The Project Gutenberg EBook of Speciation in the Kangaroo Rat, Dipodomys
ordii, by Henry W. Setzer
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
Title: Speciation in the Kangaroo Rat, Dipodomys ordii
KU. Vol 1 No 23
Author: Henry W. Setzer
Editor: E. Raymond Hall
Release Date: May 26, 2013 [EBook #42810]
Language: English
Character set encoding: ISO-8859-1
*** START OF THIS PROJECT GUTENBERG EBOOK SPECIATION IN KANGAROO RAT ***
Produced by Chris Curnow, Matthias Grammel, Joseph Cooper,
The Internet Archive for some images and the Online
Distributed Proofreading Team at http://www.pgdp.net
BY
HENRY W. SETZER
University of Kansas Publications
Museum of Natural History
Volume 1, No. 23, pp. 473-573, 27 figures in text, 7 tables
December 27, 1949
University of Kansas
LAWRENCE
1949
University of Kansas Publications, Museum of Natural History
Editors: E. Raymond Hall, Chairman, A. Byron Leonard,
Edward H. Taylor, Robert W. Wilson
Volume 1, No. 23, pp. 473-573, 27 figures in text, 7 tables
December 27, 1949
University of Kansas
Lawrence, Kansas
PRINTED BY
FERD VOILAND, JR., STATE PRINTER
TOPEKA, KANSAS
1949
22-6114
Subspeciation in the Kangaroo Rat,
Dipodomys ordii
By
HENRY W. SETZER
CONTENTS
PAGE Introduction 477 Methods and Acknowledgments 478 Paleontology 480 Phylogeny of the Species of the Genus 484 Dispersal of the Several Species 498 Subspeciation 499 Accounts of Subspecies 511 Dipodomys ordii 511 Dipodomys ordii richardsoni 511 Dipodomys ordii oklahomae 514 Dipodomys ordii compactus 515 Dipodomys ordii sennetti 517 Dipodomys ordii evexus 518 Dipodomys ordii medius 519 Dipodomys ordii obscurus 521 Dipodomys ordii terrosus 523 Dipodomys ordii fremonti 524 Dipodomys ordii uintensis 525 Dipodomys ordii sanrafaeli 526 Dipodomys ordii panguitchensis 527 Dipodomys ordii monoensis 528 Dipodomys ordii ordii 530 Dipodomys ordii luteolus 533 Dipodomys ordii extractus 534 Dipodomys ordii chapmani 536 Dipodomys ordii montanus 538 Dipodomys ordii cinderensis 540 Dipodomys ordii fetosus 541 Dipodomys ordii utahensis 543 Dipodomys ordii columbianus 544 Dipodomys ordii idoneus 546 Dipodomys ordii priscus 547 Dipodomys ordii celeripes 549 Dipodomys ordii cineraceus 550 Dipodomys ordii marshalli 551 Dipodomys ordii inaquosus 552 [Pg 476] Dipodomys ordii attenuatus 553 Dipodomys ordii fuscus 555 Dipodomys ordii longipess 556 Dipodomys ordii pallidus 558 Dipodomys ordii nexilis 559 Dipodomys ordii cupidineus 561 Dipodomys ordii palmeri 562 Conclusions 563 Tables of Measurements 565 Literature Cited 571 INTRODUCTION
The geographic range of the kangaroo rats, genus Dipodomys, extends from southern Canada south to the southern limits of the Mexican Tableland and from the Pacific Coast east to the eastern limits of the Great Plains in Kansas, Oklahoma and Nebraska. These animals are usually restricted to sandy soils in semiarid regions. The species Dipodomys ordii, with which this account is primarily concerned, is, to the best of my knowledge, almost exclusively confined to sandy areas.
Since 1841, when Gray gave the generic name Dipodomys to the kangaroo rats, basing the name on the four-toed species Dipodomys phillipsi, several other generic names have been applied. Fitzinger, in 1867, used the name Perodipus for those animals with five toes on the hind foot, designating Dipodomys agilis as the type of his genus. In 1890, Merriam proposed the generic name Dipodops with Dipodomys agilis as the type, apparently being unaware of Fitzinger's name, Perodipus. Trouessart, in 1897, through what was an apparent lapsus calami, applied the generic name Cricetodipus Peale to all of the species of the then known genera Perodipus and Dipodomys, but Trouessart later, 1904 or 1905, in his Supplementum, corrected this lapsus and used the names Dipodomys and Perodipus. Grinnell (1919:203) showed that some of the four-toed Dipodomys had five toes on one hind foot and four on the other and that Perodipus must fall as a synonym of the earlier generic name Dipodomys which was to be applied to all of the kangaroo rats.
Dipodomys ordii was named by Woodhouse in 1853, from specimens from El Paso, Texas, but between that time and 1919 the name ordii was used in combination with all of the generic names mentioned above (see synonymies under the accounts of the subspecies).
The nearest approach to a revision of the genus was Grinnell's (1922) "A Geographical Study of the Kangaroo Rats of California." Since that time, Hall and Dale (1939) revised the D. microps group and Durrant and Setzer (1945) reported upon the kangaroo rats of Utah. The present paper is a review of the species Dipodomys ordii. Some of the objectives in this review have been to learn: (1) What kinds of kangaroo rats are subspecies of the species Dipodomys ordii; (2) the limits of geographic range of this full species; (3) the extremes of color, and of size and shape of the skull [Pg 478] in this one species; (4) the significance of different sizes, shapes and colors; (5) the reasons for the existence, or formation, of selected subspecies; and (6) the relationships of Dipodomys ordii to other species in the genus.
METHODS AND ACKNOWLEDGMENTS
Available specimens were arranged according to geographic origin. These were segregated as to sex and then under each sex by age. Individual variation was next measured in each of several samples in which individuals were of like geographic origin, sex, age and season. Finally, comparable materials were arranged geographically for detection of variations of systematic worth. Following preliminary studies of material thus arranged, additional specimens were collected from critical areas.
When fully adult animals (see next paragraph) were segregated as to sex, and then measured, the degree of secondary sexual variation was found to be less than the degree of individual variation; therefore in the tables of indices, no distinction as to sex has been made.
The only external measurements of the animals used were those recorded by the collectors on the labels attached to the skins. These measurements were total length, length of tail and length of hind foot. Measurements of the ear have not been used since they were not in all instances recorded by collectors and since measurements of dry ears proved to be unsatisfactory. Only measurements of fully adult specimens have been used. The term fully adult is applied only to those specimens in which the auditory bulla is shiny and translucent, the permanent P4 is fully erupted and worn, and the tail is fully striped and penicillate. No one of these characters alone was accepted as proof of adulthood but only the three in combination.
The following measurements of the skull have been used in the tables:
Greatest length.—From the most anterior tip of the nasals to the most posterior projection of the auditory bullae.
Greatest breadth across bullae.—From the most lateral projection of the auditory bulla on one side to the corresponding position on the other bulla.
Breadth across maxillary arches.—Greatest breadth across arches in a plane perpendicular to the long axis of the skull.
Width of rostrum.—Width of the premaxillae and the nasals taken immediately anterior to the upper incisors (not greatest width of nasals which is attained farther anteriorly).
Length of nasals.—Maximum length of a nasal bone.
Least interorbital breadth.—Least width between the orbits immediately posterior to the lacrimal processes.
Basilar length.—From the anterior margin of the foramen magnum to the posterior border of the alveolus of one of the upper incisors.
Capitalized color terms are from Ridgway, "Color Standards and Color Nomenclature," Washington, D. C., 1912. Color determinations were made by comparing a masked area of pure color on the side of the animal with a masked rectangle of named color on Ridgway's plates in natural light always from the same angle.
Abbreviations used for specimens examined from the various collections are as follows:
AMNH—American Museum of Natural History.
BYU—Brigham Young University.
CNHM—Chicago Natural History Museum.
CM—Carnegie Museum.
CMNH—Colorado Museum of Natural History.
DJC—Dixie Junior College.
DRD—Donald R. Dickey Collection.
KU—Museum of Natural History, University of Kansas.
LACM—Los Angeles County Museum.
MHS—Collection of Myron H. Swenk.
MVZ—Museum of Vertebrate Zoology, University of California.
OU—Museum of Zoology, University of Oklahoma.
RH—Collection of Ross Hardy.
UM—Museum of Zoology, University of Michigan.
UN—Museum of Natural History, University of Nebraska.
USAC—Utah State Agricultural College.
USBS—United States Biological Surveys Collection.
USNM—United States National Museum.
UU—Museum of Zoology, University of Utah.
