The Project Gutenberg EBook of The Geological History of Plants, by Sir J. William Dawson This eBook is for the use of anyone anywhere in the United States and most other parts of the world at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org. If you are not located in the United States, you'll have to check the laws of the country where you are located before using this ebook. Title: The Geological History of Plants Author: Sir J. William Dawson Release Date: January 23, 2016 [EBook #51021] Language: English Character set encoding: ISO-8859-1 *** START OF THIS PROJECT GUTENBERG EBOOK THE GEOLOGICAL HISTORY OF PLANTS *** Produced by Tom Cosmas compiled from files made available at The Internet Archive.
THE INTERNATIONAL SCIENTIFIC SERIES
VOLUME LXI
THE
INTERNATIONAL SCIENTIFIC SERIES.
Each book complete in One Volume, 12mo, and bound in Cloth.
| 1. | FORMS OF WATER: A Familiar Exposition of the Origin and Phenomena of Glaciers. By J. Tyndall, LL. D., F. R. S. With 25 Illustrations. $1.50. |
| 2. | PHYSICS AND POLITICS; Or, Thoughts on the Application of the Principles of “Natural Selection” and “Inheritance” to Political Society. By Walter Bagehot. $1.50. |
| 3. | FOODS. By Edward Smith, M. D., LL. B., F. R. S. With numerous Illustrations. $1.75. |
| 4. | MIND AND BODY: The Theories of their Relation. By Alexander Bain, LL. D. With 4 Illustrations. $1.50. |
| 5. | THE STUDY OF SOCIOLOGY. By Herbert Spencer. $1.50. |
| 6. | THE NEW CHEMISTRY. By Professor J. P. Cooke, of Harvard University. With 31 Illustrations. $2.00. |
| 7. | ON THE CONSERVATION OF ENERGY. By Balfour Stewart, M.A., LL. D., F. R. S. With 14 Illustrations. $1.50. |
| 8. | ANIMAL LOCOMOTION; or, Walking, Swimming, and Flying. By J. B. Pettigrew, M. D., F. R. S., etc. With 130 Illustrations. $1.75. |
| 9. | RESPONSIBILITY IN MENTAL DISEASE. By Henry Maudsley, M.D. $1.50. |
| 10. | THE SCIENCE OF LAW. By Professor Sheldon Amos. $1.75. |
| 11. | ANIMAL MECHANISM: A Treatise on Terrestrial and Aërial Locomotion, By Professor E. J. Marey. With 117 Illustrations. $1 75. |
| 12. | THE HISTORY OF THE CONFLICT BETWEEN RELIGION AND SCIENCE. By J. W. Draper, M. D., LL. D. $1.75. |
| 13. | THE DOCTRINE OF DESCENT AND DARWINISM. By Professor Oscar Schmidt (Strasburg University). With 26 Illustrations. $1.50. |
| 14. | THE CHEMICAL EFFECTS OF LIGHT AND PHOTOGRAPHY. By Dr. Hermann Vogel (Polytechnic Academy of Berlin). Translation thoroughly revised. With 100 Illustrations. $2.00. |
| 15. | FUNGI: Their Nature, Influences, Uses, etc. By M. C. Cooke, M. A., LL. D. Edited by the Rev. M. J. Berkeley, M. A., F.L. S. With 109 Illustrations. $1.50. |
| 16. | THE LIFE AND GROWTH OF LANGUAGE. By Professor William Dwight Whitney, of Yale College. $1.50. |
| 17. | MONEY AND THE MECHANISM OF EXCHANGE. By W. Stanley Jevons, M. A., F. R. S. $1.75. « 2 » |
| 18. | THE NATURE OF LIGHT, with a General Account of Physical Optics. By Dr. Eugene Lommel. With 188 Illustrations and a Table of Spectra in Chromo-lithography. $2.00. |
| 19. | ANIMAL PARASITES AND MESSMATES. By Monsieur Van Beneden. With 83 Illustrations. $1.50. |
| 20. | FERMENTATION. By Professor Schützenberger. With 28 Illustrations. $1.50. |
| 21. | THE FIVE SENSES OF MAN. By Professor Bernstein. With 91 Illustrations. $1.75. |
| 22. | THE THEORY OF SOUND IN ITS RELATION TO MUSIC. By Professor Pietro Blaserna. With numerous Illustrations. $1.50. |
| 23. | STUDIES IN SPECTRUM ANALYSIS. By J. Norman Lockyer, F. R. S. With 6 Photographic Illustrations of Spectra, and numerous Engravings on Wood. $2.50. |
| 24. | A HISTORY OF THE GROWTH OF THE STEAM-ENGINE. By Professor R. H. Thurston. With 163 Illustrations. $2.50. |
| 25. | EDUCATION AS A SCIENCE. By Alexander Bain, LL.D. $1.75. |
| 26. | STUDENTS' TEXT-BOOK OF COLOR; Or, Modern Chromatics. With Applications to Art and Industry. By Professor Ogden N. Rood, Columbia College. New edition. With 130 Illustrations. $2.00. |
| 27. | THE HUMAN SPECIES. By Professor A. de Quatrefages, Membre de l’Institut. $2.00. |
| 28. | THE CRAYFISH: An Introduction to the Study of Zoölogy. By T. H. Huxley, F. R. S. With 82 Illustrations. $1.75. |
| 29. | THE ATOMIC THEORY. By Professor A. Wurtz. Translated by E. Cleminshaw, F. C. S. $1.50. |
| 30. | ANIMAL LIFE AS AFFECTED BY THE NATURAL CONDITIONS OF EXISTENCE. By Karl Semper. With 2 Maps and 106 Woodcuts. $2.00. |
| 31. | SIGHT: An Exposition of the Principles of Monocular and Binocular Vision. By Joseph Le Conte, LL. D. With 132 Illustrations. $1.50. |
| 32. | GENERAL PHYSIOLOGY OF MUSCLES AND NERVES. By Professor J. Rosenthal. With 75 Illustrations. $1.50. |
| 33. | ILLUSIONS: A Psychological Study. By James Sully. $1.50. |
| 34. | THE SUN. By C. A. Young, Professor of Astronomy in the College of New Jersey. With numerous Illustrations. $2.00. |
| 35. | VOLCANOES: What they Are and what they Teach. By John W. Judd, F.R. S., Professor of Geology in the Royal School of Mines. With 96 Illustrations. $2.00. « 3 » |
| 36. | SUICIDE: An Essay in Comparative Moral Statistics. By Henry Morselli, M.D., Professor of Psychological Medicine, Royal University, Turin. $1.75. |
| 37. | THE FORMATION OF VEGETABLE MOULD, THROUGH THE ACTION OF WORMS. With Observations on their Habits. By Charles Darwin, LL. D., F. R. S. With Illustrations. $1.50. |
| 38. | THE CONCEPTS AND THEORIES OF MODERN PHYSICS. By J. B. Stallo. $1.75. |
| 39. | THE BRAIN AND ITS FUNCTIONS. By J. Luys. $1.50. |
| 40. | MYTH AND SCIENCE. By Tito Vignoli. $1.50. |
| 41. | DISEASES OF MEMORY: An Essay in the Positive Psychology. By Th. Ribot, author of “Heredity.” $1.50. |
| 42. | ANTS, BEES, AND WASPS. A Record of Observations of the Habits of the Social Hymenoptera. By Sir John Lubbock, Bart., F. R. S., D. C. L., LL. D., etc. $2.00. |
| 43. | SCIENCE OF POLITICS. By Sheldon Amos. $1.75. |
| 44. | ANIMAL INTELLIGENCE. By George J. Romanes. $1.75. |
| 45. | MAN BEFORE METALS. By N. Joly, Correspondent of the Institute. With 148 Illustrations. $1.75. |
| 46. | THE ORGANS OF SPEECH AND THEIR APPLICATION IN THE FORMATION OF ARTICULATE SOUNDS. By G. H. von Meyer, Professor in Ordinary of Anatomy at the University of Zurich. With 47 Woodcuts. $1.75. |
| 47. | FALLACIES: A View of Logic from the Practical Side. By Alfred Sidgwick, B.A., Oxon. $1.75. |
| 48. | ORIGIN OF CULTIVATED PLANTS. By Alphonse de Candolle. $2.00. |
| 49. | JELLY-FISH, STAR-FISH, AND SEA-URCHINS. Being a Research on Primitive Nervous Systems. By George J. Romanes. $1.75. |
| 50. | THE COMMON SENSE OF THE EXACT SCIENCES. By the late William Kingdon Clifford. $1.50. |
| 51. | PHYSICAL EXPRESSION: Its Modes and Principles. By Francis Warner, M.D., Assistant Physician, and Lecturer on Botany to the London Hospital, etc. With 51 Illustrations. $1.75. |
| 52. | ANTHROPOID APES. By Robert Hartmann, Professor in the University of Berlin. With 63 Illustrations. $1.75. |
| 53. | THE MAMMALIA IN THEIR RELATION TO PRIMEVAL TIMES. By Oscar Schmidt. $1.50. « 4 » |
| 54. | COMPARATIVE LITERATURE. By Hutcheson Macaulay Posnett, M. A., LL. D., F. L. S., Barrister-at-Law; Professor of Classics and English Literature, University College, Auckland, New Zealand; author of “The Historical Method,” etc. $1.75. |
| 55. | EARTHQUAKES AND OTHER EARTH MOVEMENTS. By John Milne, Professor of Mining and Geology in the Imperial College of Engineering, Tokio, Japan. With 38 Figures. $1.75. |
| 56. | MICROBES, FERMENTS, AND MOULDS. By E. L. Trouessart. With 107 Illustrations. $1.50. |
| 57. | THE GEOGRAPHICAL AND GEOLOGICAL DISTRIBUTION OF ANIMALS. By Angelo Heilprin. $2.00. |
| 58. | WEATHER. A Popular Exposition of the Nature of Weather Changes from Day to Day. By Hon. Ralph Abercromby. $1.75. |
| 59. | ANIMAL MAGNETISM. By Alfred Binet and Charles Féré, Assistant Physician at the Salpêtrière. |
THE INTERNATIONAL SCIENTIFIC SERIES
THE
GEOLOGICAL HISTORY
OF PLANTS
BY
SIR J. WILLIAM DAWSON
C. M. G., LL. D., F. R. S., &c.
WITH ILLUSTRATIONS
NEW YORK
APPLETON AND COMPANY
1888
Copyright, 1888,
By D. APPLETON AND COMPANY.
PREFACE.
The object of this work is to give, in a connected form, a summary of the development of the vegetable kingdom in geological time.
To the geologist and botanist the subject is one of importance with reference to their special pursuits, and one on which it has not been easy to find any convenient manual of information. It is hoped that its treatment in the present volume will also be found sufficiently simple and popular to be attractive to the general reader.
In a work of so limited dimensions, detailed descriptions cannot be given, except occasionally by way of illustration; but references to authorities will be made in foot-notes, and certain details, which may be useful to collectors and students, will be placed in notes appended to the chapters, so as not to encumber the text.
The illustrations of this work are for the most part original; but some of them have previously appeared in special papers of the author.
J. W. D.
February, 1888.
CONTENTS.
| PAGE | ||
| CHAPTER I. | ||
| Preliminary Ideas of Geological Chronology and of the Classification of Plants | 1 | |
| CHAPTER II. | ||
| Vegetation of the Laurentian and Early Paleozoic—Questions as to Algæ | 8 | |
| CHAPTER III. | ||
| The Erian or Devonian Forests—Origin of Petroleum—The Age of Acrogens and Gymnosperms | 45 | |
| CHAPTER IV. | ||
| The Carboniferous Flora—Culmination of the Acrogens—Formation of Coal | 110 | |
| CHAPTER V. | ||
| The Flora of the Early Mesozoic—Reign of Pines and Cycads | 175 | |
| CHAPTER VI. | ||
| The Reign of Angiosperms in the Later Cretaceous and Early Tertiary or Kainozoic | 191 | |
| CHAPTER VII. « viii » | ||
| Plants from the Tertiary to the Modern Period | 219 | |
| CHAPTER VIII. | ||
| General Laws of Origin and Migrations of Plants—Relations of Recent and Fossil Floras | 237 | |
| APPENDIX. | ||
| I. | Comparative View of Paleozoic Floras | 273 |
| II. | Heer’s Latest Statements on the Greenland Flora | 281 |
| III. | Mineralisation of Fossil Plants | 284 |
| IV. | General Works on Palæobotany | 286 |
LIST OF ILLUSTRATIONS.
| PAGE | |
| Table of Chronology of Plants | (Frontispiece.) |
| Protannularia Harknessii | 21 |
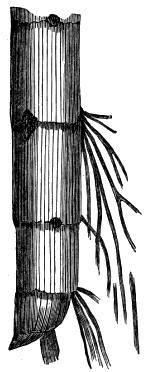
| Nematophyton Logani (three Figures) | 22, 23 |
| Trail of King-Crab | 28 |
| Trail of Carboniferous Crustacean | 28 |
| Rusichnites | 29 |
| Palæophycus | 30 |
| Astropolithon | 31 |
| Carboniferous Rill-mark | 33 |
| Cast of Shrinkage Cracks | 34 |
| Cone-in-cone | 36 |
| Buthotrephis | 37 |
| Silurian Vegetation | 40 |
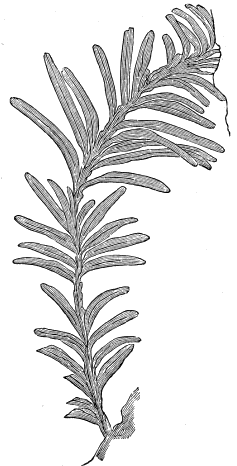
| Erian Plants | 49 |
| Protosalvinia | 54 |
| Ptilophyton (two Figures) | 62, 63 |
| Psilophyton (two Figures) | 64, 66 |
| Sphenophyllum | 65 |
| Lepidodendron | 66 |
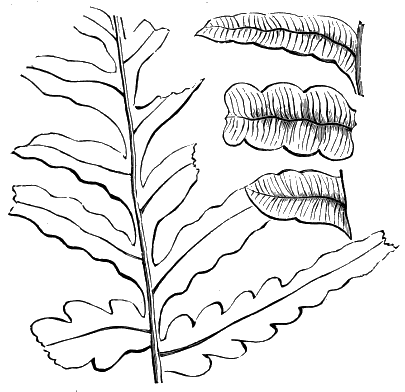
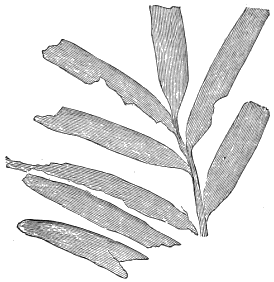
| Various Ferns | 72, 73 |
| Archæopteris | 74 |
| Caulopteris | 75 |
| Megalopteris | 76 |
| Calamites | 77 |
| Asterophyllites | 78 |
| Dadoxylon | 79 « x » |
| Cordaites | 81 |
| Erian Fruits | 82 |
| Foliage from the Coal-formation | 111 |
| Sigillariæ (five Figures) | 112-114 |
| Stigmariæ (two Figures) | 115 |
| Vegetable Tissues | 117 |
| Coals and Erect Trees (two Figures) | 118, 119 |
| Lepidodendron | 120 |
| Lepidophloios | 121 |
| Asterophyllites, &c. | 122 |
| Calamites (five Figures) | 123-125 |
| Ferns of the Coal-formation (six Figures) | 126-129 |
| Noeggerathia dispar | 130 |
| Cordaites | 131 |
| Fruits of Cordaites, &c. | 132 |
| Conifers of the Coal-formation (four Species) | 135 |
| Trigonocarpum | 136 |
| Sternbergia | 137 |
| Walchia imbricatula | 138 |
| Foliage of the Jurassic Period | 177 |
| Podozamites | 178 |
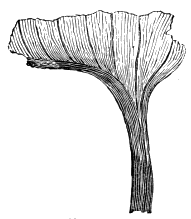
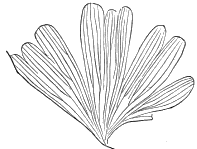
| Salisburia | 180 |
| Sequoia | 181 |
| Populus primæva | 191 |
| Stercalia and Laurophyllum | 194 |
| Vegetation of the Cretaceous Period | 195 |
| Platanus | 198 |
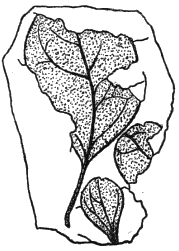
| Protophyllum | 199 |
| Magnolia | 200 |
| Liriodendron (two Figures) | 201 |
| Brasenia | 207 |
| Gaylussaccia resinosa | 228 |
| Populus balsamifera | 229 |
| Fucus | 230 |
THE
GEOLOGICAL HISTORY OF PLANTS.
PRELIMINARY IDEAS OF GEOLOGICAL CHRONOLOGY AND OF THE CLASSIFICATION OF PLANTS.
The knowledge of fossil plants and of the history of the vegetable kingdom has, until recently, been so fragmentary that it seemed hopeless to attempt a detailed treatment of the subject of this little book. Our stores of knowledge have, however, been rapidly accumulating in recent years, and we have now arrived at a stage when every new discovery serves to render useful and intelligible a vast number of facts previously fragmentary and of uncertain import.
The writer of this work, born in a district rich in fossil plants, began to collect and work at these as a boy, in connection with botanical and geological pursuits. He has thus been engaged in the study of fossil plants for nearly half a century, and, while he has published much on the subject, has endeavoured carefully to keep within the sphere of ascertained facts, and has made it a specialty to collect, as far as possible, what has been published by others. He has also enjoyed opportunities of correspondence or personal intercourse with most of « 2 » the more eminent workers in the subject. Now, in the evening of his days, he thinks it right to endeavour to place before the world a summary of facts and of his own matured conclusions—feeling, however, that nothing can be final in this matter; and that he can only hope to sketch the present aspect of the subject, and to point the way to new developments, which must go on long after he shall have passed away.
The subject is one which has the disadvantage of presupposing some knowledge of the geological history of the earth, and of the classification and structures of modern plants; and in order that all who may please to read the following pages may be placed, as nearly as possible, on the same level, this introductory chapter will be devoted to a short statement of the general facts of geological chronology, and of the natural divisions of the vegetable kingdom in their relations to that chronology.
The crust of the earth, as we somewhat modestly term that portion of its outer shell which is open to our observation, consists of many beds of rock superimposed on each other, and which must have been deposited successively, beginning with the lowest. This is proved by the structure of the beds themselves, by the markings on their surfaces, and by the remains of animals and plants which they contain; all these appearances indicating that each successive bed must have been the surface before it was covered by the next.
As these beds of rock were mostly formed under water, and of material derived from the waste of land, they are not universal, but occur in those places where there were extensive areas of water receiving detritus from the land. Further, as the distinction of land and water arises primarily from the shrinkage of the mass of the earth, and from the consequent collapse of the crust in some places and ridging of it up in others, it follows that there have, from the earliest geological periods, been deep ocean-basins, « 3 » ridges of elevated land, and broad plateaus intervening between the ridges, and which were at some times under water, and at other times land, with many intermediate phases. The settlement and crumpling of the crust were not continuous, but took place at intervals; and each such settlement produced not only a ridging up along certain lines, but also an emergence of the plains or plateaus. Thus at all times there have been ridges of folded rock constituting mountain-ranges, flat expansions of continental plateau, sometimes dry and sometimes submerged, and deep ocean-basins, never except in some of their shallower portions elevated into land.
By the study of the successive beds, more especially of those deposited in the times of continental submergence, we obtain a table of geological chronology which expresses the several stages of the formation of the earth’s crust, from that early time when a solid shell first formed on our nascent planet to the present day. By collecting the fossil remains embedded in the several layers and placing these in chronological order, we obtain in like manner histories of animal and plant life parallel to the physical changes indicated by the beds themselves. The facts as to the sequence we obtain from the study of exposures in cliffs, cuttings, quarries, and mines; and by correlating these local sections in a great number of places, we obtain our general table of succession; though it is to be observed that in some single exposures or series of exposures, like those in the great canons of Colorado, or on the coasts of Great Britain, we can often in one locality see nearly the whole sequence of beds. Let us observe here also that, though we can trace these series of deposits over the whole of the surfaces of the continents, yet if the series could be seen in one spot, say in one shaft sunk through the whole thickness of the earth’s crust, this would be sufficient for our purpose, so far as the history of life is concerned.
The evidence is similar to that obtained by Schliemann on the site of Troy, where, in digging through successive layers of débris, he found the objects deposited by successive occupants of the site, from the time of the Roman Empire back to the earliest tribes, whose flint weapons and the ashes of their fires rest on the original surface of the ground.
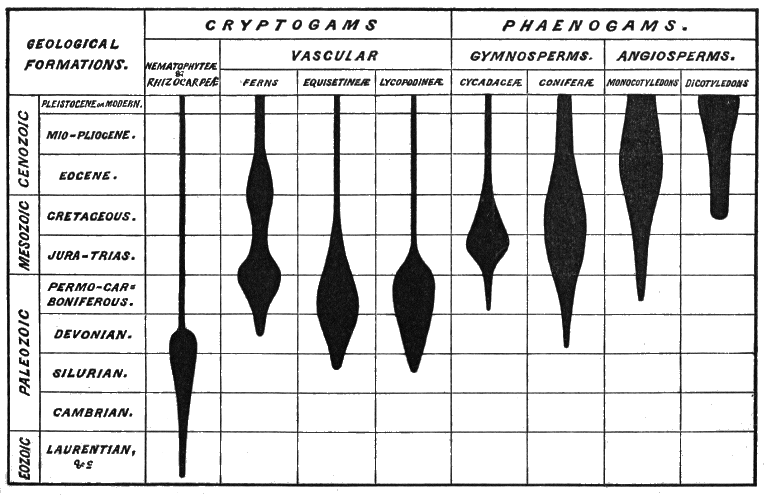
Let us now tabulate the whole geological succession with the history of animals and plants associated with it:
| ANIMALS. | SYSTEMS OF FORMATIONS. | PLANTS. | ||
| Age of Man and Mammalia. | Kainozoic. | Modern, Pleistocene, Pliocene, Miocene, Eocene. |
Angiosperms and Palms dominant. |
|
| Age of Reptiles. | Mesozoic. | Cretaceous, Jurassic, Triassic. |
Cycads and Pines dominant. |
|
| Age of Amphibians and Fishes. Age of Invertebrates. |
Palæozoic. | Permian, Carboniferous, Erian, Silurian, Ordovician, Cambrian, Huronian (Upper). |
Acrogens and Gymnosperms dominant. |
|
| Age of Protozoa. | Eozoic. | Huronian (Lower), Upper Laurentian, Middle Laurentian, Lower Laurentian. |
Protogens and Algæ. | |
It will be observed, since only the latest of the systems of formations in this table belongs to the period of human history, that the whole lapse of time embraced in the table must be enormous. If we suppose the modern period to have continued for say ten thousand years, and each of the others to have been equal to it, we shall require two hundred thousand years for the whole. There is, however, reason to believe, from the great thickness of the formations and the slowness of the deposition of many « 5 » of them in the older systems, that they must have required vastly greater time. Taking these criteria into account, it has been estimated that the time-ratios for the first three great ages may be as one for the Kainozoic to three for the Mesozoic and twelve for the Palæozoic, with as much for the Eozoic as for the Palæozoic. This is Dana’s estimate. Another, by Hull and Houghton, gives the following ratios: Azoic, 34·3 per cent.; Palæozoic, 42·5 per cent.; Mesozoic and Kainozoic, 23·2 per cent. It is further held that the modern period is much shorter than the other periods of the Kainozoic, so that our geological table may have to be measured by millions of years instead of thousands.
We cannot, however, attach any certain and definite value in years to geological time, but must content ourselves with the general statement that it has been vastly long in comparison to that covered by human history.
Bearing in mind this great duration of geological time, and the fact that it probably extends from a period when the earth was intensely heated, its crust thin, and its continents as yet unformed, it will be evident that the conditions of life in the earlier geologic periods may have been very different from those which obtained later. When we further take into account the vicissitudes of land and water which have occurred, we shall see that such changes must have produced very great differences of climate. The warm equatorial waters have in all periods, as superficial oceanic currents, been main agents in the diffusion of heat over the surface of the earth, and their distribution to north and south must have been determined mainly by the extent and direction of land, though it may also have been modified by the changes in the astronomical relations and period of the earth, and the form of its orbit.[A] We know by the evidence of « 6 » fossil plants that changes of this kind have occurred so great as, on the one hand, to permit the plants of warm temperate regions to exist within the Arctic Circle; and, on the other, to drive these plants into the tropics and to replace them by Arctic forms. It is evident also that in those periods when the continental areas were largely submerged, there might be an excessive amount of moisture in the atmosphere, greatly modifying the climate, in so far as plants are concerned.
[A] Croll, “Climate and Time.”
Let us now consider the history of the vegetable kingdom as indicated in the few notes in the right-hand column of the table.
The most general subdivision of plants is into the two great series of Cryptogams, or those which have no manifest flowers, and produce minute spores instead of seeds; and Phænogams, or those which possess flowers and produce seeds containing an embryo of the future plant.
The Cryptogams may be subdivided into the following three groups:
1. Thallogens, cellular plants not distinctly distinguishable into stem and leaf. These are the Fungi, the Lichens, and the Algæ, or sea-weeds.
2. Anogens, having stem and foliage, but wholly cellular. These are the Mosses and Liverworts.
3. Acrogens, which have long tubular fibres as well as cells in their composition, and thus have the capacity of attaining a more considerable magnitude. These are the Ferns (Filices), the Mare’s-tails (Equisetaceæ), and the Club-mosses (Lycopodiaceæ), and a curious little group of aquatic plants called Rhizocarps (Rhizocarpeæ).
The Phænogams are all vascular, but they differ much in the simplicity or complexity of their flowers or seeds. On this ground they admit of a twofold division:
1. Gymnosperms, or those which bear naked seeds not enclosed in fruits. They are the Pines and their allies, and the Cycads.
2. Angiosperms, which produce true fruits enclosing the seeds. In this group there are two well-marked subdivisions differing in the structure of the seed and stem. They are the Endogens, or inside growers, with seeds having one seed-leaf only, as the grasses and the palms; and the Exogens, having outside-growing woody stems, and seeds with two seed-leaves. Most of the ordinary forest-trees of temperate climates belong to this group.
On referring to the geological table, it will be seen that there is a certain rough correspondence between the order of rank of plants and the order of their appearance in time. The oldest plants that we certainly know are Algæ, and with these there are plants apparently with the structures of Thallophytes but the habit of trees, and which, for want of a better name, I may call Protogens. Plants akin to the Rhizocarps also appear very early. Next in order we find forests in which gigantic Ferns and Lycopods and Mare’s-tails predominate, and are associated with pines. Succeeding these we have a reign of Gymnosperms, and in the later formations we find the higher Phænogams dominant. Thus there is an advance in elevation and complexity along with the advance in geological time, but connected with the remarkable fact that in earlier times low groups attain to an elevation unexampled in later times, when their places are occupied with plants of higher type.
It is this historical development that we have to trace in the following pages, and it will be the most simple and at the same time the most instructive method to consider it in the order of time.
VEGETATION OF THE LAURENTIAN AND EARLY PALÆOZOIC—QUESTIONS AS TO ALGÆ.
Oldest of all the formations known to geologists, and representing perhaps the earliest rocks produced after our earth had ceased to be a molten mass, are the hard, crystalline, and much-contorted rocks named by the late Sir W. E. Logan Laurentian, and which are largely developed in the northern parts of North America and Europe, and in many other regions. So numerous and extensive, indeed, are the exposures of these rocks, that we have good reason to believe that they underlie all the other formations of our continents, and are even world-wide in their distribution. In the lower part of this great system of rocks which, in some places at least, is thirty thousand feet in thickness, we find no traces of the existence of any living thing on the earth. But, in the middle portion of the Laurentian, rocks are found which indicate that there were already land and water, and that the waters and possibly the land were already tenanted by living beings. The great beds of limestone which exist in this part of the system furnish one indication of this. In the later geological formations the limestones are mostly organic—that is, they consist of accumulated remains of shells, corals, and other hard parts of marine animals, which are composed of calcium carbonate, which the animals obtain directly from their food, and indirectly from the calcareous matter dissolved in the sea-water. In like « 9 » manner great beds of iron-ore exist in the Laurentian; but in later formations the determining cause of the accumulation of such beds is the partial deoxidation and solution of the peroxide of iron by the agency of organic matter. Besides this, certain forms known as Eozoon Canadense have been recognised in the Laurentian limestones, which indicate the presence at least of one of the lower types of marine animals. Where animal life is, we may fairly infer the existence of vegetable life as well, since the plant is the only producer of food for the animal. But we are not left merely to this inference. Great quantities of carbon or charcoal in the form of the substance known as graphite or plumbago exist in the Laurentian. Now, in more recent formations we have deposits of coal and bituminous matter, and we know that these have arisen from the accumulation and slow putrefaction of masses of vegetable matter. Further, in places where igneous action has affected the beds, we find that ordinary coal has been changed into anthracite and graphite, that bituminous shales have been converted into graphitic shales, and that cracks filled with soft bituminous matter have ultimately become changed into veins of graphite. When, therefore, we find in the Laurentian thick beds of graphite and beds of limestone charged with detached grains and crystals of this substance, and graphitic gneisses and schists and veins of graphite traversing the beds, we recognise the same phenomena that are apparent in later formations containing vegetable débris.
The carbon thus occurring in the Laurentian is not to be regarded as exceptional or rare, but is widely distributed and of large amount. In Canada more especially the deposits are very considerable.
The graphite of the Laurentian of Canada occurs both in beds and in veins, and in such a manner as to show that its origin and deposition are contemporaneous with « 10 » those of the containing rock. Sir William Logan states[B] that “the deposits of plumbago generally occur in the limestones or in their immediate vicinity, and granular varieties of the rock often contain large crystalline plates of plumbago. At other times this mineral is so finely disseminated as to give a bluish-grey colour to the limestone, and the distribution of bands thus coloured seems to mark the stratification of the rock.” He further states: “The plumbago is not confined to the limestones; large crystalline scales of it are occasionally disseminated in pyroxene rock, and sometimes in quartzite and in feldspathic rocks, or even in magnetic oxide of iron.” In addition to these bedded forms, there are also true veins in which graphite occurs associated with calcite, quartz, orthoclase, or pyroxene, and either in disseminated scales, in detached masses, or in bands or layers “separated from each other and from the wall-rock by feldspar, pyroxene, and quartz.” Dr. Hunt also mentions the occurrence of finely granular varieties, and of that peculiarly waved and corrugated variety simulating fossil wood, though really a mere form of laminated structure, which also occurs at Warrensburg, New York, and at the Marinski mine in Siberia. Many of the veins are not true fissures, but rather constitute a network of shrinkage cracks or segregation veins traversing in countless numbers the containing rock, and most irregular in their dimensions, so that they often resemble strings of nodular masses. It is most probable that the graphite of the veins was originally introduced as a liquid or plastic hydrocarbon; but in whatever way introduced, the character of the veins indicates that in the case of the greater number of them the carbonaceous material must have been derived from the bedded rocks traversed by these veins, to which it bears the same relation with the veins « 11 » of bitumen found in the bituminous shales of the Carboniferous and Silurian rocks. Nor can there be any doubt that the graphite found in the beds has been deposited along with the calcareous matter or muddy and sandy sediment of which these beds were originally composed.[C]
[B] “Geology of Canada,” 1863.
[C] Paper by the author on Laurentian Graphite, “Journal of London Geological Society,” 1876.
The quantity of graphite in the Lower Laurentian series is enormous. Some years ago, in the township of Buckingham, on the Ottawa River, I examined a band of limestone believed to be a continuation of that described by Sir W. E. Logan as the Green Lake limestone. It was estimated to amount, with some thin interstratified bands of gneiss, to a thickness of six hundred feet or more, and was found to be filled with disseminated crystals of graphite and veins of the mineral to such an extent as to constitute in some places one-fourth of the whole; and, making every allowance for the poorer portions, this band cannot contain in all a less vertical thickness of pure graphite than from twenty to thirty feet. In the adjoining township of Lochaber Sir W. E. Logan notices a band from twenty-five to thirty feet thick, reticulated with graphite veins to such an extent as to be mined with profit for the mineral. At another place in the same district a bed of graphite from ten to twelve feet thick, and yielding 20 per cent, of the pure material, is worked. As it appears in the excavation made by the quarrymen, it resembled a bed of coal; and a block from this bed, about four feet thick, was a prominent object in the Canadian department of the Colonial Exhibition of 1886. When it is considered that graphite occurs in similar abundance at several other horizons, in beds of limestone which have been ascertained by Sir W. E. Logan to have an aggregate thickness of thirty-five hundred feet, it is « 12 » scarcely an exaggeration to maintain that the quantity of carbon in the Laurentian is equal to that in similar areas of the Carboniferous system. It is also to be observed that an immense area in Canada appears to be occupied by these graphitic and Eozoon limestones, and that rich graphitic deposits exist in the continuation of this system in the State of New York, while in rocks believed to be of this age near St. John, New Brunswick, there is a very thick bed of graphitic limestone, and associated with it three regular beds of graphite, having an aggregate thickness of about five feet.[D]
[D] Matthew in “Quarterly Journal of the Geological Society,” vol. xxi., p. 423. “Acadian Geology,” p. 662.
It may fairly be assumed that in the present world, and in those geological periods with whose organic remains we are more familiar than with those of the Laurentian, there is no other source of unoxidized carbon in rocks than that furnished by organic matter, and that this has obtained its carbon in all cases, in the first instance, from the deoxidation of carbonic acid by living plants. No other source of carbon can, I believe, be imagined in the Laurentian period. We may, however, suppose either that the graphitic matter of the Laurentian has been accumulated in beds like those of coal, or that it has consisted of diffused bituminous matter similar to that in more modern bituminous shales and bituminous and oil-bearing limestones. The beds of graphite near St. John, some of those in the gneiss at Ticonderoga in New York, and at Lochaber and Buckingham, and elsewhere in Canada, are so pure and regular that one might fairly compare them with the graphitic coal of Rhode Island. These instances, however, are exceptional, and the greater part of the disseminated and vein graphite might rather be likened in its mode of occurrence to the bituminous matter in bituminous shales and limestones.
We may compare the disseminated graphite to that which we find in those districts of Canada in which Silurian and Devonian bituminous shales and limestones have been metamorphosed and converted into graphitic rocks not very dissimilar to those in the less altered portions of the Laurentian.[E] In like manner it seems probable that the numerous reticulating veins of graphite may have been formed by the segregation of bituminous matter into fissures and planes of least resistance, in the manner in which such veins occur in modern bituminous limestones and shales. Such bituminous veins occur in the Lower Carboniferous limestone and shale of Dorchester and Hillsborough, New Brunswick, with an arrangement very similar to that of the veins of graphite; and in the Quebec rocks of Point Levi, veins attaining to a thickness of more than a foot, are filled with a coaly matter having a transverse columnar structure, and regarded by Logan and Hunt as an altered bitumen. These palæozoic analogies would lead us to infer that the larger part of the Laurentian graphite falls under the second class of deposits above mentioned, and that, if of vegetable origin, the organic matter must have been thoroughly disintegrated and bituminised before it was changed into graphite. This would also give a probability that the vegetation implied was aquatic, or at least that it was accumulated under water.
[E] Granby, Melbourne, Owl’s Head, &c., “Geology of Canada,” 1863, p. 599.
Dr. Hunt has, however, observed an indication of terrestrial vegetation, or at least of subaërial decay, in the great beds of Laurentian iron-ore. These, if formed in the same manner as more modern deposits of this kind, would imply the reducing and solvent action of substances produced in the decay of plants. In this case such great ore-beds as that of Hull, on the Ottawa, seventy « 14 » feet thick, or that near Newborough, two hundred feet thick,[F] must represent a corresponding quantity of vegetable matter which has totally disappeared. It may be added that similar demands on vegetable matter as a deoxidising agent are made by the beds and veins of metallic sulphides of the Laurentian, though some of the latter are no doubt of later date than the Laurentian rocks themselves.
[F] “Geology of Canada,” 1863.
It would be very desirable to confirm such conclusions as those above deduced by the evidence of actual microscopic structure. It is to be observed, however, that when, in more modern sediments, Algæ have been converted into bituminous matter, we cannot ordinarily obtain any structural evidence of the origin of such bitumen, and in the graphitic slates and limestones derived from the metamorphosis of such rocks no organic structure remains. It is true that, in certain bituminous shales and limestones of the Silurian system, shreds of organic tissue can sometimes be detected, and in some cases, as in the Lower Silurian limestone of the La Cloche Mountains in Canada, the pores of brachiopodous shells and the cells of corals have been penetrated by black bituminous matter, forming what may be regarded as natural injections, sometimes of much beauty. In correspondence with this, while in some Laurentian graphitic rocks, as, for instance, in the compact graphite of Clarendon, the carbon presents a curdled appearance due to segregation, and precisely similar to that of the bitumen in more modern bituminous rocks, I can detect in the graphitic limestones occasional fibrous structures which may be remains of plants, and in some specimens vermicular lines, which I believe to be tubes of Eozoon penetrated by matter once bituminous, but now in the state of graphite.
When palæozoic land-plants have been converted into graphite, they sometimes perfectly retain their structure. Mineral charcoal, with structure, exists in the graphitic coal of Rhode Island. The fronds of ferns, with their minutest veins perfect, are preserved in the Devonian shales of St. John, in the state of graphite; and in the same formation there are trunks of Conifers (Dadoxylon Ouangondianum) in which the material of the cell-walls has been converted into graphite, while their cavities have been filled with calcareous spar and quartz, the finest structures being preserved quite as well as in comparatively unaltered specimens from the coal-formation.[G] No structures so perfect have as yet been detected in the Laurentian, though in the largest of the three graphitic beds at St. John there appear to be fibrous structures, which I believe may indicate the existence of land-plants. This graphite is composed of contorted and slickensided laminæ, much like those of some bituminous shales and coarse coals; and in these are occasional small pyritous masses which show hollow carbonaceous fibres, in some cases presenting obscure indications of lateral pores. I regard these indications, however, as uncertain; and it is not as yet fully ascertained that these beds at St. John are on the same geological horizon with the Lower Laurentian of Canada, though they certainly underlie the Primordial series of the Acadian group, and are separated from it by beds having the character of the Huronian.
[G] “Acadian Geology,” p. 535. In calcined specimens the structures remain in the graphite after decalcification by an acid.
There is thus no absolute impossibility that distinct organic tissues may be found in the Laurentian graphite, if formed from land-plants, more especially if any plants existed at that time having true woody or vascular tissues; but it cannot with certainty be affirmed that such tissues « 16 » have been found. It is possible, however, that in the Laurentian period the vegetation of the land may have consisted wholly of cellular plants, as, for example, mosses and lichens; and if so, there would be comparatively little hope of the distinct preservation of their forms or tissues, or of our being able to distinguish the remains of land-plants from those of Algæ.
We may sum up these facts and considerations in the following statements: First, that somewhat obscure traces of organic structure can be detected in the Laurentian graphite; secondly, that the general arrangement and microscopic structure of the substance corresponds with that of the carbonaceous and bituminous matters in marine formations of more modern date; thirdly, that if the Laurentian graphite has been derived from vegetable matter, it has only undergone a metamorphosis similar in kind to that which organic matter in metamorphosed sediments of later age has experienced; fourthly, that the association of the graphitic matter with organic limestone, beds of iron-ore, and metallic sulphides greatly strengthens the probability of its vegetable origin; fifthly, that when we consider the immense thickness and extent of the Eozoonal and graphitic limestones and iron-ore deposits of the Laurentian, if we admit the organic origin of the limestone and graphite, we must be prepared to believe that the life of that early period, though it may have existed under low forms, was most copiously developed, and that it equalled, perhaps surpassed, in its results, in the way of geological accumulation, that of any subsequent period.
Many years ago, at the meeting of the American Association in Albany, the writer was carrying into the room of the Geological Section a mass of fossil wood from the Devonian of Gaspé, when he met the late Professor Agassiz, and remarked that the specimen was the remains of a Devonian tree contemporaneous with his « 17 » fishes of that age. “How I wish I could sit under its shade!” was the smiling reply of the great zoölogist; and when we think of the great accumulations of Laurentian carbon, and that we are entirely ignorant of the forms and structures of the vegetation which produced it, we can scarcely suppress a feeling of disappointment. Some things, however, we can safely infer from the facts that are known, and these it may be well to mention.
The climate and atmosphere of the Laurentian may have been well adapted for the sustenance of vegetable life. We can scarcely doubt that the internal heat of the earth still warmed the waters of the sea, and these warm waters must have diffused great quantities of mists and vapours over the land, giving a moist and equable if not a very clear atmosphere. The vast quantities of carbon dioxide afterwards sealed up in limestones and carbonaceous beds must also have still floated in the atmosphere and must have supplied abundance of the carbon, which constitutes the largest ingredient in vegetable tissues. Under these circumstances the whole world must have resembled a damp, warm greenhouse, and plants loving such an atmosphere could have grown luxuriantly. In these circumstances the lower forms of aquatic vegetation and those that love damp, warm air and wet soil would have been at home.
If we ask more particularly what kinds of plants might be expected to be introduced in such circumstances, we may obtain some information from the vegetation of the succeeding Palæozoic age, when such conditions still continued to a modified extent. In this period the club-mosses, ferns, and mare’s-tails engrossed the world and grew to sizes and attained degrees of complexity of structure not known in modern times. In the previous Laurentian age something similar may have happened to Algæ, to Fungi, to Lichens, to Liverworts, and Mosses. The Algæ may have attained to gigantic dimensions, and « 18 » may have even ascended out of the water in some of their forms. These comparatively simple cellular and tubular structures, now degraded to the humble position of flat lichens or soft or corky fungi, or slender cellular mosses, may have been so strengthened and modified as to constitute forest-trees. This would be quite in harmony with what is observed in the development of other plants in primitive geological times; and a little later in this history we shall see that there is evidence in the flora of the Silurian of a survival of such forms.
It may be that no geologist or botanist will ever be able to realise these dreams of the past. But, on the other hand, it is quite possible that some fortunate chance may have somewhere preserved specimens of Laurentian plants showing their structure.
In any case we have here presented to us the strange and startling fact that the remarkable arrangement of protoplasmic matter and chlorophyll, which enables the vegetable cell to perform, with the aid of solar light, the miracle of decomposing carbon dioxide and water, and forming with them woody and corky tissues, had already been introduced upon the earth. It has been well said that no amount of study of inorganic nature would ever have enabled any one to anticipate the possibility of the construction of an apparatus having the chemical powers of the living vegetable cell. Yet this most marvellous structure seems to have been introduced in the full plenitude of its powers in the Laurentian age.
Whether this early Laurentian vegetation was the means of sustaining any animal life other than marine Protozoa, we do not know. It may have existed for its own sake alone, or merely as a purifier of the atmosphere, in preparation for the future introduction of land-animals. The fact that there have existed, even in modern times, oceanic islands rich in vegetation, yet untenanted by the higher forms of animal life, prepares us to believe « 19 » that such conditions may have been general or universal in the primeval times we are here considering.
If we ask to what extent the carbon extracted from the atmosphere and stored up in the earth has been, or is likely to be, useful to man, the answer must be that it is not in a state to enable it to be used as mineral fuel. It has, however, important uses in the arts, though at present the supply seems rather in excess of the demand, and it may well be that there are uses of graphite still undiscovered, and to which it will yet be applied.
Finally, it is deserving of notice that, if Laurentian graphite indicates vegetable life, it indicates this in vast profusion. That incalculable quantities of vegetable matter have been oxidised and have disappeared we may believe on the evidence of the vast beds of iron-ore; and, in regard to that preserved as graphite, it is certain that every inch of that mineral must indicate many feet of crude vegetable matter.
It is remarkable that, in ascending from the Laurentian, we do not at first appear to advance in evidences of plant-life. The Huronian age, which succeeded the Laurentian, seems to have been a disturbed and unquiet time, and, except in certain bands of iron-ore and some dark slates coloured with carbonaceous matter, we find in it no evidence of vegetation. In the Cambrian a great subsidence of our continents began, which went on, though with local intermissions and reversals, all through the Siluro-Cambrian or Ordovician time. These times were, for this reason, remarkable for the great abundance and increase of marine animals rather than of land-plants. Still, there are some traces of land vegetation, and we may sketch first the facts of this kind which are known, and then advert to some points relating to the earlier Algæ, or sea-weeds.
An eminent Swedish geologist, Linnarsson, has described, « 20 » under the name of Eophyton, certain impressions on old Cambrian rocks in Sweden, and which certainly present very plant-like forms. They want, however, any trace of carbonaceous matter, and seem rather to be grooves or marks cut in clay by the limbs or tails of some aquatic animal, and afterwards filled up and preserved by succeeding deposits. After examining large series of these specimens from Sweden, and from rocks of similar age in Canada, I confess that I have no faith in their vegetable nature.
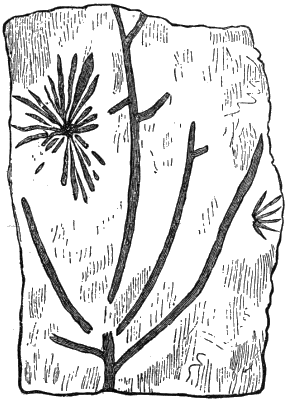
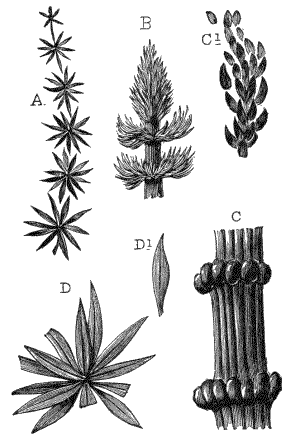
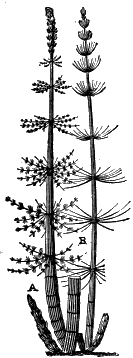
The oldest plants known to me, and likely to have been of higher grade than Algæ, are specimens kindly presented to me by Dr. Alleyne Nicholson, of Aberdeen, and which he had named Buthotrephis Harknessii[H] and B. radiata. They are from the Skiddaw rocks of Cumberland. On examining these specimens, and others subsequently collected in the same locality by Dr. Gr. M. Dawson, while convinced by their form and carbonaceous character that they are really plants, I am inclined to refer them not to Algæ, but probably to Rhizocarps. They consist of slender branching stems, with whorls of elongate and pointed leaves, resembling the genus Annularia of the coal formation. I am inclined to believe that both of Nicholson’s species are parts of one plant, and for this I have proposed the generic name Protannularia (Fig. 1). Somewhat higher in the Siluro-Cambrian, in the Cincinnati group of America, Lesquereux has found some minute radiated leaves, referred by him to the genus Sphenophyllum,[I] which is also allied to Rhizocarps. Still more remarkable is the discovery in the same beds of a stem with rhombic areoles or leaf-bases, to which the name Protostigma has been given.[J] If a plant, this may « 21 » have been allied to the club-mosses. This seems to be all that we at present know of land-vegetation in the Siluro-Cambrian. So far as the remains go, they indicate the presence of the families of Rhizocarps and of Lycopods.
[H] “Geological Magazine,” 1869.
[I] See figure in next chapter.
[J] Protostigma sigillarioides, Lesquereux.
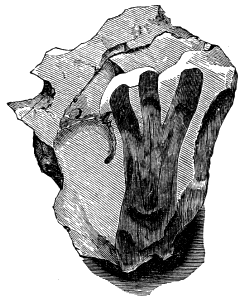
If we ascend into the Upper Silurian, or Silurian proper, the evidences of land vegetation somewhat increase. In 1859 I described, in “The Journal of the Geological Society” of London, a remarkable tree from the Lower Erian of Gaspé, under the name Prototaxites, but for which I now prefer the name Nematophyton. When in London, in 1870, I obtained permission to examine certain specimens of spore-cases or seeds from the Upper Ludlow (Silurian) formation of England, and which had been described by Sir Joseph Hooker under the name Pachytheca. In the same slabs with these I found fragments of fossil wood identical with those of the Gaspé plant. Still later I recognised similar fragments associated also with Pachytheca in the Silurian of Cape Bon Ami, New Brunswick. Lastly, Dr. Hicks has discovered similar wood, and also similar « 22 » fruits, in the Denbighshire grits, at the base of the Silurian.[K]
[K] “Journal of the Geological Society,” August, 1881.

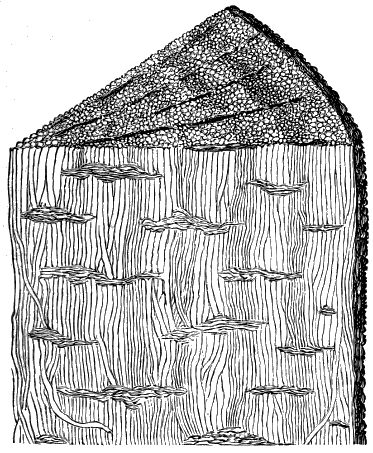
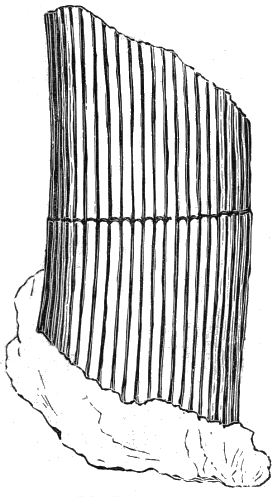
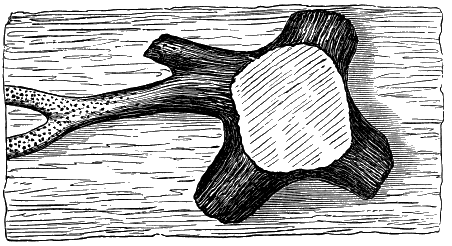
From comparison of this singular wood, the structure of which is represented in Figs. 2, 3, and 4, with the débris of fossil taxine woods, mineralised after long maceration in water, I was inclined to regard Prototaxites, or, as I have more recently named it, Nematophyton, as a primeval gymnosperm allied to those trees which Unger had described from the Erian of Thuringia, under the name Aporoxylon.[M] Later examples of more lax tissues from branches or young stems, and the elaborate examinations kindly undertaken for me by Professor Penhallow and referred to in a note to this chapter, have induced me to modify this view, and to hold that the tissues of these singular trees, which seem to have existed from the beginning « 24 » of the Silurian age and to have finally disappeared in the early Erian, are altogether distinct from any form of vegetation hitherto known, and are possibly survivors of that prototypal flora to which I have already referred. They are trees of large size, with a coaly bark and large spreading roots, having the surface of the stem smooth or irregularly ribbed, but with a nodose or jointed appearance. Internally, they show a tissue of long, cylindrical tubes, traversed by a complex network of horizontal tubes thinner walled and of smaller size. The tubes are arranged in concentric zones, which, if annual rings, would in some specimens indicate an age of one hundred and fifty years. There are also radiating spaces, which I was at first disposed to regard as true medullary rays, or which at least indicate a radiating arrangement of the tissue. They now seem to be spaces extending from the centre towards the circumference of the stem, and to have contained bundles of tubes gathered from the general tissue and extending outward perhaps to organs or appendages on the surface. Carruthers has suggested a resemblance to Algæ, and has even proposed to change the name to Nematophycus, or “thread-sea-weed”; but the resemblance is by no means clear, and it would be quite as reasonable to compare the tissue to that of some Fungi or Lichens, or even to suppose that a plant composed of cylindrical tubes has been penetrated by the mycelium or spawn of a dry-rot fungus. But the tissues are too constant and too manifestly connected with each other to justify this last supposition. That the plant grew on land I cannot doubt, from its mode of occurrence; that it was of durable and resisting character is shown by its state of preservation; and the structure of the seeds called Pachytheca, with their constant association with these trees, give countenance to the belief that they are the fruit of Nematophyton. Of the foliage or fronds of these strange plants we unfortunately know nothing. They seem, however, « 25 » to realise the idea of arboreal plants having structures akin to those of thallophytes, but with seeds so large and complex that they can scarcely be regarded as mere spores. They should perhaps constitute a separate class or order to which the name Nematodendreæ may be given, and of which Nematophyton will constitute one genus and Aporoxylon of Unger another.[N]
[M] “Palæontologie des Thuringer Waldes,” 1856.
[N] See report by the author on “Erian Flora of Canada,” 1871 and 1882, for full description of these fossils.
Another question arises as to the possible relation of these plants to other trees known by their external forms. The Protostigma of Lesquereux has already been referred to, and Claypole has described a tree from the Clinton group of the United States, with large ovate leaf-bases, to which he has given the name Glyptodendron.[O] If the markings on these plants are really leaf-bases, they can scarcely have been connected with Nematophyton, because that tree shows no such surface-markings, though, as we have seen, it had bundles of tubes passing diagonally to the surface. These plants were more probably trees with an axis of barred vessels and thick, cellular bark, like the Lepidodendron of later periods, to be noticed in the sequel. Dr. Hicks has also described from the same series of beds which afforded the fragments of Nematophyton certain carbonised dichotomous stems, which he has named Berwynia. It is just possible that these plants may have belonged to the Nematodendreæ. The thick and dense coaly matter which they show resembles the bark of these trees, the longitudinal striation in some of them may represent the fibrous structure, and the lateral projections which have been compared to leaves or leaf-bases may correspond with the superficial eminences of Nematophyton, and the spirally arranged punctures which it shows on its surface. In this case I should be disposed to regard « 26 » the supposed stigmaria-like roots as really stems, and the supposed rootlets as short, spine-like rudimentary leaves. All such comparisons must, however, in the mean time be regarded as conjectural. We seem, however, to have here a type of tree very dissimilar to any even of the later Palæozoic age, which existed throughout the Silurian, and probably further back, which ceased to exist early in the Erian age, and before the appearance of the ordinary coniferous and lepidodendroid trees. May it not have been a survivor of an old arboreal flora extending back even to the Laurentian itself?
[O] “American Journal of Science,” 1878.
Multitudes of markings occurring on the surfaces of the older rocks have been referred to the Algæ or sea-weeds, and indeed this group has been a sort of refuge for the destitute to which palæontologists have been accustomed to refer any anomalous or inexplicable form which, while probably organic, could not be definitely referred to the animal kingdom. There can be no question that some of these are truly marine plants; and that plants of this kind occur in formations older than those in which we first find land-plants, and that they have continued to inhabit the sea down to the present time. It is also true that the oldest of these Algæ closely resemble in form plants of this kind still existing; and, since their simple cellular structures and soft tissues are scarcely ever preserved, their general forms are all that we can know, so that their exact resemblance to or difference from modern types can rarely be determined. For the same reasons it has proved difficult clearly to distinguish them from mere inorganic markings or the traces of animals, and the greatest divergence of opinion has occurred in recent times on these subjects, as any one can readily understand who consults the voluminous and well-illustrated memoirs of Nathorst, Williamson, Saporta, and Delgado.
The author of this work has given much attention to these remains, and has not been disposed to claim for the « 27 » vegetable kingdom so many of them as some of his contemporaries.[P] The considerations which seem most important in making such distinctions are the following: 1. The presence or absence of carbonaceous matter. True Algæ not infrequently present at least a thin film of carbon representing their organic matter, and this is the more likely to occur in their case, as organic matters buried in marine deposits and not exposed to atmospheric oxidation are very likely to be preserved. 2. In the absence of organic matter, the staining of the containing rock, the disappearance or deoxidation of its ferruginous colouring matter, or the presence of iron pyrite may indicate the removal of organic matter by decay. 3. When organic matter and indications of it are altogether absent, and form alone remains, we have to distinguish from Algæ, trails and burrows similar to those of aquatic animals, casts of shrinkage-cracks, water-marks, and rill-marks widely diffused over the surfaces of beds. 4. Markings depressed on the upper surfaces of beds, and filled with the material of the succeeding layer, are usually mere impressions. The cases of possible exceptions to this are very rare. On the contrary, there are not infrequently forms in relief on the surfaces of rocks which are not Algæ, but may be shallow burrows arched upward on top, or castings of worms thrown up upon the surface. Sometimes, however, they may have been left by denudation of the surrounding material, just as footprints on dry snow remain in relief after the surrounding loose material has been drifted away by the wind; the portion consolidated by pressure being better able to resist the denuding agency.
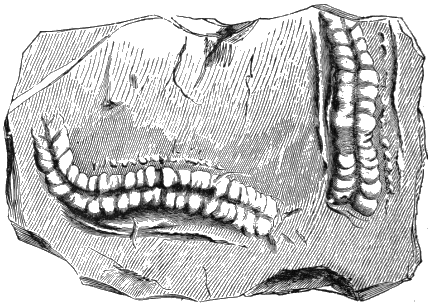
The footprints from the Potsdam sandstone in Canada, for which the name Protichnites was proposed by Owen, and which were by him referred to crustaceans probably resembling Limulus, were shown by the writer, in 1862,[Q] to correspond precisely with those of the American Limulus (Polyphemus Occidentalis) (Fig. 5). I proved by experiment with the modern animal that the recurring series of groups of markings were produced by the toes of the large posterior thoracic feet, the irregular scratches seen in Protichnites lineatus by the ordinary feet, and the central furrow by the tail. It was also shown that when the Limulus uses its swimming-feet it produces impressions of the character of those named « 29 » Climactichnites, from the same beds which afford Protichnites. The principal difference between Protichnites and their modern representatives is that the latter have two lateral furrows produced by the sides of the carapace, which are wanting in the former.
[Q] “Canadian Naturalist,” vol. vii.
I subsequently applied the same explanation to several other ancient forms now known under the general name Bilobites (Figs. 6 and 7).[R]
[R] The name Bilobites was originally proposed by De Kay for a bivalve shell (Conocardium). Its application to supposed Algæ was an error, but this is of the less consequence, as these are not true plants but only animal trails.
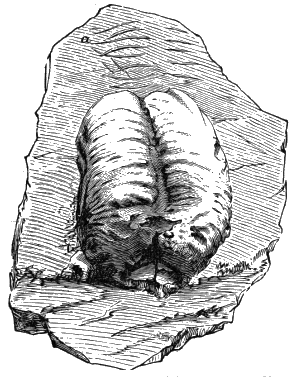
The tuberculated impressions known as Phymatoderma and Caulerpites may, as Zeiller has shown, be made by the burrowing of the mole-cricket, and fine examples occurring in the Clinton formation of Canada are probably the work of Crustacea. It is probable, however, that some of the later forms referred to these genera are really Algæ related to Caulerpa, or even branches of Conifers of the genus Brachyphyllum.
