in nearly dorsoventral planes, but at right angles
to each other. The superior (d) lies in the ridge forming the
caudal boundary of the appendicular fossa. The posterior (e)
is situated in a continuation caudad of the ridge which forms
the dorsal boundary of the internal auditory meatus. One end
of each canal is slightly enlarged to form an ampulla. The
superior (d) and posterior (e) canals join at one end and enter
the vestibule by a common opening. One end of the external
canal (c) opens into the vestibule so close to one end of the
posterior canal (e) that they appear to have a common opening,
and this is commonly stated to be the case; according to
Jayne, however, the openings are really separate. The two
remaining ends (of the superior (d) and external (c) canals) also
enter the vestibule separately, so that the three canals enter
the vestibule by five openings.
III. THE OLFACTORY ORGAN. ORGANON OLFACTUS.
The olfactory organ consists of the olfactory mucous membrane,
in the dorsocaudal part of the nasal cavity. The nasal
cavity has been described in connection with the respiratory
system.
[427]
IV. THE ORGAN OF TASTE. ORGANON GUSTUS.
The sense of taste is located in the mouth-cavity, especially
in the mucosa of the tongue and the soft palate. The proper
organs of taste are the so-called taste-buds, found on the
papillæ of the tongue, the soft palate, etc. Their exact distribution
in the cat has apparently not been worked out.
V. THE INTEGUMENT. INTEGUMENTUM COMMUNE.
The skin of the cat consists of the usual two layers, the
outer epidermis and the inner fibrous corium. The skin is
particularly thick and tough about the sides of the neck and
face.
The skin is covered almost completely with fine soft hairs.
Only the end of the nose, the pads on the soles of the feet,
and the nipples are without hair. A number of long stiff sensory
hairs (vibrissæ) are found on the upper lip, the cheek, and
above the upper eyelid.
The pads on the soles of the feet are cushion-like projections
of skin, composed largely of connective and elastic tissue
and fat. There is one large pad for each foot, and in addition
a small pad for each digit. The fore limb has besides a small
conical pad which is situated almost exactly over the pisiform
bone.
The nails are horny outgrowths of the epidermis, covering
the distal phalanges.
The muscles of the skin are the cutaneus maximus and
platysma; these have been described (page 93). The superficial
facial muscles are differentiations of the skin-muscles.
[428]
[429]
APPENDIX.
PRACTICAL DIRECTIONS.
The following appendix on the dissection and study of the cat
is not intended as a general treatise on methods of preservation and
dissection, but attempts only to give the application of well-known
methods to the dissection and study of the particular animal with
which the book deals. General methods which are referred to but
not described must be looked up, if unfamiliar, in the manuals of
general methods.
Aids to Dissection and Study of the Cat.
—A copy of Jayne’s
“Mammalian Anatomy” should be in the laboratory for reference.
Only the volume on the Bones of the Cat has been issued thus far.
Wilder and Gage’s “Anatomical Technology” will be found
very useful for methods of work.
The plates of Strauss-Durckheim’s “Anatomie du Chat” are
exceedingly valuable for the bones and muscles, and should be in
the laboratory if possible. Outline reproductions of these plates
have been published by H. S. Williams (G. P. Putnam’s Sons,
New York, 1875). The use of both the original plates and the reproductions
is made difficult by the antiquated nomenclature used
for the structures figured.
Method of Use of the Present Directions.
—Except in the case of
the Bones, the order of study of the structures is usually not the
same as the order in which they are described. This is of course
due to practical difficulties in dissection. The present directions
give the order of study, or at least the order of dissection; of course
after dissection the structures can be reviewed in the logical order
given in the descriptions. But in the first study and dissection, the
student is to use these directions for the order of work, while using
the body of the book for the descriptions. It will be necessary
therefore to keep the book opened in two places,—one in the
appendix, for the directions; one in the body of the book, for
descriptions. In studying the muscles, for example, the student will
follow the order of dissection given in the appendix, but as every
muscle is dissected he will find, confirm, and study the description of
that muscle given in the body of the book. Other organs are
studied in a similar manner.
[430]
Before commencing the study, read that portion of the preface
which explains the use of terms of direction (page VII).
Note that the page on which any figure is found may be determined
by consulting the list of figures given in the Table of Contents.
THE BONES.
The bones will of course be studied from dried specimens.
These may be prepared once for all and kept in the laboratory from
year to year. A number of mounted skeletons of the cat should be
at hand; these must be prepared by some one who has skill in such
work.
Preparation of Separate Bones.
—For preparing the separate bones
the liquid-soap process recommended by Wilder and Gage (“Anatomical
Technology,” p. 107) will be found most convenient. This
is as follows: The skin and soft parts are removed as far as possible
without injury to the bones.
Make the following “liquid-soap” mixture:
| Soft water |
2000 cc. |
| Strong ammonia |
150 cc. |
| Nitrate of potash (saltpetre) |
12 grams. |
| Hard soap |
75 grams. |
Heat these together till a homogeneous mixture is formed. Now
immerse the bone or bones to be cleaned in a liquid composed of
four parts water to one part of the above liquid-soap mixture. Boil
forty minutes; pour off the liquid and renew it. Boil about half an
hour longer; then remove soft parts with fingers, scalpel, and brush.
The bones may be boiled as much longer in the mixture as is necessary
to make the soft parts come away easily. If they are boiled too
long, however, the epiphyses will separate from the bones. This
is especially likely to occur in young skeletons, which must therefore
not be boiled so long as the bones of an old cat.
Skeletons prepared in the following rough way are useful for
many purposes. The entire body of the cat, or a single limb, is
placed in the liquid-soap mixture (after partial removal of soft parts),
and boiled till the muscles may be removed, but the ligaments,
holding the bones together, remain. Clean thoroughly without
removing the ligaments, and allow to dry. The bones are thus held
together in their natural positions. Skeletons so prepared are not
elegant, but are frequently useful.
Disarticulation of Skulls.
—The bones of the skull may be separated
as follows: Clean the skull in the liquid-soap mixture, as
above described. Then fill the cranial cavity with dry rice, beans,
corn, or some other seeds that swell much on imbibition of water.
Cork up the foramen magnum, and place the skull for twelve to
twenty-four hours in water. The swelling of the grain will partially[431]
separate the bones at the sutures. The bones may then usually be
separated completely by hand.
Entire skulls, and longitudinal sections, as represented in Fig.
43, p. 60, should be at hand.
Study of the Bones.
—The bones should be studied in the order in
which they are described. No further special directions for their
study are necessary.
Study of Ligaments.
—It will usually not be practicable for each
student to make the dissections and preparations necessary for studying
the ligaments. Rather should these be examined on demonstration
preparations preserved in the laboratory. Such preparations
may most easily be made by using fresh material, and dissecting
away muscles and other tissues, leaving only the ligaments connecting
the bones. The preparations are then preserved in one or two
per cent. formalin. It is difficult to prepare the ligaments satisfactorily
on preserved material, because they do not stand out clearly
from other tissues in such material, so that they are not easily distinguishable.
MUSCLES.
Preparation and Preservation of Material.
—Formalin forms by far
the most satisfactory preservative for anatomical material. It is much
better in almost every respect than alcohol, and has the additional
advantage of being much cheaper.
After injecting with formalin, the cat may be preserved either
immersed in a weaker solution of the same substance, or may be
kept for a long time simply wrapped in a cloth dampened with
formalin. The details of preparation differ according to which
method is to be used.
In either case, the cat should be killed with chloroform, by
placing it in a tight jar or box with a cloth or bit of cotton saturated
with chloroform. It is not necessary to bleed the specimen. When
the cat is dead, remove it to a tray, place it on its back, and tie the
limbs loosely so that they will remain outspread.
Expose the femoral artery in the thigh, as follows: Make a
median longitudinal incision through the skin of the abdomen, from
a point an inch or two caudad of the xiphoid process to the pubis.
Make an incision passing from near the middle of this obliquely
laterad onto the thigh, for about three inches. Reflect the two flaps
in the angles between these cuts, then with forceps and tracer isolate
the femoral artery just as it passes into the thigh (Fig. 127, a).
Make with scissors an oblique incision in one side of the artery, one-half
to one inch from the point where it leaves the abdominal cavity.
Introduce a canula directed toward the body, and tie it in place.
1. If the cat is to be preserved immersed in a solution of
formalin, it should now be injected with a five per cent. solution of
commercial formalin in ordinary water (commercial formalin 5 parts;[432]
water 95 parts). Into an adult cat 300 to 400 cubic centimeters of
the five per cent. formalin should be injected.
The cat may now be preserved in a one per cent. solution of
formalin. Before immersing it in the formalin it is well to remove
the hair in some way, otherwise this will hold much water and be
very inconvenient in dissection. The neatest method is to cut off
the hair with scissors; this takes some time, however. If the skin-muscles
(p. 93) are not to be dissected, the skin may be removed.
It is well in this case to leave the skin on the sides of the head and
on the feet.
It is not necessary to open the abdominal cavity, after injecting
with the five per cent. formalin. If there is at any time reason to
suspect that the viscera are not keeping well, the abdominal and
thoracic cavities should be filled with five per cent. formalin by
making a small hole through the body wall into the cavity, introducing
a canula, and injecting till the cavity is full.
The specimen should be kept in a jar or other vessel and should
be completely immersed beneath the one per cent. formalin. Any
parts projecting above the fluid may be attacked by mould. If a
part is found to be moulded, it should be submerged, when the
mould will be killed. The mould injures the specimen if allowed
to grow for a long time, but if discovered soon, no very serious harm
results.
During dissection, exposed parts of the body should be kept
from drying by covering with a damp cloth. Specimens preserved
in formalin dry out much less quickly than those preserved in
alcohol.
2. For some purposes it is much more convenient to preserve
the material without immersing it in a vessel of formalin. Specimens
injected with five per cent. formalin, as above directed, would usually
keep perfectly without further treatment, but for two difficulties:
(1) the specimen may dry; (2) it may be attacked by mould.
The drying may be prevented as follows: Use for injecting a
mixture of the five per cent. formalin with one-sixth its volume of
glycerine. This will dry only very slowly, and if pains are taken to
keep exposed parts covered with cloths dampened with the mixture
of formalin and glycerine, there will be no trouble from this source.
The skin should not be removed from such specimens except as
necessary in the course of dissection. The hair will give little
trouble, because dry.
The specimens should be kept in a tight box, that the fluid may
not evaporate rapidly from the cloths used for wrapping.
The attacks of the mould present a difficulty not so easily overcome.
As long as the specimens are in daily use for dissection, and
exposed parts kept covered with cloths saturated with the glycerine
and formalin mixture, little or no trouble is to be anticipated. But
if the specimens are left untouched for some days, and particularly if
exposed surfaces are not kept covered in the manner above recommended,[433]
mould is almost sure to attack them. Material neglected
for a week may thus be ruined.
If mould is found at any time to be attacking the material, the
attacked part should be cleaned and well wrapped in cloths saturated
in the formalin-glycerine mixture. If such means are unavailing, the
specimen should be immersed in a vessel of one per cent. formalin
and preserved for the future in this. Recourse should be had at
once also to this method of preservation if the specimen shows signs
of decomposition by having a strong odor.
It seems probable that the attacks of the mould might be
prevented by mixing some fungicide with the injecting fluid; experiments
should be made in this direction.
There are many advantages in preserving the material if possible
without immersing it. All parts retain their color and pliability
much more completely, so that the different structures are much
more easily distinguishable. The method above described, by the
use of formalin and glycerine, is to be recommended when the
material is to be worked on every day or so and is not to be kept
longer than a few weeks. It is usually not very satisfactory in warm
weather, however. If some method can be devised of entirely preventing
the attacks of the mould, this will be an excellent method of
preserving anatomical material.
The same specimens used for the study of the muscles will serve
also for the spinal cord and brain if prepared by one of the methods
above described. (For the viscera, blood-vessels, and nerves, other
specimens will have to be prepared; for these, directions are given
later.)
Muscles to be Dissected.
—It will probably usually be impossible
in a given course of study to dissect all the muscles of the cat. The
instructor will of course select such as seem most advisable to him.
The following may perhaps be recommended as a good selection
for a fairly extensive course: Muscles of the Fore Limbs (pp. 436-439);
Muscles of the Neck and the Deep Muscles of the Head (pp.
439-441); Muscles of the Wall of the Thorax and of the Abdomen
(pp. 441-442); Muscles on the Ventral Side of the Neck (p. 442).
If the superficial facial muscles (excluding those of the ear) can be
studied from a preparation, this will be a valuable addition to a
course (pp. 434-435).
DISSECTION OF MUSCLES.
General.
—In dissecting muscles a prime requisite of successful
dissection is to keep the muscles clean. Fat, connective tissue, etc.,
is to be carefully but thoroughly cleared away from the surface of
muscles and from between them. Frequently when it seems impossible
to distinguish the structures described, all difficulty will vanish
as soon as the dissection is thoroughly cleaned.
[434]
In transecting a muscle, work under it completely from one edge
to the other (except in case of very wide muscles), then introduce
the scissors or scalpel, and cut it by a smooth clean cut. Always
leave the entire origin with one-half of the muscle, the entire insertion
with the other half.
The student must be prepared to find in some cases considerable
variations from the conditions described. The descriptions attempt
to give an account of the usual structures, but specimens showing
no variations whatever are undoubtedly rare.
DIRECTIONS FOR DISSECTING THE MUSCLES OF THE CAT.
These will perhaps not usually be dissected. If they are to be
dissected, proceed as follows: Make a ventral median longitudinal
incision of the skin from the cranial end of the manubrium to a
point opposite the crest of the ilium. Connect the cranial end of this
incision with the middle of the lambdoidal ridge by a similar incision
through the skin. Make another incision from the manubrium to
the spinous process of the thirteenth thoracic vertebra. Connect
the caudal end of the first incision by an oblique cut with the root of
the tail. Make another incision from the convexity of the knee-joint
along the fold of skin which connects the hind limb and body,
as far as the incision which leads to the root of the tail. Make an
incision surrounding the base of the forelimb. Now reflect these
flaps one at a time, working in general from the ventral side toward
the dorsal middle line. Take great pains not to take up the thin
skin-muscle with the skin. In this way all of one side of the body
will be uncovered. The cutaneus maximus (Fig. 62, b) and a part
of the platysma (Fig. 62, a) will be thus exposed. Read and verify
the descriptions of these muscles (pp. 93-96). The cranial portion
of the platysma cannot be seen at this stage, but should be dissected
in connection with the muscles of the face.
II. Superficial Muscles of the Face and Head
(pp. 96-107, and
Figs. 63 and 64).
These will perhaps not usually be dissected by the student.
They should be studied on a well-dissected laboratory preparation,
if possible. If they are to be dissected, this should be done if
possible on a fresh specimen, as it is very difficult to dissect them
on preserved material. Proceed as follows:
The skin should be removed with care from the sides and top of
the face and head, without injury to the external ear.
1. The platysma (p. 95, and Fig. 62, a). Determine its
cranial attachments.
2. The intermedius scutulorum (p. 96, and Fig. 63, a).
[435]
3. The corrugator supercilii medialis (Fig. 63, b).
4. The orbicularis oculi (Fig. 63, c; Fig. 64, s).
5. The corrugator supercilii lateralis (Fig. 64, k).
6. Notice the frontoauricularis if it exists.
7. The levator auris longus (Fig. 63, g, g′). Transect and
reflect.
8. The auricularis superior (Fig. 63, k). Transect.
9. The abductor auris longus (Fig. 63, m). Transect.
10. The abductor auris brevis (Fig. 63, l). Transect.
11. The epicranius (Fig. 63, h, h′). In dissecting this muscle
the intermedius scutulorum and corrugator supercilii medialis may
be cut by an incision parallel to the middle line but a short distance
to one side of it.
12. The zygomaticus (Fig. 64, d). This and the two following
muscles may usually be best seen running beneath the fibres of the
platysma, without reflecting the latter. If necessary, however, the
platysma may be transected and reflected, though this can be done
only with great difficulty.
13. The submentalis (Fig. 64, c).
14. The depressor conchæ (Fig. 64, b).
The zygomaticus, submentalis, and depressor conchæ should
now be transected one or two centimeters from the external ear.
The corrugatores supercilii lateralis and medialis may be cut across
between the eye and the ear, and reflected. There is thus exposed:
15. The frontoscutularis. Transect near the scutiform cartilage.
16. The adductor auris inferior (Fig. 64, o). Transect near its
insertion.
The external ear is now attached to the head only by the external
auditory meatus and the tragicus lateralis muscle. Read the description
of the latter (p. 418, and Fig. 67, a), find it beneath the fat
which surrounds the proximal portion of the external ear, and cut it
as far ventrad as possible. Now cut across the cartilaginous auditory
meatus as near to the tympanic bone as possible, and remove the
external ear, preserving it for future study. (The remainder of the
muscles of the external ear may be studied in connection with the
study of the auditory organ.)
17. The orbicularis oris (p. 105 and Fig. 64, i).
18. The zygomaticus minor, if it exists (p. 105, and Fig. 64, e).
19. The quadratus labii superioris (p. 105).
a. The levator labii superioris proprius (Fig. 64, f).
b. The levator labii superioris alæque nasi (Fig. 64, g).
20. The caninus (p. 106, and Fig. 64, f′).
21. The buccinator (p. 106).
22. The myrtiformis (p. 106, and Fig. 64, h).
23. The “moustachier” (p. 107).
24. The quadratus labii inferioris (p. 107).
[436]
III. Muscles of the Fore Limb.
A. Muscles connecting the Arm with the Body.
—If the skin-muscles
have been dissected, as above, cut the cutaneus maximus
near its insertion, and reflect it toward the median dorsal line; in
this way remove it completely. Remove the platysma in the same
way. The first layer of body-muscles is thus exposed.
If the skin-muscles are not to be dissected, these may be removed
with the skin. If the skin has not yet been removed, proceed as
follows:
Beginning at the cranial end of the manubrium make two
incisions in the skin, one passing to the lambdoidal ridge and the
other to the spinous process of the thirteenth thoracic vertebra.
Raise the triangular flap thus formed, taking up the skin-muscles
with it.
1. The trapezius group of muscles (pp. 115-117, and Fig. 68,
d, h, and j) is now exposed and may be dissected. Begin with the
spinotrapezius (Fig. 68, j). Read the description of the muscle,
then raise its caudal border and work under it until its cranial border
is reached and the middle of its inner surface is free. Then transect
it, and reflect the two halves of the muscle toward the origin and
insertion, clearing the fat, etc., from the inner surface of the muscle
and the parts covered by it. Determine origin and insertion.
2. The acromiotrapezius (Fig. 68, h). Dissect in the same
manner, being careful not to injure the broad thin tendon which
connects the two muscles across the middle line.
3. The clavotrapezius (Fig. 68, d). Separate it carefully, from
the cleidomastoid (p. 120). Transect and reflect, as before.
4. The occipitoscapularis (Fig. 73, a, p. 149). Note the strong
fascia which separates it from the deeper muscles of the neck. Be
careful not to injure adjacent muscles in tracing this toward origin
and insertion. Transect.
5. The rhomboideus. Transect.
Recognize the levator scapulæ ventralis (Fig. 68, f), the sternomastoid
(Fig. 68, c; Fig. 65, g),
and the cleidomastoid (Fig. 65, h).
Then dissect—
6. The levator scapulæ ventralis (Fig. 68, f). Its origin (Fig.
72, c, c′) cannot be seen at this stage and should be left until the
cervical muscles are dissected. Be careful in transecting this muscle
to separate it well from the cleidomastoid. Transect.
7. The cleidomastoid (Fig. 65, h). Its origin cannot be fully
seen at this stage. Transect.
Make a midventral incision of the skin from the cranial end of
the manubrium to a point opposite the crest of the ilium. From the
caudal end of this incision make an incision to the root of the tail.
From the cranial end of the incision in the ventral middle line make
a cut around the base of the fore limb on its caudal side, thus connecting[437]
the ventral incision with the incision which passes between
the manubrium and the thirteenth thoracic vertebra.
Make another incision from the convexity of the knee at the joint
along the fold of the skin which connects the hind limb and body,
to the incision which leads to the root of the tail.
Reflect the flaps thus formed so as to expose the body and flank,
removing the skin-muscle with the skin.
8. The latissimus dorsi (p. 121, and Fig. 68, m). Dissect it up
and transect it. Work carefully toward its insertion and note the
origin from its outer surface in the axilla of a part of the cutaneus
maximus, and at about the same place of a thin muscle, the
epitrochlearis (p. 164, and Fig. 65, r), without determining the insertion
of the latissimus.
9. Dissect the pectoralis group of muscles (Fig. 65, and p. 144),
i.e., the pectoantibrachialis (Fig. 65, m), the pectoralis major (l),
pectoralis minor (o), and xiphihumeralis (p).
Determine the border of each muscle before lifting it, then work
it up at its middle, transect it, and work toward its ends, determining
origin and insertion.
Refer constantly to the skeleton.
10. Then recognize the teres major (p. 163, and Fig. 75, d; Fig.
77, c) and the common insertion of the teres major and latissimus
(Fig. 79, d′); find the bicipital arch (Fig. 65, t′) and determine its
composition (p. 166).
11 and 12. The serratus anterior and levator scapulæ (Fig. 73,
h and i, and p. 122). In order to expose the origin of these muscles,
the external oblique muscle (Fig. 68, p) must be recognized, and that
part of it which has its origin from the fourth to the ninth rib should
be transected about an inch and a half from its origin and reflected.
The thoracic portion of the rectus abdominis (Fig. 73, k) and of
the scalenus (Fig. 73, f-f‴) should also be recognized and then
raised at their outer borders and displaced toward the median line.
No part of any of these muscles should be removed or cut except as
directed.
The serratus anterior and levator scapulæ should be transected
and reflected to get at their origin and insertion.
B. Muscles of the Arm
(p. 156).—The arm is thus removed from
the body and the dissection may be continued on the separated arm.
13. The clavobrachial (Fig. 65, k, and p. 157). Its origin is best
seen later. Dissect it up, but do not transect it.
Search now with great care for the coracobrachialis. Consult the
figures (Fig. 79, c) and descriptions (p. 164), and be especially careful
not to injure the long head of the muscle and its tendon.
14. Dissect the coracobrachialis (Fig. 77, f). Cut and reflect it.
15. The subscapularis (Fig. 77, a, and p. 161). Near its glenoid
end, its glenoid and coracoid borders are separated by triangular
intervals from the adjacent muscles. Transect the muscle by a line
connecting the apices of these triangles. Carry an incision from the[438]
middle of this one to the middle of the vertebral border of the
scapula, and reflect the muscle on both sides of this incision,—thus
determining its area of origin. Reflect the humeral end without
injuring the capsule of the joint.
16. Dissect the supraspinatus (Fig. 75, a, and p. 159). Cut the
strong fascia free from the border of the scapula and its spine. Then
transect the muscle and reflect it, being careful not to injure the
capsule of the joint.
17. The spinodeltoideus (Fig. 75, e, and p. 156). Find its two
borders, beginning with the caudal one. In freeing the very short
cranial border do not injure the acromiodeltoideus (Fig. 75, f).
Transect and reflect the spinodeltoid.
18. The acromiodeltoideus (p. 157, and Fig. 75, f). Transect
and reflect.
19. The infraspinatus (p. 160, and Fig. 75, c). Locate the
borders in the region of the great scapular notch. It is difficult to
separate the glenoid border from the adjacent teres minor (Fig.
80, c). The separation should be begun at the humerus. Transect
the muscle at the great scapular notch and reflect. Be careful not
to injure the teres minor (Fig. 80, c).
20. The teres major (p. 163, and Fig. 75,
d; Fig. 77, c) should
be cut near its junction with the latissimus dorsi (Fig. 77, e).
21. The teres minor (Fig. 80, c, and p. 161) need not be cut.
22. The epitrochlearis (p. 164, and Fig. 65, r).
23. The triceps (p. 166). (a) The long head of the triceps (Fig.
77, i; Fig. 75, g). Find first its medial border and separate it from
the medial head. Note the union of its lateral surface with the dorsal
border of the lateral head by strong fascia, and then cut the fascia.
(b) The lateral head (Fig. 75, h).
(c) The medial head (Fig. 79, g, h, and j).
24. The anconeus (p. 170, and Fig. 80, l).
25. The brachioradialis (p. 173, and Fig. 75, k). Reflect it
carefully to its origin and insertion.
26. The biceps (Fig. 77, g, and p. 165). The capsule of the
shoulder-joint may be opened in order to expose its origin. Its
insertion cannot be seen at this stage.
27. The brachialis (Figs. 75 and 79, i, and p. 166). Work under
it near its union with the clavobrachial; cut it at that point and reflect
it.
28. Extensor carpi radialis longus (p. 173, and Fig. 75, l). Note
the oblique tendon on the radial side of the wrist near the insertion
of the brachioradialis. A second tendon passes beneath its distal
border. This tendon may be separated into a deep and superficial
portion. The latter may be traced proximad to its muscle, the
extensor carpi radialis longus.
29. Extensor carpi radialis brevis (p. 174). Its tendon is the
deeper of the two tendons seen when isolating the tendon of the
extensor longus.
[439]
30. Extensor communis digitorum (p. 174, and Fig. 75, m).
31. Extensor lateralis digitorum (p. 175, and Fig. 75, n).
32. Extensor carpi ulnaris (p. 176, and Fig. 75, o).
33. Extensor indicis (p. 176, and Fig. 85, c).
34. Pronator teres (p. 179, and Fig. 77, q).
35. Flexor carpi radialis (p. 179, and Fig. 77, r). The tendon
of the flexor carpi radialis should not be traced to its insertion until
the deep muscles of the palm of the hand have been dissected.
36. Abductor brevis pollicis (p. 184, and Fig. 77, w).
37. Flexor carpi ulnaris (p. 180, and Fig. 77, t).
38. Palmaris longus (p. 179, and Fig. 77, s). In dissecting this
muscle be careful not to injure the part of the flexor sublimis (Fig.
77, x) that rises from its surface (see flexor sublimis, p. 181). Cut
the palmaris proximad of the origin of the flexor sublimis and reflect
it.
39. Flexor sublimis digitorum (p. 181 and Fig. 77, x). Trace
one or two of the tendons through the sheaths on the ventral surfaces
of the first phalanges. Cut and reflect the radial portion.
40. Flexor profundus digitorum (p. 181, and Fig. 77, u). After
recognizing the parts cut through the tendons of the first and fifth
parts and reflect them to determine their origin. Then cut through
the common tendon so as to reflect the other three parts together.
Trace one or two of the tendons to the distal end of the digit.
41. The supinator (p. 177, and Fig. 85, b).
42. The extensor brevis pollicis (p. 178, and Fig. 85, a).
43. Pronator quadratus (p. 183, and Fig. 87, a).
The insertion of the brachialis and clavobrachial (Fig. 87, c)
and of the biceps (Fig. 87, b) should now be examined.
There remain to be dissected the small muscles of the palm of
the hand. If these have been injured on the side dissected, the
hand of the other side may be used.
44. The lumbricals (p. 184, and Fig. 88, f).
45. The flexor brevis pollicis (p. 184, and Fig. 89, a).
46. The adductor pollicis (p. 185, and Fig. 89, b).
47. The abductor digiti quinti (p. 185, and Fig. 89, i).
48. The flexor brevis digiti quinti (p. 186, and Fig. 89, h). Cut
and reflect it.
49. The opponens digiti quinti (p. 186, and Fig. 89, g). Cut
and reflect it.
50. The interossei (p. 185, and Fig. 89).
IV. Muscles of the Neck and the Deep Muscles of the Head.
If the superficial muscles of the head have been dissected according
to the directions above given (p. 434), the skin of the head and
neck, and the external ear, will have been removed, and the specimen
is ready for further study. The parotid and submaxillary glands[440]
(Fig. 65, 1
and 2) should be removed if this has not been done; also
the lymphatic glands (Fig. 65, 3).
If the superficial muscles of the head have not been dissected and
are not to be, remove the skin from the side of the neck, head, and
face to the median dorsal line, removing the thin superficial muscles
with the skin. Remove also the parotid, submaxillary, and lymphatic
glands (Fig. 65, 1,
2, and 3), and cut through the ear-muscles
and the cartilaginous auditory meatus (under direction) and remove
the external ear. Clean fat, connective tissue, etc., from the surface
of the muscles of the thorax, neck, and head; then dissect as follows:
1. The sternomastoid (p. 139, and Fig. 65, g). Transect and
reflect it.
2. The cleidomastoid (p. 120, and Fig. 65, h). This has
already been transected; examine now its origin.
Remove the large lymphatic gland beneath the sternomastoid and
close to the tympanic bulla, and clean the surface of the muscles in
this region.
3. The levator scapulæ ventralis (p. 120,
and Fig. 72, c, c′, c″).
This has already been cut; study now its origin.
4. The sternohyoid (p. 140, and Fig. 65, e). In raising it begin
at the cranial end and take care not to injure the subjacent sternothyroid
(Fig. 65, g′). Transect.
5. The sternothyroid (p. 141, and Fig. 65, g′). Transect.
6. The stylohyoid (p. 112, and Fig. 65, d). Transect.
7. The digastric (p. 107, and Fig. 65, b). Transect, and reflect
completely, to the origin and insertion.
8. The mylohyoid (p. 114, and Fig. 65, c). Transect and
reflect.
9. The geniohyoid (p. 113, and Fig. 67, g).
10. The jugulohyoid (p. 113, and Fig. 67, b).
(The extrinsic muscles of the tongue (p. 228, and Figs. 67 and
96) may be dissected at this point, if desired. (a) The styloglossus;
(b) the genioglossus; (c) the hyoglossus. They need not be cut.)
11. The masseter (p. 108, and Fig. 65, a). Cut the superficial
and middle layers near their insertions by incisions parallel to the
border of the jaw. Cut the deep layer near its origin. Look for the
origin of fibres of the temporal muscle from the inner surface of the
middle layer.
12. The temporal (p. 110, and Fig. 63, n). Clear its outer surface
completely. Cut the temporal fascia transversely and reflect it
toward the insertion of the muscle, with the superficial portion of the
muscle. Cut through the zygomatic arch at its two ends and remove
it, dissecting the fibres of the temporal free from their origin on its
inner surface. Then examine the insertion of the temporal.
(The muscles of the pharynx (p. 232) may be dissected at this
point; for directions, see page 451.)
13. The ceratohyoid (p. 115). To uncover this, the middle
constrictor of the pharynx (Fig. 67, j) must be cut and reflected.
[441]
Cut the mylohyoid, geniohyoid, genioglossus, and the mucosa
of the floor of the mouth free from the mandible on both sides, so
that the tongue and the floor of the mouth may be drawn ventrad
and turned out of the way, exposing the roof of the mouth (as in
Fig. 66, p. 112). Cut through one side of the pharynx near the
ventral surface and turn the tongue toward the uncut side.
The roof of the mouth is thus exposed. Dissect next—
14. The pterygoid muscles (Fig. 66). Remove the mucosa of
the roof of the mouth, especially at the sides of the soft palate, and
the two pterygoid muscles (p. 111) are exposed. The lower jaw
may be removed on one side to get at their origin.
(The muscles of the soft palate (p. 230, and Fig. 66, d and e),
and the superior constrictor of the pharynx (p. 233, and Fig. 66, f)
may be observed at this point, if desired.)
V. Muscles of the Wall of the Thorax and of the Abdomen.
1. The scalenus (p. 141, and Fig. 73, f-f‴). To trace its
cervical portion, cut the trachea and œsophagus just craniad of the
first rib, cut the pharynx and hyoid free from the ventral side of the
head and neck, and remove the pharynx, larynx, œsophagus, and
trachea (these should be preserved for future study of the larynx).
2. The transversus costarum (p. 150, and Fig. 73, j).
3. The serratus posterior superior (p. 148, and Fig. 73, l).
Transect this at about the middle of the muscle-bundles, and reflect
it in both directions.
4. The serratus posterior inferior (p. 148, and Fig. 73, n).
Transect and reflect as in the last.
5. The external oblique (p. 153, and Fig. 68, p). Transect it
by an incision parallel to its tendon of origin and about an inch from
it, and reflect in both directions.
6. The internal oblique (p. 154, and Fig. 73, o). Transect and
reflect as above.
7. The transversus (p. 155, and Fig. 69, l). Requires no
further dissection.
8. The rectus abdominis (p. 155, and Fig. 73, k). Open its
sheath by a longitudinal incision near the linea alba.
9. The external intercostals (p. 150, and
Fig. 73, m; Fig. 69, i).
To see these, remove the origins of the levator scapulæ and serratus
anterior; also the serratus posterior superior.
10. The internal intercostals (p. 151, and Fig. 69, k). To
uncover these, remove the external intercostals from between two or
three pairs of ribs.
11. The transversus thoracis (p. 151). To see this muscle,
which lies on the inner surface of the thoracic wall, it is necessary to
cut through the ribs on one side one or two inches from the sternum,
and open the thorax. The muscle can then be observed directly.
(The levatores costarum will be dissected later.)
[442]
VI. Muscles on the Ventral Side of the Vertebral Column in
the Neck Region (Fig. 72).
Clean the surface of these muscles, removing pharynx, larynx,
œsophagus, and trachea, if this has not been done.
1. Longus capitis (p. 142, and Fig. 72, a).
2. Longus colli (p. 144, and Fig. 72, g′, g″).
3. Levator scapulæ ventralis (p. 120,
and Fig. 72, c′, c″).
Observe its origin.
4. Rectus capitis anterior minor (p. 143, and Fig. 72, b). To
see this, cut through on one side the longus capitis and one head of
the levator scapulæ ventralis, and remove the proximal portions, as
in Fig. 72.
5. Obliquus capitis superior (p. 136, and Fig. 72, e).
6. Rectus capitis lateralis (p. 143, and Fig. 72, d).
VII. Muscles of the Back (p. 123).
Remove what remains of the dorsal portion of the latissimus dorsi,
serratus posterior superior, and serratus posterior inferior. See that
the skin is removed completely to the dorsal middle line, or even for
a little distance over onto the other side. There is thus exposed
the lumbodorsal fascia (p. 126), covering the muscles of the back.
Cut the superficial layer of the lumbodorsal fascia (that from
which the latissimus dorsi and obliquus externus abdominis have
origin) by an incision parallel to the dorsal median line, and remove
it. The muscles of the back are thus exposed (Fig. 69). Observe
in the lumbar region the longissimus dorsi (f, f′, f″); in the
thoracic region the longissimus dorsi, the spinalis dorsi (g), and the
iliocostal (h); in the cervical region the splenius (Fig. 73, b).
1. The longissimus dorsi (p. 126, and
Fig. 69, f, f′, f″).
Notice in the lumbar region the strong deep layer of the lumbodorsal
fascia, by which this muscle is covered, and from the under side of
which many of the fibres of the muscle take origin. Cut this fascia
by an incision parallel to the dorsal median line and about two
centimeters from it, from the level of the crest of the ilium as far
craniad as the fascia can be cut without cutting into the muscle-fibres
(usually to about the level of the last rib). Reflect the medial
division of the fascia to the middle line. Reflect the lateral division
of the fascia until it passes into the muscle and cannot be farther
reflected.
Observe then in the lumbar region the medial (Fig. 70, a) and
lateral (Fig. 70, b) divisions of the longissimus dorsi, and the portion
(b′) of the lateral division taking origin from the lumbodorsal fascia
(c). Dissect apart some of the muscle-fibres of the longissimus and
observe their origin and insertion.
Trace the longissimus dorsi into the thoracic and cervical regions[443]
(Fig. 69, f′, f″) noting the separation off from it on the dorsal side
of the spinalis dorsi (Fig. 69, g); on the ventral side of the iliocostal
(Fig. 69, h).
2. The extensor caudæ lateralis (p. 137, and Fig. 70, f).
Uncover this by cutting the lumbodorsal fascia over the sacral region.
3. The iliocostalis (p. 128, and Fig. 69, h). Uncover this
completely; note its connection with the longissimus dorsi at the
caudal end. Dissect apart some of its muscle-bundles, to see origin
and insertion.
4. The splenius (p. 131, and Fig. 73, b). Transect this by an
incision beginning at its lateral border about four centimeters from
the cranial end of the muscle and extending obliquely craniodorsad
to the craniomedial angle of the muscle. Be careful not to injure
the longissimus capitis (Fig. 73, g).
5. The longissimus capitis (p. 131, and Fig. 73, g). Transect.
6. The spinalis dorsi (p. 129, and Fig. 69, g). Separate some
of its muscle-fibre bundles and trace to origin and insertion.
7. The biventer cervicis (p. 132, and Fig. 69, a). Raise its
lateral border, turning it toward the middle line, and transect it near
its caudal end.
8. The complexus (p. 133, and Fig. 69, b). Raise its medial
border, and transect it near its cranial end.
9. The longus atlantis (p. 134, and Fig. 71, f).
10. The multifidus spinæ (p. 130, and Fig. 70, d). Remove a
part of the longissimus dorsi in the lumbar region, and trace the
course of the fibres of the multifidus spinæ. (If desired the entire
longissimus may be removed and the multifidus spinæ completely
uncovered.)
11. The extensor caudæ medialis (p. 136, and Fig. 70, e).
12. The semispinalis cervicis (p. 133, and Fig. 71, c). Dissect
apart some of its fibre-bundles to see origin and insertion.
13. The interspinales and intertransversarii (p. 131) may be seen
in the region from which the longissimus dorsi was removed, by
removing the bundles of the multifidus spinæ.
14. The rectus capitis posterior major (p. 134, and Fig. 71, a).
Transect.
15. The obliquus superior (p. 136, and Fig. 71, e).
16. The obliquus capitis inferior (p. 136, and Fig. 71, b).
17. The rectus capitis posterior medius (p. 135). Transect.
18. The rectus capitis posterior minor (p. 135).
19. The levatores costarum (p. 150). Remove in a part of the
thoracic region a portion of the iliocostal and longissimus dorsi, so
as to expose the dorsal ends of the ribs. The levatores costarum
will be seen as but slightly separated bundles continuous with the
external intercostals.
[444]
VIII. Muscles of the Hind Limbs (p. 186).
Remove the skin and superficial fascia from the lateral surface of
the thigh. Examine the fascia lata (p. 186, and Fig. 68, z).
Dissect:
1. The biceps femoris (p. 194, and Fig. 68, t). Work under its
cranial and caudal borders. Find the tenuissimus (Fig. 90, g) passing
obliquely beneath it and reaching its caudal border in its distal
half. Transect the biceps without injuring the tenuissimus, and
reflect its halves.
2. The tenuissimus (p. 195, and Fig. 90, g). Transect.
3. The caudofemoralis (p. 195, and Fig. 68, s). Transect.
4. The gluteus maximus (p. 187). Transect.
5. The sartorius (p. 197, and
Fig. 68, q; Fig. 91, a). Transect.
6. The tensor fasciæ latæ (p. 187,
and Fig. 68, r). Cut the fascia
lata along the distal end of the muscle-fibres and reflect the muscle.
7. The gluteus medius (p. 188, and Fig. 90, b). Work under it
carefully, separating it from underlying muscles; cut it close to its
tendon of insertion, and reflect it.
8. The pyriformis (p. 188, and Fig.
163, 7, p. 401). Cut and
reflect it.
9. The gemellus superior (p. 189, and Fig.
163, 6). This and
the next muscle will sometimes be found to be completely united.
Transect.
10. The gluteus minimus (p. 189, and Fig. 163,
5). Transect.
11. The obturator internus (p. 192, and Fig. 90, e). Cut it at
the dorsal border of the ischium in order to reflect it. The whole
origin cannot be seen at this stage.
12. The gemellus inferior (p. 190).
13. The gracilis (p. 198, and Fig. 91, b). Transect.
14. The semitendinosus (p. 196, and Figs.
90 and 91, j; Fig.
92, i). Transect.
15. The semimembranosus (p. 196, and Figs.
90 and 91, i; Fig.
92, h, h′). Transect.
16. The adductor femoris (p. 198, and Figs.
90 and 91, h; Fig.
92, g). Transect.
17. The quadratus femoris (p. 191, and Fig.
90, f). Transect.
18. The obturator externus (p. 191).
19. The adductor longus (p. 199, and Fig. 92, f). This and
the following will sometimes be found to be almost completely
united. Transect.
20. The pectineus (p. 200, and Fig. 92, e). Transect.
21. The iliopsoas (p. 193, and Fig. 91,
c; Fig. 92, d). Only
its distal part, near the insertion, can be seen at present.
22. The capsularis (p. 190). Transect.
23. The quadriceps femoris (p. 201). Isolate the rectus femoris
(Fig. 92, b) as far as its junction with the vastus
lateralis; cut it at[445]
this point and reflect its proximal portion. Turn aside the distal
end and find beneath it the transverse aponeurosis between the vastus
medialis and the vastus lateralis, and covering the vastus intermedius.
The vastus lateralis (Fig. 90, d) may be cut near the middle and
dissected up from its origin in both directions; the vastus intermedius
is thus exposed. The vastus medialis (Fig. 92, c) and the vastus
intermedius need not be cut.
24. The gastrocnemius (p. 203, and
Fig. 90, m; Fig. 91, k).
It may be raised cautiously near its middle and divided. Care
should be taken not to divide the underlying plantaris (Fig. 90, l).
After noting the connection of the outer head with the plantaris, the
latter may be divided and reflected, together with the outer head.
25. The plantaris (p. 205, and Figs. 90 and 91, l).
26. The flexor brevis digitorum (p. 212). In tracing its tendon
note the three branches of the common plantar ligament which
emerges from between the second and third tendons. The outer
branch is inserted into the fascia covering the base of the fifth digit,
while the other two spread out into the bilobed pad of the sole.
27. The soleus (p. 205, and Fig. 90, o). Cut and reflect.
28. The popliteus (p. 206, and Fig. 92, k). Cut very near the
origin and reflect.
The three following deep muscles on the ventral surface of the
shank are covered by the deep crural fascia (p. 206) which separates
them from the overlying muscles. It should be removed.
29. The flexor longus hallucis (p. 207, and Fig. 91, m). Divide
the muscle at its junction with the tendon. Open the canals on the
astragalus and calcaneus and reflect the tendon, cutting the tendon
of the flexor longus digitorum near its insertion and dividing at the
middle the quadratus plantæ (p. 213) which may be found inserted
into its ventral surface.
30. Flexor longus digitorum (p. 208, and Fig. 91, n). This
may be divided very near its origin.
31. Tibialis posterior (p. 208, and Fig. 91, o). This may be
divided near its tendon.
32. Peroneus longus (p. 209, and Fig. 90, q). The overlying
tendons may be cut in tracing its insertion.
33. Peroneus brevis (p. 210, and Fig. 90, s).
34. Peroneus tertius (p. 210, and Fig. 90, r).
35. Extensor longus digitorum (p. 210, and Fig. 90, p).
36. Tibialis anterior (p. 211, and Fig. 90, n).
37. The extensor brevis digitorum (p. 212, and Fig. 90, u).
38. The quadratus plantæ (p. 213). It was cut in dissecting the
flexor longus hallucis.
39. The lumbricales (p. 213).
40. The interossei (with the muscles of the fifth digit) (p. 214).
41. The tarsal muscles (p. 215). The overlying ligaments must
be removed in order to expose the scaphocuneiform.
[446]
IX. The Diaphragm and the Muscles on the Ventral Side of
the Vertebral Column in the Lumbar and Pelvic Regions.
Remove the ventral and lateral walls of the thorax completely,
as far back as the diaphragm,—leaving only a ring attached to the
margin of the diaphragm. Remove the abdominal walls in the same
way, as far back as the pubis, leaving the diaphragm in position.
Remove the thoracic viscera (the heart and lungs may be preserved
for future study). Remove also the abdominal viscera as far back
as the beginning of the pelvis, cutting the large intestine where it
passes into the pelvis. The dorsal wall of the abdominal cavity
should be cleared of fat, blood-vessels, etc., without injury to the
crura of the diaphragm.
1. The diaphragm (p. 151, and Fig. 74). Study by transmitted
light, to see the tendons, etc.
2. The psoas minor (p. 139, and
Fig. 162, 9, p. 398). Find its
tendon and separate it from the iliopsoas (Fig. 162, 8) as far craniad
as this can be done without tearing the fibres.
3. The iliopsoas (p. 193, and
Fig. 162, 8). Find both portions;
isolate them as far as it can be done.
4. The quadratus lumborum (p. 139). Free the lateral edge of
the iliopsoas from the abdominal wall and turn the whole muscle
mediad as far as possible. The bundles of the quadratus lumborum
will be found beneath it, against the transverse processes of the
lumbar vertebræ.
With a heavy cartilage-knife separate the two innominate bones
along the pelvic symphysis. This is easily done if the knife is
inserted exactly into the line of junction of the two bones. Divaricate
the edges some distance, so as to make a ventral opening into the
pelvic cavity. If necessary, part of the innominate bone of one side
may be removed to make the opening larger.
Find the thin sheet of fibres forming the levator ani muscle (Fig.
162, 11) at the side of the rectum, attached to the edge of the symphysis.
Carefully free the rectum and urogenital organs from this
and remove them as far as the anus, leaving the levator ani intact.
5. The iliocaudalis (p. 137, and
Fig. 162, 11′).
6. The levator ani (p. 269, and
Fig. 162, 11).
Remove with great care the levator ani and iliocaudalis of one
side, by cutting first the attachment to the symphysis and the ilium,
turning the muscle mediad, then cutting the attachment to the tail.
There are thus exposed on the lateral wall of the thorax the medial
surface of the obturator internus, craniad of it the pyriformis, caudad
of it the quadratus femoris. Dorsad of these is the abductor caudæ
internus.
7. The abductor caudæ internus (p. 137).
On the ventral surface of the tail are exposed the following:
8. The flexor caudæ longus (p. 138,
and Fig. 162, 12).
9. The flexor caudæ brevis (p. 138,
and Fig. 162, 13).
[447]
X. Muscles of the Tail (p. 136).
By removing the skin of the tail, all the muscles of the tail are
now easily seen.
1, 2, 3, 4, 5—see 5, 6, 7, 8, 9, above (IX).
6. The abductor caudæ externus (p. 137,
and Fig. 113, b; Fig.
70, g).
7. The extensor caudæ lateralis (p. 137, and Fig. 70, f).
8. The extensor caudæ medialis (p. 136, and Fig. 70, e).
THE VISCERA.
Kill the specimen with chloroform and inject with the five per
cent. formalin, or the mixture of formalin and glycerine, as for the
muscles. (It is an advantage if the preliminary examination of the
viscera can be done in a perfectly fresh, uninjected specimen; such
a specimen can be kept but a day or two, however. After one day’s
examination the skin may be removed and the specimen placed in
two per cent. formalin; it will usually be fairly well preserved by
this, though the formalin is too strong to make the later dissection
pleasant. The specimen cannot be preserved in this manner unless
the skin is removed, and a sufficient quantity of formalin used.)
Preliminary Examination of the Viscera.
(In this preliminary examination the descriptions in the text need
not be read, except when this is specially directed.)
Wet the hair along a line from the symphysis of the jaw to the
pubis. Part it and make a longitudinal incision through the skin
along the line. Make a transverse incision of the skin five centimeters
caudad of the xiphoid process and reflect the flaps. Incise the body
wall from the last rib to the pelvis along the same line as above and
also transversely. Reflect the flaps without injuring the ligaments of
the bladder. Make an incision through the pectoral muscles between
the first rib and the ventral attachment of the diaphragm and about
one centimeter from the median line. Make a second incision of the
muscles from the cranial end of the first incision along the angles of
the ribs to near the dorsolateral attachment of the diaphragm. Cut
the ribs and thoracic wall along these incisions with strong scissors;
connect the two incisions at their caudal ends, and remove the lateral
thoracic wall thus circumscribed. Note the thoracic and abdominal
cavities, separated by the diaphragm.
Note the following viscera in their fresh condition:
A. In the abdominal cavity (without injuring anything)
1. The liver, dark red and lobed, in the cranial part of the
cavity, lying against the diaphragm.
2. The gall-bladder partly imbedded in the right median lobe.
3. Displacing the liver craniad, the stomach is seen dorsad of its
left lobe. It may extend dorsad of the right lobe, its size depending
on the degree of distension.
[448]
4. The duodenum or first part of the small intestine leaves the
stomach at its right side craniad and makes a U bend, the head of
which is directed caudad; it then turns toward the middle line dorsad
of the remaining part of the small intestine.
5. The great omentum is a thin fold of peritoneum with many
bands of fat. It passes like a curtain from the great curvature of
the stomach ventrad of the coils of the small intestine (ileum). It is
tucked beneath the mass of coils at the sides and caudally, and folds
of it extend between the individual coils. Turn it craniad and note—
6. The numerous coils of the third part of the small intestine or
ileum. Turn these to one side and note—
7. The connection of the ileum with the duodenum across the
middle line. An indefinite part of the intestine between the ileum
and duodenum is called the jejunum, because in man it is found
empty after death.
8. The passage of the ileum into the side of the large intestine
which begins on the right side just caudad of the loops of the
duodenum. The first part of the large intestine is the colon. It
may be traced first toward the head (ascending colon), then sinistro-caudad
(transverse colon) and then caudad (descending colon). The
descending colon returns to the middle line and the large intestine
then continues to the anus as the rectum.
The blind pouch of the colon which lies caudad of the opening
of the ileum is the cæcum.
9. Turn the duodenum toward the left and note the pancreas,
a light red, elongated organ which stretches along the greater curvature
of the stomach and thence between the two lines of the duodenal
U to its bottom.
10. The spleen, parallel to the gastric portion of the pancreas.
It is deep red (darker than the pancreas and not so brown as the
liver) and is held against the great curvature of the stomach by a part
of the great omentum.
11. The kidneys, close against the dorsal body wall and in contact
with the caudal part of the liver.
12. The bladder, in the median line at the caudal end of the
cavity, held in place by the suspensory ligament, which passes to the
midventral line, and by the lateral ligaments.
13. The mesenteric glands, large grayish-red glands one to four
centimeters long, more numerous in the mesocolon.
14. In the female, lying against the ventral surface of the rectum,
the uterus. It divides into two horns which diverge. Near the end
of each horn the ovary.
15. The peritoneum. Study and understand the description of
it (p. 218). Dissect some of it free from the body wall. (Trace the
folds by aid of a diagram and model.)
B. In the thoracic cavity:
1. The lungs. Each is divided into lobes, three on the left, four
on the right. The caudal right lobe pushes the mediastium toward[449]
the left and thus lies in a pocket dorsocaudad of the heart and
dorsad of the vena cava inferior.
2. The pleuræ. Each pleura is a membrane like the peritoneum.
It covers the diaphragm and the thoracic walls and is reflected onto
the lungs. Hence it is divided into costal and pulmonary portions.
Read the description (p. 217) and trace their connection. Each
pleura forms a closed sac.
3. The mediastinal septum or median vertical portion formed by
the apposition of the medial walls of the two pleural sacs. It divides
the thorax into halves.
4. In the mediastinum or space between the halves of the
mediastinal septum are seen—
a. Ventrally (i.e., in the anterior mediastinum), blood-vessels
and fat, and craniad the pink thymus gland. Read the description
of the thymus (p. 254, and Fig. 107).
b. Dorsally (i.e., in the posterior mediastinum) the trachea (best
seen craniad), the œsophagus (best seen caudad), and the aorta.
c. In the middle (reckoning in a dorsoventral line) (i.e., in the
middle mediastinum) the heart, with the inferior vena cava approaching
it from the rear. Prick into the mediastinum and inflate it. Remove
the thoracic wall opposite the one already removed, so as to stretch
the mediastinum and show it more clearly. Expose the trachea
beneath the caudal end of the sternomastoidei. Note the thyroid
gland (Fig. 96, 6,
p. 229). Insert a blowpipe without injury to the
thyroid, and inflate the lungs. Cut the sternum at its caudal end and
turn it craniad, cutting the mediastinal septum and noting its double
character.
