The Project Gutenberg EBook of The Kansas University Science Bulletin, Vol. I, No. 2, February, 1902, by W. J. Baumgartner This eBook is for the use of anyone anywhere in the United States and most other parts of the world at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org. If you are not located in the United States, you'll have to check the laws of the country where you are located before using this ebook. Title: The Kansas University Science Bulletin, Vol. I, No. 2, February, 1902 Author: W. J. Baumgartner Release Date: June 8, 2017 [EBook #54868] Language: English Character set encoding: UTF-8 *** START OF THIS PROJECT GUTENBERG EBOOK KANSAS UNIVERSITY SCIENCE BULLETIN, FEB 1902 *** Produced by Larry B. Harrison, Paul Marshall and the Online Distributed Proofreading Team at http://www.pgdp.net (This book was produced from images made available by the HathiTrust Digital Library.)
(Whole Series, Vol. XI, No. 2.)
CONTENTS:
Spermatid Transformations in Gryllus assimilis,
with Special Reference to the Nebenkern.—W. J. Baumgartner.
PUBLISHED BY THE UNIVERSITY,
Lawrence, Kan.
Price of this number, 30 cents.
Entered at the post-office in Lawrence as second-class matter. [47]
Kansas University Science Bulletin.
| Vol. I, No. 2. | FEBRUARY, 1902. | { Whole Series, |
| { Vol. XI, No. 2. | ||
In Gryllus assimilis, with Special Reference to the Nebenkern.
(Thesis for the degree of Master of Arts.)
BY W. J. BAUMGARTNER.
With Plates II and III.
| I. | —Methods. |
| II. | —Material. |
| III. | —Observations. |
| (a) Nucleus. | |
| (b) Cytoplasmic Structures. | |
| 1. Nebenkern. | |
| 2. Acrosome. | |
| 3. Axial Filament. | |
| 4. Centrosome. | |
| 5. Cell Body. | |
| (c) The Spermatozoon. | |
| IV. | —Comparison of Results. |
| V. | —Summary. |
It is the object of these studies to follow carefully the spermatid transformations, and, if possible, to get a complete series of changes occurring therein. I do not pretend that I have as yet found all the stages, but in the present paper will publish observations made on Gryllus assimilis pertaining chiefly to the nebenkern, of which I have found some stages not previously described, as far as I know. I shall first describe my own findings, and then compare them with the results of others.
While other fixing agents were used, Flemming’s chromo-acetic-osmic mixture (strong) proved the best. Its fixation was satisfactory in all but a few stages, where the chemical changes going on in the cell are such that it seems difficult, probably impossible, to secure good, clear results; at least, neither Gilson’s nor Zenker’s mixture did any better on cells in these stages. [48]
Heidenhain’s iron-hæmatoxylin stain gave the best results, although Flemming’s three-color method was very satisfactory.
Most of the observations are made from sectioned material, but smear preparations were used for advanced spermatozoa.
The first observations were made in January of the present year on some material prepared by Professor McClung, to whom I wish here to make most grateful acknowledgment for proposing the line of investigation, and for many helpful suggestions during the progress of the same. The testes were those of adult insects, and showed mostly only mature or almost mature spermatozoa. Enough spermatids were seen to indicate that their transformation into spermatozoa was somewhat peculiar.
The material for the drawings was taken from specimens collected under stones and boards on and near the University campus, at Lawrence, Kan. Young nymphs of Gryllus were caught as early as March 1, but the cells of the testes were all in the spermatogonia and spermatocyte stages. During the early part of May, after the nymphs had passed their third molt, crickets were secured whose testes showed all stages of development and transformation.
Gryllus assimilis, the common black field cricket, has paired testes lying in the anterior dorsal part of the abdomen. These have a whitish transparent appearance, which becomes duller in the adult, showing sometimes a slight yellowish tinge. The shape of each is that of a somewhat conically rounded body, not unlike a flattened strawberry. Each testis consists of a central rachis about 3 mm. long, from which extend a large number of curved follicles varying in length from 1 to 3 mm.
The follicles are larger toward the blind end and taper toward the rachis. Each follicle is divided into cysts, but more often transversely than longitudinally, for frequently one cyst occupies the whole follicle in cross-section. The cysts toward the rachis end of the follicle are much longer and narrower than those of the blind end. The cells are not very large and contain twenty-four chromosomes in the spermatogonial generations; eleven and twelve were most frequently found after the reduction. [49]
The follicles of the cricket testis show the different cell generations and the same relative arrangement of them as McClung (’00) found in Hippiscus, and Sutton (’00) in Brachystola. The spermatogonia are nearest the blind end of the follicles, spermatocytes next, and the spermatids following the latter. The cells of a cyst are not, as in Anasa (Paulmier, ’99), “in the same stage of development,” but only approximately so, for some cysts show cells in the metaphase, while others have reached the telophase. Successive cysts, as in Brachystola, do not show successive stages; for, frequently, growing spermatocytes and late spermatids, or even young spermatozoa, were observed side by side. The individual cells in division pass through prophases, metaphases, anaphases, and telophases—these terms being used according to their usually accepted meanings. (See McClung, ’00.) In this paper I shall begin with the cell when it has reached the telophase of the second spermatocyte division; that is, just after the chromosomes have reached the poles of the spindle.
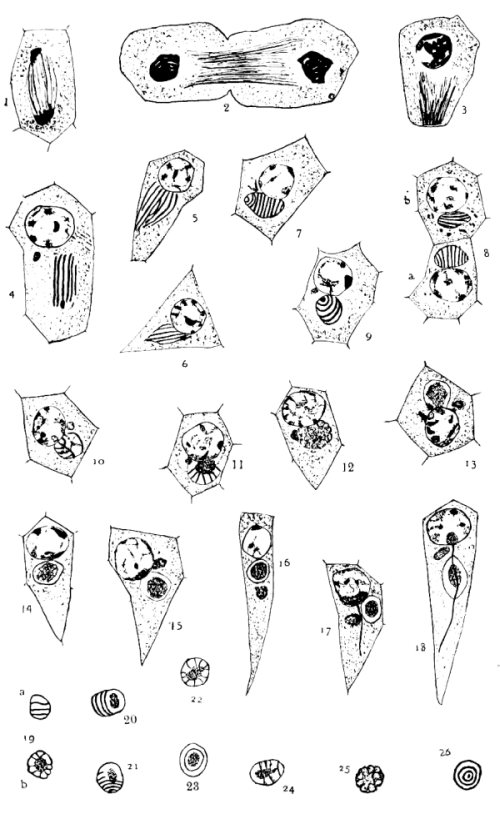
The chromatin in Gryllus behaves very much as Henking (’91) has described for Pyrrhocoris, and Paulmier (’99) for Anasa. At the end of the anaphase the chromosomes are crowded together at the poles of the spindle. (Fig. 1.) A clear space begins to be formed around them, especially on the equatorial side. (Fig. 2.) They then separate and are scattered somewhat evenly over the nuclear membrane when it is formed, as seen in figs. 4, 5, and 6. Soon protuberances appear on the surface of the individual chromosomes, making their outlines irregular. At about this time they begin to show a granular structure. The chromosomes break up more and more, and soon the chromatin appears in irregularly arranged patches or blotches of granules of various sizes. (Fig. 8.)
