The Project Gutenberg EBook of Studies of Birds Killed in Nocturnal
Migration, by Harrison B. Tordoff and Robert M. Mengel
This eBook is for the use of anyone anywhere in the United States and most
other parts of the world at no cost and with almost no restrictions
whatsoever. You may copy it, give it away or re-use it under the terms of
the Project Gutenberg License included with this eBook or online at
www.gutenberg.org. If you are not located in the United States, you'll have
to check the laws of the country where you are located before using this ebook.
Title: Studies of Birds Killed in Nocturnal Migration
Author: Harrison B. Tordoff
Robert M. Mengel
Release Date: June 20, 2016 [EBook #52382]
Language: English
Character set encoding: ISO-8859-1
*** START OF THIS PROJECT GUTENBERG EBOOK STUDIES OF BIRDS ***
Produced by Judith Wirawan, Chris Curnow, Joseph Cooper
and the Online Distributed Proofreading Team at
http://www.pgdp.net
University of Kansas Publications
Museum of Natural History
Volume 10, No. 1, pp. 1-44, 6 figures in text, 2 tables
———September 12, 1956———
BY
HARRISON B. TORDOFF AND ROBERT M. MENGEL
University of Kansas
Lawrence
1956
University of Kansas Publications, Museum of Natural History
Editors: E. Raymond Hall, Chairman, A. Byron Leonard, Robert W. Wilson
Volume 10, No. 1, pp. 1-44, 6 figures in text, 2 tables
Published September 12, 1956
University of Kansas
Lawrence, Kansas
PRINTED BY
FERD VOILAND, JR., STATE PRINTER
TOPEKA, KANSAS
1956
26-3856
Studies of Birds
Killed in Nocturnal Migration
BY
HARRISON B. TORDOFF AND ROBERT M. MENGEL
| PAGE | |
| Introduction | 4 |
| Accidents to Migrating Birds in early October, 1954 | 6 |
| General | 6 |
| Accidents at Topeka, Kansas | 6 |
| Description of WIBW-TV tower | 7 |
| Weather conditions | 7 |
| Acknowledgments | 7 |
| Notes on the Species Killed at Topeka | 8 |
| Randomness of the Sample | 17 |
| Number of Migrants | 18 |
| Differential Migration of Sex- and Age-classes | 20 |
| History of the subject | 20 |
| Differential migration of sex- and age-classes as shown by the Topeka sample | 23 |
| Molt in Relation to Migration | 29 |
| General comment | 29 |
| Molt in the Topeka sample | 30 |
| Size Differences according to Sex and Age | 31 |
| Linear measurements | 31 |
| Weights | 32 |
| Computations of Longevity and Survival | 38 |
| Processing of Samples | 38 |
| Summary | 39 |
| Literature Cited | 41 |
This paper is primarily an analysis of a sample of migrant birds killed in the autumn of 1954 by striking a television tower one mile west of Topeka, Shawnee County, Kansas. Secondarily, some aspects of migration involved in studies of this kind are discussed and historical background is presented.
Considerable interest has been occasioned in recent years in the eastern United States by large-scale accidents to night-migrating birds. Most accidents have occurred in the autumn. The widespread adoption by airports of an instrument called the ceilometer, which measures the height of cloud ceilings by reflecting from them a high-powered beam of light, has proved under certain conditions to be catastrophic to night-flying birds. Among the recent reports of such accidents are those of Spofford (1949) and Laskey (1951) for Nashville, Tennessee, Howell and Tanner (1951) for Knoxville, Tennessee, and Lovell (1952) for Louisville, Kentucky. Recently Howell, Laskey, and Tanner (1954) reviewed ceilometer "tragedies" without being able to determine the exact reason for their lethal effectiveness. Less publicized so far have been mass collisions of birds with another class of obstacles, tall radio and television towers. These slender towers, usually 500 to 1000 feet tall, are increasing rapidly in numbers and there is reason to suppose that they will take a correspondingly larger toll of bird life.
Notice has long been given by ornithologists to mass destruction of birds by more conventional solid obstructions to passage, and newspapers occasionally mention birds killed at such well-known points as the Washington Monument and the Empire State Building.
Seventy-five years ago, J. A. Allen (1880) published the results of questionnaires circulated by William Brewster to lighthouse keepers. Brewster himself (1886) described destruction of birds at a lighthouse in the Bay of Fundy, paying keen attention to behavior of the birds and the exact conditions under which nocturnal flight and accidents occurred. The subject also received attention in several countries across the Atlantic. Destruction of birds at Irish lighthouses was carefully noted over a period of years and the results were published periodically, culminating in R. M. Barrington's massive report (1900) which remains in some ways the most thorough of its type.
While conservation-minded individuals have been concerned with the tremendous mortality involved in these various events, the ill wind blows some good in that, properly used, the data provided by such accidents can shed light on many obscure aspects of bird migration. Each accidental kill of birds affords a cross-section, approaching in variable degree a random sample, of the migrants passing a given point on a given date. The types of information provided by such kills are numerous, for example: (1) information on the presence of various species and the dates of their occurrence; (2) information on the relative abundance of species; (3) quantitative data on the relative sizes of males and females, and immatures and adults (of importance to taxonomic ornithology); (4) information on the relative times of migration of males, females, adults, and young; (5) information on molts and plumages; (6) quantitative information on composition by subspecies of migrants of the same species; (7) physiological data (fat condition, etc.) pertinent to the study of migration; and probably others.
In spite of the great potential of this kind of material, the majority of ornithologists with access to such data have contented themselves with listing the species and sometimes the numbers of birds killed. A few have gone further. James T. Tanner (unpublished) attempted to compute the longevity of the Ovenbird (Seiurus aurocapillus) by analysis of ceilometer-killed birds at Knoxville, Tennessee (see below). Mention should be made of the reports of Rintoul and Baxter (1914) supplemented by Ticehurst (1916) who used rather small numbers of birds killed at Scottish lighthouses in studies of molt. However, the only effort to utilize the results of accidental kills on a large scale over a period of years appears to have been that, already mentioned, of Barrington (1900) and his co-workers in Ireland. An idea of the potentialities of the large recent kills in the United States may be obtained when it is recalled that in the 18 years of Barrington's work, which embodied some 1000 reports from lighthouse keepers, Barrington obtained for study only about 2000 specimens, many of these consisting of wings and feet only (Barrington's paper not seen in original; see J. A. Allen, 1901:205). More recently Dobben and Bruyns (1939) have analyzed the age and sex classes of some birds killed at lighthouses in Holland.
As far as we have learned, there is no previous thorough analysis in the literature of large, accidentally-killed samples of birds. On the following pages we emphasize some of the uses which can be made of such material. We think that intensive analyses of such[Pg 6] events, whenever they occur, should become a regular part of ornithological investigation and that integration of numerous studies of such incidents will provide an unprecedented mass of information on migration.
General.—The few days around the end of the first week of October, 1954, were notable for a series of accidents which occurred to migrating birds over much of eastern United States. So far as we know, these were all associated with an extensive belt of bad weather (cold fronts and stationary fronts) which covered much of the country during that period, and the accidents involved ceilometers and solid structures alike. Accidents known to us occurred as far south as Macon, Georgia (David W. Johnston, letter: Nov. 1, 1954), as far north as New York City, where many migrants were killed at the Empire State Building (New York Times, Thursday, October 7, 1954, p. 1) and elsewhere, and as far west as Smoky Hill Air Force Base at Salina, Kansas (ceilometer, October 7, some birds received at the University of Kansas). Some of the above, and incidents from a number of other localities, were mentioned in varying detail in Audubon Field Notes (vol. 9, no. 1, pp. 6, 10, 15, 17, 18, 32, February, 1955). Still other accidents occurred at Columbia, Missouri (Richard P. Grossenheider, verbal communication), and Topeka, Kansas (present paper). Some probably have escaped our notice; summaries of some of these will probably appear in ornithological journals for some time to come. At Robins Air Force Base near Macon, Georgia, at least 50,000 birds were killed, of which about 2500, representing 54 species, were picked up (Johnston, loc. cit.).
Accidents at Topeka, Kansas.—At Topeka, Shawnee County, Kansas, all birds were killed by collision with the newly-erected (1954) television transmitting tower of station WIBW-TV. This tower is one mile west of the city.
The first casualties (see Table 1 for all others) were a Sora (Porzana carolina) and a Yellow-bellied Flycatcher (Empidonax flaviventris) found on September 7. The major accidents, however, occurred on the nights of September 24-25, September 30-October 1, October 5-6, and October 6-7. Totals of birds picked up (probably over 95 per cent of birds killed) are given in Table 1, in which each date given is that of the day after the kill, i. e., the date on which the birds were collected.
All major kills occurred on cloudy and foggy nights associated with frontal weather. Throughout the period a few birds struck the tower even on fairly clear nights, and minor but appreciable "falls" occurred on the nights of October 4-5, 7-8, and 22-23. A few birds killed probably were overlooked for a time and found their way into later samples. This is especially probable in the case of some birds entered under date of October 23, as many of these were somewhat desiccated. Weights clearly altered by desiccation or mutilation were not recorded. Reports of these accidents have been published by Carson (1954 a, b, and c).
According to Carson (1954c:27), the majority of birds killed on nights of heavy flight fell "between three and four o'clock in the morning when skies were overcast and a cool front moved in from the north. Due to the cooperation of the watchmen it is thought that most of the birds that were killed were recovered. Of course some injured birds in hiding were not found and some were lost to predators."
Description of WIBW-TV TOWER.—The tower is 950 feet tall and stands on a hill approximately 1000 feet above sea level. The fact that the tower is on a hill places the top of the tower at 1010 feet above the elevation of the average local terrain. The tower is triangular in cross-section, each face seven feet wide, and is constructed of six-inch steel L-beams with three-inch cross-members every seven feet and smaller diagonal cross-members. It has no taper and bears a transmitting antenna on the top. The tower is supported by 12 guy wires, 3 wires attaching at each of 4 levels. The cables extend south, WNW, and NNE from the tower and are 1-1/2 inches in diameter. The tower is lighted by a series of red lights, some flashing and others steady. The transmitter was not in operation when the accidents took place.
Weather conditions.—All major kills at Topeka occurred when migrating birds encountered either a cold front or a stationary front lying over eastern Kansas. Typically, this frontal weather included rain, fog, and cloud ceilings down to as low as 800 to 1000 feet. Weather of this type presumably forces the migrating birds to fly below the cloud ceiling and thus brings them within the altitudinal range of the television towers.
We gratefully acknowledge our debt to the Topeka Audubon Society for making this study possible by carefully collecting birds killed at the television tower. L. B. Carson deserves special mention for his general supervision of [Pg 8]the bird collecting by the members of the Topeka Audubon Society. Members of the Society and others who picked up birds under the television tower were: Mrs. Lloyd Biggs, Elaine Carson, L. B. Carson, Jesse A. Eddy, Elizabeth Fisher, Mrs. Walter Huxman, Florence McKinney, Mrs. Charles Martin, Mrs. Fred P. Martin, T. W. Nelson, Fred Prebble, Grace Prebble, Orville Rice, Mrs. G. Warren Scholl, E. W. Senne, and Beatrice Swenson.
We received equally important assistance from students and staff of the University of Kansas in recording of data and preparation of specimens. The following helped in these ways: Rollin H. Baker, R. W. Dickerman, David L. Hardy, J. W. Hardy, Jane S. Mengel, Larry D. Mosby, Richard Van Gelder, South G. Van Hoose, and Glen E. Woolfenden. We are indebted to the Interlibrary Loan Service of the University of Kansas Library for help in securing certain reference works. Robert Sokal of the University of Kansas gave helpful advice concerning statistical procedures.
A list of numbers and kinds of birds killed is given in Table 1. Discussion of data afforded by certain species for which, large samples were available will be found below. There are additionally certain data afforded by the sample and certain comments to be made on various species which can be handled most conveniently in an annotated list. In this list we have included all weight data (still scarce for many North American birds), comments on status in Kansas of various species, results of comparisons to determine subspecies, and miscellaneous observations. Weights of birds are given in grams and were taken on a triple-beam balance. Fat condition is given in the scale proposed by McCabe (1943:556). Weight data from birds migrating at night should be especially useful because these migrants all have relatively empty crops and stomachs, thus reducing variability. Not all birds were suitable for weighing and measuring, for a variety of reasons. This accounts for discrepancies in totals between Table 1 and the annotated list.