Dissect the mediastinal septum from the thymus so as to expose
this organ.
d. The pericardium, a relatively thick-walled sac, within which the
heart lies. The pericardium is covered by the mediastinal septum
on each side, but not dorsally and ventrally. Dissect the mediastinal
septum from the pericardium. Prick and inflate the pericardium.
Open it and expose the heart.
Dissection of the Alimentary Canal.
Study and verify the descriptions of the organs as they are dissected.
I. Salivary glands (p. 223).
1. Keep the mouth open by a cork between the teeth. Find the
papillæ upon which open the sublingual and submaxillary ducts (pp.
223 and 224); remove the mucosa and enlarge the opening and
insert a black bristle into either duct. Trace the duct by the bristle
as far as possible on the floor of the mouth.
2. Find the white ridge formed on the cheek by the parotid duct
(p. 223). It is opposite the molar tooth, and at its cranial end the[450]
opening of the duct is seen. Enlarge the opening and insert a black
bristle as far as possible.
3. Remove the skin and the skin-muscles from the side of the
face, beginning at the caudal end and working craniad. At the
cranial border of the carotid on the ventral side look carefully for the
parotid duct with the bristle in it. The duct is then easily traced
by the removal of the skin and muscles. Study the parotid gland
(Fig. 131, 10, p. 322).
4. Remove the facial vein (Fig. 131, b and c) and lymphatic
gland (Fig. 131, 12)
covering the submaxillary gland (Fig. 131, 11)
and clean its outer surface. Raise it and find the submaxillary duct
leaving its inner surface. Reflect the digastric and mylohyoid
muscles and trace the duct to the oral mucosa. The remainder of
its course is shown by the bristle on the inside of the mouth.
5. The sublingual gland is uncovered in dissecting the submaxillary.
Its duct is seen by the side of the submaxillary. It may be
pricked near the gland and a bristle passed to its opening.
6. The infraorbital gland. Expose it by removal of the zygomatic
arch and masseter muscle. Find its duct by dissecting apart
the lobules at its ventral end. Prick the duct and insert a bristle to
find its inner opening.
7. The molar gland. Find it by raising the orbicularis oris of
the lower lip. By raising its ventral border and dissecting between
it and the mucosa the ducts may be found and perhaps pricked and
bristles inserted.
II. The mouth-cavity. Study the general description (p. 221);
the lips and the cheek. Then remove the masseter muscle and the
caudal part of the zygoma. Cut through the mandible between the
first premolar and the canine tooth. Then disarticulate the mandible,
cut the cheek and also the mylohyoid muscle and oral mucosa
parallel to the medial border of the mandible, and remove the mandible
with the parts thus left attached to it. The organs in the
mouth-cavity are thus left exposed and should be studied. To
demonstrate the incisive ducts (p. 222) pass a bristle into them.
The teeth (p. 224). These should be studied on a skull in
which the roots of the teeth have been laid bare by means of the
bone-forceps, as in Figs. 93 and 94.
The tongue (p. 226, and Fig. 95). Study its dorsal surface, the
papillæ, etc. Then dissect its muscles (p. 228) from the ventral side.
The stylohyoid, digastric, and mylohyoid muscles must be removed.
Then dissect (a) the styloglossus (Fig. 96, e), (b) the genioglossus
(Fig. 96, f), (c) the hyoglossus (Fig. 96, h). Make a transverse
section of the tongue to see the intrinsic muscle-fibres.
III. The thyroid gland (p. 254, and Fig. 96, 6). Clean the
sternomastoid muscles. Cut their interdigitating portions along the
median line and reflect them. Find the lateral lobe of the thyroid
gland beneath the lateral borders of the sternohyoid muscles.
Dissect it, being careful not to destroy the delicate isthmus.
[451]
IV. The pharynx (p. 231). Remove the large lymphatic gland
between the atlantal transverse process and the larynx. Clean the
outer surface of the pharynx. Dissect the muscles of the pharynx
(p. 232) as follows:
(a) The inferior constrictor (Fig. 96, k).
(b) The middle constrictor (Fig. 96, j).
(c) The glossopharyngeus (Fig. 96, i).
(d) The stylopharyngeus (Fig. 96, l).
The superior constrictor cannot well be seen at this point; it will
be examined later.
Disarticulate the cranial cornu of the hyoid from the bulla
tympani and make an incision the length of the lateral wall of the
pharynx so as to expose its cavity; study. Examine its opening into
the mouth-cavity and study the general description (p. 231). Find
and study the soft palate (p. 230). Dissect the tensor and levator
palatini muscles (p. 230, and Fig. 66,
d and e, p. 112), using, if
necessary, the specimen on which the other muscles were dissected.
Then slit the soft palate lengthwise at one side of the median line to
expose the nasopharynx (p. 231). Bring the choanæ into view by
use of a bit of mirror-glass. Pass a bristle into the nares and out at
the choanæ. Pass a bristle through the Eustachian tube into the
middle ear.
V. Œsophagus (p. 234). After completing the study of the
pharynx, leave the larynx and lungs in position and by displacing
them toward the right follow the œsophagus to its termination.
Open it to see the folds of the mucosa.
VI. Cut through the duodenum at its distal end and through the
duodenal mesentery so as to separate the stomach and duodenum,
with the liver, pancreas, and spleen, from the remainder of the
alimentary canal. Float the parts in a dish of water.
VII. Study the stomach (p. 234, and Fig. 97). Cut out the
ventral wall of the stomach and wash it out, so as to study its cavity,
and then continue the cut so as to expose the pyloric valve (p. 235).
VIII. Study the duodenum (p. 236). Cut away the ventral
walls of the duodenum far enough to expose the ampulla of Vater.
IX. Study the liver (p. 239, and Figs. 100 and 101). Expose
the hepatic duct of the left lateral lobe near its entrance into the
lobe, and follow it so as to expose it fully. If necessary, prick it and
inflate with blowpipe in order to follow it. Then expose the cystic
duct and other hepatic ducts and follow them to or from the common
bile-duct. Trace the latter to the duodenum. If the air does not
enter any duct readily, it may be made to do so by manipulating the
duct so as to break up the precipitated bile which obstructs it.
X. Study the pancreas (p. 241, and Fig. 102). Expose the
pancreatic duct near its entrance into the ampulla of Vater, by
removing the peritoneum from the pancreas just caudad of the end
of the common bile-duct, and by dissecting apart the pancreatic
lobules until the duct appears. (If possible, expose also the accessory[452]
duct in the same way on the ventral side of the pancreas, two
centimeters caudoventrad of the ampulla of Vater. Prick and inflate.
Trace its connection with the main duct. To demonstrate its opening
pass a bristle through an opening in it into the duodenum.)
(The pancreatic duct may be injected, if desired.)
XI. The ventral wall of the duodenum should now be removed.
Cut out a small piece, clean its mucosa with a fine brush and
examine the villi with a lens. Demonstrate the coats of the duodenal
wall by stripping them off with forceps.
XII. The ampulla of Vater (p. 236). Pass bristles through
openings in the common bile-duct and pancreatic duct into the
duodenum through the ampulla. Slice away the duodenal wall
parallel to the bristles until the bristles are exposed.
XIII. The spleen (p. 242, and Fig. 102).
XIV. Cut the mesentery from the small intestine and colon and
slit them both lengthwise, but do not destroy the ileocolic valve.
Wash and brush the mucous membrane clean and study the villi,
solitary glands, and Peyer’s patches. Study the ileocolic valve
(Fig. 99) and open it to study its inner surface.
DISSECTION OF THE ORGANS OF RESPIRATION.
I. The nasal cavity (p. 243). Review the description of the
nasal cavity given under the Bones (p. 59). Study the cartilaginous
framework that supports the external nose (p. 243); make a cross-section
of the framework near the end of the nose and verify the
description (p. 244, and Fig. 103). Without injuring the larynx or
tongue saw through the head in a vertical plane and a little to one
side of the median line. (If desired, the brain may first be removed
from the specimen and preserved for future study. For directions,
see page 462. The removal of the brain does not injure the head
for the study of the nasal cavity; the skull may be sawn lengthwise
in the same way as before.) Wash out the cut surfaces before
examining. Find the ventral conchæ (p. 40), the labyrinths of the
ethmoid (p. 43), the inferior meatus of the nose (p. 243), and by
bristles the lachrymal duct or canal (p. 245). The lachrymal
duct is conveniently found by passing bristles into the openings
of the lachrymal canals. One of these is to be found on the border
of each eyelid, two or three millimeters from the inner angle of the
eye (see p. 410).
Cut the œsophagus and trachea at the point where the blowpipe
was inserted into the trachea. Separate the tongue, hyoid, larynx,
œsophagus, and first part of the trachea from the adjacent parts and
remove them.
II. The larynx (p. 246). (1) Read the description, studying the
cartilages (p. 247) on a preparation (Fig. 104). Then very carefully
remove the pharynx and its constrictor muscles, the sternothyroid
and remains of sternohyoid muscles, and (2)
dissect the muscles of[453]
the larynx (p. 249, and Fig. 105). Dissect all those of one side
first. The mucosa lining the vestibule and middle portions of the
laryngeal cavity must be removed. After studying the muscles,
remove the muscles and entire mucosa by scraping carefully with a
dull but smooth scalpel, and study the cartilages (Fig. 104).
III. The trachea and lungs (p. 251, and Fig. 106). The distal
end of the trachea and lungs may now be removed with the heart
from the body. The lungs may be inflated with the bellows. The
bronchi should then be exposed, cleaned, and followed as far as
possible into the lungs. The heart should be preserved for future
study.
DISSECTION OF THE UROGENITAL ORGANS.
A. Excretory Organs.
1. Expose the kidney (p. 255, and Fig. 108) by removal of the
peritoneum and the surrounding fat, taking care not to open the
capsule of the kidney. In removing the fat from the cranial end do
not injure the suprarenal body (p. 257), which should be studied.
2. Open the capsule of the kidney and slice away its ventral wall
to expose the sinus. After dissecting the contents of the sinus open
the pelvis and study the papilla. Pass a bristle from the pelvis into
the ureter.
3. Make a median section of the kidney parallel to its ventral
surface and study structure (Fig. 109).
4. Trace the ureter to the bladder, being careful not to injure
the vas deferens. Study the bladder and its ligaments, structure of
its wall, etc. Pass a bristle through the ureter into it and then open
it to see the openings of the ureter. Trace the neck of the bladder to
the pubis.
B. Male Genital Organs.
1. Study the external genital organs (p. 257).
2. Carefully remove the integument about the anus and expose
the external sphincter ani muscle (p. 268, and Fig. 113, i). Trace
it to its origin. Remove the integument of the scrotum and dissect
the intercolumnar fascia, the levator scroti muscle (p. 271, and Fig.
113, j), the tunica vaginalis communis, and propria (p. 258).
3. The testis; the epididymis with great care, especially in
uncovering the vasa efferentia; the vas deferens and spermatic cord
(p. 259, and Figs. 110 and 111).
4. Follow the cord to the external inguinal ring (p. 259) and
expose the ring. Dissect off in order from the inguinal canal: (a)
The external oblique muscle; (b) the internal oblique muscle; (c)
the transversus. In this way the cord is followed with its tunica
communis and tunica propria to the internal inguinal ring.
5. Trace the vas deferens (p. 260, and Fig. 111) within the
abdominal cavity as far as it can be seen dorsad of the neck of the
bladder.
[454]
6. The penis (p. 262, and Figs. 111 and 113). Cut the skin
along the dorsum of the penis and reflect. Dissect the ligamentum
suspensorium penis. Then introduce a bristle into the urethra and
remove the integument from the whole penis and identify the corpora
cavernosa and corpus spongiosum and the urethra.
7. Remove the fat at the side of the rectum and find the levator
ani muscle (p. 269, and Fig. 162, 11). Transect and reflect it.
The internal sphincter ani (p. 269, and Fig. 113, h). Find the
external opening of the anal gland (p. 239); slit and examine it.
8. Dissect the ischiocavernosus (p. 269) and bulbocavernosus
(p. 271) muscles (Fig. 113), and find the bulbus of the corpus cavernosum
beneath the former. Cut it free from the ischial ramus.
9. Clean the muscles from the pelvis on the side from which the
crus penis is removed, and remove with bone forceps the body of the
pubis and the ramus of the ischium. Then trace the neck of the
bladder to its junction with the vasa deferentia, and the urethra from
that point to the external opening (Fig. 111). Study the compressor
urethræ muscle (p. 271, and Fig. 113, n). Find Cowper’s glands
(p. 261) and the prostate gland (Fig. 113). Dissect the other muscles
of the urogenital organs (pp. 268 to 273, and Fig. 113).
10. Slit the urethra on one side throughout its length and find
the veru montanum (p. 261); the openings of the vasa deferentia
(p. 261), by bristles passed into the urethra from them; the openings
of the prostate gland and the openings of Cowper’s glands.
11. Study the structure of the penis (p. 262) by making a cross-section,
and that of the glans by making a longitudinal section.
Note, if possible, the os penis.
C. Female Urogenital Organs
(p. 263, and Fig. 112). The kidney
and its ducts and the bladder are like those of the male (p. 255),
except the neck of the bladder, which will be seen in dissecting the
uterus and vagina.
a. Without cutting anything examine—
1. The ovaries (p. 264); the ligaments of the ovary (p. 264, and
Fig. 112).
2. The uterine tube. Its ostium (p. 264).
3. The body of the uterus (p. 266) as far as exposed.
4. The cornu of the uterus.
5. The broad and round ligaments of the uterus (p. 266).
6. Slit open one Fallopian tube, cornu, and body of the uterus
as far as the junction of the divisions of the body of the uterus.
b. Remove the ventral wall of the pelvis on one side only and
find the vagina and urogenital sinus (p. 267). The levator ani
muscle (p. 269, and Fig. 162, 11) must be cut and reflected.
1. Note the constrictor vestibuli, caudovaginalis, and urethralis
muscles (p. 272, and Fig. 114).
2. Introduce a probe from the uterus into the vagina and feel
with the finger for the hard cervix uteri (p. 266). Then open the
vagina on the side, but do not cut the os uteri (p. 266).
[455]
3. Examine the cervix uteri and os uteri (p. 266).
4. Introduce a probe through the neck of the bladder and note
its emergence into the urogenital sinus.
5. Find the end of the clitoris (p. 267), and its prepuce, and then
dissect the integument carefully from about the external opening of
the urogenital sinus and note an external sphincter of the sinus
(M. levator vulvæ) (p. 272, and Fig. 114), equivalent to the levator
scroti in the male and continuous with the external sphincter ani.
6. Find (if possible) the corpora cavernosa of the clitoris
(p. 267) and the ischiocavernosi muscles (p. 269).
DISSECTION OF THE CIRCULATORY SYSTEM.
Use the heart from the specimen dissected for the muscles, or
from the specimen on which the viscera were studied.
1. Study the outside (p. 275, and
Figs. 115 and 116), and learn
to recognize all parts. Find the pulmonary veins (p. 275, and Fig.
116, g, h, i) and cut them, thus separating the heart from the lungs.
2. The Pericardium (p. 279). In a specimen the thoracic contents
of which have not been injured, dissect the mediastinal septum
from the pericardial sac and remove fat about the great blood-vessels
so as to expose them all fully. Study their relations. Prick and
inflate the pericardium. Slit it lengthwise over the ventricles and
reflect it so as to expose its contents. Study the attachment of
serous and fibrous layers to the heart and their relation to one
another.
3. In dissecting the heart follow the course of the blood, studying
each cavity with the aid of the descriptions (pp. 275-279) as you
proceed. Dissect as follows:
a. Remove the dorsal wall of the right auricle (p. 275) and of its
appendage except that part of it to which the venæ cavæ are
attached.
b. Introduce the probe from the right auricle into the right ventricle,
and feel with the probe the line along which the ventricular
wall joins the septum. Cut along this line so as to turn back the
ventricular wall as a flap, which remains attached at the base of the
ventricle.
c. Introduce a probe through the conus arteriosus into the
pulmonary artery and cut along the probe.
d. Remove the dorsal wall of the left auricle and its appendage,
but do not remove that part to which the pulmonary veins are
attached.
e. Make a longitudinal incision beginning at the apex and divaricate
the lips of the cut as you pass toward the base, thus avoiding
injury to the lateral flap of the bicuspid valve. Without injury to
the flap or the columnæ carneæ, remove the heart-wall at the sides of
this incision near the base of the ventricle, as much as necessary to
expose the cavity.
[456]
f. Pass a probe into the aorta. Introduce scissors behind the
septal flap of the bicuspid valve and slit the aorta without injury to
the bicuspid valve.
II. Dissection of the Blood-vessels.
Preparation and Injection.—A new specimen must be prepared for
the dissection of the blood-vessels. The same specimen may be
used, if necessary, for the dissection of the peripheral nervous
system,—one side being used for the blood-vessels, the other for the
nerves,—but it is much better to use separate specimens for the two
systems.
Inject the femoral artery of the specimen with either five per
cent. formalin, or with the mixture of five per cent. formalin and
glycerine, exactly as for the muscles. Then, without removing the
canula or waiting, inject the artery with red starch. This is prepared
as follows:
Mix together equal volumes of vermilion (or red lead), glycerine,
and five per cent. formalin. Grind these together in a mortar, so as
to destroy the lumps; strain the resulting mixture through fine
muslin. This color mixture can be preserved in a closed bottle till
it is to be used. Then mix together one volume of dry starch, one
and one-fourth volumes of five per cent. formalin, and one-fourth
volume of the color mixture. See that no lumps are present.
Inject the above red starch mass into the femoral artery. It will
drive into the veins the formalin already injected, and the arteries
will be filled with the red starch.
The veins need not be injected, as they will be found to be filled
with the formalin mixture, colored by the blood, so that they can be
traced without great difficulty.
Dissection of Blood-vessels.—In general the arteries and veins will
be traced together. It is an excellent plan to make a sketch of each
vessel as it is dissected, showing its origin, branches, and name.
This will be a great aid to the memory and will be of much assistance
in reviewing the vessels. The sketches can later be combined
into a well-ordered drawing. The description of each vessel must
of course be studied as the vessel is dissected.
In tracing blood-vessels do not grasp them between the forceps-blades,
but handle them by taking hold with the forceps of the connective-tissue
coats of the vessels. Do not use the scalpel, but pull
away the connective tissue and fat with fine forceps, using two pairs
of forceps or one forceps and the tracer.
Variations are especially common in the blood-vessels, and the
student must not be surprised to find considerable deviations from
the conditions described and figured in the text. These variations
usually present nothing new in principle and are easily understood
by comparing them with the structures described.
Directions for Dissecting the Blood-vessels.—Make a median longitudinal[457]
incision through the skin from the symphysis of the jaw to
the caudal end of the xiphoid process. About three or four centimeters
caudad of the cranial end of the sternum make an incision at
nearly right angles to this, passing from the first incision on the
ventral side of the left arm about to the elbow. Reflect the flaps of
skin, so as to uncover the left side of the thorax and the under surface
of the arm, exposing the pectoral muscles. Isolate and transect
the pectoral muscles one at a time, cutting each near its thoracic
attachment. (The muscles (p. 145) should be reviewed at the same
time.) In this way the nerves and blood-vessels of the axilla are
exposed (Fig. 122, p. 295).
Find the axillary artery and vein (Fig. 122, f and g) emerging
from the thorax just craniad of the first rib, along with the nerves of
the brachial plexus. Remove connective tissue, etc., so that the
vessels and nerves are well isolated as they pass out of the thorax.
Take great pains not to puncture the vessels, particularly the veins.
Then remove the left side of the thorax by cutting through the
first rib near its sternal end and then near its dorsal end, without
injury to the vessels and nerves, cut the other ribs in the same way,
and take out the thoracic wall.
Now find with tracer and forceps the great blood-vessels leaving
the cranial end of the heart (see Fig. 118). Take the greatest pains
not to injure them. Find the aorta and aortic arch (p. 281); the
left subclavian artery (p. 283) (continuous with the axillary); the
innominate artery (p. 282), and the beginnings of its three branches
(see Fig. 115). Find also the superior vena cava, the innominate
veins, and the subclavian vein, continuous with the axillary vein.
I. Study the smaller branches of the thoracic aorta (p. 283, and
Fig. 118)—the intercostals, the bronchial and œsophageal arteries,
and the first pair of lumbar arteries. (The coronary arteries will be
examined later.)
II. Dissect the subclavian and its branches (p. 290) as follows:
1. The internal mammary (p. 292). Follow it onto the ventral
wall of the abdomen. Follow the vein at the same time (p. 318).
2. The vertebral artery (p. 291). Find its beginning, but do
not trace it at present.
3. The costocervical axis (p. 292). Find its beginning, and
trace the superior intercostal branch some distance The other
branches are not to be followed at present.
4. The thyrocervical axis (p. 293). Find its beginning, but do
not trace it at present.
5. The axillary artery (continuation of the subclavian) (p. 294).
Follow its branches, tracing at the same time the axillary vein
(p. 318). (Consult Fig. 122.) In tracing the blood-vessels, separate
the muscles, but do not cut them except where absolutely necessary.
(The muscles should be reviewed as the vessels are traced.)
The following notes may be of assistance in following the different
branches:
[458]
(a) The anterior thoracic was probably cut in dissecting the
pectoral muscles; it may be found, but its distal end is probably
cut off.
(b) The long thoracic is easily followed.
(c) The subscapular. Follow the main artery before dissecting
its branches. Where the subscapular disappears between the long
head of the triceps, the latissimus dorsi, and the scapula, it may be
traced and found again as follows: Remove the skin from the outer
side of the shoulder,—taking great pains to remove only the skin and
not to injure the vena cephalica (p. 319), a large vein that lies just
beneath the skin on the lateral surface of the shoulder, coming from
the elbow. The branches of the subscapular will be found appearing
on the lateral surface of the arm in the angle between the spinotrapezius,
the long head of the triceps, and the infraspinatus. The
distal branches may then be followed.
(d) The posterior circumflex (p. 296) may be traced distad in a
similar manner, by seeking it beneath the caudal border of the
spinotrapezius. (Do not injure the vena cephalica.)
(e) The other branches of the brachial artery and vein present no
difficulty till we come to the collateralis radialis superior (Fig. 122, x).
This must be traced with great care, along with the vena mediana
cubiti (p. 319, and Fig. 122, y). Remove the skin from the extensor
side of the forearm, taking great pains not to remove anything more
than the skin. The artery and vein lie beneath the skin and should
be traced to the hand (see Fig. 130).
Along with the collateral radial artery trace the vena cephalica
(p. 319, and Fig. 130, a and c). Follow it across the shoulder,
noting the branch to the posterior circumflex vein.
The remainder of the blood-vessels of the arm present no special
difficulty (see Figs. 123 and 124).
Make a diagram of the subclavian artery, as far as dissected.
III. Remove the skin from the sides of the neck, exposing the
sternomastoid muscles and the external jugular veins crossing them
(see Fig. 131). Clean the surface of the sternomastoid muscles,
without injuring the vein; separate the two muscles caudad, and cut
each close to the attachment to the sternum. Find the sternohyoid
and sternothyroid muscles, and cut them close to their attachment
to the first rib. Uncover the right side of the thorax in the same
way as the left, cut the ribs without injury to the nerves and vessels
of the right axilla, cut the internal mammary artery and vein (after
tying the latter), and thus remove the sternum with nearly the entire
thoracic wall.
The blood-vessels of the thorax may now be more completely
exposed. (If the nerves are to be dissected on the same specimen,
find the phrenic, vagus, and sympathetic nerves (Fig. 157), and take
the greatest pains not to injure them.)
1. The coronary arteries and the veins of the heart (pp. 281 and
316).
[459]
2. The superior vena cava (p. 316). Find its branches. Trace
the azygos as far back as the diaphragm without dissecting it at all.
Find the division of the innominate (p. 318) into subclavian and
external jugular.
Trace next—
3. The external jugular (p. 319). Remove the skin from the
side of the face and trace its branches (Fig. 131). The internal
jugular, vena facialis profunda, the submentalis, and the deep
terminal branches of the posterior facial cannot be followed at this
time; veins shown on Fig. 131 should all be found, however.
4. Trace the thyrocervical axis and its branches (p. 293). Add
them to your diagram of the subclavian.
5. Follow the common carotid artery (p. 283) and internal
jugular vein (p. 320). Find the division of the common carotid into
its terminal branches and then dissect its lateral branches and those
of the internal jugular (see Fig. 119).
6. The external carotid (p. 285, and Fig. 119). Follow its
branches with the exception of the internal maxillary.
7. The internal maxillary (p. 287). Find its inferior alveolar
branch first and follow it by cutting away with bone-forceps the ventral
border of the lower jaw. To follow its other branches and those
of the carotid plexus, remove the zygomatic arch, cut the temporal,
masseter, and pterygoid muscles, and cut the mandible behind the
incisor teeth and remove it. The branches which pass into the
skull are not to be followed at present. The posterior facial vein
(p. 323), the vena facialis profunda (p. 323), and the submental vein
(p. 323) may be followed at the same time.
8. The internal carotid (p. 285). Follow it to the point where
it enters the cranium.
9. Trace the other branches of the costocervical axis (p. 292).
To do this, cut the arteries and nerves of the axilla on the side on
which they have been dissected, allowing the arm to fall backward.
Then trace the branches of the costocervical axis with tracer, scalpel,
and bone-forceps, taking care not to injure the vertebral artery.
Add these branches to your diagram of the subclavian.
10. The vertebral artery (p. 291). Trace it to the foramen
transversarium of the sixth cervical vertebra. Then with bone-forceps
follow it to the atlas and into the atlantal foramen. Add this to
your diagram of the subclavian.
11. The basilar artery (p. 291) and the other arteries of the
brain (p. 289) are best studied on a preparation, similar to that
shown in Fig. 121. To obtain such a preparation it is only necessary
to remove the brain (for directions, see p. 462) of a specimen
in which the arteries have been injected.
(12. Veins of the brain and dura mater (p. 324). These can be
worked out only with much difficulty, except on specimens injected
with gelatine. The skull must be chipped away and the veins
followed without destroying them.)
[460]
13. Trace the pulmonary veins (p. 315) (filled with red injection)
and the pulmonary artery (p. 280).
IV. Vessels in the abdominal cavity.
1. Open the abdominal cavity; find the superior mesenteric vein
(p. 326, and Fig. 132) in the duodenal mesentery near the border of
the pancreas. Inject this in both directions with white starch and
then dissect the portal vein and its tributaries without injuring any
of the structures in the abdomen (p. 326, and Fig. 132).
2. Follow the inferior vena cava (p. 325) from the heart to the
diaphragm and then follow it to its tributaries in the abdominal
cavity.
3. Dissect the branches of the abdominal aorta (p. 301) and of
the inferior vena cava (Fig. 126). Make diagrams of the vessels dissected
and review as far as necessary the viscera concerned.
V. The external iliac and its branches (vessels of the hind limbs)
(pp. 309 and 329, and Figs. 127, 128, and
163).
Follow the branches of the external iliac arteries and the corresponding
veins in the same manner as the vessels of the arm were
traced, cutting the muscles only so far as absolutely necessary.
Make diagrams of the vessels dissected.
Make a diagram (a) of the arterial system as a whole; (b) of the
venous system as a whole.
THE LYMPHATIC SYSTEM (p. 330).
It will hardly be found practicable to have each student make a
dissection of the lymphatic system, and such parts of it as are to be
studied may best be shown on a specimen prepared for demonstration
purposes.
The thoracic duct and the receptaculum chyli may be demonstrated
by the following well-known method: A lean cat is fed with
milk about two hours before killing it. An egg may be beaten up
with the milk to advantage. Kill the cat with chloroform, and inject
the arteries with colored starch through the femoral, in the usual
way. The thoracic duct, the receptaculum chyli, and the lymphatics
leading to the receptaculum chyli will be colored white by the milk,
and can therefore be easily followed. For this purpose the abdomen
should be opened, and the left side of the thorax removed, as in the
dissection of the blood-vessels. The thoracic duct will be found at
the left side of the aorta and may then be traced in both directions.
For a more complete study of the lymphatics they should be
injected. This is done as follows: Make a glass canula with a small
point, and leave the point sharp. Connect this to the syringe by
means of a rubber tube. Use a saturated solution of soluble Prussian
blue as injecting fluid. Employ a freshly killed animal.
For injecting the lymphatics of the limbs, make with some pointed
instrument, as the tracer, a small hole in one of the pads on the sole
of the foot. Introduce the point of the canula into this opening and[461]
inject the fluid. This will pass into the spaces in the connective
tissue of the pad, which will swell up, and the colored fluid will pass
from the connective-tissue spaces into the lymphatics. Pressure must
be maintained with the syringe for a considerable time,—fifteen
minutes to a half-hour for a good injection of the main trunks of the
lymphatics of the limbs. The movement of the fluid should be
facilitated by pressing and manipulating the limb at the same time
with the hand,—in such a way as will tend to drive the fluid
proximad.
The lymphatics of the head may be injected in a similar manner,
the canula being introduced into the upper and lower lip, or into
the bare surface at the end of the nose.
The internal lymphatic vessels may be injected by injecting the
lymphatic glands with which they are connected. This may conveniently
be done as follows: Draw out to a fine point the tip of an
ordinary pipette or medicine-dropper. The point should be fine,
but should taper rapidly in a conical fashion, so that when the point
is inserted the part of the glass tube behind it will close up the
opening.
Fill the pipette with soluble Prussian blue; insert the point into
the gland, and inject the fluid slowly. The lymphatic vessels passing
from the glands will be filled. By injecting thus the large lymphatic
gland (“pancreas Aselli”) in the mesentery, the abdominal lymphatics,
the receptaculum chyli, and the thoracic duct may be
injected.
By using thin gelatine colored with Prussian blue as an injecting
fluid permanent preparations may be obtained; of course the
process of injection is then less simple, and should be looked up in
some manual of methods.
NERVOUS SYSTEM.
I. The Spinal Cord (p. 335).
Use the specimen on which the muscles were dissected. (Or if
the peripheral nerves are not to be dissected on the specimen used
for the blood-vessels, that may be employed.)
Make a longitudinal dorsal median incision of the skin, between
the back of the head and root of the tail. Reflect the skin for one or
two inches on each side of the incision and cut away the muscles
covering the neural arches of the vertebræ from the third cervical to
the seventh or eighth thoracic inclusive.
Remove with bone-forceps the neural arch of one of the last
cervical vertebræ and find the spinal nerve emerging from the intervertebral
foramen. Isolate the nerve for a short distance, then
proceed craniad, removing the neural arches on one side and isolating
the nerves until the third has been uncovered. The ganglion of
the second nerve should be sought among the muscles on the dorsal
surface between the atlas and axis, and after it has been isolated the[462]
arch of the axis may be removed. (The nerve may be found beneath
the clavotrapezius and traced to the ganglion.)
The ganglion of the second nerve should be isolated in or near
the atlantal foramen, the muscles to which it passes turned aside, and
the arch of the atlas removed. Having thus uncovered the first two
or more spinal ganglia, proceed caudad, removing the vertebral arches,
until the whole cord and its nerves are exposed. Then—
1. Study the cord, enlargements, filum terminale, etc. (p. 334,
and Figs. 133 and 136).
2. Slit open and reflect the dura mater (p. 337) for an inch or two.
3. Demonstrate the arachnoid by pulling it off with forceps.
4. Reflect the pia mater in the same way as the dura mater.
5. Study the fissures and grooves of the cord.
6. Cut across the cord with fine scissors at the point where it is
freed from its membranes and examine the section. Note the
arrangement of gray and white matter and the fissures and grooves,
particularly the anterior or ventral. Demonstrate the central canal
with the blowpipe.
7. Study the origin of the spinal nerves (p. 337). Count them.
Direction of exit? Carefully clean one in the thoracic region from
dura mater and connective tissue, with fine scissors, and study dorsal
and ventral roots and ganglion (see Fig. 135). Then follow it out
and find its dorsal ramus and ventral ramus and the communicating
branch of the latter with the sympathetic system. Do not trace the
peripheral branches of the nerve at present.
II. The Brain (p. 339).
The brain will usually be found to be in an entirely satisfactory
condition for study in any specimen injected with five per cent.
formalin or the glycerine and formalin mixture. The brain is a little
swollen, but all parts are well preserved, and the white and gray
matter are clearly marked off from each other. Either the specimen
used for the muscles or that employed for the blood-vessels may
therefore be used,—or if the brain was removed from the specimen
employed for the viscera, that will be satisfactory.
The following directions for removing the brain are designed for
specimens preserved as above. For removing the fresh brain the
process is essentially similar, but as the brain is then very soft, care
should be taken not to tear it. The fresh brain should be preserved
in the alcohol-formalin mixture given below, and should be allowed
to rest only on some soft substance, as absorbent cotton.
Remove the head from the body by cutting through the neck a
little craniad of the first rib if this has not already been done.
Remove all skin, muscles, and other soft parts from the head and
cervical vertebræ, as far as possible. Remove the structures in the
orbit by cutting through the zygomatic arch at each end, and
removing it. The lower jaw should also be removed, if this has not[463]
already been done. (If a fresh specimen is used, and the head is to
be employed for other purposes, the brain can be removed without
separating the head from the body, and without taking away the
lower jaw and other structures on the ventral surface of the skull.)
Have at hand dissecting-instruments and a dish containing
alcohol and formalin in the following proportions (Parker and
Floyd’s mixture):
| 95 |
per cent. alcohol |
6 parts |
| 2 |
per cent. formalin |
2 parts |
In the bottom of the dish should be placed a little absorbent
cotton, to support the brain.
In removing the brain have at hand entire and dissected skulls
and note the relations of parts on these as far as necessary before
cutting the specimen.
With bone-forceps make a small opening in the parietal bone so
as to expose the dura mater, but do not cut through the dura mater.
With some blunt instrument free the dura mater from the bone about
the opening, and continue to cut away the bone until the dorsal and
lateral faces of the cerebrum are fully exposed craniad of the
tentorium. The olfactory bulbs (Fig. 137, I) should be exposed
carefully and as fully as possible. Cut away the dorsal arch of the
atlas and carefully insert the forceps in the foramen magnum and,
working as before, remove the squamous portion of the occipital and
the parietal bones as far as the tentorium and as far ventrad as
possible. Leave the dura mater intact if possible. Free the surface
of the tentorium from the dura mater, carefully separate slightly the
cerebellum and cerebrum; insert the bone-forceps (not too far) with
the blades inclined from without ventromediad, and cut the tentorium
on each side. Remove it slowly, cutting adhesions to the dura
mater. That part of the dura mater which dips between the cerebral
hemispheres is the falx cerebri. Cut the dura mater along both sides
of the falx cerebri and remove it by turning it down at the sides and
cutting it at the level of the cut edge of the bone. Remove it also
from the cerebellum and notice how it dips down on both sides of
the tentorium and in close contact with it. Cut the falx at the
cranial end between the olfactory bulbs and cut the tentorial dura
(cut its adhesions, but do not remove with it the pineal body). The
falx and tentorial dura may then be removed.
Allow the head to hang sideways over the dish of alcohol-formalin
in such a way that the brain will tend to fall out of the cranium.
Free the olfactory bulbs from the bone. Then begin at the caudal
end and tilt the brain out with the handle of a scalpel. In doing
this note carefully and cut the cranial nerves. They should be left
with central ends as long as possible, and those on the side which is
uppermost should be cut first. In doing this refer to the foramina
in the base of the skull and to Fig. 138. Take especial pains also
not to break off the hypophysis, which is lodged in the sella turcica.
[464]
The brain falls out and rests with its dorsal surface on the cotton.
Now remove the remainder of the dura mater, carefully cutting all
adhesions to nerves. Remove also the pia mater, as far as that can be
done without pulling off at the same time parts of the brain-substance.
Preserve the brain in the alcohol-formalin mixture.
Study of the Brain.—In the study of the brain demonstration
specimens are to be used as much as or more than your own specimen.
See everything on a demonstration preparation before attempting to
expose it in your own specimen.
I. Examine the brain of a shark or of a frog. Cranial nerves may
be neglected, but the divisions of the brain should be recognized in
dorsal and ventral views and in longitudinal sections, and sketched.
II. Read the general description of the cat’s brain (pp. 339-343),
using your own specimen and a longitudinal section. Cut nothing
on your own specimen except when especially directed to do so.
Study the cavities on a preparation. Compare the diagrams (Figs.
139 and 140) and the figures of the brain.
III. Study the individual parts as follows. To avoid errors make
constant reference to preparations and figures.
1. The medulla (p. 344 and Figs.
138 and 141). Use your own
specimen and a preparation and dissect out carefully the cranial
nerves on your own specimen.
2. The cerebellum (p. 347). Study it entire, then to expose the
fourth ventricle (p. 349) slice away with a very sharp scalpel one-half
of the cerebellum by making a median longitudinal incision and then
horizontal incisions.
3. The pons (p. 347).
4. The mesencephalon (p. 351, and Figs. 141 and 142). Study
it first in a preparation. Then study the floor on your own specimen;
origin of third nerves.
5. The diencephalon (Figs. 141 and 142). Study the roof and
thalami and the pineal body on a preparation and on a longitudinal
section; the floor on your specimen.
6. The telencephalon (p. 357). (Note that only one side of this
is to be dissected.)
a. Study it externally; sulci and gyri (Figs. 145 and 146).
b. Examine a preparation showing the corpus callosum (Fig.
147). Then slice away with a very sharp scalpel the top of one
hemisphere nearly to the corpus callosum (see the preparation).
Expose the corpus callosum on this side to its cranial and caudal
borders, by tearing away the brain-substance at its side and above it.
c. Raise the corpus callosum at the side and remove it, thus
exposing the lateral ventricle in which note the septum pellucidum
and fornix, the corpus striatum, and choroid plexus of the lateral
ventricle (Fig. 148). (These are to be exposed on one side only,
the other being left intact.)
d. Expose the anterior and inferior horns of the ventricle and
find the hippocampus, the fimbria, caudal part of the fornix, the[465]
foramen of Monroe, the anterior commissure. See all these also on
a preparation (Fig. 148).
e. Remove the occipital and parietal portions of the cerebrum,
on the side already dissected, so as to expose the roof of the third
ventricle and the midbrain in your specimen, and note the pineal
body, choroid plexus of third ventricle, and structures on the roof of
the midbrain (Fig. 141).
f. Remove the choroid plexus or roof of the third ventricle and
study again the thalami (Fig. 141).
g. Make a longitudinal section of the brain, in the following
manner: Use a very sharp large scalpel, or a razor. Have this wet
with the alcohol mixture at the time of using. Place the brain
ventral surface down on a sheet of cork or a block of soft wood, the
long axis of the brain coinciding with the direction of grain of the
wood. Holding the brain firmly with one hand, place the wet knife
between the hemispheres with its edge resting on the corpus callosum.
See that it is in the median plane and parallel with the long axis of
the brain. See also that it is not inclined to one side or the other,
so that it will make on cutting a median section throughout. The
point of the knife should just reach the cork or wood between the
olfactory bulbs. Now draw the knife caudad, keeping its point
against the cork: the brain will thus be divided.
If the section is not exactly median, observe the amount of
divergence by placing the two halves together and finding the
median ventral line. Then on the half that has too much slice away
thin shavings until the cavities are exposed, showing the section to be
median. Compare with a demonstration section or Fig. 143. Draw
the section and compare with a section of shark’s brain (see Fig. 143).
h. Study a series of transverse sections, identifying parts.
Observe especially in these sections the fornix, corpus callosum, and
ventricles, and the distribution of white and gray matter (see Figs.
149-153).
III. PERIPHERAL NERVOUS SYSTEM.
(There are some advantages in dissecting the eye with its muscles
before dissecting the nerves, as a knowledge of the eye-muscles is
presupposed for dissecting some of the cranial nerves. For directions
on the eye, see p. 469.)
A new specimen should be used, if possible, for the peripheral
nervous system, though that used for the blood-vessels can be
employed, at considerable disadvantage.
Prepare as for the blood-vessels. The arteries should be injected
with red starch, to aid in tracing the nerves.
1. The Cranial Nerves (p. 369)
and Sympathetic System (p. 404).
1. Reflect the skin covering the sternomastoid muscle, and make
a longitudinal incision of the muscle so as to expose the carotid
artery. Lying along the artery find the combined trunk of the[466]
sympathetic and vagus nerves. Follow the vagus (p. 378) first
craniad; transect the muscles as necessity arises, and find its ganglion
nodosum and at the same time locate the superior cervical ganglion
of the sympathetic nerve (p. 404, and Fig. 156). Then find the
hypoglossal nerve (Fig. 156, b), passing outside of the carotid artery
to the tongue, and the accessory (Fig. 156, c), passing to the trapezius.
Cut and reflect the digastric muscle and find the small
glossopharyngeal nerve (Fig. 156, a), passing to the surface of the
bulla and then beneath the carotid artery.
2. Follow the vagus (p. 378) caudad to its termination. To do
this it is necessary to remove one side of the thorax, as in dissecting
the blood-vessels. Do not injure the nerves of the axilla, nor the
phrenic or sympathetic nerves. For the vagus in the thorax, compare
Fig. 157. Find the branches of the nerve; in dissecting them,
pull on them to make them tense. They are then more easily
visible. To dissect the abdominal portion of the vagus, open the
abdominal cavity, and compare Fig. 164 (p. 407).
3. Dissect the sympathetic (p. 404), following it and its branches
to the pelvic region (Figs. 156, 157, and 164).
4. The hypoglossal (p. 383, and Fig. 156, b).
5. The glossopharyngeal (p. 378, and Fig. 156, a).
6. The accessory nerve (p. 382, and
Fig. 156, c; Fig. 158, 1).
Cut away a portion of the tympanic bulla and the base of the
skull, sufficient to follow these nerves in the jugular foramen, to the
brain.
7. Locate the stylomastoid foramen and pick away overlying
tissue until the facial nerve is found emerging and then follow its
branches to their distribution (p. 375, and Fig. 155).
8. Expose the ventral surface of the pterygoid muscles just
mediad of the angle of the jaw. Divide and reflect them, and the
mandibular division of the fifth nerve (p. 373, and Fig. 154) will be
found dorsad of them and of the internal maxillary artery. The
chorda tympani (p. 375) passes ventrad of the artery to join the
lingual. Follow out (1) the lingual branch (p. 375) (with the chorda
tympani), and (2) the inferior alveolar (p. 375) by cutting away the
ventral border of the mandible. Then cut the mandible near the
canine tooth, and pull it to one side, and follow out the muscular
branches of the mandibular nerve.
9. Remove the mandible and find the maxillary nerve (p. 371)
emerging from the foramen rotundum. Follow its branches and find
the sphenopalatine ganglion (p. 372).
10. Remove the zygoma so as to expose the whole ventral aspect
of the orbit. Carefully pick away the fat in the orbit without injuring
any nerves, so as to expose the four recti muscles and the inferior
oblique (see p. 411, and Fig. 166). Find the abducens nerve
(p. 375, and Fig. 154), entering the dorsal edge of the lateral rectus,
and follow it back. Look on the inner surface of the inferior rectus
for the branch of the third nerve (p. 369) which supplies it. Find[467]
the branch of this nerve which runs to the inferior oblique muscle,
and on it the ciliary ganglion; find the branches to the ciliary ganglion
from the ophthalmic nerve and follow them (p. 371). Follow
also the short ciliary nerves (p. 370) to the eyeball.
11. Trace the third nerve (p. 369) to its foramen of exit and find
its branches. Where it passes between the superior and lateral recti,
find the ophthalmic nerve (p. 370) by its side and trace its branches.
12. Find the fourth nerve (p. 370), passing outside of the lateral
rectus at its origin and entering the superior oblique.
13. Follow the third, fourth, fifth, and sixth nerves into the skull
by chipping away the bone and removing the dura. Note the semilunar
or Gasserian ganglion (p. 370, and Fig. 138, k) and the origin
of the fifth nerves from it, and the relation of the ventral root of the
fifth nerve to the mandibular nerve.
2. Spinal Nerves.
The spinal nerves may be dissected on the same side used for
dissecting the cranial nerves. (If an undissected specimen is used,
remove the skin from the side of the neck, and cut the sternomastoid,
sternohyoid, and sternothyroid muscles, as directed for the vagus and
sympathetic.)
Cervical Nerves (p. 383).—The ventral rami of the cervical nerves
are to be sought as they pass out between the bundles of the
scalenus, or between the scalenus and longus capitis, in the neck.
This region has already been uncovered in dissecting the vagus and
sympathetic (Fig. 156). Dissect first the second cervical (p. 385).
Find its ventral ramus as it emerges between the levator scapulæ
ventralis and cleidomastoid (Fig. 158, 2), and follow its branches,—the
auricularis magnus (5) and cutaneus colli (6). Find its dorsal
ramus, the great occipital nerve (p. 384), by reflecting the clavotrapezius
muscle; the nerve will be found emerging from the underlying
muscles close to the craniomedial angle of the clavotrapezius,
near its origin. Trace the nerve in both directions.
The ventral ramus of the first cervical (p. 385) will be found
emerging from beneath the wing of the atlas, a little distance craniad of
the second (Fig. 156, j). Trace it. To find its short dorsal ramus,
the suboccipital nerve (p. 384), it is necessary to dissect apart the
muscles on the dorsal side of the atlas till the nerve is found passing
from the atlantal foramen.
Dissect the third, fourth, and fifth nerves (p. 385, and Fig. 158).
Brachial Plexus (p. 386, and
Figs. 159 and 160).—The brachial
plexus has been partly uncovered in dissecting the vagus and sympathetic.
(If a new specimen or the opposite side is used, reflect the
skin from the ventral surface of the thorax and arm, and cut the
pectoral muscles, thus uncovering the vessels and nerves of the
axilla.) Reflect the skin from the ventral surface of the upper arm.
(Do not use scalpel, but tear the skin from the muscles. In this[468]
way the nerves will be seen passing to the skin, while if the scalpel
were used the nerves might be cut.)
Tie the axillary vein or its two branches in two places, and cut
the vein between the tied regions. Leave the arteries as guides for
dissection, but remove the veins. Now clean thoroughly the nerves
forming the brachial plexus as they pass from the thorax or neck.
Be careful not to injure any of the fine nerves or the interconnections
of the nerves in doing this. Find and distinguish clearly the fifth,
sixth, seventh, and eighth cervical nerves and the first thoracic, as
they emerge from the neck or thorax. (Compare Fig. 159 and Fig.
157, V-VIII and I′.)
Follow out the branches of the plexus, noting the origin and
distribution of each branch, in order to determine its name. To
follow the phrenic (p. 388, and Fig. 157, f), remove a portion of the
thoracic wall. In following the other branches of the plexus, pull
back the skin wherever an exposure is to be made, and separate the
muscles. The epitrochlearis may be cut near the elbow, and the
clavobrachial near the shoulder. In following the interosseous
branches of the median nerve the fifth head of the flexor profundus,
and the extensor brevis pollicis, may be cut. As a rule it will not
be necessary to cut other muscles.
Thoracic and Lumbar Nerves.—One or two of the thoracic nerves
(p. 393) should be dissected from the outside by finding the intercostal
nerve along the caudal border of one of the ribs and tracing it
in both directions. The nerve may be exposed by removing the
external muscles covering the rib, and cutting the external intercostal
muscles. The dorsal ramus should be traced after the ventral ramus
has been studied.
The first lumbar nerve (p. 395) should be dissected in the same
way.
The other lumbar and sacral nerves (pp. 395-400) are best dissected
from within.
The alimentary canal and its appendages should be removed from
the abdomen, leaving only five or six centimeters of the caudal end
of the rectum. The kidneys and urogenital organs may be left,
to be removed during dissection. (Compare Fig. 162.)
Turn one of the kidneys to the other side, and find the second
lumbar nerve (Fig. 162, a) appearing at the lateral border of the
iliopsoas muscle. Trace it to its origin; trace it also distad, following
both branches. It will be necessary to trace the nerves through
the abdominal wall, then find them from the outside, and follow
them to their distribution.
The third nerve (Fig. 163, b) may be found by dissecting apart
the fibre-bundles of the iliopsoas and psoas minor, and following in
the same way. The kidneys, ureters, vena cava, and aorta may be
removed as occasion arises.
The remainder of the lumbar nerves may be found in order, in a
similar manner. Follow the saphenous nerve and its branches ([469]
p. 397) by removing the integument from the medial side of the leg
(see Fig. 127, p. 310). To dissect the sacral nerves (p. 399) separate
the innominate bones at the pubic symphysis and divaricate them.
Find the nerves arising from the sacral plexus, by cutting the levator
ani muscle. After N. hemorrhoidalis inferior and N. pudendus have
been traced, the rectum and urogenital organs may be removed, taking
great care not to remove more than is necessary. To dissect the
great sciatic nerve (p. 400, and Fig. 163), separate the biceps and
caudofemoralis near their proximal ends and find the large nerve-trunk
(a). Then lift the biceps away from the nerve, cut that muscle
near its middle, and reflect it. The nerve may now be followed to
its terminal branches; during the process cut the muscles only when
absolutely necessary. The inferior gluteal nerve (i) will be found
on the dorsal surface of the great sciatic; by cutting the caudofemoralis
and gluteus maximus muscles near their insertions and turning
them back the distribution of the nerve may be followed. The
superior gluteal (j) will be found at the cranial margin of the pyriformis
by reflecting the gluteus medius in the same way; the tensor
fasciæ latæ may also be cut.
SENSE-ORGANS.
I. The Eye (p. 410).
Use any specimen on which one side of the head has been left
intact. Remove the head from the body by cutting through the neck
a little craniad of the first rib.
Study the eye externally. Observe the eyelids, the conjunctiva,
nictitating membrane with its cartilage, and the Harderian
gland (Fig. 165); the two openings of the lachrymal duct, the
Meibomian glands if possible.
Remove the zygomatic arch and expose the orbit and the structures
which it contains. Study the periorbita (p. 409), and find the
lachrymal gland (p. 410).
Study the muscles of the eyeball (p. 411, and Fig. 166). The
lateral rectus on the lateral surface will perhaps be first found, and
its tendon traced beneath the inferior oblique. Use great care not
to injure the levator palpebræ superioris.
After studying the muscles, find the optic nerve. Cut it and the
muscles, and remove the eyeball for farther study.
The Eyeball (p. 412, and Fig. 167).—For an examination of the
eye it is well to have a fresh specimen and one hardened in formalin
or alcohol. The hardened specimen is more essential, however. An
eye from one of the specimens used in dissection is usually satisfactory.
All accessory portions should be trimmed from the eyeball,
leaving only the spherical ball with a short stalk formed by the optic
nerve.
Observe such features of the eye as can be seen externally: the
optic nerves, sclerotic, cornea, iris, and pupil. This should be done[470]
on a fresh specimen, if one is at hand. The changes in size and form
of the pupil can be observed in the living cat by changing it from a
light to a dark place and vice versa.
Dissection.—Examine the internal structures on a preparation.
Then with fine scissors and forceps remove from the eye about one-fourth
of the wall, in the form of a quadrant having one point at the
optic nerve, the other at the centre of the cornea. The coats of the
eye can then be studied on the piece removed, while the other structures
will be visible within the eyeball. No special directions are
necessary for observation of the structures described, unless it be the
capsule of the lens and the zonula ciliaris. The capsule of the lens
may be demonstrated by tearing a bit of it off with fine forceps.
The zonula ciliaris is easily seen by divaricating the edges of the cut
made in removing the quadrant, so as to stretch the fine fibres of
which the zonula is formed.
II. The Ear (p. 415).
(The muscles of the external ear are of little practical importance,
and will doubtless usually be omitted.)
