This eBook is for the use of anyone anywhere at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org
Title: The Organism as a Whole
From a Physicochemical Viewpoint
Author: Jacques Loeb
Release Date: June 14, 2014 [eBook #45962]
Language: English
Character set encoding: UTF-8
***START OF THE PROJECT GUTENBERG EBOOK THE ORGANISM AS A WHOLE***
| Note: | Images of the original pages are available through Internet Archive/American Libraries. See https://archive.org/details/organismaswholef00loeb |
Transcriber’s note:
In this transcription a black dotted underline indicates a hyperlink to a specific page or illustration; hyperlinks also show aqua highlighting when the cursor hovers over them. Page numbers appear in the right margin.
“He was one of those simple, disinterested, and intellectually
sterling workers to whom their own personality is as nothing in the
presence of the vast subjects that engage the thoughts of their
lives.”
John Morley.
(Article Diderot, Encyclopædia Britannica.)
It is generally admitted that the individual physiological processes, such as digestion, metabolism, the production of heat or of electricity, are of a purely physicochemical character; and it is also conceded that the functions of individual organs, such as the eye or the ear, are to be analysed from the viewpoint of the physicist. When, however, the biologist is confronted with the fact that in the organism the parts are so adapted to each other as to give rise to a harmonious whole; and that the organisms are endowed with structures and instincts calculated to prolong their life and perpetuate their race, doubts as to the adequacy of a purely physicochemical viewpoint in biology may arise. The difficulties besetting the biologist in this problem have been rather increased than diminished by the discovery of Mendelian heredity, according to which each character is transmitted independently of any other character. Since the number of Mendelian characters in each organism is large, the possibility must be faced that the organism is merely a mosaic of independent hereditary characters. If this be the case the question arises: What moulds these independent characters into a harmonious whole?
The vitalist settles this question by assuming the existence of a pre-established design for each organism and of a guiding “force” or “principle” which directs the working out of this design. Such assumptions remove the problem of accounting for the harmonious character of the organism from the field of physics or chemistry. The theory of natural selection invokes neither design nor purpose, but it is incomplete since it disregards the physicochemical constitution of living matter about which little was known until recently.
In this book an attempt is made to show that the unity of the organism is due to the fact that the egg (or rather its cytoplasm) is the future embryo upon which the Mendelian factors in the chromosomes can impress only individual characteristics, probably by giving rise to special hormones and enzymes. We can cause an egg to develop into an organism without a spermatozoön, but apparently we cannot make a spermatozoön develop into an organism without the cytoplasm of an egg, although sperm and egg nucleus transmit equally the Mendelian characters. The conception that the cytoplasm of the egg is already the embryo in the rough may be of importance also for the problem of evolution since it suggests the possibility that the genus- and species-heredity are determined by the cytoplasm of the egg, while the Mendelian hereditary characters cannot contribute at all or only to a limited extent to the formation of new species. Such an idea is supported by the work on immunity, which shows that genus- and probably species-specificity are due to specific proteins, while the Mendelian characters may be determined by hormones which need neither be proteins nor specific or by enzymes which also need not be specific for the species or genus. Such a conception would remove the difficulties which the work on Mendelian heredity has seemingly created not only for the problem of evolution but also for the problem of the harmonious character of the organism as a whole.
Since the book is intended as a companion volume to the writer’s former treatise on The Comparative Physiology of the Brain a discussion of the functions of the central nervous system is omitted.
Completeness in regard to quotation of literature was out of the question, but the writer notices with regret, that he has failed to refer in the text to so important a contribution to the subject as Sir E. A. Schäfer’s masterly presidential address on “Life” or the addresses of Correns and Goldschmidt on the determination of sex. Credit should also have been given to Professor Raymond Pearl for the discrimination between species and individual inheritance.
The writer wishes to acknowledge his indebtedness to his friends Professor E. G. Conklin of Princeton, Professor Richard Goldschmidt of the Kaiser Wilhelm Institut of Berlin, Dr. P. A. Levene of the Rockefeller Institute, Professor T. H. Morgan of Columbia University, and Professor Hardolph Wasteneys of the University of California who kindly read one or more chapters of the book and offered valuable suggestions; and he wishes especially to thank his wife for suggesting many corrections in the manuscript and the proof.
The book is dedicated to that group of freethinkers, including d’Alembert, Diderot, Holbach, and Voltaire, who first dared to follow the consequences of a mechanistic science—incomplete as it then was—to the rules of human conduct and who thereby laid the foundation of that spirit of tolerance, justice, and gentleness which was the hope of our civilization until it was buried under the wave of homicidal emotion which has swept through the world. Diderot was singled out, since to him the words of Lord Morley are devoted, which, however, are more or less characteristic of the whole group.
J. L.
The Rockefeller Institute
for Medical Research,
August, 1916
1. The physical researches of the last ten years have put the atomistic theory of matter and electricity on a definite and in all probability permanent basis. We know the exact number of molecules in a given mass of any substance whose molecular weight is known to us, and we know the exact charge of a single electron. This permits us to state as the ultimate aim of the physical sciences the visualization of all phenomena in terms of groupings and displacements of ultimate particles, and since there is no discontinuity between the matter constituting the living and non-living world the goal of biology can be expressed in the same way.
This idea has more or less consciously prevailed for some time in the explanation of the single processes occurring in the animal body or in the explanation of the functions of the individual organs. Nobody, not even a scientific vitalist, would think of treating the process of digestion, metabolism, production of heat, and electricity or even secretion or muscular contraction in any other than a purely chemical or physicochemical way; nor would anybody think of explaining the functions of the eye or the ear from any other standpoint than that of physics.
When the actions of the organism as a whole are concerned, we find a totally different situation. The same physiologists who in the explanation of the individual processes would follow the strictly physicochemical viewpoint and method would consider the reactions of the organism as a whole as the expression of non-physical agencies. Thus Claude Bernard,1 who in the investigation of the individual life processes was a strict mechanist, declares that the making of a harmonious organism from the egg cannot be explained on a mechanistic basis but only on the assumption of a “directive force.” Bernard assumes, as Bichat and others had done before him, that there are two opposite processes going on in the living organism: (1) the phenomena of vital creation or organizing synthesis; (2) the phenomena of death or organic destruction. It is only the destructive processes which give rise to the physical manifestations by which we judge life, such as respiration and circulation or the activity of glands, and so on. The work of creation takes place unseen by us in the egg when the embryo or organism is formed. This vital creation occurs always according to a definite plan, and in the opinion of Bernard it is impossible to account for this plan on a purely physicochemical basis.
There is so to speak a pre-established design of each being and of each organ of such a kind that each phenomenon by itself depends upon the general forces of nature, but when taken in connection with the others it seems directed by some invisible guide on the road it follows and led to the place it occupies. . . .
We admit that the life phenomena are attached to physicochemical manifestations, but it is true that the essential is not explained thereby; for no fortuitous coming together of physicochemical phenomena constructs each organism after a plan and a fixed design (which are foreseen in advance) and arouses the admirable subordination and harmonious agreement of the acts of life. . . .
We can only know the material conditions and not the intimate nature of life phenomena. We have therefore only to deal with matter and not with the first causes or the vital force derived therefrom. These causes are inaccessible to us, and if we believe anything else we commit an error and become the dupes of metaphors and take figurative language as real. . . . Determinism can never be but physicochemical determinism. The vital force and life belong to the metaphysical world.
In other words, Bernard thinks it his task to account for individual life phenomena on a purely physicochemical basis—but the harmonious character of the organism as a whole is in his opinion not produced by the same forces and he considers it impossible and hopeless to investigate the “design.” This attitude of Bernard would be incomprehensible were it not for the fact that, when he made these statements, the phenomena of specificity, the physiology of development and regeneration, the Mendelian laws of heredity, the animal tropisms and their bearing on the theory of adaptation were unknown.
This explanation of Bernard’s attitude is apparently contradicted by the fact that Driesch2 and v. Uexküll,3 both brilliant biologists, occupy today a standpoint not very different from that of Claude Bernard. Driesch assumes that there is an Aristotelian “entelechy” acting as directing guide in each organism; and v. Uexküll suggests a kind of Platonic “idea” as a peculiar characteristic of life which accounts for the purposeful character of the organism.
v. Uexküll supposes as did Claude Bernard and as does Driesch that in an organism or an egg the ultimate processes are purely physicochemical. In an egg these processes are guided into definite parts of the future embryo by the Mendelian factors of heredity—the so-called genes. These genes he compares to the foremen for the different types of work to be done in a building. But there must be something that makes of the work of the single genes a harmonious whole, and for this purpose he assumes the existence of “supergenes.”4 v. Uexküll’s ideas concerning the nature of a Mendelian factor and of the “supergenes” are expressed in metaphorical terms and the assumption of the “supergenes” begs the question. The writer is under the impression that this author was led to his views by the belief that the egg is entirely undifferentiated. But the unfertilized egg is not homogeneous, on the contrary, it has a simple but definite physicochemical structure which suffices to determine the first steps in the differentiation of the organism. Of course, if we suppose as do v. Uexküll and Driesch that the egg has no structure, the development of structure becomes a difficult problem—but this is not the real situation.
2. Claude Bernard does not mention the possibility of explaining the harmony or apparent design in the organism on the basis of the theory of evolution, he simply considers the problem as outside of biology. It was probably clear to him as it must be to everyone with an adequate training in physics that natural selection does not explain the origin of variation. Driesch and v. Uexküll consider the Darwinian theory a failure. We may admit that the theory of a formation of new species by the cumulative effect of aimless fluctuating variations is not tenable because fluctuating variation is not hereditary; but this would only demand a slight change in the theory; namely a replacement of the influence of fluctuating variation by that of equally aimless mutations. With this slight modification which is proposed by de Vries,5 Darwin’s theory still serves the purpose of explaining how without any pre-established plan only purposeful and harmonious organisms should have survived. It must be said, however, that any theory of life phenomena must be based on our knowledge of the physicochemical constitution of living matter, and neither Darwin nor Lamarck was concerned with this. Moreover, we cannot consider any theory of evolution as proved unless it permits us to transform at desire one species into another, and this has not yet been accomplished.
It may be of some interest to point out that we do not need to make any definite assumption concerning the mechanism of evolution and that we may yet be able to account for the fact that the surviving organisms are to all appearances harmonious. The writer pointed out that of all the 100,000,000 conceivable crosses of teleost fish (many of which are possible) not many more than 10,000, i. e., about one-hundredth of one per cent., are able to live and propagate. Those that live and develop are free from the grosser type of disharmonies, the rest are doomed on account of a gross lack of harmony of the parts. These latter we never see and this gives us the erroneous conception that harmony or “design” is a general character of living matter. If anybody wishes to call the non-viability of 9999100 per cent. of possible teleosts a process of weeding out by “natural selection” we shall raise no objection, but only wish to point out that our way of explaining the lack of design in living nature would be valid even if there were no theory of evolution or if there had never been any evolution.
3. v. Uexküll is perfectly right in connecting the problem of design in an organism with Mendelian heredity. The work on Mendelian heredity has shown that an extremely large number of independently transmissible Mendelian factors help to shape the individual. It is not yet proven that the organism is nothing but a mosaic of Mendelian factors, but no writer can be blamed for considering such a possibility. If we assume that the organism is nothing but a mosaic of Mendelian characters it is difficult indeed to understand how they can force each other into a harmonious whole6; even if we make ample allowance for the law of chance and the corresponding wastefulness in the world of the living. But it is doubtful whether this idea of the rôle of Mendelian factors is correct. The facts of experimental embryology strongly indicate the possibility that the cytoplasm of the egg is the future embryo (in the rough) and that the Mendelian factors only impress the individual (and variety) characters upon this rough block. This idea is supported by the fact that the first development—in the sea urchin to the gastrula stage inclusive—is independent of the nucleus, which is the bearer of the Mendelian factors. Not before the skeleton or mesenchyme is formed in the sea urchin egg is the influence of the nucleus noticeable. This has been shown in the experiments of Boveri in which an enucleated fragment of an egg was fertilized with a spermatozoön of a foreign species. If this is generally true, it is conceivable that the generic and possibly also the species characters of organisms are determined by the cytoplasm of the egg and not by the Mendelian factors.
In any case, we can state today that the cytoplasm contains the rough preformation of the future embryo. This would show then that the idea of the organism being a mosaic of Mendelian characters which have to be put into place by “supergenes” is unnecessary. If the egg is already the embryo in the rough we can imagine the Mendelian factors as giving rise to specific substances which go into the circulation and start or accelerate different chemical reactions in different parts of the embryo, and thereby call forth the finer details characteristic of the variety and the individual. The idea that the egg is the future embryo is supported by the fact that we can call forth a normal organism from an unfertilized egg by artificial means; while it is apparently impossible to cause the spermatozoön to develop into an organism outside the egg.
4. The influence of the whole on the parts is nowhere shown more strikingly than in the field of regeneration. It is known that pieces cut from the plant or animal may give rise to new growth which in many cases will restore somewhat the original organism. Instead of asking what is the cause of this so-called regeneration we may ask, why the same pieces do not regenerate as long as they are parts of the whole. In this form the mysterious influence of the whole over its parts is put into the foreground. We shall see that growth takes place in certain cells when certain substances in the circulation can collect there. The mysterious influence of the whole on these parts consists often merely of the fact that the circulating specific or non-specific substances—we cannot yet decide which—will in the whole be attracted by certain spots and that this will prevent them from acting on other parts of the organism. If such parts are isolated the substances can no longer flow away from these parts and the parts will begin to grow. It thus becomes utterly unnecessary to endow such organisms with a “directing force” which has to elaborate the isolated parts into a whole.
5. The same difficulty which we have discussed in regard to morphogenesis exists also in connection with those instincts which preserve the life of the organism and of the race. The reader need only be reminded of all the complicated instincts of mating by which sperm and eggs are brought together; or those by which the young are prevented from starvation to realize the apparently desperate problems in store for a mechanist, to whom the assumption of design is meaningless. And yet we are better off in regard to our knowledge of the instincts than we are in regard to morphogenesis, as in the former we can show that the apparent instincts in some cases obey simple physicochemical laws with almost mathematical accuracy. Since the validity of the law of gravitation has been proved for the solar system the idea of design in the motion of the planets has lost its usefulness, and this fact must serve us as a guide wherever we attempt to put science beyond the possibility of mysticism. As soon as we can show that a life phenomenon obeys a simple physical law there is no longer any need for assuming the action of non-physical agencies. We shall see that this has been accomplished for one group of animal instincts; namely those which determine the relation of animals to light, since these are being gradually reduced to the law of Bunsen and Roscoe. This law states that the chemical effect of light equals the product of intensity into duration of illumination. Some authors object to the tendency toward reducing everything in biology to mathematical laws or figures; but where would the theory of heredity be without figures? Figures have been responsible for showing that the laws of chance and not of design rule in heredity. Biology will be scientific only to the extent that it succeeds in reducing life phenomena to quantitative laws.
Those familiar with the theories of evolution know the extensive rôle ascribed to the adaptations of organisms. The writer in 1889 called attention to the fact that reactions to light—e. g., positive heliotropism—are found in organisms that never by any chance make use of them; and later that a great many organisms show definite instinctive reactions towards a galvanic current—galvanotropism—although no organism has ever had or ever will have a chance to be exposed to such a current except in laboratory experiments. This throws a different light upon the seemingly purposeful character of animal reactions. Heliotropism depends primarily upon the presence of photosensitive substances in the eye or the epidermis of the organism, and these substances are inherited regardless of whether they are useful or not. It is only a metaphor to call reactions resulting from the presence of photosensitive substances “adaptation.” In this book other examples are given which show that authors have too often spoken of adaptation to environment where the environment was not responsible for the phenomena. The blindness of cave animals and the resistance of certain marine animals to higher concentrations of sea water are such cases. Cuénot speaks of “preadaptation” to express this relation. The fact is that the “adaptations” often existed before the animal was exposed to surroundings where they were of use. This relieves us also of the necessity of postulating the existence of the inheritance of acquired characters, although it is quite possible that the future may furnish proof that such a mode of inheritance exists.
6. We have mentioned that according to Claude Bernard two groups of phenomena occur in the living organism: (1) the phenomena of vital creation or organizing synthesis (especially in the egg and during development); (2) the phenomena of death or organic destruction. These two processes are briefly discussed in the first and last chapters.
These introductory remarks may perhaps make it easier for the reader to retain the thread of the main ideas in the details of experiments and tables given in this book.
1. Each organism is characterized by a definite form and we shall see in the next chapter that this form is determined by definite chemical substances. The same is true for crystals, where substance and form are definitely connected and there are further analogies between organisms and crystals. Crystals can grow in a proper solution, and can regenerate their form in such a solution when broken or injured; it is even possible to prevent or retard the formation of crystals in a supersaturated solution by preventing “germs” in the air from getting into the solution, an observation which was later utilized by Schroeder and Pasteur in their experiments on spontaneous generation. However, the analogies between a living organism and a crystal are merely superficial and it is by pointing out the fundamental differences between the behaviour of crystals and that of living organisms that we can best understand the specific difference between non-living and living matter. It is true that a crystal can grow, but it will do so only in a supersaturated solution of its own substance. Just the reverse is true for living organisms. In order to make bacteria or the cells of our body grow, solutions of the split products of the substances composing them and not the substances themselves must be available to the cells; second, these solutions must not be supersaturated, on the contrary, they must be dilute; and third, growth leads in living organisms to cell division as soon as the mass of the cell reaches a certain limit. This process of cell division cannot be claimed even metaphorically to exist in a crystal. A correct appreciation of these facts will give us an insight into the specific difference between non-living and living matter. The formation of living matter consists in the synthesis of the proteins, nucleins, fats, and carbohydrates of the cells, from the split products. To give an historical example, Pasteur showed that yeast cells and other fungi could be raised on the following sterilized solution: water, 100 gm., crystallized sugar, 10 gm., ammonium tartrate, 0.2 gm. to 0.5 gm., and fused ash from yeast, 0.1 gm.7 He undertook this experiment to disprove the idea that protein or organic matter in a state of decomposition was needed for the origin of new organisms as the defenders of the idea of spontaneous generation had maintained.
2. That such a solution can serve for the synthesis of all the compounds of living yeast cells is due to the fact that it contains the sugars. From the sugars organic acids can be formed and these with ammonia (which was offered in the form of ammonium tartrate) may give rise to the formation of amino acids, the “building stones” of the proteins. It is thus obvious that the synthesis of living matter centres around the sugar molecule. The phosphates are required for the formation of the nucleins, and the work of Harden and Young suggests that they play also a rôle in the alcoholic fermentation of sugar.
Chlorophyll, under the influence of the red rays of light, manufactures the sugars from the CO2 of the air. This makes it appear as though life on our planet should have been preceded by the existence of chlorophyll, a fact difficult to understand since it seems more natural to conceive of chlorophyll as a part or a product of living organisms rather than the reverse. Where then should the sugar come from, which is a constituent of the majority of culture media and which seems a prerequisite for the synthesis of proteins in living organisms?
The investigations of Winogradsky on nitrifying,8 sulphur and perhaps also on iron bacteria have to all appearances pointed a way out of this difficulty. It seemed probable that there were specific micro-organisms which oxidized the ammonia formed in sewage or in the putrefaction of living matter, but the attempts to prove this assumption by raising such a nitrifying micro-organism on one of the usual culture media, all of which contained organic compounds, failed. Led by the results of his observations on sulphur bacteria it occurred to Winogradsky that the presence of organic compounds stood in the way of raising these bacteria, and this idea proved correct. The bacteria oxidizing ammonia to nitrites were grown on the following medium; 1 gm. ammonium sulphate, 1 gm. potassium phosphate, 1 gm. magnesium carbonate, to 1 litre of water. From this medium, which is free from sugar and contains only constituents which could exist on the planet before the appearance of life, the nitrifying bacteria were able to form sugars, fatty acids, proteins, and the other specific constituents of living matter. Winogradsky proved, by quantitative determination, that with the nitrification an increase in the amount of carbon compounds takes place. “Since this bound carbon in the cultures can have no other source than the CO2 and since the process itself can have no other cause than the activity of the nitrifying organism, no other alternative was left but to ascribe to it the power of assimilating CO2.”9 “Since the oxidation of NH3 is the only source of chemical energy which the nitrifying organism can use it was clear a priori that the yield in assimilation must correspond to the quantity of oxidized nitrogen. It turned out that an approximately constant ratio exists between the values of assimilated carbon and those of oxidized nitrogen.” This is illustrated by the results of various experiments as shown in Table I.
TABLE I
| No. 5 | No. 6 | No. 7 | No. 8 | |
| mg. | mg. | mg. | mg. | |
| Oxidized N | 722.0 | 506.1 | 928.3 | 815.4 |
| Assimilated C | 019.7 | 015.2 | 026.4 | 022.4 |
| Ratio N : C | 036.6 | 033.3 | 035.2 | 036.4 |
It is obvious that 1 part of assimilated carbon corresponds to about 35.4 parts oxidized nitrogen or 96 parts of nitrous acid.
These results of Winogradsky were confirmed in very careful experiments by E. Godlewski, Sr.10
The nitrites are further oxidized by another kind of micro-organisms into nitrates and they also can be raised without organic material.
Winogradsky had already previously discovered that the hydrogen sulphide which is formed as a reduction product from CaSO4 or in putrefaction by the activity of certain bacteria can be oxidized by certain groups of bacteria, the sulphur bacteria. Such bacteria, e. g., Beggiatoa, are also commonly found at the outlet of sulphur springs. They utilize the hydrogen sulphide which they oxidize to sulphur and afterwards to sulphates, according to the scheme:
(1) 2H2S + O2 = 2H2O + S2
(2) S2 + 3O2 + 2H2O = 2H2SO4
The sulphuric acid is at once neutralized by carbonates.
Winogradsky assumes that the oxidation of H2S by the sulphur bacteria is the source of energy which plays the same rôle as the oxidation of NH3 plays in the nitrifying bacteria, or the oxidation of carbon compounds—sugar and others—in the case of the other lower and higher organisms. Winogradsky has made it very probable that sulphur bacteria do not need any organic compounds and that their nutrition may be accomplished with a purely mineral culture medium, like that of the nitrite bacteria. On the basis of this assumption they should also be able to form sugars from the CO2 of the air.
Nathanson11 discovered in the sea water the existence of bacteria which oxidize thiosulphate to sulphuric acid. They will develop if some Na2S2O3, is added to sea water. These bacteria can only develop if CO2 from the air is admitted or when carbonates are present. For these organisms the CO2 cannot be replaced by glucose, urea, or other organic substances. Such bacteria must therefore possess the power of producing sugar and starch from CO2 without the aid of chlorophyll. Similar observations were made by Beijerinck on a species of fresh-water bacteria.12
Finally the case of iron bacteria may briefly be mentioned though Winogradsky’s views are not accepted by Molisch.
We may, therefore, consider it an established fact that there are a number of organisms which could have lived on this planet at a time when only mineral constituents, such as phosphates, K, Mg, SO4, CO2, and O2 besides NH3, or SH2, existed. This would lead us to consider it possible that the first organisms on this planet may have belonged to that world of micro-organisms which was discovered by Winogradsky.
If we can conceive of this group of organisms as producing sugar, which in fact they do, they could have served as a basis for the development of other forms which require organic material for their development.
In 1883 the small island of Krakatau was destroyed by the most violent volcanic eruption on record. A visit to the islands two months after the eruption showed that “the three islands were covered with pumice and layers of ash reaching on an average a thickness of thirty metres and frequently sixty metres.”13 Of course all life on the islands was extinct. When Treub in 1886 first visited the island, he found that blue-green algæ were the first colonists on the pumice and on the exposed blocks of rock in the ravines on the mountain slopes. Investigations made during subsequent expeditions demonstrated the association of diatoms and bacteria. All of these were probably carried by the wind. The algæ referred to were according to Euler of the nostoc type. Nostoc does not require sugar, since it can produce that compound from the CO2 of the air by the activity of its chlorophyll. This organism possesses also the power of assimilating the free nitrogen of the air. From these observations and because the Nostocaceæ generally appear as the first settlers on sand the conclusion has been drawn that they or the group of Schizophyceæ to which they belong formed the first settlers of our planet.14 This conclusion is not quite safe since in the settlement of Krakatau as well as in the first colonizing of sand areas the nature of the first settler is determined chiefly by the carrying power of wind (or waves and birds).
We may now return from this digression to the real object of our discussion, namely that the nutritive solutions of organisms must be very dilute and consist of the split products of the complicated compounds of which the organisms consist. The examples given sufficiently illustrate this statement.
The nutritive medium of our body cells is the blood, and while we take up as food the complicated compounds of plants or animals, these substances undergo a digestion, i. e., a splitting up into small constituents before they can diffuse from the intestine into the blood. Thus the proteins are digested down to the amino acids and these diffuse into the blood as demonstrated by Folin and by Van Slyke. From here the cells take them up. The different proteins differ in regard to the different types of amino acids which they contain. While the bacteria and fungi and apparently the higher plants can build up all their different amino acids from ammonia, this power is no longer found in the mammals which can form only certain amino acids in their body and must receive the others through their food. As a consequence it is usually necessary to feed young animals on more than one protein in order to make them grow, since one protein, as a rule, does not contain all the amino acids needed for the manufacture of all the proteins required for the formation of the material of a growing animal.15
3. The essential difference between living and non-living matter consists then in this: the living cell synthetizes its own complicated specific material from indifferent or non-specific simple compounds of the surrounding medium, while the crystal simply adds the molecules found in its supersaturated solution. This synthetic power of transforming small “building stones” into the complicated compounds specific for each organism is the “secret of life” or rather one of the secrets of life.
What clew have we in regard to the nature of this synthetic power? We know that the comparatively great velocity of chemical reactions in a living organism is due to the presence of enzymes (ferments) or to catalytic agencies in general. Some of these catalytic agencies are specific in the sense that a given catalyzer can accelerate the reaction of only one step in a complicated chemical reaction. While these enzymes are formed by the action of the body they can be separated from the body without losing their catalytic efficiency. It was a long time before scientists succeeded in isolating the enzyme of the yeast cell which causes the alcoholic fermentation of sugar; and this gave rise to the premature statement that it was not possible to isolate this enzyme since it was bound up with the life of the yeast cell. Such a statement was even made by a man like Pasteur, who was usually a model of restraint in his utterances, and yet the work of Buchner proved him to be wrong.
The general mechanism of the action of the hydrolyzing enzymes is known. The old idea of de la Rive, that a molecule of enzyme combines transitorily with a molecule of substrate; the further idea, which may possibly go back to Engler, that the molecule of substrate is disrupted in the “strain” of the new combination and that the broken fragments fall off or are easily knocked off by collision from the ferment molecule which is now ready to repeat the process, seems to be correct. On the assumption that the velocity of enzyme reaction is proportional to the mass of the enzyme and that de la Rive’s idea was correct, Van Slyke and Cullen were able to calculate the coefficients of the velocity of enzyme reactions for the fermentation of urea and other substances, and the agreement between calculated and observed values was remarkable.16
While the hydrolytic action of enzymes is thus clear the synthesis in the cell is still a riddle. An interesting suggestion was made by van’t Hoff, who in 1898 expressed the idea that the hydrolytic enzymes should also act in the opposite direction, namely synthetically. Thus it should not only be possible to digest proteins with pepsin but also to synthetize them from the products of digestion with the aid of the same enzyme. This expectation was based on the idea that the enzyme did not alter the equilibrium between the hydrolyzed and non-hydrolyzed part of the substrate but only accelerated the rate with which the equilibrium was reached. Van’t Hoff’s idea omitted, however, the possibility that in the transitory combination between enzyme molecule and substrate a change in the molecular configuration of the substrate or in the distribution of intramolecular strain may take place. The first apparently complete confirmation of van’t Hoff’s suggestion appeared in the form of the synthesis of maltose from grape sugar by the enzyme maltase, which decomposes maltose into grape sugar. By adding the enzyme maltase from yeast to a forty per cent. solution of glucose Croft Hill17 obtained a good yield of maltose. It turned out, however, that what he took for maltose was not this compound but an isomer, namely isomaltose, which has a different molecular configuration and cannot be hydrolyzed by the enzyme maltase.
Lactose is hydrolyzed from kephyr by an enzyme lactase into galactose and glucose; by adding this enzyme to galactose and glucose a synthesis was obtained not of lactose but of isolactose; the latter, however, is not decomposed by the enzyme lactase.
E. F. Armstrong has worked out a theory which tries to account for this striking phenomenon by assuming “that the enzyme has a specific influence in promoting the formation of the biose which it cannot hydrolyze.”18 The theory is very ingenious and seems supported by fact. This then would lead to the result that certain hydrolytic enzymes may have a synthetic action but not in the manner suggested by van’t Hoff.
The principle enunciated by Armstrong, that in the synthetic action of hydrolytic enzymes not the original compound but an isomer is formed which can not be hydrolyzed by the enzyme, may possibly be of great importance in the understanding of life phenomena. It shows us how the cell can grow in the presence of hydrolytic enzymes and why in hunger the disintegration of the cell material is so slow. It was at first thought that the formation of isomers contradicted the idea of the reversible action of enzymes, but this is not the case; on the contrary, it supports it but makes an addition which may solve the riddle of what Claude Bernard called the creative action of living matter. We shall come back to this problem in the last chapter.
Kastle and Loevenhart demonstrated the synthesis of a trace of ethylbutyrate by lipase if the latter enzyme was added to the products of the hydrolysis of ethylbutyrate, ethyl alcohol, and butyric acid by the same enzyme.19 Taylor20 obtained the synthesis of a slight amount of triolein
by the addition of the dried fat-free residue of the castor bean to a mixture of oleinic acid and glycerine. . . . No synthesis occurred with acetic, butyric, palmitic, and stearic acids with glycerine, mannite, and dulcite, and the experiments with the last two alcohols and oleinic acid likewise yielded no synthesis.
This suggests possibly a specific action of the enzyme. If this slight reversible action had any biological significance (which might be possible, since in the organism secondary favourable conditions might be at work which are lacking in vitro) there should be a parallelism between masses of lipase in different kinds of tissues and fat synthesis. Loevenhart indicated that this might be a fact, but a more extensive investigation by H. C. Bradley has made this very dubious.21
Very little is known concerning the reversible action of the hydrolytic protein enzymes. A. E. Taylor digested protamine sulphate with trypsin and found that after adding trypsin to the products of digestion a precipitate was formed after long standing; and we may also refer to experiments of Robertson with pepsin on the products of caseinogen to which we shall return in the next chapter. It therefore looks at present as if van’t Hoff’s idea of reversible enzyme action might hold in the modification offered by Armstrong. It remains doubtful, however, whether this reversibility can explain all the synthetic processes in the cell. No objection can be offered at present if any one makes the assumption that each cell has specific synthetic enzymes or some other synthetic mechanisms which are still unknown.
The mechanisms for the synthesis of proteins must have one other peculiarity: they must be specific in their action. We shall see in the next chapter that each species seems to possess one or more proteins not found in any other but closely related species. Each organism develops from a tiny microscopic germ and grows by synthetizing the non-specific building stones (amino acids) into the specific proteins of the species. This must be the work of the yet unknown synthetic enzymes or mechanisms. The elucidation of their character would seem one of the main problems of biology. Needless to say crystallography is not confronted with problems of such a nature.
The fact that the living cell grows after taking up food has given rise to curious misunderstandings. Traube has shown that drops of a liquid surrounded with a semipermeable membrane may increase in volume when put into a solution of lower osmotic pressure. This has led and is possibly still leading to the statement that the process of growth by a living cell has been imitated artificially. Only one feature has been imitated, the increase in volume; but the essential feature of the process in the living cell, i. e., the formation of the specific constituents of the living cell from non-specific products, has of course not been imitated.
4. The constant synthesis then of specific material from simple compounds of a non-specific character is the chief feature by which living matter differs from non-living matter. With this character is correlated another one; namely, when the mass of a cell reaches a certain limit the cell divides. This is perhaps most obvious in bacteria which on the proper nutritive medium take up food, grow, and divide into two bacteria, each of which takes up food, divides, and grows ad infinitum, as long as the food lasts, provided the harmful products of metabolism are removed. If it be true that specific synthetic ferments exist in each cell it follows that the cell must synthetize these also,22 as otherwise the synthesis of specific proteins would have to come to a standstill.
This problem of synthesis leads to the assumption of immortality of the living cell, since there is no a priori reason why this synthesis should ever come to a standstill of its own accord as long as enough food is available and the proper outside physical conditions are guaranteed. It is well known that Weismann has claimed immortality for all unicellular organisms and for the sex cells of metazoa, while he claimed the necessity of death for the body cells of the latter. Leo Loeb was led by his investigations on the transplantation of cancer to assume immortality not only for the cancer cell but also for the body cell of the organism. He had found in transplanting a malignant tumor from one individual to another that the tumor grew; that it was not the cells of the host but the transplanted tumor cells of the graft which grew and multiplied, and that this process could be repeated apparently indefinitely so that it was obvious that the transplanted tumor cells outlived the original animal. Such experiments have since been carried on so long that we may now say that an individual cancer cell taken from an animal and transplanted from time to time on a new host lives apparently indefinitely. Leo Loeb had found that these tumor cells are simply modified somatic cells. He therefore suggested that the somatic cells might be considered immortal with the same right as we speak of the immortality of the germ cells of such animals.23
This view receives its support first from the fact that certain trees like the Sequoia live several thousand years and may therefore be considered immortal and second, from the method of tissue culture. The method of cultivating tissue cells in a test tube, in the same way as is done for bacteria, was first proposed and carried out by Leo Loeb, in 1897,24 but his test-tube method did not permit the observation of the transplanted cell under the microscope. This was made possible by a modification of the method by Harrison, who established the fact that the axis cylinder grows out from the ganglionic cell. Harrison and Burrows then perfected the method for the cultivation of the cells of warm-blooded, animals, and with the aid of these methods Carrel succeeded in keeping connective-tissue cells of the heart of an early chick embryo alive more than four years, and these cells are still growing and dividing.25 Only very tiny masses of cells can be kept alive in this way since all the cells in the centre of a piece die on account of lack of oxygen; and every two days a few cells from the margin of the piece have to be transferred to a new culture medium.
This effect of lack of oxygen explains also why the immortality of the somatic cells is not obvious. Death in a human being consists in the stopping of heart beat and respiration, which also terminates the action of the brain or at least of consciousness. Immediately after the cessation of heart beat and respiration the cells of muscle and of the skin and probably many or most other organs are still alive and might continue to live if transferred to another body with circulation and respiration. As a consequence of the lack of oxygen supply in the dead body they will, however, die comparatively rapidly. It may be stated that hearts taken out of the body after a number of hours can still beat again when put into the proper solutions and upon receiving an adequate oxygen supply.
The idea that the body cells are naturally immortal and die only if exposed to extreme injuries such as prolonged lack of oxygen or too high a temperature helps to make one problem more intelligible. The medical student, who for the first time realizes that life depends upon that one organ, the heart, doing its duty incessantly for the seventy years or so allotted to man, is amazed at the precariousness of our existence. It seems indeed uncanny that so delicate a mechanism should function so regularly for so many years. The mysticism connected with this and other phenomena of adaptation would disappear if we could be certain that all cells are really immortal and that the fact which demands an explanation is not the continued activity but the cessation of activity in death. Thus we see that the idea of the immortality of the body cell if it can be generalized may be destined to become one of the main supports for a complete physicochemical analysis of life phenomena since it makes the durability of organisms intelligible.
5. This generalized idea of the immortality of some or possibly most or all somatic cells has a bearing upon the problem of the origin of life on our planet. The experiments of Spallanzani, Schwann, Schroeder, Pasteur, Tyndall, and all those who have worked with pure cultures of micro-organisms, have proved that no spontaneous generation of living from non-living matter can be demonstrated; and the statements to the contrary were due to experimental errors inasmuch as the new organisms formed were the offspring of others which had entered into the culture medium by mistake.
In the last chapter of that most fascinating book Worlds in the Making,26 Arrhenius discusses the possibility of life being eternal and of living germs of very small dimensions—e. g., the spores of micro-organisms—being carried through space from one planet to another or even from one solar system to another. If it be true that there is no spontaneous generation; if it be true that all cells are potentially immortal, we may indeed seriously raise the question: May not life after all be eternal? Such ideas were advocated by Richter in a rather phantastic way and more definitely by Helmholtz as well as Kelvin. The latter authors assumed that in the collision of planets or worlds on which there is life, fragments containing living organisms will be torn off and these fragments will move as seed-bearing stones through space. “If at the present instant no life existed upon this earth, one such stone falling upon it might . . . lead to its becoming covered with vegetation.” Arrhenius points out the difficulties which oppose such a view, as, e. g., the fact “that the meteorite in its fall towards the earth becomes incandescent all over its surface and any seeds on it would therefore be deprived of their germinating power.”
Arrhenius suggests another and much more ingenious idea based on the fact that for particles below a certain size the mechanical pressure produced by light waves—the radiation pressure—can overcome the attractive force of gravitation.
Bodies which according to Schwarzschild would undergo the strongest influence of solar radiation must have a diameter of 0.00016 mm. supposing them to be spherical. The first question is therefore: Are there any living seeds of such extraordinary minuteness? The reply of the botanist is that spores of many bacteria have a size of 0.0003 or 0.0002 mm., and there are no doubt much smaller germs which our microscopes fail to disclose.
This assumption is undoubtedly correct.
We will, in the first instance, make a rough calculation of what would happen if such an organism were detached from the earth and pushed out into space by the radiation pressure of our sun. The organism would first of all have to cross the orbit of Mars; then the orbits of the smaller and of the outer planets. . . . The organisms would cross the orbit of Mars after twenty days, the Jupiter orbit after eighty days, and the orbit of Neptune after fourteen months. Our nearest solar system would be reached in nine thousand years.
For the assumption of eternity of life only the transference of germs from one solar system to another would have to be considered and the question arises whether or not germs can keep their vitality so many thousands of years. Arrhenius thinks that this is possible on account of the low temperature (which must be below -220° C.) at which no chemical reaction and hence no decomposition and deterioration are possible in the spores; and on account of the absence of water vapour.
The question then arises: Have we any facts to warrant the assumption that spores may remain alive for thousands of years under such conditions and retain their power of germination? We know that seeds have a very limited vitality, and the statement that grain found in the Egyptian tombs was still able to germinate has long been recognized as a myth. Miss White27 found that in wheat grains, there appeared a well-marked drop in their germinating power after about the fourth year, reaching zero in eleven to seventeen years. In a drier climate they last longer than in a moist climate. It is of importance that the hydrolyzing enzymes in the seeds, such as diastase, erepsin, remained unimpaired even after the germinating power of the seeds had disappeared. The seeds were able to resist for two days the temperature of liquid air, though the subsequent germination was delayed by this treatment. Macfadyen28 exposed non-sporing bacteria, viz., B. typhosus, B. coli communis, Staphylococcus pyogenes aureus, and a Saccharomyces to liquid air.
The experiments showed that a prolonged exposure of six months to a temperature of about -190° has no appreciable effect on the vitality of micro-organisms. To judge by the results there appeared no reason to doubt that the experiment might have been successfully prolonged for a still longer period.
Paul Becquerel29 found that seeds which possess a very thick integument may live longer than the grain in Miss White’s experiments. The thickness of the integument prevents the exchange of gases between air and seed. Thus seeds of leguminoses (Cassia bicapsularis, Cytisus biflorus, Leucæna leucocephala, and Trifolium arvense) had retained their power of germination for eighty-seven years. Becquerel has shown that the dryness of the membrane is very essential for such a duration of life, since when dry it is impermeable for gases and the slow chemical reactions inside the grain become impossible.
In the cosmic space there is no water vapour, no atmosphere, and a low temperature, and there is hence no reason why spores should lose appreciably more of their germinating power in ten thousand years than in six months. We must therefore admit the possibility that spores may move for an almost infinite length of time through cosmic space and yet be ready for germination when they fall upon a planet in which all the conditions for germination and development exist, e. g., water, proper temperature, and the right nutritive substances dissolved in the water (inclusive of free oxygen).
While thus everything is favourable to Arrhenius’s hypothesis, Becquerel raises the objection that the spores going through space would yet be destroyed by ultraviolet light. This danger would probably exist only as long as the germ is not too far from a sun. The difficulty is a real one since the ultraviolet rays have a destructive effect even in the absence of oxygen. It is possible, however, that there are spores which can resist this effect of ultraviolet light. Arrhenius’s theory can not of course be disproved and we must agree with him that it is consistent not only with the theories of cosmogony but also with the seeming potential immortality of certain or of all cells.
The alternative to Arrhenius’s theory is that living matter did originate and still originates from non-living matter. If this idea is correct it should one day be possible to discover synthetic enzymes which are capable of forming molecules of their own kind from a simple nutritive solution. With such synthetic enzymes as a starting point the task might be undertaken of creating cells capable of growth and cell division, at least in the apparently simple form in which these phenomena occur in bacteria; viz., that after the mass has reached a certain (still microscopic) size it divides into two cells and so on. If Arrhenius is right that living matter has had no more beginning than matter in general, this hope of making living matter artificially appears at present as futile as the hope of making molecules out of electrons.
The problem of making living matter artificially has been compared to that of constructing a perpetuum mobile; this comparison is, however, not correct. The idea of a perpetuum mobile contradicts the first law of thermodynamics, while the making of living matter may be impossible though contradicting no natural law.
Pasteur’s proof that spontaneous generation does not occur in the solutions used by him does not prove that a synthesis of living from dead matter is impossible under any conditions. It is at least not inconceivable that in an earlier period of the earth’s history radio-activity, electrical discharges, and possibly also the action of volcanoes might have furnished the combination of circumstances under which living matter might have been formed. The staggering difficulties in imagining such a possibility are not merely on the chemical side—e. g., the production of proteins from CO2, and N—but also on the physical side if the necessity of a definite cell structure is considered. We shall see in the sixth chapter that without a structure in the egg to begin with, no formation of a complicated organism is imaginable; and while a bacterium may have a simple structure, such a structure as it possesses is as necessary for its existence as are its enzymes.
Attempts have repeatedly been made to imitate the structures in the cell and of living organisms by colloidal precipitates. It is needless to point out that such precipitates are of importance only for the study of the origin of structures in the living, but that they are not otherwise an imitation of the living since they are lacking the characteristic synthetic chemical processes.
1. It is a truism that from an egg of a species an organism of this species only and of no other will arise. It is also a truism that the so-called protoplasm of an egg does not differ much from that of eggs of other species when looked at through a microscope. The question arises: What determines the species of the future organism? Is it a structure or a specific chemical or groups of chemicals? In a later chapter we shall show that the egg has a simple though definite structure, but in this chapter we shall see that the egg must contain specific substances and that these substances which determine the “species” and specificity in general are in all probability proteins. Since solutions of different proteins look alike under a microscope we need not wonder that it is impossible to discriminate microscopically between the protoplasm of different eggs.
The idea of definiteness and constancy of species, a matter of daily observation in the case of man and higher animals in general, was not so readily accepted in the case of the micro-organisms, which on account of their minuteness and simplicity of structure are not so easy to differentiate. There existed for a long time serious doubt whether or not the simplest organisms, the bacteria, possessed a definite “specificity” like the higher organisms, or whether they were not endowed, as Warming put it, with an “unlimited plasticity,” which forbade classifying them according to their form into definite species as Cohn had done. An interesting episode in this discussion, which was settled about twenty-five years ago arose concerning the sulphur bacteria, which often develop in large masses on parts of decaying plants or animals along the shore. Sir E. Ray Lankester found collections of red bacteria covering putrefying animal matter in a vessel and forming a continuous membrane along its wall. These red bacteria were of very different shape, size, and grouping, but they seemed to be connected by transition forms. They had a common character, however, namely, their peach-coloured appearance. This common character, together with their association in the same habitat, led Lankester to the then justifiable belief that they all belonged to one species which was protean in character and that the different forms were only to be considered as phases of growth of this one species. The presence of the same red pigment “Bacterio-purpurin” seemed justly to indicate the existence of common chemical processes. Cohn, on the contrary, considered the different forms among these red bacteria (they are today called sulphur bacteria since they oxidize the hydrogen sulphide produced by bacteria of putrefaction to sulphur and sulphates) as definite and distinct species, in spite of their common colour and their association. Later observations showed that Cohn was right. Winogradsky30 succeeded in proving by pure culture experiments that each of these different forms of sulphur bacteria was specific and did not give rise to any of the other forms of the same colour found in the same conditions.
The method of pure line breeding inaugurated by Johannsen31 has shown that the degree of definiteness goes so far that apparently identical forms with only slight differences in size may breed true to this size; but for reasons which will become clear later on we may doubt whether they are to be considered as definite species.
The fact of specificity is supported by the fact of constancy of forms. de Vries has pointed out that regardless of the possible origin of new species by mutation the old species may persevere. Walcott has found fossils of annelids, snails, crustaceans, and algæ in a precambrian formation in British Columbia whose age (estimated on the rate of formation of radium from uranium) may be about two hundred million years and estimated on the basis of sedimentation sixty million years. And yet these invertebrates are so closely related to the forms existing today that the systematists have no difficulty in finding the genus among the modern forms into which each of these organisms belongs. W. M. Wheeler, in his investigations of the ants enclosed in amber, was able to identify some of them with forms living today, though the ants observed in the amber must have been two million years old. The constancy of species, i. e., the permanence of specificity may therefore be considered as established as far back as two or possibly two hundred millions of years. The definiteness and constancy of each species must be determined by something equally definite and constant in the egg, since in the latter the species is already fixed irrevocably.
We shall show first that species if sufficiently separated are generally incompatible with each other and that any attempt at fusing or mixing them by grafting or cross-fertilizing is futile. In the second part of the chapter we shall take up the facts which seem destined to give a direct answer to the question as to the cause of specificity. It is needless to say that this latter question is of paramount importance for the problem of evolution, as well as for that of the constitution of living matter.
2. It is practically impossible to transplant organs or tissues from one species of higher animals to another, unless the two species are very closely related; and even then the transplantation is uncertain and the graft may either fall off again or be destroyed. This specificity of tissues goes so far that surgeons prefer, when a transplantation of skin in the human is intended, to use skin of the patient or of close blood relations. The reason why the tissues of a foreign species in warm-blooded animals cannot grow well on a given host has been explained by the remarkable experiments of James B. Murphy of the Rockefeller Institute.32 Murphy discovered that it is possible to transplant successfully any kind of foreign tissue upon the early embryo of the chick. Even human tissue transplanted upon the chick embryo will grow rapidly. This shows that at this early stage the chick embryo does not yet react against foreign tissue. This lack of reaction lasts until about the twenty-first day in the life of the embryo; then the growth of the graft not only ceases but the graft itself falls off or is destroyed. Murphy noticed that this critical period coincides with the development of the spleen and of lymphatic tissue in the chick and that a certain type of migrating cells, the so-called lymphocytes, which develop in the lymphatic tissue, gather at the edge of the graft in great numbers, and he suggested that these lymphocytes (by a secretion of some substance?) rid the host of the graft. He applied two tests both of which confirmed this idea. First he showed that when small fragments of the spleen of an adult chicken are transplanted into the embryo the latter loses its tolerance for foreign grafts. The second proof is still more interesting. It was known that by treatment with Roentgen rays the lymphocytes in an animal could be destroyed. It was to be expected that an animal so treated would have lost its specific resistance to foreign tissues. Murphy found that this was actually the case. On fully grown rats in which the lymphocytes had been destroyed by X-rays (as ascertained by blood counts) tissues of foreign species grew perfectly well. These experiments have assumed a great practical importance since they can also be applied to the immunization of an animal against transplanted cancer of its own species. Murphy found that by increasing the number of lymphocytes in an animal (which can be accomplished by a mild treatment with X-rays) the immunity against foreign grafts as well as against cancer from the same species can be increased. It is quite possible that the apparent immunity to a transplantation of cancer produced by Jensen, Leo Loeb, and Ehrlich and Apolant through the previous transplantation of tissue in such an animal was due to the fact that this previous tissue transplantation led to an increase in the number of lymphocytes in the animal. The medical side, however, lies outside of our discussion, and we must satisfy ourselves with only a passing notice. The facts show that each warm-blooded animal seems to possess a specificity whereby its lymphocytes destroy transplanted tissue taken from a foreign species.
A lesser though still marked degree of incompatibility exists also in lower animals for grafts from a different species.33 The graft may apparently take hold, but only for a few days, if the species are not closely related. Joest apparently succeeded in making a permanent union between the anterior and posterior ends of two species of earthworms, Lumbricus rubellus and Allolobophora terrestris. Born and later Harrison healed pieces of tadpoles of different species together. An individual made up of two species Rana virescens and Rana palustris lived a considerable time and went through metamorphosis. Each half regained the characteristic features of the species to which it belonged. It seems, however, that if species of tadpoles of two more distant species are grafted upon each other no lasting graft can be obtained, e. g., Rana esculenta and Bombinator igneus. These experiments were made at a time when the nature and bearing of the problem of specificity was not yet fully recognized. The rôle of lymphocytes in these cases has never been investigated. The grafted piece always retained the characteristics of the species from which it was taken.
Plants possess no leucocytes and we therefore see that they tolerate a graft of foreign tissues better than is the case in animals. As a matter of fact heteroc grafting is a common practice in horticulture, although even here it is known that indiscriminate heteroplastic grafting is not feasible and that therefore the specificity is not without influence. The host is supposed to furnish only nutritive sap to the graft and in this respect does not behave very differently from an artificial nutritive solution for the raising of a plant. The law of specificity, however, remains true also for the grafted tissues: neither in animals nor in plants does the graft lose its specificity, and it never assumes the specific characters of the host, or vice versa. The apparent exceptions which Winkler believed he had found in the case of grafts of nightshade on tomatoes turned out to be a further proof of the law of specificity. Winkler, after the graft had taken, cut through the place of grafting, after which operation a callus formation occurred on the wound. In most cases either a pure nightshade or a pure tomato grew out from this callus. In some cases he obtained shoots from the place where graft and host had united, which on one side were tomato, on the other side nightshade. What really happened was that the shoots had a growing point whose cells on the one side consisted of cells of nightshade, on the other side of tomato.34 We know of no case in which the cell of a graft has lost its specificity and undergone a transformation into the cell of the host.
3. Another manifestation of the incompatibility of distant species is found in the domain of fertilization. The eggs of the majority of animals cannot develop unless a spermatozoön enters. The entrance of a spermatozoön into an egg seems also to fall under the law of specificity, inasmuch as in general only the sperm of the same or a closely related species is able to enter the egg. The writer35 has found, however, that it is possible to overcome the limitation of specificity in certain cases by physicochemical means, and by the knowledge of these means we may perhaps one day be able to more closely define the mechanism of specificity in this case. He found that the eggs of a certain Californian sea urchin, which cannot be fertilized by the sperm of starfish in normal sea water, will lose their specificity towards this type of foreign sperm if the sea water is rendered a little more alkaline, or if a little more Ca is added to the sea water, or if both these variations are effected. Godlewski has confirmed the efficiency of this method for the fertilization of sea-urchin eggs with the sperm of crinoids.
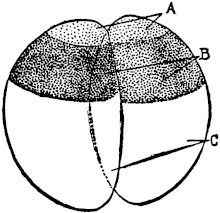
Fig. 1. Five-days-old larvæ from a sea urchin (Strongylocentrotus purpuratus) ♀ and a starfish (Asterias) ♂. (Front view.) |
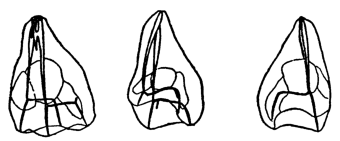
Fig. 2. Five-days-old larvæ of Strongylocentrotus purpuratus produced by artificial parthenogenesis. (Side view.) The larvæ in Figs. 1 and 2 are identical in appearance, proving that heterogeneous hybridization leads to a larva with purely maternal characters. |
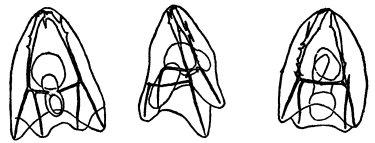
Fig. 3. Five-days-old larvæ of two closely related forms of sea urchins (S. purpuratus ♀ and S. franciscanus ♂). In this case the larva has also paternal characters as shown by the skeleton.
If such heterogeneous hybridizations are carried out, two striking results are obtained. The one is that the resulting larva has only maternal characteristics (Figs. 1 and 2), as if the sperm had contributed no hereditary material to the developing embryo. This result could not have been predicted, for if we fertilize the egg of the same Californian sea urchin, Strongylocentrotus purpuratus, with the sperm of a very closely related sea urchin, S. franciscanus, the hereditary effect of the spermatozoön is seen very distinctly in the primitive skeleton formed by the larva.36 (Fig. 3.) In the case of the heterogeneous hybridization the spermatozoön acts practically only as an activating agency upon the egg and not as a transmitter of paternal qualities.
The second striking fact is that while the sea-urchin eggs fertilized with starfish sperm develop at first perfectly normally they begin to die in large numbers on the second and third day of their development, and only a very small number live long enough to form a skeleton; and these are usually sickly and form the skeleton considerably later than the pure breed. It is not quite certain whether the sickliness of these heterogeneous hybrids begins or assumes a severe character with the development of a certain type of wandering cells, the mesenchyme cells; it would perhaps be worth while to investigate this possibility. The writer was under the impression that this sickliness might have been brought about by a poison gradually formed in the heterogeneous larvæ.
He investigated the effects of heterogeneous hybridization also in fishes, which are a much more favourable object. The egg of the marine fish Fundulus heteroclitus can be fertilized with the sperm of almost any other teleost fish, as Moenkhaus37 first observed. This author did not succeed in keeping the hybrids alive more than a day, but the writer has kept many heterogeneous hybrids alive for a month or longer,38 and found the same two striking facts which he had already observed in the heterogeneous cross between sea urchin and starfish: first, practically no transmission of paternal characters, and second, a sickly condition of the embryo which begins early and which increases with further development. The heterogeneous fish hybrids between, e. g., Fundulus heteroclitus ♀ and Menidia ♂ have usually no circulation of blood, although the heart is formed and beats and blood-vessels and blood cells are formed; the eyes are often incomplete or abnormal though they may be normal at first; the growth of the embryo is mostly retarded. In exceptional cases circulation may be established and in these a normal embryo may result, but such an embryo is chiefly maternal.
This incompatibility of two gametes from different species does not show itself in the case of heterogeneous hybridization only, but also though less often in the case of crossing between two more closely related forms. The cross between the two related forms S. purpuratus ♀ and S. franciscanus ♂ is very sturdy and shows no abnormal mortality as far as the writer’s observations go. If, however, the reciprocal crossing is carried out, namely that of S. franciscanus ♀ and S. purpuratus ♂, the development is at first normal, but beginning with the time of mesenchyme formation the majority of larvæ become sickly and die; and again the question may be raised whether or not the beginning of sickliness coincides with the development of mesenchyme cells. If we assume that the sickliness and death are due to the formation of a poison, we must assume that the poison is formed by the protoplasm of the egg, since otherwise we could not understand why the reciprocal cross should be healthy.
All of these data agree in this one point, that the fusion by grafting or fertilization of two distant species is impossible, although the mechanism of the incompatibility is not yet understood. It is quite possible that this mechanism is not the same in all the cases mentioned here, and that it may be different when two different species are mixed and when incompatibility exists between varieties, as is the case in the graft on mammals.
4. Fifty or sixty years ago surgeons did not hesitate to transfuse the blood of animals into human beings. The practice was a failure, and Landois39 showed by experiment that if blood of a foreign species was introduced into an animal the blood corpuscles of the transfused blood were rapidly dissolved and the animal into which the transfusion was made was rendered ill and often died. The result was different when the animals whose blood was used for the purpose of transfusion belonged to the same species or a species closely related to the animal into which the blood was transfused. Thus when blood was exchanged between horse and donkey or between wolf and dog or between hare and rabbit no hemoglobin appeared in the urine and the animal into which the blood was transfused remained well.40 This was the beginning of the investigations in the field of serum specificity which were destined to play such a prominent rôle in the development of medicine. Friedenthal was able to show later that if to 10 c.c. of serum of a mammal three drops of defibrinated blood of a foreign species are added and the whole is exposed in a test tube to a temperature of 38°C. for fifteen minutes the blood cells contained in the added blood are all cytolyzed; that this, however, does not occur so rapidly when the blood of a related species is used. He could thus show that human blood serum dissolves the erythrocytes of the eel, the frog, pigeon, hen, horse, cat, and even that of the lower monkeys but not that of the anthropoid apes. The blood of the chimpanzee and of the human are no longer incompatible, and this discovery was justly considered by Friedenthal as a confirmation of the idea of the evolutionists that the anthropoid apes and the human are blood relations.41
This line of investigation had in the meanwhile entered upon a new stage when Kraus, Tchistowitch, and Bordet discovered and developed the precipitin reaction, which consists in the fact that if a foreign serum (or a foreign protein) is introduced into an animal the blood serum of the latter acquired after some time the power of causing a precipitate when mixed with the antigen, i. e., with the foreign substance originally introduced into the animal for the purpose of causing the production of antibodies in the latter; while, of course, no such precipitation occurs if the serum of a non-treated rabbit is mixed with the serum of the blood of the foreign species.
In 1897 Kraus discovered that if the filtrates from cultures of bacteria (e. g., typhoid bacillus) are mixed with the serum of an animal immunized with the same serum (e. g., typhoid serum) it causes a precipitate; and that this precipitin reaction is specific. This fact was confirmed and has been extended by the work of many authors.
Tchistowitch in 1899 observed that the serum of rabbits which had received injections of horse or eel serum caused a precipitate when mixed with the serum of these latter animals.
Bordet found in 1899 that if milk is injected into a rabbit the serum of such a rabbit acquires the power of precipitating casein, and Fish found that this reaction is specific inasmuch as the lactoserum from cow’s milk can precipitate only the casein of cow’s milk but not that of human or goat milk. Wassermann and Schütze reached the same result independently of each other.