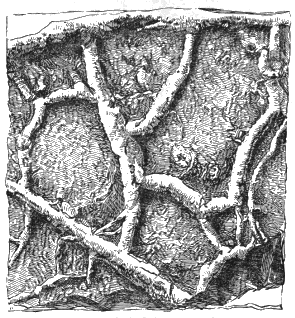
Nereites and Planulites are tracks and burrows of worms, with or without marks of setæ, and some of the markings referred to Palæochorda, Palæophycus, and Scolithus have their places here. Many examples highly illustrative of the manner of formation of the impressions are afforded by Canadian rocks (Fig. 8).
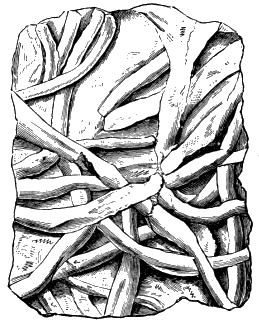
Branching forms referred to Licrophycus of Billings, and some of those referred to Buthotrephis, Hall, as well as radiating markings referable to Scotolithus, Gyrophyllites, and Asterophycus, are explained by the branching burrows of worms illustrated by Nathorst and the author. Astropolithon, a singular radiating marking of the Canadian Cambrian,[S] seems to be something organic, but of what nature is uncertain (Fig. 9).
[S] Supplement to “Acadian Geology.”
Rhabdichnites and Eophyton belong to impressions explicable by the trails of drifting sea-weeds, the tail-markings of Crustacea, and the ruts ploughed by bivalve mollusks, and occurring in the Silurian, Erian, and Carboniferous rocks.[T] Among these are the singular bilobate forms described as Rusophycus by Hall, and which are probably burrows or resting-places of crustaceans. The tracks of such animals, when walking, are the jointed impressions known as Arthrophycus and Crusiana. I have shown by the mode of occurrence « 31 » of these, and Nathorst has confirmed this conclusion by elaborate experiments on living animals, that these forms are really trails impressed on soft sediments by animals and mostly by crustaceans.
[T] “Canadian Naturalist,” 1864.
I agree with Dr. Williamson[U] in believing that all or nearly all the forms referred to Crossochorda of Schimper are really animal impressions allied to Nereites, and due either to worms or, as Nathorst has shown to be possible, to small crustaceans. Many impressions of this kind occur in the Silurian beds of the Clinton series in Canada and New York, and are undoubtedly mere markings.
[U] “Tracks from Yoredale Rocks,” “Manchester Literary and Philosophical Society,” 1885.
It is worthy of note that these markings strikingly resemble the so-called Eophyton, described by Torell from the Primordial of Sweden, and by Billings from that of Newfoundland; and which also occur abundantly in the Primordial of New Brunswick. After examining a series of these markings from Sweden shown to me by Mr. Carruthers in London, and also specimens from Newfoundland and a large number in situ at St. John, I am convinced that they cannot be plants, but must be markings of the nature of Rhabdichnites. This conclusion is based on the absence of carbonaceous matter, the intimate union of the markings with the surface of the stone, « 32 » their indefinite forms, their want of nodes or appendages, and their markings being always of such a nature as could be produced by scratches of a sharp instrument. Since, however, fishes are yet unknown in beds of this age, they may possibly be referred to the feet or spinous tails of swimming crustaceans. Salter has already suggested this origin for some scratches of somewhat different form found in the Primordial of Great Britain. He supposed them to have been the work of species of Hymenocaris. These marks may, however, indicate the existence of some free-swimming animals of the Primordial seas as yet unknown to us.
Three other suggestions merit consideration in this connection. One is that Algæ and also land-plants, drifting with tides or currents, often make the most remarkable and fantastic trails. A marking of this kind has been observed by Dr. G. M. Dawson to be produced by a drifted Laminaria, and in complexity it resembled the extraordinary Ænigmichnus multiformis of Hitchcock from the Connecticut sandstones. Much more simple markings of this kind would suffice to give species of Eophyton. Another is furnished by a fact stated to the author by Prof. Morse, namely, that Lingulæ, when dislodged from their burrows, trail themselves over the bottom like worms, by means of their cirri. Colonies of these creatures, so abundant in the Primordial, may, when obliged to remove, have covered the surfaces of beds of mud with vermicular markings. The third is that the Rhabdichnite-markings resemble some of the grooves in Silurian rocks which have been referred to trails of Gasteropods, as, for instance, those from the Clinton group, described by Hall.
Another kind of markings not even organic, but altogether depending on physical causes, are the beautiful branching rill-marks produced by the oozing of water « 33 » out of mud and sand-banks left by the tide, and which sometimes cover great surfaces with the most elaborate tracery, on the modern tidal shores as well as in some of the most ancient rocks. Dendrophycus[V] of Lesquereux seems to be an example of rill-mark, as well as Aristophycus, Clœphycus, and Zygopliycus, of Miller and Dyer, from the Lower Silurian.
[V] “Coal Flora of Pennsylvania,” vol. iii., Plate 88.
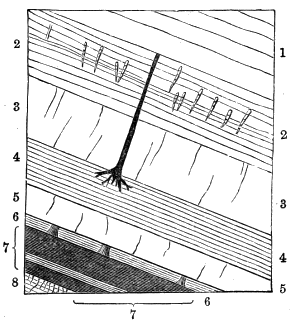
Rill-marks occur in very old rocks,[W] but are perhaps most beautifully preserved in the Carboniferous shales and argillaceous sandstones, and even more elaborately on the modern mud-banks of the Bay of Fundy.[X] Some of these simulate ferns and fronds of Laminariæ, and others resemble roots, fucoids allied to Buthotrephis, or the radiating worm-burrows already referred to (Fig. 10).
[W] “Journal of the Geological Society,” vol. xii., p. 251.
[X] “Acadian Geology,” 2d ed., p. 26.
Shrinkage-cracks are also abundant in some of the Carboniferous beds, and are sometimes accompanied with impressions of rain-drops. When finely reticulated they might be mistaken for the venation of leaves, and, when complicated with little rill-marks tributary to their sides, they precisely resemble the Dictyolites of Hall from the Medina sandstone (Fig. 11).
An entirely different kind of shrinkage-crack is that which occurs in certain carbonised and flattened plants, and which sometimes communicates to them a marvellous resemblance to the netted under surface of an exogenous leaf. Flattened stems of plants and layers of cortical matter, when carbonised, shrink in such a manner as to produce minute reticulated cracks. These become filled with mineral matter before the coaly substance has been completely consolidated. A further compression occurs, causing the coaly substance to collapse, leaving the little veins of harder mineral matter projecting. These impress their form upon the clay or shale above and below, and thus when the mass is broken open we have a carbonaceous film or thin layer covered with a network of raised lines, and corresponding minute depressed lines on the shale in contact with it. The reticulations are generally irregular, but sometimes they very closely resemble the veins of a reticulately veined leaf. One of the most curious specimens in my possession was collected by Mr. Elder in the Lower Carboniferous of Horton Bluff. The little veins which form the projecting network are in this case white calcite; but at the surface their projecting edges are blackened with a carbonaceous film.
Slickensided bodies, resembling the fossil fruits described by Geinitz as Gulielmites, and the objects believed « 35 » by Fleming and Carruthers[Y] to be casts of cavities filled with fluid, abound in the shales of the Carboniferous and Devonian. They are, no doubt, in most cases the results of the pressure and consolidation of the clay around small solid bodies, whether organic, fragmentary, or concretionary. They are, in short, local slickensides precisely similar to those found so plentifully in the coal under-clays, and which, as I have elsewhere[Z] shown, resulted from the internal giving way and slipping of the mass as the roots of Stigmaria decayed within it. Most collectors of fossil plants in the older formations must, I presume, be familiar with appearances of this kind in connection with small stems, petioles, fragments of wood, and carpolites. I have in my collection petioles of ferns and fruits of the genus Trigonocarpum partially slickensided in this way, and which if wholly covered by this kind of marking could scarcely have been recognised. I have figured bodies of this kind in my report on the Devonian and Upper Silurian plants of Canada, believing them, owing to their carbonaceous covering, to be probably slickensided fruits, though of uncertain nature. In every case I think these bodies must have had a solid nucleus of some sort, as the severe pressure implied in slickensiding is quite incompatible with a mere “fluid-cavity,” even supposing this to have existed.
[Y] “Journal of the Geological Society,” June, 1871.
[Z] Ibid., vol. x., p. 14.
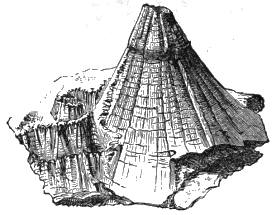
Prof. Marsh has well explained another phase of the influence of hard bodies in producing partial slickensides, in his paper on Stylolites, read before the American Association in 1867, and the application of the combined forces of concretionary action and slickensiding to the production of the cone-in-cone concretions, which occur in the coal-formation and as low as the Primordial. I have figured a very perfect and beautiful form of this « 36 » kind from the coal-formation of Nova Scotia, which is described in “Acadian Geology”[AA] (Fig. 12).
I have referred to these facts here because they are relatively more important in that older period, which may be named the age of Algæ, and because their settlement now will enable us to dispense with discussions of this kind further on. The able memoirs of Nathorst and Williamson should be studied by those who desire further information.
[AA] Appendix, p. 676, edition of 1878.
But it may be asked, “Are there no real examples of fossil Algæ?” I believe there are many such, but the difficulty is to distinguish them. Confining ourselves to the older rocks, the following may be noted:
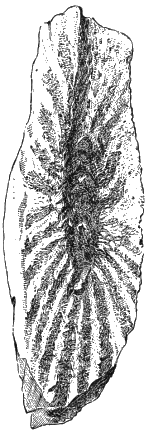
The genus Buthotrephis of Hall, which is characterised as having stems, sub-cylindric or compressed, with numerous branches, which are divaricating and sometimes leaf-like, contains some true Algæ. Hall’s B. gracilis, from the Siluro-Cambrian, is one of these. Similar plants, referred to the same species, occur in the Clinton and Niagara formations, and a beautiful species, collected by Col. Grant, of Hamilton, and now in the McGill College collection, represents a broader and more frondose type of distinctly carbonaceous character. It may be described as follows:
Buthotrephis Grantii, S. N. (Fig. 13).—Stems and « 37 » fronds smooth and slightly striate longitudinally, with curved and interrupted striæ. Stem thick, bifurcating, the divisions terminating in irregularly pinnate fronds, apparently truncate at the extremities. The quantity of carbonaceous matter present would indicate thick, though perhaps flattened, stems and dense fleshy fronds.
The species Buthotrephis subnodosa and B. flexuosa, from the Utica shale, are also certainly plants, though it is possible, if their structures and fruit were known, some of these might be referred to different genera. All of these plants have either carbonaceous matter or produce organic stains on the matrix.
The organism with diverging wedge-shaped fronds, described by Hall as Sphenothallus angustifolius, is also a plant. Fine specimens, in the collection of the Geological Survey of Canada, show distinct « 38 » evidence of the organic character of the wedge-shaped fronds. It is from the Utica shale, and elsewhere in the Siluro-Cambrian. It is just possible, as suggested by Hall, that this plant may be of higher rank than the Algæ.
The genus Palæophycus of Hall includes a great variety of uncertain objects, of which only a few are probably true Algæ. I have specimens of fragments similar to his P. virgatus, which show distinct carbonaceous films, and others from the Quebec group, which seem to be cylindrical tubes now flattened, and which have contained spindle-shaped sporangia of large size. Tortuous and curved flattened stems, or fronds, from the Upper Silurian limestone of Gaspé, also show organic matter.
Respecting the forms referred to Licrophycus by Billings, containing stems or semi-cylindrical markings springing from a common base, I have been in great doubt. I have not seen any specimens containing unequivocal organic matter, and am inclined to think that most of them, if not the whole, are casts of worm-burrows, with trails radiating from them.
Though I have confined myself in this notice to plants, or supposed plants, of the Lower Palæozoic, it may be well to mention the remarkable Cauda-Galli fucoids, referred by Hall to the genus Spirophyton, and which are characteristic of the oldest Erian beds. The specimens which I have seen from New York, from Gaspé, and from Brazil, leave no doubt in my mind that these were really marine plants, and that the form of a spiral frond, assigned to them by Hall, is perfectly correct. They must have been very abundant and very graceful plants of the early Erian, immediately after the close of the Silurian period.
We come now to notice certain organisms referred to Algæ, and which are either of animal origin, or are of higher grade than the sea-weeds. We have already discussed « 39 » the questions relating to Prototaxites. Drepanophycus, of Goeppert,[AB] I suspect, is only a badly preserved branch or stem of the Erian land-plant known as Arthrostigma. In like manner, Haliserites Dechenianus,[AC] of Goeppert, is evidently the land-plant known as Psilophyton. Sphærococcites dentatus and S. serra—the Fucoides dentatus and serra of Brongniart, from Quebec—are graptolites of two species quite common there.[AD] Dictyophyton and Uphantenia, as described by Hall and the author, are now known to be sponges. They have become Dictyospongiæ. The curious and very ancient; fossils referred by Forbes to the genus Oldhamia are perhaps still subject to doubt, but are usually regarded as Zoöphytes, though it is quite possible they may be plants. Though I have not seen the specimens, I have no doubt whatever that the plants, or the greater part of them, from the Silurian of Bohemia, described by Stur as Algæ and Characeæ,[AE] are really land-plants, some of them of the genus Psilophyton. I may say in this connection that specimens of flattened Psilophyton and Arthrostigma, in the Upper Silurian and Erian of Gaspé, would probably have been referred to Algæ, but for the fact that in some of them the axis of barred vessels is preserved.
[AB] “Fossile Flora,” 1852, p. 92, Table xli.
[AC] Ibid., p. 88, Table ii.
[AD] Brongniart, “Vegeteaux Fossiles,” Plate vi., Figs. 7 to 12.
[AE] “Proceedings of the Vienna Academy,” 1881. Hostinella, of this author, is almost certainly Psilophyton, and his Barrandiana seems to include Arthrostigma, and perhaps leafy branches of Berwynia. These curious plants should be re-examined.
It is not surprising that great difficulties have occurred in the determination of fossil Algæ. Enough, however, remains certain to prove that the old Cambrian and Silurian seas were tenanted with sea-weeds not very dissimilar from those of the present time. It is further probable that some of the graphitic, carbonaceous, and bituminous « 40 » shales and limestones of the Silurian owe their carbonaceous matters to the decomposition of Algæ, though possibly some of it may have been derived from Graptolites and other corneous Zoöphytes. In any case, such microscopic examinations of these shales as I have made, have not produced any evidence of the existence of plants of higher grade, while those of the Erian and Carboniferous periods, similar to the naked eye, abound in such evidence. It is also to be observed that, on the surfaces of beds of sandstone in the Upper Cambrian, carbonaceous débris, which seems to be the remains of either aquatic or land plants, is locally not infrequent.

Referring to the land vegetation of the older rocks, it is difficult to picture its nature and appearance. We may imagine the shallow waters filled with aquatic or amphibious Rhizocarpean plants, vast meadows or brakes of the delicate Psilophyton and the starry Protannularia and some tall trees, perhaps looking like gigantic club-mosses, or possibly with broad, flabby leaves, mostly cellular in texture, and resembling Algæ transferred to the air. Imagination can, however, scarcely realise this strange and grotesque vegetation, which, though possibly copious and luxuriant, must have been simple and monotonous in aspect, and, though it must have produced spores and seeds and even fruits, these were probably all of the types seen in the modern acrogens and gymnosperms.
“In garments green, indistinct in the twilight,
They stand like Druids of old, with voices sad and prophetic.”
Prophetic they truly were, as we shall find, of the more varied forests of succeeding times, and they may also help us to realise the aspect of that still older vegetation, which is fossilised in the Laurentian graphite; though it is not impossible that this last may have been of higher and more varied types, and that the Cambrian and Silurian may have been times of depression in the vegetable world, as they certainly were in the submergence of much of the land.
These primeval woods served at least to clothe the nakedness of the new-born land, and they may have sheltered and nourished forms of land-life still unknown to us, as we find as yet only a few insects and scorpions in the Silurian. They possibly also served to abstract from the atmosphere some portion of its superabundant carbonic acid harmful to animal life, and they stored up « 42 » supplies of graphite, of petroleum, and of illuminating gas, useful to man at the present day. We may write of them and draw their forms with, the carbon which they themselves supplied.
NOTE TO CHAPTER II.
Examination of Prototaxites (Nematophyton), by Prof. Penhallow, of McGill University.
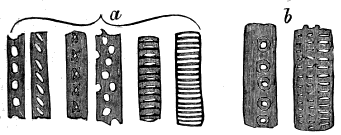
Prof. Penhallow, having kindly consented to re-examine my specimens, has furnished me with elaborate notes of his facts and conclusions, of which the following is a summary, but which it is hoped will be published in full:
"1. Concentric Layers.—The inner face of each of these is composed of relatively large tubes, having diameters from 13·6 to 34·6 micro-millimetres. The outer face has tubes ranging from 13·8 to 27·6 mm. The average diameter in the lower surface approaches to 34, that in the outer to 13·8. There is, however, no abrupt termination to the surface of the layers, though in some specimens they separate easily, with shining surfaces.
"2. Minute Structure.—In longitudinal sections the principal part of the structure consists of longitudinal tubes of indeterminate length, and round in cross-section. They are approximately parallel, but in some cases may be seen to bend sinuously, and are not in direct contact. Finer myceloid tubes, 5·33 mm. in diameter, traverse the structure in all directions, and are believed to branch off from the larger tubes. In a small specimen supposed to be a branch or small stem, and in which the vertical tubes are somewhat distant from one another, this horizontal system is very largely developed; but is less manifest in the older stems. The tubes themselves show no structure. The ray-like openings in the substance of the tissue are evidently original parts of the structure, but not of the nature of medullary rays. They are radiating spaces running outward in an interrupted manner or so tortuously that they appear to be interrupted in their course from the centre towards the surface. They show tubes turning into them, branching into them, and approximately horizontal, but tortuous. On the external surface of some specimens these radial spaces are represented by minute pits irregularly « 43 » or spirally arranged. The transverse swellings of the stem show no difference of structure, except that the tubes or cells may be a little more tortuous, and a transverse film of coaly matter extends from the outer coaly envelope inwardly. This may perhaps be caused by some accident of preservation. The outer coaly layer shows tubes similar to those of the stem.[AF] The horizontal or oblique flexures of the large tubes seem to be mainly in the vicinity of the radial openings, and it is in entering these that they have been seen to branch."
[AF] It is possible that these tubes may be merely part of the stem attached to the bark, which seems to me to indicate the same dense cellular structure seen in the bark of Lepidodendra, etc.
The conclusions arrived at by Prof. Penhallow are as follows:
"1. The plant was not truly exogenous, and the appearance of rings is independent of the causes which determine the layers of growth in exogenous plants.
"2. The plant was possessed of no true bark. Whatever cortical layer was present was in all probability a modification of the general structure,[AG]
[AG] On these points I would reserve the considerations: 1. That there must have been some relation between the mode of growth of these great stems and their concentric rings; and, 2. That the evidence of a bark is as strong as in the case of any Palæozoic tree in which the bark is, as usual, carbonised.
"3. An intimate relation exists between the large tubular cells and the myceloid filaments, the latter being a system of small branches from the former; the branching being determined chiefly in certain special openings which simulate medullary rays.
"4. The specimens examined exhibit no evidence of special decay, and the structure throughout is of a normal character.
"5. The primary structure consists of large tubular cells without apparent terminations, and devoid of structural markings, with which is associated a secondary structure of myceloid filaments arising from the former.
"6. The structure of Nematophyton as a whole is unique; at least there is no plant of modern type with which it is comparable. Nevertheless, the loose character of the entire structure; the interminable cells; their interlacing; and, finally, their branching into a secondary series of smaller filaments, point with considerable force to the true relationship of the stem as being with Algæ or other Thallophytes rather than with Grymnosperms. A more recent examination « 44 » of a laminated resinous substance found associated with the plant shows that it is wholly amorphous, and, as indicated by distinct lines of flow, that it must have been in a plastic state at a former period. The only evidence of structure was found in certain well-defined mycelia, which may have been derived from associated vegetable matter upon which they were growing, and over which the plastic matrix flowed."
I have only to add to this description that when we consider that Nematophyton Logani was a large tree, sometimes attaining a diameter of more than two feet, and a stature of at least twenty before branching; that it had great roots, and gave off large branches; that it was an aërial plant, probably flourishing in the same swampy flats with Psilophyton, Arthrostigma, and Leptophleum; that the peculiar bodies known as Pachytheca were not unlikely its fruit—we have evidence that there were, in the early Palæozoic period, plants scarcely dreamt of by modern botany. Only when the appendages of these plants are more fully known can we hope to understand them. In the mean time, I may state that there were probably different species of these trees, indicated more particularly by the stems I have described as Nematoxylon and Celluloxylon[AH] There were, I think, some indications that the plants described by Carruthers as Berwynia, may also be found to have been generically the same. The resinous matter mentioned by Prof. Penhallow is found in great abundance in the beds containing Nematophyton, and must, I think, have been an exudation from its bark.
THE ERIAN OF DEVONIAN FORESTS—ORIGIN OF PETROLEUM—THE AGE OF ACROGENS AND GYMNOSPERMS.
In the last chapter we were occupied with the comparatively few and obscure remains of plants entombed in the oldest geological formations. We now ascend to a higher plane, that of the Erian or Devonian period, in which, for the first time, we find varied and widely distributed forests.
The growth of knowledge with respect to this flora has been somewhat rapid, and it may be interesting to note its principal stages, as an encouragement to the hope that we may yet learn something more satisfactory respecting the older floras we have just discussed.
In Goeppert’s memoir on the flora of the Silurian, Devonian, and Lower Carboniferous rocks, published in 1860,[AI] he enumerates twenty species as Silurian, but these are all admitted to be Algæ, and several of them are remains which may be fairly claimed by the zoologists as zoophytes, or trails of worms and mollusks. In the Lower Devonian he knows but six species, five of which are Algæ, and the remaining one a Sigillaria, but this is of very doubtful nature. In the Middle Devonian he gives but one species, a land-plant of the genus Lepidodendron. In the Upper Devonian the number rises to fifty-seven, of which all but seven are terrestrial plants, representing a large number of the genera occurring in the succeeding Carboniferous system.
Goeppert does not include in his enumeration the plants from the Devonian of Gaspé, described by the author in 1859,[AJ] having seen only an abstract of the paper at the time of writing his memoir, nor does he appear to have any knowledge of the plants of this age described by Lesquereux in Roger’s “Pennsylvania.” These might have added ten or twelve species to his list, some of them probably from the Lower Devonian. It is further to be observed that a few additional species had also been recognised by Peach in the Old Red Sandstone of Scotland.
[AJ] “Journal of the Geological Society of London,” also “Canadian Naturalist.”
But from 1860 to the present time a rich harvest of specimens has been gathered from the Gaspé sandstones, from the shales of southern New Brunswick, from the sandstones of Perry in Maine, and from the wide-spread Erian areas of New York, Pennsylvania, and Ohio. Nearly all these specimens have passed through my hands, and I am now able to catalogue about a hundred species, representing more than thirty genera, and including all the great types of vascular Cryptogams, the Gymnosperms, and even one (still doubtful) Angiosperm. Many new forms have also been described from the Devonian of Scotland and of the Continent of Europe.
Before describing these plants in detail, we may refer to North America for illustration of the physical conditions of the time. In a physical point of view the northern hemisphere presented a great change in the Erian period. There were vast foldings of the crust of the earth, and great emissions of volcanic rock on both sides of the Atlantic. In North America, while at one time the whole interior area of the continent, as far north as « 47 » the Great Lakes, was occupied by a vast inland sea, studded with coral islands, the long Appalachian ridge had begun to assume, along with the old Laurentian land, something of the form of our present continent, and on the margins of this Appalachian belt there were wide, swampy flats and shallow-water areas, which, under the mild climate that seems to have characterised this period, were admirably suited to nourish a luxuriant vegetation. Under this mild climate, also, it would seem that new forms of plants were first introduced in the far north, where the long continuance of summer sunlight, along with great warm th, seems to have aided in their introduction and early extension, and thence made their way to the southward, a process which, as Gray and others have shown, has also occurred in later geological times.
The America of this Erian age consisted during the greater part of the period of a more or less extensive belt of land in the north with two long tongues descending from it, one along the Appalachian line in the east, the other in the region west of the Rocky Mountains. On the seaward sides of these there were low lands covered with vegetation, while on the inland side the great interior sea, with its verdant and wooded islands, realised, though probably with shallower water, the conditions of the modern archipelagoes of the Pacific.
Europe presented conditions somewhat similar, having in the earlier and middle portions of the period great sea areas with insular patches of land, and later wide tracts of shallow and in part enclosed water areas, swarming with fishes, and having an abundant vegetation on their shores. These were the conditions of the Eifel and Devonshire limestones, and of the Old Red Sandstone of Scotland, and the Kiltorcan beds of Ireland. In Europe also, as in America, there were in the Erian age great ejections of igneous rock. On both sides of the Atlantic there were somewhat varied and changing conditions of « 48 » land and water, and a mild and equable climate, permitting the existence of a rich vegetation in high northern latitudes. Of this latter fact a remarkable example is afforded by the beds holding plants of this age in Spitzbergen and Bear Island, in its vicinity. Here there seem to be two series of plant-bearing strata, one with the vegetation of the Upper Erian, the other with that of the Lower Carboniferous, though both have been united by Heer under his so-called “Ursa Stage” in which he has grouped the characteristic plants of two distinct periods. This has recently been fully established by the researches of Nathorst, though the author had already suggested it as the probable explanation of the strange union of species in the Ursa group of Heer.
In studying the vegetation of this remarkable period, we must take merely some of the more important forms as examples, since it would be impossible to notice all the species, and some of them may be better treated in the Carboniferous, where they have their headquarters. (Fig. 15.)
I may first refer to a family which seems to have culminated in the Erian age, and ever since to have occupied a less important place. It is that of the curious aquatic plants known as Rhizocarps,[AK] and referred to in the last chapter.
[AK] Or, as they have recently been named by some botanists, “Heterosporous Filices,” though they are certainly not ferns in any ordinary sense of that term.
My attention was first directed to these organisms by the late Sir W. E. Logan in 1869. He had obtained from the Upper Erian shale of Kettle Point, Lake Huron, specimens filled with minute circular discs, to which he referred, in his report of 1863, as “microscopic orbicular bodies.” Recognising them to be macrospores, or spore-cases, I introduced them into the report on the Erian « 49 » flora, which I was then preparing, and which was published in 1871, under the name Sporangites Huronensis.

In 1871, having occasion to write a communication to the “American Journal of Science” on the question then raised as to the share of spores and spore-cases in the accumulation of coal, a question to be discussed in a subsequent « 50 » chapter, these curious little bodies were again reviewed, and were described in substance as follows:
“The oldest bed of spore-cases known to me is that at Kettle Point, Lake Huron. It is a bed of brown bituminous shale, burning with much flame, and under a lens is seen to be studded with flattened disc-like bodies, scarcely more than a hundredth of an inch in diameter, which under the microscope are found to be spore-cases (or macrospores) slightly papillate externally (or more properly marked with dark pores), and sometimes showing a point of attachment on one side and a slit more or less elongated and gaping on the other. When slices of the rock are made, its substance is seen to be filled with these bodies, which, viewed as transparent objects, appear yellow like amber, and show little structure, except that the walls can be distinguished from the internal cavity, which may sometimes be seen to enclose patches of granular matter. In the shale containing them are also vast numbers of rounded, translucent granules, which may be escaped spores (microspores).” The bed containing these spores at Kettle Point was stated, in the reports of the “Geological Survey of Canada,” to be twelve or fourteen feet in thickness, and besides these specimens it contained fossil plants referable to the species Calamites inornatus and Lepidodendron primævum, and I not unnaturally supposed that the Sporangites might be the fruit of the latter plant. I also noticed their resemblance to the spore-cases of L. corrugatum of the Lower Carboniferous (a Lepidodendron allied to L. primævum), and to those from Brazil described by Carruthers under the name Flemingites, as well as to those described by Huxley from certain English coals, and to those of the Tasmanite or white coal of Australia. The bed at Kettle Point is shown to be marine by its holding the sea-weed known as Spirophyton, and shells of Lingula.
The subject did not again come under my notice till « 51 » 1882, when Prof. Orton, of Columbus, Ohio, sent me some specimens from the Erian shales of that State, which on comparison seemed undistinguishable from Sporangites Huronensis.[AL] Prof. Orton read an interesting paper on these bodies, at the meeting of the American Association in Montreal, in which were some new and striking facts. One of these was the occurrence of such bodies throughout the black shales of Ohio, extending “from the Huron River, on the shore of Lake Brie, to the mouth of the Scioto, in the Ohio Valley, with an extent varying from ten to twenty miles in breadth,” and estimated to be three hundred and fifty feet in thickness. I have since been informed by my friend Mr. Thomas, of Chicago, that its thickness, in some places at least, must be three times that amount. About the same time. Prof. Williams, of Cornell, and Prof. Clarke, of Northampton, announced similar discoveries in the State of New York, so that it would appear that beds of vast area and of great thickness are replete with these little vegetable discs, usually converted into a highly bituminous, amber-like substance, giving a more or less inflammable character to the containing rock.
[AL] These shales have been described, as to their chemical and geological relations, by Dr. T. Sterry Hunt, “American Journal of Science,” 1863, and by Dr. Newberry, in the “Reports of the Geological Survey of Ohio,” vol. i., 1863, and vol. iii., 1878.
Another fact insisted on by Prof. Orton was the absence of Lepidodendroid cones, and the occurrence of filamentous vegetable matter, to which the Sporangites seemed to be in some cases attached in groups. Prof. Orton also noticed the absence of the trigonal form, which belongs to the spores of many Lepidodendra, though this is not a constant character. In the discussion on Prof. Orton’s paper, I admitted that the facts detailed by him shook my previous belief of the lycopodiaceous character « 52 » of these bodies, and induced me to suspect, with Prof. Orton, that they might have belonged to some group of aquatic plants lower than the Lycopods.
Since the publication of my paper on Rhizocarps in the Palæozoic period above referred to, I have received two papers from Mr. Edward Wethered, F. G. S., in one of which he describes spores of plants found in the lower limestone shales of the Forest of Dean, and in the other discusses more generally the structure and origin of Carboniferous coal-beds.[AM] In both papers he refers to the occurrence in these coals and shales of organisms essentially similar to the Erian spores.
[AM] “Cotteswold Naturalists' Field Club,” 1884; “Journal of the Royal Microscopical Society,” 1885.
In the “Bulletin of the Chicago Academy of Science,” January, 1884, Dr. Johnson and Mr. Thomas, in their paper on the “Microscopic Organisms of the Boulder Clay of Chicago and Vicinity,” notice Sporangites Huronensis as among these organisms, and have discovered them also in large numbers in the precipitate from Chicago city water-supply. They refer them to the decomposition of the Erian shales, of which boulders filled with these organisms are of frequent occurrence in the Chicago clays. The Sporangites and their accompaniments in the boulder clay are noticed in a paper by Dr. G. M. Dawson, in the “Bulletin of the Chicago Academy,” June, 1885.
Prof. Clarke has also described, in the “American Journal of Science” for April, 1885, the forms already alluded to, and which he finds to consist of macrospores enclosed in sporocarps. He compares these with my Sporangites Huronensis and Protosalvinia bilobata, but I think it is likely that one of them at least is a distinct species.
I may add that in the “Geological Magazine” for 1875, Mr. Newton, F. G. S., of the Geological Survey of « 53 » England, published a description of the Tasmanite and Australian white coal, in which he shows that the organisms in these deposits are similar to my Sporangites Huronensis, and to the macrospores previously described by Prof. Huxley, from the Better-bed coal. Mr. Newton does not seem to have been aware of my previous description of Sporangites, and proposes the name Tasmanites punctatus for the Australian form.
Here we have the remarkable fact that the waste macrospores, or larger spores of a species of Cryptogamous plant, occur dispersed in countless millions of tons through the shales of the Erian in Canada and the United States.
No certain clue seemed to be afforded by all these observations as to the precise affinities of these widely distributed bodies; but this was furnished shortly after from an unexpected quarter. In March, 1883, Mr. Orville Derby, of the Geological Survey of Brazil, sent me specimens found in the Erian of that country, which seemed to throw a new light on the whole subject. These I described and pointed out their connection with Sporangites at the meeting of the American Association at Minneapolis, in 1883, and subsequently published my notes respecting them in its proceedings, and in the “Canadian Record of Science.”
Mr. Derby’s specimens contained the curious spiral sea-weed known as Spirophyton, and also minute rounded Sporangites like those obtained in the Erian of Ohio, and of which specimens had been sent to me some years before by the late Prof. Hartt. But they differed in showing the remarkable fact that these rounded bodies are enclosed in considerable numbers in spherical and oval sacs, the walls of which are composed of a tissue of hexagonal cells, and which resemble in every respect the involucres or spore-sacs of the little group of modern acrogens known as Rhizocarps, and living in shallow « 54 » water. More especially they resemble the sporocarps of the genus Salvinia. This fact opened up an entirely new field of investigation, and I at once proceeded to compare the specimens with the fructification of modern Rhizocarps, and found that substantially these multitudinous spores embedded in the Erie shales may be regarded as perfectly analogous to the larger spores of the modern Salvinia natans of Europe, as may be seen by the representation of them in Fig. 16.
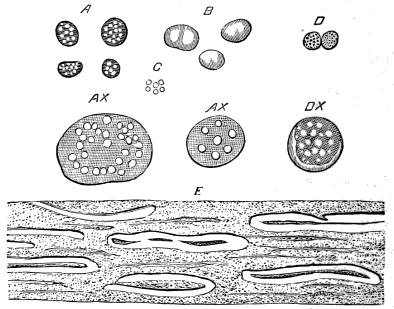
The typical macrospores from the Erian shales are perfectly circular in outline, and in the flattened state appear as discs with rounded edges, their ordinary diameter being from one seventy-fifth to one one-hundredth of an inch, though they vary considerably in size. This, however, I do not regard as an essential character. The edges, as seen in profile, are smooth, but the flat surface often presents minute dark spots, which at first I mistook « 55 » for papillæ, but now agree with Mr. Thomas in recognising them as minute pores traversing the wall of the disc, and similar to those which Mr. Newton has described in Tasmanite, and which Mr. Wethered has also recognised in the similar spores of the Forest of Dean shales. The walls also sometimes show faint indications of concentric lamination, as if they had been thickened by successive deposits.
As seen by transmitted light, and either in front or in profile, the discs are of a rich amber colour, translucent and structureless, except the pores above referred to. The walls are somewhat thick, or from one-tenth to one-twentieth the diameter of the disc in thickness. They never exhibit the triradiate marking seen in spores of Lycopods, nor any definite point of attachment, though they sometimes show a minute elongated spot which may be of this nature, and they are occasionally seen to have opened by slits on the edge or front, where there would seem to have been a natural line of dehiscence. The interior is usually quite vacant or structureless, but in some cases there are curved internal markings which may indicate a shrunken lining membrane, or the remains of a prothallus or embryo. Occasionally a fine granular substance appears in the interior, possibly remains of microspores.
The discs are usually detached and destitute of any envelope, but fragments of flocculent cellular matter are associated with them, and in one specimen from the corniferous limestone of Ohio, in Mr. Thomas’s collection, I have found a group of eight or more discs partly enclosed in a cellular sac-like membrane of similar character to that enclosing the Brazilian specimens already referred to.
The characters of all the specimens are essentially similar, and there is a remarkable absence of other organisms in the shale. In one instance only, I have observed a somewhat smaller round body with a dark centre or « 56 » nucleus, and a wide translucent margin, marked by a slight granulation. Even this, however, may indicate nothing more than a different state of preservation.
It is proper to observe here that the wall or enclosing sac of these macrospores must have been of very dense consistency, and now appears as a highly bituminous substance, in this agreeing with that of the spores of Lycopods, and, like them, having been when recent of a highly carbonaceous and hydrogenous quality, very combustible and readily admitting of change into bituminous matter. In the paper already referred to, on spore-cases in coals, I have noticed that the relative composition of lycopodium and cellulose is as follows:
| Cellulose, | C24H20O20. |
| Lycopodium, | C42H194/12NO56/10. |
Thus, such spores are admirably suited for the production of highly carbonaceous or bituminous coals, etc.
Nothing is more remarkable in connection with these bodies than their uniformity of structure and form over so great areas and throughout so great thickness of rock, and the absence of any other kind of spore-case. This is more especially noteworthy in contrast with the coarse coals and bituminous shales of the Carboniferous, which usually contain a great variety of spores and sporangia, indicating the presence of many species of acrogenous plants, while the Erian shales, on the contrary, indicate the almost exclusive predominance of one form. This contrast is well seen in the Bedford shales overlying these beds, and I believe Lower Carboniferous.[AN] Specimens of these have been kindly communicated to me by Prof. Orton, and have been prepared by Mr. Thomas. In these we see the familiar Carboniferous spores with triradiate markings called Triletes by Reinsch, and which are similar to those of Lycopodiaceous plants. Still more abundant « 57 » are those spinous and hooked spores or sporangia, to which the names Sporocarpon, Zygosporites, and Traquaria have been given, and some of which Williamson has shown to be spores of Lycopodiaceous plants.[AO]
[AN] According to Newberry, lower part of Waverly group.
[AO] Traquaria is to be distinguished from the calcareous bodies found in the corniferous limestone of Kelly’s Island, which I have described in the “Canadian Naturalist” as Saccamina Eriana, and believe to be Foraminiferal tests. They have since been described by Ulrich under a different name (Moellerina: contribution to “American Palæontology,” 1886). See Dr. Williamson’s papers in “Transactions of Royal Society of London.”
The true “Sporangites,” on the contrary, are round and smooth, with thick bituminous walls, which are punctured with minute transverse pores. In these respects, as already stated, they closely resemble the bodies found in the Australian white coal and Tasmanite. The precise geological age of this last material is not known with certainty, but it is believed to be Palæozoic.
With reference to the mode of occurrence of these bodies, we may note first their great abundance and wide distribution. The horizontal range of the bed at Kettle Point is not certainly known, but it is merely a northern outlier of the great belt of Erian shales referred to by Prof. Orton, and which extends, with a breadth of ten to twenty miles, and of great thickness, across the State of Ohio, for nearly two hundred miles. This Ohio black shale, which lies at the top of the Erian or the base of the Carboniferous, though probably mainly of Erian age, appears to abound throughout in these organisms, and in some beds to be replete with them. In like manner, in Brazil, according to Mr. Derby, these organisms are distributed over a wide area and throughout a great thickness of shale holding Spirophyton, and apparently belonging to the Upper Erian. The recurrence of similar forms in the Tasmanite and white coal of Tasmania and Australia is another important fact of distribution. To this « 58 » we may add the appearance of these macrospores in coals and shales of the Carboniferous period, though there in association with other forms.
It is also to be observed that the Erian shales, and the Forest of Dean beds described by Wethered, are marine, as shown by their contained fossils; and, though I have no certain information as to the Tasmanite and Australian white coal, they would seem, from the description of Milligan, to occur in distinctly aqueous, possibly estuarine, deposits. Wethered has shown that the discs described by Huxley and Newton in the Better-bed coal occur in the earthy or fragmentary layers, as distinguished from the pure coal. Those occurring in cannel coal are in the same case, so that the general mode of occurrence implies water-driftage, since, in the case of bodies so large and dense, wind-driftage to great distances would be impossible.
These facts, taken in connection with the differences between these macrospores and those of any known land-plant of the Palæozoic, would lead to the inference that they belonged to aquatic plants, and these vastly abundant in the waters of the Erian and Carboniferous periods.
It is still further to be observed that they are not, in the Erian beds, accompanied with any remains of woody or scalariform tissues, such as might be expected in connection with the débris of terrestrial acrogens, and that, on the other hand, we find them enclosed in cellular sporocarps, though in the majority of cases these have been removed by dehiscence or decay.
These considerations, I think, all point to the probability which I have suggested in my papers on this subject referred to above, that we have in these objects the organs of fructification of plants belonging to the order Rhizocarpeæ, or akin to it. The comparisons which I have instituted with the sporocarps and macrospores of these plants confirm this suggestion. Of the modern « 59 » species which I have had an opportunity to examine, Salvinia natans of Europe perhaps presents the closest resemblance. In this plant groups of round cellular sporocarps appear at the bases of the floating fronds. They are about a line in diameter when mature, and are of two kinds, one containing macrospores, the other microspores or antheridia. The first, when mature, hold a number of closely packed globular or oval sporangia of loose cellular tissue, attached to a central placenta. Each of these sporangia contains a single macrospore, perfectly globular and smooth, with a dense outer membrane (exhibiting traces of lamination, and showing within an irregularly vacuolated or cellular structure, probably a prothallus). I cannot detect in it the peculiar pores which appear in the fossil specimens. Each macrospore is about one-seventieth of an inch in diameter when mature. The sporocarps of the microspores contain a vastly greater number of minute sporangia, about one two-hundredths of an inch in diameter. These contain disc-like antheridia, or microspores of very minute size.
The discs from Kettle Point and from the Ohio black shale, and from the shale boulders of the Chicago clays, are similar to the macrospores of Salvinia, except that they have a thicker wall and are a little less in diameter, being about one-eightieth of an inch. The Brazilian sporocarps are considerably larger than those of the modern Salvinia, and the macrospores approach in size to those of the modern species, being one seventy-fifth of an inch in diameter. They also seem, like the modern species, to have thinner walls than those from Canada, Ohio, and Chicago. No distinct indication has been observed in the fossil species of the inner Sporangium of Salvinia. Possibly it was altogether absent, but more probably it is not preserved as a distinct structure.
With reference to the microspores of Salvinia, it is to be observed that the sporocarps, and the contained spores « 60 » or antheridia, are very delicate and destitute of the dense outer wall of the macrospores. Hence such parts are little likely to have been preserved in a fossil state; and in the Erian shales, if present, they probably appear merely as flocculent carbonaceous matter not distinctly marked, or as minute granules not well defined, of which there are great quantities in some of the shales.
The vegetation appertaining to the Sporangites has not been distinctly recognised. I have, however, found in one of the Brazilian specimens two sporocarps attached to what seems a fragment of a cellular frond, and numerous specimens of the supposed Algæ, named Spirophyton, are found in the shales, but there is no evidence of any connection of this plant with the Protosalvinia.
Modern Rhizocarps present considerable differences as to their vegetative parts. Some, like Pilularia, have simple linear leaves; others, like Marsilea, have leaves in whorls, and cuneate in form; while others, like Azolla and Salvinia, have frondose leaves, more or less pinnate in their arrangement. If we inquire as to fossils representing these forms of vegetation, we shall find that some of the plants to be noticed in the immediate sequel may have been nearly allied to the Rhizocarps. In the mean time I may state that I have proposed the generic name Protosalvinia for these curious macrospores and their coverings, and have described in the paper in the “Bulletin of the Chicago Academy of Sciences,” already quoted, five species which may be referred to this genus.
These facts lead to inquiries as to the origin of the bituminous matter which naturally escapes from the rocks of the earth as petroleum and inflammable gas, or which may be obtained from certain shales in these forms by distillation. These products are compounds of carbon and hydrogen, and may be procured from recent vegetable substances by destructive distillation. Some vegetable matters, also, are much richer in carbon and hydrogen « 61 » than others, and it is a remarkable fact that the spores of certain cryptogamous plants are of this kind, as we see in the inflammable character of the dry spores of Lycopodium; and we know that the slow putrefaction of such material underground effects chemical changes by which bituminous matter can be produced. There is, therefore, nothing unreasonable in the supposition advanced by Prof. Orton, that the spores so abundantly contained in the Ohio black shales are important or principal sources of the bituminous matter which they contain. Microscopic sections of this shale show that much of its material consists of the rich bituminous matter of these spores (Fig. 16). At the same time, while we may trace the bitumen of these shales, and of some beds of coal, to this cause, we must bear in mind that there are other kinds of bituminous rocks which show no such structures, and may have derived their combustible material from other kinds of vegetable matter, whether of marine or of land plants. We shall better understand this when we have considered the origin of coal.
The macrospores above referred to may have belonged to humble aquatic plants mantling the surfaces of water or growing up from the bottom, and presenting little aërial vegetation. But there are other Erian plants, as already mentioned, which, while of higher structure, may be of Rhizocarpean affinities.
One of these is the beautiful plant with whorls of wedge-shaped leaves, to which the name Sphenophyllum (see Fig. 20) has been given. Plants referred to this genus have been described by Lesquereux from the upper part of the Siluro-Cambrian,[AP] and a beautiful little species occurs in the Erian shales of St. John, New Brunswick.[AQ] The genus is also continued, and is still more « 62 » abundant, in the Carboniferous. Many years ago I observed, in a beautiful specimen collected by Sir W. E. Logan, in New Brunswick, that the stem of this plant had an axis of reticulated and scalariform vessels, and an outer bark.[AR] Renault and Williamson have more recently obtained more perfect specimens, and the former has figured a remarkably complex triangular axis, containing punctate and barred vessels, and larger punctate vessels filling in its angles. Outside of this there is a cellular inner bark, and this is surrounded by a thick fibrous envelope. That a structure so complex should belong to a plant so humble in its affinities is one of the strange anomalies presented by the old world, and of which we shall find many similar instances. The fruit of Sphenophyllum was borne in spikes, with little whorls of bracts or rudimentary leaves bearing round sporocarps.
[AP] “American Journal of Science.”
[AQ] Dawson, “Report on Devonian Plants,” 1870.
[AR] “Journal of the Geological Society,” 1865.
A second type of plant, which may have been Rhizocarpean in its affinities, is that to which I have given the name Ptilophyton.[AS] It consists of beautiful feathery « 63 » fronds, apparently bearing on parts of the main stem or petiole small rounded sporocarps. They are found abundantly in the Middle Erian of the State of New York, and also occur in Scotland, while one species appears to occur in Nova Scotia, as high as the Lower Carboniferous (Figs. 17, 18).
[AS] Plumalina of Hall.
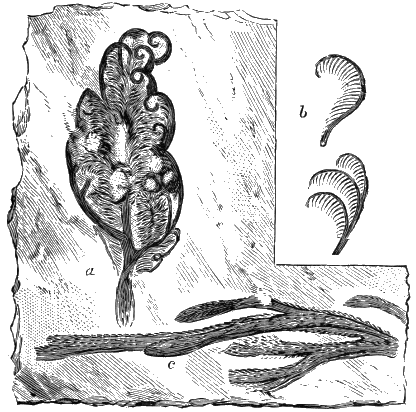
These organisms have been variously referred to Lycopods, to Algæ, or to Zoöphytes, but an extended comparison of American and Scottish specimens has led me to the belief that they were aquatic plants, more likely to have been allied to Rhizocarps than to any other group. Some evidence of this will be given in a note appended to this chapter.
Another genus, which I have named Psilophyton[AT] (Figs. 19, 21), may be regarded as a connecting link between the Rhizocarps and the Lycopods. It is so named from its resemblance, in some respects, to the curious parasitic Lycopods placed in the modern genus Psilotum. Several species have been described, and they are eminently characteristic of the Lower Erian, in which they were first discovered in Gaspé. The typical species, Psilophyton princeps, which fills many beds of shale and sandstone in Gaspé Bay and the head of the neighbouring Bay des Chaleurs with its slender stems and creeping, cord-like rhizomes, may be thus described:
[AT] “Journal of the Geological Society,” vols, xv., xviii., and xix., “Report on Devonian Plants of Canada,” 1871.
Stems branching « 65 » dichotomously, and covered with interrupted ridges. Leaves rudimentary, or short, rigid, and pointed; in barren stems, numerous and spirally arranged; in fertile stems and branchlets, sparsely scattered or absent; in decorticated specimens, represented by a minute punctate scars. Young branches circinate; rhizomata cylindrical, covered with hairs or ramenta, and having circular areoles irregularly disposed, giving origin to slender cylindrical rootlets. Internal structure—an axis of scalariform vessels, surrounded by a cylinder of parenchymatous cells, and by an outer cylinder of elongated woody cells. Fructification consisting of naked oval spore-cases, borne usually in pairs on slender, curved pedicels, either lateral or terminal.
This species was fully described by me in the papers referred to above, from specimens obtained from the rich exposures at Gaspé Bay, and which enabled me to illustrate its parts more fully, perhaps, than those of any other species of so great antiquity. In the specimens I had obtained I was able to recognise the forms of the rhizomata, stems, branches, and rudimentary leaves, and also the internal structure of the stems and rhizomata, and to illustrate the remarkable resemblance of the forms and structures to those of the modern Psilotum. The fructification was, however, altogether peculiar, consisting of narrowly ovate sporangia, borne usually in pairs, on curved and apparently rigid petioles. Under the microscope these sporangia show indications of cellular structure, and appear to have been membranous in character. In some specimens dehiscence appears to have taken place by a slit in one side, and, clay having entered into the interior, both walls of the spore-case can be seen. In other instances, being flattened, they might be mistaken « 66 » for scales. No spores could be observed in any of the specimens, though in some the surface was marked by slight, rounded prominences, possibly the impressions of the spores within. This peculiar and very simple style of spore-case is also characteristic of other species, and gives to Psilophyton a very distinct generic character. These naked spore-cases may be compared to those of such lycopodiaceous plants as Psilotum, in which the « 67 » scales are rudimentary. They also bear some resemblance, though on a much larger scale, to the spore-cases of some Erian ferns (Archæopteris), to be mentioned in the sequel. On the whole, however, they seem most nearly related to the sporocarps of the Rhizocarpeæ.
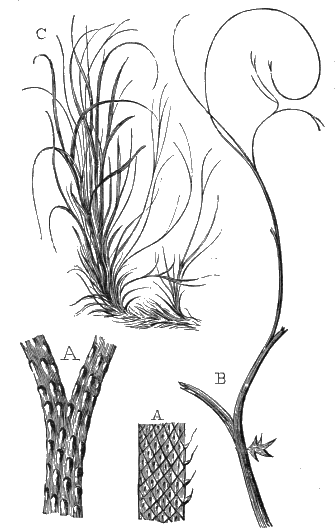
Arthrostigma, which is found in the same beds with Psilophyton, was a plant of more robust growth, with better-developed, narrow, and pointed leaves, borne in a verticillate or spiral manner, and bearing at the ends of its branches spikes of naked sporocarps, apparently similar to those of Psilophyton but more rounded in form. The two genera must have been nearly related, and the slender branchlets of Arthrostigma are, unless well preserved, scarcely distinguishable from the stems of Psilophyton.[AU]
[AU] Reports of the author on “Devonian Plants,” “Geological Survey of Canada,” which see for details as to Erian Flora of northeastern America.
If, now, we compare the vegetation of these and similar ancient plants with that of modern Rhizocarps, we shall find that the latter still present, though in a depauperated and diminished form, some of the characteristics of their predecessors. Some, like Pilularia, have simple linear leaves; others, like Marsilea, have leaves in verticils and cuneate in form; while others, like Azolla and Salvinia, have frondose leaves, more or less pinnate in their arrangement. The first type presents little that is characteristic, but there are in the Erian sandstones and shales great quantities of filamentous and linear objects which it has been impossible to refer to any genus, and which might have belonged to plants of the type of Pilularia. It is quite possible, also, that such plants as Psilophyton glabrum and Cordaites angustifolia, of which the fructification is quite unknown, may have been allied to Rhizocarps. With regard to the verticillate type, we are at once reminded of Sphenophyllum (Fig. 20), which « 68 » many palæobotanists have referred to the Marsiliacæ, though, like other Palæozoic Acrogens, it presents complexities not seen in its modern representatives. S. primævum of Lesquereux is found in the Hudson River group, and my S. antiquum in the Middle Erian. Besides these, there are in the Silurian and Erian beds plants with verticillate leaves which have been placed with the Annulariæ, but which may have differed from them in fructification. Annularia laxa, of the Erian, and Protannularia Harknessii, of the Siluro-Cambrian, may be given as examples, and must have been aquatic plants, probably allied to Rhizocarps. It is deserving of notice, also, that the two best-known species of Psilophyton (P. princeps and P. robustius), while allied to Lycopods by the structure of the stem and such rudimentary foliage as they possess, are also allied, by the form of their fructification, to the Rhizocarps, and not to ferns, as some palæobotanists have incorrectly supposed. A similar remark applies to Arthrostigma; and the beautiful pinnately leaved Ptilophyton may be taken to represent that type of foliage as seen in modern Rhizocarps, while the allied forms of the Carboniferous which Lesquereux has named Trochophyllum, seem to have had sporocarps attached to the stem in the manner of Azolla.
The whole of this evidence, I think, goes to show that in the Erian period there were vast quantities of aquatic plants, allied to the modern Rhizocarps, and that the so-called Sporangites referred to in this paper were probably the drifted sporocarps and macrospores of some of these plants, or of plants allied to them in structure and habit, of which the vegetative organs have perished. I have shown that in the Erian period there were vast swampy flats covered with Psilophyton, and in similar submerged tracts near to the sea the Protosalvinia may have filled the waters and have given off the vast multitudes of macrospores which, drifted by currents, have settled in the « 69 » mud of the black shales. We have thus a remarkable example of a group of plants reduced in modern times to a few insignificant forms, but which played a great role in the ancient Palæozoic world.
Leaving the Rhizocarps, we may now turn to certain other families of Erian plants. The first to attract our attention in this age would naturally be the Lycopods, the club-mosses or ground-pines, which in Canada and the Eastern States carpet the ground in many parts of our woods, and are so available for the winter decoration of our houses and public buildings. If we fancy one of these humble but graceful plants enlarged to the dimensions of a tree, we shall have an idea of a Lepidodendron, or of any of its allies (Figs. 15, 21). These large lycopodiaceous trees, which in different specific and generic forms were probably dominant in the Erian woods, resembled in general those of modern times in their fruit and foliage, except that their cones were large, and probably in most cases with two kinds of spores, and their leaves were also often very long, thus bearing a due proportion to the trees which they clothed. Their thick stems required, however, more strength than is necessary in their diminutive successors, and to meet this want some remarkable structures were introduced similar to those now found only in the stems of plants of higher rank. The cells and vessels of all plants consist of thin walls of woody matter, enclosing the sap and other contents of these sacs and tubes, and when strength is required it is obtained by lining their interior with successive coats of the hardest form of woody matter, usually known as lignin. But while the walls remain thin, they afford free passage to the sap to nourish every part. If thickened all over, they would become impervious to sap, and therefore unsuited to one of their most important functions. These two ends of strength and permeability are secured by partial linings of lignin, leaving portions of « 70 » the original wall uncovered. But this may be done in a great variety of ways.
The most ancient of these contrivances, and one still continued in the world of plants, is that of the barred or scalariform vessel. This may be either square or hexagonal, so as to admit of being packed without leaving vacancies. It is strengthened by a thick bar of ligneous matter up each angle, and these are connected by cross-bars so as to form a framework resembling several ladders fastened together. Hence the name scalariform, or ladder-like. Now, in a modern Lycopod there is a central axis of such barred vessels associated with simpler fibres or elongated cells. Even in Sphenophyllum and Psilophyton, already referred to as allied to Rhizocarps,[AV] there is such a central axis, and in the former rigidity is given to this by the vascular and woody elements being arranged in the form of a three-sided prism or three-rayed star. But such arrangements would not suffice for a tree, and hence in the arboreal Lycopods of the Erian age a more complex structure is introduced. The barred vessels were expanded in the first instance into a hollow cylinder filled in with pith or cellular tissue, and the outer rind was strengthened with greatly thickened cells. But even this was not sufficient, and in the older stems wedge-shaped bundles of barred tissue were run out from the interior, forming an external woody cylinder, and inside of the rind were placed bundles of tough bast fibres. Thus, a stem was constructed having pith, wood, and bark, and capable of additions to the exterior of the woody wedges by a true exogenous growth. The plan is, in short, the same with that of the stems of the exogenous trees of modern times, except that the tissues employed are less complicated. The structures of these remarkable « 71 » trees, and the manner in which they anticipate those of the true exogens of modern times, have been admirably illustrated by Dr. Williamson, of Manchester. His papers, it is true, refer to these plants as existing in the Carboniferous age, but there is every reason to believe that they were of the same character in the Erian. The plan is the same with that now seen in the stems of exogenous phænogams, and which has long ceased to be used in those of the Lycopods. In this way, however, large and graceful lycopodiaceous trees were constructed in the Erian period, and constituted the staple of its forests.
[AV] First noticed by the author, “Journal of Geological Society,” 1865; but more completely by Renault, “Comptes Rendus,” 1870.
The roots of these trees were equally remarkable with their stems, and so dissimilar to any now existing that botanists were long disposed to regard them as independent plants rather than roots. They were similar in general structure to the stems to which they belonged, but are remarkable for branching in a very regular manner by bifurcation like the stems above, and for the fact that their long, cylindrical rootlets were arranged in a spiral manner and distinctly articulated to the root after the manner of leaves rather than of rootlets, and fitting them for growing in homogeneous mud or vegetable muck. They are the so-called Stigmaria roots, which, though found in the Erian and belonging to its lycopodiaceous plants, attained to far greater importance in the Carboniferous period, where we shall meet with them again.
There were different types of lycopodiaceous plants in the Erian. In addition to humble Lycopods like those of our modern woods and great Lepidodendra, which were exaggerated Lycopods, there were thick-stemmed and less graceful species with broad rhombic scars (Leptophleum), and others with the leaf-scars in vertical rows (Sigillaria), and others, again, with rounded leaf-scars, looking like the marks on Stigmaria, and belonging to the genus Cyclostigma. Thus some variety was given to the arboreal club-mosses of these early forests. (See Fig. 15.)