(1) The External Ear. Read the description of the external ear
(p. 415), verifying it by examination and comparison of an ear still
covered with integument, in the natural condition, and of a preparation
of the isolated cartilages of the ear (Fig. 168). The latter may
be obtained by dissecting the skin and muscles from a fresh ear.
(2) The scutiform cartilage (p. 418) should be observed in the
natural position, and as isolated.
(3) The muscles of the external ear (p. 418, and Fig. 169).
Those connecting the external ear with other parts of the head have
been studied in connection with the facial muscles. The remainder
will be studied on an external ear removed according to the directions
given on page 435.
Remove the integument from the convex surface of the auricle,
sufficiently to expose the entire extent of the muscles.
1. The rotator auris. Transect.
2. The adductor auris superior (Fig. 169, 1).
3. The adductor auris medius (2).
4. The transversus auriculæ (Fig. 63, i, p. 97).
5. The auricularis externus (Fig. 169, 10).
6. The helicis (Fig. 169, 3). To expose this it will be necessary
to remove the integument from along the cranial border of the inner
surface of the auricle.
7. The antitragicus (Fig. 169, 6). Remove the integument
farther if necessary.
8. The tragicus medialis (Fig. 169,
5, 5′).
9. The conchæus externus (Fig. 169, 9).
(4) Remove the tympanic bulla and petrous bone from the rest
of the skull, by the use of bone-forceps and scalpel. Trim away all[471]
soft tissue (including the cartilaginous auditory meatus), and all
other bony parts from these, but leave them uninjured.
(5) Find the Eustachian tube and study it (p. 423).
(6) Study specimens of the bones of the middle ear (p. 423, and
Figs. 171 and 172).
(7) The middle ear should be studied on a demonstration preparation,
then dissected as follows:
Remove with bone-forceps the medial side of the tympanic bulla
(the entotympanic). Note the two cavities within the bulla, with
the shelf separating them. Observe the fenestra cochleæ. Now
remove with the forceps the shelf, first breaking through the middle
part, then removing the rest with care. Remove part also of the
membranous lining of the cavity, till the inner surface of the tympanic
membrane, with the malleus crossing it, is visible. Observe
the tensor tympani muscle (p. 424) attached to the malleus by its
small tendon. Next, with bone-forceps, fracture the thick, bony
portion uniting the caudal end of the petrous with the caudal part
of the ring of bone surrounding the external auditory meatus. These
two parts may then be separated with the fingers, leaving the petrous
bone on one side, the meatus, tympanum, and malleus on the other
(Fig. 170). The head of the malleus may then be observed, with
the incus attached to it. Note also the stapes, in the fenestra vestibuli,
with the stapedius muscle (p. 424) attached to it.
(8) The internal ear (p. 424). Note the fenestra vestibuli and
fenestra cochleæ and the promontory (p. 34). Study the cochlea and
vestibule on a demonstration preparation and compare with Fig. 173.
Then remove the wall of the promontory and find the cochlea.
Open the vestibule and find as many of the openings of the semicircular
canals as possible. Study the semicircular canals on demonstration
preparations, consulting Fig. 173. The semicircular canals
may, if desired, be exposed on your own specimen, by cutting away
the surface of the bone in places indicated in the description (p.
426), and inserting fine bristles (those from the sensory hairs on the
face of the cat are excellent for this purpose).
The membranous labyrinth (Fig. 173) may be isolated by decalcifying
the petrous bone with ten per cent. nitric acid, then dissecting
out the labyrinth. This is an operation of considerable delicacy, but
at least one or two specimens for demonstration should, if possible,
be prepared thus and kept in the laboratory.
[472]
[473]
INDEX.
- Abdominal aorta, 301
- arteries, 301
- cavity, 217, 218
- muscles, 153
- dissection, 441
- veins, 326
- Abducens nerve, 347, 375
- Abductor auris brevis, M., 100, 435; longus, M.,
100, 435
- brevis pollicis, M., 184, 439
- caudæ (coccygis) externus, M., 137, 447; internus,
M., 137, 446
- cruris, M., 195
- digiti quinti, M., 185, 439; secundi, M., 185
- medius digiti quinti, M., 215
- Accelerator urinæ, M., 271
- Accessory nerve, 346, 382
- process, 6, 8
- Acetabular bone, 76, 78
- notch, 79
- Acetabulum, 76, 78, 87
- Achilles, tendon of, 204, 206
- Acromiodeltoid muscle, 157, 438
- Acromion process, 62, 64
- Acromiotrapezius, M., 116, 436
- Adductor auris inferior, M., 104, 435; medius, M.,
419, 470; superior, M., 419, 470
- digiti secundi, M., 185
- femoris, M., 198, 444
- longus, M., 199, 444
- medius digiti quinti, M., 215
- pollicis, M., 185, 439
- Aditus laryngis, 246
- Adrenolumbar arteries, 304
- veins, 326
- Alæ magnæ, 26, 27
- parvæ, 29
- Albuginea, 260
- Alcohol as preservative, 431
- Alcohol-formalin for brains, 463
- Alimentary canal, 221; dissection of, 449
- Alisphenoids, 25, 27
- Alveolar border, 39, 47
- nerve, inferior, 374, 375
- process, 39
- Alveoli, 48
- Ampulla (ear), 426
- of Vater, 236
- Anal glands or sacs, 239; constrictors of, 269
- Anapophysis, 6
- Anastomotica magna, A., 297
- Anconeus, M., 170, 438
- Anconeus internus, M., 170
- lateralis, M., 167
- longus, M., 168
- posterior, M., 168
- Angle of rib, 18
- Angular process, 19, 48
- vein, 321
- Ankle, bones of, 82
- ligaments of, 91
- Annularis, 72
- Annular ligaments, 75, 173, 213
- Ansæ, 386
- Anterior cerebral artery, 289
- circumflex artery, 296
- commissure, 357, 365
- facial vein, 321
- fossa, 59
- perforated substance, 362
- spinal artery, 291
- superior process, 78
- Sylvian gyrus, 359
- sulcus, 359
- thoracic artery, 294; nerves, 387
- tibial artery, 313
- transverse processes, 11
- Antibrachial fascia, 172
- Antibrachium, muscles of, 172
- Anticlinal vertebra, 6
- Antihelix, 416
- Antitragicus, M., 421, 470
- Antitragus, 417
- Anus, 239
- muscles of, 268
- Aorta, 275, 279, 281, 301
- abdominalis[474], 301
- Aortic plexus, 408
- semilunar valves, 279
- sinus, 279
- Apparatus digestorius, 221
- respiratorius, 243
- urogenitalis, 255
- Appendicular fossa, 35, 58
- lobe, 348
- Appendix, 429
- Aqueduct of Sylvius, 342, 343, 350,
351, 352
- Aqueductus cerebri, 342, 343, 350,
351, 352; Fallopii, 35
- Aqueous humor, 414
- Arachnoid, 337, 369
- Arbor vitæ, 349
- Arch, bicipital, 166
- supraorbital, 37, 51
- zygomatic, 47, 49, 52
- Area elliptica, 345
- ovalis, 345
- perforata posterior, 352
- Arm, arteries of, 294
- articulations of, 73
- bones of, 64
- muscles of, 156
- dissection of, 436
- nerves of, 388
- veins of, 318
- Arteria adrenolumbalis, 304
- alveolaris inferior, 288
- anastomotica magna, 297
- anonyma, 282
- articularis genu suprema, 311
- auricularis anterior, 287; posterior, 287
- axillaris, 294
- basilaris, 291, 292
- brachialis, 296
- bronchiales, 283
- carotis communis, 282, 283; externa,
285; interna, 285
- cerebelli anterior, 292; inferior posterior, 292
- cerebri anterior, 289; media, 289;
- posterior, 292
- cervicalis ascendens, 293; profunda, 293
- circumflexa femoris lateralis, 311; humeri anterior,
296; humeri posterior, 296; scapulæ, 294
- cœliaca, 301
- colica dextra, 304; media, 303; sinistra, 306
- collateralis radialis superior, 297; ulnaris, 297
- coronariæ, 275, 279, 281
- digitales dorsales, 297; plantares, 314
- dorsalis penis, 308
- epigastrica inferior, 309
- femoralis, 307, 310
- gastrica sinistra, 303
- gastroduodenalis, 302
- gastroepiploica dextra, 303
- genu posteriores, 313
- glutea inferior, 308; superior, 308
- hemorrhoidalis media, 308; superior, 306
- hepatica, 302
- hypogastrica, 307
- ileocolica, 304
- iliaca externa, 307, 309; interna, 307
- iliolumbalis, 306
- infraorbitalis, 290
- intercostales, 283
- intercostalis suprema, 293
- interosseæ, 298, 300
- lienalis, 303
- lingualis, 285, 286
- lumbales, 283, 306
- lumboabdominalis, 304
- mammaria interna, 292
- maxillaris externa, 285, 286; interna, 287
- mesenterica inferior, 306; superior, 303
- meningea media, 289
- metatarsea, 315
- occipitalis, 285
- œsophageæ, 283
- ophthalmica, 289
- palatina descendens, 290; minor, 290
- pancreaticoduodenalis inferior, 303; superior, 303
- phrenica, 304
- poplitea, 311, 312
- princeps pollicis et indicis, 300
- profunda brachii, 296; femoris, 309
- pulmonalis, 275, 278, 280
- pylorica, 302
- radialis, 297; recurrens, 298
- renalis, 304
- sacralis lateralis, 309; media, 301, 309
- saphena, 311
- spermatica externa, 309; interna, 304
- sphenopalatina, 290
- spinalis anterior, 291
- subclavia, 282, 283, 290
- subscapularis, 294
- superior profunda, 296
- suprascapularis, 293
- suralis, 313
- tarsea medialis, 315
- temporalis superficialis, 287
- thoracica anterior, 294; longa, 294
- thoracicodorsalis, 296
- thyreoidea ima, 283, 284; superior, 284
- tibialis anterior[475], 313; posterior,
313; recurrens, 314
- transversa colli, 293; scapulæ, 293
- ulnaris, 300; recurrens, 298
- umbilicalis, 307
- uterina, 308
- ventriculi dorsalis, 303
- vertebralis, 291
- Arteries (see Arteria), 280
- abdomen, 301
- arm, 294
- bladder, 307
- brain, 289, 291
- foot, 312, 314
- hand, 300
- kidneys, 304
- intestine, 304
- limbs, 294, 309
- liver, 302
- stomach, 303
- Artery (see Arteria)
- cystic, 302
- labial, 286
- mediastinal, 282
- ovarian, 306
- submental, 286
- superior laryngeal, 285
- Articular artery, superior, 311
- circumference, 67
- facet, 4
- processes, 6, 8
- Articulation, atlanto-occipital, 17
- intervertebral, 16
- Articulations of arm, 73
- hind limb, 86
- mandible, 61
- Arytenoid cartilages, 248
- Arytenoideus transversus, M., 250
- Astragalus, 82
- Atlantal foramen, 14
- Atlanto-occipital articulation, 17
- Atlas, 14
- Atrium dextrum, 275
- sinistrum, 279
- Attollens auris. M., 100
- Auditory bulla, 32
- meatus, external, 32, 33, 52,
55, 415, 416; internal, 35,
58
- nerve, 347, 377
- organ, 415
- Auricle (ear), 415
- Auricles of heart, 274, 275, 279
- Auricula, 415
- Auricular appendage, 275, 279
- arteries, 287
- cartilage, 415, 416
- facet, 10
- impression, 77
- Auricularis anterior, M., 104, 419
- externus, M., 420, 470
- posterior, M., 99
- superior, M., 100, 435
- Auricular nerve, 379; great, 385; posterior, 377
- vein, anterior, 324; posterior, 324
- Auriculotemporal nerve, 373
- Auriculoventricular septum, 274
- Axillary artery, 294
- border, 63
- nerve, 388
- vein, 318
- Axis, 15
- Azygos vein, 316
- Backbone, 1
- Back, fascia of, 126
- muscles of, 115, 123; dissection of, 442
- Basilar artery, 291, 292
- Basioccipital bone, 22
- Basisphenoid bone, 25
- Biceps brachii, M., 165, 438, 439
- femoris, M., 194, 444
- Bicipital arch, 166
- groove, 66
- tubercle, 64
- tuberosity, 67
- Bicuspid valve, 279
- Bile duct, 241
- Biventer cervicis, M., 132, 443
- Bladder, 256
- arteries of, 307
- ligaments of, 219, 257
- Blind spot, 414
- Blood vessels, 280
- dissection of, 456
- injection of, 456
- Body cavity, 217
- Body muscles, 115
- Bones, 1
- ear, 423
- head, 21
- pelvic extremities, 76
- preparation of, 430
- sesamoid, 73, 80, 86
- study of, 431
- thoracic extremities, 62
- Bony labyrinth, 425
- Brachial artery, 296
- Brachialis, M., 166, 438, 439
- Brachial plexus, 386; dissection of, 467; vein, 318
- Brachioradialis, M., 173, 438
- Brachium conjunctivum, 348, 349
- Brachium, muscles of, 164
- pontis, 347, 348, 349
- quadrigeminum inferius, 351, 354; superius,
351, 354
- Brain, 339
- arteries of, 289, 291
- diagrams of[476], 343
- dissection of, 464
- preparation of, 462
- preservation of, 462
- removal of, 462
- study of, 464
- veins of, 324
- Breast, muscles of, 144; dissection of, 437; nerves of, 387
- Broad ligament, 264, 266
- Bronchi, 252
- Bronchial arteries, 283
- veins, 318
- Buccæ, 222
- Buccal cavity, 221
- nerves, 377
- Buccinator, M., 106, 435
- nerve, 374
- Bulbocavernosus, M., 271, 454
- Bulbourethral glands, 261
- Bulbus olfactorius, 362
- urethræ, 261
- Bulla, auditory, 32, 55
- tympanic, 32, 55
- Burdach, column of, 337, 345
- Cæcum, 238
- Calf of leg, muscles of, 203
- Calcaneometatarsalis, M., 215, 445
- Calcaneus, 82, 83
- Callosal sulcus, 362
- Canal, alimentary, 221
- anterior palatine, 41
- condyloid, 24, 58
- facial, 35, 58
- Hunter’s, 311
- hypoglossal, 24, 58
- Canalis adductorius, 311
- facialis, 35, 58
- Canal, lachrymal, 40, 54, 410
- nasolachrymal, 40, 47, 54,
410
- of Huguier, 376
- posterior palatine, 45, 46, 54,
56
- pterygoid, 27, 56, 59
- sacral, 8
- Canals, semicircular, 425, 426
- Canine teeth, 225
- Caninus, M., 100, 435
- Capitatum, os, 70
- Capitulum, 18, 66
- Capsula lentis, 414
- Capsularis, M., 190, 444
- Capsule of Glisson, 302
- of lens 414
- Caput epididymis, 260
- Cardiac nerves, 380, 405, 406
- plexus, 380, 382, 405
- Cardinal veins, 328
- Carotid artery, common, 283; external, 285; internal, 285
- groove, 26
- plexus, 287
- Carpus, 69
- Cartilage, costal, 18
- Cartilages of larynx, 247
- Cartilago auriculæ, 415, 416
- Cauda epididymis, 260
- equina, 339
- Caudal muscles, 136; dissection of, 447
- vertebræ, 1, 11
- Caudoanalis, M., 270
- Caudocavernosus, M., 271
- Caudofemoralis, M., 195, 444
- Caudorectalis, M., 270
- Caudovaginalis, M., 272
- Cavernous sinus, 325
- Cavities of the skull, 57
- Cavity, abdominal, 217
- body, 217
- buccal, 221
- mediastinal, 218
- nasal, 59, 243
- pelvic, 218
- thoracic, 217
- Cavum nasi, 243
- oris, 221
- tympani, 422
- Central nervous system, 335
- Centrale, 69
- Centrum, 3
- Cephalica, V., 319
- Cephalobrachial muscle, 118
- Cephalohumeral muscle, 118
- Ceratohyal, 49
- Ceratohyoid muscle, 115, 440
- Cerebellar arteries, 292
- fossa, 57, 58
- gyri, 348
- hemispheres, 348
- Cerebellum, 341, 347
- arteries of, 292
- Cerebral aqueduct, 342, 343, 350,
351, 352
- arteries, 289, 292
- fissures, 359
- fossa, 58
- gyri, 359
- hemispheres, 341, 357
- peduncles, 351
- sulci, 359
- vein, inferior, 325
- Cerebri magna, V., 324
- Cerebrum, 341, 357
- arteries of, 289, 292
- Ceruminous glands, 416
- Cervical ganglia of sympathetic, 404, 405
- lymphatic glands, 332
- nerves, 383
- plexus, 386
- vertebræ[477], 1,
11, 15
- Cervicalis ascendens, A., 293; V., 320; profunda, A., 293
- Cervicofacial muscle, 95
- Cervix uteri, 266
- Chambers of eye, 414
- Cheek, muscles of, 105
- Cheeks, 222
- Chevron bones, 11
- Chiasma, optic, 342, 354, 355, 369
- Chiasmatic groove, 29, 59
- Choanæ, 60, 232, 244
- Chordæ tendineæ, 278
- Chorda tympani nerve, 375, 376
- Choroid, 413
- plexus of fourth ventricle, 350; of lateral ventricles,
353, 363, 367; of third ventricle,
354, 355
- Ciliary body, 413
- folds, 413
- ganglion, 369, 371
- nerve, long, 371; short, 370
- Circle of Willis, 289, 292
- Circulatory system, 274; dissection of, 455
- Circulus arteriosus, 289, 292
- Circumference, articular, 67
- Circumflexa femoris lateralis, A., 311
- humeri anterior, A., 296; posterior, A., 296
- scapulæ, A., 294
- Circumflex nerve, 388
- Circumvallate papillæ, 227
- Clava, 345, 349
- Clavicle, 64
- Clavicula, 64
- Clavobrachial muscle, 157, 437, 439
- Clavotrapezius muscle, 118, 436
- Cleidomastoid muscle, 120, 436, 440
- Clinoid processes, 26, 29
- Clitoris, 264, 267
- Coats of the eyeball, 413
- of the testis, 258
- Coccygeal nerves, 404
- Coccygeus, M., 137
- Cochlea, 425
- Cœliac artery, 301
- ganglion, 406
- plexus, 406
- Cœlom, 217
- Colica dextra, A., 304
- media, A., 303
- sinistra, A., 306
- Collateral ligaments, 74, 88
- radial artery, 297
- ulnar artery, 297
- Colliculi inferiores, 349, 351
- superiores, 351
- Collum of rib, 19
- Colon, 237
- Colon, ligaments of, 219
- Columnæ carneæ, 278
- Columns of spinal cord, 336
- Commissura anterior, 357, 365
- labiorum, 222
- posterior, 355
- Commissure, middle, 354
- Common bile-duct, 241
- carotid artery, 282, 283
- iliac veins, 328
- Communicating branches of sympathetic, 406
- Complexus, M., 133, 443
- Compound terms, viii
- Compressor urethræ membranaceæ, M., 271, 454
- Concha auris, 415, 416
- nasalis superior, 42
- ventral nasal, 40
- Conchæus externus, M., 422, 470
- Condyle, occipital, 23, 52, 55
- Condyles of femur, 80
- tibia, 80
- Condyloid canal, 24, 58
- process, 48
- Conjunctiva, 409, 410
- Constrictor cunni, M., 272
- pharyngis inferior, M., 232, 451; medius, M.,
233, 451; superior, M., 233
- vestibuli, M., 272
- Constrictors of anal glands, 269
- Conus arteriosus, 277
- Cor, 274
- Coracobrachialis, M., 164
- Coracoid border, 63, 64
- process, 63, 64
- Coracovertebral angle, 63
- Cord, spinal, 335
- Corium, 427
- Cornea, 412, 413
- Cornetoconchæus, M., 422
- Cornua of hyoid, 49
- of uterus, 266
- Coronal sulcus, 360
- Coronaria ventriculi, V., 326
- Coronary artery, 275, 279, 281
- ligament of liver, 221
- sinus, 277, 316
- sulcus, 274
- veins, 277, 315
- Coronoid fossa, 48, 66
- process, 48, 69
- Corpora mammillaria, 356
- quadrigemina, 351
- Corpus callosum, 357, 363
- cavernosum clitoridis, 267; penis, 263; urethræ,
261, 263
- ciliare, 413
- geniculatum laterale, 354; mediale, 351, 354
- luteum[478], 264
- of sternum, 20
- of vertebræ, 3
- pineale, 351, 355
- restiforme, 346, 348, 349
- spongiosum, 261
- striatum, 363, 367
- vitreum, 415
- Corrugator supercilii lateralis, M., 98, 435; medialis, M.,
97, 435
- Cortical portion of kidney, 256
- Costæ, 18
- spuriæ, 19
- veræ, 19
- Costal cartilage, 18
- facet, 3, 5
- Costocervical axis, 292
- vein, 318
- Cowper’s glands, 261
- Cranial cavity, 57
- nerves, 346, 369; dissection of, 465
- portion of skull, 21
- Cranium, 21
- Cremaster muscle, 258
- Cremasteric fascia, 258
- Crest, external occipital, 24, 52
- nasal, 40, 42
- of ilium, 78
- of tibia, 81
- sagittal, 25, 50
- Cribriform plate, 43
- Cricoarytenoideus lateralis, M., 251
- posterior, 249
- Cricoid cartilage, 248
- Cricothyroid muscle, 249
- Crista helicis, 417
- Crucial ligaments, 88, 89
- Cruciate sulcus, 360, 361
- Crura cerebelli, 341, 348
- of diaphragm, 152
- of penis, 263
- Crural arches, 154
- fascia, deep, 206
- muscle, 202
- Crus cerebelli ad medullam, 346
- Crus of diaphragm, 152
- Crus, muscles of, 203
- Crystalline lens, 414
- Cuboid, 82, 84
- Cuneatus, fasciculus, 337, 345, 346
- Cuneiform bones, 70, 80, 84, 85
- Cutaneous nerve, external, 388
- Cutaneus colli, N., 385
- femoralis lateralis, N., 396
- femoris posterior, N., 403
- maximus, M., 93, 434
- medialis, N., 390
- Cystic artery, 302
- duct, 241
- Deciduous teeth, 226
- Deltoid muscle, 156
- ridge, 66
- Demifacets, costal, 3
- Dens, 14, 15
- Dental arteries, 288
- nerves, inferior, 375
- Dentes, 224
- Depressor conchæ, M., 103, 435
- Dermal muscles, 93
- Diaphragm, 151; dissection of, 446
- Diastema, 225
- Diencephalon, 342, 343, 344, 352
- Digastric muscle, 107, 440
- Digestive apparatus, 221
- Digital arteries, 297, 314
- Digital fossa, 79
- Digits, 72
- Directions for dissection, 429
- Disarticulation of skulls, 430
- Dissection of alimentary canal, 449
- blood-vessels, 456
- brain, 464
- circulatory system, 455
- cranial nerves, 465
- diaphragm, 446
- ear, 470
- eye, 469
- ligaments, 431
- lymphatics, 460
- muscles, 433
- nervous system, 461, 465
- pelvic muscles, 446
- respiratory organs, 452
- sense organs, 469
- spinal nerves, 467
- sympathetic system, 465
- urogenital organs, 453
- viscera, 447
- Dorsalis penis, A., 308; N., 403
- Dorsum sellæ, 26, 59
- Drying of material, 432
- Ductus Botalli, 280
- communis choledochus, 241
- Ducts of salivary glands, 222, 223
- Duodenohepatic ligament, 220
- Duodenorenal ligament, 219
- Duodenum, 236; ligaments of, 219, 220
- Dura mater, 337, 368
- veins of, 324
- venous sinuses of, 324
- Ear, 415
- Ear bones, 423
- dissection of, 470
- external, removal of, 435
- muscles of, 96, 99, 103,
104, 418
- Ectocuneiform, 84
- Ectopectoralis, M., 145
- Ectosylvian gyrus[479], 360
- sulcus, 359
- Ectotympanic bone, 32
- Eighth cranial nerve, 347, 377
- Elbow-joint, 74
- Eleventh cranial nerve, 346, 382
- Eminence, iliopectineal, 78
- Eminentia conchæ, 416
- Encephalon, 339
- Entocuneiform, 85
- Entopectoralis, M., 146
- Entotympanic bone, 32
- Eparterial bronchus, 252
- Epicondyles, 67, 80
- Epicranius, M., 101, 435
- Epidermis, 427
- Epididymis, 260
- Epigastric artery, inferior, 309
- Epiglottic cartilage, 248
- Epiglottis, 246
- Epihyal, 49
- Epimeralis, M., 190
- Epiphysis, 351, 355
- of bones, 3
- Epistropheus, 15
- Epitrochlea, 67
- Epitrochlearis, M., 164, 438
- Ethmoidal artery, 289
- foramen, 38
- nerve, 371
- Ethmoid bone, 42
- cells, 44
- Ethmoturbinals, 42
- Eustachian tube, 232, 423
- Excretory organs, 255; dissection of, 453
- Exoccipital bones, 22
- Extensor antibrachii longus, M., 164
- brevis digitorum, M., 212, 445; pollicis, M.,
178, 439
- carpi radialis brevis, M., 174, 438; longus, M.,
173, 438; ulnaris, M., 176, 439
- caudæ lateralis, M., 137, 443,
447; medialis, M., 130, 136,
443, 447
- communis digitorum, M., 174, 439
- digitorum lateralis, M., 175, 439; longus,
210, 445
- dorsi communis, M., 124
- indicis proprius, M., 176, 439
- muscles of arm, 173
- ossis metacarpi pollicis, M., 178
- External carotid artery, 285
- ear, 415
- iliac artery, 307, 309; vein, 328,
329
- intercostal muscles, 150
- jugular vein, 319
- maxillary artery, 285, 286
- oblique muscle, 153
- occipital crest, 24, 52; protuberance, 24
- pterygoid muscle, 111
- sternocostal muscle, 150
- Extremities, pelvic, bones of, 76
- thoracic, bones of, 62
- Eye, 409, 412
- Eyeball, 412
- Eyelids, 409; muscles of, 97, 410, 412
- Eye, muscles of, 411
- Face, muscles of, 96
- veins of, 321
- Facet, articular, 4
- auricular, 10
- costal, 3, 5
- transverse costal, 4
- tubercular, 4
- Facial bones, 21
- canal, 35, 58
- muscles, 96; dissection of 434
- nerve, 347, 375
- portion of skull, 21, 49
- vein, anterior, 321
- vein, deep, 323; posterior, 323
- Fallopian tubes, 263, 264
- False ribs, 19
- vocal cords, 246
- Fascia, antibrachial, 172
- cremasteric, 258
- deep crural, 206
- endothoracica, 217
- intercolumnar, 258
- lata, 186
- lumbodorsal, 126
- of back, 126
- of thigh, 186
- propria, 258
- Fasciculus cuneatus, 337, 345, 346
- cuneatus lateralis, 346
- gracilis, 336, 345
- Fauces, pillars of, 230
- Female genital organs, 263
- Femoral artery, 307, 310
- nerve, 397
- Femur, 79
- Fenestra cochleæ, 34, 425
- ovalis, 34
- rotunda, 34
- vestibuli, 34, 424, 425
- Fibrocartilages, intervertebral, 16
- Fibrocartilagines intervertebrales, 16
- Fibula, 82
- Fifth cranial nerve, 347, 379
- Filiform papillæ, 227
- Filum terminale, 335
- Fingers, arteries of, 297
- bones of, 72
- muscles of, 185
- First cranial nerve, 369
- Fissura cerebri lateralis, 358
- Fissure of Sylvius, 358
- orbital[480], 27,
54, 59
- Fissures of cerebrum, 359
- medulla oblongata, 344, 345
- spinal cord, 335
- Flexor brevis digiti quinti, M., 186, 439; digiti secundi, M.,
185; digitorum, M., 205, 212,
445; pollicis, M., 184, 439
- carpi radialis, M., 179, 439
- carpi ulnaris, M., 180, 439
- caudæ brevis, M., 138, 446; longus, M.,
138, 446; longus digitorum, M., 207,
208, 445; hallucis, M., 207, 445
- muscles of forearm, 179
- perforans, M., 181
- perforatus, M., 181
- profundus digitorum, M., 181, 439
- sublimis digitorum, M., 181, 439
- Floating ribs, 19
- Follicles, Graafian, 264
- solitary, 237
- Foot, arteries of, 312, 314
- bones of, 82
- joints and ligaments of, 91
- muscles of, 212; dissection of, 445
- nerves of, 402
- veins of, 329
- Foramen, anterior palatine, 57
- atlantal, 14
- cæcum, 345
- epiploicum, 220
- ethmoidal, 38
- hypoglossal, 24, 56, 58
- incisivum, 41, 57
- infraorbital, 40, 54
- interventricular, 356, 365, 368
- intervertebral, 4
- jugular, 24, 33, 56, 58
- lacerum, 26, 59; anterius, 54
- magnum, 22, 52, 55, 58
- mandibular, 48
- mental, 47
- obturator, 76
- of Monroe, 356, 365, 368
- of Winslow, 220
- optic, 29, 54, 59
- ovale, 27, 54, 56, 59
- ovale of heart, 277
- rotundum, 27, 54, 56, 59
- sacral, 9, 10
- sphenopalatine, 46, 54
- stylomastoid, 33, 34, 52,
55
- supracondyloid, 67
- transversarium, 13
- vertebral, 1
- Forearm, muscles of, 172
- Fore limbs, bones of, 62
- muscles of, 156; dissection, 436
- lymphatics of, 332
- Formalin-alcohol mixture for brains, 463
- Formalin as preservative, 431
- Formalin-glycerine mixture for injection, 432
- Formula for teeth, 224
- Fornicate gyrus, 362
- Fornix, 358, 363, 364
- Fossa, anterior, 59
- appendicular, 35, 58
- cerebellar, 57, 58
- cerebral, 58
- coronoid, 48, 66
- digital, 79
- external pterygoid, 30, 53
- glenoid, 63
- iliopectineal, 310
- infraspinous, 63
- intercondyloid, 80
- internal pterygoid, 28, 54
- mandibular, 32, 52
- masseteric, 48
- middle, 59
- olecranon, 66
- olfactory, 59
- orbital, 53
- ovalis, 277
- radial, 66
- rhomboidea, 349
- subscapular, 63
- supraspinous, 63
- temporal, 51, 53
- trochanteric, 79
- Fourth cranial nerve, 350, 370
- ventricle, 341, 343, 344,
349
- Frenulum glossoepiglottica, 227
- linguæ, 222, 227
- of epiglottis, 246
- of lips, 221
- penis, 257
- Frontal bone, 37
- lobe of brain, 358
- muscle, 101
- nerve, 370
- plate, 37
- process, 39, 40, 47, 51
- sinuses, 38, 61
- spine, 37
- vein, 321
- Frontoauricular muscle, 99, 435
- Frontoscutularis, M., 104, 435
- Fundus of stomach, 234
- Fungiform papillæ, 227
- Funiculi of spinal cord, 336
- Galea aponeurotica, 101
- Gall-bladder, 241
- Ganglia, sympathetic, 404, 405, 406,
408
- thoracic, 406
- Ganglion cervicale inferius, 405
- cervicale mediale, 405; superius, 404
- cœliac[481], 406
- Gasserian, 347, 370
- geniculi, 375
- inferior mesenteric, 408
- jugulare, 378
- nodosum, 378
- of the root, 378
- of the trunk, 378
- ophthalmic, 369
- petrosum, 378
- semilunar, 347, 370, 406
- sphenopalatine, 372
- superior mesenteric, 407
- superius, 378
- Gasserian ganglion, 347, 370
- Gastrica sinistra, A., 303
- Gastric plexuses, 382
- Gastrocnemius, M., 203, 445
- Gastroduodenal ligament, 235
- artery, 302
- Gastroepiploica dextra, A., 303; V., 327
- Gastrohepatic ligament, 220, 245
- Gastrolienal ligament, 235
- omentum, 220
- Gastrosplenic omentum, 220
- vein, 327
- Gemellus inferior, M., 190, 444
- superior, M., 189, 444
- Geniculate ganglion, 375
- Genioglossus, M., 228, 440, 450
- Geniohyoid muscle, 113, 440
- Genital organs, 257; dissection of, 453, 454
- Genitofemoral nerve, 396
- Genu (brain), 364
- Gland, bulbourethral, 261
- Cowper’s, 261
- infraorbital, 224
- lachrymal, 410
- molar, 224
- orbital, 224
- parotid, 223
- prostate, 261
- sublingual, 224
- submaxillary, 223
- thymus, 254
- thyroid, 254
- Glands, Harderian, 410
- inguinal, 334
- lymphatic, 330
- Meibomian, 409
- of mouth, 223
- of urethra, 261
- tarsal, 409
- Glandulæ oris, 223
- suprarenales, 257
- Glans penis, 257, 261
- Glenoid angle, 62, 64
- border, 63
- fossa, 63
- Glenovertebral angle, 63
- Glisson, capsule of, 302
- Glossoepiglotticus, M., 250
- Glossopharyngeal nerve, 347, 378
- Glossopharyngeus, M., 232, 451
- Glottis, 246
- Gluteal arteries, 308
- nerve, inferior, 399; superior, 399
- vein, inferior, 329
- Gluteus maximus, M., 187, 444
- medius, M., 188, 444
- minimus, M., 189, 444
- quartus, M., 190
- Glycerine-formalin mixture for injecting, 432
- Goll, column of, 337, 345
- Graafian follicles, 264
- Gracilis, fasciculus, 336, 345; M., 198,
444
- Gray matter, 336
- Great auricular vein, 324
- Greater curvature of stomach, 234
- Great omentum, 219, 235
- scapular notch, 64
- sciatic nerve, 400
- sciatic notch, 77
- sigmoid cavity, 69
- transverse fissure, 363
- trochanter, 79
- tuberosity, 65
- Gyri of cerebellum, 348
- cerebrum, 359
- Gyrus, anterior Sylvian, 359
- compositus posterior, 360
- ectosylvius, 360
- fornicatus, 362
- marginalis, 360, 361
- orbitalis, 360
- posterior Sylvian, 359
- sigmoideus, 360
- suprasylvius, 360
- Habenulæ, 355
- Hæmal processes, 11
- Hair, distribution of, 427
- Hamatum, os, 70
- Hamular process, 28
- Hamulus, 28, 54
- Hand, arteries of, 300
- bones of, 71
- joints and ligaments of, 75
- muscles of, 184; dissection of, 439
- Harderian glands, 410
- Head, bones of, 21
- muscles of, 96; dissection, 434, 440
- nerves of, 369
- Heart, 274; dissection of, 455
- veins of, 315
- Heel, 83
- Helicis, M., 421, 470
- Helix[482], 417
- Hemispheres of cerebellum, 348
- cerebrum, 341, 357
- Hemorrhoidal arteries, 306, 308
- nerve, inferior, 404
- nerve, middle, 403
- veins, 329
- Hepar, 239
- Hepatic artery, 302
- ducts, 241
- plexus, 408
- veins, 326
- Hepatorenal ligament, 221
- Hiatus facialis, 35
- Hilus, 255
- Hind brain, 344
- Hind limb, bones of, 76
- muscles of, 186; dissection, 444
- Hip-joint, 87
- Hip, muscles of, 186
- Hippocampal sulcus, 362
- Hippocampus, 365
- Horns of uterus, 266
- Huguier, canal of, 376
- Humerus, 64
- Humor, aqueous, 414
- vitreous, 415
- Hunter’s canal, 311
- Hyoepiglotticus, M., 250
- Hyoglossus, M., 228, 440, 450
- Hyoid bone, 49
- muscles of, 112
- Hyparterial bronchi, 252
- lobes of lungs, 281
- Hypogastric arteries, 307
- vein, 328, 329
- Hypoglossal canal, 24, 58
- foramen, 24, 56, 58
- nerve, 346, 383
- Hypophysis, 355, 356
- Iliac arteries, 307, 309
- glands, 333
- veins, 328, 329
- Ileocolic artery, 304
- valve, 237
- Ileum, 237
- Iliocaudalis, M., 137, 446
- Iliocostalis, M., 124, 128, 443
- Iliohypogastric nerve, 395
- Ilioinguinal nerve, 395
- Iliolumbar arteries, 306
- veins, 326
- Iliopectineal eminence, 78
- fossa, 310
- line, 78
- Iliopsoas, M., 138, 193, 444, 446
- Iliosacral ligaments, 86, 87
- Ilium, 76, 77
- Incisive duct, 222
- foramen, 41, 57
- Incisor teeth, 225
- Incisura acetabuli, 79
- scapulæ, 64
- Incus, 424
- Index, 72
- Index, muscles of, 185
- Inferior alveolar artery, 288
- cerebral vein, 325
- cervical ganglion, 405
- dental artery, 288
- epigastric artery, 309
- gluteal arteries, 308
- maxillary bone, 47
- mesenteric artery, 306; vein, 328
- thyroid artery, 283, 284
- vena cava, 325
- vesical artery, 307
- Infraorbital artery, 290
- foramen, 40, 54
- gland, 224
- nerve, 372
- Infraspinatus muscle, 160, 438
- Infraspinous fossa, 63
- Infratrochlear nerve, 371
- Infundibulum, 355
- Inguinal canal, 259
- glands, 334
- ligament, 153
- rings, 259
- Injection mass, 456
- of blood-vessels, 456
- of formalin, 431
- of lymphatics, 460
- Innominate arteries, 282
- bone, 76
- veins, 318
- Insula, 359
- Integument, 427
- muscles, 93
- Integumentum commune, 427
- Intercarpal ligaments, 75
- Intercolumnar fascia, 258
- Intercondyloid fossa, 80
- Intercostal arteries, 283
- artery, superior, 293
- Intercostales externi, M., 150, 441
- interni, 151, 441
- Intercostal veins, 316
- Internal carotid artery, 285
- ear, 424
- iliac artery, 307; vein, 329
- intercostals, 151
- jugular vein, 320
- mammary artery, 292; vein, 318
- maxillary artery, 287
- oblique muscle, 154
- pterygoid muscle, 111
- sternocostal muscle, 151
- Intermediate cuneiform, 84
- Intermedius scutulorum, M.[483], 96, 443
- Interossei, Mm., 185, 214, 439, 445
- Interosseous arteries, 298, 300
- ligaments, 75
- membrane, 75, 90
- nerve, dorsal, 392; posterior, 390
- Interparietal bone, 25
- Interspinalis, M., 124, 131, 136, 443
- Intertransversarii, M., 124, 131, 136,
443
- Intertrochanteric line, 79
- Interventricular foramen, 356, 365, 368
- Intervertebral fibrocartilages, 16
- foramina, 4
- Intestine, 236
- arteries of, 303
- Intestine, large, 237
- small, 236
- Intestinum crassum, 237
- tenue, 236
- Intrinsic muscles of tongue, 229
- Iris, 413, 414
- Ischiadicus, N., 400
- Ischiocavernosus, M., 269, 454, 455
- Ischium, 76, 78
- Isthmus faucium, 221, 230
- rhombencephali, 350
- Jacobson, organ of, 222, 244
- Jaw, lower, bone of, 47
- upper, bones of, 39, 41
- Jejunum, 237
- Joints of pelvic limbs, 86
- of skull, 61
- of thoracic limbs, 73
- Jugal bone, 47
- Jugular foramen, 24, 33, 56, 58
- ganglion, 378
- notch, 24
- process, 24, 52, 55
- vein, external, 319; internal, 320
- Jugulohyoid muscle, 113, 440
- Kidney, 255
- Kidneys, arteries to, 304
- Knee-joint, 87, 89
- Knee, ligaments of, 87, 89
- Labial arteries, 286
- veins, 321
- Labia oris, 221
- Labrum glenoidale, 87
- Labyrinth, 424, 425
- Labyrinths of ethmoid, 42, 43
- Lachrymal apparatus, 410
- bone, 46
- canal, 40, 54, 410
- gland, 410
- nerve, 371
- Lambdoidal ridge, 24, 50, 52
- Lamina, 4
- chorioidea epithelialis, 355, 363, 367
- cribrosa, 43
- fusca, 413
- papyracea, 44
- perpendicularis, 42, 43
- spiralis, 425
- terminalis, 352, 357
- Large intestine, 237
- Laryngeal arteries, 285
- Laryngeal muscles, 249; dissection of, 452
- nerve, inferior, 380; superior, 380
- veins, 321
- Larynx, 246
- cartilages of, 247
- muscles of, 249
- Lateral angle (scapula), 63
- cuneiform, 84
- fissure of cerebrum, 358
- ligaments of bladder, 257
- mass of atlas, 14; of sacrum, 10
- sulcus, 360
- ventricles, 341, 343, 362,
368
- Latissimus dorsi, M., 121, 437
- Leg, arteries of, 309
- bones of, 79
- ligaments of, 86
- lower, muscles of, 203
- muscles, 186; dissection, 444
- veins of, 329
- Lens, crystalline, 414
- Lenticular bone, 424
- process, 424
- Lesser curvature of stomach, 234
- omentum, 220
- palatine artery, 290
- sciatic notch, 78
- trochanter, 79
- tuberosity of humerus, 65
- Levator anguli oris, M., 106
- ani, M., 137, 269, 446,
454
- auris longus, M., 99, 435
- claviculæ, M., 120
- Levatores costarum, M., 150, 443
- Levator labii superioris alæque nasi, M., 105, 435;
superioris proprius, M., 106, 435
- palpebræ superioris, M., 412
- scapulæ, M., 122, 123, 437
- scapulæ dorsalis, M., 118
- scapulæ ventralis, M., 120, 436, 440,
443
- scroti, M., 258, 271, 453
- veli palatini, M., 230, 451
- vulvæ, M., 272, 455
- Lien, 242
- Ligament (see also Ligamentum)
- annular, 75, 173, 213
- anterior longitudinal, 16
- broad[484], 264, 266
- cervical supraspinous, 17
- coronary, 221
- duodenohepatic, 220
- duodenorenal, 219
- gastroduodenal, 235
- gastrohepatic, 220, 235
- gastrolienal, 235
- hepatorenal, 221
- inguinal, 153
- lateral, of atlas, 17
- orbital, 47
- of peritoneum, 218
- posterior longitudinal, 16
- Poupart’s, 153
- pulmonary, 253
- round, 87, 220, 267
- stylomandibular, 62, 228
- supraspinous, 16
- suspensory, 219, 220, 257
- transverse, 18, 87, 91,
172
- triangular, 221
- Ligaments, collateral, 74, 88
- crucial, 89
- iliosacral, 86, 87
- intercarpal, 75
- interosseous, 75
- of bladder, 219, 257
- of hip-joint, 87
- of liver, 220
- of malleoli, 91
- of menisci, 90
- of ovary, 264
- of pelvic limbs, 86
- of ribs, 19
- of skull, 61
- of tarsus, 91
- of thoracic limbs, 73
- of uterus, 266
- of vertebral column, 16
- preparation of, 431
- study of, 431
- vaginal, 173
- Ligamentum (a)
- alaria, 18
- arteriosum, 280
- Botalli, 280
- collaterale, 74, 88
- cruciatum, 89
- denticulatum, 337
- dorsalia, 92
- flava, 17
- interossea, 92
- interspinalia, 17
- intertransversaria, 17
- lateralia, 92
- longitudinale anterius, 16; posterius, 16
- nuchæ, 17
- ovarii, 264
- patellæ, 81, 88
- plantaria, 92
- supraspinale, 16
- suspensorium penis, 262
- teres, 87, 220
- transversum cruris, 91
- Limbs, arteries of, 294, 309
- bones of, 62, 76
- lymphatics of, 332, 334
- muscles of, 156, 186
- veins of, 318, 328
- Linea aspera, 80
- Line, iliopectineal, 78
- intertrochanteric, 79
- spiral, 79
- Lingua, 226
- Lingual artery, 285, 286
- nerve, 375
- vein, 323
- Lingula, 28
- Lips, 221
- muscles of, 105
- Liquid soap, for bones, 430
- Liver, 239
- arteries of, 302
- ligaments of, 220
- veins of, 326
- Lobes of cerebrum, 358
- of liver, 239
- Lobus pyriformis, 361, 362
- Longissimus capitis, M., 128, 131, 443
- cervicis, M., 128, 134
- dorsi, M., 124, 126, 134,
442
- Long saphenous nerve, 397
- thoracic artery, 294
- Longus atlantis, M., 134, 443
- capitis, M., 142, 442
- colli, M., 144, 442
- Lumbar aponeurosis, 154
- arteries, 283, 306
- nerves, 394
- plexus, 395
- veins, 326
- vertebræ, 1, 7
- Lumboabdominal arteries, 304
- Lumbodorsal fascia, 126
- Lumboinguinal nerve, 396
- Lumbosacral cord, 399
- plexus, 396
- Lumbricales, Mm., 184, 213, 439, 445
- Lunare, 69
- Lungs, 252
- Lymph, 330
- Lymphatic duct, right, 331, 332
- glands, 330
- Lymphatics, 330
- demonstration of, 460
- injection of, 460
- Magnum, os, 70
- Malar bone[485], 47
- Male genital organs, 257
- Malleoli, ligaments of, 91
- Malleolus, lateral, 82
- medial, 81
- Malleus, 423
- Mammary artery, internal, 292
- glands, 268
- Mammillary bodies, 356
- process, 6, 8
- Mandible, 47
- articulations of, 61
- Mandibular foramen, 48
- fossa, 32, 52
- nerve, 370, 373
- Manubrium, 20
- Manus, bones of, 71
- Marginal gyrus, 360, 361
- sulcus, 361
- Margin, supraorbital, 37
- Massa intermedia, 354, 357
- Masseter, M., 108, 440
- Masseteric fossa, 48
- nerve, 374
- Mastication, muscles of, 107
- Mastoid portion of temporal, 34, 35
- process, 35, 55
- Maxilla, 39
- Maxillary arteries, 285, 286, 287
- Maxillary bone, 39
- inferior, 47
- nerve, 370, 371
- spine, 45
- Maxilloturbinal, 40
- Meati of nose, 44, 60, 243, 244,
245
- Meatus auditorius externus, 32, 33, 52, 55,
415, 416
- external auditory, 32, 33, 52,
55, 415, 416
- inferior, 60
- internal auditory, 35, 58
- superior, 44
- Medial cuneiform, 85
- Mediana cubiti, V., 319
- Median nerve, 390
- Mediastinal artery, 282
- cavity, 218
- lobe of lung, 253
- Mediastinum, 218
- Medius, 72
- Medulla oblongata, 340, 344
- Medullary portion of kidney, 256
- Medulla spinalis, 335
- Meibomian glands, 409
- Membrana nictitans, 410
- tympani, 422
- Membrane, anterior atlanto-occipital, 17
- interosseous, 75, 90
- mucous, 221
- posterior atlanto-occipital, 17
- Membranes of brain, 368
- spinal cord, 337
- Membranous labyrinth, 425; preparation of, 471
- Meningeal artery, middle, 289
- Menisci, 87; ligaments of, 90
- Mental foramen, 47
- nerves, 375
- Mesencephalon, 341, 343, 344, 351
- Mesenteric arteries, 303, 306
- Mesenteric ganglion, inferior, 408; superior, 407
- glands, 333
- plexus, inferior, 408; superior, 408
- vein, inferior, 328; superior, 328
- Mesentery, 218, 219, 236
- Mesocolon, 219
- Mesocuneiform, 84
- Mesogastrium, 219
- Mesorectum, 219
- Metacarpals, 71
- muscles of, 185
- Metacarpus, 71
- Metacromion, 64
- Metapophysis, 6
- Metatarsal artery, 315
- Metatarsals, 85
- Metatarsus, 85
- Metencephalon, 341, 343, 344, 347
- Midbrain, 341, 343, 344, 351
- Middle cerebral artery, 289
- cervical ganglion, 405
- Middle ear, 422; muscles of, 424
- commissure, 354
- fossa, 59
- meningeal artery, 289
- sacral artery, 301, 309; vein, 328
- Minimus, 72; muscles of, 185
- Mitral valve, 279
- Modiolus, 425
- Molar gland, 224
- teeth, 225
- Monroe, foramen of, 356, 365, 368
- Mould, attacks of, 432
- Moustachier, M., 107, 435
- Mouth, 221; glands of, 223
- Mucosa, 221
- Mucous membrane, 221
- Multangulum majus, os, 71
- minus, os, 71
- Multifidus spinæ, 124, 130, 443
- Muscle, see Musculus
- Muscles, 93; dissection of, 434
- abdomen, 153; dissection, 441
- antibrachium, 172
- anus, 268
- arm, 156; dissection of, 437
- back, 115, 123; dissection, 442
- body, 115
- breast, 144; dissection, 437
- of crus[486], 203
- digits, 184, 213
- ear, 96, 99, 103, 104,
418; dissection, 470, 471
- external ear, 418
- eye, 411; dissection of, 469
- eyelid, 97, 410, 412
- face, 96; dissection, 434
- fingers, 185
- foot, 212; dissection, 445
- forearm, 172
- fore limb, 156; dissection, 436
- hand, 184; dissection, 439
- head, 96; dissection, 434, 440
- hind limbs, 186; dissection, 444
- hip, 186
- hyoid, 112
- index finger, 185
- larynx, 249; dissection of, 452
- lips, 105
- lower leg, 203
- mastication, 107
- middle ear, 424
- minimus, 185
- neck, 131, 139; dissection, 439,
442
- orbit, 411
- palate, 230; dissection, 451
- pectoral, 144
- pelvic limbs, 186
- pelvis, dissection, 446
- penis, 271
- pharynx, 232; dissection, 451
- preparation of, 431
- shoulder, 115, 156
- skin, 93, 427; dissection, 434
- tail, 136; dissection, 447
- tarsus, 215
- thigh, 194
- thoracic limbs, 156; dissection, 436
- thoracic wall, 148
- thorax, 144; dissection, 441
- thumb, 184
- toes, 214
- tongue, 228; dissection, 450
- urethra, 271
- urogenital organs, 268; dissection, 454
- vagina, 272
- vertebral column, 123, 138; dissection, 442,
446
- Musculi pectinati, 276
- Musculocutaneous nerve, 388
- Musculospiral nerve, 392
- Musculus (i)
- abductor auris brevis, 100, 435; longus, 100,
435
- brevis pollicis, 184, 439
- caudæ (coccygis) externus, 137, 447
- internus, 137, 446
- cruris, 195
- digiti quinti, 185, 439; secundi, 185
- medius digiti quinti, 215
- accelerator urinæ, 271
- acromiodeltoideus, 157, 438
- acromiotrapezius, 116, 436
- adductor auris inferior, 104, 435; medius, 419,
470; superior, 419, 470
- femoris, 198, 444
- digiti secundi, 185
- longus, 199, 444
- medius digiti quinti, 215
- pollicis, 185, 439
- anconeus, 170, 438; internus, 170;
lateralis, 167; longus, 168; posterior, 168
- antitragicus, 421, 470
- arytenoideus transversus, 250
- attollens auris, 100
- auricularis anterior, 104, 419; externus, 420,
470; posterior, 99; superior, 100, 435
- biceps brachii, 165, 438, 439; femoris,
194, 444
- biventer cervicis, 132, 443
- brachialis, 166, 438, 439
- brachioradialis, 173, 438
- buccinator, 106, 435
- bulbocavernosus, 271, 454
- calcaneometatarsalis, 215, 445
- caninus, 106, 435
- capsularis, 190, 444
- caudoanalis, 270
- caudocavernosus, 271
- caudofemoralis, 195, 444
- caudorectalis, 270
- caudovaginalis, 272
- ceratohyoideus, 115, 440
- cervicofacialis, 95
- clavobrachialis, 157, 437, 439
- clavotrapezius, 118, 436
- cleidomastoideus, 120, 436, 440
- coccygeus, 137
- complexus, 133, 443
- compressor urethræ membranaceæ, 271, 454
- conchæus externus, 422, 470
- constrictor cunni, 272
- pharyngis inferior, 232, 451; medius, 233,
451; superior, 233
- vestibuli, 272
- coracobrachialis, 164
- cornetoconchæus, 422
- corrugator supercilii lateralis, 98, 435
- corrugator supercilii medialis, 97, 435
- cremaster, 258
- cricoarytenoideus lateralis, 251; posterior, 249
- cricothyreoideus, 249
- cruralis, 202
- cutaneus maximus[487], 93,
434
- deltoideus, 156
- depressor conchæ, 103, 435
- digastricus, 107, 440
- ectopectoralis, 145
- entopectoralis, 146
- epicranius, 101, 435
- epimeralis, 190
- epitrochlearis, 164, 438
- extensor antibrachii longus, 164
- brevis digitorum, 212, 445; pollicis, 178,
439
- carpi radialis brevis, 174, 438; longus, 173,
438
- carpi ulnaris, 176, 439
- caudæ lateralis, 137,443, 447; medialis,
130, 136, 443, 447
- communis digitorum, 174, 439
- digitorum lateralis, 175, 439; longus, 210,
445
- dorsi communis, 124
- indicis proprius, 176, 439
- ossis metacarpi pollicis, 178
- flexor brevis digiti quinti, 186, 439; secundi,
185
- digitorum, 205, 212, 445
- pollicis, 184, 439
- carpi radialis, 179, 439; ulnaris, 180,
439
- caudæ brevis, 138, 446; longus, 138,
446
- longus digitorum, 207, 208, 445; hallucis,
207, 445
- perforans, 181
- perforatus, 181
- profundus digitorum, 181, 439
- sublimis digitorum, 181, 439
- frontalis, 101
- frontoauricularis, 99, 435
- frontoscutularis, 104, 435
- gastrocnemius, 203, 445
- gemellus inferior, 190, 444; superior, 189,
444
- genioglossus, 228, 440, 450
- geniohyoideus, 113, 440
- glossoepiglotticus, 250
- glossopharyngeus, 232, 451
- gluteus maximus, 187, 444; medius, 188,
444; minimus, 189, 444; quartus, 190
- gracilis, 198, 444
- helicis, 421, 470
- hyoepiglotticus, 250
- hyoglossus, 228, 440, 450
- iliocaudalis, 137, 446
- iliocostalis, 124, 128, 443
- iliopsoas, 138, 193, 444,
446
- incisivi, 107
- infraspinatus, 160, 438
- intercostales externi, 150, 441; interni,
151, 441
- intermedius scutulorum, 96, 434
- interossei, 185, 214, 439,
445
- interspinalis, 124, 131, 136,
443
- intertransversarii, 124, 131, 136,
443
- ischiocavernosus, 269, 454, 455
- jugulohyoideus, 113, 440
- latissimus dorsi, 121, 437
- levator anguli oris, 106
- ani, 137, 269, 446,
454
- auris longus, 99, 435
- claviculæ, 120
- levatores costarum, 150, 443
- levator labii superioris alæque nasi, 105, 435
- labii superioris proprius, 106, 435
- palpebræ superioris, 412
- scapulæ, 122, 123, 437; dorsalis,
118; ventralis, 120, 436, 440,
443
- scroti, 258, 271, 453
- veli palatini, 230, 451
- vulvæ, 272, 455
- longissimus capitis, 128, 131, 443; cervicis,
128, 134; dorsi, 124, 126,
134, 442
- longus atlantis, 134, 443; capitis, 142,
442; colli, 144, 442
- lumbricales, 184, 213, 439,
445
- masseter, 108, 440
- moustachier, 107, 435
- multifidus spinæ, 124, 130, 443
- mylohyoideus, 114, 440
- myrtiformis, 106, 435
- nasalis, 106
- obliqui, 411
- obliquus abdominis externus, 153, 441; internus,
154, 441
- capitis inferior 136; superior, 136, 442, 443
- inferior, 412, 443
- superior, 412
- obturator externus, 191, 444; internus, 192,
444
- occipitalis, 101
- occipitofrontalis, 101
- occipitoscapularis, 118, 436
- opponens digiti quinti, 186, 215, 439
- orbicularis oculi, 98, 435; oris, 105,
435
- palmaris longus, 179, 439
- parameralis, 195
- pectineus, 200, 444
- pectoantibrachialis, 145, 437
- pectoralis major, 145, 437; minor, 146,
437
- pedis perforatus, 205, 212
- peroneus brevis, 210, 445; longus, 209,
445; tertius, 210, 445
- plantaris, 205, 445
- platysma[488], 95, 434
- popliteus, 206, 445
- pronator quadratus, 183, 439
- teres, 179, 439
- psoas minor, 139, 446
- pterygoideus externus, 111, 441; internus, 111,
441
- pterygopharyngeus, 233
- pubiocaudalis, 269
- pyriformis, 188, 444
- quadratus femoris, 191, 444
- labii inferioris, 107, 435; superioris, 105,
435
- lumborum, 139, 446
- plantæ, 213, 445
- quadriceps femoris, 201, 444
- recti, 411
- rectocavernosus, 271
- rectovaginalis, 272
- rectus abdominis, 155, 441
- capitis anterior major, 142; minor, 143, 442
- lateralis, 143, 442
- posterior major, 134, 443; medius, 135,
443; minor, 135, 443
- femoris, 201, 444
- inferior, 411; lateralis, 411; medialis,
411; superior, 411
- retractor oculi, 411; penis, 271
- rhomboideus, 119, 436; capitis, 118
- rotator auris, 419, 470
- rotatores, 131
- sartorius, 197, 444
- scalenus, 141, 441
- scalenus anterior, 141, 142
- medius, 141; posterior, 141
- scaphocuneiformis, 216, 445
- scutuloauricularis inferior, 419
- semimembranosus, 196, 444
- semispinalis capitis, 130, 132
- cervicis, 130, 133, 443
- semitendinosus, 196, 444
- serratus anterior, 122, 437
- posterior inferior, 148, 441; superior, 148,
441
- soleus, 205, 445
- sphincter ani externus, 268, 453; internus, 269,
454
- spinalis dorsi, 124, 128, 129,
133, 443
- spinodeltoideus, 156, 438
- spinotrapezius, 115, 436
- splenius, 131, 443
- stapedius, 424, 471
- sternocleidomastoideus, 120, 139
- sternocostalis externus, 150; internus, 151
- sternohyoideus, 140, 440
- sternomastoideus, 139, 440
- sternothyreoideus, 141, 440
- styloglossus, 228, 440, 450
- stylohyoideus, 112, 440
- stylopharyngeus, 233, 451
- submentalis, 103, 435
- subscapularis, 161, 437
- supercervicocutaneus, 95
- supinator, 177, 439; longus, 173
- supraspinatus, 159, 438
- temporalis, 110, 440
- tensor fasciæ latæ, 187, 444
- tympani, 424, 471
- veli palatini, 230, 451
- tenuissimus, 195, 444
- teres major, 163, 437, 438; minor,
161, 438
- thyreoarytenoideus, 251
- thyreohyoideus, 249
- tibialis anterior, 211, 445; posterior, 208,
445
- trachelomastoideus, 131
- tragicus lateralis, 418, 435; medialis, 421,
470
- transversus abdominis, 155, 441; auriculæ,
420, 470; costarum, 150, 441; perinei,
269; thoracis, 151, 441
- trapezius, 115, 436
- triangularis sterni, 151
- triceps brachii, 166, 438
- suræ, 206
- urethralis, 273
- vastus intermedius, 203, 445; lateralis, 201,
445; medialis, 202, 445
- xiphihumeralis, 147, 437
- zygomaticus (major), 101, 435; minor, 105,
435
- Myelencephalon, 340, 343, 344
- Mylohyoid muscle, 114, 440
- Myrtiformis, M., 106, 435
- Nails, 427
- Names of organs, v
- Names of vertebral muscles, 124
- Nares, 51, 60, 244
- posterior, 60
- Nasal bone, 42
- cavity, 59, 243
- concha, superior, 42; ventral, 40
- crest, 40, 42
- Nasalis, M., 106
- Nasal nerve, posterior, 372
- process, 41
- spine, 37; posterior, 45
- veins, external, 321
- Nasolachrymal canal, 40, 47, 54, 410
- duct, 40, 47, 410
- Nasopharynx, 231
- Nasoturbinal bone, 42
- Navicular, 69, 82
- Naviculare pedis, os, 84
- Neck, arteries of[489], 283, 293
- muscles of, 131, 139
- dissection, 439, 442
- nerves of, 383
- veins of, 320
- Nerve, see Nervus.