Myers and later Uhlenhuth showed that if white of egg from a hen’s egg is injected into a rabbit, precipitins for white of egg are found in the serum of the latter, and Uhlenhuth42 found, by trying the white of egg of different species of birds, that the precipitin reaction called forth by the blood of the immunized animal is specific, inasmuch as the proteins from a hen’s egg will call forth the formation of precipitins in the blood of the rabbit which will precipitate only the white of egg of the hen or of closely related birds.
To Nuttall43 belongs the credit of having worked out a quantitative method for measuring the amount of precipitate formed, and in this way he made it possible to draw more valid conclusions concerning the degree of specificity of the precipitin reaction. He found by this method that when the immune serum is mixed with the serum or the protein solution used for the immunization a maximum precipitate is formed, but if it is mixed with the serum of related forms a quantitatively smaller precipitate is produced. In this way the degree of blood relationship could be ascertained. He thus was able to show that when the blood of one species, e. g., the human, was injected into the blood of a rabbit, after some time the serum of the rabbit was able to cause a precipitate not only with the serum of man, or chimpanzee, but also of some lower monkeys; with this difference, however, that the precipitate was much heavier when the immune serum was added to the serum of man. The method thus shows the existence of not an absolute but of a strong quantitative specificity of blood serum. This statement may be illustrated by the following table from Nuttall. The antiserum used for the precipitin reaction was obtained by treating a rabbit with human blood serum. The forty-five bloods tested had been preserved for various lengths of time in the refrigerator with the addition of a small amount of chloroform.
TABLE II
Quantitative Tests with Anti-Primate Sera
Tests with Antihuman Serum
Among the Primate bloods that of the Chimpanzee gave too high a figure, owing to the precipitum being flocculent and not settling well, for some reason which could not be determined. The figure given by the Ourang is somewhat too low, and the difference between Cynocephalus sphinx and Ateles is not as marked as might have been expected in view of the qualitative tests and the series following. The possibilities of error must be taken into account in judging of these figures; repeated tests should be made to obtain something like a constant. Other bloods than those of Primates give small reactions or no reactions at all. The high figures (10%) obtained with two Carnivore bloods can be explained by the fact that one gave a loose precipitum, and the other was a somewhat concentrated serum.44
We have mentioned that even the proteins of the egg are specific according to Uhlenhuth. Graham Smith, one of Nuttall’s collaborators, applied the latter’s quantitative method to this problem and confirmed the results of Nuttall. A few examples may serve as an illustration.
TABLE III
Tests with Anti-Duck’s-Egg Serum
| Material tested | Amount of precipitum | Percentage | |
|---|---|---|---|
| Duck’s | egg-albumin | .0384 | 100 |
| Pheasant’s | " | .0328 | 185 |
| Fowl’s | " | .0234 | 161 |
| Silver Pheasant’s | " | .0140 | 136 |
| Blackbird’s | " | .0065 | 115 |
| Crane’s | " | .0051 | 114 |
| Moorhen’s | " | .0046 | 112 |
| Thrush’s | " | .0046 | 112 |
| Emu’s | " | .0018 | 105 |
| Hedge-Sparrow’s | " | trace | 10? |
| Chaffinch’s | " | · | 100 |
| Tortoise serum | trace | 10? | |
| Turtle serum | " | 10? | |
| Alligator serum | · | 100 | |
Frog, Amphiuma, and Dogfish sera, as well as Tortoise and Dogfish egg-albumins, were also tested, with negative results.
TABLE IV
Tests with Anti-Fowl’s-Egg Serum
| Material tested | Amount of precipitum | Percentage | |
|---|---|---|---|
| Fowl’s | egg-albumin (old) | .0159 | 100 |
| Fowl’s | " (fresh) | .0140 | 188 |
| Silver Pheasant’s | " | .0075 | 147 |
| Pheasant’s | " | .0075 | 147 |
| Crane’s | " | .0046 | 129 |
| Blackbird’s | " | .0046 | 129 |
| Duck’s | " | .0037 | 123 |
| Moorhen’s | " | .0028 | 118 |
Thrush, Emu, Greenfinch, and Hedge-sparrow egg-albumins were tested and gave traces of precipita, as also did Tortoise and Turtle sera. The egg-albumins of the Tortoise, Frog, Skate, and two species of Dogfish did not react. Alligator, Frog, Amphiuma, and Dogfish sera also yielded no results.45
By improving the quantitative method in various ways, Welsh and Chapman46 were able to explain why the precipitin reaction with egg-white was not strictly specific but gave also, though quantitatively weaker, results with the egg-white of related birds. They found that by a new method devised by them “it is possible to indicate in an avian egg-white antiserum the presence of a general avian antisubstance (precipitin) together with the specific antisubstance.”
The Bordet reaction was not only useful in indicating the specificity and blood relationship for animals but also among plants. Thus Magnus and Friedenthal47 were able to demonstrate with Bordet’s method the relationship between yeast (Saccharomyces cerevisiæ) and truffle (Tuber brumale).
5. We must not forget, while under the spell of the problem of immunity, that we are interested at the moment in the question of the nature of the specificity of living organisms. It is only logical to conclude that the fossil forms of invertebrate animals and of algæ and bacteria, which Walcott found in the Cambrian and which may be two hundred million years old, must have had the same specificity at that time as they or their close relatives have today; and this raises the question: What is the nature of the substances which are responsible for and transmit this specificity? It is obvious that a definite answer to this question brings us also to the very problem of evolution as well as that of the constitution of living matter.
There can be no doubt that on the basis of our present knowledge proteins are in most or practically all cases the bearers of this specificity. This has been found out not only with the aid of the precipitin reaction but also with the anaphylaxis reaction, by which, as the reader may know, is meant that when a small dose of a foreign substance is introduced into an animal a hypersensitiveness develops after a number of days or weeks, so that a new injection of the same substance produces serious and in some cases fatal effects. This hypersensitiveness, which was first analysed by Richet,48 is specific for the substance which has been injected. Now all these specific reactions, the precipitin reaction as well as the anaphylactic reaction, can be called forth by proteins. Thus Richet, in his earliest experiments, showed that only the protein-containing part of the extract of actinians, by which he called forth anaphylaxis, was able to produce this phenomenon, and later he showed that it was generally impossible to produce anything resembling anaphylaxis by non-protein substances, e. g., cocain or apomorphin.49 Wells isolated from egg-white four different proteins (three coagulable proteins and one non-coagulable) which can be distinguished from each other by the anaphylaxis reaction, although all come from the same biological object.50 Michaelis as well as Wells found that the split products of the protein molecule are no longer able to call forth the anaphylaxis reaction. Since peptic digestion has the effect of annihilating the power of proteins to call forth anaphylaxis, we are forced to the conclusion that the first cleavage products of proteins have already lost the power of calling forth immunity reactions.
A pretty experiment by Gay and Robertson51 should be mentioned in this connection. Robertson had shown
that a substance closely resembling paranucleins both in its properties and its C, H, and N content can be formed from the filtered products of the complete peptic hydrolysis of an approximately four per cent. neutral solution of potassium caseinate by the action of pure pepsin at 36°C.
He considered this a case of a real synthesis of proteins from the products of its hydrolytic cleavage. This interpretation was not generally accepted and received a different interpretation by Bayliss and other workers. Gay and Robertson were able to show that paranuclein when injected into an animal will sensitize guinea-pigs for anaphylactic intoxication for either paranuclein or casein and apparently indiscriminately. The products of complete peptic digestion of casein had no such effect, but the synthetic product of this digestion obtained by Robertson’s method has the same specific antigenic properties as paranuclein, thus making it appear that Robertson had indeed succeeded in causing a synthesis of paranuclein with the aid of pepsin from the products of digestion of casein by pepsin.
There are a few statements in the literature to the effect that the specificity of organisms might be due to other substances than proteins. Thus Bang and Forssmann claimed that the substances (antigens) responsible for the production of hemolysis were of a lipoid nature, but their statements have not been confirmed, and Fitzgerald and Leathes52 reached the conclusion that lipoids are non-antigenic. Ford claims to have obtained proof that a glucoside contained in the poisonous mushroom Amanita phalloides can act as an antigen. But aside from this one fact we know that proteins and only proteins can act as antigens and are therefore the bearers of the specificity of living organisms.
Bradley and Sansum53 found that guinea-pigs sensitized to beef or dog hemoglobin fail to react or react but slightly to hemoglobin of other origin. The hemoglobins tried were dog, beef, cat, rabbit, rat, turtle, pig, horse, calf, goat, sheep, pigeon, chicken, and man.
6. It would be of the greatest importance to show directly that the homologous proteins of different species are different. This has been done for hemoglobins of the blood by Reichert and Brown,54 who have shown by crystallographic measurements that the hemoglobins of any species are definite substances for that species.
The crystals obtained from different species of a genus are characteristic of that species, but differ from those of other species of the genus in angles or axial ratio, in optical characters, and especially in those characters comprised under the general term of crystal habit, so that one species can usually be distinguished from another by its hemoglobin crystals. But these differences are not such as to preclude the crystals from all species of a genus being placed in an isomorphous series (p. 327).
As far as the genus is concerned it was found that the hemoglobin crystals of any genus are isomorphous.
In some cases this isomorphism may be extended to include several genera, but this is not usually the case, unless as in the case of dogs and foxes, for example, the genera are very closely related.
The most important question for us is the following: Are the differences between the corresponding hemoglobin crystals of different species of the same genus such as to warrant the statement that they indicate chemical differences? If this were the case we might say that blood reactions as well as hemoglobin crystals indicate that differences in the constitution of proteins determine the species specificity and, perhaps, also species heredity. The following sentences by Reichert and Brown seem to indicate that this may be true for the crystals of hemoglobin.
The hemoglobins of any species are definite substances for that species. But upon comparing the corresponding substances (hemoglobins) in different species of a genus it is generally found that they differ the one from the other to a greater or less degree; the differences being such that when complete crystallographic data are available the different species can be distinguished by these differences in their hemoglobins. As the hemoglobins crystallize in isomorphous series the differences between the angles of the crystals of the species of a genus are not, as a rule, great; but they are as great as is usually found to be the case with minerals or chemical salts that belong to an isomorphous group (p. 326).
As Professor Brown writes me, the difficulty in answering the question definitely, whether or not the hemoglobins of different species are chemically different, lies in the fact that there is as yet no criterion which allows us to discriminate between a species and a Mendelian mutation except the morphological differences. It is not impossible that while species differ by the constitution of some or most of their proteins, Mendelian heredity has a different chemical basis.
It is regrettable that work like that of Reichert and Brown cannot be extended to other proteins, but it seems from anaphylaxis reactions that we might expect results similar to those in the case of the hemoglobins. The proteins of the lens are an exception inasmuch as, according to Uhlenhuth, the proteins of the lens of mammals, birds, and amphibians cannot be discriminated from each other by the precipitin reaction.55
7. The serum of certain humans may cause the destruction or agglutination of blood corpuscles of certain other humans. This fact of the existence of “isoagglutinins” seems to have been established for man, but Hektoen states that he has not been able to find any isoagglutinins in the serum of rabbits, guinea-pigs, dogs, horses, and cattle. Landsteiner found the remarkable fact that the sera of certain individuals of humans could hemolyze the corpuscles of certain other individuals, but not those of all individuals. A systematic investigation of this variability led him to the discovery of three distinct groups of individuals, the sera of each group acting in a definite way towards the corpuscles of the representatives of each other group. Later observers, for example Jansky and Moss, established four groups. These groups are, according to Moss,56 as follows:
Group 1. Sera agglutinate no corpuscles.
Corpuscles agglutinated by sera of Groups 2, 3, 4.
Group 2. Sera agglutinate corpuscles of Groups 1, 3.
Corpuscles agglutinated by sera of Groups 3, 4.
Group 3. Sera agglutinate corpuscles of Groups 1, 2.
Corpuscles agglutinated by sera of Groups 2, 4.
Group 4. Sera agglutinate corpuscles of Groups 1, 2, 3.
Corpuscles agglutinated by no serum.
The relative frequency of the four groups follows from the following figures. Of one hundred bloods tested by Moss in series of twenty there were found:
10 belonging to Group 1.
40 belonging to Group 2.
7 belonging to Group 3.
43 belonging to Group 4.
Groups 2 and 4 are in the majority and in overwhelming numbers, which indicates that, as a rule, the sera agglutinate the blood corpuscles of individuals of the other groups, but not those of individuals belonging to the same group. The phenomenon that a serum agglutinates no corpuscles (Group 1), or that the corpuscles are agglutinated by no serum (Group 4), are the exceptions. It is obvious that, as far as our problem is concerned, only Groups 2 and 3 are to be considered. There is no Mendelian character which refers only to one half of the individuals except sex. Since nothing is said about a relation of Groups 2 and 3 to sex such a relation probably does not exist.
8. The facts thus far reported imply the suggestion that the heredity of the genus is determined by proteins of a definite constitution differing from the proteins of other genera. This constitution of the proteins would therefore be responsible for the genus heredity. The different species of a genus have all the same genus proteins, but the proteins of each species of the same genus are apparently different again in chemical constitution and hence may give rise to the specific biological or immunity reactions.
We may consider it as established by the work of McClung, Sutton, E. B. Wilson, Miss Stevens, Morgan, and many others, that the chromosomes are the carriers of the Mendelian characters. These chromosomes occur in the nucleus of the egg and in the head of the sperm. Now the latter consists, in certain fish, of lipoids and a combination of nucleinic acid and protamine or histone, the latter a non-coagulable protein, more resembling a split product of one of the larger coagulable proteins.
A. E. Taylor57 found that if the spermatozoa of the salmon are injected into a rabbit, the blood of the animal acquires the power of causing cytolysis of salmon spermatozoa. When, however, the isolated protamines or nucleinic acid or the lipoids prepared from the same sperm were injected into a rabbit no results of this kind were observed. H. G. Wells more recently tested the relative efficiency of the constituents of the testes of the cod (which in addition to the constituents of the sperm contained the proteins of the testicle). From the testicle he prepared a histone (the protein body of the sperm nucleus), a sodium nucleinate, and from the sperm-free aqueous extract of the testicles a protein resembling albumin was separated by precipitation.58
The albumin behaved like ordinary serum albumin or egg albumin, producing typical and fatal anaphylactic reactions and being specific when tried against mammalian sera. The nucleinate did not produce any reactions when guinea-pigs were given small sensitizing and larger intoxicating doses (0.1 gm.) in a three weeks’ interval; a result to be expected, since no protein is present in the preparation. The histone was so toxic that its anaphylactic properties could not be studied.
It is not impossible that protamines and histones might be found to act as specific antigens if they were not so toxic. The positive results which Taylor observed after injection of the sperm might have been due to the proteins contained in the tail of the spermatozoa, which in certain animals at least does not enter the egg and hence can have no influence on heredity.
It is thus doubtful whether or not any of the constituents of the nucleus contribute to the determination of the species. This in its ultimate consequences might lead to the idea that the Mendelian characters which are equally transmitted by egg and spermatozoön, determine the individual or variety heredity, but not the genus or species heredity. It is, in our present state of knowledge, impossible to cause a spermatozoön to develop into an embryo,59 while we can induce the egg to develop into an embryo without a spermatozoön. This may mean that the protoplasm of the egg is the future embryo, while the chromosomes of both egg and sperm nuclei furnish only the individual characters.
1. We have become acquainted with two characteristics of living matter: the specificity due to the specific proteins characteristic for each genus and possibly species and the synthesis of living matter from the split products of their main constituents instead of from a supersaturated solution of their own substance, as is the case in crystals. We are about to discuss in this and the next chapter a third characteristic, namely, the phenomenon of fertilization. While this is not found in all organisms it is found in an overwhelming majority and especially the higher organisms, and of all the mysteries of animated nature that of fertilization and sex seems to be the most captivating, to judge from the space it occupies in folklore, theology, and “literature.” Bacteria, when furnished the proper nutritive medium, will synthetize the specific material of their own body, will grow and divide, and this process will be repeated indefinitely as long as the food lasts and the temperature and other outside conditions are normal. It is purely due to limitation of food that bacteria or certain species of them do not cover the whole planet. But, as every layman knows, the majority of organisms grow only to a certain size, then die, and the propagation takes place through sex cells or gametes: a female cell—the egg—containing a large bulk of protoplasm (the future embryo) and reserve material; and the male cell which in the case of the spermatozoön contains only nuclear material and no cytoplasmic material except that contained in the tail which in some and possibly many species does not enter the egg. The male element—the spermatozoön—enters the female gamete—the egg—and this starts the development. In the case of most animals the egg cannot develop unless the spermatozoön enters. The question arises: How does the spermatozoön activate the egg? And also how does it happen that the spermatozoön enters the egg? We will first consider the latter question. These problems can be answered best from experiments on forms in which the egg and the sperm are fertilized in sea water. Many marine animals, from fishes down to lower forms, shed their eggs and sperm into the sea water where the fertilization of the egg takes place, outside the body of the female.
The first phenomenon which strikes us in this connection is again a phenomenon of specificity. The spermatozoön can, as a rule, only enter an egg of the same or a closely related species, but not that of one more distantly related. What is the character of this specificity? The writer was under the impression that a clue might be obtained if artificial means could be found by which the egg of one species might be fertilized with a distant species for which this egg is naturally immune. Such an experiment would mean that the lack of specificity had been compensated by the artificial means. It is well known that the egg of the sea urchin cannot as a rule be fertilized with the sperm of a starfish in normal sea water. The writer tried whether this hybridization could not be accomplished provided the constitution of the sea water were changed. He succeeded in causing the fertilization of a large percentage of the eggs of the Californian sea urchin, Strongylocentrotus purpuratus, with the sperm of various starfish (e. g., Asterias ochracea) and Holothurians by slightly raising the alkalinity of the sea water, through the addition of some base (NaOH or tetraethylammoniumhydroxide or various amines), the optimum being reached when 0.6 c.c. N/10 NaOH is added to 50 c.c. of sea water. It is a peculiar fact that this solution is efficient only if both egg and sperm are together in the hyperalkaline sea water. If the eggs and sperm are treated separately with the hyperalkaline sea water and are then brought together in normal sea water no fertilization takes place as a rule, while with the same sperm and eggs the fertilization is successful again if both are mixed in the hyperalkaline solution. From this the writer concluded that the fertilizing power depends on a rapidly reversible action of the alkali on the surface of the two gametes. It was found that an increase of the concentration of calcium in the sea water also favoured the entrance of the Asterias sperm into the egg of purpuratus; and that if CCa was increased it was not necessary to add as much NaOH.
The spermatozoön enters the egg through the so-called fertilization cone, i. e., a protoplasmic process comparable to the pseudopodium of an amœboid cell. The analogy of the process of phagocytosis—i. e., the taking up of particles by an amœboid cell—and that of the engulfing of the spermatozoön by the egg presents itself. We do not know definitely the nature of the forces which act in the case of phagocytosis—although surface tension forces and agglutination have been suggested; both are surface phenomena and are rapidly reversible.
We should then say that the specificity in the process of fertilization consists in a peculiarity of the surface of the egg and spermatozoön which in the case of S. purpuratus ♀ and Asterias ♂ can be supplied by a slight increase in the COH or CCa.
By this method fifty per cent. or more of the eggs of purpuratus could be fertilized with the sperm of the starfish Asterias ochracea, capitata, Ophiurians, and Holothurians, while with the sperm of another starfish, Pycnopodia spuria, only five per cent., and with the sperm of Asterina only one per cent. could be fertilized.60 Godlewski succeeded by the same method in fertilizing the eggs of a Naples starfish with the sperm of a crinoid.61 The writer did not succeed in bringing about the fertilization of the egg of another sea urchin in California, Strongylocentrotus franciscanus, with the sperm of a starfish. Although these eggs formed a membrane in contact with the sperm, the latter did not enter the egg; nor has the writer as yet succeeded in causing the sperm of Asterias to enter the egg of Arbacia.
Kupelwieser62 observed that the spermatozoön of molluscs may occasionally enter into the egg of S. purpuratus in normal sea water and later, at Naples, he observed the same for the sperm of annelids. In these cases no development took place. In teleost fishes the spermatozoön can enter the eggs of widely different species but with rare exceptions all the embryos will die in an early stage of development.63
2. The fact that an increase in the alkalinity or in the concentration of calcium allowed foreign sperm to enter the egg of the sea urchin, suggested the idea that a diminution of alkalinity or calcium in the sea water might block the entrance of the sperm of sea urchin into eggs of their own species. This was found to be correct; when we put eggs and sperm of the same species of sea urchin into solutions whose concentration of Ca or of OH is too small, the sperm, although it may be intensely active, cannot enter the egg.
For the purpose of these experiments the ovaries and testes of the sea urchins were not put into sea water, but instead into pure m/2 NaCl and after several washings in this solution were kept in it (they remain alive for several days in pure m/2 NaCl). Several drops of such sperm and one drop of eggs were in one series of experiments put into 2.5 c.c. of a neutral mixture of m/2 NaCl and 3⁄8 m MgCl2 in the proportion in which these two salts exist in the sea water. In such a neutral solution eggs of Arbacia or purpuratus are not fertilized no matter how long they remain in it, although the spermatozoa swim around the eggs very actively. That no spermatozoön enters the eggs can be shown by the fact that the eggs do not divide (although they can segment in such a solution if previously fertilized in sea water or some other efficient solution). When, however, eggs and sperm are put into 2.5 c.c. of the same solution of NaCl+MgCl2, containing in addition one drop of a N/100 solution of NaOH (or NH3 or benzylamine or butylamine) or eight drops of m/100 NaHCO3, most, and often practically all of the eggs at once form fertilization membranes and segment at the proper time, indicating that fertilization has been accomplished. The same result can be obtained if the eggs are transferred into a neutral mixture of NaCl+MgCl2+CaCl2 (in the proportion in which these salts exist in the sea water) or into a neutral mixture of NaCl+MgCl2+KCl+CaCl2. In such neutral mixtures the eggs form fertilization membranes and begin to segment. The eggs are not fertilized in a neutral solution of NaCl or of NaCl+KCl.64
It is, therefore, obvious that if we diminish the alkalinity of the solution surrounding the egg and deprive this solution of CaCl2 we establish the same block to the entrance of the spermatozoön of Arbacia into the egg of the same species as exists in normal sea water for the entrance of the sperm of the starfish into the egg of purpuratus.
The “block” created in this way, to the entrance of the sperm of Arbacia into the egg of the same species is also rapidly reversible.
We reach the conclusion, therefore, that the specificity which allows the sperm to enter an egg is a surface effect which can be increased or diminished by an increase or diminution in the concentration of OH as well as of Ca. The writer has shown that an increase in the concentration of both substances may cause an agglutination of the spermatozoa of starfish to the jelly which surrounds the egg of purpuratus.65 It is thus not impossible that the specificity which favours the entrance of a spermatozoön into an egg of its own species may consist in an agglutination between spermatozoön and egg protoplasm (or its fertilization cone); and that this agglutination is favoured if the COH or CCa or both are increased within certain limits.
Godlewski discovered a very interesting form of block to the entrance of the spermatozoön into the egg which takes place if two different types of sperm are mixed. He had found that the sperm of the annelid Chætopterus is able to enter the egg of the sea urchin and that in so doing it causes membrane formation. The egg, however, does not develop but dies rapidly, as is the case when we induce artificial membrane formation, as we shall see in the next chapter.
Godlewski found that if the sperm of Chætopterus and the sperm of sea urchins are mixed the mixture is not able to induce development or membrane formation, since now neither spermatozoön can enter; blood has the same inhibiting effect as the foreign sperm. The mixture does not interfere with the development of the eggs if they are previously fertilized.66
The phenomenon was further investigated by Herlant67 who found that if the sperm of a sea urchin is mixed with the sperm of certain annelids (Chætopterus) or molluscs, and if after some time the eggs of the sea urchin are added to the mixture of the two kinds of sperm no egg is fertilized. If, however, the solution is subsequently diluted with sea water or if the egg that was in this mixture is washed in sea water, the same sperm mixture in which the egg previously remained unfertilized will now fertilize the egg. From these and similar observations Herlant draws the conclusion that the block existed at the surface of the egg, inasmuch as a reaction product of the two types of sperm is formed after some time which alters the surface of the egg and thereby prevents the sperm from entering. This view is supported not only by all the experiments but also by the observation of the writer that foreign sperm or blood is able to cause a real agglutination after some time if mixed with the sperm of a sea urchin or a starfish.68 We can imagine that the precipitate forms a film around the egg and acts as a block for the agglutination between egg and spermatozoön. The block can be removed mechanically by washing.
3. The fact has been mentioned that the most motile sperm will not be able to enter into the egg if certain other conditions (specificity or COH or CCa) are not fulfilled. On the other hand, living but immobile sperm cannot enter the egg under any conditions. If we add a trace of KCN to the sperm of Arbacia so that the spermatozoön becomes immobile no egg is fertilized even if the eggs and the sperm are thoroughly shaken together; while the same spermatozoa will fertilize these eggs as soon as the HCN has evaporated and they again become motile. It was formerly thought that the spermatozoön had to bore itself into the egg, being propelled by the movements of the flagellum. It is, however, more probable that only a certain energy of vibration is needed on the part of the spermatozoön to make the latter stick to the surface of the egg and agglutinate and that later forces of a different character bring the spermatozoön into the egg. The fact that under normal conditions a very slight degree of motility on the part of the spermatozoön allows it to enter the egg of its own species seems to favour such a view.
It is a common experience that spermatozoa become very active when they reach the neighbourhood of an egg. v. Dungern assumed that only foreign sperm became thus active, but F. R. Lillie69 has pointed out that this may be a specific effect. The writer tested this idea on the sperm and eggs of two species of starfish and of sea urchins. It should be mentioned that the eggs of the starfish used in this experiment were completely immature and could not be fertilized, while the eggs of the sea urchins were mature. The testicles and ovaries had been kept in NaCl and all the sperm was immotile. Eggs and sperm were mixed together in a pure m/2 NaCl solution where the sperm was only rendered motile by the proximity of eggs. The following table gives the result.70
TABLE V
Specificity of Activation of Sperm by Eggs
| Asterias♂ | Asterina♂ | Franciscanus♂ | Purpuratus♂ | |
|---|---|---|---|---|
| Asterias♀ (immature) | Immediately very motile. | No activation. | Moderately active. | Slight effect in immediate contact with egg. |
| Asterina♀ (immature) | Not motile. | Violent activity. | Violent activity. | Slight effect only near the egg. |
| Franciscanus♀ (mature) | Slightly motile. | No motility. | Immediately active. | Immediately active. |
| Purpuratus♀ (mature) | Slightly motile after some time. | Slight effect in immediate contact with eggs. | Immediately active. | Immediately active. |
The spermatozoa of starfish show a marked specificity inasmuch as they are strongly activated only by the eggs of their own species, although in this experiment these were immature, and to a slight degree only by the eggs of the sea urchin purpuratus. But it is also obvious that the specificity is far from exclusive since the immature eggs of Asterina activate the sperm of the sea urchin franciscanus as powerfully as is done by the mature eggs of the sea urchin purpuratus and franciscanus. In studying these results the reader must keep in mind first that all these experiments were made in a NaCl solution and second that it requires a stronger influence to activate the spermatozoa of the starfish, which are not motile at first even in sea water, than the sea urchin spermatozoa which are from the first very active in such sea water, and which may therefore be considered as being at the threshold of activity in pure NaCl solution.
Wasteneys and the writer (in experiments not yet published) did not succeed in demonstrating an activating effect of the eggs of various marine teleosts upon sperm of the same species.
4. F. R. Lillie71 has studied the very striking phenomenon of transitory sperm agglutination which takes place when the sperm of a sea urchin or of certain annelids is put into the supernatant sea water of eggs of the same species. If we put one or more drops of a very thick sperm suspension of the Californian sea urchin S. purpuratus carefully into the centre of a dish containing 3 c.c. of ordinary sea water and let the drop stand for one-half to one minute and then by gentle agitation mix the sperm with the sea water the mass of thick sperm which is at first rather viscous is distributed equally in sea water in a few seconds and the result is a homogeneous sperm suspension. When, however, the same experiment is made with the sea water which has been standing for a short time over a large mass of eggs of the same species, the thick drop of sperm seems to be less miscible and instead of a homogeneous suspension we get, as a result, the formation of a large number of distinct clusters which are visible to the naked eye and which may possess a diameter of 1 or 2 mm. The rest of the sea water is almost free from sperm. These clusters of spermatozoa may last for from two to ten minutes and then dissolve by the gradual detachment of the spermatozoa from the periphery of the cluster.
This phenomenon seems to occur in sea urchins and annelids. The writer has vainly looked for it in different forms of the Californian starfish or molluscs and in fish at Woods Hole. Lillie failed to find it in the starfish at Woods Hole.
The writer found that the sperm of the Californian sea urchin Strongylocentrotus purpuratus will form clusters with the egg sea water of purpuratus but not with that of franciscanus; while the sperm of franciscanus will agglutinate with the egg sea water of both species, but the clusters last a little longer with the eggs of its own species.
He also found that the clusters are more durable in a neutral than in a slightly alkaline solution and that the agglutination disappears the more rapidly the more alkaline the solution. The presence of bivalent cations, especially Ca, also favours the agglutination.
It was also found that this agglutination occurs only when the spermatozoa are very motile; thus if a trace of KCN is added to a mass of thick sea-urchin sperm so that the spermatozoa become immotile a drop of this sperm will not agglutinate when put in egg sea water of the same species; while later, after the HCN has evaporated, the same sperm will agglutinate when put into such sea water.
The writer suggests the following explanation of the phenomenon. The egg sea water contains a substance which forms a precipitate with a substance on the surface of the spermatozoön whereby the latter becomes slightly sticky. This precipitate is slowly soluble in sea water and the more rapidly the more alkaline (within certain limits). Only when the spermatozoa run against each other with a certain impact will they stick together, as Lillie suggested. Lillie assumes that this agglutinating substance contained in egg sea water is required to bring about fertilization and he therefore calls it “fertilizin.”72 But this assumption seems to go beyond the facts inasmuch as the existence of such an agglutinating substance can only be proved in a few species of animals (sea urchins and annelids); and as, moreover, sea-urchin sperm can fertilize eggs which will not cause the sperm to agglutinate, e. g., the egg of franciscanus can be fertilized by sperm of purpuratus, although the egg sea water of franciscanus causes no agglutination of the sperm of purpuratus. When the jelly surrounding the egg of the Californian sea urchin S. purpuratus is dissolved with acid and the eggs are washed, the eggs will not cause any more sperm agglutination; and yet one hundred per cent. of such eggs can be fertilized by sperm.73
5. It is well known that if an egg is once fertilized it becomes impermeable for other spermatozoa. This cannot well be due to the fact that the egg develops; for the writer found some time ago that eggs of Strongylocentrotus purpuratus which are induced to develop by means of artificial parthenogenesis can be fertilized by sperm. The following observation leaves no doubts in this respect. When the unfertilized eggs of purpuratus are put for two hours into hypertonic sea water (50 c.c. of sea water+8 c.c. 21⁄2 m NaCl) and then transferred into sea water it occasionally happens that a certain percentage of the eggs will begin to divide into 2, 4, 8 or more cells, without developing any further. When to such eggs after they have remained in the resting stage for a number of hours or a day, sperm is added, some or all of the blastomeres form a fertilization membrane and now begin to develop into larvæ; and if the spermatozoön gets into a blastomere of the 2- or 4-cell stage normal plutei will result. When the sperm is added while the eggs are in active parthenogenetic cell division the individual blastomeres into which a spermatozoön enters will also form a fertilization membrane, but such blastomeres perish very rapidly. It is not yet possible to state why it should make such a difference for the possibility of development whether the spermatozoön enters into a blastomere when at rest or when it is in active nuclear division, although the idea presents itself that in the latter case an abnormal mix-up and separation of chromosomes and other constituents may be responsible for the fatal result. Whatever may be the explanation of this phenomenon it proves to us that it is not the process of development in itself which acts as a block to the entrance of a spermatozoön into an egg which is already fertilized.74
When the spermatozoön enters the egg of the sea urchin it calls forth the formation of a membrane—the fertilization membrane. It might be considered possible that this membrane formation or the alteration underlying or accompanying it is responsible for the fact that an egg once fertilized becomes immune against a spermatozoön. We shall see in the next chapter that it is possible to call forth the membrane in an unfertilized sea-urchin egg by treating it with butyric acid. This membrane is so tough in the egg of Strongylocentrotus that no spermatozoön can get through it; in the egg of Arbacia the membrane is occasionally replaced by a soft gelatinous film. If no second treatment is given to such eggs they will disintegrate in a comparatively short time, but when sperm is added some or most of the eggs will develop in the way characteristic of fertilized eggs.75 When the membrane is too tough to allow the spermatozoön to enter the egg it can be shown that if the membrane is torn mechanically the egg can still be fertilized by sperm.
Should it be possible that the spermatozoön can no longer agglutinate with the fertilized egg or that those phagocytotic reactions which we suppose to play a rôle in the entrance of the spermatozoön into the egg are no longer possible after a spermatozoön has entered? The mere fact of development is apparently not the cause which bars a spermatozoön from entering an egg already fertilized by sperm.
Lillie assumes that the egg loses its “fertilizin” in the process of membrane formation since the sea water containing such eggs no longer gives the agglutinin reaction with sperm, and he believes that the lack of “fertilizin” in the fertilized egg or in the egg after membrane formation is the cause of the block in the fertilized egg. But we have seen that the artificial membrane formation does not create such a block although it puts an end to the “fertilizin” reaction. In the egg of purpuratus the “fertilizin” reaction ceases when the jelly surrounding the egg is dissolved by an acid and the eggs are repeatedly washed; yet such eggs can easily be fertilized by sperm.
Lillie does not assume that the “fertilizin” causes an agglutination between egg and spermatozoön—we should assent to such an assumption—but that the “fertilizin” acts like an “amboceptor” between egg and spermatozoön, the latter being the complement, the former the antigen. The pathologist would probably object to this interpretation since no “amboceptor” is needed for agglutination. The writer has had some doubts concerning the value of Ehrlich’s side-chain theory which, besides, can only be applied in a metaphorical sense to the mechanism of the entrance of the spermatozoön into the egg.76
6. The reason that an egg once fertilized with sperm cannot be fertilized again may be found in a group of facts which we will now discuss, namely, the self-sterility of many hermaphrodites. The fact that hermaphrodites are often self-sterile, while their eggs can be fertilized with sperm from a different individual of the same species has played a great rôle in the theories of evolution. We are here only concerned with the mechanism which determines the block to the entrance of a spermatozoön into an egg of the same hermaphroditic individual.
Castle77 observed and studied the phenomenon of self-sterility in an Ascidian, Ciona intestinalis, which is hermaphroditic. Animals which were kept isolated discharged both eggs and sperm into the surrounding sea water. Often no egg was fertilized, but in some cases five, ten, or as many as fifty per cent. of the eggs could be successfully fertilized with sperm from the same individual; while if several individuals were put into the same dish as a rule one hundred per cent. of the eggs which were discharged segmented. Morgan78 found that the eggs of various females differ in their power of being fertilized by sperm of the same individual while one hundred per cent. could usually be fertilized with sperm of a different individual. He found in addition that if the eggs of Ciona are put for about ten minutes into a two per cent. ether solution in sea water in a number of cases the percentage of eggs fertilized by sperm of the same individual shows a slight increase. Fuchs79 has reported results similar to those of Castle and Morgan.
A new point of attack has been introduced into the work of self-sterility in plants by the consideration of heredity. Darwin found that in Reseda which is monœcious (or hermaphroditic) certain individuals are either completely self-sterile or completely self-fertile; and Compton showed that apparently self-fertility is a Mendelian dominant to self-sterility.80
According to Jost this self-sterility in hermaphroditic plants is due to the fact that if pollen of the same plant is used the normal growth of the pollen tube is inhibited, while this inhibition does not exist for pollen from a different individual. Correns calls these substances which prevent the adequate growth of pollen, “inhibitory” substances, and finds that they can apparently be transmitted to the offspring. He made experiments on Cardamine pratensis which is self-sterile.81 He fertilized two individuals of Cardamine crosswise and raised sixty plants of the first generation. He compared the fertility of these F1 plants toward (a) their parents, and (b) foreign plants. All the fertilizations with the foreign plants were successful, but the fertilizations with the parents were only partly successful. According to their reaction they could be divided into four groups:
(A) fertile with both parents. Type bg
(B) fertile with one (B), sterile with the other parent (G).
(a) fertile with B, sterile with G. Type bG
(b) fertile with G, sterile with B. Type Bg
(C) sterile with both parents. Type BG
It was found that approximately fifteen of the sixty children belonged to each of the four groups. This should be expected if the inhibitory substance to each parent is transmitted to the children independently. Half of the children will thus inherit the inhibitory substance of one parent and the other half will inherit the inhibitory substance of the other parent. This agrees with the assumption that there are definite determiners for the inhibitory substances in the children which will be transmitted to half of the children. Rather complicated assumptions are needed to explain all the facts observed by Correns on this basis and since the subject is still under investigation we need not go further into the details.
To us the assumption and experimental support of the idea that self-sterility is caused by the presence of a substance inhibitory to the entrance of a spermatozoön is important. Should it be possible that the block created by the entrance of a spermatozoön into the egg is also due to an inhibitory substance carried by a spermatozoön into the egg; and furthermore that the effect of the inhibitory substance should be the prevention of further agglutination of the spermatozoön with the egg or of the growth of the pollen tube in plants? On such an assumption self-sterility would be due to a lack of agglutination between the egg of a hermaphrodite and a spermatozoön of the same individual. The experiments on the agglutinins have shown that while isoagglutinins (i. e., agglutinins for other individuals of the same species) are common auto-agglutinins (i. e., agglutinins for cells of the same individual) rarely if ever occur.
7. A positive chemotropism of the spermatozoa toward an egg of the same species has been demonstrated in a few cases, but it seems that this phenomenon is not determined by that type of substances which give rise to species specificity. The famous experiment of Pfeffer on the spermatozoa of ferns inaugurates this line of investigation. He found that such spermatozoa when moving in a straight line through the water will be deviated in their course if they come near an archegonium; they will then turn toward it, enter it, and enter the egg. Pfeffer showed that 0.01 per cent. malic acid if put into a capillary tube will attract the spermatozoa of ferns.
When the liquid in the tube contains only 0.01 per cent. malic acid the spermatozoa of ferns very soon move toward the opening of the capillary tube and within from five to ten minutes many hundreds of spermatozoa may accumulate in the tube. The malic acid acts as well in the form of a free acid as in the form of salts.82
These experiments were continued and amplified by Shibata. Bruchmann83 found that the spermatozoa of Lycopodium are positively chemotactic to citric acid and salts of this acid, although no citric acid could be shown in the contents of the archegonia. They are also positively chemotactic to the watery extract from archegonia.
Dewitz, Buller, and the writer have vainly tried to prove the existence of a positive chemotropism of spermatozoa to eggs of the same species. Lillie claims to have proved a positive chemotropism of the sperm of sea urchins to “fertilizin,” but such a conclusion is only justified if a method similar to that of Pfeffer’s with capillary tubes, gives positive results; such a method was not used in Lillie’s experiments. It seems that the fertilization of the egg by sperm is rendered possible by two facts; first that where fertilization takes place outside the body egg and sperm are shed simultaneously by the two sexes. This can be easily observed in the case of fish. But it is also the case in invertebrates. Thus the writer has observed that the sea urchins Strongylocentrotus purpuratus at the shore of Pacific Grove all spawn simultaneously. The examination extended over several miles of shore. At such spawning seasons the sea water becomes a suspension of sperm.
The second fact guaranteeing the fertilization of the eggs is the overwhelming excess of spermatozoa over eggs. The enormous waste in animated nature is in agreement with the idea of a lack of purpose; since in this case the laws of chance must play a great rôle; and the origin of durable organisms by laws of chance is only comprehensible on the basis of an enormous wastefulness, for which evidence is not lacking.
1. The majority of eggs cannot develop unless they are fertilized, that is to say, unless a spermatozoön enters into the egg. The question arises: How does the spermatozoön cause the egg to develop into a new organism? The spermatozoön is a living organism with a complicated structure and it is impossible to explain the causation of the development of the egg from the structure of the spermatozoön. No progress was possible in this field until ways were found to replace the action of the living spermatozoön by well-known physicochemical agencies.84 Various observers such as Tichomiroff, R. Hertwig, and T. H. Morgan had found that unfertilized eggs may begin to segment under certain conditions, but such eggs always disintegrated in their experiments without giving rise to larvæ. In 1899 the writer succeeded in causing the unfertilized eggs of the sea urchin Arbacia to develop into swimming larvæ, blastulæ, gastrulæ, and plutei, by treating them with hypertonic sea water of a definite osmotic pressure for about two hours. When such eggs were then put back into normal sea water many segmented and a certain percentage developed into perfectly normal larvæ, blastulæ, gastrulæ, and plutei.85 Soon afterward this was accomplished by other methods for the unfertilized eggs of a large number of marine animals, such as starfish, molluscs, and annelids. None of these eggs can develop under normal conditions unless a spermatozoön enters. These experiments furnished proof that the activating effect of the spermatozoön upon the egg can be replaced by a purely physicochemical agency.86
The first method used in the production of larvæ from the unfertilized eggs did not lend itself to an analysis of the activating effect of the spermatozoön upon the egg, since nothing was known about the action of a hypertonic solution, except that it withdraws water from the egg; and there was no indication that the entrance of the spermatozoön causes the egg to lose water. No further progress was possible until another method of artificial parthenogenesis was found. When a spermatozoön enters the egg of a sea urchin or starfish or certain annelids, the surface of the egg undergoes a change which is called membrane formation; and which consists in the appearance of a fine membrane around the egg, separated from the latter by a liquid (Figs. 4 and 5). O. and R. Hertwig and Herbst had observed that such a membrane could be produced in an unfertilized egg if the latter was put into chloroform or xylol, but such eggs perished at once. It was generally assumed, moreover, that the process of membrane formation was of no significance in the phenomenon of fertilization, except perhaps that the fertilization membrane guarded the fertilized egg against a further invasion by sperm. However, since the fertilized egg is protected against this possibility by other means the membrane is hardly needed for such a purpose.
 |
 |
| Fig. 4 | Fig. 5 |
| Fig. 4. Unfertilized egg surrounded by spermatozoa (whose flagellum is omitted in the drawing). | |
| Fig. 5. The same egg after a spermatozoön has entered. The fertilization membrane is separated from the egg by a clear space. | |
In 1905 the writer found that membrane formation, or rather the change of the surface of the egg underlying the membrane formation, is the essential feature in the activation of the egg by a spermatozoön. He observed that when unfertilized eggs of the Californian sea urchin Strongylocentrotus purpuratus are put for from one and a half to three minutes into a mixture of 50 c.c. of sea water+2.6 c.c. N/10 acetic or propionic or butyric or valerianic acid and are then put into normal sea water all or the majority of the eggs form membranes; and that such eggs when the temperature is very low will segment once or repeatedly and may even—if the temperature is as low as 4°C. or less—develop into swimming blastulæ87; but they will then disintegrate. On the other hand, if they are kept at room temperature they will develop only as far as the aster formation and nuclear division and then begin to disintegrate. It should be mentioned that the time which elapses between artificial membrane formation and nuclear division is greater than that between the entrance of a spermatozoön and nuclear division.
It was obvious, therefore, that artificial membrane formation induced by butyric acid initiates the processes underlying development of the egg but that for some reason the egg is sickly and perishes rapidly.
When, however, such eggs were given a short treatment with hypertonic sea water or with lack of oxygen or with KCN they developed into normal larvæ. This new or improved method of artificial parthenogenesis is as follows: The eggs are put for from two to four minutes into 50 c.c. sea water containing a certain amount of N/10 butyric acid (2.6 c.c. in the case of S. purpuratus in California and 2.0 c.c. in the case of Arbacia in Woods Hole). Ten or fifteen minutes later the eggs are put into hypertonic sea water (50 c.c. sea water+8 c.c. 21⁄2 m NaCl or Ringer solution or cane sugar) in which they remain, at 15° C. from thirty-five to sixty minutes in the case of purpuratus, and from 171⁄2 minutes to 221⁄2 minutes at 23° in the case of Arbacia at Woods Hole. If the eggs are then transferred to normal sea water they will develop. In making these experiments, which have been repeated and confirmed by numerous investigators, it should be remembered that this effect of the hypertonic solution has a high temperature coefficient (about two for 10° C.) and that a slight overexposure to the hypertonic sea water injures the eggs so that development is abnormal. By this method it is possible to imitate the activating effect of the living spermatozoön upon the egg in every detail and eggs treated in this way will develop in large numbers into perfectly normal larvæ. We shall see later that they can also be raised to the adult state.
2. The next task was to find out the nature of the action of the two agencies upon the development of the egg. It soon became obvious that the membrane formation (or the alteration underlying membrane formation) was the more important of the two, since in the eggs of starfish and annelids this was sufficient for the production of larvæ, and that the second treatment had only the corrective effect, of overcoming the sickly condition in which mere membrane formation had left the eggs. It was, therefore, of great interest to ascertain what substances or agencies caused membrane formation in the egg, since it now became clear that the spermatozoön could only cause membrane formation by carrying one such substance into the egg. These investigations led the writer to the result that all those substances and agencies which are known to cause cytolysis or hemolysis (see Chapter III) will also induce membrane formation, and that the essential feature in the causation of development is a cytolysis of the superficial or cortical layer of the egg. As soon as this layer is destroyed the development of the egg can begin.
The substances and agencies which cause cytolysis and hence, if their action is restricted to the surface of the egg, will induce development are, besides the fatty acids: (1) saponin or solanin or bile salts; (2) the solvents of lipoids, benzol, toluol, amylene, chloroform, aldehyde, ether, alcohols, etc.; (3) bases; (4) hypertonic or hypotonic solutions; (5) rise in temperature, and (6) certain salts, e. g., BaCl2 and SrCl2 in the case of the egg of purpuratus, and according to R. Lillie, NaI or NaCNS in the egg of Arbacia. Whenever we submit an unfertilized sea-urchin egg to any of these agencies and restrict the cytolysis to the superficial or cortical layer of the egg (i. e., if we transfer the egg to normal sea water before the cytolytic agent has had time to diffuse into the main egg) the egg will form a membrane and behave as if the membrane formation had been called forth by a fatty acid, with this difference only, that the various agencies are not all equally harmless for the egg.88
If the idea was correct that the change underlying membrane formation was essentially a cytolysis of the cortical layer of the egg, it was to be expected (from the data contained in Chapter III) that the blood serum or the cell extracts of foreign species would also cause membrane formation and thus induce the development of the unfertilized egg, while serum of animals of the same species or genus would have no such effects. This was found to be correct. In 1907 the writer showed that the blood serum of a Gephyrean worm, Dendrostoma, was able to cause membrane formation in the egg of the sea urchin. When added in a dilution of 1 c.c. of serum to 500 or 1000 c.c. of sea water to eggs of purpuratus a certain number formed fertilization membranes. It was found later that the serum and tissue extracts of a large number of animals, especially of mammals (rabbit, pig, ox, etc.), had the same effect, though it was necessary to use higher concentrations, one-half sea water and one-half isotonic blood serum. The eggs of every female sea urchin, however, did not give the reaction and not all the eggs even of sensitive females formed membranes. The writer found, however, that it was possible to increase the susceptibility of the eggs against foreign blood serum by putting them into a 3⁄8 m solution of SrCl2 for from five to ten minutes (or possibly a little longer) before exposing them to the foreign blood serum. BaCl2 acts similarly. The fact that SrCl2 alone can cause membrane formation in unfertilized eggs if they are left long enough in the solution suggests that the sensitizing effect of the substance consists in a modification of the cortical layer similar to that underlying membrane formation; and that the subliminal effect of a short treatment with SrCl2 and the subliminal effect of the foreign serum when combined suffice to bring about the membrane formation.
Not only the watery extract of foreign cells but also that of foreign sperm, induces membrane formation in the sea-urchin egg. The watery extract of sperm of starfish is especially active, but the degree of activity varies considerably with the species of starfish from which the sperm is taken. The eggs of different species of sea urchins also show a different degree of susceptibility for the sperm of foreign species. Thus the eggs of Strongylocentrotus purpuratus require a higher concentration of sperm extract than the eggs of S. franciscanus. For the latter the amount of foreign cell constituents which suffices to call forth membrane formation is so small that contact with almost any foreign living spermatozoön produces this effect; and as a rule no previous sensitizing action of SrCl2 is required. When we bring the unfertilized eggs of S. franciscanus into contact with the living sperm of starfish or shark or even of fowl, the eggs form a fertilization membrane without previous sensitization. A specific substance from the foreign spermatozoön causes membrane formation before the spermatozoön has time to enter the egg. The effect is the same as if artificial membrane formation had been called forth with butyric acid, i. e., they begin to develop and then disintegrate unless they receive a second short treatment.
When, however, we treat the eggs with the watery extracts from the cells of their own or closely related species we find that these extracts are utterly inactive, even if used in comparatively strong concentrations. This agrees with the results given in Chapter III.
These phenomena lead to a very paradoxical result; namely that while in the case of foreign sperm we can cause membrane formation by both the living and the dead spermatozoön, only the living spermatozoön of the same species can induce membrane formation. This might find its explanation on the assumption that the active substance contained in the foreign sperm or serum is water-soluble and a protein, while the activating or membrane-forming substance in the spermatozoön is insoluble in water but soluble in the egg (or in lipoids). If this assumption is correct the two substances are essentially different.
Robertson89 has succeeded in extracting a substance from the sperm of the sea urchin which causes membrane formation of the sea-urchin egg after the latter has been sensitized by a treatment with SrCl2. It seems to the writer that if the substance extracted by Robertson were the real fertilizing agent contained in the spermatozoön it should fertilize the egg without a previous sensitization of the egg with SrCl2 being required.
3. The action of acids in the mechanism of artificial parthenogenesis provides some interesting physiological problems. When unfertilized sea-urchin eggs are left in sea water containing any of the lower fatty acids up to capronic, the eggs will form no membranes, while in such sea water, and they will show no outer signs of cytolysis (swelling). When, however, the eggs are left in sea water containing any of the fatty acids from heptylic upward the eggs will form membranes while in the acid sea water and soon afterward will cytolyze completely and swell enormously. In solutions of the mineral acids no membranes are formed and none are formed as a rule when the eggs are transferred back to sea water. When both a mineral and a lower fatty acid, e. g., butyric, are added to sea water the mineral acid acts as if it were not present, i. e., the eggs form membranes when transferred back to sea water if the concentration of the butyric acid is high enough. All these data are comprehensible if we assume that only that part of the acid causes membrane formation which is lipoid soluble, while the water soluble part is not involved in the process of membrane formation; and that the cytolysis or swelling of the whole egg can only take place in the higher fatty acids (heptylic or above) which are little soluble in water and very soluble in lipoids, while the lower fatty acids, whose water solubility is comparatively high, can only bring about a cytolysis and swelling in the cortical layer but not in the rest of the egg. This makes it appear as though the part undergoing an alteration in membrane formation was a lipoid; and this would harmonize with the assumption that the specific membrane-inducing substance in the spermatozoön is not soluble in water, but soluble in fat.
4. These and other observations led the writer to the view that the essential process which causes development might be an alteration of the surface of the egg, in all probability an alteration of the superficial layer probably of the nature of a superficial cytolysis. The question remains: What could be the physicochemical nature of this cytolysis? The writer had suggested in former papers that in the cytolysis underlying membrane formation lipoids were dissolved, and he supposed that the substance to be dissolved might be a calcium-lipoid compound which might form a continuous layer under the surface of the egg.90 v. Knaffl, working on the cytolysis of eggs in the writer’s laboratory, gave the following idea of the process:
Protoplasm is rich in lipoids; probably it is mainly an emulsion of these and proteins. Any physical or chemical stimulus which can liquefy the lipoids causes cytolysis of the egg. The protein of the egg can really only swell or be dissolved if the condition of aggregation of the lipoid is altered by chemical or physical agencies. The mechanism of cytolysis consists in the liquefaction of the lipoids and thereupon the lipoid-free protein swells or is dissolved by taking up water. . . . Hence this supports Loeb’s view that membrane formation is induced by the liquefaction of lipoids.91
The writer suggested that the destruction of an emulsion in the cortical layer might possibly be the essential feature of the alteration leading to membrane formation and development. It had been long observed that unfertilized starfish eggs may begin to develop apparently without any outside “stimulus,” and A. P. Mathews found that slight mechanical agitation of these eggs in sea water increased the number which developed. It has been shown in numerous experiments by Delage, R. S. Lillie, and the writer, that the substances causing development in the starfish egg are identical or closely related to those which bring about this effect in the egg of the sea urchin and in both cases the development is preceded by a membrane formation.
But how can membrane formation be produced by mere agitation? It seems to me that this can be understood if we suppose that it depends upon the destruction of an emulsion in the cortical layer of the egg. It is conceivable that in the egg of certain forms the stability of this emulsion is so small that mere shaking would be enough to destroy it and thus induce membrane formation and development.92
The durability of emulsions varies, and where an emulsion is very durable shaking has no effect, while where it is at the critical point of separating into two continuous phases a slight shaking will bring about the separation, and where the emulsion is still less durable we observe the phenomenon of a “spontaneous” parthenogenesis. Eggs like those of most sea urchins belong to the former, eggs like those of some starfish and annelids belong to the second or third type.
It is impossible to state at present whether the fertilization membrane is preformed in the fertilized egg and merely lifted off from the egg or whether its formation is due to the hardening of a colloidal substance separated from the emulsion (or excreted) and hardened in touch with sea water. But we can be sure of one thing, namely, that the liquid between egg and membrane contains some colloidal substance which determines the tension and spherical shape of the membrane. The membrane is obviously permeable not only to water but also to dissolved crystalloids, while it is impermeable to colloids. When we add some colloidal solution (e. g., white of egg, blood serum, or tannic acid) to the sea water containing fertilized eggs of purpuratus, the membrane collapses and lies close around the egg; while if the eggs are put back into sea water or a sugar solution the membrane soon assumes its spherical shape. This is intelligible on the assumption that in the process of membrane formation (or in the destruction of the emulsion in the cortical layer) a colloidal substance goes into solution which cannot diffuse into the sea water since the membrane is impermeable to the colloidal particles. The membrane is, however, permeable to the constituents of sea water or to sugar. Consequently sea water will diffuse into the space between membrane and egg until the tension of the membrane equals the osmotic pressure of the colloid dissolved in the space between egg and the membrane. If we add enough colloid to the outside solution so that its osmotic pressure is higher than that of the colloidal solution inside the membrane the latter will collapse.
It should also be stated that the unfertilized eggs of many marine animals are surrounded by a jelly (chorion) which is dissolved when the egg is fertilized.93 The writer has shown that the same chemical substances which will induce membrane formation and artificial parthenogenesis will as a rule also cause a swelling and liquefaction of the chorion.
We have devoted so much space to the mechanism of membrane formation since it is likely to give a clearer insight into the physicochemical nature of physiological processes than the phenomena of muscular stimulation and contraction or nerve stimulation, upon which the majority of physiologists base their conclusions concerning the mechanism of life phenomena.
Before we come to the discussion of the second factor in the activation of the egg it should be stated more definitely that for the eggs of some forms the first factor, the process underlying membrane formation, suffices for the development of the egg into a larva and that no second factor is required in these cases. This is true for the eggs of starfish and certain annelids. Thus in 1901 Loeb94 and Neilson showed that a short treatment with HCl and HNO3 sufficed to cause some eggs of Asterias in Woods Hole to develop into larvæ without a second treatment being needed, and Delage95 showed the same for CO2; and in 1905 the writer found that the eggs of the Californian starfish Asterina can be induced to form a membrane by butyric acid treatment and that ten per cent. of these eggs developed into normal larvæ. Quite recently R. S. Lillie observed that the eggs of Asterias at Woods Hole can be caused to form membranes and develop into larvæ by a treatment with butyric acid and that the time of exposure required to get a maximal number of larvæ varies approximately inversely with the concentration of the acid, within a range of 0.0005 to 0.006 N butyric acid. If the exposure is too short membrane formation will occur without normal development.96
All this leads us to the conclusion that the main effect of the spermatozoön in inducing the development of the egg consists in an alteration of the surface of the latter which is apparently of the nature of a cytolysis of the cortical layer. Anything that causes this alteration without endangering the rest of the egg may induce its development. The spermatozoön, therefore, causes the development of the egg by carrying a substance into the latter which effects an alteration of its surface layer.
5. We will now discuss the action of the second, corrective factor, in the inducement of development. When we cause membrane formation in a sea-urchin egg by the proper treatment with butyric acid it will commence to develop and segment but will disintegrate rapidly if kept at room temperature and the more rapidly the higher the temperature. If, however, the eggs are treated afterward for a certain length of time (from thirty-five to sixty minutes at 15° C. for purpuratus and 171⁄2 to 221⁄2 minutes for Arbacia at 23° C.) in a solution which is isosmotic with 50 c.c. sea water+8 c.c. 21⁄2 m NaCl,97 they will develop into larvæ, many of which may be normal. Any hypertonic solution of this osmotic pressure, sea water, sugar, or a single salt, will suffice provided the solution does not contain substances that are too destructive for living matter. The hypertonic solution produces its corrective effect only if the egg contains free oxygen; and in a slightly alkaline medium more rapidly than in a neutral medium. The time of exposure in the hypertonic solution diminishes in certain limits with the concentration of OH ions in the solution.
It is strange that in the eggs of purpuratus the corrective effect can also be brought about by exposing the eggs after the artificial membrane formation for about three hours to normal sea water free from oxygen; or to sea water in which the oxidations have been retarded by the addition of KCN. This method is not so reliable as the treatment with hypertonic solution.
What does the hypertonic solution do to prevent the disintegration of the egg after the artificial membrane formation? The writer suggested in 1905 that the artificial membrane formation alone starts the development but leaves the eggs usually in a sickly condition and that the hypertonic solution or the lack of oxygen allows them to recuperate from such a condition. The second factor is, according to this view, merely a corrective or curative factor. The following observations will explain the reasons for such an assumption.