TCWC—Texas Coöperative Wildlife Collection.This study is based on 3,732 specimens which were assembled at the Museum of Natural History, University of Kansas, or studied at other institutions. For the loan of this material and for the opportunity afforded for its study, I am extremely grateful to the authorities of each of these institutions and to the owners of the private collections.
Acknowledgement is made to the Office of Research and Inventions of the United States Navy for assistance with the field work which permitted the acquisition of essential specimens from several of the critical geographic areas while the author was research assistant on a larger over-all project (N6 ori-164-T02) of which the determination of the geographic range of this rodent species, a potential host of tularemia, was one facet. Tularemia was not detected in this genus.
I extend my thanks also to Professor Stephen D. Durrant, of the University of Utah, for helpful corrections in the preparation of the manuscript; to Mrs. Virginia Cassell Unruh for the preparation of the drawings; to Professor E. Raymond Hall, of the University of Kansas, for guidance in the study and critical assistance with the manuscript; to Professors H. H. Lane and Worthie H. Horr for valued suggestions; to Mr. J. R. Alcorn for providing specimens for dissection when he was working under the University of Kansas endowment fund; and to the other friends and associates who have given of their time and criticism.
PALEONTOLOGY
The family Heteromyidae was defined by Wood (1935:81) essentially as follows: Cheek teeth brachydont to hypsodont and even rootless; usually six cusps per molar, three on each loph; enamel rarely divided into two plates, never reduced to one; skull light, thin and papery; mastoids inflated, mastoidal breadth often greatest, never appreciably less than zygomatic breadth; interorbital space wider than rostrum; palate nearly horizontal and little if any below level of zygomata; nasals extended beyond incisors; zygomata slender, with greatly reduced malar, almost, or quite, abutting against tympanic; frontals and parietals broad, with latter reaching, or nearly reaching, orbits; frontal trapezoidal; parietal quadrate, to pentagonal or triangular; interparietal primitively large, secondarily reduced; squamosal mostly or entirely confined to orbit; tympanic vesicular and inflated, in some forms highly inflated; mastoids inflated and bullous, reaching top of skull, and forming part of occipital surface; occipitals contracted and limited in area on occiput, but extending onto dorsum of skull; coronoid processes small, inclined caudad and lying below level of condyle; jaw small and weak with large, everted angle; tail as long as, or longer than, head and body; claws of manus elongate, fossorial, but forelimb slender; pelage usually coarse and frequently spinose; ears and eyes large; body murine in form; locomotion in many forms saltatorial.
This characterization of the family includes all of the members of the subfamilies Perognathinae, Heteromyinae and Dipodomyinae as well as the genus Microdipodops which I am disinclined to place with any of the three subfamilies. Apparently it is more closely related to the subfamily Perognathinae.
The subfamily Dipodomyinae, which contains the genera Dipodomys, Prodipodomys and Cupidinimus, might be characterized [Pg 481] after Coues' (1875) original description of the subfamily as follows: Cheek teeth progressively hypsodont, in Dipodomys becoming ever-growing; enamel progressively interrupted, eventually reduced to anterior and posterior plates; upper and lower third molars reduced in size; tooth pattern rapidly destroyed, leaving only an enamel oval; upper incisors smooth (some fossils) or grooved (living forms); progressive expansion of the auditory bullae and increase in saltatorial ability; pterygoid fossa double; calcaneal-navicular or even calcaneal-cuneiform articulation; tail tufted.
Owing to the fact that so little paleontological material is known and because even that is fragile and not easily accessible for study, knowledge of the fossil representatives has been drawn primarily from the literature, especially from Wood's (1935) account.
Heteromyids are known from the Chadron formation, of early Oligocene age, in which a single tooth was found. In the Orellan stage of the mid-Oligocene where the genus Heliscomys occurs, it is notably generalized, in comparison with other members of the family, but it may not be ancestral at all. The lower premolar is tricuspidate and the first and second molars are quadritubercular with a broad cingulum. The teeth are bunodont and brachydont, with the cusps not uniting to form lophs. Wood (1935:78) shows Mookomys formicorum (from the Arikeean) as the next heteromyid in the evolutionary sequence and postulates that this species arose from Heliscomys gregoryi. Mookomys is judged by Wood to be the common ancestral form of the perognathines and the dipodomyines.
Cupidinimus, the genus next in line, is characterized by smooth upper incisors; lower molars with incipient H-pattern; cheek teeth progressively hypsodont and lophate (but always rooted); and calcaneal-navicular articulation.
The time range of this genus is from the late Miocene (Niobrara River, Local Fauna) of Nebraska to the medial Pliocene, Thousand Creek (Hemphillian) of Nevada.
Hibbard (1937:462) described Dipodomys kansensis from the Ogallala formation (Hemphillian age) of Kansas. He redescribed his species, and made it the type of the new genus Prodipodomys (Hibbard, 1939:458), differentiating it from Dipodomys on the basis of the three-rooted p4, double-rooted m1 and m2 and the single rooted m3. It is shown to be closely allied to Dipodomys by the form and position of a large foramen posterior and labial to m3, and by the development of the masseteric ridge.
The next youngest heteromyid fossils which have been described are of the genus Prodipodomys? from Arizona. Gidley (1922:123) described Dipodomys minor from the Benson (Blancan) which Gazin (1942:486) refers to the genus Prodipodomys?. Wood (1935:156) described Dipodomys gidleyi from the Curtis (Pleistocene). Both of these species are primitive as regards dentition; that is to say, the enamel ring of the tooth is complete and lacks any sign of a break. The limb bones of D. gidleyi show lesser saltatorial ability, and therefore appear to be more primitive, than those of any living Dipodomys.
Several heteromyids which have not been assigned to any genus are known. Wilson (1939:36-37) recognized some from the Avawatz (Clarendonian) and the Ricardo (Clarendonian). Another, possibly of the genus Diprionomys?, from the Barstow (Barstovian) was described by Wood (1935:197) as follows: "The general shape of the tooth as figured strongly suggests either one of the most advanced species of Dipodomys or else a Geomyid.... It is much more advanced than are any known contemporary heteromyids, and compares fairly well with such late Tertiary and Pleistocene geomyids as have been described. It certainly is not referable to any known heteromyid genus other than Dipodomys, and should probably be called a Geomyid." Wilson (loc. cit.) refers to these specimens as Dipodomyine (?) n. gen. and sp. If these specimens referred to by Wood and Wilson are true heteromyids then a change in the phylogenetic scheme proposed by Wood (1935) would be necessary. Wilson (loc. cit.) says, referring to the Avawatz specimen, "The cheek teeth are very hypsodont but are apparently not persistent in growth,... Wide enamel breaks are present in M/1 dividing the enamel into anterior and posterior bands. The enamel of P/4 is complete in the present stage of wear, but an examination of the tooth indicates that breaks would develop with additional attrition at the buccal and lingual margins of the metalophid, and at the buccal border of the protolophid. The incisor is of the slender heteromyid type."
Wood (1935:118) in referring to the ancestry of Cupidinimus with regard to the grooving of the incisors says: "The philosophy of evolution which would prohibit its derivation from Mookomys, because of the grooved incisors in the latter genus, would require a separate line leading back at least to the Lower Miocene."
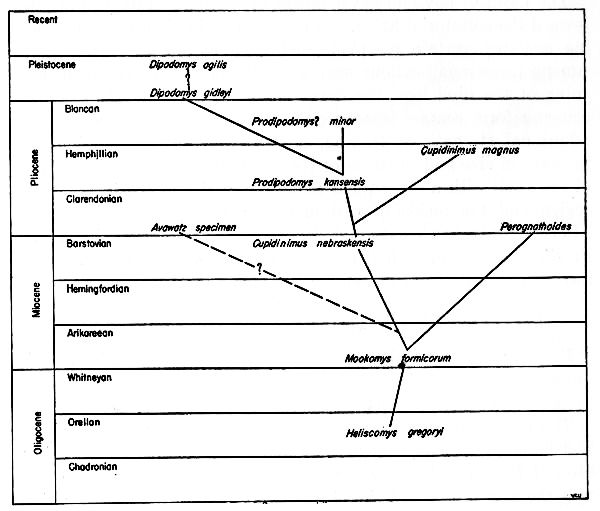
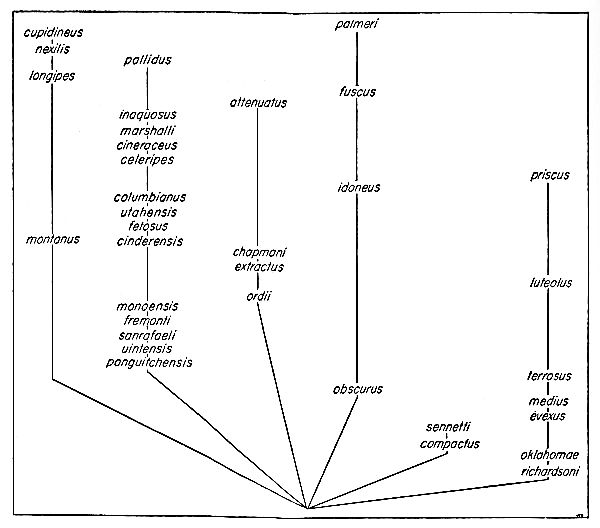
In view of the above statements, it is conceivable that additional material will be found carrying the dipodomyine line back into the [Pg 483] early Miocene. Perhaps the line involving Mookomys and Cupidinimus which was regarded by Wood as the line of descent, is merely an aberrant side branch that parallels in its structures the main line of evolution of the dipodomyines (Figure 1).