- Nerves, cervical, 383
- coccygeal, 404
- cranial, 369
- lumbar, 394
- of foot, 402
- of head, 369
- of neck, 383
- of palm, 393
- of shoulder, 387
- of tail, 404
- of taste, 378
- sacral, 399
- spinal, 337, 383
- sympathetic, 404
- thoracic, 393
- Nervous system, 335; dissection, 461, 465; preparation,
461; sympathetic, 404
- Nervus abducens, 347, 375
- accessorius, 346, 382
- acusticus, 347, 377
- alveolaris inferior, 374, 375
- auricularis, 379
- auricularis magnus, 385; posterior, 377
- auriculotemporalis, 373
- axillaris, 388
- buccalis, 377
- buccinatorius, 374
- canalis pterygoidii, 372
- cerebrales, 369
- cardiacus, 380, 405, 406
- chorda tympani, 375, 376
- ciliaris brevis, 370
- ciliaris longus, 371
- circumflexus, 388
- cutaneus colli, 385
- femoris lateralis, 396; posterior, 403
- medialis, 390
- dorsalis penis, 403
- ethmoidalis, 371
- facialis, 347, 375
- femoralis, 397
- frontalis, 370
- genitofemoralis, 396
- glossopharyngeus, 347, 378
- gluteus inferior, 399; superior, 399
- hemorrhoidalis inferior, 404; medius, 403
- hypoglossus, 346, 383
- iliohypogastricus, 395
- ilioinguinalis, 395
- infraorbitalis, 372
- infratrochlearis, 371
- interosseus dorsalis, 392; posterior, 390
- ischiadicus, 400
- lachrymalis, 371
- laryngeus inferior, 380; superior, 380
- lingualis, 375
- lumboinguinalis, 396
- mandibularis, 370, 373
- massetericus, 374
- maxillaris, 370, 371
- medianus, 390
- mentalis, 375
- musculocutaneus, 388
- nasalis posterior, 372
- obturatorius, 397
- occipitalis major, 384
- oculomotorius, 352, 369
- olfactorius, 369
- ophthalmicus, 370
- opticus, 342, 354, 355,
369, 412
- palatinus major, 372; minor, 372
- patheticus (trochlearis), 350, 370
- peroneus communis, 400; profundus, 402; superficialis,
402
- petrosus profundus, 373, 405; superficialis major,
373, 375
- pharyngeus, 379
- phrenicus, 388
- plantaris lateralis, 403; medialis, 403
- pterygoideus, 374
- pudendus, 403
- radialis, 392; superficialis, 392
- saphenus, 397
- sphenopalatinus, 372
- splanchnicus major, 406; minor, 407
- subcutaneus malæ, 372
- suboccipitalis, 384
- subscapulares, 388
- suprascapularis, 388
- suralis, 400
- temporalis profundus, 374
- thoracicus anterior, 387; posterior,388
- tibialis, 402
- trifacialis, 370
- trigeminus, 347, 370
- trochlearis, 350, 370
- ulnaris, 393
- vagus, 346, 378
- vidianus, 372
- zygomaticus, 372, 377
- Neurapophysis, 4
- Nictitating membrane, 410
- Ninth cranial nerve, 347, 378
- Nipples, 268
- Nomenclature, v
- Nose, 243
- Notch, jugular, 24
- Number of bones, 1
- Oblique muscles of eye, 412
- Obliquus abdominis externus, M.[490], 153,
441; internus, M., 154, 441; capitis inferior, M.,
136, 443; superior, M., 136, 442,
443
- Obturator externus, M., 191, 444
- foramen, 76
- internus, M., 192, 444
- nerve, 397
- Occipital artery, 285
- bone, 22
- condyle, 23, 52, 55
- crest, external, 24, 52
- lobe, 358
- muscle, 101
- nerve, great, 384
- protuberance, 24
- tubercle, external, 52
- vein, 321
- Occipitofrontal muscle, 101
- Occipitoscapularis, M., 118, 436
- Oculomotor nerve, 352, 369
- Odontoid process, 14, 15
- Œsophageal arteries, 283
- veins, 318
- Œsophagus, 234
- Olecranon, 69
- fossa, 66
- Olfactory bulb, 341, 362
- fossa, 59
- mucosa, 245
- nerve, 369
- organ, 245, 426
- tract, 362
- Oliva, 345
- Omental sac, 219
- Omentum, gastrolienal, 220
- gastrosplenic, 220
- great, 219, 235
- lesser, 220
- Ophthalmic artery, 289
- ganglion, 369
- nerve, 370
- Opponens digiti quinti, M., 186, 215, 439
- Optic chiasma, 342, 354, 355, 369
- foramen, 29, 54, 59
- nerves, 342, 354, 355,
369, 412
- tract, 354, 355
- Ora serrata, 414
- Orbicularis oculi, M., 98, 435
- oris, M., 105, 435
- Orbit, 409
- muscles of, 411
- Orbital fissure, 27, 54, 59
- fossa, 53
- gland, 224
- gyrus, 360
- ligament, 47
- plate, 37
- process, 47
- Orbitosphenoid, 29
- Organon auditus, 415
- gustus, 427
- olfactus, 426
- visus, 409
- Os calcis, 83
- capitatum, 70
- coxæ, 76
- cuboideum, 82, 84
- cuneiforme primum, 85; secundum, 84; tertium, 84
- ethmoidale, 42
- frontale, 37
- hamatum, 70
- hyoideum, 49
- incisivum, 41
- innominatum, 76
- interparietale, 25
- lachrymale, 46
- lenticulare, 424
- magnum, 70
- multangulum majus, 71; minus, 71
- nasale, 42
- naviculare pedis, 84
- occipitale, 22
- palatinum, 45
- parietale, 36
- penis, 263
- pisiforme, 70
- presphenoidale, 29
- sacrum, 8
- scapholunaris, 70
- sphenoidale, 25
- temporale, 30
- triquetrum, 70
- uteri, 266
- zygomaticum, 47
- Ossa sesamoidea, 73, 80, 86
- Ossicula auditus, 423
- Ostium tubæ abdominale, 264
- Ovarian artery, 306
- Ovary, 263, 264
- Oviducts, 263, 264
- Pads of feet, 427
- Palate, hard, 56, 222
- muscles, 230; dissection, 451
- soft, 222, 229
- Palatine arteries, 290
- bones, 45
- canal, anterior, 41; posterior, 45, 46,
54, 56
- foramen, anterior, 57
- grooves, 41, 56
- nerve, greater, 372; lesser, 372
- process, 40
- Palatum durum, 56, 222
- Palmar arch, 300
- interosseæ, 300
- Palmaris longus, M., 179, 439
- Palpebræ[491], 409
- Palpebral vein, inferior, 321; superior,321
- Pancreas, 241
- Aselli, 333
- Pancreatic ducts, 242
- Pancreaticoduodenalis inferior, A., 303; superior, A., 303; V.,
326
- Papillæ of tongue, 227
- Papilla of kidney, 256
- Parameralis, M., 195
- Parietal bone, 36
- eminence, 36, 51
- layer of peritoneum, 217, 218
- tubercle, 36, 51
- Parker and Floyd’s mixture, 463
- Parotid duct, 222, 223
- gland, 223
- Patella, 80
- patellar ligament, 88
- surface of femur, 80
- Patheticus (trochlearis), N., 350, 370
- Pectineus, M., 200, 444
- Pectoantibrachialis, M., 145, 437
- Pectoralis major, M., 145, 437; minor, M., 146,
437
- Pectoral muscles, 144, 437
- ridge, 66
- Pedicle, 4
- Pedis perforatus, M., 205, 212
- Peduncles of cerebrum, 351
- Pedunculus cerebelli, 346
- cerebri, 351
- Pelvic arch, 76
- cavity, 218
- extremities, bones of, 76
- girdle, 76
- limbs, joints and ligaments of, 86
- muscles, 186; dissection, 444
- symphysis, 87
- Pelvis, bones of, 76
- of kidney, 256
- ligaments of, 86
- Penis, 257, 262
- arteries of, 308
- muscles of, 271
- nerves of, 403
- Pericardium, 279
- Perineum, 268
- Periorbita, 409
- Peripheral nervous system, 369; dissection, 465
- Peritoneal cavity, 218; lesser, 219
- Peritoneum, 218
- Peroneal groove, 84
- tubercle, 84
- Peroneus brevis, M., 210, 445
- communis, N., 400
- longus, M., 209, 445
- profundus, N., 402
- superficialis, N., 402
- tertius, M., 210, 445
- Pes, 85
- Petrosal ganglion, 378
- nerve, deep, 373, 405; great superficial, 373,
375
- sinus, inferior, 325
- Petrous portion of temporal bone, 33
- Peyer’s glands, 237, 238
- patches, 237, 238
- Phalanges, 72, 86; ligaments of, 76
- Pharyngeal nerve, 379
- Pharynx, 231
- muscles, 232; dissection, 451
- Phrenic artery, 304
- nerve, 388
- vein, 326
- Pia mater, 337, 369
- Pillars of fauces, 230
- fornix, 363, 364
- Pineal body, 351, 355
- Pinna, 415
- Pisiform bone, 70
- Pituitary body, 356
- Plantar arch, 314, 330
- ligaments, 92
- nerves, 403
- Plantaris, M., 205, 445
- Platysma, M., 95, 434
- Pleura, 217
- Plexus, anterior gastric, 382
- aortic, 408
- brachial, 386
- cardiac, 380, 382, 405
- carotid, 287
- cervical, 386
- cœliac, 406
- hepatic, 408
- inferior mesenteric, 408
- lumbar, 395
- lumbosacral, 396
- posterior gastric, 382
- pulmonary, 382
- renal, 408
- sacral, 399
- solar, 406
- splenic, 408
- superior mesenteric, 408
- suprarenal, 408
- Plica aryepiglottica, 246
- glossoepiglottica, 227, 246
- semilunaris, 410
- Pollex, 72
- Plicæ ciliares, 413
- Pons, 341, 347
- Popliteal artery, 311, 312
- bone, 206
- gland, 334
- notch, 81
- vein, 330
- Popliteus, M.[492], 206, 445
- Portal vein, 326
- Portio minor (of fifth nerve), 373
- Posterior auricular artery, 287
- circumflex artery, 296
- commissure, 355
- facial vein, 323
- inferior spine, 77
- perforated area, 352
- sulcus, 359
- Sylvian gyrus, 359
- thoracic nerve, 388
- tibial artery, 313
- Postglenoid process, 32
- Postmandibular process, 32, 33
- Postorbital process, 38
- Postzygapophysis, 4
- Poupart’s ligament, 153
- Practical directions, 429
- Præsylvian sulcus, 360
- Premaxilla, 41
- Premaxillary bone, 41
- Premolar teeth, 225
- Preparation of bones, 430
- ligaments, 431
- of viscera, 447
- Prepuce, 257, 262, 267
- Preservation of material, 431
- Preservatives for anatomical material, 431
- Presphenoid bone, 25, 29
- Prezygapophyses, 4
- Princeps pollicis et indicis, A., 300
- Profunda brachii, A., 296
- femoris, A., 309
- Process, accessory, 6, 8
- acromion, 62, 64
- alveolar, 39
- angular, 19, 48
- anterior clinoid, 29; superior, 78; transverse,
11
- articular, 4, 6, 8
- condyloid, 48
- coracoid, 63, 64
- coronoid, 48, 69
- frontal, 39, 40, 47, 51
- hæmal, 11
- hamular, 28
- jugular, 24, 52, 55
- lenticular, 424
- mammillary, 6, 8
- mastoid, 35, 55
- nasal, 41
- odontoid, 14, 15
- orbital, 47
- palatine, 40
- posterior clinoid, 26
- postglenoid, 32
- postmandibular, 32, 53
- postorbital, 38
- pseudotransverse, 8, 9
- pterygoid, 26, 28
- spinous, 3, 6, 8
- styliform, 33, 56
- transverse, 4, 6, 8, 12
- xiphoid, 20
- zygomatic, 31, 32, 38,
40, 47, 51
- Processus costarius, 13
- uncinatus, 417
- Promontory, 34
- Pronator muscles of forearm, 179
- quadratus, M., 183, 439
- teres, M., 179, 439
- Prosencephalon, 343, 344, 352
- Prostate gland, 261
- Protuberance, external occipital, 24
- Pseudotransverse process, 8, 9
- Psoas minor, M., 139, 446
- Pterygoid bone, 25
- canal, 27, 56, 59; nerve of,
372
- Pterygoideus externus, M., 111, 441; internus, M., 111,
441
- Pterygoid fossa, external, 30, 53; internal, 28,
54
- muscles, 111
- nerves, 374
- process, 26, 28
- Pterygopharyngeus, M., 233
- Pubic tubercle, 78
- Pubiocaudalis, M., 269
- Pubis, 76, 78
- Pudendus, N., 403
- Pulmonary artery, 275, 278, 280
- ligament, 253
- plexus, 382
- sinuses, 278
- veins, 275, 279, 315
- Pulmones, 252
- Pulvinar, 354
- Pupil, 413, 414
- Pyloric artery, 302
- valve, 235
- Pyramidal tracts, 345
- Pyramides, 345
- Pyriformis, M., 188, 444
- Quadratus femoris. M., 191, 44
- labii inferioris, M., 107, 435; superioris, M.,
105, 435
- lumborum, M., 139, 446
- plantæ, M., 213, 445
- Quadriceps femoris, M., 201, 444
- Radial artery, 297
- fossa, 66
- nerve, 392; superficial, 392
- notch, 69
- recurrent artery, 298
- sesamoid, 73
- Radio ulnar articulation, 75
- Radius[493], 67
- Radix, 4; of lungs, 252
- Rami of spinal nerves, 339
- Ramus of ischium, 78
- of mandible, 47, 48
- of pubis, 78
- Receptaculum chyli, 333; demonstration of, 460
- Rectocavernosus, M., 271
- Rectovaginalis, M., 272
- Rectovesical pouch, 266
- Rectum, 239; ligaments of, 219
- Rectus abominis, M., 155, 441
- capitis anterior major, M., 142; minor, 143,
442
- lateralis, M., 143, 442
- posterior major, M., 134, 443; medius, M.,
135, 443; minor, M., 135, 443
- femoris, M., 201, 444
- muscles of eye, 411
- Reference works, 429
- Removal of brain, 462
- Ren, 255
- Renal artery, 304
- plexus, 408
- veins, 326
- Respiratory nerve, internal, 388
- organs, 243; dissection, 452
- Retina, 414
- Retractor oculi, M., 411
- penis, M., 271
- Rhinencephalon, 362
- Rhombencephalon, 341, 344
- Rhomboideus, M., 119, 436
- capitis, M., 118
- Ribs, 18
- false, 19
- floating, 19
- true, 19
- Ridge deltoid, 66
- lambdoidal, 24, 50, 52
- pectoral, 66
- supracondyloid, 67
- Rolando, column of, 346
- Roots of spinal nerves, 338
- Rotator auris, M., 419, 470
- Rotatores, Mm., 131
- Round ligament of femur, 87, 220, 267
- Sacral arteries, 301, 309
- canal, 8
- foramina, 9, 10
- nerves, 399
- plexus, 399
- vein, middle, 328
- vertebræ, 1, 8
- Sacrum, 1, 8
- Sagittal crest, 25, 50
- sinus, 36, 324
- Salivary glands, 223
- Santorini, duct of, 242
- Saphenous artery, 311
- nerve, 397
- veins, 329, 330
- Sartorius, M., 197, 444
- Scalæ, 425
- Scala tympani, 425
- vestibuli, 425
- Scalenus, M., 141, 441
- Scapha, 416
- Scaphocuneiformis, M., 216, 445
- Scapholunar bone, 70
- Scaphoid bone, 69, 82, 84
- Scapula, 62; muscles of, 160
- Scarpa’s triangle, 310
- Sciatic nerve, great, 400
- notch, great, 77; lesser, 78
- Sclera, 413
- Sclerotic, 412, 413
- Scrotum, 257, 258
- Scutiform cartilage, 418
- Scutuloauricularis inferior, M., 419
- Second cranial nerve, 342, 354, 369,
412
- Sella turcica, 26, 59
- Semicircular canals, 425, 426; demonstration, 471
- Semilunar cartilages, 87
- ganglion, 347, 370, 406
- notch, 69
- tendon, 151
- valve, 278, 279
- Semimembranosus, M., 196, 444
- Semispinalis (meaning), 124
- capitis, M., 130, 132
- cervicis, M., 130, 133, 443
- Semitendinosus, M., 196, 444
- Sense-organs, 409; dissection, 469
- Serous covering, 219
- Serratus anterior, M., 122, 437
- Serratus posterior inferior, M., 148, 441; superior, M., 148,
441
- Sesamoid bones, 73, 80, 86
- Seventh cranial nerve, 374, 375
- Shoulder-joint, 73
- muscles of, 115, 156
- nerves of, 387
- Sight, organ of, 409
- Sigmoid cavity, great, 69
- gyrus, 360
- Sinus cavernosus, 325
- columnæ vertebralis, 325
- Sinuses, frontal, 38, 61
- of dura mater, 324
- sphenoidal, 29, 61
- Sinus of kidney, 255
- of vertebral column, 325
- petrosus inferior, 325
- sagittalis superior, 324
- transversus[494], 324
- Sixth cranial nerve, 347, 375
- Skeleton, 1; preparation, 430
- Skin, 427
- muscles, 93, 427; dissection, 434
- Skull, 21, 49
- disarticulation of, 430
- joints and ligaments of, 61
- sutures of, 61
- Small intestine, 236
- Smell, organ of, 426
- Soap-mixture for bones, 430
- Soft palate, 229; muscles of, 230
- Soleus, M., 205, 445
- Sole of foot, muscles of, 212
- Solar plexus, 406
- Solitary glands, 237
- Spermatic artery, external, 309; internal, 304
- cord, 258
- vein, internal, 326
- Sphenoid bone, 25
- Sphenoidal sinus, 29, 61
- Sphenopalatine artery, 290
- foramen, 46, 54
- ganglion, 372
- nerve, 372
- Sphincter ani externus, M., 268, 453; internus, M., 269,
454
- Spigelian lobe of liver, 241
- Spinal accessory nerve, 346, 382
- artery, anterior, 291
- column, 1
- cord, 335; dissection, 461
- Spinalis, 124
- dorsi, M., 124, 128, 129,
133, 443
- Spinal ganglia, 338
- nerves, 337, 383; dissection of, 467
- veins, 325
- Spine, frontal, 37
- maxillary, 45
- nasal, 37
- of helix, 417
- ischium, 77, 78
- scapula, 62, 63
- tibia, 81
- posterior inferior, 77
- posterior nasal, 45
- Spinodeltoid muscle, 156, 438
- Spinotrapezius, M., 115, 436
- Spinous processes, 3, 6, 8
- Spiral line or ridge, 79
- Splanchnic nerves, 406, 407
- Spleen, 242
- Splenial sulcus, 361, 362
- Splenic artery, 303
- plexus, 408
- Splenius, M., 131, 443
- Squama occipitalis, 22
- temporalis, 31
- Stapedius, M., 424, 471
- Stapes, 424
- Starch injection mass, 456
- Stenon’s duct, 223
- Steno’s duct, 222, 223
- Stenson’s duct, 222
- Sternocleidomastoid muscle, 120, 139
- Sternocostalis externus, M., 150
- internus, M., 151
- Sternohyoid muscle, 140, 440
- Sternomastoid muscle, 139, 440
- Sternothyroid muscle, 141, 440
- Sternum, 20
- Stomach, 234 arteries of, 303
- Striæ medullares, 349, 354
- Study, order of, 429
- Styliform process, 33, 56
- Styloglossus, M., 228, 440, 450
- Stylohyal, 49
- Stylohyoid muscle, 112, 440
- Styloid process, 68, 69
- Stylomandibular ligament, 62, 228
- Stylomastoid foramen, 33, 34, 52, 55
- Stylopharyngeus, M., 233, 451
- Subclavian artery, 282, 283, 290
- vein, 318
- Subcutaneus malæ nerve, 372
- Sublingual gland, 224
- Submaxillary gland, 223
- Submental artery, 286
- Submentalis, M., 103, 435
- Submental vein, 321, 323
- Suboccipital nerve, 384
- Subscapular artery, 294
- fossa, 63
- Subscapularis, M., 161, 437
- Subscapular nerves, 388
- Substantia perforata anterior, 362; posterior, 352
- Sulci of cerebellum, 348
- cerebrum, 359
- medulla oblongata, 345
- spinal cord, 335
- Sulcus ansatus, 360
- anterior, 359
- basilaris, 347
- callosalis, 362
- coronalis, 360
- coronarius, 274
- cruciatus, 360, 361
- ectosylvius, 359
- falcialis, 361
- hippocampal, 362
- hypothalamicus, 354
- intertubercularis, 66
- lateralis, 360
- marginalis, 361
- palatinus, 41, 56
- posterior, 359
- postrhinalis, 361
- præsylvius[495], 360
- rhinalis, 360; posterior, 361, 362
- splenialis, 361, 362
- supracallosalis, 362
- supraorbitalis, 360
- suprasylvius, 359
- Supercervicocutaneus, M., 95
- Superficial temporal artery, 287
- Superior cervical ganglion, 404
- Supinator, M., 177, 439; longus, M., 173
- Supracallosal sulcus, 362
- Supracondyloid foramen, 67
- ridge, 67
- Supraglenoidal tubercle, 64
- Supraoccipital bone, 22
- Supraorbital arch, 37, 51
- sulcus, 360
- Suprarenal body, 257
- plexus, 408
- Suprascapular artery, 293
- nerve, 388
- notch, 64
- Supraspinatus, M., 159, 438
- Supraspinous fossa, 63
- ligament, 16
- Suprasylvian gyrus, 360
- sulcus, 259
- Sural artery, 313
- nerve, 400
- Suspensory ligament, 219, 220, 257
- Sustentaculum tali, 84
- Sutures of skull, 61
- Sylvius, aqueduct of, 342, 343, 350,
351, 352
- fissure of, 358
- Sympathetic nervous system, 404; dissection, 465
- Symphysis of jaw, 47, 61
- menti, 47, 61
- pelvis, 76
- pubis, 76, 78
- Systema lymphaticum, 330
- nervorum sympatheticum, 404
- Tail, arteries of, 309
- muscles of, 136
- Tail muscles, dissection of, 447
- Tail, nerves of, 404
- veins of, 328
- vertebræ of, 11
- Talus, 82
- Tapetum, 413
- Tarsal glands, 409
- Tarsea medialis, A., 315
- Tarsus, 82
- joints and ligaments of, 91
- muscles of, 215
- Taste-buds, 427
- Taste, nerve of, 378
- organs of, 427
- Teeth, 224
- deciduous, 226
- Tegmen tympani, 35
- Telencephalon, 341, 343, 344, 357
- Temporal arteries, 287
- Temporal bone, 30
- Temporal fossa, 51, 53
- Temporal lobe, 358
- Temporal muscle, 110, 440
- Temporal nerve, deep, 374
- Temporal nerves, 373, 374, 377
- Temporal vein, superficial, 324
- Tendo calcaneus, 204, 206
- Tendon of Achilles, 204, 206
- Tensor fasciæ latæ M., 187, 444
- tympani, M., 424, 471
- veli palatini, M., 230, 451
- Tenth cranial nerve, 346, 378
- Tentorium, 36, 58
- Tenuissimus, M. 195, 444
- Teres major, M., 163, 437, 438
- Teres minor, M., 161, 438
- Terms of direction, vii
- Testes, 258, 259
- Thalamus, 352, 353
- Thebesius, valve of, 277
- Thigh, bones of, 79
- fascia of, 186
- muscles of, 194
- Third cranial nerves, 352, 369
- ventricle, 342, 343, 356
- Thoracic aorta, 281
- cavity, 217
- extremities, bones of, 62
- duct, 320, 331, 333 460
- ganglia, 406
- limbs, muscles of, 156; joints and ligaments, 73
- nerves, 393
- nerves, anterior, 387
- nerve, posterior, 388
- Thoracicodorsalis, A., 296
- Thoracic vertebræ, 1
- wall, muscles of, 148
- Thorax, bones of, 18
- lymphatics of, 333
- muscles of, 144
- Thumb, muscles of, 184
- Thymus gland, 254
- Thyroarytenoid muscle, 251
- Thyrocervical axis, 293
- Thyrohyal, 49
- Thyrohyoid muscle, 249
- Thyroid arteries, 284
- Thyroid cartilage, 247
- gland, 254
- Tibia, 80
- Tibial artery, anterior, 313
- Tibialis anterior, M., 211, 445; posterior, M., 208,
445
- Tibial nerve[496], 402
- recurrent artery, 314
- Toes, bones of, 86
- muscles of, 214
- Tongue, 226
- muscles of, 228; dissection, 450
- Tonsils, 230
- Tooth formula, 224
- Topographical terms, vii
- Trabeculae, 277
- carneæ, 278
- Trachea, 251
- Trachelomastoid muscle, 131
- Tracts, pyramidal, 345
- Tractus olfactorius, 362
- opticus, 354, 355
- postrhinalis, 361, 362
- transversus peduncularis, 351, 352
- Tragicus lateralis, M., 418, 435
- medialis, M., 421, 470
- Tragus, 417
- Transversa, V., 321
- colli, A., 293
- scapulæ, A., 293; V., 320
- Transverse costal facet, 4
- Transverse ligament, 18, 87, 91, 172
- process, 4, 6, 8, 12
- sinus, 324
- Transversospinalis, 124
- Transversus abdominis, M., 155, 441
- auriculæ, M., 420, 470
- costarum, M., 150, 441
- perinei, M., 269
- thoracis, M., 151, 441
- Trapezium, 71
- (brain), 345
- Trapezius, M., 115, 436
- Trapezoid, 71
- Triangle, Scarpa’s, 310
- Triangularis sterni, M., 151
- Triangular ligament, 221
- Triceps brachii, M., 166, 438
- suræ, M., 206
- Tricuspid valve, 277
- Trifacial nerve, 370
- Trigeminal nerve, 347, 370
- Triquetrum, os, 70
- Trochanter, great, 79
- Trochanteric fossa, 79
- Trochanter, lesser, 79
- Trochlea (astragalus), 83
- (humerus), 66
- Trochlearis nerve, 350, 370
- True ribs, 19
- Truncus costocervicalis, 292
- thyrocervicalis, 293
- Tuba auditiva, 56, 423
- Tuber cinereum, 355
- Tubercle, external occipital, 52
- peroneal, 84
- pubic, 78
- of rib, 19
- Tubercular facet, 4
- Tuberculum anterius, 14
- articulare, 32
- sellæ, 26
- Tunica albuginea, 260
- dartos, 258
- fibrosa, 255
- vaginalis communis, 258; propria, 258
- Tuberosity, great, 65
- lesser, 65
- of ischium, 78
- of scapula, 63
- of tibia, 80
- Turbinals, 40, 42
- Twelfth cranial nerve, 346, 383
- ’Tween brain, 342, 352
- Tympanic bone, 31, 32
- bulla, 31, 32, 55, 422
- cavity, 422
- membrane, 422
- Tympanohyal, 49
- Ulna, 68
- Ulnar artery, 300
- nerve, 393
- recurrent artery, 298
- Umbilical artery, 307
- vein, 329; fœtal, 220
- Unciform bone, 70
- Ureter, 256
- Urethra, 257, 261; muscles of, 271
- Urethralis, M., 273
- Urogenital organs, 255; dissection of, 453; muscles of, 268
- sinus, 263, 267
- system, 255
- Uterine artery, 308
- tubes, 263, 264
- Uterus, 263, 266
- Uvea, 414
- Vagina, 263, 267
- Vaginal ligaments, 173
- Vagina, muscles of, 272
- Vagus, N., 346, 378
- Valsalva, sinuses of, 278, 279
- Variations, 328, 434, 456
- Vertebral muscles, 123, 138; dissection, 442,
446
- Vasa efferentia testis, 260
- Vas deferens, 260
- Vastus intermedius, M., 203, 445
- lateralis, M., 201, 445
- medialis, M., 202, 445
- Vater, ampulla of, 236
- Vein, see Vena
- Veins, 315
- of abdomen[497], 326
- arm, 318
- brain, 324
- face, 321
- foot, 329
- heart, 315
- larynx, 321
- leg, 329
- lips, 321
- liver, 326
- spinal cord, 325
- Velum medullare anterius, 350, 351; posterius, 344,
349
- palatinum, 222, 229; muscles of, 230
- Venæ, 315
- Vena adrenolumbalis, 326
- angularis, 321
- anonyma, 318
- auricularis anterior, 324; magna, 324; posterior,
324
- axillaris, 318
- azygos, 316
- brachialis, 318
- cardinales, 328
- cava inferior, 274, 275, 325;
superior, 274, 275, 316
- cephalica, 319
- cerebri inferior, 325; magna, 324
- cervicalis ascendens, 320
- cordis magna, 316
- coronaria ventriculi, 326
- costocervicalis 318
- facialis anterior, 331; posterior, 323;
- profunda, 323
- frontalis, 321
- gastroepiploica, 327
- gastrolienalis, 327
- glutea inferior, 329
- hemorrhoidalis media, 329
- hepatica, 326
- hypogastrica, 328, 329
- ilia cacommunis, 328; externa, 328, 329;
interna, 328, 329
- iliolumbalis, 326
- intercostales, 316
- jugularis externa, 319; interna, 320
- lingualis, 323
- lumbalis, 326
- mammaria interna, 318
- mediana cubiti, 319
- mesenterica inferior, 328; superior, 328
- nasalis externa, 321
- occipitalis, 321
- pancreaticoduodenalis, 326
- phrenica, 326
- poplitea, 330
- portæ, 326
- pulmonales, 275, 279, 315
- renalis, 326
- sacralis media, 328
- saphena magna, 329; parva, 329, 330
- spermatica interna, 326
- subclavia, 318
- submentalis, 321, 323
- temporalis superficialis, 324
- transversa, 321
- transversa scapulæ, 320
- umbilicalis, 329
- vertebralis, 318
- Venous sinuses of dura mater, 324
- Ventral arch (atlas), 14
- Ventricles of brain, 340, 362; of heart, 274,
277
- Ventriculi dorsalis, A., 303
- Ventriculus, 234, 246
- Vermis, 348
- Vertebræ, 1
- caudales, 11
- cervicales, 11
- lumbales, 7
- sacrales, 8
- thoracales, 1
- Vertebral arch, 3
- artery, 291
- column, 1; ligaments of, 16; muscles of, 123,
138
- foramen, 1
- vein, 318
- Vertebrarterial canal, 291
- Veru montanum, 261
- Vesical arteries, 307
- Vesica urinaria, 256
- Vestibule (ear), 425
- (larynx), 246
- (mouth), 221
- Vestibulum oris, 221
- of uterine tube, 264
- Vibrissæ, 427
- Vidian nerve, 372
- Villi, 236, 237
- Viscera, 217
- Visceral skeleton, 21
- Vitreous body, 415
- humor, 415
- Vocal cords, 246, 249
- false, 246
- Vomer, 44
- Vomeronasal organ, 222, 244
- Vulva, 267
- Wharton’s duct, 223
- White matter, 336
- zone of eye, 413
- Wilder and Gage’s soap mixture for bones, 430
- Willis, circle of, 289, 292
- Winslow, foramen of, 220
- Wirsung, duct of, 242
- Wrist[498], 75; bones of, 69
- Xiphihumeralis, M., 147, 437
- Xiphoid process, 20
- Zonula Arnoldi, 345
- ciliaris, 414
- Zinni, 414
- Zygomatic arch, 47, 49, 51, 52
- bone, 47
- nerves, 372, 377
- process, 31, 32, 38, 40,
47, 51
- Zygomaticus (major), M., 101, 435; minor, M., 105,
435
Transcriber’s Notes
Inconsistent spelling, hyphenation, formatting, etc. have not been standardised, except as mentioned under Changes.
The differences (in wording and structure) between the Table of Contents and the headings in the text have not been rectified.
The inconsistencies in the use, numbering and typography of section headings in the text have not been standardised, except as
mentioned below. There is an entry for Muscles of the Soft Palate in the Table of Contents, but no such sub-section heading in
the text.
p. vii, In compounding these terms of direction ...: the hyphenation does not always comply with this statement.
p. 372, N. canalis pterygoidii: possibly an error for N. canalis pterygoidei.
Changes made:
In several instances, the ditto symbol has been replaced by the dittoed text.
Some minor obvious typographical errors and missing punctuation have been corrected silently.
p. v: Leop-Carol. changed to Leop.-Carol
p. xv: entry Index added to Table of Contents
p. 142: (Fig. 72, c″) changed to (Fig. 72, c)
p. 293: Truncus thyreocervicalis changed to Truncus thyrocervicalis
p. 356: corpu callosum changed to corpus callosum
p. 475: Brachium conjuctivum changed to Brachium conjunctivum
p. 481: entry Glottis moved to proper place
p. 484: (Ligamentum) cruciata changed to (Ligamentum) cruciatum
p. 487: palbebræ superioris changed to palpebræ superioris
p. 492: Postzygapophyses changed to Postzygapophysis
End of the Project Gutenberg EBook of Anatomy of the Cat, by
Jacob Reighard and H. S. Jennings
*** END OF THIS PROJECT GUTENBERG EBOOK ANATOMY OF THE CAT ***
***** This file should be named 58394-h.htm or 58394-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/5/8/3/9/58394/
Produced by deaurider, Harry Lamé and the Online Distributed
Proofreading Team at http://www.pgdp.net (This file was
produced from images generously made available by The
Internet Archive)
Updated editions will replace the previous one--the old editions will
be renamed.
Creating the works from print editions not protected by U.S. copyright
law means that no one owns a United States copyright in these works,
so the Foundation (and you!) can copy and distribute it in the United
States without permission and without paying copyright
royalties. Special rules, set forth in the General Terms of Use part
of this license, apply to copying and distributing Project
Gutenberg-tm electronic works to protect the PROJECT GUTENBERG-tm
concept and trademark. Project Gutenberg is a registered trademark,
and may not be used if you charge for the eBooks, unless you receive
specific permission. If you do not charge anything for copies of this
eBook, complying with the rules is very easy. You may use this eBook
for nearly any purpose such as creation of derivative works, reports,
performances and research. They may be modified and printed and given
away--you may do practically ANYTHING in the United States with eBooks
not protected by U.S. copyright law. Redistribution is subject to the
trademark license, especially commercial redistribution.
START: FULL LICENSE
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full
Project Gutenberg-tm License available with this file or online at
www.gutenberg.org/license.
Section 1. General Terms of Use and Redistributing Project
Gutenberg-tm electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or
destroy all copies of Project Gutenberg-tm electronic works in your
possession. If you paid a fee for obtaining a copy of or access to a
Project Gutenberg-tm electronic work and you do not agree to be bound
by the terms of this agreement, you may obtain a refund from the
person or entity to whom you paid the fee as set forth in paragraph
1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this
agreement and help preserve free future access to Project Gutenberg-tm
electronic works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the
Foundation" or PGLAF), owns a compilation copyright in the collection
of Project Gutenberg-tm electronic works. Nearly all the individual
works in the collection are in the public domain in the United
States. If an individual work is unprotected by copyright law in the
United States and you are located in the United States, we do not
claim a right to prevent you from copying, distributing, performing,
displaying or creating derivative works based on the work as long as
all references to Project Gutenberg are removed. Of course, we hope
that you will support the Project Gutenberg-tm mission of promoting
free access to electronic works by freely sharing Project Gutenberg-tm
works in compliance with the terms of this agreement for keeping the
Project Gutenberg-tm name associated with the work. You can easily
comply with the terms of this agreement by keeping this work in the
same format with its attached full Project Gutenberg-tm License when
you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are
in a constant state of change. If you are outside the United States,
check the laws of your country in addition to the terms of this
agreement before downloading, copying, displaying, performing,
distributing or creating derivative works based on this work or any
other Project Gutenberg-tm work. The Foundation makes no
representations concerning the copyright status of any work in any
country outside the United States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other
immediate access to, the full Project Gutenberg-tm License must appear
prominently whenever any copy of a Project Gutenberg-tm work (any work
on which the phrase "Project Gutenberg" appears, or with which the
phrase "Project Gutenberg" is associated) is accessed, displayed,
performed, viewed, copied or distributed:
This eBook is for the use of anyone anywhere in the United States and
most other parts of the world at no cost and with almost no
restrictions whatsoever. You may copy it, give it away or re-use it
under the terms of the Project Gutenberg License included with this
eBook or online at www.gutenberg.org. If you are not located in the
United States, you'll have to check the laws of the country where you
are located before using this ebook.
1.E.2. If an individual Project Gutenberg-tm electronic work is
derived from texts not protected by U.S. copyright law (does not
contain a notice indicating that it is posted with permission of the
copyright holder), the work can be copied and distributed to anyone in
the United States without paying any fees or charges. If you are
redistributing or providing access to a work with the phrase "Project
Gutenberg" associated with or appearing on the work, you must comply
either with the requirements of paragraphs 1.E.1 through 1.E.7 or
obtain permission for the use of the work and the Project Gutenberg-tm
trademark as set forth in paragraphs 1.E.8 or 1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any
additional terms imposed by the copyright holder. Additional terms
will be linked to the Project Gutenberg-tm License for all works
posted with the permission of the copyright holder found at the
beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including
any word processing or hypertext form. However, if you provide access
to or distribute copies of a Project Gutenberg-tm work in a format
other than "Plain Vanilla ASCII" or other format used in the official
version posted on the official Project Gutenberg-tm web site
(www.gutenberg.org), you must, at no additional cost, fee or expense
to the user, provide a copy, a means of exporting a copy, or a means
of obtaining a copy upon request, of the work in its original "Plain
Vanilla ASCII" or other form. Any alternate format must include the
full Project Gutenberg-tm License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works
provided that
* You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is owed
to the owner of the Project Gutenberg-tm trademark, but he has
agreed to donate royalties under this paragraph to the Project
Gutenberg Literary Archive Foundation. Royalty payments must be paid
within 60 days following each date on which you prepare (or are
legally required to prepare) your periodic tax returns. Royalty
payments should be clearly marked as such and sent to the Project
Gutenberg Literary Archive Foundation at the address specified in
Section 4, "Information about donations to the Project Gutenberg
Literary Archive Foundation."
* You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or destroy all
copies of the works possessed in a physical medium and discontinue
all use of and all access to other copies of Project Gutenberg-tm
works.
* You provide, in accordance with paragraph 1.F.3, a full refund of
any money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days of
receipt of the work.
* You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project
Gutenberg-tm electronic work or group of works on different terms than
are set forth in this agreement, you must obtain permission in writing
from both the Project Gutenberg Literary Archive Foundation and The
Project Gutenberg Trademark LLC, the owner of the Project Gutenberg-tm
trademark. Contact the Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
works not protected by U.S. copyright law in creating the Project
Gutenberg-tm collection. Despite these efforts, Project Gutenberg-tm
electronic works, and the medium on which they may be stored, may
contain "Defects," such as, but not limited to, incomplete, inaccurate
or corrupt data, transcription errors, a copyright or other
intellectual property infringement, a defective or damaged disk or
other medium, a computer virus, or computer codes that damage or
cannot be read by your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium
with your written explanation. The person or entity that provided you
with the defective work may elect to provide a replacement copy in
lieu of a refund. If you received the work electronically, the person
or entity providing it to you may choose to give you a second
opportunity to receive the work electronically in lieu of a refund. If
the second copy is also defective, you may demand a refund in writing
without further opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS', WITH NO
OTHER WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT
LIMITED TO WARRANTIES OF MERCHANTABILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of
damages. If any disclaimer or limitation set forth in this agreement
violates the law of the state applicable to this agreement, the
agreement shall be interpreted to make the maximum disclaimer or
limitation permitted by the applicable state law. The invalidity or
unenforceability of any provision of this agreement shall not void the
remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in
accordance with this agreement, and any volunteers associated with the
production, promotion and distribution of Project Gutenberg-tm
electronic works, harmless from all liability, costs and expenses,
including legal fees, that arise directly or indirectly from any of
the following which you do or cause to occur: (a) distribution of this
or any Project Gutenberg-tm work, (b) alteration, modification, or
additions or deletions to any Project Gutenberg-tm work, and (c) any
Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of
computers including obsolete, old, middle-aged and new computers. It
exists because of the efforts of hundreds of volunteers and donations
from people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future
generations. To learn more about the Project Gutenberg Literary
Archive Foundation and how your efforts and donations can help, see
Sections 3 and 4 and the Foundation information page at
www.gutenberg.org
Section 3. Information about the Project Gutenberg Literary Archive Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg Literary
Archive Foundation are tax deductible to the full extent permitted by
U.S. federal laws and your state's laws.
The Foundation's principal office is in Fairbanks, Alaska, with the
mailing address: PO Box 750175, Fairbanks, AK 99775, but its
volunteers and employees are scattered throughout numerous
locations. Its business office is located at 809 North 1500 West, Salt
Lake City, UT 84116, (801) 596-1887. Email contact links and up to
date contact information can be found at the Foundation's web site and
official page at www.gutenberg.org/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
[email protected]
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To SEND
DONATIONS or determine the status of compliance for any particular
state visit www.gutenberg.org/donate
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations. To
donate, please visit: www.gutenberg.org/donate
Section 5. General Information About Project Gutenberg-tm electronic works.
Professor Michael S. Hart was the originator of the Project
Gutenberg-tm concept of a library of electronic works that could be
freely shared with anyone. For forty years, he produced and
distributed Project Gutenberg-tm eBooks with only a loose network of
volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as not protected by copyright in
the U.S. unless a copyright notice is included. Thus, we do not
necessarily keep eBooks in compliance with any particular paper
edition.
Most people start at our Web site which has the main PG search
facility: www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.