Meanwhile the nucleus has slowly increased in size. It reaches its maximum, which is probably about treble its original diameter, before the cell begins to elongate. The granules have been breaking up into finer and finer pieces, until, a little after the cell has reached its maximum, the chromatin has largely disappeared, or, to be more [50] exact, has lost its affinity for stains. (Fig. 16.) As the cell begins to elongate, the nucleus goes to one end, namely, that toward the periphery of the cyst. (Figs. 14 and 15.) The chromatin again frequently appears in patches as the cell decreases in size. (Figs. 33 and 36.) In many cells the patches never disappear. When it has diminished to less than half its maximum diameter, the nucleus begins to elongate, becoming elliptical, and finally forms the long, tube-like head of the spermatozoon. (Figs. 43 and 45.)
I found here that the nucleus was hollow, or, better, that it showed a clear space within. For a long time it contains the many chromatin granules, as seen in figs. 39, 40, and 41; but at last the walls seem to attract all these granular masses, and the center is entirely clear, as in fig. 45. In fig. 44 are shown cross-sections of spermatozoa heads, of the stage of fig. 43. In fig. 45 we have a mature spermatozoon head.
The accessory chromosome in Gryllus lags behind during the spermatogonial divisions, just as in Xiphidium (McClung, ’99), and retains its identity all through the growth period, in many cases becoming quite large. At first it cannot be distinguished in the spermatid, but soon its stronger stain and exemption from disintegration make it apparent. (Fig. 4.) It is flattened against the nuclear wall and, unlike Paulmier’s “small chromosome,” it does not break up but remains intact, as Henking and McClung have described. (Figs. 14, 18, 32, 37, and others.) It gets larger and then smaller, and is finally lost in the rest of the chromatin, as the latter condenses to form the spermatozoon head. I was not able to see that it occupied one side of the elongating head, as McClung describes it in Xiphidium. The darker stain and regular contour, and sometimes its greater size, made it in most stages quite evident.
It is in the cytoplasmic structures that Gryllus shows some things that have not been described, so far as I know; yet I almost hesitate to enter the discussion concerning the nebenkern, the mitosome, the idiozome, the archoplasm, the attraction sphere, the acrosome, the “mitochondrion körper” and other bodies which have been described in the extra-nuclear parts of the germ-cell. But because I have been unable to find in any of the papers a description of a structure like the one in Gryllus, I shall proceed to describe the nebenkern as I find it. I shall interpret the appearances as far as possible, leaving to others, or to later efforts, the complete harmonizing of this element with the structures of other species. [51]
Before I proceed, I would add my protest against the name “nebenkern,” as voiced by Calkins (’95), Erlanger (’96), Meves (’00), and others. It is inapt and not at all descriptive or distinctive. But since we have the word, and investigators will use it to mean something, I think it would be best to restrict the word, as Calkins and Erlanger suggest, to the remaining spindle fibers and connective fibers which go to each spermatid and which have an important part in forming the tail membrane. I shall retain the name for a further reason, which St. George (’97) gives for first using it, “weil sie die Frage nach der Herkunft dieses Gebildes offen lässt.” A comparison of the literature will convince any one that the origin of this element is still a much-disputed question.
There are, in Gryllus spermatids, two quite permanent and prominent extra-nuclear bodies. The larger of these I shall call nebenkern, because I believe it originates from the spindle remains, and goes to form the tail covering. Its behavior is as follows: At the beginning of the telophase the fibers which had connected the centrosomes with the chromosomes bulge out like barrel staves, as Henking (’91) has described. These break loose from the chromosomes and centrosomes as the clear space begins to form around the chromatin mass (fig. 2); but they are not separated into two rings, as in Anasa and Pyrrhocoris. As the dividing cell membrane is formed, the middle portions of the fibers are drawn together and the so-called “mid-body” is formed. Some of the fibers unite, producing thicker ones, as seen in figs. 1, 2, and 3. In the last-named figure some of the central fibers seem to be still united with those of the companion spermatids.
As the daughter-cell shortens and the nucleus takes a more central position these fibers unite still more, shorten, bend together (figs. 4, 5, and 6), and finally assume the shape shown in fig. 7. I shall call it for the present the “striated condition,” or simply “striated nebenkern.” The shape resembles that of an egg with rather sharply drawn-out ends. The surface is occupied at intervals with deeply [52] staining lines which look much like hoops. In most cases the long axis of the nebenkern is at right angles to the axis of the spindle from which it was formed, while the striæ, or dark lines, are in planes parallel to it, as is seen in figs. 7 and 8a. In a few cases the lines are parallel to the long axis of the nebenkern, as shown in fig. 8b. Fig. 9 exhibits a partial end view of the nebenkern, while fig. 26 represents a polar view in all planes.
The fibers in figs. 4 and 5 are not all in the same plane, giving frequently, with changing focus, the impression that the lines are the edges of plates. Even in figs. 7 and 8, the appearances shown made me hesitate for some time before I dared say that the lines represent striæ on the surface and not plates extending through the body. But the end views (figs. 9 and 26) and the two drawings in fig. 19 show clearly that we are dealing with encircling hoops and not dividing plates. Just how or why the fibers bend and assume the shape of figs. 7 and 8 I am not able to say, but I am sure it is not an artefact, as might be supposed, because I have found them occupying whole cysts of several testes, and nearly all of them were as regular and definite as fig. 7.[A]
I have found the nebenkern in its perfect condition (fig. 7) only in material fixed in Flemming; although Zenker’s fixation showed some of the breaking-up stages. This may be due partly to poorer fixation in other mixtures and partly to chance, as not so many testes fixed in other agents were examined, and the appearance evidently represents a very brief stage. These fibers now begin to break up, and we get figures that remind one of Meves’s “mitochondrion körper”; that is, a darker staining center and radiating lines to the outer ring. (Fig.11.) Sometimes stages a little later recall Paulmier’s “blackberry stage.” (Figs. 12 and 25.) Fig. 19a is a surface view and fig. 19b an optical section of a nebenkern that is in the process of breaking up. Figs. 20 and 21 have a part of one end cut off, while fig. 23 is a polar view. Fig. 22 is probably a forming blackberry stage. [53]
All the above are brief stages, and we soon get the appearance of figs. 13-16. Here we have the darkly staining central ball surrounded by a clear space, enclosed by a ring, as Meves (’00) found it in Pygæra. The central ball sometimes shows vacuoles (fig. 16), but most frequently stains as one mass. In some cysts with cells resembling fig. 16, it is difficult to distinguish between nucleus and nebenkern, yet the latter’s stain is never as intense as that of chromatin. The stages shown in figs. 13 and 14 persist for a long time, and it is probable that this is the end of the telophase and the beginning of the spermatid transformations. I have treated the above more in detail and shown more drawings than its importance may deserve, but it is in these stages that Gryllus seems to differ from other species, and I have tried to describe and illustrate the appearances and changes fully.