As Wilson (1939:37) says: "Indeed it is hard to recognize such a form as Cupidinimus nebraskensis as directly ancestral to Dipodomys in view of the occurrence of the much more advanced Avawatz specimen in deposits that are at most only slightly later than those in which the former is found. The kangaroo rats were apparently much farther along in their development by lower Pliocene time than heretofore supposed."
Wood (1935:78) suggested that Dipodomys gidleyi gave rise to Dipodomys spectabilis and Dipodomys ordii, and Dipodomys minor gave rise to Dipodomys compactus. However, my own study indicates that Dipodomys compactus is conspecific with Dipodomys ordii and should stand as Dipodomys ordii compactus. Consequently a different phyletic arrangement than that proposed by Wood (loc. [Pg 484] cit.) is required. Since D. compactus is more closely allied to Prodipodomys? minor than D. ordii is to D. gidleyi, it is possible that P.? minor gave rise to D. ordii and that D. spectabilis is the end product of the phyletic trend of D. gidleyi (Figure 1).
The trend of phyletic development in the dipodomyines has been toward the saltatorial habit. To acquire this habit from a scampering ancestor, certain morphological modifications were necessary. Among these modifications were a lengthening of the tail, a lengthening of the hind legs, the development of a calcaneal-navicular-ectocuneiform contact instead of a calcaneal-navicular contact for additional strength in leaping, a shortening of the forelimb, an increase in size and inflation of the mastoid and tympanic portions of the skull with a consequent reduction in size of the interparietal region and the fusion of certain of the cervical vertebrae. Late Miocene (Cupidinimus) and Pliocene (Avawatz specimen and Prodipodomys) forms had acquired certain of these morphological modifications that are present in the modern genus Dipodomys.
PHYLOGENY OF THE SPECIES OF THE GENUS
Representatives of nine species of Dipodomys were dissected in an attempt to determine the degree of specialization and the relative systematic position of each species.
The myology was found to agree in detail as to origin, insertion and innervation with that of Dipodomys spectabilis as reported by Howell (1932). The only variation noted in the muscular system was the size of the individual muscles in those animals of widely divergent body size.
Dipodomys ordii is the most generalized and Dipodomys deserti is the most specialized of the kangaroo rats (see Table 1), as judged by the osteology. Information gained by the study of the viscera of the various species supports this judgment. The visceral mass is relatively loose in D. ordii, but is markedly compact in D. deserti. This compactness appears to be brought about by the foreshortening of the mesenteries which support the entire gut and by the closer apposition of the large intestine to the caecum; both the intestine and caecum occupy a ventral position in the abdominal cavity. In Dipodomys ordii the entire visceral mass is loosely interconnected and the caecum is relatively small as compared to the tightly compact viscera and the large caecum in Dipodomys deserti. Another striking feature is the size, proportion and position of the liver. In all animals dissected, even in D. deserti, the right lobe of the liver [Pg 485] descends and forms a capsule around the anterior end of the right kidney. In the Ord kangaroo rat, the bulk of the liver lies on the right side of the body cavity. That is to say, there is a greater bulk of the liver on the right side and it is situated more dorsad than in any of the other species examined. In the most specialized condition, as in Dipodomys deserti, the bulk of the liver is almost equal on the right and left sides, and instead of having the greater bulk situated dorsally as in D. ordii it is cup-shaped, with the dorsal and ventral parts of approximately equal size and situated on almost the same transverse plane. The entire mass of the liver is concave posteriorly.
TABLE 1
Skeletal Indices of Dipodomys
Humeroradial Intermembral Crural Tibioradial Femorotarsal-metatarsal Humero-Cranial ordii 144.5 57.2 127.75 60.55 88.4 63.4 microps 138.5 56.17 132.3 57.27 90.95 60.8 panamintinus 146.1 55.3 132.0 57.5 90.5 60.8 agilis 147.0 56.05 133.65 57.25 94.55 62.65 heermanni 142.9 54.2 135.9 55.35 92.2 60.93 ingens 142.9 54.1 130.6 56.2 89.65 66.2 spectabilis 140.9 53.05 133.9 54.2 95.6 64.6 phillipsii 163.4 55.05 137.85 58.97 101.5 64.5 merriami 160.75 53.85 137.5 57.36 99.75 63.9 nitratoides 155.0 54.1 137.4 57.0 98.25 65.5 deserti 149.5 53.4 139.4 54.9 96.6 67.6 The right kidney is variable in position in reference to the left. In all species the right kidney lies anterior to the left but in some, D. deserti and D. ingens, it is markedly anterior.
In Dipodomys agilis, D. merriami and D. deserti there are small to large patches of lymphoid tissue on the caecum. These patches were not noted in any of the other species examined and I do not know their function. In the three above mentioned species, however, [Pg 486] the large intestine is shorter in proportion to the small intestine than in any other species except D. heermanni (see Table 2) and with the exception of D. heermanni, D. venustus and D. ordii the actual measurements are less.
Inasmuch as little is known of the food habits of the various species of kangaroo rats, any ascription of adaptive significance to the varying proportions of the digestive system would be only speculative.
Midgley (1938) describes the visceral anatomy of D. ordii and D. microps. Except for the differences here noted the description of the viscera as given by Midgley (loc. cit.) applies to the rest of the species studied.
TABLE 2
Visceral Measurements (in Millimeters) of Dipodomys
heermanni merriami agilis deserti ordii spectabilis venustus panamintinus ingens Large intestine 432 290 464 397 237 413 374 419 430 Small intestine 165 126 220 195 131 228 207 255 274 Percent of small to
large intestine38.2 43.4 47.5 49.2 55.2 55.3 55.4 60.9 63.7 From the differences noted in the skeleton, in the entire visceral mass, and in the shape and position of the liver it appears that as a saltator becomes more specialized skeletally, there is a concurrent compacting and aligning of the viscera into a more or less bilaterally balanced mass. It seems that this alignment is for a stabilization in leaping. It seems reasonable that the individual that has a loose and unconsolidated visceral mass, or in which the viscera or at least the heaviest part of the viscera is relatively unilateral, would be thrown slightly off balance at the end of the jump. This would place the animal at a slight disadvantage before being able to make the next jump. Howell (1944:40) comments on the fact that kangaroo rats often land off balance, "owing apparently to clumsy use of the tail." Possibly the unilaterality of the visceral mass plus a shorter tail and a more clumsy use of that organ accounts for the off balance landings which Howell has observed.
The skeleton, particularly of the appendages, shows the most [Pg 487] modification, ranging from a relatively generalized to a specialized condition. Skeletal indices, as established by Howell (1944:199) have been used in estimating the amount of such specialization.
These indices are obtained by dividing the length of one segment of a limb by the length of another segment and are expressed in percentages. The Femorotarsal-metatarsal and Cranial indices are not from Howell (loc. cit.).
The Humeroradial index (radius/humerus ×100) in the generalized animal is theoretically 100 because the humerus and radius are of the same length. In kangaroo rats, which are saltators, the index rises to more than 100 owing to the lengthening of the radial component.
The Intermembral index (humerus and radius/femur and tibia ×100) in a generalized animal is theoretically 100, but, as Howell (1944:205) points out, the index in generalized mammals is probably nearer 75. If the hind leg elongates at the expense of the forelimb the animal will be a better saltator and the skeletal elements will yield a lower intermembral index.
The Femorotibial or Crural index (tibia/femur ×100) expresses the development of the tibia as an adaptation to the saltatorial habit and in generalized animals would be expected to be 100. As an adaptation to saltation the tibia would elongate at the expense of the femur and the index would be more than 100. The degree of divergence from 100 would be an expression of the degree of saltatorial ability.
The Tibioradial index (radius/tibia ×100) in the generalized animal also would be expected to approximate 100 but it is doubtful if any living mammals, except brachiating kinds, yield an index of more than 75. In saltators, the index is low because of the elongation of the hind appendages, whereas the forelimbs do not change their length or are shortened.