Another group of plants which attained to great development in the Erian age is that of the Ferns or Brackens. The oldest of these yet known are found in the Middle Erian. The Eopteris of Saporta, from the Silurian, at one time supposed to carry this type much further back, has unfortunately been found to be a mere imitative form, consisting of films of pyrites of leaf-like shapes, and produced by crystallisation. In the Middle Erian, however, more especially in North America, many species have been found (Figs. 22 to 24).[AW] I have myself recorded more than thirty species from the Middle Erian of Canada, and these belong to several of the genera found in the Carboniferous, though some are peculiar to the Erian. Of the latter, the best known are perhaps those of the genus Archæopteris (Fig. 24), so abundant in the plant-beds of Kiltorcan in Ireland, as well as in North America. In this genus the fronds are large and luxuriant, with broad obovate pinnules decurrent, on the leaf-stalk, and with simple sac-like spore-cases borne on modified pinnæ. Another very beautiful fern found with Archæopteris is that which I have named Platyphyllum, and which grew on a creeping stem or parasitically on stems of other plants, and had marginal fructification.[AX] Another very remarkable fern, which some botanists have supposed may belong to a higher group than the ferns, is Megalopteris (Fig. 26).
[AW] For descriptions of these ferns, see reports cited above.
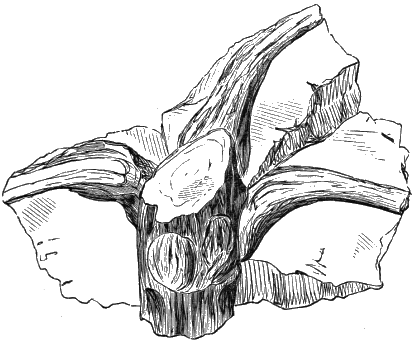
Some of the Erian ferns attained to the dimensions of tree-ferns. Large stems of these, which must have floated out far from land, have been found by Newberry in the marine limestone of Ohio (Caulopteris antiqua and C. peregrina, Newberry),[AY] and Prof. Hall has found in the Upper Devonian of Gilboa, New York, the remains of a forest of tree-ferns standing in situ with their great masses of aërial roots attached to the soil in which they grew (Caulopteris Lockwoodi, Dn.).[AZ]
[AY] “Journal of the Geological Society,” 1871.
[AZ] Ibid.
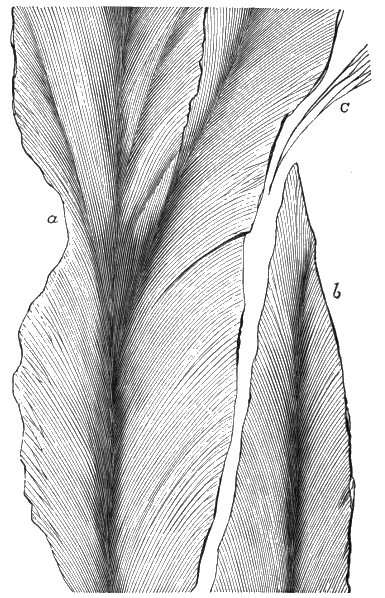
These aërial roots introduce us to a new contrivance for strengthening the stems of plants by sending out into the soil multitudes of cord-like cylindrical roots from « 76 » various heights on the stem, and which form a series of stays like the cordage of a ship. This method of support still continues in the modern tree-ferns of the tropics and the southern hemisphere. In one kind of tree-fern « 77 » stem from the Erian of New York, there is also a special arrangement for support, consisting of a series of peculiarly arranged radiating plates of scalariform vessels, not exactly like those of an exogenous stem, but doing duty for it (Asteropteris)[BA] Similar plants have been described from the Erian of Falkenberg, in Germany, and of Saalfeld, in Thuringia, by Goeppert and Unger, and are referred to ferns by the former, but treated as doubtful by the latter,[BB] This peculiar type of tree-fern is apparently a precursor of the more exogenous type of Heterangium, recently described and referred to ferns by Williamson. Here, again, we have a mechanical contrivance now restricted to higher plants appropriated by these old cryptogams.
[BA] “Journal of the Geological Society,” London, 1881.
[BB] “Sphenopteris Refracta,” Goeppert; “Flora des Uebergangsgebirges.” “Cladoxylon Mirabile,” Unger; “Palæontologie des Thuringer Waldes.”
The history of the ferns in geological time is remarkably different from that of the Lycopods; for while the latter have long ago descended from their pristine eminence to a very humble place in nature, the former still, in the southern hemisphere at least, retain their arboreal dimensions and ancient dominance.
The family of the Equisetaceæ, or mare’s-tails, was also represented by large species of Calamites and by Asterophyllites in the Erian; but, as its headquarters are in the Carboniferous, we may defer its consideration till the next chapter. (Figs. 27, 28.)
Passing over these for the present, we find that the flowering plants are represented in the Erian forests by at least two types of Gymnosperms, that of Taxineæ or yews, and an extinct family, that of the Cordaites (Figs. 30, 31). The yew-trees are closely allied to the pines and spruces, and are often included with them in the family of Coniferæ. They differ, however, in the habit of producing berries or drupe-like fruits instead of cones, and there is some reason to believe that this was the habit of the Erian trees of this group, though their wood in some instances resembles rather that of the Araucaria, or Norfolk « 79 » Island pine, than that of the modern yews. These trees are chiefly known to us by their mineralised trunks, which are often found like drift-wood on modern sand-banks embedded in the Erian sandstones or limestones. It often shows its structure in the most perfect manner in specimens penetrated by calcite or silica, or by pyrite, and in which the original woody matter has been resolved into anthracite or even into graphite. These trees have true woody tissues presenting that beautiful arrangement of pores or thin parts enclosed in cup-like discs, which is characteristic of the coniferous trees, and which is a great improvement on the barred tissue already referred to, affording a far more strong, tough, and durable wood, such as we have in our modern pines and yews (Fig. 29).
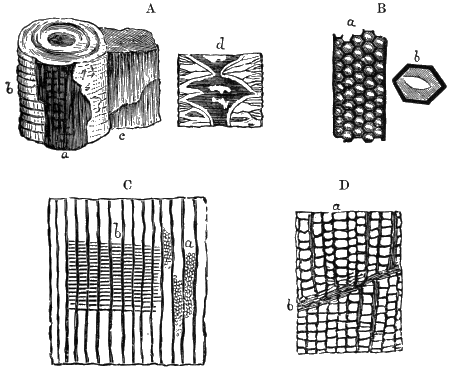
These primitive pines make their appearance in the Middle Erian, in various parts of America, as well as in Scotland and Germany, and they are represented by wood indicating the presence of several species. I have myself indicated and described five species from the Erian of Canada and the United States. From the fact that these trees are represented by drifted trunks embedded in sandstones and marine limestones, we may, perhaps, infer that they grew on the rising grounds of the Erian land, and that their trunks were carried by river-floods into the sea. No instance has yet certainly occurred of the discovery of their foliage or fruit, though there are some fan-shaped leaves usually regarded as ferns which may have belonged to such trees. These in that case would have resembled the modern Gingko of China, and some of the fruits referred to the genus Cardiocarpum may have been produced by them. Various names have been given to these trees. I have preferred that given by Unger, Dadoxylon, as being more non-committal as to affinities than the others.[BC] Many of these trees had very long internal pith-cylinders, with curious transverse tubulæ, and which, when preserved separately, have been named Sternbergia.
[BC] Araucarites, Goeppert; Araucarioxylon, Kraus.
Allied to these trees, and perhaps intermediate between them and the Cycads, were those known as Cordaites (Fig. 30), which had trunks resembling those of Dadoxylon, but with still larger Sternbergia piths and an internal axis of scalariform vessels, surrounded by a comparatively thin woody cylinder. Some of them have leaves over a foot in length, reminding one of the leaves of broad-leaved grasses or iridaceous plants. Yet their flowers and fruit seem to have been more nearly allied to the yews than to any other plants (Fig. 31). Their stems were less woody « 81 » and their piths larger than in the true pines, and some of the larger-leaved species must have had thick, stiff branches. They are regarded as constituting a separate family, intermediate between pines and cycads, and, beginning in the Middle Devonian, they terminate in the Permian, where, however, some of the most gigantic species occur. In so far as the form and structure of the leaves, stems, and fruit are concerned, there is marvellously little difference between the species found in the Erian and the Permian. They culminated, however, in the Carboniferous period, and the coal-fields of southern France have proved so far the richest in their remains.
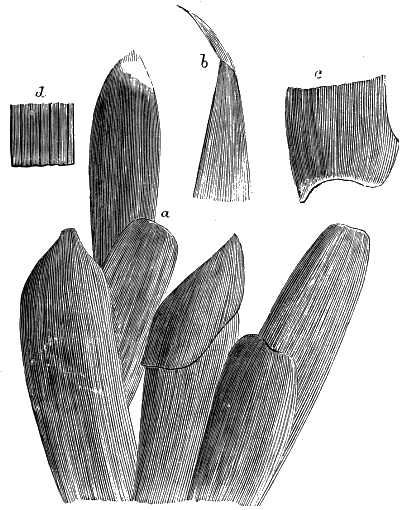
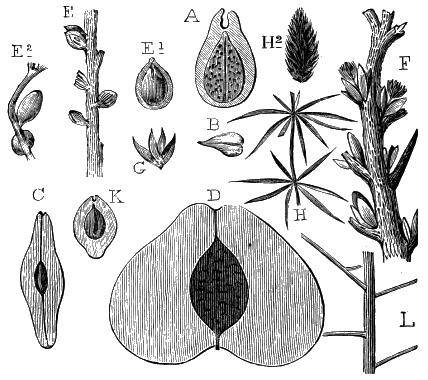
Lastly, a single specimen, collected by Prof. James Hall, of Albany, at Eighteen-mile Creek, Lake Erie, has the structure of an ordinary angiospermous exogen, and has been described by me as Syringoxylon mirabile.[BD] This unique example is sufficient to establish the fact of the existence of such plants at this early date, unless some accident may have carried a specimen from a later formation « 83 » to be mixed with Erian fossils. It is to be observed, however, that the non-occurrence of any similar wood in all the formations between the Upper Erian and the Middle Cretaceous suggests very grave doubt as to the authenticity of the specimen. I record the fact, waiting further discoveries to confirm it. Of the character of the specimen which I have described I entertain no doubt.
[BD] “Journal of the Geological Society,” vol. xviii.
We shall be better able to realise the significance and relations of this ancient flora when we have studied that of the succeeding Carboniferous. We may merely remark here on the fact that, in these forests of the Devonian and in the marshes on their margins, we find a wonderful expansion of the now modest groups of Rhizocarps and Lycopods, and that the flora as a whole belongs to the highest group of Cryptogams and the lowest of Phænogams, so that it has about it a remarkable aspect of mediocrity. Further, while there is evidence of some variety of station, there is also evidence of much equality of climate, and of a condition of things more resembling that of the insular climates of the temperate portions of the southern hemisphere than that of North America or Europe at present.
The only animal inhabitants of these Devonian woods, so far as known, were a few species of insects, discovered by Hartt in New Brunswick, and described by Dr. Scudder. Since, however, we now know that scorpions as well as insects existed in the Silurian, it is probable that these also occurred in the Erian, though their remains have not yet been discovered. All the known insects of the Erian woods are allies of the shad-flies and grasshoppers (Neuroptera and Orthoptera), or intermediate between the two. It is probable that the larvæ of most of them lived in water and fed upon the abundant vegetable matter there, or on the numerous minute crustaceans and worms. There were no land vertebrates, so far as known, but there were fishes (Dipterus, etc.), allied to the modern « 84 » Barramunda or Ceratodus of Australia, and with teeth suited for grinding vegetable food. It is also possible that some of the smaller plate-covered fishes (Placoganoids, like Pterichthys) might have fed on vegetable matter, and, in any case, if they fed on lower animals, the latter must have subsisted on plants. I mention these facts to show that the superabundant vegetation of this age, whether aquatic or terrestrial, was not wholly useless to animals. It is quite likely, also, that we have yet much to learn of the animal life of the Erian swamps and woods.
I.—Classification of Sporangites.
It is, of course, very unsatisfactory to give names to mere fragments of plants, yet it seems very desirable to have some means of arranging them. With respect to the organisms described above, which were originally called by me Sporangites, under the supposition that they were Sporangia rather than spores, this name has so far been vindicated by the discovery of the spore-cases belonging to them, so that I think it may still be retained as a provisional name; but I would designate the whole as Protosalviniæ, meaning thereby plants with rhizocarpean affinities, though possibly when better understood belonging to different genera. We may under these names speak of their detached discs as macrospores and of their cellular envelopes as sporocarps. The following may be recognized as distinct forms:
1. Protosalvinia Huronensis, Dawson, Syn., Sporangites Huronensis, “Report on Erian Flora of Canada,” 1871.—Macrospores, in the form of discs or globes, smooth and thick-walled, the walls penetrated by minute radiating pores. Diameter about one one-hundredth of an inch, or a little more, When in situ several macrospores are contained in a thin cellular sporocarp, probably globular in form. From the Upper Erian, and perhaps Lower Carboniferous shales of Kettle Point, Lake Huron, of various places in the State of Ohio, and in the shale boulders of the boulder clay of Chicago and vicinity. First collected at Kettle Point by Sir W. E. Logan, and « 85 » in Ohio by Prof. Edward Orton, and at Chicago by Dr. H. A. Johnson and Mr. B. W. Thomas, also in New York by Prof. J. M. Clarke.
The macrospores collected by Mr. Thomas from the Chicago clays and shales conform closely to those of Kettle Point, and probably belong to the same species. Some of them are thicker in the outer wall, and show the pores much more distinctly. These have been called by Mr. Thomas S. Chicagoensis, and may be regarded as a varietal form. Specimens isolated from the shale and mounted dry, show what seems to have been the hilum or scar of attachment better than those in balsam.
Sections of the Kettle Point shale show, in addition to the macrospores, wider and thinner shreds of vegetable matter, which I am inclined to suppose to be remains of the sporocarps.
2. Protosalvinia (Sporangites) Braziliensis, Dawson, “Canadian Record of Science,” 1883.—Macrospores, round, smooth, a little longer than those of the last species, or about one seventy-fifth of an inch in diameter, enclosed in round, oval, or slightly reniform sporocarps, each containing from four to twenty-four macrospores. Longest diameter of sporocarps three to six millimetres. Structure of wall of sporocarps hexagonal cellular. Some sporocarps show no macrospores, and may possibly contain microspores. The specimens are from the Erian of Brazil. Discovered by Mr. Orville Derby. The formation, according to Mr. Derby, consists of black shales below, about three hundred feet thick, and containing the fucoid known as Spirophyton, and probably decomposed vegetable matter. Above this is chocolate and reddish shale, in which the well-preserved specimens of Protosalvinia occur. These beds are very widely distributed, and abound in Protosalvinia and Spirophyton.
3. Protosalvinia (Sporangites) bilobata, Dawson, “Canadian Record of Science,” 1883.—Sporocarps, oval or reniform, three to six millimetres in diameter, each showing two rounded prominences at the ends, with a depression in the middle, and sometimes a raised neck or isthmus at one side connecting the prominences. Structure of sporocarp cellular. Some of the specimens indicate that each prominence or tubercle contained several macrospores. At first sight it would be easy to mistake these bodies for valves of Beyrichia.
Found in the same formations with the last species, though, in so far as the specimens indicate, not precisely in the same beds. Collected by Mr. Derby.
4. Protosalvinia Clarkei, Dawson, P. bilobata, Clarke, “American Journal of Science.”—Macrospores two-thirds to one millimetre in « 86 » diameter. One, two, or three contained in each sporocarp, which is cellular. The macrospores have very thick walls with radiating tortuous tubes. Unless this structure is a result of mineral crystallisation, these macrospores must have had very thick walls and must have resembled in structure the thickened cells of stone fruits and of the core of the pear, or the tests of the Silurian and Erian seeds known as Pachytheca, though on a smaller scale.
It is to be observed that bodies similar to these occur in the Boghead earthy bitumen, and have been described by Credner.
I have found similar bodies in the so-called “Stellar coal” of the coal district of Pictou, Nova Scotia, some layers of which are filled with them. They occur in groups or patches, which seem to be enclosed in a smooth and thin membrane or sporocarp. It is quite likely that these bodies are generically distinct from Protosalvinia.
5. Protosalvinia punctata, Newton, “Geological Magazine,” New Series, December 2d, vol. ii.—Mr. Newton has named the discs found in the white coal and Tasmanite, Tasmanites, the species being Tasmanites punctatus, but as my name Sporangites had priority, I do not think it necessary to adopt this term, though there can be little doubt that these organisms are of similar character. The same remark may be made with reference to the bodies described by Huxley and Newton as occurring in the Better-bed coal.
In Witham’s “Internal Structure of Fossil Vegetables,” 1833, Plate XI, are figures of Lancashire cannel which shows Sporangites of the type of those in the Erian shales. Quekett, in his “Report on the Torbane Hill Mineral,” 1854, has very well figured similar structures from the Methel coal and the Lesmahagow cannel coal. These are the earliest publications on the subject known to me; and Quekett, though not understanding the nature of the bodies he observed, holds that they are a usual ingredient in cannel coals.
II.—The Nature and Affinities of Ptilophyton.
(Lycopodites Vanuxemii of “Report on Devonian and Upper Silurian Plants,” Part I., page 35, L. plumula of “Report on Lower Carboniferous Plants,” page 24, Plate I., Figs. 7, 8, 9.) In the reports above referred to, these remarkable pinnate, frond-like objects were referred to the genus Lycopodites, as had been done by Goeppert in his description of the European species Lycopodites pennæformis, which is very near to the American Erian form. Since 1871, however, there have been many new specimens obtained, and very various opinions expressed as to their affinities. While Hall has named some of them Plumalina, and has regarded them as animal « 87 » structures, allied to hydroids, Lesquereux has described some of the Carboniferous forms under the generic name Trochophyllum, which is, however, more appropriate to plants with verticillate leaves which are included in this genus. Before I had seen the publications of Hall and Lesquereux on the subject, I had in a paper on “Scottish Devonian Plants”[BE] separated this group from the genus Lycopodites, and formed for it the genus Ptilophyton, in allusion to the feather-like aspect of the species. My reasons for this, and my present information as to the nature of these plants, may be stated as follows: Schimper, in his “Palæontologie Vegetale” (possibly from inattention to the descriptions or want of access to specimens), doubts the lycopodiaceous character of species of Lycopodites described in my published papers on plants of the Devonian of America and in my Report of 1871. Of these, L. Richardsoni and L. Matthewi are undoubtedly very near to the modern genus Lycopodium. L. Vanuxemii is, I admit, more problematical; but Schimper could scarcely have supposed it to be a fern or a fucoid allied to Caulerpa had he observed that both in my species and the allied L. pennæformis of Goeppert, which he does not appear to notice, the pinnules are articulated upon the stem, and leave scars where they have fallen off. When in Belfast in 1870, my attention was again directed to the affinities of these plants by finding in Prof. Thomson’s collection a specimen from Caithness, which shows a plant apparently of this kind, with the same long narrow pinna? or leaflets, attached, however, to thicker stems, and rolled up in a circinate manner. It seems to be a plant in vernation, and the parts are too much crowded and pressed together to admit of being accurately figured or described; but I think I can scarcely be deceived as to its true nature. The circinate arrangement in this case would favour a relationship to ferns; but some lycopodiaceous plants also roll themselves in this way, and so do the branches of the plants of the genus Psilophyton. (Fig. 17, supra.)
[BE] “Canadian Naturalist,” 1878.
The specimen consists of a short, erect stem, on which are placed somewhat stout alternate branches, extending obliquely outward and then curving inward in a circinate manner. The lower ones appear to produce on their inner sides short lateral branchlets, and upon these, and also upon the curved extremities of the branches, are long, narrow, linear leaves placed in a crowded manner. The specimen is thus not a spike of fructification, but a young stem or branch in vernation, and which when unrolled would be of the form of those « 88 » peculiar pinnate Lycopodites of which L. Vanuxemii of the American Devonian and L. pennæformis of the European Lower Carboniferous are the types, and it shows, what might have been anticipated from other specimens, that they were low, tufted plants, circinate in vernation. The short stem of this plant is simply furrowed, and bears no resemblance to a detached branch of Lycopodites Milleri which lies at right angles to it on the same slab. As to the affinities of the singular type of plants to which this specimen belongs, I may quote from my “Report on the Lower Carboniferous Plants of Canada,” in which I have described an allied species, L. plumula:
“The botanical relations of these plants must remain subject to doubt, until either their internal structure or their fructification can be discovered. In the mean time I follow Goeppert in placing them in what we must regard as the provisional genus Lycopodites. On the one hand, they are not unlike the slender twigs of Taxodium and similar Conifers, and the highly carbonaceous character of the stems gives some colour to the supposition that they may have been woody plants. On the other hand, they might, so far as form is concerned, be placed with Algæ of the type of Brongniart’s Chondrites obtusus, or the modern Caulerpa plumaria. Again, in a plant of this type from the Devonian of Caithness to which I have referred in a former memoir, the vernation seems to have been circinate, and Schimper has conjectured that these plants may be ferns, which seems also to have been the view of Shumard.”
On the whole, these plants are allied to Lycopods rather than to ferns; and as they constitute a small but distinct group, known only, so far as I am aware, in the Lower Carboniferous and Erian or Devonian, they deserve a generic name, and I proposed for them in my “Paper on Scottish Devonian Plants,” 1878, that of Ptilophyton, a name sufficiently distinct in sound from Psilophyton, and expressing very well their peculiar feather-like habit of growth. The genus was defined as follows:
“Branching plants, the branches bearing long, slender leaves in two or more ranks, giving them a feathered appearance; vernation circinate. Fruit unknown, but analogy would indicate that it was borne on the bases of the leaves or on modified branches with shorter leaves.”
The Scottish specimen above referred to was named Pt. Thomsoni, and was characterised by its densely tufted form and thick branches. The other species known are: Pt. pennæformis, Goeppert, L. Carboniferous; Pt. Vanuxemii, Dawson, Devonian; Pt. plumula, Dawson, L. Carboniferous.
Shumard’s Filicites gracilis, from the Devonian of Ohio, and Stur’s Pinites antecedens, from the Lower Carboniferous of Silesia, may possibly belong to the same genus. The Scottish specimen referred to is apparently the first appearance of this form in the Devonian of Europe.
I have at a still later date had opportunities of studying considerable series of these plants collected by Prof. Williams, of Cornell University, and prepared a note in reference to them for the American Association, of which, however, only an abstract has been published. I have also been favoured by Prof. Lesquereux and Mr. Lacoe, of Pittston, with the opportunity of studying the specimens referred to Trochophyllum.
Prof. Williams’s specimens occur in a dark shale associated with remains of land-plants of the genera Psilophyton. Rhodea, &c., and also marine shells, of which a small species of Rhynchonella is often attached to the stems of the Ptilophyton. Thus these organisms have evidently been deposited in marine beds, but in association with land-plants.
The study of the specimens collected by Prof. Williams develops the following facts: (1) The plants are not continuous fronds, but slender stems or petioles, with narrow, linear leaflets attached in a pinnate manner. (2) The pinnules are so articulated that they break off, leaving delicate transverse scars, and the lower parts of the stems are often thus denuded of pinnæ for the length of one or more inches. (3) The stems curve in such a manner as to indicate a circinate vernation. (4) In a few instances the fronds were observed to divide dichotomously toward the top; but this is rare. (5) There are no indications of cells in the pinnules; but, on the other hand, there is no appearance of fructification unless the minute granules which roughen some of the sterns are of this nature. (6) The stems seem to have been lax and flexuous, and in some instances they seem to have grown on the petioles of ferns preserved with them in the same beds. (7) The frequency of the attachment of small brachiopods to the specimens of Ptilophyton would seem to indicate that the plant stood erect in the water. (8) Some of the specimens show so much carbonaceous matter as to indicate that the pinnules were of considerable consistency. All these characters are those rather of an aquatic plant than of an animal organism or of a land-plant.
The specimens communicated by Prof. Lesquereux and Mr. Lacoe are from the Lower Carboniferous, and evidently represent a different species with similar slender pitted stems, often partially denuded of pinnules below; but the pinnules are much broader and « 90 » more distant. They are attached by very narrow bases, and apparently tend to lie on a plane, though they may possibly have been spirally arranged. On the same slabs are rounded sporangia or macrospores like those of Lepidodendron, but there is no evidence that these belonged to Trochophyllum. On the stems of this plant, however, there are small, rounded bodies apparently taking the places of some of the pinnules. These may possibly be spore-cases; but they may be merely imperfectly developed pinnules. Still the fact that similar small granules appear on the stems of the Devonian species, favours the idea that they may be organs of fructification.
The most interesting discovery, however, which results from the study of Mr. Lacoe’s specimens, is that the pinnules were cylindrical and hollow, and probably served to float the plant. This would account for many of the peculiarities in the appearance and mode of occurrence of the Devonian Ptilophyton, which are readily explained if it is supposed to be an aquatic plant, attaching itself to the stems of submerged vegetable remains and standing erect in the water by virtue of its hollow leaves. It may well, however, have been a plant of higher organisation than the Algæ, though no doubt cryptogamous.
The species of Ptilophyton will thus constitute a peculiar group of aquatic plants, belonging to the Devonian and Lower Carboniferous periods, and perhaps allied to Lycopods and Pillworts in their organisation and fruit, but specially distinguished by their linear leaves serving as floats and arranged pinnately on slender stems. The only species yet found within the limits of Canada is Pt. plumula, found by Dr. Honeyman in the Lower Carboniferous of Nova Scotia; but as Pt. Vanuxemii abounds in the Erian of New York, it will no doubt be found in Canada also.
III.—Tree-Ferns of the Erian Period.
As the fact of the occurrence of true tree-ferns in rocks so old as the Middle Erian or Devonian has been doubted in some quarters, the following summary is given from descriptions published in the “Journal of the Geological Society of London” (1871 and 1881), where figures of the species will be found:
Of the numerous ferns now known in the Middle and Upper Devonian of North America, a great number are small and delicate species, which were probably herbaceous; but there are other species which may have been tree-ferns. Little definite information, however, has, until recently, been obtained with regard to their habit of growth.
The only species known to me in the Devonian of Europe is the Caulopteris Peachii of Salter, figured in the “Quarterly Journal of the Geological Society” for 1858. The original specimen of this I had an opportunity of seeing in London, through the kindness of Mr. Etheridge, and have no doubt that it is the stem of a small arborescent fern, allied to the genus Caulopteris, of the coal formation.
In my paper on the Devonian of Eastern America (“Quarterly Journal of the Geological Society,” 1862), I mentioned a plant found by Mr. Richardson at Perry, as possibly a species of Megaphyton, using that term to denote those stems of tree-ferns which have the leaf-scars in two vertical series; but the specimen was obscure, and I have not yet obtained any other.
More recently, in 1869, Prof. Hall placed in my hands an interesting collection from Gilboa, New York, and Madison County, New York, including two trunks surrounded by aërial roots, which I have described as Psaronius textilis and P. Erianus, in my “Revision of the Devonian Flora,” read before the Royal Society.[BF] In the same collection were two very large petioles, Rhachiopteris gigantea and R. palmata, which I have suggested may have belonged to tree-ferns.
[BF] Abstract in “Proceedings of the Royal Society,” May, 1870; also “Report on Erian Plants of Canada,” 1871.
My determination of the species of Psaronius, above mentioned, has recently been completely confirmed by the discovery on the part of Mr. Lockwood, of Gilboa, of the upper part of one of these stems, with its leaf-scars preserved and petioles attached, and also by some remarkable specimens obtained by Prof. Newberry, of New York, from the Corniferous limestone of Ohio, which indicate the existence there of three species of tree-ferns, one of them with aërial roots similar to those of the Gilboa specimens. The whole of these specimens Dr. Newberry has kindly allowed me to examine, and has permitted me to describe the Gilboa specimen, as connected with those which I formerly studied in Prof. Hall’s collections. The specimens from Ohio he has himself named, but allows me to notice them here by way of comparison with the others. I shall add some notes on specimens found with the Gilboa ferns.
It may be further observed that the Gilboa specimens are from a bed containing erect stumps of tree-ferns, in the Chemung group of the Upper Devonian, while those from Ohio are from a marine limestone, belonging to the lower part of the Middle Devonian.
1. Caulopteris Lockwoodi, Dawson.—Trunk from two to three « 92 » inches in diameter, rugose longitudinally. Leaf-scars broad, rounded above, and radiatingly rugose, with an irregular scar below, arranged spirally in about five ranks; vascular bundles not distinctly preserved. Petioles slender, much expanded at the base, dividing at first in a pinnate manner, and afterwards dichotomously. Ultimate pinnæ with remains of numerous, apparently narrow pinnules.
This stem is probably the upper part of one or other of the species of Psaronius found in the same bed (P. Erianus, Dawson, and P. textilis, Dawson).[BG] It appears to have been an erect stem embedded in situ in sandstone, and preserved as a cast. The stem is small, being only two inches, or a little more, in diameter. It is coarsely wrinkled longitudinally, and covered with large leaf-scars, each an inch in diameter, of a horseshoe-shape. The petioles, five of which remain, separate from these scars with a distinct articulation, except at one point near the base, where probably a bundle or bundles of vessels passed into the petiole. They retain their form at the attachment to the stem, but a little distance from it they are flattened. They are inflated at the base, and somewhat rapidly diminish in size. The leaf-scars vary in form, and are not very distinct, but they appear to present a semicircular row of pits above, largest in the middle. From these there proceed downward a series of irregular furrows, converging to a second and more obscure semicircle of pits, within or below which is the irregular scar or break above referred to. The attitude and form of the petioles will be seen from Fig. 24, supra.
[BG] Memoir on Devonian Flora, “Proceedings of the Royal Society,” May, 1870.
The petioles are broken off within a few inches of the stem; but other fragments found in the same beds appear to show their continuation, and some remains of their foliage. One specimen shows a series of processes at the sides, which seem to be the remains of small pinnæ, or possibly of spines on the margin of the petiole. Other fragments show the division of the frond, at first in a pinnate manner, and subsequently by bifurcation; and some fragments show remains of pinnules, possibly of fertile pinnules. These are very indistinct, but would seem to show that the plant approached, in the form of its fronds and the arrangement of its fructification, to the Cyclopterids of the sub-genus Aneimites, one of which (Aneimites Acadica), from the Lower Carboniferous of Nova Scotia, I have elsewhere described as probably a tree-fern,[BH] The « 93 » fronds were evidently different from those of Archæopteris[BI] a genus characteristic of the same beds, but of very different habit of growth. This accords with the fact that there is in Prof. Hall’s collection a mass of fronds of Cyclopteris (Archæopteris) Jacksoni, so arranged as to make it probable that the plant was an herbaceous fern, producing tufts of fronds on short stems in the ordinary way. The obscurity of the leaf-scars may render it doubtful whether the plant above described should be placed in the genus Caulopteris or in Stemmatopteris; but it appears most nearly allied to the former. The genus is at present, of course, a provisional one; but I have thought it only justice to the diligent labours of Mr. Lockwood to name this curious and interesting fossil Caulopteris Lockwoodi.
[BH] “Quarterly Journal of the Geological Society,” 1860.
[BI] The genus to which the well-known Cyclopteris (Adiantites) Hibernicus of the Devonian of Ireland belongs.
I have elsewhere remarked on the fact that trunks, and petioles, and pinnules of ferns are curiously dissociated in the Devonian beds—an effect of water-sorting, characteristic of a period in which the conditions of deposition were so varied. Another example of this is, that in the sandstones of Gaspé Bay, which have not as yet afforded any example of fronds of ferns, there are compressed trunks, which Mr. Lockwood’s specimens allow me at least to conjecture may have belonged to tree-ferns, although none of them are sufficiently perfect for description.
Mr. Lockwood’s collection includes specimens of Psaronius textilis; and in addition to these there are remains of erect stems somewhat different in character, yet possibly belonging to the higher parts of the same species of tree-fern. One of these is a stem crushed in such a manner that it does not exhibit its form with any distinctness, but surrounded by smooth, cylindrical roots, radiating from it in bundles, proceeding at first horizontally, and then curving downward, and sometimes terminating in rounded ends. They resemble in form and size the aërial roots of Psaronius Erianus; and I believe them to be similar roots from a higher part of the stem, and some of them young and not prolonged sufficiently far to reach the ground. This specimen would thus represent the stem of P. Erianus at a higher level than those previously found. We can thus in imagination restore the trunk and crown of this once graceful tree-fern, though we have not the detail of its fronds. Mr. Lockwood’s collections also contain a specimen of the large fern-petiole which I have named Rhachiopteris punctata. My original specimen was obtained by Prof. Hall from the same horizon in New York. « 94 » That of Mr. Lockwood is of larger size, but retains no remains of the frond. It must have belonged to a species quite distinct from Caulopteris Lockwoodi, but which may, like it, have been a tree-fern.
2. Caulopteris antiqua, Newberry.—This is a flattened stem, on a slab of limestone, containing Brachiopods, Trilobites, &c., of the Corniferous limestone. It is about eighteen inches in length, and three and a half inches in average breadth. The exposed side shows about twenty-two large leaf-scars arranged spirally. Each leaf, where broken off, has left a rough fracture; and above this is a semicircular impression of the petiole against the stem, which, as well as the surface of the bases of the petioles, is longitudinally striated or tuberculated. The structures are not preserved, but merely the outer epidermis, as a coaly film. The stem altogether much resembles Caulopteris Peachii, but is of larger size. It differs from C. Lockwoodi in the more elongated leaf-bases, and in the leaves being more remotely placed; but it is evidently of the same general character with that species.
3. Caulopteris (Protopteris) peregrina, Newberry.—This is a much more interesting species than the last, as belonging to a generic or subgeneric form not hitherto recognised below the Carboniferous, and having its minute structure in part preserved.
The specimens are, like the last, on slabs of marine limestone of the Corniferous formation, and flattened. One represents an upper portion of the stem with leaf-scars and remains of petioles; another a lower portion, with aërial roots. The upper part is three inches in diameter, and about a foot in length, and shows thirty leaf-scars which are about three-fourths of an inch wide, and rather less in depth. The upper part presents a distinct rounded and sometimes double marginal line, sometimes with a slight depression in the middle. The lower part is irregular, and when most perfect shows seven slender vascular bundles, passing obliquely downward into the stem. The more perfect leaf-bases have the structure preserved, and show a delicate, thin-walled, oval parenchyma, while the vascular bundles show scalariform vessels with short bars in several rows, in the manner of many modern ferns. Some of the scars show traces of the hippocrepian mark characteristic of Protopteris; and the arrangement of the vascular bundles at the base of the scars is the same as in that genus, as are also the general form and arrangement of the scars. On careful examination, the species is indeed very near to the typical P. Sternbergii, as figured by Corda and Schimper.[BJ]
The genus Protopteris of Sternberg, though the original species (P. punctata) appears as a Lepidodendron in his earlier plate (Plate 4), and as a Sigillaria (S. punctata) in Brongniart’s great work, is a true tree-fern; and the structure of one species (P. Cottai) has been beautifully figured by Corda. The species hitherto described are from the Carboniferous and Permian.
The second specimen of this species represents a lower part of the stem. It is thirteen inches long and about four inches in diameter, and is covered with a mass of flattened aërial roots lying parallel to each other, in the manner of the Psaronites of the coal-formation and of P. Erianus of the Upper Erian or Devonian.
4. Asteropteris noveboracensis, gen. and sp. n.—The genus Asteropteris is established for stems of ferns having the axial portion composed of vertical radiating plates of scalariform tissue embedded in parenchyma, and having the outer cylinder composed of elongated cells traversed by leaf-bundles of the type of those of Zygopteris.
The only species known to me is represented by a stem 2·5 centimetres in diameter, slightly wrinkled and pitted externally, perhaps by traces of aërial roots which have perished. The transverse section shows in the centre four vertical plates of scalariform or imperfectly reticulated tissue, placed at right angles to each other, and united in the middle of the stem. At a short distance from the centre, each of these plates divides into two or three, so as to form an axis of from ten to twelve radiating plates, with remains of cellular tissue filling the angular interspaces. The greatest diameter of this axis is about 1·5 centimetre. Exterior to the axis the stem consists of elongated cells, with somewhat thick walls, and more dense toward the circumference. The walls of these cells present a curious reticulated appearance, apparently caused by the cracking of the ligneous lining in consequence of contraction in the process of carbonization. Embedded in this outer cylinder are about twelve vascular bundles, each with a dumb-bell-shaped group of scalariform vessels enclosed in a sheath of thick-walled fibres. Each bundle is opposite to one of the rays of the central axis. The specimen shows about two inches of the length of the stem, and is somewhat bent, apparently by pressure, at one end.
This stem is evidently that of a small tree-fern of a type, so far as known to me, not before described,[BK] and constituting a very complex and symmetrical form of the group of Palæozoic ferns allied « 96 » to the genus Zygopteris of Schimper. The central axis alone has a curious resemblance to the peculiar stem described by Unger (“Devonian Flora of Thuringia”) under the name of Cladoxylon mirabile; and it is just possible that this latter stem may be the axis of some allied plant. The large aërial roots of some modern tree-ferns of the genus Angiopteris have, however, an analogous radiating structure.
[BK] Prof. Williamson, to whom I have sent a tracing of the structure, agrees with me that it is new.
The specimen is from the collection of Berlin H. Wright, Esq., of Penn Yan, New York, and was found in the Portage group (Upper Erian) of Milo, New York, where it was associated with large petioles of ferns and trunks of Lepidodendra, probably L. Chemungense and L. primævum.
The occurrence of this and other stems of tree-ferns in marine beds has recently been illustrated by the observation of Prof. A. Agassiz that considerable quantities of vegetable matter can be dredged from great depths in the sea on the leeward side of the Caribbean Islands. The occurrence of these trunks further connects itself with the great abundance of large petioles (Rhachiopteris) in the same beds, while the rarity of well-preserved fronds is explained by the coarseness of the beds, and also by the probably long maceration of the plant-remains in the sea-water.
In connection with this I may refer to the remarkable facts recently stated by Williamson[BL] respecting the stems known as Heterangium and Lyginodendron. It would seem that these, while having strong exogenous peculiarities, are really stems of tree-ferns, thus placing this family in the same position of advancement with the Lycopods and Equisetaceæ of the Coal period.
[BL] “Proceedings of the Royal Society,” January 6, 1887.
IV.—On Erian Trees of the Genus Dadoxylon, Unger.
(Araucarites of Goeppert, Araucarioxylon of Kraus.)
Large woody trunks, carbonised or silicified, and showing wood-cells with hexagonal areoles having oval pores inscribed in them, occur abundantly in some beds of the Middle Erian of America, and constitute the most common kind of fossil wood all the way to the Trias. They have in the older formations, generally, several rows of pores on each fibre, and medullary rays composed of two or more series of cells, but become more simple in these respects in the Permian and Triassic series. The names Araucarites and Araucarioxylon are perhaps objectionable, inasmuch as they suppose affinities to Araucaria which may not exist. Unger’s name, which is non-committal, « 97 » is therefore, I think, to be preferred. In my “Acadian Geology,” and in my “Report on the Geology of Prince Edward Island,” I have given reasons for believing that the foliage of some at least of these trees was that known as Walchia, and that they may have borne nutlets in the manner of Taxine trees (Trigonocarpum, &c). Grand d’Eury has recently suggested that some of them may have belonged to Cordaites, or to plants included in that somewhat varied and probably artificial group.
The earliest discovery of trees of this kind in the Erian of America was that of Matthew and Hartt, who found large trunks, which I afterwards described as Dadoxylon Ouangondianum, in the Erian sandstone of St. John, New Brunswick, hence named by those geologists the “Dadoxylon sandstone.” A little later, similar wood was found by Prof. Hall and Prof. Newberry in the Hamilton group of New York and Ohio, and the allied wood of the genus Ormoxylon was obtained by Prof. Hall in the Portage group of the former State. These woods proved to be specifically distinct from that of St. John, and were named by me D. Halli, D. Newberryi, and Ormoxylon Erianum. The three species of Dadoxylon agreed in having composite medullary rays, and would thus belong to the group Palæoxylon of Brongniart. In the case of Ormoxylon this character could not be very distinctly ascertained, but the medullary rays appeared to be simple.
I am indebted to Prof. J. M. Clarke, of Amherst College, Massachusetts, for some well-preserved specimens of another species from the Genesee shale of Canandaigua, New York. They show small steins or branches, with a cellular pith surrounded with wood of coniferous type, showing two to three rows of slit-formed, bordered pores in hexagonal borders. The medullary sheath consists of pseudo-scalariform and reticulated fibres; but the most remarkable feature of this wood is the structure of the medullary rays, which are very frequent, but short and simple, sometimes having as few as four cells superimposed. This is a character not before observed in coniferous trees of so great age, and allies this Middle Erian form with some Carboniferous woods which have been supposed to belong to Cordaites or Sigillaria. In any case this structure is new, and I have named the species Dadoxylon Clarkii, after its discoverer. The specimens occur, according to Prof. Clarke, in a calcareous layer which is filled with the minute shells of Styliola fissurella of Hall, believed to be a Pteropod; and containing also shells of Goniatites and Gyroceras. The stems found are only a few inches in diameter, but may be branches of larger trees.
It thus appears that we already know five species of Coniferous trees of the genus Dadoxylon in the Middle Erian of America, an interesting confirmation of the facts otherwise known as to the great richness and variety of this ancient flora. The late Prof. Goeppert informed me that he had recognised similar wood in the Devonian of Germany, and there can be no doubt that the fossil wood discovered by Hugh Miller in the Old Red Sandstone of Scotland, and described by Salter and McNab, is of similar character, and probably belongs to the genus Dadoxylon. Thus this type of Coniferous tree seems to have been as well established and differentiated into species in the Middle Devonian as in the succeeding Carboniferous.
I may here refer to the fact that the lower limit of the trees of this group coincides, in America, with the upper limit of those problematical trees which in the previous chapter I have named Protogens (Nematophyton, Celluloxlyon,[BM] Nematoxylon[BN]), though Aporoxylon of Unger extends, in Thuringia, up to the Upper Devonian (Cypridina schists).
[BM] “Journal of the Geological Society,” May, 1881.
[BN] Ibid., vol. xix, 1863.
V.—Scottish Devonian Plants of Hugh Miller and others.
(Edinburgh Geological Society, 1877.)
Previously to the appearance of my descriptions of Devonian plants from North America, Hugh Miller had described forms from the Devonian of Scotland, similar to those for which I proposed the generic name Psilophyton; and I referred to these in this connection in my earliest description of that genus.[BO] He had also recognised what seemed to be plants allied to Lycopods and Conifers. Mr. Peach and Mr. Duncan had made additional discoveries of this kind, and Sir J. Hooker and Mr. Salter had described some of these remains. More recently Messrs. Peach, Carruthers, and McNab have worked in this field, and still later[BP] Messrs. Jack and Etheridge have summed up the facts and have added some that are new.
[BO] “Journal of the Geological Society,” London, 1859.
[BP] Ibid., 1877.
The first point to which I shall refer, and which will lead to the other matters to be discussed, is the relation of the characteristic Lepidodendron of the Devonian of eastern America, L. Gaspianum, to L. nothum of Unger and of Salter. At the time when I described this species I had not access to Scottish specimens of Lepidodendron « 99 » from the Devonian, but these had been well figured and described by Salter, and had been identified with L. nothum of Unger, a species evidently distinct from mine, as was also that figured and described by Salter, whether identical or not with Unger’s species. In 1870 I had for the first time an opportunity to study Scottish specimens in the collection of Mr. Peach; and on the evidence thus afforded I stated confidently that these specimens represented a species distinct from L. Gaspianum, perhaps even generically so.[BQ] It differs from L. Gaspianum in its habit of growth by developing small lateral branches instead of bifurcating, and in its foliage by the absence or obsolete character of the leaf-bases and the closely placed and somewhat appressed leaves. If an appearance of swelling at the end of a lateral branch in one specimen indicates a strobile of fructification, then its fruit was not dissimilar from that of the Canadian species in its position and general form, though it may have differed in details. On these grounds I declined to identify the Scottish species with L. Gaspianum. The Lepidodendron from the Devonian of Belgium described and figured by Crepin,[BR] has a better claim to such identification, and would seem to prove that this species existed in Europe as well as in America. I also saw in Mr. Peach’s collection in 1870 some fragments which seemed to me distinct from Salter’s species, and possibly belonging to L. Gaspianum.[BS]
[BQ] “Report on Devonian Plants of Canada,” 1871.
[BR] “Observations sur quelques Plantes Fossiles des dépôts Devoniens.”
[BS] “Proceedings of the Geological Society of London,” March, 1871.
In the earliest description of Psilophyton I recognised its probable generic affinity with Miller’s “dichotomous plants,” with Salter’s “rootlets,” and with Goeppert’s Haliserites Dechenianus, and stated that I had “little doubt that materials exist in the Old Red Sandstone of Scotland for the reconstruction of at least one species of this genus.” Since, however, Miller’s plants had been referred to coniferous roots, and to fucoids, and Goeppert’s Haliserites was a name applicable only to fucoids, and since the structure and fruit of my plants placed them near to Lycopods, I was under the necessity of giving them a special generic name, nor could I with certainty affirm their specific identity with any European species. The comparison of the Scottish specimens with woody rootlets, though incorrect, is in one respect creditable to the acumen of Salter, as in almost any state of preservation an experienced eye can readily perceive that branchlets of Psilophyton must have been woody rather « 100 » than herbaceous, and their appearance is quite different from that of any true Algæ.
The type of Psilophyton is my P. princeps, of which the whole of the parts and structures are well known, the entire plant being furnished in abundance and in situ in the rich plant-beds of Gaspé. A second species, P. robustius, has also afforded well-characterised fructification. P. elegans, whose fruit appears as “oval scales,” no doubt bore sac-like spore-cases resembling those of the other species, but in a different position, and perfectly flattened in the specimens procured. The only other Canadian species, P. glabrum, being somewhat different in appearance from the others, and not having afforded any fructification, must be regarded as uncertain.
The generic characters of the first three species may be stated as follows:
Stems dichotomous, with rudimentary subulate leaves, sometimes obsolete in terminal branchlets and fertile branches; and in decorticated specimens represented only by punctiform scars. Young branches circinate. Rhizomata cylindrical, with circular root-areoles. Internal structure of stem, an axis of scalariform vessels enclosed in a sheath of imperfect woody tissue and covered with a cellular bark more dense externally. Fruit, naked sac-like spore-cases, in pairs or clusters, terminal or lateral.
The Scottish specimens conform to these characters in so far as they are known, but not having as yet afforded fruit or internal structure, they cannot be specifically determined with certainty. More complete specimens should be carefully searched for, and will no doubt be found.
In Belgium, M. Crepin has described a new species from the Upper Devonian of Condroz under the name P. Condrusianum (1875). It wants, however, some of the more important characters of the genus, and differs in having a pinnate ramification, giving it the aspect of a fern. In a later paper (1876) the author considers this species distinct from Psilophyton, and proposes for it a new generic name Rhacophyton.
The characters given by Mr. Carruthers, in his paper of 1873, for the species P. Dechenianum, are very few and general: “Lower branches short and frequently branching, giving the plant an oblong circumscription.” Yet even these characters do not apply, so far as known, to Miller’s fucoids or Salter’s rootlets or Goeppert’s Haliserites. They merely express the peculiar mode of branching already referred to in Salter’s Lepidodendron nothum. The identification of the former plants with the Lepidodendron and Lycopodites, indeed, « 101 » rests only on mere juxtaposition of fragments, and on the slight resemblance of the decorticated ends of the branches of the latter plants to Psilophyton. It is contradicted by the obtuse ends of the branches of the Lepidodendron and Lycopodites, and by the apparently strobilaceous termination of some of them.
Salter’s description of his Lepidodendron nothum is quite definite, and accords with specimens placed in my hands by Mr. Peach: “Stems half an inch broad, tapering little, branches short; set on at an acute angle, blunt at their terminations. Leaves in seven to ten rows, very short, not a line long, and rather spreading than closely imbricate.” These characters, however, in so far as they go, are rather those of the genus Lycopodites than of Lepidodendron, from which this plant differs in wanting any distinct leaf-bases, and in its short, crowded leaves. It is to be observed that they apply also to Salter’s Lycopodites Milleri, and that the difference of the foliage of that species may be a result merely of different state of preservation. For these reasons I am disposed to place these two supposed species together, and to retain for the species the name Lycopodites Milleri. It may be characterised by the description above given, with merely the modification that the leaves are sometimes nearly one-third of an inch long and secund (Fig. 17, supra, lower figure).
Decorticated branches of the above species may no doubt be mistaken for Psilophyton, but are nevertheless quite distinct from it, and the slender branching dichotomous stems, with terminations which, as Miller graphically states, are “like the tendrils of a pea,” are too characteristic to be easily mistaken, even when neither fruit nor leaves appear. With reference to fructification, the form of L. Milleri renders it certain that it must have borne strobiles at the ends of its branchlets, or some substitute for these, and not naked spore-cases like those of Psilophyton.
The remarkable fragment communicated by Sir Philip Egerton to Mr. Carruthers,[BT] belongs to a third group, and has, I think, been quite misunderstood. I am enabled to make this statement with some confidence, from the fact that the reverse or counterpart of Sir Philip’s specimen was in the collection of Sir Wyville Thomson, and was placed by him in my hands in 1870. It was noticed in my paper on “New Devonian Plants,” in the “Journal of the Geological Society of London,” and referred to my genus Ptilophyton, as stated above under Section II., page 86 et seq.
Mr. Salter described, in 1857,[BU] fragments of fossil wood from the Scottish Devonian, having the structure of Dadoxylon, though very imperfectly preserved; and Prof. McNab has proposed[BV] the generic name Palæopitys for another specimen of coniferous wood collected by Hugh Miller, and referred to by him in the “Testimony of the Rocks.” From Prof. McNab’s description, I should infer that this wood may, after all, be generically identical with the woods usually referred to Dadoxylon of Unger (Araucarioxylon of Kraus). The description, however, does not mention the number and disposition of the rows of pores, nor the structure of the medullary rays, and I have not been able to obtain access to the specimens themselves. I have described five species of Dadoxylon from the Middle and Upper Erian of America, all quite distinct from the Lower Carboniferous species. There is also one species of an allied genus, Ormoxylon. All these have been carefully figured, and it is much to be desired that the Scottish specimens should be re-examined and compared with them.
[BU] “Journal of the London Geological Society.”
[BV] “Transactions of the Edinburgh Botanical Society,” 1870.
Messrs. Jack and Etheridge have given an excellent summary of our present knowledge of the Devonian flora of Scotland, in the Journal of the London Geological Society (1877). From this it would appear that species referable to the genera Calamities, Lepidodendron, Lycopodites, Psilophyton, Arthrostigma, Archæopteris, Caulopteris, Palæopitys, Araucarioxylon, and Stigmaria have been recognised.
The plants described by these gentlemen from the Old Red Sandstone of Callender, I should suppose, from their figures and descriptions, to belong to the genus Arthrostigma, rather than to Psilophyton. I do not attach any importance to the suggestions referred to by them, that the apparent leaves may be leaf-bases. Long leaf-bases, like those characteristic of Lepidofloyos, do not occur in these humbler plants of the Devonian. The stems with delicate “horizontal processes” to which they refer may belong to Ptilophyton or to Pinnularia.
In conclusion, I need scarcely say that I do not share in the doubts expressed by some British palæontologists as to the distinctness of the Devonian and Carboniferous floras. In eastern America, where these formations are mutually unconformable, there is, of course, less room for doubt than in Ireland and in western America, where they are stratigraphically continuous. Still, in passing « 103 » from the one to the other, the species are for the most part different, and new generic forms are met with, and, as I have elsewhere shown, the physical conditions of the two periods were essentially different.[BW]
[BW] “Reports on Devonian Plants and Lower Carboniferous Plants of Canada.”
It is, however, to be observed that since—as Stur and others have shown—Calamities radiatus, and other forms distinctively Devonian in America, occur in Europe in the Lower Carboniferous, it is not unlikely that the Devonian flora, like that of the Tertiary, appeared earlier in America. It is also probable, as I have shown in the “Reports” already referred to, that it appeared earlier in the Arctic than in the temperate zone. Hence an Arctic or American flora, really Devonian, may readily be mistaken for Lower Carboniferous by a botanist basing his calculations on the fossils of temperate Europe. Even in America itself, it would appear, from recent discoveries in Virginia and Ohio, that certain Devonian forms lingered longer in those regions than farther to the northeast;[BX] and it would not be surprising if similar plants occurred in later beds in Devonshire or in the south of Europe than in Scotland. Still, these facts, properly understood, do not invalidate the evidence of fossil plants as to geological age, though errors arising from the neglect of them are still current.
[BX] Andrews, “Palæontology of Ohio,” vol. ii.; Meek, “Fossil Plants from Western Virginia,” Philosophical Society, Washington, 1875.
VI.—Geological Relations of some Plant-bearing Beds of
Eastern Canada.
(“Report on Erian Plants,” 1871.)
The Gaspé sandstones have been fully described by Sir W. E. Logan, in his “Report on the Geology of Canada,” 1863. He there assigns to them a thickness of seven thousand and thirty-six feet, and shows that they rest conformably on the Upper Silurian limestones of the Lower Helderberg group (Ludlow), and are in their turn overlaid unconformably by the conglomerates which form the base of the Carboniferous rocks of New Brunswick. I shall add here merely a few remarks on points in their physical character connected with the occurrence of plants in them.
Prototaxites (Nematophyton) Logani and other characteristic Lower Erian plants occur in the base of the sandstones at Little Gaspé. This fact, along with the occurrence, as stated in my paper of 1863, of rhizomes of Psilophyton preserving their scalariform « 104 » structure, in the upper part of the marine Upper Silurian limestones,[BY] proves the flora of the Devonian rocks to have had its beginning at least in the previous geological period, and to characterise the lower as well as the upper beds of the Devonian series. In this connection I may state that, from their marine fossils, as well as their stratigraphical arrangement, Sir W. E. Logan and Mr. Billings regard the lower portions of the Gaspé sandstones as the equivalents of the Oriskany sandstone of New York. On the other hand, the great thickness of this formation, the absence of Lower Devonian fossils from its upper part, and the resemblance of the upper beds to those of the newer members of the Devonian elsewhere, render it probable that the Gaspé sandstones, though deficient in the calcareous members of the system, seen farther to the westward, represent the whole of the Devonian period.
[BY] The marine fossils of these beds have been determined by Mr. Billings. They are Upper Silurian, with an intermixture of Lower Devonian in the upper part. Fragments of Nematophyton occur in beds of the same age in the Bay des Chaleurs, at Cape Bon Ami.
The Gaspé sandstones, as their name imports, are predominantly arenaceous, and often coarsely so, the sandstones being frequently composed of large grains and studded with quartz-pebbles. Grey and buff are prevalent colours, but red beds also occur, more especially in the upper portion. There are also interstratified shaly beds, sometimes occurring in groups of considerable thickness, and associated with fine-grained and laminated argillaceous sandstone, the whole having in many places the lithological aspect of the coal-measures. At one place, near the middle of the series, there is a bed of coal from one inch to three inches in thickness, associated with highly bituminous shales abounding in remains of plants, and also containing fragments of crustaceans and fishes (Pterygotus, Ctenacanthus ? &c). The beds connected with this coal are grey sandstones and grey and dark shales, much resembling those of the ordinary coal formation. The coal is shining and laminated, and both its roof and floor consist of laminated bituminous shale with fragments of Psilophyton. It has no true under-clay, and has been, I believe, a peaty mass of rhizomes of Psilophyton. It occurs near Tar Point, on the south side of Gaspé Bay, a place so named from the occurrence of a thick dyke of trap holding petroleum in its cavities. The coal is of considerable horizontal extent, as in its line of strike a similar bed has been discovered on the Douglas River, about four miles distant. It has not been recognised on the north « 105 » side of the bay, though we find there beds, probably on very nearly the same horizon, holding Psilophyton in situ.
As an illustration of one of the groups of shaly beds, and of the occurrence of roots of Psilophyton, I may give the following sectional list of beds seen near “Watering Brook,” on the north shore of the bay. The order is descending:
| FT. | IN. | ||
| 1. | Grey sandstones and reddish pebbly sandstone of great thickness | ||
| 2. | Bright-red shale | 8 | 0 |
| 3. | Grey shales with stems of Psilophyton, very abundant but badly preserved | 0 | 5 |
| 4. | Grey incoherent clay, slickensided, and with many rhizomes and roots of Psilophyton | 0 | 3 |
| 5. | Hard grey clay or shale, with fragments and roots of Psilophyton | 4 | 0 |
| 6. | Red shale | 8 | 0 |
| 7. | Grey and reddish crumbling sandstone |
Groups of beds similar to the above, but frequently much more rich in fossils, occur in many parts of the section, and evidently include fossil soils of the nature of under-clays, on which little else appears to have grown than a dense herbage of Psilophyton, along with plants of the genus Arthrostigma.
In addition to these shaly groups, there are numerous examples of beds of shale of small thickness included in coarse sandstones, and these beds often occur in detached fragments, as if the remnants of more continuous layers partially removed by currents of water. It is deserving of notice that nearly all these patches of shale are interlaced with roots or stems of Psilophyton, which sometimes project beyond their limits into the sandstone, as if the vegetable fibres had preserved the clay from removal. In short, these lines of patches of shale seem to be remnants of soils on which Psilophyton has flourished abundantly, and which have been partially swept away by the currents which deposited the sand. Some of the smaller patches may even be fragments of tough swamp soils interwoven with roots, drifted by the agency of the waves or possibly by ice; such masses are often moved in this way on the borders of modern swamps on the sea-coast.
The only remaining point connected with local geology to which I shall allude is the admirable facilities afforded by the Gaspé coast both for ascertaining the true geological relations of the beds, and for studying the Devonian plants, as distinctly exposed on large surfaces « 106 » of rock. On the coast of the river St. Lawrence, at Cape Rozier and its vicinity, the Lower Silurian rocks of the Quebec group are well exposed, and are overlaid unconformably by the massive Upper Silurian limestones of Cape Gaspé, which rise into cliffs six hundred feet in height, and can be seen filled with their characteristic fossils on both sides of the cape. Resting upon these, and dipping at high angles toward Gaspé Bay, are the Devonian sandstones, which are exposed in rugged cliffs slightly oblique to their line of strike, along a coast-line of ten miles in length, to the head of the bay. On the opposite side of the bay they reappear; and, thrown into slight undulations by three anticlinal curves, occupy a line of coast fifteen miles in length. The perfect manner in which the plant-bearing beds are exposed in these fine natural sections may serve to account for the completeness with which the forms and habits of growth of the more abundant species can be described.