The writer found that if we keep the unfertilized eggs after artificial membrane formation in sea water deprived of oxygen the disintegration of the egg following artificial membrane formation is prevented for a day at least. The same result can be obtained by adding ten drops of 1⁄10 per cent. KCN to 50 c.c. of sea water, and certain narcotics, e. g., chloral hydrate, act in the same way. Wasteneys and the writer found that chloral hydrate (and other narcotics) in the concentration required do not suppress or even lower the oxidations in the egg to any considerable extent,98 but they prevent the processes of cell division. Hence it seems that the egg disintegrates so rapidly after artificial membrane formation because it is killed by those processes leading to nuclear division or cell division which are induced by the artificial membrane formation. If we suppress these phenomena of development (for not too long a time) we give the egg a chance to recover and if now the impulse to develop is still active we notice a perfectly normal development. If the egg is kept too long without oxygen it suffers for other reasons and cannot develop; the writer has shown that if eggs fertilized by sperm are kept for too long a time without oxygen they also will no longer be able to develop normally. The short treatment with a hypertonic solution supplies the corrective factor required, so that the egg can then undergo cell division at room temperature without disintegrating.
The correctness of this interpretation, which is in reality mainly a statement of observations, is proved by the two following groups of facts. The older observers had already noticed that the unfertilized eggs of the sea urchin when lying in sea water will die after a day or more, and that occasionally such eggs show nuclear division or even the beginning of cell division shortly before disintegration sets in. The writer has studied this phenomenon in the unfertilized eggs of purpuratus and found that only the eggs of certain females show this cell division before disintegration and that the cell division is preceded by an atypical form of membrane formation; the eggs surrounding themselves by a fine gelatinous film comparable to that produced in the egg of Arbacia by a treatment with butyric acid. It is difficult to state what induces the alteration of the surface in the eggs that lie so long in sea water. It may be due to the CO2 formed by the eggs—since we know that CO2 may induce membrane formation—or it may be due to the alkalinity of the sea water or to a substance originating from the jelly surrounding the eggs. It was found that if such eggs are kept without oxygen their disintegration (and cell division) will be delayed considerably. The presumable explanation for this is that the lack of oxygen prevents the internal changes underlying cell division and thus prevents the disintegration of the egg. The direct proof that an egg in the process of cell division is more endangered by abnormal solutions than an egg at rest has been furnished by numerous observations of the writer. He showed in 1906 that the fertilized egg of purpuratus dies rather rapidly in a pure m/2 NaCl or any other abnormal isotonic solution, while the unfertilized egg can live for days in such solutions.99 In a series of papers, beginning in 1905, he showed that the fertilized egg will live longer in hypertonic, hypotonic, and otherwise abnormally constituted solutions when the cell divisions are suppressed by lack of oxygen or by the addition of KCN or of chloral hydrate.100 It is thus obvious that coincident with the changes underlying nuclear division or cell division alterations occur in the sensitiveness of the egg to salt solutions of abnormal concentration or constitution, e. g., NaCl+CaCl2 isotonic with sea water, hypertonic, or hypotonic solutions.
We must, therefore, conclude that artificial membrane formation induces development but that it leaves the egg in a sickly condition in which the very processes leading to cell division bring about its destruction; that if it is given time it can recover from this condition and that the treatment with the hypertonic solution also brings about this recovery rapidly and reliably.
Herlant101 suggested that the corrective effect of the hypertonic solution consisted in the proper development of the astrospheres required for cell division. According to this author mere membrane formation does not lead to the formation of sufficiently large astrospheres and hence cell division may remain impossible.102 The writer has no a priori objection to this suggestion which agrees with earlier observations by Morgan except that it is at present difficult to harmonize it with all the facts. Why should it be possible to replace the treatment with the hypertonic solution by a suspension of the oxidations in the egg for three hours while we know that lack of oxygen suppresses the formation of astrospheres in the fertilized eggs? What becomes of the astrospheres if the treatment with the hypertonic solution precedes the membrane formation by a number of hours or a day (which is possible as we shall see), and why do they not induce cell division, if Herlant’s idea is correct? Nevertheless the suggestion of Herlant deserves to be taken into serious consideration.
6. How can an alteration of the surface of the egg—e. g., a cytolytic or other destruction of the cortical layer—lead to a beginning of development? The answer is possibly given in the relation of oxidation to development. The writer found in 1895 that if oxygen is withdrawn from the fertilized sea-urchin egg it can not segment and this seems to be the case for eggs in general.103 In 1906 he found that the rapid disintegration of the eggs of the sea urchin which follows artificial membrane formation could be prevented when the eggs were deprived of oxygen or when the oxidations were suppressed in the eggs by KCN. This suggested a connection between the disintegration of the egg after artificial membrane formation and the increase in the rate of oxidations; and he found further that the formation of acid is greater in the fertilized than in the unfertilized egg. He, therefore, expressed the view in 1906 that the essential feature (or possibly one of the essential features) of the process of fertilization was the increase of the rate of oxidations in the egg and that this increase was caused by the membrane formation alone.104 These conclusions have been since amply confirmed by the measurements of O. Warburg as well as those of Loeb and Wasteneys, both showing that the entrance of the spermatozoön into the egg raises the rate of oxidations from 400 to 600 per cent., and that membrane formation alone brings about an increase of similar magnitude. Loeb and Wasteneys found that the hypertonic solution does not increase the rate of oxidations in a fertilized egg. It does do so, however, in an unfertilized egg without membrane formation, but merely for the reason that in such an egg the hypertonic solution brings about the cytolytic change in the cortex of the egg underlying membrane formation.105 According to Warburg it is probable that the oxidations occur mainly if not exclusively at the surface of the egg since NaOH, which does not diffuse into the egg, raises the rate of oxidations more than NH4OH which does diffuse into the egg. And finally, the same author showed that the oxidations in the sea-urchin egg are due to a catalytic process in which iron acts as a catalyzer.106 In view of all these facts and their harmony with the methods of artificial parthenogenesis the suggestion is justifiable that the alteration or cytolysis of the cortical layer of the egg is in some way connected with the increased rate of oxidations.
The question remains then: How can membrane formation or the alteration of the cortical layer underlying membrane formation cause an increase in the rate of oxidations? One possibility is that the iron (or whatever the nature of the catalyzer may be) exists in the cortex of the egg in a masked condition—or in a condition in which it is not able to act—while the alteration of the cortical layer makes the iron active. It might be that either the iron or the oxidizable substrate is contained in the lipoid layer in the unfertilized condition of the egg and that the destruction or cytolysis of the cortical layer brings both the iron and the oxidizable substrate into the watery phase in which they can interact.
Another possibility is that the act of fertilization increases the permeability of the egg. This idea, which seems attractive, was first suggested and discussed by the writer in 1906.107 He had found that when fertilized and unfertilized eggs were put into abnormal salt solutions, e. g., pure solutions of NaCl, the fertilized eggs died more rapidly than the unfertilized eggs and he pointed out that these experiments suggested the possibility that fertilization increases the permeability of the egg for salts. The reason for his hesitation to accept this interpretation was, that the fertilized egg is also more easily injured by lack of oxygen than the unfertilized egg and in this case the greater sensitiveness of the fertilized egg was obviously due to its greater rate of metabolism. Later experiments by the writer showed that the fertilized egg can be made more resistant to abnormal salt solutions if its development is suppressed by lack of oxygen or by KCN or by certain narcotics. With our present knowledge it does not seem very probable that lack of oxygen diminishes the permeability of the egg, but we know that it inhibits the developmental processes. Warburg has made it appear very probable that the fertilized egg is impermeable for NaOH and if this is the case it should also be impermeable for NaCl.108
The idea that fertilization and membrane formation cause an increase in the permeability of the egg was later accepted and elaborated by R. Lillie. This author assumes that the unfertilized egg cannot develop because it contains too much CO2 but that the CO2 can escape from the egg as soon as its permeability is increased through the destruction of the cortical layer of the egg.109 After the CO2 has escaped, the excessive permeability must be restored to its normal value and this is the rôle of the hypertonic treatment. It is, however, difficult to harmonize the assumption of an impermeability of the unfertilized egg for CO2 with the fact that if the unfertilized sea-urchin egg is cut into two, as is done in merogony, no development takes place, while such pieces will develop when a spermatozoön enters. The cortical layer is removed along the cut surface and there is no reason why the CO2 should not escape. Besides, the experiments of Godlewski and the writer prove that the cortical layer of the unfertilized sea-urchin egg is apparently very permeable for CO2 since the latter causes membrane formation if contained in the sea water in sufficiently high concentration.
Lillie assumes that the hypertonic treatment restores the permeability raised to excess by the butyric acid treatment, but this assumption is not in harmony with the following facts. The writer has shown that it is immaterial whether the eggs are treated first with the hypertonic solution and then with butyric acid or the reverse, if only the eggs remain longer in the hypertonic solution when the hypertonic treatment precedes the butyric acid treatment. It was stated in the beginning of this chapter that the development of the egg can be induced by hypertonic sea water, and we know the reason since hypertonic sea water is a cytolytic agency. The writer found that when we expose unfertilized eggs of purpuratus for from two to two and a half-hours to hypertonic sea water they will often not develop and only a few eggs will undergo the first cell divisions, then going into a condition of rest. When these eggs, both the segmented and unsegmented, were treated twenty-four or thirty-six hours later with butyric acid, so that they formed a membrane, they all developed into larvæ without further treatment. It is impossible to apply Lillie’s theory to these facts, for the simple reason that the treatment with hypertonic sea water was just long enough to induce development in some eggs and hence according to Lillie’s ideas must have increased the permeability of these eggs. Yet these same eggs were induced to develop normally when subsequently treated with butyric acid, which according to Lillie also acts by increasing the permeability. Nothing indicates that the treatment of the eggs with a hypertonic solution diminishes their permeability; the reverse would be much more probable.
Lillie’s theory also fails to explain that mere treatment of the eggs with a hypertonic solution can bring about their development into larvæ. This, however, is intelligible on the assumption that the hypertonic solution in this case has two different effects, first a cytolysis of the cortical layer of the egg and second an entirely different effect, possibly upon the interior of the egg, which represents the second or corrective effect.
McClendon110 has shown that the electrical conductivity of the egg is increased after fertilization, and J. Gray111 has found that this increase in conductivity is only transitory and disappears in fifteen minutes. This might indicate that the egg becomes transitorily more permeable for salts after the entrance of the spermatozoön or after membrane formation; although an increase in conductivity might be caused by other changes than a mere increase in permeability of the egg. The writer is of the opinion that it is necessary to meet all these and other difficulties before we can state that the alteration of the cortical layer, which is the essential feature of development, acts chiefly or exclusively by an increase in the permeability of the egg.112
7. When the experiments on artificial parthenogenesis were first published they aroused a good deal of antagonism not only among reactionaries in general but also among a certain group of biologists. O. Hertwig had defined fertilization as consisting in the fusion of two nuclei, the egg nucleus and the sperm nucleus. No such fusion of two nuclei takes place in artificial parthenogenesis since no spermatozoön enters the egg, and it became necessary, therefore, to abandon Hertwig’s definition as wrong. The objection raised that the phenomena are limited to a few species soon became untenable since it has been possible to produce artificial parthenogenesis in the egg of plants (Fucus, according to Overton) as well as of animals, from echinoderms up to the frog; and it may possibly one day be accomplished also in warm-blooded animals. A second objection was that the eggs caused to develop by the methods of artificial parthenogenesis could never reach the adult stage and that hence the phenomenon was merely pathological. There was no basis for such a statement, except that it is extremely difficult to raise marine invertebrates. Delage113 was courageous enough to make an attempt to raise parthenogenetic larvæ of the sea urchin beyond the larval stage and he succeeded in one case in carrying the animal to the mature stage. It proved to be a male.
Better opportunities were offered when a method was discovered which induced the development of the unfertilized eggs of the frog. In 1907, Guyer made the surprising observation that if he injected lymph or blood into the unfertilized eggs of frogs he succeeded in starting development and he even obtained two free-swimming tadpoles. “Apparently the white rather than the red corpuscles are the stimulating agents which bring about development, because injections of lymph which contains only white corpuscles produce the same effects as injections of blood.” Curiously enough, Guyer thought that probably the cells which he introduced and not the egg were developing. In 1910, Bataillon showed that a mere puncture of the egg with a needle could induce development but he believes that for the full development the introduction of a fragment of a leucocyte is required. Bataillon has called attention to the analogy with the writer’s results on lower forms, the puncturing of the egg corresponding to the cytolysis of the surface layer of the egg and the introduction of a leucocyte as the analogue of the second or corrective factor. The method of producing artificial parthenogenesis by puncturing the egg has thus far been successful only in the egg of the frog. The writer has tried it in vain on the eggs of many other forms. He has at present seven parthenogenetic frogs over a year old, produced by merely puncturing the eggs with a fine needle (Fig. 6). These frogs have reached over half the size of the adult frog. They can in no way be distinguished from the frogs produced by fertilization with a spermatozoön. This makes the proof conclusive that the methods of artificial parthenogenesis can result in the production of normal organisms which can reach the adult stage.
Bancroft and the writer tried to determine the sex of a parthenogenetic tadpole and of a frog just carried through metamorphosis. Since in early life the sex glands of both sexes in the frog contain eggs it is not quite easy to determine the sex, except that in the male the eggs gradually disappear and from this and other criteria we came to the conclusion that both parthenogenetic specimens, which were four months old, were males.
The writer has recently examined the gonads of a ten months old parthenogenetic frog. Here no doubt concerning the sex was possible since the gonads were well-developed testicles containing a large number of spermatozoa of normal appearance, and no eggs.114 (Figs. 7 and 8.) This would indicate that the frog belongs to those animals in which the male is heterozygous for sex.
8. The fact that the egg of so high a form as the frog can be made to develop into a perfect and normal animal without a spermatozoön—although normally the egg of this form does not develop unless a spermatozoön enters—corroborates the idea expressed in previous chapters that the egg is the future embryo and animal; and that the spermatozoön, aside from its activating effect, only transmits Mendelian characters to the egg. The question arises: Is it possible to cause a spermatozoön to develop into an embryo? The idea has been expressed that the egg was only the nutritive medium on which the spermatozoön developed into an embryo, but this idea has been rendered untenable by the experiments on artificial parthenogenesis. Nevertheless the question whether or not the spermatozoön can develop into an embryo on a suitable culture medium remains, and it can only be decided by direct experiments. It was shown by Boveri, Morgan, Delage, Godlewski, and others, that if a spermatozoön enters an enucleated egg or piece of egg it can develop into an embryo, but since the cytoplasm of the egg is the future embryo this experiment proves only that the egg nucleus may be replaced by the sperm nucleus; and also that the sperm nucleus carries into the egg, the substances which induce development. Incidentally these experiments on merogony also prove that the mere mechanical tearing of the cortical layer,—which must happen in the separation of the unfertilized egg into parts with and without a nucleus,—by dissection or by shaking, is not sufficient to start development in the sea-urchin egg.
J. de Meyer put the spermatozoa of sea urchins into sea water containing an extract of the eggs of the same species but found only that the spermatozoa swell in such a solution. Loeb and Bancroft made extensive experiments in cultivating spermatozoa of fowl in vitro on suitable culture media. In yolk and white of egg the head of the spermatozoön underwent transformation into a nucleus, but no mitosis or aster formation was observed.115 These experiments should be continued.
1. The writer in a former book (Dynamics of Living Matter, 1906, p. 1), defined living organisms as chemical machines consisting chiefly of colloidal material and possessing the peculiarity of preserving and reproducing themselves. Some authors like Driesch, and v. Uexküll seem to find it impossible to account for the development of such machines from an undifferentiated egg on a purely physicochemical basis. A study of Driesch’s very interesting and important book116 shows that he assumes the eggs of certain animals, e. g., the sea urchin, to consist of homogeneous material; and he concludes that nature has solved, in the formation of highly differentiated organisms from such undifferentiated material, a problem which does not seem capable of a solution by physicochemical agencies alone. But the supposition of a structureless egg is wrong, since Boveri has demonstrated the existence of a very simple but definite structure in the unfertilized egg of the sea urchin; and a similar simple structure has been demonstrated by other authors, especially Conklin, in the eggs of other forms.
In this chapter we shall attempt the task among others of showing how, on the basis of the simple physicochemical structure of the unfertilized egg, the main organ of self-preservation of the organism, the intestine, is formed through the mere process of cell division and growth. Cell division is the most general of the specific functions of living matter and it is the basis underlying the differentiation of the comparatively simple structure of the egg into a more complex organism. If cell division and growth were equal in all parts of the egg no differentiation would be possible, but the different regions of the unfertilized egg contain different constituents and these, probably on account of their chemical difference, do not all begin to grow or divide simultaneously and equally.
Fig. 9
Boveri117 found that in the unfertilized egg of the sea urchin Strongylocentrotus lividus at Naples a definite structure is indicated by the fact that the yellowish-red pigment is not equally distributed over the whole surface of the egg but is arranged in a wide ring from the equator almost to one of the poles. Thus three zones can be recognized in the egg (Fig. 9), a small clear cap A at one pole, a pigmented ring B, and the rest again unpigmented C. Observation has shown that each one of these regions gives rise to a definite constituent of the egg: A furnishes the mesenchyme from which the skeleton and the connective tissue originate; B is the material for the formation of the intestine, and C gives rise to the ectoderm.
The pigment is only at the surface of the egg, and its collection at B indicates only that the material in B differs physicochemically from A and C. The real determiners of the three different groups of organs are three different groups of substances whose distribution is approximately but probably not wholly identical with the regions indicated by distribution of pigment. The intestine-forming material is probably not entirely lacking in C but is contained here in a lower concentration and probably the more so the greater the distance from B; and the same may probably be said for the substances determining mesenchyme and ectoderm formation. Hence the unfertilized egg contains already a rough preformation of the embryo inasmuch as the main axis of the embryo and the arrangement of its first organs are determined.
Fig. 11 |
After the egg is fertilized the cell divisions begin. The first division is as a rule at right angles to the stratification of the egg, each of the two cells contains one-half of the pigment ring (and of each of A and C) (Fig. 10), and after the next division each contains one-fourth of the pigmented part. Each of the four cells is a diminutive whole egg since each contains the three layers in the normal arrangement (Fig. 11). The next divisions bring about an unequal division of the material. Four cells will be formed of ectoderm material C and only little intestine material B, the other four cells containing B and A. These latter form at the next division four very small colourless cells, the so-called micromeres, A (Fig. 12), from which the mesenchyme, skeleton, and connective tissue are formed, four larger cells, B, from which the intestine is formed, and eight cells, C, from which the ectoderm will arise. The separation of the three groups of substances is probably not as complete as our purely diagrammatic drawing (Fig. 12) indicates.
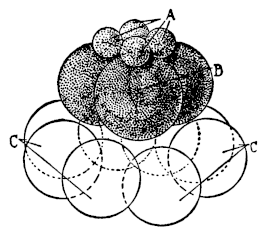
Fig. 12
The cell division proceeds and the cells become smaller and smaller and all gather at the surface of the egg, thus forming a hollow sphere. It is not known what brings about this gathering of the cells at the surface, whether it is protoplasmic creeping or streaming or whether the cells are held by a jelly-like layer which covers the surface of the egg (hyaline membrane) (Fig. 13). Then the cilia are formed at the external surface of these cells and the egg begins to swim; we say it has reached the first larval, the so-called blastula stage. This happens according to Driesch after the tenth series of cell divisions, when the number of cells is theoretically 1024, in reality not quite so many (between 800 and 900). The next step consists in the cells derived from the material A (mesenchyme and micromeres) gliding into the hollow sphere, where they form a ring, the physicochemical process responsible for this gliding being yet unknown. At the opening of this ring an active growing of the cells of the entoderm into the hollow sphere takes place and the hollow cylinder formed by this growth is the intestine (Fig. 14). Why the cells grow into the hollow sphere and not into the opposite direction is unknown. The next step is the formation of a skeleton by the formation of crystals consisting of the CaCO3 by the mesenchyme cells surrounding the intestine. For the establishment of the principle in which we are interested the description of morphogenesis need not be carried farther.
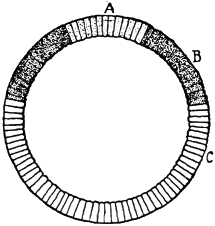
Fig. 13 |
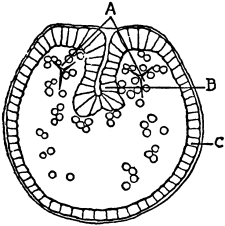
Fig. 14 |
This principle which is under discussion here is the development of a purposeful arrangement of organs out of the egg. If we assume that the egg consists of homogeneous material we are indeed confronted with a riddle. Since the facts contradict such an assumption but show, as Boveri has pointed out, a prearrangement which allows us to indicate in the unfertilized egg already the exact spot where the intestine will grow into the blastula cavity, we are on solid physicochemical ground, although many questions of detail cannot yet be answered. Such a preformation as Boveri has demonstrated is only conceivable if the material of the egg has not too high a degree of fluidity; we may consider it as consisting essentially of a semi-solid gel which is not homogeneous throughout the egg but divided into three strata.
2. Lyon118 tried to ascertain whether by centrifuging the sea-urchin egg it was possible to modify its structure and thereby affect the later embryo. He and subsequent experimenters found that it only is possible to change the position of the nucleus and the distribution of the pigment in the egg. It follows from this that the nucleus and the pigment are suspended in rather fluid material, the former in the centre, the pigment at or near the surface. The position of the nucleus determines the first plane of segmentation, since the nuclear division precedes the division of the cytoplasm of the egg and the plane of nuclear division becomes also the plane of the division of the whole egg—a point which need not be discussed here. It was found, however, by Lyon and the subsequent investigators that the place where the micromeres are formed and where the intestine of the embryo later originates is little influenced by the centrifuging of the egg. The localization of this spot must therefore be determined by a structure sufficiently solid not to be shifted by the centrifugal force. The intestinal stratum in the egg contains the forerunners of the tissues which secrete hydrolyzing enzymes, e. g., trypsin into the digestive tract.
When the surrounding solution is altered in constitution or when the temperature is too high, the intestine instead of growing into the hollow sphere grows outside, we get an evagination instead of an invagination of the intestine. Such larvæ may live for a few days but they cannot grow into a living organism. The forces which make the intestine grow into the hollow sphere are unknown; it may possibly be only the difference between the tension on the external and internal surfaces of the hollow sphere; under normal conditions, the resistance on the inner surface being smaller, the intestine grows into the hollow sphere.
The intestine is one of the organs required for the self-preservation of a more complicated organism, in fact a higher organism without a digestive tract is not capable of living for any length of time. In the gastrula—i. e., the blastula with an intestine—we have an organism which is durable, but the processes leading up to the formation of the intestine are so simple that it is difficult to understand why the assumption of a “supergene” should be required in this case.
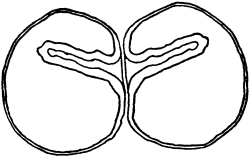
Fig. 15
3. Driesch119 was the first to show that if we isolate one of the first two cells of a dividing egg each develops into a whole embryo of half size. This is perfectly intelligible, since each of the two cells contains all the three layers in the normal arrangement (Fig. 10). The cells divide and the cells having the tendency to creep to the surface of the mass arrange themselves in a hollow sphere, the blastula. Since micromeres and intestine material are present and in their normal position an intestine will grow into the blastula and a whole organism will result. All of this is as necessary as is the formation of one embryo from the whole egg material. Yet the two half-embryos betray their origin from two cleavage cells of the same egg, in that the two gastrulæ formed are often if not always symmetrical to each other (Fig. 15), as the writer had a chance to observe in the egg of Strongylocentrotus purpuratus120 in the following experiment. The eggs of the sea urchin Strongylocentrotus purpuratus are put soon after fertilization into solutions which differ from sea water in two points; namely that they are neutral or very faintly acid (through the CO2 absorbed from the air) instead of being faintly alkaline, and second, that one of the following three constituents of the sea water is lacking; namely: K, Na, or Ca. When the eggs are allowed to segment in such a solution the first two cleavage cells are as a rule in a large percentage of cases—often as many as ninety per cent.—separated from each other, and when the eggs are put into normal sea water (about twenty minutes after the cell division) each cell develops into a normal embryo. In a number of cases the embryos remained inside the egg membrane and did not move until after the invagination of the intestine was far advanced; in such cases it was found quite often that the invagination began at the plane of cleavage at symmetrical points of the two embryos, and the growth of the intestine was symmetrical in both embryos.
This symmetry is probably due to the following fact: the first cleavage plane goes through that spot where the intestine grows into the blastula cavity. If the micromere material does not change its position after the two cleavage cells are separated and the new blastulæ do not become completely spherical the symmetry which we observed is bound to occur. The occurrence is a confirmation of Boveri’s observation. It is natural that Driesch also found that each cell in the four-cell stage should give rise to a full embryo, since each of these cells is in reality a diminutive egg containing the three strata in the right arrangement. When, however, the cells of the eight- or sixteen-cell stage were isolated Driesch’s results were different. In this case the isolated cells from the ectoderm material did no longer all form a gastrula; when such a cell still formed a gastrula it was probably due to the fact that it contained some entoderm material; while the cells taken from the entoderm region all formed embryos and therefore contained ectoderm material.121 The isolated ectoderm cells of a blastula could no longer form an intestine; they were lacking the entoderm material. It looks as if a gradual migration of all the entoderm material from the ectoderm into the entoderm took place during the blastula formation.
When the contents of the egg are displaced by pressure the result will be determined by the location of the main mass of the intestine-forming material; where the main mass of this body is located the invagination of the intestine will take place. In his earlier work Driesch assumed from pressure experiments that the egg had a great power of “regulation.” In a later paper122 he expressed to a large extent his agreement with Boveri who denied this power of “regulation” and showed that the existence of the structure of the egg—i. e., a division into three strata, one forming the ectoderm, the second the entoderm, and the third the mesoderm—was sufficient to explain the various phenomena of apparent “regulation.” Driesch’s idea of a regulation in this case has often been used to insist upon the non-explicability of the phenomena of development from a purely physicochemical viewpoint. It is, therefore, only fair to point out that Boveri123 has furnished the facts for a simpler explanation, which seems to have escaped the notice of antimechanists.124
The objection may be raised that in accepting Boveri’s facts and interpretation we pushed the miracle only one step farther and that we now have to explain the origin of the structure in the unfertilized egg. This Boveri has done by showing that the egg grows from the wall of the ovary and that that part of the egg which is connected with the wall of the ovary gives rise to the ectoderm layer, while the opposite part gives rise to the mesenchyme and the intestine. This shows a connection between the orientation of the egg in the wall of the ovary and its stratification. While this does not solve the problem of stratification in the egg it gives the clue to its solution.
The ultimate origin of stratification probably goes back to the fact of the presence of watery and water-immiscible substances, such as fats. The experiments by Beutner and the writer have shown that the electromotive forces which are observed in living tissues originate at the boundaries between a watery and a water-immiscible phase, like oleic acid or lecithin.125 In his earlier writings126 the writer had thought that the colloids had special significance and this idea seems to prevail today; but the actual observations have shown that the phase boundary fat-water is of greater importance. Needless to say the fats if not present in the cell from the beginning can be formed in the metabolism.
4. All the “regulation” in the egg is of a purely physicochemical character; it consists essentially of a flow of material. If this idea is correct, the apparent power of “regulation” of the blastomeres should differ according to the degree of fluidity and the possibility of different layers separating, and this assumption is apparently supported by facts. The first plane of segmentation of the egg is usually the plane of symmetry of the later organism and where the degree of fluidity is less than in the sea-urchin egg, a separation of the two first blastomeres should easily result in the formation of two half-embryos instead of two whole embryos.
This is the case for the frog’s egg as Roux showed in a classical experiment. Roux destroyed one of the two first cleavage cells of a frog’s egg with a hot needle and found that as a rule the surviving cell developed into only a half-embryo.127 The frog’s egg consists of two substances, a lighter one which is on top and a heavier one below. Although viscous, the two substances are not too viscous to prevent a flow if the egg is turned upside down. O. Schultze found that if a normal egg is turned upside down in the two-cell stage and held in that position, two full embryos arise, one from each of the two blastomeres. Through the flow of the lighter liquid in the egg upwards the two halves of the protoplasm on top become separated and develop independently into two whole embryos instead of into two half-embryos. In Roux’s experiment this flow of protoplasm was avoided. Morgan showed that if Roux’s experiment is repeated with the modification that the egg is put upside down after the destruction of the one cell, the intact cell will give rise not to a half but to a whole embryo.128 These experiments prove that each of the first two cleavage cells of the frog’s egg represents one-half of the embryo and that a whole embryo can develop from each half only when a redistribution of material takes place, which in the egg of the frog can be brought about by gravitation since the egg consists of a lighter and a heavier mass.
When, therefore, in the egg of the sea urchin each of the first two blastomeres naturally gives rise to a whole embryo it is due to a greater degree of fluidity of the protoplasm and not to a lack of preformation of the embryo in the cytoplasm. This idea is confirmed by the observations on the egg of Ctenophores whose cytoplasm seems to be more solid than that of most other eggs. Chun found that the isolated blastomere of the first cell division produced a half-larva, possessing only four instead of the eight locomotor plates of the normal animal.
It seems that in the egg of molluscs, also, the simple symmetry relations of the body are already preformed. It is well known that there are shells of snails which turn to the right while others turn in the opposite direction. The shells of Lymnæus turn to the right, those of Planorbis to the left. It was observed by Crampton129, Kofoid, and Conklin that the eggs of right-wound snails do not segment in a symmetrical, but in a spiral, order, and that in left-handed snails the direction of the spiral segmentation is the reverse of that of the segmentation in the right-handed snails. Conklin was able to show that the asymmetrical spiral structure is already preformed in the egg before cleavage. The asymmetry of the body in snails is therefore already preformed in the egg.130
E. B. Wilson131 has found a marked differentiation in the eggs of some annelids and molluscs. He isolated the first two blastomeres of the egg of Lanice, an Annelid. These two blastomeres are somewhat different in size; from the larger one of the first two blastomeres, the segmented trunk of the worm originates. Wilson found that
when either cell of the two-cell stage is destroyed, the remaining cell segments as if it still formed a part of an entire embryo.132 The later development of the two cells differs in an essential respect, and in accordance with what we should expect from a study of the normal development. The posterior cell develops into a segmented larva with a prototroch, an asymmetrical pre-trochal or head region, and a nearly typical metameric seta-bearing trunk region, the active movements of which show that the muscles are normally developed. The pre-trochal or head region bears an apical organ, but is more or less asymmetrical, and, in every case observed, but a single eye was present, whereas the normal larva has two symmetrically placed eyes. The development of the anterior cell contrasts sharply with that of the posterior. This embryo likewise produces a prototroch and a pre-trochal region, with an apical organ, but produces no post-trochal region, develops no trunk or setæ, and does not become metameric. Except for the presence of an apical organ, these anterior embryos are similar in their general features to the corresponding ones obtained in Dentalium. None of the individuals observed developed a definite eye, though one of them bore a somewhat vague pigment spot.
This result shows that from the beginning of development the material for the trunk region is mainly localized in the posterior cell; and, furthermore, that this material is essential for the development of the metameric structure. The development of this animal is, therefore, to this extent, at least, a mosaic work from the first cleavage onward—a result that is exactly parallel to that which I earlier reached in Dentalium, where I was able to show that the posterior cell contains the material for the mesoblast, the foot, and the shell; while the anterior cell lacks this material. I did not succeed in determining whether, as in Dentalium, this early localization in Lanice pre-exists in the unsegmented egg. The fact that the larva from the posterior cell develops but a single eye, suggests the possibility that each of the first two cells may be already specified for the formation of one eye; but this interpretation remains doubtful from the fact that the larva from the anterior cell did not, in the five or six cases observed, produce any eye.
Conklin has established the existence of a definite structure in the unfertilized eggs of Ascidians, Amphioxus, and many molluscs. In all cases the results of the isolation of the first blastomeres seem to agree with the demonstrable structure of the unfertilized egg.
5. These examples may suffice to show that the egg has from the beginning a simple structure, and we will now point out by which means further differentiation may come about. Sachs suggested that all differentiation and the formation of every organ presupposes the previous existence of specific substances responsible for the formation. These substances which are now called internal secretions or hormones develop gradually during embryonic development. What exists first is a jelly-like block of protoplasmic material with a varying degree of viscosity and with just enough differentiation to indicate head and tail end, a right and left, and a dorsal and ventral side of the future embryo.
Aside from such simple differences phenomena of protoplasmic streaming contribute to the further differentiation. Such streaming begins, according to Conklin,133 in the egg just before fertilization when the surface layer of the egg protoplasm
streams to the point of entrance of the sperm, and these movements may lead to the segregation of different kinds of plasma in different parts of the egg and to the unequal distribution of these substances in different regions of the egg.
One of the most striking cases of this is found in the Ascidian Styela in which there are four or five different kinds of substances in the egg which differ in colour, so that their distribution to different regions of the egg and to different cleavage cells may be easily followed and even photographed while in the living condition. The peripheral layer of protoplasm is yellow and when it gathers at the lower pole of the egg where the sperm enters it forms a yellow cap. This yellow substance then moves following the sperm nucleus, up to the equator of the egg on the posterior side and there forms a yellow crescent extending around the posterior side of the egg just below the equator. On the anterior side of the egg a grey crescent is formed in a somewhat similar manner and at the lower pole between these two crescents is a slate-blue substance, while at the upper pole is an area of colourless protoplasm. The yellow crescent goes into cleavage cells which become muscle and mesoderm, the grey crescent into cells which become nervous system and notochord, the slate-blue substance into endoderm cells, and the colourless substance into ectoderm cells.
Thus within a few minutes after the fertilization of the egg and before or immediately after the first cleavage, the anterior and posterior, dorsal and ventral, right and left poles are clearly distinguishable, and the substances which will give rise to ectoderm, endoderm, mesoderm, muscles, notochord, and nervous system are plainly visible in their characteristic positions.134
We may finally allude briefly to the fact that when once a number of tissues are differentiated each one may influence the other by calling forth tropistic reactions. Thus the writer showed that in the yolk sac of the fish Fundulus the pigment cells lie at first without any definite order but that they gradually are compelled to creep entirely on the blood-vessels and form a sheath around them with the result that the yolk sac assumes a tiger-like marking.135 Driesch136 has pointed out that the mesenchyme cells are directed in their migration; and it seems that the direction of the growth of the axis cylinder is determined by the tissues into which it grows. The idea of tropistic reactions in the formation of organs has been discussed by Herbst.137
6. As a consequence of further changes definite anlagen or buds originate later in the embryo which are destined to give rise to definite organs. Thus in the tadpole early mesenchyme cells are formed which are the anlagen for the four legs, which will grow out under the proper conditions. These anlagen are specific inasmuch as from the anlage of a foreleg only a foreleg, and from the anlage for a hindleg only a hindleg, will develop. Braus138 has proved this by transplanting the anlage of a foreleg to different parts of the body. No matter into which part of the body they are transplanted the mesenchyme cells for the foreleg will give rise to a foreleg only; even if they are transplanted into the spot from which the hindlegs grow out under natural conditions. There is therefore nothing to indicate “regulation.”
The same is true for the formation of the eye and probably in general. We have to consider the formation of the various organs of the body as being due to the development of specific cells in definite locations in the organisms which will grow out into definite organs no matter into which part of the organism they are transplanted. It is at present unknown what determines the formation of these specific anlagen. They may lie dormant for a long time and then begin to grow at definite periods of development. We shall see later that we know more about the conditions which cause them to grow.
7. The fact that the egg, and probably every cell, has a definite structure should determine the limits of the divisibility of living matter. In most cases the complete destruction of a cell means the cessation of life phenomena. A brain or kidney which has been ground to a pulp is no longer able to perform its functions; yet we know that such pulps can still perform some of the characteristic chemical processes of the organ; e. g., the alcoholic fermentation characteristic of yeast can be caused by the press juice from yeast; or characteristic oxidations can be induced by the ground pulp of organs. The question arises as to how far the divisibility of living matter can be carried without interfering with the total of its functions. Are the smallest particles of living matter which still exhibit all its functions of the order of magnitude of molecules and atoms, or are they of a different order? The first step toward obtaining an answer to this question was taken by Moritz Nussbaum,139 who found that if an infusorian be divided into two pieces, one with and one without a nucleus, only the piece with a nucleus will continue to live and perform all the functions of self-preservation and development which are characteristic of living organisms. This shows that at least two different structural elements, nucleus and cytoplasm, are needed for life. We can understand to a certain extent from this why an organ after being reduced to a pulp, in which the differentiation into nucleus and protoplasm is definitely and permanently lost, is unable to accomplish all its functions.140
The observations of Nussbaum and those who repeated his experiments showed that although two different structures are required, not the whole mass of an infusorian is needed to maintain its life. The question then arose: How small a fraction of the original cell is required to permit the full maintenance of life? The writer tried to decide this question in the egg of the sea urchin. He had found a simple method by which the eggs of the sea urchin (Arbacia) can easily be divided into smaller fragments immediately after fertilization. When the egg is brought from five to ten minutes after fertilization (long before the first segmentation occurs) into sea water which has been diluted by the addition of equal parts of distilled water, the egg takes up water, swells, and causes the membrane to burst. Part of the protoplasm then flows out, in one egg more, in another less. If these eggs are afterward brought back into normal sea water those fragments which contain a nucleus begin to divide and develop.141 It was found that the degree of development which such a fragment reaches is a function of its mass; the smaller the piece, the sooner as a rule its development ceases. The smallest fragment which is capable of reaching the pluteus stage possesses the mass of about one-eighth of the whole egg. Boveri has since stated that it was about one twenty-seventh of the whole mass. Inasmuch as only the linear dimensions are directly measurable, a slight difference in measurement will cause a great discrepancy in the calculation of the mass. Driesch’s results disagree with the statement of Boveri and support the observation of the writer.
If we raise the question why such a limit exists in regard to the divisibility of living matter, it seems probable that only those fragments of an egg are capable of development into a pluteus which contain a sufficient amount of material of each of the three layers. If this be correct, it would certainly not suffice to mix the chemical constituents of the egg in order to produce a normal embryo; this would require besides the proper chemical substances a definite arrangement or structure of this material. The limits of divisibility of a cell seem therefore to depend upon its physical structure and must for this reason vary for different organisms and cells. The smallest piece of a sea-urchin egg that can reach the pluteus stage is still visible with the naked eye, and is therefore considerably larger than bacteria or many algæ, which also may be capable of further division.
8. The most important fact which we gather from these data is that the cytoplasm of the unfertilized egg may be considered as the embryo in the rough and that the nucleus has apparently nothing to do with this predetermination. This must raise the question suggested already in the third chapter whether it might not be possible that the cytoplasm of the eggs is the carrier of the genus or even species heredity, while the Mendelian heredity which is determined by the nucleus adds only the finer details to the rough block. Such a possibility exists, and if it should turn out to be true we should come to the conclusion that the unity of the organism is not due to a putting together of a number of independent Mendelian characters according to a “pre-established plan,” but to the fact that the organism in the rough existed already in the cytoplasm of the egg before the egg was fertilized. The influence of the hereditary Mendelian factors or genes consisted only in impressing the numerous details upon the rough block and in thus determining its variety and individuality; and this could be accomplished by substances circulating in the liquids of the body as we shall see in later chapters.
1. The action of the organism as a whole seems nowhere more pronounced than in the phenomena of regeneration, for it is the organism as a whole which represses the phenomena of regeneration in its parts, and it is the isolation of the part from the influence of the whole which sets in action the process of regeneration. The leaf of the Bermuda “life plant”—Bryophyllum calycinum—behaves like any other leaf as long as it is part of a healthy whole plant, while when isolated it gives rise to new plants. The power of so doing was possessed by the leaf while a part of the whole, and it was the “whole” which suppressed the formative forces in the leaf. When a piece is cut from the branch of a willow it forms roots near the lower end and shoots at the upper end, so that a tolerably presentable “whole” is restored. How does the “whole” prevent the basal end of the shoot from forming roots as long as it is part of the plant? A certain fresh-water flatworm has the mouth and pharynx in the middle of the body. When a piece is excised between the head and the pharynx a new head is formed at the oral end, a new tail at the opposite end, and in the middle of the remaining old tissue a new mouth and pharynx is formed. How does the “whole” suppress all this formative power in the part before the latter is isolated? It almost seems as if the isolation itself were the emancipation of the part from the tyranny of the whole. The explanation of this tyranny or of the correlation of the parts in the whole is to be found, however, in a different influence. The earlier botanists, Bonnet, Dutrochet, and especially Sachs,142 pointed out that the phenomena of correlation are determined by the flow of sap in the body of a plant. These authors formulated the idea that the formation of new organs in the plant is determined by the existence of specific substances which are carried by the ascending or descending sap. Specific shoot-producing substances are carried to the apex, while specific root-producing substances are carried to the base of a plant. When a piece is cut from a branch of willow the root-forming substances must continue to flow to the basal end of the piece, and since their further progress is blocked there they induce the formation of roots at the basal end. Goebel143 and de Vries have accepted this view and the writer made use of it in his first experiments on regeneration and heteromorphosis in animals.144 At that time the idea of the existence of such specific organ-forming substances was received with some scepticism, but since then so many proofs for their existence have been obtained that the idea is no longer questioned. Such substances are known now under the name of “internal secretions” or “hormones”; their connection with the theory of Sachs was forgotten with the introduction of the new nomenclature.
It may be well to enumerate some of the cases in which the influence of specific substances circulating in the blood upon phenomena of growth has been proven. One of the most striking observations in this direction is the one made by Gudernatsch on the growth of the legs of tadpoles of frogs and toads.145 The young tadpoles have no legs, but the mesenchyme cells from which the legs are to grow out later are present at an early stage. From four months to a year or more may elapse before the legs begin to grow. Gudernatsch found that legs can be induced to grow in tadpoles at any time, even in very young specimens, by feeding them with the thyroid gland (no matter from what animal). No other material seems to have such an effect. The thyroid contains iodine, and Morse146 states that if instead of the gland, iodized amino acids are fed to the tadpole the same result can be produced. We must, therefore, draw the conclusion that the normal outgrowth of legs in a tadpole is due to the presence in the body of substances similar to the thyroid in their action (it may possibly be thyroid substance) which are either formed in the body or taken up in the food.
Thus we see that the mesenchyme cells giving rise to legs may lie dormant for months or a year but will grow out when a certain type of substances, e. g., thyroid, circulates in the blood. There may exist an analogy between the activating effect of the thyroid substance and the activating effect of the spermatozoön or butyric acid (or other parthenogenetic agencies) upon the egg, but we cannot state that the thyroid substance activates the mesenchyme cells by altering their cortical layer.
The fact that the substance of the thyroid may induce general growth in the human is too well known to require more than an allusion in this connection. When growth stops in children as a consequence of a degeneration of the thyroid, feeding of the patient with thyroid again induces growth. It may also suffice merely to call attention to the connection between acromegaly and the hypophysis.
It was formerly believed that the nervous system acted as a regulator of the phenomena of metamorphosis in animals, but it was possible to show by simple experiments that the central nervous system does not play this rôle and that the regulator must be the blood or substances contained therein. In the metamorphosis of the Amblystoma larva the gills at the head and tail undergo changes simultaneously, the gills being absorbed completely. The writer showed that in larvæ in which the spinal cord was cut in two, no matter at which level,—the sympathetic nerves were in all probability also cut—the two organs continued to undergo metamorphosis simultaneously.147 Uhlenhuth found that if the eye of a salamander larva is transplanted into another larva the transplanted eye undergoes its metamorphosis into the typical eye of the adult form, simultaneously with the normal eyes of the individual into which it was transplanted.148 These and other observations of a similar character leave no doubt that substances circulating in the blood and not the central nervous system are responsible for the phenomena of growth and metamorphosis.
An interesting observation on the rôle of internal secretion in growth was made by Leo Loeb.149 When the fertilized ovum comes in contact with the wall of the uterus it calls forth a growth there, namely the formation of the maternal placenta (decidua). This author showed that the corpus luteum of the ovary gives off a substance to the blood which alters the tissues in the uterus in such a way that contact with any foreign body induces this deciduoma formation. The case is of interest since it indicates that the substance given off by the corpus luteum does not induce growth directly, but that it allows mechanical contact with a foreign body to do so while without the intervention of the corpus luteum substance no such effect of the mechanical stimulus would be observable. The action of the substance of the corpus luteum is independent of the nervous system, since in a uterus which has been cut out and retransplanted the same phenomenon can be observed.
Bouin and Ancel150 have shown that the corpus luteum, which in the case of pregnancy continues to exist for a long time, is responsible for the changes in the mammary gland in the first half of pregnancy, when an active cell proliferation takes place in the gland. This process can be interrupted by destroying the corpus luteum artificially. During the second half of gravidity no further cell proliferation takes place, but the cells begin to secrete milk while during the period of cell proliferation such secretions do not occur.
Claude Bernard and Vitzou had shown that the period of growth and moulting of the higher crustacea is accompanied by a heaping up of glycogen in the liver and subdermal connective tissue. Smith151 found that during the period between two moultings, when there is no growth, the storage cells are seen to be filled with large and numerous fat globules instead of with glycogen. He also found that in the Cladocera “the period of active growth is accompanied by glycogen—as opposed to fat—metabolism.” He observed, moreover, that if Cladocera are crowded at a low temperature the fat metabolism (with inhibition to growth) is favoured, while at high temperatures and with no crowding of individuals the glycogen metabolism is favoured. In the latter case a purely parthenogenetic mode of propagation is observed, while in the former sexual reproduction takes place. The effect of crowding of individuals is possibly due to products of excretion, which then act on growth and reproduction indirectly by changing the “glycogen metabolism” to “fat metabolism.”
All these cases agree in this, that apparently specific substances induce or favour growth, not in the whole body, but in special parts of the body. Sachs suggested that there must be in each organism as many specific organ-forming substances as there are organs in the body.
We will now show that the assumption of the existence of such “organ-forming” substances (which may or may not be specific) and of their flow in definite channels explains the inhibitory influence of the whole on the parts as well as the unbridled regeneration of the isolated parts.
2. We have seen that the resting egg can be aroused to development and growth by substances contained in a spermatozoön or by certain other substances mentioned in the preceding chapter. We will assume that plants contain a large number of cells or buds which are comparable to the resting egg cell, but which can be aroused to action by certain substances circulating in the sap; and that the same is effected for animal cells by substances in the blood. In plants the cells which can be aroused to new growth have very often a rather definite location while in lower animals they are more ubiquitous. For experimental purposes organisms where these buds have a definite location are more favourable, since we are better able to study the mechanism underlying the process of activation and inhibition (correlation). When a leaf of the plant Bryophyllum calycinum is cut off and put on moist sand or into water or even into air saturated with water vapour, new plants will arise from notches of the leaf. This is the usual way of propagating the plant and in no other part of the leaf except the notches will new plants arise. These notches therefore contain cells comparable to seeds or to unfertilized eggs or to the mesenchyme cells which give rise to legs in the tadpole of the frog. The question arises: Why do notches in the leaf never begin to grow while the leaf is attached to an intact plant, and why do they grow when the leaf is isolated? To this we are inclined to give an answer in the sense of Bonnet, Sachs, de Vries, and Goebel, namely that the flow of (specific?) substances in the plant determines when and where dormant buds or anlagen shall begin to grow. Such substances may originate or may be present in the leaf; but as long as it is connected with a normal plant they will be carried by the circulation to the growing points of the stem and of the roots and they cannot reach the notches; while when we detach the leaf, either a new distribution or a new flow of liquids will be established whereby the substances reach some of the notches; and in these notches new roots and a new shoot will be formed. When we cut off a leaf and put it into moist air, not all but only a few of the notches will, as a rule, grow out (Fig. 16); but when we isolate each notch leaving as much of the rest of the leaf as possible attached to it, each notch will give rise to a new plant.152 (Fig. 17.) We see, therefore, that it does not even require a whole plant to cause inhibition but that we may observe the tyranny of the whole over the parts in a single leaf. The explanation is as follows: When we isolate a leaf, some of the notches will commence to grow into new plants and this growth will arrest the development of the other notches of the leaf in the same way as their development was suppressed by the whole plant.

Fig. 16. Growth of roots and shoots in a few notches of an isolated leaf of Bryophyllum calycinum |
Fig. 17. If all the notches of a leaf are isolated from each other each notch will give rise to roots and a shoot, but the growth will be less rapid than in Fig. 16. Figs. 16 and 17 were two leaves taken from the same node of a plant. |
The explanation is the same; those notches which begin to grow first will attract the flow of substances to themselves, thus preventing the other notches from getting those substances. This idea is supported by the fact that if all the notches are isolated from the leaf each notch will give rise to a slowly growing plant, while if the leaf is not cut into pieces, and a few notches only grow out, their growth is much more rapid.
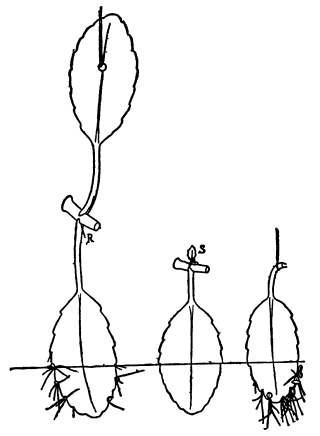
Fig. 18 Fig. 19 Fig. 20
In all these experiments the idea that the “isolation” in itself is responsible for the growth still presents itself. It can be disposed of by the following experiment which never fails. Three leaves of Bryophyllum calycinum are suspended in an atmosphere saturated with water vapour but their tips are submersed in water (Figs. 18, 19, 20). The first leaf, Fig. 20, is entirely separated from its stem, the second leaf, Fig. 19, remains connected with the adjacent piece of stem, and the third leaf, Fig. 18, remains also connected with this piece of stem but the latter still possesses both leaves. The first leaf, Fig. 20, produces new roots and shoots in the submerged part in a few days; the second leaf, Fig. 19, produces no roots or shoots for a long time. This might find its explanation by the assumption that the first leaf, being more isolated than the second, regenerates more quickly. But this explanation becomes untenable owing to the fact that the third leaf, Fig. 18, being less isolated than both (possessing a second leaf in addition to the stem), forms new roots and shoots also more quickly than the second leaf. The phenomena become intelligible in the following way. The fact that in the second leaf shoots and roots are formed very late, if at all, finds its explanation not in the lessened isolation of this leaf, but in the fact that the formation of a new shoot or of a callus in the piece of stem takes place more quickly than the formation of roots and shoots in the notches of a completely isolated leaf. The stem acts therefore as a centre of suction for the flow of substances from the leaf and this prevents or retards the formation of roots and shoots in the notches. In the isolated leaf of Bryophyllum calycinum no callus formation takes place and hence no flow of the sap away from the leaf will occur. This will allow one or more of the notch buds of this leaf to grow out and then a flow will be established towards these growing buds.
In the third specimen, Fig. 18, the presence of two leaves suppresses or, as a rule, retards the growth of a shoot on the stem and possibly also the flow from one leaf may block to some extent the flow from the opposite leaf if the piece of stem is very short. This puts the leaves in a condition not as good as that in leaf Fig. 20, but better than in leaf Fig. 19.153
In the normal plant the buds in the notches of the leaf remain dormant since the flow of the “stimulating” substances takes place towards the tips of the stem and root, and because these substances are retained there in excess. This is probably the real basis of the mysterious dominance of the “whole” over its “parts” or of the anlagen of the tip of the stem over those farther below. When a piece of the stem of Bryophyllum is cut off and its leaves are removed, the two apical buds will grow out first. This “dominance” finds its explanation probably in the anatomical structure and the mechanism of sap flow which tend to bring the “stimulating” substances first to the anlagen in the tip. In Laminaria Setchell has been able to show directly that regeneration always starts from that tissue which conducts nutritive material.
When we cut out a piece of a stem of Bryophyllum, and remove all the leaves, new shoots will be formed from the two apical buds of the stem, and roots will arise from the most basal nodes; provided that the stem is suspended in air saturated with water vapour. The growth in such a stem deprived of all leaves is slow. If we remove all the leaves on such a piece of stem except the two at the apical end, the stem will form only roots, but these will develop much more rapidly than on a stem without leaves. If we remove all the leaves except the two at the basal end, the stem will only form shoots (at the apical end) but these will develop much more rapidly than in a leafless stem. Hence the leaves accelerate the growth of roots towards the basal end and inhibit it towards the apical end; and they favour the growth of shoots towards the apical end and inhibit it in the nodes located nearer the base.
We thus see that while the stem inhibits the growth of the leaves connected with it, the latter accelerate the growth in the stem. Both facts can probably be explained on the same basis; namely, on the assumption that it is the flow of substances from the leaf to the stem which inhibits the growth of the notches and accelerates the growth of the buds in the stem. On this assumption it would also follow that the leaves send root-forming substances towards the basal and shoot-forming substances towards the apex of the stem. It also seems to follow from recent as yet unpublished experiments by the writer that the root-forming substances are associated or identical with the substances which cause geotropic curvature in the stem.
These observations show that the phenomena of correlation or of the influence of the whole over the parts is due to peculiarities of circulation or the flow of sap; and that the isolation prevents the sap from flowing away to other parts of the plant. There is no need for assuming the existence of a mysterious force which directs the piece to grow into a whole.

Fig. 21 |

Fig. 22 |
3. Phenomena of inhibition or correlation such as we have described in Bryophyllum are not lacking in the regeneration of animals, as experiments on Tubularia show.154 Tubularia mesembryanthemum (Fig. 21) is a hydroid consisting of a long stem terminating at one end in a stolon which attaches itself to solid bodies such as rocks, at the other end in a polyp. The writer found that if we cut a piece from a stolon and suspend it in an aquarium it forms as a rule a polyp at either end (Fig. 22), but the velocity with which the two polyps are formed is not the same, the polyp at the oral end of the piece being formed much more rapidly—a day or one or two weeks sooner—than the aboral polyp. The process of polyp regeneration at the aboral pole could, however, be accelerated and its velocity made equal to that of the regeneration of the oral polyp by suppressing the formation of the latter. This was accomplished by depriving the oral pole of the oxygen necessary for regeneration, e. g., by merely putting the oral end of the piece of stem into the sand. It was, therefore, obvious that the formation of the oral polyp retarded the formation of the aboral polyp. This inhibition might have been due to the fact that a specific organ-forming material needed for the formation of a polyp existed in sufficient quantity in the stem for the formation of one polyp only at a time. This idea, however, was found to be incorrect since when the stem was cut into two or more pieces each piece formed a polyp at once at its oral pole and regenerated the aboral polyps also, but again with the usual delay. It seemed more probable then that the cause of the difference in the rapidity of polyp formation at both ends lay in the fact that certain material flowed first to the oral pole and induced polyp formation here but that this flow was reversed as soon as the polyp at the oral pole was formed or as soon as the formation of the oral polyp was inhibited by lack of oxygen. The partial or full completion of the formation of the oral polyp acted as an inhibition to the further flow of material to this pole. This idea was supported by an observation made independently by Godlewski and the writer that if a piece of stem be cut out of a Tubularia, and if the piece be ligatured somewhere between the two ends, the oral and the aboral polyps are formed simultaneously. This would be comprehensible on the assumption that the retarding effect which the formation of the oral has on the aboral polyp was indeed of the nature of a flow of material towards the oral pole.

Fig. 23
Miss Bickford155 found that the difference in time between the formation of the two polyps disappears also when the piece cut from the stem becomes so small that it is of the order of magnitude of a single polyp. In that case two incomplete polyps are formed simultaneously at each end (Fig. 23). The new head in the regeneration of Tubularia arises, as Miss Bickford observed, from the tissue near the wound. At some distance from the wound in the old tissue two rows of tentacles arise, which are noticeable as rows of longitudinal lines inside the stem before the head is formed. Driesch noticed that the newly formed head is the smaller the smaller the whole piece. (This is true, however, only in rather small pieces.) There is, therefore, in small pieces a rough proportionality between size of head and size of regenerating piece. Driesch156 uses this interesting fact to prove the existence of an entelechy, while we are inclined to see in it an analogue to the observation of Leo Loeb, that the velocity of the process of healing in the case of a deficiency of the epithelium decreases when the size of the uncovered area diminishes. While we do not wish to offer any suggestion concerning the mechanism of these quantitative phenomena—they may be related in some way with the velocity of certain chemical reactions—we see no reason for assuming that they cannot be explained on a purely physicochemical basis.
The writer noticed that certain pigmented cells from the entoderm of the organism always gather at that end where a new polyp is about to be formed. These red or yellowish cells always collect first at the oral end of a piece of stem. It may be that certain substances given off by the pigmented cells at the cut end are responsible for the polyp formation, but this is only a surmise.
Another suggestion made by Child,157 is that there exists an axial gradient in the stem whereby the cells regenerate the more quickly the nearer they are to the oral pole. If this were correct, and we cut a long piece from the stem of a Tubularia and bisect the piece, the oral pole of the anterior half should regenerate more quickly than the oral pole of the posterior half. According to the writer’s observations on a Tubularian (T. crocea) growing in the estuaries near Oakland, California, both oral ends regenerate equally fast in such cases.
4. The phenomena of regeneration in Cerianthus membranaceus, a sea anemone, can be easily understood from the experiments on Tubularians, if we imagine the body wall of Cerianthus to consist of a series of longitudinal elements running parallel to the axis of symmetry of the animal from the tentacles to the foot. The number of these elements may be supposed to correspond to the number of tentacles in the outer row of the normal animal. Each such element behaves like a Tubularian, with this difference, however, that the elements in Cerianthus are more strongly polarized than in Tubularia, and that each one is able to form a tentacle at its oral pole only. This fact can be nicely illustrated in the following way: if a square or oblong piece (a b c d, Fig. 24) be cut from the body wall of a Cerianthus in such a way that one side, a c, of the oblong is parallel to the longitudinal axis of the animal, tentacles will grow on one of the four sides only; namely, on the side a b.158 (Fig. 25.) The other three free edges are not able to produce tentacles. If an incision be made in the body wall of a Cerianthus, tentacles will grow on the lower edge of the incision (Fig. 26).