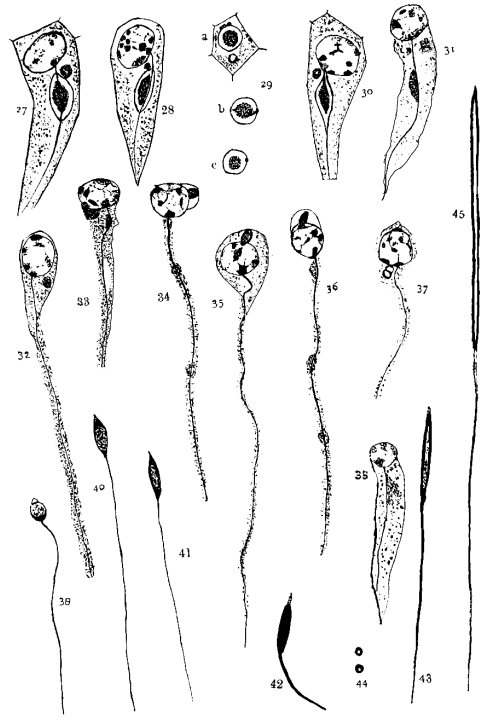
As the cell elongates, the axial filament grows out, the nebenkern approaches it, and a junction is effected in such a way that the axial filament runs over the surface of the nebenkern. (Figs. 18, 27, 28, and 30.) At first the nebenkern is still almost round, but it begins to elongate and the dark inner ball sends out a protrusion to the nucleus, and sometimes back to the point of separation between the external envelope and the axial filament. The outside ring disappears and the dark mass moves down the axial filament. (Fig. 31.) In many cases it breaks up into several small drops, which appear at intervals on the axial filaments. (Figs. 34 and 36.) In fig. 38 is seen such a hanging drop. In this way the nebenkern material is distributed over the axial filament and forms a sheath around it.
[A] Professor McClung, Mr. Sutton, and Mr. Blackman, of Kansas University, and Doctor Child and Mr. Harper, of Chicago University, have seen my preparations, and all confirm my statement that the appearance is not an artefact.
The second extra-nuclear body I shall call the acrosome, as it forms the point of the spermatozoon head. I could not trace its origin, because the fixation was not definite in some of the early stages. Fig. 4 shows a small, darkly staining body, which may be a centrosome, but more likely is the beginning of the acrosome, of which figs. 7, 9 and 10 show developing stages, and figs. 11, 13 and 15 more advanced ones. Sometimes the central portion stains darkest (figs. 11, 32, and 36), but more frequently there is a small clearer space in the center surrounded by a ring or band of darker material. (Figs. 10, 13, 27, and 30.) This ring usually stains darker on one side, very often on the side toward the nucleus. [54]
The position, which is, with few exceptions, in the angle between the nucleus and the nebenkern, induces me to consider the differently staining bodies as the same organ of the cell. For some time after the nebenkern has disappeared the acrosome keeps its position; then it approaches the nuclear wall, flattens against it, and later wanders to the apical end, where it forms the tip of the spermatozoon. (See figs. 32-36 and 39-42.) Fig. 16 shows the acrosome back of the nebenkern. It is not an isolated example, still, I think, it is an abnormality. Fig. 37 shows an apparent division of the acrosome. I did not see enough instances to consider it a regular occurrence.
As the cell is elongating, the axial filament is seen apparently growing out of the nucleus. I do not mean to say that it grows out of the nuclear substance, but in almost every cell observed, where the axial filament was incomplete, there was rather a large mass of chromatin gathered at the place where the axial filament was attached to the nucleus. It may be that the smaller one of the extra-nuclear bodies in figs. 4, 7 and 9 is a centrosome which is passing to the equatorial region of the nucleus, where it later develops the axial filament. As already noted, the axial filament does not pass through the nebenkern, but only over its surface. Fig. 29, a, b, and c, shows different cross-sections through the elongating cell. In b the axial filament was cut at an angle.
I have not followed the centrosome through its migrations. In fig. 2 one of the two bodies is the centrosome, but I did not trace its changes farther.
The cells, up to the stage of figs. 13 or 14, have been scattered promiscuously through the cyst; but as the cell begins to elongate, the one end becomes the anterior-nuclear end, and it shifts to the periphery of the cyst. The central part of the cyst now shows the so-called central lumen. As the lengthening goes on the heads of the forming spermatozoa are all turned toward the distal end of the cyst. The cyst becomes very narrow and long, apparently preparing to contain the long, slender spermatozoa. Near the rachis the spermatozoa seem much twisted, and I surmise that they turn so as to have the head foremost when set free into the vas deferens. Fig. 42 may indicate some such behavior. [55]
The mature spermatozoon consists: (1) of a sharp, spear-like point; (2) a tube-like head about .02 mm. long, composed of a densely staining outer wall and a clearer central cavity; and (3) a filamentous tail about .5 mm. long. The point comes from the acrosome, the head from the nucleus, and the tail from the axial filament and nebenkern. While I do not deny the presence of a middle piece, my observations do not warrant me in describing one, although fig. 40 would suggest it.
Dr. R. von Erlanger (’97, 1, and ’97, 2) and Dr. F. Meves (’00) have given excellent discussions of the literature on the nebenkern. I cannot do better than direct the attention of other investigators to them. Hence, I shall discuss only such authors and such points as have special bearing on my own results. The spermatid changes in the Gryllidæ have been studied, so far as I know, only by St. George (’67) and by vom Rath (’92).