In the Bay des Chaleurs, similar rocks exist with some local variations. In the vicinity of Campbellton are calcareous and magnesian breccia or agglomerate, hard shales, conglomerates and sandstones of Lower Devonian age. The agglomerate and lower shales contain abundant remains of fishes of the genera Cephalaspis, Coccosteus, Ctenacanthus, and Homacanthus, and also fragments of Pterygotus. The shales and sandstones abound in remains of Psilophyton, with which are Nematophyton, Arthrostigma, and Leptophleum of the same species found in the Lower Devonian of Gaspé Bay. These beds near Campbellton dip to the northward, and the Restigouche River here occupies a synclinal, for on the opposite side, at Bordeaux Quarry, there are thick beds of grey sandstone dipping to the southward, and containing large silicified trunks of Prototaxites, in addition to Psilophyton. These beds are all undoubtedly Lower Erian, but farther to the eastward, on the north side of the river, there are newer and overlying strata. These are best seen at Scaumenac Bay, opposite Dalhousie, between Cape Florissant and Maguacha Point, where they consist of laminated and fine-grained sandstone, with shales of grey colours, but holding some reddish beds at top, and overlaid unconformably by a great thickness of Lower Carboniferous red conglomerate and sandstone. In these beds numerous fossil fishes have been found, among which Mr. Whiteaves recognises species of Pterichthys, Glyptolepis, Cheirolepis, &c. With these are found somewhat plentifully four species of fossil ferns, all of Upper Erian types, of which one is peculiar to this locality; but the others are found in the Upper Erian of Perry, in Maine, or in the Cat skill group of New York.
In order that distinct notions may be conveyed as to the geological horizons of the species, I may state that the typical Devonian or Erian series of Canada and New York may be divided in descending order into—1. The Chemung group, including the Chemung and Portage sandstones and shales. 2. The Hamilton group, including the Genesee, Hamilton, and Marcellus shales. 3. The Corniferous limestone and its associated beds. 4 The Oriskany sandstone. As the Corniferous limestone, which is the equivalent of the Lower Carboniferous limestone in the Carboniferous period, is marine, and affords scarcely any plants, we may, as is usually done for like purposes in the Carboniferous, group it with the Oriskany under the name Lower Erian. The Hamilton rocks will then be Middle Erian, and the Chemung group Upper Erian. In the present state of our knowledge, the series may be co-ordinated with the rocks of Gaspé, New Brunswick, and Maine, as in the following table:
| Subdivisions. | New York and Western Canada. |
Gaspé and Bay des Chaleurs. |
Southern New Brunswick. |
Coast of Maine. |
| Upper Devonian or Erian. |
Chemung Group. |
Upper Sandstones. Long Cove, &c. Scauminac Beds. |
Mispec Group. Shale, Sandstone, and Conglomerate. |
Perry Sandstones. |
| Middle Devonian or Erian. |
Hamilton Group. |
Middle Sandstones. Bois Brulé, Cape Oiseau, &c. |
Little R. Group (including Cordaite Shales and Dadoxylon Sandstone). |
|
| Lower Devonian or Erian. |
Corniferous and Oriskany groups. |
Lower Sandstones. Gaspé Basin, Little Gaspé, &c. Campbellton Beds. |
Lower Conglomerates, &c. |
It may be proper, before closing this note, to state the reasons which have induced me to suggest in the following pages the use of the term “Erian,” as equivalent to “Devonian,” for the great system of formations intervening between the Upper Silurian and the Lower Carboniferous in America. I have been induced to adopt this course by the following considerations: 1. The great area of « 108 » undisturbed and unaltered rocks of this age, including a thickness in some places of eighteen thousand feet, and extending from east to west through the Northern States of the Union and western Canada for nearly seven hundred miles, while it spreads from north to south from the northern part of Michigan far into the Middle States, is undoubtedly the most important Devonian area now known to geologists. 2. This area has been taken by all American geologists as their typical Devonian region. It is rich in fossils, and these have been thoroughly studied and admirably illustrated by the New York and Canadian Surveys. 3. The rocks of this area surround the basin of Lake Erie, and were named, in the original reports of the New York Survey, the “Erie Division” 4. Great difficulties have been experienced in the classification of the European Devonian, and the uncertainties thus arising have tended to throw doubt on the results obtained in America in circumstances in which such difficulties do not occur.
These reasons are, I think, sufficient to warrant me in holding the great Erie Division of the New York geologists as the typical representative of the rocks deposited between the close of the Upper Silurian and the beginning of the Carboniferous period, and to use the term Erian as the designation of this great series of deposits as developed in America, in so far at least as their flora is concerned. In doing so, I do not wish to introduce a new name merely for the sake of novelty; but I hope to keep before the minds of geologists the caution that they should not measure the Erian formations of America, or the fossils which they contain, by the comparatively depauperated representatives of this portion of the geological scale in the Devonian of western Europe.
VII.—On the Relations of the so-called “Ursa Stage” of Bear Island
with the Palæozoic Flora of North America.
The following note is a verbatim copy of that published by me in 1873, and the accuracy of which has now been vindicated by the recent observations of Nathorst:
The plants catalogued by Dr. Heer, and characterising what he calls the “Ursa Stage,” are in part representatives of those of the American flora which I have described as the “Lower Carboniferous Coal-Measures” (Subcarboniferous of Dana), and whose characteristic species, as developed in Nova Scotia, I noticed in the “Journal of the Geological Society” in 1858 (vol. xv.). Dr. Heer’s list, however, includes some Upper Devonian forms; and I would suggest that « 109 » either the plants of two distinct beds, one Lower Carboniferous and the other Upper Devonian, have been near to or in contact with each other and have been intermixed, or else that in this high northern latitude, in which (for reasons stated in my “Report on the Devonian Flora”[BZ]) I believe the Devonian plants to have originated, there was an actual intermixture of the two floras. In America, at the base of the Carboniferous of Ohio, a transition of this kind seems to occur; but elsewhere in northeastern America the Lower Carboniferous plants are usually unmixed with the Devonian.
[BZ] “Geological Survey of Canada,” 1871.
Dr. Heer, however, proceeds to identify these plants with those of the American Chemung, and even with those of the Middle Devonian of New Brunswick, as described by me—a conclusion from which I must altogether dissent, inasmuch as the latter belong to beds which were disturbed and partially metamorphosed before the deposition of the lowest Carboniferous or “Subcarboniferous” beds.
Dr. Heer’s error seems to have arisen from want of acquaintance with the rich flora of the Middle Devonian, which, while differing in species, has much resemblance in its general facies, and especially in its richness in ferns, to that of the coal-formation.
To geologists acquainted with the stratigraphy and the accompanying animal fossils, Dr. Heer’s conclusions will of course appear untenable; but they may regard them as invalidating the evidence of fossil plants; and for this reason it is, I think, desirable to give publicity to the above statements.
I consider the British equivalent of the lower coal-measures of eastern America to be the lower limestone shales, the Tweedian group of Mr. Tate (1858), but which have sometimes been called the “Calciferous Sandstone” (a name preoccupied for a Cambrian group in America). This group does not constitute “beds of passage” to the Devonian, more especially in eastern America, where the lower coal-formation rests unconformably on the Devonian, and is broadly distinguished by its fossils.
The above notes would not have been extended to so great length, but for the importance of the Erian flora as the precursor of that of the Carboniferous, and the small amount of attention hitherto given to it by geologists and botanists.
THE CARBONIFEROUS FLORA—CULMINATION OF THE ACROGENS—FORMATION OF COAL.
Ascending from the Erian to the Carboniferous system, so called because it contains the greatest deposits of anthracite and bituminous coal, we are still within the limits of the Palæozoic period. We are still within the reign of the gigantic club-mosses, cordaites, and taxine pines. At the close of the Erian there had been over the whole northern hemisphere great changes of level, accompanied by active volcanic phenomena, and under these influences the land flora seems to have much diminished. At length all the old Erian species had become extinct, and their place was supplied by a meagre group of lycopods, ferns, and pines of different species from those of the preceding Erian. This is the flora of the Lower Carboniferous series, the Tweedian of England, the Horton series of Nova Scotia, the lower coal-measures of Virginia, the culm of Germany. But the land again subsided, and the period of the marine limestone of the Lower Carboniferous was introduced. In this the older flora disappeared, and when the land emerged we find it covered with the rich flora of the coal-formation proper, in which the great tribes of the lycopods and cordaites attained their maxima, and the ferns were continued as before, though under new generic and specific forms.

There is something very striking in this succession of a new plant world without any material advance. It is like passing in the modern world from one district to another, in which we see the same forms of life, only represented by distinct though allied species. Thus, when the voyager crosses the Atlantic from Europe to America, he meets with pines, oaks, birches, poplars, and beeches of the same genera with those he had left behind; but the species are distinct. It is something like this that meets us in our ascent into the Carboniferous world of plants. Yet we know that this is a succession in time, that all our old Erian friends are dead and buried long ago, and that these are new forms lately introduced (Fig. 32).
Conveying ourselves, then, in imagination forward to the time when our greatest accumulations of coal were formed, and fancying that we are introduced to the American or European continent of that period, we find ourselves in a new and strange world. In the Devonian age, and even in the succeeding Lower Carboniferous, there was in the interior of America a wide inland sea, with forest belts clinging to its sides or clothing its islands. But in the coal period this inland sea had given « 112 » place to vast swampy flats, and which, instead of the oil-bearing shales of the Erian, were destined to produce those immense and wide-spread accumulations of vegetable matter which constitute our present beds of bituminous and anthracite coal. The atmosphere of these great swamps is moist and warm. Their vegetation is most exuberant, but of forms unfamiliar to modern eyes, and they swarm with insects, millipedes, and scorpions, and with batrachian reptiles large and small, among which we look in vain for representatives of the birds and beasts of the present day.

Fig. 34.—Sigillaria Lorwayana, Dawson. a, Zones of fruit-scars. b, Leaf-scar
enlarged, c, Fruit-scar enlarged. See appended note. |
|
Prominent among the more gigantic trees of these swampy forests are those known to us as Sigillariæ (Fig. 33). They have tall, pillar-like trunks, often several feet in diameter, ribbed like fluted columns, but in the reverse way, and spreading at the top into a few thick branches, which are clothed with long, grass-like leaves. They resemble in some respects the Lepidodendra of the Erian age, but are more massive, with ribbed instead of scaly trunks, and longer leaves. If we approach one of them more closely, we are struck with the regular ribs of its trunk, dotted with rows of scars of fallen leaves, from which it receives its name Sigillaria, or seal-tree (Figs. 34-37). If we cut into its stem, we find that, instead of « 113 » the thin bark and firm wood with which we are familiar in our modern trees, it has a hard external rind, then a great thickness of cellular matter with rope-like bands of fibres, constituting an inner bark, while in the centre is a firm, woody axis of comparatively small diameter, and somewhat intermediate in its structures between that of the Lepidodendra and those of the cycads and the taxine conifers. Thus a great stem, five feet in diameter, may consist principally of cellular and bast fibres with very little true woody matter. The roots of this tree are « 114 » perhaps its most singular feature. They usually start from the stem in four main branches, then regularly bifurcate several times, and then run out into great cylindrical cables, running for a long distance, and evidently intended to anchor the plant firmly in a soft and oozy soil. They were furnished with long, cylindrical rootlets placed regularly in a spiral manner, and so articulated that when they dropped off they left regular rounded scars. They are, in short, the Stigmariæ, which we have already met with in the Erian (Figs. 38, 39). In Fig. 33 I have endeavoured to restore these strange trees. It is not wonderful that such plants have caused much botanical controversy. It was long before botanists could be convinced that « 115 » their roots are properly roots at all, and not stems of some aquatic plant. Then the structure of their stems is most puzzling, and their fruit is an enigma, for while some have found connected with them cones supposed to resemble those of lycopods, others attribute to them fruits like those of yew-trees. For years I have been myself gathering materials from the rich coal-formation deposits of Nova Scotia in aid of the solution of these questions, and in the mean time Dr. Williamson, of Manchester, and Renault and other botanists in France, have been amassing and studying stores of specimens, and it is still uncertain who may finally be the fortunate discoverer to set all controversies at rest. My present belief is, that the true solution consists in the fact that there are many kinds of Sigillariæ. While in the modern forests « 116 » of America and Europe the species of any of our ordinary trees, as oaks, birches, or maples, may almost be counted on one’s fingers, Schimper in his vegetable palæontology enumerates about eighty species of Carboniferous Sigillariæ; and while on the one hand many of these are so imperfectly known that they may be regarded as uncertain, on the other hand many species must yet remain to be discovered.[CA] Now, in so vast a number of species there must be a great range of organisation, and, indeed, it has already been attempted to subdivide them into several generic groups. The present state of the question appears to me to be this, that in these Sigillariæ we have a group divisible into several forms, some of which will eventually be classed with the Lepidodendra as lycopods, while others will be found to be naked-seeded phænogams, allied to the pines and cycads, and to a remarkable group of trees known as Cordaites, which we must shortly notice.
[CA] In a recent memoir (Berlin, 1887) Stur has raised the number of species in one subdivision of the Sigillariæ (the Favulariæ) to forty-seven!
Before considering other forms of Carboniferous vegetation, let us glance at the accumulation of coal, and the agency of the forests of Sigillariæ therein. Let us imagine, in the first instance, such trees as those represented in the figures, growing thickly together over vast swampy flats, with quantities of undergrowth of ferns and other plants beneath their shade, and accumulating from age to age in a moist soil and climate a vast thickness of vegetable mould and trunks of trees, and spores and spore-cases, and we have the conditions necessary for the growth of coal. Many years ago it was observed by Sir William Logan that in the coal-field of South Wales it was the rule with rare exceptions that, under every bed of coal, there is a bed of clay filled with roots of the Stigmaria, already referred to as the root of Sigillaria. This discovery « 117 » has since been extended to all the coal-fields of Europe and America, and it is a perfectly conclusive fact as regards the origin of coal. Each of these “under-clays,” as they are called, must, in fact, have been a soil on which grew, in the first instance, Sigillariæ and other trees having stigmaria-roots. Thus, the growth of a forest of Sigillariæ was the first step toward the accumulation of a bed of coal. More than this, in some of the coarser and more impure coals, where there has been sufficient earthy matter to separate and preserve impressions of vegetable forms, we can see that the mass of the coal is made up of flattened Sigillariæ, mixed with vegetable débris of all kinds, including sometimes vast quantities of lepidodendroid spores, and the microscopic study of the coal gives similar results (Fig. 40). Further, on the surfaces of many coals, and penetrating the shales or sandstones which form their roofs, we find erect stumps of sigillaria and other trees, showing that the accumulation of the coal terminated as it had begun, by a forest-growth. I introduce here a section of a few of the numerous beds of coal exposed in the cliffs of the South Joggins, in Nova Scotia, in illustration of these facts. We can thus see how in the slowly subsiding areas of the coal-swamps successive beds of coal were accumulated, alternating with beds of sandstone and shale (Figs. 41, 42). For other details of this kind I must refer to papers mentioned in the sequel.
Returning to the more special subject of this work, I may remark that the lepidodendroid trees and the ferns, both the arborescent and herbaceous kinds, are even more richly represented in the Carboniferous than in the preceding Erian, I must, however, content myself with merely introducing a few representatives of some of the more common kinds, in an appended note, and here give a figure of a well-known Lower Carboniferous lepidodendron, with its various forms of leaf-bases, and its foliage and fruit (Fig. 43), and a similar illustration of an allied generic form, that known as Lepidophloios[CB] (Fig. 44).
[CB] For full descriptions of these, see “Acadian Geology.”
Another group which claims our attention is that of the Calamites. These are tall, cylindrical, branchless stems, with whorls of branchlets, bearing needle-like leaves and spreading in stools from the base, so as to form dense thickets, like Southern cane-brakes (Fig. 46). They bear, in habit of growth and fructification, a close « 119 » relation to our modern equisetums, or mare’s-tails, but, as in other cases we have met with, are of gigantic size and comparatively complex structure. Their stems, in cross-section, show radiating bundles of fibres, like those of exogenous woods, yet the whole plan of structure presents some curious resemblances to the stems of their humble successors, the modern mare’s-tails. It would seem, from the manner in which dense brakes of these Calamites have been preserved in the coal-formation of Nova Scotia, that they spread over low and occasionally inundated flats, and formed fringes on the seaward sides of the great Sigillaria forests. In this way they no doubt contributed to prevent the invasion of the areas of coal accumulation by the muddy waters of inundations, and thus, though they may not have furnished much of the material of coal, they no doubt contributed to its purity. Many beautiful plants of the genera Asterophyllites and Annularia are supposed to have been allied to the Calamites, or to have connected them with the Rhizocarps. The stems and fruit of these plants have strong points of resemblance to those of Sphenophyllum, and the leaves are broad, and not narrow and angular like those of the true Calamites (Fig. 45).

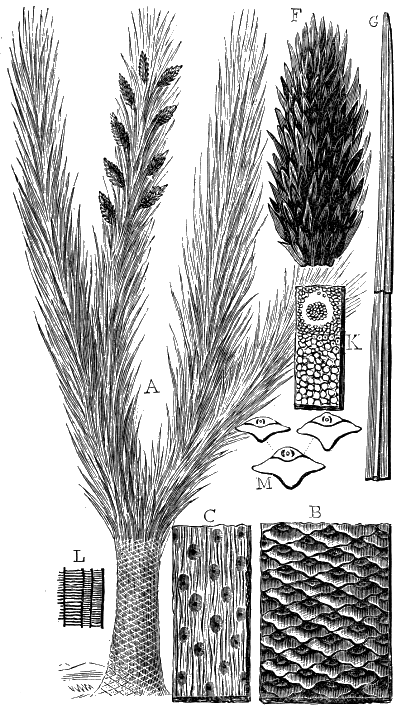
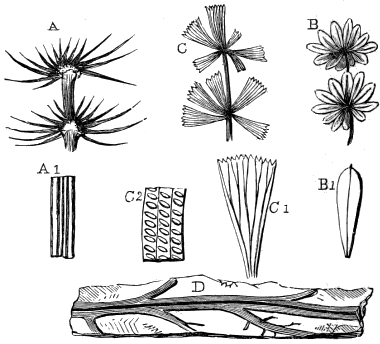
No one has done more than my friend Dr. Williamson, of Manchester, to illustrate the structure of Calamites, and he has shown that these plants, like other cryptogams of the Carboniferous, had mostly stems with regular fibrous wedges, like those of exogens. The structure of the stem is, indeed, so complex, and differs so much in different stages of growth, and different states of preservation, that we are in danger of falling into the greatest confusion in classifying these plants. Sometimes what we call a Calamite is a mere cast of its pith showing longitudinal striæ and constrictions at the nodes. Sometimes we have the form of the outer surface of the woody cylinder, showing longitudinal ribs, nodes, and marks of the emission of the branchlets. Sometimes we have the outer surface of the plant covered with a smooth bark showing flat ribs, or almost smooth, and having at the nodes regular articulations with the bases of the verticillate « 123 » branchlets, or on the lower part of the stem the marks of the attachment of the roots. The Calamites grew in dense clumps, budding off from one another, sometimes at different levels, as the mud or sand accumulated about their stems, and in some species there were creeping rhizomata or root-stocks (Figs. 46 to 49).
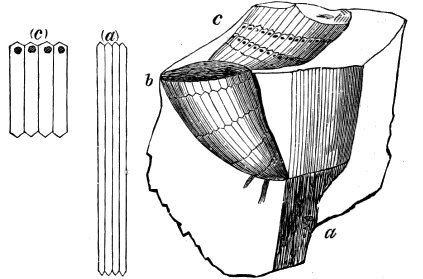
But all Calamites were not alike in structure. In a recent paper[CC] Dr. Williamson describes three distinct structural types. What he regards as typical Calamites has in its woody zone wedges of barred vessels, with thick bands of cellular tissue separating them. A second type, which « 124 » he refers to Calamopitus, has woody bundles composed of reticulated or multiporous fibres, with their porous sides parallel to the medullary rays, which are better developed than in the previous form. The intervening cellular masses are composed of elongated cells. This is a decided advance in structure, and is of the type of those forms having the most woody and largest stems, which Brongniart named Calamodendron (Fig. 50). A third form, to which Dr. Williamson seems to prefer to assign this last name, has the tissue of the woody wedges barred, as in the first, but the medullary rays are better developed than in the second. In this third form the intermediate tissue, or primary medullary rays, is truly fibrous, and with secondary medullary rays traversing it. My own observations lead me to infer that there was a fourth type of calamitean stem, less endowed with woody matter, and having a larger fistulous or cellular cavity than any of those described by Dr. Williamson.
[CC] “Memoirs of the Philosophical Society,” Manchester, 1886-'87.
There is every reason to believe that all these various « 125 » and complicated stems belonged to higher and nobler types of mare’s-tails than those of the modern world, and that their fructification was equisetaceous and of the form known as Calamostachys.
We have already seen that noble tree-ferns existed in the Erian period, and these were continued, and their number and variety greatly extended, in the Carboniferous. In regard to the structure of their stems, and the method of supporting these by aërial roots, the tree-ferns of all ages have been nearly alike, and the form and structure of the leaves, except in some comparatively rare and exceptional types, has also been much the same. Any ordinary observer examining a collection of coal-formation ferns recognises at once their kinship to the familiar brackens of our own time. Their fructification is, unfortunately, rarely preserved, so that we are not able, in the case of many species, to speak confidently of their affinities with modern forms; but the knowledge of this subject has been constantly extending, and a sufficient amount of information has been obtained to enable us to say something as to their probable relationships. (Figs. 51 to 55.)
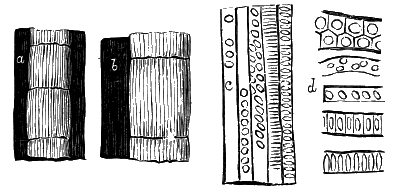
The families into which modern ferns are divided are, it must be confessed, somewhat artificial, and in the case « 126 » of fossil ferns, in which the fructification is for the most part wanting, it is still more so, depending in great part on the form and venation of the divisions of the fronds. Of about eight families into which modern ferns are divided, seven are found in a fossil state, and of these, four at least, the Cyathaceæ, the Ophioglosseæ, the Hymenophyllaceæ, and the Marattiaceæ, go back to the coal-formation.[CD]
[CD] Mr. R. Kidston has recently described very interesting forms of fern fructification from the coal-formation of Great Britain, and much has been done by European palæobotanists, and also by Lesquereux and Fontaine in America.

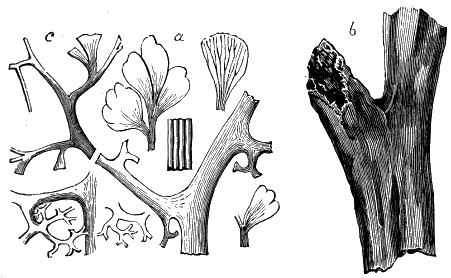
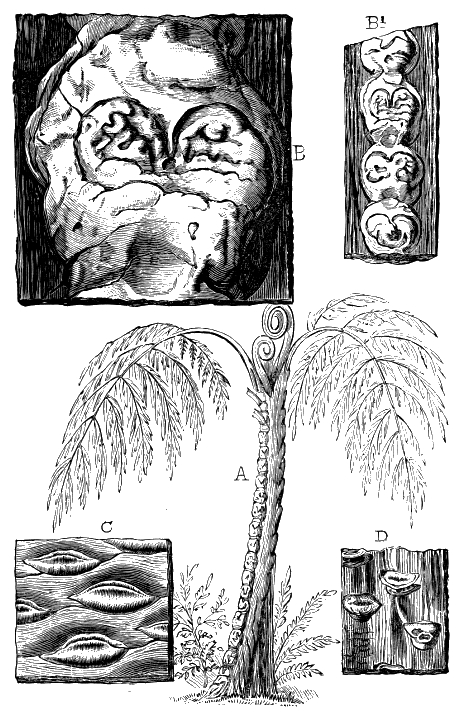
Some of these ferns have the more complex kind of spore-case, with a jointed, elastic ring. It is to be observed, however, that those forms which have a simple spore-case, either netted or membranous, and without annulus, are most common in the Devonian and lowest Carboniferous. Some of the forms in these old rocks are somewhat difficult to place in the system. Of these, the species of Archæopteris, of the Upper and Middle Erian, are eminent as examples. This type, however, scarcely extends as high as the coal-formation.[CE] Some of the tree-ferns of the Carboniferous present very remarkable features. One of these, of the genus Megaphyton, seems to have two rows of great leaves, one at each side of the stem, which was probably sustained by large bundles of aërial roots (Fig. 56).
[CE] The pretty little ferns of the genus Botrychium (moonwort), so common in American and European woods, seem to be their nearest modern allies.
In the Carboniferous, as in the Erian, there are leaves which have been referred to ferns, but are subject to doubt, as possibly belonging to broad-leaved taxine trees allied to the gingko-tree of China. One of these, represented in Fig. 57, has been found in the coal-formation of Nova Scotia, and referred to the doubtful genus Noeggerathia. Fontaine has proposed for similar leaves found in Virginia the new generic name Saportea.
Ferns, as might be inferred from their great age, are at the present time dispersed over the whole world; but their headquarters, and the regions to which tree-ferns are confined, are the more moist climates of the tropics and of the southern hemisphere. The coal-swamps of the northern hemisphere seem to have excelled even these favoured regions of the present world as a paradise for ferns.
I have already stated that the Carboniferous constitutes the headquarters of the Cordaites (Fig. 58), of which a large number of species have been described, both in « 131 » Europe and America. We sometimes, though rarely, find their stems showing structure. In this case we have a large cellular pith, often divided by horizontal partitions into flat chambers, and constituting the objects which, when detached, are called Sternbergiæ (Fig. 62). These Sternbergia piths, however, occur in true conifers as well, as they do in the modern world in some trees, like our common butternut, of higher type; and I showed many years ago that the Sternbergia type may be detected in the young twigs of the balsam-fir (Abies balsamifera). The pith was surrounded by a ring of scalariform or barred tissue, often of considerable thickness, and in young stems so important as to have suggested lycopodiaceous affinities. But as the stem grew in size, a regular ring of woody wedges, with tissue having rounded or hexagonal pores or discs, like those of pines, was developed. Outside this was a bark, often apparently of some thickness. This structure in many important points resembles that of cycads, and also approaches to the structure of Sigillaria, while in its more highly developed forms it approximates to that of the conifers.
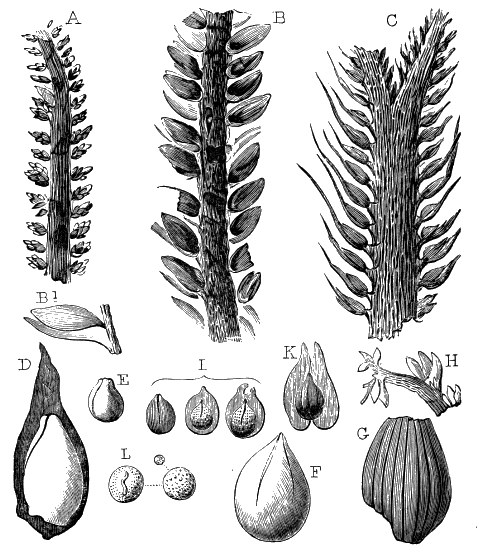
On the stems so constructed were placed long and often broad many-nerved leaves, with rows of stomata or breathing-pores, and attached by somewhat broad bases to the stem and branches. The fruit consisted of racemes, or clusters of nutlets, which seem to have been provided « 133 » with broad lateral wings for flotation in the air, or in some cases with a pulpy envelope, which flattens into a film. There seem to have been structures of both these kinds, though in the state of preservation of these curious seeds it is extremely difficult to distinguish them. In the first case they must have been intended for dissemination by the wind, like the seeds of spruces. In the latter case they may have been disseminated like the fruits of taxine trees by the agency of animals, though what these were it would be difficult to guess. These trees had very great reproductive power, since they produced numerous seeds, not singly or a few together, as in modern yews, but in long spikes or catkins bearing many seeds (Fig. 59).
It is to be observed that the Cordaites, or the Cordaitinæ, as they have been called, as a family,[CF] constitute another of those intermediate groups with which we have already become familiar. On the one hand they approach closely to the broader-leaved yews like Gingko, Phyllocladus, and Podocarpus, and, on the other hand, they have affinities with Cycadaceæ, and even with Sigillariæ. They were beautiful and symmetrical trees, adding something to the variety of the rather monotonous Palæozoic forests. They contributed also somewhat to the accumulation of coal. I have found that some thin beds are almost entirely composed of their leaves, and the tissues of their wood are not infrequent in the mineral charcoal of the larger coal-seams. There is no evidence that their roots were of the stigmaroid type, though they evidently grew in the same swampy flats with the Sigillariæ and Calamites.
[CF] Engler; Cordaitées of Renault.
It may, perhaps, be well to say here that I believe there was a considerably wide range of organisation in the Cordaitinæ as well as in the Calamites and Sigillariæ, and that it will eventually be found that there were three lines « 134 » of connection between the higher cryptogams and the phænogams, one leading from the lycopods by the Sigillariæ, another leading by the Cordaites, and the third leading from the Equisetums by the Calamites. Still further back the characters afterward separated in the club-mosses, mare’s-tails, and ferns, were united in the Rhizocarps, or, as some now, but I think somewhat unreasonably, prefer to call them, the “heterosporous Filicinæ.” In the more modern world, all the connecting links have become extinct and the phænogams stand widely separated from the higher cryptogams. I do not make these remarks in a Darwinian sense, but merely to state what appear to be the lines of natural affinity and the links wanting to give unity to the system of nature.
Of all the trees of the modern world, none are perhaps so widely distributed as the pines and their allies. On mountain-tops and within the Arctic zone, the last trees that can struggle against the unfavourable conditions of existence are the spruces and firs, and in the warm and moist islands of the tropics they seem equally at home with the tree-ferns and the palms. We have already seen that they are a very ancient family, and in the sandstones of the coal-formation their great trunks are frequently found, infiltrated with calcareous or silicious matter, and still retaining their structure in the greatest perfection (Fig. 60). So far as we know, the foliage of some of them which constitutes the genera Walchia and Araucarites of some authors (Figs. 60, 63) was not dissimilar from that of modern yews and spruces, though there is reason to believe that some others had broad, fern-like leaves like those of the gingko. None of them, so far as yet certainly known, were cone-bearing trees, their fruit having probably been similar to that of the yews (Fig. 61). The minute structures of their stems are nearer to those of the conifers of the islands of the southern hemisphere than to that of those in our northern climes—a correlation, no doubt, to the equable climate of the period. There is not much evidence that they grew with the Sigillariæ in the true coal-swamps, though some specimens have been found in this association. It is more likely that they were in the main inland and upland trees, and that in consequence they are mostly known to us by drifted trunks borne by river inundations into the seas and estuaries.
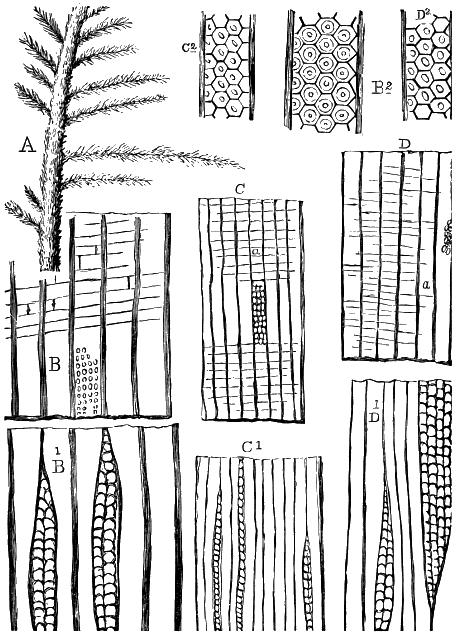
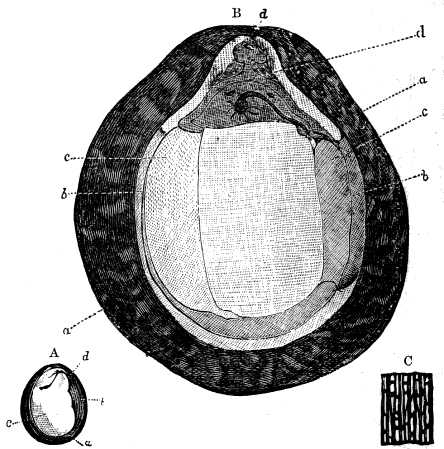
A remarkable fact in connection with them, and showing also the manner in which the most durable vegetable structures may perish by decay, is that, like the Cordaites, they had large piths with transverse partitions, a structure « 137 » which, as I have already mentioned, appears on a minute scale in the twigs of the fir-tree, and that sometimes casts of these piths in sandstone appear in a separate form, constituting what have been named Sternbergiæ or Artisiæ. As Renault well remarks with reference to Cordaites, the existence of this chambered form of pith implies rapid elongation of the stem, so that the Cordaites and conifers of the coal-formation were probably quickly growing trees (Fig. 62).
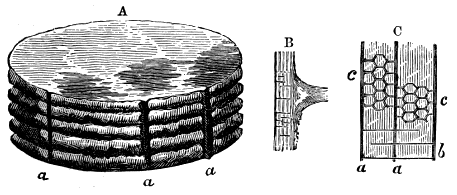
The same general statements may be made as to the coal-vegetation as in relation to that of the Erian. In the coal period we have found none of the higher exogens, and there are only obscure and uncertain indications of the presence of endogens, which we may reserve for a future chapter; but gymnosperms abound and are highly characteristic. On the other hand, we have no mosses or lichens, and very few Algæ, but a great number of ferns and Lycopodiaceæ or club-mosses (Fig. 63). Thus, the coal-formation period is botanically a meeting-place of the lower phænogams and the higher cryptogams, and presents many forms which, when imperfectly known, have puzzled botanists in regard to their position in one or other series. In the present world, the flora most akin « 138 » to that of the coal period is that of warm, temperate regions in the southern hemisphere. It is not properly a tropical flora, nor is it the flora of a cold region, but rather indicative of a moist and equable climate. Still, we must bear in mind that we may often be mistaken in reasoning as to the temperature required by extinct species of plants, differing from those now in existence. Further, we must not assume that the climatal conditions of the northern hemisphere were in the coal period at all similar to those which now prevail. As Sir Charles Lyell has shown, a less amount of land in the higher latitudes would greatly modify climates, and there is every reason to believe that in the coal period there was less land than now. Further, it has been shown by Tyndall that a very small additional amount of carbonic acid in the atmosphere would, by obstructing the radiation of heat from the earth, produce almost the effect of a glass roof or conservatory, extending over the whole world. Again, there is much in the structure of the leaves of the coal-plants, as well as in the vast amount of carbon which they accumulated in the form of coal, and the characteristics of the animal life of the period, to indicate, on independent « 139 » grounds, that the carboniferous atmosphere differed from that of the present world in this way, or in the presence of more carbonic acid—a substance now existing in the very minute proportion of one thousandth of the whole—a quantity adapted to the present requirements of vegetable and animal life, but probably not to those of the coal period.
Thus, if we inquire as to any analogous distribution of plants in the modern world, we find this only in the warmer insular climates of the southern hemisphere, where ferns, lycopods, and pines appear under forms somewhat akin to those of the Carboniferous, but mixed with other types, some of which are modern, others allied to those of the next succeeding geological ages of the Mesozoic and Tertiary; and under these periods it will be more convenient to make comparisons.
The readers of recent English popular works on geology will have observed the statement reiterated that a large proportion of the material of the great beds of bituminous coal is composed of the spore-cases of lycopodiaceous plants—a statement quite contrary to that resulting from my microscopical examinations of the coal of more than eighty coal-beds in Nova Scotia and Cape Breton, as stated in “Acadian Geology” (page 463), and more fully in my memoir of 1858 on the “Structures in Coal,”[CG] and that of 1866, on the “Conditions of Accumulation of Coal.”[CH] The reason of this mistake is, that an eminent English naturalist, happening to find in certain specimens of English coal a great quantity of remains of spores and spore-cases, though even in his specimens they constitute only a small portion of the mass, and being apparently unacquainted with what others had done in this field, wrote a popular article for the “Contemporary Review,” in which he extended an isolated and « 140 » exceptional fact to all coals, and placed this supposed origin of coal in a light so brilliant and attractive that he has been followed by many recent writers. The fact is, as stated in “Acadian Geology,” that trunks of Sigillariæ and similar trees constitute a great part of the denser portion of the coal, and that the cortical tissues of these rather than the wood remain as coal. But cortical or epidermal tissues in general, whether those of spore-cases or other parts of plants, are those which from their resistance to water-soakage and to decay, and from their highly carbonaceous character, are best suited to the production of coal. In point of fact, spore-cases, though often abundantly present, constitute only an infinitesimal part of the matter of the great coal-beds. In an article in “The American Journal of Science,” which appeared shortly after that above referred to, I endeavoured to correct this error, though apparently without effect in so far as the majority of British geological writers are concerned. From this article I have taken with little change the following passages, as it is of importance in theoretical geology that such mistakes, involving as they do the whole theory of coal accumulation, should not continue to pass current. The early part of the paper is occupied with facts as to the occurrence of spores and spore-cases as partial ingredients in coal. Its conclusions are as follows: It is not improbable that sporangites, or bodies resembling them, may be found in most coals; but it is most likely that their occurrence is accidental rather than essential to coal accumulation, and that they are more likely to have been abundant in shales and cannel coals, deposited in ponds or in shallow waters in the vicinity of lycopodiaceous forests, than in the swampy or peaty deposits which constitute the ordinary coals. It is to be observed, however, that the conspicuous appearance which these bodies, and also the strips and fragments of epidermal tissue, which resemble them in « 141 » texture, present in slices of coal, may incline an observer, not having large experience in the examination of coals, to overrate their importance; and this I think has been done by most microscopists, especially those who have confined their attention to slices prepared by the lapidary. One must also bear in mind the danger arising from mistaking concretionary accumulations of bituminous matter for sporangia. In sections of the bituminous shales accompanying the Devonian coal above mentioned, there are many rounded yellow spots, which on examination prove to be the spaces in the epidermis of Psilophyton through which the vessels passing to the leaves were emitted. To these considerations I would add the following, condensed from the paper above referred to (p. 139), in which the whole question of the origin of coal is fully discussed:[CI]
[CG] “Journal of the Geological Society,” vol. xv.
[CH] Ibid., vol. xxii.
[CI] See also “Acadian Geology,” 2d ed., pp. 138, 461, 493.
1. The mineral charcoal or ‘mother coal’ is obviously woody tissue and fibres of bark, the structure of the varieties of which, and the plants to which it probably belongs, I have discussed in the paper above mentioned.
2. The coarser layers of coal show under the microscope a confused mass of fragments of vegetable matter belonging to various descriptions of plants, and including, but not usually in large quantities, sporangites.
3. The more brilliant layers of the coal are seen, when separated by thin laminæ of clay, to have on their surfaces the markings of Sigillariæ and other trees, of which they evidently represent flattened specimens, or rather the bark of such specimens. Under the microscope, when their structures are preserved, these layers show cortical tissues more abundantly than any others.
4. Some thin layers of coal consist mainly of flattened layers of leaves of Cordaites or Pychnophyllum.
5. The Stigmaria under-clays and the stumps of « 142 » Sigillaria in the coal-roofs equally testify to the accumulation of coal by the growth of successive forests, more especially of Sigillariæ. There is, on the other hand, no necessary connection of sporangite-beds with Stigmarian soils. Such beds are more likely to be accumulated in water, and consequently to constitute bituminous shales and cannels.
6. Lepidodendron and its allies, to which the spore-cases in question appear to belong, are evidently much less important to coal accumulation than Sigillaria, which cannot be affirmed to have produced spore-cases similar to those in question, even though the observation of Goldenberg as to their fruit can be relied on; the accuracy of which, however, I am inclined to doubt.
On the whole, then, while giving due credit to those who have advocated the spore-theory of coal, for directing attention to this curious and no doubt important constituent of mineral fuel, and admitting that I may possibly have given too little attention to it, I must maintain that sporangite-beds are exceptional among coals, and that cortical and woody matters are the most abundant ingredients in all the ordinary kinds; and to this I cannot think that the coals of England constitute an exception.
It is to be observed, in conclusion, that the spore-cases of plants, in their indestructibility and richly carbonaceous character, only partake of qualities common to most suberous and epidermal matters, as I have explained in the publications already referred to. Such epidermal and cortical substances are extremely rich in carbon and hydrogen, in this resembling bituminous coal. They are also very little liable to decay, and they resist more than other vegetable matters aqueous infiltration—properties which have caused them to remain unchanged, and to continue free from mineral additions more than other vegetable tissues. These qualities are well seen in the bark of our American white birch. It is no wonder that « 143 » materials of this kind should constitute considerable portions of such vegetable accumulations as the beds of coal, and that when present in large proportion they should afford richly bituminous beds. All this agrees with the fact, apparent on examination of the common coal, that the greater number of its purest layers consist of the flattened bark of Sigillariæ and similar trees, just as any single flattened trunk embedded in shale becomes a layer of pure coal. It also agrees with the fact that other layers of coal, and also the cannels and earthy bitumens, appear under the microscope to consist of finely comminuted particles, principally of epidermal tissues, not only from the fruits and spore-cases of plants, but also from their leaves and stems. These considerations impress us, just as much as the abundance of spore-cases, with the immense amount of the vegetable matter which has perished during the accumulation of coal, in comparison with that which has been preserved.
I am indebted to Dr. T. Sterry Hunt for the following very valuable information, which at once places in a clear and precise light the chemical relations of epidermal tissue and spores with coal. Dr. Hunt says: "The outer bark of the cork-tree, and the cuticle of many if not all other plants, consists of a highly carbonaceous matter, to which the name of suberin has been given. The spores of Lycopodium also approach to this substance in composition, as will be seen by the following, one of two analyses by Duconi,[CJ] along with which I give the theoretical composition of pure cellulose or woody fibre, according to Payen and Mitscherlich, and an analysis of the suberin of cork, from Quercus suber, from which the ash and 2·5 per cent of cellulose have been deducted.[CK]
[CJ] Liebig and Kopp, “Jahresbuch,” 1847-'48.
| Cellulose. | Cork. | Lycopodium. | |
| Carbon | 44·44 | 65·73 | 64·80 |
| Hydrogen | 6·17 | 8·33 | 8·73 |
| Nitrogen | .... | 1·50 | 6·18 |
| Oxygen | 49·39 | 24·44 | 20·29 |
| Total | 100·00 | 100·00 | 100·00 |
"This difference is not less striking when we reduce the above centesimal analyses to correspond with the formula of cellulose, C24H20O20, and represent cork and Lycopodium as containing twenty-four equivalents of carbon. For comparison I give the composition of specimens of peat, brown coal, lignite, and bituminous coal:[CL]
[CL] “Canadian Naturalist,” vi., 253.
| Cellulose | C24H20O20 |
| Cork | C24H182/10O67/10 |
| Lycopodium | C24H194/10NO56/10 |
| Peat (Vaux) | C24H144/10O10 |
| Brown coal (Schröther) | C24H143/10O106/10 |
| Lignite (Vaux) | C24H113/10O64/10 |
| Bituminous coal (Regnault) | C24H10O33/10 |
“It will be seen from this comparison that, in ultimate composition, cork and Lycopodium are nearer to lignite than to woody fibre, and may be converted into coal with far less loss of carbon and hydrogen than the latter. They in fact approach closer in composition to resins and fats than to wood, and, moreover, like those substances repel water, with which they are not easily moistened, and thus are able to resist those atmospheric influences which effect the decay of woody tissue.”
I would add to this only one further consideration. The nitrogen present in the Lycopodium spores, no doubt, belongs to the protoplasm contained in them, a substance which would soon perish by decay; and subtracting this, the cell-walls of the spores and the walls of the spore-cases « 145 » would be most suitable material for the production of bituminous coal. But this suitableness they share with the epidermal tissue of the scales of strobiles, and of the stems and leaves of ferns and lycopods, and, above all, with the thick, corky envelope of the stems of Sigillariæ and similar trees, which, as I have elsewhere shown,[CM] from its condition in the prostrate and erect trunks contained in the beds associated with coal, must have been highly carbonaceous and extremely enduring and impermeable to water. In short, if, instead of “spore-cases,” we read “epidermal tissues in general, including spore-cases,” all that has been affirmed regarding the latter will be strictly and literally true, and in accordance with the chemical composition, microscopical characters, and mode of occurrence of coal. It will also be in accordance with the following statement, from my paper on the “Structures in Coal,” published in 1859:
[CM] “Vegetable Structures in Coal,” “Journal of Geological Society,” xv., 626. “Conditions of Accumulation of Coal,” ibid., xxii., 95. “Acadian Geology,” 197, 464.
“A single trunk of Sigillaria in an erect forest presents an epitome of a coal-seam. Its roots represent the Stigmaria under-clay; its bark the compact coal; its woody axis the mineral charcoal; its fallen leaves (and fruits), with remains of herbaceous plants growing in its shade, mixed with a little earthy matter, the layers of coarse coal. The condition of the durable outer bark of erect trees concurs with the chemical theory of coal, in showing the especial suitableness of this kind of tissue for the production of the purer compact coals. It is also probable that the comparative impermeability of the bark to mineral infiltration is of importance in this respect, enabling this material to remain unaffected by causes which have filled those layers, consisting of herbaceous materials and decayed wood, with pyrites and other mineral substances.”
We need not go far in search of the uses of the coal vegetation, when .we consider the fact that the greatest civilised nations are dependent on it for their fuel. Without the coal of the Carboniferous period and the iron-ore which is one of the secondary consequences of coal accumulation, just as bog-ores of iron occur in the subsoils of modern peats, it would have been impossible either to sustain great nations in comfort in the colder climates of the northern hemisphere or to carry on our arts and manufactures. The coal-formation yields to Great Britain alone about one hundred and sixty million tons of coal annually, and the miners of the United States extract mainly from the same formation nearly a hundred million tons, while the British colonies and dependencies produce about five million tons; and it is a remarkable fact that it is to the English race that the greatest supply of this buried power and heat and light has been given.
The great forests of the coal period, while purifying the atmosphere of its excess of unwholesome carbonic acid, were storing up the light and heat of Palæozoic summers in a form in which they could be recovered in our human age, so that, independently of their uses to the animals which were their contemporaries, they are indispensable to the existence of civilised man.
Nor can we hope soon to be able to dispense with the services of this accumulated store of fuel. The forests of to-day are altogether insufficient for the supply of our wants, and though we are beginning to apply water-power to the production of electricity, and though some promising plans have been devised for the utilisation of the direct heat and light of the sun, we are still quite as dependent as any of our predecessors on what has been done for us in the Palæozoic age.
In the previous pages I have said little respecting the physical geography of the Carboniferous age; but, as may « 147 » be inferred from the vegetation, this in the northern hemisphere presented a greater expanse of swampy flats little elevated above the sea than we find in any other period. As to the southern hemisphere, less is known, but the conditions of vegetation would seem to have been essentially the same.
Taking the southern hemisphere as a whole, I have not seen any evidence of a Lower Devonian or Upper Silurian flora; but in South Africa and Australia there are remains of Upper Devonian or Lower Carboniferous plants. These were succeeded by a remarkable Upper Carboniferous or Permian group, which spread itself all over India, Australia, and South Africa,[CN] and contains some forms (Vertebraria, Phyllotheca, Glossopteris, &c. ) not found in rocks of similar age in the northern hemisphere, so that, if the age of these beds has been correctly determined, the southern hemisphere was in advance in relation to some genera of plants. This, however, is to be expected when we consider that the Triassic and Jurassic flora of the north contains or consists of intruders from more southern sites. These beds are succeeded in India by others holding cycads, &c., of Upper Jurassic or Lower Cretaceous types (Rajmahal and Jabalpur groups).
[CN] Wyley, “Journal Geol. Society,” vol. xxiii., p. 172; Daintree, ibid., vol. xxviii.; also Clarke and McCoy.
Blanford has shown that there is a very great similarity in this series all over the Australian and Indian region.[CO] Hartt and Darby have in like manner distinguished Devonian and Carboniferous forms in Brazil akin to those of the northern hemisphere. Thus the southern hemisphere would seem to have kept pace with the northern, and according to Blanford there is evidence there of cold conditions in the Permian, separating the Palæozoic « 148 » flora from that of the Mesozoic, in the same manner that Ramsay has supposed a similar period of cold to have done north of the equator. This would imply a very great change of climate, since we have evidence of the extension of the Lower Carboniferous flora at least as far north as Spitzbergen. The upper coal-formation we cannot, however, trace nearly so far north; so that a gradual refrigeration may have been going on before the Permian. Thus in both hemispheres there was a general similarity in the later Palæozoic flora, and perhaps similar conditions leading to its extinction and to its replacement by that to be described in the next chapter.
[CO] “Journal Geol. Society,” vol. xxxi.
NOTES TO CHAPTER IV.
I. Characters and Classification of Palæozoic Plants.
In the space available in this work it would be impossible to enter fully into the classification of Palæozoic plants; but it may be well to notice some important points for the guidance of those who may desire to collect specimens; more especially as much uncertainty exists as to affinities and very contradictory statements are made. The statements below may be regarded as the results of actual observation and of the study of specimens in situ in the rocks, as well as in the cabinet and under the microscope.
Gymnospermeæ.
Family Coniferæ; Genus Dadoxylon, Endlicher; Araucarites, Goeppert; Araucarioxylon, Kraus.
The trunks of this genus occur from the Middle Devonian to the Permian inclusive, as drift-logs calcified, silicified, or pyritised. The only foliage associated with them is of the type of Walchia and Araucarites—viz., slender branches with numerous small spiral acicular leaves. Two of the coal-formation species, D. materiarum and another, had foliage of this type. That of the others is unknown. They are all distinct from the wood of Cordaites, for which see under that genus.
The following are North American species:
| Trunks. | |||
| Dadoxylon Ouangondianum, Dn. | M. Erian | Report, 1871.[CP] | |
| D. Halli, Dn. | ” | ” | |
| D. Newberryi, Dn. | ” | ” | |
| D. Clarkii, Dn. (Cordæoxylon ?) | ” | Report, 1882. | |
| D. Acadianum, Dn. | Coal-formation and millstone grit. |
Acadian Geology. | |
| D. Materiarum, Dn. | Do. and Permo-Carb. | ” | |
| D. (Palæoxylon) antiquius, Dn. | L. Carboniferous. | ” | |
| D. annulatum, Dn. | Coal-formation. | ” | |
| Ormoxylon Erianum, Dn. | Erian. | Report, 1871. | |
| Foliage. | |||
| Araucarites gracilis, Dn. | N. Coal-formation and Permian. | ” | |
| Walchia robusta, Dn. | Permian. | Report on Prince Edward Island. |
|
| W. imbricatula, Dn. | ” | ||
[CP] “Geological Survey of Canada: Fossil Plants of Erian and Upper Silurian Formations,” by J. W. Dawson.
All of the above can be vouched for as good species based upon microscopic examination of a very large number of trunks from different parts of North America. The three Erian species of Dadoxylon and D. antiquius from the Lower Carboniferous have two or more rows of cells in the medullary rays. The last named has several rows, and is a true Palæoxylon allied to D. Withami of Great Britain. D. materiarium is specially characteristic of the upper coal-formation and Permian, and to it must belong one or both of the species of foliage indicated above. D. Clarkii has very short, simple medullary rays of only a few cells superimposed, and has an inner cylinder of scalariform vessels, approaching in these points to Cordaites. Ormoxylon has a very peculiar articulated pith and simple medullary rays.
Witham in 1833 described several Carboniferous species of pine-wood, under the generic name Pinites, separating under the name Pitus species which appeared to have the discs on the cell-walls « 150 » separate and in transverse lines. Witham’s name was changed by Goeppert to Araucarites, to indicate the similarity of these woods to Araucaria, Pinites being reserved for trees more closely allied to the ordinary pines. Endlicher, restricting Araucarites to foliage, etc., of Araucaria-like trees, gave the name Dadoxylon to the wood; and this, through Unger’s “Genera and Species,” has gained somewhat general acceptance. Endlicher also gave the name Pissadendron to the species which Witham had called Pitus; but Brongniart proposed the name Palæoxylon to include all the species with thick and complex medullary rays, whatever the arrangement of the discs. In Schimper’s new work Kraus substitutes Araucarioxylon for Endlicher’s Dadoxylon, and includes under Pissadendron all the species placed by Brongniart in Palæoxylon.
To understand all this confusion, it may be observed that the characters available in the determination of Palæozoic coniferous wood are chiefly the form and arrangement of the wood-cells, the character of the bordered pores or discs of their walls, and the form and composition of the medullary rays.
The character on which Witham separated his genus Pitus from Pinites is, as I have ascertained by examination of slices of one of his original specimens kindly presented to me by Mr. Sanderson, of Edinburgh, dependent on state of preservation, the imperfectly preserved discs or areolations of the walls of the fibre presenting the appearance of separate and distinct circles, while in other parts of the same specimens these discs are seen to be contiguous and to assume hexagonal forms, so that in this respect they do not really differ from the ordinary species of Dadoxylon. The true character for subdividing those species which are especially characteristic of the Carboniferous, is the composite structure of the medullary rays, which are thick and composed of several radial piles of cells placed side by side. This was the character employed by Brongniart in separating the genus Palæoxylon, though he might with convenience have retained Witham’s name, merely transferring to the genus the species of Witham’s Pinites which have complex medullary rays. The Erian rocks present the greatest variety of types, and Palæoxylon is especially characteristic of the Lower Carboniferous, while species of Dadoxylon with two rows of bordered pores and simple medullary rays are especially plentiful in the upper coal-formation and Permo-Carboniferous.
The following table will clearly show the distinctive characters and relations of the genera in question, as held by the several authors above referred to:
Wood of Palæozoic Conifers.
| Woody fibres. | Medullary rays and pith. | Generic names. | Geological age. | ||
| No discs. | One or two series of cells. | Aporoxylon, Unger. | Devonian (Erian). | ||
| Discs in one series contiguous, or in several series spirally arranged. |
Complex, or of two or more series of cells. Pith Sternbergian. |
|
Middle and Lower Carboniferous and Devonian. |
||
| Simple, or of one row of cells. Pith Sternbergian. |
|
Upper Carboniferous and Permian. |
|||
| Pith in spherical chambers. | Ormoxylon,[CQ] Dn. | Devonian. | |||
| Medullary sheath scalariform. Medullary rays frequent, simple, short. |
Dadoxylon (Cordæoxylon),[CR] Dn. | Devonian. |
[CQ] Type O. Erianum, Dn., “Report on Canadian Plants,” 1871.
[CR] Type D. Clarkii, Dn., “Report on Canadian Plants,” 1882. This may be wood of Cordaites, to which it approaches very closely.
Family Cordaites, Genus Cordaites, Brongniart.
Trunks marked by transverse scars of attachment of bases of leaves; leaves broad, with many parallel veins, and attached by a broad base; pistillate and staminate catkins of the nature of Antholithes. Fruit winged or pulpy, of the kind known as Cardiocarpum. Stem with a Sternbergia pith, usually large, surrounded by a ring of pseudo-scalariform vessels, and with a cylinder usually narrow, of woody wedges, with bordered pores in one or more series, and with simple medullary rays.
From specimens kindly presented to me by Prof. Renault, I have been able to ascertain that the stems of some at least of these plants (Eucordaites) are distinct in structure from all the species of Dadoxylon, above mentioned, except D. Clarkii, of the Erian. They may be regarded as intermediate between those of conifers and cycads, which is indeed the probable position of these remarkable plants.
Grand d’Eury has divided the Cordaites into sub-genera, as follows:
1. Eucordaites.—Leaves spatulate, obovate, elliptical, or lanceolate, « 152 » sessile, entire, with rounded apices and of leathery consistency. The leaves are from twenty to ninety centimetres in length. The nerves are either equally or unequally strong.
2. Dorycordaites.—Leaves lanceolate, with sharp points; nerves numerous, fine, and equal in strength. The leaves attain a length of from forty to fifty centimetres.
3. Poacordaites.—Leaves narrow, linear, entire, blunt at the point, with nerves nearly equally strong. The leaves are as much as forty centimetres in length.
To these Renault and Zeiller have added a fourth group, Scutocordaites.
Genus Sternbergia.
This is merely a provisional genus intended to receive casts of the pith cylinders of various fossil trees. Their special peculiarity is that, as in the modern Cecropia peltata, and some species of Ficus, the pith consists of transverse dense partitions which, on the elongation of the internodes, become separated from each other, so as to produce a chambered pith cavity, the cast of which shows transverse furrows. The young twigs of the modern Abies balsamifera present a similar structure on a minute scale. I have ascertained and described such pith-cylinders in large stems of Dadoxylon Ouangondianum, and D. materiarium. They occur also in the stems of Cordaites and probably of Sigillariæ. I have discussed these curious fossils at length in “Acadian Geology” and in the “Journal of the Geological Society of London,” 1860. The following summary is from the last-mentioned paper:
a. As Prof. Williamson and the writer have shown, many of the Sternbergia piths belong to coniferous trees of the genus Dadoxylon.
b. A few specimens present multiporous tissue, of the type of Dictyoxylon, a plant of unknown affinities, and which, according to Williamson, has a Sternbergia pith.
c. Other examples show a true scalariform tissue, comparable with that of Lepidodendron or Sigillaria, but of finer texture. Corda has shown that plants of the type of the former genus (his Lomatophloios) had Sternbergia piths. Some plants of this group are by external characters loosely reckoned by botanists as ribless Sigillariæ (Clathraria); but I believe that they are not related even ordinally to that genus.
d. Many Carboniferous Sternbergiæ show structures identical with those described above as occurring in Cordaites, and also in some of the trees ordinarily reckoned as Sigillariæ.
Genus Cardiocarpum.