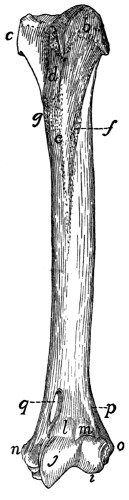
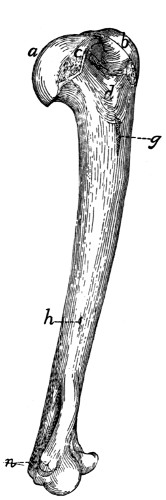
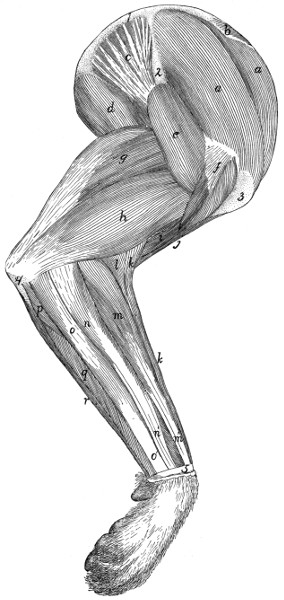
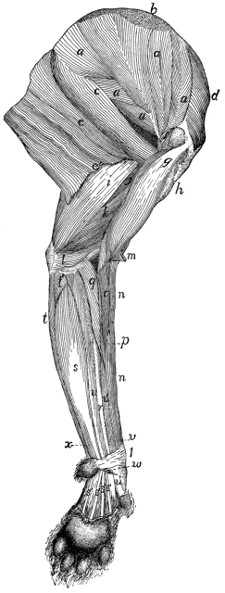
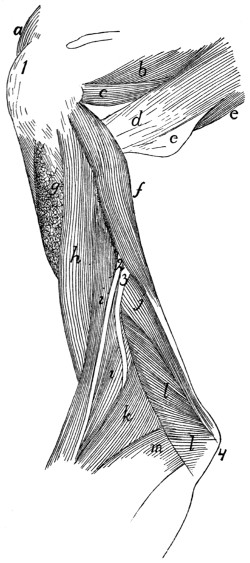
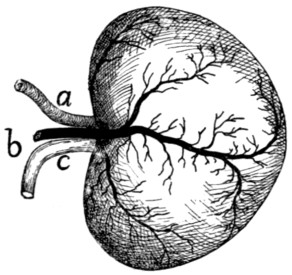
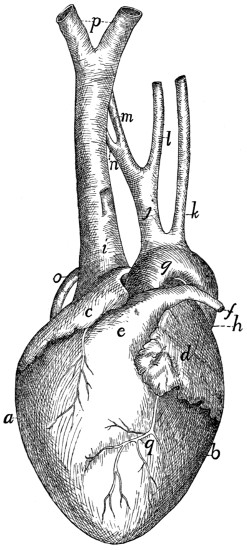
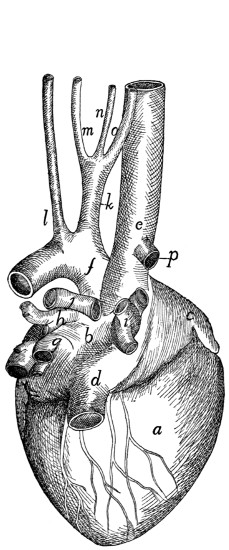
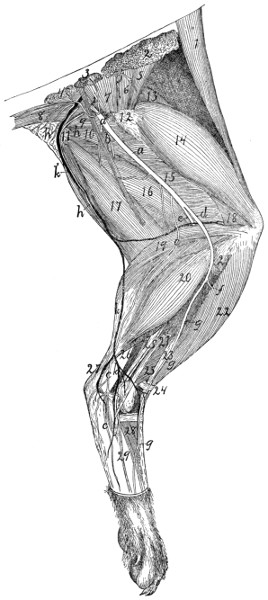
{kind=link}
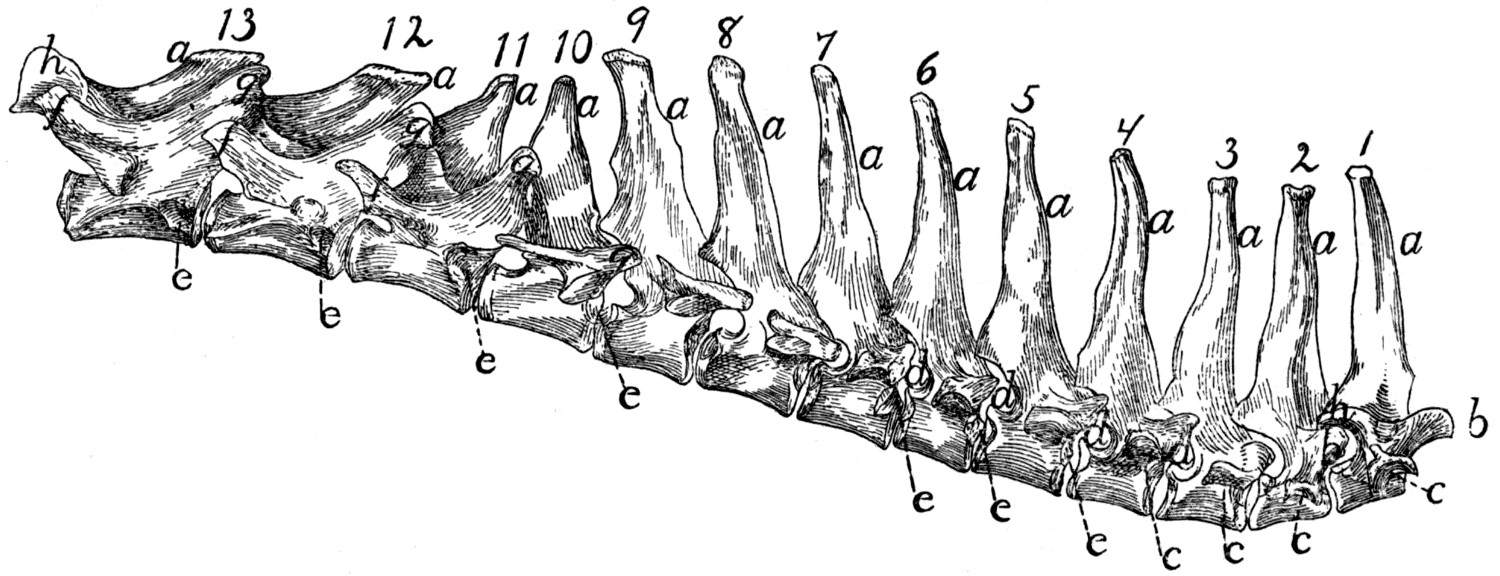{kind=link}
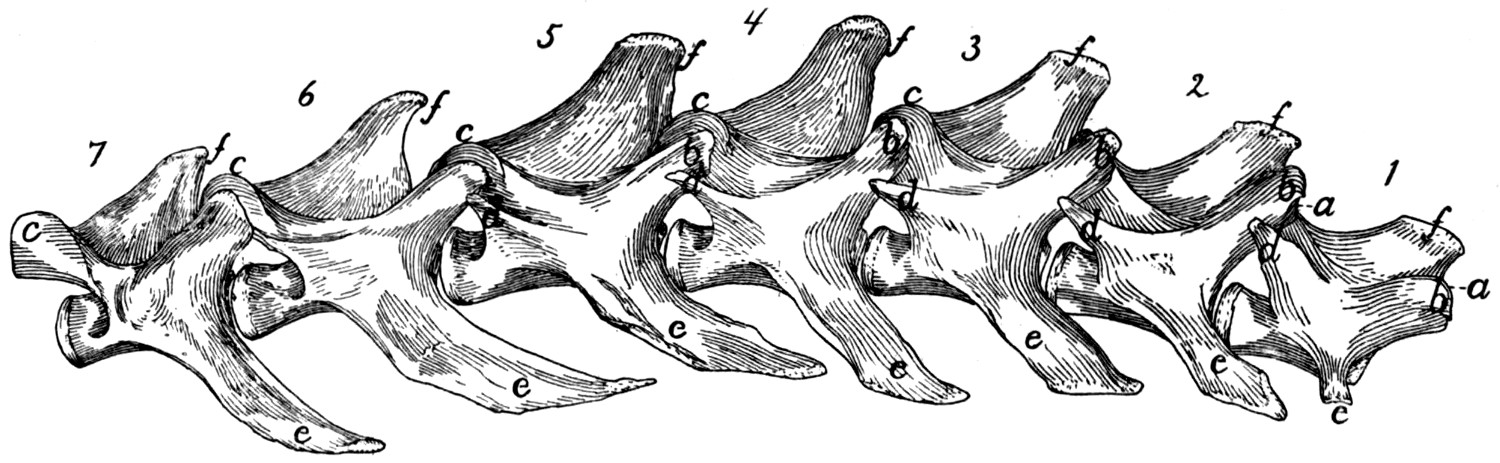{kind=link}
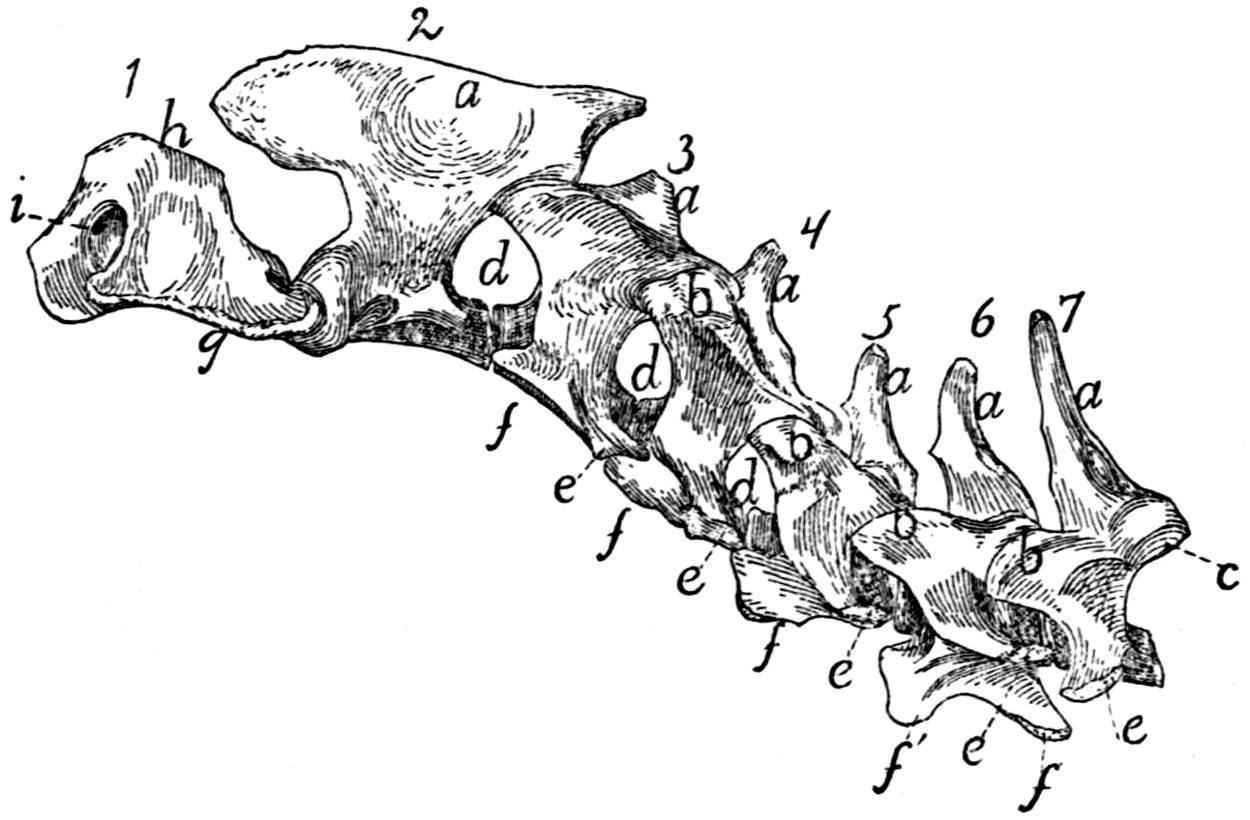{kind=link}
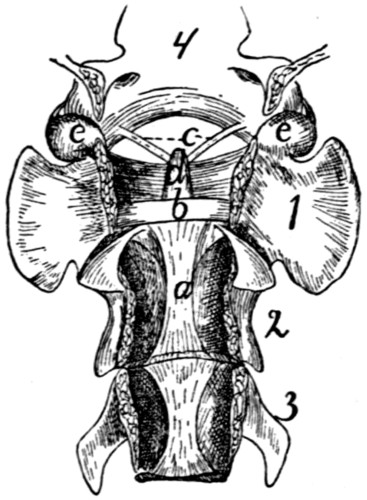{kind=link}
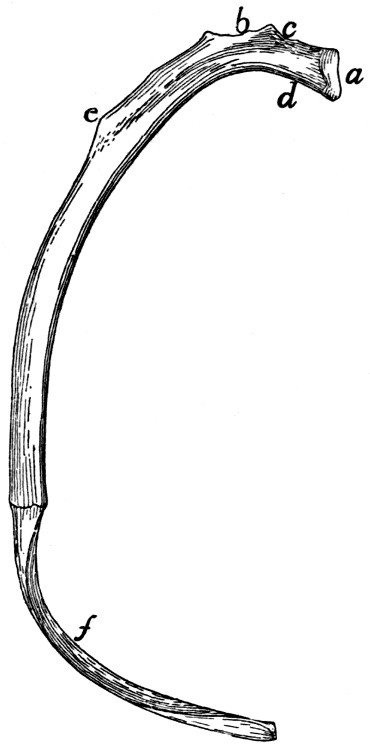{kind=link}
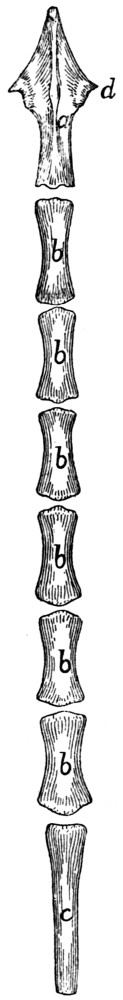{kind=link}
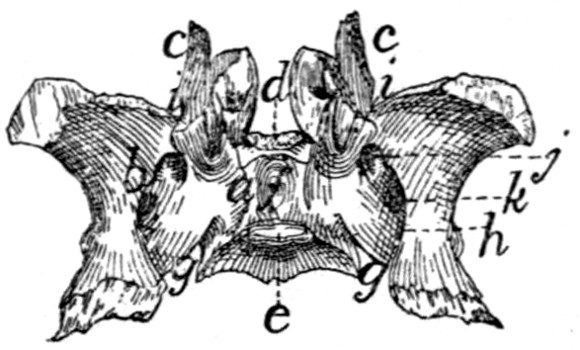{kind=link}
{kind=link}
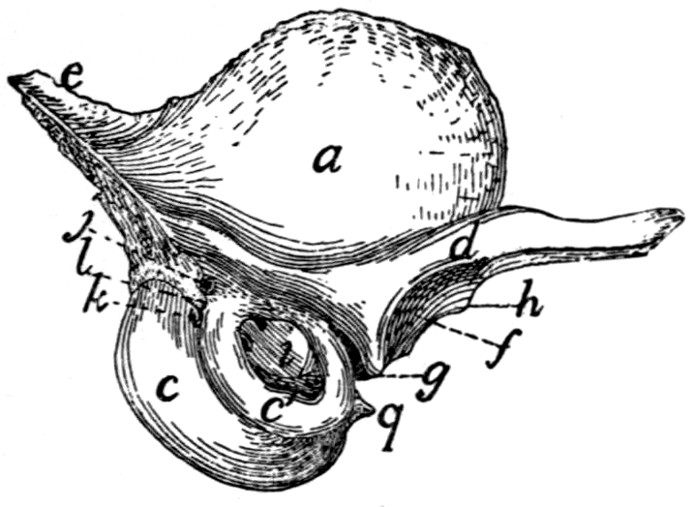{kind=link}
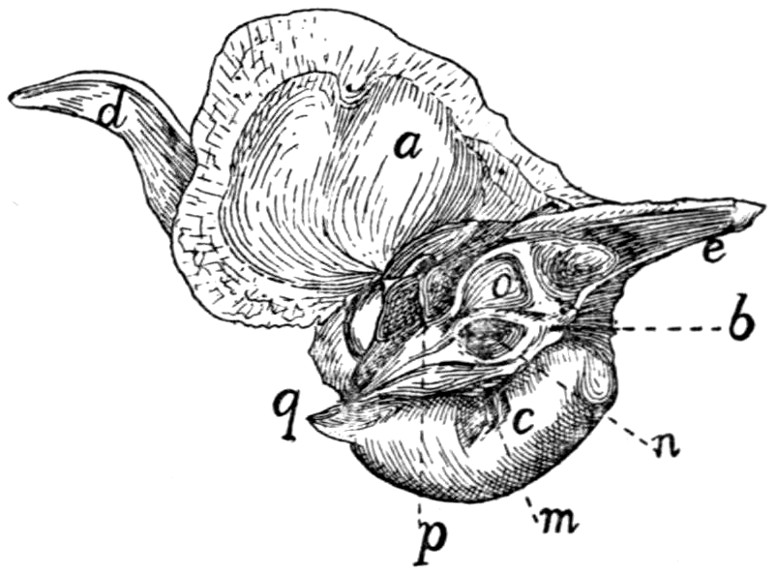{kind=link}
{kind=link}
{kind=link}
{kind=link}
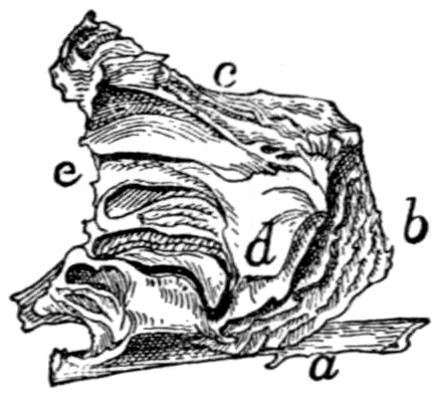{kind=link}
{kind=link}
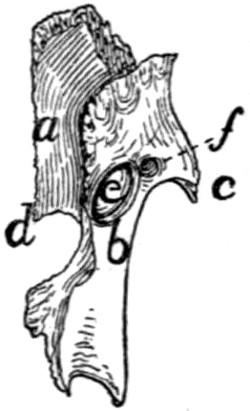{kind=link}
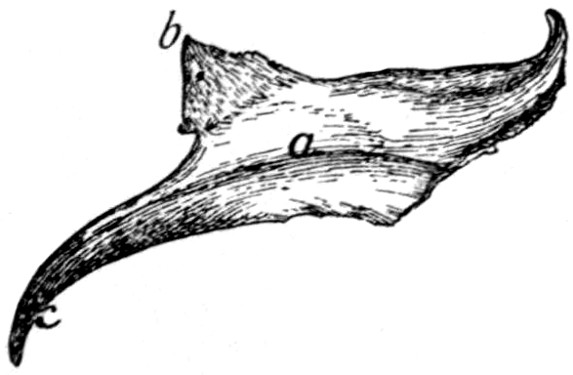{kind=link}
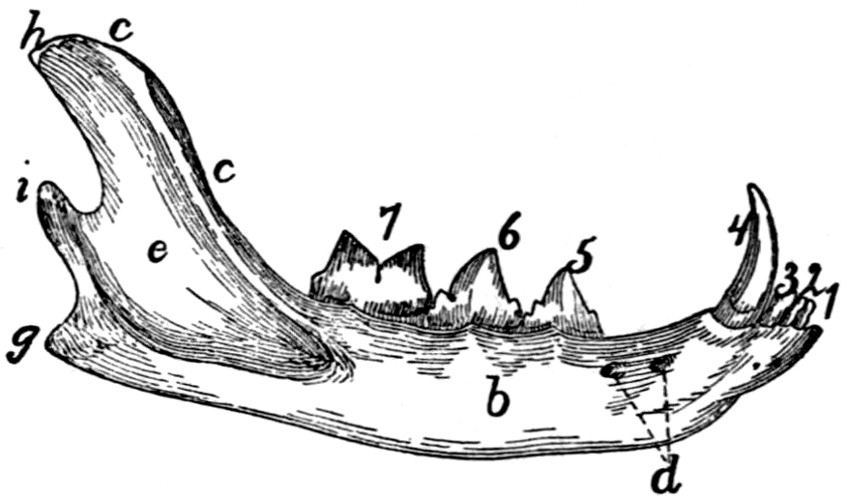{kind=link}
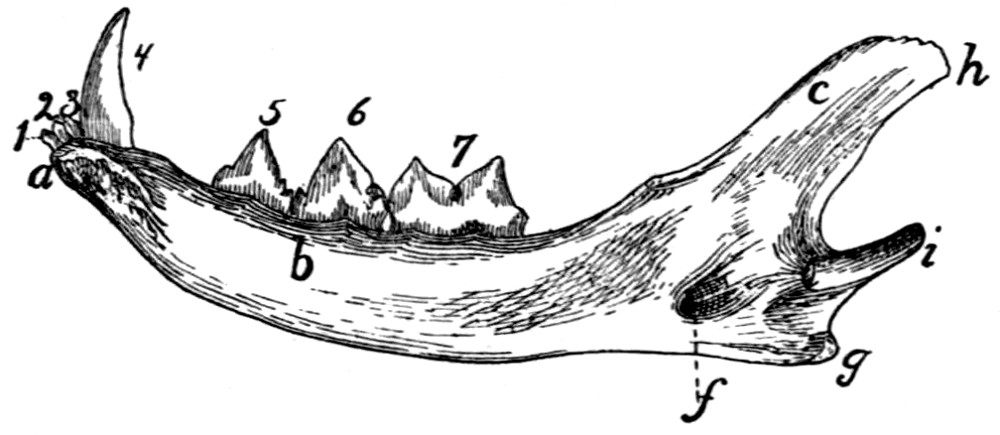{kind=link}
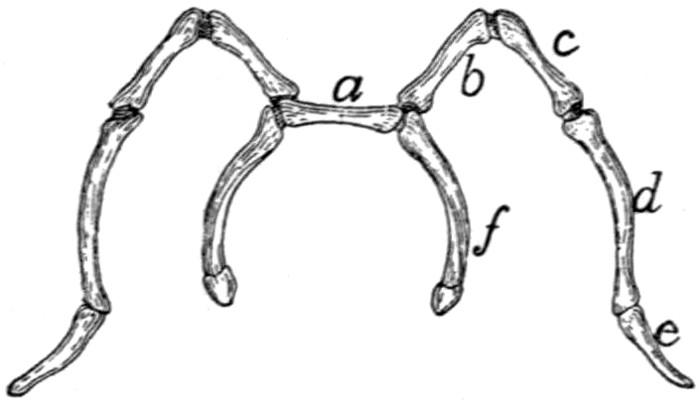{kind=link}
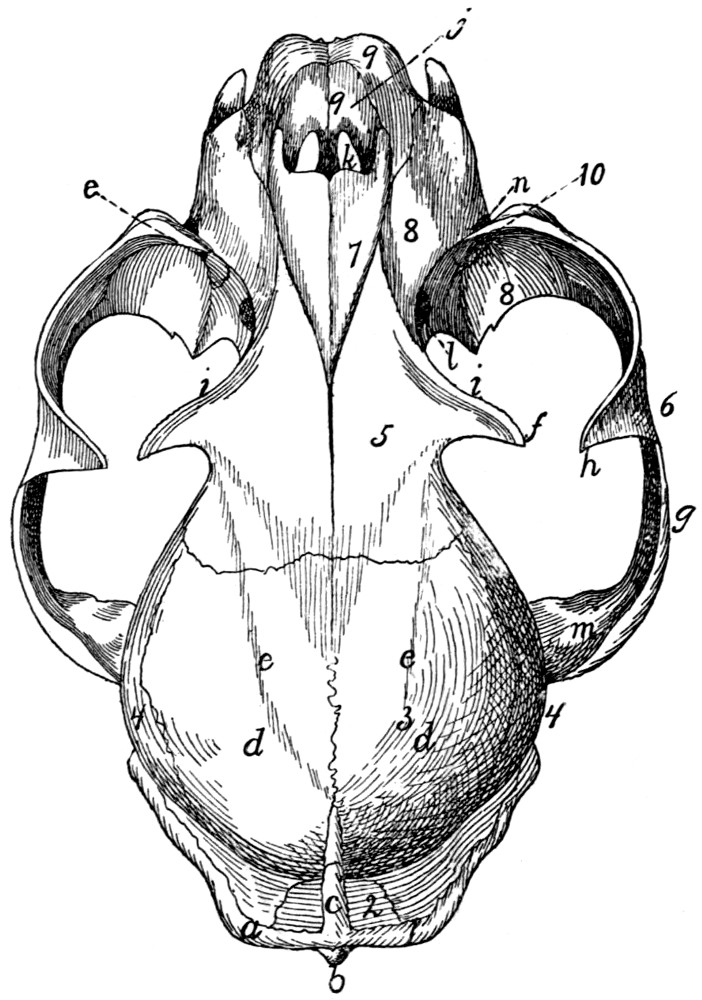{kind=link}
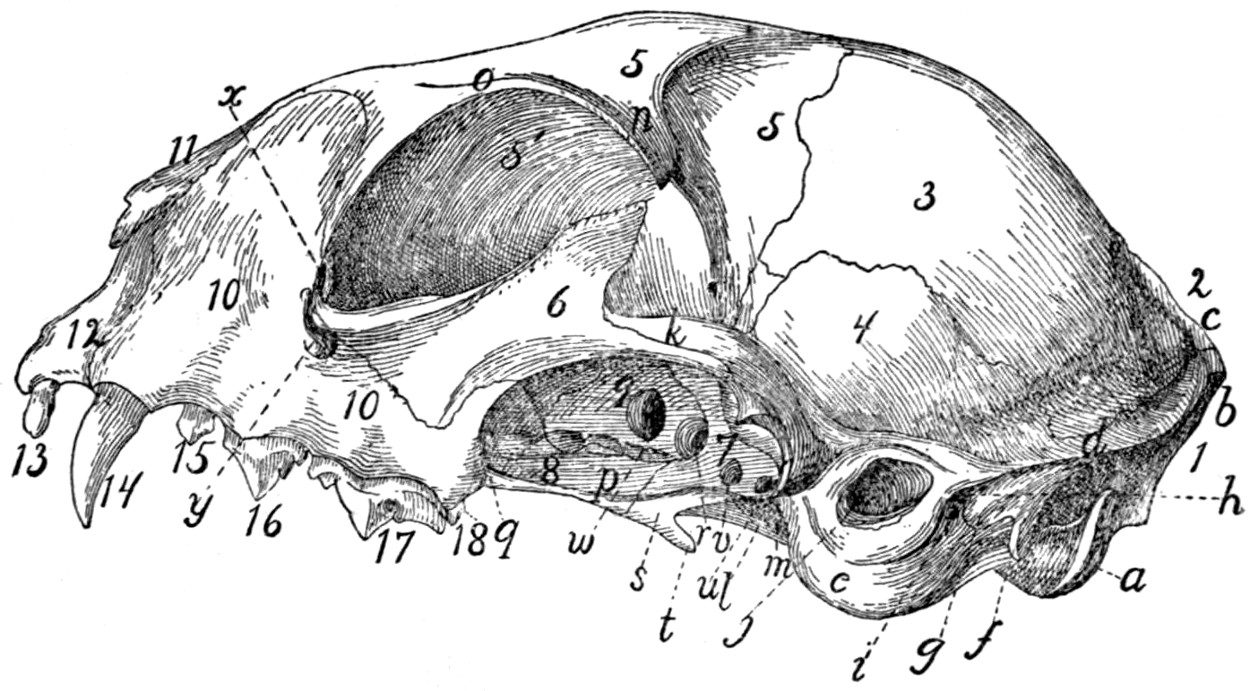{kind=link}
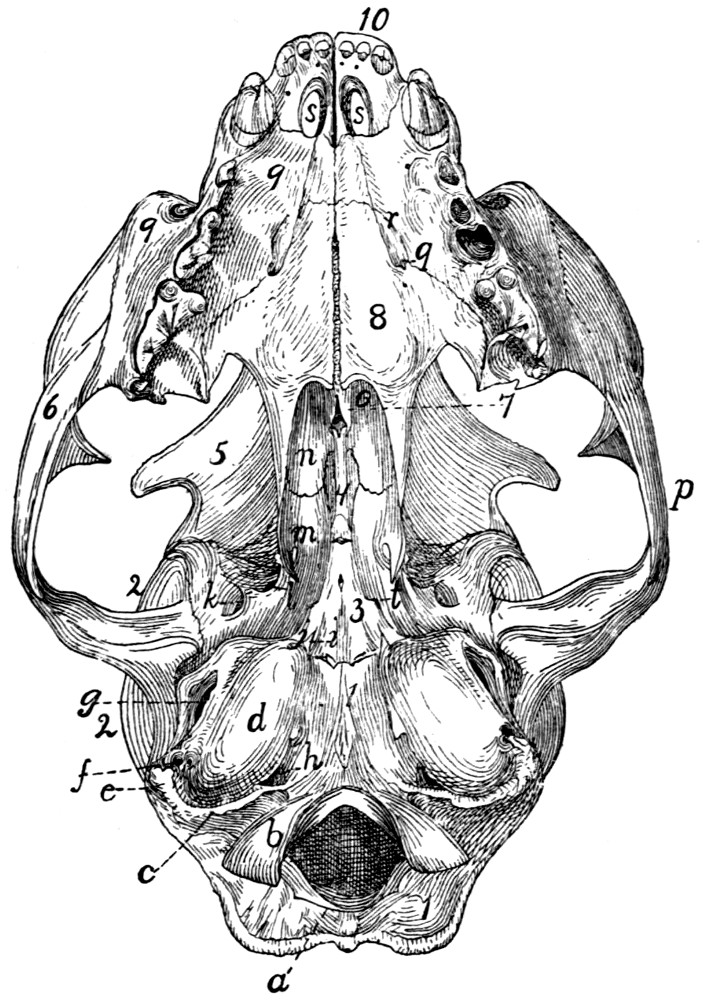{kind=link}
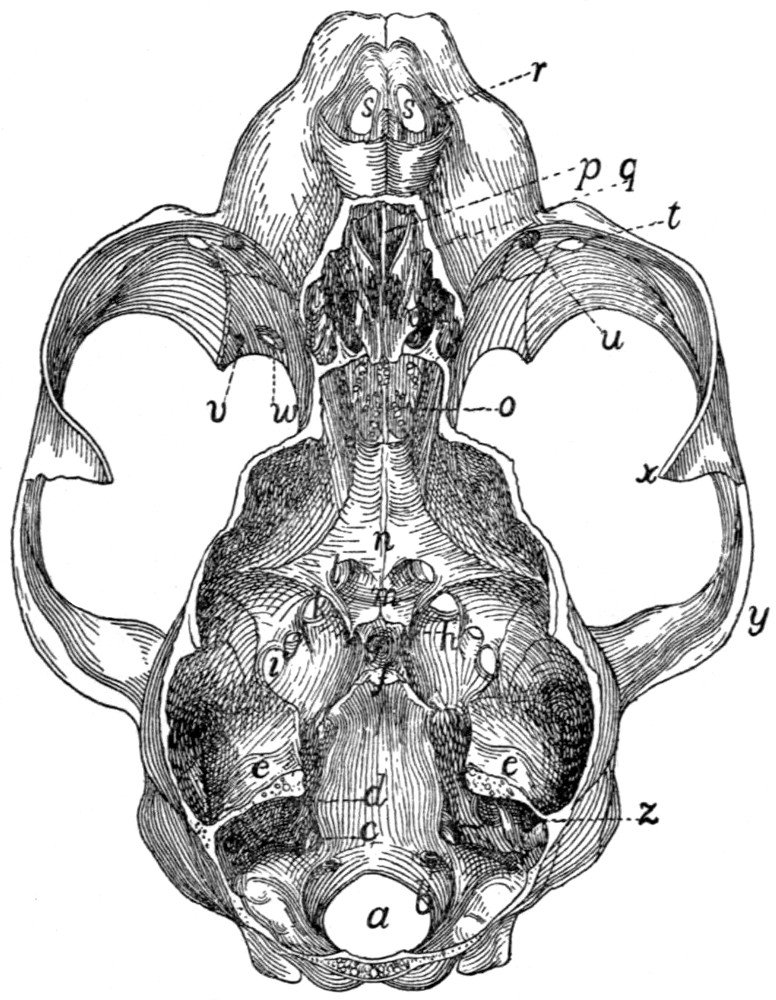{kind=link}
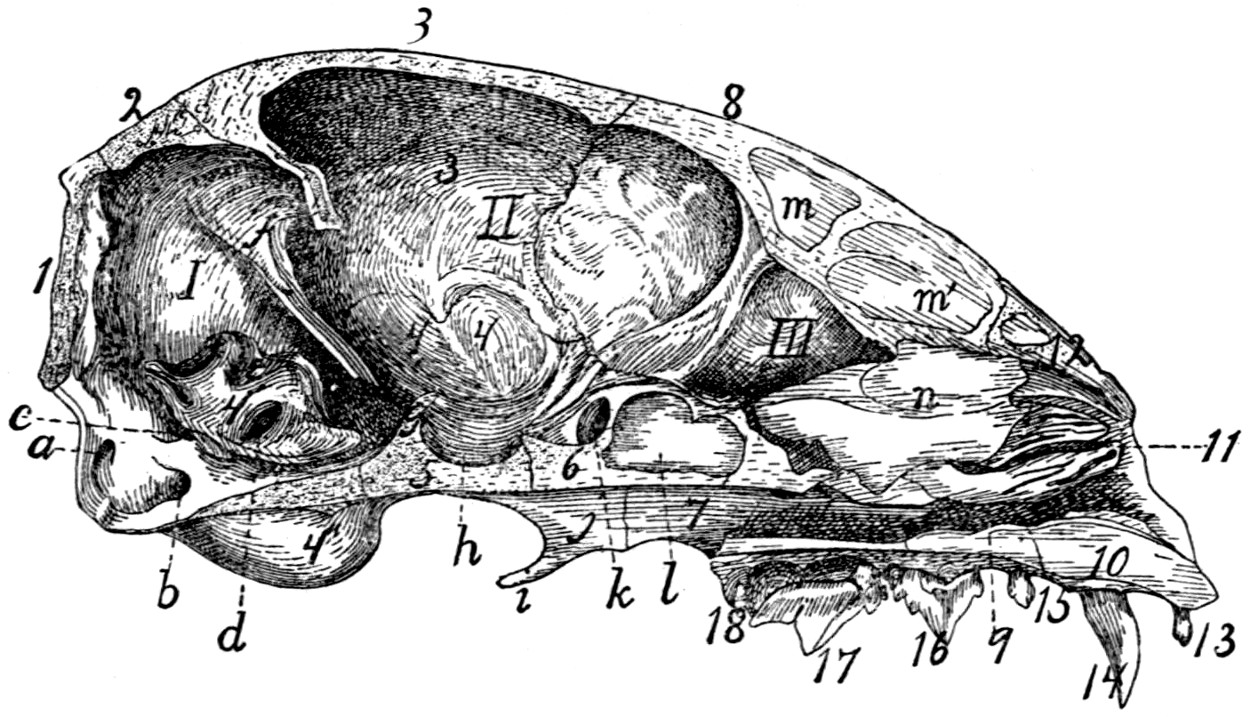{kind=link}