All passerine species were aged by noting the degree of ossification of the skull. In no case, of the more than a thousand passerines aged by examination of the skull, did we find difficulty in determining whether an individual was a bird of the year or an adult. We found no specimens in which ossification of the skull was nearing completion. In the several species in our sample with distinctive first-winter plumages, we found complete agreement in age as shown by plumage and by condition of the skull. We think this is further proof, if such is needed, that this method of aging is thoroughly reliable in early autumn for the passerine species included in our sample and for others with similar breeding seasons.
Table 1.—Birds Killed at a Television Tower at Topeka, Kansas, in 1954
See annotated list for division into sex- and age-classes. Where discrepancies exist between totals given here and totals given in the annotated list, these result from the fact that some specimens could not be sexed and aged.
| Sept. 25 | Oct. 1 | Oct. 3 | Oct. 4 | Oct. 5 | Oct. 6 | Oct. 7 | Oct. 8 | Oct. 9 | Oct. 10 | Oct. 23 | Totals | |
| Pied-billed Grebe | 1 | 1 | 1 | 3 | ||||||||
| Green Heron | 1 | 1 | ||||||||||
| Blue-winged Teal | 1 | 1 | 6 | 8 | ||||||||
| Virginia Rail | 3 | 1 | 4 | |||||||||
| Sora | 1 | 6 | 1 | 1 | 1 | 10 | ||||||
| American Coot | 3 | 3 | ||||||||||
| Mourning Dove | 8 | 1 | 1 | 10 | ||||||||
| Yellow-billed Cuckoo | 1 | 1 | ||||||||||
| Black-billed Cuckoo | 1 | 1 | ||||||||||
| Yellow-shafted Flicker | 3 | 3 | ||||||||||
| Yellow-bellied Flycatcher | 1 | 1 | ||||||||||
| House Wren | 2 | 3 | 1 | 2 | 1 | 1 | 10 | |||||
| Long-billed Marsh Wren | 1 | 1 | 1 | 3 | ||||||||
| Short-billed Marsh Wren | 1 | 2 | 1 | 4 | ||||||||
| Catbird | 1 | 28 | 1 | 1 | 6 | 6 | 43 | |||||
| Brown Thrasher | 1 | 1 | 1 | 3 | ||||||||
| Wood Thrush | 3 | 3 | ||||||||||
| Hermit Thrush | 1 | 1 | ||||||||||
| Olive-backed Thrush | 14 | 1 | 1 | 16 | ||||||||
| Golden-crowned Kinglet | 1 | 5 | 6 | |||||||||
| Ruby-crowned Kinglet | 2 | 1 | 8 | 1 | 1 | 1 | 14 | |||||
| Yellow-throated Vireo | 1 | 1 | ||||||||||
| Blue-headed Vireo | 1 | 19 | 1 | 2 | 5 | 8 | 3 | 1 | 40 | |||
| Red-eyed Vireo | 18 | 36 | 2 | 13 | 2 | 3 | 1 | 75 | ||||
| Philadelphia Vireo | 3 | 9 | 12 | |||||||||
| Warbling Vireo | 8 | 19 | 1 | 4 | 1 | 1 | 34 | |||||
| Black and White Warbler | 1 | 1 | 3 | 5 | ||||||||
| Tennessee Warbler | 1 | 1 | 2 | 1 | 5 | |||||||
| Orange-crowned Warbler | 7 | 14 | 1 | 4 | 19 | 5 | 1 | 1 | 52 | |||
| Nashville Warbler | 7 | 94 | 4 | 3 | 39 | 27 | 5 | 1 | 1 | 181 | ||
| Parula Warbler | 1 | 1 | 2 | |||||||||
| Yellow Warbler | 3 | 3 | 1 | 1 | 8 | |||||||
| Magnolia Warbler | 1 | 2 | 3 | |||||||||
| Black-throated Blue Warbler | 2 | 1 | 3 | |||||||||
| Myrtle Warbler | 1 | 1 | ||||||||||
| Black-throated Green Warbler | 1 | 1 | ||||||||||
| Chestnut-sided Warbler | 1 | 1 | 2 | |||||||||
| Bay-breasted Warbler | 1 | 2 | 3 | |||||||||
| Palm Warbler | 3 | 1 | 4 | |||||||||
| Oven-bird | 4 | 21 | 2 | 3 | 1 | 1 | 32 | |||||
| Northern Water-thrush | 5 | 1 | 6 | |||||||||
| Mourning Warbler | 15 | 64 | 2 | 11 | 2 | 1 | 95 | |||||
| Yellow-throat | 10 | 115 | 2 | 4 | 25 | 18 | 1 | 1 | 176 | |||
| Yellow-breasted Chat | 1 | 1 | ||||||||||
| Wilson Warbler | 1 | 2 | 3 | |||||||||
| Canada Warbler | 2 | 2 | ||||||||||
| American Redstart | 1 | 1 | ||||||||||
| Bobolink | 4 | 4 | ||||||||||
| Rose-breasted Grosbeak | 2 | 2 | ||||||||||
| Indigo Bunting | 1 | 2 | 3 | 1 | 7 | |||||||
| Dickcissel | 31 | 1 | 3 | 1 | 36 | |||||||
| Savannah Sparrow | 1 | 6 | 1 | 1 | 5 | 1 | 1 | 16 | ||||
| Grasshopper Sparrow | 7 | 2 | 3 | 3 | 1 | 1 | 1 | 18 | ||||
| Leconte Sparrow | 3 | 3 | ||||||||||
| Sharp-tailed Sparrow | 1 | 1 | 1 | 3 | ||||||||
| Slate-colored Junco | 1 | 1 | ||||||||||
| Clay-colored Sparrow | 11 | 1 | 2 | 1 | 15 | |||||||
| Fox Sparrow | 1 | 1 | ||||||||||
| Lincoln Sparrow | 41 | 7 | 5 | 22 | 3 | 1 | 3 | 82 | ||||
| Swamp Sparrow | 1 | 1 | 2 | 4 | ||||||||
| Song Sparrow | 2 | 2 | ||||||||||
| Total—species | 22 | 41 | 6 | 3 | 13 | 31 | 29 | 16 | 10 | 8 | 15 | 61 |
| Total—individuals | 94 | 585 | 16 | 3 | 26 | 146 | 147 | 31 | 10 | 8 | 24 | 1090 |
[Pg 10]The annotated list may be consulted for further data in connection with the species listed in Table 1. As is indicated below, we regard the figures of this sample as unreliable to an unknown degree in comparing the relative abundance of one species with another. Accumulation of such data from various localities, however, should prove useful in another type of comparison. Samples of the same species killed in the same way at about the same time at different localities should be directly comparable. Eventually, this should provide us with a means of determining relative abundance of a species in different parts of its migratory route.
Approximately 200 of the most interesting specimens were preserved as study skins and are in the University of Kansas Museum of Natural History. An effort was made to preserve at least one of each species, and we fell only a few short of this goal. All of the forms rare in Kansas are represented by skins. We could see no reason to list the preserved specimens in detail here. Species of which no study skins were made, however, are so marked.
So far as we can tell, no truly western subspecies (from west of the Great Plains) occurred in the Topeka sample. Probably most or all of the birds came from areas more or less directly north of eastern Kansas.
In critical areas where different subspecies of the same species occur together in migration, data from samples of this kind should prove enlightening. In future analyses, conducted in such areas, it might be possible to preserve all specimens of some of the variable species, or at least to measure all individuals of species in which[Pg 11] size is the most important variable character. Quantitative study could then be made of the different geographic variants occurring, their proportions in the migrant population determined, and their origins deduced. In studying populations of Painted Buntings (Passerina ciris) wintering in Mexico, Storer (1951) has provided an interesting demonstration of methods which can be applied to such samples.
A few bats killed at the tower provided a surprise. They will be discussed separately by Richard Van Gelder.
Podilymbus p. podiceps. Pied-billed Grebe.—Weights: male, 394.8 (all weights in grams); females, 332.5, 289.7; all fat.
Butorides v. virescens. Green Heron.—Weight: 1 (unsexed), 168.6.
Anas discors. Blue-winged Teal.—Weights: 4 males, mean 421.2 (391.3-458.1); 3 females, 367.7, 371.6, 393.2; all fat.
Rallus limicola. Virginia Rail.—Weights: 3 males, 73.7, 83.2, 90.5; 1 female, 67.3; moderately fat to fat.
Porzana carolina. Sora.—Weights: 4 males, mean 76.8 (68.7-89.9); 3 females, 62.6, 63.2, 63.5; moderately fat to very fat.
Fulica americana. American Coot.—Weights: 2 females, 385.3, 530.0, both fat. None preserved.
Zenaidura macroura marginella. Mourning Dove.—Weights: 2 adult males, 121.8, 140.2; 3 immature males, 113.1, 126.1, 130.0; 3 adult females, 122.5, 126.9, 136.0; 2 immature females, 129.4, 132.7; moderately fat to very fat. The presence of Mourning Doves in the sample is interesting as these birds are not generally regarded as night migrants. Conceivably the specimens were local birds going to roost. None preserved.
Colaptes auratus luteus. Yellow-shafted Flicker.—Weights: 2 males, 126.0, 139.4, little fat. Flickers have several times been recorded as night migrants.
Empidonax flaviventris. Yellow-bellied Flycatcher.—Weight: 1 immature male, 11.9, moderately fat. This is a rare species in Kansas, the present being the ninth preserved specimen for the State.
Troglodytes aëdon parkmanii. House Wren.—Weights: 4 adult males, mean 10.5 (9.8-10.9), 2 immature males, 9.0, 11.3; 1 adult female, 9.9, 1 immature female, 7.0; no fat (im. ♀) to fat.
Telmatodytes palustris dissaëptus. Long-billed Marsh Wren.—Weights: 1 adult male, 10.8; 1 adult female, 9.2; both moderately fat. The specimens are moderately bright and rufescent above, being typical of the populations of the central plains.
Cistothorus platensis stellaris. Short-billed Marsh Wren.—Weights: 1 immature male, 8.2; 1 adult female, 8.1; immature female, 8.2; all fat.
Dumetella carolinensis. Catbird.—Weights: 6 adult males, mean 37.5 (34.1-42.5), little fat to very fat; 14 immature males, mean 37.57 ± .94 (standard error), S. D. (standard deviation) 3.37, little fat to fat; 11 adult females, mean 39.09 ± .94, S. D. 2.97, little fat to fat; 12 immature females, mean 38.42 ± .83, S. D. 2.74, moderately fat to fat.
Toxostoma r. rufum. Brown Thrasher.—Weight: 1 immature male, 60.2, little fat.
Hylocichla mustelina. Wood Thrush.—Weights: 1 adult male, 54.2, moderately fat; 2 adult females, 44.6, 45.7, little fat and fat, respectively.
Hylocichla ustulata swainsonii. Olive-backed Thrush.—Weights: 6 immature males, mean 31.0 (28.1-33.2), little fat to fat; 6 adult females, mean 29.6 (27.1-35.0), moderately fat to fat; 3 immature females, 27.1, 33.8, 35.8, little fat to fat. The absence of adult males in our sample of 15 birds is noteworthy but inexplicable with our few data.
Regulus s. satrapa. Golden-crowned Kinglet.—Weights: 1 adult male, 6.7, moderately fat; 2 immature males, 6.5, 7.4, moderately fat and fat; 2 adult females, 7.3, 7.4, moderately fat and fat; 1 immature female, 7.2, moderately fat.
Regulus c. calendula. Ruby-crowned Kinglet.—Weights: 3 adult males, 6.2, 7.6, 8.2, little fat to fat; 1 immature male, 6.6, fat; 4 adult females, mean 6.1 (5.6-6.7), moderately fat to fat; 3 immature females, 5.8, 6.6, 7.0, moderately fat to fat.
Vireo flavifrons. Yellow-throated Vireo.—Weight: 1 immature male, 21.5, very fat.
Vireo s. solitarius. Blue-headed Vireo.—Weights: 9 adult males, mean 17.7 (16.6-19.5), little fat to very fat; 17 immature males, mean 17.53 ± .46, S. D. 1.83, no fat (13.8) to very fat (21.3); 7 adult females, mean 17.6 (15.0-21.6), moderately fat to very fat; 6 immature females, mean 17.0 (14.5-18.9), moderately fat to fat. Surprisingly numerous in the sample.
Vireo olivaceus. Red-eyed Vireo.—Weights: 1 adult male, 16.1, moderately fat; 38 immature males, mean 21.21 ± .43, S. D. 2.60, little fat (1 specimen) to excessively fat, mostly moderately fat or fat; 2 adult females, 18.1, 18.1, both fat; 23 immature females, mean 19.28 ± .46, S. D. 2.16, little (2 specimens) to very fat, mostly fat.