I have found at least eight species of these fruits in the Erian and Carboniferous of New Brunswick and Nova Scotia, all of which are evidently fruits of gymnospermous trees. They agree in having a dense coaly nucleus of appreciable thickness, even in the flattened specimens, and surrounded by a thin and veinless wing or margin. They have thus precisely the appearance of samaras of many existing forest-trees, some of which they also resemble in the outline of the margin, except that the wings of samaras are usually veiny. The character of the nucleus, and the occasional appearance in it of marks possibly representing cotyledons or embryos, forbids the supposition that they are spore-cases. They must have been fruits of phænogams. Whether they were winged fruits or seeds, or fruits with a pulpy envelope like those of cycads and some conifers, may be considered less certain. The not infrequent distortion of the margin is an argument in favour of the latter view, though this may also be supposed to have occurred in samaras partially decayed. On the other hand, their being always apparently flattened in one plane, and the nucleus being seldom, if ever, found denuded of its margin, are arguments in favour of their having been winged nutlets or seeds. Until recently I had regarded the latter view as more probable, and so stated the matter in the second edition of “Acadian Geology.” I have, however, lately arrived at the conclusion that the Cardiocarpa of the type of C. cornutum were gymnospermous seeds, having two cotyledons embedded in an albumen and covered with a strong membranous or woody tegmen surrounded by a fleshy outer coat, and that the notch at the apex represents the foramen or micropyle of the ovule. The structure was indeed very similar to that of the seeds of Taxus and of Salisburia. With respect to some of the other species, however, especially those with very broad margins, it still appears likely that they were winged.
The Cardiocarpa were borne in racemes or groups, and it seems certain that some of them at least are the seeds of Cordaites. The association of some of them and of those of the next genus with Sigillariæ is so constant that I cannot doubt that some of them belong to plants of that genus, or possibly to taxine conifers. The great number of distinct species of these seeds, as compared with that of known trees which could have produced them, is very remarkable.
Genus Trigonocarpum.
These are large angled nuts contained in a thick envelope, and showing internal structures resembling those of the seeds of modern « 154 » Taxineæ. There are numerous species, as well as allied seeds referred to the provisional genera Rhabdocarpus and Carpolithes. In Trigonocarpum Hookeri I have described the internal structure of one of those seeds, and many fine examples from the coal-field of St. Etienne, in France, have been described by Brongniart, so that their internal structure is very well known.
Genus Antholithes.
This is also a provisional genus, to include spikes of floral organs, some of which are known to have belonged to Cordaites, others probably to Sigillariæ.
Family Sigillariaceæ.
Under this name palæobotanists have included a great number of trees of the Carboniferous system, all of which are characterised by broad leaf-sears, with three vascular scars, and usually arranged in vertical rows, and by elongated three-nerved leaves, and roots of the stigmaria type—that is, with rounded pits, marking the attachment of rootlets spirally arranged. These trees, however, collected in the genus Sigillaria by arbitrary characters, which pass into those of the Lepidodendroid trees, have been involved in almost inextricable confusion, to disentangle which it will be necessary to consider: 1. The external characters of Sigillariæ, and trees confounded with them. 2. Subdivision of Sigillariæ by external markings. 3. The microscopic character of their stems. 4. What is known of their foliage and fruit.
1. Characters of Sigillaroid and Lepidodendroid Trunks.
It may be premised that the modes of determination in fossil botany are necessarily different from those employed in recent botany. The palæobotanist must have recourse to characters derived from the leaves, the scars left by their fall, and the internal structures of the stem. These parts, held in little esteem by botanists in describing modern plants, and much neglected by them, must hold the first place in the regard of the fossil botanist, whereas the fructification, seldom preserved, and generally obscure, is of comparatively little service. It is to be remarked also that in such generalised plants as those of the Palæozoic, remarkable rather for the development of the vegetative than of the reproductive organs, the former rise in importance as compared with their value in the study of modern plants.
In Sigillariæ, Lepidodendra, &c., the following surfaces of the stem may be presented to our inspection:
1. The outer surface of the epidermis without its leaves, but with the leaf-bases and leaf-scars more or less perfectly preserved. On this surface we may recognise: (1) Cellular swellings or projections of the bark to which the leaves are attached. These may be called leaf-bases, and they are sometimes very prominent. (2) The actual mark of the attachment of the leaf situated in the most prominent part of the leaf-base. This is the leaf-scar. (3) In the leaf-scar when well preserved we can see one or more minute punctures or prominences which are the points where the vascular bundles passing to the leaf found exit. These are the vascular scars.
When the leaves are attached, the leaf-scars and vascular scars cannot be seen, but the leaf-bases can be made out. Hence it is important, if possible, to secure specimens with and without the leaves. In flattened specimens the leaf-bases are often distorted by pressure and marked with furrows which must not be mistaken for true structural characters. The leaf-bases, which are in relief on the outer surface of the stem, of course appear as depressions on the mould in the containing rock, in which the markings often appear much more distinctly than on the plant itself.
2. The outer surface of the epidermis may have been removed or may be destroyed by the coarseness of the containing rock. In this case the leaf-bases are usually preserved on the surface of the outer or corky bark, but the leaf-scars and vascular scars have disappeared. This gives that condition of Lepidodendroid trees to which the name Knorria has been applied. When plants are in this state careful inspection may sometimes discover traces of the leaf-scars on portions of the stem, and thus enable the Knorria to be connected with the species to which it belongs.
3. The outer or corky bark may be removed, exposing the surface of the inner or fibrous and cellular bark, which in the plants in question is usually of great thickness. In this case neither the leaf-bases nor the scars are seen, but punctures or little furrows or ridges appear where the vascular bundles entered the inner bark. Specimens in this state are usually said to be decorticated, though only the outer bark is removed. It is often difficult to determine plants in this condition, unless some portion of the stem can be found still retaining the bark; but when care is taken in collecting, it will not infrequently be found that the true outer surface can be recovered from the containing rock, especially if a coaly layer representing the outer bark intervenes between this and the inner impression. Specimens « 156 » of this kind, taken alone, have been referred to the genera Knorria, Bothrodendron, and Halonia.
4. In some cases, though not frequently, the outer surface of the ligneous cylinder is preserved. It almost invariably presents a regularly striated or irregularly wrinkled appearance, depending upon the vertical woody wedges, or the positions of the medullary rays or vascular bundles. Specimens of this kind constituted some of the Endogenites of the older botanists, and the genus Schizodendron of Eichwald appears to include some of them. Many of them have also been incorrectly referred to Calamites.
5. In some cases the cast of the medullary cylinder or pith may alone be preserved. This may be nearly smooch or slightly marked by vertical striæ, but more usually presents a transverse striation, and not infrequently the transverse constrictions and septa characteristic of the genus Sternbergia. Loose Sternbergiæ afford little means of connecting them with the species to which they belong, except by the microscopic examination of the shreds of the ligneous cylinder which often cling to them.[CS]
[CS] See my paper, “Journal of Geological Society,” vol. xxvii.
These facts being premised, the following general statements may be made respecting some of the more common Palæozoic genera, referring, however, principally to the perfect markings as seen on the epidermis:
Sigillaria.—Leaf-bases hexagonal or elongated, or confluent on a vertical ridge. Leaf-scars hexagonal or shield-shaped. Vascular scars three, the two lateral larger than the central. This last character is constant, depending on the fact that the leaves of Sigillaria have two or more vascular bundles. All so-called Sigillariæ having the central vascular scar largest, or only one vascular bundle, should be rejected from this genus. In young branches of branching Sigillariæ the leaf-scars sometimes appear to be spiral, but in the older stems they form vertical rows; interrupted, however, by transverse rows or bands of fruit-scars, each with a single large central vascular scar, and which have borne the organs of fructification. Arthrocaulis of McCoy is founded on this peculiarity.
Syringodendron.—Differs from Sigillaria in the leaf-scars, which are circular and with a single vascular bundle. It is a matter of doubt whether these plants were of higher rank than Sigillaria tending toward the pines, or of lower rank tending toward Cyclostigma. Their leaf-bases form vertical ridges.
Lepidodendron.—Leaf-bases rhombic, oval, or lanceolate, moderately « 157 » prominent. Leaf-scars rhombic or sometimes shield-shaped or heart-shaped, in the middle or upper part of the leaf-base. Vascular scars three—the middle one always largest and corresponding to the single nerve of the leaf; the lateral ones sometimes obsolete.
In older stems three modes of growth are observed. In some species the expansion of the bark obliterates the leaf-bases and causes the leaf-scars to appear separated by wide spaces of more or less wrinkled bark, which at length becomes longitudinally furrowed and simulates the ribbed character of Sigillaria. In others the leaf-bases grow in size as the trunk expands, so that even in large trunks they are contiguous though much larger than those on the branches. In others the outer bark, hardening at an early age, is incapable of either of the above changes, and merely becomes cleft into deep furrows in the old trunks.
Lepidophloios.—Leaf-bases transverse and prominent—often very much so. Leaf-scars transversely rhombic or oval with three vascular scars, the central largest. Leaves very long and one-nerved. Large strobiles or branchlets borne in two ranks or spirally on the sides of the stem, and leaving large, round scars (cone-scars), often with radiating impressions of the basal row of scales.
Species with long or drooping leaf-bases have been included in Lepidophloios and Lomatophloios, Species with short leaf-bases and cone-scars in two rows have been called Ulodendron, and some of them have been included in Sigillaria (sub-genus Clathraria). Decorticated stems are Bothrodendron and Halonia. Some of the species approach near to the last genus, especially to the Lepidodendra with rhombic leaf-bases like L. tetragonum.
Cyclostigma.—Leaf-bases undeveloped. Leaf-scars circular or horseshoe-shaped, small, with a central vascular scar. In old trunks of Cyclostigma the leaf-scars become widely separated, and sometimes appear in vertical rows. Young branches of Lepidodendron sometimes have the leaf-scars similar to those of Cyclostigma.
Leptophleum.—Leaf-bases flat, rhombic; leaf-scars obsolete; vascular scar single, central. The last two genera are characteristically Devonian.
In contradistinction from the trees above mentioned, the following general statements may be made respecting other groups:
In conifers the leaf-bases are usually elongated vertically, often scaly in appearance, and with the leaf-scar terminal and round, oval, or rhombic, and with a single well-marked vascular scar.
In Calamites, Calamodendron, and Asterophyllites the scars of the branchlets or leaves are circular or oval, with only a single vascular « 158 » scar, and situated in verticils at the top of well-marked nodes of the stem.
In tree-ferns the leaf-bases are large and usually without a distinct articulating surface. The vascular bundles are numerous. Protopteris has rounded leaf-scars with a large horseshoe-shaped bundle of vessels above and small bundles below. Caulopteris has large elliptic or oval leaf-scars with vascular scars disposed concentrically. Palæopteris,[CT] of Geinitz, has the leaf-scars transversely oval and the vascular bundles confluent in a transverse band with an appendage or outlying bundle below. Stemmatopteris has leaf-scars similar to those of Caulopteris, but the vascular bundles united into a horseshoe-shaped band.
[CT] This name, preoccupied by Geinitz, has been inadvertently misapplied to the Devonian ferns of the genus Archæopteris.
2. Subdivision of Sigillariæ in Accordance with their Markings.
The following groups may be defined in this way; but, being based on one character only, they are of course in all probability far from natural:
1. Sigillaria, Brongniart. Type, Sigillaria reniformis, Brongniart, or S. Brounii, Dawson.—Stem with broad ribs, usually much broader than the usually oval or elliptical tripunctate areoles, but disappearing at base, owing to expansion of the stem. Leaves narrow, long, three-nerved.
2. Rhytidolepis, Sternberg. Type, S. scutellata, Brongniart.—Ribs narrow, and often transversely striate. Areoles large, hexagonal or shield-shaped, tripunctate. Leaves as in last group. Kings of rounded scars on the stems and branches mark attachment of fruit. It is possible that some of the smaller stems of this group may be branches of trees of group first.
3. Syringodendron, Sternberg. Type, S. organum, L. and H., S. oculata, Brongniart.—Stems ribbed; areoles small and round, and apparently with a single scar, or three closely approximated. These are rare, and liable to be confounded with decorticated examples of other groups; but I have some specimens which unquestionably represent the external surface.
4. Favularia, Sternberg. Type, Sigillaria elegans of Brongniart.—Leaf-bases hexagonal, or in young branches elliptical, in vertical rows, but without distinct ribs, except in old or decorticated stems. Fruit borne in verticils on the branches bearing transverse rows of rounded scars. Leaves somewhat broad and longitudinally striate.
5. Leioderma, Goldenberg. Type, S. Sydnensis, Dawson.—Ribs obsolete. Cortical and ligneous surfaces striate. Vascular scars double, elongate longitudinally, and alike on cortical and inner surfaces. Areoles in rows and distinct; stigmaria-roots striate, with small and distinct areoles.
6. Clathraria, Brongniart. Type, S. Menardi, Brongniart.—Areoles hexagonal, not in distinct rows, but having a spiral appearance. Some of the plants usually referred to this group are probably branches of Favularia. Others are evidently fragments of plants of the genus Lepidophloios.
3. Internal Structures of Sigillaria-Stems.
I long ago pointed out, on the evidence of the external markings and mode of growth, that the stems of Sigillariæ must have been exogenous, and this conclusion has now been fully confirmed by the microscopic researches of Williamson, not only in the case of Sigillariæ, but of Lepidodendra and Calamodendra as well. Confining myself to my own observations, three types of Sigillariæ are known to me by their internal structures, though I cannot certainly correlate all of these with the external markings referred to above.
1. Diploxylon, in which the stem consists of a small internal axis surrounded by a very thick inner bark and a dense outer cortex. A fine example from the South Joggins is thus described:[CU]
[CU] “Journal of the Geological Society of London,” November, 1877.
"The axis of the stem is about six centimetres in its greatest diameter, and consists of a central pith-cylinder and two concentric coats of scalariform tissue. The pith-cylinder is replaced by sandstone, and is about one centimetre in diameter. The inner cylinder of scalariform tissue is perfectly continuous, not radiated, and about one millimetre in thickness. Its vessels are somewhat crushed, but have been of large diameter. Its outer surface, which readily separates from that of the outer cylinder, is striated longitudinally. The outer cylinder, which constitutes by much the largest part of the whole, is also composed of scalariform tissue; but this is radially arranged, with the individual cells quadrangular in cross-section. The cross-bars are similar on all the sides and usually simple and straight, but sometimes branching or slightly reticulated. The wall intervening between the bars has extremely delicate longitudinal waving lines of ligneous lining, in the manner first described by Williamson as occurring in the scalariform tissue of certain Lepidodendra. A few small radiating spaces, partially « 160 » occupied with pyrites, obscurely represent the medullary rays, which must have been very feebly developed. The radiating bundles passing to the leaves run nearly horizontally; but their structure is very imperfectly preserved. The stem being old and probably long deprived of its leaves, they may have been partially disorganised before it was fossilised. The outer surface of the axis is striated longitudinally, and in some places marked with impressions of tortuous fibres, apparently those of the inner bark. In the cross-section, where weathered, it shows concentric rings; but under the microscope these appear rather as bands of compressed tissue than as proper lines of growth. They are about twenty in number. This tree has an erect, ribbed trunk, twelve feet in height and fifteen inches in diameter, swelling to about two feet at the base."
2. Favularia Type.—This has been well described by Brongniart and by Renault,[CV] and differs from the above chiefly in the fact that the outer exogenous woody zone is composed of reticulated instead of scalariform tissue, and the inner zone is of the peculiar form which I have characterised as pseudo-scalariform.
[CV] “Botanique Fossile,” Paris, 1881.
3. Sigillaria Proper.—This I have illustrated in my paper in the “Journal of the Geological Society” for May, 1871, and it appears to represent the highest and most perfect type of the larger ribbed Sigillaria. This structure I have described as follows, basing my description on a very fine axis found in an erect stem, and on the fragments of the woody axis found in the bases of other erect stems:
a. A dense cellular outer bark, usually in the state of compact coal—but when its structure is preserved, showing a tissue of thickened parenchymatous cells.
b. A very thick inner bark, which has usually in great part perished, or been converted into coal, but which, in old trunks, contained a large quantity of prosenchymatous tissue, very tough and of great durability. This “bast-tissue” is comparable with that of the inner bark of modern conifers, and constitutes much of the mineral charcoal of the coal-seams.
c. An outer ligneous cylinder, composed of wood-cells, either with a single row of large bordered pores,[CW] in the manner of pines « 161 » and cycads, or with two, three, or four rows of such pores sometimes inscribed in hexagonal areoles in the manner of Dadoxylon. This woody cylinder is traversed by medullary rays, which are short, and composed of few rows of cells superimposed. It is also traversed by oblique radiating bundles of pseudo-scalariform tissue proceeding to the leaves. In some Sigillariæ this outer cylinder was itself in part composed of pseudo-scalariform tissue, as in Brongniart’s specimen of S. elegans; and in others its place may have been taken by multiporous tissue, as in a case above referred to; but I have no reason to believe that either of these variations occurred in the typical ribbed species now in question. The woody fibres of the outer cylinder may be distinguished most readily from those of conifers, as already mentioned, by the thinness of their walls, and the more irregular distribution of the pores. Additional characters are furnished by the medullary rays and the radiating bundles of scalariform tissue when these can be observed.
[CW] These are the same with the wood-cells elsewhere called discigerous tissue, and to which I have applied the terms uniporous and multiporous. The markings on the walls are caused by an unlined portion of the cell-wall placed in a disk or depression, and this often surrounded by an hexagonal rim of thickened wall; but in all cases these structures are less pronounced than in Dadoxylon, and less regular in the walls of the same cell, as well as in different layers of the tissues of the axis.
d. An inner cylinder of pseudo-scalariform tissue. I have adopted the term pseudo-scalariform for this tissue, from the conviction that it is not homologous with the scalariform ducts of ferns and other acrogens, but that it is merely a modification of the discigerous wood-cells, with pores elongated transversely, and sometimes separated by thickened bars, corresponding to the hexagonal areolation of the ordinary wood-cells. A similar tissue exists in cycads, and is a substitute for the spiral vessels existing in ordinary exogens.
e. A large medulla, or pith, consisting of a hollow cylinder of cellular tissue, from which proceed numerous thin diaphragms towards the centre of the stem.
These structures of the highest type of Sigillaria are on the one hand scarcely advanced beyond those of Calamopitus, as described by Williamson, and on the other approach to those of Cordaites, as seen in specimens presented to me by Renault.
Finally, as to the fruit of Sigillariæ, I have no new facts to offer. The strobiles or spikes associated with these trees have been variously described as gymnospermous (Renault) or cryptogamous (Groldenberg and Williamson). 1 have never seen them in place. Two considerations, however, have always weighed with me in reference to this subject. One is the constant abundance of Trigonocarpa « 162 » and Cardiocarpa in the soil of the Sigiliaria forests, as I have studied this at the South Joggins. The other is that the rings of fruit-scars on the branches of Sigiliaria are homologous with leaf-scars, not with branches, and therefore should have borne single carpels and not cones or spikes of inflorescence. These are merely suggestions, but I have no doubt they will be vindicated by future discoveries, which will, I have no doubt, show that in the family Sigillariaceæ we have really two families, one possibly of gymnospermous rank, or at least approaching to this, the other allied to the Lepidodendra.
Cryptogamia.
(Acrogenes.)
Family Lepidodendreæ; Genus Lepidodendron, Sternberg.
These are arboreal Lycopods having linear one-nerved leaves, stems branching dichotomously, and with ovate or rhombic leaf-bases bearing rhombic leaf-scars, often very prominent. The fruit is in scaly strobiles, terminal or lateral, and there are usually, if not always, macrospores and microspores in each strobile. The young branches and stems have a central pith, a cylinder of scalariform tubes sending out ascending bundles to the leaves through a thick cellular and fibrous inner bark, and externally a dense cortex confluent with or consisting of the leaf-bases. Older stems have a second or outer layer of scalariform fibres in wedges with medullary rays, and strengthening the stem by a true exogenous growth, much as in the Diploxylon type of Sigiliaria. The development of this exogenous cylinder is different in amount and rate in different species.[CX] This different development of the exogenous axis is accompanied with appropriate external appearances in the stems, and the changes which take place in their markings. These are of three kinds. In some species the areoles, at first close together, become, in the process of the expansion of the stem, separated by intervening spaces of bark in a perfectly regular manner; so that in old stems, while widely separated, they still retain their arrangement, while in young stems they are quite close to one another. This is the case in L. corrugatum. In other species the leaf-scars or bases increase in size in the old stems, still retaining their forms and their contiguity to each other. This is the case in L. undulatum, and generally in those Lepidodendra which have large leaf-bases. In these species the « 163 » continued vitality of the bark is shown by the occasional production of lateral strobiles on large branches, in the manner of the modern red pine of America. In other species the areoles neither increase in size nor become regularly separated by growth of the intervening bark; but in old stems the bark splits into deep furrows, between which may be seen portions of bark still retaining the areoles in their original dimensions and arrangement. This is the case with L. Pictoense. This cracking of the bark no doubt occurs in very old trunks of the first two types, but not at all to the same extent.
[CX] See “Memoirs of Dr. Williamson,” in “Philosophical Transactions,” for ample details.
As a type of Lepidodendron, I may describe one of the oldest Carboniferous species characteristic of the Lower Carboniferous in America, and corresponding to L. Veltheimianum of Europe.
Lepidodendron Corrugatum, Dawson.—(See Fig. 43, supra.) “Quarterly Journal of Geological Society,” vol. xv.; “Acadian Geology,” page 451.
Habit of Growth.—Somewhat slender, with long branches and long, slender leaves having a tendency to become horizontal or drooping.
Markings of Stem.—Leaf-bases disposed in quincunx or spirally, elongate, ovate, acute at both ends, but more acute and slightly oblique at the lower end; most prominent in the upper third, and with a slight vertical ridge. Leaf-scars small, rounded, and showing only a single punctiform vascular scar. The leaf-scar on the outer surface is in the upper third of the base; but the obliquity of the vascular bundle causes it to be nearly central on the inside of the epidermis. In young succulent shoots the leaf-scars are contiguous and round as in Cyclostigma, without distinct leaf-bases. In this state it closely resembles L. Olivieri, Eichwald.[CY]
[CY] Lethæa Rossica, Plate Y, Figs. 12, 13.
In the ordinary young branches the leaf-scars are contiguous, and closely resemble those of L. elegans, Brongt. (Fig. 43 C). As the branches increase in diameter the leaf-scars slightly enlarge and sometimes assume a verticillate appearance (Fig. 43 D). As they still further enlarge they become separated by gradually increasing spaces of bark, marked with many waving striæ or wrinkles (Fig. 43 I, N). At the base of old stems the bark assumes a generally wrinkled appearance without distinct scars.
Knorria or Decorticated States.—Of these there is a great variety, depending on the state of preservation, and the particular longitudinal ridges. Fig. 43 D shows a form in which the vascular bundles appear as cylindrical truncate projections. Other forms show « 164 » the leaf-bases prominent, or have an appearance of longitudinal ribbing produced by the expansion of the bark.
Structure of Stem.—This is not perfectly preserved in any of my specimens, but one flattened specimen shows a central medulla with a narrow ring of scalariform vessels surrounding it, and constituting the woody axis. The structure is thus similar to that of L. Harcourtii, which I regard as probably the same with the closely allied European species L. Veltheimianum.
Leaves.—These are narrow, one-nerved, curving somewhat rapidly outward (Figs. 43, B, C, D). They vary from one to two inches in length.
Roots.—I have not seen these actually attached, but they occur very abundantly in the under-clays of some erect forests of these plants at Horton Bluff, and are of the character of Stigmariæ (Figs. 30, 31). In some of the under-clays the long, flattened rootlets are excessively abundant, and show the mark of a central vascular bundle.
Fructification.—Cones terminal, short, with many small, acute imbricate scales. Spore-cases globular, smooth (Fig. 43 C). On the surface of some shales and sandstones at Horton there are innumerable round spore-cases of this tree about the size of mustard-seed (Fig. 43 F). Large slabs are sometimes covered with these, and thin layers of shale are filled with flattened specimens.
This is the characteristic species of the Lower Carboniferous coal-measures, occurring in great profusion at Horton Bluff and its vicinity, also at Sneid’s Mills near Windsor, Noel and Five-Mile River, at Norton Creek and elsewhere in New Brunswick (Matthew’s collection), and at Antigonish (Honeyman’s collection).
I have received from the lowest Carboniferous beds of Ohio specimens of this species.[CZ] According to Rogers and Lesquereux similar forms occur in the Vespertine of Pennsylvania and in the Lower Carboniferous of Illinois. L. Veltheimianum of western Europe and L. glincanum of Russia are closely allied Lower Carboniferous species.[DA]
[CZ] “Journal of Geological Society,” November, 1862, p. 313.
[DA] For comparisons of these see “Report on Plants of Lower Carboniferous of Canada,” p. 21.
A very different type is furnished by a new species from the middle coal-formation of Clifton, New Brunswick.
Lepidodendron Cliftonense, Dawson.—Habit of Growth.—Robust, with thick branches, and leaves several inches in length. Terminal branches becoming slender, with shorter leaves.
Markings of Stem.—Leaf-bases long oval, pointed at ends, enlarging with growth of stem. Leaf-scars central, rhombic, transverse.
Leaves.—One-nerved, acutely pointed, from four inches in length on the larger branches to one inch or less on the branchlets.
Fructification.—Cones large, cylindrical or long oval, with large scales of trigonal form, and not elongated but lying close to the surface. Borne on lateral, slender branchlets, with short leaves.
Genus Lepidophloios, Sternberg; Ulodendron, L. and H.; Lomatophloios, Corda.
Lepidophloios.—Under this generic name, established by Sternberg, I include those lycopodiaceous trees of the coal-measures which have thick branches, transversely elongated leaf-scars, each with three vascular points and placed on elevated or scale-like protuberances, long one-nerved leaves, and large lateral strobiles in vertical rows or spirally disposed. Their structure resembles that of Lepidodendron, consisting of a Sternbergia pith, a slender axis of large scalariform vessels, giving off from its surface bundles of smaller vessels to the leaves, a very thick cellular bark, and a thin dense outer bark, having some elongated cells or bast-tissue on its inner side. In these trees the exogenous outer cylinder is less developed than in the Lepidodendra, and is sometimes wanting in stems or branches of some thickness.
Regarding L. laricinum of Sternberg as the type of the genus, and taking in connection with this the species described by Goldenberg, and my own observations on numerous specimens found in Nova Scotia, 1 have no doubt that Lomatophloios crassicaulis of Corda, and other species of that genus described by Goldenberg, Ulodendron and Bothrodendron of Lindley, Lepidodendron ornatissimum of Brongniart, and Halonia punctata of Geinitz, all belong to this genus, and differ from each other only in conditions of growth and preservation. Several of the species of Lepidostrobus and Lepidophyllum also belong to Lepidophloios.
The species of Lepidophloios are readily distinguished from Lepidodendron by the form of the areoles, and by the round scars on the stem, which usually mark the insertion of the large strobiles, though in barren stems they may also have produced branches; still, the fact of my finding the strobiles in situ in one instance, the accurate resemblance which the scars bear to those left by the cones of the red pine when borne on thick branches, and the actual impressions of the radiating scales in some specimens, leave no doubt in my « 166 » mind that they are usually the marks of cones; and the great size of the cones of Lepidophloios accords with this conclusion.
The species of Lepidophloios are numerous, and individuals are quite abundant in the coal formation, especially toward its upper part. Their flattened bark is frequent in the coal-beds and their roofs, affording a thin layer of pure coal, which sometimes shows the peculiar laminated or scaly character of the bark when other characters are almost entirely obliterated. The leaves also are nearly as abundant as those of Sigillaria in the coal-shales. They can readily be distinguished by their strong, angular midrib.
The markings of Lepidophloios may easily be mistaken for those of the Clathraria type of Sigillaria. When the stem only is seen, they can be distinguished by the length of the leaf-bases in Lepidophloios, and by the dominant central vascular scar; also by the one-nerved and ribbed leaves. Where the large, round marks of the cones are present, these are an infallible guide, never being present in Sigillaria. As the cones grew on the upper sides of the branches, the impression of the lower side often shows no cone-scars, or only two lateral rows, whereas on the upper side of the same branch they appear spirally arranged. I may describe as an example—
Lepidophloios Acadianus, Dawson. Leaf-bases broadly rhombic, or in old stems regularly rhombic, prominent, ascending, terminated by very broad rhombic scars having a central point and two lateral obscure points. Outer bark laminated or scaly. Surface of inner bark with single points or depressions. Leaves long, linear, with a strong keel on one side, five inches or more in length. Cone-scars sparsely scattered on thick branches, either in two rows or spirally, both modes being sometimes seen on the same branch. Scalariform axis scarcely an inch in diameter in a stem five inches thick. Fruit, an ovate strobile with numerous acute scales covering small globular spore-cases. This species is closely allied to Ulodendron majus and Lepidophloios laricinus, and presents numerous varieties of marking. Coal-formation, Nova Scotia.
Family Calamiteæ; Genus Calamites, Suckow.
The plants of this genus are unquestionably allied to the modern Equisetaceæ, but excel these so much in variety of form and structure, and are so capricious in their states of preservation, and so liable to be mistaken for parts of plants generically different, that they have given rise to much controversy. The following considerations will enable us to arrive at some certainty.
The genus Calamites was originally founded in the longitudinally « 167 » ribbed and jointed stems so frequent in the coal-formation, and of which the common C. Suckovii is a typical form. The most perfect of these stems represent the outer surface immediately within the epidermis, in which case transverse lines or constrictions mark the nodes, and at the nodes there are rounded spots, sometimes indicating radial processes of the pith, first described by Williamson; in other cases, the attachment of branchlets, or in some specimens both. But some specimens show the outer surface of the epidermis, in which case the transverse nodal lines are usually invisible, though the scars of branchlets may appear. In still other examples the whole of the outer tissues have perished, and the so-called Calamite is a cast of the interior of the stem, showing merely longitudinal ribbing and transverse nodal constrictions. In studying these plants in situ in the erect Calamite brakes of the coal-formation of Nova Scotia, one soon becomes familiar with these appearances, but they are evidently unknown to the majority of palæobotanists, though described in detail more than twenty years ago.
When the outer surface is preserved it is sometimes seen to bear verticils of long needle-like leaves (C. Cistii), or of branchlets with secondary whorls of similar leaves (C. Suckovii and C. undulatus). No Calamite known to me bears broad one-nerved leaves like those of Asterophyllites and Annularia, though the larger stems of these plants have been described as Calamites, and the term Calamocladus has been used to include both groups. The base of the Calamite stem usually terminates in a blunt point, and may be attached to a rhizome, or several stems may bud out from each other in a group or stool. The roots are long and cylindrical, sometimes branching. The fruit consists of spikes of spore-cases, borne in whorls and subtended by linear floral leaves. To these strobiles the name Calamostachys has been given.
Williamson has shown that the stem of Calamites consists of a central pith or cavity of large size surrounded by a cylinder consisting of alternate wedges of woody and cellular matter, with vertical canals at the inner sides of the wedges, and slender medullary rays. The thick cellular wedges intervening between the woody wedges he calls primary medullary rays; the smaller medullary rays in the wedges, secondary medullary rays. There is thus a highly complex exogenous stem based on the same principle with the stem of a common Equisetum, but with much greater strength and complexity.
Williamson has also shown that there are different sub-types of these stems. More especially he refers to the three following:
(a) Calamites proper, which has the woody wedges of scalariform or barred tissue with thin medullary rays, and the thick primary medullary rays are cellular.
(b) Calamopitus has reticulated or multiporous tissue in the woody wedges with medullary rays, and the primary medullary wedges are composed of elongated cells.
(c) Calamodendron has the woody wedges of barred tissue as in a, with medullary rays, but has the intervening medullary wedges of an elongated tissue approaching to woody fibre, and also with medullary rays.
To these I would add a fourth type, which I have described, from the coal-formation of Nova Scotia.[DB]
[DB] “Quarterly Journal of the Geological Society,” 1871.
(d) Eucalamodendron differs from Calamodendron in having true bordered pores or pseudo-scalariform slit-pored tissue, and corresponds to the highest type of calamitean stem.
I would also add that under a and b there are some species in which the woody cylinder is very thin in comparison to the size of the stem. In c and d the woody cylinder is thick and massive, and the stems are often large and nodose.
As an example of an ordinary Calamite in which the external surface and foliage are preserved, I may quote the following from my report on the “Flora of the Lower Carboniferous and Millstone Grit,” 1873:
Calamites Undulatus, Brongniart.—This species is stated by Brongniart to be distinguished from the C. Suckovii, the characteristic Calamite of the middle coal-formation, by its undulated ribs marked with peculiar cellular reticulation. He suggests that it may be merely a variety of C. Suckovii, an opinion in which Schimper coincides; but since I have received large additional collections from Mr. Elder, containing not only the stems and branches, but also the leaves and rhizomes, I am constrained to regard it as a distinct though closely allied species.
The rhizomata are slender, being from one to two inches in diameter, and perfectly flattened. They are beautifully covered with a cellular reticulation on the thin bark, and show occasional round areoles marking the points of exit of the rootlets. I have long been familiar with irregular flattened stems thus reticulate, but have only recently been able to connect them with this species of Calamite.
The main stems present a very thin carbonaceous bark reticulated like the rhizomes. They have flat, broad ribs separated by deep « 169 » and narrow furrows, and undulated in a remarkable manner even when the stems are flattened. This undulation is, however, perhaps an indication of vertical pressure while the plant was living, as it seems to have had an unusually thin and feeble cortical layer, and the undulations are apparently best developed in the lower part of the stem. At the nodes the ribs are often narrowed and gathered together, especially in the vicinity of the rounded radiating marks which appear to indicate the points of insertion of the branches. At the top of each rib we have the usual rounded areole, probably marking the insertion of a primary branchlet.
The branches have slender ribs and distant nodes, from which spring secondary branchlet s in whorls, these bearing in turn small whorls of acicular leaflets much curved upward, and which are apparently round in cross section and delicately striate. They are much shorter than the leaves of Calamites Suckovii, and are less dense and less curved than those of C. nodosus, which I believe to be the two most closely allied species.
Lesquereux notices this species as characteristic of the lower part of the Carboniferous in Arkansas.
It will be observed that I regard the striated and ribbed stems not as internal axes, but as representing the outer surface of the plants. This was certainly the case with the present species and with C. Suckovii and C. nodosus. Other species, and especially those which belonged to Calamodendron, no doubt had a smooth or irregularly wrinkled external bark; but this gives no good ground for the manner in which some writers on this subject confound Calamites with Calamodendra, and both with Asterophyllites and Sphenophyllum. With this no one who has studied these plants, rooted in their native soils, and with their appendages still attached, can for a moment sympathise. One of the earliest geological studies of the writer was a bed of these erect Calamites, which he showed to Sir C. Lyell in 1844, and described in the “Proceedings of the Geological Society” in 1851, illustrating the habit of growth as actually seen well exposed in a sandstone cliff. Abundant opportunities of verifying the conclusions formed at that time have since occurred, the results of which have been summed up in the figures in Acadian Geology, which, though they have been treated by some botanists as merely restorations, are in reality representations of facts actually observed.
On these subjects, without entering into details, and referring for these to the elaborate discussions of Schimper, Williamson, and McNab, and to my paper on the subject, “Journal of the Geological Society,” vol. xxvii, p. 54, I may remark:
1. That the aërial stems of ordinary Calamites had a thin cortical layer, with lacunæ and fibrous bundles and multiporous vessels—the whole not differing much from the structure of modern Equiseta.
2. Certain arborescent forms, perhaps allied to the true Calamites, as well as possibly the old underground stems of ordinary species[DC] assumed a thick-walled character in which the tissues resembled the wedges of an exogen, and abundance of pseudo-scalariform fibres were developed, while the ribbing of the external surface became obsolete or was replaced by a mere irregular wrinkling.
[DC] Williamson, “Transactions of the Royal Society.” McNab, in “Proceedings of the Edinburgh Botanical Society.”
3. Sufficient discrimination has not been exercised in separating casts of the internal cavities of Calamites and Calamodendron from those representing other surfaces and the proper external surface.
4. There is no excuse for attributing to Calamites the foliage of Annularia, Asterophyllites, and Sphenophyllum, since these leaves have not been found attached to true Calamite stems, and since the structure of the stems of Asterophyllites as described by Williamson, and that of Sphenophyllum as described by the writer,[DD] are essentially different from those of Calamites.
[DD] “Journal of the Geological Society,” 1866.
5. As the species above described indicates, good external characters can be found for establishing species of this genus, and these species are of value as marks of geological age.
Genus Archæocalamites, Sternberg.
This genus has been established to include certain Calamites of the Devonian and Lower Carboniferous, in which the furrows on the stem do not alternate at the nodes or joints, and the leaves in one species at least bifurcate. C. radiatus, Brongniart, is the typical species. In North America it occurs in the Erian, probably as low as the Middle Erian. In Europe it has so far been recognised in the Lower Carboniferous only. I have, however, seen stems from alleged Devonian beds in Devonshire which may have belonged to this species.
Family Asterophylliteæ; Genus Asterophyllites, Brongniart.
Stems ribbed and jointed like the Calamites, but with inflated nodes and a stout internal woody cylinder, which has been described by Williamson. From the joints proceeded whorls of leaves or of branchlets, bearing leaves which differed from those of Calamites in their having a distinct middle rib or vein. The fructification consisted « 171 » of long slender cones or spikes, having whorls of scales bearing the spore-cases. Some authors speak of Asterophyllites as only branches and leaves of Calamites; but though at first sight the resemblance is great, a close inspection shows that the leaves of Asterophyllites have a true midrib, which is wanting in Calamites.
Genus Annularia.—It is perhaps questionable whether these plants should be separated from Asterophyllites, The distinction is that they produce branches in pairs, and that their whorls of leaves are one-sided and usually broader than those of Asterophyllites, and united into a ring at their insertion on the stem. One little species, A. sphenophylloides, is very widely distributed.
Pinnularia—a provisional genus—-includes slender roots or stems branching in a pinnate manner, and somewhat irregularly. They are very abundant in the coal shales, and were probably not independent plants, but aquatic roots belonging to some of the plants last mentioned. The probability of this is farther increased by their resemblance in miniature to the roots of Calamites. They are always flattened, but seem originally to have been round, with a slender thread-like axis of scalariform vessels, enclosed in a soft, smooth, cellular bark.
Family Rhizocarpeæ; Genus Sphenophyllum.
Leaves in whorls, wedge-shaped, with forking veins. Fructification on spikes, with verticils of sporocarps. These plants are by some regarded as allied to the Calamiteæ and Asterophylliteæ, by others as a high grade of Rhizocarps of the type of Marsilia. The stem had a star-shaped central bundle of scalariform or reticulato-scalariform vessels.
Genus Sporangites. (Sporocarpon, Williamson.)
Under this name we may provisionally include those rounded spherical bodies found in the coal and its accompanying beds, and also in the Erian, which may be regarded as Macrospores or Sporocarps of Protosalvinia, or other Rhizocarpean plants akin to those described above in Chapter III, which see for description.
Genus Protosalvinia.—Under this we include sporocarps allied to those of Salvinia, as described in Chapter III.
Family Filices.
Under this head I shall merely refer to a few groups of special interest, and to the provisional arrangement adopted for the fronds of ferns when destitute of fructification.
The external appearances of trunks of tree-ferns have been already referred to.
With respect to tree ferns, the oldest known examples are those from the Middle Devonian of New York and Ohio, which I have described in the “Journal of the Geological Society,” 1871 and 1881. As these are of some interest, I have reproduced their descriptions in a note appended to Chapter III, which see.
The other forms most frequently occurring in the Carboniferous are Caulopteris, Palæopteris, and Megaphyton[DE] Stems showing merely masses of aërial roots are known by the name Psaronius.
[DE] See my “Acadian Geology,” also below.
With reference to the classification of Palæozoic ferns, this has hitherto been quite arbitrary, being based on mere form and venation of fronds, but much advance has recently been made in the knowledge of their fructification, warranting a more definite attempt at classification. The following are provisional genera usually adopted:
1. Cyclopteris, Brongniart.—Leaflets more or less rounded or wedge-shaped, without midrib, the nerves spreading from the point of attachment. This group includes a great variety of fronds evidently of different genera, were their fructification known; and some of them probably portions of fronds, the other parts of which may be in the next genus.
2. Neuropteris, Brongniart.—Fronds pinnate, and with the leaflets narrowed at the base; midrib often not distinct, and disappearing toward the apex. Nervures equal, and rising at an acute angle. Ferns of this type are among the most abundant in the coal-formation.
3. Odontopteris, Brongniart.—In these the frond is pinnate, and the leaflets are attached by their whole base, with the nerves either proceeding wholly from the base, or in part from an indistinct midrib, which soon divides into nervures.
4. Dictyopteris, Gutbier.—This is a beautiful style of fern, with leaflets resembling those of Neuropteris, but the veins arranged in a network of oval spaces. Only a few species are known in the coal-formation.
5. Lonchopteris, Brongniart.—Ferns with netted veins like the above, but with a distinct midrib, and the leaflets attached by the whole base. Of this, also, we can boast but few species.
6. Sphenopteris, Brongniart.—These are elegant ferns, very numerous in species, and most difficult to discriminate. Their most « 173 » distinctive characters are leaflets narrowed at the base, often lobed, and with nervures dividing in a pinnate manner from the base.
7. Phyllopteris, Brongniart.—These are pinnate, with long lanceolate pinnules, having a strong and well-defined midrib, and nerves proceeding from it very obliquely, and dividing as they proceed toward the margin. The ferns of this genus are for the most part found in formations more recent than the Carboniferous; but I have referred to it, with some doubt, one of our species.
8. Alethopteris, Brongniart.—This genus includes many of the most common coal-formation ferns, especially the ubiquitous A. lonchitica, which seems to have been the common brake of the coal-formation, corresponding to Pteris aquilina in modern Europe and America. These are brake-like ferns, pinnate, with leaflets often long and narrow, decurrent on the petiole, adherent by their whole base, and united at base to each other. The midrib is continuous to the point, and the nervures run off from it nearly at right angles. In some of these ferns the fructification is known to have been marginal, as in Pteris.
9. Pecopteris, Brongniart.—This genus is intermediate between the last and Neuropteris. The leaflets are attached by the whole base, but not usually attached to each other; the midrib, though slender, attains to the summit; the nervures are given off less obliquely than in Neuropteris. This genus includes a large number of our most common fossil ferns.
10. Beinertia, Goeppert.—A genus established by Goeppert for a curious Pecopteris-like fern, with flexuous branching oblique nervures becoming parallel to the edge of the frond.
11. Hymenophyllites, Goeppert.—These are ferns similar to Sphenopteris, but divided at the margin into one-nerved lobes, in the manner of the modern genus Hymenophyllum.
12. Palæopteris, Geinitz.—This is a genus formed to include certain trunks of tree-ferns with oval transverse scars of leaves.
13. Caulopteris, Lindley and Hutton.—Is another genus of fossil trunks of tree-ferns, but with elongate scars of leaves.
14. Psaronius, Cotta.—Includes other trunks of tree-ferns with alternate scars or thick scales, and ordinarily with many aërial roots grouped round them, as in some modern tree-ferns.
15. Megaphyton, Artis.—Includes trunks of tree-ferns which bore their fronds, which were of great size, in two rows, one on each side of the stem. These were very peculiar trees, less like modern ferns than any of the others. My reasons for regarding them as ferns are stated in the following extract from a recent paper:
“Their thick stems, marked with linear scars and having two rows of large depressed areoles on the sides, suggest no affinities to any known plants. They are usually ranked with Lepidodendron and Ulodendron, but sometimes, and probably with greater reason, are regarded as allied to tree-ferns. At the Joggins a very fine species (M. magnificum) has been found, and at Sydney a smaller species (M. humile); but both are rare and not well preserved. If the large scars bore cones and the smaller bore leaves, then, as Brongniart remarks, the plant would much resemble Lepidophloios, in which the cone-scars are thus sometimes distichous. But the scars are not round and marked with radiating scales as in Lepidophloios; they are reniform or oval, and resemble those of tree-ferns, for which reason they may be regarded as more probably leaf-scars; and in that case the smaller linear scars would indicate ramenta, or small aërial roots. Further, the plant described by Corda as Zippea disticha is evidently a Megaphyton, and the structure of that species is plainly that of a tree-fern of somewhat peculiar type. On these grounds I incline to the opinion of Geinitz that these curious trees were allied to ferns, and bore two rows of large fronds, the trunks being covered with coarse hairs or small aërial roots. At one time I was disposed to suspect that they may have crept along the ground; but a specimen from Sydney shows the leaf-stalks proceeding from the stem at an angle so acute that the stem must, I think, have been erect. From the appearance of the scars it is probable that only a pair of fronds were borne at one time at the top of the stem; and, if these were broad and spreading, it would be a very graceful plant. To what extent plants of this type contributed to the accumulation of coal I have no means of ascertaining, their tissues in the state of coal not being distinguishable from those of ferns and Lycopodiaceæ.”
16. For descriptions of the genus Archæopteris and other Erian ferns, see Chapter III.
THE FLORA OF THE EARLY MESOZOIC.
Great physical changes occurred at the close of the Carboniferous age. The thick beds of sediment that had been accumulating in long lines along the primitive continents had weighed down the earth’s crust. Slow subsidence had been proceeding from this cause in the coal-formation period, and at its close vast wrinklings occurred, only surpassed by those of the old Laurentian time. Hence in the Appalachian region of America we have the Carboniferous beds thrown into abrupt folds, their shales converted into hard slates, their sandstones into quartzite and their coals into anthracite, and all this before the deposition of the Triassic Red Sandstones which constitute the earliest deposit of the great succeeding Mesozoic period. In like manner the coal-fields of Wales and elsewhere in western Europe have suffered similar treatment, and apparently at the same time.
This folding is, however, on both sides of the Atlantic limited to a band on the margin of the continents, and to certain interior lines of pressure, while in the middle, as in Ohio and Illinois in America, and in the great interior plains of Europe, the coal-beds are undisturbed and unaltered. In connection with this we have an entire change in the physical character of the deposits, a great elevation of the borders of the continents, and probably a considerable deepening of the seas, leading to the establishment of general geographical conditions which still remain, though they have been temporarily modified by subsequent subsidences and re-elevations.
Along with this a great change was in progress in vegetable and animal life. The flora and fauna of the Palæozoic gradually die out in the Permian and are replaced in the succeeding Trias by those of the Mesozoic time. Throughout the Permian, however, the remains of the coal-formation flora continue to exist, and some forms, as the Calamites, even seem to gain in importance, as do also certain types of coniferous trees. The Triassic, as well as the Permian, was marked by physical disturbances, more especially by great volcanic eruptions discharging vast beds and dykes of lava and layers of volcanic ash and agglomerate. This was the case more especially along the margins of the Atlantic, and probably also on those of the Pacific. The volcanic sheets and dykes associated with the Red Sandstones of Nova Scotia, Connecticut, and New Jersey are evidences of this.
At the close of the Permian and beginning of the Trias, in the midst of this transition time of physical disturbance, appear the great reptilian forms characteristic of the age of reptiles, and the earliest precursors of the mammals, and at this time the old Carboniferous forms of plants finally pass away, to be replaced by a flora scarcely more advanced, though different, and consisting of pines, cycads, and ferns, with gigantic equiseta, which are the successors of the genus Calamites, a genus which still survives in the early Trias. Of these groups the conifers, the ferns, and the equiseta are already familiar to us, and, in so far as they are concerned, a botanist who had studied the flora of the Carboniferous would have found himself at home in the succeeding period. The cycads are a new introduction. The whole, however, come within the limits of the cryptogams and the gymnosperms, so that here we have no advance.[DF]
[DF] Fontaine’s “Early Mesozoic Flora of Virginia” gives a very good summary of this flora in America.
As we ascend, however, in the Mesozoic, we find new and higher types. Even within the Jurassic epoch, the next in succession to the Trias, there are clear indications of the presence of the endogens, in species allied to the screw-pines and grasses; and the palms appear a little later, while a few exogenous trees have left their remains in the Lower Cretaceous, and in the Middle and Upper Cretaceous these higher plants come in abundantly and in generic forms still extant, so that the dawn of the modern flora belongs to the Middle and Upper « 178 » Cretaceous. It will thus be convenient to confine ourselves in this chapter to the flora of the earlier Mesozoic.
Passing over for the present the cryptogamous plants already familiar in older deposits, we may notice the new features of gymnospermous and phænogamous life, as they present themselves in this earlier part of the great reptilian age, and as they extended themselves with remarkable uniformity in this period over all parts of the world. For it is a remarkable fact that, if we place together in our collections fossil plants of this period from Australia, India, China, Siberia, Europe, or even from Greenland, we find wonderfully little difference in their aspect. This uniformity we have already seen prevailed in the Palæozoic flora; and it is perhaps equally marked in that of the Mesozoic. Still we must bear in mind that some of the plants of these periods, as the ferns and pines, for example, are still world-wide in their distribution; but this does not apply to others, more especially the cycads (Fig. 65).
The cycads constitute a singular and exceptional type in the modern world, and are limited at present to the warmer climates, though very generally distributed in these, as they occur in Africa, India, Japan, Australia, Mexico, Florida, and the West Indies. In the Mesozoic age, however, they were world-wide in their distribution, and are found as far north as Greenland, though most of the species found in the Cretaceous « 179 » of that country are of small size/ and may have been of low growth, so that they may have been protected by the snows of winter. The cycads have usually simple or unbranching stems, pinnate leaves borne in a crown at top, and fruits which, though somewhat various in structure and arrangement, are all of the simpler form of gymnospermous type. The stems are exogenous in structure, but with slender wood and thick bark, and barred tissue, or properly as tissue intermediate between this and the disc-bearing fibres of the pines.
Though the cycads have a considerable range of organisation and of fructification, and though some points in reference to the latter might assign them a higher place, on the whole they seem to occupy a lower position than the conifers or the cordaiteæ of the Carboniferous. In the Carboniferous some of the fern-like leaves assigned to the genus Noeggerathia have been shown by Stur and Weiss to have been gymnosperms, probably allied to cycads, of which they may be regarded at least as precursors. Thus the cycadean type does not really constitute an advance in grade of organisation in the Mesozoic, any further than that, in the period now in question, it becomes much more developed in number and variety of forms. But the conifers would seem to have had precedence of it for a long time in the Palæozoic, and it replaces in the Mesozoic the Cordaites, which in many respects excelled it in complexity.
The greater part of the cycads of the Mesozoic age would seem to have had short stems and to have constituted the undergrowth of woods in which conifers attained to greater height. An interesting case of this is the celebrated dirt-bed of the quarries of the Isle of Portland, long ago described by Dean Buckland. In this fossil soil trunks of pines, which must have attained to great height, are interspersed with the short, thick stems of cycads, of the genus named Cycadoidea by Buckland, « 180 » and which from their appearance are called “fossil birds' nests” by the quarrymen. Some, however, must have attained a considerable height so as to resemble palms.
The cycads, with their simple, thick trunks, usually marked with rhombic scars, and bearing broad spreading crowns of large, elegantly formed pinnate leaves, must have formed a prominent part of the vegetation of the northern hemisphere during the whole of the Mesozoic period. A botanist, had there been such a person at the time, would have found this to be the case everywhere from the equator to Spitzbergen, and probably in the southern hemisphere as well, and this throughout all the long periods from the Early Trias to the Middle Cretaceous. In a paper published in the “Linnæan Transactions” for 1868, Dr. Carruthers enumerates twenty species of British Mesozoic cycads, and the number might now be considerably increased.
The pines present some features of interest. We have already seen their connection with the broad-leaved Cordaites, and in the Permian there are some additional types of broad-leaved coniferæ. In the Mesozoic we have great numbers of beautiful trees, with those elegant fan-shaped leaves characteristic of but one living species, the Salisburia, or gingko-tree of China. It is curious that this tree, though now limited to eastern Asia, will grow, though it rarely fruits, in most parts of temperate Europe, and in America as far north as Montreal, and that in the Mesozoic period it occupied all these regions, and even Siberia and Greenland, and with many and diversified species (Fig. 66).
Salisburia belongs to the yews, but an equally curious fact applies to the cypresses. The genus Sequoia, limited at present to two species, both Californian, and one of them the so-called “big tree,” celebrated for the gigantic size to which it attains, is represented by species found as far back at least as the Lower Cretaceous, and in every part of the northern hemisphere.[DG] It seems to have thriven in all these regions throughout the Mesozoic and early Kainozoic, and then to have disappeared, leaving only a small remnant to represent it in modern days. A number of species have been described from the Mesozoic and Tertiary, all of them closely related to those now existing (Fig. 67).
[DG] In the Eocene of Australia.
The following notice of these trees is for the most part translated, with some modifications and abridgment, from a paper read by the late Prof. Heer before the Botanical Section of the Swiss Natural History Society:
The name itself deserves consideration. It is that of an Indian of the Cherokee tribe, Sequo Yah, who invented an alphabet without any aid from the outside world of culture, and taught it to his tribe by writing it upon « 182 » leaves. This came into general use among the Cherokees, before the white man had any knowledge of it; and afterward, in 1828, a periodical was published in this character by the missionaries. Sequo Yah was banished from his home in Alabama, with the rest of his tribe, and settled in New Mexico, where he died in 1843.
When Endlicher was preparing his synopsis of the conifers, in 1846, and had established a number of new genera, Dr. Jacbon Tschudi, then living with Endlicher, brought before his notice this remarkable man, and asked him to dedicate this red-wooded tree to the memory of a literary genius so conspicuous among the red men of America. Endlicher consented to do so, and only endeavored to make the name pronounceable by changing two of its letters.
Endlicher founded the genus on the redwood of the Americans, Taxodium sempervirens of Lamb; and named the species Sequoia sempervirens. These trees form large forests in California, which extend along the coast as far as Oregon. Trees are there met with of 300 feet in height and 20 feet in diameter. The seeds have been brought to Europe a number of years ago, and we already see in upper Italy and around the Lake of Geneva, and in England, high trees; but, on the other hand, they have not proved successful around Zurich.
In 1852, a second species of Sequoia was discovered in California, which, under the name of big tree, soon attained a considerable celebrity. Lindley described it, in 1853, as Wellingtonia gigantea; and, in the following year, Decaisne and Torrey proved that it belonged to Sequoia, and that it accordingly should be called Sequoia gigantea.
While the Sequoia sempervirens, in spite of the destructiveness of the American lumbermen, still forms large forests along the coast, the Sequoia gigantea is confined to the isolated clumps which are met with inland at « 183 » a height of 5,000 to 7,000 feet above sea-level, and are much sought after by tourists as one of the wonders of the country. Reports came to Europe concerning the largest of them which were quite fabulous, but we have received accurate accounts of them from Prof. Whitney. The tallest tree measured by him has a height of 325 feet, and in the case of one of the trees the number of the rings of growth indicated an age of about 1,300 years. It had a girth of 50 to 60 feet.
We know only two living species of Sequoia, both of which are confined to California. The one (S. sempervirens) is clothed with erect leaves, arranged in two rows, very much like our yew-tree, and bears small, round cones; the other (S. gigantea) has smaller leaves, set closely against the branches, giving the tree more the appearance of the cypress. The cones are egg-shaped, and much larger. These two types are therefore sharply defined.
Both of these trees have an interesting history. If we go back into the Tertiary, this same genus meets us with a long array of species. Two of these species correspond to those living at present: the S. Langsdorfii to the S. sempervirens, and the S. Couttsiæ to the S. gigantea.[DH] But, while the living species are confined to California, in the Tertiary they are spread over several quarters of the globe.
[DH] S. Couttsiæ has leaves like S. gigantea, and cones like those of 8. sempervirens.
Let us first consider the Sequoia Langsdorfii. This was first discovered in the lignite of Wetterau, and was described as Taxites langsdorfii. Heer found it in the upper Rhone district, and there lay beside the twigs the remains of a cone, which showed that the Taxites Langsdorfii of Brongniart belonged to the Californian genus Sequoia established by Endlicher. He afterward « 184 » found much better preserved cones, together with seeds, along with the plants of east Greenland, which fully confirmed the determination. At Atanekerdluk in Greenland (about 70° north latitude) this tree is very common. The leaves, and also the flowers and numerous cones, leave no doubt that it stands very near to the modern redwood. It differs from it, however, in having a much larger number of scales in the cone. The tree is also found in Spitzbergen at nearly 78° north latitude, where Nordenskiöld has collected, at Cape Lyell, wonderfully preserved branches. From this high latitude the species can be followed down through the whole of Europe as far as the middle of Italy (at Senegaglia, Gulf of Spezia). In Asia, also,, we can follow it to the steppes of Kirghisen, to Possiet, and to the coast of the Sea of Japan, and across to Alaska and Sitka. It is recognized by Mr. Starkie Gardner as one of the species found in the Eocene of Mull in the Hebrides.[DI] It is thus known in Europe, Asia, and America, from 43° to 78° north latitude, while its most nearly related living species, perhaps even descended from it, is now confined to California.
[DI] It is Fareites Campbelli of Forbes.
With this S. Langsdorfii, three other Tertiary species are nearly related (S. brevifolia, Hr., S. disticha, Hr., and S. Nordenskiöldi, Hr.). These have been met with in Greenland and Spitzbergen, and one of them has lately been found in the United States. Three other species, in addition to these, have been described by Lesquereux, which appear to belong to the group of the S. Langsdorfii, viz., S. longifolia, Lesq., S. angustifolia, and S. acuminata, Lesq. Several species also occur in the Cretaceous and Eocene of Canada.
These species thus answer to the living Sequoia sempervirens; but we can also point to Tertiary representatives « 185 » of the S. gigantea. Their leases are stiff and sharp-pointed, are thinly set round the branches, and lie forward in the same way: the egg-shaped cones are in some cases similar.
There are, however, in the early Tertiary six species, which fill up the gap between S. sempervirens and S. gigantea. They are the S. Couttsiæ, S. affinis, Lesq., S. imbricata, Hr., S. sibirica, Hr., S. Heerii, Lesq., and S. biformis, Lesq. Of these, S. Couttsiæ, Hr., is the most common and most important species. It has short leaves, lying along the branch, like S. gigantea, and small, round cones, like S. Langsdorfii and sempervirens. Bovey Tracey in Devonshire has afforded splendid specimens of cones, seeds, and twigs, which have been described in the “Philosophical Transactions.” More lately, Count Saporta has described specimens of cones and twigs from Armissan. Specimens of this species have also been found in the older Tertiary of Greenland, so that it must have had a wide range. It is very like to the American S. affinis, Lesq.
In the Tertiary there have been already found fourteen well-marked species, which thus include representatives of the two living types, S. sempervirens and S. gigantea.