Fig. 24 |
Fig. 25 |
Fig. 26 |
The writer tried whether or not by tying a ligature around the middle of a piece of an Actinian this polarity could be suppressed; but the experiments did not succeed, inasmuch as the cells compressed by the ligature died, and were liquefied through bacterial action so that the pieces in front and behind the ligature fell apart. It is therefore impossible to decide whether or not a current or a flow of substances in a certain direction through these elements is responsible for this polarity, though this may be possible. The writer found, however, that one condition is necessary for the growth and regeneration of tentacles which also plays a rôle in the corresponding phenomena in plants, namely turgidity. The tentacles of Cerianthus are hollow cylinders closed at the tip, and by liquid being pressed into them they can be stretched and appear turgid. If, however, an incision is made in the body, the tentacles above the incision can no longer be stretched out. In one experiment the oral disk of a Cerianthus was cut off; very soon new tentacles began to grow at the top, and after having reached a certain size, an incision was made in the animal. The tentacles above the incision collapsed in consequence and ceased to grow, while growth of the others continued. On the lower edge of the incision new tentacles began to grow.
Fig. 27 |
Fig. 28 |
Fig. 29 |
It seems also possible that Morgan’s well-known experiment on regeneration in Planaria can be explained by a flow of substances. He159 found that if a piece a c d b be cut out of a fresh-water Planarian at right angles to the longitudinal axis (Fig. 27), at the front end a new normal head, at the back end a new tail, will be regenerated (Fig. 28); but that if a piece a c d b be cut from a Planarian obliquely (Fig. 29) instead of at right angles to the longitudinal axis a tiny head is formed at the foremost corner of the piece a and a tiny tail at the hindmost corner b (Fig. 30). Why is it that in the oblique piece the head is formed in the corner and not all along the cut surface as is the case when the cut is made at right angles to the longitudinal axis? The writer is inclined to believe that the right answer to this question has been given by Bardeen.160 This author has pointed out the apparent rôle that the circulatory (or so-called digestive) canals in Planarians play in the localization of the phenomena of regeneration, inasmuch as the new head always forms symmetrically at the opening of the circulatory vessel or branch which is situated as much as possible at the foremost end of the regenerating piece of worm. He assumes that through muscular action the liquids of the body are forced to stream toward this end, and that this fact has some connection with the formation of a new head. There can be no doubt that the facts here mentioned agree with Bardeen’s suggestion. The oblique pieces in Morgan’s experiments which at first have the heads and tails outside the line of symmetry of the middle piece, gradually assume a normal position (Figs. 31, 32). The writer is inclined to believe that this is due to mechanical conditions. The head a e c of such an oblique piece is asymmetrical, the one side a e being less stretched than the other e c. The higher tension of the piece e c will have the effect of bringing e nearer c, since we know that acid formation and hence energy production increases in proportion to surface, i. e., it must be the greater the more it is stretched. The reverse is true for the tail d f b, and the effect here will be that f will be pulled nearer d. In this way purely mechanical conditions are responsible for the fact that the soft tissues of the animal are gradually restored to their true orientation.
Fig. 31 |
Fig. 32 |
As a final possible example of the influence of internal secretion or substances contained in the blood may be mentioned the following curious observation of Przibram.161 In a crustacean, Alpheus, the two chelæ (pincers) are not equal in size and form, one being very much larger than the other. Przibram found that when he cut off the larger pincer in such crustaceans the remaining pincer assumes in the next moulting the size and shape of the removed large pincer; while in place of the removed pincer one of the small type is produced. Hence a reversal of the two pincers is thus brought about. If later on the large pincer is again cut off the process is repeated and the original dissymmetry is restored. Przibram was able to show that the nervous system has no connection with this phenomenon.
The elements which have entered into the discussion thus far are, first, the flow of substances in preformed channels; second, the existence of general or specific substances required for the growing or regenerating organ. A third element is to be added; namely the “suction” effect upon these substances of a developing organ. Thus we see that if one or a few of the notches in a leaf of Bryophyllum grow out the other notches of the leaf are inhibited from growing. There is enough material present in the leaf for all the notches to grow into shoots as is proved by the fact that all will grow out if they are isolated from each other. This was explained on the assumption that the notches of a whole which happen to develop first, create a flow of these substances from the rest of the leaf to themselves and thus prevent any getting to the other notches. We stated that this is supported by the fact that the few notches growing out in an undivided leaf grow more rapidly than the many shoots growing from each notch of a divided leaf. But why should a growing shoot or a growing point in general produce such a suction? I think this may be possible on the assumption that the consumption of these substances by the growing organs causes a low osmotic pressure of these substances in the growing region and this fall of osmotic potential will act as a cause for the further flow. This brings about the apparent “suction” effect of the growing elements upon the flow of substances.
5. We mentioned that when a piece is cut from a Planaria between pharynx and head a new mouth is formed in the middle. It should also be mentioned that according to Child the piece after regeneration is smaller than it was before.162 This indicates that material in the old cells has been digested or has undergone hydrolysis in order to furnish the nutritive material for the new head and tail, since the piece cannot take up any food from the outside before a mouth is formed. These phenomena of autodigestion—the process itself will be discussed in the last chapter—seem to occur in many (if not all) phenomena of regeneration. It may be that the collecting of red cells at the end in a Tubularian where regeneration is about to begin has to do with the furnishing of material by self-digestion, since these cells are partly at least destroyed in the process. It is of interest to look for more examples of autodigestion accompanying phenomena of regeneration.
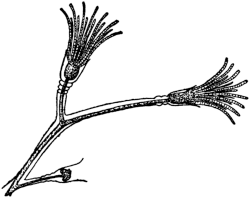
Fig. 33 |
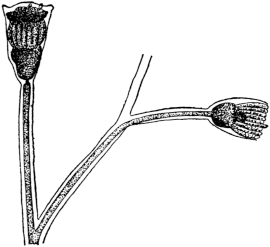
Fig. 34 |
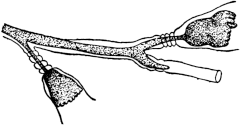
Fig. 35
The writer has observed more closely the transformation of an organ into more undifferentiated material in Campanularia (Fig. 33), a hydroid.163 This organism shows a remarkable stereotropism. Its stolons attach themselves to solid bodies, and the stems appear on the side of the stolon exactly opposite the point or area of contact with the solid body. The stems grow, moreover, exactly at right angles to the solid surface element to which the stolon is attached. If such a stem be cut and put into a watch glass with sea water, it can be observed that those polyps which do not fall off go through a series of changes which make it appear as if the differentiated material of the polyp were transformed into undifferentiated material. The tentacles are first put together like the hairs of a camel’s-hair brush (Fig. 34), and gradually the whole fuses to a more or less shapeless mass which flows back into the periderm (Fig. 35). It follows from this that in this process certain solid constituents of the polyp, e. g., the cell walls, must be liquefied. This undifferentiated material formed from the polyp may afterward flow out again, giving rise to a stolon or a polyp; to the former where it comes in contact with a solid body, to the latter where it is surrounded by sea water. These observations suggest the idea of reversibility of the process of differentiation of organs and tissues, in certain forms at least. We have to imagine that some of the cells or interstitial tissue is digested and that as a consequence the organ loses its characteristic shape.
Giard and Caullery have found that a regressive metamorphosis occurs in Synascidians, and that the animals hibernate in this condition. The muscles of the gills of these animals are decomposed into their individual cells. The result is the formation of a parenchyma which consists of single cells and of cell aggregates resembling a morula.164
Driesch,165 experimenting on the regeneration of an Ascidian, found that when he cut off the gills and siphons of the animal the portion removed was able to regenerate a whole animal. The gill-piece excised contained no heart, no intestine, and no stolon, and all these organs were regenerated from the gills. In a number of cases the regeneration took place by bud formation at the edge of the wound, but in other cases the gills were transformed into an undifferentiated mass of tissue from which the missing parts of the animals arose by budding and new gills were formed.
It is probable that the two cases are only quantitatively different. In both, autodigestion of certain cell constituents and possibly of whole cells must take place in order to obtain material for the formation of the lost part of the Ascidian. If an interstitial tissue is digested it becomes a question of how much of this tissue undergoes hydrolysis. If there is little destroyed the old shape of the gills remains, if too much is digested the old gills become a shapeless mass in which a certain number of the old cells are maintained and give rise to the new animal by cell division. The material for the new organs must of course be furnished from old cells which have been digested.
If regeneration takes place in pieces which take up no food the newly formed organs must originate from material absorbed from cells of the animal which are hydrolyzed and whose material serves as food for those cells which grow. Very often this process of digestion takes place without loss of the total form of the organ and is overlooked by the pure morphologists. In Campanularia also the process of collapse described above is only apparent in a fraction of the cases as in Driesch’s observations on Clavellina.166 It is also possible that the red and yellow entoderm cells which gather at the end where the new polyp forms furnish the material which is utilized for the process of growth of the cells from which the tentacles arise (with or without giving off specific “hormones” besides).
6. We have mentioned the ideas concerning a design, or “entelechy,” acting as a guide to the developing egg and have shown that this revival of Platonic and Aristotelian philosophy in biology was due to a misconception; namely, that the egg consisted of homogeneous material which was to be differentiated into an organism. For this supernatural task supernatural agencies seemed required. But we have seen that the unfertilized egg is already differentiated in a way which makes the further differentiation a natural affair. This idea of a quasi superhuman intelligence presiding over the forces of the living is met with in the field of regeneration, and here again it is based upon a misconception. The lens of the eye is formed in the embryo from the epithelium lying above the so-called optic cup (the primitive retina). Where this retina touches the epithelium the latter begins to grow into the cup, the ingrowing piece of epithelium is cut off and forms the lens, which probably under the influence of substances secreted by the optic cup becomes transparent. Certain animals like the salamander are able to form a new lens when the old one has been removed by operation, but the new lens is formed in an entirely different way; namely, from the upper edge of the iris. G. Wolf, who observed this regeneration used it to endow the organism with a knowledge of its needs; the idea of a Platonic preconceived plan or an Aristotelian purpose suggested itself. But it can be shown that the organism does in this case what it is compelled to do by its physical and chemical structure.
Uhlenhuth167 has shown by way of tissue culture that the cells of the iris cannot grow and divide as long as they are full of pigment granules as they normally are. When the fine superficial membrane of the iris is torn the pigment granules fall out and the cells can now grow and multiply. If the lens is taken out of the eye of the salamander the fine membrane of the iris is torn and the pigment cells at the edge (especially the upper edge) lose their pigment granules which fall down on account of their specific gravity. As soon as this happens the cells will proliferate. A spherical mass of cells is formed which become transparent and which will cease to grow as soon as they reach a certain size. The unanswered question is: Why does the mass of cells become transparent so that it can serve as a lens? The answer is that young cells when put into the optic cup always become transparent no matter what their origin; it looks as if this were due to a chemical influence exercised by the optic cup or by the liquid it contains. Lewis has shown that when the optic cup is transplanted into any other place under the epithelium of a larva of a frog the epithelium will always grow into the cup where the latter comes in contact with the epithelium; and that the ingrowing part will always become transparent. This leaves us then with one puzzle still: Why is the growth of the lens limited? The limitation in the growth of organs is one of the most important problems in growth and organ formation, though unfortunately our knowledge of this topic is inadequate.
7. The botanist J. Sachs was the first to definitely state that in each species the ultimate size of a cell is a constant, and that two individuals of the same species but of different size differ in regard to the number, but not in regard to the size of their cells.168 Amelung, a pupil of Sachs, determined the correctness of Sachs’s theory by actual counts. Sachs, in addition, recognized that wherever there were large masses of protoplasm, e. g., in siphoneæ and other cœloblasts, many nuclei were scattered throughout the protoplasm. He inferred from this that “each nucleus is only able to gather around itself and control a limited mass of protoplasm.”169 He points out that in the case of the animal egg the reserve material—fat granules, proteins, and carbohydrates—are partly transformed into the chromatin substances of the nuclei, and that the cell division of the egg results in the cells reaching a final size in which each nucleus has gathered around itself that mass of protoplasm which it is able to control. Morgan170 and Driesch171 tested and confirmed the idea of Sachs for the eggs of Echinoderms. We stated in the previous chapter that Driesch produced artificially larvæ of sea urchins of one-eighth, one-fourth, and one-half their normal size by isolating a single cleavage cell in one of the first stages of segmentation of the fertilized sea-urchin egg. He counted in each of the dwarf gastrulæ resulting from these partial eggs the number of mesenchyme cells and found that the larvæ from a one-half blastomere possessed only one-half, those from a one-fourth blastomere only one-fourth, and those from a one-eighth blastomere only one-eighth of the number of cells which a normal larva developing from a whole egg possessed. Moreover, he could show that when two eggs were caused to fuse so as to produce a single larva of double size, the gastrulæ of such larvæ had twice the number of mesenchyme cells. Driesch drew the conclusion from his observations that each morphogenetic process in an egg reaches its natural end when the cells formed in the process have reached their final size.
Since each daughter nucleus of a dividing blastomere has the same number of chromosomes as the original nucleus of the egg, it is clear that in a normally fertilized egg each nucleus has twice the mass of chromosomes that is contained in the nucleus of a merogonic egg, i. e., an enucleated fragment of protoplasm into which a spermatozoön has entered and which is able to develop. Such a fragment has only the sperm nucleus. This phenomenon of merogony was discovered by Boveri and was elaborated by Delage.172 Boveri, in comparing the final size of the cells in normal and merogonic eggs after the cell divisions had come to a standstill, found that this size is always in proportion to the original mass of the chromatin contained in the egg; the cells of the merogonic embryo, e. g., the mesenchyme cells, are only half the size of the same cells in the normally fertilized embryo. Driesch furnished a further proof of Boveri’s law, that the final ratio of the mass of the chromatin substance in a nucleus to the mass of protoplasm is a constant in a given species. Driesch compared the size of the mesenchyme cells in a sea-urchin embryo produced by artificial parthenogenesis with those of a normally fertilized egg and found them half of the size of the latter. When the fertilized eggs and the parthenogenetic eggs are equal in size from the start,—which is practically the case if eggs of the same female are used,—the process of the formation of mesenchyme cells comes to a standstill when their number in the normally fertilized eggs is half as large as the final number in the parthenogenetic egg.173 Boveri’s results as well as those of Driesch were obtained by counting the cells formed by eggs of equal size and not by simply measuring the size of the cells. It is most remarkable that certain apparent exceptions to Boveri’s law which Driesch has actually found had been predicted by Boveri.
These facts show that the growth of an organ comes to a standstill when a certain size is reached or a certain number of cells are formed. We cannot yet state why this should be, but we are able to add that the formation of a lens of normal size in the regeneration of the eye is in harmony with the phenomena in the embryo. There seems therefore no reason for stating that the regeneration of the lens cannot be explained on a purely physicochemical basis. The only justification for such a statement on the part of Wolf is that he was not in possession of the more complete set of facts now available through the work of Fischel and Uhlenhuth.
The healing of a wound is a process essentially similar to the regeneration of the lens. Normally the cells which begin to proliferate after a wound is made in the skin lie dormant, inasmuch as they neither grow nor divide. When a wound is made certain layers of epidermal cells undergo rapid cell division. Leo Loeb174 has studied this case extensively. He found that if the skin is removed anywhere, epidermis cells from the wound edge creep upon the denuded spot and form a covering. This may be a tropism (stereotropism) or it may be a mere surface tension phenomenon. Next a rapid process of cell division begins in the cells adjacent to the wound these cells having been heretofore dormant. He is inclined to attribute this increase in the rate of cell division to the stretching of the epithelial cells, and he is supported in this reasoning by the observation that the larger the wound the more rapid the process of healing.175 During wound healing the mitoses first increase markedly in the old epithelium. With the closure of the wound a sudden fall in the mitoses takes place. The closure of the wound causes an increase in the number of epithelial rows over the defect. This increase is therefore reached at an earlier period in the larger wound since the process of mitosis is more rapid here. Leo Loeb thinks that the pressure of the epithelial cells upon each other leads to a rapid diminution in the mitotic proliferation.176
Should it be possible that this is more generally the case, e. g., also in the lens after it has reached a certain size? The conditions limiting growth require further investigation.
It is hardly necessary to point out that in these cases we are seemingly dealing with cases of the inhibition of growth which cannot be explained by the tyranny of the whole over the parts, and that there must be conditions at work other than the mere flow of substances which can cause a cessation of growth. This can be illustrated by certain observations on the egg.
8. The history of the egg shows a reversible condition of rest and of activity. The primordial egg cell multiplies actively until a large number of eggs are formed in the ovary which may reach into the millions in the case of sea urchins or certain annelids. These cell divisions then stop and the egg goes into the resting stage in which it deposits the reserve material for the development of the embryo. From this condition it can only be called into activity again by the spermatozoön or the agencies of artificial parthenogenesis.
It seemed of interest to find out whether or not the development of the egg may be reversed once more after it has been activated. From all that has been said in the chapter on artificial parthenogenesis, such a reversal should take place in the cortical layer. The result of these experiments seems to be that if a complete destruction or change in the cortical layer has once taken place—such as that caused by the entrance of a spermatozoön into the egg—no reversal is possible; although the development of the fertilized egg may be suppressed for a long time by either low temperature or lack of oxygen, or, in the case of seeds and spores, by lack of water. But as soon as the conditions for the chemical reactions in the egg are normal again, the development may go on unless the egg has suffered by the methods used to prevent development or by the long duration of the suppression. With an incomplete destruction of the cortical layer both development as well as reversal of development are possible. Thus the writer has shown that in the egg of Arbacia the effect of the cortical alteration of the egg induced by the butyric acid treatment or by the treatment with bases can be reversed. When unfertilized eggs of Arbacia are put for from two to five minutes into 50 c.c. sea water + 2.0 c.c. N/10 butyric acid they will all form a gelatinous, somewhat atypical fertilization membrane; when put back into normal sea water all will perish in a few hours unless they are submitted to the short treatment with a hypertonic solution mentioned in the previous chapter, while if submitted to this treatment they will develop. If, however, these eggs are transferred from the butyric acid sea water not into normal sea water but into sea water containing some NaCN (10 drops of 1⁄10 per cent. NaCN or KCN in 50 c.c. sea water), and if they remain here for some time (e. g. overnight) they will not perish when subsequently transferred back to normal sea water. Such eggs will develop when fertilized with sperm. The activating effect of the membrane formation has, therefore, been reversed and the eggs have gone back into the resting stage.177 Wasteneys has found that the rate of oxidation which was raised considerably by the artificial membrane formation goes back to the value characteristic for the resting eggs after the reversal of their developmental tendency.178 Similar results were obtained in eggs activated with NH4OH. It appears from this as though the change in the cortical layer which leads to the development of the egg and the increase in the rate of oxidations were reversible in the egg of Arbacia.179
The writer had previously noticed that eggs of Strongylocentrotus purpuratus, which had been treated for two hours with hypertonic sea water, not infrequently began to divide into two, four, or eight cells (and sometimes more) and then went back into the resting state (except that they possessed the second factor required for development as stated in Chapter V). It may be remarked incidentally that such eggs at the time of cell division contained the centrosomes and astrospheres, and yet went back into a resting state, thus showing that the centrosomes are only transitory organs or organs which are only active under certain conditions. It is quite possible that in these phenomena of reversal not the whole of the cortical layer has undergone alteration.
The writer must leave it undecided whether the changes from the resting to the active state in body cells can also be explained in analogy with these experiments.
9. In the formation of the lens we have already noticed an instance where the adjacent organ influences growth inasmuch as the optic cup controlled the formation of the lens. Such influences are quite commonly observed. A piece of Tubularia when cut out from a stem and suspended in water will regenerate at the aboral pole not a stolon but a polyp, so that we have an animal terminating at both ends of its body in a head. The writer called such cases in which an organ is replaced by an organ of a different kind heteromorphosis.

Fig. 36
Contact with a solid body favours the formation of stolons. Fig. 36 shows a piece of a stem of Pennaria another hydroid, which was lying on the bottom of an aquarium and which formed stolons at both ends a and b. In Margelis, another hydroid, the writer observed that without any operation the apical ends of branches which were in contact with solid bodies continued to grow as stolons, while those surrounded by sea water continued to grow as stems.
Herbst discovered a very interesting form of heteromorphosis in certain crustaceans; namely, that in the place of an eye which was cut off, an entirely different organ could be formed, namely, an antenna. He showed that the experimenter has it in his power to determine whether the crustacean shall regenerate an eye or an antenna in place of the eye. The latter will take place when the optic ganglion is removed with the eye, the former when it is not removed. These experiments were carried out successfully on Palæmon, Palæmonetes, Sicyonia, Palinurus, and other crustaceans.
The influence of gravitation is very familiar in plants; in stems of Bryophyllum placed horizontally the roots usually come out from the lower end of the callus. Such phenomena are not often found in animals but they exist here too as the following observation shows.
Fig. 37
Fig. 38
If we cut a piece a b (Fig. 37), from the stem s s of Antennularia antennina (Fig. 38), a hydroid, and put it into the water in a horizontal position, new stems c d (Fig. 37) may arise on its upper side. The small branches on the under side of the old stem a b begin suddenly to grow vertically downward.180 In appearance and function these downward-growing elements are entirely different from the branches of the normal Antennularia; they are roots. In order to understand better the transformation which thus occurs in these branches, it may be stated that under normal conditions they have a limited growth (see Fig. 38), are directed upward, and have polyps on their upper side. The parts which grow down (Fig. 37) have no polyps, but attach themselves like true roots to solid bodies. Thus the changed position of the stem alone, without any operation, suffices to transform the lateral branches, whose growth is limited, into roots with unlimited growth. The lateral branches on the upper side of the stem do not undergo such a transformation into roots except in the immediate surroundings of the place where a new stem arises. It seems that the formation of a new stem also causes an excessive growth of roots, possibly because the formation of new branches causes the removal of substances which naturally inhibit the formation of roots. If a piece from the stem be put vertically into the water with top downward, the uppermost point may continue to grow as a stem, while the lowest point may give rise to roots. In this case, therefore, a change in the orientation of organs has the effect of changing the character of organs.
There are only two ways by which we can account for these influences of gravitation. Either certain substances flow to the lowest level and collecting there induce growth and possibly changes in the character of growth (as in Antennularia) or if the cells have elements of different specific gravity the relative position of these elements may possibly change and influence in this way the conditions for growth. The influence of gravitation as well as of contact upon life phenomena are at present little understood.
In all these cases of heteromorphosis the original form is not restored. It is needless to say that they are incompatible with the theory of natural selection.
The reader will have noticed that in this chapter one term has not been mentioned which is commonly met with in the literature, namely the “wound stimulus.” As the writer had indicated in a former publication,181 the word “stimulus” is generally used to disguise our ignorance of (and also our lack of interest in) the causes which underlie the phenomena which we investigate. Regeneration very often does not take place near the wound but at some distance from it. But even when the regeneration takes place at the edge of the wound the latter only serves to create conditions for regeneration, and these conditions cannot be expressed by the word “stimulus.”
While our knowledge of the rôle of the whole in regeneration is incomplete in a great many details it seems that the known facts warrant the statement that the phenomena of regeneration belong as much to the domain of determinism as those of any of the partial phenomena of physiology.
1. It is a general fact that both sexes appear in approximately equal numbers, provided a sufficiently large number of cases are examined. This fact has furnished the clue for the discovery of the mechanism which determines the relative number of the two sexes. The honour of having pointed the way to the solution of the problem belongs to McClung.182 It has been known that certain insects, e. g., Hemiptera and Orthoptera, possess two kinds of spermatozoa but only one kind of eggs. The two kinds of spermatozoa differ in regard to a single chromosome, which is either lacking or different in one-half of the spermatozoa.
The first one to recognize the existence of two kinds of spermatozoa was Henking, who stated that in Pyrrhocoris (a Hemipteran) one-half of the spermatozoa of each male possessed a nucleolus, while in the other half it was lacking. Montgomery afterward showed that Henking’s nucleolus was an accessory chromosome. McClung was the first to recognize the importance of this fact for the problem of sex determination. He observed an accessory chromosome in one-half of the spermatozoa of two forms of Orthoptera, Brachystola and Hippiscus, and reached the following conclusion:
A most significant fact, and one upon which almost all investigators are united in opinion, is that the element is apportioned to but one-half of the spermatozoa. Assuming it to be true that the chromatin is the important part of the cell in the matter of heredity, then it follows that we have two kinds of spermatozoa that differ from each other in a vital matter. We expect, therefore, to find in the offspring two sorts of individuals in approximately equal numbers, under normal conditions, that exhibit marked differences in structure. A careful consideration will suggest that nothing but sexual characters thus divides the members of a species into two well-defined groups, and we are logically forced to the conclusion that the peculiar chromosome has some bearing upon the arrangement.
N. M. Stevens and E. B. Wilson183 have not only proved the correctness of this idea for a number of animals but have laid the foundation of our present knowledge of the subject. Wilson showed that in those cases where there are two types of spermatozoa, one with and one without an accessory or as it is now called an X chromosome, all the cells of the female have one chromosome more than the cells of the male. From this he concludes correctly that in such species a female is produced when the egg is fertilized by a spermatozoön containing an X chromosome, while a male is produced when a spermatozoön without an X chromosome enters the egg.
Such a form is Protenor, one of the Hemiptera. Wilson made sure that all the eggs are alike in the number of chromosomes, each egg containing an X chromosome in addition to the six chromosomes characteristic of the species Protenor. There are two types of spermatozoa in equal numbers in this species, each with six chromosomes, but one with, the other without, an X chromosome. The two possible chromosome combinations between egg and spermatozoa are therefore as follows (see the diagrammatic Fig. 39):
| Egg | Spermatozoön | Result |
| (1) 6 + X | + 6 | = 12 + X = Male |
| (2) 6 + X | + 6 + X | = 12 + 2X = Female |
The egg which receives a spermatozoön without an X chromosome has after fertilization 12+X chromosomes and develops into a male; while the egg into which a spermatozoön with an X chromosome enters gives rise to a female. Since all the body cells arise from the fertilized egg by nuclear division and the chromosomes remain constant in number in all cells, the consequence is that all the cells of a female Protenor have two X chromosomes; while all the cells of a male Protenor have only one X chromosome.
The chromosome situation in Protenor is a somewhat extreme case, inasmuch as one X chromosome is entirely lacking in the male. In other forms of Hemiptera, e. g., Lygæus, there are also two types of spermatozoa appearing in equal numbers differing in regard to the X chromosome, but here it is only a difference in size; one-half of the spermatozoa having a large X chromosome, the other half instead a smaller chromosome. Calling this latter the Y chromosome, the sex determination in this form is as follows: leaving aside the chromosomes which are equal in both egg and spermatozoön we may say that there is one type of egg containing one large X chromosome; there are two types of spermatozoa in equal numbers, one possessing a large X chromosome, the other possessing a small Y chromosome. Wilson showed by a study of the chromosomes in males and females that when one of the spermatozoa containing a large X chromosome enters the egg, the egg will develop into a female; while when one of the spermatozoa containing a small Y chromosome enters it will give rise to a male. Leaving aside the common chromosomes of both sexes, a fertilized egg containing XX gives rise to a female, while one containing XY gives rise to a male. There is in this case as in that of Protenor a preponderance of chromosome material in the female, but this quantitative difference is not essential for the determination of sex, since in some species the Y chromosome may be as large as the X chromosome.
The main fact is that the female cells have the chromatin composition XX, the male cells the composition XY, where Y is apparently qualitatively different and often, but not necessarily, smaller than X, or entirely lacking.
It may be mentioned in passing that indirect evidence exists indicating that in man there are also two kinds of spermatozoa and one kind of egg, and that sex depends on whether a male determining or a female determining spermatozoön enters the egg.
2. This mode of sex determination holds only for those animals in which there is one type of egg and two types of spermatozoa. Experimental evidence furnished first by Doncaster in 1908 on a moth, Abraxas, indicated that a number of other forms exists in which matters are reversed, inasmuch as there are two types of eggs and one type of spermatozoa. This condition of affairs exists not only in the moth Abraxas, but also in the fowl as shown by Pearl. In these forms it is assumed that all the spermatozoa have one sex chromosome X, while there are two types of eggs, one possessing the sex chromosome X, the other possessing Y. When a spermatozoön enters an egg with an X chromosome, the egg will give rise to a male, while if it enters a Y egg, a female will arise. The evidence pointing toward this result is chiefly contained in experiments on sex-limited or more correctly sex-linked heredity; i. e., a form of heredity which follows the sex in a peculiar way. Thus colour-blindness is a case of sex-linked inheritance, since this abnormality appears overwhelmingly in the male offspring of a colour-blind person. Doncaster crossed two varieties of Abraxas differing in one character which was sex-linked, and the results of his crossings indicated that in this form there are two types of eggs and one type of spermatozoa.184
These observations on sex-linked heredity confirm the idea that the sex chromosomes determine the sex. The most extensive and conclusive experiments along this line are those by Morgan on the fruit fly Drosophila. In this form there are two kinds of spermatozoa and one kind of eggs; the egg has one X chromosome, while one-half of the spermatozoa has an X the other a Y chromosome; the entrance of the latter into an egg gives rise to a male, of the former to a female.
While the eyes of the wild fruit fly Drosophila ampelophila are red, Morgan185 noticed in one of his cultures a male that had white eyes. This white-eyed male was mated to a red-eyed female. The offspring, the F1 generation, were all red eyed, males as well as females. These were inbred and now gave in the F2 generation the following three types of offspring:
| (1) 50 per cent. females, all with red eyes. | ||
| (2) 50 per cent. males | 25 per cent. with red eyes. | |
| 25 per cent. with white eyes. | ||
The character white eye was therefore transmitted only to half the grandsons; it was a sex-linked character. It is known from a study of the pedigrees of colour-blind individuals that if the corresponding experiment had been carried out with them, instead of with white-eyed flies, the same proportions of normal and colour-blind would have been found: namely, normal colour vision in the F1 generation, in both males and females, and half of the males of the F2 generation colour-blind, the other half and all the females with normal vision. Of course, in man, intermarriage between two different F1 strains would have been required in place of the inbreeding of the F1 generation, which took place in Morgan’s experiments. Morgan interprets his experiments as follows. The normal red-eyed Drosophila has one kind of eggs, each possessing one X chromosome. This X chromosome has also the factor for the development of red-eye pigment. The white-eyed male has two kinds of spermatozoa, one with an X chromosome, the other with a Y chromosome, both lacking the factor for red-eye pigment. If we designate the X chromosome with the factor for red-eye pigment by X and the X and Y chromosomes lacking the factor for redness with X and Y the following combinations must result if we cross a normal red-eyed female with a white-eyed male:
| Eggs | Sperm | Result |
| X | X | XX red-eyed female |
| X | Y | XY red-eyed male |
It is obvious that all the offspring of the first generation (the F1 generation) must be red eyed, since all the eggs have one X chromosome with the factor for red. According to the results obtained from cytological studies which will be explained in the next chapter, the females with the chromatin constitution XX will form two types of eggs in equal numbers: namely, eggs with an X and eggs with an X, i. e., all eggs have one X chromosome, but in fifty per cent. of the eggs the X has the factor for red, in fifty per cent. this factor is lacking (X). The males having the chromosome constitution XY form two types of spermatozoa, one with an X possessing the factor for red pigment and one, the Y chromosomes, lacking this factor. If inbred the next F2 generation will give rise to the following four types of offspring: (1) XX, (2) XX, (3) XY, (4) XY, all four types in equal numbers.
(1) and (2) give females, both red eyed, since both contain a red-factored X chromosome. (3) and (4) give males, (3) giving rise to red-eyed males, since it contains a red-factored X chromosome, (4) producing males with white eyes since this X chromosome is lacking the factor for red eyes. Since all four combinations must appear in equal numbers (provided the experimental material is ample enough, which was the case in these experiments), in the F1 generation both males and females should have red eyes and in the F2 generation all the females should have red eyes and half of the males should have red, half white eyes. These results were obtained.
The experiments were carried further. No white-eyed females had appeared thus far. On the same assumptions of the relation of the X, X, and Y chromosomes to the heredity of sex as well as to eye colour it was possible to predict under what conditions and in which proportions white-eyed females should arise. Thus if a red-eyed female of the F1 generation (a cross between white-eyed male and normal female) be mated with a white-eyed male the result should be an equal number of white-eyed males and white-eyed females if the chromosome theory of sex determination were correct. The reasoning would be as follows:
The red-eyed female, having the chromosome constitution XX should form two kinds of eggs in equal numbers with the constitution X and X; the white-eyed male having the chromosome constitution XY should form two kinds of spermatozoa X and Y. The following four types of individuals must then be produced in equal numbers:
(1) XX, (2) XX, (3) XY, and (4) XY.
In this case (2) must give rise to white-eyed females and (4) to white-eyed males, while (1) must give rise to red-eyed females and (3) to red-eyed males. Hence white-eyed males and females and red-eyed males and females are to be expected in this case in equal numbers, and this was actually observed.
The numerical agreement in this and the other experiments between the expected and observed result cannot well be an accident. The fact that the inheritance of sex-linked characters in man follows the same laws as in Drosophila is a strong argument in favour of the assumption that in man, also, sex is determined by two kinds of spermatozoa.
Morgan and his students discovered no less than thirty-six sex-linked characters in Drosophila, and each behaved in a similar way to the red and white eye colour in regard to sex-linked inheritance, so that the chromosome theory of sex determination rests on a safe basis. That sex is merely determined by the number of X chromosomes, not by the Y chromosome, is proved by the facts that the Y chromosome may be completely absent as in Protenor and that Bridges186 has found a type of female Drosophila with a chromosome formula XXY whose sex was not affected by the supernumerary Y.
3. On the basis of all these experiments and theories it is comparatively easy to explain a number of phenomena concerning sex ratios which before had been very puzzling. In bees it had been shown many years ago by Dzierzon that the males develop from unfertilized eggs while the females, queens and workers, develop from fertilized eggs. This is intelligible on the assumption that the unfertilized egg contains only one X chromosome while the spermatozoön carries into the egg the second X chromosome. But if the male bee produces two types of spermatozoa we should expect that only one-half of the fertilized eggs should be females, the other half males. But it happens that of the two types of spermatozoa only one is formed since in one of the cell divisions which lead to the formation of spermatozoa one viable spermatozoön only is formed while the other one perishes. It is, therefore, quite possible that it is the female-producing spermatozoön which survives while the male-producing spermatozoön dies.
It is occasionally observed that an insect shows one sex on one side of its body and the opposite sex on the other side. Boveri suggested that this phenomenon of gynandromorphism is due to the fact that the spermatozoön for some unknown reason does not fuse with the egg nucleus until after the egg has undergone its first cell division. In this case it fuses with the nucleus of one of the two cells into which the egg divides (or in some cases even one of the later cells?). As a consequence the one-half of the embryo which arises from the cell which was not fertilized would have only one X chromosome and in a case like the bee would develop parthenogenetically, while the other half of the body, developing from the cell into which a spermatozoön has penetrated, would be fertilized. The latter half of the body would be female, the former male. In his last paper before his untimely death, Boveri has given proof for the correctness of this interpretation as far as gynandromorphism in the bee is concerned.187
It seems to be generally true that where sexual reproduction leads only to the formation of females the case finds its explanation in the fact that the male-producing spermatozoa perish and only the female-producing spermatozoa survive. Such an observation was made by Morgan on a certain species of phylloxerans.
The slight preponderance in the number of one sex which is occasionally found—an excess of six per cent. males over females in the human race—may well find its explanation on the assumption of a slightly greater mortality of the female-determining spermatozoa.
In certain forms parthenogenetic and sexual reproduction may alternate in a cycle, e. g., in plant lice, Daphnia, and rotifers. In plant lice it has been observed for a long time that when the plant is normal and the weather warm the aphides remain wingless, reproduce parthenogenetically, and only females exist, and this may last for years and for more than fifty generations; but that when the plant is allowed to dry out both sexes appear.
Here we are dealing with a limited determination of sex inasmuch as the experimenter has it in his power to prevent or allow the production of males. The facts do not in all probability contradict the statements made concerning the rôle of the X chromosomes in the determination of sex. We have seen that where sex is determined by two types of spermatozoa one type of eggs is produced which possesses only one X chromosome. Such eggs might produce males if not fertilized (as they do in bees), but they cannot produce females because for that purpose they must have two X chromosomes. It has been shown for certain cases, and it may be true generally, that if eggs of this type give rise to parthenogenetic females they may do so because they have for some reason two X chromosomes. Usually such an egg loses one of the X chromosomes in a process of nuclear division (the so-called reduction division) which usually precedes fertilization. If this reduction division is omitted the egg has two X chromosomes and if such an egg develops parthenogenetically it gives rise to a female. These cases do not, therefore, contradict the connection between X chromosomes and sex determination established by cytological observations and breeding experiments, on the contrary, they confirm it. The question remains: How can external conditions bring it about that the reduction division is omitted? To this question no definite answer can be given at present.
We may in passing mention the well-known observation that twins which originate from the same egg always have the same sex; while twins arising from different eggs show the usual variation as to sex. Twins coming from one egg have the same chorion and can thereby be diagnosed as such. They can be produced as we have stated in Chapter V by a separation of the first two cleavage cells of the egg, each one giving rise to a full embryo. It harmonizes with all that has been said above that the sex of two such individuals must be the same since they have the same number of X chromosomes, the latter being determined in the human race by the nature of the spermatozoön which enters the egg.
4. While thus far all the facts agree with the dominating influence of certain chromosomes upon sex determination, one group of facts has not yet been explained: namely, hermaphroditism. By hermaphroditism is meant the existence of complete and separate sets of female and male gonads in the same individual. This condition exists regularly not only in definite groups of animals, e. g., certain snails, leeches, tape-worms, but also, as everybody knows, in flowering plants. While in some forms both kinds of sex cells, male and female, are formed and mature simultaneously, as, e. g., in the Ascidian Ciona (see Chapter IV), in others they are formed successively, very often the spermatozoa appearing first (protandric hermaphroditism). In the long tapeworm Tænia each ring has testes and ovaries, but the young rings are only male while in the older rings the testes disappear and the ovaries are formed. The same ring is in succession male and female. How can we reconcile the facts of hermaphroditism with the chromosome theory of sex determination? Rhabdonema nigrovenosum, a parasite living in the lungs of the frog, is hermaphroditic, but its eggs produce not a hermaphroditic generation but one with the two separate sexes; this generation is not parasitic and lives in the soil. The generation produced by these separate males and females gives rise again to a hermaphrodite which migrates into the lungs of the frogs. According to Boveri and Schleip188 the cells of the hermaphrodite have twelve chromosomes. It produces two types of spermatozoa with six and five chromosomes respectively (one-half of the cells losing one chromosome which is left at the line of cleavage between the two cells); and one type with six chromosomes. In this way separate males and females are produced by the hermaphrodite, females with twelve and males with eleven chromosomes.
The males produce again two kinds of spermatozoa, male and female producing, but the male-producing spermatozoa become functionless. This fusion of the other spermatozoön containing six chromosomes with an egg having six chromosomes leads again to the formation of the hermaphrodite with twelve chromosomes. It is obvious that in this case the cause for the hermaphroditism is not disclosed. If chromosomes have anything to do with hermaphroditism there must be an undiscovered element in the chromosomes which may explain why the female as well as the hermaphrodite have the same chromosome constitution; or we are forced to look for another determinant outside the X chromosomes or the chromosomes altogether. This seems to be the only cytological work on the problem of hermaphroditism. Experimental work has been begun by Correns189 and by Shull on the determination of hermaphroditism in plants but lack of space forbids us to give details.
5. As stated at the beginning of this chapter, the chromosome theory of sex determination explained only one feature of the problem, namely, the relative numbers in which both sexes or only one sex, as the case may be, are produced; and in this respect the evidence is so complete that we must accept it. But with all this, the problem of sex determination is not exhausted, since a physiological solution of the problem of sex determination demands an account of how the sex chromosomes can induce the formation not only of ovaries and testes but also of the other sex characters. For the solution of this problem biology will have to depend largely on experiments in which it is possible to influence the formation of sex characters and of the sex glands themselves.
The most striking observations in this direction were made by Baltzer on a marine worm, Bonellia. In this animal the two sexes are very different, the male being a tiny parasite, a few millimetres in length, which spends its life in the uterus of the female, whose size is about five centimetres. A female carries as a rule several and often a large number of the male parasites in its uterus, which indicates that the males prevail numerically. The fertilized eggs of the animals are laid in the sea water where the larvæ hatch. At the time of hatching all larvæ are alike. The differentiation of the larvæ into the dwarf males and the giant females can be determined at will. The larvæ have a tendency to attach themselves to the proboscis of the female as soon as they hatch. If given a chance to do so and if they stick to the proboscis for more than three days they will develop into males, which soon afterwards creep into the female where they continue their parasitic existence. If, however, no adult female Bonellia is put into the aquarium in which the larvæ hatch, about ninety per cent. of the larvæ will, after a period of rest, develop into females; the rest develop into males. Those which develop into females will often show a primary maleness which may manifest itself in the production of sperm or of other secondary male sexual characters. This tendency is stronger the longer the period of rest lasts. If the larvæ are allowed to settle on the proboscis of the adult female but are removed too early hermaphrodites are produced having male and female characters mixed.
Baltzer has suggested on the basis of some observations that the larvæ while on the proboscis of the female absorb some substance secreted by the proboscis, and this substance accelerates the further development into a male and suppresses the female tendency. If this substance from the proboscis does not reach the larvæ the tendency to become males is gradually suppressed in the majority and only a few develop into pure males or protandric hermaphrodites, while the female characters are given a chance to develop. Baltzer assumes, therefore,—as it seems to us correctly—that in all larvæ the tendency for both sexual characters is present, that they are, in other words, hermaphrodites, but the chance for the suppression of one and the development of the other group of characters can be influenced by certain chemical substances which the larva may take up.190
Giard has studied the effects of a curious form of castration brought about by parasites, which is followed by a change in the sexual character of the castrated animal. The phenomenon is very striking in certain forms of crabs when they are attacked by a parasitic crustacean, Sacculina. The two sexes differ in the crab Carcinus mænas by the form of the abdomen, but when a male is attacked by the parasite its abdomen assumes the female shape. Smith observed in another crab that in such cases even the abdominal appendages of the male may be transformed into those of a female. The transformation is so complete that the older observers had reached the conclusion that the parasite attacked only the females, since they overlooked the fact that the castration by the parasite transformed the secondary sexual characters of the male into those of a female.
Giard observed that in a diœcious plant, Lychnis dioica, a parasitic fungus brings about the transformation of the host into a hermaphrodite.
G. Smith has discovered a fact which shows that chemical changes must underlie these morphological transformations of primary or secondary sexual characters. He noticed that in male crabs the presence of the parasite Sacculina changes the contents of the fatty constituents in the blood, making them equal to that of the female. Vaney and Meignon had previously shown that during the chrysalid stage the female silkworms have always more glycogen and less fat than the males. The castration by parasites is paralleled by what Caullery calls the castration by senility.191 In certain birds and also in mammals at the time when the sexual glands cease to function certain secondary sexual characters of the other sex make their appearance. The most common case is that certain secondary male characters appear in the old female (exceptionally also in the young female with abnormal ovaries) (arrhenoidy). Thus old female pheasants assume the plumage of the male, and in the human female after the menopause and especially among sterile women a beard may begin to grow. The opposite phenomenon, the old male assuming female characters, is not so common. Very interesting observations on changes in the plumage of castrated fowl have recently been made by Goodale.192
It had long been observed by cattle breeders that in the case of twins of different sex the female—the so-called free-martin—is usually sterile. F. Lillie193 has recently discovered the cause of this interesting phenomenon. Such twins originate from two different eggs since the mother has two corpora lutea, one in each ovary. In normal single pregnancies in cattle there is never more than one corpus luteum present. The two eggs begin to develop separately in each horn of the uterus.
The rapidly elongating ova meet and fuse in the small body of the uterus at some time between the 10 mm. and the 20 mm. stage. The blood-vessels from each side then anastomose in the connecting part of the chorion; a particularly wide arterial anastomosis develops, so that either fetus can be injected from the other. The arterial circulation of each also overlaps the venous territory of the other, so that a constant interchange of blood takes place. If both are males or both are females no harm results from this; but if one is male and the other female, the reproductive system of the female is largely suppressed, and certain male organs even develop in the female. This is unquestionably to be interpreted as a case of hormone action.
The reproductive system of these sterile females is for the most part of the female type, though greatly reduced. The gonad is the part most affected; so much so that most authors have interpreted it as testis.
It should be added, however, that this result cannot at present be generalized, since in the hermaphrodites the specific hormones of both sexes must circulate without suppressing each other’s efficiency.
All these facts indicate that certain substances secreted by the ovaries or testes may inhibit the development of certain sexual characters of the opposite sex. When these inhibitions are partly or entirely removed the secondary sexual characters of the opposite sex may appear. This fact may also be interpreted as an indication of a latent hermaphroditism and if this be correct the real and latent hermaphrodites differ only by the degree of inhibition for one sex, this inhibition being lacking or less complete in the real than in the latent hermaphrodite.
In the light of this conclusion the observations on the regeneration of both ovaries and testicles which Janda observed in a hermaphroditic worm, Criodrilus lacuum,194 is no longer so mysterious. This worm normally possesses in the segments near the head a pair of ovaries and several pairs of testes. Janda found that if the anterior parts containing the gonads of these worms are cut off a complete regeneration takes place, including both types of gonads, ovaries as well as testes. As a rule, more than one pair of ovaries appear in the regenerated piece. This important experiment shows that in a hermaphrodite both types of sex organs can be produced from body cells or from latent buds resembling body cells. This phenomenon would be intelligible on the assumption that in the body of a hermaphrodite substances circulate which favour the development of both types of sex organs, while in a diœcian animal probably only one type of sex organ would be developed; the formation of the other being inhibited.
Richard Goldschmidt has discovered in his breeding experiments on the gipsy-moth (Lymantria dispar) a phenomenon which will probably throw much light on the physiology of sex determination. He found that certain crosses between the Japanese and the European gipsy-moth do not give pure sexes, males or females, but mixtures of the sexual characters of both sexes, and this mixture is a very definite one for definite crosses. These differences are such that it is possible to grade the hybrids according to their manifestations of maleness or femaleness, both in morphological characters and instincts. Goldschmidt calls this peculiar phenomenon intersexualism, and its essential feature is that the various degrees of intersexualism can be produced at will by the right combination of races.
Female intersexualism begins with animals which show feathered antennæ of medium size (feathered antennæ are a male character), but which are otherwise entirely female in appearance except that they produce a smaller number of eggs which are fertilized normally. In the next stage patches of the brown male pigment appear on the white female wings in steadily increasing quantity. The instincts are still female, the males are attracted and copulate. But the characteristic egg sponge laid by the animal contains nothing but anal hairs in spite of the fact that the abdomen is filled with ripe eggs. In the next stage whole sections of the wings show male colouration, with cuneiform female sectors between, the abdomen becomes smaller, contains fewer ripe eggs, the instincts are only slightly female, the males are attracted very little, and reproduction is impossible. In the next stage the male pigment covers practically the whole wing, the abdomen is almost male, but still contains ovaries with a few ripe eggs, the instincts are intermediate between male and female. Then follow very male-like animals which still show in different organs their female origin and have rudimentary ovaries. . . . The end of the series is formed by males, which show in some minor characters, such as the shape of wings, still some traces of their female origin.
The series of the male intersexes starts with males showing a few white female spots on their wings. These become larger and larger, the amount of brown pigment correspondingly decreasing. . . . Hand in hand with this the abdomen increases in size, reaching in the most extreme cases two-thirds of the female size (without containing eggs). The same is true for the instincts which become more and more female.
(And also for the copulatory organs which also become more and more female.)
As stated above, the main fact that every desired degree of intersexualism can be produced at will by properly combining the races for breeding, and the intersexual potencies of the different races has been worked out by Goldschmidt.195
6. The relation between chemical substances circulating in the body—either derivatives of food taken up from without or of chemical compounds formed naturally inside the body—and the production of sexual characters is best shown in the polymorphism found among the social ants, bees, and wasps. Here we have, as a rule, in addition to the two sexes a third one, the workers, which are in reality rudimentary and for that reason sterile females. They differ more or less markedly from both the typical male and female in their external form, and, as a rule cannot copulate owing to their deficient structure. This third sex, the sterile neuters, can be transformed at desire into sexual females in certain species, as P. Marchal has demonstrated. He worked with a form of social wasps in which the workers are sterile and smaller than the real females. In such a society of wasps all the males and workers die in the fall and only the fertilized females survive, each one founding a new nest in the following spring. From the first eggs laid, workers arise, small in stature and sterile; these workers are nourished by their mother. Then these workers take care of the feeding of all those larvæ which arise from the eggs which their mother continues to lay. Throughout the spring only workers arise from the eggs. The males appear in the summer, the real females towards the end of the season when the sexes copulate.
Marchal isolated a number of the sterile workers, providing them with food but giving them no larvæ to raise. He found that the workers which thus far had been sterile became fertile, producing, however, only males. This latter fact is easily understood from what has been said regarding the bees, namely, that the female produces only one type of eggs, hence the unfertilized egg can give rise only to males. The astonishing or important point is that the ovaries of the workers begin to develop as soon as they no longer have a chance to nourish the larvæ, provided the food which would have been given to the larvæ is now at their disposal. In other words, the development of their ovaries is the outcome of eating the food which under normal conditions they would have given to the larvæ. The food must, therefore, contain a substance which induces the development of eggs. The natural sterility of the neuters or workers is, therefore, to use P. Marchal’s expression, a case of “food castration,” (“castration nutriciale”).196 The workers originate from fertilized eggs and are therefore females, but for the full development of the ovaries and the other sexual characters something else besides the XX chromosomes is needed and this is supplied in this case by the quantity or quality of the food. May we not conclude that the same thing may happen generally, except that these substances are formed by the body under the normal conditions of nutrition through the influence of constituents of the second X chromosome?
It is known that the future queens among the bees receive also a special type of food which the workers do not receive. Again the idea of “food castration” of the latter is suggested.
In rotifers Whitney197 has shown that the cycle in the production of males and females can be regulated by the food. In some species a scanty supply of green flagellates produced purely female offspring, while a copious diet of the same green flagellates produced a predominance of male grandchildren, sometimes as high as ninety-five per cent. This was confirmed by Shull and Ladoff.198
7. The effects of the removal of the ovaries or testes upon the development of secondary sexual characters differ for different species. In insects the secondary sexual characters are not altered by an operative removal of the sexual glands as in the caterpillar, e. g., Ocneria dispar, according to Oudemans. This result has been invariably confirmed by all subsequent workers, especially by Meisenheimer. Crampton grafted the heads of pupæ of butterflies upon the bodies of other specimens of the opposite sex, but the sexual characters of the head remained unaltered.
In vertebrates, however, there exists a distinct influence of a secretion from the sexual glands upon the development of certain of the secondary sexual characters, which do not develop until sexual maturity. In a way the observations on arrhenoidy and thelyidy referred to above are indications of this influence.
Bouin and Ancel had already suggested that the sexual glands of mammals have two independent constituents, the sexual cells and the interstitial tissue; and that the latter tissue is responsible for the development of the secondary sexual character. This has been proved definitely by Steinach,199 who showed that when young rats are castrated certain secondary sexual characters are not fully developed. The seminal vesicles and the prostate remain rudimentary and the penis develops incompletely. Such animals when adult recognize the female and seem to follow it, but do not persist in their attention and neither erection nor cohabitation occurs. When, however, the testes are retransplanted into the muscles of the castrated young animal (so that they are no longer connected with their nerves) seminal vesicles, prostate, and penis develop normally, and these animals show normal sexual ardour and cohabitate with a female although the female cannot become pregnant since the males cannot ejaculate any sperm. When the retransplanted testes were examined it was found that all the sperm cells had perished, only the interstitial tissue of the testes remaining. It was, therefore, proved that the development of the seminal vesicles, the prostate, the penis, and the normal sexual instincts and activities depends upon the internal secretions from this interstitial tissue and not upon the sex cells proper. This agrees with the conclusions at which Bouin and Ancel had arrived by ligaturing the vasa deferentia of male animals.
Steinach in another series of experiments castrated young male rats and transplanted into them the ovaries of young females. These ovaries did not disintegrate, the eggs remaining, and corpora lutea were formed. In such feminized individuals the seminal vesicles, prostate, and penis did not reach their normal development, and it was thereby proved that the internal secretions from the ovary do not promote the growth of the secondary sexual male characters. On the contrary, Steinach was able to show that the growth of the penis was directly inhibited by the ovary, since in the feminized males this organ remained smaller than in the merely castrated animals. On the other hand the infantile uterus and tube when transplanted into the young male with the ovaries grow in a normal way, and Steinach thinks that pregnancy in such feminized males is possible if sperm be injected into the uterus. In some regards the feminized males showed the morphological habitus of females. Soon after the transplantation of ovaries into a castrated male the nipples of its mammary glands begin to grow to the large size which they have in the female and by which the two sexes can easily be discriminated. In addition the stronger longitudinal growth of the body in the male does not occur in the feminized specimens, the body growth becomes that of a female; and likewise the fat and hair of the feminized male resemble that of a real female.
While the castrated males show an interest in the females, the feminized males are absolutely indifferent to females and behave like them when put together with normal males; and, what is more interesting, they are treated by normal males like normal females. The sexual instincts have, therefore, also been reversed in the feminized males by the substitution of ovaries for testes.
The inhibition of the growth of the penis by the ovary is of importance; it supports the idea already expressed that in hermaphrodites this inhibition of the growth of the secondary organs of the other sex is only feeble or does not exist at all.
We may finally ask whether there is any connection between the cytological basis of sex determination by special sex chromosomes and the physiological basis of sex determination by specific substances or internal secretions. It is possible that the sex chromosomes determine or favour, in a way as yet unknown, the formation of the specific internal secretion discussed in the second part of this chapter. In this way all the facts of sex determination might be harmonized, and it may become clear that when it is possible to modify secretions by outside conditions or to feed the body with certain as yet unknown specific substances the influence of the sex chromosomes upon the determination of sex may be overcome.
1. The scientific era of the investigation of heredity begins with Mendel’s paper on plant hybridization which was not appreciated by his contemporaries. Mendel invented a method for the quantitative study of heredity which consisted essentially in crossing two forms of peas differing only in one well-defined hereditary character; and in following statistically and separately the results of this crossing and that of the inbreeding of the second and third generations of hybrids. This led him to the recognition of one essential feature of heredity; namely, that while the hybrids of the first generation are all alike, each hybrid produces two types of sex cells in equal numbers, one for each of the pure breeds which has been used for the crossing. This takes place not only when the forms used for the crossing differ in regard to one character only but also if they differ for two or more characters. The statement made is Mendel’s law of heredity, or, more correctly, Mendel’s law of the segregation of the hereditary characters of the parents in the sex cells of the hybrids.201 Mendel’s law allows us to tabulate and calculate beforehand the relative number of different forms which appear if the offspring of a mating of two varieties are bred among themselves.
In order to do this it must be remembered also that while in some cases the hybrid is an intermediate between the two parent forms, in other cases it cannot be discriminated from one of the two parent forms. In such cases the character which appears in the hybrid was called by Mendel the dominant character and the one which disappeared the recessive character. According to Bateson, who was the first to systematize the phenomena of Mendelian heredity, recessiveness means generally the absence of a character which is present in the dominant type. When, e. g., the cross between a tall and a dwarf form of pea gives in the first generation only tall peas, on the basis of the presence and absence theory the dominant form contains a factor for growth which is lacking in the dwarf form. While this theory fits many cases it meets with difficulties in others. Thus the presence of a factor for pigment should be dominant over the absence of such a factor, which is usually the case, inasmuch as the cross of a coloured rat or rabbit with an albino is black or coloured. There is, however, also a case where whiteness is dominant over colour, as we shall see later. This fact does not necessarily contradict the presence and absence theory.202
When two pure breeds of parents differ in one character, e. g., two varieties of beans, one with a violet the other with a white flower, the cross between the two species (the F1 generation) has pale violet flowers, approximately intermediate between the two parents. If these hybrids are bred among themselves the offspring is called the F2 generation. According to Mendel’s law the hybrids of the first F1 generation all have two kinds of eggs in equal numbers, one kind representing the pure breed of the parents with violet, the other of the pure breed with white flowers. The same is true for the pollen cells. Hence the following possible combinations must appear in the offspring when the pale violet hybrids are inbred:
The four possible combinations are: (1) violet—violet; (2) violet—white; (3) violet—white; (4) white—white. The first will result in pure violet flowers, the fourth in pure white, and the second and third in pale violet flowers. Since all four combinations will appear in equal numbers when the number of crossings is sufficiently large the numerical result will be:
violet : pale violet : white = 1 : 2 : 1
Fifty per cent. of the F2 generation will be pale violet, 25 per cent. violet, and 25 per cent. white. The violets and whites each will breed true when bred among themselves since they are pure, and produce only one type of eggs and pollen. The pale violets are hybrids and will again produce the two types of eggs and pollen, that is, if bred among themselves will again give violets, pale violets, and whites in the ratio 1:2:1. This the experiment confirms.
As has been stated, it not infrequently happens that all the hybrids of the first generation are alike. In such cases the one character is “recessive,” i. e., overshadowed or covered by the other the “dominant” character, which alone appears in the hybrids. Thus when Mendel crossed peas having round seeds with peas having angular seeds all the hybrids had round seeds. The round form is dominant, the angular recessive, i. e., all the hybrids have round seeds. When these hybrids were bred among themselves the next generation produced round and angular seeds in the ratio of 3:1 (5474 round to 1850 angular). The explanation is as follows. Let R denote round, A angular character; the pure breeds of parents have the gametic constitution RR and AA respectively. When crossed, all the offsprings have the constitution RA and since A is recessive this hybrid generation resembles the pure RR parents. The F1 generation produces two kinds of eggs R and A and two kinds of pollen R and A in equal numbers, and these if inbred give the following four combinations in equal numbers:
RR, RA, AR, AA.
Since RA, AR, and RR all give round seeds the F2 generation produces round seeds to angular seeds in the ratio of 3:1. The two organisms with the gametic constitution RR and RA look alike, yet they are different in regard to heredity. The gametically pure form RR is called homozygous, the impure form RA heterozygous.