Wing length: 1 adult male, 79.1; 38 immature males, mean 78.05 ± .30, S. D. 1.80; 2 adult females, 76.3, 79.0, 23 immature females, mean 75.83 ± .42, S. D. 1.99.
As mentioned below, the presence of only 3 adults in the sample of 64 Red-eyed Vireos is highly significant and their occurrence only in the earlier samples is strong evidence of early migration by the adults.
Vireo philadelphicus. Philadelphia Vireo.—Weights: 2 adult males, 12.1, 15.9, moderately fat and very fat; 2 immature males, 11.1, 13.2, fat and very fat; 2 adult females, 13.1, 14.2, both fat; 5 immature females, mean 14.1 (12.0-15.2), moderately fat to very fat.
This species previously has been collected in Kansas only twice. Both records are from Doniphan County in September, 1922. Field observers occasionally record the Philadelphia Vireo in eastern Kansas. Long (1940:450) calls it a "very rare migrant in the extreme east." Our sample of 12 birds killed on two nights (and probably after the peak of migration of this species) leads us to think that this vireo is actually a regular, but overlooked, migrant in fair numbers.
Vireo g. gilvus. Warbling Vireo.—Weights: 12 adult males, mean 15.92 ± .43, S. D. 1.44, moderately fat to very fat; 8 immature males, mean 16.64 (14.2-17.8), fat to very fat; 5 adult females, mean 16.1 (13.7-18.0), fat to very fat; 5 immature females, mean 15.4 (14.1-17.8), little fat to fat.
Wing length: 12 adult males, mean 73.08 ± .49, S. D. 1.64; 8 immature [Pg 13]males, mean 71.15 (69.9-72.8); 5 adult females, mean 70.0 (69.2-71.0); 5 immature females, mean 68.4 (67.7-70.3).
Tail length: 12 adult males, mean 53.33 ± .53, S. D. 1.77; 8 immature males, mean 50.03 (47.1-51.3); 4 adult females, mean 48.6 (47.7-49.8); 5 immature females, mean 49.2 (47.3-53.0).
There is no indication that western birds (V. g. swainsonii) make up any part of this sample.
The sample of 34 Warbling Vireos is too small to show the significance, if any, of the 2:1 ratio of males to females in the sample. Adequate samples of this species, taken at intervals, would add interesting information on time of migration of the four sex- and age-classes.
Mniotilta varia. Black and White Warbler.—Weights: 1 adult male, 12.5, fat; 2 adult females, 10.0, 10.0, little fat, fat.
Vermivora peregrina. Tennessee Warbler.—Weights: 1 adult male, 10.9, very fat; 1 immature male, 12.9, very fat; 2 adult females, 9.1, 12.5, moderately fat and very fat. The relative scarcity of Tennessee Warblers in the sample is surprising. They are common in the area in spring.
Vermivora c. celata. Orange-crowned Warbler.—Weights: 9 adult males, mean 8.8 (7.7-10.9), little fat to fat; 13 immature males, mean 8.92 ± .15, S. D. .53, little fat to fat; 5 adult females, mean 8.8 (8.3-10.3), little fat to moderately fat; 17 immature females, mean 9.13 ± .08, S. D. .72, little fat to fat. Of the 19 Orange-crowned Warblers killed on October 7, 11 had little fat, 6 were moderately fat, and only 2 were fat. No one-night sample of any other warbler killed at Topeka had less fat than this group of warblers. Furthermore, our sample (including 11 males) from October 7 (all sex- and age-classes) averaged 8.81 grams; the sample of 13 (including only 4 males) from October 1 averaged 9.1 grams. If one can assume, for any one species, that individuals undertake nocturnal migration only when they are physiologically ready, and this includes a certain amount of fat as a fuel source (Wolfson, 1954), then this further assumption seems justified: birds killed in migration with little fat must have flown longer or farther or both than birds killed with more fat. No further speculation on this point is permissible with our data, but the possibilities for study of future large kills, especially where actual time of death of the birds is known, are obvious.
Vermivora r. ruficapilla. Nashville Warbler.—More Nashville Warblers were picked up at Topeka than any other species and they are discussed in detail elsewhere in this report. The four sex- and age-classes can be identified with fair accuracy on plumage characteristics alone. Adult males have a large amount of reddish-brown in the crown, not completely veiled by the gray tips of the crown feathers. Immature males have a smaller but distinct crown patch, usually completely veiled. All males, compared with females, are grayer on the sides of the head, have a more nearly white eye-ring, and show clearer yellow on the throat. Adult females differ from immature females in that they more often have a trace of rufous in the crown and tend to be brighter below than the immatures. Of 177 specimens, 20 were very fat, 108 were fat, 46 were moderately fat, and 3 had little fat.
Parula americana. Parula Warbler.—Weight: 1 adult female, 7.9, fat.
Dendroica petechia aestiva. Yellow Warbler.—Weights: 1 immature male, [Pg 14]10.2, fat; 3 adult females, 8.8, 9.5, 10.1, moderately fat; 2 immature females, 9.0, 9.4, little fat and fat.
Dendroica magnolia. Magnolia Warbler.—Weights: 1 adult female, 9.0, moderately fat; 2 immature females, 7.9, 10.3, moderately fat and fat.
Dendroica c. caerulescens. Black-throated Blue Warbler.—Weights: 2 immature males, 13.8, 14.1, excessively fat; 1 immature female, 11.4, fat. This species is rare in Kansas. Although its breeding range is almost entirely east and north of Kansas, records in files at the University of Kansas show that more specimens have been taken in western than in eastern Kansas.
Dendroica c. coronata. Myrtle Warbler.—Weight: 1 immature female, 11.6, fat.
Dendroica pensylvanica. Chestnut-sided Warbler.—Weights: 2 immature females, 8.1, 10.0, little fat. Only one specimen from Kansas had been preserved previously although the species is a regular transient in small numbers throughout the state.
Dendroica castanea. Bay-breasted Warbler.—Weights: 1 adult male, 19.2, excessively fat; 1 adult female, 11.7, little fat; 1 immature female, 11.2, moderately fat. Only 5 specimens of this warbler have been taken previously in Kansas, 4 in spring (Ruth, 1952:18-19) and 1 in fall.
Dendroica p. palmarum. Palm Warbler.—Weights: 2 immature males, 9.9, 10.9, moderately fat; 2 unsexed immatures, 9.1, 9.4, moderately fat. This species has been taken in fall in Kansas only once before (KU 26353, taken by Wetmore, at Lawrence, on October 5, 1907), but probably occurs regularly in both spring and fall migration.
Seiurus a. aurocapillus. Oven-bird.—Weights: 2 adult males, 22.5, 23.8, fat and very fat; 14 immature males, mean 21.89 ± .66, S. D. 2.46, fat to very fat; 8 adult females, mean 21.4 (18.3-25.7), moderately fat to fat; 6 immature females, mean 18.2 (15.6-20.0), moderately fat to fat.
Seiurus noveboracensis notabilis. Northern Water-thrush.—Weights: 3 immature males, 18.1, 18.6, 22.2, moderately fat to fat; 1 immature female, 22.2, fat. Referring these birds to notabilis is a somewhat arbitrary procedure. They display some intermediacy of characters and probably stem from a population, intermediate between notabilis and noveboracensis, occupying much of central North America (cf. McCabe and Miller, 1933).
Oporornis philadelphia. Mourning Warbler.—Weight data presented elsewhere. The birds killed at Topeka provide the latest fall dates for this species in Kansas. Fifteen were killed on September 25, 64 on October 1, 2 on October 5, 11 on October 6, 2 on October 7, and 1 on October 8. We find no other records later than September 15. Of 93 specimens examined, 1 was excessively fat, 22 were very fat, 45 were fat, 21 were moderately fat, and 4 had little fat. The abundance of this secretive species in the sample was a great surprise. It had previously been considered a rather rare migrant in this area.
Geothlypis trichas occidentalis [>brachidactyla?]. Yellow-throat.—Weight data presented elsewhere. This species was second in numbers only to the Nashville Warbler in the total kill at Topeka. Of 167 birds examined, 29 were very fat, 114 were fat, 23 were moderately fat, and 1 had little fat.
The Yellow-throats are greatly in need of meaningful and comprehensive revision, which to date has been restricted to the western subspecies (Behle, 1950). Since the appearance of the 1931 A. O. U. Check-List a great deal [Pg 15]of scattered taxonomic work on the species, as yet unsynthesized, has made the picture of its geographic variation a blurry one so far as the details are concerned. Made in the absence of adequate comparative material, the above identification is to be regarded as tentative. Also, it is, unfortunately, based only on those 6 of our 176 specimens preserved as skins. Five of these are adult males, the sixth being an immature female. Compared with a series of Kentucky specimens regarded as typical brachidactyla, these birds are paler and brighter above (tending toward gray-green rather than brownish olive), brighter and more extensively yellow below, with broader, more nearly white superciliary stripes above their black masks (in males). In size they are close to occidentalis (see Behle, 1950:202). Five males have an average wing-length of 56.6 mm. (53-59); one female measures 53. Six males from Kentucky: 55.1 (53-56); four females, 51.1 (48-56). Our birds may be assumed to have stemmed from a population to the north and west which, if not occidentalis (or campicola Behle and Aldrich, of which no comparative material is at hand), is intermediate between brachidactyla and more western birds. Judging from Behle's map (1950:fig. 32), these birds may have come from an area near the confluence of three subspecies (campicola, occidentalis, brachidactyla). Long (1940:452) reports three subspecies breeding in Kansas (brachidactyla, northeast; occidentalis, west; trichas, southeast). The occurrence in Kansas of G. t. trichas as currently understood is completely out of the question.
Icteria v. virens. Yellow-breasted Chat.—Weight: 1 unsexed immature, 29.7, moderately fat.
Wilsonia p. pusilla. Wilson Warbler.—Weights: 2 adult females, 7.5, 7.8, fat, moderately fat; 1 unsexed adult, 8.3, fat.
Wilsonia canadensis. Canada Warbler.—Weight: 1 immature female, 10.0, little fat. We know of only five other specimens from Kansas, although this warbler seems to be a regular migrant in small numbers in the state.
Setophaga r. ruticilla. American Redstart.—Weight: 1 immature female, 9.1, moderately fat.
Dolichonyx oryzivorus. Bobolink.—Weights: 2 adult females, 39.5, 42.9; 2 immature females, 38.8, 42.0; all excessively fat. Specimens of the Bobolink previously have been taken in fall in Kansas only on September 20 and 24, 1933, near Lawrence, by Long and Preble (Long, 1934).
Pheucticus ludovicianus. Rose-breasted Grosbeak.—Weights: 1 adult male, 50.4, fat; one immature male, 54.5, very fat.
Passerina cyanea. Indigo Bunting.—Weights: 1 adult male, 18.4, fat; 2 immature males, 17.2, 17.2, fat and very fat; 2 adult females, 14.3, 16.9, moderately fat and very fat; 1 immature female, 13.4, little fat. The sample was carefully checked for Lazuli Buntings (Passerina amoena); none was found.
Spiza americana. Dickcissel.—Weight data presented elsewhere in this paper. Dickcissels were picked up at the television tower on October 1 (31), 5 (1), 6 (3), and 7 (1). These birds, together with an adult female taken 3 miles east and 3 miles south of Lawrence, on October 11, 1953, by Tordoff, are the only specimens of this species taken as late as October in Kansas. The Dickcissel becomes inconspicuous in late summer and many observers here and elsewhere have thought the species disappeared much earlier than it really does (see Ganier, 1949). Of 34 specimens, 20 were very fat and 14 were fat.
Passerculus sandwichensis nevadensis. Savannah Sparrow.—Weights: 1 adult male, 19.4, fat; 2 immature males, 18.3, 19.0, moderately fat; 5 adult females, mean 17.2 (14.8-19.5), little fat to fat; 4 immature females, mean 18.0 (16.9-19.6), moderately fat to fat. Many of the Savannah Sparrows migrating through Kansas have in the past been referred to the subspecies P. s. anthinus (= alaudinus of the 1931 A. O. U. Check-List) by various workers (see Long, 1940:454). As Peters and Griscom (1938:464-5) have shown, true anthinus, breeding in the far northwest, ordinarily occurs in migration only in the western part of the country, the breeding Savannah Sparrows of a large part of the central continental region (east to southern Wisconsin) being P. s. nevadensis as now understood. Migrants of this pale, clay-colored subspecies should be abundant in Kansas, and all of the specimens in the present sample are referable to it.