The Femorotarsalmetatarsal index (tarsometatarus/femur ×100) in the generalized condition would be less than 50 and an index approaching 100 would indicate a specialization for saltation owing to the elongation of the tarsometatarsal elements.
The Cranial index (breadth across bullae/length of skull ×100) reflects the development of the auditory or mastoid region of the skull as an adaptation for more acute hearing and possibly for more delicate balance. In heteromyids, the generalized condition would be represented by an index of 50 or less, and as the width across the bullae increases, the index rises toward 100.
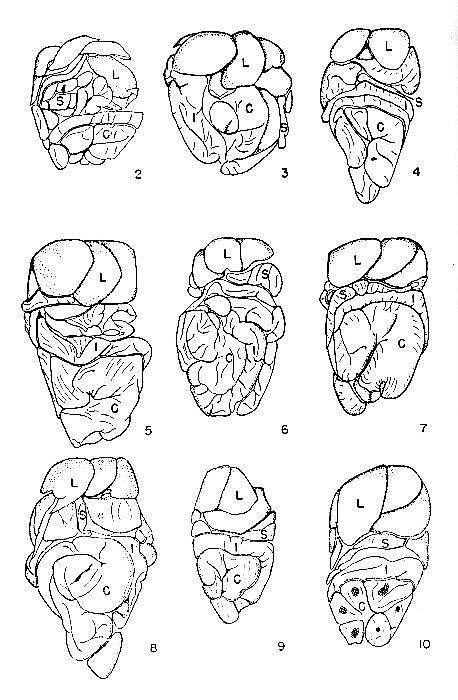
Figs. 2-10. Showing the compacting of the visceral mass; liver at the top, small intestine and caecum at the bottom. All figures approximately × 1. Fig. 2. Dipodomys ordii inaquosus, [M], adult, no. 23365, KU; 7 mi. W Fallon, Churchill County, Nevada; trapped 27 October, 1945. Fig. 3. Dipodomys panamintinus mohavensis, [M], adult, no. 22094, KU; 1-1/2 mi. N Mojave, Kern County, California; 3 February, 1948. Fig. 4. Dipodomys heermanni morroensis, [M], adult, no. 22082, KU; S side Morro Bay, 4 mi. S Morro, San Luis Obispo County, California; 25 January, 1948. Fig. 5. Dipodomys ingens, [F], adult, no. 22069, KU; 25 mi. SW Mendota, San Benito County, California; 2 February, 1948. Fig. 6. Dipodomys agilis perplexus, [F], adult, no. 22091, KU; 1-3/10 mi. N Monolith, Kern County, California; 3 February, 1948. Fig. 7. Dipodomys venustus sanctiluciae, [M], adult, no. 22071, KU; 1-1/2 mi. S Jolon, Monterey County, California; 26 January, 1948. Fig. 8. Dipodomys spectabilis spectabilis, [F], adult, no. 22110, KU; 5 mi. NE Willcox, Cochise County, Arizona; 19 January, 1948. Fig. 9. Dipodomys merriami merriami, [F], adult, no. 23366, KU; E side Carson Lake, Churchill County, Nevada; 2 October, 1945. Fig. 10. Dipodomys deserti deserti, [M], adult, no. 23364, KU; 15 mi. WSW Fallon, Churchill County, Nevada; 3 November, 1945. TABLE 3
Relative Specializations of the Species for each Index
Humeroradial Intermembral Crural Tibioradial Femorotarsal-metatarsal Cranial Average ordii 5 1 1 1 1 5 2.33 microps 1 2 4 5 4 2 3.0 panamintinus 6 3 3 3 3 1 3.1 agilis 7 4 5 6 6 4 5.3 heermanni 3 6 7 9 5 3 5.5 ingens 4 7 2 8 2 10 5.5 spectabilis 2 11 6 11 7 8 7.5 phillipsii 11 5 10 2 11 7 7.6 merriami 10 9 9 4 10 6 8.0 nitratoides 9 8 8 7 9 9 8.5 deserti 8 10 11 10 8 11 9.6 The figure 1 represents the least specialized condition for the index, while the figure 11 represents the most specialized condition. The remainder of the numbers indicate the relative degree of specialization of each species for each index.
The species that have been examined are listed in Table 3 in increasing order of specialization from top to bottom.
Usually animals of extreme morphological specialization are much restricted environmentally. Attempts to correlate the relative evolutionary position of the various species, as indicated by the degree of specialization interpreted from the indices with that of habitus has proven unsuccessful. For example, Dipodomys merriami, which is third from the top in the list as arranged above, is neither restricted to loose sandy soil as is D. deserti nor to brush as are D. agilis and D. venustus. D. merriami does, nevertheless, inhabit a variety of habitats from loose sandy soils to rather hard rocky ground. Throughout the genus there is, however, a general trend toward increased specialization as the animals adopt the more open desert environment, as is indicated by the elongation of the tail and hind appendage and increase in size of the auditory region of the skull. A marked difference is noted in the size of the pinna of the ear in the various species. Generally, those species having small pinnae inhabit open desert country while those with large pinnae inhabit brushy country. This is in direct contradistinction to the hares and rabbits in which the small-eared kinds are brush dwellers whereas the large-eared kinds are inhabitants of open country. Three possible explanations for hares and rabbits having this specialization of the pinnae are: (1) To enable the open-dwelling animals with the larger pinnae to hear more readily the approach of an enemy when it is yet far away, while the brush-living forms, which rely for escape on a short dash into cover, do not need so large a "funnel"; (2) large pinnae have been developed by those animals which live in the open desert as an aid in dissipating the body heat; (3) large pinnae in brush-dwelling animals would be a decided disadvantage in rapid movement through the brush. Grinnell (1922:20) points out that animals with large pinnae usually have small auditory bullae and conversely, animals with small pinnae have large bullae. This compensatory factor, implying an auditory function, appears to be inoperative in D. panamintinus mohavensis which has small ears and small bullae and in D. elephantinus which has large ears and large auditory bullae. Grinnell (loc. cit.) suggests that several additional factors enter into the problem, such as the amount of digging each species must do to gain safety, the texture of the soil for burrowing, the extent of forage area and the type of cover in connection with the mode of attack of predators. Of these factors, perhaps the most important are the two first mentioned.
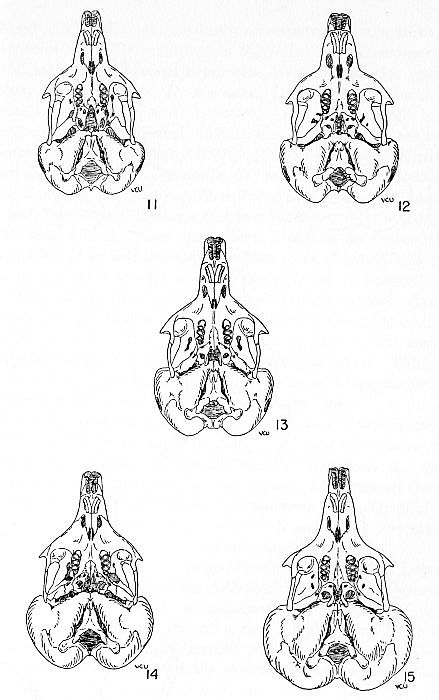
Figs. 11-15. Ventral views of skulls showing the degree of development of the auditory bullae and the configuration of the pterygoid fossae. All figures approximately × 1. Fig. 11. Dipodomys ordii compactus, [M], adult, no. 646, TCWC; 19 mi. S Port Aransas, Mustang Island, Nueces County, Texas; 24 April, 1939. Fig. 12. Dipodomys ordii oklahomae, [F], adult, no. 265456, USBS; 2-1/4 mi. S Norman, Cleveland County, Oklahoma; 21 March, 1934. Fig. 13. Dipodomys ordii richardsoni, [F], adult, no. 15995, KU; 1 mi. S Lamar, Prowers County, Colorado; 8 September, 1945. Fig. 14. Dipodomys ordii nexilis, [F], adult, no. 149941, USBS; 5 mi. W. Naturita, Montrose County, Colorado; 20 July, 1907. Fig. 15. Dipodomys deserti deserti, [F], adult, no. 18670, KU; 14 mi. WSW Fallon, Churchill County, Nevada; 3 November, 1945.