2. W. S. Sutton203 was the first to show that the behaviour of the chromosomes furnishes an adequate basis on which to account for Mendel’s law of the segregation of the characters in the sex cells of the hybrids. If we disregard the cases of parthenogenesis and the X chromosomes, we may state that each species is characterized by a definite number of chromosomes, e. g.204
| man (probably) | 24 | corn | 20 |
| mouse | 20 | evening primrose | 7 |
| snail (Helix hortensis) | 22 | nightshade | 36 |
| potato beetle | 18 | tobacco | 24 |
| cotton | 28 | tomato | 12 |
| four o’clock | 16 | wheat | 8 |
| garden pea | 7 |
In the fertilization of the egg the number of chromosomes is doubled (if we disregard for the moment the complication caused by the X and Y chromosomes which was considered in the previous chapter). It was noticed by Montgomery that each chromosome had a definite size and individuality, and he suggested that homologous chromosomes existed in sperm and egg and that in fertilization the homologous chromosomes of egg and sperm always joined and fused in the special stage designated as synapsis, which will interest us later. On the basis of this suggestion Sutton developed the chromosome theory of the mechanism of Mendelian heredity or segregation.
According to this theory, all the cells of an individual (inclusive of the egg cells and sperm cells) have two sets of homologous chromosomes, one from the father, the other from the mother. Before the egg and sperm are ready for the production of a new individual, each loses one set of homologous chromosomes in the so-called reduction division, but the lost set is made up indiscriminately of maternal as well as paternal chromosomes, so that while one egg retains the maternal chromosome A the other will retain the paternal one, and so on. If before the reduction division all the eggs had the chromosome constitution AA1, BB1, CC1, DD1 (where A B C D are the paternal and A1 B1 C1 D1 the maternal chromosomes), after the reduction division each daughter cell has a full set of four chromosomes, but maternal and paternal mixed. Thus the one cell may have AB1CD1, the other A1B1C1D1, etc. This, according to Sutton, is the basis of the Mendelian heredity. Suppose the determiner of a certain character (violet colour of flower in the bean) is located in a chromosome A of this species. The homologous chromosome in beans with white colour may be designated as a. According to the chromosome theory of Mendelian heredity a differs from A in one point, though this difference is probably only of a chemical character and not visible.
If an egg with A is fertilized by a pollen with a (or vice versa), after fertilization the chromosome constitution of the fertilized egg is Aa. All the other homologous chromosomes are identical and therefore need not be considered. All the nuclei of the F1 generation have the chromosome constitution Aa. All will form eggs and pollen with nuclei of the same chromosome constitution Aa, but all these sex cells will go through the maturation division before they are fertilized; and this reduction division leads to the existence of two kinds of eggs in equal numbers, one containing only the A, the other only the a chromosome; and the same happens in the pollen. When therefore the hybrids F1 are mated among themselves, the following four chromosome combinations will be produced:
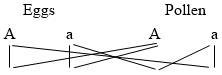
Possible combinations in fertilized eggs AA, Aa, aa, in the ratio 1:2:1.
Now this is exactly the ratio of Mendelian heredity in the F2 generation. The plant with the chromosome constitution AA will form violet flowers, those with the chromosome constitution Aa will form pale violet flowers, and those with the chromosome constitution aa will form white flowers.
To quote Sutton’s words:
The result would be expressed by the formula AA: Aa: aa which is the same as that given for any character in a Mendelian case. Thus the phenomena of germ cell division and of heredity are seen to have the same essential features viz., purity of units (chromosomes, characters) and the independent transmission of the same; while as a corollary it follows in each case that each of the two antagonistic units (chromosomes, characters) is contained by exactly half the gametes produced.
It is obvious that Sutton by this idea did for heredity in general what McClung had done for sex determination or sex heredity, that is, he showed that the numerical results obtained in Mendelian heredity can be accounted for on the basis that factors for hereditary characters are carried by definite chromosomes. The cytological basis of sex determination becomes only a special case of the cytological basis of Mendelian heredity. In the examples quoted the plants giving rise to violet and to white flowers are homozygous for the colour of flower having the chromosome constitution AA and aa respectively; while the plants with pale violet flowers are heterozygous, having the chromosome constitution Aa in their nuclei. The former give rise to identical sex cells A and A or a and a; while the heterozygous plants give rise to different sex cells A and a.
From this point of view in Drosophila (and very probably also in man) the female is homozygous for sex having in all its cells the critical chromosome constitution XX and giving rise to one type of eggs only, each with one X chromosome; while the male in these forms is heterozygous for sex having in all its cells the chromosome constitution XY and forming two different types of spermatozoa in equal numbers X and Y. In Abraxas and in the fowl the female is heterozygous for sex and the male homozygous.
3. If the chromosomes are the vehicle for Mendelian heredity it should be possible to show that the various hereditary characters which follow Mendel’s law must be distributed over the various chromosomes; and it should be possible to find out which characters are contained in the same chromosome. It has already been stated that sex-linked heredity is intelligible on the assumption that the X chromosome carries the sex-linked characters. T. H. Morgan and his pupils have shown with the greatest degree of probability that corresponding linkages occur in the other chromosomes and that there are in Drosophila exactly as many groups of linkage as there are different chromosomes, namely four.205
Mendel had found that when he crossed two species of peas differing in regard to two pairs of characters, he obtained in the F2 generation results which he calculated on the assumption that the segregation of the two pairs of characters in the sex cells of the hybrids took place independently of each other. To illustrate by an example: When crossing a yellow round pea with a green wrinkled variety in which the characters round and yellow are dominant, green and wrinkled recessive, all the hybrids of the F1 generation had the characters round and yellow. When these were inbred the F2 generation produced four types of seed in the ratio 9: 3: 3: 1, namely:
(1) yellow round (315 seeds)
(2) yellow wrinkled (101 seeds)
(3) green round (108 seeds)
(4) green wrinkled (32 seeds)
The explanation according to Mendel’s theory is as follows: Since the segregation of each pair of characters occurs independently, there must be 3 yellow to 1 green and also 3 round to 1 wrinkled in the F2 generation. The yellow will, therefore, be round and wrinkled in the ratio of 3:1, which will give 9 yellow round to 3 yellow wrinkled. The green will also be round and wrinkled in the ratio of 3:1, which will give 3 green round to 1 green wrinkled, which is the ratio of 9: 3: 3: 1 found by Mendel.
On the basis of the chromosome theory the following explanation could be given of this numerical relation. The peas with yellow round seeds have sex cells with a factor for both yellow and for round in two different chromosomes; these two different chromosomes we will designate with Y and R. The peas with green and wrinkled seeds will have in their sex cells factors for these characters in two homologous chromosomes g and w, where g is the homologue of Y and w of R. The cells of the hybrids of the F1 generation will have the chromosome constitution Yg Rw, where Y and g and R and w are homologous chromosomes which will lie alongside each other YRgw. In the formation of sex cells a reduction of these four chromosomes to two takes place whereby, according to the theory of Sutton, the following two types of separation can take place: YR and gw, or gR and Yw. (A separation into Yg and Rw is impossible since the division takes place only between homologous chromosomes.) Hence there will be four types of eggs, YR, gw, gR, and Yw and the same four types of pollen cells. The F2 generation will produce the sixteen possible combinations in equal numbers: namely,
YRYR YRgw YRgR YRYw
gwYR gwgw gwgR gwYw
gRYR gRgw gRgR gRYw
YwYR Ywgw YwgR YwYw
Since w and g are recessives and therefore disappear when in combination with their respective dominants Y and R the result will be 9 YR (yellow round), 3 Yw (yellow wrinkled), 3 Rg (round green), and 1 gw (green wrinkled) as Mendel actually observed and as all investigators since have confirmed.
Bateson made the discovery that these Mendelian ratios 9: 3: 3: 1 did not always occur when forms differing in two characters were crossed. He found typical and very constant deviations from this ratio in definite cases and these cases he interpreted as being due to “gametic coupling.”
These phenomena demonstrate the existence of a complex interrelation between the factorial units. This interrelation is such that certain combinations between factors may be more frequent than others. The circumstances in which this interrelation is developed and takes effect we cannot as yet distinguish, still less can we offer with confidence any positive conception as to the mode in which it is exerted.206
Morgan has given an ingenious explanation of these deviations on the basis of the chromosome theory of Mendelian heredity. He assumes that they occur in those cases where the two or more characters are contained in the same chromosome. In that case the two factors lying in the same chromosome should generally be found together. Such was the case for instance in the experiments with flies having red eyes and yellow body colour versus white eyes and grey body colour, the character for white eyes and yellow body being located in the X chromosome (see preceding chapter), or in the experiments on Abraxas. These phenomena are called linkage, and the numerical results of linkage were given in the preceding chapter in connection with the crossing of sex-linked characters.
We have already mentioned that before the maturation division occurs the homologous maternal and paternal chromosomes fuse—the so-called synapsis of the cytologists—and afterward separate again. It had been observed by Janssens that in this stage of fusion and subsequent separation a partial twisting and a partial exchange between two chromosomes may take place. Morgan assumes that this exchange accounts for certain deviations in the ratio of linkage. If in Fig. 40 the white and black signify two homologous chromosomes I and I1 containing the two pairs of homologous factors AB and ab respectively, the synapsis state would be as in Fig. 41. If the separation were complete, either I or its homologue I1 might be lost in the maturation division of the egg. If, however, the synapsis is slightly irregular, as in Fig. 42, where the chromosomes are slightly twisted, I and I1 will not separate completely but an exchange will take place, part of I1 and I becoming exchanged. This would result in the formation of two mixed chromosomes Ab and aB (Fig. 42). This partial exchange of homologous chromosomes, which Morgan calls “crossing over,” occurs, as he found in Drosophila, in the egg only, not in the maturation division of the sperm. He informs me that in the silkworm moth Tanaka found that it occurs only in the male, while in Primula it takes place both in the ovules and in the pollen as shown by Gregory.

Fig. 40 |
Fig. 41 |
Fig. 42 |
Morgan and his fellow-workers have put this theory to numerous tests by breeding experiments and the results have fully supported it. According to the chromosome theory linkage should occur only when factors lie in the same chromosome. Hence it should be possible, on the basis of this linkage theory, to foretell how many linkage groups there may occur in a species; namely, as many as there are chromosomes. In Drosophila there are four pairs of chromosomes, and Morgan and his fellow-workers found only four groups of linked characters.207 This agreement can be no mere accident.
Carrying the assumption still farther, these authors were able to show that each individual character has in all probability a definite location in the chromosome, so that it seems as if each individual chromosome consisted of a series of smaller chromosomes, each of which may be a factor in the determination of a hereditary character which is transmitted according to Mendel’s law of segregation. Biology has thus reached in the chromosome theory of Mendelian heredity an atomistic conception, according to which independent material determiners for hereditary characters exist in a linear arrangement in the chromosomes.
4. We are not concerned in this volume with the many applications of the theory of heredity to the breeding of plants, animals, and man; the reader will find a discussion of these topics in the numerous writings of the special workers on genetics.208 We are, however, interested in the bearing this work has on the conception of the organism. Two questions present themselves: Is the organism nothing but a mosaic of hereditary characters determined essentially by definite elements located in the chromosomes; and if this be true, what makes a harmonious whole organism out of this kaleidoscopic assortment? We call it a kaleidoscopic assortment since a glance at the list of hereditary characters found in one chromosome, according to Morgan, shows that there is apparently no physiological or chemical connection between them, and second: How can a factor contained in the chromosome determine a hereditary character of the organism? To the first question we venture to offer the answer which has been already suggested in various chapters of this book, that the cytoplasm of the egg is the future embryo in the rough; and that the factors of heredity in the sperm only act by impressing the details upon the rough block. This metaphor will receive a more definite meaning by the answer to the second question. The characters which follow Mendelian heredity are morphological features as well as instincts. For the former we have already had occasion to show in previous chapters to what extent they depend upon the internal secretions or the existence of specific compounds in the circulation, and the same is true for the instincts (Chapters VIII and X). This then leads us to the suggestion that these determiners contained in the chromosomes give rise each to the formation of one or more specific substances which influence various parts of the body. We probably do not notice all the effects in each case, but when a special organ is affected in a conspicuous way, we connect the factor with this organ or the special feature of the organ which is altered, and speak of a determiner or factor for that organ, or for one of its characters. We also understand in this way why outside conditions should be able to overcome the hereditary tendency in certain cases, for instance why the influence of certain hereditary factors for pigmentation should depend upon temperature as E. Baur observed.
The view, according to which the determiners in the chromosomes only tend to give special characters to the embryo or to the adult while the cytoplasm of the egg may be considered the real embryo, receives some support from the fact that the first development of the egg is purely maternal, even if the egg nucleus has been replaced by sperm of a different species. If an egg of a sea urchin be cut into two pieces, one with and one without a nucleus, and the enucleated piece be fertilized with the sperm of a different species of sea urchin, the blastula and gastrula stages are purely maternal and only the skeleton of the pluteus stage begins to betray the influence of the foreign sperm inasmuch as this skeleton is purely paternal, according to Boveri. In all experiments on hybridization it has been found that the rate of cell division of the egg is a purely maternal character. Thus when fish eggs of a species, in which the rate of first segmentation of the egg is about eight hours, are fertilized with sperm of a species for which the same process requires about thirty minutes or less at the same temperature, the rate of segmentation is again about eight hours. There is then no chromosome influence noticeable in the early development.
When two forms of sea urchins, Strongylocentrotus franciscanus and purpuratus,209 are crossed, certain features of the skeleton of the embryo, e. g., the so-called cross-bars, are a dominant, inasmuch as they are found in purpuratus and both the crosses, while they are absent in franciscanus. The development prior to the formation of the skeleton is purely maternal. These observations again lend support to the idea that the Mendelian factors of heredity must have the embryo to work on and that the organism is not to be considered a mere mosaic of Mendelian factors. This is further supported by the idea that the species specificity resides in the proteins of the unfertilized egg (see Chapter III), and it is quite likely that this species specificity decides which type of animal should arise from an egg.
The idea had been suggested that the factors which determine the future character might be ferments or enzymes, or substances from which such ferments develop. A. R. Moore210 pointed out that the cross-bars in the skeleton of the hybrid between S. purpuratus and franciscanus develop more slowly than in the pure breed and that this should be expected if the determiners were enzymes. Since the pure purpuratus has two determiners for the development of the cross-bars (from both egg and sperm), the hybrids only one (from either egg or sperm), the pure purpuratus should have twice the enzyme mass of the hybrid. It is known that the velocity of a chemical reaction increases in proportion with the mass (or in some cases in proportion with the square root of the mass) of the enzyme; the cross-bars should therefore develop faster in the pure than in the hybrid breeds, as was observed by Moore. It was, however, not possible to obtain quantitative data.
On the other hand, it is obvious that this reasoning would not hold for all cases. Thus when beans with violet flowers are crossed with white-flowered beans the hybrids are pale blue, which indicates that the hybrids have less pigment than the pure violet. Now we know that the mass of enzyme does not influence the chemical equilibrium but only the velocity of the reaction. The hybrids and pure violets differ, however, in the mass of violet pigment formed, that is to say, in regard to the equilibrium. Hence the idea that the determiners are enzymes or give rise to enzymes is probably not applicable to cases of this type.
The experiments on the heredity of pigments are at present almost the only ones which can be used for an analysis of the chemical nature of the character and its possible determiner. The important work of G. Bertrand211 and of Chodat212 on the production of black pigment in the cells of animals and plants with the aid of enzymes has paved the way for such work. Bertrand has shown that tyrosine (p-oxyphenylaminopropionic acid) is transformed into a black pigment by an enzyme tyrosinase which occurs in numerous organisms and is obviously the cause of pigment and colouration in a great number of species. This discovery was utilized in the study of the heredity of pigments by Miss Durham, Gortner,213 and very recently by Onslow.214 The latter showed that from the skins of certain coloured rabbits and mice a peroxidase can be extracted which behaves like a tyrosinase toward tyrosine in the presence of hydrogen peroxide. This peroxidase was found in the skins of black agouti, chocolate and blue rabbits, but not in yellow or orange rabbits. The recessive whiteness in rabbits and mice according to this author is due to the lack of the peroxydase. There exists a dominant whiteness in the English rabbit which is due to a tyrosinase inhibitor which destroys the activity of the tyrosinase “and the dominant white bellies of yellow and agouti rabbits are due to the same cause.” “Variations in coat colour are probably due to a quantitative rather than to a qualitative difference in the pigment present.”
One point might still be mentioned since it may help to overcome a difficulty in visualizing the connection between the localization of a factor in the chromosome and the production of a comparatively large quantity of a specific chemical compound, e. g., a chromogen or a tyrosinase. We must remember that all the cells of an organism have identical chromosomes, so that a factor for an enzyme like tyrosinase is contained in every cell throughout the whole body. It is likely, however, that the same factor (which we may conceive to be a definite chemical compound) will find a different chemical substrate to work on in the cells of different organs of the body, since the different organs differ in their chemical composition. Thus it is conceivable that in the production of tyrosinase or of tyrosine not a single chromomere of one single cell is engaged, but the sum total of all these individual chromomeres of all the cells in one or several organs of the body. The writer has added this remark especially in consideration of the fact that some authors seem to feel that the chromosome conception of heredity is incompatible with a physicochemical view of this process.
Since we have mentioned this difficulty which some writers seem to find in the chromosome theory of Mendelian heredity, it may be added that a single factor may suffice to determine a series of complicated reflexes. Thus the heliotropic reactions of animals are due to the presence of photosensitive substances, and it suffices for the hereditary transmission of the complicated purposeful reactions based on these tropisms that a factor for the formation of the photosensitive substance should exist.215
5. Another point should be emphasized, namely that for Mendelian heredity it is immaterial whether the character is introduced by the spermatozoön or by the egg. This fact which Mendel himself already recognized is in full harmony with the conclusion that the chromosomes and not the cytoplasm are the bearers of Mendelian heredity, since only in respect to the chromosome constitution are egg and sperm alike, while they differ enormously in regard to the mass of protoplasm they carry. We can, therefore, be tolerably sure that wherever we deal with a hereditary factor which is determined by the egg alone the cytoplasm of the latter is partly or exclusively responsible for the result.
We have already mentioned the fact that the rate of segmentation of the egg is such a character. Yet this character is as definite as any Mendelian character, and it would be as easy to discriminate two species of eggs by the time required from insemination to the beginning of cell division as it would be by any Mendelian character of their parents.
The application of our modern knowledge of heredity to human affairs has been discussed in a very original way by Bateson in his address before the British Association in Sydney to which the reader may be referred.216
1. The idea that the organism as a whole cannot be explained from a physicochemical viewpoint rests most strongly on the existence of animal instincts and will. Many of the instinctive actions are “purposeful,” i. e., assisting to preserve the individual and the race. This again suggests “design” and a designing “force,” which we do not find in the realm of physics. We must remember, however, that there was a time when the same “purposefulness” was believed to exist in the cosmos where everything seemed to turn literally and metaphorically around the earth, the abode of man. In the latter case, the anthropo- or geocentric view came to an end when it was shown that the motions of the planets were regulated by Newton’s law and that there was no room left for the activities of a guiding power. Likewise, in the realm of instincts when it can be shown that these instincts may be reduced to elementary physicochemical laws the assumption of design becomes superfluous.
If we look at the animal instincts purely as observers we might well get the impression that they cannot be explained in mechanistic terms. We need only consider what mysticism apparently surrounds all those instincts by which the two sexes are brought together and by which the entrance of the spermatozoön into the egg is secured; or the remarkable instincts which result in providing food and shelter for the young generation.
We have already had occasion to record some cases of instincts which suggest the possibility of physicochemical explanation; for example the curious experiment of Steinach on the reversal of the sexual instincts of the male whose testes had been exchanged for ovaries. There is little doubt that in this case the sexual activities of each sex are determined by specific substances formed in the interstitial tissue of the ovary and testes. The chemical isolation of the active substances and an investigation of their action upon the various parts of the body would seem to promise further progress along this line.
Marchal’s observations on the laying of eggs by the naturally sterile worker wasps are a similar case. The fact that such workers lay eggs when the queen is removed or when they are taken away from the larvæ may be considered as a manifestation of one of those wonderful instincts which form the delight of readers of Maeterlinck’s romances from insect life. Imagine the social foresight of the sterile workers who when the occasion demands it “raise” eggs to preserve the stock from extinction! And yet what really happens is that these workers, when there are no larvæ, can consume the food which would otherwise have been devoured by the larvæ; and some substance contained in this food induces the development of eggs in the otherwise dormant ovaries. What appeared at first sight as a mysterious social instinct is revealed as an effect comparable to that of thyroid substance upon the growth of the legs of tadpoles in Gudernatsch’s experiment (Chapter VII).
2. If we wish to show in an unmistakable way the mechanistic character of instincts we must be able to reduce them to laws which are also valid in physics. That instinct, or rather that group of instincts, for which this has been accomplished are the reactions of organisms to light. The reader is familiar with the tendency of many insects to fly into the flame. It can be shown that many species of animals, from the lowest forms up to the fishes, are at certain stages—very often the larval stage—of their existence, slaves of the light. When such animals, e. g., the larvæ of the barnacle or certain winged plant lice or the caterpillars of certain butterflies, are put into a trough or test-tube illuminated from one side only, they will rush to the side from which the light comes and will continue to do this whenever the orientation of the trough or test-tube to the light is changed; while they will be held at the window side of the vessel if the light or the position of the vessel remains unchanged. This instinct to get to the source of light is so strong that, e. g., the caterpillars of Porthesia chrysorrhœa die of starvation on the window side of the vessel, with plenty of food close behind. This powerful “instinct” is, as we intend to show, in the last analysis, the expression of the Bunsen-Roscoe law of photochemical reactions. A large number of chemical reactions are induced or accelerated by light, and the Bunsen-Roscoe law shows that the chemical effect is in these cases, within certain limits, equal to the product of the intensity into the duration of illumination.
The “attraction” or “repulsion” of animals by the light had been explained by the biologists in an anthropomorphic way by ascribing to the animals a “fondness” for light or for darkness. Thus Graber, who had made the most extensive experiments, gave as a result the statement that animals which are fond of light are also fond of blue while they hate the red, and those which are fond of the “dark” are fond of red and hate the blue.218 In 1888 the writer published a paper in which he pointed out that the so-called fondness of animals for light and blue and for dark and red was simply a case of an automatic orientation of animals by the light comparable to the turning of the tips of a plant towards the window of the room in which the plant is raised.219
The phenomenon of a plant bending or growing to the source of light is called positive heliotropism (while we speak of negative heliotropism in all cases in which the plant turns away from the light, as is observed in many roots). The writer pointed out that animals which go to the light are positively heliotropic (or phototropic) and do so because they are compelled automatically by the light to move in this direction, while he called those animals which move away from the light negatively heliotropic; they are automatically compelled by the light to move away from it. What the light does is to direct the motions of the animals and to explain this the following theory was proposed. Animals possess photosensitive elements on the surface of their bodies, in the eyes, or occasionally also in epithelial cells of their skin. These photosensitive elements are arranged symmetrically in the body and through nerves are connected with symmetrical groups of muscles. The light causes chemical changes in the eyes (or the photosensitive elements of the skin). The mass of photochemical reaction products formed in the retina (or its homologues) influences the central nervous system and through this the tension or energy production of the muscles. If the rate of photochemical reaction is equal in both eyes this effect on the symmetrical muscles is equal, and the muscles of both sides of the body work with equal energy; as a consequence the animal will not be deviated from the direction in which it was moving. This happens when the axis or plane of symmetry of the animal goes through the source of light, provided only one source of light be present. If, however, the light falls sidewise upon the animal, the rate of photochemical reaction will be unequal in both eyes and the rate at which the symmetrical muscles of both sides of the body work will no longer be equal; as a consequence the direction in which the animal moves will change. This change will take place in one of two ways, according as the animal is either positively or negatively heliotropic; in the positively heliotropic animal the resulting motion will be toward, in the negatively heliotropic from, the light. Where we have no central nervous system, as in plants or lower animals, the tension of the contractile or turgid organs is influenced in a different way, which we need not discuss here.
The reader will perceive that according to the writer’s theory two agencies are to be considered in these reactions: first, the symmetrical arrangement of the photosensitive and the contractile organs, and second, the relative masses of the photochemical reaction products produced in both retinæ or photosensitive organs at the same time. If a positively heliotropic animal is struck by light from one side, the effect on tension or energy production of muscles connected with this eye will be such that an automatic turning of the head and the whole animal towards the source of light takes place; as soon as both eyes are illuminated equally the photochemical reaction velocity will be the same in both eyes, the symmetrical muscles of the body will work equally, and the animal will continue to move in this direction. In the case of the negatively heliotropic animal the picture is the same except that if only one eye is illuminated the muscles connected with this eye will work less energetically. The theory can be nicely tested for negatively heliotropic animals in the larvæ of the blowfly when they are fully grown, and for positively heliotropic animals on the larvæ of Balanus, and many other organisms.
Fig. 43 |

Fig. 44 |
One of the difficulties in identifying the motions of animals to or from the light with the positive and negative heliotropism of plants consisted in the fact that plants are mostly sessile (and respond to a one-sided illumination with heliotropic curvatures to or from the light), while most animals are free moving and respond to the one-sided illumination by being turned and compelled to move to or from the light. This difficulty was overcome by the observation that sessile animals like the hydroid Eudendrium (Fig. 43) or the tube worm Spirographis (Fig. 44) react to a one-sided illumination also with heliotropic curvatures like sessile plants.220 On the other hand, it had been found before by Strassburger that free-swimming plant organisms like the swarmspores of algæ move to or from the source of light as do free-swimming animals.
3. The writer suggested in 1897221 that the light acts chemically in the heliotropic reactions and in 1912 that the heliotropic reactions probably follow the law of Bunsen and Roscoe,222 and it was possible to confirm this idea by direct experiments.223 This law states that the photochemical effect of light equals i t where i is the intensity of the light and t the duration of illumination. The experiments were carried out on young regenerating polyps of Eudendrium by measuring the time required to cause fifty per cent. of the polyps to bend to the source of light. The intensity of light was varied by altering the distance of the source of light from the polyps. Table VI gives the result.
TABLE VI
| Distance between Polyps and Source of Light | Time Required to Cause Fifty Per Cent. of the Polyps to Bend towards the Source of Light | |
|---|---|---|
| Observed | Calculated from Bunsen-Roscoe Law | |
| Metres | Minutes | Minutes |
| 0.25 | 110 | |
| 0.50 | between 35 and 40 | 140 |
| 1.00 | 150 | 160 |
| 1.50 | between 360 and 420 | 360 |
We must therefore conclude that the heliotropic curvature of the polyps is determined by a photochemical action of the light. The light brings about or accelerates a chemical reaction which follows the Bunsen-Roscoe law. As soon as the product of this reaction on one side of the polyp exceeds that on the other by a certain quantity, the bending occurs. When the product i t is the same for symmetrical spots of the organism no bending can result. This is what our theory suggested.
It is very difficult to prove directly the applicability of the Bunsen-Roscoe law for free-moving animals, but it can be shown that intermittent light is as effective as constant light of the same intensity, provided that the total duration of the illumination by the intermittent light is equal to that of the constant light, and the duration of the intermission is sufficiently small (Talbot’s law). Talbot’s law is in reality only a modification of the Bunsen-Roscoe law. Ewald has proved in a very elegant way the applicability of Talbot’s law to the orientation of the eyestalk of Daphnia.224 This makes it probable that the law of Bunsen-Roscoe underlies generally the heliotropic reaction of animals.
It is of importance for the theory of the identity of the heliotropism of animals and plants that in the latter organisms the law of Bunsen and Roscoe is also applicable. This had been shown previously by Fröschel225 and by Blaauw.226 In the following table are given the results of Blaauw’s experiments on the applicability of the Bunsen-Roscoe law for the heliotropic curvature of the seedlings of oats (Avena sativa). The time required to cause heliotropic curvatures for intensities of light varying from 0.00017 to 26520 metre-candles was measured. The product i t, namely metre-candles-seconds, varies very little (between 16 and 26).
TABLE VII
| I Duration of Illumination | II Metre-Candles | III Metre-Candles-Seconds | I Duration of Illumination | II Metre-Candles | III Metre-Candles-Seconds | |||
|---|---|---|---|---|---|---|---|---|
| 43 | hours | 0.00017 | 26.3 | 25 | seconds | 1 | .0998 | 27.5 |
| 13 | " | 0.000439 | 20.6 | 8 | " | 3 | .02813 | 24.2 |
| 10 | " | 0.000609 | 21.9 | 4 | " | 5 | .456 | 21.8 |
| 6 | " | 0.000855 | 18.6 | 2 | " | 8 | .453 | 16.9 |
| 3 | " | 0.001769 | 19.1 | 1 | " | 18 | .94 | 18.9 |
| 100 | minutes | 0.002706 | 16.2 | 2⁄5 | " | 45 | .05 | 18.0 |
| 60 | " | 0.004773 | 17.2 | 2⁄25 | " | 308 | .7 | 24.7 |
| 30 | " | 0.01018 | 18.3 | 1⁄25 | " | 511 | .4 | 20.5 |
| 20 | " | 0.01640 | 19.7 | 1⁄55 | " | 1255 | 22.8 | |
| 15 | " | 0.0249 | 22.4 | 1⁄100 | " | 1902 | 19.0 | |
| 8 | " | 0.0498 | 23.9 | 1⁄400 | " | 7905 | 19.8 | |
| 4 | " | 0.0898 | 21.6 | 1⁄800 | " | 13094 | 16.4 | |
| 40 | seconds | 0.6156 | 24.8 | 1⁄1000 | " | 26520 | 26.5 | |
It is, therefore, obvious that the blind instinct which forces animals to go to the light, e. g., in the case of the moth, is identical with the instinct which makes a plant bend to the light and is a special case of the same law of Bunsen and Roscoe which also explains the photochemical effects in inanimate nature; or in other words, the will or tendency of an animal to move towards the light can be expressed in terms of the Bunsen-Roscoe law of photochemical reactions.
The writer had shown in his early publications on light effects that aside from the heliotropic reaction of animals, which as we now know depends upon the product of the intensity and duration of illumination, there is a second reaction which depends upon the sudden changes in the intensity of illumination. These latter therefore obey a law of the form: Effect = f (didt).227 Jennings has maintained that the heliotropic reactions of unicellular organisms are all of this kind, but investigations by Torrey and by Bancroft228 on Euglena have shown that Jennings’s statements were based on incomplete observations.
4. In these experiments only one source of light was applied. “When two sources of light of equal intensity and distance act simultaneously upon a heliotropic animal, the latter puts its median plane at right angles to the line connecting the two sources of light.”229 This fact has been amply verified by Bohn, by Parker and his pupils, and especially by Bradley Patten, who used it to compare the relative efficiency of two different lights.
The behaviour of the animals under the influence of two lights is a confirmation of our theory of heliotropism inasmuch as the animal moves in such a direction that the symmetrical elements of the surface of the body are struck by light of the same intensity at the same angle, so that as a consequence equal masses of photosensitive substances are produced in symmetrical elements of their eyes or skin in equal times. The effect on the symmetrical muscles will be identical. As soon as one of the lights is a little stronger the animal will deviate towards this light, in case it is positively heliotropic and towards the weaker light if it is negatively heliotropic. This deviation again is not the product of chance but follows a definite law as Patten230 has recently shown. He used the negatively heliotropic larvæ of the blowfly. These larvæ were made to record their trail while moving under the influence of the two lights. The results of the measurements of 2500 trails showing the progressive increase in angular deviation of the larvæ (from the perpendicular upon the line connecting the two lights), with increasing differences between the lights, are given in the following table. Since the deviation or angular deflection of the larvæ is towards the weaker of the two lights it is marked negative.
TABLE VIII
| Percentage Difference in the Intensity of the Two Lights | Average Angular Deflection of the Two Paths of the Larvæ towards the Weaker Light | |
|---|---|---|
| Per Cent. | Degrees | |
| 0 | 0-0.09 | |
| 8 | 1⁄3 | 0-2.77 |
| 16 | 2⁄3 | 0-5.75 |
| 25 | 0-8.86 | |
| 33 | 1⁄3 | -11.92 |
| 50 | -20.28 | |
| 66 | 2⁄3 | -30.90 |
| 83 | 1⁄3 | -46.81 |
| 100 | -77.56 | |
Let us assume that the negatively heliotropic animal is at an equal distance from the two unequal lights and placed so that at the beginning of the experiment its median plane is at right angles to the line connecting the two lights, but with its head turned away from them. In that case the velocity of reaction in the symmetrical photosensitive elements of the eyeless larvæ is greater on the side of the stronger light. Since the animal is negatively heliotropic this will result in a greater relaxation or a diminution of the energy production of the muscles turning the head of the animal towards the side of the stronger light. Hence the animal will automatically deviate from the straight line towards the side of the weaker light. By the alteration of the position of its body the photosensitive elements exposed to the stronger of the two lights will be put at a less efficient angle and hence the rate of photochemical reaction on this side will be diminished. The deviation from the perpendicular in which the animal will ultimately move will be such that as a consequence, the rate of photochemical reaction in symmetrical elements is again equal. The ultimate direction of motion will, according to our theory always be such that the mass of chemical products formed under the influence of light in symmetrical photosensitive elements during the same time is equal.
Patten also investigated the question whether the same difference of percentage between two lights would give the same deviation, regardless of the absolute intensities of the lights used. The absolute intensity was varied by using in turn from one to five glowers. The relative intensity between the two lights varied in succession by 0, 81⁄3, 162⁄3, 25, 331⁄3, and 50 per cent. Yet the angular deflections were within the limits of error identical for each relative difference of intensity of the two lights no matter whether, 1, 2, 3, 4, or 5 glowers were used. The following table shows the result.
TABLE IX
A Table Based on the Measurements of 2700 Trails Showing the Angular Deflections at Five Different Absolute Intensities
| Number of Glowers | Difference of Intensity between the Two Lights | |||||
|---|---|---|---|---|---|---|
| 0 per cent. | 81⁄3 per cent. | 162⁄3 per cent. | 25 per cent. | 331⁄3 per cent. | 50 per cent. | |
| Deflection in Degrees | ||||||
| 1 | -0.550 | -2.32 | -5.270 | -9.04 | -11.86 | -19.46 |
| 2 | -0.100 | -3.05 | -6.120 | -8.55 | -11.92 | -22.28 |
| 3 | +0.450 | -2.60 | -5.650 | -8.73 | -13.15 | -20.52 |
| 4 | -0.025 | -2.98 | -6.600 | -9.66 | -11.76 | -19.88 |
| 5 | -0.225 | -2.92 | -5.125 | -8.30 | -10.92 | -19.28 |
| Average | -0.090 | -2.77 | -5.750 | -8.86 | -11.92 | -20.28 |
Such constancy of quantitative results is only possible where we are dealing with purely physicochemical phenomena or where life phenomena are unequivocally determined by purely physicochemical conditions.
5. It seems difficult for some biologists, even with the validity of the Bunsen-Roscoe law proven, to imagine that the movements of the animals under the influence of light are not voluntary (or not dictated by the mysterious “trial and error” method of Jennings).231 But one wonders how it is possible on such an assumption to account for the fact that the angle of deflection of the larva of the fly when under the influence of two lights of different intensities should be always the same for a given difference in intensity; or why the time for curvature in Eudendrium should vary inversely with the intensity of illumination. It is, however, possible to complete the case for the purely physicochemical analysis of these instincts. John Hays Hammond, Jr., has succeeded in constructing heliotropic machines which in the dark follow a lantern very much in the manner of a positively heliotropic animal. The eyes of this heliotropic machine consist of two lenses in whose focus is situated the “retina” consisting of selenium wire. The two eyes are separated from each other by a projecting piece of wood which represents the nose and allows one eye to receive light while the other is shaded. The galvanic resistance of selenium is altered by light; and when one selenium wire is shaded while the other is illuminated, the electric energy (supplied by batteries inside the machine) which makes the wheels turn (these take the place of the legs of the normal animal) no longer flows symmetrically to the steering wheel, and the machine turns towards the light. In this way the machine follows a lantern in a dark room in a way similar to that of a positively heliotropic animal. Here we have a model of the heliotropic animal whose purely mechanistic character is beyond suspicion, and we may be sure that it is not “fondness” for light or for brightness nor will-power nor a method of “trial and error” which makes the machine follow the light.
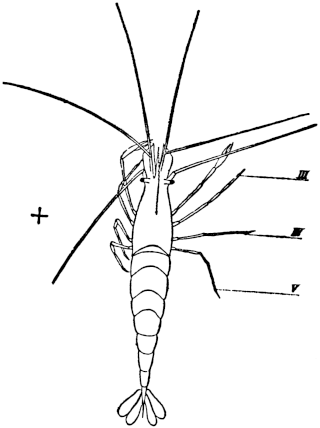
Fig. 45
6. It may also be of interest to know that in heliotropism the motions of the legs are automatically controlled by the chemical changes taking place in symmetrical elements of the retina. In order to prove this point we will turn to the phenomenon of galvanotropism. The galvanic current forces certain animals to move in the direction of one of the two electrodes just as the light forces the heliotropic animals to move towards (or from) the source of light. The change in the concentration of the ions at the boundary of the various organs, especially the nerves, determines the galvanotropic reactions. When the shrimp Palæmonetes is put into a trough with dilute salt solution through which a current of a certain intensity flows, the animal is compelled to move towards the anode.232 It can walk forwards, backwards, or sidewise. Here we can observe directly that the effect of the current consists in altering the tension of the muscles of the legs in such a way as to make it easy for the animal to move toward the anode and difficult to move toward the cathode. Thus if the current be sent sidewise through the animal, say from left to right (Fig. 45), the legs of the left side assume the flexor position, those of the right the extensor position. With this position of its legs the animal can easily move to the left, i. e., the anode, and only with difficulty to the right, i. e., the cathode. This change in the position of the legs occurs when the animal is not moving at all, thus showing that the galvanotropic movements take place not because the animal intends to go to the anode, but that the animal goes to the anode because its legs are practically prevented by the galvanic current from working in any other way. This is exactly what happens in the heliotropic motions of animals.233
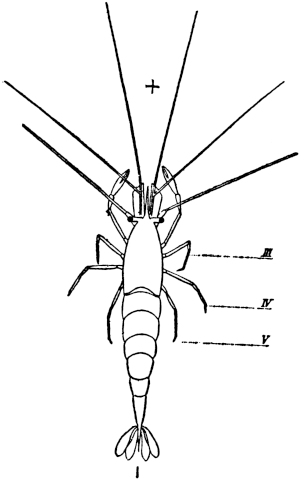
Fig. 46
To understand what happens when the current goes lengthwise through the body it should be stated that Palæmonetes uses the third, fourth, and fifth pairs of legs for its locomotion. The third pair pulls in the forward movement, and the fifth pair pushes. The fourth pair generally acts like the fifth, and requires no further attention. If a current be sent through the animal longitudinally, from tail to head, and the strength be increased gradually, a change soon takes place in the position of the legs (Fig. 46). In the third pair the tension of the flexors predominates, in the fifth the tension of the extensors. The animal can thus move easily with the pulling of the third and the pushing of the fifth pairs of legs, that is to say, the current changes the tension of the muscles in such a way that the forward motion is rendered easy, the backward motion is difficult. Hence it can easily move toward the anode, but only with difficulty toward the cathode. If a current be sent through the animal in the opposite direction, namely, from head to tail, the third pair of legs is extended, the fifth pair bent; that is, the third pair can push, and the fifth pair pull. The animal will thus move backward easily and forward with difficulty, and it is thus driven to the anode again.
The explanation which Loeb and Maxwell proposed for this influence of the current on the legs assumes that there are three groups of ganglion cells in the central nervous system of these animals which are oriented according to the three main axes of the body; (1) right-left and left-right, (2) backward, and (3) forward. It depends upon whether the ganglion cells or the nerve elements are in anelectrotonus, which muscles are bent and which relaxed. It would lead us too far to recapitulate the theory in this place, and the reader who is interested in it is referred to Loeb and Maxwell’s paper.234 The importance of the observations lies in the fact that they show that any element of will or choice on the part of the animal in these motions is eliminated, that the animal moves where its legs carry it, and not that the legs carry the animal where the latter “wishes” to go.
7. This may be the place to dispel an error which has sometimes crept into the discussion of the tropistic reactions of animals. It has been stated occasionally that it is the energy gradient and not the automatic orientation of the animal by the light which makes the positively heliotropic animal move towards the source of light and the negatively heliotropic away from it. Thus the positively heliotropic animal would be compelled to move towards the source of light as a consequence of the fact that the intensity of the light increases the more the nearer the animal approaches the source of light. If the source of light be the reflected sky-light the difference of intensity at both ends of a microscopic organism is so slight that it is beneath the limit capable of influencing the motions.
Fig. 47
A simple experiment published by the writer in 1889 suffices to dispel the idea that the energy gradient determines the direction of the motion of an animal in tropistic reactions. Let direct sunlight (S, Fig. 47) fall through the upper half of a window (w w) upon a table, and diffused daylight (D) through the lower half of the window on the same table. A test-tube a c is placed on the table in such a way that its long axis is at right angles to the plane of the window; and one half a b is in the direct sunlight, the other half in the shade. If at the beginning of the experiment the positively heliotropic animals are in the direct sunlight at a, they promptly move toward the window, gathering at the window end c of the tube, although by so doing they go from the sunshine into the shade.235 This experiment is in harmony with our idea that the effect of light consists in turning the head of the animal and subsequently its whole body toward the source of light. By going from the strong light into the shade the reaction velocity in both eyes is diminished equally and hence there is no reason for the animal to change its orientation, though its progressive motion may be stopped for an instant by the change. But at the boundary between sunlight and daylight a sudden change from strong to weak light occurs. If the energy gradient determined the direction of the positively heliotropic animal, the motion should stop at the boundary from strong to weak light, which may happen for an instant but which will not interfere with the progressive motion of the animal.
8. Graber had found that when animals are put into a trough covered half with blue and half with red glass, those that are “fond” of light go under the blue, those that are “fond” of darkness go under the red glass. The writer pointed out that this result should be expected on the basis of his theory of heliotropism, if the assumption be correct that the red light is considerably less efficient than light which goes through blue glass (such glass also allows green rays to go through). The botanists had already shown that red glass is impermeable for the rays which cause heliotropic reactions of plants, and the writer was able to show the same for the heliotropic reactions of animals. Red glass acts, therefore, almost like an opaque body for these animals.
A closer examination of the most efficient rays for the heliotropic reactions of different organisms has revealed the fact that for some organisms a region in the blue λ = 460–490 µµ, for others a region in the yellowish-green, near about λ = 520–530 µµ is the most efficient.236 For many plants and for some animals, like Eudendrium and the larvæ of the worm Arenicola, a region in the blue is most efficient; for certain, if not most, animals a region in the yellow-green is most efficient. Among unicellular green algæ, Chlamydomonas, has its maximal efficiency in the yellowish-green and Euglena in the blue. According to observations by Mast, some green unicellular organisms like Pandorina, Eudorina, and Spondylomorum seem to behave more like Chlamydomonas, while certain others behave more like Euglena.237 Wasteneys and the writer suggested that there are two groups of heliotropic substances, one with a maximum of photosensitiveness in the blue, the other in the yellowish-green; and that the latter group may or may not be related or identical with the visual purple which is most rapidly bleached by light of a wave length near λ = 520–530 µµ.
The ophthalmologist Hess238 has utilized the heliotropic reactions of animals in an attempt to prove that all animals from the lowest invertebrates up to the fishes inclusive suffer from total colour-blindness. This statement was based on the observation that for most positively heliotropic animals the region in the yellowish-green near λ = 520 µµ seems the most efficient. Since this region of the spectrum appears also as the brightest to a totally colour-blind man, he concluded that all these animals are totally colour-blind. There is no reason why heliotropic reactions should be used as an indicator for colour sensations; if totally colour-blind human beings were possessed of an irresistible impulse to run into a flame Hess’s assumption might be considered, but no such phenomenon exists in colour-blind man. Moreover, v. Frisch239 has shown by experiments on the influence of the background on the colouration of fish as well as by experiments on bees and on Daphnia that the reactions of these animals to light of different wave-lengths indicate different effects besides those of mere intensity. Thus v. Frisch could train bees to go to a blue piece of cardboard distributed among many cardboards of different shades of grey. Bees thus trained would alight on any blue object even if it contained no food. It would be impossible to do this with totally colour-blind organisms.
9. Heliotropic reactions play a great rôle in the preservation of individuals as well as of species. In order to understand this rôle it must be stated that the photosensitive substances appear often only under certain conditions and that their effect is inhibited under other conditions. Thus among ants the winged males and females alone show positive heliotropism,240 while the wingless workers are free from this reaction. This positive heliotropism becomes violent at the time of the nuptial flight and this phenomenon itself seems to be a heliotropic phenomenon since it takes place in the direction of the light. When the queen founds her nest she loses her wings and becomes negatively heliotropic again. Kellogg241 has shown that the nuptial flight of the bees is also a purely heliotropic phenomenon. When a part of the hive remote from the entrance is illuminated the bees rush to the light and can thus be prevented from swarming. These phenomena suggest that the presence of some substance secreted by the sex glands may cause the intensification of the heliotropism which leads to the nuptial flight.
In certain species of Daphnia, fresh-water copepods, and of Volvox, a trace of CO2 suffices to make negatively heliotropic or indifferent specimens violently positively heliotropic.242 Certain forms of marine copepods and the larvæ of Polygordius can be made positively heliotropic by lowering the temperature243 and the larvæ of the barnacle can be made negatively heliotropic by strong light.244 It is quite possible that a change in the sense of heliotropism by temperature and light is to some extent at least responsible for the periodic depth migrations of heliotropic animals. Many if not all positively heliotropic animals can be made negatively heliotropic by exposure to ultraviolet light.245
A most interesting example of the rôle of heliotropism in the preservation of a species is shown in the caterpillars of Porthesia chrysorrhœa. The butterfly lays its eggs upon a shrub. The larvæ hatch late in the fall and hibernate in a nest on the shrub, as a rule not far from the ground. As soon as the temperature reaches a certain height, they leave the nest; under natural conditions, this happens in the spring when the first leaves have begun to form on the shrub. (The larvæ can, however, be induced to leave the nest at any time in the winter provided the temperature is raised sufficiently.) After leaving the nest, they crawl directly upward on the shrub where they find the leaves on which they feed. Should the caterpillars move down the shrub, they would starve, but this they never do, always crawling upward to where they find their food. What gives the caterpillar this never-failing certainty which saves its life, and for which a human being might envy the little larva? Is it a dim recollection of experiences of former generations? It can be shown that it is the light reflected from the sky which guides the animal upward. When we put these animals into a horizontal test-tube in a room, they all crawl toward the window, or toward a lamp; the animal is positively heliotropic. It is this positive heliotropism which makes them move upward where they find their food, when the mild air of the spring calls them forth from their nest. At the top of the branch, they come in contact with a leaf, and chemical or tactile influences set the mandibles of the young caterpillar into activity. If we put these larvæ into closed test-tubes which lie with their longitudinal axes at right angles to the window, they will all migrate to the window end, where they stay and starve, even if their favourite leaves are close behind them. They are slaves of the light.
The few young leaves on top of a twig are quickly eaten by the caterpillar. The light, which saved its life by making it creep upward where it finds food, would cause it to starve could it not free itself from the bondage of positive heliotropism. The animal, after having eaten, is no longer a slave of the light, but can and does creep downward. It can be shown that a caterpillar, after having been fed, loses its positive heliotropism almost completely and permanently. If we submit unfed and fed caterpillars of the same nest contained in two different test-tubes to the same artificial or natural source of light, the unfed will creep to the light and stay there until they die, while those that have eaten will pay little or no attention to the light. Their sensitiveness to light has disappeared; after having eaten they become independent of light and can creep in any direction. The restlessness which accompanies the condition of hunger makes the animal creep downward—which is the only direction open to it—where it finds new young leaves on which it can feed. The wonderful hereditary instinct, upon which the life of the animal depends, is its positive heliotropism in the unfed condition and its loss of this heliotropism after having eaten. The latter phenomenon is in harmony with the experiments which show that the heliotropism of certain species of Daphnia disappears when the water becomes neutral.
And finally it may be pointed out that the majority of green plants could not exist if their stems were not positively, their roots negatively, heliotropic. It is the positive heliotropism which makes the top grow toward the light, which enables the leaves to get the light necessary for assimilation, and the roots to grow into the soil where they find the water and nutritive salts.
10. While we do not wish to deal here with the different tropisms it should be stated that aside from heliotropism, chemotropism as well as stereotropism play the most essential rôle in the so-called instinctive actions of animals. It is a problem of orientation by the diffusion of molecules from a centre when a male butterfly is deviated from its flight and alights on the wooden box in which is enclosed a female of the same species. We have already alluded to certain phenomena of chemotropism in Chapter IV. Certain organisms have a tendency to bring their bodies as much as possible on all sides in contact with solid bodies; thus the butterfly Amphipyra, which is a fast runner, will come to rest under a glass plate when the plate is put high enough above the ground so that it touches the back of the butterfly. The animals which live under stones or underground or in caves are as a rule both negatively heliotropic and positively stereotropic. Their tropisms predestine or force them into the life they lead.
The sensitive area which forms the basis of tropisms is as a rule developed not in the whole organism but only in certain segments of the body. Thus the eyes are located in the head. But when the action of one segment becomes overpowering the whole organism follows the segment. It has been customary among physiologists to speak of reflexes in such cases. Thus, e. g., the arms of the male frog develop a powerful positive stereotropism on their ventral surface during the spawning season. It would avoid confusion to realize that there is nothing gained in applying to this tropism the meaningless term “reflex”; it is better to call them tropisms since the organism as a whole is involved. If necessary we might speak of segmental tropisms. The act of seeking the female as well as that of cohabitation are in many cases combinations of chemotropism and stereotropism. The development of these tropisms depends upon the presence of certain specific substances in the body, a fact emphasized already in the case of heliotropism. In case of the development of the segmental stereotropism of the male frog at the time of spawning it has been shown that it depends on an internal secretion from the testes.
It has been suggested by some authors that the tropistic reactions are determined by some feeling or emotion on the part of the organism. We have no means of judging the emotions of lower animals (except by “intuition”). The writer suggested in 1899 in his book on brain physiology that emotions may be determined by specific substances which also determine the tropistic reaction (as well as phenomena of organ formation, although this latter phenomenon has nothing to do with the subject of instincts); and the excellent work of Cannon246 has shown the rôle of adrenalin in the expression of fear. It is, therefore, both unwarranted and unnecessary to state that hypothetical emotions determine the tropistic reactions.
1. The term environment in relation to an organism may easily assume a mystic rôle if we assume that it can modify the organisms so that they become adapted to its peculiarities. Such ideas are difficult to comprehend from a physicochemical viewpoint, according to which environment cannot affect the living organism and non-living matter in essentially different ways. Of course we know that proteins will as a rule coagulate at temperatures far below the boiling point of water and that no life is conceivable for any length of time at temperatures above 100° C., but heat coagulation of proteins occurs as well in the test-tube as in the living organism. If we substitute for the indefinite term environment the individual physical and chemical forces which constitute environment it is possible to show that the influence of each of these forces upon the organism finds its expression in simple physicochemical laws and that there is no need to introduce any other considerations.
We select for our discussion first the most influential of external conditions, namely temperature. The reader knows that there is a lower as well as an upper temperature limit for life. Setchell has ascertained that in hot springs whose temperature is 43° C., or above, no animals or green alga are found.247 In hot springs whose temperature is above 43° he found only the Cyanophyceæ, whose structure is more closely related to that of the bacteria than to that of the algæ, inasmuch as they have neither definitely differentiated nuclei nor chromophores. The highest temperature at which Cyanophyceæ occurred was 63° C. Not all the Cyanophyceæ were able to stand temperatures above 43° C., but only a few species. The other Cyanophyceæ were found at a temperature below 40° C., and were no more able to stand higher temperatures than the real algæ or animals. The Cyanophyceæ of the hot springs were as a rule killed by a temperature of 73°. From this we must conclude that they contain proteins whose coagulation temperature lies above that of animals and green plants, and may be as high as 73°. Among the fungi many forms can resist a temperature above 43° or 45°; the spores can generally stand a higher temperature than the vegetative organs. Duclaux found that certain bacilli (Tyrothrix) found in cheese are killed in one minute at a temperature of from 80° to 90°; while for the spores of the same bacillus a temperature of from 105° to 120° was required.248
Duclaux has called attention to a fact which is of importance for the investigation of the upper temperature limit for the life of organisms. According to this author it is erroneous to speak of a definite temperature as a fatal one; instead we must speak of a deadly temperature zone. This is due to the fact that the length of time which an organism is exposed to a higher temperature is of importance. Duclaux quotes as an example a series of experiments by Christen on the spores of soil and hay bacilli. The spores were exposed to a stream of steam and the time determined which was required at the various temperatures to kill the spores.
| It took at 100° | over sixteen hours |
| " " " 105–110° | two to four hours |
| " " " 115° | thirty to sixty minutes |
| " " " 125–130° | five minutes or more |
| " " " 135° | one to five minutes |
| " " " 140° | one minute |
In warm-blooded animals 45° is generally considered a temperature at which death occurs in a few minutes; but a temperature of 44°, 43°, or 42° is also to be considered fatal with this difference only, that it takes a longer time to bring about death. This fact is to be considered in the treatment of fever.
It is generally held that death in these cases is due to an irreversible heat coagulation of proteins. According to Duclaux, it can be directly observed in micro-organisms that in the fatal temperature zone the normally homogeneous, or finely granulated, protoplasm is filled with thick, irregularly arranged bodies, and this is the optical expression of coagulation. The fact that the upper temperature limit differs so widely in different forms is explained by Duclaux through differences in the coagulation temperature of the various proteins. It is, e. g. known that the coagulation temperature varies with the amount of water of the colloid. According to Cramer, the mycelium of Penicillium contains 87.6 water to 12.4 dry matter, while the spores have 38.9 water and 61.1 dry substance. This may explain why the mycelium is killed at a lower temperature than the spores. According to Chevreul, with an increase in the amount of water, the coagulation temperature of albuminoids decreases. The reaction of the protoplasm influences the temperature of coagulation, inasmuch as it is lower when the reaction is acid, higher when the reaction is alkaline. The experiments of Pauli show also a marked influence of salts upon the temperature of coagulation of colloids.
The process of heat coagulation of colloids is also a function of time. If the exposure to high temperature is not sufficiently long, only part of the colloid coagulates; in this case an organism may again recover.
Inside of these upper and lower temperature limits we find that life phenomena are influenced by temperature in such a way that their rate is about doubled for an increase of the temperature of 10° C., and that this temperature coefficient for 10°, Q10, very often steadily diminishes from the lower to the higher temperature; so that near the lower temperature limit it becomes often considerably greater than 2 and near the higher temperature limit it becomes very often less than 2.249 This influence of temperature is so general that we are bound to associate it with an equally general feature of life phenomena; and such a feature would be most likely the chemical reactions. It is known through the work of Berthelot, van’t Hoff, and Arrhenius that the temperature coefficient for the velocity of chemical reactions is also generally of about the same order of magnitude; namely ≧2 for a difference of 10°. In chemical reactions there is also a tendency for Q10 to become larger for lower temperature, and coefficients of Q10 about 5 or 6 have repeatedly been found for purely chemical reactions between 0° and 10°, e. g., for the inversion of cane sugar by the hydrogen ion. The temperature coefficient for the reaction velocity of ferments shows the same diminution of Q10 with rising temperature which is also noticed in most life phenomena. Thus Van Slyke and Cullen250 found that the reaction rate of the enzyme urease “is nearly doubled by every 10° rise in temperature between 10° and 50°. Within this range the temperature coefficient is nearly constant and averages 1.91. From O° to 10° it is 2.80, from 50° to 60° it is only 1.09. The optimum is at about 55°.” The rapid fall of the temperature coefficient for enzyme action at the upper temperature limit has been ascribed by Tammann to a progressive destruction of the active mass of enzyme by the higher temperature (by hydrolysis). This will, however, not account for the high value of the coefficient near the lower limit. But is it not imaginable that at low temperature an aggregation of the enzyme particles exists which is also equivalent to a diminution of the active mass of the enzyme and that this aggregation is gradually dispersed by the rising temperature? This would account for the fact that at a temperature near 0°C life phenomena stop because the enzymes are all in a state of aggregation or gelation; that then more and more are dissolved and the rate of chemical reaction increases since the mass of enzyme particles increases until all the enzyme molecules are dissolved or rendered active. Under this assumption three processes are superposed in the variation of the value of Q10 with temperature: (1) the supposed increase in the number of available ferment molecules with increasing temperature near the lower temperature limit; (2) the temperature coefficient of the reaction velocity which is nearly = 2 for 10°C.; (3) the diminution of the number of available ferment molecules by hydrolysis or some other action of the increasing temperature. This latter is noticeable near the upper temperature limit. The reason that 1 and 3 interfere more strongly in life phenomena than in the chemical reactions of crystalloid substances may possibly be accounted for by the fact that the enzymes and most of the constituents of living matter are colloidal, i. e., consist of particles of a considerably greater order of magnitude than the molecules of crystalloids.251
We will now show the rôle of the temperature coefficient upon phenomena of development. F. R. Lillie and Knowlton252 first determined the influence of temperature upon the development of the egg of the frog and showed that it was of the same nature as that of a chemical reaction. These experiments were repeated a year later by O. Hertwig.253
The time required for the eggs to reach definite stages was measured for different temperatures and it was found that the temperature coefficient Q10 between 2.5° and 6° was equal to 10 or more; between 6° and 15° it was between 2.6 and 4.5; between 10° and 20° it was 2.9 to 3.3, and between 20° and 24° it was between 1.4 and 2.0. To anybody who has worked on this problem it is obvious that no exact figures can be obtained in this way, since the point when a certain stage of development is reached is not so sharply defined as to exclude a certain latitude of arbitrariness. The writer found that very exact figures can be obtained on the influence of temperature upon development of the sea-urchin egg by measuring the time from insemination to the first cell division. Such experiments were carried out in a cold-water form Strongylocentrotus purpuratus and a form living in warmer water, Arbacia.254 The figures on Arbacia have been verified by different observers in different years.