Ammodramus savannarum perpallidus. Grasshopper Sparrow.—Weights: 3 adult males, 16.4, 17.6, 20.6, moderately fat, fat, fat; 5 immature males, mean 18.1 (16.0-20.2), little fat to fat; 5 adult females, mean 17.9 (16.8-18.9), moderately fat to very fat; 5 immature females, mean 18.1 (16.8-20.6), fat to very fat.
Passerherbulus caudacutus. Leconte Sparrow.—Weights: 1 immature male, 11.2, moderately fat; 1 immature female, 12.2, moderately fat.
Ammospiza caudacuta nelsoni. Sharp-tailed Sparrow.—Weights: 2 adult males, 15.2, 17.1, moderately fat and very fat; 1 adult female, 13.3, little fat. Five specimens of this species have been taken previously in Kansas, all in October in the eastern part of the state. Additionally, several observers have reported birds seen but not collected. The three birds from Topeka were picked up on October 6, 7, and 10 and are the only specimens taken since 1907. Possibly our specimens from Topeka struck the tower on the same night. Tordoff noticed, upon preparation, that the specimens from October 7 and 10 showed progressive drying of the extremities and spoilage as compared with the bird picked up on October 6.
Junco hyemalis cismontanus. Slate-colored Junco.—Weight: 1 immature female, 16.4, little fat. Juncos of hybrid type, whether J. h. hyemalis × J. oreganus subsp. or true J. h. cismontanus, are fairly common in eastern Kansas.
Spizella pallida. Clay-colored Sparrow.—Weights: 2 adult males, 11.6, 12.2, both fat; 1 immature male, 11.8, fat; 1 adult female, 12.5, fat; 7 immature females, mean 11.1 (9.7-12.5), little fat to fat.
Passerella iliaca iliaca. Fox Sparrow.—Weight: 1 adult female, 29.4, little fat. A trifle grayer above than any of several Kentucky specimens, this bird nevertheless seems well within the range of variation of iliaca.
Melospiza l. lincolnii. Lincoln Sparrow.—Weights and measurements are discussed elsewhere. Of 81 specimens, 15 were very fat, 47 were fat, 12 were moderately fat, and 7 had little fat. Interestingly, there is no evidence that the large southern montane subspecies (M. l. alticola) has contributed to the present sample. No bimodality is evident in the curve of wing-length in our birds, the largest of which barely approach, the small extreme recorded for alticola by Miller and McCabe (1935:156).
Melospiza georgiana ericrypta. Swamp Sparrow.—Weights: 3 immature females, 14.3, little fat, 16.7, 17.0, moderately fat. Swamp Sparrows examined were all more or less brightly colored and seem to belong to this northern subspecies.
Melospiza melodia juddi. Song Sparrow.—Weights: 1 adult female, 19.4, [Pg 17]little fat; 1 unsexed immature, 16.0, little fat. A large proportion of the migrant and wintering Song Sparrows in eastern Kansas probably originate from the range of this subspecies in the northern plains. Melospiza melodia euphonia.—One immature female (not weighed) was picked up below the tower on October 27, 1954, and thus does not appear in Table 1. The specimen proved typical of this generally more eastern subspecies upon comparison with a large series from Kentucky. For what it may be worth we refer the single specimen to this subspecies. Long (1940:456) reported two eastern subspecies from Kansas ("beata," melodia). All Kansas specimens genuinely of eastern origin probably originate from the range of euphonia, as now understood.
The reliability of certain of the conclusions which might be drawn from data of the kind presented herein depends largely on the randomness of the sample. To what degree does this sample provide a true cross-section of the nocturnal migrants present over the area on a given night or succession of nights? As far as the relative abundance of species in the sample is concerned, there is little doubt that it is not at all random. The absence of such species as the Gray-cheeked Thrush (Hylocichla minima), among the passerines, and many of the shorebirds known to be migrating through the area at the time is evidence for this statement. Quite possibly many seminocturnal species did not strike the tower at all for the simple reason that they could see it, and certain large-eyed diurnal species (such as thrushes and shorebirds) may avoid collision to some extent, thus not appearing in the sample in proportion to their actual numbers. Finally, some or all of the species concerned probably migrate partly by day. The sample may to some degree reflect the true relative abundance of closely related species. For example, there is little doubt that, as shown by the sample, Nashville Warblers are more numerous locally at this season than Tennessee Warblers, a fact that can readily be corroborated by ordinary field observation. Also, the sample is useful in suggesting the actual abundance of species which are furtive and/or difficult to identify under normal field conditions, for example, the Mourning Warbler and Philadelphia Vireo. It is obvious that the sample should reflect the true relative abundance at one place and time of any two species with equal tendency to migrate by night and equal tendency to strike the tower. Since the facts in regard to both tendencies are at present unknown for most species, we think that interspecific comparisons should be avoided or approached with extreme caution.
In respect to the relative abundance of the various sex- and age-[Pg 18]classes within a given species, the sample is, we think, as close to random as is possible to obtain. Certainly it is greatly superior to samples obtained by field collecting, where possible differences in habits, wariness, and experience of the birds, and subconscious (if not conscious) selection by collectors can all bias the results. Dwight (1900:128-9) believed that the greater wariness of adult birds was almost entirely responsible for the seemingly disproportionate number of immatures in autumn and gave some observational evidence in favor of his views. The large percentage of adults in some of the samples here treated tends to reinforce Dwight's position. To a somewhat lesser extent, this advantage in randomness of accidental kills over routine collecting may be supposed to apply also in demonstrating the composition by subspecies of a single migrant species.
So far as particulars already mentioned are concerned, the present sample or other samples of tower-killed birds would seem to be in no way superior (that is, more nearly random) to samples obtained in connection with lighthouses and other lighted objects, and ceilometers. In one important respect, however, it is probably somewhat superior to these as the dimly red-lighted structure has not been shown to have any important collecting or attracting influence. Thus, in computations intended to estimate the over-all abundance of migrants, the sample should be more reliable than samples involving bright light with its possible attracting effect.
If it can be assumed that nocturnally migrating birds are approximately uniformly spaced across the sky and that the red lights did not attract birds which would otherwise have missed the tower, it is possible to compute the volume of migration from the sample killed. In regard to the first assumption, both Stone (1906:250-251) and Lowery (1951:409-413) have presented evidence of fairly uniform distribution of nocturnal migrants. We have no information on the second assumption beyond the facts that birds do not strike the high towers on clear nights or lower towers even on stormy nights.
On nights when large numbers of birds struck the 950 foot Topeka tower, only a few struck a 500 foot radio tower, also lighted with red lights, at Lawrence, 24 miles east, under similar weather conditions. Most of the birds found at Topeka were fairly close to the base of the tower, indicating that they struck the tower itself[Pg 19] or that they were flying high enough to strike guy wires only fairly close to the tower. The scarcity of birds under the guy wires some distance from the tower at Topeka and at the radio tower at Lawrence causes us to think that most of the birds were flying more than 450 feet above the ground. On this basis, we have computed numbers of migrants passing through a plane one mile long and 500 feet high (2,640,000 square feet), intersecting the assumed path of migration at right angles. Vertically, the theoretical plane begins at 450 feet above ground and has its top edge at 950 feet. The solid (discounting spaces between girders, etc.) cross-sectional area of the tower intersecting this plane was computed by actual measurement to be 1685 square feet. On the night of September 30-October 1, 585 birds were killed. By computation (585/1685 = X/2,640,000), approximately 916,000 birds passed through the mile-long plane that night. On each of the nights of October 5-6 and October 6-7, approximately 230,000 birds passed through this plane. By comparison, Lowery (1951:436) recorded maximum station densities in one night in spring of 63,600 birds at Tampico, Mexico, and 51,600 at Lawrence, Kansas, as determined by moon-watching. Lowery's figures refer to numbers of birds crossing any part of a circle one mile in diameter and are roughly comparable to ours if, as we think, most of the birds at Topeka were flying at altitudes between 450 and 950 feet above the ground.
It must be realized that these figures are only approximations. One variable ignored is the frontal extent (or area, viewed from the front, subject to damage by striking an obstruction) of the birds themselves. Since practically all birds killed showed head or trunk injuries, rather than a high proportion with only broken wings, we chose to disregard frontal extent of the birds in our calculations. If our figures are inaccurate by as much as 50 per cent in either direction, which seems unlikely to us, they still give some idea of the tremendous volume of nocturnal migration under some conditions.
It may be more meaningful to compute numbers of migrants by species. This can be done readily by making appropriate substitutions from Table 1 in the equation given above. For example, on the night of September 30-October 1, approximately 147,000 Nashville Warblers passed through the mile-long plane and on the same night, 100,000 Mourning Warblers and 14,000 Philadelphia Vireos. Neither of the last two species would be judged to be abundant migrants in autumn in eastern Kansas by ordinary field[Pg 20] observations; the television tower sample, however, indicates that these as well as other species must often be overlooked when they do stop in Kansas.
History of the Subject.—For a long time it has been known in a general way that old and young birds and males and females of some species do not always migrate at the same times, by the same routes, or even to the same places. This is a subject about which much has been written. Reading the summaries of some general texts, it is easy to acquire the impression that the facts of the matter are well known. On the contrary, they are poorly known and much remains to be learned before differential migration is understood. This can best be indicated by a brief survey of the literature.
The importance of the subject was emphasized by Meinertzhagen (1930:52) in one of the later reviews of differential migration: "The main problem concerns the Cause of Migration, the Stimulus which compels Migration and the Origin of the Migratory Habit.... There is, however, a minor problem which affords valuable evidence in helping us to solve the major problem, bearing very directly on it, namely, the order of sex and age on migration."
The mystery of how birds, especially the young, find their way in migration has fascinated students since the earliest times. The quite natural though purely anthropomorphic conclusion of early scholars was that the old birds led the young on migration. This attractive idea persisted long after ornithology began to grow into a science. The classic theory was restated by Palmén (1876:267), in one of the first thorough reviews of the subject of migration, as follows: "Directe Beobachtungen in der Natur ergeben, dass die Schaaren von ziehenden Vögeln allgemein ältere und stärkere Individuen als Anführer des Zuges haben." Variously modified, this view continued to crop up for some time and still found support in the 1890's (see Dixon, 1892:69). Gätke (1895:101) correctly questioned the credibility of Palmén's "direct observations."
With the gradual abandonment of the unsupportable classic theory, diametrically opposed views were adopted by workers on opposite sides of the Atlantic. The American stand was ably expressed by Brewster (1886), who went to great pains to state his case and give evidence, and who was later supported by Allen (1896:144-147; 1909:17). The Americans held that adult birds nearly always preceded the young in migration, and this was based on much evidence, whether or not correctly interpreted. Dwight[Pg 21] (1900:127) also gave evidence in favor of this theory. Equally definite, if, as has later been shown, somewhat vaguely documented, was the famous work of Gätke (1895:see pp. 100-113), who after many years' observation of migrant birds in Heligoland concluded the exact opposite, that young in general precede adults (see critiques of Allen, 1896:144-147; Wiegold, 1926:5). Gätke's dissenting opinion was for a time supported enthusiastically by British workers (Gurney, 1923:579-580).
As so often happens, neither extreme has withstood the test of time, and more recent summaries (Meinertzhagen, 1930:55-56; Thomson, 1926, 1936:488-489; Wiegold, 1926) have tended to compromise. Many exceptions to Gätke's extreme conclusion have been detected. Exceptions to the Brewster-Allen stand have also been discovered, although work along these lines on the American side has lagged somewhat. Rowan (1926) has given further evidence on the migration of certain shorebirds, and some evidence has accrued in relation to particular species and groups as a result of life-history and banding studies (see Pitelka, 1946). Authors of major works on migration, however, have either been preoccupied with other phases of migration or avoided the issue. In an able study (one of several on related subjects) of the composition by sex and age of migrant populations in north Germany, Drost (1935:177) did not go into the question of order on migration.