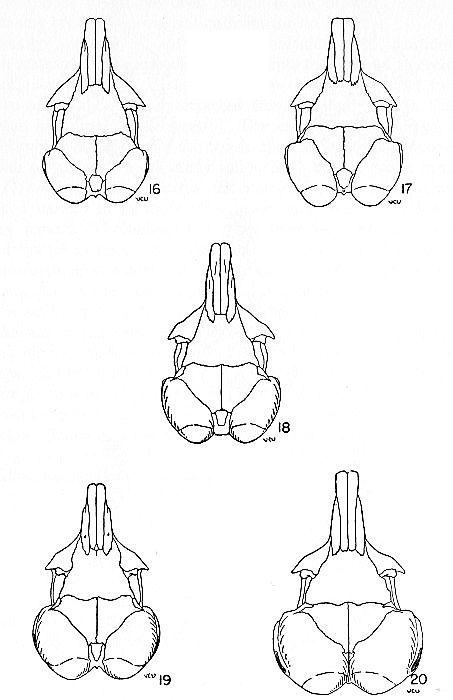
Figs. 16-20. Dorsal views of skulls showing the degrees of inflation of the auditory bullae and the correlation of large bullae with small interparietal. All figures approximately × 1. Fig. 16. Dipodomys ordii compactus, for data see Fig. 11. Fig. 17. Dipodomys ordii oklahomae, for data see Fig. 12. Fig. 18. Dipodomys ordii richardsoni, for data see Fig. 13. Fig. 19. Dipodomys ordii nexilis, for data see Fig. 14. Fig. 20. Dipodomys deserti deserti, for data see Fig. 15. Wood (1935:155), on the basis of structure of the teeth, listed the species which he examined in the following increasing order of specialization: Dipodomys compactus (now Dipodomys ordii compactus), D. nitratoides, D. merriami, D. ordii, D. agilis, D. herrmanni, D. spectabilis, and D. deserti. This arrangement is at variance with that of Grinnell (1922) who listed the species in order of increasing specialization as: Dipodomys herrmanni, D. panamintinus, D. ingens, D. spectabilis, D. merriami, D. nitratoides, D. ordii, D. agilis, D. venustus, D. microps and D. deserti. As noted, the only agreement between the two arrangements is the placing of D. deserti as the most specialized. Relying on skeletal indices alone, I would accord the same position to D. deserti but would not arrange the other species as have either Wood or Grinnell.
In this study, the amount of specialization of each species, as indicated by the skeleton, was determined by assigning consecutive numbers from 1 to 11 to each species in its place in each index, and then totaling and averaging these arbitrary numbers (Table 3). It will be noted that there is a tendency for each species to occupy the same relative position in each of the indices.
It is felt, however, that a more nearly correct arrangement, according to degree of specialization, is obtained by using the six skeletal indices plus the information obtained from the study of the viscera. On this basis the species may be arranged from least to most specialized as follows: Dipodomys ordii, D. microps, D. panamintinus, D. agilis, D. herrmanni, D. ingens, D. spectabilis, D. phillipsii, D. merriami, D. nitratoides and D. deserti.
Grinnell (1922:95-96) arranged the Recent species of Dipodomys in nine groups. Davis (1942:332) also proposed an arrangement of nine groups in which he combined the Compactus and Ordii groups of Grinnell, established a new Elator group by removing Dipodomys elator from Grinnell's Phillipsii group, and in linear arrangement, Davis shifted the Spectabilis and what remained of the Phillipsii groups to new positions. Burt (1936:152) arranged Grinnell's groups into three (unnamed) groups solely on the basis of the structure of the baculum. In the arrangement proposed by Grinnell, two of his nine groups contained only one species each, one other, the Microps group, has since been shown to contain only one species and another, the Compactus group, contained only kinds which are, by me, regarded only as subspecies of Dipodomys ordii. To my mind neither Davis nor Burt added to or fundamentally changed the basic concepts as set forth in 1922 by Grinnell. Owing [Pg 494] to the paucity of material at that time, especially from areas of intergradation, Grinnell's groupings and arrangement were as nearly natural as could be expected. With the accumulation of additional material and with the knowledge that certain kinds treated by Grinnell as full species are in actuality subspecies, it is felt that the several species of kangaroo rats can best be arranged in six groups which, from the least to the most specialized, are as follows:
Ordii Group.—Composed of the subspecies of Dipodomys ordii and Dipodomys microps. Grinnell placed these two species in separate groups; Burt on characters of the baculum alone placed D. microps with Dipodomys deserti and Dipodomys spectabilis. Within the single species D. ordii, I find that the difference in shape and size of the baculum between the subspecies of D. ordii is as great as the difference which Burt (1936:154-155) found between the full species D. agilis and D. microps. The characters of the baculum are an aid, but not in and of themselves an adequate basis, for determining the natural relationships of the groups of species. Certainly the remainder of the morphological differences between D. deserti and D. microps are so great that I doubt that the similarity in the baculum is significant, at least in this one instance. The chisel-shaped lower incisors of D. microps appear to be a specialization. They may enable D. microps to utilize more woody types of vegetation than can D. ordii. Both species occupy the same territory over much of their geographic range, probably because they eat different kinds of food.
Panamintinus Group.—Composed of all the now known subspecies of Dipodomys panamintinus and the species Dipodomys stephensi, if the latter is a full species. This group was included by Grinnell in the Heermanni group, with which it agrees in broadness of the maxillary arches and the configuration of the penis bone, but on the basis of the degree of specialization, as indicated by the indices (see Table 1), I feel that the Panamintinus group is more properly placed after the Ordii group and should be separated from the Heermanni group. Actually, animals in the Panamintinus group are intermediate between those of the Ordii and Heermanni groups.
Heermanni Group.—Composed of the subspecies of Dipodomys heermanni and Dipodomys agilis, the species Dipodomys ingens, Dipodomys venustus and Dipodomys elephantinus. D. [Pg 495] ingens even though larger in linear measurements than any of the other kinds included in this group, has almost the same degree of specialization as does D. heermanni. D. agilis, even though somewhat less specialized than the other kinds placed in this group, by the general nature of the indices, by the form of the visceral mass and to some degree by the shape of the baculum, shows itself properly to belong with this group. The species D. venustus, judged by characters of the visceral anatomy, also belongs here rather than with some other group or as a separate group. From the appearance of the visceral mass, D. venustus is somewhat more specialized than either D. heermanni or D. agilis, but D. venustus does show its affinities with this group. The species D. elephantinus has not been examined as thoroughly as have the other species but the external morphology and the configuration of the cranium place it with this group.
Spectabilis Group.—Composed of the subspecies of Dipodomys spectabilis. In two of the six indices, D. spectabilis shows a high degree of specialization toward saltation, but in the other four indices it shows a relatively low degree of specialization or is average for the genus. Burt (1936:155) placed D. spectabilis with D. deserti on the basis of the baculum alone. I have not examined D. nelsoni but place it with this group as did also Grinnell and Davis.
Merriami Group.—Composed of the subspecies of Dipodomys merriami, Dipodomys nitratoides and Dipodomys phillipsii, and the species Dipodomys platycephalus, Dipodomys margaritae, Dipodomys insularis, Dipodomys mitchelli, Dipodomys ornatus and Dipodomys elator. I have not examined five of these species. However, the indices and characters of the viscera indicate that the three species first mentioned are closely allied. Owing to the lack of known intergradation between the three, I judge that they should be retained as full species, but the difference in degree of morphological specialization is no more than would be expected between subspecies. I have examined no specimens of Dipodomys elator but from what I know of its morphology, I think that Grinnell better indicated its relations in allying it with D. phillipsii than did Davis in erecting a new group for it on the basis of linear measurements.
Deserti Group.—Composed of Dipodomys deserti which has only two subspecies. In all morphological respects, D. deserti is the most specialized species in the genus as shown by the [Pg 496] reduced number (4) of toes on the hind foot, the bilateral arrangement of the viscera, the extreme development of the auditory region of the skull and by developing, early in life, the hiatus in the enamel wall of each molariform tooth.
The parallel arrangement below emphasizes the differences and similarities between Grinnell's (1922) arrangement and the one proposed in the present paper.