TABLE X
Influence of Temperature upon the Time (in Minutes) Required From Insemination to the First Cell Division
| Temperature | Arbacia | Strongylocentrotus purpuratus | ||||
|---|---|---|---|---|---|---|
| Loeb and Wasteneys 1911 | Loeb and Chamberlain 1915 | |||||
| °C. | Minutes | Minutes | Minutes | |||
| 3 | 532 | |||||
| 4 | 469 | |||||
| 5 | 352 | |||||
| 6 | 275 | |||||
| 7 | 498 | 291 | ||||
| 8 | 410 | 411 | 210 | |||
| 9 | 308 | 297 | .5 | 159 | ||
| 10 | 217 | 208 | 143 | |||
| 11 | 175 | 175 | ||||
| 12 | 147 | 148 | 131 | |||
| 13 | 129 | |||||
| 14 | 116 | 121 | ||||
| 15 | 100 | 100 | 100 | |||
| 16 | 85 | .5 | ||||
| 17 | 70 | .5 | ||||
| 18 | 68 | 68 | 187 | |||
| 19 | 65 | 178 | ||||
| 20 | 56 | 56 | 175 | |||
| 21 | 53 | .3 | 178 | |||
| 22 | 47 | 46 | 175 | |||
| 23 | 45 | .5 | Upper temperature limit | |||
| 24 | 42 | |||||
| 25 | 40 | 39 | .5 | |||
| 26 | 33 | .5 | ||||
| 27 | .5 | 34 | ||||
| 30 | 33 | |||||
| 31 | 37 | |||||
These figures permitted the determination of the temperature coefficients Q10 with a sufficient degree of accuracy (see next table). It seemed of importance to attempt to decide what the chemical reaction underlying these reaction velocities is (if it is a chemical reaction). Loeb and Wasteneys255 investigated the temperature coefficient for the rate of oxidations in the newly fertilized egg of Arbacia and found that the temperature coefficient Q10 for that process does not vary in the same way as the temperature coefficient for cell division.
TABLE XI
Temperature Coefficients Q10 for the Rate of Segmentation and Oxidations in the Eggs of Strongylocentrotus AND Arbacia
| Temperature | Q10 for Rate of Segmentation in | Q10 for Rate of Oxidations in Arbacia | ||
|---|---|---|---|---|
| Strongylocentrotus | Arbacia | |||
| °C. | ||||
| 3– | 13 | 3.91 | 2.18 | |
| 4– | 14 | 3.88 | ||
| 5– | 15 | 3.52 | 2.16 | |
| 7– | 17 | 3.27 | 7.3 | 2.00 |
| 8– | 18 | 6.0 | ||
| 9– | 19 | 2.04 | 4.7 | |
| 10– | 20 | 1.90 | 3.8 | 2.17 |
| 11– | 21 | 3.3 | ||
| 12– | 22 | 1.74 | 3.1 | |
| 13– | 23 | 2.8 | 2.45 | |
| 15– | 25 | 2.5 | 2.24 | |
| 16– | 26 | 2.6 | ||
| 17.5– | 27.5 | 2.2 | 2.00 | |
| 20– | 30 | 1.7 | 1.96 | |
It is obvious that the temperature coefficient of the rate of oxidations is remarkably constant, about 2 for 10°, for various temperatures and does not show the variation from 7 or more to 2.2 for Q10 for the rate of segmentation.
Kanitz256 has shown that in a graph in which the logarithms of the segmentation velocities are drawn as ordinates and the temperatures as abscissæ the logarithms form two straight lines which are joined at an angle. According to the law of van’t Hoff and Arrhenius concerning the influence of temperature upon velocities of chemical reactions the logarithms should lie in a straight line. We are dealing therefore in these cases with two exponential curves, one representing in Arbacia the interval 7–13° and the second from 13–26°; in Strongylocentrotus between 3–9° and 9–20°.
It was found in these experiments that if measurements of the Q10 of later stages of development are attempted the variations due to unavoidable difficulties become too great to permit an equal degree of reliability in the determinations.
The vast importance of this influence of temperature upon the rate of development is seen in the fact that in addition to the food supply the rate of the maturing of plants and animals depends on this factor.
2. This influence of temperature upon development has been used to find the conditions determining fluctuating variation. The reader knows that by this expression are understood the differences between individuals of a pure strain or breed. These variations are not inherited, a fact contrary to the idea of Darwin, who assumed that by the selection of extreme cases of fluctuating variation new varieties could develop. What is the basis of this fluctuating variation? The writer concluded that if fluctuating variations were due to a slight variation in the quantity of a specific substance—in some cases an enzyme—required for the formation of a hereditary character, the temperature coefficient might be used to test the idea. We have just seen that the time required from insemination until the cell division of the first egg occurs is very sharply defined for each temperature. If a large number e. g. one hundred or more eggs are under observation simultaneously in a microscopic field it can be seen that they do not all segment at the same time but in succession; this is the expression of fluctuating variation. Miss Chamberlain and the writer have measured the time which elapses between the moment the first egg of such a group segments and the moment the last egg begins its segmentation, and found that this latitude of variation is also very definite for each temperature, and that its temperature coefficient is for each interval of 10° practically identical with the temperature coefficient of the segmentation for the same interval.257 The slight deviations are practically all in the same sense and accounted for by a slight deficiency in the nature of the experiments. The two following tables give the latitude of variations for different temperatures for the first segmentation in Arbacia and the temperature coefficient for this latitude and the rate of segmentation. These two latter coefficients are practically identical.
TABLE XII
| Temperature | Latitude of Variation | Temperature | Latitude of Variation |
|---|---|---|---|
| °C. | Minutes | °C. | Minutes |
| 19 | 52.5 | 18 | 12.0 |
| 10 | 39.5 | 19 | 12.5 |
| 11 | 26.0 | 20 | 19.6 |
| 12 | 22.5 | 21 | 18.0 |
| 13 | 19.2 | 22 | 17.8 |
| 14 | 17.5 | 23 | 18.0 |
| 15 | 13.0 | 24 | 18.0 |
| 25 | 15.0 |
TABLE XIII
| Temperature Interval | temperature coefficient of | |
|---|---|---|
| Latitude of Variation | Segmentation | |
| °C. | ||
| 19–19 | 4.2 | 4.7 |
| 10–20 | 3.9 | 3.8 |
| 11–21 | 3.2 | 3.3 |
| 12–22 | 2.8 | 3.1 |
| 13–23 | 2.4 | 2.8 |
| 14–24 | 2.3 | 2.8 |
| 15–25 | 2.6 | 2.5 |
If we assume that the temperature coefficient for the segmentation of the egg is that of a chemical reaction (other than oxidation) underlying the process of segmentation, the fluctuating variation in the time of the segmentations of the various eggs fertilized at the same time is due to the fact that the mass of the enzyme controlling that reaction varies within definite limits in different eggs. The first egg segmenting at a given temperature has the maximal, the last egg segmenting has the minimal mass of enzyme. It should be added that the time of the first segmentation is determined by the cytoplasm and is not a Mendelian character, as was stated in a previous chapter.
3. The point of importance to us is that the influence of temperature upon the organism is so constant that if disturbing factors are removed it would be possible to use the time from insemination to the first segmentation of an egg of Arbacia as a thermometer on the basis of the table on page 295.
Facts of this character should dispose of the idea that the organism as a whole does not react with that degree of machine-like precision which we find in the realm of physics and chemistry. Such an idea could only arise from the fact that biologists have not been in the habit of looking for quantitative laws, chiefly, perhaps, because the difficulties due to disturbing secondary factors were too great. The worker in physics knows that in order to discover the laws of a phenomenon all the disturbing factors which might influence the result must first be removed. When the biologist works with an organism as a whole he is rarely able to accomplish this since the various disturbing influences, being inseparable from the life of the organism, can often not be entirely removed. In this case the biologist must look for an organism in which by chance this elimination of secondary conditions is possible. The following example may serve as an illustration of this rather important point in biological work. Although all normal human beings have about the same temperature, yet if the heart-beats of a large number of healthy human beings are measured the rate is found to vary enormously. Thus v. Körösy found among soldiers under the most favourable and most constant conditions of observations—the soldiers were examined early in the morning before rising—variations in the rate of heart-beat between 42 and 108. In view of this fact, those opposed to the idea that the organism as a whole obeys purely physicochemical laws might find it preposterous to imagine that the rate of heart-beat could be used as a thermometer. Yet if we observe the influence of temperature on the rate of the heart-beat of a large number of embryos of the fish Fundulus, while the embryos are still in the egg, we find that at the same temperature each heart beats at the same rate, the deviations being only slight and such as the fluctuating variations would demand.258 This constancy is so great that the rate of heart-beat of these embryos could in fact be used as a rough thermometer. The influence of temperature upon the rate of heart-beat is completely reversible so that when we measure the rate for increasing as well as for decreasing temperatures we get approximately the same values as the following table shows.
TABLE XIV
| Temperature | Time Required for Nineteen Heart-beats in the Embryo of Fundulus | |
|---|---|---|
| °C. | Seconds | |
| 30 | 6. | 25 |
| 25 | 8. | 5 |
| 20 | 11. | 5 |
| 15 | 19. | 0 |
| 10 | 32. | 5 |
| 15 | 61. | 0 |
| 10 | 33. | 5 |
| 15 | 18. | 8 |
| 20 | 12. | 0 |
| 25 | 10. | 0 |
| 30 | 6. | 0 |
Why does each embryo have the same rate of heart-beat at the same temperature in contradistinction to the enormous variability of the same rate in man? The answer is, on account of the elimination of all secondary disturbing factors. In the embryo of Fundulus the heart-beat is a function almost if not exclusively of two variables, the mass of enzymes for the chemical reactions underlying the heart-beat and the temperature. By inheritance the mass of enzymes is approximately the same and in this way all the embryos beat at the same rate (within the limits of the fluctuating variation) at the same temperature. This identity exists, however, only as long as the embryo is relatively quiet in the egg. As soon as the embryo begins to move this equality disappears since the motion influences the heart-beat and the motility of different embryos differs.
In man the number of disturbing factors is so great that no equality of the rate for the same temperature can be expected. Differences in emotions or the internal secretions following the emotions, differences in previous diseases and their after-effects, differences in metabolism, differences in the use of narcotics or drugs, and differences in activity are only some of the number of variables which enter.
4. As stated above the temperature influences practically all life phenomena in a similar characteristic way, e. g., the production of CO2 in seeds259 and the assimilation of CO2 by green plants.260 The writer would not be surprised if even the aberrations in the colour of butterflies under the influence of temperature turned out to be connected with the temperature coefficient. The experiments of Dorfmeister, Weismann, Merrifield, Standfuss, and Fischer, on seasonal dimorphism and the aberration of colour in butterflies have so often been discussed in biological literature that a short reference to them will suffice. By seasonal dimorphism is meant the fact that species may appear at different seasons of the year in a somewhat different form or colour. Vanessa prorsa is the summer form, Vanessa levana the winter form of the same species. By keeping the pupæ of Vanessa prorsa several weeks at a temperature of from 0° to 1° Weismann succeeded in obtaining from the summer chrysalids specimens which resembled the winter variety, Vanessa levana.
If we wish to get a clear understanding of the causes of variation in the colour and pattern of butterflies, we must direct our attention to the experiments of Fischer, who worked with more extreme temperatures than his predecessors, and found that almost identical aberrations of colour could be produced by both extremely high and extremely low temperatures. This can be seen clearly from the following tabulated results of his observations. At the head of each column the reader finds the temperature to which Fischer submitted the pupæ, and in the vertical column below are found the varieties that were produced. In the vertical column A are given the normal forms:
TABLE XV
| 0° to -20°C. | 0° to +10°C. | A (Normal Forms) | +35° to +37°C. | +36° to +41°C. | +42° to +46°C. |
|---|---|---|---|---|---|
| ichnusoides (nigrita) | polaris | urticæ | ichnusa | polaris | ichnusoides (nigrita) |
| antigone (iokaste) | fischeri | io | —— | fischeri | antigone (iokaste) |
| testudo | dixeyi | polychloros | erythromelas | dixeyi | testudo |
| hygiæa | artemis | antiopa | epione | artemis | hygiæa |
| elymi | wiskotti | cardui | —— | wiskotti | elymi |
| klymene | merrifieldi | atalanta | —— | merrifieldi | klymene |
| weismanni | porima | prorsa | —— | porima | weismanni |
The reader will notice that the aberrations produced at a very low temperature (from 0° to -20°C.) are absolutely identical with the aberrations produced by exposing the pupæ to extremely high temperatures (42° to 46°C.). Moreover, the aberrations produced by a moderately low temperature (from 0° to 10°C.) are identical with the aberrations produced by a moderately high temperature (36° to 41°C.).
From these observations Fischer concludes that it is erroneous to speak of a specific effect of high and of low temperatures, but that there must be a common cause for the aberration found at the high as well as at the low temperature limits. This cause he seems to find in the inhibiting effects of extreme temperatures upon development.
If we try to analyse such results as Fischer’s from a physicochemical point of view, we must realize that what we call life consists of a series of chemical reactions, which are connected in a catenary way; inasmuch as one reaction or group of reactions (a) (e. g., hydrolyses) causes or furnishes the material for a second reaction or group of reactions (b) (e. g., oxidations). We know that the temperature coefficient for physiological processes varies slightly at various parts of the scale; as a rule it is higher near 0° and lower near 30°. But we know also that the temperature coefficients do not vary equally for the various physiological processes. It is, therefore, to be expected that the temperature coefficients for the group of reactions of the type (a) will not be identical through the whole scale with the temperature coefficients for the reactions of the type (b). If therefore a certain substance is formed at the normal temperature of the animal in such quantities as are needed for the catenary reaction (b), it is not to be expected that this same perfect balance will be maintained for extremely high or extremely low temperatures; it is more probable that one group of reactions will exceed the other and thus produce aberrant chemical effects, which may underlie the colour aberrations observed by Fischer and other experimenters.
It is important to notice that Fischer was also able to produce aberrations through the application of narcotics. Wolfgang Ostwald has produced experimentally, through variation of temperature, dimorphism of form in Daphnia.
5. Next or equal in importance with the temperature is the nature of the medium in which the cells are living.
It has often been pointed out that the marine animals and the cells of the body of metazoic animals are surrounded by a medium of similar constitution, the sea water and the blood or lymph, both media being salt solutions differing in concentration but containing the three salts NaCl, KCl, and CaCl2 in about the same relative concentration, namely 100 molecules NaCl : 2.2 molecules of KCl : 1.5 molecules of CaCl2. This has suggested to some authors the poetical dream that our home was once the ocean, but we cannot test the idea since unfortunately we cannot experiment with the past. Plants, unicellular fresh-water algæ, and bacteria do not demand such a medium for their existence.
Herbst had shown that when sea-urchin larvæ were raised in a medium in which only one of the constituents of the sea water was lacking (not only NaCl, KCl, or CaCl2, but also Na2SO4, NaHCO3, or Na2HPO4), the eggs could not develop into plutei; from which he concluded that every constituent of the sea water was necessary. This would indicate a case of extreme adaptation to all the minutiæ of the external medium.
Experiments on a much more favourable animal for this purpose, namely, the eggs of the marine fish Fundulus, gave altogether different results. The eggs of this marine fish develop naturally in sea water but they develop just as well in fresh or in distilled water, and the young fish when they are made to hatch in distilled water will continue to live in this medium. This proves that these eggs require none of the salts of the sea water for their development. When these eggs are put immediately after fertilization into a pure solution of NaCl of that concentration in which this salt exists in the sea water practically all the eggs die without forming an embryo; but if a small quantity of CaCl2 is added every egg is able to form one, and these embryos will develop into fish and the latter will hatch. This led the writer to the conclusion that these fish (and perhaps marine animals in general) need the Ca of the sea water only to counteract the injurious effects which a pure NaCl solution has if it is present in too high a concentration.261 When we raise the eggs in a pure NaCl solution of a concentration ≦m/8 practically every egg will develop; and even in a m/4 or 3⁄8 m many or some eggs will form embryos without adding Ca; it may be that a trace of Ca present in the membrane of the egg may suffice to counter-balance the injurious action of a weak salt solution.
The concentration of the NaCl in the sea water at Woods Hole (where these experiments were made) is about m/2, and as soon as this concentration of NaCl is reached the eggs are all killed as a rule before they can form an embryo, unless a small but definite amount of Ca is added. It was found that the eggs can be raised in much higher concentrations of NaCl, but in that case more Ca must be added. The following table gives the minimal amount of CaCl2 which must be added in order to allow fifty per cent. of the eggs to form embryos. (The eggs were put into the solution an hour or two after fertilization.)
TABLE XVI
| Concentration of NaCl | Cc. m/16 CaCl2 Required for 50 c.c. NaCl Solution | ||
|---|---|---|---|
| m. | |||
| 3⁄ | 8 | 0. | 1 |
| 4⁄ | 8 | 0. | 3 |
| 5⁄ | 8 | 0. | 5 |
| 6⁄ | 8 | 0. | 6 |
| 7⁄ | 8 | 0. | 9 |
| 8⁄ | 8 | 1.2– | 1.4 |
| 9⁄ | 8 | 1.8– | 2.0 |
| 10⁄ | 8 | 2.0– | 2.5 |
| 11⁄ | 8 | 2. | 0? |
| 12⁄ | 8 | 3.0– | 3.5 |
| 13⁄ | 8 | 6. | 0 |
This indicates that the quantity of CaCl2 required to counteract the injurious effects of a pure solution of NaCl increases approximately in proportion to the square of the concentration of the NaCl solution.262 The reader will notice that the eggs can survive and develop in a solution of three times the concentration of sea water, provided enough Ca is added.
It was found also that not only Ca but a large number of other bivalent metals were able to counteract the injurious action of an excessive NaCl solution; namely Mg, Sr, Ba, Mn, Co, Zn, Pb, and Fe;263 only Hg and Cu could not be used since they are themselves too toxic. The antagonistic efficiency of the bivalent cations other than Ca was, however, smaller than that of Ca. The following table gives the highest concentration of NaCl solution in which the newly fertilized eggs of Fundulus can still form an embryo.264
50 c.c. 10⁄8 m NaCl+4 c.c. m/1 MgCl2
50 c.c. 14⁄8 m NaCl+1 c.c. m/1 CaCl2
50 c.c. 11⁄8 m NaCl+1 c.c. m/1 SrCl2
50 c.c. 7⁄8 m NaCl+1 c.c. m/1 BaCl2
On the other hand it was seen that in all the chlorides with a univalent cation, LiCl, KCl, RbCl, CsCl, NH4Cl, the eggs could form embryos up to a certain concentration of the salt; but that this concentration could be raised by the addition of Ca.
TABLE XVII
Concentrations at which the Eggs no longer Are Able to Form Embryos
| In the Pure Salts | In the Same Salts with the Addition of 1 c.c. m CaCl2 to 50 c.c. Solution | |||
|---|---|---|---|---|
| LiCl | about 6/ | 32 m | >5/ | 8 m |
| NaCl | m/ | 2 | >14/ | 8 m |
| KCl | >11/ | 16 m | >8/ | 8 m |
| <6/ | 8 m | |||
| RbCl | >8/ | 8 m | >9/ | 8 m |
| <7/ | 8 m | |||
| CsCl | >3/ | 8 m | >8/ | 8 m |
| <4/ | 8 m | |||
In short it turned out that the injurious action of the pure solution of any chloride (or any other anion) with a univalent metal could be counteracted to a considerable extent by the addition of small quantities of a salt with a bivalent metal. It was also found in the early experiments of the writer that the bivalent or polyvalent anions had no such antagonistic effect upon the injurious action of the salts with a univalent cation.
We therefore see that what at first sight appeared in the experiments of Herbst a necessity, namely, the presence of each constituent of the sea water, turns out as a special case of a more general law; the salts with univalent ions are injurious if their concentration exceeds a certain limit and this injurious action is diminished by a trace of a salt with a bivalent cation.
Why was it not possible to prove this fact for the eggs of the sea urchin? Before we answer this question, we wish to enter upon the discussion of the nature of the injurious action of a pure NaCl solution of a certain concentration and of the annihilation of this action by the addition of a small quantity of Ca. The writer suggested in 1905 that the injurious action of a pure NaCl solution consisted in rendering the membrane of the egg permeable for NaCl, whereby the germ inside the membrane is killed; while the addition of a small amount of Ca (or any other bivalent metal) prevents the diffusion of Na into the egg,265 possibly, as T. B. Robertson266 suggested, by forming a precipitate with some constituent of the membrane, whereby the latter becomes more impermeable. The correctness of this idea can be demonstrated in the following way. When eggs of Fundulus, which are three or four days old and contain an embryo, are put into a test-tube containing 3 m NaCl they will float on this solution for about three or four hours; after that they will sink to the bottom. Before this happens the egg will shrink and when it ceases to float the embryo is usually dead. This is intelligible on the assumption that the NaCl solution entered the egg, increased its specific gravity so that it could not float any longer and killed the embryo. When we add, however, 1 c.c. 10⁄8 m CaCl2 to 50 c.c. 3 m NaCl the eggs will float, the heart will continue to beat normally and the embryo will continue to develop for three days or more, because the calcium prevents the NaCl from entering into the egg.267 For if we put a newly hatched embryo into 50 c.c. NaCl+1 c.c. 10⁄8 m CaCl2 it will die almost instantly; hence the membrane must have acted for three or more days as a shield which prevented the NaCl from diffusing into the egg in the presence of CaCl2.
The same experiments cannot be demonstrated in the sea-urchin egg, first, because it can live neither in distilled water nor in very dilute nor very concentrated solutions; and second, because it is not separated as is the germ of the Fundulus egg from the surrounding solution by a membrane which is under proper conditions practically impermeable for water and salts.
Nevertheless it can be shown that the results at which we arrived in our experiments on Fundulus are of a general application. Osterhout268 has shown that plants which grow in the soil or in fresh water are readily killed by a pure NaCl solution of a certain concentration, while they can resist the same concentration of NaCl if some CaCl2 is added. Wo. Ostwald269 has shown the same for a species of Daphnia. We, therefore, come to the conclusion that the injurious action following an alteration in the constitution of the sea water is in some of the cases due to an increase in the permeability of the membranes of the cell, whereby substances can diffuse into the cell which when the proper balance prevails cannot diffuse. For this balance the ratio of the concentration of the salts with univalent cation Na and K over those with bivalent cation Ca and Mg CNa+K saltsCCa+Mg salts is of the greatest importance.
6. The importance of this quotient appears in the so-called “behaviour” of marine animals. We have mentioned the newly hatched larvæ of the barnacle in connection with heliotropism. These larvæ swim in a trough of normal sea water at the surface, being either strongly positively or negatively heliotropic. They collect as a rule in two dense clusters, one at the window and one at the room side of the dish. If such animals are put into a solution of NaCl+KCl (in the proportion in which these salts exist in the sea water), they will fall to the bottom unable to rise to the surface. They will, however, rise to the surface and swim energetically to or from the window if a certain quantity of any of the chlorides of a bivalent metal, Mg, Ca, or Sr, is added, but these movements will last only a few minutes when only one of these three salts is added; and then the animals will fall to the bottom again. If, however, two salts, e. g., MgCl2 and CaCl2, are added the animals will stay permanently at the surface and react to light as they would have done in normal sea water. These animals also can resist comparatively large changes in the concentration of the sea water, and it seemed of interest to find out whether the quotient CNaCl+KClCMgCl2+CaCl2, which just allowed all the animals to swim at the surface, had a constant value. The MgCl2+CaCl2 solution was 3⁄8 m and contained the two metals in the proportion in which they exist in the sea water; namely, 11.8 molecules MgCl2 to 1.5 molecules CaCl2. The next table gives the result.270 Since these experiments lasted a day or more each, usually two different concentrations of NaCl+KCl of the ratio 1 : 2 or 1 : 4 were compared in one experiment.
TABLE XVIII
These experiments indicate that the ratio of CNa+KCCa+Mg remains very nearly constant with varying concentrations of CNa+K.
In former experiments on jellyfish the writer had shown that there exists an antagonism between Mg and Ca271, and this observation was subsequently confirmed by Meltzer and Auer272 for mammals. It was observed that in a solution of NaCl+KCl+MgCl2 the larvæ of the barnacle were also not able to remain at the surface for more than a few minutes, while an addition of some CaCl2 made them swim permanently at the surface. Various quantities of MgCl2 were added to a mixture of m/4 or m/2 NaCl+KCl, to find out how much CaCl2, was required to allow them to swim permanently at the surface.
TABLE XIX
| C.c. of m/16 CaCl2 Necessary to Induce the Majority of the Larvæ to Swim in | ||||
|---|---|---|---|---|
| m/2 (Na+K) | m/4 (Na+K) | |||
| 50 c.c. NaCl+KCl+0.75 c.c. 3⁄8 m MgCl2 | 0. | 2 | ||
| 50 c.c. NaCl+KCl+ 1.5 c.c. 3⁄8 m MgCl2 | 0. | 4 | 0. | 3 |
| 50 c.c. NaCl+KCl+ 2.5 c.c. 3⁄8 m MgCl2 | 0. | 4 | 0. | 4 |
| 50 c.c. NaCl+KCl+ 5.0 c.c. 3⁄8 m MgCl2 | 0.7– | 0.8 | 0.7– | 0.8 |
| 50 c.c. NaCl+KCl+10.0 c.c. 3⁄8 m MgCl2 | 1. | 6 | 1. | 6 |
| 50 c.c. NaCl+KCl+15.0 c.c. 3⁄8 m MgCl2 | 1. | 8 | ||
| 50 c.c. NaCl+KCl+20.0 c.c. 3⁄8 m MgCl2 | 1. | 8 | ||
In order to interpret these figures correctly we must remember that we are dealing with two different antagonisms, one between the salts with univalent and bivalent metals and the other between Mg and Ca. The former antagonism is satisfied by the addition of Mg, inasmuch as enough Mg was present for this purpose in all solutions. What was lacking was the balance between Mg and Ca. The experiments in Table XIX therefore answer the question of the ratio between Mg and Ca. If we consider only the concentrations of Mg between 2.5 and 10.0 c.c. 3⁄8 m MgCl2—which are those closest to the normal concentration of Mg in the sea water—we notice that CCa must vary in proportion to CMg. If we now combine the results of this and the previous paragraph we may express them in the form of the theory of physiologically balanced salt solutions, by which we mean that in the ocean (and in the blood or lymph) the salts exist in such ratio that they mutually antagonize the injurious action which one or several of them would have if they were alone in solution.273 This law of physiologically balanced solutions seems to be the general expression of the effect of changes in the constitution of the salt solutions for marine or all aquatic organisms.
This chapter would not be complete without an intimation of the rôle of buffers in the sea water and the blood, by which the reaction of these media is prevented from changing in a way injurious to the organism. These buffers are the carbonates and phosphates. Instead of saying that the organisms are adapted to the medium, L. Henderson has pointed out the fitness of the environment for the development of organisms and one of these elements of fitness are the buffers against alterations of the hydrogen ion concentration.274 The ratio in which the salts of the different metals exist in the sea water is another. It is obvious that the quantitative laws prevailing in the effect of environment upon organisms leave no more room for the interference of a “directing force” of the vitalist than do the laws of the motion of the solar system.
1. It is assumed by certain biologists that the environment influences the organism in such a way as to increase its adaptation. Were this correct it would not contradict a purely physicochemical conception of life; it would only call for an explanation of the mechanism by which the adaptation is brought about. There are striking cases on record which warn us against the universal correctness of the view that the environment causes an adaptive modification of the organism. Thus the writer pointed out in 1889 that positive heliotropism occurs in organisms which have no opportunity to make use of it,275 e. g., Cuma rathkii, a crustacean living in the mud, and the caterpillars of the willow borer living under the bark of the trees. We understand today why this should be so, since heliotropism depends upon the presence of photosensitive substances, and it can readily be seen that the question of use or disuse has nothing to do with the production of certain harmless chemical compounds in the body. A much more striking example is offered in the case of galvanotropism. Many organisms show the phenomenon of galvanotropism, yet, as the writer pointed out years ago, galvanotropism is purely a laboratory product and no animal has ever had a chance or will ever have a chance to be exposed to a constant current except in the laboratory of a scientist. This fact is as much of a puzzle to the selectionist and to the Lamarckian (who would be at a loss to explain how outside conditions could have developed this tropism) as to the vitalist who would have to admit that the genes and supergenes indulge occasionally in queer freaks and lapses. The only consistent attitude is that of the physicist who assumes that the reactions and structures of animals are consequences of the chemical and physical forces, which no more serve a purpose than those forces responsible for the solar systems. From this viewpoint it is comprehensible why utterly useless tropisms or structures should occur in organisms.
2. A famous case for the apparent adaptation of animals to environment has been the blind cave animals. It is known that in caves blind salamanders, blind fishes, and blind insects are common, while such forms are comparatively rare in the open. This fact has suggested the idea that the darkness of the cave was the cause of the degeneration of the eyes. A closer investigation leads, however, to a different explanation. Eigenmann has shown that of the species of salamanders living habitually in North American caves, two have apparently quite normal eyes. They are Spelerpes maculicauda and Spelerpes stejnegeri. Two others living in caves have quite degenerate eyes, Typhlotriton spelæus and Typhlomolge rathbuni. If disuse is the direct cause of blindness we must inquire why Spelerpes is not blind.
Another difficulty arises from the fact that a blind fish Typhlogobius is found in the open (on the coast of southern California) in shallow water, where it lives under rocks in holes occupied by shrimps. The question must again be raised: How can it happen that in spite of exposure to light Typhlogobius is blind?
The most important fact is perhaps the one found by Eigenmann in the fishes of the family of Amblyopsidæ. Six species of this group live permanently in caves, are not found in the open, and have abnormal eyes, while one lives permanently in the open, is never found in caves, and one comes from subterranean springs. The one form which is found only in the open, Chologaster cornutus, has a simplified retina as well as a comparatively small eye, in other words, its eye is not normal. This indicates the possibility that the other representatives which are found only in caves also might have abnormal eyes even if they had never lived in caves.
Through these facts the old idea becomes questionable, namely, that the cave animals had originally been animals with normal eyes which owing to disuse had undergone a gradual hereditary degeneration.
Recent experiments made on the embryos of the fish Fundulus have yielded the result that it is possible to produce blindness in fish by various means other than lack of light.276 Thus the writer found that by crossing the egg of Fundulus with the sperm of a widely different species, namely, Menidia, blind embryos were produced very frequently; that is to say such embryos had the degenerate eyes characteristic of blind cave fishes. Very often no other external trace of an eye, except a gathering of pigment, could be found, while a close histological examination would possibly have resulted in the demonstration of rudiments of a lens and other tissues of the eye.
Another method of producing blind fish embryos consists in exposing the egg immediately, or soon after fertilization, to a temperature between 0° and 2° C. for a number of hours. Many embryos are killed by this treatment, but those which survive behave very much like the hybrids between Fundulus and Menidia, i. e., a number of them have quite degenerated eyes. If the eggs have once formed an embryo they can be kept at the temperature of 0° for a month or more without giving rise to blind animals. Occasionally such rudimentary eyes were also observed when eggs were kept in a solution containing a trace of KCN. Stockard has succeeded in producing cyclopean eyes in Fundulus by adding an excess of magnesium salt to the sea water in which the eggs developed or by adding alcohol, and McClendon has confirmed and added to these results.
The writer tried repeatedly, but in vain, to produce Fundulus with deficient eyes by keeping the embryos in the dark. Sperm and egg were not allowed to be exposed to the light yet the embryos without exception had normal eyes.
F. Payne raised sixty-nine successive generations of a fly Drosophila in the dark, but the eyes and the reaction of the insects to light remained perfectly normal.
Uhlenhuth has recently demonstrated in a very striking way that the development of the eyes does not depend upon the influence of light or upon the eyes functioning. He transplanted the eyes of young salamanders into different parts of their bodies where they were no longer connected with the optic nerves. The eyes after transplantation underwent a degeneration which was followed by a complete regeneration. He showed that this regeneration took place in complete darkness and that the transplanted eyes remained normal in salamanders kept in the dark for fifteen months. Hence the eyes which were no longer in connection with the central nervous system, which had received no light, and could not have functioned, regenerated and remained normal. The degeneration which took place in the eyes immediately after being transplanted was apparently due to the interruption of the circulation in the eye, and the regeneration commenced in all probability with the re-establishment of the circulation in the transplanted organ.
In our own experiments it can be shown that the circulation in the embryo was deficient in all cases where the eyes degenerated. The hybrids between Fundulus and Menidia have often a beating heart but rarely a circulation (although they form blood); and the same phenomenon occurred in the embryos which were exposed to a low temperature at an early period of their lives. Hence all the facts agree that conditions which lead to an abnormal circulation (and consequently also to an abnormal or inadequate nutrition of the embryonic eye) may prevent development and lead to the formation of blind fishes. Eigenmann states that no blood-vessels enter the eye of the blind cave salamander Typhlotriton. The presence or absence of light does not usually interfere with the circulation or nutrition of the embryonic eye, and hence does not as a rule lead to the formation of degenerated eyes.
This would lead us to the assumption that the blind fish owe their deficiency not to lack of light but to a condition which interferes with the circulation in the embryonic eye. Such a condition might be brought about by an anomaly in the germ plasm or in one chromosome, the nature and cause of which we are not able to determine at present; but which, since it occurs in the germ plasm or the chromosomes, must be hereditary. This would explain why it is, that animals with perfect eyes may occur in caves and why perfectly blind animals may occur in the open. It leaves, however, one point unexplained; namely, the greater frequency of blind species in caves or in the dark and the relative scarcity of such forms in the open.
Eigenmann has shown that all those forms which live in caves were adapted to life in the dark before they entered the cave.277 These animals are all negatively heliotropic and positively stereotropic, and with these tropisms they would be forced to enter a cave whenever they are put at the entrance. Even those among the Amblyopsidæ which live in the open have the tropisms of the cave dweller. This eliminates the idea that the cave adapted the animals for the life in the dark.
Only those animals can thrive in caves which for their feeding and mating do not depend upon visual mechanisms; and conversely, animals which are not provided with visual mechanisms can hold their own in the open, where they meet the competition of animals which can see, only under exceptional conditions. This seems to account for the fact that in caves blind species are comparatively more prevalent than in the open.
In other words, the adaptation of blind animals to the cave is only apparent; they were adapted to cave life before they entered the cave. Many animals are obviously burdened with a germinal abnormality giving rise to imperfection and smallness of the eye—the hereditary factor involved may have to do with the development of the blood-vessels and lymphatics of the eye. Such mutants can survive more easily in the cave, where they do not have to meet the competition of seeing forms, than in the open. In man also an hereditary form of blindness is known, the so-called hereditary glaucoma. It has nothing to do with light, but the disease seems to be due to an hereditary anomaly of the circulation in the eye.
Kammerer278 has recently reported that by keeping the blind European cave salamander Proteus anguinus under certain conditions of illumination he succeeded in producing two specimens with larger eyes. According to him the eyes of Proteus may develop to a certain point and then retrogress again. He states that by keeping young salamanders alternately for a week or two in sunlight and in a dark room where they were exposed to red incandescent light, two males formed somewhat larger eyes. The first year no alteration was visible. In the second year a slight increase in the size of the eyes was noticeable under the skin. In the third year the eye protruded slightly and this increased somewhat in the fourth year.
There is thus far only one case on record in animal biology in which the light influences the formation of organs. The writer found that the regeneration of the polyps of the hydroid Eudendrium does not take place if the animals are kept in the dark, while the polyps will regenerate if exposed to the light;279 and the time of exposure may be rather short according to Goldfarb.280 It is possible that Proteus resembles in this respect Eudendrium; it should be stated, however, that of many different forms tried by the writer over a number of years, Eudendrium was the only one which gave evidence of such an influence of light. Of course it is not impossible that the light might influence reflexly the development of blood-vessels in the eyes of certain animals, e. g., Proteus, and thus allow the eyes of Proteus to grow a little larger.
We therefore come to the conclusion that it is not the cave that made animals blind but that animals with a hereditary tendency towards a degeneration of the eyes can survive in a cave while they can only exceptionally survive in the open. The cause of the degeneration is a disturbance in the circulation and nutrition of the eye, which is as a rule independent of the presence or absence of light.
We may by way of a digression stop for a moment to consider the most astonishing and uncanny case of adaptation; namely, the formation of the transparent refractive media, especially the lens in front of the retina. It is due to these media that the rays which are sent out by a luminous point can be united to an image point on the retina. One part of this process is understood; namely, the formation of a lens. Wherever the optic cup of the embryo is transplanted under the epithelium the latter will be transformed into a transparent lens. When the upper edge of the iris is injured in the salamander so that the cells can multiply, the mass of newly formed cells also becomes transparent and a lens is formed. This indicates the existence of a substance in the optic cup which makes the epithelial cells transparent; and which also limits the size of the lens which is formed. The lens is not always a perfect optical instrument, on the contrary, it is as a rule somewhat defective. Of course, a great many details concerning the process of lens regeneration have still to be worked out.
3. It is well known that most marine animals die if put into fresh water and vice versa; and in salt lakes or ponds with a concentration of salt so high that most marine animals would succumb if suddenly transferred to such a solution we have a limited fauna and flora. The common idea is that marine animals become adapted to fresh water or vice versa; or to the conditions in salt lakes; especially if the changes take place gradually. Yet it can be shown that the existence of these different faunas can be explained without the assumption of an adaptive effect of the environment. The writer has worked with a marine fish Fundulus whose eggs develop naturally in sea water which, however, will develop just as well in distilled water; and the young fish hatching in distilled water live and grow in this medium. Most of the adult fish die after several days, when put suddenly into distilled water, but they can live in fresh water which contains only a trace of salt. They can also live in very concentrated sea water, e. g., twice the normal concentration. Suppose that a bay of the ocean containing such fish should suddenly become landlocked and the concentration of the sea water be thus raised to twice its natural amount; the majority of forms would die and only Fundulus and possibly a few other species with the same degree of resistance would survive. An investigator examining the salinity of the water and not knowing the natural resistance of Fundulus to changes in concentration would be inclined to assume that he had before him an instance of a gradual adaptation of the fish to a higher concentration of the sea water; whereas the fish was already immune to this high concentration before coming in contact with it.
This fish seemed a favourable object from which to find out how far an adaptation to the environment really existed; and the result was surprising. By changing the concentration of the sea water gradually it is possible to raise the natural resistance of the fish only a trifle, not much over ten per cent. The concentration of the natural sea water is a little over that of a m/2 solution of NaCl+KCl+CaCl2 in the proportion in which these three salts exist in the sea water. When adult Fundulus are put into a 10⁄8 m solution of NaCl+KCl+CaCl2 in the proportion in which these salts occur in sea water they die in less than a day, but when put from sea water directly into a 8⁄8 m or 9⁄8 m solution they can live indefinitely. It was found281 that if the concentration of the sea water was raised gradually (by m/8 a day) the fish on the fifth day could resist a 10⁄8 m solution of NaCl+KCl+CaCl2 for a month (or possibly indefinitely; the experiment was discontinued after that period). When a 10⁄8 m solution was allowed to become more concentrated slowly by evaporation (at room temperature) all the fish died rapidly when the concentration was 12⁄8 m or even below. In higher concentrations they can live only a day or two. These experiments show that while the fish is naturally immune to a 9⁄8 m NaCl+KCl+CaCl2 solution, by the method of slowly raising the concentration it may be made to tolerate a 10⁄8 m or 11⁄8 m solution, but not more. These fish when once adapted to a 10⁄8 m solution can be put suddenly into a very weak solution, e. g., a m/80 NaCl, without suffering and when brought back into a 10⁄8 m solution of NaCl+KCl+CaCl2 they will continue to live. If they remain for several days in the weak solution their power of resistance to 10⁄8 m NaCl+KCl+CaCl2 solution is weakened.
What change takes place when the fish is made more resistant and why is its normal resistance so great? The answer based on the writer’s experiments seems to be as follows: Fundulus is comparatively resistant to sudden changes in the concentration of the sea water between m/80 and 9⁄8 m because it possesses a comparatively impermeable skin whose permeability is not seriously altered by sudden changes within these limits of concentration; while if these limits are exceeded and the fish are brought suddenly into too high a concentration the skin becomes permeable and the fish dies, the gills becoming unfit for use or nerves being injured by the salt which diffuses into the fish.
The fact, that by slowly raising the concentration to 10⁄8 m the fish may resist this limit, is in reality no adaptation. There is no sharp limit between the injurious and non-injurious concentration. We have seen that the fish is naturally immune to a 9⁄8 m solution. It is also naturally immune to a 10⁄8 m or 11⁄8 m solution if we give it time to compensate the injurious effects of a 10⁄8 m solution by the repairing action of its blood or kidneys. Beyond this no rise is possible. In reality adaptation does not exist in this case.
In former experiments the writer had shown that a pure NaCl solution of that concentration in which this fish naturally lives kills it very rapidly, while it lives in such a solution indefinitely if a little CaCl2 is added. The explanation of this fact is that the pure NaCl solution is able to diffuse into the tissues of the animal while the addition of a trace of CaCl2 renders the membrane practically impermeable to NaCl. The question then arose whether it was possible to make the fish more resistant to a pure NaCl solution of sufficiently high concentration and how this could be done. On the basis of the idea of an adaptive effect of the environment we should expect that by gradually raising the concentration of a pure NaCl solution the latter would gradually alter the animal and make it more resistant. The method of procedure suggested was therefore to put the fish first in low and gradually into increasing concentrations of NaCl. This method was tried and found futile for the purpose. Fundulus when put from sea water (after having been washed) into a 6⁄8 m NaCl solution die in about four hours. When kept previously in a weaker NaCl solution they die if anything more quickly. But it is possible to make them live longer in a 6⁄8 m solution of NaCl; we have to proceed, however, by a method which is in contrast with the ideas of the adaptive influence of the environment. When the fish are first treated with sea water (or with a mixture of NaCl+KCl+CaCl2) of a higher concentration so that they become adapted to a 10⁄8 m solution of NaCl+KCl+CaCl2 or to 10⁄8 m sea water, they become also more resistant to an otherwise toxic solution of NaCl. Fish taken directly from sea water were killed in less than four hours when put into a 6⁄8 m NaCl solution, while fish of the same lot previously adapted to 10⁄8 m sea water in the manner described above lived two or three days in a 6⁄8 m NaCl solution.282
It is not impossible that it was the high concentration of calcium in the 10⁄8 m sea water which rendered the fish more immune to a subsequent treatment with NaCl. We know why a pure NaCl solution kills them and we also know why the addition of CaCl2 protects them against this pernicious effect. It is rather strange that where the conditions of the experiments are clear we find nothing to indicate an adaptive effect of the environment.
4. Ehrlich’s work on trypanosomes seems to indicate a remarkable power of adaptation on the part of organisms to certain poisons. If the writer understands these experiments correctly they consisted in infecting a mouse with a certain strain of trypanosomes, and treating it with a certain arsenic compound, which inhibited somewhat the propagation of the parasites but did not kill them all. Four or five days later trypanosomes from this mouse were transmitted to another mouse and after twenty-four hours this mouse was treated with a stronger dose of the same arsenic compound; and this process was repeated. After the third transmission or later, the trypanosomes can resist considerably higher doses of the same poison than at first and this resistance is retained for years. Ehrlich seems to have taken it for granted that he had succeeded in transforming the surviving trypanosomes into a type which is permanently more resistant to the arsenic compound than was the original strain.
The writer is not entirely convinced that in these experiments a possibility was sufficiently considered which is suggested by Johannsen’s experiments on the importance of pure lines in work on heredity. According to this author a strain of trypanosomes taken at random should, in all likelihood, contain a population consisting of strains with different degrees of resistance. If a high but not the maximal concentration of an arsenic compound is repeatedly injected into the infected mice the weaker populations of trypanosomes are killed and only the more resistant survive. These of course continue to retain their resistance if transplanted to hosts of the same species. According to this interpretation the arsenic-fast strain may possibly have existed before the experiments were made, and Ehrlich’s treatment consisted only in eliminating the less resistant strains.
On the other hand, it has been shown that if an arsenic-fast strain of trypanosomes is carried through a tsetse fly it loses its arsenic-fastness. This fact may possibly eliminate the applicability of the pure line theory to a discussion of the nature of the arsenic-fastness, but it seems that further experiments are desirable.
5. Dallinger stated that he succeeded in adapting certain protozoans to a temperature of 70° C. by gradually raising their temperature during several years. It is desirable that this statement be verified; until this is done doubts are justified. Schottelius found that colonies of Micrococcus prodigiosus when transferred from a temperature of 22° to that of 38° no longer formed pigment and trimethylamine. After the cocci had been cultivated for ten or fifteen generations at 38° they failed to form pigment even when transferred back to 22° C. Dieudonné283 used Bacillus fluorescens for similar purposes. At 22° it forms a fluorescing pigment and trimethylamine, but not at 35°. By constantly cultivating this bacillus at 35° Dieudonné found that after the fifteenth generation had been cultivated at 35° the bacillus produced pigment and trimethylamine at 35°. Davenport and Castle284 found that tadpoles of a frog kept at 15° went into heat rigour at 40.3° C., while those kept for twenty-eight days at 25° were not affected by this temperature but went into heat rigour at 43.5°. When the latter tadpoles were put back for seventeen days to a temperature of 15° they had lost their resistance to high temperature partially, but not completely, since they went into heat rigour at 41.6°. The authors suggest that this adaptation to a higher temperature is due to a loss of water on the part of protoplasm, whereby the latter becomes more resistant to an increase in temperature. This idea was put to a test by Kryž285, who found that the coagulation temperature of their muscle plasm is not altered by keeping cold-blooded animals at different temperatures.
Loeb and Wasteneys286 found that Fundulus taken from a low temperature of 10° C. die in less than two hours when suddenly transferred to sea water of 29° C.; and in a few minutes if suddenly transferred to a temperature of 35° C. If, however, the fish were transferred to a temperature of 27° C. for forty hours they could live indefinitely in sea water of 35°. By exposing the fish each day two hours to a gradually rising temperature they could render them resistant to a temperature of 39°. The remarkable fact was that fish if once made resistant to a high temperature (35°) did not lose this resistance when kept for four weeks at from 10° to 14° C. Control fish taken from the same temperature died in from two to four minutes; immunized fish taken from 10° and put directly to 35° C. lived for many hours or indefinitely. They will even retain this immunity when kept for two weeks at a temperature of 0.4° C.
Why is it that an animal can in general resist a high temperature better if the latter is raised gradually than when it is raised suddenly? Physics offers us an analogy to this phenomenon in the experience that glass vessels which burst easily when their temperature is raised suddenly, remain intact when the temperature is raised gradually. Glass is a poor conductor of heat and when the temperature is raised suddenly inside a glass cylinder the inner layer of the cylinder expands while the outer layer on account of the slowness of conduction of heat does not expand equally and the cylinder may burst. We might assume that the sudden increase in temperature brings about certain changes in the cells (e. g., an increase in permeability or destruction of the surface layer?). If the rise of temperature occurs gradually the blood or lymph or the cell sap may have time to repair the damage, and this repair seems to be irreversible, at least for some time, as the experiments on Fundulus seem to indicate. If the temperature rises too rapidly the damage cannot be repaired quickly enough by the cell or body liquids.
It is also to be considered that substances might be formed in the body at a higher temperature which do not exist at a lower temperature, and vice versa, and this might explain results like those of Schottelius or Dieudonné and many others.
6. The theory of an adapting effect of the environment has often been linked with the assumption of the inheritance of acquired characters. The older claims of the hereditary transmission of acquired characters, such as Brown-Séquard’s epilepsy in guinea pigs after the cutting of the sciatic nerve, have been shown to be unjustified or have found a different and more rational explanation. Recently P. Kammerer has claimed to have proven by new experiments that by environmental changes, hereditary changes can be produced.
It has been mentioned already that the mature male frogs and toads possess during the breeding season lumps on the thumbs or arms which are pigmented and which bear numerous minute horny black spines; these secondary sexual characters serve the male frog in holding the females in the water during copulation. There is one species which does not possess this sexual character, namely the male of the so-called midwife toad (Alytes obstetricans). In this species the animals copulate on land, and it is natural to connect the lack of this secondary sexual character in the male with its different breeding habit. Kammerer now forced such toads to copulate in water instead of on land (by keeping the animals in a terrarium with a high temperature). He makes the statement that by forcing the parents to lay their eggs during successive spawning periods in water he finally obtained offspring which under normal temperature conditions lay their eggs naturally in water; in other words, they have changed their habits. We will not discuss this part of his statement since the breeding habits of animals in captivity are liable to be abnormal. But Kammerer makes the further important statement287 that the male offspring of such couples will in the third generation produce the swelling on the thumb and the usual roughness, and in the fourth generation black pads and hypertrophy of the muscles of the forearm will appear. In other words, he reports having succeeded in producing an inheritance of an acquired morphological character which has never been known to occur in this species. Bateson, on account of the importance of the case, wished to examine it more closely and I will quote his report.
The systematists who have made a special study of Batrachia appear to be agreed that Alytes in nature does not have these structures; and when individuals possessing them can be produced for inspection it will, I think, be time to examine the evidence for the inheritance of acquired characters more seriously. I wrote to Dr. Kammerer in July, 1910, asking him for the loan of such a specimen and on visiting the Biologische Versuchsanstalt in September of the same year I made the same request, but hitherto none has been produced. In matters of this kind much generally depends on interpretations made at the time of observation; here, however, is an example which could readily be attested by preserved material.288
More recently the same author has reported another hereditary morphological change brought about by outside conditions.289 A certain salamander (Salamandra maculosa) has yellow spots on a generally dark skin. Kammerer states that if such salamanders are kept on a yellow ground they become more yellow, not by an extension of the chromatophores (which would not be surprising) but by actual multiplication and growth of the yellow pigment cells; while the black skin is inhibited in its growth. The reverse is true if these salamanders are kept on black soil; in this case according to Kammerer the growth of the yellow cells of the skin is inhibited while the black part of the skin grows. Curiously enough, according to him, these induced changes are hereditary. Here again we are dealing with the inheritance of an acquired morphological character.
Megusar290 has repeated Kammerer’s experiments on salamanders but contradicts him by stating that the colour of the soil has no influence on the colouration of salamanders. Of course, we know the phenomenon of colour adaptation in which the animal changes its colour pattern according to the environment. This is an effect of the retina image on the skin and has been interpreted by the writer as a case of colour telephotography, for which no physical explanation has yet been found.291 This phenomenon, however, does not lead to any hereditary change of colour.
Kammerer makes many statements on the heredity of acquired modifications of instinct; indeed he claims that an interest in music on the part of parents produces offspring with musical talent. In such claims much depends upon the subjective interpretation of the observer.
The writer is not aware that there is at present on record a single adequate proof of the heredity of an acquired character. We have records of changes in the offspring by poisoning the germ plasm by alcohol given to parents—as in Stockard’s well-known experiments—or by exposing butterflies to extreme temperatures, but in these cases the germ cells were poisoned or altered by the alcohol or by chemical compounds produced at very low or very high temperatures. This is of course an entirely different thing from stating that by inducing the midwife toad to lay its eggs in the water the male offspring acquires the pads and horns of other species of frogs on its thumb; or that by keeping black salamanders on yellow paper the offspring is more yellow. Yet if there is an inheritance of acquired characters which can in any way throw light on the so-called phenomena of adaptation it must consist in results such as Kammerer claims to have obtained.
While the writer does not decline to accept Ehrlich’s interpretation of the arsenic-fast strains of trypanosomes or Kammerer’s statements in regard to the inheritance of acquired character, he feels that more work should be done before they can be used for our problem.
7. This attitude leaves us in a quandary. The whole animated world is seemingly a symphony of adaptation. We have mentioned already the eye with its refractive media so well curved and placed that a more or less perfect image of the outside objects is focussed exactly on the retina; and this in spite of the fact that lens and retina develop independently; we have mentioned and discussed the cases of instincts or automatic arrangements which are required to perpetuate life—the attraction of the two sexes and the automatic mechanisms by which sperm and egg are brought together; the maternal instincts by which the young are taken care of; and all those adaptations by which animals get their food and the suitable conditions of preservation. Can we understand all these adaptations, without a belief in the heredity of acquired characters? As a matter of fact the tenacity with which some authors cling to such a belief is dictated by the idea that this is the only alternative to the supra-naturalistic or vitalistic ideas. The writer is of the opinion that we do not need to depend upon the assumption of the heredity of acquired characters, but that physiological chemistry is adequate for this purpose.
The earlier writers explained the growth of the legs in the tadpole of the frog or toad as a case of an adaptation to life on land. We know through Gudernatsch that the growth of the legs can be produced at any time even in the youngest tadpole, which is unable to live on the land, by feeding the animal with the thyroid gland. As we have stated in Chapter VII, it is quite possible that in nature the legs of the tadpole begin to grow when enough of the thyroid or a similar compound has been formed or is circulating in the animal.
It might justly be claimed as a case of adaptation that the egg attaches itself to the wall of the uterus and calls forth the formation of the decidua. We have mentioned the observation of Leo Loeb that the corpus luteum of the ovary gives off a substance to the blood which alters the tissues in the uterus in such a way that contact with any foreign body (e. g., the egg) induces this decidua formation. Again what appeared as adaptation when unknown turns out to be a result of the action of a definite chemical substance circulating in the body.
It appears as a case of adaptation that the eggs of the majority of animals cannot develop without a spermatozoön, and yet we can imitate the activating effect of a spermatozoön on the egg by definite chemical compounds, which leads to the suggestion that the activating effect of the spermatozoön on the egg might be due to the fact that it carries such a compound.
The wonderful adaptations exhibited in the mating instincts seem to be due to definite substances secreted by the sex glands, as was shown by Steinach (Chapter VII). Here, again, the process as popularly conceived, is the reverse of the truth; those survive that have the equipment,—they did not acquire the equipment under the influence of environment.
It is absolutely imperative for green plants that their stems and leaves be exposed to the light since only in this way are they able to form carbohydrates; and it is equally essential that the roots should grow into the soil so that the plant may get the nitrates and phosphates required to build up its proteins and nucleins. This result is, in the language of adaptationists, brought about by an adaptive response of the plant to the light. In reality this adaptive response is due (Chapter X) to the presence of a photosensitive substance present in almost all green plants.
Lewis has shown that if the optic cup is transplanted under the skin of a young larva into any part of the body the skin in contact with the optic cup will form a lens; it looks as if a chemical substance from the optic cup were responsible for the formation of the lens.
These examples might be multiplied indefinitely. They all indicate that apparent morphological and instinctive adaptations are merely caused by chemical substances formed in the organism and that there is no reason for postulating the inheritance of acquired characters. We must not forget that there are just as many cases where chemical substances circulating in the body lead to indifferent or harmful results. As an example of the first type, we may mention the existence of heliotropism in animals living in the dark, of the latter type, the inheritance of deficiencies like colour-blindness or glaucoma.
While it is possible for forms with moderate disharmonies to survive, those with gross disharmonies cannot exist and we are not reminded of their possible existence. As a consequence the cases of apparent adaptation prevail in nature.
The following observation may serve to give an idea how small is the number of existing or durable forms compared with the number of forms incapable of existence. We have mentioned the fact observed by Moenkhouse, the writer, and Newman, that it is possible to fertilize the eggs of each marine bony fish with the sperm of practically every other marine bony fish. The number of teleosts at present in existence is about ten thousand. If we accomplish all possible hybridizations, one hundred million different crosses will result. Of these only a small fraction of one per cent. can live (see Chapter I), and it is generally the lack of a proper circulation which inhibits them from reaching maturity. It is, therefore, no exaggeration to state that the number of species existing today is only an extremely small fraction of those which can and possibly do originate, but which escape our notice and disappear because they cannot live or reproduce. If we consider these facts we realize that the mere laws of chance are adequate to account for the fact of the apparently purposeful adaptations; as they are adequate to account for the Mendelian numbers.
Darwin’s work has been compared to that of Copernicus and Galileo inasmuch as all these men freed the mind from the incubus of Aristotelian philosophy which, with the efficient co-operation of the church and the predatory system of economics, caused the stagnation, squalor, immorality, and misery of the Middle Ages. Copernicus and Galileo were the first to deliver the intellect from the idea of a universe created for the purpose of man; and Darwin rendered a similar service by his insistence that accidental and not purposeful variations gave rise to the variety of organisms. In this struggle for intellectual freedom the names of Huxley and Haeckel must be gratefully remembered, since without them Darwin’s idea would not have conquered humanity.
Darwin assumed that the small fluctuating variations could accumulate to larger variations and thus cause new forms to originate.
It was the merit of de Vries292 to have pointed out that fluctuating variations are not hereditary and hence could not have played the rôle assigned to them by Darwin, while discontinuous variations as they appear in the so-called “sports” or mutations are inherited. This was an important step in the history of the theory of evolution. It did not touch the foundation of Darwin’s work, namely the substitution of the idea of an accidental evolution for that of a purposeful creation; it only modified the conception of the possible mechanism of evolution. According to de Vries, there are special species or groups of species which are in a state of mutation. He considers the evening primrose on which he made his observations as one of these forms. Morgan and his pupils have observed over 130 mutations in a fly Drosophila. From our present limited knowledge we must admit the possibility that the tendency toward the production of mutants is not equally strong in different forms. Whether this part of de Vries’s idea is or is not correct there can be no doubt that variations occur which consist in the loss and apparently, though in rarer cases, in the gain or a modification of a Mendelian factor. If we wish to visualize the basis of such a change we may do so by imagining well-defined chemical constituents in one or more of the chromomeres undergoing a chemical change.
This way of looking at the origin of variation has had the effect of putting an end to the vague speculations concerning the evolution of one form from another. We demand today the experimental test when such a statement is made and as a consequence the amount of mere speculation in this field has diminished considerably.
It is possible that any further progress concerning evolution must come by experimental attempts to bring about at will definite mutations. Such attempts have been reported but they are not all beyond the possibility of error.293 The most remarkable among them are those by Tower who by a very complicated combination of effects of temperature and moisture claims to have produced definite mutations in the potato beetle. The conditions for these experiments are so expensive and complicated that a repetition by other investigators has not yet been possible.