One is left with the impression that the whole subject is still unsettled. While earlier workers sought to reduce the entire matter to law, the latest disclaim the possibility of generalization. After summarizing Brewster's and Gätke's opinions, Thomson (1926:79) wrote: "It would seem, in any event, that no general rule can be laid down." Meinertzhagen's summation (1930:56-57) still represents fairly well the status of our knowledge: "Order of sex and age on autumn passage is very difficult to arrive at, as evidence is conflicting. But, on the whole, it seems that birds flock together, old and young, preparatory to moving south, and do in many cases initiate migration in company.... But once movement is initiated, among birds which do not habitually fraternise in flocks, adults, and especially males, will naturally outstrip the less virile females and still less virile offspring.... The consequence is that any observer at an intermediate station such as Heligoland is, in noting birds of the year as first arrivals, has not had an opportunity of noting the flocks of adults which have passed without alighting. On the other hand, there is very definite evidence to show that among certain species, adults follow their offspring on[Pg 22] migration. The reason for different behaviour among different types of birds remains obscure." We regard much of this as still theory.
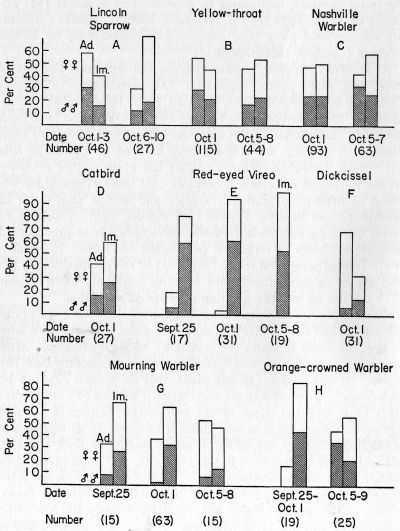 Fig. 1. Composition by age and sex as found in one or more series of
each of eight species of birds included in the Topeka sample. Each separate
series is represented by a single histogram, the histograms for a species
being grouped with the earliest series on the left. Each histogram expresses
the numbers of adults (left-hand column) and immatures (right-hand
column) in terms of percentage of the whole series. Thus the two bars
of each couplet add up to 100 per cent. The hatched portion of each bar
represents males, the clear portion females.
Fig. 1. Composition by age and sex as found in one or more series of
each of eight species of birds included in the Topeka sample. Each separate
series is represented by a single histogram, the histograms for a species
being grouped with the earliest series on the left. Each histogram expresses
the numbers of adults (left-hand column) and immatures (right-hand
column) in terms of percentage of the whole series. Thus the two bars
of each couplet add up to 100 per cent. The hatched portion of each bar
represents males, the clear portion females.
It would be difficult to imagine a better way of resolving the problems and uncertainties just reviewed than by the detailed analy[Pg 23]sis of large samples of migratory birds killed at random at various points and times. An analysis of the sample of birds accidentally killed at Topeka is presented here as an initial step in this direction. Although the conclusions based on this sample are tentative and may in time be altered, the data themselves are definite. If this general type of analysis is repeated again and again—applied to samples taken on many dates and in many localities—a mass of hitherto unparalleled evidence for the study of migration will emerge.
Differential Migration OF Sex- and Age-classes as shown by the Topeka Sample.—Smaller samples have not been treated. Species affording samples seemingly large enough to justify at least preliminary analysis were: Catbird, Red-eyed Vireo, Mourning Warbler, Dickcissel, Nashville Warbler, Orange-crowned Warbler, Yellow-throat, and Lincoln Sparrow (Fig. 1). For all of these except the Catbird and Dickcissel, at least two samples from a week or more apart were available for comparison in an effort to detect trends in migration. Fig. 1 shows the actual ratios of sex- and age-classes observed in samples of the species listed above. Each of the last four species provided two separate samples, of sufficient size to warrant an attempt at measuring the statistical significance of the observed changes in adult-immature ratios (Table 2).
Table 2.—Statistics of the Ratios of Adults to Immatures in Four Species
| Species | Dates of samples | Total number | Number and percentage of adults[1] | Difference (in %)[2] | P[3] |
| Nashville Warbler | Oct. 1 (93) Oct. 5-7 (63) | 156 | 45 (.484) 26 (.413) | .071 | .36 |
| Orange-crowned Warbler | Sept. 25-Oct. 1 (19) Oct. 5-9 (25) | 44 | 3 (.158) 11 (.440) | .282 | .05 |
| Yellow-throat | Oct. 1 (115) Oct. 5-8 (44) | 159 | 62 (.540) 20 (.455) | .085 | .34 |
| Lincoln Sparrow | Oct. 1-3 (44) Oct. 6-10 (27) | 71 | 27 (.614) 8 (.296) | .318 | .01 |
[1] Percentage of immatures equals 1.000 minus percentage of adults.
[2] Standard error of the difference between ratios was computed by the formula
| σe = | √ | Pe Qe ( | 1 | + | 1 | ) | , |
| N1 | N2 |
where Pe equals percentage of adults and Qe equals percentage of immatures in the entire sample.
[3] Probability of error; i. e., a P of .01 means there is one chance in 100 that the difference observed does not represent an actual difference in nature.
Upon the application of statistical methods it soon became evident that, unless changes in ratio between two samples are marked, large samples would be required in order to reach conclusions of high statistical significance in a single study of the present type. In this case (see Table 2), the Lincoln Sparrow and Orange-crowned Warbler, though represented by only moderate-sized series, show marked changes in age composition over the period studied, and the statistical treatment indicates a high degree of probability that these changes are real. Assurance that the lesser changes observed in the Nashville Warbler and Yellow-throat are real, on the other hand, is much less, even though the samples are larger. Few if any of the samples here discussed are as large as might be desired. Therefore, conclusions based upon them (see below) are to be regarded as tentative. Many other, future, samples will perhaps also be insufficient in size in themselves. There are, however, statistical advantages to repetition which will serve to make the repeated analysis even of small samples significant and valuable.
Certain of the samples not treated statistically show ratios that can be seen by inspection to be probably significant. For example the almost complete absence of adults from the three samples of Red-eyed Vireos (Fig. 1E) cannot be disregarded in view of the size of the whole sample of the species. The same applies to the high percentage of adult females and the near absence of adult males in the sample of the Dickcissel (Fig. 1F). The continuity in direction of changes observed in the three samples of the Mourning Warbler (Fig. 1G) and Red-eyed Vireo is likewise probably significant, even though some of the samples compared are small. It seems to us that the application of statistical methods to these species should await the accumulation of more material. For anyone desiring to treat them statistically now, the data are inherent in this paper.
We have not computed the standard errors of the ratios of sexes within age groups (except experimentally in a few cases). This can easily be done, however, and the significance of a given ratio determined, on the assumption (perhaps sometimes dubiously justifiable) that the sex-ratio in the species concerned is one:one. Obviously there is no point in computation of the standard errors of adult-immature ratios in single samples (such as that of the Dickcissel) until the actual ratio prevailing in the species in nature at the season in question is known for comparison with the observed ratio. Our formal statistical treatment, therefore, has been limited[Pg 25] to an examination of the significance of the changes between adult-immature ratios in samples of the same species taken a number of days apart.
The samples suggest several patterns of differential migration of sex- and age-classes. Indeed, the important consideration brought out—in our opinion not hitherto sufficiently emphasized in literature—seems to be that in generalizing about adults and immatures, one must be careful to take sexes into account, and conversely, in generalizing about males and females, one must consider also age. In other words, there are really four classes to be considered. This poses additional problems in analysis and introduces the need for still larger samples in order to reach significant conclusions. To illustrate: an adult-immature ratio of 40:20 (N = 60) may be satisfactorily significant, while within the 40 adults a ratio of 25 males:15 females may not be. Were the original sample 80:40 (N = 120) with male adults 50 and female adults 30, it is obvious that the significance of the latter ratio would be greater. The same applies in reverse if the greater emphasis is placed on sex and the lesser on age. Because of the moderate size of the samples this problem has been felt in the present study in respect to sex ratios within age groups, many of which must at present be regarded as of tentative significance.
In short, what the earlier ornithologists regarded as a simple problem is in reality a complex one. There are only two patterns in what may be called the Brewster-Gätke argument: adults first or immatures first (with of course the further possibility of both at the same time). Both patterns occur, as is now known, at least to some extent. But actual patterns, as suggested by our samples, are more complex when all classes are considered. It will readily be seen that, if adult males, immature males, adult females, and immature females be regarded as units, each with certain migratory characteristics, the combinations of these units in various orders of migratory precedence are potentially numerous. In fact, of course, they do not behave strictly as units (or perhaps very rarely so), but our data strongly indicate that the tendency exists in many cases. This may be stated another way. The present samples may be reduced to two basic patterns, fitting the classic early American (adults first) and early European (immatures first) theories. But, either such simple arrangement is compounded in some, perhaps in truth in all, instances by differential migration of the sexes within each age class. This proposition can also be stated backwards:[Pg 26] the samples show differential times of migration of the sexes, compounded by differential times of migration of the age groups within each sex. The order in which these matters are approached depends on what one is trying to find out. Influenced by the literature, in which most emphasis has been placed on age, we have approached the problem from that standpoint. The data and figures here given, however, can be juggled if one wishes to place first emphasis on the order of sexes in migration.
Bearing in mind what has just been said, particularly in respect to sizes of samples necessary for significance, let us consider the patterns of migration suggested by the Topeka sample. These are as follows:
(1) An early migration largely composed of adults, giving way later on to a preponderance of immatures. Regardless of variations among them, samples showing this basic pattern are in line with the opinions of Brewster (1886) and his followers. This pattern is here shown by the Lincoln Sparrow, Yellow-throat, Nashville Warbler, Catbird (one sample only), and Red-eyed Vireo (Fig. 1, A, B, C, D, E). The evidence of these and all other samples would admittedly be more conclusive if the samples were further apart in time or, better still, were there more of them. There is evidence that differences in migration of the sexes, within age classes, influence this pattern, sharply in some instances. In the later samples of Lincoln Sparrow, Yellow-throat, and Red-eyed Vireo (Fig. 1, A, B, E) there are relatively fewer males, both adult and immature, than in the earlier samples and this may be true also of the Catbird, judging from the single sample. The Red-eyed Vireo (Fig. 1, E) is characterized by small number, or absence of, males in each sample but the samples are not significantly different, and can be regarded as one. Although the samples of the Dickcissel and Mourning Warbler (Fig. 1, F, G) show a somewhat different over-all pattern and are discussed further on, they also contain few adult males. Since these samples are from a period that is near the end of the migration of Red-eyed Vireos, Mourning Warblers, and Dickcissels, it may be assumed tentatively that the adult males have already migrated. Meinertzhagen (1930:56) postulated that in many species there is an earlier or more rapid migration of adults, particularly males, and the data for the above species in our sample tend to support his assumption. But our data suggest in addition that in some species immature males migrate earlier, or more rapidly, than do immature females, just as adult males precede adult[Pg 27] females in some instances. Within this general pattern (adults first) another variation is shown by the Nashville Warbler (Fig. 1, C) in which the later sample of adults is heavily weighted towards males, even though an increasing over-all proportion of immatures is evidenced. In this case, and contrary to Meinertzhagen's suggestion, it would seem that adult females have preceded or outstripped adult males in migration.
(2) An early preponderance of immatures, followed by a preponderance of adults. The several species of birds at Topeka that display this pattern conform with the conclusions of Gätke and other early Old World ornithologists that in most species immatures precede adults in migration. In the present sample two variations of this pattern occur.
(a) In the Dickcissel (Fig. 1, F) and the Mourning Warbler (Fig. 1, G), immatures decrease more markedly than adults (visible in samples of Mourning Warbler; inferred in Dickcissel), leaving the adults in the majority in the closing phase of migration. The distinctive and interesting feature in each of these two species is the ascendancy in numbers of adults despite the almost complete disappearance of adult males. The relative increase of adults is here caused by a retarded migration of adult females, which linger conspicuously behind all other classes. Something of this nature was suggested, in theory, by Dixon (1892:70) who thought that adult females are delayed by "maternal duties." It was hinted at also by Dwight (1900:127) who thought that in some species females molted later than males as a result of prolongation of parental responsibilities. As mentioned already, there is need for caution in interpreting the present samples because the Dickcissel is represented only by one sample and two of the three samples of Mourning Warblers are small. In the case of the Mourning Warbler, the samples may be regarded as one, nearly lacking in adult males. The progressive increase of adult females, however, may be significant; at least there are enough of these to make division of the birds into three samples enlightening. There is, of course, some chance that the majority of adult males have not yet migrated, or are migrating by a different route. This seems unlikely in both cases. October 1 is late in the migration of the Dickcissel and it seems that large-scale migration would not occur much later, and in the case of the Mourning Warbler adult males are rare in all three samples, extending over a considerable period and reaching late into the probable migration period of the species. It is inter[Pg 28]esting to conjecture just when and where adult male Mourning Warblers do migrate in autumn. Brewster (1886:16) wrote: "This species arrives at Cambridge [Massachusetts] about September 12, and during the remainder of the month is ... abundant.... The adults, however, are so very uncommon that I have never known them [to] represent more than five per cent of the total number of individuals. They do not seem to be more numerous in the earlier flights than towards the close of the month, and I am very sure that they cannot be found in this locality before the young begin to appear." While the present samples show an abundance of adult females of this species (could Brewster have failed to recognize these as adults?) the whereabouts of the adult males remains a mystery.