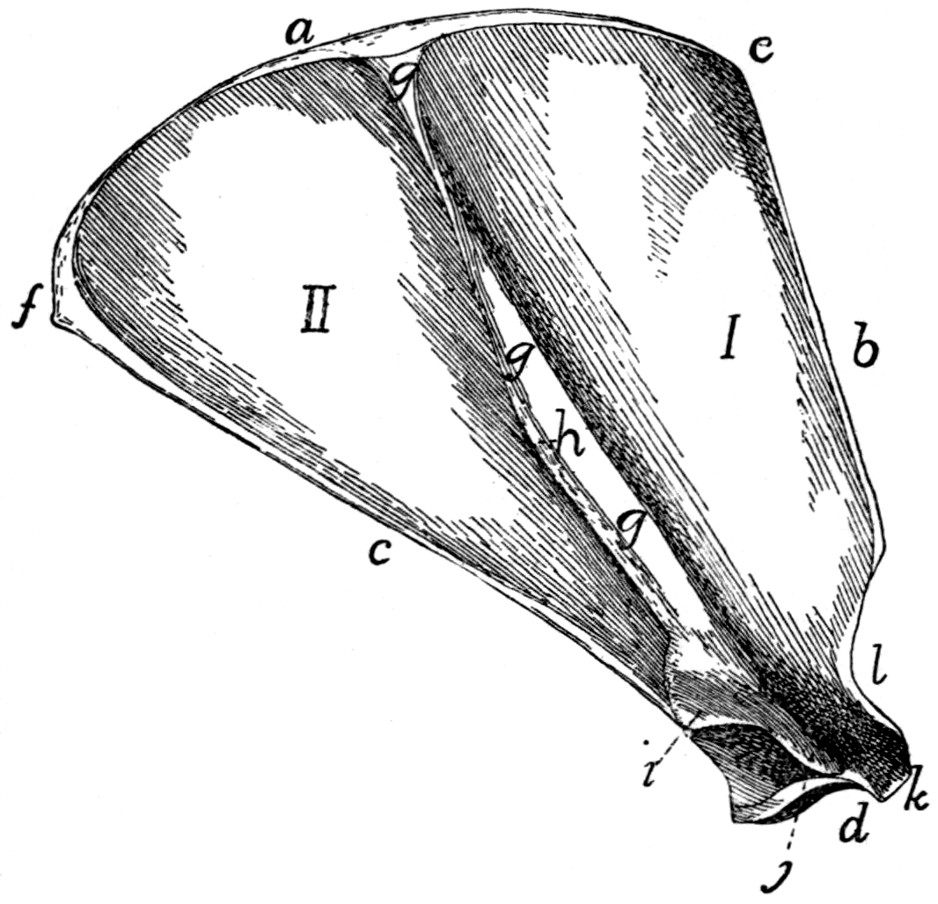{kind=link}
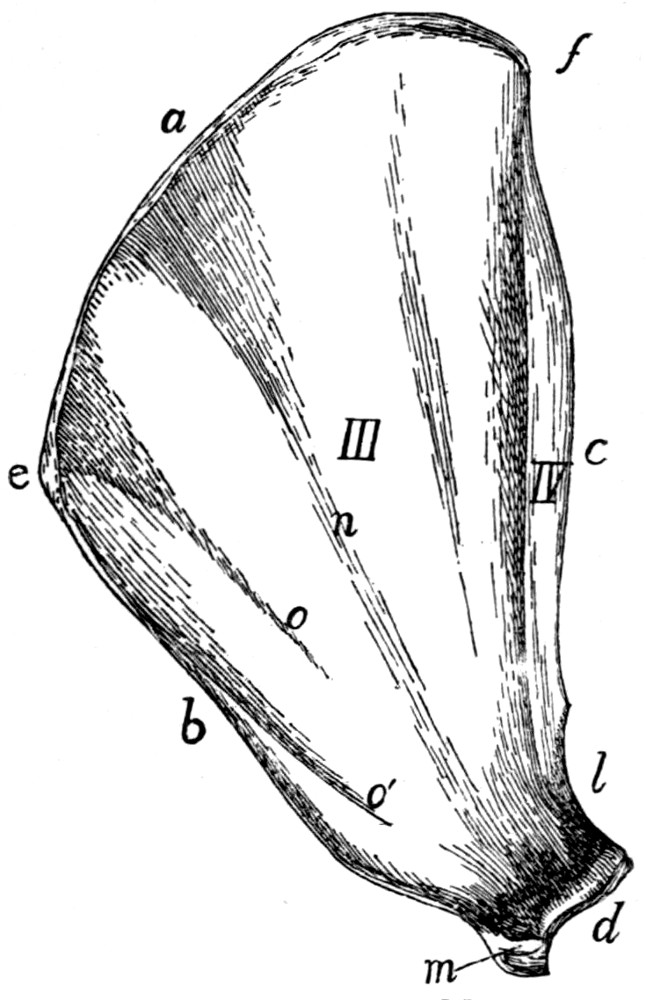{kind=link}
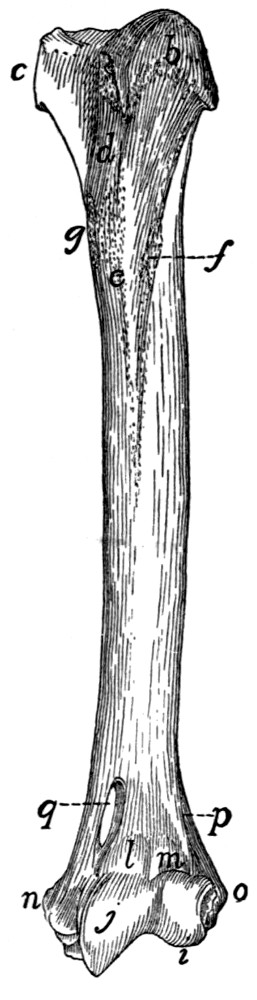{kind=link}
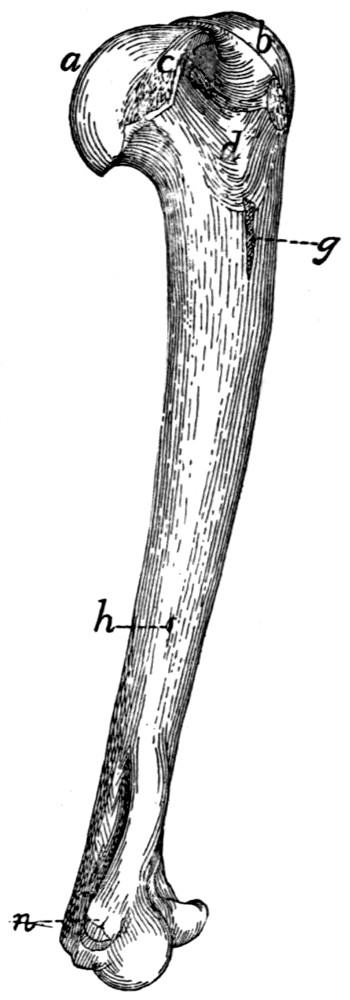{kind=link}
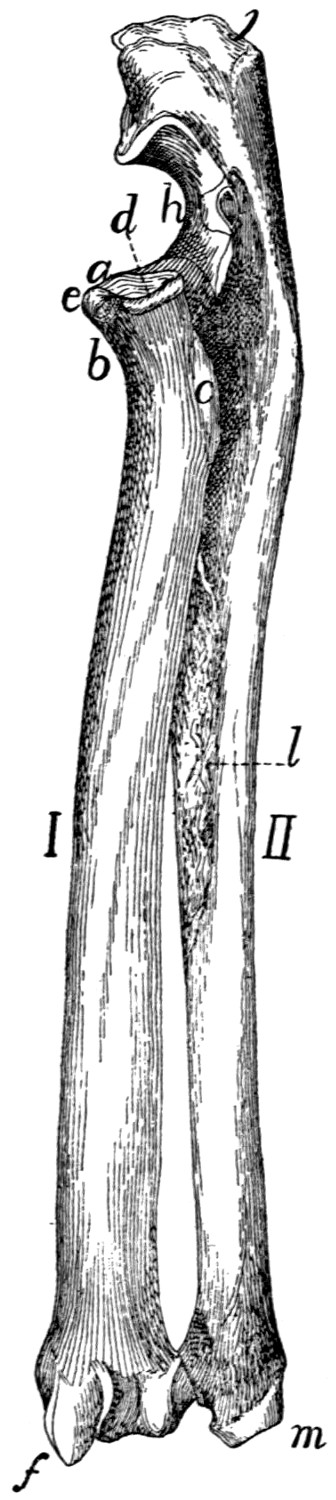{kind=link}
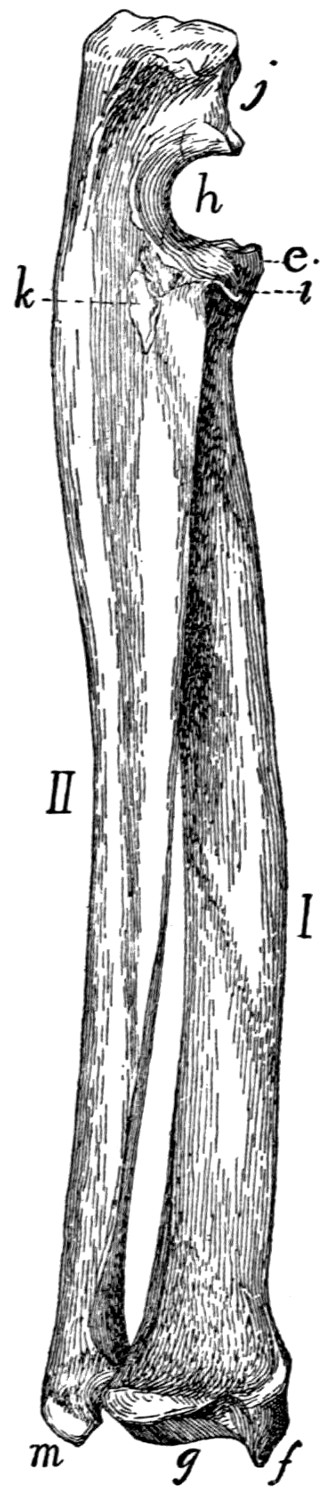{kind=link}
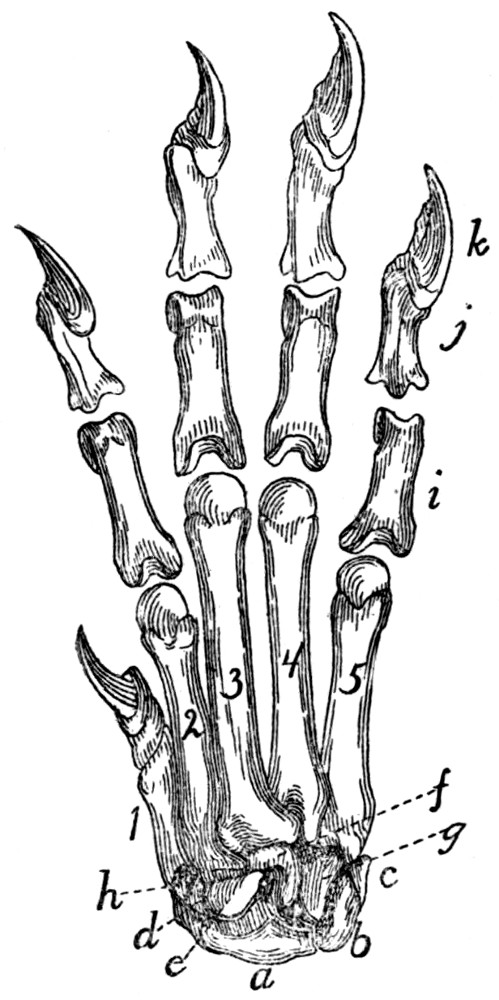{kind=link}
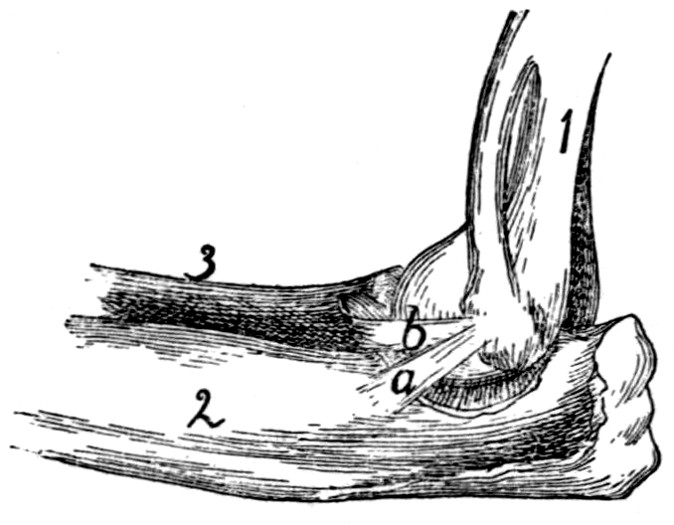{kind=link}
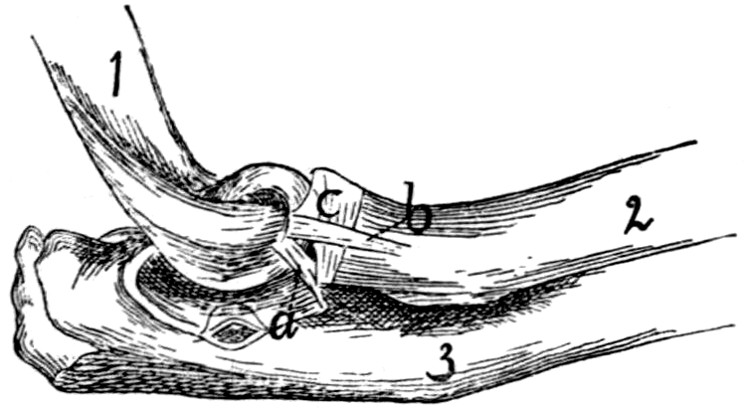{kind=link}
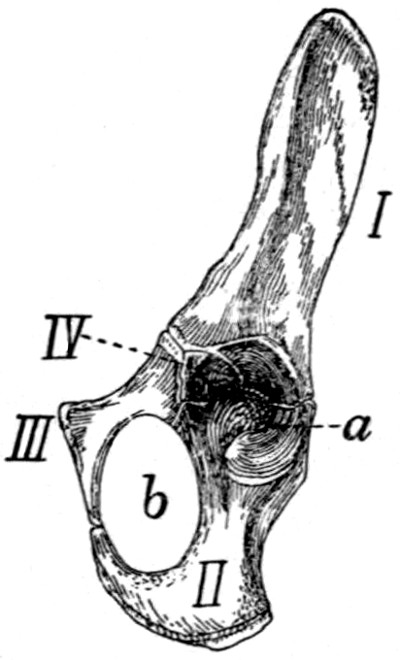{kind=link}
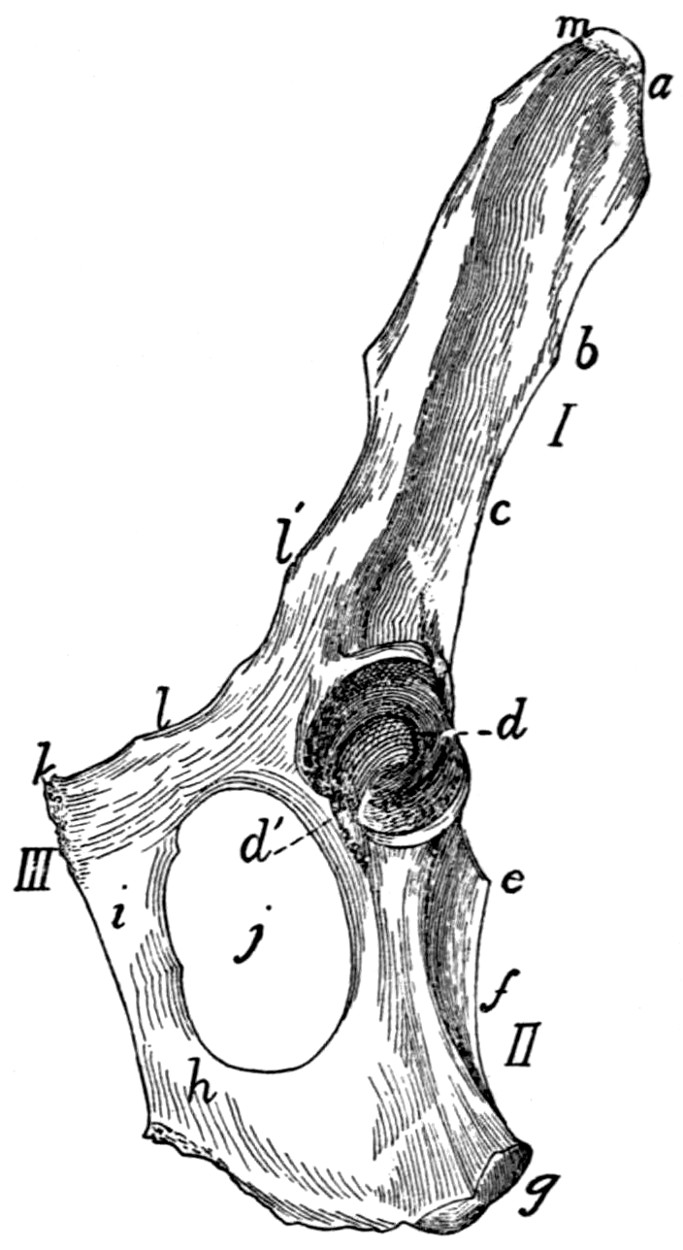{kind=link}
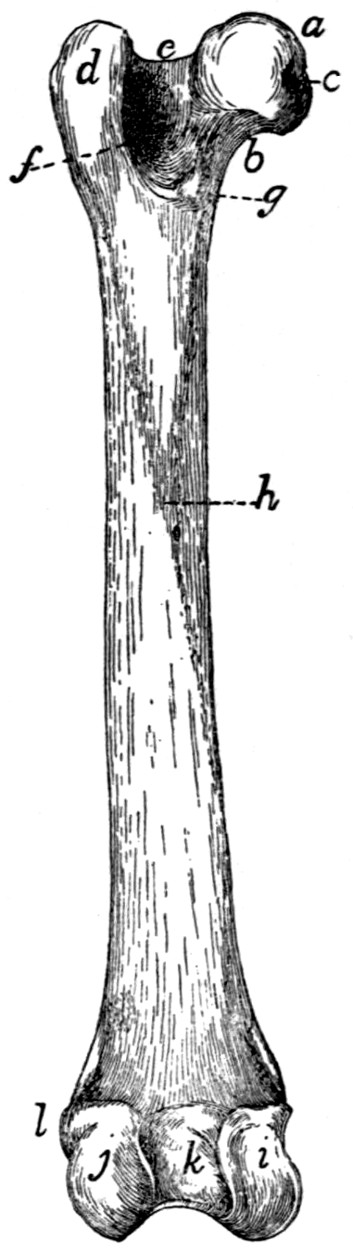{kind=link}
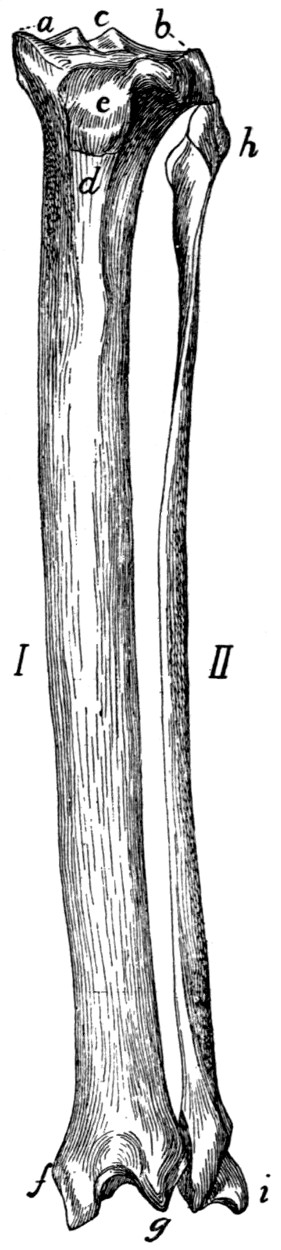{kind=link}
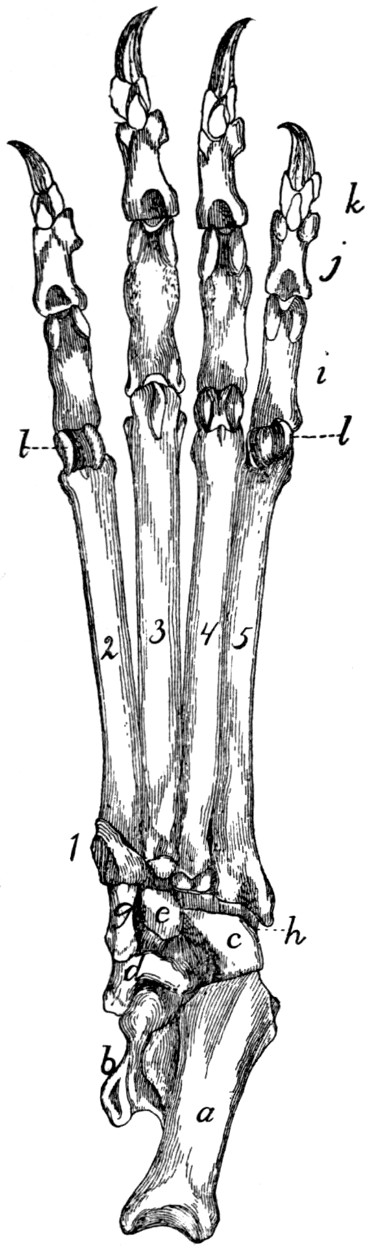{kind=link}
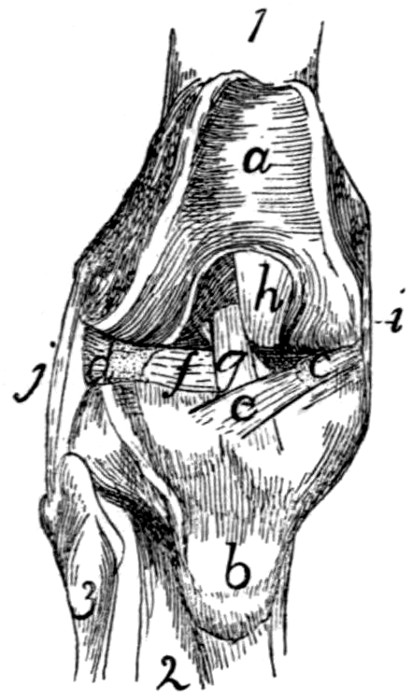{kind=link}
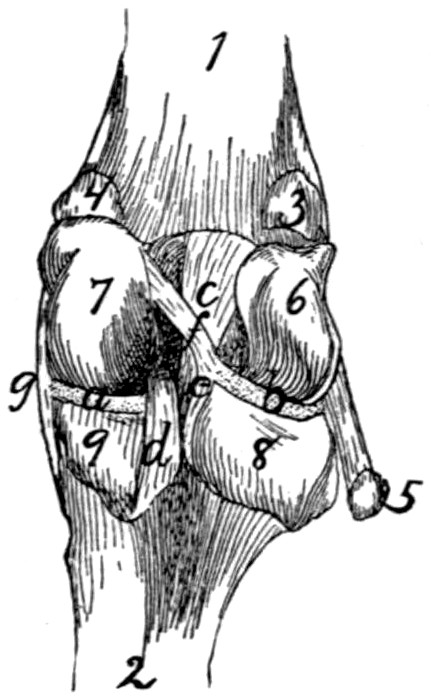{kind=link}
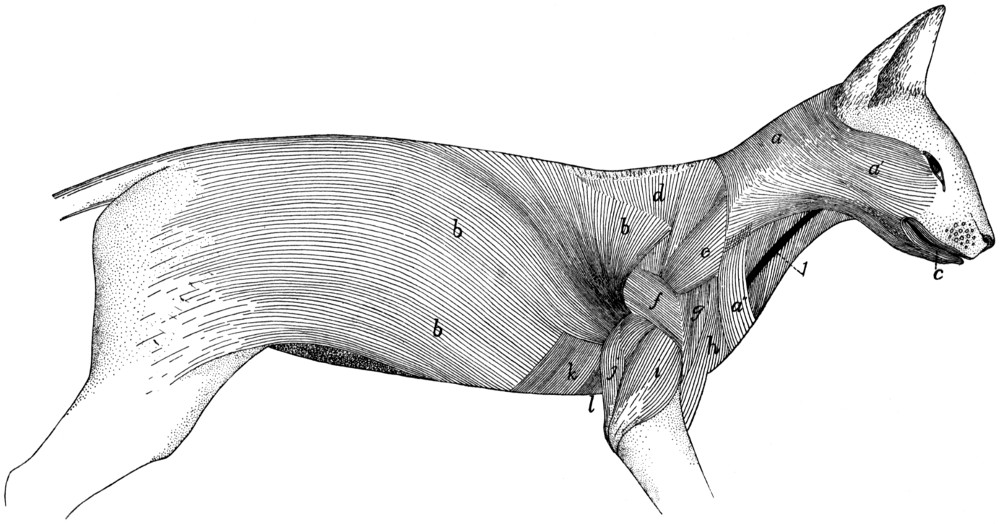{kind=link}
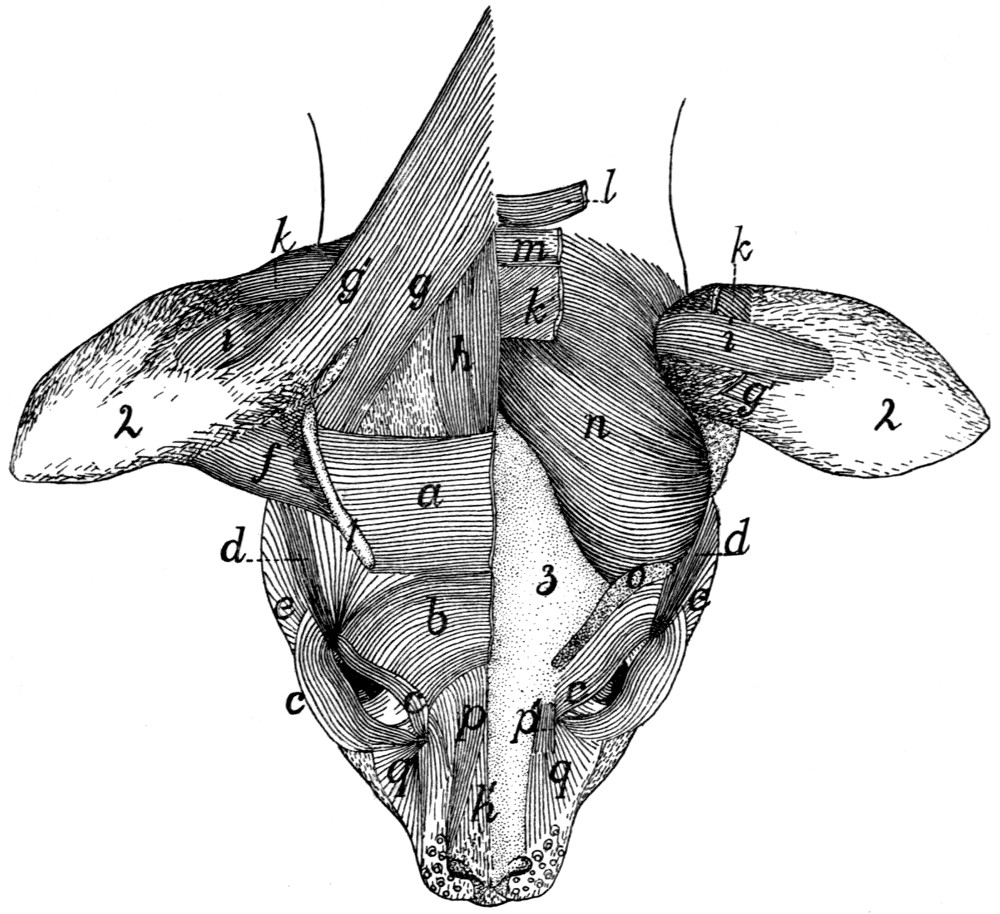{kind=link}
{kind=link}
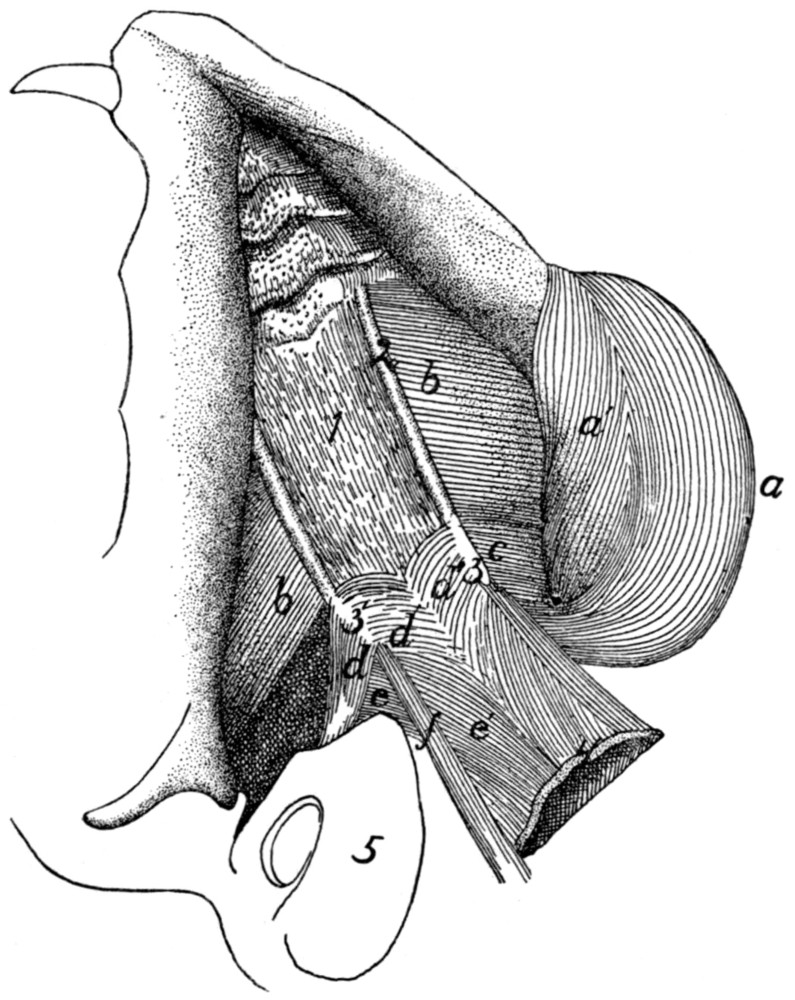{kind=link}
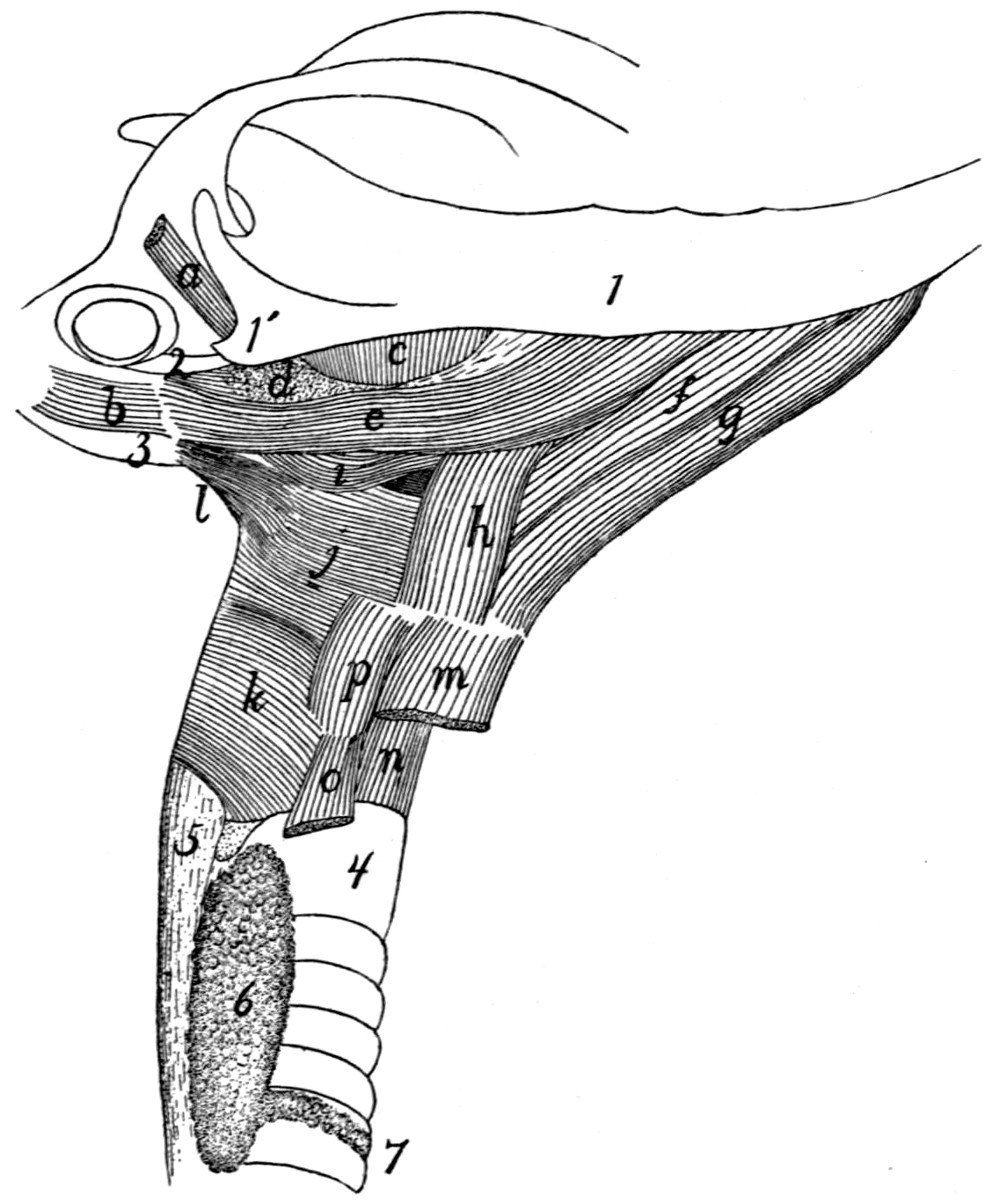{kind=link}
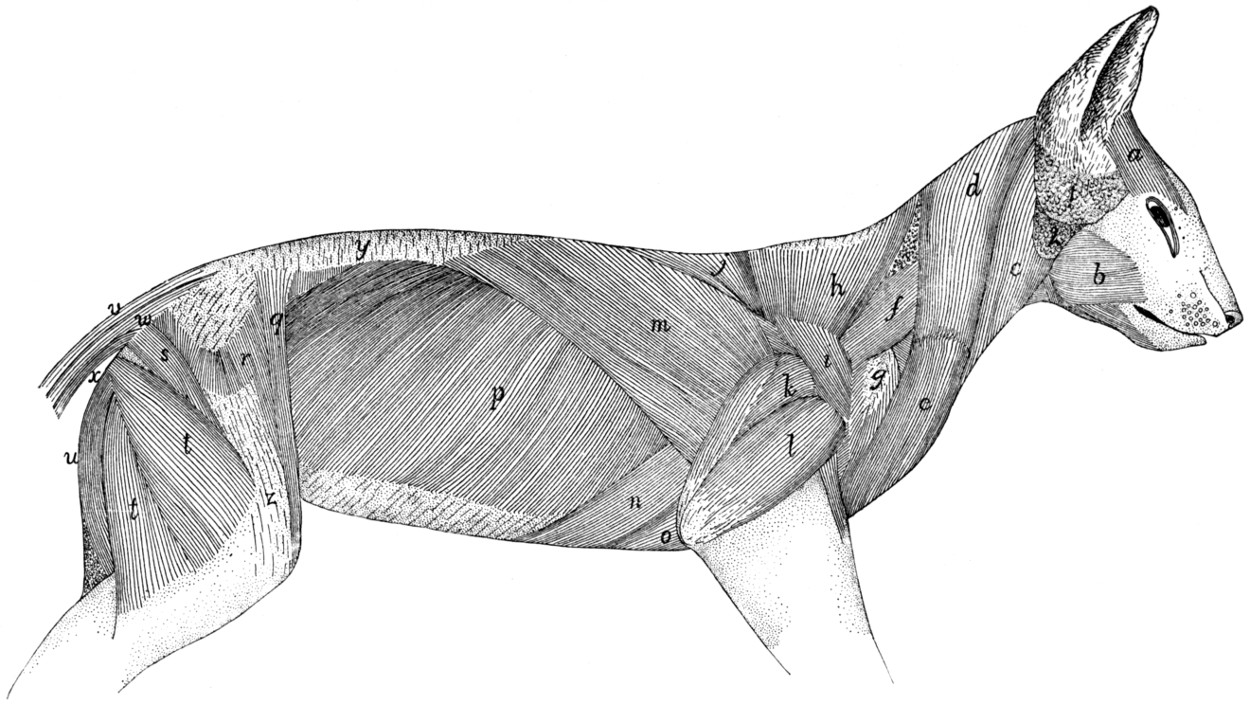{kind=link}
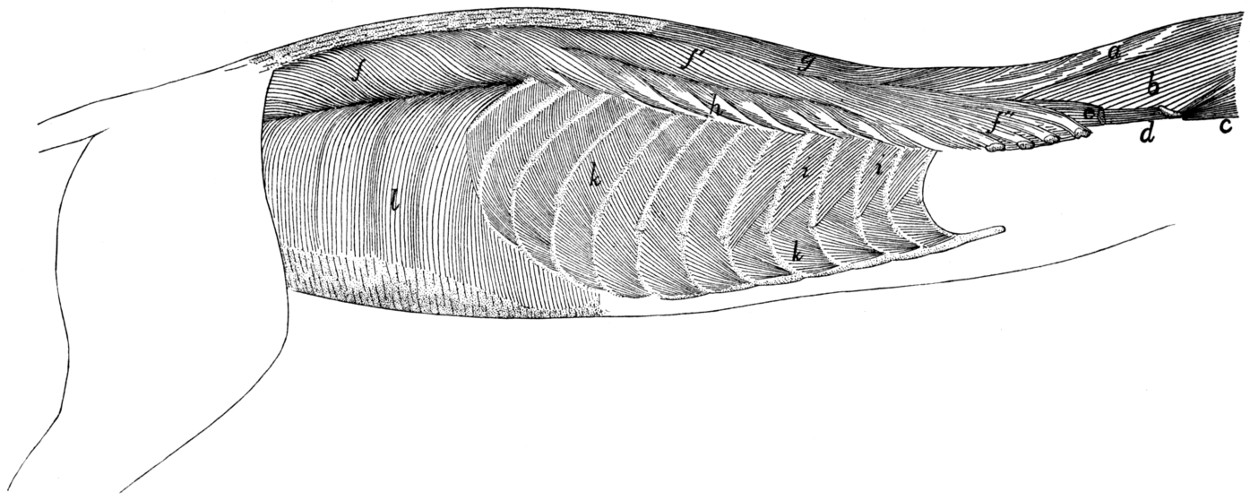{kind=link}
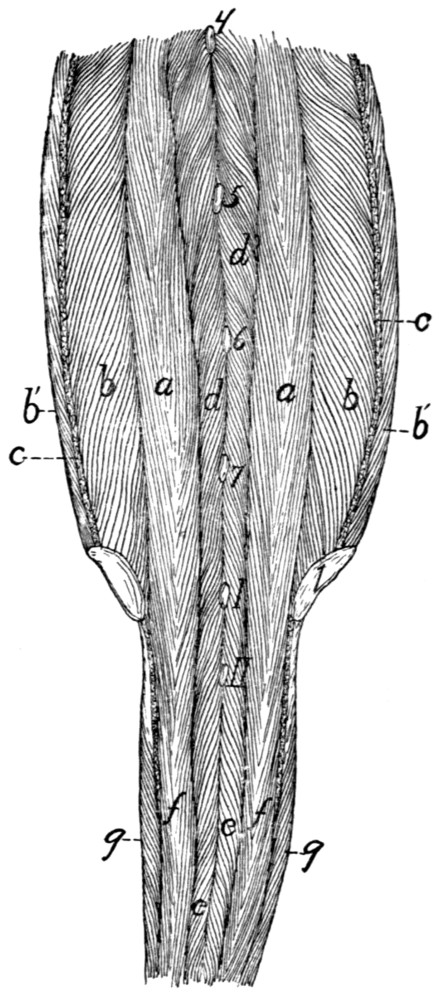{kind=link}
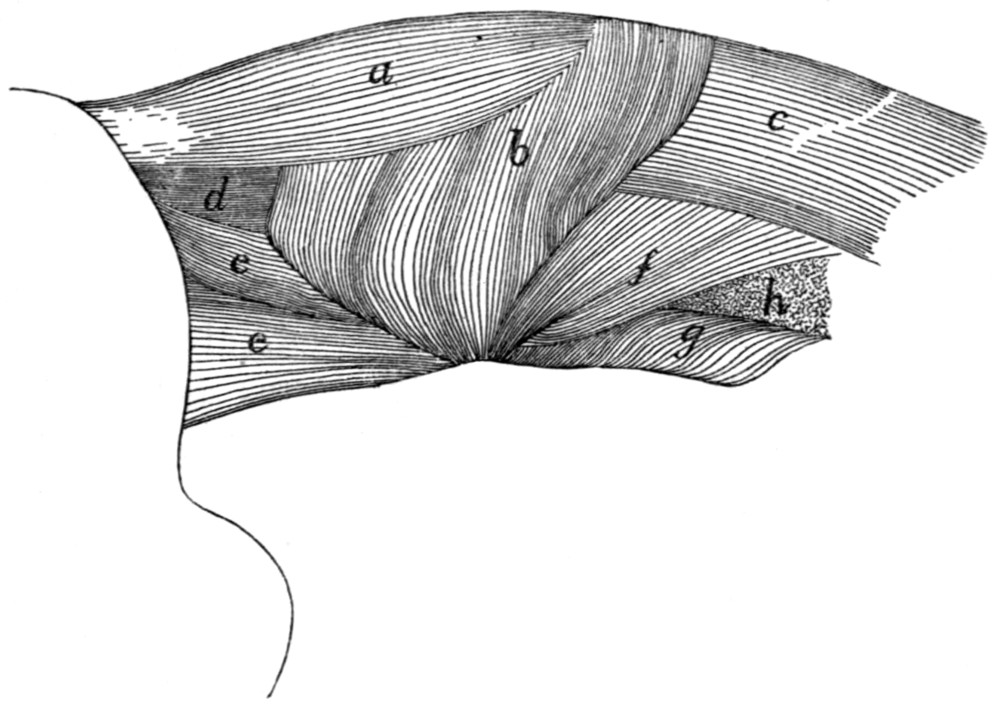{kind=link}
{kind=link}
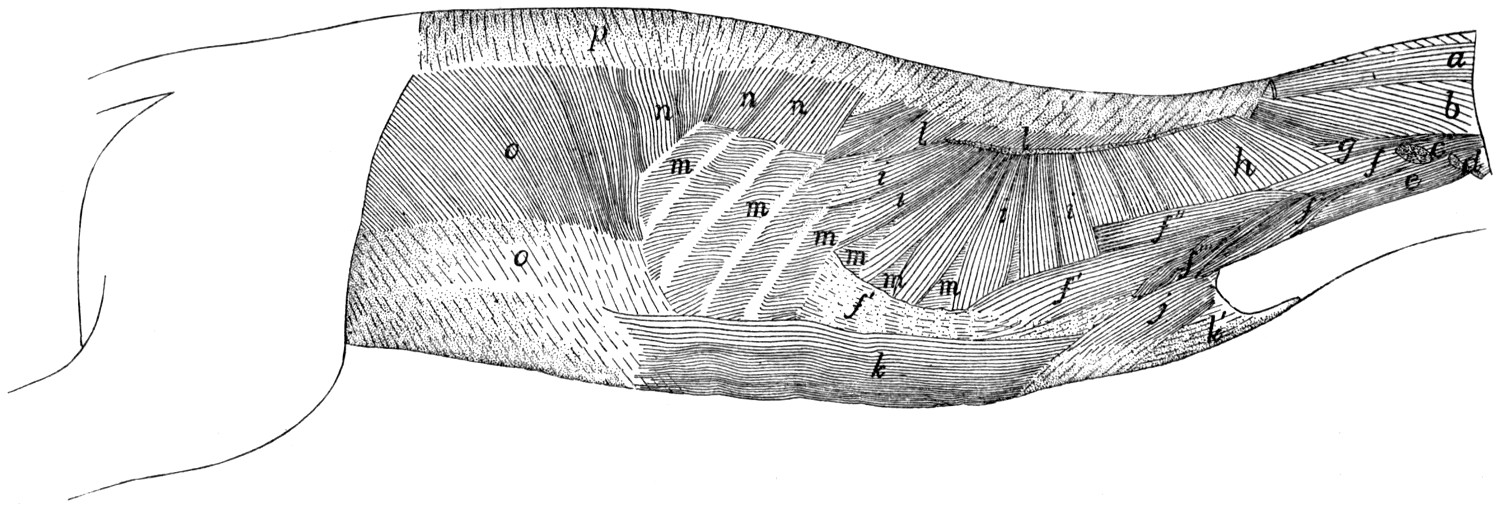{kind=link}
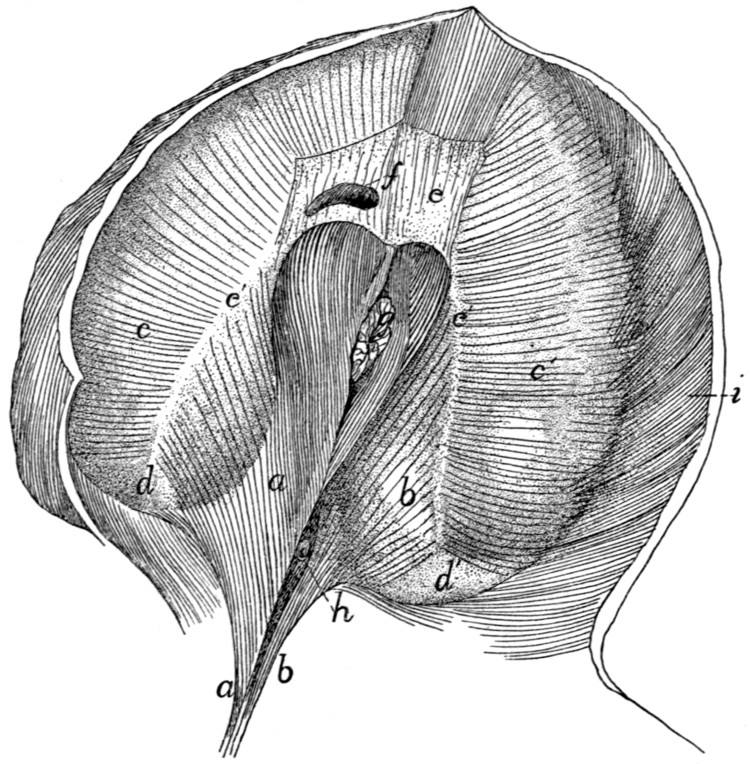{kind=link}
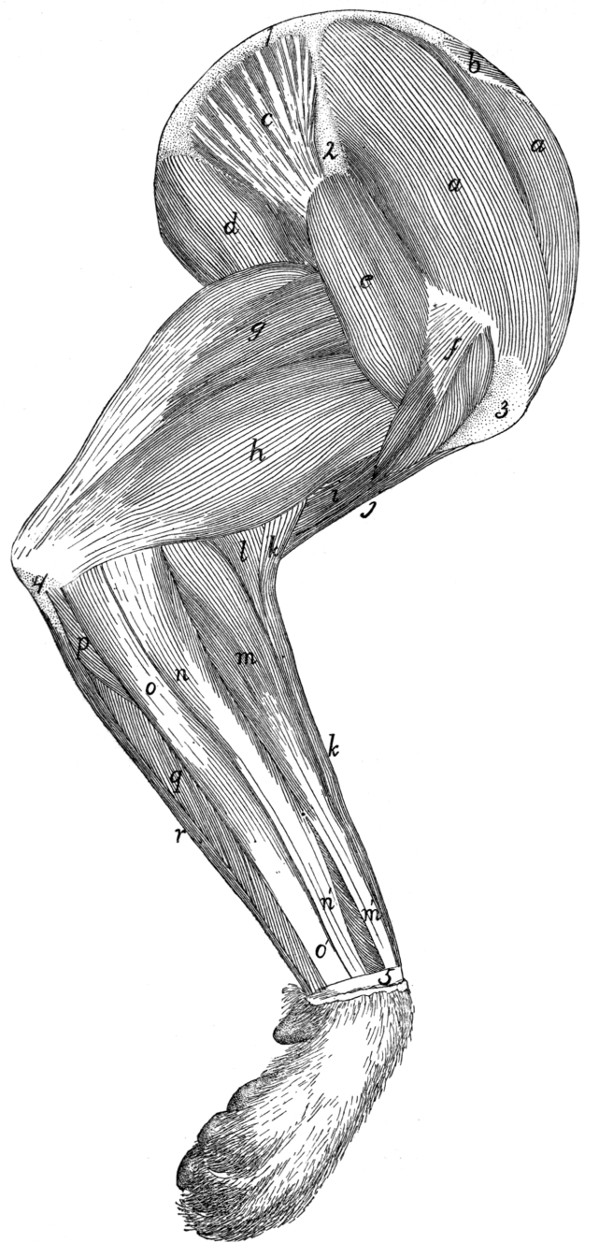{kind=link}