It is, however, still uncertain whether the mere addition or loss of Mendelian characters can lead to the origin of new species. Species specificity is determined by specific proteins (Chapter III.), while some Mendelian characters at least seem to be determined by hormones or substances which need neither be proteins nor specific for the species.
1. It is an old saying that we cannot understand life unless we understand death. The dead body, if its temperature is not too low and if it contains enough water, undergoes rapid disintegration. It was natural to argue that life is that which resists this tendency to disintegration. The older observers thought that the forces of nature determined the decay, while the vital force resisted it. This idea found its tersest expression in the definition of Bichat, that “life is the sum total of the forces which resist death.” Science is not the field of definitions, but of prediction and control. The problem is: first, how does it happen that as soon as respiration has ceased only for a few minutes the human body is dead, that is to say, will commence to undergo disintegration, and second, what protects the body against this decay while the respiration goes on, although temperature and moisture are such as to favour decay?
The earlier biologists had already raised the question why it was that the stomach and intestine did not digest themselves. The hydrochloric acid and the pepsin in the stomach and the trypsin in the intestine digest proteins taken in in the form of food; why do they not digest the proteins of the cells of the stomach and the intestine? They will promptly digest the stomach as soon as the individual is dead, but not during life. A self-digestion may also be caused if the arteries of the stomach are ligatured. Claude Bernard and others suggested that the layer of mucus protected the cells of the stomach and of the intestine from the digestive enzymes; or that the epithelial layer had a protective effect. Pavy suggested that the alkali of the blood had a protective action. All these theories became untenable when Fermi showed that all kinds of living organisms, protozoans, worms, arthropods, are not digested in solutions of trypsin as long as they are alive, while they are promptly digested in the same solution when dead.294 This is in harmony with the fact that many parasites live in the intestine without being digested as long as they are alive. Fermi concluded that the living cell cannot be attacked by the digestive ferments, while with death a change occurs by which they can be attacked. But what is this change? Fermi seems to be inclined to think that the “living molecule” of protein is not hydrolysable (perhaps because the enzyme cannot attach itself to it?), while a change in the constitution or configuration of the proteins takes place after respiration has ceased. The fact that the living cell resists the digestive action of trypsin and pepsin has found two other modes of explanation, first, that the cells are surrounded by a membrane or envelope through which the enzyme cannot diffuse, and second, that the living cells possess antiferments. But the so-called antiferments are also said to exist after the death of the cell, whereas after death the cell is promptly digested. Frédéricq, as well as Klug, has shown that worms which are not attacked by trypsin are digested by this enzyme when they are cut into small pieces; although the pieces of course contain the antienzyme. The other suggestion that a membrane impermeable for trypsin protects the cells would explain why living protozoa are not digested by trypsin, but it leaves another fact unexplained, namely, the autodigestion of all the cells after death by enzymes contained in the cells themselves.
2. The disintegration of the body after death is not caused exclusively or even chiefly by the digestive enzymes of the intestinal tract or the micro-organisms entering the dead body from the outside, but by the enzymes contained in the cells themselves. This phenomenon of autolysis295 was first characterized by Hoppe-Seyler.296
All organs suffering death within the organism, in the absence of oxygen, undergo softening and dissolution in a manner resembling that of putrefaction. In the course of that process, albuminous matter gives rise to leucin and tyrosin, fat to free acids and soaps. This maceration, identical with the pathological conception of softening, is accomplished without giving rise to ill odour and is a process similar to the one resulting from the action of water, acids, and digestive enzymes.
In work of this kind, rigid asepsis is required to exclude the possibility of bacterial infection and this was first done by Salkowski, who showed that in aseptically kept tissues like liver and muscle the amount of substances that can be extracted with hot water increases considerably. By the work of others, especially Martin Jacoby and Levene, it was established that the power of self-digestion is shared by all organs. Analysis of the products of the autodigestion of tissues shows that, e. g., the amino acids, which constitute the proteins, are produced. Dakin, Jones, and Levene demonstrated the hydrolytic products of the nucleins, in the case of the self-digestion of tissues.297
Again the question arises: Why do the tissues not undergo autolysis during lifetime and what protects them, and the answer is that self-digestion is a consequence of the lack of oxidations. The presence of antiferments must continue after death and cannot be the cause which prevents the self-digestion during life, since nothing indicates the destruction of the hypothetical antidigestive enzymes through lack of oxygen. The recent work of Bradley and Morse298 and of Bradley299 has thrown some light on the problem. These authors found that proteins of the liver which are indigestible can be made digestible by the liver enzymes if an acid salt or a trace of acid is added to the mixture. A m/200 HCl solution gives marked acceleration of the autodigestion of the liver. This would explain why autodigestion takes place after oxidations cease. In many if not all the cells, acids are constantly formed during lifetime, e. g., lactic acid, which through oxidation are turned to CO2, and this diffuses into the blood so that the H ion concentration in the cells does not rise materially. If, however, the oxidations cease, as is the case after death, the formation of lactic acid continues, but the acid is not oxidized to CO2 and thus removed, and as a consequence the H ion concentration increases in the cells and the self-digestion of proteins, which the digestive enzymes contained in the cells themselves could not attack formerly, becomes possible. Acid increases the digestibility of a protein, probably by salt formation. Theoretically we should not be surprised that while in the liver an increase in the CH favours autolysis in other tissues the same result is produced by the reverse effect. We might say that the preservation of a certain CH probably at or near the point of neutrality during life prevents self-digestion, while the gross alteration of the CH in either direction after death (or after the cessation of oxidations in the tissues) induces autolysis. Bradley indeed suggests that many of the phenomena of autolysis during lifetime, such as atrophy, necrosis, involution, might be due to an increase in the CH in the tissues.
These facts agree with the suggestion of Fermi that in the living cell the proteins cannot be attacked by the digestive enzymes but relieves us of the necessity of making the monstrous assumption of a “living molecule” of proteins as distinct from a “dead” molecule. The difference between life and death is not one between living and dead molecules, but more likely between the excess of synthetic over hydrolytic processes.
In the second chapter we mentioned the interesting idea of Armstrong that when a synthesis is brought about by a digestive enzyme (e. g., maltase) not the original substrate is formed (e. g., maltose) but an isomer, in this case isomaltose; and this isomer is not attacked by the enzyme maltase. We thus get a rational understanding of the statement which Claude Bernard used to make but which remained at his time mysterious: la vie, c’est la création. During life, when nutritive material is abundant, through the reversible action of certain enzymes, synthetic compounds are formed from the building stones furnished by the blood. These synthetic isomers cannot be hydrolyzed by the enzymes by which they are formed and hence on account of the isomeric structure are immune against destruction. It is not impossible that the increase of the concentration of acid in the cells after death transforms the isomers into that form in which they can be digested by the enzymes contained in the cell. Another possibility is that the increase in digestibility brought about by an increase in CH in the cell is due to the hydrating effect of acids on proteins with a subsequent increase in digestibility. Whatever the answer may be, the work done since Claude Bernard has removed that cloud of obscurity which in his days surrounded the prevalence of synthetic action in the living and of disintegration in the dead tissues.
3. We have already referred to the connection between the lack of oxygen and the onset of autolysis and disintegration of tissues in the body. It is of interest that there are cells in which the disintegration under the influence of lack of oxygen is so rapid that it can be followed under the microscope. The writer has observed that certain cells undergo complete irreversible dissolution in a very short time under the influence of lack of oxygen, e. g., the first segmentation cells of the egg of a teleost fish Ctenolabrus.300
Fig. 48 |
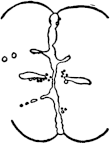
Fig. 49 | |
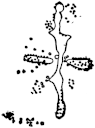
Fig. 50 |
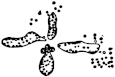
Fig. 51 |
When these eggs are deprived of oxygen at the time they reach the eight- or sixteen-cell stage, it can be noticed that the membranes of the blastomeres are transformed into small droplets within half an hour or more, according to the temperature. These droplets begin to flow together, forming larger drops. [Figures 48 to 51 show the successive stages of this process.] When the eggs are exposed to the air in time, segmentation can begin again; but if a slightly longer time is allowed to elapse, the process becomes irreversible and life becomes extinct. Such clear structural changes cannot often be observed in the eggs of other animals under the same conditions. Are these changes of structure (apparently liquefaction of solid elements) responsible for death under such conditions? In order to obtain an answer to this question, the writer investigated the effect of the lack of oxygen upon the heart-beat of the embryo of the same fish Ctenolabrus. This egg is perfectly transparent and the heart-beat can easily be watched. When these eggs are put into an Engelmann gas chamber and a current of pure hydrogen is sent through, the heart may cease to beat in fifteen or twenty minutes; it stops beating suddenly, before the number of heart-beats has diminished noticeably, and ceases beating before all the free oxygen can have had time to diffuse from the egg. In one case the heart beat ninety times per minute before the hydrogen was sent through; four minutes after the current of hydrogen had passed through the gas chamber, the rate of the heart-beat was eighty-seven per minute, three minutes later it was seventy-seven, and then the beats stopped suddenly. It is hard to believe that this cessation could have been caused by lack of energy. Hydrolytic processes alone could furnish sufficient energy to maintain the heart-beat for some time, even if all the oxygen had been used up. The suddenness of the standstill at a time when the rate had hardly diminished seems to be more easily explained by a sudden collapse of the machine; it might be that liquefaction or some other change of structure occurs in the heart or its ganglion cells, comparable to that which we mentioned before. In another fish Fundulus, where the cleavage cells undergo no visible changes in the case of lack of oxygen, the heart of the embryo can continue to beat for about twelve hours in a current of hydrogen. In this case the rate of the heart-beat sinks during the first hour in the hydrogen current from about one hundred to twenty or ten per minute; then it continues to beat at this rate for ten hours or more. In this case one might believe that during the period of steady diminution of the tension of oxygen in the heart (during the first hour), the heart-beat sinks steadily while it keeps up at a low but steady rate as long as the energy for the beat is supplied solely by hydrolytic processes; but there is certainly no change in the physical structure of the cells noticeable in Fundulus, and consequently there is no sudden standstill of the heart.
Budgett has observed that in many infusorians visible changes of structure occur in the case of lack of oxygen301; as a rule the membrane of the infusorian bursts or breaks at one point, whereby the liquid contents flow out. Hardesty and the writer found that Paramœcium becomes more strongly vacuolized when deprived of oxygen, and at last bursts. Amœbæ likewise become vacuolized and burst under these conditions. Budgett found that a number of poisons, such as potassium cyanide, morphine, quinine, antipyrine, nicotine, and atropine, produce structural changes of the same character as those described for lack of oxygen. As far as KCN is concerned, Schoenbein had already observed that it retards the oxidation in the tissues, and Claude Bernard and Geppert confirmed this observation. For the alkaloids, W. S. Young has shown that they are capable of retarding certain processes of autoxidation. This accounts for the fact that the above-mentioned poisons produce changes similar to those observed in the case of lack of oxygen.302
The phenomenon of rapid disintegration when deprived of oxygen (or in the presence of KCN) seems to be general as Child303 has shown in extensive experiments. Child has used it to show that younger animals disintegrate more rapidly than older or larger ones, and he uses this fact for a theory of senescence. He connects the more rapid disintegration of the young animal with a greater metabolism.304 Without wishing to doubt Child’s interesting observations the writer is not quite certain whether the more rapid disintegration of the younger forms is not a result of the fact that the walls of membranes in the young are softer than those of the older animals, and hence are more readily liquefied. Such a difference could be due to mere chemical constitution, e. g., the increase in Ca in the membrane with the increase in age. In old age in man the deposit of Ca in the blood-vessels is a frequent occurrence.
These facts may help us to understand the nature of death and dissolution of the body in higher animals. Death in these animals is due to cessation of oxidations, but the surprising fact is that if the oxidations have been interrupted but a few minutes life cannot be restored even by artificial respiration. This suggests that the respiratory ganglia in the medulla oblongata suffer an irreparable injury or an irreversible change (comparable to that just described in the cells of Ctenolabrus) even when deprived of oxygen for only a short time. As a consequence of the irreversible injury to the medulla the respirations cease permanently, the heart-beat must also cease, and gradually the different tissues must undergo the dissolution characteristic of death. While all the cells may be immortal they are only so in the presence of oxygen and the nutritive solution which the circulating blood furnishes. With the proper supply of oxygen cut off they can no longer live.
4. It is an unquestionable fact that each form has a quite definite duration of life. Unicellular organisms are immortal; but for the higher organisms with sexual reproduction the duration of life is almost as characteristic as any morphological peculiarity of a species. No species can exist unless the natural life of its individuals outlasts the period of sexual maturity; and unless the average duration of life is long enough to allow as many offspring to be brought into the world as will compensate for loss by death. The male bee dies before it is a year old, while the queen may live several years. In a certain species of butterflies, the Psychidæ, the parthenogenetic female lays its eggs while still in the cocoon and then dies without ever leaving the cocoon. The imago of the ephemera leaves the water in the evening, copulates, lets its eggs fall into the water, and is dead the next morning. The imperfect condition of their mandibles and alimentary canal makes them unfit for a long duration of life. The males of the rotifers which are devoid of organs of digestion live but a few days.
In the Zoölogical Station at Naples in 1906, an actinian, Actinia equina, was alive after having been in captivity fifteen years, and another one, Cerianthus, had been observed for twenty-four years. Korschelt kept earthworms for as long as ten years. The fresh-water mussel may reach the age of sixty years or more and crayfish may live for over twenty years. The differences in the duration of life of mammals are too well known to need discussion. If the cells and tissues are immortal, how does it happen that the duration of life is so characteristic for each species?
Metchnikoff305 has recently investigated the cause of “natural” death in the butterfly of the silkworm. The butterfly in this species lacks the organs necessary for taking up food, like the male rotifer or the ephemeridæ and hence is already, by this fact, condemned to a short life. Metchnikoff observed that these butterflies could live twenty-three days, but the average duration of life was 15.6 for the males and 16.6 days for the females; and that seventy-five per cent. of them contained no parasitic fauna or flora in their intestine. They lose considerably in weight during their lives, but the males still contain the fat body at the time of death. None of the changes accompanying “old age” in man are found in the tissues of these butterflies before death. Metchnikoff is inclined to believe that the animal is poisoned by some excretion retained in the body; namely, the urine, and that this poison also causes the symptoms of weakness which characterize the animal. He could prove the toxic character of their urine on other animals. This combined with starvation could sufficiently account for the short duration of life. The facts of the case show that it is due to an imperfection in the construction of the organism such as one would expect to find more or less in each animal if one discards the idea of purposefulness and divine wisdom in nature. Only a slight, perhaps an infinitesimal, fraction, of those species which are theoretically possible and which at one time or another arise can survive. Those which are durable show all transitions from the grossest disharmonies to an apparent lack of such shortcomings.
5. Minot had tried to prove that the death of metazoa is due to the greater differentiation and specialization of their tissues. Admitting the immortality of the unicellular organisms he argues that death is the price metazoa pay for the higher differentiation of their cells. This is of course purely metaphorical, but we may put it into a form in which it is capable of discussion in physicochemical terms, by assuming that death is a necessary stage in the development of a species. We are inclined, however, to follow Metchnikoff and suspect some poison accidentally or unavoidably formed in the body or some structural shortcoming as the cause of “natural” death.
An unusually favourable object for the study of natural death is the animal egg. The egg of the starfish Asterias forbesii when taken out of the body is usually immature, but in the spawning season it ripens in sea water. The writer306 observed that eggs which ripen disintegrate very rapidly when not fertilized. This disintegration may be due to a process of autolysis, which sets in only after the egg has extruded the two polar bodies. The writer found that by preventing the maturation of the egg either by withdrawing the oxygen or by replacing the alkaline sea water by a neutral solution or by exposing the eggs for some time to acidulated sea water, the disintegration could also be prevented.
Further experiments showed that even in the mature egg rapid disintegration could be prevented by lack of oxygen, and similar results were obtained by Mathews. When the egg is fertilized it does not disintegrate in the presence of oxygen but it gradually dies in the absence of oxygen. One is almost tempted to say that while the fertilized egg is a strict aërobe the mature unfertilized egg is an anaërobe. This latter statement, however, becomes doubtful since the presence of oxygen may help the disintegration only indirectly by allowing certain changes to go on in the egg. The important points for us are that duration of life in the mature unfertilized egg is comparatively short and that the entrance of a spermatozoön or the process of artificial parthenogenesis saves the life of the egg. Loeb and Lewis found that the life of the unfertilized sea-urchin egg (which is usually mature when removed from the ovaries) can also be prolonged when its oxidations are suppressed. The decay of the unfertilized egg seems to be due to the fact that those alterations in the cortical layer which underlie the membrane formation and which are responsible for the starting of development gradually take place. In such a condition the egg will die quickly unless deprived of oxygen. This view is supported by the observation of Wasteneys that unfertilized eggs of Arbacia show an increased rate of oxidations when allowed to remain for some time in sea water; we have seen in Chapter V that such an increase also accompanies artificial membrane formation.
6. If the limited duration of life of an organism is determined by one or more definite harmful chemical processes, we should expect to find a temperature coefficient for the duration of life or at least be able to show that if all other conditions are the same the duration of life is for a given organism a function of temperature. The writer307 investigated the duration of life of fertilized and unfertilized eggs of Strongylocentrotus purpuratus for the upper temperature limits.
TABLE XX
| Temperature | Duration of life of the eggs of S. purpuratus | |||
|---|---|---|---|---|
| Unfertilized | Fertilized | |||
| °C. | Minutes | Minutes | ||
| 32 | > 11⁄6 | 11⁄2 | ||
| < 2 | ||||
| 31 | > 21⁄4 | |||
| < 3 | ||||
| 30 | > 3 | > 4 | ||
| < 5 | < 5 | |||
| 29 | > 6 | |||
| < 7 | ||||
| 28 | > 8 | > 11 | ||
| < 10 | < 13 | |||
| 27 | about 18 | > 20 | ||
| < 22 | ||||
| 26 | > 35 | > 35 | ||
| < 40 | < 40 | |||
| 25 | > 76 | |||
| < 81 | ||||
| 24 | > 168 | > 192 | ||
| < 200 | < 209 | |||
| Hours | ||||
| 22 | 101⁄5 | |||
| 21 | 24 | |||
| 20 | 72 | |||
These observations show a very high temperature coefficient near the upper temperature limit, and this may account at least partly for the fact that in tropical seas the pelagic fauna is so much more limited than in polar seas.308 It is quite probable that the high temperature coefficients at the utmost limits are only an expression of the coagulation time of certain proteins.
P. and N. Rau state that in the cold certain butterflies live longer, and similar statements exist for the silkworm, but these statements are not based on exact experiments, which are difficult. Dr. Northrop and the writer have started experiments on the influence of temperature on the duration of life of the fly Drosophila. Newly hatched flies were kept first without food except water and air at 34°, 28°, 24°, 19°, 14°, and 10°, and second with cane sugar. The average duration of life was as follows:
| With water | days | With cane sugar | days | |
| 34° | ....... | 2.1 | .......... | 6.2 |
| 28° | ....... | 2.4 | .......... | 7.2 |
| 24° | ....... | 2.4 | .......... | 9.4 |
| 19° | ....... | 4.1 | .......... | 12.3 |
| 14° | ....... | 8.3 | ||
| 10° | ....... | 11.9 |
These experiments show that there is a definite temperature coefficient for the duration of life and that this coefficient is of the order of magnitude of that of a chemical reaction. We are continuing these experiments with animals in the presence of food. It should, however, be remembered that the fly carries with it a good deal of reserve material from the larval period. We have carried on simultaneously determinations of the temperature coefficients of the duration of the larval and pupa stage of these organisms at the same temperatures and found ratios similar to those given above for the duration of life with water only.
7. Metchnikoff309 has furnished the scientific facts for our understanding of senescence. He has demonstrated that the changes in tissue which give rise to phenomena of senility are due to the action of phagocytes. Thus the ganglion cells are altered (digested?) and destroyed by “neuronophags” and this is the main cause of mental senility. Definite phagocytic cells, the osteoclasts, slowly dissolve the bones (by the excretion of an acid?) and this leads to the known fragility of the bones in old age. The whiteness of the hair is due to the action of phagocytes; in the muscles in old age the contractile elements are destroyed by the sarcoplasm, and so on. It agrees with these facts that where organs are absorbed in the embryonic development of an animal, as e. g., the tail of the tadpole in metamorphosis, the phenomenon is due to a process of phagocytosis (and autolysis). We have mentioned the fact that in the larva of the Amblystoma the absorption of the gills and of the tail occurs simultaneously and that both must be caused by a constituent of the blood. Such a constituent may be responsible for phagocytosis and autolysis in the organs undergoing absorption. Metchnikoff calls attention to the fact that certain infectious diseases, e. g., syphilis, may bring about precocious senility; and he mentions also the senile appearance of young cretins which is due to the diseased thyroid. “It is no mere analogy to suppose that human senescence is the result of a slow but chronic poisoning of the organism.” He assumes that in man this poisoning is caused by the products of fermentation in the large intestine and that the micro-organisms responsible for these fermentations may therefore be regarded as the real cause of senility in man. Parrots which are long-lived birds have a limited flora of microbes in their intestine, while cows and horses which are short-lived in comparison with man possess an extraordinary richness of the intestinal flora. But, needless to say, it is not the quantity of microbes alone which is to be considered, the nature of the microbes is of much greater importance.
Certain plants like the Californian Sequoia gigantea may be considered as practically immortal since they live several thousands of years; other plants, the annuals, die after fructification. Metchnikoff quotes from a letter by de Vries that this author prolonged the life of Œnotheras by cutting the flowers before fertilization.
Under ordinary conditions the stem dies after producing from forty to fifty flowers, but if cutting be practised new flowers are produced until the winter cold intervenes. By cutting the stem sufficiently early the plants are induced to develop new buds at the base and these buds survive winter and resume growth in the following spring.
Metchnikoff suggests that it is a poison formed in the plant (in connection with fructification?) which kills the annuals, while it is not formed or is less harmful in the perennials. He compares the situation to the death of the lactic acid bacilli if the lactic acid is allowed to accumulate. This hypothesis is certainly worthy of consideration, and it is quite possible that in addition to structural shortcomings poisons formed by certain organs of the body as well as poisons formed by bacteria account for the phenomenon of death in metazoa.
Abraxas, 203, 238, 241
Acquired characters, inheritance of, 337 ff.
Actinia equina, 361
Adaptation, 12, 318 ff.;
to life in caves, 319 ff.;
fresh and salt water, 327 ff.;
poisons, 332 ff.;
temperature, 334 ff.;
caused by hormones, 342
Addison, W. H. F., 188
Agglutination, of corpuscles by sera, 67 ff.;
of sperm, 78, 82 ff.
Allolobophora terrestris, 46
Alpheus, 176
Alytes obstetricans, 337, 338
Amanita phalloides, 63
Amblystoma, 157, 368
Amelung, 184
Amphipyra, 283
Analogies between living and dead matter, 14 ff.
Anaphylaxis reaction, 61 ff.
Ancel, 158, 225 ff.
Antagonistic salt action. See Balanced salt solutions.
Antennularia antennina, 194, 196
Apes, blood relationship to man, 54, 56 ff.
Apolant, 45
Arbacia, 75 ff., 96, 99, 101, 111, 114, 150, 190 ff., 293 ff., 298, 299, 364
Arenicola, 277
Armstrong, E. F., 26, 28, 354
Arrhenius, S., 33 ff., 88, 290, 296
Arrhenoidy, 218, 225
Artificial parthenogenesis, 95 ff.;
in sea urchins, 95 ff.;
new method of, 98, 99;
by blood, 101 ff.;
by sperm extract, 103;
by acids, 105;
by mechanical agitation, 107;
in starfish, 110;
rôle of hypertonic solution, 112, 115, 116;
and oxidation, 116, 117, 118;
and permeability, 119 ff.;
in frogs, 124;
and determination of sex, 125
Artificial production of life, 38–39
Assimilation of CO2 without chlorophyll, 17 ff.
Asterias, 49, 81, 110, 363;
ochracea, 73 ff.;
capitata, 74
Asterina, 75, 81, 110
Astrospheres, 115 ff., 192
Auer, J., 315
Autolysis, 351 ff.
Avena, 263
B. coli communis, 36;
typhosus, 36;
fluorescens, 334
Bacteria, growth of, 15 ff., 29, 71 ff.;
specificity in, 41 ff.
“Bacterio-purpurin,” 41
Balanced salt solutions, 307–317;
theory of, 317;
and adaptation, 331 ff.
Balanus, 259
Baltzer, F., 215 ff.
Bancroft, F. W., 70, 125, 127, 264, 269 ff.
Bang, 63
Bardeen, C. R., 174 ff.
Barnacle, larvæ of, 313 ff.
Bataillon, 124
Bateson, W., 230, 240 ff., 338, 348
Batrachia, 338
Baur, E., 48, 246
Bayliss, 63
Becquerel, P., 36 ff.
Beggiatoa, 19
Beijerinck, M., 20
Berkeley, Lord, 111
Bernard, Claude, 2 ff., 26, 159, 350, 354, 355, 358
Berthelot, 290
Bertrand, G., 248 ff.
Beutner, R., 140
Bichat, 2, 349
Bickford, E. E., 169
Blaauw, H. A., 263
Blackman, F. F., 302
Blastomeres, 141 ff.
Blind animals, 319 ff.
Blood, transfusion of, 53 ff.
Blood relationship, established by transfusion, 53, 54 ff.;
precipitin reaction, 55 ff.;
anaphylaxis reaction, 61 ff.;
hemoglobin crystals, 64 ff.
Blood serum, precipitin reaction of, 54 ff.;
effect of, on unfertilized eggs, 101 ff., 124
Blowfly, heliotropism of larvæ of, 265 ff.
Bohn, G., 253, 264, 269
Bombinator igneus, 46
Bonellia, 215
Bonnet, 154, 161
Bordet, 54 ff., 60
Bouin, 158, 225 ff.
Boveri, Th., 8, 126, 128 ff., 134, 138 ff., 150 ff., 186 ff., 209 ff., 246
Brachystola, 199
Bradley, H. C., 27, 64, 353, 354
Brandt, 366
Braus, H., 147
Bridges, C. B., 208, 229, 231 ff.
Brown, A. P., 64 ff.
Bruchmann, H., 93
Bryophyllum calycinum, 153, 160 ff., 177
Buchner, 24
Budgett, 358
Buller, 93
Bunsen-Roscoe, law of, 11, 256 ff., 261, 263, 264
Burrows, 31
Campanularia, 178, 181
Cannon, W. B., 285
Carcinus mænas, 217
Cardamine pratensis, 90
Carrel, 31
Cassia bicapsularis, 37
Castle, W. E., 89 ff., 335
Caullery, M., 158, 180, 217
Cave animals, 319 ff.
Cell division, 15, 29, 129 ff.;
suppression of, 113 ff.
Cells, nutritive media of, 15 ff;
immortality of, 30 ff.;
migrating, 44;
mesenchyme, 51 ff., 130 ff., 147, 155 ff.
Cerianthus membranaceus, 171 ff., 188, 361
Chætopterus, 78 ff.
Chamberlain, M. M., 293, 297
Chapman, H. G., 60
Chemotropism of spermatozoa, 92 ff.
Chevreul, 289
Child, C. M., 7, 170, 177, 358
Chlamydomonas, 277
Chodat, R., 248
Chologaster, 320
Christen, 288
Chromosomes, rôle of, in sex determination, 198 ff.;
theory of Mendelian heredity, 233
Chun, 142
Ciona intestinalis, 89 ff., 212
Cladocera, 159
Clausen, H., 302
Clavellina, 181
Cohen, E., 292
Cohn, 41 ff.
Compton, 90
Conklin, E. G., 129, 134, 143, 145 ff.
Constancy of species, 40–43
Copernicus, 346
Corpus luteum, action of, 157–158
Correlation, 154, 167
Correns, C., 90 ff., 214
Cramer, 289
Crampton, H. E., 143, 225
Criodrilus lacuum, 219–220
Crossing over of chromosomes, 241 ff.
Crystals, differences between living organisms and, 14 ff.
Ctenolabrus, 355, 357, 359
Ctenophores, 142
Cuénot, L., 12, 324
Cullen, G. E., 24, 291
Cuma rathkii, 318
Cyanophyceæ, 287
Cytisus biflorus, 37
Cytoplasm of eggs as future embryo, 8, 9, 70, 126, 151 ff.
Dakin, 352
Dallinger, 334
Daphnia, 210, 262, 279, 280, 282, 306, 312
Darbishire, A. D., 347
Darwin, 90, 297, 346 ff.
Darwinian theory, 5 ff.
Davenport, C. B., 244, 335
Death, 349 ff.;
natural, cause of, 364, 369
Decidua formation induced by corpus luteum, 157–158
Delage, Y., 107, 110, 111, 123, 126, 186
de la Rive, 24
de Meyer, J., 127
Dendrostoma, 101
Dentalium, 144
Design, 4, 5
Determination of sex, in bees, 208 ff.;
in phylloxerans, 210;
in Bonellia, 215
Development of egg, 127 ff.
de Vries, H., 6, 42, 154, 161, 347, 369
Dewitz, 93
Dieudonné, C., 334, 337
“Directive force,” 2
Disharmonies, 7
Divisibility of living matter, limits of, 148–151
Dominance, 230
Doncaster, L., 203
Dorfmeister, 303
Driesch, H., 4 ff., 128, 133, 136, 138 ff., 147, 150, 169 ff., 180 ff., 184 ff.
Drosophila ampelophila, 204 ff., 237, 243, 322, 347, 366
Duclaux, E., 288, 289
v. Dungern, 80
Duration of life, 360 ff.
Durham, 249
Dutrochet, 154
Dzierzon, 208
Ectoderm formation, 130 ff.
Egg, as the future embryo, 8, 9, 70, 126, 151;
artificial parthenogenesis of, 95 ff.;
organisms from, 128 ff.;
determining unity of organism, 151–152;
chromosomes in, 198 ff.
Egg structure, 129 ff.;
influence of centrifugal force on, 135;
and regulation, 139, 140, 141;
and fluidity of protoplasm, 141
Ehrlich, 45, 322, 332 ff., 341;
side-chain theory of, 88, 188
Eigenmann, 320, 323 ff.
Electromotive forces, origin in living organs, 140
Engelmann, 357
Engler, 24
Entelechy, 4, 170, 182
Environment, influence of, 286 ff.;
temperature, 288 ff., 344 ff.;
salinity, 306;
adaptation to, 319
Enzyme action, 23 ff., 297, 302
Ernst, A., 21
Eternity of life, 34 ff., 360
Eudendrium, 260, 261, 269, 277, 278, 326
Eudorina, 277
Euglena, 264, 269, 272, 277
Euler, H., 21
Evolution, 346 ff.;
and mutation, 348
Ewald, W. F., 261 ff., 269, 280, 301
Farmer, J. B., 347
Fermi, 350, 354
Fertilization, heterogeneous, 48 ff., 51, 73 ff.;
specificity in, 71 ff.;
and oxidation, 117 ff.;
and permeability, 119 ff.
“Fertilizin,” 84, 87 ff., 93
Fischel, 187
Fischer, 303 ff.
Fish, 55
Fitness of environment, 317
Fitzgerald, J. G., 63
Flow of substances and regeneration in Bryophyllum, 161 ff.
Fluctuating variations, 6, 297 ff., 346 ff.
Folin, 22
Food, influence on polymorphism in wasps, 222 ff.
Food castration, 224;
influence on sexual cycle in rotifers, 224;
on metamorphosis in tadpoles, 155
Ford, 63
Forssmann, 63
Frédéricq, 351
Free-martin, cause of sterility, 218–219
Friedenthal, H., 53 ff., 60
Frisch, K., 278, 279
Fröschel, P., 263
Fuchs, H. M., 90
Fucus, 123
Fundulus heteroclitus, 51, 116, 147, 300, 301, 302, 307 ff., 321 ff., 328 ff., 335, 337, 357 ff.
Galileo, 346
Galvanotropism, 11, 270 ff., 319
Gay, F. P., 62 ff.
Generation, spontaneous, 14 ff., 34
Genes, 4 ff., 152, 319
Genus and species, chemical basis of, 40 ff.
Geppert, 358
Germination in seeds, 35 ff.
Giard, 180, 216 ff.
Godlewski, E., 48, 75, 78, 120, 126, 169
Godlewski, E., Sr., 18
Goebel, K., 154, 161
Goldfarb, A. J., 326
Goldschmidt, R., 220 ff.
Goodale, H. D., 218
Gortner, R., 249
Graber, V., 256, 276
Grafting, heteroplastic, in animals, 46;
in plants, 47
Gravitation, influence on organ formation in Antennularia, 194 ff.;
on the egg of the frog, 141
Gray, J., 122
Gregory, 243
Groom, T. T., 280
Growth, termination of, 184;
influence of cell size, 187
Gudernatsch, J. F., 155, 255, 342
Guyer, 124
Gynandromorphism, 209
Haeckel, 346
Half-embryos and whole embryos, 141, 142
Hammond, J. H., Jr., 269
Harden, 16
Hardesty, 358
Harmonious character of organism, 5, 6, 318 ff., 341 ff.
Harrison, 31
Hartley, 111
Healing of wound, 187
Hektoen, 66
Heliotropism, 11 ff., 257 ff., 318;
heredity of, 250 ff.;
change of, 279, 280 ff.;
and adaptation, 318
Helmholtz, 34
Hemoglobins, crystallographic measurements of, 64 ff.
Henderson, L., 317
Henking, 198 ff.
Herbst, C., 97, 147, 193, 306, 310
Heredity, of genus and species, 40 ff., 70, 151, 152;
Mendelian, 70, 151 ff., 229 ff., 348;
of sex, 198;
sex-linked, 203 ff., 238 ff.;
and evolution, 348
Herlant, M., 78 ff., 115 ff.
Hermaphroditism, 89 ff., 212 ff., 216, 219 ff.
See also Inhibition and Regeneration.
Hertwig, O., 97, 123, 292
Hertwig, R., 95, 97
Hess, C., 278
Heterogeneous hybrids, purely maternal, 49, 50
Heterogeneous transplantation, Murphy’s experiments on, 44 ff.;
limitation of, 46
Heteromorphosis, 155, 193–196
Hill, C., 25
Hippiscus, 199
Holmes, S. J., 269
Hoppe-Seyler, 351
Hormones, 145, 155, 181, 219;
and Mendelian heredity, 245 ff., 348;
and adaptation, 342.
See also Organ-forming substances.
Huxley, 346
Hybridization, heterogeneous, in sea urchins, 48 ff., 73 ff.;
in fishes, 51;
in plants (Mendel’s), 230 ff.
Hydrolytic enzymes, action of, 24;
reversible action of, 24 ff.
Hypertonic solution, 99, 111 ff.
Imitation of cell structures by colloids, 39
Immortality, of cancer cells, 30;
of somatic cells, 30 ff.;
of life in general, 34 ff.
Inheritance, of colour-blindness, 203, 204, 205;
of eye pigment in Drosophila, 204 ff.;
of pigments, 248 ff.;
of acquired characters, 337 ff.
Inhibition of regeneration in Bryophyllum, 162 ff.
Inhibition of sexual characters of opposite sex, in pheasants, 218;
lack of in hermaphrodites, 219;
in Bonellia, 226
Instincts, 10 ff., 253 ff.;
sexual, 198 ff.
Intersexualism, 221
Intestine, formation of, 130 ff.
Isoagglutinins, 66 ff., 92
Isolation of blastomeres, 136 ff.
Jacoby, 352
Janda, V., 219 ff.
Jansky, 67
Janssens, 242
Jennings, H. S., 264 ff.
Jensen, 45
Joest, 46
Johannsen, W., 42, 333
Jones, 352
Jost, 90
Kammerer, P., 325, 337 ff.
Kanitz, A., 290, 292, 296
Kastle, J. H., 26 ff.
Kellogg, V. L., 279
Kelvin, 34
King, W. O. R., 50, 247
Klug, 351
v. Knaffl, E., 106
Knowlton, E. P., 292
Kofoid, C. A., 143
v. Körösy, 300
Korschelt, 361
Krakatau, 21
Kraus, 54 ff.
Krogh, 292
Kryž, F., 335
Kupelwieser, H., 75
Lack of oxygen, influence on disintegration of tissue, 355 ff.
Ladoff, S., 224
Lamarck, 6
Laminaria, 165
Landois, L., 53
Landsteiner, 66
Lanice, 143 ff.
Lankester, E. R., 41
Leathes, J. B., 63
Leucæna leucocephala, 37
Levene, 351, 352
Lewis, 183, 344, 364
Light, influence on organ formation, in cave animals, 319 ff.;
in Proteus, 325;
in Eudendrium, 326.
See also Heliotropism.
Lillie, F. R., 80, 82 ff., 87 ff., 93, 134, 191, 218, 292
Lillie, R. S., 101, 107, 110, 120 ff.
Lipase, synthetic action of, 26
Living and dead matter, specific differences between, 14 ff.
Lloyd, D. J., 111
Localization of Mendelian characters in individual chromosomes, 243, 244
Loeb, Leo, 30 ff., 45, 157, 170, 187 ff., 342
Loevenhart, A. S., 26 ff.
Lumbricus rubellus, 46
Lychnis dioica, 217
Lycopodium, 93
Lygæus, 201
Lymantria dispar, 220
Lymnæus, 142
Lymphocytes, rôle of, 45 ff.
Lyon, E. P., 134 ff.
Macfadyen, A., 36
Maeterlinck, 255
Magnus, W., 60
Maltase, synthetic action of, 25
Marchal, P., 222 ff., 254
Margelis, 192
Mass of chromatin and of cytoplasm, 186
Mast, 269, 277
Mathews, A. P., 107, 363
Matthaei, G. L. C., 302
Maxwell, S. S., 270, 274, 277
McClendon, J. F., 122, 322
McClung, C. E., 68, 198 ff., 237
Megusar, 340
Meignon, 217
Meisenheimer, 225
Meltzer, S.J., 315
Membrane formation, 86 ff.;
artificial, 98 ff.
Mendel, G., 23, 229 ff.
Mendelian characters, and evolution, 70, 348;
and internal secretions, 243, 348;
and enzymes, 247, 248, 249
Mendelian, factors of heredity, 4 ff., 68, 151 ff.;
mutation, 66;
dominant, 90;
segregation, 229 ff.
See also Non-Mendelian inheritance.
Mendelian heredity, mechanism of, 229 ff.;
and chromosomes, 233 ff.;
and hormones, 245 ff., 348;
and enzymes, 247 ff.
Menidia, 51, 321, 323
Merogony, 120, 126, 186
Merrifield, 303
Mesenchyme formation, 130 ff.
Metamorphosis of tadpoles induced by thyroid, 155, 156
Metchnikoff, 361 ff., 367 ff.
Michaelis, L., 62, 317
Micrococcus prodigiosus, 334
Micromeres, 132 ff.
Minot, 362
Moenkhaus, W. J., 51, 344
Molisch, 20
Montgomery, 199, 234
Moore, A. R., 50, 247 ff., 280
Morgan, T. H., 46, 68, 89 ff., 95, 116, 126, 134, 141 ff., 173, 175, 184, 204 ff., 229 ff., 241 ff., 244, 347
Morse, M., 156, 353
Morton, J. J., 44
Moss, W. L., 67
Muller, H. J., 229, 231 ff.
Murphy, J. B., 44 ff.
Mutation, 6, 42, 243; and evolution, 347, 348
Myers, 55
Nathanson, 19
Natural death, 361 ff.
Neilson, 110
Newman, 344
Newton’s Law, 253
Nitrifying bacteria, 16 ff.
Non-Mendelian inheritance, genus and species characters, 70, 151, 251;
rate of segmentation, 246;
first development, 247
Northrop, 366
Nostocaceæ, 21
Nussbaum, M., 149
Nuttall, G. H. F., 56 ff.
Ocneria dispar, 225
Œnotherus, 369
Onslow, H., 249
Organ-forming substances or hormones in regeneration, 154 ff.;
causing metamorphosis in tadpoles, 155–157;
decidua formation, 158;
development of milk glands, 158;
Sachs’s theory of, 159
Organisms from eggs, 128 ff.
Origin of life, 14 ff., 33 ff.
Osborne, 23
Osterhout, W. J. V., 312
Ostwald, Wo., 29, 305, 312
Oudemans, 225
Overton, 123
Palæmon, 193
Palæmonetas, 193;
geotropism of, 270
Palinurus, 193
Pandorina, 277
Parker, G. H., 264, 269
Parthenogenesis, artificial, 95 ff.;
“spontaneous,” 107
Pasteur, 14 ff., 24, 33, 38
Patten, B., 264 ff
Pauli, W., 289
Pavy, 350
Payne, F., 322
Pearl, R., 203, 244
Penicillium, 289
Pennaria, 192
Pepsin, synthetic action of, 28, 62, 63
Pfeffer, 92 ff.
Phagocytosis, 367
Planaria, 173 ff., 177
Planorbis, 142
Plants, heteroplastic grafting in, 47 ff.;
regeneration in, 160 ff.
Polygordius, 280
Polymorphism, 222
Porthesia, 256, 280 ff.
Preadaptation, 12, 324
Precipitin reaction, 54 ff.
Preformation of organism in egg, 128 ff., 142–145
Presence and absence theory, 230 ff.
Primula, 243
Proteins, specific reactions of, 54 ff.;
and species specificity, 68;
and evolution, 70, 348
Protenor, 200 ff., 208
Proteus, 325 ff.
Przibram, H., 176
Pure lines, 333, 334
Pycnopodia spuria, 74
Pyrrhocoris, 198
Radiation pressure, rôle in transmission of spores through interstellar space, 34 ff.
Rana, esculenta, 46;
palustris, 46;
virescens, 46
Rate of segmentation, a non-Mendelian hereditary character, 246
Rau, 366
Reaction, tropistic, 11 ff., 92 ff., 147, 178, 187, 255 ff.;
precipitin, 54 ff.;
anaphylaxis, 61 ff.
Regeneration, 9 ff., 153 ff.;
in plants, 160 ff.;
in Bryophyllum, 161–167;
in animals, 167 ff.;
in Tubularia, 167–170;
in Cerianthus, 171 ff.;
in Planarians, 173–176;
in Alpheus, 176;
and autolysis, 178–181;
of lens, 182, 183;
external influences on, 192 ff.;
of gonads in hermaphrodites, 219
Regulation, 139, 140, 141;
in regeneration, see Regeneration.
Reichert, E. T., 64 ff.
Reseda, 90
Resistance of spores, 36;
seeds, 36 ff.
Reversibility of development, in Campanularia, 178 ff.;
in Ascidians, 180;
in egg, 189 ff.;
in Antennularia, 194
Rhabdonema nigrovenosum, 213
Richet, C., 61
Richter, 34
Ringer solution, 99
Robertson, T. B., 28 ff., 62 ff., 104, 311
Roentgen rays, 45
Roscoe, see Bunsen
Rotifers, determination of sexual cycle by food, 224
Roux, W., 141 ff.
Saccharomyces, 36;
cerevisiæ, 60
Sacculina, 216 ff.
Sachs, 88
v. Sachs, J., 145, 154 ff., 159, 161, 184
Salamandra maculosa, 339
Salkowski, 352
Salts required for life, 306 ff.
Sansum, W. D., 64
Schizophyceæ, 21
Schleip, W., 213
Schoenbein, 358
Schottelius, 334, 337
Schroeder, 14, 33
Schultze, O., 141
Schütze, 55
Schwann, 33
Schwarzschild, 34
Secretions, internal, 145, 155, 157
Self-digestion, 350 ff.
Self-sterility, 89 ff.
Senescence, 367
Sequoia, 31, 368
Setchell, W. A., 165, 287
Sex, of parthenogenetic frogs, 125;
of twins, 211
Sex chromosome, 199 ff.
Sex determination, cytological basis of, 198 ff.;
physiological basis of, 214 ff.
Sexual characters, 198 ff.
Shibata, 93
Shull, A. F., 214, 224
Sicyonia, 193
Side-chain theory, 88, 188
Smith, Geoffrey, 159, 217
Smith, Graham, 58
Spain, K. C., 188
Spallanzani, 33
Species, chemical basis of, 40 ff.;
specificity of, 41 ff.;
incompatibility of, not closely related, 44 ff.
Species specificity, determined by proteins, 63, 68, 348;
apparently not by nucleins, 69
Specificity, of grafted tissues, 47;
of spermatozoa, 48;
of blood sera, 53 ff.;
in fertilization, 71 ff.;
of activation of sperm by eggs, 80 ff.
Spelerpes, 320
Spermatozoa, fertilization of eggs by, 72 ff.;
activation by eggs of, 80 ff.;
agglutination of, 82 ff.;
cluster formation of, 83;
chemotropism of, 92 ff.;
cultivating of, 126 ff.;
chromosomes of, 198 ff.
Spirographis, 260
Spondylomorum, 277
Spontaneous generation, 33, 38
Spooner, G. B., 134
Standfuss, 303
Staphylococcus pyogenes aureus, 36
Steffenhagen, K., 55
Steinach, E., 225 ff., 254, 343
Stereotropism, 178, 187, 283
Stevens, Miss, 68, 199
Stimulus, 196
Stockard, 322, 340
Strassburger, 260
Streaming as means of egg differentiation, 145, 146
Strongylocentrotus franciscanus, 50, 52, 75, 81 ff., 103, 247
Strongylocentrotus lividus, 129
Strongylocentrotus purpuratus, 52, 73 ff., 81 ff., 94, 98 ff., 103, 108, 109, 111 ff., 137, 191, 246 ff., 293 ff., 364;
larvæ of, 49 ff.
Sturtevant, A. H., 229 ff.
Styela, 146
Sulphur bacteria, 19 ff.
Supergenes, 5, 9, 136, 319
Sutton, W. S., 68, 233 ff.
Synthesis of living matter, by micro-organisms, 15 ff.;
by enzymes, 24 ff.
Synthetic action of enzymes, 23 ff., 38
Tænia, 212
Talbot, 262
Tammann, 291
Tanaka, 243
Taylor, A. E., 27, 69 ff.
Tchistowitch, 54 ff.
Teleost fish, crosses of, 6 ff., 345
Temperature, effect on heliotropism, 280;
upper limit for organisms, 287 ff.;
effect on life, 288 ff.;
on butterflies, 303 ff.;
adaptation to, 334 ff.
Temperature coefficient, 290 ff., 305;
for enzyme, 291;
for development, 292 ff.;
for oxidations, 295;
and fluctuating variation, 296 ff.;
for heart-beat, 300 ff.;
for duration of life, 366
Thatcher, Miss, 181
Thyroid inducing metamorphosis in tadpoles, 155, 156
Tichomiroff, 95
Tissue culture of spermatozoa, 127
Tissues, transplantation of, 30 ff., 44 ff.;
cultivation of, 31 ff.;
specificity of, 44 ff.
Torrey, H. B., 264, 269
Tower, 348
Transfusion of blood, 53
Transplantation, of tissues, 30 ff., 44 ff.;
of cancers, 45;
of anlagen, 148;
of eye of salamander, 157;
of testes, 226;
of ovaries, 227
Traube, 28
Treub, 21
Trial and error, 268, 270
Trifolium arvense, 37
Tropisms, 11 ff., 92 ff., 147, 178, 187, 253 ff.;
and instincts, 253;
theory of, 257 ff.
Tropisms, in embryonic development, 147;
of cave animals, 324
Trypanosomes, 332 ff.
Trypsin, synthetic action of, 27
Tuber brumale, 60
Tubularia crocea, 171
Tubularia mesembryanthemum, 167, 169, 192
Twins, origin of, 136 ff.;
sex of, 211
Tyndall, 33
Typhlogobius, 320
Typhlomolge, 320
Typhlotriton, 320, 323
Tyrosinase, 249, 250
Tyrosine, 249, 250
v. Uexküll, J., 4 ff., 128, 139
Uhlenhuth, E., 157, 183, 187
Uhlenhuth, P., 55, 58, 66, 322
Underhill, F. P., 23
Vanessa, prorsa, 303;
levana, 303
Vaney, 217
Van Slyke, D. D., 22, 24, 291
van’t Hoff, 24 ff., 290, 292, 296
Variation, 6, 297 ff., 346–348
Vitzou, 159
Volvox, 280
Walcott, 42, 61
Warburg, O., 117 ff.
Warming, 41
Wasps, polymorphism in, 222–224;
sex determination, 255 ff.
Wassermann, 55
Wasteneys, H., 29, 82, 87, 112, 113, 117, 191, 277, 293, 295, 335, 364
Weiggert, 188
Weismann, 7, 30, 303
Wells, H. G., 62, 69
Welsh, D. A., 60
Werner, F., 340
Wheeler, W. M., 43
White, J., 36
Whitney, D. D., 224
Wilson, E. B., 68, 143, 199 ff.
Winkler, 47
Winogradsky, S., 16 ff., 42
Wolf, G., 182, 187
Yeast cells, cultivation of, 15 ff.
Young, 16, 358
A Selection from the Catalogue of
G. P. PUTNAM’S SONS
Complete Catalogues sent on application
1.—The Study of Man. By Professor A. C. Haddon, M.A., D.Sc., M.R.I.A. Fully illustrated. 8o.
“A timely and useful volume. . . . The author wields a pleasing pen and knows how to make the subject attractive. . . . The work is calculated to spread among its readers an attraction to the science of anthropology. The author’s observations are exceedingly genuine and his descriptions are vivid.”—London Athenæum.
2.—The Groundwork of Science. A Study of Epistemology. By St. George Mivart, F.R.S. 8o.
“The book is cleverly written and is one of the best works of its kind ever put before the public. It will be interesting to all readers, and especially to those interested in the study of science.”—New Haven Leader.
3.—Rivers of North America. A Reading Lesson for Students of Geography and Geology. By Israel C. Russell, Professor of Geology, University of Michigan, author of “Lakes of North America,” “Glaciers of North America,” “Volcanoes of North America,” etc. Fully illustrated. 8o.
“There has not been in the last few years until the present book any authoritative, broad résumé on the subject, modified and deepened as it has been by modern research and reflection, which is couched in language suitable for the multitude. . . . The text is as entertaining as it is instructive.”—Boston Transcript.
4.—Earth Sculpture; or, The Origin of Land-Forms. By James Geikie, LL.D., D.C.L., F.R.S., etc., Murchison Professor of Geology and Mineralogy in the University of Edinburgh; author of “The Great Ice Age,” etc. Fully illustrated. 8o.
“This volume is the best popular and yet scientific treatment we know of of the origin and development of land-forms, and we immediately adopted it as the best available text-book for a college course in physiography. . . . The book is full of life and vigor, and shows the sympathetic touch of a man deeply in love with nature.”—Science.
5.—Volcanoes: Their Structure and Significance. By T. G. Bonney, F.R.S., University College, London. Fully illustrated. 8o. Revised and Enlarged Edition. Illustrated.
“It is not only a fine piece of work from a scientific point of view, but it is uncommonly attractive to the general reader, and is likely to have a larger sale than most books of its class.”—Springfield Republican.
6.—Bacteria: Especially as they are related to the economy of nature, to industrial processes, and to the public health. By George Newman, M.D., F.R.S. (Edin.), D.P.H. (Camb.), etc., Demonstrator of Bacteriology in King’s College, London. With 24 micro-photographs of actual organisms and over 70 other illustrations. 8o.
“Dr. Newman’s discussions of bacteria and disease, of immunity, of antitoxins, and of methods of disinfection, are illuminating, and are to be commended to all seeking information on these points. Any discussion of bacteria will seem technical to the uninitiated, but all such will find in this book popular treatment and scientific accuracy happily combined.”—The Dial.
7.—A Book of Whales. By F. E. Beddard, M.A., F.R.S. Illustrated. 8o.
“Mr. Beddard has done well to devote a whole volume to whales. They are worthy of the biographer who has now well grouped and described these creatures. The general reader will not find the volume too technical, nor has the author failed in his attempt to produce a book that shall be acceptable to the zoölogist and the naturalist.”—N. Y. Times.
8.—Comparative Physiology of the Brain and Comparative Psychology. With special reference to the Invertebrates. By Jacques Loeb, M.D., Professor of Physiology in the University of Chicago. Illustrated. 8o.
“No student of this most interesting phase of the problems of life can afford to remain in ignorance of the wide range of facts and the suggestive series of interpretations which Professor Loeb has brought together in this volume.”—Joseph Jastrow, in the Chicago Dial.
9.—The Stars. By Professor Simon Newcomb, U.S.N., Nautical Almanac Office, and Johns Hopkins University. 8o. Illustrated.
“The work is a thoroughly scientific treatise on stars. The name of the author is sufficient guarantee of scholarly and accurate work.”—Scientific American.
10.—The Basis of Social Relations. A Study in Ethnic Psychology. By Daniel G. Brinton, A.M., M.D., LL.D., Sc.D., Late Professor of American Archaeology and Linguistics in the University of Pennsylvania; Author of “History of Primitive Religions,” “Races and Peoples,” “The American Race,” etc. Edited by Livingston Farrand, Columbia University. 8o.
“Professor Brinton his shown in this volume an intimate and appreciative knowledge of all the important anthropological theories. No one seems to have been better acquainted with the very great body of facts represented by these sciences.”—Am. Journal of Sociology.
11.—Experiments on Animals. By Stephen Paget. With an Introduction by Lord Lister. Illustrated. 8o.
“To a large class of readers this presentation will be attractive, since it gives to them in a nut-shell the meat of a hundred scientific dissertations in current periodical literature. The volume has the authoritative sanction of Lord Lister.”—Boston Transcript.
12.—Infection and Immunity. With Special Reference to the Prevention of Infectious Diseases. By George M. Sternberg, M.D., LL.D., Surgeon-General U. S. Army (Retired). Illustrated. 8o.
“A distinct public service by an eminent authority. This admirable little work should be a part of the prescribed reading of the head of every institution in which children of youths are gathered. Conspicuously useful.”—N. Y. Times.
13.—Fatigue. By A. Mosso, Professor of Physiology in the University of Turin. Translated by Margaret Drummond, M.A., and W. B. Drummond, M.B., C.M., F.R.C.P.E.; extra Physician, Royal Hospital for Sick Children, Edinburgh; Author of “The Child, His Nature and Nurture.” Illustrated. 8o.
“A book for the student and for the instructor, full of interest, also for the intelligent general reader. The subject constitutes one of the most fascinating chapters in the history of medical science and of philosophical research.”—Yorkshire Post.
14.—Earthquakes. In the Light of the New Seismology. By Clarence E. Dutton, Major, U. S. A. Illustrated. 8o.
“The book summarizes the results of the men who have accomplished the great things in their pursuit of seismological knowledge. It is abundantly illustrated and it fills a place unique in the literature of modern science.”—Chicago Tribune.
15.—The Nature of Man. Studies in Optimistic Philosophy. By Élie Metchnikoff, Professor at the Pasteur Institute. Translation and introduction by P. Chambers Mitchell, M.A., D.Sc. Oxon. Illustrated. 8o.
“A book to be set side by side with Huxley’s Essays, whose spirit it carries a step further on the long road towards its goal.”—Mail and Express.
16.—The Hygiene of Nerves and Mind in Health and Disease. By August Forel, M.D., formerly Professor of Psychiatry in the University of Zurich. Authorized Translation. 8o.
A comprehensive and concise summary of the results of science in its chosen field. Its authorship is a guarantee that the statements made are authoritative as far as the statement of an individual can be so regarded.
17.—The Prolongation of Life. Optimistic Essays. By Élie Metchnikoff, Sub-Director of the Pasteur Institute. Author of “The Nature of Man,” etc. 8o. Illustrated. Net, $2.50. Popular Edition. With an introduction by Prof. Charles S. Minot.
In his new work Professor Metchnikoff expounds at greater length, in the light of additional knowledge gained in the last few years, his main thesis that human life is not only unnaturally short but unnaturally burdened with physical and mental disabilities. He analyzes the causes of these disharmonies and explains his reasons for hoping that they may be counteracted by a rational hygiene.
18.—The Solar System. A Study of Recent Observations. By Prof. Charles Lane Poor, Professor of Astronomy in Columbia University. 8o. Illustrated.
The subject is presented in untechnical language and without the use of mathematics. Professor Poor shows by what steps the precise knowledge of to-day has been reached and explains the marvellous results of modern methods and modern observations.
19.—Heredity. By J. Arthur Thomson, M.A., Professor of Natural History in the University of Aberdeen; Author of “The Science of Life,” etc. 8o. Illustrated.
The aim of this work is to expound, in a simple manner, the facts of heredity and inheritance as at present known, the general conclusions which have been securely established, and the more important theories which have been formulated.
20.—Climate—Considered Especially in Relation to Man. By Robert Decourcy Ward, Assistant Professor of Climatology in Harvard University. 8o. Illustrated.
This volume is intended for persons who have not had special training in the technicalities of climatology. Climate covers a wholly different field from that included in the meteorological text-books. It handles broad questions of climate in a way which has not been attempted in a single volume. The needs of the teacher and student have been kept constantly in mind.
21.—Age, Growth, and Death. By Charles S. Minot, James Stillman Professor of Comparative Anatomy in Harvard University, President of the Boston Society of Natural History, and Author of “Human Embryology,” “A Laboratory Text-book of Embryology,” etc. 8o. Illustrated.
This volume deals with some of the fundamental problems of biology, and presents a series of views (the results of nearly thirty years of study), which the author has correlated for the first time in systematic form.
22.—The Interpretation of Nature. By C. Lloyd Morgan, LL.D., F.R.S. Crown 8vo.
Dr. Morgan seeks to prove that a belief in purpose as the causal reality of which nature is an expression is not inconsistent with a full and whole-hearted acceptance of the explanations of naturalism.
23.—Mosquito Life. The Habits and Life Cycles of the Known Mosquitoes of the United States; Methods for their Control; and Keys for Easy Identification of the Species in their Various Stages. An account based on the investigation of the late James William Dupree, Surgeon-General of Louisiana, and upon the original observations by the Writer. By Evelyn Groesbeeck Mitchell, A.B., M.S. With 64 Illustrations. 8o.