Grinnell's arrangement Present arrangement Heermanni Group Heermanni Group Dipodomys heermanni Dipodomys heermanni Dipodomys morroensis Dipodomys agilis Dipodomys mohavensis Dipodomys ingens Dipodomys leucogenys Dipodomys venustus Dipodomys panamintinus Dipodomys elephantinus Dipodomys stephensi Dipodomys ingens Spectabilis Group Spectabilis Group Dipodomys spectabilis Dipodomys nelsoni Dipodomys spectabilis Dipodomys nelsoni Phillipsii Group Now in Merriami Group Below Dipodomys phillipsii Dipodomys perotensis Dipodomys ornatus Dipodomys elator Merriami Group Merriami Group Dipodomys merriami Dipodomys merriami Dipodomys nitratoides Dipodomys nitratoides Dipodomys platycephalus Dipodomys platycephalus Dipodomys margaritae Dipodomys margaritae Dipodomys insularis Dipodomys insularis Dipodomys mitchelli Dipodomys mitchelli Dipodomys phillipsii Dipodomys ornatus Dipodomys elator Ordii Group Ordii Group Dipodomys ordii Dipodomys ordii Dipodomys microps Compactus Group Now in Ordii Group Above Dipodomys compactus Dipodomys sennetti Agilis Group Now in Heermanni Group Above Dipodomys agilis Dipodomys venustus Dipodomys elephantinus Microps Group Now in Ordii Group Above Dipodomys microps Dipodomys levipes Deserti Group Deserti Group Dipodomys deserti Dipodomys deserti Were in Heermanii Group Above Panamintinus Group Dipodomys panamintinus Dipodomys stephensi Names of the subspecies are omitted from the groups named above and only the names of full species, as understood by Grinnell and as understood now, have been used. It will be noted that the phylogenetic order follows that of Grinnell rather than the one proposed herein.
The fossil record of the kangaroo rats is so scanty that one can but speculate on the evolutionary sequence. Wood (1935) presented a diagnosis of the early phyletic history up to and through Cupidinimus; this is probably as correct as can be made. I cannot, however, share his view that the recent species of Dipodomys have originated from a descendant of Cupidinimus nebraskensis; instead, I think that the Recent species originated from some unknown ancestor in the southwest.
Fig. 21. Diagrammatic representation of the relationships and history of the Recent species Dipodomys.
In view of the foregoing evidence it seems best to estimate the relationships and history of the various species and groups of species only as far back as the early Pleistocene (see Figure 21). Inasmuch as faunas of fossil mammals from the mid-Pleistocene contain few, if any, Recent species (see Hibbard, 1937:193), the living species of Dipodomys have probably had a geologic history no longer than the period of time which has elapsed since the middle Pleistocene, or at the earliest the early Pleistocene. Of the Recent species, only Dipodomys agilis is known as a fossil; it was found in the late [Pg 498] Pleistocene tar pits of California. Under the heading "Dipodomys near ingens," however, Schultz (1938:206) recorded remains of kangaroo rats from the tar seeps of McKittrick in the San Joaquin Valley of California.
DISPERSAL OF THE SEVERAL SPECIES
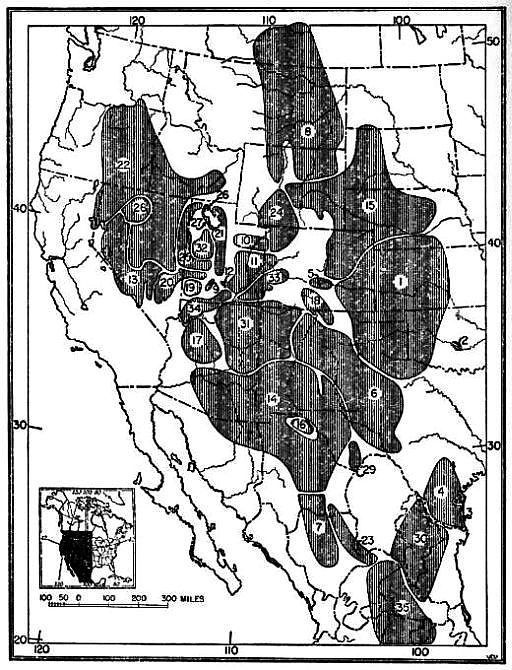
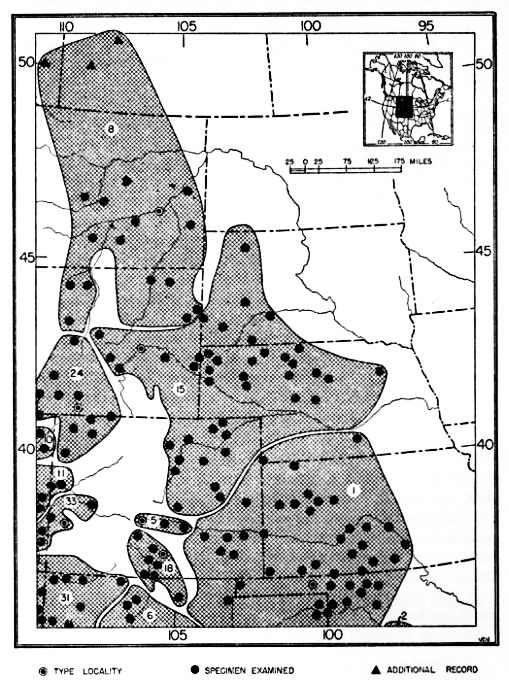
If we assume the region of origin and center of dispersal of a group of animals to be the one in which the greatest numbers of the most specialized species of a given genus are found, then the northern Tableland of Mexico and the adjoining region of the United States in southeastern California and southwestern Nevada is the region of origin and the center of dispersal for the genus Dipodomys. Dipodomys deserti, Dipodomys merriami, Dipodomys panamintinus, Dipodomys microps, Dipodomys phillipsii and Dipodomys ordii are found in the region mentioned. That the aforementioned region may be the center of differentiation for this genus is further indicated by: First, the finding, in this region, of saline deposits of Cenozoic (Miocene) age, indicating aridity, which is thought to have been one of the essential stimuli for the development of the saltatorial habit in the genus Dipodomys; second, the recovery of the advanced heteromyids from the Avawatz and Ricardo of the Clarendonian (Pliocene) of this same region; and third, the present abundance and diversification of kangaroo rats in this same geographic region which has been more or less arid since Miocene time.
A secondary center of evolution has been the low, hot interior valleys and adjacent foothills of central California where Dipodomys ingens, Dipodomys heermanni, Dipodomys venustus, Dipodomys agilis, Dipodomys elephantinus and Dipodomys nitratoides are now found. Although there are as many species as in the principal center of origin, the amount of specialization and adaptive radiation in California is not so great. Probably during the Quaternary, when the process of mountain building was actively under way the animals that had reached central California from the parental center became isolated by the emergence of the Tehachapi Mountains. This mountain range separated the California animals from populations farther south and east. As a result, D. nitratoides was differentiated from D. merriami, and D. heermanni underwent an evolution of its own which resulted in animals having either four or five toes on the hind foot. At the same time Dipodomys ingens developed there and has since been undergoing an evolution parallel to that of the large-sized species, Dipodomys spectabilis. The two species have paralleled each other not only in large size but to some [Pg 499] extent in habits such as building large mounds that are kept free of vegetation and in occupying areas of rather hard clayey soil. Structurally, however, D. ingens has not yet become quite so specialized as D. spectabilis, probably because D. ingens has had less time in which to become so. A second species, if it is a full species, Dipodomys elephantinus, has also been isolated in central California but has not attained so high a degree of specialization as D. ingens. It is interesting to note that in each of the two stocks, two large-sized species have been evolved. In the parental stock the two species are Dipodomys deserti and Dipodomys spectabilis; the former is the most specialized species in the genus. In the stock isolated in California, however, even though two large species have been formed they are still below the average in degree of specialization for the genus. As noted elsewhere in this paper, the species from these low, hot valleys, excepting D. nitratoides, are all closely related one to another. Dipodomys venustus and Dipodomys elephantinus are either closely related species or possibly only subspecies of one species, Dipodomys agilis.
It is worthy of note that as the distance away from the center of differentiation increases, the number of species decreases. For example, in the northern Great Basin there are only two species (Dipodomys ordii and Dipodomys microps) and farther eastward, on the eastern side of the Rocky Mountains, there is only the one species, Dipodomys ordii. In north-central Texas, Dipodomys elator, perhaps a relict species, is found occupying an area farther east than that occupied by Dipodomys ordii at that latitude.
Dipodomys ordii, Dipodomys phillipsii and Dipodomys merriami occupy the southern portion of the range of the genus. Instead of being generalized at this southern part of the periphery of the range as are the kinds found on the other parts of the periphery of the range of the genus, these three southern kinds are notably specialized there in the south. The subspecies D. o. palmeri which occurs in the area, is the most specialized of the species Dipodomys ordii; and Dipodomys phillipsii and Dipodomys merriami stand high in the scale of specialization with respect to the other species of the genus. The reason for this is not clear.