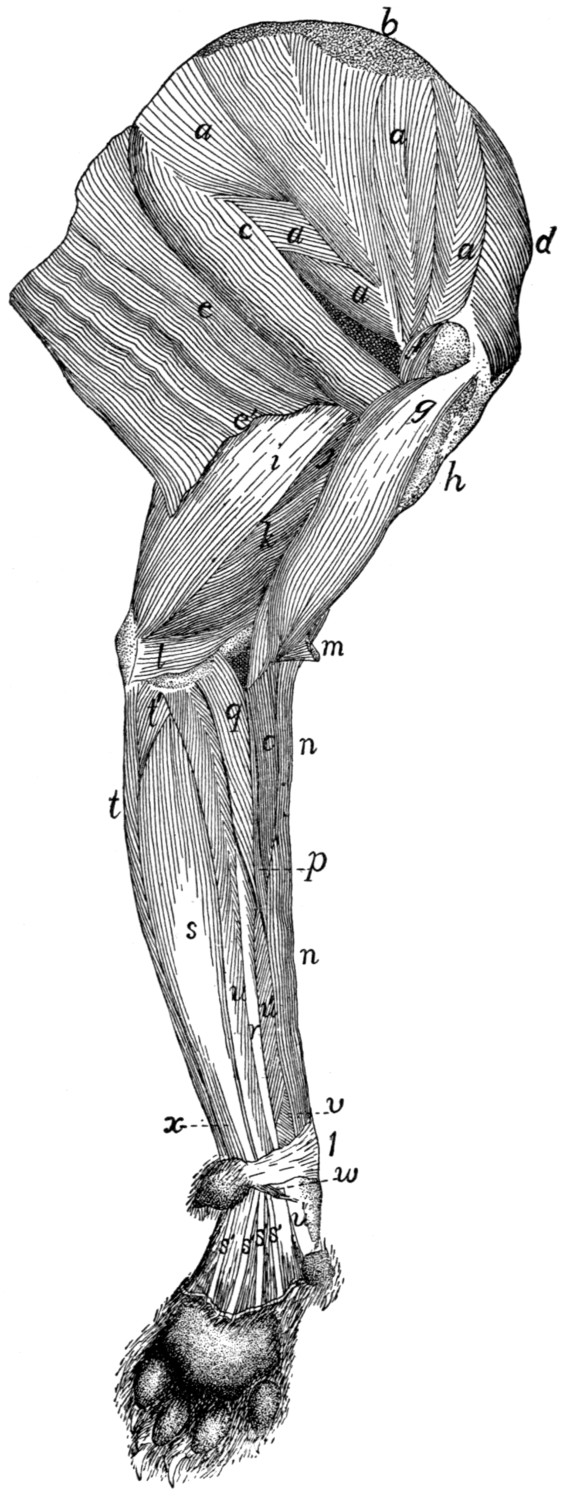{kind=link}
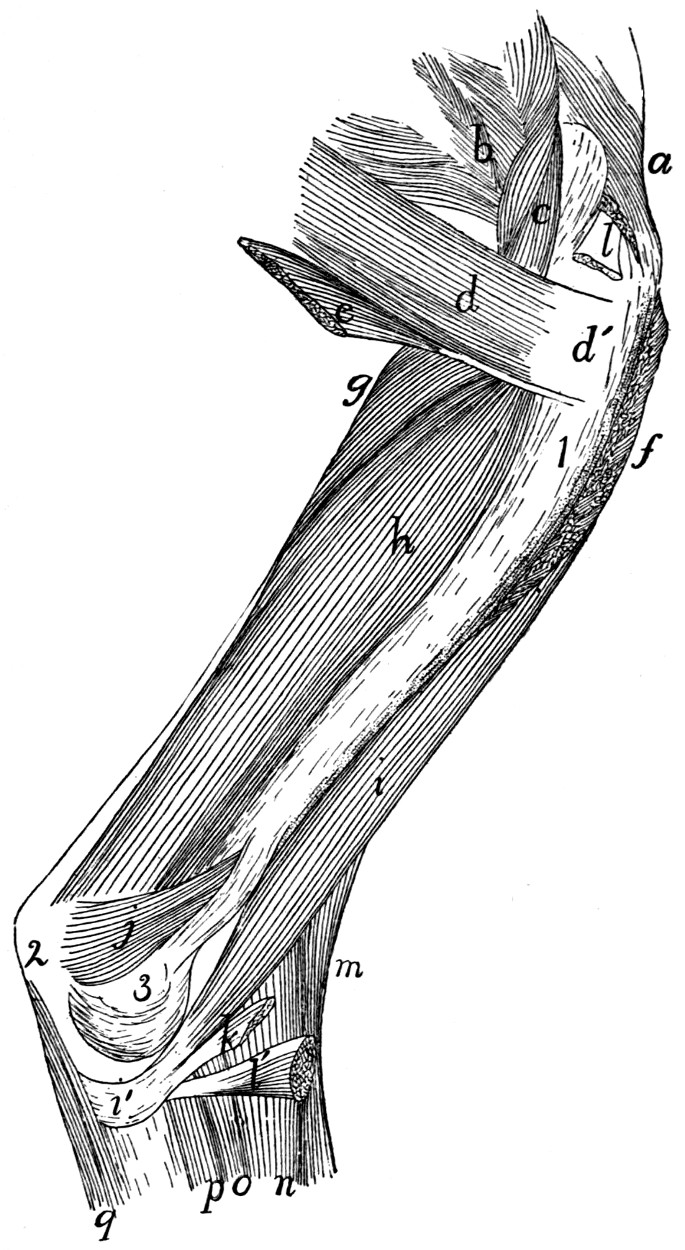{kind=link}
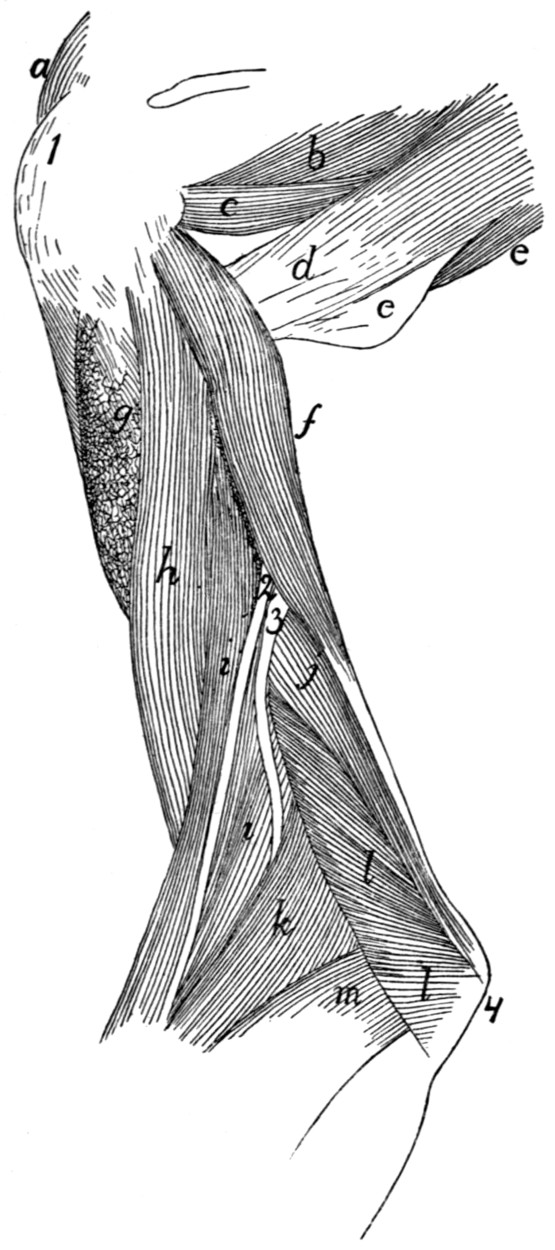{kind=link}
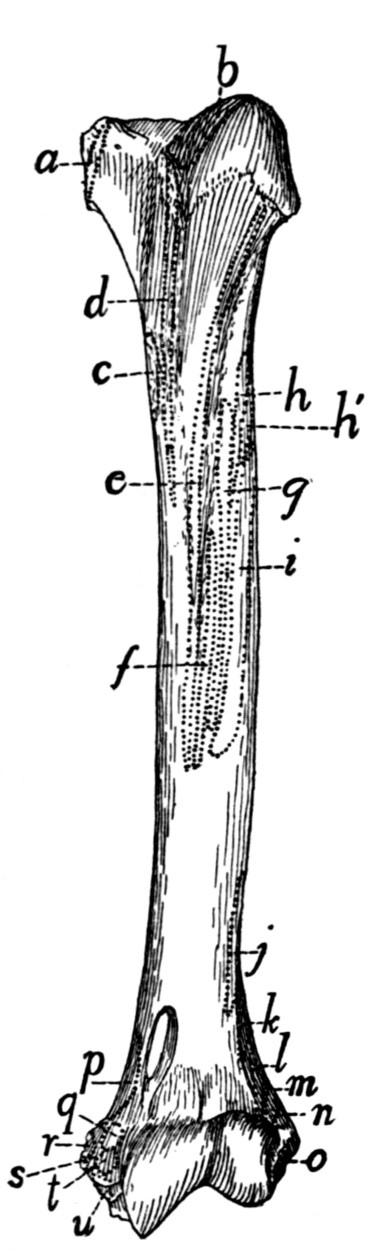{kind=link}
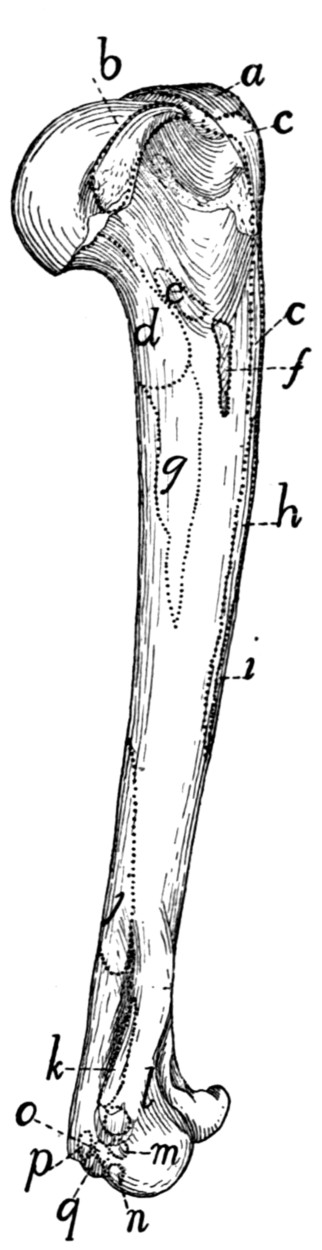{kind=link}
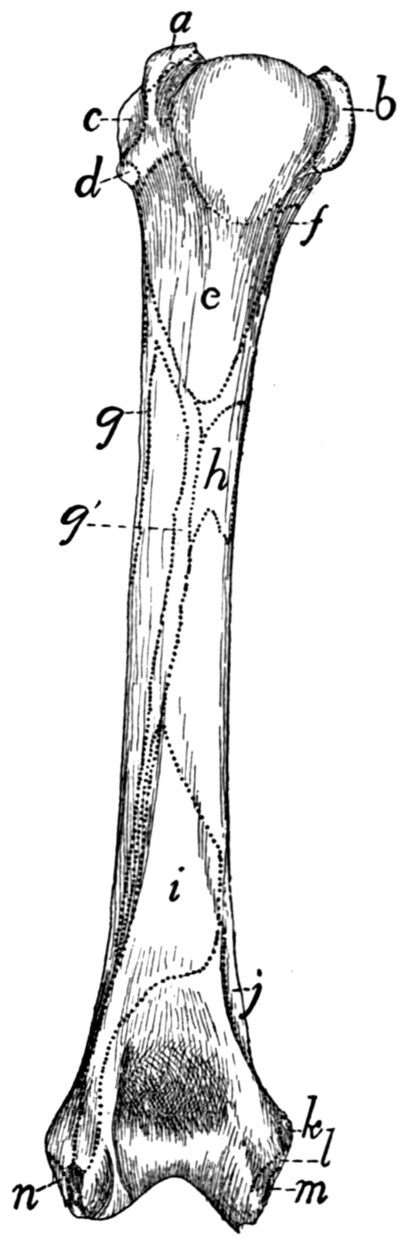{kind=link}
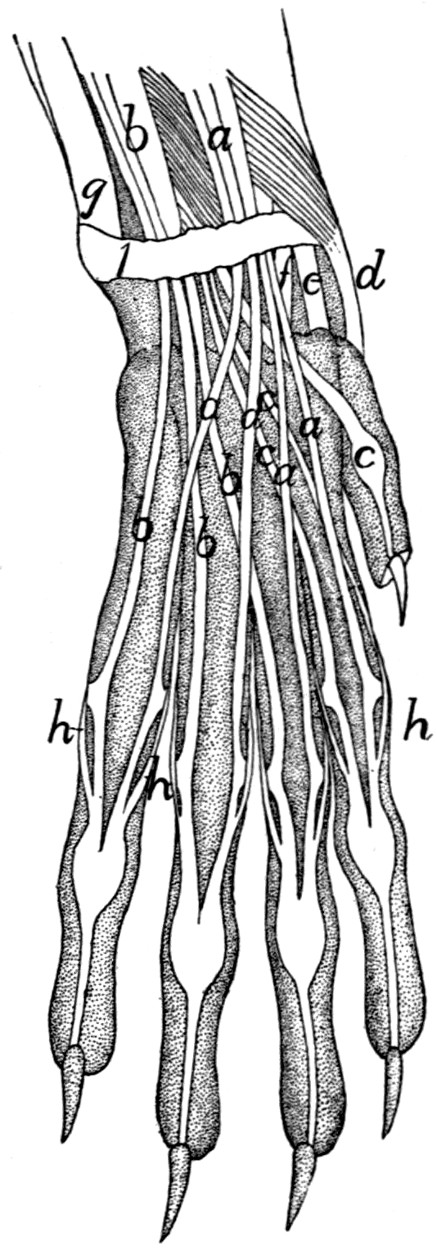{kind=link}
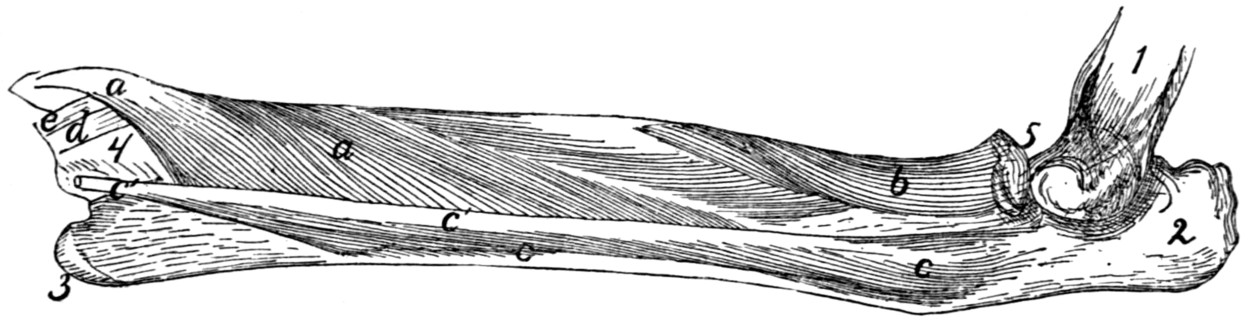{kind=link}
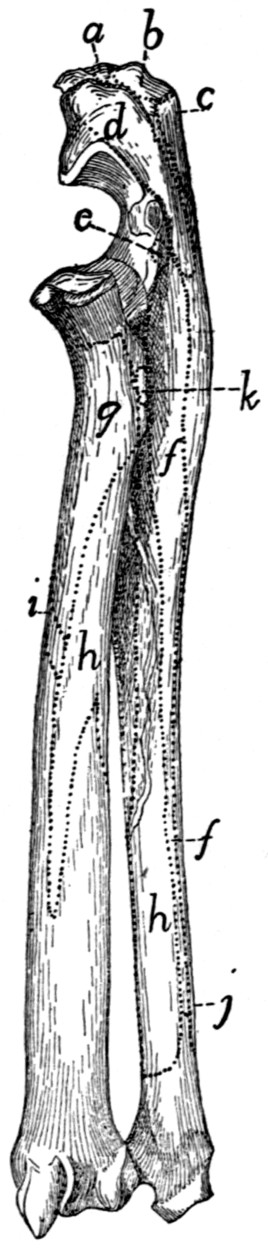{kind=link}
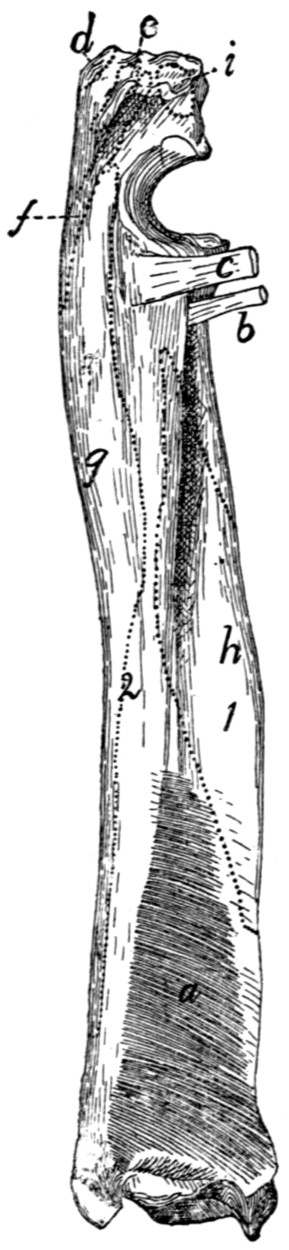{kind=link}
{kind=link}
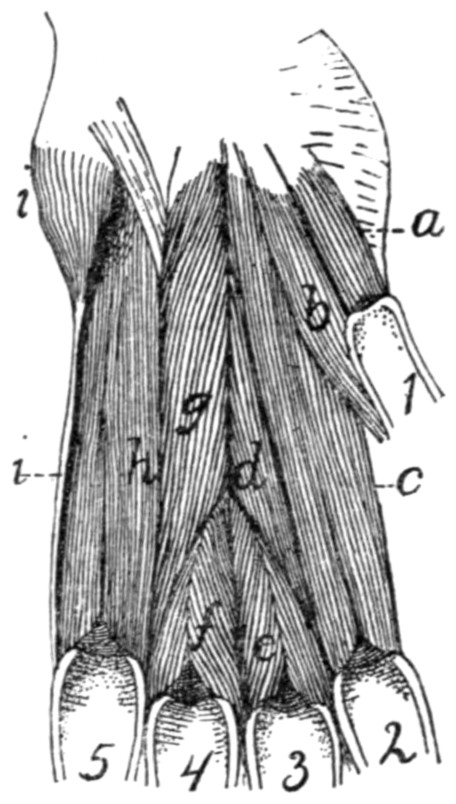{kind=link}
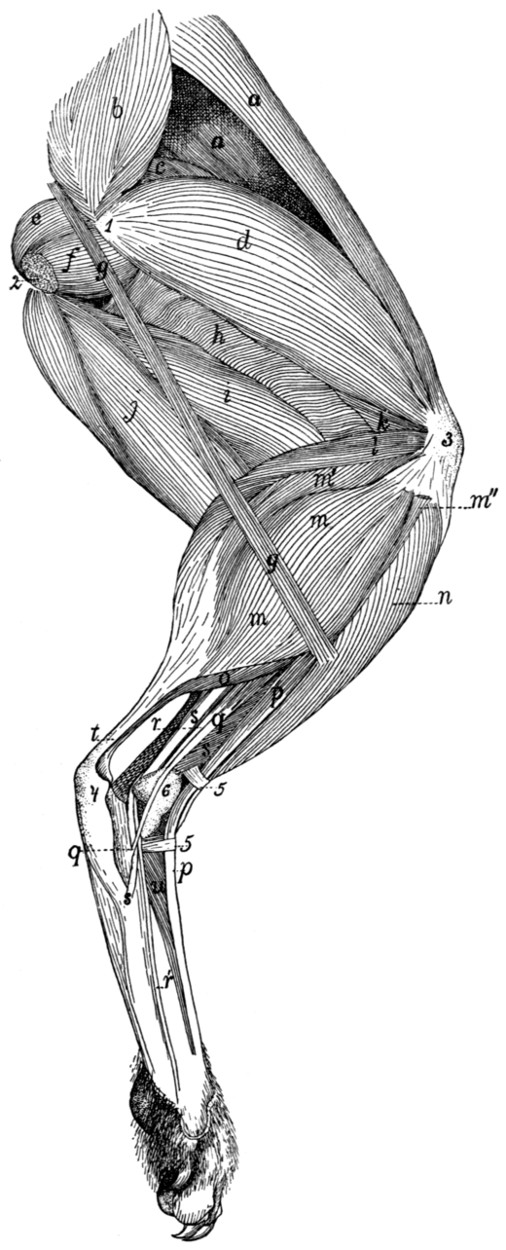{kind=link}
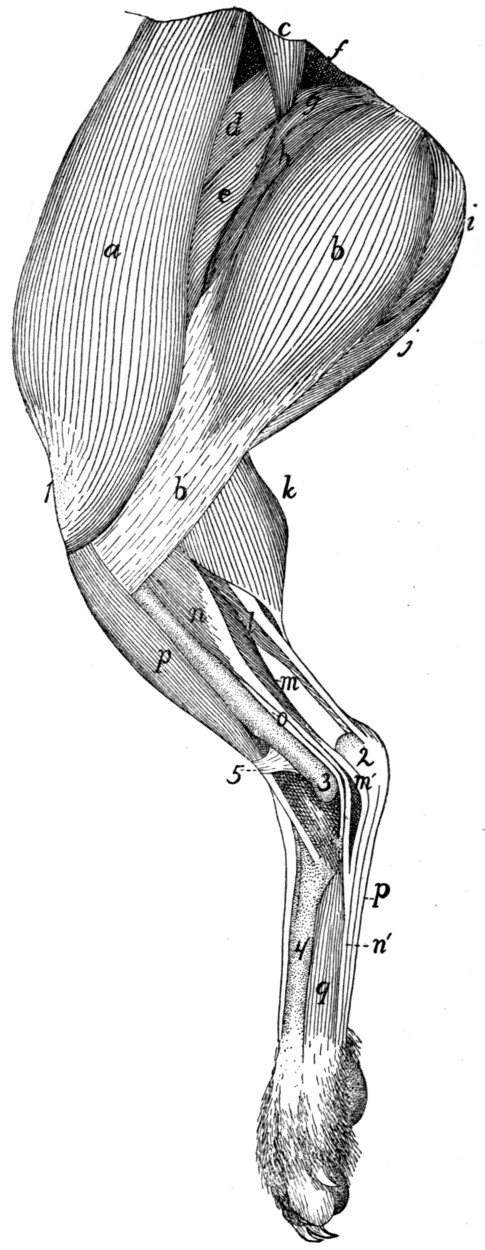{kind=link}
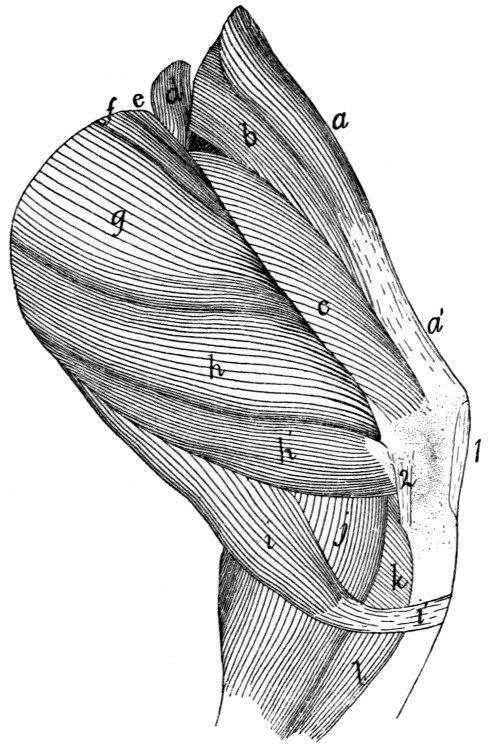{kind=link}
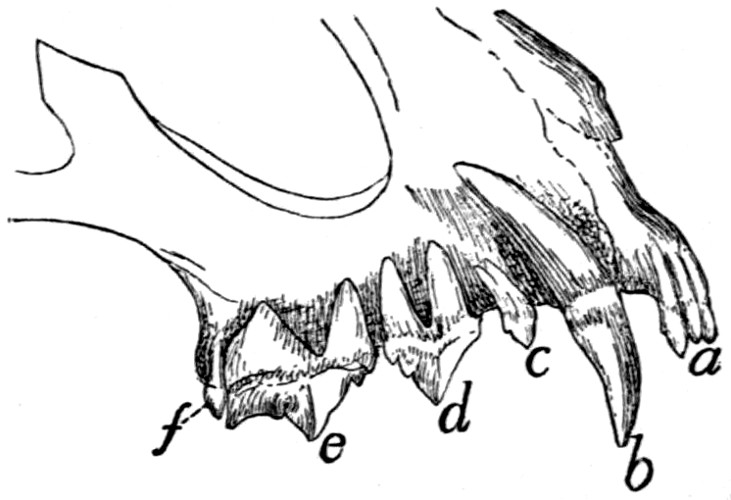{kind=link}
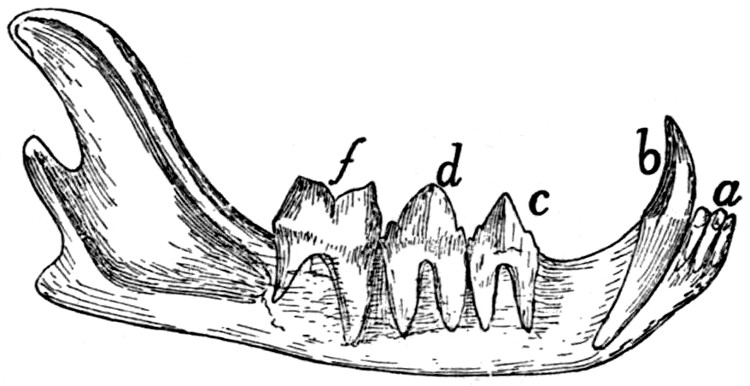{kind=link}
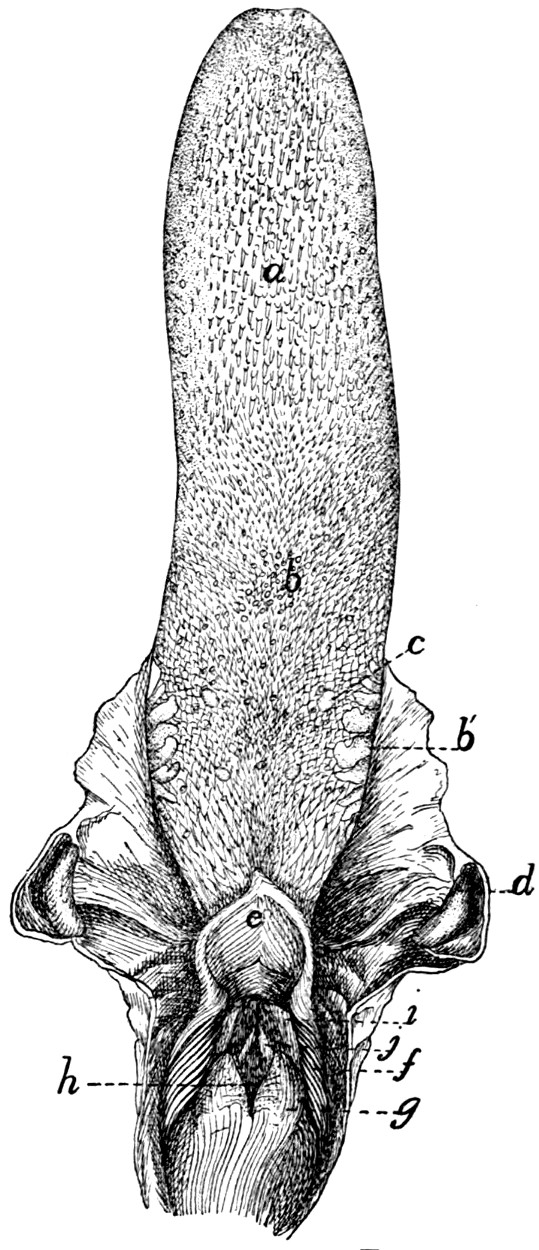{kind=link}
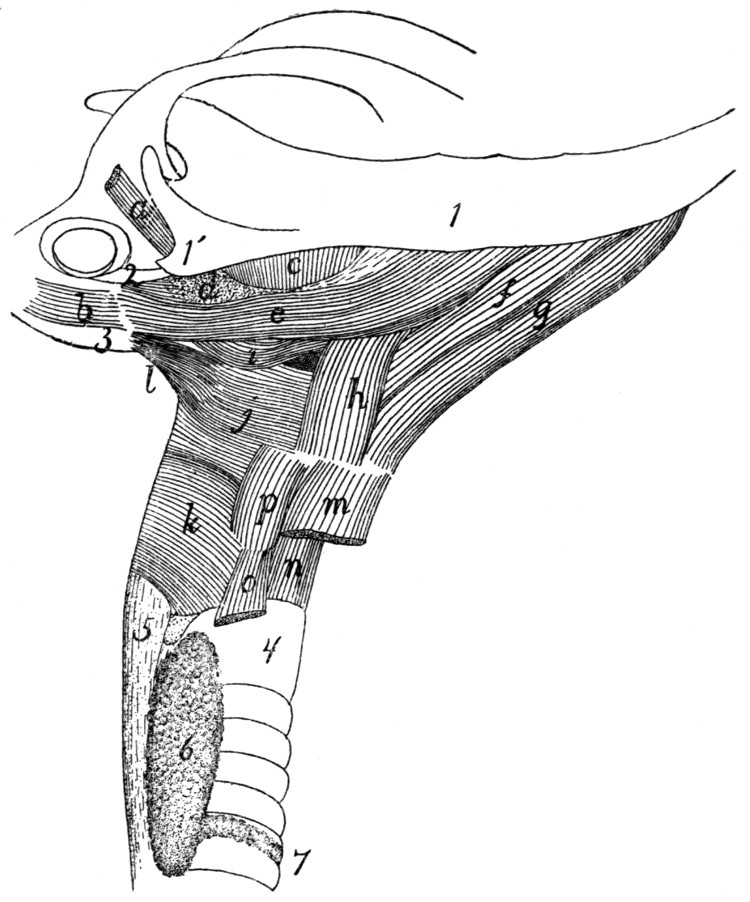{kind=link}
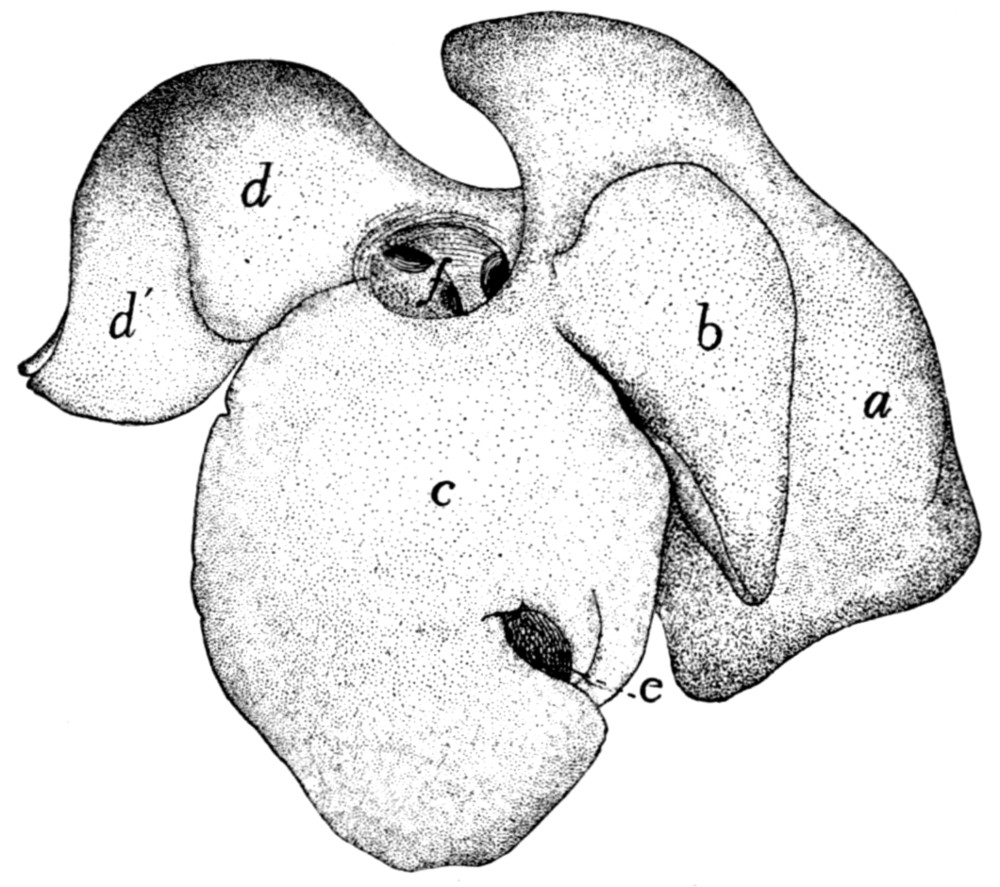{kind=link}
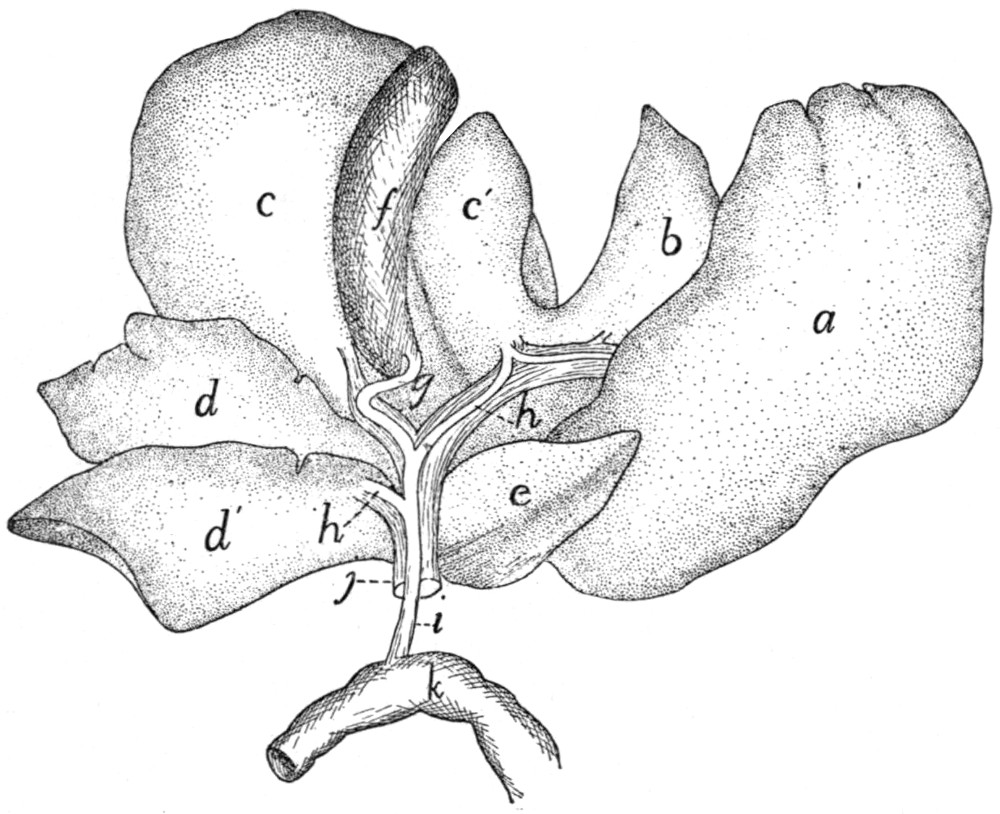{kind=link}
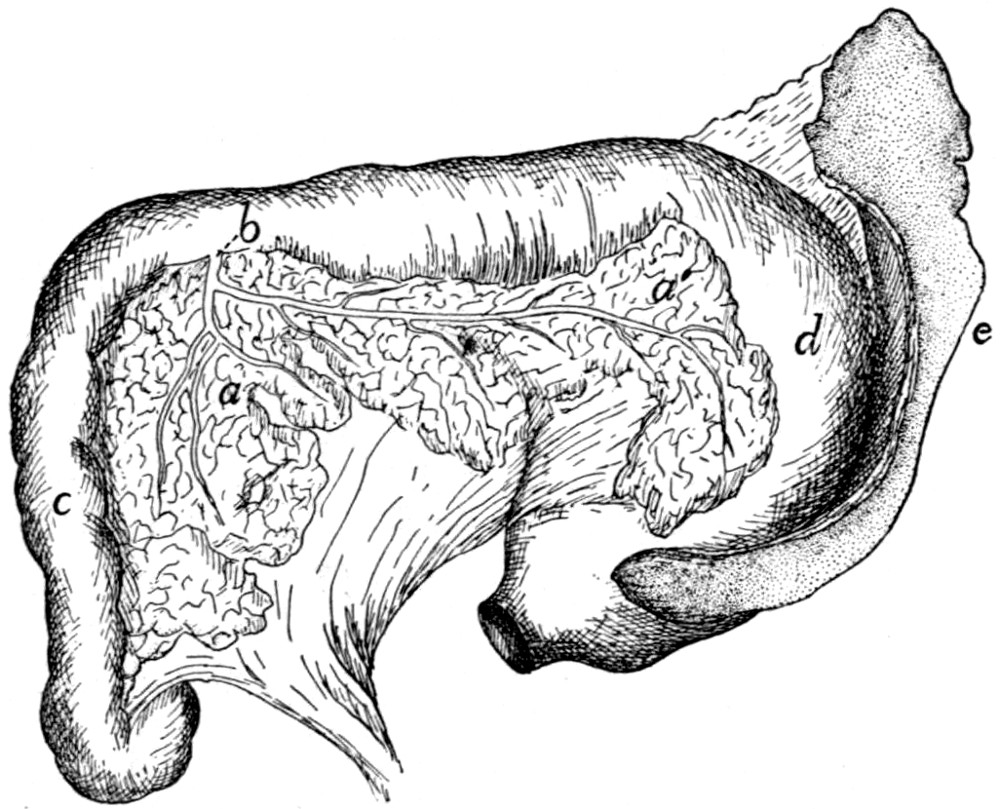{kind=link}
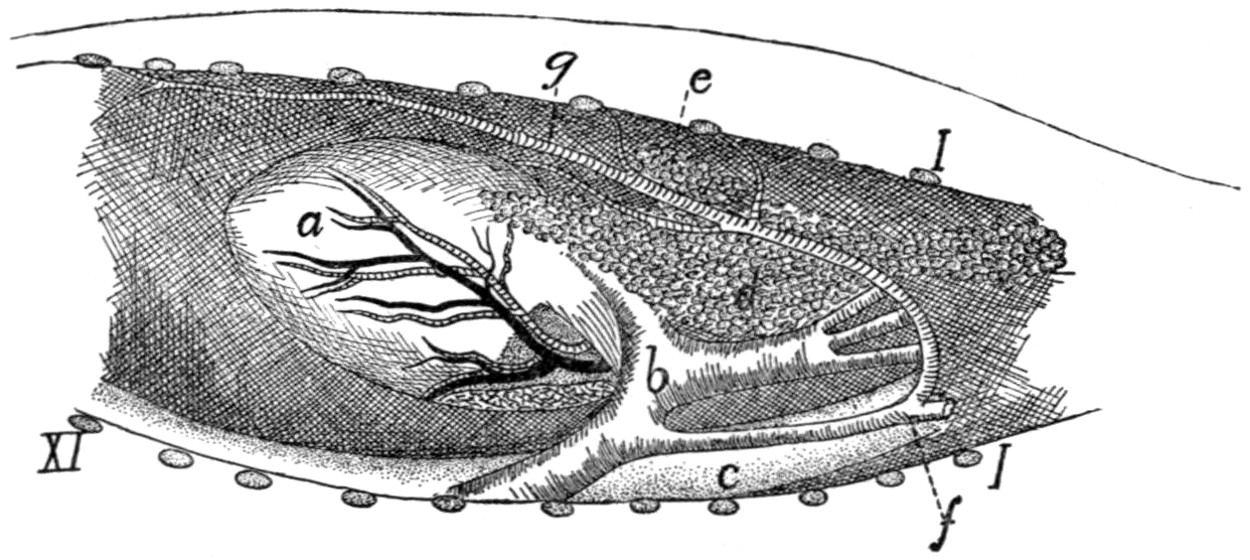{kind=link}
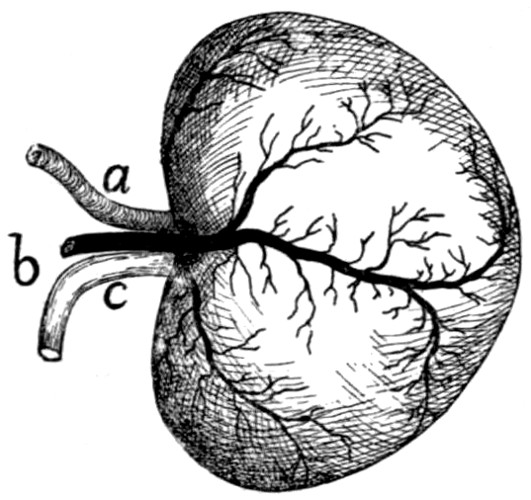{kind=link}
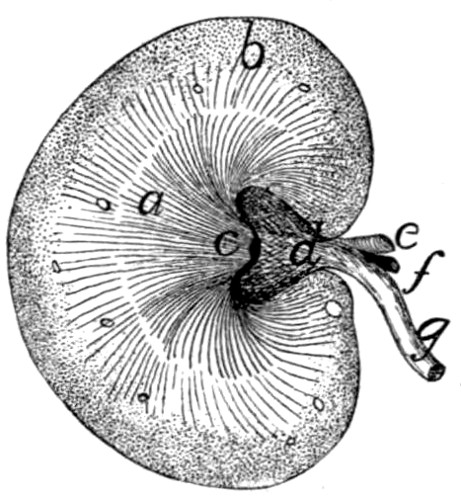{kind=link}
{kind=link}
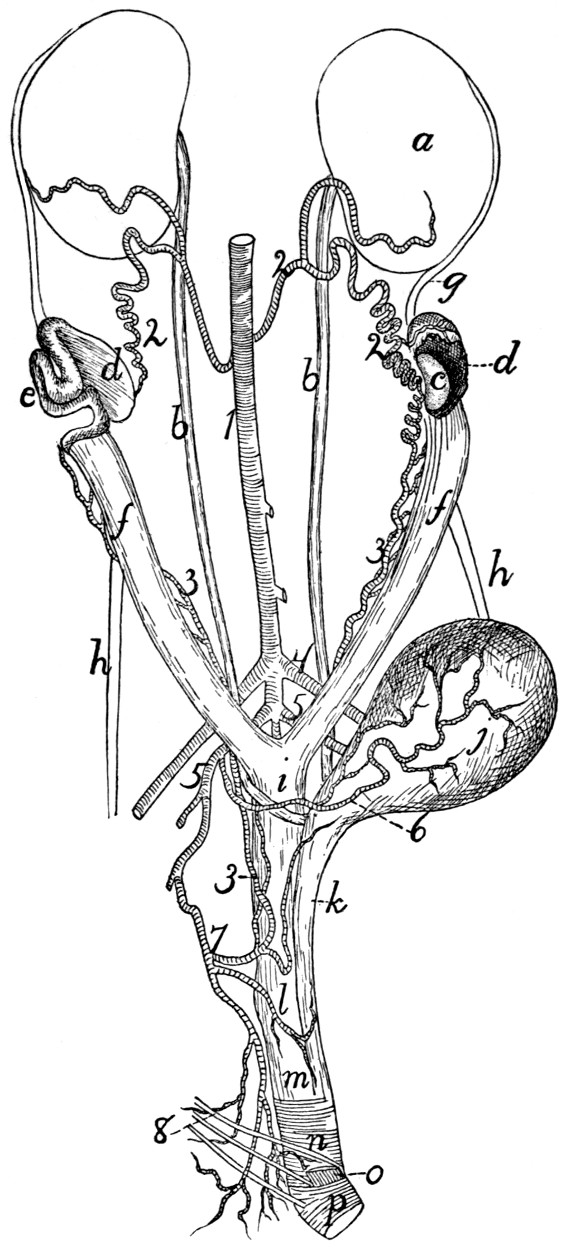{kind=link}
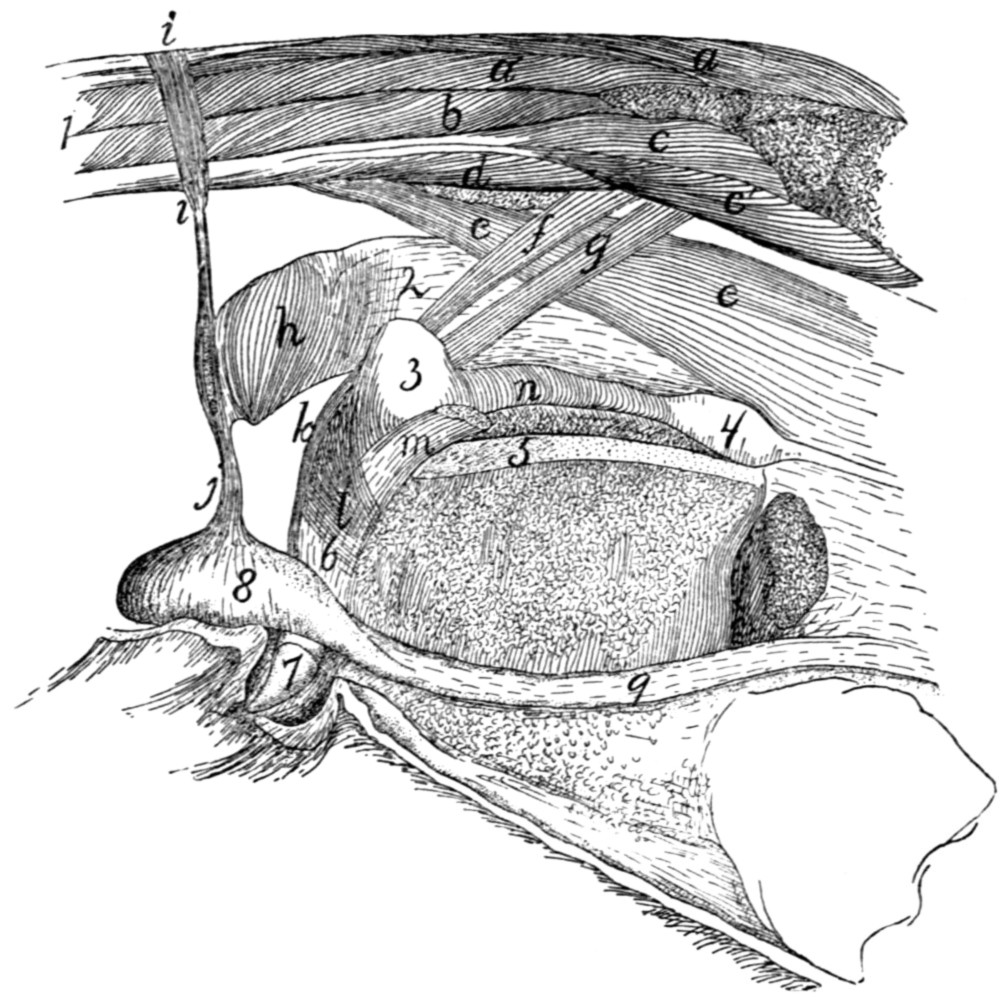{kind=link}
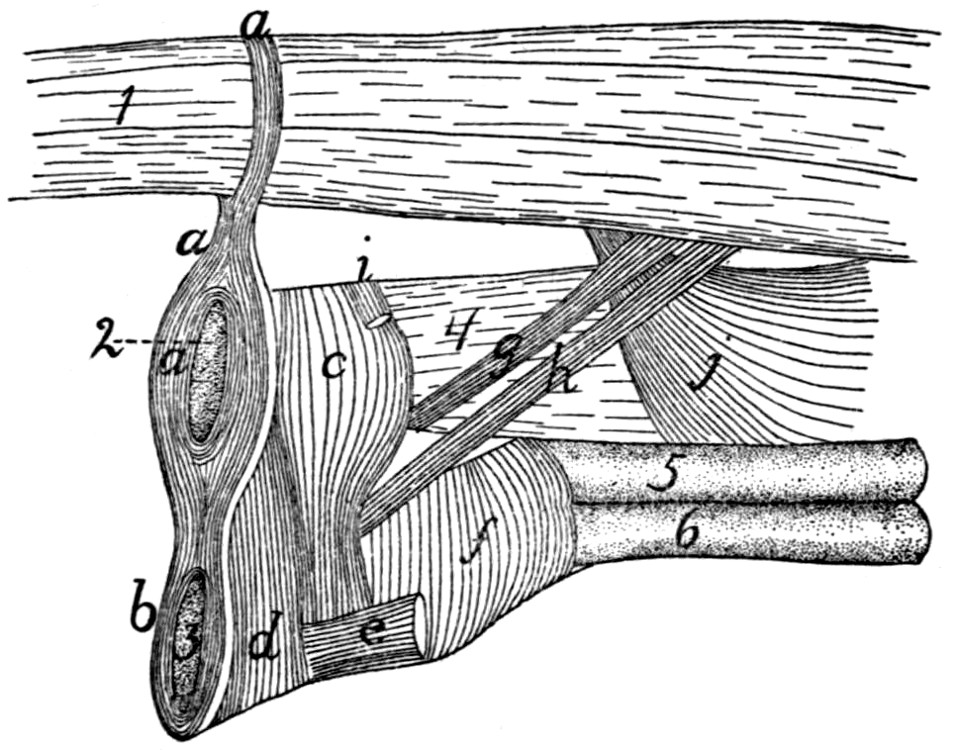{kind=link}
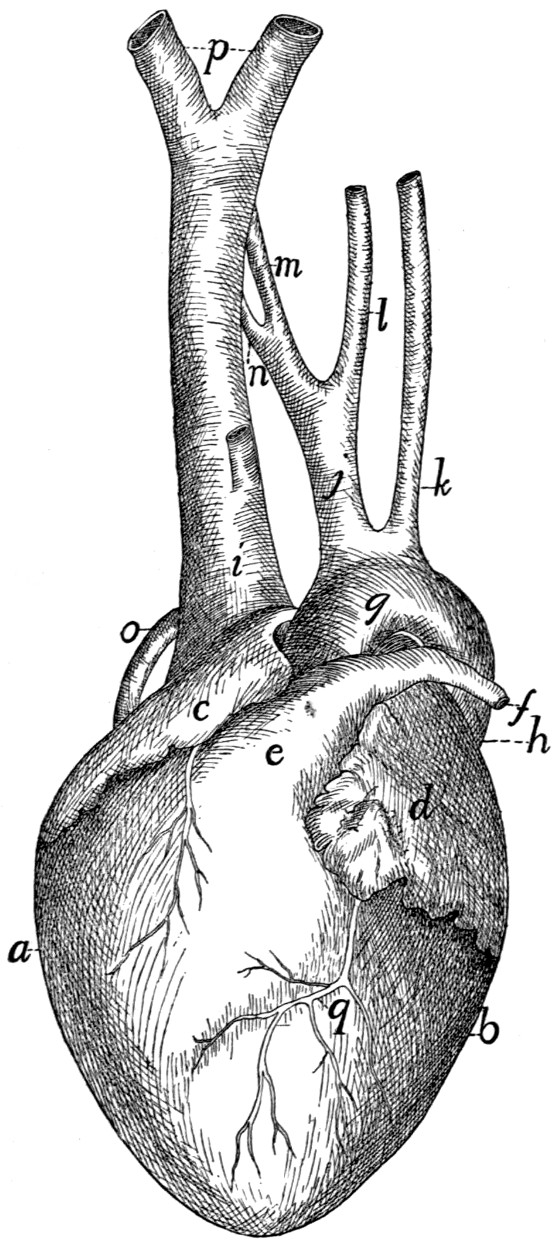{kind=link}