This volume has been designed to meet the demand of the constantly increasing number of students for a work presenting in compact form the essential facts so far made known by scientific investigation in regard to the different phases of this, as is now conceded, important and highly interesting subject. While aiming to keep within reasonable bounds, that it may be used for work in the field and in the laboratory, no portion of the work has been slighted, or fundamental information omitted, in the endeavor to carry this plan into effect.
24.—Thinking, Feeling, Doing. An Introduction to Mental Science. By E. W. Scripture, Ph.D., M.D., Assistant Neurologist Columbia University, formerly Director of the Psychological Laboratory at Yale University. 189 Illustrations. 2d Edition, Revised and Enlarged. 8o.
“The chapters on Time and Action, Reaction Time, Thinking Time, Rhythmic Action, and Power and Will are most interesting. This book should be carefully read by every one who desires to be familiar with the advances made in the study of the mind, which advances, in the last twenty-five years, have been quite as striking and epoch-making as the strides made in the more material lines of knowledge.”—Jour. Amer. Med. Ass’n., Feb. 22, 1908.
25.—The World’s Gold. By L. de Launay, Professor at the École Supérieure des Mines. Translated by Orlando Cyprian Williams. With an Introduction by Charles A. Conant, author of “History of Modern Banks of Issue,” etc. 8o.
M. de Launay is a professor of considerable repute not only in France, but among scientists throughout the world. In this work he traces the various uses and phases of gold; first its geology; secondly, its extraction; thirdly, its economic value.
26.—The Interpretation of Radium. By Frederick Soddy, Lecturer in Physical Chemistry in the University of Glasgow. Third Edition, rewritten, with data brought down to 1912. 8o With 33 Diagrams and Illustrations.
As the application of the present-day interpretation of Radium (that it is an element undergoing spontaneous disintegration) is not confined to the physical sciences, but has a wide and general bearing upon our whole outlook on Nature, Mr. Soddy has presented the subject in non-technical language, so that the ideas involved are within reach of the lay reader. No effort has been spared to get to the root of the matter and to secure accuracy, so that the book should prove serviceable to other fields of science and investigation, as well as to the general public.
27.—The Social Evil. With Special Reference to Conditions Existing in the City of New York. A Report Prepared in 1902 under the Direction of the Committee of Fifteen. Second Edition, Revised, with New Material Covering the Years 1902–1911. Edited by Edwin R. A. Seligman, LL.D., McVickar Professor of Political Economy in Columbia University. 8vo.
A study that is far from being of merely local interest and application. The problem is considered in all its aspects and, for this purpose, reference has been made to conditions prevailing in other communities and to the different attempts foreign cities have made to regulate vice.
28.—Microbes and Toxins. By Etienne Burnet, of the Pasteur Institute, Paris. With an Introduction by Élie Metchnikoff, Sub-Director of the Pasteur Institute, Paris. With about 71 Illustrations.
A well-known English authority said in recommending the volume: “Incomparably the best book there is on this tremendously important subject. In fact, I am assured that nothing exists which gives anything like so full a study of microbiology.” In the volume are considered the general functions of microbes, the microbes of the human system, the form and structure of microbes, the physiology of microbes, the pathogenic protozoa, toxins, tuberculin and mallein, immunity, applications of bacteriology, vaccines and serums, chemical remedies, etc.
29.—Problems of Life and Reproduction. By Marcus Hartog, D. Sc., Professor of Zoology in University College, Cork. 8vo.
The author uses all the legitimate arms of scientific controversy in assailing certain views that have been widely pressed on the general public with an assurance that must have given many the impression that they were protected by the universal consensus of biologists. Among the subjects considered are: “The Cellular Pedigree and the Problem of Heredity”; “The Relation of Brood-Formation to Ordinary Cell-Division”; “The New Force, Mitokinetism”; “Nuclear Reduction and the Function of Chroism”; “Fertilization”; “The Transmission of Acquired Characters”; “Mechanism and Life”; “The Biological Writings of Samuel Butler”; “Interpolation in Memory”; “The Teaching of Nature Study.”
30.—Problems of the Sexes. By Jean Finot, Author of “The Science of Happiness,” etc. Translated under authority by Mary J. Safford. 8vo.
A masterly presentation of the attitude of the ages toward women and an eloquent plea for her further enfranchisement from imposed and unnatural limitations. The range of scholarship that has been enlisted in the writing may well excite one’s wonder, but the tone of the book is popular and its appeal is not to any small section of the reading public but to all the classes and degrees of an age that, from present indications, will go down in history as the century of Woman.
31.—The Positive Evolution of Religion. Its Moral and Social Reaction. By Frederic Harrison. 8vo.
The author has undertaken to estimate the moral and social reaction of various forms of Religion—beginning with Nature Worship, Polytheism, Catholicism, Protestantism, and Deism. The volume may be looked upon as the final word, the summary of the celebrated author’s philosophy—a systematic study of the entire religious problem.
32.—The Science of Happiness. By Jean Finot, Author of “Problems of the Sexes,” etc. Translated from the French by Mary J. Safford. 8o.
In this work, which was crowned by the Academy, the author considers a subject, the solution of which offers more enticement to the well-wisher of the race than the gold of the Incas did to the treasure-seekers of Spain, who themselves doubtless looked upon the coveted yellow metal, however mistakenly, as a key to the happiness which all are trying to find. “Amid the noisy tumult of life, amid the dissonance that divides man from man,” remarks M. Finot, “the Science of Happiness tries to discover the divine link which binds humanity to happiness through the soul and through the union of souls.” The author considers the nature of happiness and the means of its attainment, as well as many allied questions.
33.—Genetic Theory of Reality. Being the Outcome of Genetic Logic as Issuing in the Æsthetic Theory of Reality Called Pancalism. By James Mark Baldwin, Ph.D., D.Sc., LL.D., Foreign Correspondent of the Institute of France, Author of “History of Psychology,” etc.
The author here states the general results of the extended studies in genetic and social science and anthropology made by him and others, and gives a critical account of the history of the interpretation of nature and man, both racial and philosophical.
The book offers an Introduction to Philosophy from a new point of view. It contains, also, a valuable glossary of the terms employed in these and similar discussions.
34.—Mosquito Control in Panama. The Eradication of Malaria and Yellow Fever in Cuba and Panama. By J. A. Le Prince, C.E., A. M., Chief Sanitary Inspector, Isthmian Canal Commission, 1904–1914, and A. J. Orenstein, M.D., Assistant Chief Sanitary Inspector, Isthmian Canal Commission. With an introduction by L. O. Howard, Ph.D., Entomologist and Chief, Bureau of Entomology, United States Department of Agriculture. 8o. 95 illustrations.
Mr. Le Prince’s books will be not only of great practical importance as a guide to future work of the same character, especially in the Tropics, but also of permanent historic value.
35.—The Organism as a Whole. From a Physico-Chemical Viewpoint. By Jacques Loeb, Author of “Comparative Physiology of the Brain.” 8o.
The author accounts for the harmonious character of the organism on a purely physico-chemical basis, without the assumption of design on the one hand, and without the formulation of too definite a theory of evolution on the other. The book contains, in addition to the text, all the necessary illustrations.
1 Bernard C., Leçons sur les Phénomènes de la Vie. Paris, 1885, i., 22–64.
2 Driesch, H., The Science and Philosophy of the Organism. 2 vols. The Gifford Lectures, 1907 and 1908.
3 v. Uexküll, J., Bausteine zu einer biologischen Weltanschauung. München, 1913.
4 v. Uexküll, J., Bausteine zu einer biologischen Weltanschauung. München, 1913, p. 216.
5 de Vries, H., Die Mutationstheorie. Leipzig, 1901.
6 This difficulty is also felt by mechanistic writers like Child, who on page 12 of his recent book on Senescence and Rejuvenescence (Chicago, 1915) makes the following remarks: “These theories of Weismann do not account satisfactorily for the peculiarly constant course and character of development and morphogenesis. If we follow them to their logical conclusion, which their authors have not done, we find ourselves forced to assume the existence of some sort of controlling and co-ordinating principle outside the units themselves and superior to them. If the units constitute the physicochemical basis of life, as their authors maintain, then this controlling principle, since it is an essential feature of life, must of necessity be something which is not physicochemical in nature. In short these theories lead us in the final analysis to the same conclusion as that reached by the neovitalists. If we are not content to accept this conclusion we must reject the theories.” These last sentences do not exhaust all the possibilities, since the writer is trying to show in this book that the widest acceptance of the chromosome theory of heredity is compatible with a consistent physicochemical conception of the organism as a whole.
7 Pasteur, L., Annal. d. Chim. et d. Physique, 1862, 3 sér., lxiv., 1.
8 Winogradsky, S., “Die Nitrification,” Handb. d. tech. Mykol., 1904–06, iii., 132.
9 Winogradsky, loc. cit., p. 163 and ff.
10 Godlewski, E., Anz. d. Akad. d. Wissensch. in Krakau, 1892, 408; 1895, 178.
11 Nathanson, Mitteil. d. zool. Station, Neapel, 1902.
12 Beijerinck, M., Folia Microbiologica, 1914, iii., 91.
13 Ernst, A., The New Flora of the Volcanic Island of Krakatau, Cambridge, 1908.
14 Euler, H., Pflanzenchemie, 1909, ii. and iii., 140.
15 This fact was thoroughly established by Mendel and Osborne. A summary of their work is given in Underhill, F. P., Physiology of the Amino Acids, 1916.
16 Van Slyke, D. D., and Cullen, G. E., Jour. Biol. Chem., 1914, xix., 141.
17 Hill, C., Jour. Chem. Soc., 1898, lxxiii., 634.
18 Armstrong, E. F., Proc. Royal Soc., 1905, B. lxxvi., 592.
19 Kastle, J. H., and Loevenhart, A. S., Am. Chem. Jour., 1900, xxiv., 491.
20 Taylor, A. E., Univ. Cal. Pub., 1904, Pathology, i., 33; Jour. Biol. Chem., 1906, ii., 87.
21 Bradley, H. C., Jour. Biol. Chem., 1913, xiii., 407.
22 This would lead to the idea that the enzymes in the cell also synthetize molecules of their own kind, or that, in other words, the synthetic processes in the cell are of the nature of autocatalysis. Loeb, Der chemische Character des Befruchtungsvorgangs, Leipzig, 1908. Robertson, T. B., Arch. f. Entwicklngsmech., 1908, xxv., 581; xxvi., 108; 1913, xxxvii., 497; Am. Jour. Physiol., 1915, xxxvii., 1; Robertson and Wasteneys, H., Arch. f. Entwicklngsmech., 1913, xxxvii., 485. Ostwald, Wo., Über die zeitlichen Eigenschaften der Entwicklungsvorgänge, Leipzig, 1908.
23 Loeb, Leo, Jour. Med. Res., 1901, vi., 28; Arch. f. Entwicklngsmech., 1907, xxiv., 655.
24 Loeb, Leo, Über die Entstehung von Bindegewebe, Leucocyten und rothen Blutkörperchen aus Epithel und über eine Methode isolierte Gewebsteile zu züchten. Chicago, 1897.
25 While this has been demonstrated thus far only for connective-tissue cells it may be true also for other cells.
26 Arrhenius, S., Worlds in the Making, London and New York, 1908, p. 212.
27 White, J., Proc. Roy. Soc., 1909, B, lxxxi., 417.
28 Macfadyen, A., Proc. Roy. Soc., 1903, lxxi., 76.
29 Becquerel, P., Revue générale des Sciences, 1914, xxv., 559.
30 Winogradsky, S., Beiträge zur Morphologie und Physiologie der Bacterien. Leipzig, 1888.
31 Johannsen, W., Elemente der exacten Erblichkeitslehre. 2d ed., 1913.
32 Murphy, J. B., Jour. Exper. Med., 1913, xvii., 482; 1914, xix., 181; xix., 513; Murphy and Morton, J. J., Jour. Exper. Med., 1915, xxii., 204.
33 The reader is referred to Morgan’s book on Regeneration (New York, 1901), for the literature on this subject.
34 Baur, E., Einführung in die experimentelle Vererbungslehre. Berlin, 1911, p. 232.
35 Literature on this subject in Chapter IV.
36 Loeb, J., King, W. O. R., and Moore, A. R., Arch. f. Entwicklngsmech., 1910, xxix., 354.
37 Moenkhaus, W. J., Am. Jour. Anat., 1904, iii., 29.
38 Loeb, J., Jour. Morphol., 1912, xxiii., 1.
39 Landois, L., Zur Lehre von der Bluttransfusion. Leipzig, 1875.
40 This is probably true only within the limits of exactness used in these experiments.
41 Friedenthal, H., “Experimenteller Nachweis der Blutverwandtschaft.” Arch. f. Physiol., 1900, 494.
42 Uhlenhuth, P., and Steffenhagen, K., Kolle-Wassermann, Handb. d. pathol. Mikroorg., 2nd Ed., 1913, iii., 257.
43 Nuttall, George H. F., Blood Immunity and Blood Relationship, Cambridge Univ. Press, 1904.
44 Nuttall, Blood Immunity and Blood Relationship, pp. 319 and 320.
45 Nuttall, pp. 345 and 346.
46 Welsh, D. A., and Chapman, H. G., Jour. Hygiene, 1910, x., 177.
47 Magnus, W., and Friedenthal, H., Ber. d. deutsch. bot. Gesellsch., 1906, xxiv., 601.
48 Richet, C., L’anaphylaxie. Paris, 1912.
49 Quoted from Wells, H. G., Jour. Infect. Diseases, 1908, v., 449.
50 Ibid., 1911, ix., 147.
51 Gay, F. P., and Robertson, T. B., Jour. Biol. Chem., 1912, xii., 233.
52 Fitzgerald, J. G., and Leathes, J. B., Univ. Cal. Pub., 1912, “Pathology,” ii., 39.
53 Bradley, H. C., and Sansum, W. D., Jour. Biol. Chem., 1914, xviii., 497.
54 Reichert, E. T., and Brown, A. P., “The Differentiation and Specificity of Corresponding Proteins and other Vital Substances in Relation to Biological Classification and Organic Evolution.” Carnegie Institution Publication No. 116, Washington, 1909.
55 Uhlenhuth, Das biologische Verfahren zur Erkennung und Unterscheidung von Menschen und Tierblut, Jena, 1905, p. 102.
56 Moss, W. L., Johns Hopkins Hospital Bulletin, 1910, xxi., 62.
57 Taylor, A. E., Jour. Biol. Chem., 1908, v., 311.
58 Wells, H. G., Jour. Infect. Diseases, 1911, ix., 166.
59 Loeb, J., and Bancroft, F. W., Jour. Exper. Zoöl., 1912, xii., 381.
60 Loeb, J., Arch. f. d. ges. Physiol., 1903, xcix., 323; 1904, civ., 325; Arch. f. Entwcklngsmech., 1910, xxx., II., 44; 1914, xl., 310; Science, 1914, xl., 316.
61 Godlewski, E., Arch. f. Entwcklngsmech., 1906, xx., 579.
62 Kupelwieser, H., Arch. f. Entwcklngsmech., 1909, xxvii., 434; Arch. f. Zellforsch., 1912, viii., 352.
63 See Chapter II.
64 Loeb, J., Science, 1914, xl., 316; Am. Naturalist, 1915, xlix., 257.
65 Loeb, Arch. f. Entwcklngsmech., 1914, xl., 310.
66 Godlewski, E., Arch. f. Entwcklngsmech., 1911, xxxiii., 196.
67 Herlant, M., Anat. Anzeiger, 1912, xlii., 563.
68 Loeb, J., Jour. Exper. Zoöl., 1914, xvii., 123.
69 Lillie, F. R., Jour. Exper. Zoöl., 1914, xvi., 523.
70 Loeb, J., Am. Naturalist, 1915, xlix., 257.
71 Lillie, F. R., Science, 1913, xxxviii., 524; Jour. Exper. Zoöl., 1914, xvi., 523; Biol. Bull., 1915, xxviii., 18.
72 Lillie, F. R., loc. cit.
73 Loeb, J., Jour. Exper. Zoöl., 1914, xvii., 123; Am. Naturalist, 1915, xlix., 257.
74 Loeb, J., Arch. f. Entwcklngsmech., 1907, xxii., 479; Artificial Parthenogenesis and Fertilization, Chicago, 1913, p. 240.
75 Loeb, J., Science, 1913, xxxviii., 749; Arch. f. Entwcklngsmech., 1914, xxxviii., 277; Wasteneys, H., Jour. Biol. Chem., 1916, xxiv., 281.
76 Loeb, J., Am. Naturalist, 1915, xlix., 257.
The writer may be permitted to illustrate by a special case his reason for declining to accept Ehrlich’s side-chain theory. Ehrlich and Sachs found that if to a given mass of toxin small quantities of antitoxin are added successively the first fraction added neutralized more than the later fractions; and on the basis of this reasoning Ehrlich concluded that ten different toxins were contained in the diphtheria toxin. Arrhenius showed that the same phenomenon can be obtained when a weak base like NH4OH is neutralized by a weak acid (e. g., boric acid); hence we should assume that NH4OH consists of ten different forms of ammonia. Both cases, the saturation of toxin with antitoxin and ammonia with boric acid are equilibrium phenomena. (Arrhenius, S., Quantitative Laws in Biological Chemistry, London, 1915.)
77 Castle, W. E., Bull. Mus. Comp. Zoöl., Harvard, 1896, xxvii., 203.
78 Morgan, T. H., Jour. Exper. Zoöl., 1904, i., 135; Arch. f. Entwcklngsmech., 1910, xxx., 206.
79 Fuchs, H. M., Jour. Genet., 1915, iv., 215.
80 Quoted from Fuchs.
81 Correns, C., Biol. Centralbl., 1913, xxxiii., 389.
82 Pfeffer, Untersuchungen aus dem botanischen Institut zu Tübingen, 1881–1885, i., 363.
83 Bruchmann, H., Flora, 1909, ic., 193.
84 The substitution of well-known physicochemical agencies for the mysterious action of the spermatozoön was the task the writer set himself in this work and not the explanation of natural parthenogenesis, as the author of a recent text-book seems to assume.
85 Loeb, J., Am. Jour. Physiol., 1899, iii., 135; 1900, iii., 434.
86 Loeb, J., Artificial Parthenogenesis and Fertilization, Chicago, 1913. The reader is referred to this book for the literature on the subject.
87 The reader will find a description of the development of this egg in the next chapter.
88 The reader is referred for details to the writer’s book on the subject.
89 Robertson, T. B., Arch. f. Entwcklngsmech., 1912, xxxv., 64.
90 Loeb, J., Über den chemischen Charakter des Befruchtungsvorgangs, etc., Leipzig, 1908.
91 v. Knaffl, E., Arch. f. d. ges. Physiol., 1908, cxxiii., 279.
92 Loeb, J., Artificial Parthenogenesis and Fertilization, p. 255.
93 It has been stated by several writers that the eggs of the sea urchin can no longer form the fertilization membrane when the jelly surrounding the egg is dissolved. The writer has found that if the jelly surrounding the eggs of Strongylocentrotus purpuratus is dissolved by acid the eggs still form a fertilization membrane upon the entrance of a spermatozoön.
94 Loeb, J., Artificial Parthenogenesis and Fertilization, 1913, p. 250 and ff.
95 Delage, Y., Arch. d. Zoöl. expér. et gén., 1902, x., 213; 1904, ii., 27; 1905, iii., 104.
96 Lillie, R. S., Jour. Biol. Chem., 1916, xxiv., 233.
97 It is necessary to call attention to the fact that sugar solutions of a high concentration (e. g., m solutions) have a much higher osmotic pressure than that which they should have theoretically (Lord Berkeley and Hartley). Delage by ignoring this fact has misinterpreted his experiments with sugar solutions. See Lloyd, D. J., Arch. f. Entwcklngsmech., 1914, xxxviii., 402.
98 Loeb, J., and Wasteneys, H., Jour. Biol. Chem., 1913, xiv., 517; Biochem. Ztschr., 1913, lvi., 295.
99 Loeb, J., Biochem. Ztschr., 1906, ii., 81.
100 Loeb, J., Arch. f. d. ges. Physiol., 1906, cxiii., 487; Biochem. Ztschr., 1910, xxvi., 279, 289; xxvii., 304; xxix., 80; Arch. f. Entwcklngsmech., 1914, xl., 322.
101 Herlant, M., Arch. de Biol., 1913, xxviii., 505.
102 It is also important to remember that the formation of astrospheres after mere membrane formation occurs considerably more slowly than if the egg has also received a treatment with a hypertonic solution.
103 The writer found that the eggs of Fundulus will segment a number of times even if all the oxygen has apparently been removed.
104 Loeb, J., Biochem. Ztschr., 1906, ii., 183.
105 Thus the treatment of an unfertilized egg without membrane with a hypertonic solution combines two effects, first the general cytolytic alteration of the cortical layer of the membrane and the corrective effect of the hypertonic solution. The former effect raises the rate of oxidations in the egg, the latter does not.
106 Warburg, O., Sitzungsber. d. Heidelberger Akad. d. Wissnsch., B. 1914.
107 Loeb, J., Biochem. Ztschr., 1906, ii., 87.
108 Unless the egg is left so long in the pure NaCl solution that its permeability is increased.
109 Lillie, R. S., Jour. Morphol., 1911, xxii., 695; Am. Jour. Physiol., 1911, xxvii., 289.
110 McClendon, J. F., Publications of the Carnegie Institution, No. 183, 125; Am. Jour. Physiol., 1910, xxvii., 240.
111 Gray, J., Proc. Cambridge Philosophical Society, 1913, xvii., 1.
112 R. Lillie has recently shown that in a hypotonic solution water diffuses more rapidly into a fertilized than into an unfertilized egg. This is exactly what one should expect since the unfertilized egg is not only surrounded by the cortical layer but also by a thick layer of jelly both of which are lacking in the fertilized egg. It is difficult to understand how this observation can throw any light on the mechanism of development, since water diffuses rapidly enough into the unfertilized egg.
113 Delage, Y., Compt. rend. Acad. Sc., 1909, cxlviii., 453.
114 Since this was written, two more of the parthenogenetic frogs over a year old died. Both were males.
115 Loeb, J., Artificial Parthenogenesis and Fertilization, Chicago, 1913.
116 Driesch, H., Science and Philosophy of the Organism. London, 1908 and 1909.
117 Boveri, Th., Verhandl. d. physik.-med. Gesellsch., Würzburg, 1901, xxxiv., 145.
118 Lyon, E. P., Arch. f. Entwcklngsmech., 1907, xxiii., 151; Morgan, T. H., and Spooner, G. B., ibid., 1909, xxviii., 104; Morgan, Jour. Exper. Zoöl., 1910, ix., 594; Conklin, E. G., ibid., 1910, ix., 417; Lillie, F. R., Biol. Bull., 1909, xvi., 54.
119 Driesch, H., Ztschr. f. wissnsch. Zoöl., 1891, liii., 160.
120 Loeb, J., Arch. f. Entwcklngsmech., 1909, xxvii., 119.
121 Driesch, H., Arch. f. Entwcklngsmech., 1900, x., 361.
122 Driesch, H., Arch. f. Entswcklngsmech., 1902, xiv., 500.
123 Boveri, Th., Verhandl. d. physik. med. Gesellsch., Würzburg, N.F., 1901, xxxiv., 145.
124 v. Uexküll makes in his last book (Bausteine zu einer biologischen Weltanschauung, München, 1913, p. 24) the following statement: “Driesch succeeded in showing that the germ cell has no trace of a machine-like structure but consists entirely of equivalent parts.” This is not correct.
125 Loeb, J., and Beutner, R., Biochem. Ztschr., 1912, xli., 1; xliv., 303; 1913, li., 288; li., 300; 1914, lix., 195.
126 Loeb, J., The Dynamics of Living Matter. New York, 1906. Introductory Remarks.
127 Roux, W., Virchow’s Archiv, 1888, cxiv., 113.
128 Morgan, T. H., Embryology of the Frog. New York.
129 Crampton, H. E., New York Academy of Sciences, 1894; Kofoid, C. A., Proc. Am. Acad. Arts and Sciences, 1894, xxix.
130 Conklin, E. G., Anat. Anzeig., 1903, xxiii., 577; Heredity and Environment in the Development of Man. Princeton, 1915, p. 171.
131 Wilson, E. B., Science, 1904, xx., 748; Jour. Exper. Zoöl., 1904, i., 1, 197.
132 The reader will notice the absence of “regulation.”
133 Conklin, E. G., Heredity and Environment in the Development of Man. Princeton University Press, 1915. The reader is referred to this book for the literature and main facts on the structure of the egg; it should also be stated that Conklin’s book is one of the best introductions to modern biology in the English literature.
134 Conklin, E. G., loc. cit., p. 117.
135 Loeb, J., Jour. Morphol., 1893, xiii., 161; The Mechanistic Conception of Life. Chicago, 1912, p. 106.
136 Driesch, H., Science and Philosophy of the Organism, i., p. 104.
137 Herbst, C., Formative Reize in der tierischen Ontogenese. Leipzig, 1901.
138 Braus, H., Münchener Med. Wochnschr., 1903, 1 (II.), No. 47, p. 2076.
139 Nussbaum, M., Arch. f. mikroscop. Anat., 1886, xxvi., 485.
140 It must not be overlooked that in bacteria and the blue algæ no distinct differentiation into nucleus and protoplasm can be shown. To these organisms, therefore, the experiments of Nussbaum cannot be applied.
141 Loeb, J., Arch. d. f. ges. Physiol., 1893, lv., 525.
142 v. Sachs, J., “Stoff und Form der Pflanzenorgane,” Gesammelte Abhandlungen, 1892, ii., 1160. Arbeiten a. d. bot. Inst. Würzburg, 1880–82.
143 Goebel, K., Einleitung in die experimentelle Morphologie der Pflanzen, 1908.
144 Loeb, J., Untersuchungen zur physiologischen Morphologie der Tiere. I. Heteromorphose. Würzburg, 1891. II. Organbildung und Wachsthum. 1892. Reprinted in Studies in General Physiology. Chicago, 1906.
145 Gudernatsch, J. F., Zentralbl. f. Physiol., 1912, xxvi., 323; Arch. f. Entwcklngsmech., 1912, xxxv., 457; Am. Jour. Anat., 1914, xv., 431.
146 Morse, M., Jour. Biol. Chem., 1914, xix., 421.
147 Loeb, J., Arch. f. Entwcklngsmech., 1897, iv., 502.
148 Uhlenhuth, E., ibid., 1913, xxxvi., 211.
149 Loeb, Leo, Zentralbl. f. allg. Path. u. path. Anat., 1907, xviii., 563; Zentralbl. f. Physiol., 1908, xxii., 498; 1909, xxiii., 73; 1910, xxiv., 203; Arch. f. Entwcklngsmech., 1909, xxvii., 89, 463; Jour. Am. Med. Assoc., 1908, l., 1897; 1909, liii., 1471.
150 Quoted from M. Caullery, Les Problèmes de la Sexualité, Paris, 1913, p. 126.
151 Smith, Geoffrey, Proc. Roy. Soc., B. 1915, lxxxviii., 418.
152 Loeb, J., Bot. Gazette, 1915, lx., 249.
153 With larger leaves the experiment may also succeed in moist air.
154 Loeb, J., Untersuchungen zur physiologischen Morphologie. I. Heteromorphose. 1891. II. Organbildung und Wachsthum. Würzburg, 1892.
155 Bickford, E. E., Jour. Morphol., 1894, ix., 417.
156 Driesch, H., Science and Philosophy of the Organism, i., 127.
157 Child, C. M., “Die physiologische Isolation von Teilen des Organismus,” Roux’s Vorträge und Aufsätze, Leipzig, 1911.
158 Loeb, J., “Untersuchungen zur physiologischen Morphologie der Tiere.”
159 Morgan, T. H., Regeneration, New York, 1901.
160 Bardeen, C. R., Am. Jour. Physiol., 1901, v., 1; Arch. f. Entwcklngsmech., 1903, xvi., 1.
161 Przibram, H., Arch. f. Entwcklngsmech., 1901, xi., 329.
162 Child, C. M., Senescence and Rejuvenescence. Chicago, 1915.
163 Loeb, J., Am. Jour. Physiol., 1900, iv., 60.
164 The writer quotes this after Driesch.
165 Driesch, H., Arch. f. Entwcklngsmech., 1902, xiv., 247.
166 One author, Miss Thatcher, in trying to repeat these observations, did not notice the total collapse of the tissues and concluded that my observations must have been wrong. The writer is fairly certain that his observations were correct.
167 Not yet published.
168 v. Sachs, J., “Physiologische Notizen,” vi., Flora, 1893.
169 Ibid., ix., 425, Flora, 1895.
170 Morgan, T. H., Arch. f. Entwcklngsmech., 1895, ii., 81; 1901, xiii., 416; 1903, xvi., 117.
171 Driesch, H., Arch. f. Entwcklngsmech., 1898, vi., 198; 1900, x., 361.
172 Delage, Y., Arch. Zoöl. expér., 1899, vii., 383.
173 Driesch, H., Arch. f. Entwcklngsmech., 1905, xix., 648.
174 Loeb, Leo, Arch. f. Entwcklngsmech., 1898, vi., 297.
175 Spain, K. C., and Loeb, Leo, Jour. Exper. Med., 1916, xxiii., 107; Loeb, L., and Addison, W. H. F., Arch. f. Entwcklngsmech., 1911, xxxii., 44; 1913, xxxvii., 635.
176 The excessive formation of epithelial cells in the healing of wounds has led the older pathologists to the generalization that if something is removed in the body an excessive compensation will take place. The formation of antibodies has even been explained on this basis by Weiggert and Ehrlich in their side-chain theory. As a matter of fact, this generalization is entirely incorrect and in regeneration of starfish, actinians, flatworms, annelids, and possibly in all forms the reverse is true; e. g., if we cut off the anterior half of the body in Cerianthus less is reproduced than was cut away namely only tentacles and the mouth, but not the missing piece of the body. Weiggert’s conception of regeneration was probably based on the phenomenon of the healing of wounds, but the excessive epithelium formation in this case is not the expression of a general law of regeneration but of the peculiar mechanical conditions which lead to mitoses. It would be a very strange coincidence indeed if a theory of antibody formation based on such an erroneous generalization should be correct.
177 Loeb, Arch. f. Entwcklngsmech., 1914, xxxviii., 277.
178 Wasteneys, H., Jour. Biol. Chem., 1916, xxiv., 281.
179 F. Lillie thinks that the KCN in this experiment merely inhibits the change of the cortical layer necessary for development. This is contradicted by two facts: first, the writer has shown in 1906 that KCN does not inhibit the membrane formation, and, second, the eggs will not return to the resting stage when put back into sea water too soon; in that case they will disintegrate. This shows that in the KCN something more happens than the mere block to disintegration.
180 Loeb, J., Untersuchungen zur physiologischen Morphologie der Tiere. II. Organbildung und Wachsthum. Würzburg, 1892.
181 Loeb, J., Die chemische Entwicklungserregung des tierischen Eies. Berlin, 1909.
182 McClung, C. E., “The Accessory Chromosome—Sex Determinant?” Biol. Bull., 1902, iii., 43.
183 Wilson, E. B., “Studies on Chromosomes,” Jour. Exper. Zoöl., 1905, ii., 371, 507; 1906, iii., 1; 1909, vi., 69, 147; 1910, ix., 53; 1912, xiii., 345. “Croonian Lecture,” 1914, Proc. Roy. Soc., B. lxxxviii., 333.
184 Doncaster, L., The Determination of Sex. Cambridge, 1914.
185 Morgan, T. H., Heredity and Sex. New York, 1913.
186 Bridges, C. B., Genetics, 1916, i., 1.
187 Boveri, Th., Arch. f. Entwcklngsmech., 1915, xlii., 264.
188 Boveri, Th., Verhand. d. phys.-med. Gesellsch. Würzburg, 1911, xli., 85. Schleip, W., Ber. d. naturf. Gesellsch., Freiburg i. Br., 1911, xix.
189 Correns, C., Biol. Centralbl., 1916, xxxvi., 12.
190 Baltzer, F., Mitteil. d. zoölog. Station, Neapel, 1914, xxii.
191 Caullery, M., Les Problèmes de la Sexualité. Paris, 1913.
192 Goodale, H. D., Biol. Bull., 1916, xxx., 286.
193 Lillie, F., Science, 1916, xliii., 611.
194 Janda, V., Arch. f. Entwcklngsmech., 1912, xxxiii., 345; xxxiv., 557.
195 Goldschmidt, R., Proc. Nat. Acad. Sc., 1916, ii., 53; Ztschr. induct. Abstammungslehre, 1912, vii., and 1914, xi.
196 This account of Marchal’s beautiful experiments is taken from Caullery, M., Les Problèmes de la Sexualité. Paris, 1913.
197 Whitney, D. D., Science, 1916, xliii., 176.
198 Shull, A. F., and Ladoff, S., Science, 1916, xliii., 177.
199 Steinach, E., Zentralbl. f. Physiol., 1910, xxiv., 551; Arch. f. d. ges. Physiol., 1912, cxliv., 72.
200 For the literature on the subject the reader is referred to Morgan, T. H., Sturtevant, A. H., Muller, H. J., and Bridges, C. B., The Mechanism of Mendelian Heredity. New York, 1915.
201 Mendel, G., “Experiment in Plant-Hybridization,” translated in W. Bateson’s classical book on Mendel’s Principles of Heredity. Cambridge, 1909.
202 The reader will find a critical discussion of the presence and absence theory on page 220 of Morgan, Sturtevant, Muller, and Bridges, The Mechanism of Mendelian Heredity. New York, 1915.
203 Sutton, W. S., “The Chromosomes in Heredity,” Biol. Bull., 1904, iv., 231.
204 Morgan, T. H., Sturtevant, A. H., Muller, H. J., and Bridges, C. B., Mechanism of Mendelian Heredity. New York, 1915, p. 26.
205 Morgan, T. H., Sturtevant, A. H., Muller, H. J., and Bridges, C. B., The Mechanism of Mendelian Heredity. New York, 1915.
206 Bateson, W., loc. cit., p. 157.
207 The number of hereditary characters examined to test the theory was over 130.
208 Bateson, W., Mendel’s Principles of Heredity, 3d ed., 1913; Davenport, Chas. B., Heredity in Relation to Eugenics, 1911. Pearl, R., Modes of Research in Genetics.
209 Loeb, J., King, W. O. R., and Moore, A. R., Arch. f. Entwcklngsmech., 1910, xxix., 354. These experiments have been repeated at different seasons of the year and in different years and have been found to be constant.
210 Moore, A. R., Arch. f. Entwcklngsmech., 1912, xxxiv., 168.
211 Bertrand, G., Ann. d. l’Inst. Pasteur, 1908, xxii., 381; Bull. Soc. Chim., 1896, xv., 791.
212 Chodat, R., Arch. d. Sc. phys. et nat., 1915, xxxix., 327.
213 Gortner, R. A., Trans. Chem. Soc., 1910, xcvii., 110.
214 Onslow, H., Proc. Roy. Soc., 1915, B. lxxxix., 36.
215 Loeb, J., “Egg Structure and the Heredity of Instincts,” The Monist, 1897, vii., 481.
216 Bateson, W., Nature, 1916, xciii., 674.
217 Ideas similar to those expressed in this chapter may be found in the writer’s former book Comparative Physiology of the Brain and Comparative Psychology, New York, 1900, and in the books by George Bohn, La Naissance de l’Intelligence, Paris, 1909, and La nouvelle Psychologie animale, Paris, 1911.
218 Graber, V., Grundlinien zur Erforschung des Helligkeits- und Farbensinnes der Tiere. Prag, 1884.
219 Loeb, J., Sitzungsber. d. physik.-med. Gesellsch. Würzburg, 1888. Der Heliotropismus der Tiere und seine Übereinstimmung mit dem Heliotropismus der Pflanzen. Würzburg, 1889. Arch. f. d. ges. Physiol., 1897, lxvi., 439.
220 Loeb, J., Arch. f. d. ges. Physiol., 1890, xlvii., 391; 1896, lxiii., 273.
221 Loeb, J., Arch. f. d. ges. Physiol., 1897, lxvi., 439.
222 Loeb, J., The Mechanistic Conception of Life, Chicago, 1912, p. 27.
223 Loeb, J., and Ewald, W. F., Zentralbl. f. Physiol., 1914, xxvii., 1165.
224 Ewald, W. F., Science, 1913, xxxviii., 236.
225 Fröschel, P., Sitzungsber. d. k. Akad. d. Wissensch., Wien, 1908, cxvii.
226 Blaauw, H. A., Rec. d. travaux botaniques Neérlandais, 1909, v., 209.
227 Loeb, J., Arch. f. d. ges. Physiol., 1893, liv., 81; Jour. Exper. Zoöl., 1907, iv., 151.
228 Bancroft, F. W., Jour. Exper. Zoöl., 1913, xv., 383.
229 Loeb, J., Studies in General Physiology, Chicago, 1905, p. 2.
230 Patten, Bradley M., Am. Jour. Physiol., 1915, xxxviii., 313.
231 According to this theory the animal is not directly oriented by the outside force, e. g. the light, but selects among its random movements the one which is most “suited” and keeps on moving in this direction. This idea is untenable for most if not all the cases of tropisms and has been refuted by practically all the workers in this field, e. g., Parker and his pupils, Bohn, H. B. Torrey, Holmes, Bancroft, Ewald, and others. It is only upheld by Jennings and Mast; and is accepted among those to whom the idea of a physicochemical explanation of life phenomena does not appeal. Torrey and Bancroft (for the literature the reader is referred to Bancroft’s paper, Jour. Exper. Zoöl., 1913, xv., 383) have shown directly that the theory of trial and error is not even correct for the organism for which Jennings has developed this idea; namely Euglena.
232 Loeb, J., and Maxwell, S. S., Arch. f. d. ges. Physiol., 1896, lxiii., 121.
233 That the mechanisms by which heliotropic and galvanotropic orientation is brought about are identical was shown by Bancroft in Euglena (Bancroft, loc. cit.).
234 Loeb, J., and Maxwell, S. S., Arch. f. d. ges. Physiol., 1896, lxiii., 121.
235 Loeb, J., Dynamics of Living Matter, p. 126.
236 Loeb, J., and Maxwell, S. S., Univ. Cal. Pub., 1910, Physiol., iii., 195; Loeb and Wasteneys, Proc. Nat. Acad. Sc., 1915, i., 44; Science, 1915, xli., 328; Jour. Exper. Zoöl., 1915, xix., 23; 1916, xx., 217.
237 Mast, S. O., Proc. Nat. Acad. Sc., 1915, i., 622.
238 Hess, C., “Gesichtssinn,” Winterstein’s Handb. d. vergl. Physiol., 1913, iv.
239 v. Frisch, K., “Der Farbensinn und Formensinn der Biene,” Zoöl. Jahrb. Abt. f. allg. Zoöl. u. Physiol., 1914, xxxv. See also Ewald, W. F., Ztschr. f. Sinnesphysiol., 1914, xlviii., 285.
240 Loeb, J., Der Heliotropismus der Tiere, 1889.
241 Kellogg, V. L., Science, 1903, xviii., 693.
242 Loeb, J., Arch. f. d. ges. Physiol., 1906, cxv., 564.
243 Ibid., 1893, liv., 81.
244 Groom, Theo. T., and Loeb, J., Biol. Centralbl., 1890, x., 160; Ewald, W. F., Jour. Exper. Zoöl., 1912, xiii., 591.
245 Loeb, J., Arch. f. d. ges. Physiol., 1906, cxv., 564; Moore, A. R., Jour. Exper. Zoöl., 1912, xiii., 573.
246 Cannon, W. B., Bodily Changes in Pain, Hunger, Fear, and Rage, New York, 1915.
247 Setchell, W. A., Science, 1903, xxvii., 934.
248 Duclaux, E., Traité de microbiol., 1898, i., 280.
249 A full discussion of the literature on temperature coefficients is given in A. Kanitz’s book on Temperatur and Lebensvorgänge, Berlin, 1915.
250 Van Slyke, D. D., and Cullen, G. E., Jour. Biol. Chem., 1914, xix., 141.
251 These considerations may meet the objections of Krogh to the application of the van’t Hoff rule of temperature effect on reaction velocity to life phenomena. See also the discussion of this subject in Kanitz’s book.
252 Lillie, F. R., and Knowlton, E. P., Zoöl. Bull., 1897, i.
253 Hertwig, O., Arch. mikrosk. Anat., 1898, li., 319. See also E. Cohen, Vorträge für Aerste über physikalische Chemie. 2d ed. Leipzig, 1907.
254 Loeb, J., Arch. f. d. ges. Physiol., 1908, cxxiv., 411; Loeb J., and Wasteneys, H., Biochem. Ztschr., 1911, xxxvi., 345; Loeb J., and Chamberlain, M. M., Jour. Exper. Zoöl., 1915, xix., 559.
255 Loc. cit.
256 Kanitz, A., loc. cit., p. 123.
257 Loeb, J., and Chamberlain, M. M., Jour. Exper. Zoöl., 1915, xix., 559.
258 Loeb, J., and Ewald, W. F., Biochem. Ztschr., 1913, lviii., 179.
259 Clausen, H., Landwirtschaftl. Jahrb., 1890, xix., 893.
260 Matthaei, G. L. C., Trans. Philosoph. Soc., 1904, cxcvii., 47; Blackman, F. F., Ann. of Bot., 1905, xix., 281.
261 Loeb, J., “The Poisonous Character of a Pure NaCl Solution.” Am. Jour. Physiol., 1900, iii., 329; Arch. f. d. ges. Physiol., 1901, lxxxviii., 68; Am. Jour. Physiol., 1902, vi., 411; Biochem. Zischr., 1906, ii., 81.
262 Loeb, J., Jour. Biol. Chem., 1915, xxiii., 423.
263 Loeb, J., “On the Physiological Effects of the Valency and Possibly the Electrical Charges of Ions,” Am. Jour. Physiol., 1902, vi., 411.
264 Loeb, J., Jour. Biol. Chem., 1914, xix., 431.
265 Loeb, J., Arch. f. d. ges. Physiol., 1905, cvii., 252.
266 Robertson, T. B., Ergeb. d. Physiol., 1910, x., 216.
267 Loeb, J., Biochem. Ztschr., 1912, xlvii., 127.
268 Osterhout, W. J. V., Bot. Gazette, 1906, xlii., 127; 1907, xliv., 257; Jour. Biol., Chem., 1906, i., 363.
269 Ostwald, Wo., Arch. f. d. ges. Physiol., 1905, cvi., 568.
270 Loeb, J., Jour. Biol. Chem., 1915, xxiii., 423.
271 Loeb, J., Jour. Biol. Chem., 1905–06, i., 427.
272 Meltzer, S. J., and Auer, J., Am. Jour. Physiol., 1908, xxi., 400.
273 This theory was first expressed by the writer in Am. Jour. Physiol., 1900, iii., 434.
274 Henderson, L., The Fitness of the Environment. See also Michaelis, L., Die Wasserstoffionenconzentration. Berlin. 1914.
275 Loeb, J., Der Heliotropismus der Tiere and seine Übereinstimmung mit dem Heliotropismus der Pflanzen. Würzburg, 1890 (appeared in 1889).
276 Loeb, J., Biol. Bull., 1915, xxix., 50.
277 Cuénot has proposed the term preadaptation for such cases and this term expresses the situation correctly. Cuénot, L., La Génèse des Espèces animales. Paris, 1911.
278 Kammerer, P., Arch. f. Entwcklngsmech., 1912, xxxiii., 349.
279 Loeb, J., Arch. d. f. ges. Physiol., 1896, lxiii., 273.
280 Goldfarb, A. J., Jour. Exper. Zoöl., 1906, iii., 129; 1910, viii., 133.
281 Loeb, J., Biochem. Ztschr., 1913, liii., 391.
282 Loeb, J., Biochem. Ztschr., 1913, liii., 391.
283 Dieudonné, A., Arb. a. d. kais. Gesndhtsmt., 1894, ix., 492.
284 Davenport, C. B., and Castle, W. E., Arch. f. Entwcklngsmech., 1896, ii., 227.
285 Kryž, F., Arch. f. Entwcklngsmech., 1907, xxiii., 560.
286 Loeb, J., and Wasteneys, H., Jour. Exper. Zoöl., 1912, xii., 543.
287 Kammerer, P., Arch. f. Entwcklngsmech., 1909, xxviii., 448.
288 Bateson, W., Problems of Genetics, pp. 201–202. Yale University Press, 1913.
289 Kammerer, P., Arch. f. Entwcklngsmech., 1913, xxxvi., 4.
290 Werner, F., Biol. Centralbl., 1915, xxxv., 176.
291 Loeb, J., The Mechanistic Conception of Life. Chicago, 1912.
292 de Vries, H., The Mutation Theory, translated by Farmer, J. B., and Darbishire, A. D., Chicago, 1909. Species and Varieties. Chicago, 1906. Gruppenweise Artbildung. Berlin, 1913.
293 For a critical discussion of the details, see Bateson, W., Problems of Genetics, New Haven, 1913, Chapter X.
294 Fermi, C., Centralbl. f. Bacteriologie, Abt. 1, 1910, lvi., 55.
295 Levene, P. A., Autolysis. The Harvey Lectures, 1905–1906, p. 73, gives a full account of the work on this subject up to 1905.
296 Hoppe-Seyler, F., Tübinger med.-chem. Untersuchungen, 1871, P. 499.
297 Levene, P. A., Am. Jour. Physiol., 1904, xii., 276.
298 Bradley, H. C., and Morse, M., Jour. Biol. Chem., 1915, xxi., 209.
299 Bradley, H. C., ibid., 1915, xxii., 113.
300 Loeb, J., Arch. f. d. ges. Physiol., 1895, lxii., 249.
301 Budgett, S. P., Am. Jour. Physiol., 1898, i., 210.
302 Loeb, J., The Dynamics of Living Matter, New York, 1906, pp. 19–21.
303 Child, C. M., Senescence and Rejuvenescence, Chicago, 1915.
304 It is a fact that in the early cells of Ctenolabrus the dissolution of the cell walls through lack of O precedes death, since when oxygen is admitted early enough the cells recover again. In infusorians the bursting of the animal due to lack of O occurs suddenly, while the animal is still moving, and this bursting is the cause of death, and not the reverse.
305 Metchnikoff, E., Ann. d. l’Inst. Pasteur, 1915, xxix., 477.
306 Loeb, J., Biol. Bull., 1902, iii., 295.
307 Loeb, J., Arch. f. d. ges. Physiol., 1908, cxxiv., 411.
308 K. Brandt (“Über den Nitratgehalt des Ozeanwassers and seine biologische Bedeutung,” Abh. d. kais. Leop. Carol. deutsch. Akad. d. Naturfoscher., 1915) accounts for this fact by the assumption that through the greater activity of the denitrifying bacteria in the tropical waters the amount of available nitrates is here comparatively smaller than in the polar oceans. The writer fully appreciates the importance of this fact but nevertheless is inclined also to see a limiting factor in the enormously rapid decline of the duration of life at the upper temperature limits.
309 Metchnikoff, E., The Prolongation of Life. New York, 1907.
A small number of spelling anomalies were noted and these have mostly been corrected, but a few that possibly represent authentic contemporary alternatives have been left unchanged. A list of anomalies is given below.
Corrections
spermatozoon —> spermatozoön
i.e. —> i. e.
e.g. —> e. g.
nermaphrodite —> hermaphrodite
suceeded —> succeeded
ôf —> of
tryosinase —> tyrosinase
in-as-much —> inasmuch
ultra-violet —> ultraviolet
view-point —> viewpoint
Fredericq —> Frédéricq
Korösy —> Körösy
Sitzngsber —> Sitzungsber
negaceros —> megaceros
Variants
clew/clue
Entswcklngsmech/Entwcklngsmech/Entwicklngsmech
peroxidase/peroxydase (latter spelling in quoted text.)
20° C/20°C (spaced and unspaced temperature specifications)
8vo/8^o (octavo paper size in the publications listed in the end matter)
***END OF THE PROJECT GUTENBERG EBOOK THE ORGANISM AS A WHOLE***
******* This file should be named 45962-h.txt or 45962-h.zip *******
This and all associated files of various formats will be found in:
http://www.gutenberg.org/4/5/9/6/45962
Updated editions will replace the previous one--the old editions will be renamed.
Creating the works from public domain print editions means that no one owns a United States copyright in these works, so the Foundation (and you!) can copy and distribute it in the United States without permission and without paying copyright royalties. Special rules, set forth in the General Terms of Use part of this license, apply to copying and distributing Project Gutenberg-tm electronic works to protect the PROJECT GUTENBERG-tm concept and trademark. Project Gutenberg is a registered trademark, and may not be used if you charge for the eBooks, unless you receive specific permission. If you do not charge anything for copies of this eBook, complying with the rules is very easy. You may use this eBook for nearly any purpose such as creation of derivative works, reports, performances and research. They may be modified and printed and given away--you may do practically ANYTHING with public domain eBooks. Redistribution is subject to the trademark license, especially commercial redistribution.
To protect the Project Gutenberg-tm mission of promoting the free distribution of electronic works, by using or distributing this work (or any other work associated in any way with the phrase "Project Gutenberg"), you agree to comply with all the terms of the Full Project Gutenberg-tm License available with this file or online at www.gutenberg.org/license.
1.A. By reading or using any part of this Project Gutenberg-tm electronic work, you indicate that you have read, understand, agree to and accept all the terms of this license and intellectual property (trademark/copyright) agreement. If you do not agree to abide by all the terms of this agreement, you must cease using and return or destroy all copies of Project Gutenberg-tm electronic works in your possession. If you paid a fee for obtaining a copy of or access to a Project Gutenberg-tm electronic work and you do not agree to be bound by the terms of this agreement, you may obtain a refund from the person or entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be used on or associated in any way with an electronic work by people who agree to be bound by the terms of this agreement. There are a few things that you can do with most Project Gutenberg-tm electronic works even without complying with the full terms of this agreement. See paragraph 1.C below. There are a lot of things you can do with Project Gutenberg-tm electronic works if you follow the terms of this agreement and help preserve free future access to Project Gutenberg-tm electronic works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation" or PGLAF), owns a compilation copyright in the collection of Project Gutenberg-tm electronic works. Nearly all the individual works in the collection are in the public domain in the United States. If an individual work is in the public domain in the United States and you are located in the United States, we do not claim a right to prevent you from copying, distributing, performing, displaying or creating derivative works based on the work as long as all references to Project Gutenberg are removed. Of course, we hope that you will support the Project Gutenberg-tm mission of promoting free access to electronic works by freely sharing Project Gutenberg-tm works in compliance with the terms of this agreement for keeping the Project Gutenberg-tm name associated with the work. You can easily comply with the terms of this agreement by keeping this work in the same format with its attached full Project Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern what you can do with this work. Copyright laws in most countries are in a constant state of change. If you are outside the United States, check the laws of your country in addition to the terms of this agreement before downloading, copying, displaying, performing, distributing or creating derivative works based on this work or any other Project Gutenberg-tm work. The Foundation makes no representations concerning the copyright status of any work in any country outside the United States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate access to, the full Project Gutenberg-tm License must appear prominently whenever any copy of a Project Gutenberg-tm work (any work on which the phrase "Project Gutenberg" appears, or with which the phrase "Project Gutenberg" is associated) is accessed, displayed, performed, viewed, copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived from the public domain (does not contain a notice indicating that it is posted with permission of the copyright holder), the work can be copied and distributed to anyone in the United States without paying any fees or charges. If you are redistributing or providing access to a work with the phrase "Project Gutenberg" associated with or appearing on the work, you must comply either with the requirements of paragraphs 1.E.1 through 1.E.7 or obtain permission for the use of the work and the Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or 1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted with the permission of the copyright holder, your use and distribution must comply with both paragraphs 1.E.1 through 1.E.7 and any additional terms imposed by the copyright holder. Additional terms will be linked to the Project Gutenberg-tm License for all works posted with the permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm License terms from this work, or any files containing a part of this work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this electronic work, or any part of this electronic work, without prominently displaying the sentence set forth in paragraph 1.E.1 with active links or immediate access to the full terms of the Project Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary, compressed, marked up, nonproprietary or proprietary form, including any word processing or hypertext form. However, if you provide access to or distribute copies of a Project Gutenberg-tm work in a format other than "Plain Vanilla ASCII" or other format used in the official version posted on the official Project Gutenberg-tm web site (www.gutenberg.org), you must, at no additional cost, fee or expense to the user, provide a copy, a means of exporting a copy, or a means of obtaining a copy upon request, of the work in its original "Plain Vanilla ASCII" or other form. Any alternate format must include the full Project Gutenberg-tm License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying, performing, copying or distributing any Project Gutenberg-tm works unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing access to or distributing Project Gutenberg-tm electronic works provided that
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm electronic work or group of works on different terms than are set forth in this agreement, you must obtain permission in writing from both the Project Gutenberg Literary Archive Foundation and Michael Hart, the owner of the Project Gutenberg-tm trademark. Contact the Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable effort to identify, do copyright research on, transcribe and proofread public domain works in creating the Project Gutenberg-tm collection. Despite these efforts, Project Gutenberg-tm electronic works, and the medium on which they may be stored, may contain "Defects," such as, but not limited to, incomplete, inaccurate or corrupt data, transcription errors, a copyright or other intellectual property infringement, a defective or damaged disk or other medium, a computer virus, or computer codes that damage or cannot be read by your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right of Replacement or Refund" described in paragraph 1.F.3, the Project Gutenberg Literary Archive Foundation, the owner of the Project Gutenberg-tm trademark, and any other party distributing a Project Gutenberg-tm electronic work under this agreement, disclaim all liability to you for damages, costs and expenses, including legal fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a defect in this electronic work within 90 days of receiving it, you can receive a refund of the money (if any) you paid for it by sending a written explanation to the person you received the work from. If you received the work on a physical medium, you must return the medium with your written explanation. The person or entity that provided you with the defective work may elect to provide a replacement copy in lieu of a refund. If you received the work electronically, the person or entity providing it to you may choose to give you a second opportunity to receive the work electronically in lieu of a refund. If the second copy is also defective, you may demand a refund in writing without further opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth in paragraph 1.F.3, this work is provided to you 'AS-IS', WITH NO OTHER WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO WARRANTIES OF MERCHANTABILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied warranties or the exclusion or limitation of certain types of damages. If any disclaimer or limitation set forth in this agreement violates the law of the state applicable to this agreement, the agreement shall be interpreted to make the maximum disclaimer or limitation permitted by the applicable state law. The invalidity or unenforceability of any provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the trademark owner, any agent or employee of the Foundation, anyone providing copies of Project Gutenberg-tm electronic works in accordance with this agreement, and any volunteers associated with the production, promotion and distribution of Project Gutenberg-tm electronic works, harmless from all liability, costs and expenses, including legal fees, that arise directly or indirectly from any of the following which you do or cause to occur: (a) distribution of this or any Project Gutenberg-tm work, (b) alteration, modification, or additions or deletions to any Project Gutenberg-tm work, and (c) any Defect you cause.
Project Gutenberg-tm is synonymous with the free distribution of electronic works in formats readable by the widest variety of computers including obsolete, old, middle-aged and new computers. It exists because of the efforts of hundreds of volunteers and donations from people in all walks of life.
Volunteers and financial support to provide volunteers with the assistance they need are critical to reaching Project Gutenberg-tm's goals and ensuring that the Project Gutenberg-tm collection will remain freely available for generations to come. In 2001, the Project Gutenberg Literary Archive Foundation was created to provide a secure and permanent future for Project Gutenberg-tm and future generations. To learn more about the Project Gutenberg Literary Archive Foundation and how your efforts and donations can help, see Sections 3 and 4 and the Foundation information page at www.gutenberg.org
The Project Gutenberg Literary Archive Foundation is a non profit 501(c)(3) educational corporation organized under the laws of the state of Mississippi and granted tax exempt status by the Internal Revenue Service. The Foundation's EIN or federal tax identification number is 64-6221541. Contributions to the Project Gutenberg Literary Archive Foundation are tax deductible to the full extent permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S. Fairbanks, AK, 99712., but its volunteers and employees are scattered throughout numerous locations. Its business office is located at 809 North 1500 West, Salt Lake City, UT 84116, (801) 596-1887. Email contact links and up to date contact information can be found at the Foundation's web site and official page at www.gutenberg.org/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
[email protected]
Project Gutenberg-tm depends upon and cannot survive without wide spread public support and donations to carry out its mission of increasing the number of public domain and licensed works that can be freely distributed in machine readable form accessible by the widest array of equipment including outdated equipment. Many small donations ($1 to $5,000) are particularly important to maintaining tax exempt status with the IRS.
The Foundation is committed to complying with the laws regulating charities and charitable donations in all 50 states of the United States. Compliance requirements are not uniform and it takes a considerable effort, much paperwork and many fees to meet and keep up with these requirements. We do not solicit donations in locations where we have not received written confirmation of compliance. To SEND DONATIONS or determine the status of compliance for any particular state visit www.gutenberg.org/donate
While we cannot and do not solicit contributions from states where we have not met the solicitation requirements, we know of no prohibition against accepting unsolicited donations from donors in such states who approach us with offers to donate.
International donations are gratefully accepted, but we cannot make any statements concerning tax treatment of donations received from outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation methods and addresses. Donations are accepted in a number of other ways including checks, online payments and credit card donations. To donate, please visit: www.gutenberg.org/donate
Professor Michael S. Hart was the originator of the Project Gutenberg-tm concept of a library of electronic works that could be freely shared with anyone. For forty years, he produced and distributed Project Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed editions, all of which are confirmed as Public Domain in the U.S. unless a copyright notice is included. Thus, we do not necessarily keep eBooks in compliance with any particular paper edition.
Most people start at our Web site which has the main PG search facility: www.gutenberg.org
This Web site includes information about Project Gutenberg-tm, including how to make donations to the Project Gutenberg Literary Archive Foundation, how to help produce our new eBooks, and how to subscribe to our email newsletter to hear about new eBooks.