SUBSPECIATION
Dipodomys ordii is, without question, a valid species if one accepts Mayr's (1942:120) definition that "Species are groups of actually or potentially interbreeding natural populations, which are reproductively isolated from other such groups." D. ordii is not [Pg 500] known to hybridize with other species where their geographic ranges are adjacent or overlap. The first part of the definition "actually or potentially interbreeding populations" is substantiated by the 35 recognizable subspecies which can be defined as "a complex of interbreeding and completely fertile individuals which are morphologically identical or vary only within the limits of individual, ecological and seasonal variability. The typical characters of this group of individuals are genetically fixed and no other geographic race of the same species occurs within the same range" (after Rensch, 1934; from Mayr, 1942:106). Thus we find that certain populations of individuals differ from others and that in geographic areas between two of these populations, individuals (intergrades) are found which resemble those of both populations. In another instance, a population may be geographically isolated yet in its characters it may be recognizable as a subspecies without actual intergradation because of slight degrees of difference, or a group may be different from another without being geographically separated and may or may not show intergradation.
Subspeciation in Dipodomys ordii almost certainly has been effected, by means of mutations, under the influence of natural selection. Natural selection enhanced by geographic and ecologic isolation, probably has retained mutations of evolutionary significance, thus permitting the development of the many recognizable subspecies.
In the subspecies of Dipodomys ordii the color ranges from pale to dark. The difference in color is as pronounced as that between the full species D. deserti and D. heermanni. The lightest-colored subspecies are Dipodomys ordii celeripes, D. o. extractus and D. o. compactus; the darkest are D. o. obscurus and D. o. palmeri.
There is a marked tendency for intergrades between a light-colored subspecies, such as D. o. celeripes, and a dark-colored kind, such as D. o. utahensis, to show varying degrees of blending in color. The insular population, D. o. compactus, has, however, two distinct color phases, a light phase and a slightly darker phase, and shows no tendency toward blending. In other kinds of mammals, blending of color is known to be the result of the action of multiple alleles, but in the insular kangaroo rat (D. o. compactus) the color appears to be the result of either a reduced multiple allelomorphic complex or even a unit factor. The two color phases of this insular subspecies, which might be an expression of a unit factor, more probably are specializations in which the multiple alleles for color [Pg 501] have been reduced. The probability that there is either a unit factor or a reduced number of alleles at work is suggested by the taking of more dark-colored than light-colored animals and by the absence of blending of color. This insular population has undoubtedly been derived from the mainland kangaroo rat, D. o. sennetti, which has the usual range of variation but, to my knowledge, there are no individuals of D. o. sennetti so light as the darkest animals of D. o. compactus from the islands.
Populations from a given locality are remarkably stable in color except the animals from Samalayuca, Chihuahua, which vary in color from individuals almost as light as D. o. compactus to animals that approach D. o. ordii in darkness of pelage.
The subspecies of D. ordii show no noticeable variation in the extent of the hip stripe, supraorbital and postauricular spots, basal white ring of the tail, lateral stripes of the tail or the extent of white on the venter and feet. There is, however, variation in the degree and extent of the arietiform facial markings. In Dipodomys ordii utahensis, D. o. cupidineus, D. o. obscurus and D. o. fuscus these markings are pronounced. In D. o. celeripes, D. o. pallidus, D. o. compactus and D. o. attenuatus these markings are either obliterated or nearly so.
In Dipodomys ordii, color does not seem to be correlated with amount of moisture or geography, but rather with color of soil. For example, all animals from the Bonneville Basin of western Utah, are light colored as are the soils; animals from the San Rafael Desert of eastern Utah are reddish, as is the soil. More striking extremes of this are shown by D. o. compactus of Padre and Mustang islands, Texas, which is pale-colored as is the sand on which it lives, and D. o. medius from east-central New Mexico and western Texas, which is reddish as is the soil there, which is derived from Permian rocks. In localities where alkaline soils are present, kangaroo rats may be found with a roseaceous cast to the pelage as a result of the action of the alkaline salts on the pigment of the hair. The roseaceous color is lost when the animal sheds the old pelage.
In the dorsal and ventral stripes of the tail, I find as much variation in the species D. ordii as Grinnell (1922:Fig. E, p. 14) recorded in the whole genus. In D. o. obscurus, D. o. fuscus and D. o. utahensis the stripes are complete to the distal end of the tail and dark, whereas in D. o. pallidus and D. o. celeripes the ventral stripe is either absent or nearly so and the dorsal stripe is pale.
Color as a taxonomic character is valuable in a broad sense, and [Pg 502] is useful in placing an individual or a group of individuals in the subspecies to which they pertain. In most subspecies studied, color was quite uniform throughout the range of the animals, but in D. o. ordii and D. o. columbianus color is so variable that cranial features were relied on almost exclusively for the final diagnosis.
Among the subspecies of Dipodomys ordii there is relatively little variation in the length of the head and body. The smallest measurement is 95.5 mm. in D. o. idoneus and the largest is 118.3 mm. in D. o. richardsoni. The shortest tail is found to be 112.0 mm. in D. o. celeripes and the longest is 154.7 mm. in D. o. terrosus. The length of the hind foot varies from 35.0 mm. in D. o. idoneus to 44.5 mm. in D. o. nexilis.
Allen's Rule is not operative in the species D. ordii. According to this rule, shorter tails and smaller feet in conjunction with a large body would be expected as the more northerly limits of the species are approached, and conversely, smaller body and larger appendages would be expected as the southerly limits of the species are approached. This is not the case, however, since the subspecies D. o. terrosus ranges farthest north and has the longest tail, whereas D. o. celeripes, found in the central part of the range of the species, has the shortest tail. Again, in regard to the hind foot, the shortest is found in D. o. idoneus which is at the extreme south of the range of the species, whereas the longest hind foot is found in D. o. nexilis which occupies a nearly central position in the range.
Long tail and long hind foot would seem to be specializations for saltation and the two would be expected to be correlated. Actually there is no significant correlation in D. ordii. D. o. celeripes, in which the hind foot is near the mean for the species (39.8 as opposed to the mean of 40.7), has the shortest tail. D. o. compactus has a short tail (117.0 mm.) but a medium-sized hind foot. D. o. nexilis and D. o. terrosus have both a long hind foot and long tail.
Cranial measurements vary less, probably because one person can measure a series with a uniformly subjective error. External measurements, however, are liable to a greater degree of subjective error. The total length of the skull varies from 35.4 mm. in D. o. attenuatus to 41.3 mm. in D. o. terrosus. In no one series of adults from one locality, however, is the variation so marked as it is for the species as a whole. The usual range of variation in length of skull in any given series is not, as a rule, more than 2.5 mm.
Cranial indices (breadth across bullae/length of skull × 100) as established for random samples of the different species of the genus [Pg 503] (exclusive of D. ordii) ranged from 60.8 to 67.6. In the subspecies of D. ordii the same index varies from 59.7 to 65.2 with an average of 63.4. In other words, the degree of specialization indicated by this one index, in a few subspecies of D. ordii, is almost as great as that in Dipodomys deserti, which on the basis of total morphology appears to be the most specialized species in the genus. Also, on the basis of this same index, some subspecies of D. ordii are more generalized than is any other species in the genus.
There is a general tendency for the nasals to decrease in length and the rostrum to decrease in width as the southern limits of the range of D. ordii are approached. In ascertaining the decrease in length of the nasals an index was obtained as follows: nasals/interorbital width × 100 (see Table 4). The width of the rostrum, however, does not decrease in the same degree, nor at the same rate, as does the length of the nasals. This decrease in length of the nasals and in width of the rostrum may be correlated with the mean annual relative humidity of the environment. It is known (Howell and Gersh, 1936:8) that desert rodents, more exactly kangaroo rats, have a water retention mechanism in the kidneys and walls of the urinary bladder which enables them more efficiently to conserve metabolic water. The significance of the decrease of the area of the nasal mucosa, which seems to be related to relative aridity, is not yet properly understood.
In no cranial feature other than shortened nasals and narrowed rostrum, does Dipodomys ordii show a gradation such that it might be termed a cline. Other parts of the skull that were measured do not vary greatly.
Perhaps the greatest amount of variation in the skull is in features which are not readily measurable by the usual physical means. The shape and size of the pterygoid fossae vary from almost round to rather ovoid in a given series of animals from one locality; the size and configuration of the zygomatic arches vary from slender to robust and from straight to curved laterally; the size of the lacrimal processes varies much in any given series, as do also the degree of expansion of the distal end of the nasals, the convexity of the braincase and the curvature of the upper incisors. In all instances where these features varied much, one size or shape was more pronounced in the series than any other size or shape. Thus, when comparisons were made, the size and certain shapes were the criteria used in assigning the animals under consideration to a given subspecies.
Subspeciation in Dipodomys ordii seems to have been influenced [Pg 504] by water barriers. It is known (Grinnell, 1922:28) that kangaroo rats lack the ability to swim. Large stable rivers such as the Colorado, Snake and Columbia serve as effective barriers to further dispersal of kangaroo rats. Streams that freeze over in the winter months, however, are not efficient barriers. This is indicated by the "blending" of morphological characters of D. o. nexilis and D. o. sanrafaeli along the Green River which freezes over.