St. George, in his early paper, described the transformation process in the “Hausgrille” along with other insects. He found the “Nebenkörper,” and saw that it took a part in forming the envelope of the axial filament, a small particle remaining near the nucleus and other drops of it appearing at intervals on the tail. In his paper on Blatta (’86, 1) he accepted Bütchli’s name, “nebenkern,” and traced its origin to the spindle remains. In his paper on Phratora (’86, 2) he takes up the description of the nebenkern in the spermatocytes and spermatogonia. In the spermatid, he describes the blackberry stage in these words: “Während das andere (not nucleus) Körperchen weniger lebhaft gefärbt als ein kleiner Fadenknäuel mit unregelmässigem Contour erscheint.”
There can be no true nebenkern in the spermatogonia and spermatocytes, as St. George first used the word, and as I have limited its application. But St. George found the object, traced its origin and fate, and described several of the stages as they appear in Gryllus.
Vom Rath (’92) found Gryllotalpa very poor material for following the spermatid changes. However, he described an oval body which he found varied in appearance, depending on the method of fixation. He calls it the “nebenkern” and thinks it goes to help form the tail.
The description is too incomplete for comparison. Judging from Gryllus, I question its being very poor material. [56]
Bütschli (’71) studied the spermatid transformations in Acrididæ and Locustidæ along with that of other animals. He saw the object formerly described by St. George and named it “nebenkern.” He saw it divide into halves, elongate, and form the spermatozoon tail.
The dividing of the nebenkern into halves seems to be an appearance quite common in insect spermatogenesis. I have myself seen it in several genera of Acrididæ—Hippiscus, Arphia, Melanoplus, and Brachystola; besides Bütschli, St. George, Henneguy, Platner, Paulmier and others have described it.
Platner in his studies has given special attention to the nebenkern. In his first paper on “Pulmonates” (’85) he did not trace the origin of the nebenkern, but described it as consisting of four to six rods of different lengths and irregularly bent. These were connected, forming an irregular polygon. In its later stages he saw it with a mass of protoplasm pass down along the primary tail—an early protrusion of protoplasm. Finally it is lost.
In his succeeding paper (’86, 1) he studied the “nebenkern” spindle remains—in the spermatogonia and spermatocytes of pulmonates. In the spermatids, as in the former generations of cells, the nebenkern grows out of the nucleus, where it, with the chromatin, had formed the spireme. It appears as a loop, which becomes larger, twisted, and entangled, and finally breaks loose from the nucleus. Later it goes to form the spiral covering of the primary tail, changing it to the axial filament and true tail.
In his next paper (’86, 2) he describes the changes when the dividing of the protoplasm lags behind in the spermatocyte divisions. His description agrees in so many points with my own, that I shall quote his exact words:
“Die Spindelfasern hingegen contrahiren sich mehr und mehr nach dem Equator hin, wobei sie mit ein ander verschmelzen und merkwürdiger Weise je weiter dieser Verdichtungsprocess fort schreitet um so mehr an Tinctions fähigkeit speciell gegenüber dem Hämatoxylin gewinnen. Sie stellen jetzt zwei dreieckige oder hakenförmige Gebilde dar, die mit der Spitze noch im Equator zusammenhängen mit der breiten Seite sind sie den zugehörigen Zellen zugewendet. Hier sind ihre Grenzen undeutlicher, verwaschen und zeigen hier auch noch häufig eine streifige Beschaffenheit, welche auf ihren Ursprung hin weist. Zuweilen lassen sich einzelne Fäden noch eine beträchtliche Strecke weit in das Protoplasma hinein verfolgen, welches zwischen ihnen und den sich ausbildenden Zellkernen liegt.”
The “häkenformige Gebilde” moves away from the periphery, its sides elongate, break, and unite at the nuclear end, thus form the polygonal nebenkern. “Derselbe geht also in diesem Falle direckt aus den Spindelfasern hervor.” When the protoplasm does divide a similar process takes place. The spindle remains divide at an equatorial line and each half forms a nebenkern, and, as he says: “Also auch hier geht der Nebenkern direckt aus den Spindelfasern hervor. Vielleicht geht in den Spermatiden der Nebenkern überhaupt immer aus den Spindelfasern hervor, in dem die langfädige Verbindüngsbrücke, die ihn oft mit dem Kern verbindet, sich wohl als ein noch einige zeit persisttirender Rest der esteren deuten lässt.”
In his paper on Limax (’89, 1) he followed the nebenkern through all the divisions and thinks it a constant organ of the cell. In the second part of the paper on Helix and Paludina the nebenkern was considered as formed from the remains of the spindlepole and the [57] centrosome. Later Platner (’89, 2) found the nebenkern in the pancreas cells. In reports upon Pygæra and Sphinx he changed the name nebenkern to mitosome. The centrosome lies in front of the nucleus and forms the point of the head. This he calls the nebenkern. From the spindle remains arise two bodies. A large, fibrous one from the equatorial end has a clear space around it and the axial filament passes through it. It soon elongates and forms the tail. This is the large mitosome. The other is much smaller and arises from the polar end of the spindle fibers. It takes its place in the angle between the large mitosome and the nucleus. Here it persists till the nucleus begins to elongate, when it lengthens and surrounds the basal end of the spermatozoon tail. This is the small mitosome.
Platner (’85) saw a true nebenkern. I have already criticized the use of the word “nebenkern,” as the name for the spindle remains in the spermatocytes. Platner himself later denied the nuclear origin of the nebenkern. His results (’86, 2) agree with mine concerning the uniting, converging, staining and bending of the spindle-fiber remains and their passing to the tail. Again, Platner (’89, 1) probably followed the centrosome and attraction sphere, as well as the nebenkern, in the second part of the paper on Helix and Paludina. In the pancreas the body is a result of secretion, and is not a nebenkern. In Pygæra Platner’s large mitosome is the real nebenkern, as shown by its fibrous structure, its surrounding clear space and destiny. His small mitosome is what I have described as the acrosome, and he is mistaken as to its final use. The body he called centrosome in the nucleus is the persisting accessory chromosome. Such would be my interpretation of his figures.
Henking (’91) has followed the spermatid changes in Pyrrhocoris quite carefully. He finds that the fibers between the chromatin masses are separated into peripheral fibers and central spindle fibers. The first, a part of the second and the yolk mass, forms the nebenkern, which passes down over the axial filament. The rest of the central fibers form the mitosome. This takes its position at the angle between the nebenkern and the nucleus. On the surface of the nucleus it passes to the anterior end, then back to its original position. A piece now is constricted off and disappears, while the rest, increasing its affinity for stains, again wanders to the anterior pole of the nucleus, and becomes the acrosome.
The large amount of yolk substance is a disturbing element in Pyrrhocoris, but the nebenkern agrees with that of Gryllus in having the same origin and destiny, as does also the acrosome in fate and position. Besides, there is much similarity in the stages, as seen by comparing Henking’s fig. 63 with my fig. 25, and his fig. 85 with my fig. 32.