We can follow this genus still further back. If we go back to the Cretaceous age, we find ten species, of which five occur in the Urgon of the Lower Cretaceous, two in the Middle, and three in the Upper Cretaceous. Among these, the Lower Cretaceous exhibits the two types of the Sequoia sempervirens and S. gigantea. To the former the S. Smithiana answers, and to the latter, the Reichenbachii, Gein. The S. Smithiana stands indeed uncommonly near the S. Langsdorfii, both in the appearance of the leaves on the twigs and in the shape of the cones. These are, however, smaller, and the leaves do not become narrower toward the base. The S. pectina, Hr., of the Upper Cretaceous, has its leaves arranged in two rows, and « 186 » presents a similar appearance. The S. Reichenbachii is a type more distinct from those now living and those in the Tertiary. It has indeed stiff, pointed leaves, lying forward, but they are arcuate, and the cones are smaller. This tree has been known for a long time, and it serves in the Cretaceous as a guiding star, which we can follow from the Urgonian of the Lower Cretaceous up to the Cenomanian. It is known in France, Belgium, Bohemia, Saxony, Greenland, and Spitzbergen (also in Canada and the United States). It has been placed in another genus—Geinitzia—but we can recognise, by the help of the cones, that it belongs to Sequoia.
Below this, there is found in Greenland a nearly related species, the S. ambigua, Hr., of which the leaves are shorter and broader, and the cones round and somewhat smaller.
The connecting link between S. Smithiana and Reichenbachii is formed by S. subulata, Hr., and S. rigida, Hr., and three species (S. gracilis, Hr., S. fastigiata and S. Gardneriana, Carr.), with leaves lying closely along the branch, and which come very near to the Tertiary species S. Couttsiæ. We have therefore in the Cretaceous quite an array of species, which fill up the gap between the S. sempervirens and gigantea, and show us that the genus Sequoia had already attained a great development in the Cretaceous. This was still greater in the Tertiary, in which it also reached its maximum of geographical distribution. Into the present world the two extremes of the genus have alone continued; the numerous species forming its main body have fallen out in the Tertiary.
If we look still further back, we find in the Jura a great number of conifers, and, among them, we meet in the genus Pinus with a type which is highly developed, and which still survives; but for Sequoia we have till now looked in vain, so that for the present we can not place the rise of the genus lower than the Urgonian of the Cretaceous, « 187 » however remarkable we may think it that in that period it should have developed into so many species; and it is still more surprising that two species already make their appearance which approach so near to the living Sequoia sempervirens and S. gigantea.
Altogether, we have become acquainted, up to the present time, with twenty-six species of Sequoia. Fourteen of these species are found in the Arctic zone, and have been described and figured in the “Fossil Flora of the Arctic Regions.” Sequoia has been recognised by Ettingshausen even in Australia, but there in the Eocene.
This is, perhaps, the most remarkable record in the whole history of vegetation. The Sequoias are the giants of the conifers, the grandest representatives of the family, and the fact that, after spreading over the whole northern hemisphere and attaining to more than twenty specific forms, their decaying remnant should now be confined to one limited region in western America and to two species constitutes a sad memento of departed greatness.[DJ] The small remnant of S. gigantea still, however, towers above all competitors, as eminently the “big trees ”; but, had they and the allied species failed to escape the Tertiary continental submergences and the disasters of the glacial period, this grand genus would have been to us an extinct type. In like manner the survival of the single gingko of eastern Asia alone enables us to understand that great series of taxine trees with fern-like leaves of which it is the sole representative.
[DJ] The writer has shown that much of the material of the great lignite beds of the Canadian Northwest consists of wood of Sequoia of both the modern types.
Besides these peculiar and now rare forms, we have in the Mesozoic many others related closely to existing yews, cypresses, pines, and spruces, so that the conifers were probably in greater abundance and variety than they are at this day.
In this period, also, we find the earliest representatives of the endogenous plants. It is true that some plants found in the coal-formation have been doubtfully referred to these, but the earliest certain examples would seem to be some bamboo-like and screw-pine-like plants occurring in the Jurassic rocks. Some of these are, it is true, doubtful forms, but of others there seems to be no question. The modern Pandanus or screw-pine of the tropical regions, which is not a pine, however, but a humble relation of the palms, is a stiffly branching tree, of a candelabra-like form, and with tufts of long leaves on its branches, and nuts or great hard berries for fruit, borne sometimes in large masses, and so protected as to admit of their drifting uninjured on the sea. The stems are supported by masses of aërial roots like those which strengthen the stems of tree-ferns. These structures and habits of growth fit the Pandanus for its especial habitat on the shores of tropical islands, to which its masses of nuts are drifted by the winds and currents, and on whose shores it can establish itself by the aid of its aërial roots.
Some plants referred to the cycads have proved veritable botanical puzzles. One of these, the Williamsonia gigas of the English oölite, originally discovered by my friend Dr. Williamson, and named by him Zamia gigas, a very tall and beautiful species, found in rocks of this age in various parts of Europe, has been claimed by Saporta for the Endogens, as a plant allied to Pandanus. Some other botanists have supposed the flowers and fruits to be parasites on other plants, like the modern Rafflesia of Sumatra, but it is possible that after all it may prove to have been an aberrant cycad.
The tree-palms are not found earlier than the Middle Cretaceous, where we shall notice them in the next chapter. In like manner, though a few Angiosperms occur in rocks believed to be Lower or Lower Middle Cretaceous in Greenland and the northwest territory of Canada, and « 189 » in Virginia, these are merely precursors of those of the Upper Cretaceous, and are not sufficient to redeem the earlier Cretaceous from being a period of pines and cycads.
On the whole, this early Mesozoic flora, so far as known to us, has a monotonous and mean appearance. It no doubt formed vast forests of tall pines, perhaps resembling the giant Sequoias of California; but they must for the most part have been dark and dismal woods, probably tenanted by few forms of life, for the great reptiles of this age must have preferred the open and sunny coasts, and many of them dwelt in the waters. Still we must not be too sure of this. The berries and nuts of the numerous yews and cycads were capable of affording much food. We know that in this age there were many great herbivorous reptiles, like Iguanodon and Hadrosaurus, some of them fitted by their structure to feed upon the leaves and fruits of trees. There were also several kinds of small herbivorous mammals, and much insect life, and it is likely that few of the inhabitants of the Mesozoic woods have been preserved as fossils. We may yet have much to learn of the inhabitants of these forests of ferns, cycads, and pines. We must not forget in this connection that in the present day there are large islands, like New Zealand, destitute of mammalia, and having a flora comparable with that of the Mesozoic in the northern hemisphere, though more varied. We have also the remarkable example of Australia, with a much richer flora than that of the early Mesozoic, yet inhabited only by non-placental mammals, like those of the Mesozoic.
The principal legacy that the Mesozoic woods have handed down to our time is in some beds of coal, locally important, but of far less extent than those of the Carboniferous period. Still, in America, the Richmond coal-field in Virginia is of this age, and so are the anthracite beds of the Queen Charlotte Islands, on the west coast of Canada, and the coal of Brora in Sutherlandshire. Valuable « 190 » beds of coal, probably of this age, also exist in China, India, and South Africa; and jet, which is so extensively used for ornament, is principally derived from the carbonised remains of the old Mesozoic pines.
In the next chapter we have to study a revolution in vegetable life most striking and unique, in the advent of the forest-trees of strictly modern types.
NOTE TO CHAPTER V.
I append to this chapter a table showing the plant-bearing series of the Cretaceous and Laramie of North America, from a paper in “Trans. R. S. C,” 1885, which see for further details:
(In Descending Order.)
| Periods. | Floras and sub-floras. | References. | |
| Transition Eocene to Cretaceous. |
Upper Laramie or Porcupine Hill. Fort Union group, U. S. territory. |
Platanus beds of Souris River and Calgary. Report of Geol. Survey of Canada for 1879, and Memoir of 1885. |
|
| Upper Cretaceous (Danian and Senonian). |
Middle Laramie or Willow Creek beds. | ||
| Lower Laramie or St. Mary River. | Lemna and Pistia beds of bad lands of 49th parallel, Red Deer River, &c., with lignites. Report 49th Parallel and Memoir of 1885. |
||
| Fox Hill series | Marine. | ||
| Fort Pierre series | Marine. | ||
| Belly River | Sequoia and Brasenia beds of S. Saskatchewan, Belly River, &c. with lignites. Memoir of 1885. |
||
| Coal measures of Nanaimo, B.C., probably here. |
Memoir of 1883. Many dicotyledons, palms, &c. |
||
| Middle Cretaceous (Turonian and Cenomanian). |
Dunvegan series of Peace River. Dakota group, U. S. Amboy clays, U. S. |
Memoir of 1883. Many dicotyledons, cycads, &c. |
|
| Mill Creek beds of Rocky Mountains. | Dicotyledonous leaves, similar to Dakota group of the U. S. Memoir of 1885. |
||
| Lower Cretaceous (Neocomian.&c). |
Suskwa River beds and Queen Charlotte Island coal series. Intermediate beds of Rocky Mountains. Potomac series of Virginia. |
Cycads, pines, a few dicotyledons. Report Geol.Survey. Memoir of 1885. |
|
| Kootanie series of Rocky Mountains. |
Cycads, pines, and ferns. Memoir of 1885. |
||
THE REIGN OF ANGIOSPERMS IN THE LATER CRETACEOUS AND KAINOZOIC.
It is a remarkable fact in geological chronology that the culmination of the vegetable kingdom antedates that of the animal. The placental mammals, the highest group of the animal kingdom, are not known till the beginning of the Eocene Tertiary. The dicotyledonous Angiosperms, which correspond to them in the vegetable kingdom, occur far earlier—in the beginning of the Upper Cretaceous or close of the Lower Cretaceous. The reign of cycads and pines holds throughout the Lower Cretaceous, but at the close of that age there is a sudden incoming of the higher plants, and a proportionate decrease, more especially of the cycads.
I have already referred to the angiospermous wood supposed to be Devonian, but I fear to rest any conclusion on this isolated fact. Beyond this, the earliest indications of plants of this class have been found in the Lower Cretaceous. Many years ago Heer described and figured the leaves of a poplar (Populus primæva) from « 192 » the supposed Lower Cretaceous of Komé, in Greenland (Fig. 68). Two species, a Sterculia and a Laurus or Salix, occur among fossils described by me in the upper part of the Kootanie series of the Rocky Mountains, and Fontaine has recently found in the Potomac group of Virginia—believed to be of Neocomian age—several angiospermous species (Sassafras, Menispermites, Sapindus, Aralia, Populus, &c.) mixed with a rich flora of cycads and pines. These are the early forerunners of the modern angiospermous flora; but so far as known they do not occur below the Cretaceous, and in its lower portions only very rarely. When, however, we ascend into the Upper Cretaceous, whether of Europe or America, there is a remarkable incoming of the higher plants, under generic forms similar to those now existing. This is, in truth, the advent of the modern flora of the temperate regions of the earth. A very interesting tabular view of its early distribution is given by Ward, in the “American Journal of Science” for 1884, of which the following is a synopsis, with slight emendations. I may add that the new discoveries made since 1884 would probably tend to increase the proportionate number of dicotyledons in the newer groups.
Dicotyledonous Trees in the Cretaceous.
| Upper Senonian (Fox Hill group of America.) |
179 | species. |
| Lower Senonian Upper white chalk of Europe; Fort Pierre group of America; coal-measures of Nanaimo? |
81 | species. |
| Turonian Lower white chalk; New Jersey marls; Belly R. group. |
20 | species. |
| Cenomanian. (Chalk-marl, greensand, and Gault, Niobrara and Dakota groups of America); Dunvegan group of Canada; Amboy clays of New Jersey. |
357 | species. |
| Neocomian (Lower greensand and Speeton clay, Wealden and Hastings sands, Kootanie and Queen Charlotte groups of Canada.) |
20 | species.[DK] |
Thus we have a great and sudden inswarming of the higher plants of modern types at the close of the Lower Cretaceous. In relation to this, Saporta, one of the most enthusiastic of evolutionists, is struck by this phenomenon of the sudden appearance of so many forms, and some of them the most highly differentiated of dicotyledonous plants. The early stages of their evolution may, he thinks, have been obscure and as yet unobserved, or they may have taken place in some separate region, or mother country as yet undiscovered, or they may have been produced by a rapid and unusual multiplication of flower-haunting insects! Or it is even conceivable that the apparently sudden elevation of plants may have been due to causes still unknown. This last seems, indeed, the only certain inference in the case, since, as Saporta proceeds to say in conclusion: “Whatever hypothesis one may prefer, the fact of the rapid multiplication of dicotyledons, and of their simultaneous appearance in a great number of places in the northern hemisphere at the beginning of the Cenomanian epoch, cannot be disputed.”[DL]
[DL] “Monde des Plantes,” p. 197.
The leaves described by Heer, from the Middle Cretaceous of Greenland, are those of a poplar (P. primæva). Those which I have described from a corresponding horizon in the Rocky Mountains are a Sterculites (S. vetustula), probably allied to the mallows, and an elongated leaf, Laurophyllum (L. crassinerve) (Fig. 69), which may, however, have belonged to a willow rather than a laurel. These are certainly older than the Dakota group « 194 » of the United States and the corresponding formations in Canada. On the eastern side of the American continent, in Virginia, the Potomac series is supposed to be of Lower Cretaceous age, and here Fontaine, as already stated, has found an abundant flora of cycads, conifers, and ferns, with a few angiospermous leaves, which have not yet been described.
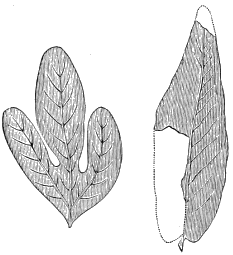
In the Canadian Rocky Mountains, a few hundreds of feet above the beds holding the beforementioned species, are the shales of the Mill Creek series, rich in many species of dicotyledonous leaves, and corresponding in age with the Dakota group, whose fossils have been so well described, first by Heer and Capellini, and afterward by Lesquereux. We may take this Dakota group and the quader-sand stone of Germany as types of the plant-bearing Cenomanian, and may notice the forms occurring in them.
In the first place, we recognise here the successors of our old friends, the ferns and the pines, the latter represented by such genera as Taxites, Sequoia, Glyptostrobus, Gingko, and even Pinus itself. We also have a few cycads, but not so dominant as in the previous ages. The fan-palms are well represented, both in America and in the corresponding series in Europe, especially by the genus Sabal, which is the characteristic American type of fan-palm, and there is one genus which Saporta regards as intermediate between the fan-palms and the pinnately leaved species. There are also many fragments of stems « 195 » and leaves of carices and grasses, so that these plants, now so important to the nourishment of man and his companion animals, were already represented.
But the great feature of the time was its dicotyledonous forests, and I have only to enumerate the genera supposed to be represented in order to show the richness of the time in plants of this type. It may be necessary to explain here that the generic names used are mostly based on leaves, and consequently cannot be held as being « 196 » absolutely certain, since we know that at present one genus may have considerable variety in its leaves, and, on the other hand, that plants of different genera may be very much alike in their foliage. There is, however, undoubtedly a likeness in plan or type of structure in leaves of closely allied plants, and, therefore, if judiciously studied, they can be determined with at least approximate certainty.[DM] More especially we can attain to much certainty when the fruits as well as the leaves are found, and when we can obtain specimens of the wood, showing its structure. Such corroboration is not wanting, though unfortunately the leaves of trees are generally found drifted away from the other organs once connected with them. In my own experience, however, I have often found determinations of the leaves of trees confirmed by the discovery of their fruits or of the structure of their stems. Thus, in the rich cretaceous plant-beds of the Dunvegan series we have beech-nuts associated in the same beds with leaves referred to Fagus. In the Laramie beds I determined many years ago nuts of the Trapa or water-chestnut, and subsequently Lesquereux found, in beds in the United States, leaves which he referred to the same genus. Later, I found in collections made on the Red Deer River of Canada my fruits and Lesquereux’s leaves on the same slab. The presence of trees of the genera Carya and Juglans in the same formation was inferred from their leaves, and specimens have since been obtained of silicified wood, with the microscopic structure of the modern butternut. Still we are willing to admit that determinations from leaves alone are liable to doubt.
[DM] Great allowance has to be made for the variability of leaves of the same species. The modern hazel (C. rostrata) is a case in point. Its leaves, from different parts of the same plant, are so dissimilar in form and size that they might readily be regarded as of different species.
In the matter of names of fossil leaves, I sympathise very strongly with Dr. Nathorst, of Stockholm, in his « 197 » objection to the use of modern generic names for mere leaves, and would be quite content to adopt some non-committal termination, as that of “phyllum” or “ites” suggested by him. I feel, however, that almost as much is taken for granted if a plant is called Corylophyllum or Corylites, as if called Corylus. In either case a judgment is expressed as to its affinities, which if wrong under the one term is wrong under the other; and after so much has been done by so many eminent botanists, it seems inexpedient to change the whole nomenclature for so small and questionable an advantage. I wish it, however, to be distinctly understood that plants catalogued on the evidence of leaves alone are for the most part referred to certain genera on grounds necessarily imperfect, and their names are therefore subject to correction, as new facts may be obtained.
The more noteworthy modern genera included in the Dakota flora, as catalogued by Lesquereux, are the following: Liquidambar, the sweet-gum, is represented both in America and Europe, the leaves resembling those of the modern species, but with entire edges, which seems to be a common peculiarity of Cretaceous foliage.[DN] Populus (poplar), as already stated, appears very early in Greenland, and continues with increasing number of species throughout the Cretaceous and Tertiary. Salix (willow) appears only a little later and continues. Of the family Cupuliferæ we have Fagus (beech), Quercus (oak), and Castanea (chestnut), which appear together in the Dakota group and its equivalents. Fruits of some of the species are known, and also wood showing structure. « 198 »Betula (birch) is represented by a few species, and specimens of its peculiar bark are also common. Alnus (alder) appears in one species at least. The genus Plat anus (Fig. 71), that of the plane-trees, represented at present by one European and one American species, has several species in the Cretaceous, though the plane-trees seem to culminate in the early part of the succeeding Eocene, where there are several species with immense leaves. The large « 199 » leaves, known as Credneria, found in the Cenomanian of Europe, and those called Protophyllum (Fig. 72) in America, appear to be nearer to the plane-trees than to any others, though representing an extinct type. The laurels are represented in this age, and the American genus Sassafras, which has now only one species, has not one merely but several species in the Cretaceous. Diospyros, the persimmon-tree, was also a Cretaceous genus.
[DN] With reference to this, something may be learned from the leaves of modern trees. In these, young shoots have leaves often less toothed and serrated than those of the adult tree. A remarkable instance is the Populus grandidentatus of America, the young shoots of which have entire leaves, quite unlike except in venation those of the parent tree, and having an aspect very similar to that of the Cretaceous poplars.
The single species of the beautiful Liriodendron, or tulip-tree, is a remnant of a genus which had several Cretaceous species (Figs. 74, 75). The magnolias, still well represented in the American flora, were equally plentiful in the Cretaceous (Fig. 73). The walnut family were well represented by species of Juglans (butternut) and Carya, or hickory. In all, no less than forty-eight genera are present belonging to at least twenty-five families, running through the whole range of the dicotyledonous exogens. This is a remarkable result, indicating a sudden profusion « 200 » of forms of these plants of a very striking character. It is further to be observed that some of the genera have many species in the Cretaceous and dwindle toward the modern. In others the reverse is the case—they have expanded in modern times. In a number there seems to have been little change.
Dr. Newberry has given, in the “Bulletin of the Torrey Botanical Club” an interesting résumé of the history of the beautiful Liriodendron, or tulip-tree, which may be taken as an example of a genus which has gone down in importance in the course of its geological history.
"The genus Liriodendron, as all botanists know, is represented in the present flora by a single species, ‘the tulip-tree’ which is confined to eastern America, but grows over all the area lying between the Lakes and the Gulf, the Mississippi and the Atlantic. It is a magnificent tree, on the « 201 » whole, the finest in our forests. Its cylindrical trunk, sometimes ten feet in diameter, carries it beyond all its associates in size, while the beauty of its glossy, lyre-shaped leaves and tulip-like flowers is only surpassed by the flowers and foliage of its first cousin, Magnolia grandiflora. That a plant so splendid should stand quite alone in the vegetation of the present day excited the wonder of the earlier botanists, but the sassafras, the sweet-gum, and the great Sequoias of the far West afford similar examples of isolation, and the latter are still more striking illustrations of solitary grandeur." (Figs. 74 and 75.)
"Three species of Liriodendron are indicated by leaves found in the Amboy clays—Middle Cretaceous—of New Jersey, and others have been obtained from the Dakota group in the West, and from the Upper Cretaceous strata of Greenland. Though differing considerably among themselves in size and form, all these have the deep sinus of the upper extremity so characteristic of the genus, and the nervation is also essentially the same. Hence, we must conclude that the genus Liriodendron, now represented « 202 » by a single species, was in the Cretaceous age much more largely developed, having many species, and those scattered throughout many lands. In the Tertiary age the genus continued to exist, but the species seem to have been reduced to one, which is hardly to be distinguished from that now living. In many parts of Europe leaves of the tulip-tree have been found, and it extended as far south as Italy. Its presence there was first made known by Unger, in his ‘Synopsis,’ page 232, and in his ‘Genera et Species,’ page 443, where he describes it under the name of Liriodendron procaccinii. The genus has also been noticed in Europe by Massalongo, Heer, and Ettingshausen, and three species have been distinguished. All these are, however, so much like the living species that they should probably be united with it. We here have a striking illustration of the wide distribution of a species which has retained its characters both of fruit and leaf quite unchanged through long migrations and an enormous lapse of time.
“In Europe the tulip-tree, like many of its American associates, seems to have been destroyed by the cold of the Ice period, the Mediterranean cutting off its retreat, but in America it migrated southward over the southern extension of the continent and returned northward again with the amelioration of the climate.”
Leaves of Liriodendron have been recognised in the Cretaceous of Greenland, though it is now a tree of the warm temperate region, and Lesquereux describes several species from the Dakota group. But the genus has not yet been recognised in the Laramie or in the Upper Cretaceous of British Columbia. In the paper above quoted, Newberry describes three new species from the Amboy clays, one of which he considers identical with a Greenland form referred by Heer to L. Meekii of the Dakota group. Thus, if all Lesquereux’s species are to be accepted, the genus begins « 203 » in the Middle Cretaceous with at least nine American species.
In New Jersey the Amboy clays are referred to the same age with the Dakota beds of the West. In these Dr. Newberry has found a rich flora, including many angiosperms. The following is condensed from a preliminary notice in the “Bulletin of the Torrey Botanical Club”:[DO]
[DO] March, 1886.
"The flora of the Amboy clays is closely related to that of the Dakota group—most of the genera and some of the species being identical—so that we may conclude they were nearly contemporaneous, though the absence in New Jersey of the Fort Benton and Niobrara groups of the upper Missouri and the apparent synchronism of the New Jersey marls and the Pierre group indicate that the Dakota is a little the older.
“At least one-third of the species of the Amboy clays seem to be identical with leaves found in the Upper Cretaceous clays of Greenland and Aachen (Aix la Chapelle), which not only indicates a chronological parallelism, but shows a remarkable and unexpected similarity in the vegetation of these widely separated countries in the middle and last half of the Cretaceous age. The botanical character of the flora of the Amboy clays will be seen from the following brief synopsis:
“Algæ.—A small and delicate form, allied to Chondrites.
“Ferns.—Twelve species, generally similar and in part identical with those described by Heer from the Cretaceous beds of Greenland, and referred to the genera Dicksonia, Gleichenia, and Aspidium.
“Cycads.—Two species, probably identical with the forms from Greenland described by Heer under the names of Podozamites marginatus and P. tenuinervis.
“Conifers.—Fourteen species, belonging to the genera Moriconia, Brachyphyllum, Cunninghamites, Pinus, Sequoia, and others referred by Heer to Juniperus, Libocedrus, Frenelopsis, Thuya, and Dammara. Of these, the most abundant and most interesting are Moriconia cyclotoxon—the most beautiful of conifers—and Cunninghamites elegans, both of which occur in the Cretaceous clays of Aachen, Prussia, and Patoot, Greenland. The Brachyphyllum was a large and strong species, with imbricated cones, eight inches in length.
“The angiosperms form about seventy species, which include three of Magnolia, four of Liriodendron, three or four of Salix, three of Celastrophyllum (of which one is identical with a Greenland species), one Celastrus (also found in Greenland), four or five Aralias, two Sassafras, one Cinnamomum, one Hedera; with leaves that are apparently identical with those described by Heer as belonging to Andromeda, Cissites, Cornus, Dewalquea, Diospyros, Eucalyptus, Ficus, Ilex, Juglans, Laurus, Menispermites, Myrica, Myrsine, Prunus, Rhamnus, and others not yet determined.
“Some of the Aralias had palmately-lobed leaves, nearly a foot in diameter, and two of the tulip-trees (Liriodendron) had leaves quite as large as those of the living species. One of these had deeply lobed leaves, like those of the white oak. Of the other, the leaves resembled those of the recent tulip-tree, but were larger. Both had the peculiar emargination and the nervation of Liriodendron.
“Among the most interesting plants of the collection are fine species of Bauhinia and Hymenæa. Of these, the first is represented by a large number of leaves, some of which are six or seven inches in diameter. They are deeply bilobed, and have the peculiar and characteristic form and nervation of the leaves of this genus. Bauhinia is a leguminous genus allied to Cercis, and now inhabits « 205 » tropical and warm temperate climates in both hemispheres. Only one species occurs in the United States, Bauhinia lunarioides, Gray, found by Dr. Bigelow on the Rio Grande.
“Hymenæa is another of the leguminosæ, and inhabits tropical America. A species of this genus has been found in the Upper Cretaceous of France, but quite different from the one before us, in which the leaves are much larger, and the leaflets are united in a common petiole, which is winged; this is a modification not found in the living species, and one which brings it nearer to Bauhinia.
“But the most surprising discovery yet made is that of a number of quite large helianthoid flowers, which I have called Palæanthus. These are three to four inches in diameter, and exhibit a scaly involucre, enclosing what much resembles a fleshy receptacle with achenia. From the border of this radiate a number of ray florets, one to two inches in length, which are persistent and must have been scarious, like those of Helichrysum. Though these flowers so much resemble those of the compositæ, we are not yet warranted in asserting that such is certainly their character. In the Jurassic rocks of Europe and India some flowers not very unlike these have been found, which have been named Williamsonia, and referred to cycads by Carruthers. A similar fossil has been found in the Cretaceous rocks of Greenland, and named by Heer Williamsonia cretacea, but he questions the reference of the genus to the Cycadeæ, and agrees with Nathorst in considering all the species of Williamsonia as parasitic flowers, allied to Brugmansia or Rafflesia. The Marquis of Saporta regards them as monocotyledons, similar to Pandanus. More specimens of the flowers now exhibited will perhaps prove—what we can now only regard as probable—that the Compositæ, like the Leguminosæ, Magnoliaceæ, Celastraceæ, and other highly organised plants, formed part « 206 » of the Cretaceous flora. No composite flowers have before been found in the fossil state, and, as these are among the most complex and specialised forms of florescence, it has been supposed that they belonged only to the recent epoch, where they were the result of a long series of formative changes.”
The above presents some interesting new types not heretofore found in the Middle Cretaceous. More especially the occurrence of large flowers of the composite type presents a startling illustration of the early appearance of a very elevated and complex form. Great interest also attaches to these Amboy beds, as serving, with those of Aix and Greenland, to show that the margins of the Atlantic were occupied with a flora similar to that occurring at the same time in the interior plateau of North America and on the Pacific slope.
The beds at Aix-la-Chapelle are, however, probably somewhat newer than the Dakota or Amboy beds, and correspond more nearly in age with those of the Cretaceous coal-field of Vancouver Island, where there is a very rich Upper Cretaceous flora, which I have noticed in detail in the “Transactions of the Royal Society of Canada.”[DP] In these Upper Cretaceous beds there are fan-palms as far north at least as the latitude of 49°, indicating a very mild climate at this period. This inference is corroborated by the Upper Cretaceous flora of Atané and Patoot in Greenland, as described by Heer.
[DP] Vol. ii., 1884.
The dicotyledonous plants above referred to are trees and shrubs. Of the herbaceous exogens of the period we know less. Obviously their leaves are less likely to find their way into aqueous deposits than the leaves of trees. They are, besides, more perishable, and in densely wooded countries there are comparatively few herbaceous plants. I have examined the beds of mud deposited at the mouth « 207 » of a woodland streamlet, and have found them stored with the fallen leaves of trees, but it was in vain to search for the leaves of herbaceous plants.
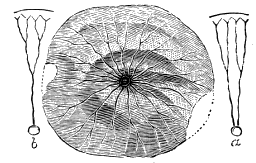
The climate of North America and Europe, represented by the Cenomanian vegetation, is not tropical but warm temperate; but the flora was more uniform than at present, indicating a very equable climate and the possibility of temperate genera existing within the Arctic circle, and it would seem to have become warmer toward the close of the period.
The flora of the Cenomanian is separated in most countries from that of the Senonian, or uppermost Cretaceous, by a marine formation holding few plants. This depends on great movements of elevation and depression, to which we must refer in the sequel. In a few regions, however, as in the vicinity of the Peace River in Canada, there are plant-bearing beds which serve to bridge over the interval between the Early Cenomanian and the later Cretaceous.[DQ]
[DQ] See paper by the author in the “Transactions of the Royal Society of Canada,” 1882.
To this interval also would seem to belong the Belly River series of western Canada, which contains important beds of Coal, but is Closely associated with the marine Fort Pierre series. A very curious herbaceous plant of this group, which I have named Brasenia antiqua, occurs in the beds associated with one of the coals. It is a close ally of the modern B. peltata, an aquatic plant which occurs in British Columbia and in eastern « 208 » America, and is also said to be found in Japan, Australia, and India, a width of distribution appropriate to so old a type (Fig. 76).
In so far as vegetable life is concerned, the transition from the Upper Cretaceous to the Tertiary or Kainozoic is easy, though in many parts of the world, and more especially in western Europe, there is a great gap in the deposits between the upper Chalk and the lowest Eocene. With reference to fossil plants, Schimper recognises in the Kainozoic, beginning with the oldest, five formations—Palæocene, Eocene, Oligocene, Miocene, and Pliocene. Throughout these a flora, similar to that of the Cretaceous on the one hand and the modern on the other, though with important local peculiarities, extends. There is evidence, however, of a gradual refrigeration, so that in the Pliocene the climates of the northern hemisphere were not markedly different from their present character.
In the first instance an important error was committed by palæobotanists, in referring to the Miocene many deposits really belonging to the Eocene. This arose from the early study of the rich plant-bearing Miocene beds of Switzerland, and from the similarity of the flora all the way from the Middle Cretaceous to the later Tertiary. The differences are now being worked out, and we owe to Mr. Starkie Gardner the credit of pointing these out in England, and to the Geological Survey of Canada that of collecting the material for exhibiting them in the more northern part of America.
In the great interior plain of America there rests on the Cretaceous a series of clays and sandstones with beds of lignite, some of them eighteen feet in thickness. This was formerly known as the lignitic or lignite Tertiary, but more recently as the Laramie series. These beds were deposited in fresh or brackish water, in an internal sea or group of lakes and swamps, when the continent was lower than at present. They have been « 209 » studied both in the United States[DR] and Canada; and, though their flora was originally referred by mistake to the Miocene, it is now known to be Eocene or Palæocene, or even in part a transition group between the latter and the Cretaceous. The following remarks, taken chiefly from recent papers by the author,[DS] will serve to illustrate this:
[DR] See more especially the elaborate and valuable reports by Lesquereux and Newberry, and a recent memoir by Ward on “Types of the Laramie Flora,” “Bulletins of the United States Geological Survey,” 1887.
[DS] “Transactions of the Royal Society of Canada,” 1886-'87.
On the geological map of Canada the Laramie series, formerly known as the lignitic or lignite Tertiary, occurs, with the exception of a few outliers, in two large areas west of the 100th meridian, and separated from each other by a tract of older Cretaceous rocks, over which the Laramie beds may have extended, before the later denudation of the region.
The most eastern of these areas, that of the Souris River and Wood Mountain, extends for some distance along the United States boundary, between the 102d and 109th meridians, and reaches northward to about thirty miles south of the “elbow” of the South Saskatchewan River, which is on the parallel of 51° north. In this area the lowest beds of the Laramie are seen to rest on those of the Fox Hill group of the Upper Cretaceous, and at one point on the west they are overlaid by beds of Miocene Tertiary age, observed by Mr. McConnell, of the Geological Survey, in the Cypress Hills, and referred by Cope, on the evidence of mammalian remains, to the White River division of the United States geologists, which is regarded by them as Lower Miocene.[DT] The age of the Laramie beds is thus stratigraphically determined to be between the Fox Hill Cretaceous and the Lower « 210 » Miocene. They are also undoubtedly continuous with the Fort Union group of the United States geologists on the other side of the international boundary, and they contain similar fossil plants. They are divisible into two groups—a lower, mostly argillaceous, and to which the name of “Bad Lands beds” may be given, from the “bad lands” of Wood Mountain, where they are well exposed, and an upper, partly arenaceous member, which may be named the Souris River or Porcupine Creek division. In the lower division are found reptilian remains of Upper Cretaceous type, with some fish remains more nearly akin to those of the Eocene.[DU] Neither division has as yet afforded mammalian remains.
[DT] “Report of the Geological Survey of Canada,” 1885.
[DU] Cope, in Dr. G. M. Dawson’s “Report on the 49th Parallel.”
The western area is of still larger dimensions, and extends along the eastern base of the Rocky Mountains from the United States boundary to about the 55th parallel of latitude, and stretches eastward to the 111th meridian. In this area, and more especially in its southern part, the officers of the Geological Survey of Canada have recognised three divisions, as follows: (1) The Lower Laramie or St. Mary River series, corresponding in its character and fossils to the Lower or Bad Lands division of the other area. (2) A middle division, the Willow Creek beds, consisting of clays, mostly reddish, and not recognised in the other area. (3) The Upper Laramie or Porcupine Hills division, corresponding in fossils, and to some extent in mineral character, to the Souris River beds of the eastern area.
The fossil plants collected by Dr. G. M. Dawson in the eastern area were noticed by the author in an appendix to Dr. Dawson’s report on the 49th parallel, in 1875, and a collection subsequently made by Dr. Selwyn was described in the “Report of the Geological Survey of Canada” for 1879-'80. Those of the western area, and « 211 » especially collections made by myself near Calgary in 1883, and by officers of the Geological Survey in 1884, have been described in the “Transactions of the Royal Society of Canada” vols. iii. and iv.
In studying these fossil plants, I have found that there is a close correspondence between those of the Lower and Upper Laramie in the two areas above referred to respectively, and that the flora of the Lower Laramie is somewhat distinct from that of the Upper, the former being especially rich in certain aquatic plants, and the latter much more copious on the whole, and much more rich in remains of forest-trees. This is, however, possibly an effect rather of local conditions than of any considerable change in the flora, since some Upper Laramie forms recur as low as the Belly River series of the Cretaceous, which is believed on stratigraphical grounds to be considerably older than the Lower Laramie.
With reference to the correlation of these beds with those of the United States, some difficulty has arisen from the tendency of palæobotanists to refer the plants of the Upper Laramie to the Miocene age, although in the reports of Mr. Clarence King, the late director of the United States Geological Survey, these beds are classed, on the evidence of stratigraphy and animal fossils, as Upper Cretaceous. More recently, however, and partly perhaps in consequence of the views maintained by the writer since 1875, some change of opinion has occurred, and Dr. Newberry and Mr. Lesquereux seem now inclined to admit that what in Canada we recognise as Upper Laramie is really Eocene, and the Lower Laramie either Cretaceous or a transition group between this and the Eocene. In a recent paper [DV] Dr. Newberry gives a comparative table, in which he correlates the Lower « 212 » Laramie with the Upper Cretaceous of Vancouver Island and the Faxoe and Maestricht beds of Europe, while he regards the Upper Laramie as equivalent to European Eocene. Except in so far as the equivalence of the Lower Laramie and Vancouver Island beds is concerned, this corresponds very nearly with the conclusions of the writer in a paper published last year[DW]—namely, that we must either regard the Laramie as a transition Cretaceo-Eocene group, or must institute our line of separation in the Willow Creek or Middle Laramie division, which has, however, as yet afforded no fossil plants. I doubt, however, the equivalence of the Vancouver beds and the Lower Laramie, except perhaps in so far as the upper member of the former is concerned. I have also to observe that in the latest report of Mr. Lesquereux he still seems to retain in the Miocene certain formations in the West, which from their fossil plants I should be inclined to regard as Eocene.[DX]
[DV] Newberry, “Transactions of the New Fork Academy,” February, 1886.
[DW] “Transactions of the Royal Society of Canada,” vol. ii.
[DX] While these sheets were going through the press I received a very valuable report of Mr. Lester F. Ward upon the Laramie of the United States. I have merely had time to glance at this report, but can see that the views of the author agree closely with those above expressed.
Two ferns occurring in these beds are remarkable as evidence of the persistence of species, and of the peculiarities of their ancient and modern distribution. Onoclea sensibilis, the very common sensitive fern of eastern America, is extremely abundant in the Laramie beds over a great area in the West. Mr. Starkie Gardner and Dr. Newberry have also shown that it is identical with the Filicites Hebridicus of Forbes, from the early Eocene beds of the Island of Mull, in Scotland. Thus we have a species once common to Europe and America, but now restricted to the latter, and which has continued to exist over all the vast ages between the Cretaceous and the present day. In the Laramie beds I have found associated « 213 » with this species another and more delicate fern, the modern Davallia (Stenloma) tenuifolia, but this, unlike its companion, no longer occurs in America, but is found in the mountains of Asia. This is a curious illustration of the fact that frail and delicate plants may be more ancient than the mountains or plains on which they live.
There are also some very interesting and curious facts in connection with the conifers of the Laramie. One of the most common of these is a Thuja or arbor vitæ (the so-called “cedar” of Canada). The Laramie species has been named T. interrupta by Newberry, but it approaches very closely in its foliage to T. occidentalis, of eastern Canada, while its fruit resembles that of the western species, T. gigantea.
Still more remarkable are the Sequoias to which we have already referred, but which in the Laramie age seem to have been spread over nearly all North America. The fossil species are of two types, representing respectively the modern S. gigantea and S. sempervirens, and their wood, as well as that of Thuja, is found in great abundance in the lignites, and also in the form of silicified trunks, and corresponds with that of the recent species. The Laramie contains also conifers of the genera Glyptostrobus, Taxodium, and Taxus; and the genus Salisburia or gingko—so characteristic of the Jurassic and Cretaceous—is still represented in America as well as in Europe in the early Eocene.
We have no palms in the Canadian or Scottish Palæocene, though I believe they are found further south. The dicotyledonous trees are richly represented. Perhaps the most conspicuous were three species of Platanus, the leaves of which sometimes fill the sandstones, and one of which, P. nobilis, Newberry, sometimes attains the gigantic size of a foot or more in diameter of its blade. The hazels are represented by a large-leaved species, « 214 » C. Macquarrii, and by leaves not distinguishable from those of the modern American species, C. Americana and C. rostrata. There are also chestnuts and oaks. But the poplars and willows are specially abundant, being represented by no less than six species, and it would seem that all the modern types of poplar, as indicated by the forms and venation of the leaves, existed already in the Laramie, and most of them even in the Upper Cretaceous. Sassafras is represented by two species, and the beautiful group of Viburnum,, to which the modern tree-cranberry belongs, has several fine species, of some of which both leaves and berries have been found. The hickories and butternuts are also present, the horse-chestnut, the Catalpa and Sapindus, and some curious leaves which seem to indicate the presence of the modern genus Symphorocarpus, the snow-berry tribe.
The above may suffice to give an idea of the flora of the older Eocene in North America, and I may refer for details to the works of Newberry, Lesquereux, and Ward, already cited. I must now add that the so-called Miocene of Atanekerdluk, Greenland, is really of the same age, as also the “Miocene” of Mull, in Scotland, of Antrim, in Ireland, and of Bovey Tracey, in the south of England, and the Gelinden, or “Heersian” beds, of Belgium, described by Saporta. In comparing the American specimens with the descriptions given by Gardner of the leaf-beds at Ardtown, in Mull, we find, as already stated, Onoclea sensibilis, common to both. The species of Sequoia, Gingko, Taxus, and Glyptostrobus are also identical or closely allied, and so are many of the dicotyledonous leaves. For example, Platanoides Hebridicus is very near to P. nobilis, and Corylus Macquarrii is common to both formations, as well as Populus Arctica and P. Richardsoni. I may add that ever since 1875-'76, when I first studied the Laramie plants, I have maintained their identity with those of the Fort Union group « 215 » of the United States, and of the so-called Miocene of McKenzie River and Greenland, and that the whole are Paleocene; and this conclusion has now been confirmed by the researches of Gardner in England, and by the discovery of true Lower Miocene beds in the Canadian northwest, overlying the Laramie or lignite series.
In a bulletin of the United States Geological Survey (1886), Dr. White has established in the West the continuous stratigraphical succession of the Laramie and the Wahsatch Eocene, thus placing the Laramie conformably below the Lower Eocene of that region. Cope has also described as the Puerta group a series of beds holding vertebrate fossils, and forming a transition from the Laramie to the Wahsatch. White also testifies that a number of fresh-water mollusks are common to the Wahsatch and the Laramie. This finally settles the position of the Laramie so far as the United States geologists are concerned, and shows that the flora is to be regarded as Eocene if not Upper Cretaceous, in harmony with what has been all along maintained in Canada. An important résumé of the flora has just been issued by Ward in the bulletins of the United States Geological Survey (1887).
Before leaving this part of the subject, I would deprecate the remark, which I see occasionally made, that fossil plants are of little value in determining geological horizons in the Cretaceous and Tertiary. I admit that in these periods some allowance must be made for local differences of station, and also that there is a generic sameness in the flora of the northern hemisphere, from the Cenomanian to the modern, yet these local differences and general similarity are not of a nature to invalidate inferences as to age. No doubt, so long as palæobotanists seemed obliged, in deference to authority, and to the results of investigations limited to a few European localities, to group together, without distinction, all the floras of the later Cretaceous and earlier Tertiary, « 216 » irrespective of stratigraphical considerations, the subject lost its geological importance. But, when a good series has been obtained in any one region of some extent, the case becomes different. Though there is still much imperfection in our knowledge of the Cretaceous and Tertiary floras of Canada, I think the work already done is sufficient to enable any competent observer to distinguish by their fossil plants the Lower, Middle, and Upper Cretaceous, and the latter from the Tertiary; and, with the aid of the work already done by Lesquereux and Newberry in the United States, to refer approximately to its true geological position any group of plants from beds of unknown age in the West.
An important consequence arising from the above statements is that the period of warm climate which enabled a temperate flora to exist in Greenland was that of the later Cretaceous and early Eocene rather than, as usually stated, the Miocene. It is also a question admitting of discussion whether the Eocene flora of latitudes so different as those of Greenland, Mackenzie River, northwest Canada, and the United States, were strictly contemporaneous, or successive within a long geological period in which climatal changes were gradually proceeding. The latter statement must apply at least to the beginning and close of the period; but the plants themselves have something to say in favour of contemporaneity. The flora of the Laramie is not a tropical but a temperate flora, showing no doubt that a much more equable climate prevailed in the more northern parts of America than at present. But this equability of climate implies the possibility of a great geographical range on the part of plants. Thus it is quite possible and indeed highly probable that in the Laramie age a somewhat uniform flora extended from the Arctic seas through the great central plateau of America far to the south, and in like manner along the western coast of « 217 » Europe. It is also to be observed that, as Gardner points out, there are some differences indicating a diversity of climate between Greenland and England, and even between Scotland and Ireland and the south of England, and we have similar differences, though not strongly-marked, between the Laramie of northern Canada and that of the United States. When all our beds of this age from the Arctic sea to the 49th parallel have been ransacked for plants, and when the palæobotanists of the United States shall have succeeded in unravelling the confusion which now exists between their Laramie and the Middle Tertiary, the geologist of the future will be able to restore with much certainty the distribution of the vast forests which in the early Eocene covered the now bare plains of interior America. Further, since the break which in western Europe separates the flora of the Cretaceous from that of the Eocene does not exist in America, it will then be possible to trace the succession from the Mesozoic flora of the Trias and of the Queen Charlotte Islands and Kootanie series of the Lower Cretaceous up to the close of the Eocene; and to determine, for America at least, the manner and conditions under which the angiospermous flora of the later Cretaceous succeeded to the pines and cycads which characterised the beginning of the Cretaceous period. In so far as Europe is concerned, this may be more difficult, since the want of continuity of land from north to south seems there to have been fatal to the continuance of some plants during changes of climate, and there were also apparently in the Kainozoic period invasions at certain times of species from the south and east, which did not occur to the same extent in America.
In recent reports on the Tertiary floras of Australia and New Zealand,[DY] Ettingshausen holds that the flora of « 218 » the Tertiary, as a whole, was of a generalised character; forms now confined to the southern and northern hemispheres respectively being then common to both. It would thus seem that the present geographical diversities must have largely arisen from the great changes in climate and distribution of land and water in the later Tertiary.
[DY] “Geological Magazine,” August, 1887.
The length of our discussion of the early angiospermous flora does not permit us to trace it in detail through the Miocene and Pliocene, but we may notice the connection through these in the next chapter, and may refer to the magnificent publications of Heer and Lesquereux on the Tertiary floras of Europe and America respectively.
PLANTS FROM THE TERTIARY TO THE MODERN PERIOD.
It may be well to begin this chapter with a sketch of the general physical and geological conditions of the period which was characterised by the advent and culmination of the dicotyledonous trees.
In the Jurassic and earliest Cretaceous periods the prevalence, over the whole of the northern hemisphere and for a long time, of a monotonous assemblage of gymnospermous and acrogenous plants, implies a uniform and mild climate, and facility for intercommunication in the north. Toward the end of the Jurassic and beginning of the Cretaceous, the land of the northern hemisphere was assuming greater dimensions, and the climate probably becoming a little less uniform. Before the close of the Lower Cretaceous period the dicotyledonous flora seems to have been introduced, under geographical conditions which permitted a warm temperate climate to extend as far north as Greenland.
In the Cenomanian or Middle Cretaceous age we find the northern hemisphere tenanted with dicotyledonous trees closely allied to those of modern times, though still indicating a climate much warmer than that which at present prevails. In this age, extensive but gradual submergence of land is indicated by the prevalence of chalk and marine limestones over the surface of both continents; but a circumpolar belt seems to have been maintained, protecting the Atlantic and Pacific basins from « 220 » floating ice, and permitting a temperate flora of great richness to prevail far to the north, and especially along the southern margins and extensions of the circumpolar land. These seem to have been the physical conditions which terminated the existence of the old Mesozoic flora and introduced that of the Middle Cretaceous.
As time advanced the quantity of land gradually increased, and the extension of new plains along the older ridges of land was coincident with the deposition of the great Laramie series, and with the origination of its peculiar flora, which indicates a mild climate and considerable variety of station in mountain, plain, and swamp, as well as in great sheets of shallow and weedy fresh water.
In the Eocene and Miocene periods, the continents gradually assumed their present form, and the vegetation became still more modern in aspect. In that period of the Eocene, however, in which the great nummulitic limestones were deposited, a submergence of land occurred on the eastern continent which must have assimilated its physical conditions to those of the Middle Cretaceous. This great change, affecting materially the flora of Europe, was not equally great in America, which also by the north and south extension of its mountain-chains permitted movements of migration not possible in the Old World. From the Eocene downward, the remains of land-animals and plants are found chiefly in lake-basins occupying the existing depressions of the land, though more extensive than those now remaining. It must also be borne in mind that the great foldings and fractures of the crust of the earth which occurred at the close of the Eocene, and to which the final elevation of such ranges as the Alps and the Rocky Mountains belongs, permanently modified and moulded the forms of the continents.
These statements raise, however, questions as to the precise equivalence in time of similar floras found in different « 221 » latitudes. However equable the climate, there must have been some appreciable difference in proceeding from north to south. If, therefore, as seems in every way probable, the new species of plants originated on the Arctic land and spread themselves southward, this latter process would occur most naturally in times of gradual refrigeration or of the access of a more extreme climate—that is, in times of the elevation of land in the temperate latitudes, or, conversely, of local depression of land in the Arctic, leading to invasions of northern ice. Hence, the times of the prevalence of particular types of plants in the far north would precede those of their extension to the south, and a flora found fossil in Greenland might be supposed to be somewhat older than a similar flora when found farther south. It would seem, however, that the time required for the extension of a new flora to its extreme geographical limit is so small, in comparison with the duration of an entire geological period, that, practically, this difference is of little moment, or at least does not amount to antedating the Arctic flora of a particular type by a whole period, but only by a fraction of such period.
It does not appear that, during the whole of the Cretaceous and Eocene periods, there is any evidence of such refrigeration as seriously to interfere with the flora, but perhaps the times of most considerable warmth are those of the Dunvegan group in the Middle Cretaceous, and those of the later Laramie and oldest Eocene.
It would appear that no cause for the mild temperature of the Cretaceous needs to be invoked, other than those mutations of land and water which the geological deposits themselves indicate. A condition, for example, of the Atlantic basin in which the high land of Greenland should be reduced in elevation, and at the same time the northern inlets of the Atlantic closed against the invasion of Arctic ice, would at once restore climatic conditions « 222 » allowing of the growth of a temperate flora in Greenland. As Dr. Brown has shown,[DZ] and as I have elsewhere argued, the absence of light in the Arctic winter is no disadvantage, since, during the winter, the growth of deciduous trees is in any case suspended; while the constant continuance of light in the summer is, on the contrary, a very great stimulus and advantage.
[DZ] “Florula Discoana.”
It is a remarkable phenomenon in the history of genera of plants in the later Mesozoic and Tertiary, that the older genera appear at once in a great number of specific types, which become reduced as well as limited in range down to the modern. This is, no doubt, connected with the greater differentiation of local conditions in the modern; but it indicates also a law of rapid multiplication of species in the early life of genera. The distribution of the species of Salisburia, Sequoia, Platanus, Sassafras, Liriodendron, Magnolia, and many other genera, affords remarkable proofs of this.
Gray, Saporta, Heer, Newberry, Lesquereux, and Starkie Gardner have all ably discussed these points; but the continual increase of our knowledge of the several floras, and the removal of error as to the dates of their appearance, must greatly conduce to clearer and more definite ideas. In particular, the prevailing opinion that the Miocene was the period of the greatest extension of warmth and of a temperate flora into the Arctic, must be abandoned in favour of the later Cretaceous and Eocene; and, if I mistake not, this will be found to accord better with the evidence of general geology and of animal fossils.
In these various revolutions of the later Cretaceous and Kainozoic periods, America, as Dr. Gray has well pointed out, has had the advantage of a continuous stretch of high land from north to south, affording a more sure « 223 » refuge to plants in times of submergence, and means of escape to the south in times of refrigeration. Hence, the greater continuity of American vegetation and the survival of genera like Sequoia and Liriodendron, which have perished in the Old World. Still, there are some exceptions to this, for the gingko-tree is a case of survival in Asia of a type once plentiful in America, but now extinct there. Eastern Asia has had, however, some considerable share of the same advantage possessed by America, with the addition, referred to by Gray, of a better and more insular climate.
But our survey of these physical conditions can not be considered complete till we shall have considered the great Glacial age of the Pleistocene. It is certain that throughout the later Miocene and Pliocene the area of land in the northern hemisphere was increasing, and the large and varied continents were tenanted by the noblest vegetation and the grandest forms of mammalian life that the earth has ever witnessed. As the Pliocene drew to a close, a gradual diminution of warmth came on, and more especially a less equable climate, and this was accompanied with a subsidence of the land in the temperate regions and with changes of the warm ocean-currents. Thus gradually the summers became cooler and the winters longer and more severe, the hill-tops became covered with permanent snows, glaciers ploughed their way downward into the plains, and masses and fields of floating ice cooled the seas. In these circumstances the richer and more delicate forms of vegetation must have been chilled to death or obliged to remove farther south, and in many extensive regions, hemmed in by the advance of the sea on the one hand and land-ice on the other, they must have altogether perished.
Yet even in this time vegetation was not altogether extinct. Along the Gulf of Mexico in America, and in the Mediterranean basin in Europe, there were still some « 224 » remains of a moderate climate and certain boreal and arctic forms moving southward continued to exist here and there in somewhat high latitudes, just as similar plants now thrive in Grinnell Land within sight of the snows of the Greenland mountains. A remarkable summary of some of these facts as they relate to England was given by an eminent English botanist, Mr. Carruthers, in his address as President of the Biological Section of the British Association at Birmingham in 1886. At Cromer, on the coast of Norfolk, the celebrated forest-bed of newer Pliocene age, and containing the remains of a copious mammalian fauna, holds also remains of plants in a state admitting of determination. These have been collected by Mr. Reid, of the Geological Survey, and were reported on by Carruthers, who states that they represent a somewhat colder temperature than that of the present day. I quote the following details from the address.
With reference to the plants of the forest-bed or newer Pliocene he remarks as follows:
"Only one species (Trapa natans, Willd.) has disappeared from our islands. Its fruits, which Mr. Reid found abundantly in one locality, agree with those of the plants found until recently in the lakes of Sweden. Four species (Prunus speciosa, L., Œnanthe Tichenalii, Sm., Potamogeton pterophyllus, Sch., and Pinus abies, L.) are found at present only in Europe, and a fifth (Potamogeton trichoides, Cham.) extends also to North America; two species (Peucedanum palustre, Moench, and Pinus sylvestris, L.) are found also in Siberia, while six more (Sanguisorba officinalis, L., Rubus fruticosus, L., 5 Cornus sanguinea, L., Euphorbia amygdaloides, L., Quercus robur, L., and Potamogeton crispus, L.) extend into western Asia, and two (Fagus sylvatica, L., and Alnus glutinosa, L.) are included in the Japanese flora. Seven species, while found with the others, enter also into the Mediterranean flora, extending to North Africa: these « 225 » are Thalictrum minus, L., Thalictrum flavum, L., Ranunculus repens, L., Stellaria aquatica, Scop., Corylus avellana, L., Yannichellia palustris, L., and Cladium mariscus, Br. With a similar distribution in the Old World, eight species (Bidens tripartita, L., Myosotis cæspitosa, Schultz, Suæda maritima, Dum., Ceratophyllum demersum,, L., Sparganium ramosum, Huds., Potamogeton pectinatus, L., Carex paludosa, Good., and Osmunda regalis, L.) are found also in North America. Of the remainder, ten species (Nuphar luteum, Sm., Menyanthes trifoliata, L., Stachys palustris, L., Rumex maritimus, L., Rumex acetosella, L., Betula alba, L., Scirpus pauciflorus, Lightf., Taxus baccata, L., and Isoetes lacustris, L.) extend round the north temperate zone, while three (Lycopus europæus, L., Alisma plantago, L., and Phragmites communis, Trim), having the same distribution in the north, are found also in Australia, and one (Hippuris vulgaris, L.) in the south of South America. The list is completed by Ranunculus aquatilis, L., distributed over all the temperate regions of the globe, and Scirpus lacustris, L., which is found in many tropical regions as well."
He remarks that these plants, while including species now very widely scattered, present no appreciable change of characters.
Above this bed are glacial clays, which hold other species indicating an extremely cold climate. They are few in number, only Salix polaris, a thoroughly arctic species, and its ally, S. cinerea, L., and a moss, Hypnum turgescens, Schimp., no longer found in Britain, but an Alpine and arctic species. This bed belongs to the beginning of the Glacial period, the deposits of which have as yet afforded no plants in England. But plants occur in post-glacial and upper-glacial beds in different parts of England, to which Carruthers thus refers:
"The period of great cold, during which arctic ice « 226 » extended far into temperate regions, was not favorable to vegetable life. But in some localities we have stratified clays with plant-remains later than the Glacial epoch, yet indicating that the great cold had not then entirely disappeared. In the lacustrine beds at Holderness is found a small birch (Betula nana, L.), now limited in Great Britain to some of the mountains of Scotland, but found in the arctic regions of the Old and New World and on Alpine districts in Europe, and with it Prunus padus, L., Quercus robur, L., Corylus avellana, L., Alnus glutinosa, L., and Pinus sylvestris, L. In the white clay-beds at Bovey Tracey of the same age there occur the leaves of Arctostaphylos uva-ursi, L., three species of willow, viz., Salix cinerea, L., S. myrtilloides, L., and S. polaris, Wahl., and in addition to our Alpine Betula nana, L., the more familiar B. alba, L. Two of these plants have been lost to our flora from the change of climate that has taken place, viz., Salix myrtilloides, L., and S. polaris, Wahl.; and Betula nana, L., has retreated to the mountains of Scotland. Three others (Dryas octopetala, L., Arctostaphylos uva-ursi, L., and Salix herbacea, L.) have withdrawn to the mountains of northern England, Wales, and Scotland, while the remainder are still found scattered over the country. Notwithstanding the diverse physical conditions to which these plants have been subjected, the remains preserved in these beds present no characters by which they can be distinguished from the living representatives of the species."
One of the instances referred to is very striking. At Bovey Tracey the arctic beds rest directly on those holding the rich, warm temperate flora of the Eocene; so that here we have the evidence of fossil plants to show the change from the climate of the Eocene to that of arctic lands, and the modern vegetation to indicate the return of a warm temperature.
In Canada, in the Pleistocene beds known as the Leda clays, intervening between the lower boulder clay and the Saxicava sand, which also holds boulders, there are beds holding fossil plants, in some places intermixed with sea-shells and bones of marine fishes, showing that they were drifted into the sea at a time of submergence. These remains are boreal rather than arctic in character, and with the remains of drift-wood often found in the boulder deposits serve to indicate that there were at all times oases of hardy life in the glacial deserts, just as we find these in polar lands at the present day. I condense from a paper on these plants[EA] the following facts, with a few additional notes:
[EA] “Canadian Naturalist,” 1866.
The importance of all information bearing on the temperature of the Post-pliocene period invests with much interest the study of the land-plants preserved in deposits of this age. Unfortunately, these are few in number, and often not well preserved. In Canada, though fragments of the woody parts of plants occasionally occur in the marine clays and sands, there is only one locality which has afforded any considerable quantity of remains of their more perishable parts. This is the well-known deposit of Leda clay at Green’s Creek, on the Ottawa, celebrated for the perfection in which the skeletons of the capelin and other fishes are preserved in the calcareous nodules imbedded in the clay. In similar nodules, contained apparently in a layer somewhat lower than that holding the ichthyolites, remains of land-plants are somewhat abundant, and, from their association with shells of Leda glacialis, seem to have been washed down from the land into deep water. The circumstances would seem to have been not dissimilar from those at present existing in the northeast arm of Gaspé Basin, where I have dredged from mud now being deposited in deep water, living « 228 » specimens of Leda limatula, mixed with remains of land-plants.