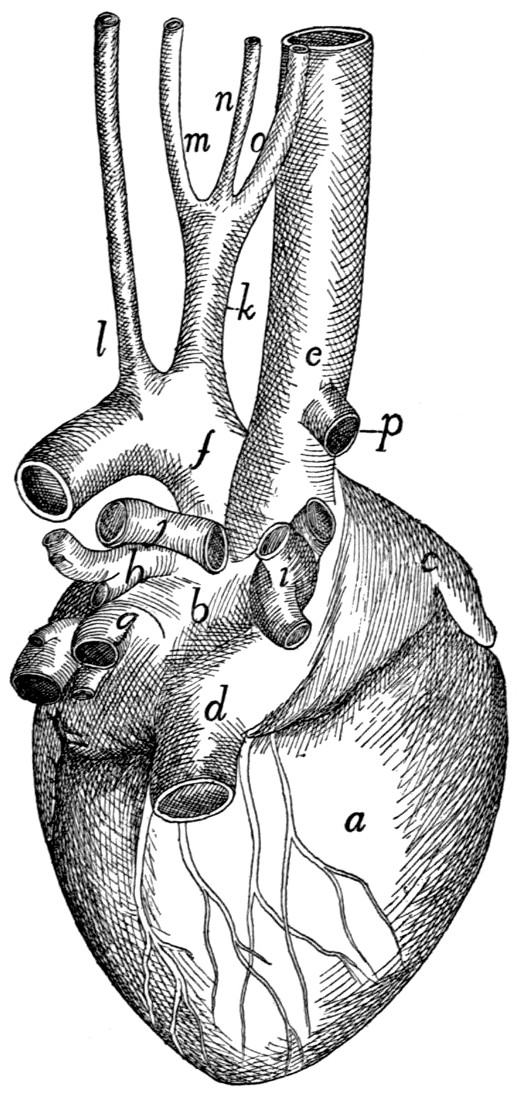{kind=link}
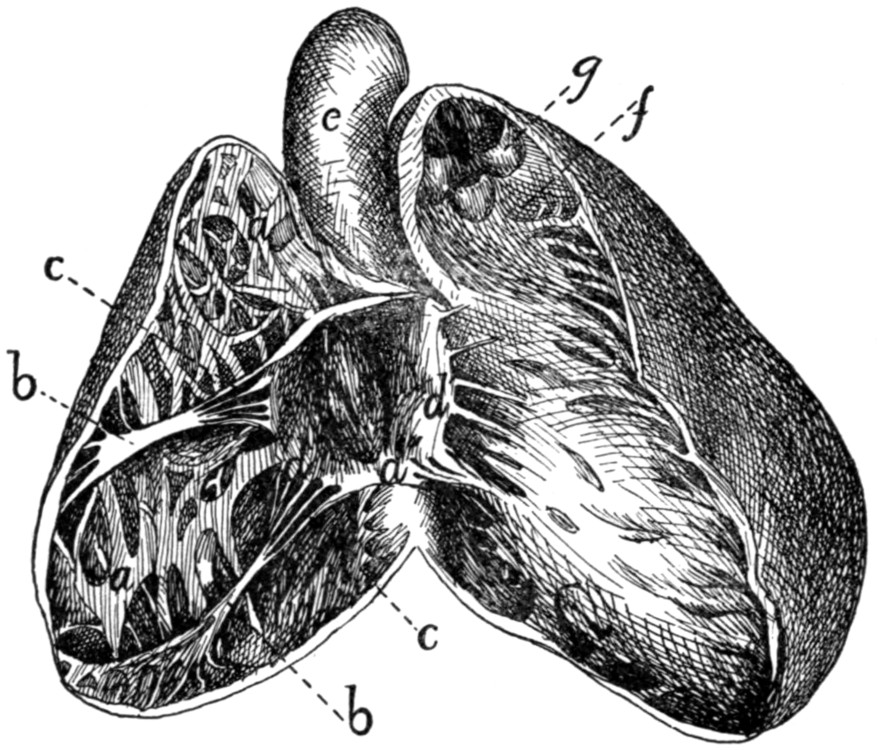{kind=link}
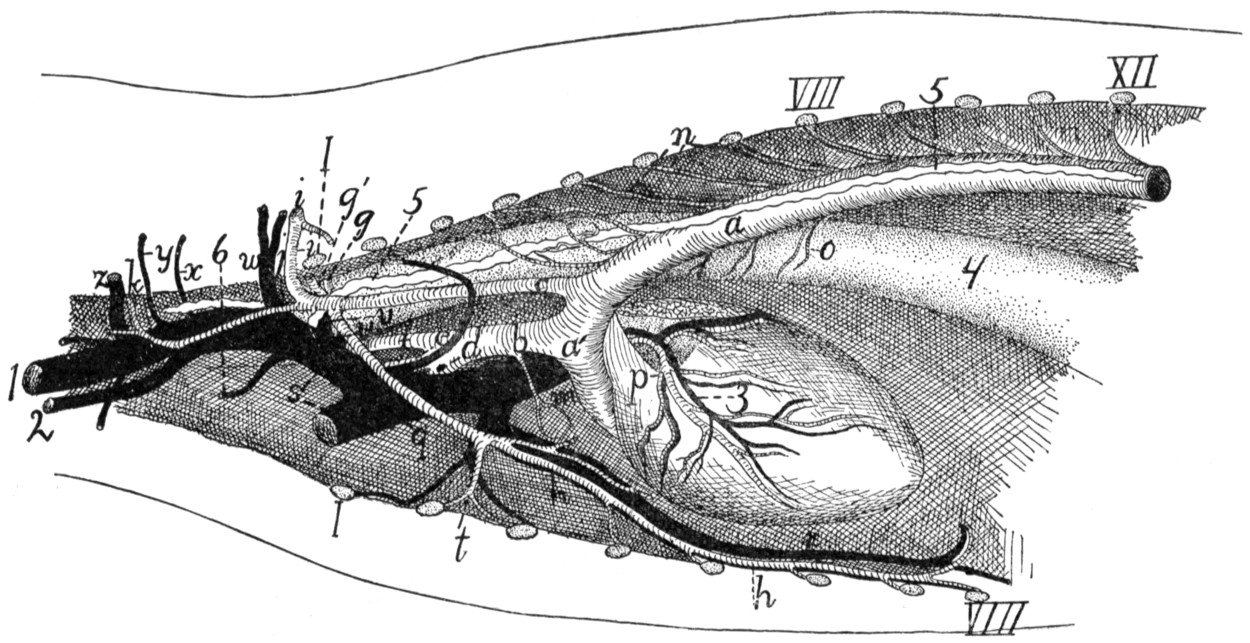{kind=link}
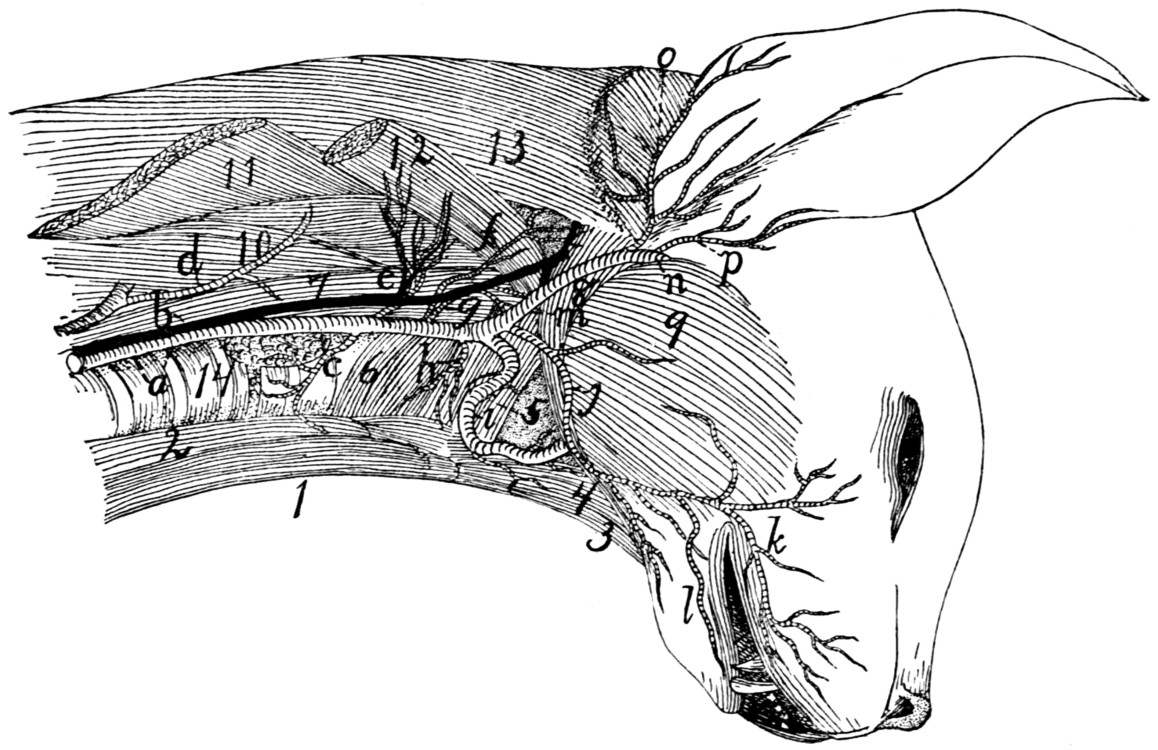{kind=link}
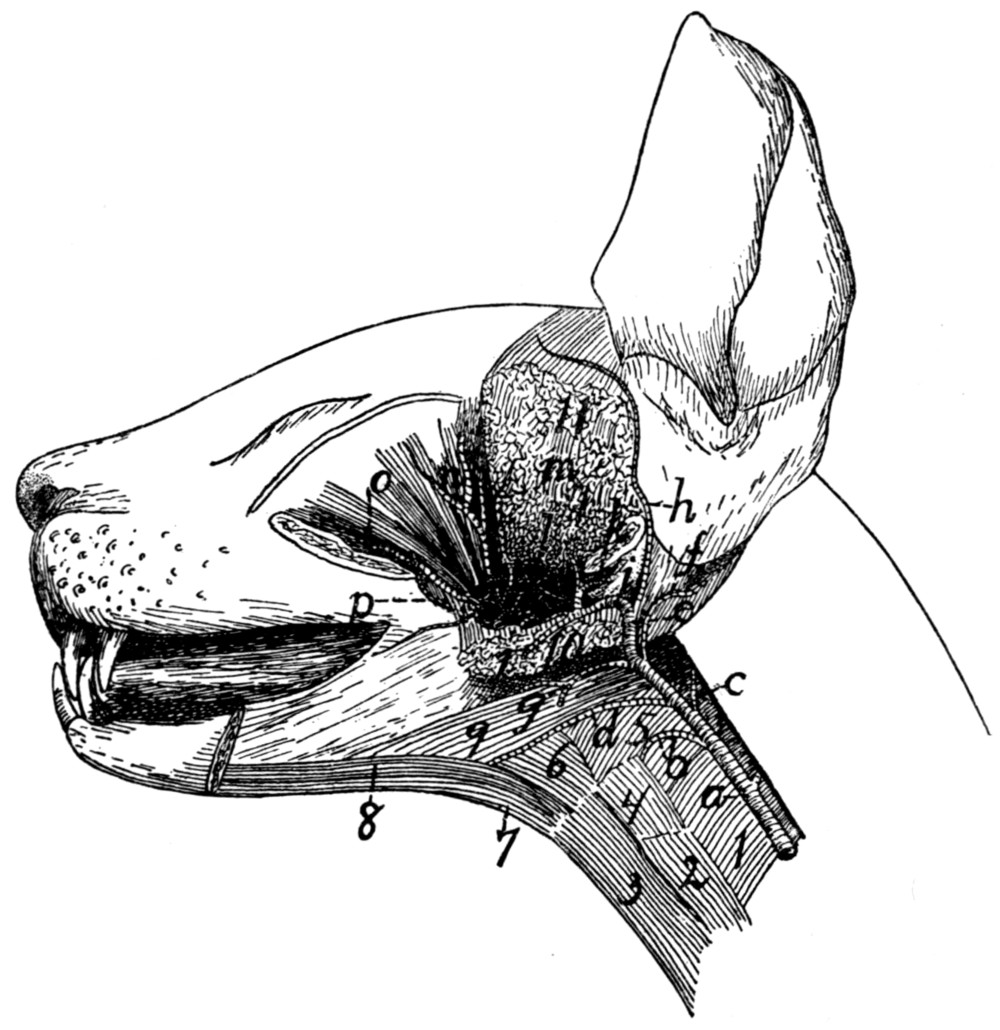{kind=link}
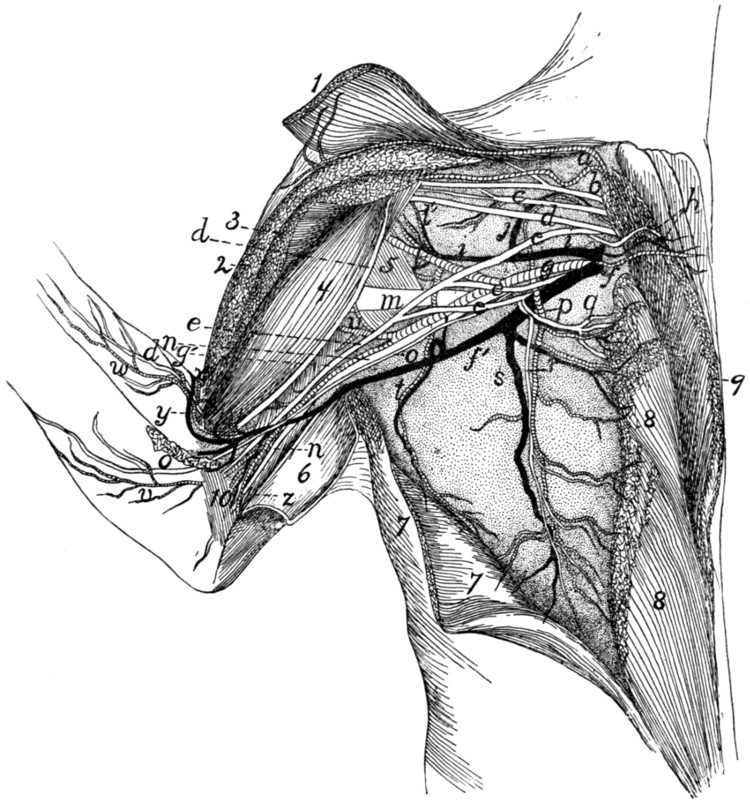{kind=link}
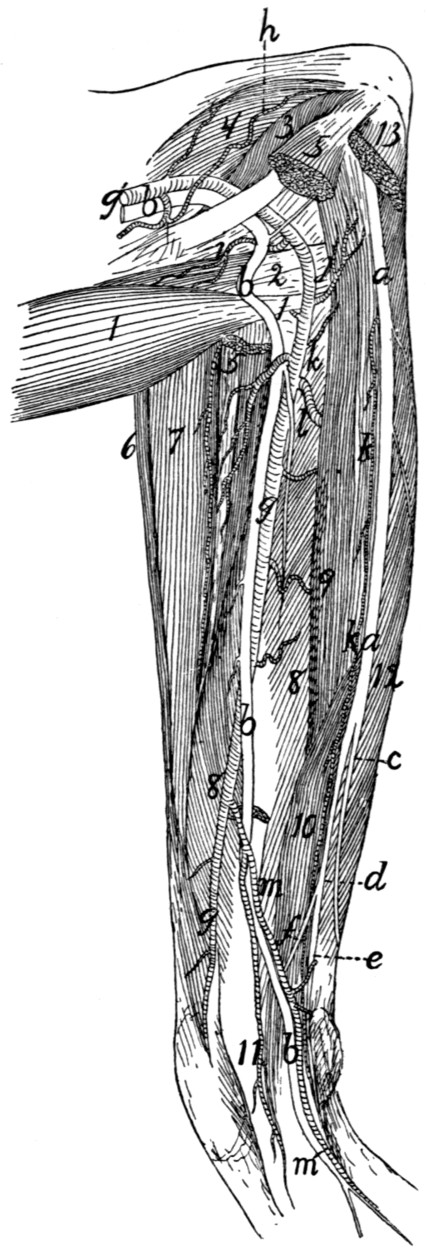{kind=link}
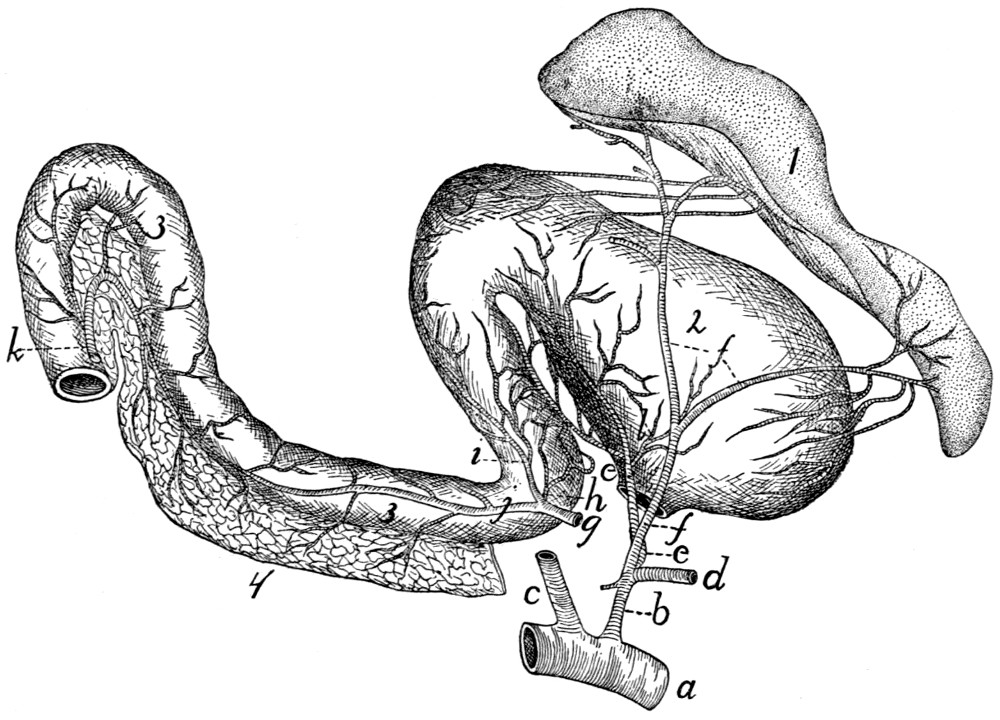{kind=link}
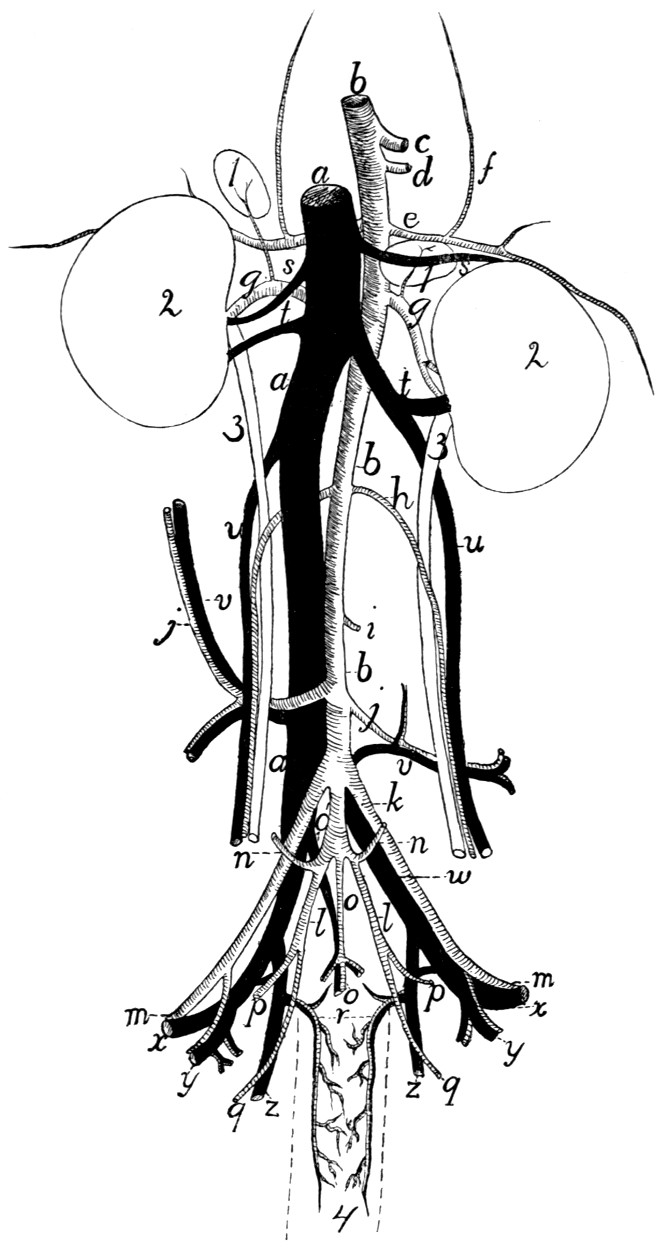{kind=link}
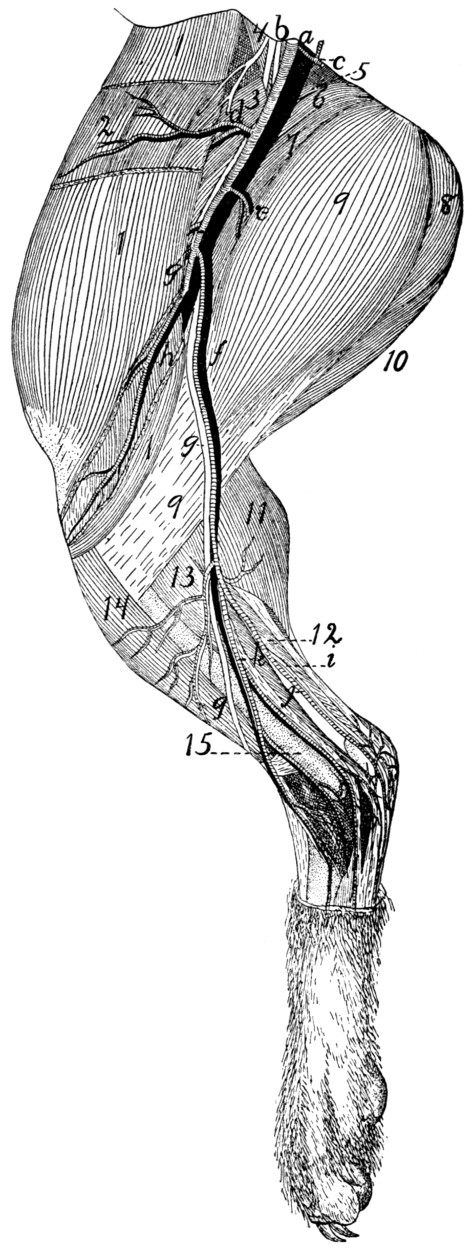{kind=link}
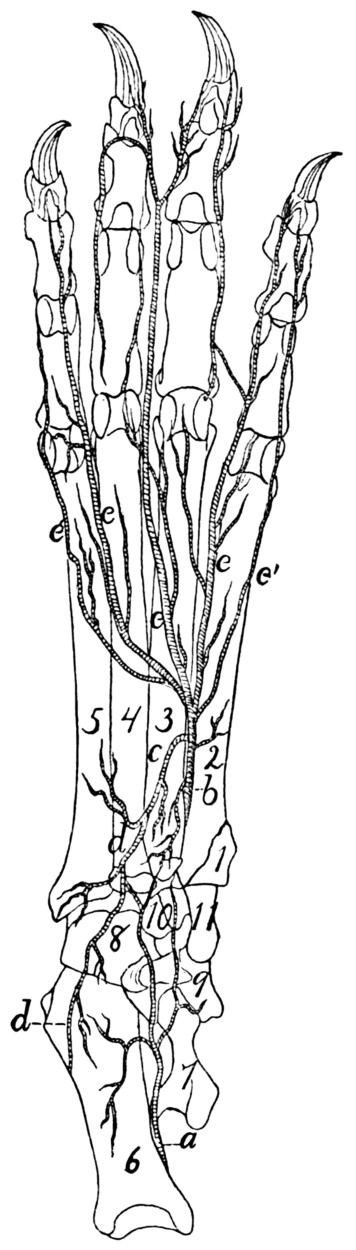{kind=link}
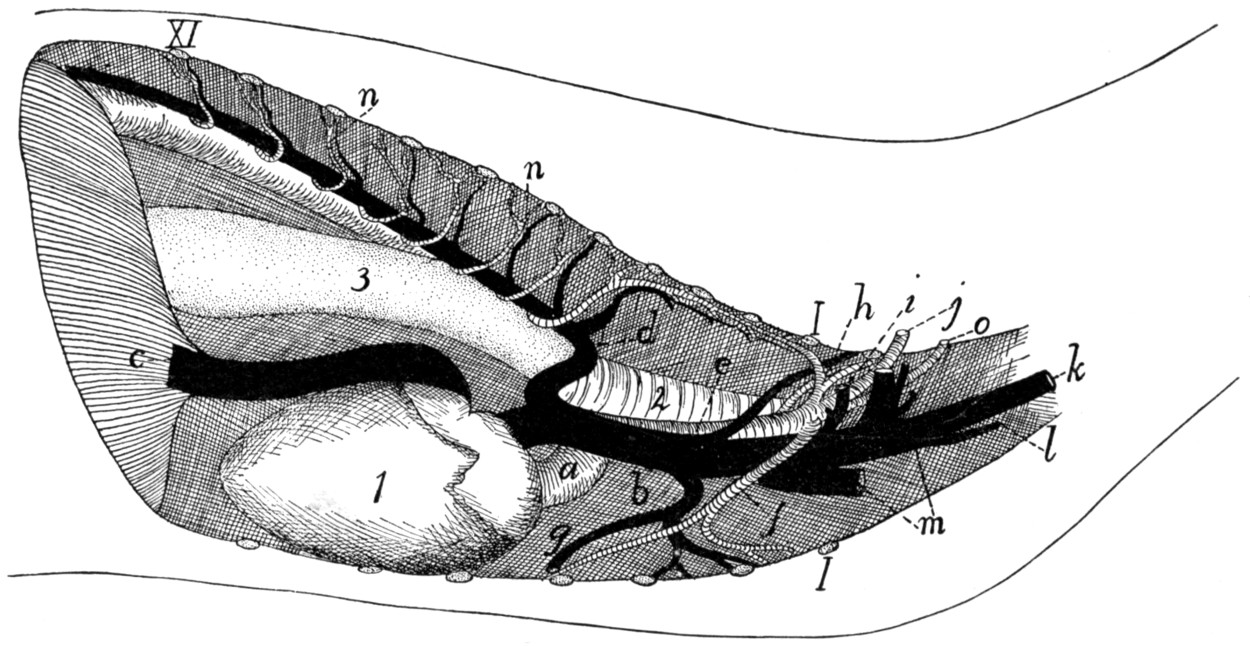{kind=link}
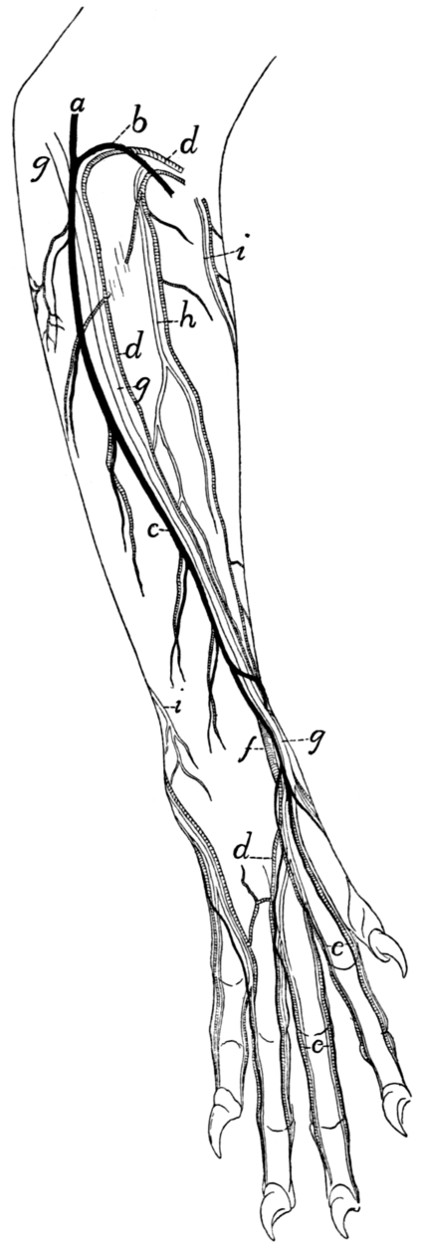{kind=link}
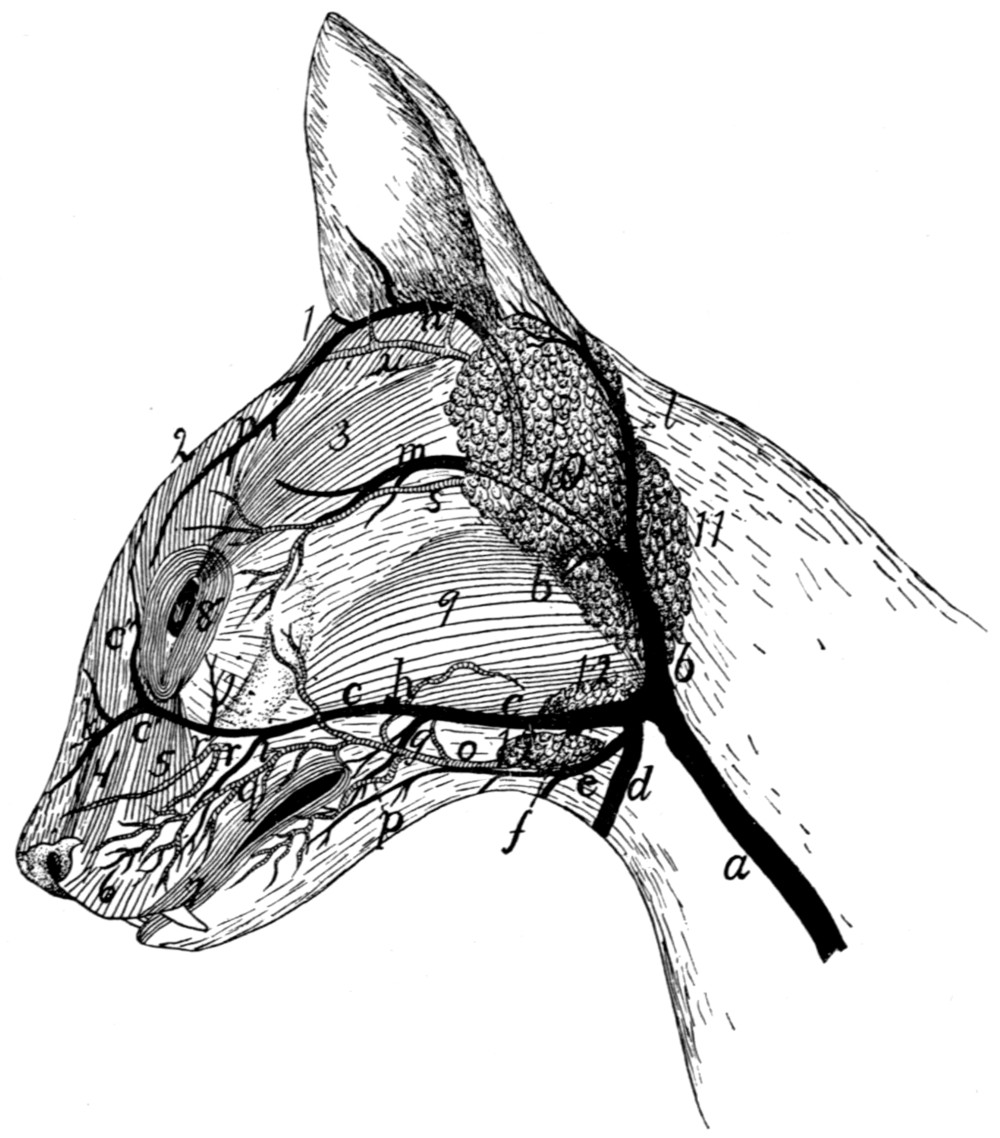{kind=link}
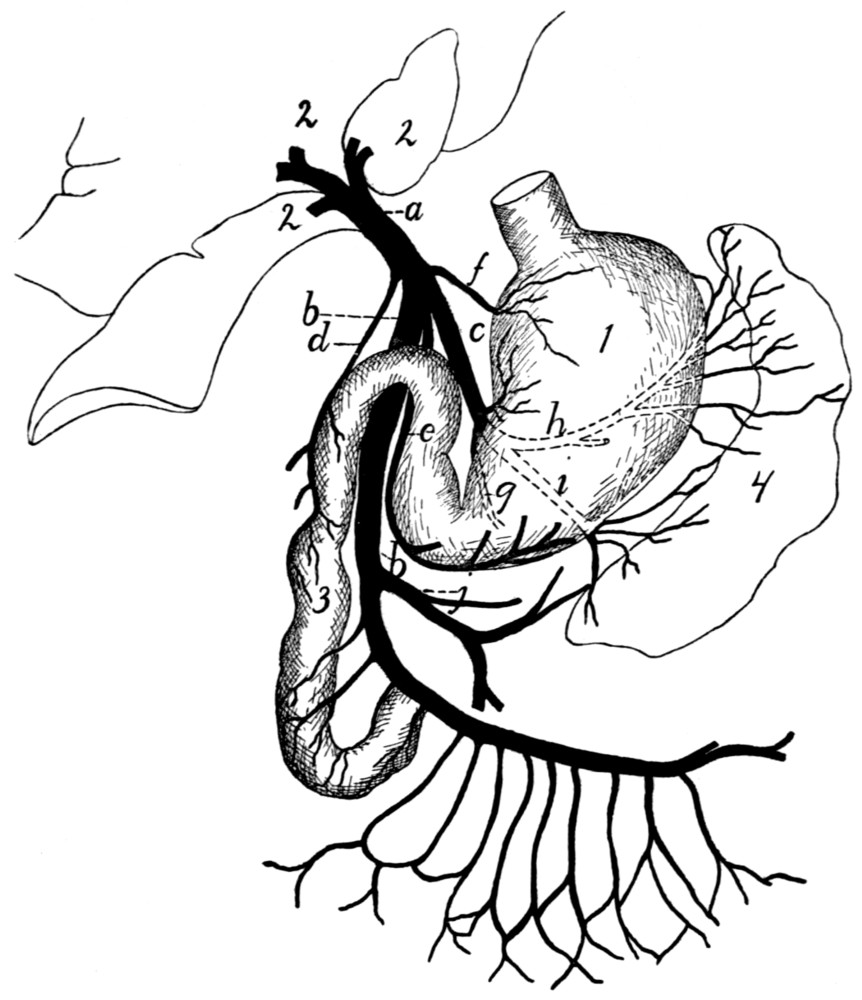{kind=link}
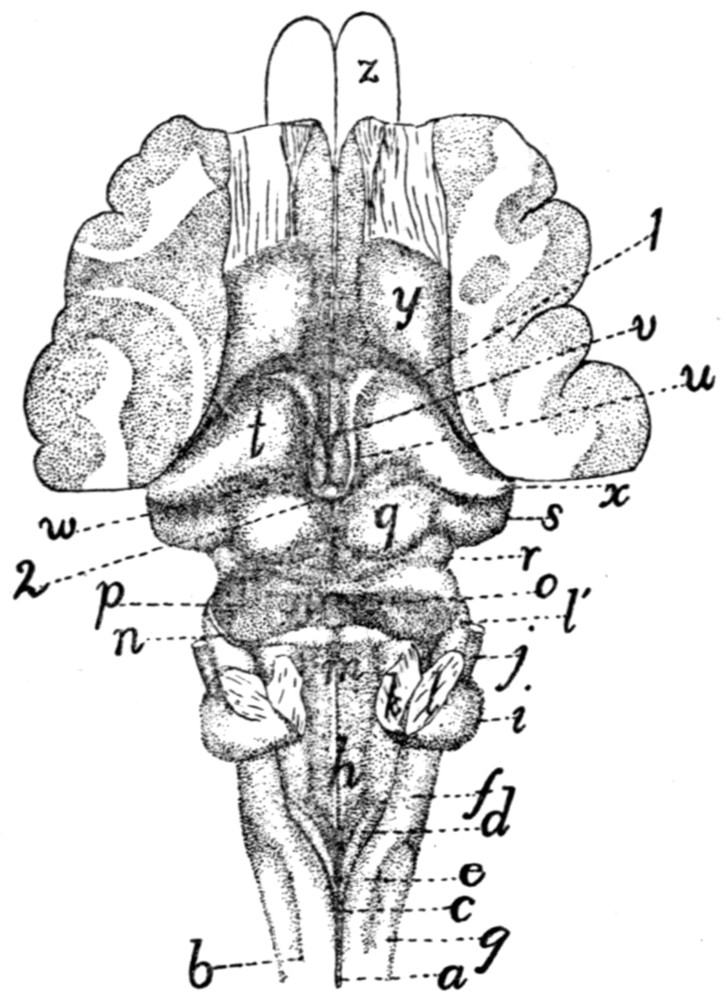{kind=link}
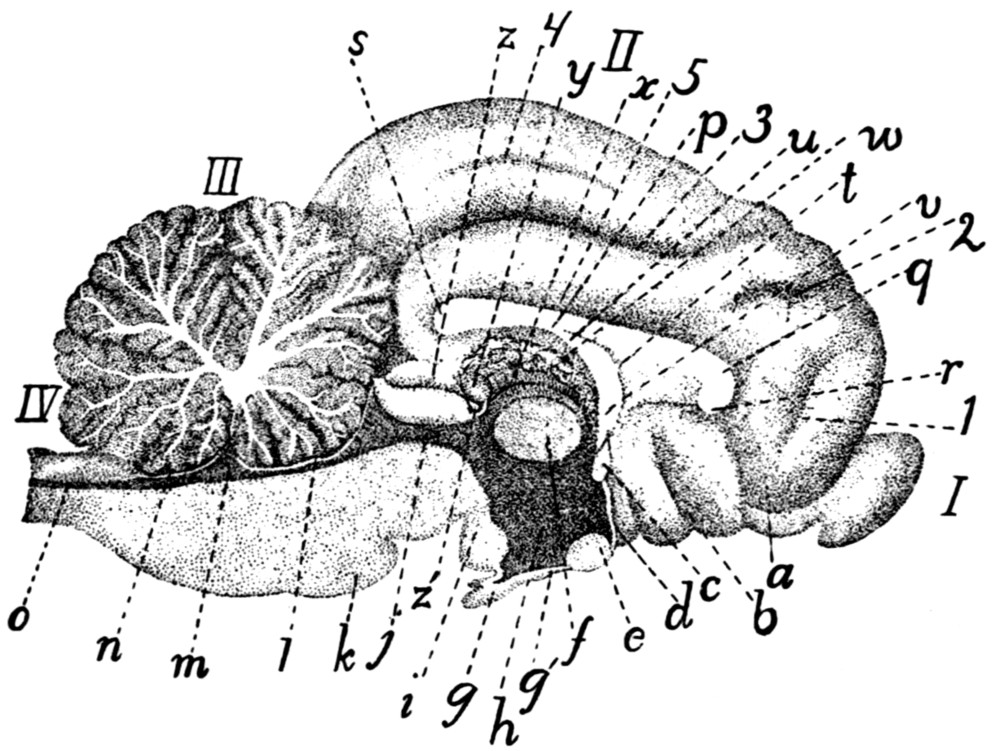{kind=link}
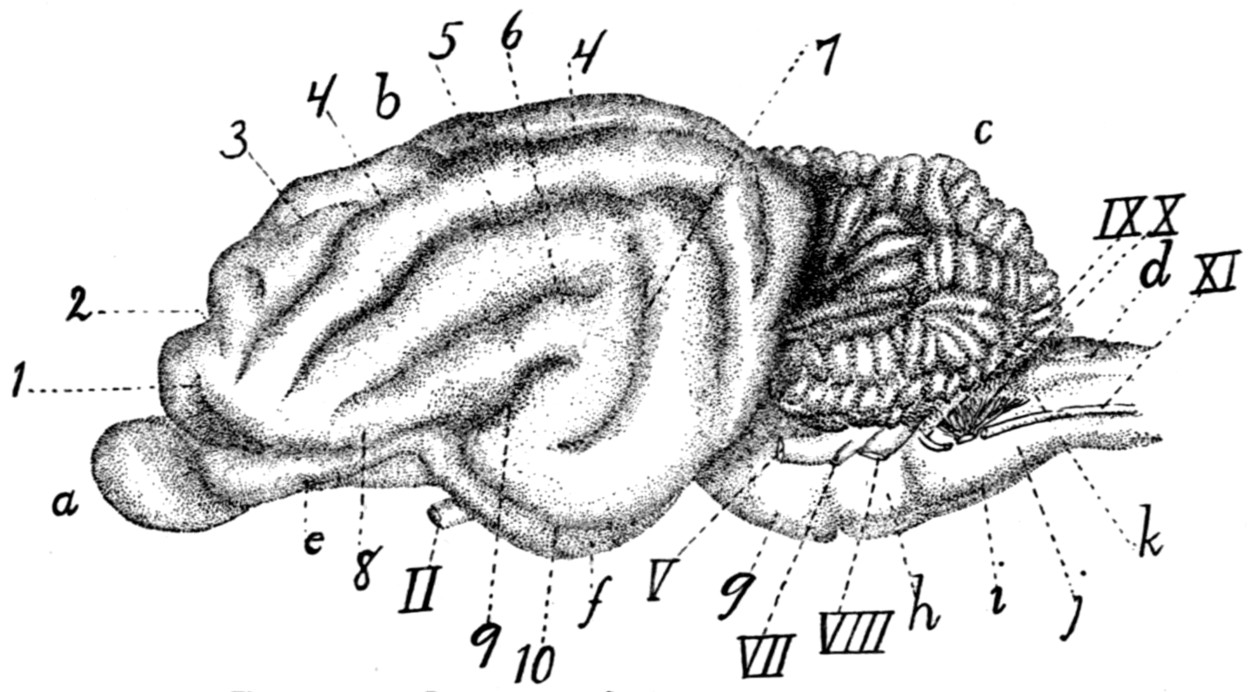{kind=link}
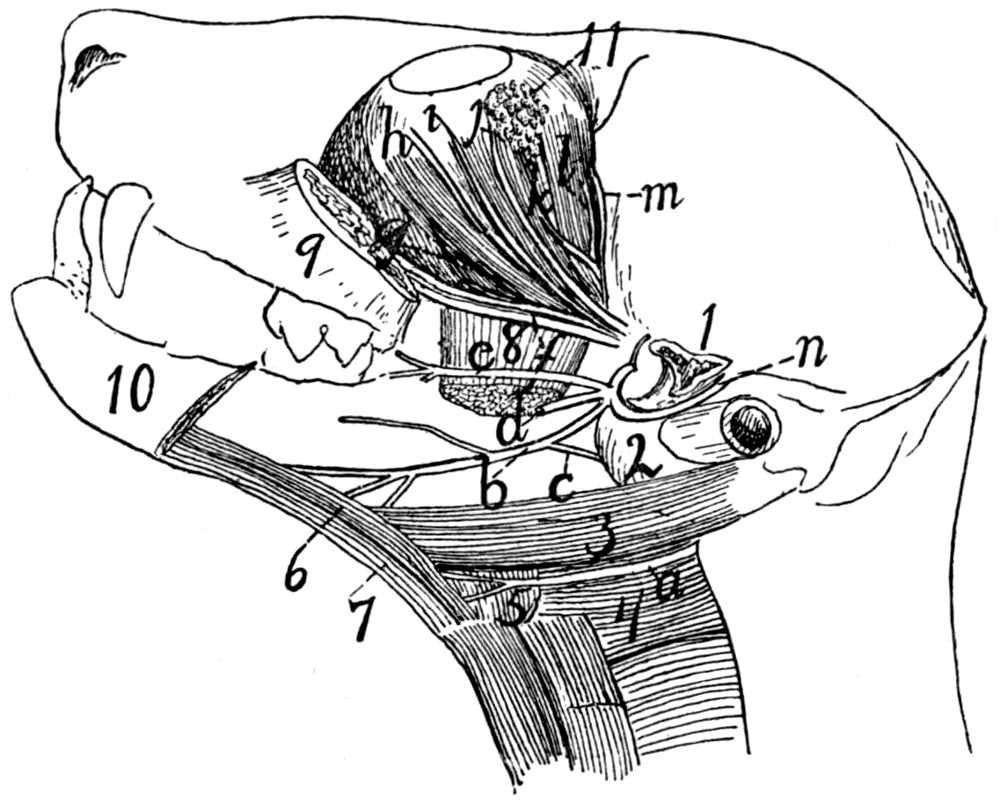{kind=link}
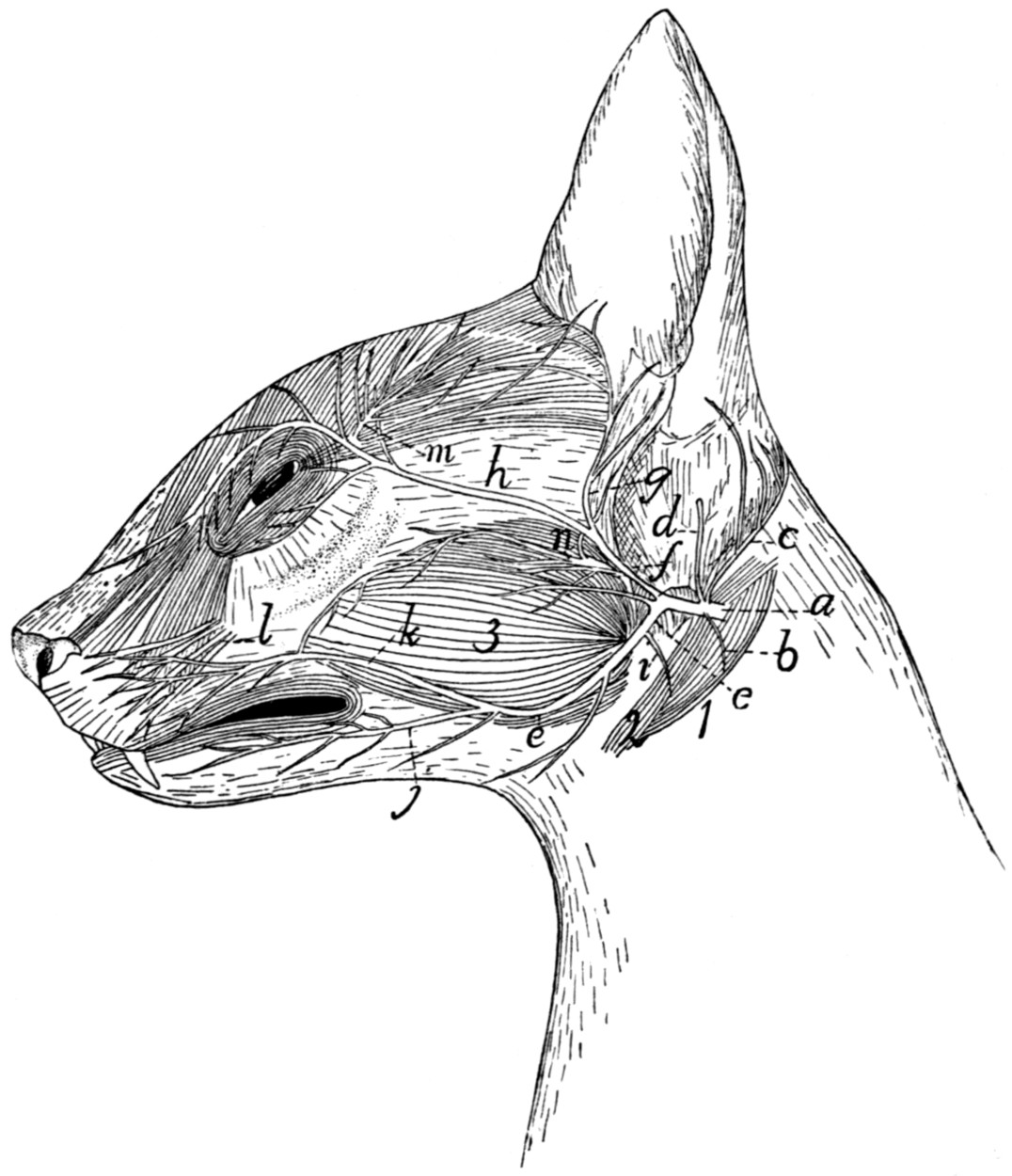{kind=link}
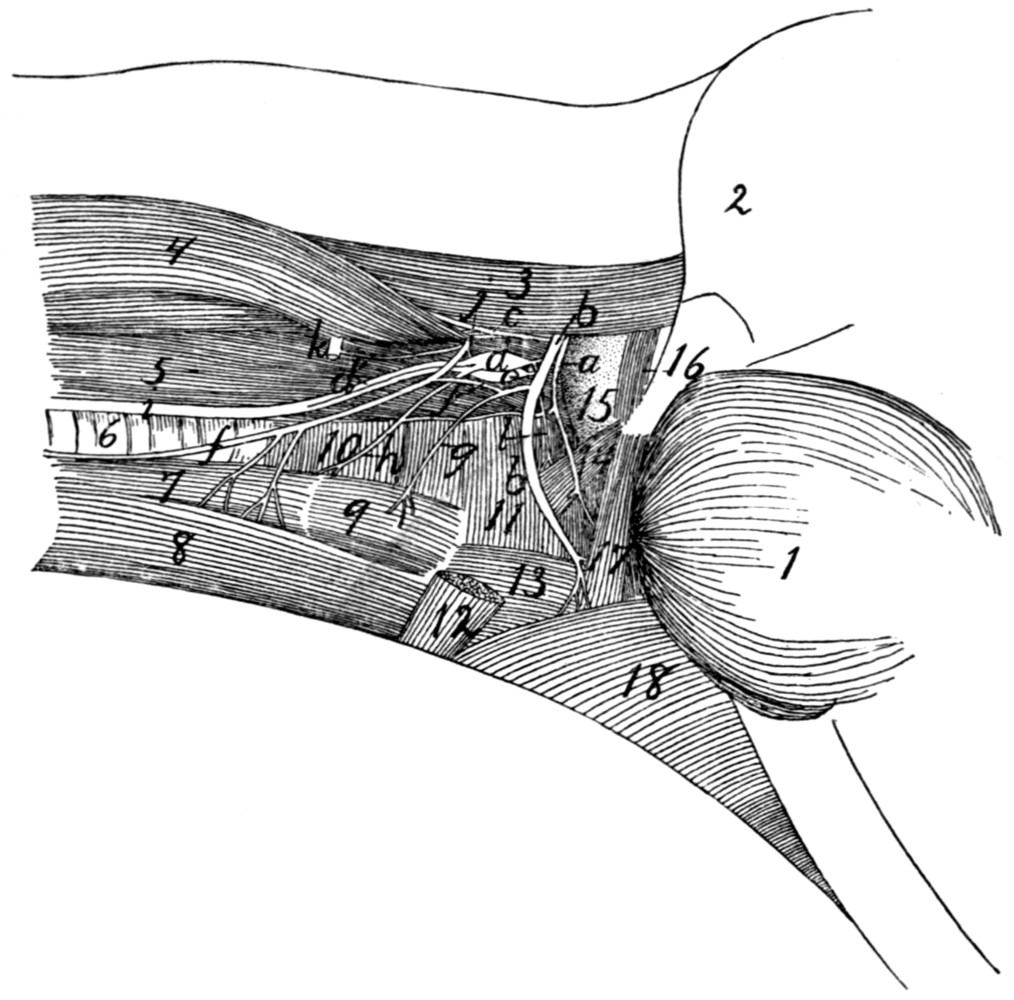{kind=link}
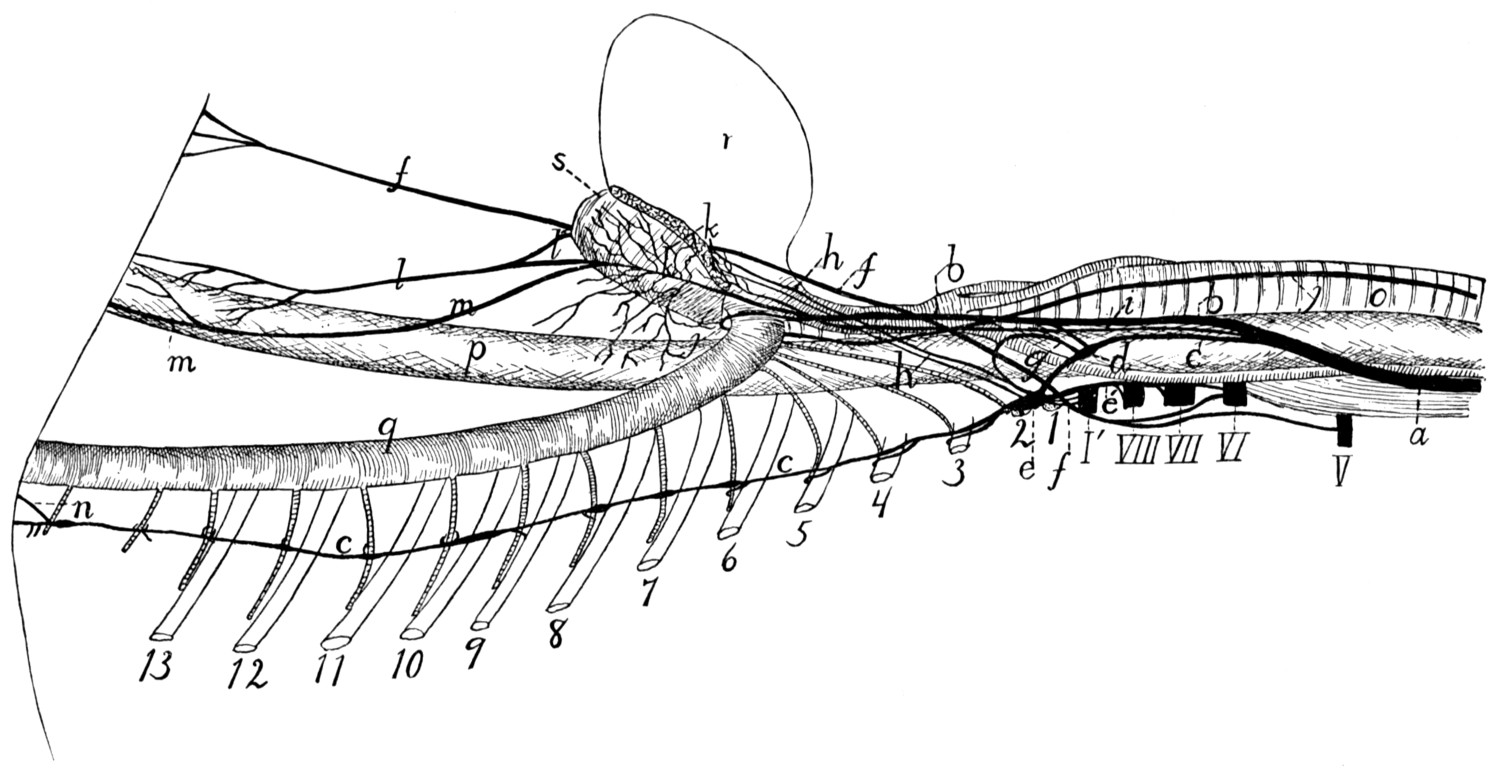{kind=link}
{kind=link}
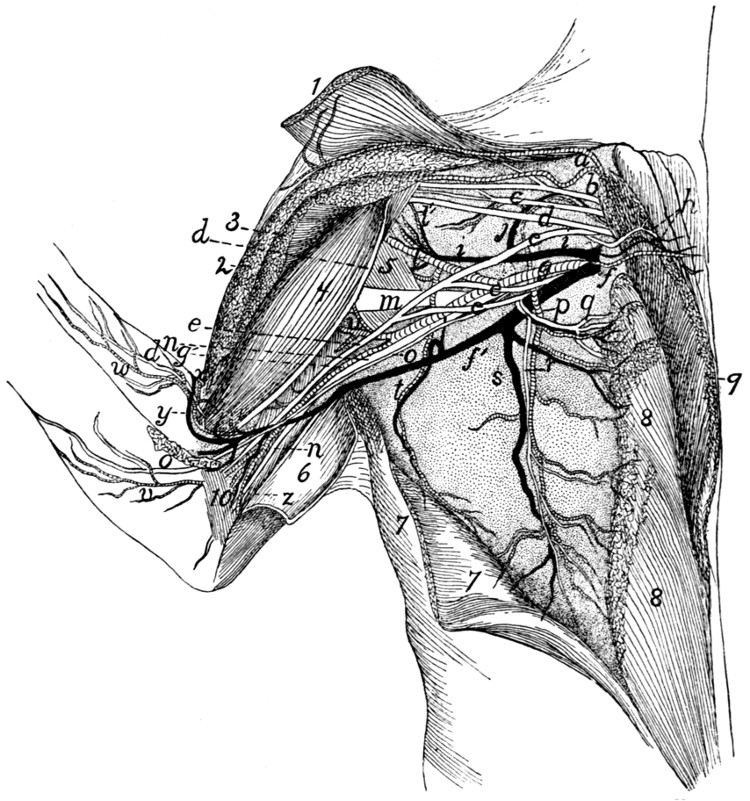{kind=link}
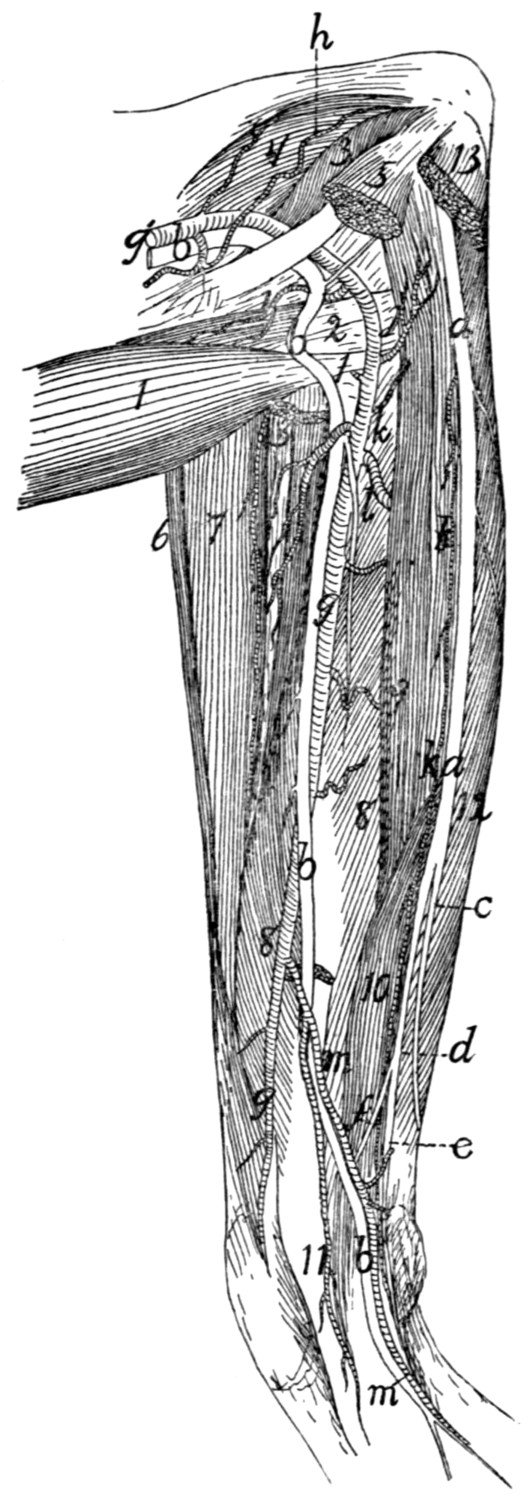{kind=link}
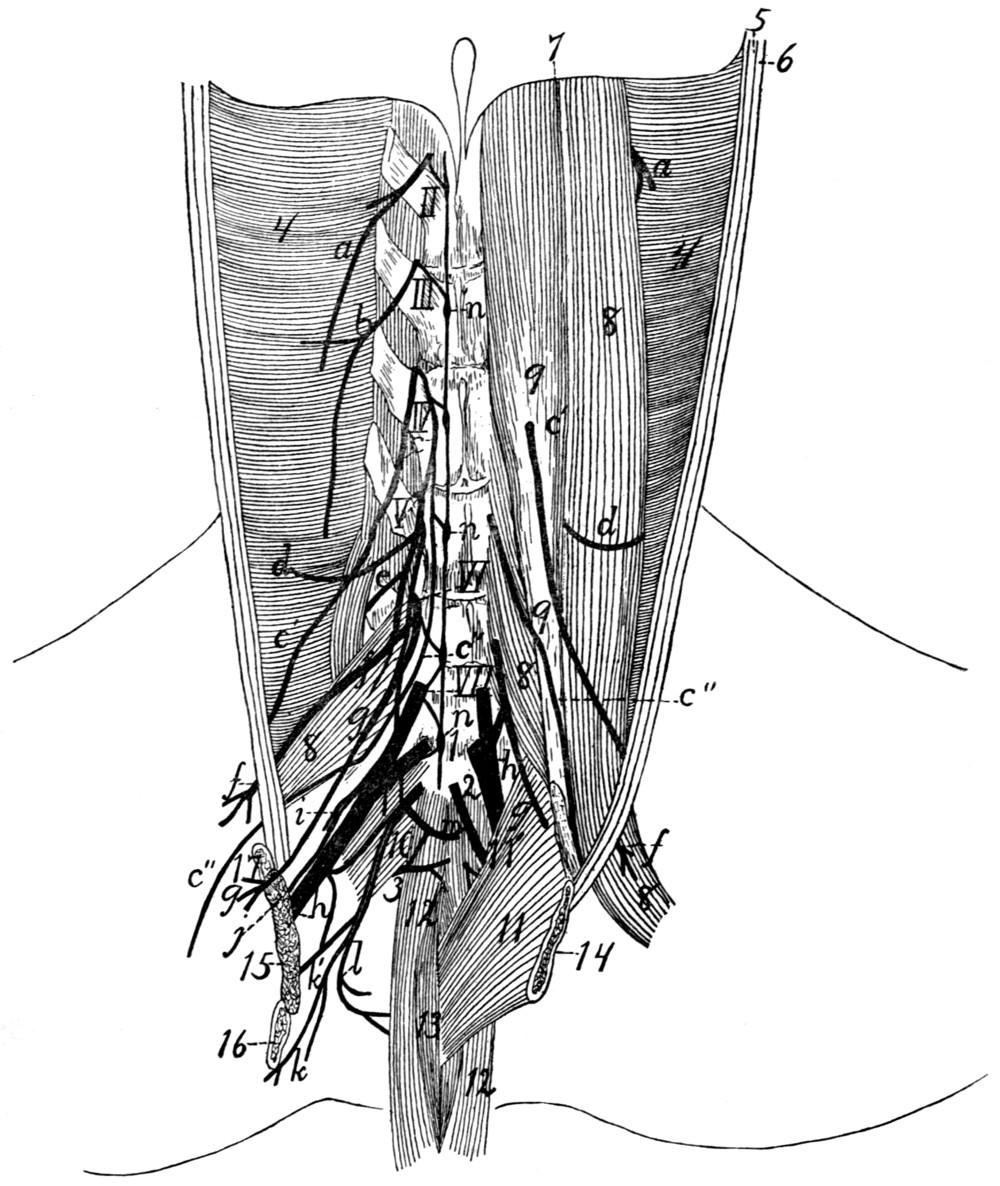{kind=link}
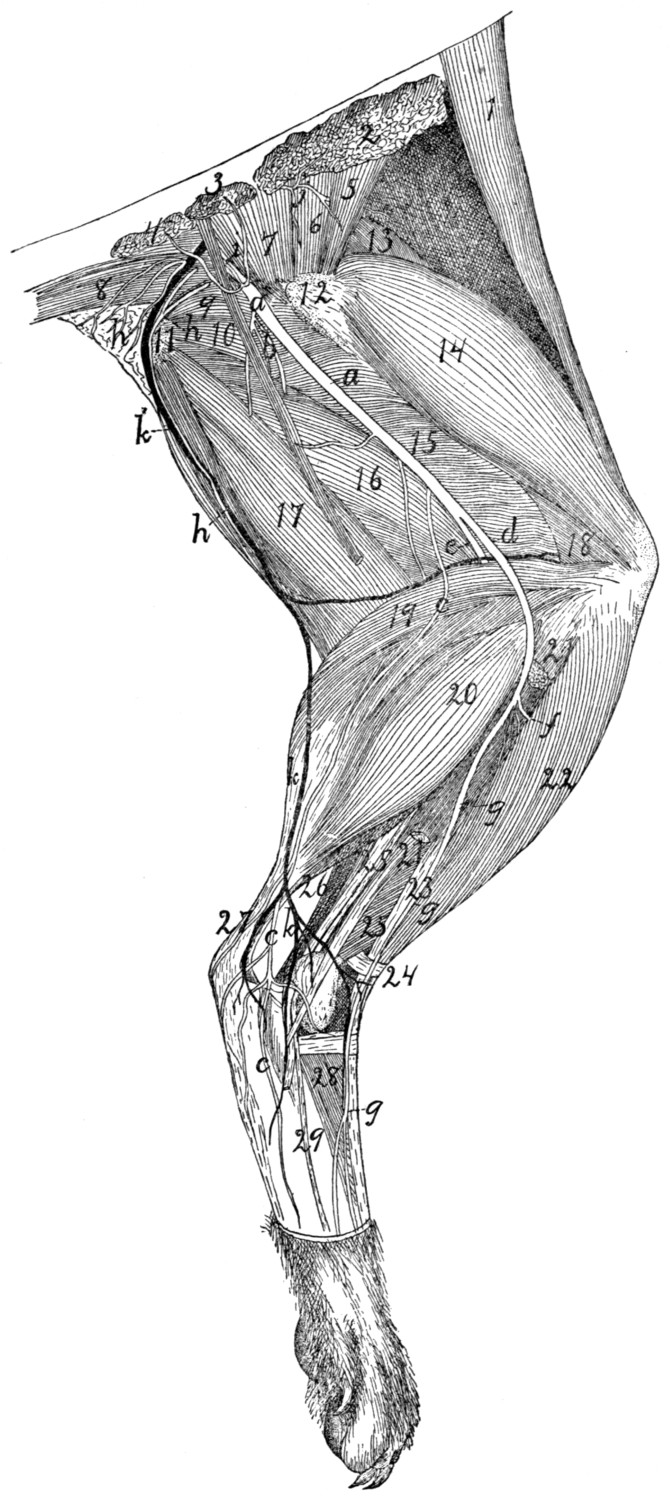{kind=link}
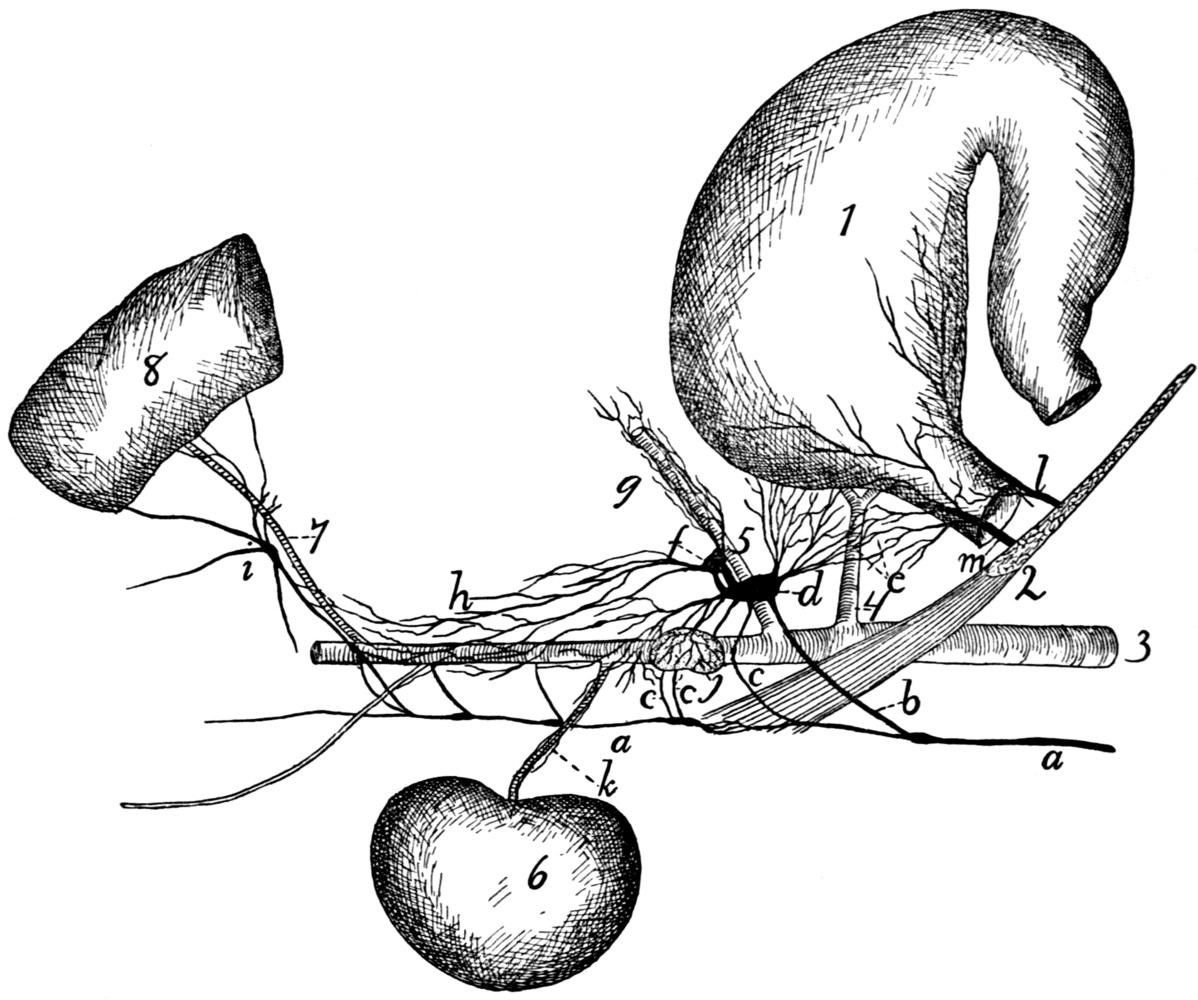{kind=link}
{kind=link}
{kind=link}
{kind=link}