Wilcox (’95 and ’96) described the spermatid metamorphosis in Caloptenus. The interzonal fibers, a long, striated body composed very plainly of distinct fibers, contracts longitudinally, and the corners round themselves off, and this forms the nebenkern. It loses its fibrous structure, comes close to the nucleus, and then grows out into the axial filament. The centrosome moves half way around the nucleus and lies between the latter and the nebenkern. Later the centrosome becomes the middle piece.
[58] This description does not give much detail nor do the drawings show the stages clearly. As far as given, the formation of the nebenkern is the same as in Gryllus; but in Gryllus the axial filament is not formed from the nebenkern, as can be plainly seen from fig. 17.
Erlanger (’96), in a short paper, discusses the use of the term “nebenkern,” and suggests limiting it as is done in this paper. He opposes St. George’s opinion, that the nebenkern comes from the cytomicrosomes. In Blatta the cytomicrosomes are preserved during the whole process of mitosis, and have no connection with the spindle fibers, but during the telophase they collect in reduced numbers around the daughter nuclei.
In 1897 he called the collection of granules around the centrosome, the centrodeutoplasm. He considers them to be identical with St. George’s cytomicrosomes and the archoplasm (or attraction sphere) of other writers. In order to harmonize results, he suggests that, since the centrosome sometimes wanders around the nucleus, the centrodeutoplasm (or sphere) may unite with the spindle remains in some cases to form one body, as shown by the descriptions of Henking, Henneguy, Meves, and others. He later (’97, 2) describes the so-called “sphere,” and distinguishes between it and the true nebenkern.
I would strongly commend his excellent discussion of literature and his careful comparison of the results of investigators. He has shown clearly that the nebenkern comes from the spindle remains.
Calkins (’96) finds that the nebenkern comes from the spindle fibers and is useless in the cell. But Lumbricus is peculiar in having the nebenkern simply disintegrate, for, in many cases, he admits that the nebenkern has an important function. I do not have access to Henneguy’s or Bolles-Lee’s or Toyama’s works, yet I should judge from Erlanger’s and Meves’s criticism that all of these have the nebenkern originate from the spindle remains, and Henneguy describes it as having a “fibrillar appearance,” and Bolles-Lee as “fibrillar structure.” Accordingly, I think that each of these has discovered the correct origin of the structure, and I do not doubt that there is, at least in the first two, a more or less direct change from the spindle remains to the nebenkern.
Paulmier (’99) finds that in Anasa the nebenkern comes from the yolk mass and remains of the spindle fibers. A part of this mass separates off, while the whole is still in a confused condition, and forms the acrosome. The nebenkern forms the tail sheath, while the acrosome forms the point to the head.
[59] A comparison of Paulmier’s fig. 42 with my fig. 4 suggests the thought that they are the same stage, and his fibers are remains of the spindle. His fig. 43, of course, agrees with my fig. 25; and fig. 43 may correspond to figs. 26 or 14, only that his stain is weaker.
Meves’s investigations are the most extensive of the recent ones on the nebenkern. He has used Paludina and Pygæra as his objects. The consideration of the mitochondrion in the spermatocytes I shall pass over, as it is not within the bounds of this paper; yet I hope to study the earlier generations of my material, and shall then compare the results. In Meves’s description of Paludina, I find but one point in which it agrees with Gryllus. The head of the young spermatozoon in Paludina has a clear space in the center filled with nuclear fluid, which remains till the head begins to stretch. In Gryllus it is hollow until maturity.
In Paludina the nebenkern in one kind of spermatids is formed from threads made up of mitochondria—small, round bodies identical with St. George’s cytomicrosomes and Erlanger’s centrodeutoplasm. These threads change to vesicles, which, reduced to four, closely surround the centrosome as it lengthens into the middle piece. At first they form a four-cleft cylinder, but finally a single sheath. An idiozome and spindle remains are seen in the spermatid. They persist for awhile, and the former changes into the acrosome. In Pygæra Meves finds two kinds of spermatids, distinguished by a small difference in size. The larger forms the typical spermatozoon. The spindle remains form a “Spindelrest körper,” which is soon lost. The ends of the mitochondrion chains form a ring of dark mass surrounding a clear space. The ring is broken by radiating clear spaces. These spaces collect and unite into larger vacuoles, which surround the darker center. Finally there is only a dark ball with a surrounding clear space shut in by a ring. The centrosome with attached axial filament fastens itself to the nucleus; then both begin to grow longer. The axial filament passes over the surface of the ring surrounding the ball—the “mitochondrion körper” or nebenkern. This body elongates a great deal then the darker mass puts forth numerous threads which surround the axial filament.
As to Pygæra, the peculiarity is the complete agreement of his “mitochondrion körper” and my nebenkern in appearance and behavior for a part of the transformation and their complete disagreement in the other part. The question with me is, “Do the two bodies whose final stages are so similar originate so differently, or has one of us mistaken the origin of the body?”
Since reading Meves’s paper I have carefully reexamined my material, and I am positive that I am right as to the origin of the nebenkern; but, on the other hand, I would not say that Meves is wrong in his observations, as in doing so I should fall into the same error which [60] I think Meves himself has made. In his discussion of the literature he has forced every description to agree with his ideas, or has declared that the author has described some extra-nuclear organ as a nebenkern which is not a nebenkern. Thus by implication, if not by direct statement, he says that a nebenkern never comes from the spindle remains. With due respect to his ability and long experience as an investigator, I must say that Meves is mistaken in this. St. George himself traced the cytomicrosomes back to the spindle remains. Besides, the many investigators whom I have cited above cannot be mistaken as to the origin of the nebenkern. In my own material I am positive that there is a direct passing of the interzonal fibers over into the earlier stages of the nebenkern.
That the body which forms the tail covering does not come from the spindle remains in all cases, I am willing to admit. Meves has cited many investigators, especially on vertebrates, whose results favor such an opinion. The spindle remains do not change into a nebenkern even in all Arthropods, as Blackman (’01) finds no nebenkern, nor anything in anywise resembling it, in Scolopendra.
From my study of the results of other investigators, it is evident to me that there are at least two general methods for the formation of the covering for the spermatozoon tail. One of these plans will harmonize Meves’s mitochondrion körper, Erlanger’s centrodeutoplasm, Heidenhain’s pseudo-chromosomes, and other similar structures. The other will show that Platner’s large mitosome, Paulmier’s blackberry stage and my striated nebenkern are only different stages of the spindle remains changing into the tail covering.