The following are the species of plants recognised in these nodules:
1. Drosera rotundifolia, Linn. In a calcareous nodule from Green’s Creek, the leaf only preserved. This plant is common in bogs in Canada, Nova Scotia, and Newfoundland, and thence, according to Hooker, to the Arctic circle. It is also European.
2. Acer spicatum, Lamx. (Acer montanum, Aiton.) Leaf in a nodule from Green’s Creek. Found in Nova Scotia and Canada, also at Lake Winnipeg, according to Richardson.
3. Potentilla Canadensis, Linn. In nodules from Green’s Creek; leaves only preserved. I have had some difficulty in determining these, but believe they must be referred to the species above named, or to P. simplex, Michx., supposed by Hooker and Gray to be a variety. It occurs in Canada and New England, but I have no information as to its range northward.
4. Gaylussaccia resinosa, Torrey and Gray. Leaf in nodule at Green’s Creek. Abundant in New England and in Canada, also on Lake Huron and the Saskatchewan, according to Richardson (Fig. 77).
5. Populus balsamifera, Linn. Leaves and branches in nodules at Green’s Creek. This is by much the most common species, and its leaves are of small size, as if from trees growing in cold and exposed situations. The species is North American and Asiatic, and abounds in New England and Canada. It extends to the Arctic circle, and is « 229 » abundant on the shores of the Great Slave Lake and on the McKenzie River, and according to Richardson constitutes much of the drift timber of the Arctic coast (Fig. 78).
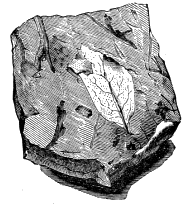
Fig. 77.—Gaylussaccia resinosa.
Pleistocene, Canada.
|
Fig. 78.—Populus balsamifera. Pleistocene, Canada.
|
6. Thuja occidentalism Linn. Trunks and branches in the Leda clay at Montreal. This tree occurs in New England and Canada, and extends northward into the Hudson Bay territories. It is a northern though not arctic species in its geographical range. According to Lyell it occurs associated with the bones of Mastodon in New Jersey. From the great durability of its wood, it is one of the trees most likely to be preserved in aqueous deposits.
7. Potamogeton perfoliatus, Linn. Leaves and seeds in nodules at Green’s Creek. Inhabits streams of the Northern States and Canada, and according to Richardson extends to Great Slave Lake.
8. Potamogeton pusillus. Quantities of fragments which I refer to this species occur in nodules at Green’s Creek. They may possibly belong to a variety of P. hybridus which, together with P. natans, now grows in « 230 » the river Ottawa, where it flows over the beds containing these fossils.
9. Cariceæ and Gramineæ. Fragments in nodules from Green’s Creek appear to belong to plants of these groups, but I cannot venture to determine their species.
10. Equisetum scirpoides, Michx. Fragments in nodules, Green’s Creek. This is a widely distributed species, occurring in the Northern States and Canada.
11. Fontinalis. In nodules at Green’s Creek there occur, somewhat plentifully, branches of a moss apparently of the genus Fontinalis.
12. Algæ. With the plants above mentioned, both at Green’s Creek and at Montreal, there occur remains of sea-weeds (Fig. 79). They seem to belong to the genera Fucus and Ulva, but I cannot determine the species. A thick stem in one of the nodules would seem to indicate a large Laminaria. With the above there are found at Green’s Creek a number of fragments of leaves, stems, and fruits, which I have not been able to refer to their species, principally on account of their defective state of preservation.
None of the plants above mentioned is properly arctic in its distribution, and the assemblage may be characterised as a selection from the present Canadian flora of some of the more hardy species having the most northern range. Green’s Creek is in the central part of Canada, near to the parallel of 46°, and an accidental selection « 231 » from its present flora, though it might contain the same species found in the nodules, would certainly include with these, or instead of some of them, more southern forms. More especially the balsam poplar, though that tree occurs plentifully on the Ottawa, would not be so predominant. But such an assemblage of drift-plants might be furnished by any American stream flowing in the latitude of 50° to 55° north. If a stream flowing to the north, it might deposit these plants in still more northern latitudes, as the McKenzie River does now. If flowing to the south, it might deposit them to the south of 50°. In the case of the Ottawa, the plants could not have been derived from a more southern locality, nor probably from one very far to the north. We may therefore safely assume that the refrigeration indicated by these plants would place the region bordering the Ottawa in nearly the same position with that of the south coast of Labrador fronting on the Gulf of St. Lawrence at present. The absence of all the more arctic species occurring in Labrador should perhaps induce us to infer a somewhat milder climate than this.
The moderate amount of refrigeration thus required would in my opinion accord very well with the probable conditions of climate deducible from the circumstances in which the fossil plants in question occur. At the time when they were deposited the sea flowed up the Ottawa valley to a height of 200 to 400 feet above its present level, and the valley of the St. Lawrence was a wide arm of the sea, open to the arctic current. Under these conditions the immense quantities of drift-ice from the northward, and the removal of the great heating surface now presented by the low lands of Canada and New England, must have given for the Ottawa coast of that period a summer temperature very similar to that at present experienced on the Labrador coast, and with this conclusion the marine remains of the Leda clay, as well as the few « 232 » land molluscs whose shells have been found in the beds containing the plants, and which are species still occurring in Canada, perfectly coincide.
The climate of that portion of Canada above water at the time when these plants were imbedded may safely be assumed to have been colder in summer than at present, to an extent equal to about 5° of latitude, and this refrigeration may be assumed to correspond with the requirements of the actual geographical changes implied. In other words, if Canada was submerged until the Ottawa valley was converted into an estuary inhabited by species of Leda, and frequented by capelin, the diminution of the summer heat consequent on such depression would be precisely suitable to the plants occurring in these deposits, without assuming any other cause of change of climate.
I have arranged elsewhere the Post-pliocene deposits of the central part of Canada, as consisting of, in ascending order: (1) The boulder clay; (2) a deep-water deposit, the Leda clay; and (3) a shallow-water deposit, the Saxicava sand. But, although I have placed the boulder clay in the lowest position, it must be observed that I do not regard this as a continuous layer of equal age in all places. On the contrary, though locally, as at Montreal, under the Leda clay, it is in other places and at other levels contemporaneous with or newer than that deposit, which itself also locally contains boulders.
At Green’s Creek the plant-bearing nodules occur in the lower part of the Leda clay, which contains a few boulders, and is apparently in places overlaid by large boulders, while no distinct boulder clay underlies it. The circumstances which accumulated the thick bed of boulder clay near Montreal were probably absent in the Ottawa valley. In any case we must regard the deposits of Green’s Creek as coeval with the Leda clay of Montreal, and with the period of the greatest abundance of « 233 » Leda glacialis, the most exclusively arctic shell of these deposits. In other words, I regard the plants above mentioned as probably belonging to the period of greatest refrigeration of which we have any evidence, of course not including that mythical period of universal incasement in ice, of which, as I have elsewhere endeavoured to show, in so far as Canada is concerned, there is no evidence whatever.[EB]
[EB] Notes on Post-Pliocene of Canada, “Canadian Naturalist,” 1872.
The facts above stated in reference to Post-pliocene plants concur, with all the other evidence I have been able to obtain, in the conclusion that the refrigeration of Canada in the Post-pliocene period consisted of a diminution of the summer heat, and was of no greater amount than that fairly attributable to the great depression of the land and the different distribution of the ice-bearing arctic current.
In connection with the plants above noticed, it is interesting to observe that at Green’s Creek, at Pakenham Mills, at Montreal, and at Clarenceville on Lake Champlain, species of Canadian Pulmonata have been found in deposits of the same age with those containing the plants. The species which have been noticed belong to the genera Lymnea and Planorbis.
The Glacial age was, fortunately, not of very long duration, though its length has been much exaggerated by certain schools of geologists,[EC] It passed away, and a returning cosmic spring gladdened the earth, and was ushered in by a time of great rainfall and consequent denudation and deposit, which has been styled the “Pluvial Period” The remains of the Pliocene forests then returned—with somewhat diminished numbers of species—from « 234 » the south and again occupied the land, though they have not been able, in their decimated condition, to restore the exuberance of the flora of the earlier Tertiary. In point of fact, as we shall see in the next chapter, it is the floras originating within the polar circle and coming down from the north that are rich and copious. Those that, after periods of cold or submergence, return from the south, are comparatively poor. Hence the modern flora is far inferior to that of the Middle Kainozoic. In America, however, and in eastern Asia, for reasons already stated, the return was more abundant than in Europe.
[EC] This I have long maintained on grounds connected with Pleistocene fossils, amount of denudation and deposit, &c., and I am glad to see that Prestwich, the best English authority on such subjects, has recently announced similar conclusions, based on independent reasons.
Simultaneously with the return of the old temperate flora, the arctic plants that had overspread the land retreated to mountain-tops, now bared of ice and snow, and back to the polar lands whence they came; and so it happens that, on the White Mountains, the Alps, and the Himalayas, we have insular patches of the same groups of plants that exist around the pole.
These changes need not have required a very long time, for the multiplication and migration of plants are very rapid, especially when aided by the agency of migratory animals. Many parts of the land must, indeed, have been stocked with plants from various sources, and by agencies—as that of the sea—which might at first sight seem adverse to their distribution. The British Islands, for example, have no indigenous plants. Their flora consists mainly of Germanic plants, which must have migrated to Britain in that very late period of the Post-glacial when the space now occupied by the North Sea was mostly dry land. Other portions of it are Scandinavian plants, perhaps survivors of the Glacial age, or carried by migratory birds; and still another element consists of Spanish plants, brought north by spring migrants, and establishing themselves in warm and sheltered spots, just as the arctic plants do on the bleak hill-tops. « 235 » The Bermudas, altogether recent islands, have one hundred and fifty species of native plants, all of which are West Indian and American, and must have been introduced by the sea-currents or by migratory birds.
And so the earth became fitted for the residence of modern man. Yet it is not so good or Edenic a world as it once was, or as it may yet become, were another revolution to restore a mild climate to the arctic regions, and to send down a new swarm of migratory species to renew the face of the earth and restore it to its pristine fertility of vegetable life.
Thus closes this long history of the succession of plants, reaching from the far back Laurentian to the present day. It has, no doubt, many breaks, and much remains to be discovered. Yet it may lead us to some positive conclusions regarding the laws of the introduction of plants.
One of these, and perhaps the most remarkable of all, is that certain principles were settled very far back, and have remained ever since. We have seen that in the earliest geological periods all that pertains to the structure, powers, and laws of the vegetable cell was already fixed and settled. When we consider how much this implies of mechanical structure and chemical and vital property, the profound significance of this statement becomes apparent. The relations in these respects between the living cell and the soil, the atmosphere and the sunshine, were apparently as perfect in the early Palæozoic as in any subsequent time. The same may be said of the structures of the leaf and of the stem. In such old forms as Nematophyton these were, it is true, peculiar and rudimentary, but in the Devonian and Carboniferous the structure of leaves and stems embodied all the parts and principles that we find at present. In regard to fructification there has been more progress, for, so far as we know, the highest and most complex forms of flowery, « 236 » fruits, and seeds belong to the more recent periods, and simpler forms were at least dominant in the older times. Yet even in this respect the great leading laws and structures of bisexual reproduction were perfected in the early Palæozoic, and the improvements introduced in the gymnosperm and the angiosperm of later periods have consisted mainly in additions of accessory parts, and in modifications and refinements suited to the wants of the higher and more complex types.
GENERAL LAWS OF ORIGIN AND MIGRATIONS OF PLANTS.—RELATIONS OF RECENT AND FOSSIL FLORAS.
The origination of the successive floras which have occupied the northern hemisphere in geological time, not, as one might at first sight suppose, in the sunny climes of the south, but under the arctic skies, is a fact long known or suspected. It is proved by the occurrence of fossil plants in Greenland, in Spitzbergen, and in Grinnell Land, under circumstances which show that these were their primal homes. The fact bristles with physical difficulties, yet is fertile of the most interesting theoretical deductions, to reach which we may well be content to wade through some intricate questions. Though not at all a new fact, its full significance seems only recently to have dawned on the minds of geologists, and within the last few years it has produced a number of memoirs and addresses to learned societies, besides many less formal notices.[ED]
[ED] Saporta, “Ancienne Végétation Polaire”; Hooker, “Presidential Address to Royal Society,” 1878; Thistleton Dyer, “Lecture on Plant Distribution”; Mr. Starkie Gardner, “Letters in ‘Nature,’” 1878, &c. The basis of most of these brochures is to be found in Heer’s “Flora Fossilis Arctica.”
The earliest suggestion on the subject known to the writer is that of Prof. Asa Gray, in 1867, with reference to the probable northern source of the related floras of North America and eastern Asia. With the aid of the new facts disclosed by Heer and Lesquereux, Gray returned « 238 » to the subject in 1872, and more fully developed this conclusion with reference to the Tertiary floras,[EE] and he has recently still further discussed these questions in an able lecture on “Forest Geography and Archæology.”[EF] In this he puts the case so well and tersely that we may quote the following sentences as a text for what follows:
[EE] Address to American Association.
[EF] “American Journal of Science,” xvi., 1818.
“I can only say, at large, that the same species (of Tertiary fossil plants) have been found all round the world; that the richest and most extensive finds are in Greenland; that they comprise most of the sorts which I have spoken of, as American trees which once lived in Europe—magnolias, sassafras, hickories, gum-trees, our identical southern cypress (for all we can see of difference), and especially Sequoias, not only the two which obviously answer to the two big-trees now peculiar to California, but several others; that they equally comprise trees now peculiar to Japan and China, three kinds of gingko-trees, for instance, one of them not evidently distinguishable from the Japan species which alone survives; that we have evidence, not merely of pines and maples, poplars, birches, lindens, and whatever else characterise the temperate zone forests of our era, but also of particular species of these, so like those of our own time and country that we may fairly reckon them as the ancestors of several of ours. Long genealogies always deal more or less in conjecture; but we appear to be within the limits of scientific inference when we announce that our existing temperate trees came from the north, and within the bounds of nigh probability when we claim not a few of them as the originals of present species. Remains of the same plants have been found fossil in our temperate region as well as in Europe.”
Between 1860 and 1870 the writer was engaged in working out all that could be learned of the Devonian plants of eastern America, the oldest known flora of any richness, and which consists almost exclusively of gigantic, and to us grotesque, representatives of the club-mosses, ferns, and mares'-tails, with some trees allied to the cycads and pines. In this pursuit nearly all the more important localities were visited, and access was had to the large collections of Prof. Hall and Prof. Newberry, in New York and Ohio, and to those made in the remarkable plant-bearing beds of New Brunswick by Messrs. Matthew and Hartt. In the progress of these researches, which developed an unexpectedly rich assemblage of species, the northern origin of this old flora seemed to be established by its earlier culmination in the northeast, in connection with the growth of the American land to the southward, which took place after the great Upper Silurian subsidence, by elevations beginning in the north while those portions of the continent to the southwest still remained under the sea. The same result was indicated by the persistence in the Carboniferous of the south and west of old Erian forms, like Megalopteris.
When, in 1870, the labours of those ten years were brought before the Royal Society of London, in the Bakerian lecture of that year, and in a memoir illustrating no less than one hundred and twenty-five species of plants older than the great Carboniferous system, these deductions were stated in connection with the conclusions of Hall, Logan, and Dana, as to the distribution of sediment along the northeast side of the American continent, and the anticipation was hazarded that the oldest Palæozoic floras would be discovered to the north of Newfoundland. Mention was also made of the apparent earlier and more copious birth of the Devonian flora in America than in Europe, a fact which is itself connected with the greater northward extension of this continent.
The memoir containing these results was not published by the Royal Society, but its publication was secured in a less complete form in the reports of the “Geological Survey of Canada.” The part of the memoir relating to Canadian fossil plants, with a portion of the theoretical deductions, was published in a report issued in 1871.[EG] In this report the following language was used:
[EG] “Fossil Plants of the Devonian and Upper Silurian Formations of Canada,” pp. 92, twenty plates, Montreal, 1871.
"In eastern America, from the Carboniferous period onward, the centre of plant distribution has been the Appalachian chain. From this the plants and sediments extended westward in times of elevation, and to this they receded in times of depression. But this centre was nonexistent before the Devonian period, and the centre for this must have been to the northeast, whence the great mass of older Appalachian sediment was derived. In the Carboniferous period there was also an eastward distribution from the Appalachians, and links of connection in the Atlantic bed between the floras of Europe and America. In the Devonian such connection can have been only far to the northeast. It is therefore in Newfoundland, Labrador, and Greenland that we are to look for the oldest American flora, and in like manner on the border of the old Scandinavian nucleus for that of Europe.
"Again, it must have been the wide extension of the sea of the corniferous limestone that gave the last blow to the remaining flora of the Lower Devonian; and the re-elevation in the middle of that epoch brought in the Appalachian ridges as a new centre, and established a connection with Europe which introduced the Upper Devonian and Carboniferous floras. Lastly, from the comparative richness of the later Erian[EH] flora in eastern America, especially in the St. John beds, it might be a « 241 » fair inference that the northeastern end of the Appalachian ridge was the original birthplace or centre of creation of what we may call the later Palæozoic flora, or of a large part of that flora."
[EH] See pages 107 and 108.
When my paper was written I had not seen the account published by the able Swiss palæobotanist Heer, of the remarkable Devonian flora of Bear Island, near Spitzbergen.[EI] From want of acquaintance with the older floras of America and western Europe, Heer fell into the unfortunate error of regarding the whole of Bear Island plants as Lower Carboniferous, a mistake which his great authority has tended to perpetuate, and which has even led to the still graver error of some European geologists, who do not hesitate to regard as Carboniferous the fossil plants of the American deposits from the Hamilton to the Chemung groups inclusive, though these belong to formations underlying the oldest Carboniferous, and characterised by animal remains of unquestioned Devonian age. In 1872 I addressed a note to the Geological Society of London on the subject of the so-called “Ursa stage” of Heer, showing that, though it contained some forms not known at so early a date in temperate Europe, it was clearly, in part at least, Devonian when tested by North American standards; but that in this high latitude, in which, for reasons stated in the report above referred to, I believed the Devonian plants to have originated, there might be an intermixture of the two floras. But such a mixed group should in that latitude be referred to a lower horizon than if found in temperate regions. Dr. Nathorst, as already stated, has recently obtained new facts which go to show that plants of two distinct horizons may have been intermixed in the collections submitted to Heer.
[EI] “Transactions of the Swedish Academy” 1871; “Journal of the London Geological Society,” vol. xxviii.
Between 1870 and 1873 my attention was turned to the two sub-floras intermediate between those of the Devonian and the coal-formation, the floras of the Lower Carboniferous (Subcarboniferous of some American geologists) and the Millstone Grit, and in a report upon these[EJ] similar deductions were expressed. It was stated that in Newfoundland the coal-beds seem to belong to the Millstone Grit series, and as we proceed southward they belong to progressively newer portions of the Carboniferous system. The same fact is observed in the coal-beds of Scotland, as compared with those of England, and it indicates that the coal-formation flora, like that of the Devonian, spread itself from the north, and this accords with the somewhat extensive occurrence of Lower Carboniferous rocks and fossils in the Parry Islands and elsewhere in the arctic regions.
[EJ] “Fossil Plants of Lower Carboniferous and Millstone Grit Formations of Canada,” pp. 47, ten plates, Montreal, 1873.
Passing over the comparatively poor flora of the earlier Mesozoic, consisting largely of cycads, pines, and ferns, and as yet little known in the arctic, and which may have originated in the south, though represented, according to Heer, by the supposed Jurassic flora of Siberia, we find, especially at Komé and Atané in Greenland, an interesting occurrence of those earliest precursors of the truly modern forms of plants which appear in the Cretaceous, the period of the English chalk and of the New Jersey greensands. There are two plant-groups of this age in Greenland; one, that of Komé, consists almost entirely of ferns, cycads, and pines, and is of decidedly Mesozoic aspect. This is called Lower Cretaceous. The other, that of Atané, holds remains of many modern temperate genera, as Populus, Myrica, Ficus, Sassafras, and Magnolia. This is regarded as Upper Cretaceous. Resting upon these Upper Cretaceous beds, without the intervention « 243 » of any other formation,[EK] are beds rich in plants of much more modern appearance, and referred by Heer to the Miocene period, a reference, as we have seen, not warranted by comparison with the Tertiary plants of Europe or of America. Still farther north this so-called Miocene assemblage of plants appears in Spitzbergen and Grinnell Land; but there, owing to the predominance of trees allied to the spruces, it has a decidedly more boreal character than in Greenland, as might be anticipated from its nearer approach to the pole.[EL]
[EK] Nordenskiöld, “Expedition to Greenland,” “Geological Magazine,” 1872.
[EL] Yet even here the bald cypress (Taxodium distichum), or a tree nearly allied to it, is found, though this species is now limited to the Southern States. Fielden and De Ranee, “Journal of the Geological Society,” 1878.
If now we turn to the Cretaceous and Tertiary floras of western America, as described by Lesquereux, Newberry, and others, we find in the lowest Cretaceous rocks there known—those of the Dakota group—which may be in the lower part of the Middle Cretaceous, a series of plants[EM] essentially similar to those of the so-called Upper Cretaceous of Greenland. They occur in beds indicating land and fresh-water conditions as prevalent at the time over great areas of the interior of America. But overlying this plant-bearing formation we have an oceanic limestone (the Niobrara), corresponding in many respects to the European chalk, and extending far north into the British territory,[EN] indicating that the land of the Lower Cretaceous was replaced by a vast Mediterranean Sea, filled with warm water from the equatorial currents, and not invaded by cold waters from the north. This is succeeded by thick Upper Cretaceous deposits of clay and sandstone, with marine remains, though very sparsely « 244 » distributed; and these show that further subsidence or denudation in the north had opened a way for the arctic currents, killing out the warm-water animals of the Niobrara group, and rilling up the Mediterranean of that period. Of the flora of these Upper Cretaceous periods, which must have been very long, we know something in the interior regions, from the discovery of a somewhat rich flora in the Dunvegan beds of the Peace River district, on the northern shore of the great Cretaceous Mediterranean;[EO] and on the coast of British Columbia we have the remarkable Cretaceous coal-field of Vancouver Island, which holds the remains of plants of modern genera, and, indeed, of almost as modern aspect as those of the so-called Miocene of Greenland. They indicate, however, a warmer climate as then prevalent on the Pacific coast, and in this respect correspond with a peculiar transition flora, intermediate between the Cretaceous and Eocene or earliest Tertiary of the interior regions, and which is described by Lesquereux as the Lower Lignitic.
[EM] Lesquereux, “Report on Cretaceous Flora.”
[EN] G. M. Dawson, “Report on Forty-ninth Parallel.”
[EO] “Reports of Dr. G. M. Dawson, Geological Survey of Canada.” Also, “Transactions of the Royal Society of Canada,” vol. i.
Immediately above these Upper Cretaceous beds we have the great Lignite Tertiary of the West—the Laramie group of recent American reports—abounding in fossil plants, at one time regarded as Miocene, but now known to be Lower Eocene, though farther south extending upward toward the Miocene age.[EP] These beds, with their characteristic plants, have been traced into the British territory north of the forty-ninth parallel, and it has been shown that their fossils are identical with those of the « 245 » McKenzie River valley, described by Heer as Miocene, and probably also with those of Alaska, referred to the same age.[EQ] Now this truly Eocene flora of the temperate and northern parts of America has so many species in common with that called Miocene in Greenland that its identity can scarcely be doubted. These facts have led to scepticism as to the Miocene age of the upper plant-bearing beds of Greenland, and more especially Mr. J. Starkie Gardner has ably argued, from comparison with the Eocene flora of England and other considerations, that they are really of that earlier date.[ER]
[EP] Lesquereux’s “Tertiary Flora”; “White on the Laramie Group”; Stevenson, “Geological Relations of Lignitic Groups,” American Philosophical Society, June, 1875; Dawson, “Transactions of the Royal Society of Canada,” vol. iv.; Ward, “Bulletin of United States Geological Survey.”
[EQ] G. M. Dawson, “Report on the Geology of the Forty-ninth Parallel,” where full details on these points may be found. “Transactions of the Royal Society of Canada,” vol. iv.
[ER] “Nature,” December 12, 1878.
In looking at this question, we may fairly assume that no climate, however equable, could permit the vegetation of the neighbourhood of Disco in Greenland to be exactly identical with that of Colorado and Missouri, at a time when little difference of level existed in the two regions. Either the southern flora migrated north in consequence of a greater amelioration of climate, or the northern flora moved southward as the climate became colder. The same argument, as Gardner has ably shown, applies to the similarity of the Tertiary plants of temperate Europe to those of Greenland. If Greenland required a temperature of about 50°, as Heer calculates, to maintain its Eocene flora, the temperature of England and that of the Southwestern States must have been higher, though probably more equable, than at present.
We cannot certainly affirm anything respecting the migrations of these floras, but there are some probabilities which deserve attention. The ferns and cycads of the so-called Lower Cretaceous of Greenland are nothing but a continuation of the previous Jurassic flora. Now this was established at an equally early date in the Queen « 246 » Charlotte Islands,[ES] and still earlier in Virginia,[ET] The presumption is, therefore, that it came from the south. It has, indeed, the facies of a southern hemisphere and insular flora, and probably spread itself northward as far as Greenland, at a time when our northern continents were groups of islands, and when the ocean currents were carrying warm water far toward the arctic regions. The flora which succeeds this in the sections at Atané has no special affinities with the southern hemisphere, and is of a more temperate and continental character.[EU] It is not necessarily Upper Cretaceous, since it is similar to that of the Dakota group farther south, and this is at least Middle Cretaceous. This flora must have originated either somewhere in temperate America or within the Arctic circle, and it must have replaced the older one by virtue of increasing coolness and continental character of climate. It must, therefore, have been connected with that elevation of the land which took place at the beginning of the Cretaceous. During this elevation it spread over all western America at one time or another, and, as the land again subsided under the sea of the Niobrara chalk, it assumed an aspect more suited to a warm climate, but still held its place on such islands as remained above water along the Pacific coast and in the north, and it continued to exist on these islands till the colder seas « 247 » of the Upper Cretaceous had again given place to the warm plains and land-locked brackish seas or fresh-water lakes of the Laramie period (Eocene). Thus the true Upper Cretaceous marks a cool period intervening between the so-called Upper Cretaceous (really Middle Cretaceous) and the so-called Miocene (really Lower Eocene) floras of Greenland.
[ES] “Reports of the Geological Survey of Canada.”
[ET] Fontaine has well described the Mesozoic flora of Virginia, “American Journal of Science,” January, 1879, and “Report on Early Mesozoic Floras.”
[EU] In the “Proceedings of the Royal Society of Tasmania,” 1887, Mr. R. M. Johnston, F. L. S., states that in the Miocene beds of Tasmania trees of European genera abound. The Mesozoic flora of that island is of the usual conifero-cycadean type. Ettingshausen makes a similar statement in the “Geological Magazine” respecting the Tertiary flora of Australia and New Zealand, stating that, like the Tertiary floras of Europe, they have a mixed character, being partly of types now belonging to the northern hemisphere.
This latter established itself in Greenland, and probably all around the Arctic circle, in the warm period of the earliest Eocene, and, as the climate of the northern hemisphere became gradually reduced from that time till the end of the Pliocene, it marched on over both continents to the southward, chased behind by the modern arctic flora, and eventually by the frost and snow of the Glacial age. This history may admit of correction in details; but, so far as present knowledge extends, it is in the main not far from the truth.
Perhaps the first great question which it raises is that as to the causes of the alternations of warm and cold climates in the north, apparently demanded by the vicissitudes of the vegetable kingdom. Here we may set aside the idea that in former times plants were suited to endure greater cold than at present. It is true that some of the fossil Greenland plants are of unknown genera, and many are species new to us; but we are on the whole safe in affirming that they must have required conditions similar to those necessary to their modern representatives, except within such limits as we now find to hold in similar cases among existing plants. Still we know that at the present time many species found in the equable climate of England will not live in Canada, though species to all appearance similar in structure are native here. There is also some reason to suppose that species when new may have greater hardiness and adaptability than when in old age and verging toward extinction. In any case these facts can account for but a small part of the phenomena, which « 248 » require to be explained by physical changes affecting the earth as a whole, or at least the northern hemisphere. Many theoretical views have been suggested on this subject, and perhaps the most practical way of disposing of these will be first to set aside a number which are either precluded by the known facts, incapable of producing the effects, or altogether uncertain as to their possible occurrence.
1. In this class we may place the theory that the poles of the earth have changed their position. Independently of astronomical objections, there is good geological evidence that the poles of the earth must have been nearly in their present places from the dawn of life until now. From the Laurentian upward, those organic limestones which mark the areas where warm and shallow equatorial water was spreading over submerged continents are so disposed as to prove the permanence of the poles. In like manner all the great foldings of the crust of the earth have followed lines which are parts of great circles tangent to the existing polar circles. So, also, from the Cambrian age the great drift of sediment from the north has followed the line of the existing Arctic currents from the northeast to the southwest, throwing itself, for example, along the line of the Appalachian uplifts in eastern America, and against the ridge of the Cordilleras in the west.
2. Some of the above considerations, along with astronomical evidence, prevent us from assuming any considerable change in the obliquity of the axis of the earth during geological time.
3. That the earth and the sun have diminished in heat during geological time seems probable; but physical and geological facts alike render it certain that this influence could have produced no appreciable effect, even in the times of the earliest floras, and certainly not in the case of Tertiary vegetation.
4. It has been supposed that the earth may have at different times traversed more or less heated zones of space, giving alternations of warm and cold temperature. No such differences in space are, however, known, nor does there seem any good ground for imagining their existence.
5. The heat of the sun is known to be variable, and the eleven years' period of sun-spots has recently attracted much attention as producing appreciable effects on the seasons. There may possibly be longer cycles of solar energy, or the sun may be liable, like some variable stars, to paroxysms of increased energy. Such changes are possible, and may fairly be taken into the account, provided that we fail to find known causes sufficient to account for the phenomena.
Of well-known causes there seem to be but three. These are: First, that urged by Lyell—viz., the varying distribution of land and water along with that of marine currents; secondly, the varying eccentricity of the earth’s orbit, along with the precession of the equinoxes, and the effects of this on oceanic circulation, as illustrated by Croll; thirdly, the different conditions of the earth’s atmosphere with reference to radiation, as argued by Tyndall and Hunt. As these causes are all founded on known facts, and not exclusive of each other, we may consider them together. I shall take the Lyellian theory first, regarding it as the most important, and the best supported by geological facts.
We know that the present distribution of land and water greatly influences climate, more especially by affecting that of the ocean currents and of the winds, and by the different action of land as compared with water in the reception and radiation of heat. The present distribution of land gives a large predominance to the arctic and sub-arctic regions, as compared with the equatorial and with the antarctic; and we might readily imagine « 250 » other distributions that would give very different results. But this is not an imaginary case. We know that, while the forms and positions of the great continents have been fixed from a very early date, they have experienced many great submergences and re-elevations, and that these have occurred in somewhat regular sequence, as evidenced by the cyclical alternations of organic limestones and earthy sediments in successive geological formations.
An example bearing on our present subject may serve to illustrate this. In the latter part of the Upper Silurian period (the Lower Helderberg age), vast areas of the American continent[EV] were covered with an ocean in which were deposited organic limestones whose fossils show that this great interior sea was pervaded by equatorial waters bringing food and warmth, while the incipient ranges of the Appalachians on the east, and the Cordilleras on the west, and the Laurentian axis on the north, fenced off from it the colder arctic waters. How different must the climate of America and of the region north of it have been in these circumstances from that which prevails at present, or from that which prevailed in certain other periods, when it was open to the incursions of the arctic ice-laden currents, bearing loads of fine sediment![EW] It was in these circumstances, and in the similar circumstances in which the great Corniferous limestone of the Devonian was deposited—a limestone showing in its rich coral fauna even warmer waters than those of the Lower Helderberg—that the Devonian flora « 251 » took its origin in the north and advanced southward over new lands in process of emergence from the sea. The somewhat similar condition evidenced by the Lower Carboniferous limestone preceded the advent of the great and rich flora of the coal-formation.
[EV] See a memoir and map by Prof. Hall, “Reports of the Regents of New York,” 1874-'75.
[EW] It seems certain that the faunæ of the old limestones, like the Trenton, Niagara, Lower Helderberg, and Corniferous, belong to warm and sheltered sea areas, and that those rich in graptolites and trilobites, enclosed in muddy sediments, belong to the colder arctic waters. Such arctic faunæ are those of the Quebec group and of the Utica shale, and to some extent that of the Hamilton group.
Lyell’s theory on this subject has, I think, in some recent publications, been somewhat misapprehended. It is true that he stated hypothetically two contrasted conditions of distribution, in one of which all the land was equatorial, in another all polar; but he did not suppose that these conditions had actually occurred; and even in his earlier editions, before the recent discoveries and discussions as to ocean currents, he was always careful to attach due value to these in connection with subsidences and elevations.[EX] In his later editions he introduced more full references to current action, and also stated Croll’s theory, but still maintained the validity of his original conclusions.
[EX] See “Principles of Geology,” edition of 1840, chapter vii.
The sufficiency of this Lyellian theory to account for the facts, in so far as plants are concerned, may, I think, be inferred from the course of the isothermal lines at present. The south end of Greenland is on the latitude of Christiania in Norway on the one hand, and of Fort Liard in the Peace River region on the other; and while Greenland is clad in ice and snow, wheat and other grains, and the ordinary trees of temperate climates, grow at the latter places,[EY] It is evident, therefore, that only exceptionally unfavourable circumstances prevent the Greenland area from still possessing a temperate flora, and these unfavourable circumstances possibly tell even on the localities with which we have compared it. Further, the mouth of the McKenzie River is in the same latitude with « 252 » Disco, near which are some of the most celebrated localities of fossil Cretaceous and Tertiary plants. Yet the mouth of the McKenzie River enjoys a much more favourable climate and has a much more abundant flora than Disco. If north Greenland were submerged, and low land reaching to the south terminated at Disco, and if from any cause either the cold currents of Baffin’s Bay were arrested, or additional warm water thrown into the North Atlantic by the Gulf Stream, there is nothing to prevent a mean temperature of 45° Fahr. from prevailing at Disco; and the estimate ordinarily formed of the requirements of its extinct floras is 50°,[EZ] which is probably above rather than below the actual temperature required.
[EY] See “Macoun’s Report,” “Geological Survey of Canada,” and Richardson’s “Boat Voyage.”
[EZ] Heer. See, also, papers by Prof. Haughton and by Gardner in “Nature” for 1878.
Since, then, geological facts assure us of mutations of the continents much greater than those apparently required to account for the changes of climate implied in the existence of the ancient arctic floras, it does not seem absolutely necessary to invoke any others.[FA] If, however, there are other true causes which might either aid or counteract those above referred to, it may be well to consider them.
[FA] Sir William Thomson, “Transactions of the Geological Society of Glasgow,” February 22, 1878.
Mr. Croll has, in his valuable work “Climate and Time” and in various memoirs, brought forward an ingenious astronomical theory to account for changes of climate. This theory, as stated by himself in a recent paper,[FB] is that when the eccentricity of the earth’s orbit is at a high value, and the northern winter solstice is in perihelion, agencies are brought into operation which make the southeast trade-winds stronger than the northeast, and compel them to blow over upon the northern « 253 » hemisphere as far as the Tropic of Cancer. The result is that all the great equatorial currents of the ocean are impelled into the northern hemisphere, which thus, in consequence of the immense accumulation of warm water, has its temperature raised, so that ice and snow must to a great extent disappear from the arctic regions. In the prevalence of the converse conditions, the arctic zone becomes clad in ice, and the southern has its temperature raised.
[FB] “Cataclysmic Theories of Geological Climate,” “Geological Magazine,” May, 1878.
At the same time, according to Croll’s calculations, the accumulation of ice on either pole would tend, by shifting the earth’s centre of gravity, to raise the level of the ocean and submerge the land on the colder hemisphere. Thus a submergence of land would coincide with a cold condition, and emergence with increasing warmth. Facts already referred to, however, show that this has not always been the case, but that in many cases submergence was accompanied with the influx of warm equatorial waters and a raised temperature, this apparently depending on the question of local distribution of land and water; and this in its turn being regulated not always by mere shifting of the centre of gravity, but by foldings occasioned by contraction, by equatorial subsidences resulting from the retardation of the earth’s rotation, and by the excess of material abstracted by ice and frost from the arctic regions, and drifted southward along the lines of arctic currents. This drifting must in all geological times have greatly exceeded, as it certainly does at present, the denudation caused by atmospheric action at the equator, and must have tended to increase the disposition to equatorial collapse occasioned by retardation of rotation.[FC]
[FC] Croll, in “Climate and Time,” and in a note read before the British Association in 1876, takes an opposite view; but this is clearly contrary to the facts of sedimentation, which show a steady movement of débris toward the south and southwest.
While such considerations as those above referred to « 254 » tend to reduce the practical importance of Mr. Croll’s theory., on the other hand they tend to remove one of the greatest objections against it—namely, that founded on the necessity of supposing that glacial periods recur with astronomical regularity in geological time. They cannot do so if dependent on other causes inherent in the earth itself, and producing important movements of its crust.
The third great cause of warmer climates in the past is the larger proportion of carbon dioxide, or carbonic-acid gas, in the atmosphere in early geological times, as proved by the immense amount of carbon now sealed up in limestone and coal, and which must at one time have been in the air. It has been shown that a very small additional quantity of this substance would so obstruct radiation of heat from the earth as to act almost like a glass roof. If, however, the quantity of carbonic acid, great at first, was slowly and regularly removed, even if, as suggested by Hunt, small additional supplies were gradually added from space, this cause could have affected only the very oldest floras. But it is known that some comets and meteorites contain carbonaceous matter, and this allows us to suppose that accessions of carbon may have been communicated at irregular intervals. If so, there may have been cycles of greater and less abundance of this substance, and an atmosphere rich in carbon dioxide might at one and the same time afford warmth and abundance of food to plants.
It thus appears that the causes of ancient vicissitudes of climate are somewhat complex, and when two or more of them happened to coincide very extreme changes might result, having most important bearings on the distribution of plants.
This may help us to deal with the peculiarities of the great Glacial age, which may have been rendered exceptionally severe by the combination of several of the causes of refrigeration. We must not suppose, however, that « 255 » the views of those extreme glacialists who suppose continental ice-caps reaching half way to the equator are borne out by facts. In truth, the ice accumulating round the pole must have been surrounded by water, and there must have been tree-clad islands in the midst of the icy seas, even in the time of greatest refrigeration. This is proved by the fact that, in the Leda clay of eastern Canada, which belongs to the time of greatest submergence, and whose fossil shells show sea-water almost at the freezing-point, there are leaves of poplars and other plants which must have been drifted from neighbouring shores. Similar remains occur in clays of like origin in the basin of the great lakes and in the West. These have been called “interglacial,” but there is no evidence to prove that they are not truly glacial. Thus, while we need not suppose that plants existed within the Arctic circle in the Glacial age, we have evidence that those of the cold temperate and sub-arctic zones continued to exist pretty far north. At the same time the warm temperate flora would be driven to the south, except where sustained in insular spots warmed by the equatorial currents. It would return northward on the re-elevation of the land and the renewal of warmth.
If, however, our modern flora is thus one that has returned from the south, this would account for its poverty in species as compared with those of the early Tertiary. Groups of plants descending from the north have been rich and varied. Returning from the south they are like the shattered remains of a beaten army. This, at least, has been the case with such retreating floras as those of the Lower Carboniferous, the Permian, and the Jurassic, and possibly that of the Lower Eocene of Europe.
The question of the supply of light to an arctic flora is much less difficult than some have imagined. The long summer day is in this respect a good substitute for a longer season of growth, while a copious covering of « 256 » winter snow not only protects evergreen plants from those sudden alternations of temperature which are more destructive than intense frost, and prevents the frost from penetrating to their roots, but, by the ammonia which it absorbs, preserves their greenness. According to Dr. Brown, the Danish ladies of Disco long ago solved this problem.[FD] He informs us that they cultivate in their houses most of our garden flowers—as roses, fuchsias, and geraniums—showing that it is merely warmth and not light that is required to enable a sub-tropical flora to thrive in Greenland. Even in Canada, which has a flora richer in some respects than that of temperate Europe, growth is effectually arrested by cold for nearly six months, and though there is ample sunlight there is no vegetation. It is, indeed, not impossible that in the plans of the Creator the continuous summer sun of the arctic regions may have been made the means for the introduction, or at least for the rapid growth and multiplication, of new and more varied types of plants.
[FD] “Florula Discoana,” Botanical Society of Edinburgh, 1868.
Much, of course, remains to be known of the history of the old floras, whose fortunes I have endeavoured to sketch, and which seem to have been driven like shuttle-cocks from north to south, and from south to north, especially on the American continent, whose meridional extension seems to have given a field specially suited for such operations.
This great stretch of the western continent, from north to south, is also connected with the interesting fact that, when new floras are entering from the arctic regions, they appear earlier in America than in Europe, and that in times when old floras are retreating from the south old genera and species linger longer in America. Thus, in the Devonian and Cretaceous new forms of those periods appear in America long before they are recognized « 257 » in Europe, and in the modern epoch forms that would be regarded in Europe as Miocene still exist. Much confusion in reasoning as to the geological ages of the fossil floras has arisen from want of attention to this circumstance.
What we have learned respecting this wonderful history has served strangely to change some of our preconceived ideas. We must now be prepared to admit that an Eden can be planted even in Spitzbergen, that there are possibilities in this old earth of ours which its present condition does not reveal to us; that the present state of the world is by no means the best possible in relation to climate and vegetation; that there have been and might be again conditions which could convert the ice-clad arctic regions into blooming paradises, and which at the same time would moderate the fervent heat of the tropics. We are accustomed to say that nothing is impossible with God; but how little have we known of the gigantic possibilities which lie hidden under some of the most common of his natural laws!
These facts have naturally been made the occasion of speculations as to the spontaneous development of plants by processes of varietal derivation. It would, from this point of view, be a nice question to calculate how many revolutions of climate would suffice to evolve the first land-plant; what are the chances that such plant would be so dealt with by physical changes as to be preserved and nursed into a meagre flora like that of the Upper Silurian or the Jurassic; how many transportations to Greenland would suffice to promote such meagre flora into the rich and abundant forests of the Upper Cretaceous, and to people the earth with the exuberant vegetation of the early Tertiary. Such problems we may never be able to solve. Probably they admit of no solution, unless we invoke the action of an Almighty mind, operating through long ages, and correlating with boundless power and wisdom all the energies inherent in inorganic and organic « 258 » nature. Even then we shall perhaps be able to comprehend only the means by which, after specific types have been created, they may, by the culture of their Maker, be “sported” into new varieties or subspecies, and thus fitted to exist under different conditions or to occupy higher places in the economy of nature.
Before venturing on such extreme speculations as some now current on questions of this kind, we would require to know the successive extinct floras as perfectly as those of the modern world, and to be able to ascertain to what extent each species can change either spontaneously or under the influence of struggle for existence or expansion under favourable conditions, and under arctic semi-annual days and nights, or the shorter days of the tropics. Such knowledge, if ever acquired, it may take ages of investigation to accumulate.
As to the origin and mode of introduction of successive floras, I am, for the reasons above stated, not disposed to dogmatise, or to adopt as final any existing theory of the development of the vegetable kingdom. Still, some laws regulating the progress of vegetable life may be recognised, and I propose to state these in connection with the Palæozoic floras, to which my own studies have chiefly related.
Fossil plants are almost proverbially uncertain with reference to their accurate determination, and have been regarded as of comparatively little utility in the decision of general questions of palæontology. This results principally from the fragmentary condition in which they have been studied, and from the fact that fragments of animal structures are more definite and instructive than corresponding portions of plants.
It is to be observed, however, that our knowledge of fossil plants becomes accurate in proportion to the extent to which we can carry the study of specimens in the beds in which they are preserved, so as to examine more perfect « 259 » examples than those usually to be found in museums. When structures are taken into the account, as well as external forms, we can also depend more confidently on our results. Further, the abundance of specimens to be obtained in particular beds often goes far to make up for their individual imperfection. The writer of these pages has been enabled to avail himself very fully of these advantages; and on this account, if on no other, feels entitled to speak with some authority on theoretical questions.
It is an additional encouragement to pursue the subject, that, when we can obtain definite information as to the successive floras of any region, we thereby learn much as to climate and vicissitudes in regard to the extent of land and water; and that, with reference to such points, the evidence of fossil plants, when properly studied, is, from the close relation of plants to those stations and climates, even more valuable than that of animal fossils.
It is necessary, however, that in pursuing such inquiries we should have some definite views as to the nature and permanence of specific forms, whether with reference to a single geological period or to successive periods; and I may be excused for stating here some general principles, which I think important for our guidance.
1. Botanists proceed on the assumption, vindicated by experience, that, within the period of human observation, species have not materially varied or passed into each other. We may make, for practical purposes, the same assumption with regard to any given geological period, and may hold that for each such period there are specific types which, for the time at least, are invariable.
2. When we inquire what constitutes a good species for any given period, we have reason to believe that many names in our lists represent merely varietal forms or erroneous determinations. This is the case even in the modern flora; and in fossil floras, through the poverty of specimens, their fragmentary condition, and various states « 260 » of preservation, it is still more likely to occur. Every revision of any group of fossils detects numerous synonyms, and of these many are incapable of detection without the comparison of large suites of specimens.
3. We may select from the flora of any geological period certain forms, which I shall call specific types, which may for such period be regarded as unchanging. Having settled such types, we may compare them with similar forms in other periods, and such comparisons will not be vitiated by the uncertainty which arises from the comparison of so-called species which may, in many cases, be mere varietal forms, as distinguished from specific types. Our types may be founded on mere fragments, provided that these are of such a nature as to prove that they belong to distinct forms which cannot pass into each other, at least within the limits of one geological period.
4. When we compare the specific types of one period with those of another immediately precedent or subsequent, we shall find that some continue unchanged through long intervals of geological time, that others are represented by allied forms regarded either as varietal or specific, and as derived or otherwise, according to the view which we may entertain as to the permanence of species. On the other hand, we also find new types not rationally deducible on any theory of derivation from those known in other periods. Further, in comparing the types of a poor period with those of one rich in species, we may account for the appearance of new types in the latter by the deficiency of information as to the former; where many new types appear in the poorer period this conclusion seems less probable. For example, new types appearing in poor formations, like the Lower Erian and Lower Carboniferous, have greater significance than if they appeared in the Middle Erian or in the Coal Measures.
5. When specific types disappear without any known successors, under circumstances in which it seems unlikely « 261 » that we should have failed to discover their continuance, we may fairly assume that they have become extinct, at least locally; and where the field of observation is very extensive, as in the great coal-fields of Europe and America, we may esteem such extinction as practically general, at least for the northern hemisphere. When many specific types become extinct together, or in close succession, we may suppose that such extinction resulted from physical changes; but where single types disappear, under circumstances in which others of similar habit continue, we may not unreasonably conjecture that, as Pictet has argued in the case of animals, such types may have been in their own nature limited in duration, and may have died out without any external cause.
6. With regard to the introduction of specific types we have not as yet a sufficient amount of information. Even if we freely admit that ordinary specific forms, as well as mere varieties, may result from derivation, this by no means excludes the idea of primitive specific types originating in some other way. Just as the chemist, after analysing all compounds and ascertaining all allotropic forms, arrives at length at certain elements not mutually transmutable or derivable, so the botanist and zoölogist must expect sooner or later to arrive at elementary specific types, which, if to be accounted for at all, must be explained on some principle distinct from that of derivation. The position of many modern biologists, in presence of this question, may be logically the same with that of the ancient alchemists with reference to the chemical elements, though the fallacy in the case of fossils may be of more difficult detection. Our business at present, in the prosecution of palæobotany, is to discover, if possible, what are elementary or original types, and, having found these, to enquire as to the law of their creation.
7. In prosecuting such questions geographical relations must be carefully considered. When the floras of « 262 » two successive periods have existed in the same region, and under circumstances that render it probable that plants have continued to grow on the same or adjoining areas throughout these periods, the comparison becomes direct, and this is the case with the Erian and Carboniferous floras in northeastern America. But, when the areas of the two formations are widely separated in space as well as in time, any resemblances of facies that we may observe may have no connection whatever with an unbroken continuity of specific types.
I desire, however, under this head, to affirm my conviction that, with reference to the Erian and Carboniferous floras of North America and of Europe, the doctrine of “homotaxis,” as distinct from actual contemporaneity, has no place. The succession of formations in the Palæozoic period evidences a similar series of physical phenomena on the grandest scale throughout the northern hemisphere. The succession of marine animals implies the continuity of the sea-bottoms on which they lived. The headquarters of the Erian flora in America and Europe must have been in connected or adjoining areas in the North Atlantic. The similarity of the Carboniferous flora on the two sides of the Atlantic, and the great number of identical species, proves a still closer connection in that period. These coincidences are too extensive and too frequently repeated to be the result of any accident of similar sequence at different times, and this more especially as they extend to the more minute differences in the features of each period, as, for instance, the floras of the Lower and Upper Devonian, and of the Lower, Middle, and Upper Carboniferous.
8. Another geographical question is that which relates to centres of dispersion. In times of slow subsidence of extensive areas, the plants inhabiting such areas must be narrowed in their range and often separated from one another in detached spots, while, at the same time, important « 263 » climatal changes must also occur. On the re-emergence of the land such of these species as remained would again extend themselves over their former areas of distribution, in so far as the new climatal and other conditions would permit. We would naturally suppose that the first of the above processes would tend to the elimination of varieties, the second, to their increase; but, on the other I hand, the breaking up of a continental flora into that of distinct islets, and the crowding together of many forms, might be a process fertile in the production of some varieties if fatal to others.
Further, it is possible that these changes of subsidence may have some connection with the introduction, as well as with the extinction, even of specific types. It is certain, at least, in the case of land-plants, that such types come in most plentifully immediately after elevation, though they are most abundantly preserved in periods of slow subsidence. I do not mean, however, that this connection is one of cause and effect; there are, indeed, indications that it is not so. One of these is, that in some cases the enlargement of the area of the land seems to be as injurious to terrestrial species as its diminution.
9. Another point on which I have already insisted, and which has been found to apply to the Tertiary as well as to the Palæozoic floras, is the appearance of new types within the arctic and boreal areas, and their migration southward. Periods in which the existence of northern land coincided with a general warm temperature of the northern hemisphere seem to have been those most favourable to the introduction of new forms of land-plants. Hence, there has been throughout geological time a general movement of new floras from the Palæarctic and Nearctic regions to the southward.
Applying the above considerations to the Erian and Carboniferous floras of North America, we obtain some data which may guide us in arriving at general conclusions. « 264 » The Erian flora is comparatively poor, and its types are in the main similar to those of the Carboniferous. Of these types a few only reappear in the middle coal-formation under identical forms; a great number appear under allied forms; some altogether disappear. The Erian flora of New Brunswick and Maine occurs side by side with the Carboniferous of the same region; so does the Erian of New York and Pennsylvania with the Carboniferous of those States. Thus we have data for the comparison of successive floras in the same region. In the Canadian region we have, indeed, in direct sequence, the floras of the Upper Silurian, the Lower, Middle, and Upper Erian, and the Lower, Middle, and Upper Carboniferous, all more or less distinct from each other, and affording an admirable series for comparison in a region whose geographical features are very broadly marked. All these floras are composed in great part of similar types, and probably do not indicate very dissimilar general physical conditions, but they are separated from each other by the great subsidences of the Corniferous limestone and the Lower Carboniferous limestone, and by the local but intense subterranean action which has altered and disturbed the Erian beds toward the close of that period. Still, these changes were not universal. The Corniferous limestone is absent in Gaspé, and probably in New Brunswick, where, consequently, the Erian flora could continue undisturbed during that long period. The Carboniferous limestone is absent from the slopes of the Appalachians in Pennsylvania, where a retreat may have been afforded to the Upper Erian and Lower Carboniferous floras. The disturbances at the close of the Erian were limited to those eastern regions where the great limestone-producing subsidences were unfelt, and, on the other hand, are absent in Ohio, where the subsidences and marine conditions were almost at a maximum.
Bearing in mind these peculiarities of the area in question, we may now group in a tabular form the distinct specific types recognised in the Erian system, indicating, at the same time, those which are represented by identical species in the Carboniferous, those represented by similar species of the same general type, and those not represented at all. For example, Calamites cannæformis extends as a species into the Carboniferous; Asterophyllites latifolia does not so extend, but is represented by closely allied species of the same type; Nematophyton disappears altogether before we reach the Carboniferous.
Table of Erian and Carboniferous Specific Types.
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Of the above forms, fifty-one in all, found in the Erian of eastern America, all, except the last four, are certainly distinct specific types. Of these only four reappear in the Carboniferous under identical species, but no less than twenty-six reappear under representative or allied forms, some at least of which a derivationist might claim as modified descendants. On the other hand, nearly one half of the Devonian types are unknown in the Carboniferous, while there remain a very large number of Carboniferous types not accounted for by anything known in the Devonian. Further, a very poor flora, including only two or three types, is the predecessor of the Erian flora in the Upper Silurian, and the flora again becomes poor in the Upper Devonian and Lower Carboniferous. Every new species discovered must more or less modify the above statements, and the whole Erian flora of America, as well as the Carboniferous, requires a thorough comparison with that of Europe before general conclusions can be safely drawn. In the mean time I may indicate the direction in which the facts seem to point by the following general statements:
1. Some of the forms reckoned as specific in the Devonian and Carboniferous may be really derivative races. There are indications that such races may have originated in one or more of the following ways: (1) By a natural tendency in synthetic types to become specialised in the direction of one or other of their constituent elements. In this way such plants as Arthrostigma and Psilophyton may have assumed new varietal forms. (2) By embryonic retardation or acceleration,[FE] whereby certain species may have had their maturity advanced or postponed, thus giving them various grades of perfection in reproduction and complexity of structure. The fact that so many Erian and Carboniferous plants seem to be on the confines « 267 » of the groups of Acrogens and Gymnosperms may be supposed favourable to such exchanges. (3) The contraction and breaking up of floras, as occurred in the Middle Erian and Lower Carboniferous, may have been eminently favourable to the production of such varietal forms as would result from what has been called the “struggle for existence.” (4) The elevation of a great expanse of new land at the close of the Middle Erian and the beginning of the coal period would, by permitting the extension of species over wide areas and fertile soils, and by removing the pressure previously existing, be eminently favourable to the production of new, and especially of improved, varieties.
[FE] In the manner illustrated by Hyatt and Cope.
2. Whatever importance we may attach to the above supposed causes of change, we still require to account for the origin of our specific types. This may forever elude our observation, but we may at least hope to ascertain the external conditions favourable to their production. In order to attain even to this it will be necessary to inquire critically, with reference to every acknowledged species, what its claims to distinctness are, so that we may be enabled to distinguish specific types from mere varieties. Having attained to some certainty in this, we may be prepared to inquire whether the conditions favourable to the appearance of new varieties were also those favourable to the creation of new types, or the reverse—whether these conditions were those of compression or expansion, or to what extent the appearance of new types may be independent of any external conditions, other than those absolutely necessary for their existence. I am not without hope that the further study of fossil plants may enable us thus to approach to a comprehension of the laws of the creation, as distinguished from those of the continued existence of species.
3. In the present state of our knowledge we have no good ground either to limit the number of specific types « 268 » beyond what a fair study of our material may warrant, or to infer that such primitive types must necessarily have been of low grade, or that progress in varietal forms has always been upward. The occurrence of such an advanced and specialised type as that of Dadoxylon in the Middle Devonian should guard us against these errors. The creative process may have been applicable to the highest as well as to the lowest forms, and subsequent deviations must have included degradation as well as elevation. I can conceive nothing more unreasonable than the statement sometimes made that it is illogical or even absurd to suppose that highly organised beings could have been produced except by derivation from previously existing organisms. This is begging the whole question at issue, depriving science of a noble department of inquiry on which it has as yet barely entered, and anticipating by unwarranted assertions conclusions which may perhaps suddenly dawn upon us through the inspiration of some great intellect, or may for generations to come baffle the united exertions of all the earnest promoters of natural science. Our present attitude should not be that of dogmatists, but that of patient workers content to labour for a harvest of grand generalisations which may not come till we have passed away, but which, if we are earnest and true to Nature and its Creator, may reward even some of us.
Within the human period great changes of distribution of plants have occurred, chiefly through the agency of man himself, and we have had ample evidence that plants are able to establish themselves and prosper in climates and conditions to which unaided they could not have transported themselves, as, for instance, in the case of European weeds naturalised in Australia and New Zealand. There is, however, no reason to believe that any specific change has occurred to any plant within the Pleistocene or modern period.
In a recent address, delivered to the biological section of the British Association, Mr. Carruthers has discussed this question, and has shown that the earliest vegetable specimens described by Dr. Schweinfurth from the Egyptian tombs present no appearance of change. This fact appears also in the leaves and other organs of plants preserved in the nodules in the Pleistocene clays of the Ottawa, and in specimens of similar age found in various places in Britain and the continent of Europe.[FF]
[FF] “Proceedings British Association,” 1886, “Pleistocene Plants of Canada,” Canadian Naturalist, 1866.
The difficulties attending the ordinary theories of evolution as applied to plants have been well set forth by the same able botanist in his “Presidential Address to the Geological Association in 1877,” a paper which deserves careful study. One of his illustrations is that ancient willow, Salix polaris, referred to in a previous chapter, which now lives in the arctic regions, and is found fossil in the Pleistocene beds at Cromer and at Bovey Tracey.