(b) Another variation is displayed by the Orange-crowned Warbler (Fig. 1, H). Here also there is an increase of adults towards the end of migration, but this increase is marked by a growing percentage not of females but of males. Locally this species is a late migrant compared with most others of the Parulidae. Thus the first sample, composed of birds taken September 25-October 1, may be regarded as fairly early in the fall migration. Immature birds compose 84.2 per cent of this sample, there being no adult males at all. By October 5-9 the picture has changed markedly, the sample being composed of 44 per cent adults (82 per cent of which, in turn, are males) and 56 per cent immatures. In view of this trend one can not help suspecting that a still later sample would show a majority of adults, perhaps nearly all males. This of course does not necessarily follow; the migration of immatures could simply be more protracted, and could have commenced earlier, than that of adults.
Little imagination is required to see how enlightening it might be could we analyze thoroughly the patterns of all migrating species. When the detailed facts are available, it seems likely that general trends will emerge which may be of great significance to the study of migration in general. A final point which must eventually be clarified is determination of the extent of variability in the pattern of each species from year to year and locality to locality.
Once patterns of precedence in migration of different classes are established, search into the life-histories of the species concerned may help to explain the peculiarities discovered. In the present case, for instance, we find a possible clue to the reason for the high proportion of adult females of the Dickcissel late in migration, as[Pg 29] shown by our sample. Gross (1921:14-15) presented evidence that adult female Dickcissels molt considerably later than their mates, and we have independent evidence that individuals of this species are at times almost flightless when molting the remiges!
General comment.—The exact relationship between molt and migration seems not to have been definitely established. The subject has received cursory attention in the literature and conflicting opinions have been expressed. Dwight (1900:126-128) believed that molt is completed or nearly completed before migration in nearly all passerine species that occur in New York save for certain swallows and flycatchers. Molt has since been found to precede migration of at least one of the flycatchers (Empidonax virescens) considered by Dwight to be an exception to this rule (Mengel, 1952). In Great Britain the subject of molt in migration was considered in some detail by Rintoul and Baxter (1914) and Ticehurst (1916), who arrived at conclusions similar to Dwight's. These workers also found certain swallows to be exceptions to the rule.
The above authors and others have shown that, at least among passerines, some body molt is frequently found in migrating individuals but that molt of tail feathers is much less often found and molt of remiges almost nonexistent. Baxter and Rintoul noted only four cases of molting wing feathers among hundreds of migrants. Among the diverse non-passerine orders the picture seems to be more complicated, as might be expected. We do not, however, comprehend the reasoning which led Meinertzhagen (1930:56) to summarize: "... on the whole it can be said that though birds seldom migrate when flight feathers are in quill, moult in general does not influence migration." This seems to us an obvious non sequitur. Meinertzhagen (loc. cit.) went on to say: "Males and females of one species are believed to moult simultaneously [see, however, Dwight, 1900:127], and there is no doubt that in some cases the two sexes migrate at slightly different times, and occasionally prefer different winter quarters. Birds of the year never moult their quills previous to their first autumn migration [Consultation of Dwight, 1900, who gives many examples of this, would have spared Meinertzhagen this error.], and yet they frequently follow adults on passage and sometimes precede them. There are no grounds for believing that adults have moulted their quills before birds of the year are prepared to migrate [but there are, in [Pg 30]many cases; cf. Dwight, 1900:127], in the case where adults precede the latter. Neither is there any evidence to show that adults have not moulted their quills till after their offspring are ready for passage, in the case where they follow their offspring. It does not, therefore, appear that moult is an important factor."
Comments interpolated above show our feeling that this summary is inadequate and misleading. To us it seems that the extreme rarity in migration of birds with remiges in molt is strong evidence that molt does influence at least the time of migration. It is immaterial whether this molt occurs before or after migration, although in the majority of cases it seems to take place before. Much more needs to be known of the migration pattern and molt of each species before generalizations can safely be made.
Analysis of samples of migrants can show only the presence and nature or the absence of molt in birds actually migrating. In the present instance shortage of time and manpower for preserving some and processing all of the sample resulted in incomplete data being kept on molt. We include this section to emphasize uncertainties still prevalent and to stimulate further work.
Molt in the Topeka sample.—Our limited findings coincide with those of Rintoul and Baxter (1914). Body molt was noted in a number of individuals and species. When present, this molt almost invariably was in its final stages. One immature male Rose-breasted Grosbeak (October 1) was in heavy body molt. It is perhaps worthy of mention here that this grosbeak evidently migrates at times in extensive molt. An adult male (RMM 1102) taken by Mengel near Henderson, Kentucky, on September 9, 1949, was molting plumage of body, wings, and tail, no feather of the last being longer than one half inch. This remarkable specimen had only five primaries on one side and four on the other fully functional. The outermost on the left and two outermost on the right were from the previous plumage, not yet dropped; the three innermost of each wing were new and full-length.
In the present sample molt of remiges was noted in one specimen, an adult female Indigo Bunting (October 1) with outer primaries sheathed and with molt in progress in the body plumage. The one (immature) Yellow-breasted Chat in the sample (October 1) had all of its tail feathers nearly full-length but in quill, possibly as a result of accident, and two feathers were being replaced also in the tail of an immature Clay-colored Sparrow (October 6), which was also in body molt and had some, juvenal feathers on the belly and flanks.
Body molt near completion was further noted as follows: immature male Yellow-throated Vireo (October 1), adult male Blue-headed Vireo (October 1), immature female Leconte Sparrow (October 23), several Lincoln Sparrows (various dates).
Linear measurements.—Taxonomists long have recognized in many species that males differ in size from females. Less attention, until recently, has been paid to the relative sizes of adult and immature birds. Many taxonomists, however, seem to have had an uneasy suspicion that immature birds are "untrustworthy" in comparison with adults, and immatures have often been excluded from samples when recognizable. Since, however, there are still relatively few reliably aged specimens in collections, for the most part only those immature birds immediately recognizable as such by obvious plumage differences (which are often present only in juvenal plumage) have been excluded from series. The majority of birds in first winter plumage so closely resemble adults that the two ages have been included in series for measurement. In most passerines these younger birds still bear the juvenal feathers in wing and tail and are, in size of these important parts, quite as "untrustworthy" as birds still in juvenal body plumage. Even if a complete postjuvenal molt occurs we still should not assume that first winter feathers are as long as adult winter feathers without first determining that this is so. Although aware of this problem, systematists until recently seemingly have been more or less content to disregard it, or forced to do so for practical reasons. Miller (1941:179) had little choice but to hope that size differences between adult and immature juncos were unimportant. Behle (1942:217) wrote of Horned Larks, Eremophila alpestris: "... the plumages of first-year birds and adults seem indistinguishable, though I have never quite satisfied myself that there are no differences in lengths of rectrices and remiges." He added, with logic confusing to us: "Since it is a difficult problem to determine the ages of horned larks that have passed the postjuvenal molt, this similarity of plumages is fortunate for the systematist."
In recent years, some workers have analyzed size differences between adults and immatures. Sibley (1950:115) showed that adult Red-eyed Towhees (Pipilo erythrophthalmus) had notably longer wings and tails than immatures, and the same was demonstrated in Red Crossbills (Loxia curvirostra) by Tordoff (1952). In work [Pg 32]with jays (Aphelocoma), Pitelka (1951:199) found that: "... in comparisons of dimensions of sex and age groups within a given sample, although magnitude of difference varies from one character to another, most of the averages are successively smaller for first-year males and adult and first-year females." He listed exceptions and concluded: "Segregation [of sex and age classes] has proved to be of extreme significance in an interpretation of individual and geographic variation."
Much along these lines can be learned by examination of large random samples such as that afforded by the Topeka accident. Although only a few species in this sample were measured, the results secured seem to show further the need for segregation of age classes in taxonomic work with some species.
Figure 2 shows the variation in the lengths of wing and tail in the Nashville Warbler. It is evident from the figure that in both of these characters the four sex- and age-classes differ significantly. An accurate idea of geographic variation in this species could not be obtained without separating these classes in comparisons. Age classes in spring and summer, long after the skull is completely ossified, can be segregated only if differences in plumage can be found. In the Nashville Warbler, such differences occur in fall (see annotated list) but these probably are obliterated by the partial prenuptial molt. These facts emphasize the importance, for taxonomic studies, of aged specimens collected in late summer or early fall on their breeding ground and in fresh winter plumage. Figure 3 shows the variation in length of wing in the Lincoln Sparrow in which age seems to be of much less importance than in the Nashville Warbler. Males and females of the Lincoln Sparrow differ significantly in wing-length, but adults and immatures are of nearly the same size. It would seemingly not be necessary to separate age classes in studies of geographic variation in the Lincoln Sparrow. Size data for some other species are given in the annotated list.
Weights.—Little seems to have been done to determine the correlation of weights with sex- and age-classes. Weight may be the best single index of over-all size and is especially useful to students of the physiology of migration. Weight, however, is subject to the considerable variable imposed by fat condition. In large and comparable series, this variable is probably removed insofar as comparability of means is concerned, but the high variability of weight in most cases naturally results in more overlap (less separability) between populations with means close together than is found with most linear measurements.
 Fig. 2. Statistics of variation in length of wing and tail in the Nashville
Warbler. The solid lines represent the observed ranges in millimeters.
The stippled boxes represent two standard errors (σm) to each side of
the means (vertical lines). The open boxes represent one standard deviation
(σ) to each side of the means.
Fig. 2. Statistics of variation in length of wing and tail in the Nashville
Warbler. The solid lines represent the observed ranges in millimeters.
The stippled boxes represent two standard errors (σm) to each side of
the means (vertical lines). The open boxes represent one standard deviation
(σ) to each side of the means.
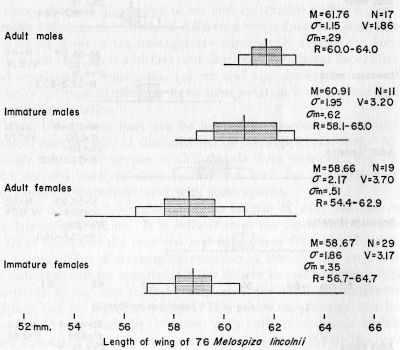 Fig. 3. Statistics of variation in length of wing in the Lincoln Sparrow.
Fig. 3. Statistics of variation in length of wing in the Lincoln Sparrow.
Figures 4-6 show variation in weight in the samples of Nashville Warbler, Mourning Warbler, Yellow-throat, Dickcissel, and Lincoln Sparrow. Each figure is essentially self-explanatory. It will be seen that in the Nashville Warbler and Lincoln Sparrow, weight is roughly proportional to wing-length (shown in figs. 2 and 3), giving about equally good separation of sex- and age-classes in the latter and poorer separation in the former. Data for these and other species indicate a generally greater weight of males than of females, but less difference according to age. In some other species, for example the Yellow-throat, immatures seem to be a little heavier on the average than adults. It is unfortunate that wing-lengths are not at present available for these.
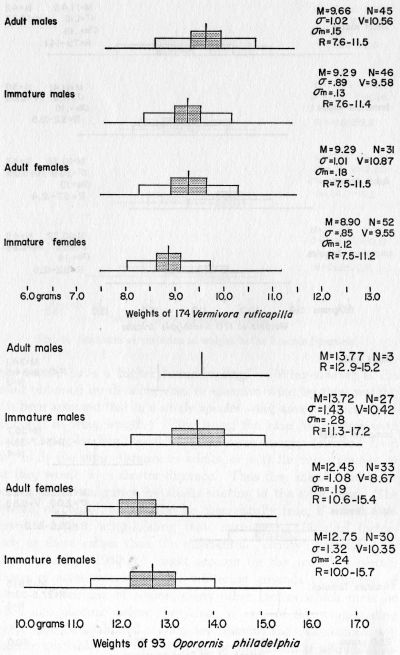 Fig. 4. Statistics of variation in weight in the Nashville Warbler and
Mourning Warbler.