1. The chromosomes of the second spermatocytes break up and the chromatin becomes diffused all through the nucleus. Later the chromatin collects in granules again and finally forms the walls of the tube-like spermatozoon head.
2. The spindle fibers break loose as the clear space is formed around the chromatin mass. They unite and contract, becoming fewer, thicker, and shorter. These bend and form the “striated nebenkern.” The fibers break up and sometimes show a blackberry appearance. Soon there is a collection of darker material at the center, surrounded by a clear [61] space, which is shut off from the cytoplasm by a darkly staining membrane which in sections appears as a ring. This stage persists for some time; then the nebenkern moves against the axial filament, elongates, loses the ring, and the dark mass passes down along the axial filament. Often it appears in several small drops.
3. The axial filament does not come from the nebenkern nor from the acrosome. It comes apparently from the nucleus; probably from the centrosome closely attached to the nucleus. It never passes through the nebenkern—only over its surface.
4. The acrosome occupies a position in the angle between the nebenkern and the nucleus. It shows a central clear space surrounded by a darker mass which stains more intensely on one side. Later it passes to the front of the nucleus and forms the point of the spermatozoon head.
5. The mature spermatozoon consists of a sharp point, a slender, tube-like head filled with a clear fluid, and a long, thread-like tail.
Laboratory of Zoology and Histology,
University of Kansas.
September 27, 1901.
1. Blackman, M. W., ’01: Spermatogenesis of the Myriapods. Kans. Univ. Quart., vol. 10.
2. Bütschli, O., ’71, 1: Vorläufige Mittheilung über Bau und Entwickelung, der Inseckten und Crustaceen. Zeitschr. f. wiss. Zool., Bd. 21.
3. —— ’71, 2: Nähere Mittheilung über die Entwickelung und den Bau der Samenfäden der Inseckten. Zeitschr. f. wiss. Zool., Bd. 21.
4. Calkins, G. N., ’95: The Spermatogenesis of Lumbricus. Journ. of Morph., vol. 11.
5. Erlanger, R., ’96, 1: Ueber den sogenannten Nebenkern in den männlichen Geschlechtszellen der Inseckten. Zool. Anz., Bd. 19.
6. —— ’96, 2: Die Entwickelung der männlichen Geschlechtszellen. Zool. Centralbl., Jahrg. 3.
7. —— ’97, 1: Ueber Spindelreste und den echten Nebenkern in den Hodenzellen. Zool. Centralbl., Jahrg. 4.
8. —— ’97, 2: Ueber die sogenannten Sphäre in den männlichen Geschlechtszellen. Zool. Centralbl., Jahrg. 4.
9. Heidenhain, M., ’00: Ueber die Centralkapseln und Pseudochromosomen in den Samenzellen von Proteus, sowie über ihr verhältniss zu den Idiozomen, Chondromiten und Archoplasmaschleifen. Anat. Anz., Bd. 18.
10. Henking, H., ’91: Ueber Spermatogenese und deren Beziehung zur Entwickelung bei Pyrrhocoris apterus. Zeitschr. f. wiss. Zool., Bd. 51.
11. McClung, C. E., ’99: A Peculiar Nuclear Element in the Male Reproductive Cells of Insects. Zool. Bull., vol. 2.
12. —— ’00: The Spermatocyte Divisions of the Acrididæ. Kans. Univ. Quart., vol. 9.
13. Meves, F., ’00: Ueber den von v. la Valette St. George endeckten Nebenkern (Mitochondrion Körper) der Samenzellen. Arch. f. mikr. Anat., Bd. 56.
14. Paulmier, F. C., ’99: The Spermatogenesis of Anasa tristis. Journ. of Morph., vol. 15, supplement.
15. Platner, G., ’85: Ueber die Spermatogenese bei den Pulmonaten. Arch. f. mikr. Anat., Bd. 25.
16. —— ’86, 1: Ueber die Enstehung des Nebenkerns und seine Beziehung zur Kerntheilung. Arch. f. mikr. Anat., Bd. 26.
17. —— ’86, 2: Zur Bildung der Geschlechtsproduckte bei den Pulmonaten. Arch. f. mikr. Anat., Bd. 26.
18. —— ’89, 1: Beiträge zur Kenntniss der Zelle und ihrer Theilungserscheinungen, I und II. Arch. f. mikr. Anat., Bd. 33.
19. —— ’89, 2: Beiträge zur Kenntniss der Zelle und ihrer Theilung, IV und V. Arch. f. mikr. Anat., Bd. 33.
20. Vom Rath, O., ’92: Zur Kenntniss der Spermatogenese von Gryllotalpa vulgaris. Arch. f. mikr. Anat., Bd. 42.
21. St. George, v. la Valette, ’67: Ueber die Genese der Samen Körper. Arch. f. mikr. Anat., Bd. 3.
22. —— ’86, 1: Spermatologische Beiträge, Zweite Mittheilung. Arch. f. mikr. Anat., Bd. 27.
23. —— ’86, 2: Spermatologische Beiträge, Vierte Mittheilung. Arch. f. mikr. Anat., Bd. 28.
24. —— ’87: Zelltheilung und Samenbildung bei Forficula auricularia. Festschrift f. A. v. Kölliker.
25. —— ’97: Zur Samen- und Eibildung beim Seidenspinner (Bombyx mori). Arch. f. mikr. Anat., Bd. 50.
[B] 26. Sutton, W. S., ’00: Spermatogonial Divisions in Brachystola magna. Kans. Univ. Quart., vol. 9.
27. Wilcox, E. V., ’95: Spermatogenesis of Caloptenus femur-rubrum and Cicada tibicen. Bull. of Mus. of Comp. Zool. Harvard., vol. 27.
28. —— ’96: Further Studies on the Spermatogenesis of Caloptenus femur-rubrum. Bull. of Mus. of Comp. Zool. Harvard., vol. 29.
[B] Nos. 12 and 26 were used when describing the material only and not the spermatid transformations.
All drawings were made by the author with the aid of a camera lucida. All figures except 2, 3, 4 and 45 were made with a B. & L. 1⁄16 objective and one inch eye-piece, producing a magnification of 1340 diameters. Figs. 2, 3, 4 and 45 were drawn with a Leitz 1⁄16 objective and a Zeiss compensating ocular No. 12, giving a magnification of 2325 diameters. The drawings were not reduced in the photo-mechanical reproduction.
Fig. 1. An early telophase, in which the fibers appear like barrel staves. Some of the fibers have thickened.
Fig. 2. A later stage, in which the fibers are uniting, the clear space is forming, and the chromosomes are beginning to separate.