He notes the fact that the genus Salix is a very variable one, including 19 subgeneric groups and 160 species, with no less than 222 varieties and 70 hybrids. Salix polaris belongs to a subgeneric group containing 29 species, which are arranged in four sections, that to which S. polaris belongs containing six species. Now it is easy to construct a theoretical phylogeny of the derivation of the willows from a supposed ancestral source, but when we take our little S. polaris we find that this one twig of our ancestral tree takes us back without change to the Glacial period. The six species would take us still farther, and the sections, sub-genera, and genus at the same rate would require an incalculable amount of past time. He concludes the inquiry in the following terms:
"But when we have reached the branch representing the generic form we have made but little progress in the phylogenesis of Salix. With Populus this genus forms a small order, Salicineæ, The two genera are closely allied, yet separated by well-marked characters; it is not, however, difficult to conceive of both having sprung from a generalised form. But there is no record of such a form. The two genera appear together among the earliest known dicotyledons, the willows being represented by six and the poplars by nine species. The ordinal form, if it ever existed, must necessarily be much older than the period of the Upper Cretaceous rocks, that is, than the period to which the earliest known dicotyledons belong.
“The Salicineæ are related to five other natural orders, in all of which the apetalous flowers are arranged in catkins. These different though allied orders must be led up by small modifications to a generalised amentiferous type, and thereafter the various groups of apetalous plants by innumerable eliminations of differentiating characters until the primitive form of the apetalous plant is reached. Beyond this the uncurbed imagination will have more active work in bridging over the gap between Angiosperms and Gymnosperms, in finding the intermediate forms that led up to the vascular cryptogams, and on through the cellular plants to the primordial germ. Every step in this phylogenetic tree must be imagined. The earliest dicotyledon takes us not a step farther back in the phylogenetic history of Salix than that supplied by existing vegetation. All beyond the testimony of our living willows is pure imagination, unsupported by a single fact. So that here, also, the evidence is against evolution, and there is none in favour of it.”
It is easy to see that similar difficulties beset every attempt to trace the development of plants on the principle of slow and gradual evolution, and we are driven « 271 » back on the theory of periods of rapid origin, as we have already seen suggested by Saporta in the case of the Cretaceous dicotyledons. Such abrupt and plentiful introduction of species over large areas at the same time, by whatever cause effected—and we are at present quite ignorant of any secondary causes—becomes in effect something not unlike the old and familiar idea of creation. Science must indeed always be baffled by questions of ultimate origin, and, however far it may be able to trace the chain of secondary causation and development, must at length find itself in the presence of the great Creative Mind, who is “before all things and in whom all things consist.”
I.—COMPARATIVE VIEW OF THE SUCCESSIVE PALÆOZOIC
FLORAS OF NORTHEASTERN AMERICA AND
GREAT BRITAIN.
In eastern Canada there is a very complete series of fossil plants, extending from the Silurian to the Permian, and intermediate in its species between the floras of interior America and of Europe. I may use this succession, mainly worked out by myself,[FG] to summarise the various Palæozoic floras and sub-floras, in order to give a condensed view of this portion of the history of the vegetable kingdom, and to direct attention to the important fact, too often overlooked, that there is a definite succession of fossil plants as well as of animals, and that this is important as a means of determining geological horizons. A British list for comparison has been kindly prepared for me by Mr. R. Kidston, F. Gr. S. For lists referring to the western and southern portions of America, I may refer to the reports of Lesquereux and Fontaine and White.[FH]
[FG] “Acadian Geology,” “Reports on Fossil Plants of Canada,” Geological Survey of Canada.
[FH] “Geological Surveys of Pennsylvania, Ohio, and Illinois.”
In this connection I am reminded, by an excellent little paper of M. Zeiller,[FI] on Carboniferous plants from the region of the Zambesi, in Africa, that the flora which in the Carboniferous period extended over the temperate portions of the northern hemisphere and far into the arctic, also passed across the equator and prevailed in the southern hemisphere. Of eleven species brought from the Zambesi by M. Lapierre and examined by M. Zeiller, all were identical with European « 274 » species of the upper coal-formation, and the same fact has been observed in the coal flora of the Cape Colony.[FJ] These facts bear testimony to the remarkable uniformity of climate and vegetation in the coal period, and I perfectly agree with Zeiller that they show, when taken in connection with other parallelisms in fossils, an actual contemporaneousness of the coal flora over the whole world.
[FI] Paris, 1883.
[FJ] Grey, “Journal of the Geological Society,” vol. xxvii.
1. Carboniferous Flora.
(1) Permo-Carboniferous Sub-Flora:
This occurs in the upper member of the Carboniferous system of Nova Scotia and Prince Edward Island, originally named by the writer the Newer Coal-formation, and more recently the Permo-Carboniferous, and the upper beds of which may not improbably be contemporaneous with the Lower Permian or Lower Dyas of Europe. In this formation there is a predominance of red sandstones and shales, and it contains no productive beds of coal. Its fossil plants are for the most part of species found in the Middle or Productive Coal-formation, but are less numerous, and there are a few new forms akin to those of the European Permian. The most characteristic species of the upper portion of the formation, which has the most decidedly Permian aspect, are the following:
| Dadoxylon materiarium, Dawson. | |
| * | Walchia (Araucarites) robusta, Dn. |
| * | W. (A.) gracilis, Dn. |
| * | W. imbricatula, Dn. |
| Calamites Suckovii, Brongt. | |
| C. Cistii, Brongt. | |
| * | C. gigas, Brongt. |
| Neuropteris rarinervis, Bunbury. | |
| Alethopteris nervosa, Brongt. | |
| Pecopteris arborescens, Brongt. | |
| * | P. rigida, Dn. |
| P. oreopteroides, Brongt. | |
| * | Cordaites simplex, Dn. |
Of these species, those marked with an asterisk have not yet been found in the middle or lower members of the Carboniferous system. They will be found described, and several of them figured, in my “Report on the Geology of Prince Edward Island.”[FK] The others are « 275 » common and widely diffused Carboniferous species, some of which have extended to the Permian period in Europe as well. From the upper beds, characterised by these and a few other species, there is a gradual passage downward into the productive coal-measures, and a gradually increasing number of true coal-formation species.
[FK] 1871.
It is worthy of remark here that the association in the Permo-Carboniferous of numerous trunks of Dadoxylon with the branches of Walchia and with fruits of the character of Trigonocarpa, seems to show that these were parts of one and the same plant.
This formation represents the Upper Barren Measures of West Virginia, which are well described by Fontaine and White,[FL] and the reasons which these authors adduce for considering the latter equivalent to the European Permian will apply to the more northern and eastern deposits as well, though these have afforded fewer species of plants, and are apparently less fully developed.
[FL] “Report on the Permian Flora of Western Virginia and South Pennsylvania,” 1880.
(2) Coal-formation Sub-Flora:
The Middle or Productive Coal-formation, containing all the beds of coal which are mined in Nova Scotia and Cape Breton, is the headquarters of the Carboniferous flora. From this formation I have catalogued[FM] one hundred and thirty-five species of plants; but, as several of these are founded on imperfect specimens, the number of actual species may be estimated at one hundred and twenty. Of these more than one half are species common to Europe and America. No less than nineteen species are Sigillariæ, and about the same number are Lepidodendra. About fifty are ferns and thirteen are Calamites, Asterophyllites, and Sphenophylla. The great abundance and number of species of Sigillariæ, Lepidodendra, and ferns are characteristic of this sub-flora; and among the ferns certain species of Neuropteris, Pecopteris, Alethopteris, and Sphenopteris greatly preponderate.
[FM] “Acadian Geology,” and “Report on Flora of Lower Carboniferous,” 1873.
These beds are the equivalents of the Middle Coal-measures, or Productive Coal-measures of Pennsylvania, Ohio, &c., and of the coal-formation proper of various European countries. Very many of the species are common to Nova Scotia and Pennsylvania; but in proceeding westward the number of identical species seems to diminish.
(3) The Millstone Grit Sub-Flora:
In this formation the abundance of plants and the number of species are greatly diminished.[FN] Trunks of coniferous trees of the species Dadoxylon Acadianum, having wide wood-cells with three or more series of discs and complex medullary rays, become characteristic. Calamites undulatum is abundant and seems to replace C. Suckovii, though C. cannæformis and C. cistii continue. Sigillariæ become very rare, and the species of Lepidodendron are few, and mostly those with large leaf-bases. Lepidophloios still continues, and Cordaites abounds in some beds. The ferns are greatly reduced, though a few characteristic coal-formation species occur, and the genus Cardiopteris appears. Beds of coal are rare in this formation; but where they occur there is in connection with them a remarkable anticipation of the rich coal-formation flora, which would thus seem to have existed locally in the Millstone Grit period, but to have found itself limited by generally unfavorable conditions. In America, as in Europe, it is in the north that this earlier development of the coal-flora occurs, while in the south there is a lingering of old forms in the newer beds. In Newfoundland and Cape Breton, for instance, as well as in Scotland, productive coal-beds and a greater variety of species of plants occur in this formation.
[FN] “Report on Fossil Plants of the Lower Carboniferous and Millstone Grit of Canada,” 1873.
The following would appear to be the equivalents of this formation, in flora and geological position:
1. The Seral Conglomerate of Rogers in Pennsylvania, &c.
2. The Lower Coal-formation Conglomerate and Chester groups of Illinois (Worthen).
3. The Lower Carboniferous Sandstone of Kentucky, Alabama, and Virginia.
4. The Millstone Grit and Yoredale rocks of northern England, and the Culmiferous of Devonshire.
5. The Moor rock and Lower Coal-measures of Scotland.
6. Flagstones and Lower Shales of the south of Ireland, and Millstone Grit of the north of Ireland.
7. The Jüngste Grauwacke of the Hartz, Saxony, and Silesia.
(4) The Carboniferous Limestone Series:
This affords few fossil plants in eastern America, and in so far as known they are similar to those of the next group. In Scotland it is richer in plants, but, according to Mr. Kidston, these are largely « 277 » similar to those of the underlying beds, though with some species which extend upward into the Millstone Grit. In Scotland the alga named Spirophyton and Archæocalamites radiatus—which in America are Erian—appear in this formation.
(5) The Lower Carboniferous Sub-Flora:
This group of plants is best seen in the shales of the Horton series, under the Lower Carboniferous marine limestones. It is small and peculiar. The most characteristic species are the following:
Dadoxylon (Palæoxylon) antiquius, Dn.—A species with large medullary rays of three or more series of cells.
Lepidodendron corrugatum, Dn.—A species closely allied to L. Veltheimianum of Europe, and which is its American representative. This is perhaps the most characteristic plant of the formation. It is very abundant, and presents very protean appearances, in its old stems, branches, twigs, and Knorria forms. It had well-characterised stigmaria roots, and constitutes the oldest erect forest known in Nova Scotia.
Lepidodendron tetragonum, Sternberg.
L. obovatum, Sternb.
L. aculeatum, Sternb.
L. dichotomum, Sternb.
The four species last mentioned are comparatively rare, and the specimens are usually too imperfect to render their identification certain, but Lepidodendra are especially characteristic trees of this horizon.
Cyclopteris (Aneimites) Acadica, Dn.—A very characteristic fern, allied in the form of its fronds to C. tenuifolia of Goeppert, to C. nana of Eichwald, and to Adiantites antiquus of Stur. Its fructification, however, is nearer to that of Aneimia than to that of Adiantum.
Ferns of the genera Cardiopteris and Hymenophyllites also occur, though rarely.
Ptilophyton plumula, Dn.—This is the latest appearance of this Erian genus, which also occurs in the Lower Carboniferous of Europe and of the United States.
Cordaites borassifolia, Brongt.
On the whole, this small flora is markedly distinct from that of the Millstone Grit and true coal-formation, from which it is separated by the great length of time required for the deposition of the marine limestones and their associated beds, in which no land-plants « 278 » have been found; nor is this gap filled up by the conglomerates and coarse arenaceous beds which, as I have explained in “Acadian Geology,” in some localities take the place of the limestones, as they do also in the Appalachian region farther south.
The palæobotanical and stratigraphical equivalents of this series abroad would seem to be the following:
1. The Vespertine group of Rogers in Pennsylvania.
2. The Kinderhook group of Worthen in Illinois.
3. The Marshall group of Winchell in Michigan.
4. The Waverley sandstone (in part) of Ohio.
5. The Lower or False Coal-measures of Virginia.
6. The Calciferous sandstones of McLaren, or Tweedian group of Tate in Scotland.
7. The Lower Carboniferous slate and Coomhala grits of Jukes in Ireland.
8. The Culm and Culm Grauwacke of Germany.
9. The Graywacke or Lower Coal-measures of the Vosges, as described by Schimper.
10. The Older Coal-formation of the Ural, as described by Eichwald.
11. The so-called “Ursa Stage” of Heer includes this, but he has united it with Devonian beds, so that the name cannot be used except for the local development of these beds at Bear Island, Spitsbergen. The Carboniferous plants of arctic America, Melville Island, &c., as well as those of Spitzbergen, appear all to be Lower Carboniferous.[FO]
[FO] “Notes on Geological Map of the Northern Portion of the Dominion of Canada,” by Dr. G. M. Dawson, 1887.
All of the above groups of rocks are characterised by the prevalence of Lepidodendra of the type of L. corrugatum, L. Veltheimianum, and L. Glincanum; pines of the sub-genus Pitus of Witham, Palæoxylon of Brongniart, and peculiar ferns of the genera Cyclopteris, Cardiopteris, Triphyllopteris, and Sphenopteris. In all the regions above referred to they form the natural base of the great Carboniferous system.
In Virginia, according to Fontaine and White, types, such as Archæopteris, which in the north are Upper Erian, occur in this group. Unless there have been some errors in fixing the lower limit of the Vespertine, this would indicate a longer continuance of old forms in the south.
2. Erian Flora.
(1) Upper Erian Sub-Flora:
This corresponds to the Catskill and Chemung of the New York series, and to the Upper Devonian of Europe.
The flora of this formation, which consists mostly of sandstones, is not rich. Its most distinctive species on both sides of the Atlantic seem to be the ferns of the genus Archæopteris, along with species referred to the genus Cyclopteris, but which, in so far as their barren fronds are concerned, for the most part resemble Archæopteris.
The characteristic American species are Archæopteris Jacksoni, A. Rogersi, and A. Gaspiensis. Cyclopteris obtusa and C. (Platyphyllum) Brownii are also very characteristic species. In Europe, Archæopteris Hibernica is a prevalent species.
Leptophleum rhombicum and fragments of Psilophyton are also found in the Upper Erian. There is evidence of the existence of vast numbers of Rhizocarps in this period, in the deposits of spore-cases (Sporangites Huronensis) in the shales of Kettle Point, Lake Huron; and in deposits of similar character in Ohio and elsewhere in the West.
The Upper Erian flora is thus very distinct from that of the Lower Carboniferous, and the unconformable relation of the beds in the Northeast may perhaps indicate a considerable lapse of time. Still, even in localities where there appears to be a transition from the Carboniferous into the Devonian, as in the Western States and in Ireland, the characteristic flora of each formation may be distinguished, though, as already stated, there is apparently some mixture in the South.
(2) Middle Erian Sub-Flora:
Both in Canada and the United States that part of the great Erian system which may be regarded as its middle division, the Hamilton and Marcellus shales of New York, the Cordaites shales of St. John, New Brunswick, and the middle shales and sandstones of the Gaspé series, presents conditions more favourable to the abundant growth of land-plants than either the upper or lower member. In the St. John beds, in particular, there is a rich fern flora, comparable with that of the coal-formation, and numerous stipes of ferns and trunks of tree-ferns have been found in the Hamilton and Corniferous series in the West, as well as trunks of Dadoxylon. It is, however, distinguished by a prevalence of small and delicate species, and by such forms as Hymenophyllites and the smaller Sphenopterids, and also by some peculiar ferns, as Archæopteris and Megalopteris. « 280 » In addition to ferns, it has small Lepidodendra, of which L. Gaspianum is the chief. Calamiteæ occur, Archæocalamites radiatus being the dominant species. This plant, which in Europe appears to reach up into the Lower Carboniferous, is so far strictly Erian in northeast America. Sigillariæ scarcely appear, but Cordaites is abundant, and the earliest known species of Dadoxylon appear, while the Psilophyton, so characteristic of the Lower Erian, still continues, and the remarkable aquatic plants of the genus Ptilophyton are locally abundant.
(3) Lower Erian Sub-Flora:
This belongs to the Lower Devonian sandstones and shales, and is best seen in that formation at Gaspé and the Bay des Chaleurs. It is equivalent to the Oriskany sandstone, so far as its animal fossils and mineral character are concerned. It is characterised by the absence of true ferns, Calamites and Sigillariæ, and by the presence of such forms as Psilophyton, Arthrostigma, Leptophleum, and Nematophyton. Lepidodendron Gaspianum and Leptophleum already occur, though not nearly so abundant as Psilophyton.
The Lower Erian plants have an antique and generalised aspect which would lead us to infer that they are near the beginning of the land-flora, or perhaps in part belong to the close of an earlier flora still in great part unknown and few indications of land-plants have been found earlier.
At Campbellton and Scaumenac Bay, on the Bay des Chaleurs, fossil fishes of genera characteristic of the Lower and Upper Devonian horizons respectively, occur in association with fossil plants of these horizons, and have been described by Mr. Whiteaves.[FP]
[FP] “Transactions of the Royal Society of Canada.”
It is interesting to note that, as Fontaine and White have observed, certain forms which are Erian in the northeast are found in the Lower members of the Carboniferous in West Virginia, indicating the southward march of species in these periods.
3. The Silurian Flora and still Earlier Indications of Plants.
In the upper beds of the Silurian, those of the Helderberg series, we still find Psilophyton and Nematophyton; but below these we know no land-plants in Canada. In the United States, Lesquereux and Claypole have described remains which may indicate the existence of lycopodiaceous and annularian types as far back as the beginning « 281 » of the Upper Silurian, or even as low as the Hudson River group, and Hicks has found Nematophyton and Psilophyton in beds about as old in Wales, along with the uncertain stems named Berwynia. In the Lower Silurian the Protannularia of the Skiddaw series in England may represent a land-plant, but this is uncertain, and no similar species has been found in Canada.
The Cambrian rocks are so far barren of land-plants; the so-called Eophyton being evidently nothing but markings, probably produced by crustaceans and other aquatic animals. In the still older Laurentian the abundant beds of graphite probably indicate the existence of plants, but whether aquatic or terrestrial it is impossible to decide at present.
It would thus appear that our certain knowledge of land-vegetation begins with the Upper Silurian or the Silurio-Cambrian, and that its earliest forms were Acrogens allied to Lycopods, and prototypal trees, forerunners of the Acrogens or the gymnosperms. In the Lower Devonian little advance is made. In the Middle Devonian this meagre flora had been replaced by one rivalling that of the Carboniferous, and including pines, tree-ferns, and arboreal forms of Lycopods and of equisetaceous plants, as well as numerous herbaceous plants. At the close of the Erian the flora again became meagre, and continued so in the Lower Carboniferous. It again became rich and varied in the Middle Carboniferous, to decay in the succeeding Permian.
II.—HEER’S LATEST RESULTS IN THE GREENLAND FLORA.
A very valuable report of Prof. Steenstrup, published in Copenhagen in 1883, the year in which Heer died, contains the results of his last work on the Greenland plants, and is so important that a summary of its contents will be interesting to all students of fossil botany or of the vicissitudes of climate which the earth has undergone.[FQ]
[FQ] Meddelelser om Gronland, Hefte V., Copenhagen, 1883.
The plant-bearing beds of Greenland are as follows, in ascending order:
1. Cretaceous.
1. The Komé series, of black shales resting on the Laurentian gneiss. These beds are found at various other localities, but the « 282 » name above given is that by which they are generally known. Their flora is limited to ferns, cycads, conifers, and a few endogens, with only Populus primæva to represent the dicotyledons. These beds are regarded as Lower Cretaceous (Urgonian), but the animal fossils would seem to give them a rather higher position. They may be regarded as equivalent to the Kootanie and Queen Charlotte beds in Canada, and the Potomac series in Virginia.
2. The Atané series. These also are black shales with dark-coloured sandstones. They are best exposed at Upernavik and Waigat. Here dicotyledonous leaves abound, amounting to ninety species, or more than half the whole number of species found. The fossil plants resemble those of the Dakota series of the United States and the Dunvegan series of Canada, and the animal fossils indicate the horizon of the Fort Pierre or its lower part. They may be regarded as representing the lower part of the Upper Cretaceous. The genera Populus, Myrica, Quercus, Ficus, Platanus, Sassafras, Laurus, Magnolia, and Liriodendron are among those represented in these beds, and the peculiar genera Macclintockia and Credneria are characteristic. The genus Pinus is represented by five species, Sequoia by five, and Salisburia by two, with three of the allied genus Baiera. There are many ferns and cycads.
3. The Patoot series. These are yellow and red shales, which seem to owe their colour to the spontaneous combustion of pyritous lignite, in the manner observed on the South Saskatchewan and the Mackenzie rivers. Their age is probably about that of the Fox-Hill group or Senonian, and the Upper Cretaceous of Vancouver Island, and they afford a large proportion of dicotyledonous leaves. The genera of dicotyledons are not dissimilar from those of Atané, but we now recognise Betula and Alnus, Comptonia, Planera, Sapotacites, Fraxinus, Viburnum, Cornus, Acer, Celastrus, Paliurus, Ceanothus, Zizyphus, and Cratægus as new genera of modern aspect.
On the whole there have been found in all these beds 335 species, belonging to 60 families, of which 36 are dicotyledonous, and represent all the leading types of arborescent dicotyledons of the temperate latitudes. The flora is a warm temperate one, with some remarkable mixtures of sub-tropical forms, among which perhaps the most remarkable are Kaidocarpum referred to the Pandaneæ, and such exogens as Ficus and Cinnamomum.
2. Tertiary.
4. The Unartok series. This is believed to be Eocene. It consists of sandstone, which appears on the shores of Disco Island, and « 283 » possibly at some other places on the coast. The beds rest directly and apparently conformably on the Upper Cretaceous, and have afforded only eleven species of plants. Magnolia is represented by two species, Laurus by two, Platanus by two, and one of these said to be identical with a species found by Lesquereux in the Laramie,[FR] Viburnum, Juglans, Quercus, each by one species; the ubiquitous Sequoias by S. Langsdorfii. This is pretty clearly a Lower Laramie flora.
[FR] Viburnum marginatum of Lesquereux.
5. The Atanekerdluk series, consisting of shaly beds, with limestone intercalated between great sheets of basalt, much like the Eocene of Antrim and the Hebrides. These beds have yielded 187 species, principally in bands and concretions of siderite, and often in a good state of preservation. They are referred to the Lower Miocene, but, as explained in the text, the flora is more nearly akin to that of the Eocene of Europe and the Laramie of America. The animal fossils are chiefly fresh-water shells. Onoclea sensibilis, several conifers, as Taxites Olriki, Taxodium distichum, Glyptostrobus Europæus, and Sequoia Langsdorfii, and 42 of the dicotyledons are recognised as found also in American localities. Of these, a large proportion of the more common species occur in the Laramie of the Mackenzie River and elsewhere in northwest Canada, and in the western United States. It is quite likely also that several species regarded as distinct may prove to be identical.
It would seem that throughout the whole thickness of these Tertiary beds the flora is similar, so that it is probable it belongs altogether to the Eocene rather than to the Miocene.
No indication has been observed of any period of cold intervening between the Lower Cretaceous and the top of the Tertiary deposits, so that, in all the vast period which these formations represent, the climate of Greenland would seem to have been temperate. There is, however, as is the case farther south, evidence of a gradual diminution of temperature. In the Lower Cretaceous the probable mean annual temperature in latitude 71° north is stated as 21° to 22° centigrade, while in the early Tertiary it is estimated at 12° centigrade. Such temperatures, ranging from 71° to 53° of Fahrenheit, represent a marvellously warm climate for so high a latitude. In point of fact, however, the evidence of warm climates in the arctic regions, in the Palæozoic as well as in the Mesozoic and early Tertiary, should perhaps lead us to conclude that, relatively to the whole of geological time, the present arctic climate is unusually severe, and « 284 » that a temperate climate in the arctic regions has throughout geological time been the rule rather than the exception.
III.—MINERALISATION OF FOSSIL PLANTS.
The state of preservation of fossil plants has been referred to incidentally in several places in the text; but the following more definite statements may be of service to the reader.
I. Organic remains imbedded in aqueous deposits may occur in an unchanged condition, or only more or less altered by decay. This is often the case with such enduring substances as bark and wood, and even with leaves, which appear as thin carbonaceous films when the layers containing them are split open. In the more recent deposits such remains occur little modified, or perhaps only slightly changed by partial decay of their more perishable parts. In the older formations, however, they are usually found in a more or less altered condition, in which their original substance has been wholly or in part changed into coaly, or bituminous, or anthracitic or graphitic matter, so that leaves are sometimes represented by stains of graphite, as if drawn on stone with a lead-pencil. Yet even in this case some portion of the original substance remains, and without any introduction of foreign material.
II. On the other hand, such remains are often mineralised by the filling of their pores or the replacement of their tissues with mineral matter, so that they become hard and stony, and sometimes retain little or nothing of their original substance. The more important of these changes, in so far as they affect fossil plants, may be arranged under the following heads:
(a) Infiltration of mineral matter which has penetrated the pores of the fossil in a state of solution. Thus the pores of fossil wood are often filled with calcite, quartz, oxide of iron, or sulphide of iron, while the woody walls of the cells and vessels remain in a carbonised state, or converted into coaly matter. When wood is preserved in this way it has a hard and stony aspect; but we can sometimes dissolve away the mineral matter, and restore the vegetable tissue to a condition resembling that before mineralisation. This is especially the case when calcite is the mineralising substance. We sometimes find, on microscopic examination, that even cavities so small as those of vegetable cells and vessels have been filled with successive coats of different kinds of mineral matter.
(b) Organic matters may be entirely replaced by mineral substances. In this case the cavities and pores have been first filled, « 285 » and then—the walls or solid parts being removed by decay or solution—mineral matter, either similar to that filling the cavities, or differing in colour or composition, has been introduced. Silicified wood often occurs in this condition. In the case of silicified wood, it sometimes happens that the cavities of the fibers have been filled with silica, and the wood has been afterward removed by decay, leaving the casts of the tubular fibers as a loose filamentous substance. Some of the Tertiary coniferous woods of California are in this state, and look like asbestus, though they show the minute markings of the tissue under the microscope. In the case of silicified or agatized woods, it would seem that the production of carbon dioxide from the decaying wood has caused the deposition of silica in its place, from alkaline solutions of that substance, and thus the carbon has been replaced, atom by atom, by silicon, until the whole mass has been silicified, yet retaining perfectly its structure.
(c) The cavities left by fossils which have decayed may be filled with clay, sand, or other foreign matter, and this, becoming subsequently hardened into stone, may constitute a cast of the fossils. Trunks of trees, roots, &c., are often preserved in this way, appearing as stony casts, often with the outer bark of the plant forming a carbonaceous coating on their surfaces. In connection with this state may be mentioned that in which, the wood having decayed, an entire trunk has been flattened so as to appear merely as a compressed film of bark, yet retaining its markings; and that in which the whole of the vegetable matter having been removed, a mere impression of the form remains.
Fossils preserved in either of the modes, (a) or (b), usually show more or less of their minute structures under the microscope. These may be observed:—(1) By breaking off small splinters or flakes and examining them, either as opaque or as transparent objects. (2) By treating the material with acids, so as to dissolve out the mineral matters, or portions of them. This method is especially applicable to fossil woods mineralised with calcite or pyrite. (3) By grinding thin sections. These are first polished on one face on a coarse stone or emery hone, and then on a fine hone, then attached by the polished face to glass slips with a transparent cement or Canada balsam, and ground on the opposite face until they become so thin as to be translucent. In most cities there are lapidaries who prepare slices of this kind; but the amateur can readily acquire the art by a little practice, and the necessary appliances can be obtained through dealers in minerals or in microscopic materials. Very convenient cutting and polishing machines, some of them quite small and portable, are « 286 » now made for the use of amateurs. In the case of exogenous woods, three sections are necessary to exhibit the whole of the structures. One of these should be transverse and two longitudinal, the latter in radial and tangential planes.
IV.—GENERAL WORKS ON PALÆOBOTANY.
In the text frequent reference has been made to special memoirs and reports on the fossil plants of particular regions or formations. There are, however, some general books, useful to students, which may be mentioned here. Perhaps the most important is Schimper’s “Traité de Paléontologie Végétale.” Very useful information is also contained in Renault’s “Cours de Botanique Fossile,” and in Balfour’s “Introduction to Palæontological Botany,” and Nicholson’s “Palæontology.” Unger’s “Genera et Species,” Brongniart’s “Histoire des Végétaux Fossiles,” and Lindley and Button’s “Fossil Flora,” are older though very valuable works. Williamson’s “Memoirs,” in the “Philosophical Transactions,” have greatly advanced our knowledge of the structures of Palæozoic plants. Lastly, the “Palæophytology” of Schenk, now in course of publication in German and French, in connection with Zittel’s “Palæontology,” is an important addition to manuals of the subject.
Acer, 228.
Acrogens, 6.
Agassiz, Prof., 16.
Alaska, Flora of, 245.
Algæ, real and spurious, 26, 230.
Amboy clays, Flora of, 203.
America, Cretaceous of, 190.
Angiosperms, 6.
Annularia, 122.
Anogens, 6.
Antholithes, 132.
Aporoxylon, 25.
Araucarioxylon, 148.
Araucarites, 134.
Archæocalamites, 170.
Archæopteris, 77, 85.
Arctic origin of plants, 221, 238.
Arthrophycus, 30.
Arthrostigma, 67.
Asterophyllites, 78, 122, 170.
Asteropteris, 77, 85.
Astropolithon, 30.
Atané, Plants of, 242, 281.
Atanekerdluk, Plants of, 283.
Australia, Palæozoic flora of, 147.
Tertiary flora of, 217.
Bauhinia, 204.
Bear Island, 241.
Betula, 198.
Bilobites, 28.
Bovey Tracey, Plants of, 226.
Brasenia, 207.
Buckland, Dr., 179.
Buthotrephis, 37.
Calamites, 77, 123, 166.
Calamodendron, 125.
Cambrian flora, 20.
Canada, Erian of, 103.
Carboniferous of, 110.
Laramie of, 209.
Pleistocene of, 227.
Carbon in Laurentian, 9.
Carboniferous flora, 110.
Carboniferous, Climate of, 138.
of Southern Hemisphere, 147.
Cardiocarpum, 82, 153.
Carruthers, Mr., 24, 98, 180.
On modifications of modern plants, 225, 269.
Carya, 196.
Cauda-galli fucoid, 105.
Caulerpites, 29.
Caulopteris, 75, 94.
Clarke, Prof., 51.
Climate, Causes of, 247.
Climate and plants, 216, 220, 232.
of Carboniferous, 138.
of Cretaceous and Eocene, 216.
of Devonian, 47.
« 288 »
of Early Mesozoic, 178.
Climate and plants of Laurentian, 17.
of Pleistocene, 227, 230.
of Pliocene, 223.
Coal, origin of, 117, 139.
Comparison of floras, 272.
Composite, 266.
Cone-in-cone, 36.
Coniferæ, Erian, 78, 96.
Carboniferous, 134, 148.
Mesozoic, etc., 181.
Cope, Mr., 215.
Cordaites, 78, 130, 151.
Corylus, 213.
Crepin, M., 99.
Cretaceous, Flora of, 190.
Climate of, 216.
Croll on climate, 252.
Cromer, Plants of, 224.
Cycads, Mesozoic, 178.
Cyclostigma, 157.
Dadoxylon, 96, 134, 148.
Dawson, Dr. G. M., 52, 210.
Delgado, Prof., 26.
Dendrophycus, 33.
Derby, Orville, 53.
Devonian flora, 45.
Devonian or Erian, 107, 279.
Climate of, 47.
Dicotyledons, Cretaceous, 192.
Table of, 192.
Dictyolites, 33.
Dictyospongia, 39.
Disco, Exotic plants at, 256.
Flora of, 245, 282.
Drepanophycus, 39.
Drosera, 228.
Dunvegan beds, 244.
Eocene, Flora of, 208, 214.
Climate of, 216.
Eophyton, 31.
Eopteris, 72.
Eozoon of Laurentian, 9.
Equisetum, 176, 230.
Erian flora, 45, 279.
Climate of, 47.
Erian or Devonian, 107.
Ettingshausen, Dr., 187, 215.
Exogens, Cretaceous, 192.
Tertiary, 213, 224.
Fagus, 196, 197.
Ferns, Erian, 72.
Carboniferous, 126, 171.
Fructification of, 128.
Stems of, 90, 129.
Tertiary, 212.
Filices, 72, 126, 171.
Flora of Cambrian, 26.
of Carboniferous, 110, 274.
of Cretaceous, 190.
of Early Mesozoic, 175.
of Erian, 45, 279.
of Jurassic, 177, 186.
of Laramie, 209.
of Laurentian, 8.
of Miocene, 220, 223.
of Modern, 219.
of Permian, 274.
of Pleistocene, 223, 227.
of Tertiary, 191, 208, 214, 219.
Fontaine, Prof., 130, 176.
Fontinalis, 230.
Fort Union beds, 210.
Fucoids, 27.
Gardner, Mr. Starkie, 212.
Geinitz, Dr., 174.
Geological formations, Table of, 4.
Glossopteris, 147.
Glyptodendron, 25.
Glyptostrobus, 194.
Goeppert, Dr., 99.
Grant, Col., 36.
« 289 »
Graphite from plants, 8.
Gray, Dr., Origin of floras, 223, 237.
Greenland, Climate of, 216.
Fossil flora of, 247.
Gulielmites, 35.
Gymnosperms, 6.
Haliserites, 39.
Hartt, Prof., 53.
Heer, Dr., 108, 181.
Helderberg period, Sea of, 250.
Heterangium, 77.
Hicks, Dr., 21.
Hunt, Dr. Sterry, 13, 143.
Huxley, Prof., 53.
Hymenæa, 204.
Insects, Erian, 83.
Juglans, 196.
Jurassic flora, 177.
Kainozoic flora, 191, 208, 214, 219.
Kidston, Mr. R., 128, 273.
King, Mr. Clarence, 211.
Komé, Plants of, 242, 281.
Laramie flora, 209, 215.
Laurentian plants, 8.
Laurentian, Climate of, 17.
Laurophyllum, 193.
Laws of introduction of plants, 237, 266.
Leda clay, Flora of, 232.
Lepidodendron, 120, 156, 162.
Lepidophloios, 121, 157, 165.
Leptophleum, 157.
Lesquereux, Mr. L., 169, 214.
Licrophycus, 30.
Lignitic series of America, 208.
Liquidambar, 197.
Liriodendron, 199.
Lower Carboniferous flora, 277.
Logan, Sir W., 48.
Lyell on climate, 249.
Magnolia, 200.
McConnell, Mr., 209.
McNab, Prof., 169.
Megalopteris, 76.
Megaphyton, 129.
Mesozoic flora, 175.
Climate of, 178.
Migrations of plants, 240, 245.
Miller, Hugh, 98.
Miocene flora, 220.
Miocene, Supposed, 242.
Modern flora, 219.
Modern plants, how modified, 269.
Modifications of plants, 266.
Nathorst, Dr., 26, 196.
Nematodendreæ, 25.
Nematophycus, 23.
Nematophyton, 21, 22, 42.
Newberry, Dr., 200, 203, 214.
Newfoundland, Fossil plants of, 242.
Newton, Mr., 52.
Nicholson, Dr. A., 20.
Niobrara series, 243, 246.
Noeggerathia, 130.
Northern origin of plants, 238.
Origin of plants, 237.
Orton, Prof., 51.
Pachytheca, 21.
Palæanthus, 205.
Palæochorda, 30.
Palæophycus, 30, 38.
Palæozoic floras compared, 273.
Palms, 188, 194.
Pandanus, 188.
Patoot beds, 282.
Peach, Mr., 98.
Petroleum, Origin of, 56.
Phymatoderma, 29.
Plants, Classification of, 6.
Platanus, 198.
Platyphyllum, 74.
« 290 »
Pleistocene climate, 227, 230.
Pleistocene flora, 223, 227.
Pliocene climate, 223.
Podozamites, 178.
Poles, Supposed change of, 248.
Populus, 191, 228.
Potamogeton, 229.
Potentilla, 228.
Protannularia, 21.
Protichnites, 27.
Protophyllum, 199.
Protosalvinia, 52.
Protostigma, 20.
Prototaxites, 21.
Psaronius, 93.
Psilophyton, 64.
Ptilophyton, 62, 86.
Quercus, 197.
Rhizocarps, 48.
Rill-marks, 33.
Rusichnites, 28.
Saccamina, 57.
Salisburia, 180.
Salter, Mr., 98.
Salvinia, 54.
Saporta, Count de, 26, 193.
Saportea, 57.
Sassafras, 199.
Scalariform tissue, 70.
Schimper, Dr., 116, 169, 208.
Scolithus, 30.
Scottish Devonian, 98.
Sequoia, 181.
Shrinkage cracks, 33.
Sigillaria, 71, 112, 154.
Southern Hemisphere, 217, 273.
Carboniferous in, 147.
Tertiary in, 217.
Sphenophyllum, 61, 122, 171.
Spirophyton, 38.
Spitzbergen, 241.
Sterculites, 193.
Sternbergia, 137, 152.
Stigmaria, 115.
Stur, Dr., on Sigillaria, 116.
Symphorocarpus, 214.
Syringodendron, 156.
Syringoxylon, 82.
Table of formations, 4.
Tasmania, Fossil plants of, 217, 246.
Tasmanite, 57.
Tertiary period, Flora of, 191, 208, 214, 219.
Tertiary of Australia, 217.
Thallogens, 6.
Thomas, Mr., 51.
Thuja, 213, 229.
Time, Geological, 5.
Trapa, 196.
Tree-ferns, 90, 129.
Triassic flora, 176.
Trigonocarpum, 136, 153.
Tyndall, Prof., 138.
Ulrich, Prof., 57.
Unartok beds, 281.
Ursa stage of Heer, 108, 241.
Walchia, 134, 138.
Ward, Mr. L. T., 192, 212, 215.
Wethered, Mr. E., 52.
White, Dr., 215.
Williams, Prof., 51.
Williamson, Dr., 26, 31, 71, 167.
Williamsonia, 188.
THE END.
D. APPLETON & CO.'S PUBLICATIONS.
CHARLES DARWIN’S WORKS.
ORIGIN OF SPECIES BY MEANS OF NATURAL SELECTION, OR THE PRESERVATION OF FAVORED RACES IN THE STRUGGLE FOR LIFE. Revised edition, with Additions. 12mo. Cloth, $2.00.
DESCENT OF MAN, AND SELECTION IN RELATION TO SEX. With many Illustrations. A new edition. 12mo. Cloth, $3.00.
JOURNAL OF RESEARCHES INTO THE NATURAL HISTORY AND GEOLOGY OF COUNTRIES VISITED DURING THE VOYAGE OF H. M. S. BEAGLE ROUND THE WORLD. New edition. 12mo. Cloth, $2.00.
EMOTIONAL EXPRESSIONS OF MAN AND THE LOWER ANIMALS. 12 mo. Cloth, $3.50.
THE VARIATIONS OF ANIMALS AND PLANTS UNDER DOMESTICATION. With a Preface, by Professor Asa Gray. 2 vols. Illustrated. Cloth, $5.00.
INSECTIVOROUS PLANTS. 12mo. Cloth, $2.00.
MOVEMENTS AND HABITS OF CLIMBING PLANTS. With Illustrations. 12mo. Cloth, $1.25.
THE VARIOUS CONTRIVANCES BY WHICH ORCHIDS ARE FERTILIZED BY INSECTS. Revised edition, with Illustrations. 12mo. Cloth, $1.75.
THE EFFECTS OF CROSS AND SELF FERTILIZATION IN THE VEGETABLE KINGDOM. 12mo. Cloth, $2.00.
DIFFERENT FORMS OF FLOWERS ON PLANTS OF THE SAME SPECIES. With Illustrations. 12mo. Cloth, $1.50.
THE POWER OF MOVEMENT IN PLANTS. By Charles Darwin, LL. D., F. R. S., assisted by Francis Darwin. With Illustrations. 12mo. Cloth, $2.00.
THE FORMATION OF VEGETABLE MOULD THROUGH THE ACTION OF WORMS. With Observations on their Habits. With Illustrations. 12mo. Cloth, $1.50.
New York: D. APPLETON & CO., 1, 3, & 5 Bond Street.
D. APPLETON & CO.'S PUBLICATIONS.
THOMAS H. HUXLEY’S WORKS.
SCIENCE AND CULTURE, AND OTHER ESSAYS. 12mo. Cloth, $1.50.
THE CRAYFISH: AN INTRODUCTION TO THE STUDY OF ZOÕLOGY. With 82 Illustrations. 12mo. Cloth, $1.75.
SCIENCE PRIMERS: INTRODUCTORY. 18mo. Flexible cloth, 45 cents.
MAN’S PLACE IN NATURE. 12mo. Cloth, $1.25.
ON THE ORIGIN OF SPECIES. 12mo. Cloth, $1.00.
MORE CRITICISMS ON DARWIN, AND ADMINISTRATIVE NIHILISM. 12mo. Limp cloth, 50 cents.
MANUAL OF THE ANATOMY OF VERTEBRATED ANIMALS. Illustrated. 12mo. Cloth, $2.50.
MANUAL OF THE ANATOMY OF INVERTEBRATED ANIMALS. 12mo. Cloth, $2.50.
LAY SERMONS, ADDRESSES, AND REVIEWS. 12mo. Cloth, $1.75.
CRITIQUES AND ADDRESSES. 12mo. Cloth, $1.50.
AMERICAN ADDRESSES; WITH A LECTURE ON THE STUDY OF BIOLOGY. 12mo. Cloth, $1.25.
PHYSIOGRAPHY: AN INTRODUCTION TO THE STUDY OF NATURE. With Illustrations and Colored Plates. 12mo. Cloth, $2.50.
HUXLEY AND YOUMANS’S ELEMENTS OF PHYSIOLOGY AND HYGIENE. By T. H. Huxley and W. J. Youmans. 12mo. Cloth, $1.50.
New York: D. APPLETON & CO., 1, 3, & 5 Bond Street.
D. APPLETON & CO.'S PUBLICATIONS.
JOHN TYNDALL’S WORKS.
ESSAYS ON THE FLOATING MATTER OF THE AIR, in Relation to Putrefaction and Infection. 12mo. Cloth, $1.50.
ON FORMS OF WATER, in Clouds, Rivers, Ice, and Glaciers. With 35 Illustrations. 12mo. Cloth, $1.50.
HEAT AS A MODE OF MOTION. New edition. 12mo. Cloth, $2.50.
ON SOUND: A Course of Eight Lectures delivered at the Royal Institution of Great Britain. Illustrated. 12mo. New edition. Cloth, $2.00.
FRAGMENTS OF SCIENCE FOR UNSCIENTIFIC PEOPLE. 12mo. New revised and enlarged edition. Cloth, $2.50.
LIGHT AND ELECTRICITY. 12mo. Cloth, $1.25.
LESSONS IN ELECTRICITY, 1875-'76. 12mo. Cloth, $1.00
HOURS OF EXERCISE IN THE ALPS. With Illustrations. 12mo. Cloth, $2.00.
FARADAY AS A DISCOVERER. A Memoir. 12mo. Cloth, $1.00.
CONTRIBUTIONS TO MOLECULAR PHYSICS in the Do main of Radiant Heat. $5.00.
SIX LECTURES ON LIGHT. Delivered in America in 1872-'73. With an Appendix and numerous Illustrations. Cloth, $1.50.
FAREWELL BANQUET, given to Professor Tyndall, at Delmonico’s, New York, February 4, 1873. Paper, 50 cents.
ADDRESS delivered before the British Association, assembled at Belfast. Revised with Additions, by the author, since the Delivery 12mo. Paper, 50 cents.
New York: D. APPLETON & CO., 1, 3, & 5 Bond Street.
D. APPLETON & CO.'S PUBLICATIONS.
ERNST HAECKEL’S WORKS.
THE HISTORY OF CREATION; OR, THE DEVELOPMENT OF THE EARTH AND ITS INHABITANTS BY THE ACTION OF NATURAL CAUSES. A Popular Exposition of the Doctrine of Evolution in general, and of that of Darwin, Goethe, and Lamarck in particular. From the German of Ernst Haeckel, Professor in the University of Jena. The translation revised by Professor E. Ray Lankester, M. A., F. R, S., Fellow of Exeter College, Oxford. Illustrated with Lithographic Plates. In two vols., 12mo. Cloth, $5.00.
THE EVOLUTION OF MAN, A Popular Exposition of the Principal Points of Human Ontogeny and Phylogeny. From the German of Ernst Haeckel, Professor in the University of Jena, author of “The History of Creation,” etc. With numerous Illustrations. In two vols., 12mo. Cloth. Price, $5.00.
“In this excellent translation of Professor Haeckel’s work, the English reader has access to the latest doctrines of the Continental school of evolution, in its application to the history of man. It is in Germany, beyond any other European country, that the impulse given by Darwin twenty years ago to the theory of evolution has influenced the whole tenor of philosophical opinion. There may be, and are, differences in the degree to which the doctrine may be held capable of extension into the domain of mind and morals; but there is no denying, in scientific circles at least, that as regards the physical history of organic nature much has been done toward making good a continuous scheme of being.”—London Saturday Review.
FREEDOM IN SCIENCE AND TEACHING. From the German of Ernst Haeckel. With a Prefatory Note by T. H. Huxley, F. R. S. 12mo. $1.00.
New York: D. APPLETON & CO., 1, 3, & 5 Bond Street.
D. APPLETON & CO.'S PUBLICATIONS.
SIR JOHN LUBBOCK’S (Bart.) WORKS.
THE ORIGIN OF CIVILIZATION AND THE PRIMITIVE CONDITION OF MAN, MENTAL AND SOCIAL CONDITION OF SAVAGES. Fourth edition, with numerous Additions. With Illustrations, 8vo. Cloth, $5.00.
“The first edition of this work was published in the year 1870. The work has been twice revised for the press in the interval, and now appears in its fourth edition enlarged to the extent of nearly two hundred pages, including a full index.”
“This interesting work—for it is intensely so in its aim, scope, and the ability of its author—treats of what the scientists denominate anthropology, or the natural history of the human species; the complete science of man, body and soul, including sex, temperament, race, civilization, etc.”—Providence Press.
PREHISTORIC TIMES, AS ILLUSTRATED BY ANCIENT REMAINS AND THE MANNERS AND CUSTOMS OF MODERN SAVAGES. Illustrated. Entirely new revised edition. 8vo. Cloth, $5.00.
The book ranks among the noblest works of the interesting and important class to which it belongs. As a résumé of our present knowledge of prehistoric man, it leaves nothing to be desired. It is not only a good book of reference but the best on the subject.
“This is, perhaps, the best summary of evidence now in our possession concerning the general character of prehistoric times. The Bronze Age, The Stone Age, The Tumuli, The Lake Inhabitants of Switzerland, The Shell Mounds, The Cave Man, and The Antiquity of Man, are the titles of the most important chapters.”—Dr. C. K. Adams’s Manual of Historical Literature.
ANTS, BEES, AND WASPS. A Record of Observations on the Habits of the Social Hymenoptera. With Colored Plates. 12mo. Cloth, $2.00.
“This volume contains the record of various experiments made with ants, bees, and wasps during the last ten years, with a view to test their menial condition and powers of sense. The principal point in which Sir John’s mode of experiment differs from those of Huber, Forel, McCook, and others, is that he has carefully watched and marked particular insects, and has had their nests under observation for long periods—one of his ants' nests having been under constant inspection ever since 1874 His observations are made principally upon ants, because they show more power and flexibility of mind; and the value of his studies is that they belong to the department of original research.”
“We have no hesitation in saying that the author has presented us with the most valuable series of observations on a special subject that has ever been produced, charmingly written, full of logical deductions, and, when we consider his multitudinous engagements, a remarkable illustration of economy of time. As a contribution to insect psychology, it will be long before this book finds a parallel.”—London Athenæum.
New York: D. APPLETON & CO., 1, 3, & 5 Bond Street.
D. APPLETON & CO.'S PUBLICATIONS.
Br. H. ALLEYNE NICHOLSON’S WORKS.
TEXT-BOOK OF ZOÖLOGY, for Schools and Colleges. 12mo. Half roan, $1.60.
MANUAL OF ZOÖLOGY, for the Use of Students, with a General Introduction to the Principles of Zoölogy. Second edition. Revised and enlarged, with 243 Woodcuts. 12mo. Cloth, $2 50.
TEXT-BOOK OF GEOLOGY, for Schools and Colleges. 12mo. Half roan, $1.25.
INTRODUCTION TO THE STUDY OF BIOLOGY, Illustrated. 12mo. Cloth, 60 cents.
THE ANCIENT LIFE-HISTORY OF THE EARTH. A Comprehensive Outline of the Principles and Leading Facts of Palæontological Science. 12mo. Cloth, $2.00.
“A work by a master in the science who understands the significance of every phenomenon which he records, and knows how to make it reveal its lessons. As regards its value there can scarcely exist two opinions. As a text-book of the historical phase of palæontology is will be indispensable to students, whether specially pursuing geology or biology; and without it no man who aspires even to an outline knowledge of natural science can deem his library complete.”—The Quarterly Journal of Science.
“The Professor of Natural History in the University of St. Andrews has, by his previous works on zoology and paleontology, so fully established his claim to be an exact thinker and a close reasoner, that scarcely any recommendation of ours can add to the interest with which all students in natural history will receive the present volume. It is, as its second title expresses it; a comprehensive outline of the principles and leading facts of palæontoiogical science. Numerous woodcut illustrations very delicately executed, a copious glossary, and an admirable index, add much to the value of this volume.”—Athenæum.
New York: D. APPLETON & CO., 1, 3, & 5 Bond Street.
Transcriber Notes
All images were moved so as to not split paragraphs.
End of the Project Gutenberg EBook of The Geological History of Plants, by
Sir J. William Dawson
*** END OF THIS PROJECT GUTENBERG EBOOK THE GEOLOGICAL HISTORY OF PLANTS ***
***** This file should be named 51021-h.htm or 51021-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/5/1/0/2/51021/
Produced by Tom Cosmas compiled from files made available
at The Internet Archive.
Updated editions will replace the previous one--the old editions will
be renamed.
Creating the works from print editions not protected by U.S. copyright
law means that no one owns a United States copyright in these works,
so the Foundation (and you!) can copy and distribute it in the United
States without permission and without paying copyright
royalties. Special rules, set forth in the General Terms of Use part
of this license, apply to copying and distributing Project
Gutenberg-tm electronic works to protect the PROJECT GUTENBERG-tm
concept and trademark. Project Gutenberg is a registered trademark,
and may not be used if you charge for the eBooks, unless you receive
specific permission. If you do not charge anything for copies of this
eBook, complying with the rules is very easy. You may use this eBook
for nearly any purpose such as creation of derivative works, reports,
performances and research. They may be modified and printed and given
away--you may do practically ANYTHING in the United States with eBooks
not protected by U.S. copyright law. Redistribution is subject to the
trademark license, especially commercial redistribution.
START: FULL LICENSE
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full
Project Gutenberg-tm License available with this file or online at
www.gutenberg.org/license.
Section 1. General Terms of Use and Redistributing Project
Gutenberg-tm electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or
destroy all copies of Project Gutenberg-tm electronic works in your
possession. If you paid a fee for obtaining a copy of or access to a
Project Gutenberg-tm electronic work and you do not agree to be bound
by the terms of this agreement, you may obtain a refund from the
person or entity to whom you paid the fee as set forth in paragraph
1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this
agreement and help preserve free future access to Project Gutenberg-tm
electronic works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the
Foundation" or PGLAF), owns a compilation copyright in the collection
of Project Gutenberg-tm electronic works. Nearly all the individual
works in the collection are in the public domain in the United
States. If an individual work is unprotected by copyright law in the
United States and you are located in the United States, we do not
claim a right to prevent you from copying, distributing, performing,
displaying or creating derivative works based on the work as long as
all references to Project Gutenberg are removed. Of course, we hope
that you will support the Project Gutenberg-tm mission of promoting
free access to electronic works by freely sharing Project Gutenberg-tm
works in compliance with the terms of this agreement for keeping the
Project Gutenberg-tm name associated with the work. You can easily
comply with the terms of this agreement by keeping this work in the
same format with its attached full Project Gutenberg-tm License when
you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are
in a constant state of change. If you are outside the United States,
check the laws of your country in addition to the terms of this
agreement before downloading, copying, displaying, performing,
distributing or creating derivative works based on this work or any
other Project Gutenberg-tm work. The Foundation makes no
representations concerning the copyright status of any work in any
country outside the United States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other
immediate access to, the full Project Gutenberg-tm License must appear
prominently whenever any copy of a Project Gutenberg-tm work (any work
on which the phrase "Project Gutenberg" appears, or with which the
phrase "Project Gutenberg" is associated) is accessed, displayed,
performed, viewed, copied or distributed:
This eBook is for the use of anyone anywhere in the United States and
most other parts of the world at no cost and with almost no
restrictions whatsoever. You may copy it, give it away or re-use it
under the terms of the Project Gutenberg License included with this
eBook or online at www.gutenberg.org. If you are not located in the
United States, you'll have to check the laws of the country where you
are located before using this ebook.
1.E.2. If an individual Project Gutenberg-tm electronic work is
derived from texts not protected by U.S. copyright law (does not
contain a notice indicating that it is posted with permission of the
copyright holder), the work can be copied and distributed to anyone in
the United States without paying any fees or charges. If you are
redistributing or providing access to a work with the phrase "Project
Gutenberg" associated with or appearing on the work, you must comply
either with the requirements of paragraphs 1.E.1 through 1.E.7 or
obtain permission for the use of the work and the Project Gutenberg-tm
trademark as set forth in paragraphs 1.E.8 or 1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any
additional terms imposed by the copyright holder. Additional terms
will be linked to the Project Gutenberg-tm License for all works
posted with the permission of the copyright holder found at the
beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including
any word processing or hypertext form. However, if you provide access
to or distribute copies of a Project Gutenberg-tm work in a format
other than "Plain Vanilla ASCII" or other format used in the official
version posted on the official Project Gutenberg-tm web site
(www.gutenberg.org), you must, at no additional cost, fee or expense
to the user, provide a copy, a means of exporting a copy, or a means
of obtaining a copy upon request, of the work in its original "Plain
Vanilla ASCII" or other form. Any alternate format must include the
full Project Gutenberg-tm License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works
provided that
* You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is owed
to the owner of the Project Gutenberg-tm trademark, but he has
agreed to donate royalties under this paragraph to the Project
Gutenberg Literary Archive Foundation. Royalty payments must be paid
within 60 days following each date on which you prepare (or are
legally required to prepare) your periodic tax returns. Royalty
payments should be clearly marked as such and sent to the Project
Gutenberg Literary Archive Foundation at the address specified in
Section 4, "Information about donations to the Project Gutenberg
Literary Archive Foundation."
* You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or destroy all
copies of the works possessed in a physical medium and discontinue
all use of and all access to other copies of Project Gutenberg-tm
works.
* You provide, in accordance with paragraph 1.F.3, a full refund of
any money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days of
receipt of the work.
* You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project
Gutenberg-tm electronic work or group of works on different terms than
are set forth in this agreement, you must obtain permission in writing
from both the Project Gutenberg Literary Archive Foundation and The
Project Gutenberg Trademark LLC, the owner of the Project Gutenberg-tm
trademark. Contact the Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
works not protected by U.S. copyright law in creating the Project
Gutenberg-tm collection. Despite these efforts, Project Gutenberg-tm
electronic works, and the medium on which they may be stored, may
contain "Defects," such as, but not limited to, incomplete, inaccurate
or corrupt data, transcription errors, a copyright or other
intellectual property infringement, a defective or damaged disk or
other medium, a computer virus, or computer codes that damage or
cannot be read by your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium
with your written explanation. The person or entity that provided you
with the defective work may elect to provide a replacement copy in
lieu of a refund. If you received the work electronically, the person
or entity providing it to you may choose to give you a second
opportunity to receive the work electronically in lieu of a refund. If
the second copy is also defective, you may demand a refund in writing
without further opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS', WITH NO
OTHER WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT
LIMITED TO WARRANTIES OF MERCHANTABILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of
damages. If any disclaimer or limitation set forth in this agreement
violates the law of the state applicable to this agreement, the
agreement shall be interpreted to make the maximum disclaimer or
limitation permitted by the applicable state law. The invalidity or
unenforceability of any provision of this agreement shall not void the
remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in
accordance with this agreement, and any volunteers associated with the
production, promotion and distribution of Project Gutenberg-tm
electronic works, harmless from all liability, costs and expenses,
including legal fees, that arise directly or indirectly from any of
the following which you do or cause to occur: (a) distribution of this
or any Project Gutenberg-tm work, (b) alteration, modification, or
additions or deletions to any Project Gutenberg-tm work, and (c) any
Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of
computers including obsolete, old, middle-aged and new computers. It
exists because of the efforts of hundreds of volunteers and donations
from people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future
generations. To learn more about the Project Gutenberg Literary
Archive Foundation and how your efforts and donations can help, see
Sections 3 and 4 and the Foundation information page at
www.gutenberg.org
Section 3. Information about the Project Gutenberg Literary Archive Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg Literary
Archive Foundation are tax deductible to the full extent permitted by
U.S. federal laws and your state's laws.
The Foundation's principal office is in Fairbanks, Alaska, with the
mailing address: PO Box 750175, Fairbanks, AK 99775, but its
volunteers and employees are scattered throughout numerous
locations. Its business office is located at 809 North 1500 West, Salt
Lake City, UT 84116, (801) 596-1887. Email contact links and up to
date contact information can be found at the Foundation's web site and
official page at www.gutenberg.org/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
[email protected]
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To SEND
DONATIONS or determine the status of compliance for any particular
state visit www.gutenberg.org/donate
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations. To
donate, please visit: www.gutenberg.org/donate
Section 5. General Information About Project Gutenberg-tm electronic works.
Professor Michael S. Hart was the originator of the Project
Gutenberg-tm concept of a library of electronic works that could be
freely shared with anyone. For forty years, he produced and
distributed Project Gutenberg-tm eBooks with only a loose network of
volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as not protected by copyright in
the U.S. unless a copyright notice is included. Thus, we do not
necessarily keep eBooks in compliance with any particular paper
edition.
Most people start at our Web site which has the main PG search
facility: www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.