Fig. 4. Statistics of variation in weight in the Nashville Warbler and
Mourning Warbler.
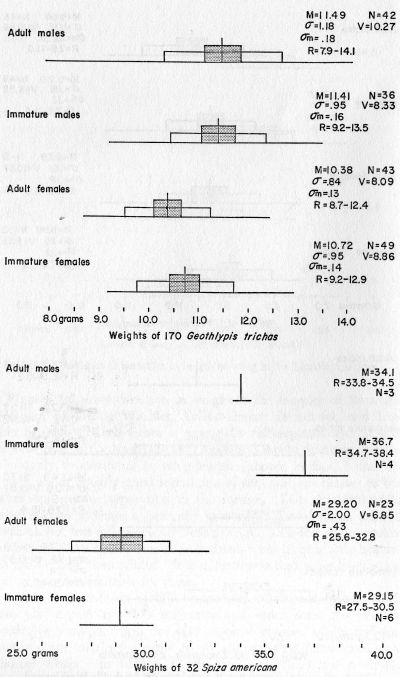 Fig. 5. Statistics of variation in weight in the Yellow-throat and Dickcissel.
Fig. 5. Statistics of variation in weight in the Yellow-throat and Dickcissel.
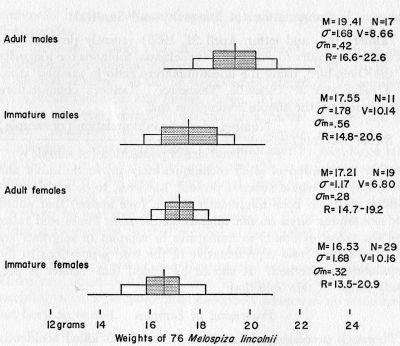 Fig. 6. Statistics of variation in weight in the Lincoln Sparrow.
Fig. 6. Statistics of variation in weight in the Lincoln Sparrow.
These comments on weight suggest an additional factor which
may play a part in rate of migration and which some day may be
profitably studied. Suppose that in some species adults and immatures
are nearly the same in weight but that immatures have
shorter wings. In such a species the immatures are relatively
shorter-winged for their weight than adults. In aerodynamic terms,
[Pg 35]
[Pg 36]
[Pg 37]they would have a higher "wing-loading." (Wing-loading is the
result obtained by dividing area of effective wing by total weight;
it is here assumed that in a single species wing area is directly proportional
to wing length.) This being the case, immatures with
higher "wing-loading" would require more energy (derived from
fat) to fly the same distance as adults, or with the same amount of
fat they would fly a shorter distance. Thus they might tend to be
outstripped in migration by adults starting at the same time. The
reverse, of course, would also be theoretically true, if adults possessed
a higher wing-loading than immatures. Physical factors
such as these rather than the differential "virility" postulated by
Meinertzhagen (1930:56) might account for the arrival of certain
classes of some species on the wintering grounds in advance of
others. There are, of course, many other factors which must be
taken into account before the effect, if any, of the wing-loading
factor can be evaluated. Data for illuminating calculations will
become available, however, with the accumulation of abundant information
on weights, measurements, and migration patterns.
Tanner (ms., and letter, April 21, 1955) recently devoted considerable ingenuity to computing by actuarial methods the longevity of the Oven-bird, using the adult-immature ratio in samples killed at a ceilometer at Knoxville, Tennessee. Tanner's computations were based on the simple assumption that
| S (survival rate) = | Number of adults in population (or sample) |
| Total size of population (or sample). |
Further application of such techniques may prove desirable and rewarding. It would seem at present, however, to be a risky procedure, as it has been abundantly shown (see above) that adults and immatures often do not migrate at the same times and rates, and the ratios of adults to immatures in samples of migrants are likely to be far from representative of the true proportions in the populations concerned. It should be added that Tanner is perfectly aware of this objection.
Thorough processing of large samples of birds killed accidentally is time-consuming. We were fortunate in having considerable assistance; even so, all desirable data could not be obtained from the 1090 birds of the present sample. As aids to others conducting studies of this kind we should mention a few points which may be of assistance.
Birds should be picked up as soon as possible after death, certainly by the end of the day after the accident and preferably much sooner. They should be weighed as soon as possible after collection (weights decrease rapidly, even under refrigeration), and the weights (in grams, to one tenth of a gram) written on tags attached to a leg of each specimen. The sample should then be sorted by species or groups of species of approximately equal size (to avoid crushing of smaller birds by larger ones), placed in boxes, paper bags, or better, air-tight containers clearly marked with date, locality, and other necessary particulars, and relegated to a deep-freeze compartment. For all but the smallest samples, such freezing units are indispensable to complete study. Once frozen, the birds may be selected for study at leisure, but time is still important, as, even when frozen, gonads may eventually deteriorate, and birds[Pg 39] eventually become desiccated which is a disadvantage if skins are to be made.
In the cases of large kills, or limited manpower, or both, it may be impossible to process all birds, however desirable this might be. If possible, however, all should be collected, identified, the numbers and species recorded, and rarities saved. Further, partial analysis, or more properly, complete analysis of a partial sample, can be made. Analyses which for any reason require randomness of sample pose a special problem. We think that in very large kills the best way to solve this problem is probably to make one or more transects across the area where dead birds are found. These transects should cross both the areas of greatest and least density (to allow for fast and slow flying species). Their width may be adjusted to give the desired number of birds, that is, the number that can be adequately processed. Another alternative would be to decide to study certain abundant species and pick up all of these. There are other possibilities, but in any event the method of sampling should be thoroughly described wherever all birds have not been processed.
The foregoing paper discusses accidents in which large numbers of night-migrating birds are killed. A brief historical review of ornithological interest in such occurrences is given, and the types of data provided by the accidents are listed and discussed. In particular, recent accidents occurring in early October, 1954, through much of eastern United States are cited, and detailed analysis is presented of a sample of 1090 birds killed one mile west of Topeka, Shawnee County, Kansas, between September 25 and October 23, 1954.
At Topeka during the period mentioned, 1090 birds representing 61 species were collected and were processed at the University of Kansas. For all specimens, weight, sex, age, and fat condition were recorded, and certain species were measured as well. Some notes on molt were taken. A total of 193 birds was preserved as study skins, and 49 as skeletons. Comments on weight, size, sex, age, subspecific identity, and status in Kansas are presented in an annotated list.
Randomness of this and other similar samples is discussed. A theoretical computation is given for several nights of the numbers[Pg 40] of migrants passing through a plane one mile in width, from 450 to 950 feet above ground level, and oriented to face the arriving migrants. The computed totals give some idea of the tremendous volume of nocturnal migration under some conditions. Potentialities of further study of such events are discussed and a comparison is made with lunar observations.
Differential migration of sex- and age-groups as shown by the larger samples taken at Topeka (Catbird, Red-eyed Vireo, Nashville Warbler, Yellow-throat, Mourning Warbler, Dickcissel, Lincoln Sparrow) is discussed and the history of theories on this subject reviewed. It is shown that age and sex must both be taken into account in studies of differential migration. Several patterns of migration are displayed by the species analyzed, adults migrating earlier than immatures in some instances, young earlier than adults in others, but all seemingly being complicated to varying degrees by differential migration of sexes within age groups. It is suggested that explanations of these patterns may be sought in the life histories of the species involved.
Molt in relation to migration is discussed briefly, and it is held that there is an important relationship between molt and time of migration. Specimens noted to be in molt are listed.
Size differences, in wing length, tail length, and weight are discussed in relation to sex and age, and variation in one or more of these characters is analyzed as found in the Topeka samples of Nashville Warbler, Mourning Warbler, Yellow-throat, Dickcissel, and Lincoln Sparrow. It seems that in some instances significant size differences prevail between adults and immatures and that these age classes should be separated in taxonomic work with species where differences in size are known to exist. When the facts are not known they should be determined, and the large samples collected in accidents to nocturnal migrants present excellent opportunities for ascertaining the facts.
Other uses of material obtained in large migration accidents are discussed, such as computations of longevity and the problems of processing large, accidentally-killed samples. Care should be taken to select samples as nearly random as possible when all birds cannot be processed.
Repeated and thorough analysis of accidental kills should provide a mass of valuable data bearing on many questions and problems which have thus far been little studied.
Allen, J. A.
1880. Destruction of birds by light-houses. Bull. Nuttall Orn. Club, 5(3):131-138, July.
1896. Gätke's 'Heligoland.' Auk, 13(2):137-153, April.
1901. Barrington's 'The Migration of Birds at Irish Light Stations.' [Review.] Auk, 18(2):205-206, April.
1909. An American's views of bird migration. Brit. Birds, 3(1):12-19, June 1.
Barrington, R. M.
1900. The migration of birds as observed at Irish lighthouses and lightships [etc.]. London, R. H. Porter; Dublin, Edward Ponsonby. Pp. XXV, 285, 667. (Not seen; citation from Mullens and Swann, Bibliogr. Brit. Orn., 1917, p. 43 and Zimmer, Cat. Ayer Coll., I, p. 40, 1926; see also reviews, Ibis, 1900:677-679, Auk, 1901:205-206; some sources list pagination as XXV + 667, possibly in error.)
Behle, W. H.
1942. Distribution and variation of the Horned Larks (Otocoris alpestris) of western North America. Univ. California Publ. Zoöl., 46(3):205-316, May 20.
1950. Clines in the Yellow-throats of western North America. Condor, 52(5):193-219, September-October (September 25).
Brewster, W.
1886. Bird migration. Mem. Nuttall Orn. Club, No. I, pp. 1-22, Cambridge, March.
Carson, L. B.
1954a. [Destruction of birds at a television tower at Topeka, Kansas.] Topeka Aud. News, 9(1):pp. = 1-2, July, August, September [published October]. [Unsigned, unpaged, and untitled article by L. B. Carson, ed.]
1954b. [Further destruction of birds at a television tower at Topeka, Kansas.] Topeka Aud. News, 9(2):pp. = 5-7, October, November, December [published December]. [Unsigned, unpaged, and untitled article by L. B. Carson, ed.]
1954c. New records for fall migrants in eastern Kansas. Kansas Orn. Soc. Bull., 5(4):27-29, December.
Dixon, C.
1892. The migration of birds. London, Chapman and Hall. Pp. XVI + 300.
Dobben, W. W. H. van, and M. F. Bruyns
1939. Zug nach Alter und Geschlecht an niederländischen Leuchttürmen. Ardea, 28:61-79, December (not seen; see Auk, 1940:271).
Drost, R.
1935. Ueber das Zahlenverhältnis von Alter und Geschlecht auf dem Herbst-und Frühjahrszuge. Vogelzug, 6(4):177-182, October.
Dwight, J., Jr.
1900. The sequence of plumages and moults of the passerine birds of New York. Annals New York Acad. Sci., 13(1):73-360, October 19.
Ganier, A. F.
1949. The late summer Dickcissel departure. Migrant, 20(3):52-53, September.
Gätke, H.
1895. Heligoland as an ornithological observatory. Translated by Rudolph Rosenstock. Edinburgh, David Douglas. Pp. X + II + 599 + 11 II. (advt.).
Gross, A. O.
1921. The Dickcissel (Spiza americana) of the Illinois prairies. Auk, 38(1):1-26, January 18 (first of two parts).
Gurney, J. H.
1923. Bird migration as observed on the east coast of England. Ibis, 11th ser., 5(4):573-603, October 3.
Howell, J. C., Laskey, A. R., and J. T. Tanner
1954. Bird mortality at airport ceilometers. Wilson Bull., 66(3):207-215, September [published October 29].
Howell, J. C., and J. T. Tanner
1951. An accident to migrating birds at the Knoxville airport. Migrant, 22(4):61-62, December.
Laskey, A. R.
1951. Another disaster to migrating birds at the Nashville airport. Migrant, 22(4):57-60, December.
Long, W. S.
1934. Notes from eastern Kansas. Auk, 51(2):255, April 4.
1940. Check-list of Kansas birds. Trans. Kansas Acad. Sci., 43:433-456.
Lovell, H. B.
1952. Catastrophe to birds at a Louisville airport. Kentucky Warbler, 28(1):5-6, February.
Lowery, G. H., Jr.
1951. A quantitative study of the nocturnal migration of birds. Univ. Kansas Publ., Mus. Nat. Hist., 3(2):361-472, June 29.
McCabe, T. T.