Fig. 3. A daughter-cell, showing the chromosomes separating and the fibers thickening.
Fig. 4. The chromosomes are scattered over the nuclear membrane; the fibers appear as a few thick rods. The beginning of the acrosome is seen. The accessory chromosome can be distinguished.
Fig. 5. Same as fig. 4, with some of the fibers curved and no acrosome.
Fig. 6. Same as fig. 5, but with the acrosome visible.
Fig. 7. The chromosomes are granular; the fibers have bent and formed the round “striated nebenkern.” The acrosome is present.
Fig. 8a. Same as fig. 7, but the acrosome is not present.
Fig. 8b. The fibers run parallel with the long axis of the nebenkern.
Fig. 9. Here is shown a partial end view of the nebenkern. The accessory chromosome and the acrosome are both prominent.
Fig. 10. The nebenkern shows some of the fibers on the under side. It is beginning to break up.
Fig. 11. The nebenkern shows the beginning of the dark center; the fibers extend from it to the ring. The acrosome has a peculiar appearance.
Fig. 12. The fibers have broken up and the whole has assumed a vesicular appearance, resembling a blackberry. The acrosome is present.
Fig. 13. This shows a persisting spermatid stage. The chromatin appears in patches. The nebenkern is in the ball-and-ring stage. The acrosome shows its characteristic clear center surrounded by a ring darker on one side. The two dark bodies on the nuclear surface are probably artefacts.
Fig. 14. A stage a little later than 13. The dark central ball shows some vacuoles. The cell is beginning to elongate.
Fig. 15. Same as fig. 14, with the ball denser.
Fig. 16. The nucleus has become almost clear. The acrosome is behind the nebenkern.
Fig. 17. This shows the axial filament apparently growing out from the nucleus. There is an aggregation of chromatin at its base.
Fig. 18. The axial filament passes over the nebenkern, which has slightly elongated. The accessory chromosome is very plain.
Fig. 19. a shows a surface view, and b an optical section, of a nebenkern which is in process of breaking up.
Figs. 20 and 21 have part of the surface fibers cut off and show the dark mass forming within.
Fig. 22. Shows a nebenkern passing from the blackberry stage into the ring stage.
Fig. 23. This is an end view of the stage shown in figs. 20 and 21.
Fig. 24. The fainter lines are the fibers on the under side of the nebenkern.
Fig. 25. The blackberry stage, showing the mass of vesicles.
Fig. 26. This represents a nebenkern of the stage shown in fig. 7, drawn from a polar view, with adjusting focus.
Fig. 27. This shows the nebenkern elongating. The axial filament passes over its surface.
Fig. 28. A little later than fig. 27.
Fig. 29. Shows cross-sections of elongating nebenkern; a shows axial filament, nebenkern and acrosome; in b the axial filament is cut at an angle.
Fig. 30. A little later than fig. 28; the axial filament not so plainly shown.
Fig. 31. The ring has disappeared, and the dark mass is passing down along the filament. The acrosome is indistinct.
Fig. 32. The nebenkern has disappeared, but the acrosome has kept its position.
Fig. 33. The acrosome has flattened against the nucleus. Some remains of the nebenkern are seen.
Fig. 34. The acrosome has moved to the side of the nucleus. Some nebenkern remains are shown.
Fig. 35. The acrosome is in front of the nucleus.
Fig. 36. A little later stage than fig. 35.
Fig. 37. Shows a divided acrosome. It is not a frequent appearance.
Fig. 38. The nebenkern appears as a hanging drop on the side of the axial filament.
Fig. 39. The nucleus has condensed, the walls have thickened, and the chromatin appears in granules. The acrosome is pointed.
Fig. 40. A little later stage than 39.
Fig. 41. A little later stage than 40.
Fig. 42. The head is stained intensely, while the acrosome is lighter. Probably a middle piece indicated.
Fig. 43. The head shows the clear space.
Fig. 44. Cross-sections of the head, of the stage shown in fig. 43.
Fig. 45. The head and part of the tail of a mature spermatozoon.
Transcriber's Notes:
The cover image was created by the transcriber, and is in the public domain.
Typographical errors have been silently corrected but other variations in spelling and punctuation remain unaltered.
The quotations in German appear to have some errors, but these quotations have been reproduced exactly as in the original paper.
End of the Project Gutenberg EBook of The Kansas University Science
Bulletin, Vol. I, No. 2, Febr, by W. J. Baumgartner
*** END OF THIS PROJECT GUTENBERG EBOOK KANSAS UNIVERSITY SCIENCE BULLETIN, FEB 1902 ***
***** This file should be named 54868-h.htm or 54868-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/5/4/8/6/54868/
Produced by Larry B. Harrison, Paul Marshall and the Online
Distributed Proofreading Team at http://www.pgdp.net (This
book was produced from images made available by the
HathiTrust Digital Library.)
Updated editions will replace the previous one--the old editions will
be renamed.
Creating the works from print editions not protected by U.S. copyright
law means that no one owns a United States copyright in these works,
so the Foundation (and you!) can copy and distribute it in the United
States without permission and without paying copyright
royalties. Special rules, set forth in the General Terms of Use part
of this license, apply to copying and distributing Project
Gutenberg-tm electronic works to protect the PROJECT GUTENBERG-tm
concept and trademark. Project Gutenberg is a registered trademark,
and may not be used if you charge for the eBooks, unless you receive
specific permission. If you do not charge anything for copies of this
eBook, complying with the rules is very easy. You may use this eBook
for nearly any purpose such as creation of derivative works, reports,
performances and research. They may be modified and printed and given
away--you may do practically ANYTHING in the United States with eBooks
not protected by U.S. copyright law. Redistribution is subject to the
trademark license, especially commercial redistribution.
START: FULL LICENSE
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full
Project Gutenberg-tm License available with this file or online at
www.gutenberg.org/license.