1943. An aspect of collectors' technique. Auk, 60(4):550-558, October 7.
McCabe, T. T., and A. H. Miller
1933. Geographic variation in the Northern Water-thrushes. Condor, 35(5):192-197, September-October (September 15).
Meinertzhagen, R.
1930. Nicoll's birds of Egypt. Vol. I (of 2). London, Hugh Rees Ltd. Pp. XVI + 348.
Mengel, R. M.
1952. Certain molts and plumages of Acadian and Yellow-bellied flycatchers. Auk, 69(3):273-283, July 7.
Miller, A. H.
1941. Speciation in the avian genus Junco. Univ. California Publ. Zoöl., 44(3):173-434, May 24.
Miller, A. H., and T. T. McCabe
1935. Racial differentiation in Passerella (Melospiza) lincolnii. Condor, 37(3):144-160, May-June (May 15).
Palmén, J. A.
1876. Ueber die Zugstrassen der Vögel. Leipzig, Wilhelm Engelmann. Pp. VI + 292 + I.
Peters, J. L., and L. Griscom
1938. Geographical variation in the Savannah Sparrow. Bull. Mus. Comp. Zoöl., 80(13):445-478, January.
Pitelka, F. A.
1946. Age in relation to migration in the Blue Jay. Auk, 63(1):82-84, January 25.
1951. Speciation and ecologic distribution in American jays of the genus Aphelocoma. Univ. California Publ. Zoöl., 50(3):195-464, July 20.
Rintoul, L. J., and E. V. Baxter
1914. Notes on some passerine birds found migrating in moult. Scottish Naturalist, no. 35, pp. 245-252, November.
Rowan, W.
1926. Notes on Alberta waders included on the British list. Part II. Brit. Birds, 20(2):34-42, June 1.
Ruth, E. L.
1952. The Bay-breasted Warbler in Kansas. Kansas Orn. Soc. Bull., 3(3):18-19.
Sibley, C. G.
1950. Species formation in the Red-eyed Towhees of Mexico. Univ. California Publ. Zoöl., 50(2):109-194, November 24.
Spofford, W. R.
1949. Mortality of birds at the ceilometer of the Nashville airport. Wilson Bull., 61(2):86-90, June.
Stone, W.
1906. Some light on night migration. Auk, 23(3):249-252, July.
Storer, R. W.
1951. Variation in the Painted Bunting (Passerina ciris), with special reference to wintering populations. Occas. Papers Mus. Zool., Univ. Michigan, no. 532, pp. 1-12, June 29.
Thomson, A. L.
1926. Problems of bird-migration. Boston and New York, Houghton Mifflin Company. Pp. XV + I + 350.
1936. Recent progress in the study of bird-migration: a review of the literature, 1926-35. Ibis, 13th ser., 6(3):472-530, July 1.
Ticehurst, C. B.
1916. Notes on migrants and moult, with special reference to the moults of some of our summer visitants. Scottish Naturalist, no. 50, pp. 29-38, February.
Tordoff, H. B.
1952. Notes on plumages, molts, and age variation of the Red Crossbill. Condor, 54(4):200-203, July-August.
Wiegold, H.
1926. Masse, Gewichte und Zug nach Alter und Geschlecht bei Helgoländer Zugvögeln. Wissenschaftliche Meeresuntersuchungen, Abt. Helgoland, Neue Folge, 15ter Band, Heft 3, No. 17, Lipsius & Tischer, Kiel und Leipzig, pp. 1-73.
Wolfson, A.
1954. Weight and fat deposition in relation to spring migration in transient White-throated Sparrows. Auk, 71(4):413-434.
Transmitted June 30, 1955.
Obvious printer's errors have been repaired, other inconsistent spellings have been kept, for example inconsistent use of hyphen (e.g. "age-classes" and "age classes") and diacritical mark (e.g. "Zool." and "Zoöl.")
End of the Project Gutenberg EBook of Studies of Birds Killed in Nocturnal
Migration, by Harrison B. Tordoff and Robert M. Mengel
*** END OF THIS PROJECT GUTENBERG EBOOK STUDIES OF BIRDS ***
***** This file should be named 52382-h.htm or 52382-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/5/2/3/8/52382/
Produced by Judith Wirawan, Chris Curnow, Joseph Cooper
and the Online Distributed Proofreading Team at
http://www.pgdp.net
Updated editions will replace the previous one--the old editions will
be renamed.
Creating the works from print editions not protected by U.S. copyright
law means that no one owns a United States copyright in these works,
so the Foundation (and you!) can copy and distribute it in the United
States without permission and without paying copyright
royalties. Special rules, set forth in the General Terms of Use part
of this license, apply to copying and distributing Project
Gutenberg-tm electronic works to protect the PROJECT GUTENBERG-tm
concept and trademark. Project Gutenberg is a registered trademark,
and may not be used if you charge for the eBooks, unless you receive
specific permission. If you do not charge anything for copies of this
eBook, complying with the rules is very easy. You may use this eBook
for nearly any purpose such as creation of derivative works, reports,
performances and research. They may be modified and printed and given
away--you may do practically ANYTHING in the United States with eBooks
not protected by U.S. copyright law. Redistribution is subject to the
trademark license, especially commercial redistribution.
START: FULL LICENSE
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full
Project Gutenberg-tm License available with this file or online at
www.gutenberg.org/license.
Section 1. General Terms of Use and Redistributing Project
Gutenberg-tm electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or
destroy all copies of Project Gutenberg-tm electronic works in your
possession. If you paid a fee for obtaining a copy of or access to a
Project Gutenberg-tm electronic work and you do not agree to be bound
by the terms of this agreement, you may obtain a refund from the
person or entity to whom you paid the fee as set forth in paragraph
1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this
agreement and help preserve free future access to Project Gutenberg-tm
electronic works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the
Foundation" or PGLAF), owns a compilation copyright in the collection
of Project Gutenberg-tm electronic works. Nearly all the individual
works in the collection are in the public domain in the United
States. If an individual work is unprotected by copyright law in the
United States and you are located in the United States, we do not
claim a right to prevent you from copying, distributing, performing,
displaying or creating derivative works based on the work as long as
all references to Project Gutenberg are removed. Of course, we hope
that you will support the Project Gutenberg-tm mission of promoting
free access to electronic works by freely sharing Project Gutenberg-tm
works in compliance with the terms of this agreement for keeping the
Project Gutenberg-tm name associated with the work. You can easily
comply with the terms of this agreement by keeping this work in the
same format with its attached full Project Gutenberg-tm License when
you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are
in a constant state of change. If you are outside the United States,
check the laws of your country in addition to the terms of this
agreement before downloading, copying, displaying, performing,
distributing or creating derivative works based on this work or any
other Project Gutenberg-tm work. The Foundation makes no
representations concerning the copyright status of any work in any
country outside the United States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other
immediate access to, the full Project Gutenberg-tm License must appear
prominently whenever any copy of a Project Gutenberg-tm work (any work
on which the phrase "Project Gutenberg" appears, or with which the
phrase "Project Gutenberg" is associated) is accessed, displayed,
performed, viewed, copied or distributed:
This eBook is for the use of anyone anywhere in the United States and
most other parts of the world at no cost and with almost no
restrictions whatsoever. You may copy it, give it away or re-use it
under the terms of the Project Gutenberg License included with this
eBook or online at www.gutenberg.org. If you are not located in the
United States, you'll have to check the laws of the country where you
are located before using this ebook.
1.E.2. If an individual Project Gutenberg-tm electronic work is
derived from texts not protected by U.S. copyright law (does not
contain a notice indicating that it is posted with permission of the
copyright holder), the work can be copied and distributed to anyone in
the United States without paying any fees or charges. If you are
redistributing or providing access to a work with the phrase "Project
Gutenberg" associated with or appearing on the work, you must comply
either with the requirements of paragraphs 1.E.1 through 1.E.7 or
obtain permission for the use of the work and the Project Gutenberg-tm
trademark as set forth in paragraphs 1.E.8 or 1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any
additional terms imposed by the copyright holder. Additional terms
will be linked to the Project Gutenberg-tm License for all works
posted with the permission of the copyright holder found at the
beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including
any word processing or hypertext form. However, if you provide access
to or distribute copies of a Project Gutenberg-tm work in a format
other than "Plain Vanilla ASCII" or other format used in the official
version posted on the official Project Gutenberg-tm web site
(www.gutenberg.org), you must, at no additional cost, fee or expense
to the user, provide a copy, a means of exporting a copy, or a means
of obtaining a copy upon request, of the work in its original "Plain
Vanilla ASCII" or other form. Any alternate format must include the
full Project Gutenberg-tm License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works
provided that
* You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is owed
to the owner of the Project Gutenberg-tm trademark, but he has
agreed to donate royalties under this paragraph to the Project
Gutenberg Literary Archive Foundation. Royalty payments must be paid
within 60 days following each date on which you prepare (or are
legally required to prepare) your periodic tax returns. Royalty
payments should be clearly marked as such and sent to the Project
Gutenberg Literary Archive Foundation at the address specified in
Section 4, "Information about donations to the Project Gutenberg
Literary Archive Foundation."
* You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or destroy all
copies of the works possessed in a physical medium and discontinue
all use of and all access to other copies of Project Gutenberg-tm
works.
* You provide, in accordance with paragraph 1.F.3, a full refund of
any money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days of
receipt of the work.
* You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project
Gutenberg-tm electronic work or group of works on different terms than
are set forth in this agreement, you must obtain permission in writing
from both the Project Gutenberg Literary Archive Foundation and The
Project Gutenberg Trademark LLC, the owner of the Project Gutenberg-tm
trademark. Contact the Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
works not protected by U.S. copyright law in creating the Project
Gutenberg-tm collection. Despite these efforts, Project Gutenberg-tm
electronic works, and the medium on which they may be stored, may
contain "Defects," such as, but not limited to, incomplete, inaccurate
or corrupt data, transcription errors, a copyright or other
intellectual property infringement, a defective or damaged disk or
other medium, a computer virus, or computer codes that damage or
cannot be read by your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium
with your written explanation. The person or entity that provided you
with the defective work may elect to provide a replacement copy in
lieu of a refund. If you received the work electronically, the person
or entity providing it to you may choose to give you a second
opportunity to receive the work electronically in lieu of a refund. If
the second copy is also defective, you may demand a refund in writing
without further opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS', WITH NO
OTHER WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT
LIMITED TO WARRANTIES OF MERCHANTABILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of
damages. If any disclaimer or limitation set forth in this agreement
violates the law of the state applicable to this agreement, the
agreement shall be interpreted to make the maximum disclaimer or
limitation permitted by the applicable state law. The invalidity or
unenforceability of any provision of this agreement shall not void the
remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in
accordance with this agreement, and any volunteers associated with the
production, promotion and distribution of Project Gutenberg-tm
electronic works, harmless from all liability, costs and expenses,
including legal fees, that arise directly or indirectly from any of
the following which you do or cause to occur: (a) distribution of this
or any Project Gutenberg-tm work, (b) alteration, modification, or
additions or deletions to any Project Gutenberg-tm work, and (c) any
Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of
computers including obsolete, old, middle-aged and new computers. It
exists because of the efforts of hundreds of volunteers and donations
from people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future
generations. To learn more about the Project Gutenberg Literary
Archive Foundation and how your efforts and donations can help, see
Sections 3 and 4 and the Foundation information page at
www.gutenberg.org Section 3. Information about the Project Gutenberg
Literary Archive Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg Literary
Archive Foundation are tax deductible to the full extent permitted by
U.S. federal laws and your state's laws.
The Foundation's principal office is in Fairbanks, Alaska, with the
mailing address: PO Box 750175, Fairbanks, AK 99775, but its
volunteers and employees are scattered throughout numerous
locations. Its business office is located at 809 North 1500 West, Salt
Lake City, UT 84116, (801) 596-1887. Email contact links and up to
date contact information can be found at the Foundation's web site and
official page at www.gutenberg.org/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
[email protected]
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To SEND
DONATIONS or determine the status of compliance for any particular
state visit www.gutenberg.org/donate
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations. To
donate, please visit: www.gutenberg.org/donate
Section 5. General Information About Project Gutenberg-tm electronic works.
Professor Michael S. Hart was the originator of the Project
Gutenberg-tm concept of a library of electronic works that could be
freely shared with anyone. For forty years, he produced and
distributed Project Gutenberg-tm eBooks with only a loose network of
volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as not protected by copyright in
the U.S. unless a copyright notice is included. Thus, we do not
necessarily keep eBooks in compliance with any particular paper
edition.
Most people start at our Web site which has the main PG search
facility: www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.