Section 1. General Terms of Use and Redistributing Project
Gutenberg-tm electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or
destroy all copies of Project Gutenberg-tm electronic works in your
possession. If you paid a fee for obtaining a copy of or access to a
Project Gutenberg-tm electronic work and you do not agree to be bound
by the terms of this agreement, you may obtain a refund from the
person or entity to whom you paid the fee as set forth in paragraph
1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this
agreement and help preserve free future access to Project Gutenberg-tm
electronic works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the
Foundation" or PGLAF), owns a compilation copyright in the collection
of Project Gutenberg-tm electronic works. Nearly all the individual
works in the collection are in the public domain in the United
States. If an individual work is unprotected by copyright law in the
United States and you are located in the United States, we do not
claim a right to prevent you from copying, distributing, performing,
displaying or creating derivative works based on the work as long as
all references to Project Gutenberg are removed. Of course, we hope
that you will support the Project Gutenberg-tm mission of promoting
free access to electronic works by freely sharing Project Gutenberg-tm
works in compliance with the terms of this agreement for keeping the
Project Gutenberg-tm name associated with the work. You can easily
comply with the terms of this agreement by keeping this work in the
same format with its attached full Project Gutenberg-tm License when
you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are
in a constant state of change. If you are outside the United States,
check the laws of your country in addition to the terms of this
agreement before downloading, copying, displaying, performing,
distributing or creating derivative works based on this work or any
other Project Gutenberg-tm work. The Foundation makes no
representations concerning the copyright status of any work in any
country outside the United States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other
immediate access to, the full Project Gutenberg-tm License must appear
prominently whenever any copy of a Project Gutenberg-tm work (any work
on which the phrase "Project Gutenberg" appears, or with which the
phrase "Project Gutenberg" is associated) is accessed, displayed,
performed, viewed, copied or distributed:
This eBook is for the use of anyone anywhere in the United States and
most other parts of the world at no cost and with almost no
restrictions whatsoever. You may copy it, give it away or re-use it
under the terms of the Project Gutenberg License included with this
eBook or online at www.gutenberg.org. If you are not located in the
United States, you'll have to check the laws of the country where you
are located before using this ebook.
1.E.2. If an individual Project Gutenberg-tm electronic work is
derived from texts not protected by U.S. copyright law (does not
contain a notice indicating that it is posted with permission of the
copyright holder), the work can be copied and distributed to anyone in
the United States without paying any fees or charges. If you are
redistributing or providing access to a work with the phrase "Project
Gutenberg" associated with or appearing on the work, you must comply
either with the requirements of paragraphs 1.E.1 through 1.E.7 or
obtain permission for the use of the work and the Project Gutenberg-tm
trademark as set forth in paragraphs 1.E.8 or 1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any
additional terms imposed by the copyright holder. Additional terms
will be linked to the Project Gutenberg-tm License for all works
posted with the permission of the copyright holder found at the
beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including
any word processing or hypertext form. However, if you provide access
to or distribute copies of a Project Gutenberg-tm work in a format
other than "Plain Vanilla ASCII" or other format used in the official
version posted on the official Project Gutenberg-tm web site
(www.gutenberg.org), you must, at no additional cost, fee or expense
to the user, provide a copy, a means of exporting a copy, or a means
of obtaining a copy upon request, of the work in its original "Plain
Vanilla ASCII" or other form. Any alternate format must include the
full Project Gutenberg-tm License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works
provided that
* You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is owed
to the owner of the Project Gutenberg-tm trademark, but he has
agreed to donate royalties under this paragraph to the Project
Gutenberg Literary Archive Foundation. Royalty payments must be paid
within 60 days following each date on which you prepare (or are
legally required to prepare) your periodic tax returns. Royalty
payments should be clearly marked as such and sent to the Project
Gutenberg Literary Archive Foundation at the address specified in
Section 4, "Information about donations to the Project Gutenberg
Literary Archive Foundation."
* You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or destroy all
copies of the works possessed in a physical medium and discontinue
all use of and all access to other copies of Project Gutenberg-tm
works.
* You provide, in accordance with paragraph 1.F.3, a full refund of
any money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days of
receipt of the work.
* You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project
Gutenberg-tm electronic work or group of works on different terms than
are set forth in this agreement, you must obtain permission in writing
from both the Project Gutenberg Literary Archive Foundation and The
Project Gutenberg Trademark LLC, the owner of the Project Gutenberg-tm
trademark. Contact the Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
works not protected by U.S. copyright law in creating the Project
Gutenberg-tm collection. Despite these efforts, Project Gutenberg-tm
electronic works, and the medium on which they may be stored, may
contain "Defects," such as, but not limited to, incomplete, inaccurate
or corrupt data, transcription errors, a copyright or other
intellectual property infringement, a defective or damaged disk or
other medium, a computer virus, or computer codes that damage or
cannot be read by your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium
with your written explanation. The person or entity that provided you
with the defective work may elect to provide a replacement copy in
lieu of a refund. If you received the work electronically, the person
or entity providing it to you may choose to give you a second
opportunity to receive the work electronically in lieu of a refund. If
the second copy is also defective, you may demand a refund in writing
without further opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS', WITH NO
OTHER WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT
LIMITED TO WARRANTIES OF MERCHANTABILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of
damages. If any disclaimer or limitation set forth in this agreement
violates the law of the state applicable to this agreement, the
agreement shall be interpreted to make the maximum disclaimer or
limitation permitted by the applicable state law. The invalidity or
unenforceability of any provision of this agreement shall not void the
remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in
accordance with this agreement, and any volunteers associated with the
production, promotion and distribution of Project Gutenberg-tm
electronic works, harmless from all liability, costs and expenses,
including legal fees, that arise directly or indirectly from any of
the following which you do or cause to occur: (a) distribution of this
or any Project Gutenberg-tm work, (b) alteration, modification, or
additions or deletions to any Project Gutenberg-tm work, and (c) any
Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of
computers including obsolete, old, middle-aged and new computers. It
exists because of the efforts of hundreds of volunteers and donations
from people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future
generations. To learn more about the Project Gutenberg Literary
Archive Foundation and how your efforts and donations can help, see
Sections 3 and 4 and the Foundation information page at
www.gutenberg.org
Section 3. Information about the Project Gutenberg Literary Archive Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg Literary
Archive Foundation are tax deductible to the full extent permitted by
U.S. federal laws and your state's laws.
The Foundation's principal office is in Fairbanks, Alaska, with the
mailing address: PO Box 750175, Fairbanks, AK 99775, but its
volunteers and employees are scattered throughout numerous
locations. Its business office is located at 809 North 1500 West, Salt
Lake City, UT 84116, (801) 596-1887. Email contact links and up to
date contact information can be found at the Foundation's web site and
official page at www.gutenberg.org/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
[email protected]
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To SEND
DONATIONS or determine the status of compliance for any particular
state visit www.gutenberg.org/donate
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations. To
donate, please visit: www.gutenberg.org/donate
Section 5. General Information About Project Gutenberg-tm electronic works.
Professor Michael S. Hart was the originator of the Project
Gutenberg-tm concept of a library of electronic works that could be
freely shared with anyone. For forty years, he produced and
distributed Project Gutenberg-tm eBooks with only a loose network of
volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as not protected by copyright in
the U.S. unless a copyright notice is included. Thus, we do not
necessarily keep eBooks in compliance with any particular paper
edition.
Most people start at our Web site which has the main PG search
facility: www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.