Any mountain which has vegetational belts above the Transition Life-zone would serve as a barrier to the dispersal of these animals. The Uinta Mountains, lying in an east-west direction, are interposed between the ranges of D. o. priscus and D. o. uintensis. The high Wasatch Mountains and their associated outliers, lying in a north-south direction in Utah, serve as an efficient barrier to the east-west movement of kangaroo rats and as a result, the subspecies east of the mountain mass are remarkably different from those to the west.
TABLE 4
Proportionate Decrease of Nasals
Width of rostrum Length of nasals Least interorbital width Nasals
Interorbital × 100terrosus 4.1 14.75 13.5 91.6 luteolus 4.35 13.9 12.95 93.1 evexus 4.3 14.35 13.75 94.8 montanus 4.1 13.5 12.65 93.8 ordii 3.5 13.3 13.0 97.7 idoneus 3.7 13.2 13.75 103.5 palmeri 3.3 12.8 13.0 101.1 The first three columns represent the actual measurements of the various elements; the fourth column is the index established.
Six different complexes (groups) of subspecies of D. ordii have probably arisen as a result of geographical separation.
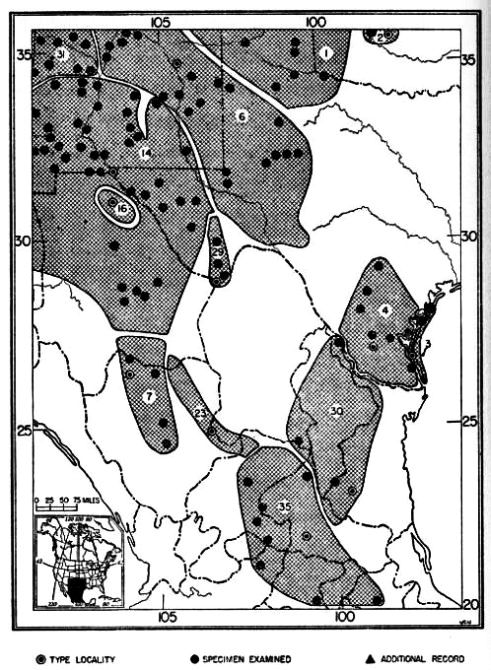
The Great Plains complex consisting of D. o. richardsoni, D. o. oklahomae, D. o. evexus, D. o. terrosus, D. o. luteolus, D. o. priscus[Pg 505] and D. o. medius are, with the exception of D. o. priscus, inhabitants of the high plains grassland habitat. D. o. priscus inhabits the red Desert of Wyoming.
The Gulf Coast complex, comprising D. o. sennetti and D. o. compactus are separable from all others by small auditory bullae and short tail. D. o. compactus probably has differentiated from D. o. sennetti since the cutting off, by wave action, from the mainland, of the islands on which D. o. compactus lives.
The Mexican complex consisting of D. o. obscurus, D. o. fuscus, D. o. idoneus and D. o. palmeri have probably differentiated by natural selection acting on fortuitous variations, but I lack first hand knowledge of the region concerned.
The southwestern complex consists of D. o. chapmani, D. o. extractus, D. o. attenuatus and D. o. ordii. D. o. attenuatus and D. o. chapmani are subspecifically distinct owing to geographic isolation, although both kinds show intergradation where their ranges approach that of D. o. ordii.
The western desert complex, composed of D. o. monoensis, utahensis, cineraceus, columbianus, cinderensis, fetosus, celeripes, marshalli, inaquosus, pallidus, panguitchensis and fremonti was isolated from the other complexes of D. ordii by the Quaternary uplift of the Wasatch Mountain mass, consisting of the Wasatch, Fish Lake and San Pitch mountains and the Wasatch, Aquarius, Paunsaugunt and Kaiparowits plateaus, and the concurrent reëstablishment of drainage systems. The drainages are those of the Colorado and Columbia rivers and that of the Snake River from Blackfoot, Idaho, to the junction with the Columbia. D. o. fremonti has been isolated on the upper reaches of the Fremont River which arises from the eastern side of the Wasatch Divide. D. o. panguitchensis has been isolated in Panguitch Valley as a result of the canyons formed by the Sevier River in Utah. D. o. cineraceus, although its subspecific and insular status are in doubt, appears to have been isolated on Dolphin Island, Great Salt Lake, Utah.
The intermontane complex consisting of D. o. montanus, longipes, cupidineus, nexilis, sanrafaeli and uintensis, like the western desert complex, has become separated from the remainder of the subspecies of the species D. ordii by Quaternary geologic events. D. o. cupidineus has been cut off by the gorges of the Colorado River to the south and the Virgin River to the north. D. o. sanrafaeli is separated from D. o. uintensis by the Tavaputs Plateau and by the Roan and Book cliffs, and is separated from the range of D. o. nexilis by [Pg 506] the Colorado River although there is intergradation between D. o. nexilis and D. o. sanrafaeli. D. o. longipes has been separated from the rest of this intermontane complex by the San Juan and Colorado rivers, but to the east it intergrades freely with adjacent subspecies. D. o. montanus has been relatively isolated in the San Luis Valley of Colorado and New Mexico, but in the southern part of its range it does show intergradation with other subspecies.
The complexes mentioned above are represented graphically in Figure 22, in a way that expresses some of my ideas as to their genetic relationships.
The indices used to determine the amount of specialization that each complex of subspecies has undergone are as follows:
The Body index (head and body/length of tail × 100) is the expression of the elongation of the tail as an organ of balance while the length of the head and body remain relatively constant. As the tail elongates the index decreases and as the tail becomes shorter the index increases.
The Pedal index (hind foot/head and body × 100) is the expression of the development of the hind foot as an element essential for the saltatorial habit. As the hind foot elongates the index will increase; elongation of the hind foot is interpreted as a specialization.
The Cranial index (breadth across bullae/length of the skull × 100) reflects the degree of development of the tympanic or mastoid region, or both, and is thought to be an adaptation for more acute hearing and possibly for more delicate balance. Inflation of the tympanic bullae is thought to be a specialization. As the auditory bullae become more inflated, the index increases toward 100.
The Bullar index (width of maxillary arches/breadth across bullae × 100) also expresses the degree of inflation of the auditory bullae. In a generalized mammal, at least in the heteromyids, the index would be 100, but as the auditory bullae become larger the index will decrease from 100.
In attempting to arrange the subspecies of D. ordii according to degree of specialization, the geographic positions of the subspecies have been considered along with the information provided by the above-mentioned indices. These indices were used in the same way as were the indices for the species of the genus. In Tables 5 and 6 and in the accounts and maps the subspecies are arranged from the least specialized to the most specialized.
TABLE 5
Indices for the Subspecies of DIPODOMYS ORDII
Body Pedal Cranial Bullar richardsoni 88.85 34.35 60.95 88.25 oklahomae 86.75 35.5 61.7 90.25 compactus 127.7 37.25 59.75 88.35 sennetti 94.25 34.0 62.85 85.95 evexus 80.1 35.7 60.5 92.9 medius 80.4 33.7 63.55 85.9 obscurus 62.95 86.4 terrosus 75.25 35.05 61.6 86.85 fremonti 80.55 34.7 62.9 85.5 uintensis 77.2 35.3 62.3 86.0 monoensis 85.4 36.4 63.4 85.6 ordii 79.05 37.6 62.75 86.9 luteolus 75.0 37.05 62.35 86.3 extractus 83.65 34.35 64.3 84.25 chapmani 75.05 36.35 62.9 85.65 montanus 80.4 36.15 64.25 82.5 cinderensis 85.1 37.2 65.15 84.75 fetosus 81.8 38.85 63.95 83.95 utahensis 80.2 36.95 64.45 84.35 columbianus 78.5 37.55 64.25 84.9 idoneus 72.3 36.6 64.2 85.0 priscus 74.9 39.45 62.3 84.95 celeripes 91.85 38.75 65.0 84.25 cineraceus 75.5 39.1 63.9 84.8 marshalli 81.5 37.3 65.2 83.0 inaquosus 78.05 37.9 64.25 83.05 attenuatus 73.5 37.35 64.0 83.4 fuscus 79.8 39.0 64.3 83.2 longipes 75.7 37.1 64.3 82.75 pallidus 76.9 40.75 64.35 84.65 nexilis 77.1 40.7 64.95 78.45 cupidineus 73.15 39.1 64.1 80.85 palmeri 72.25 37.15 65.1 80.45 TABLE 6
Numerals (derived from Table 5) are Indicative of the Relative Degree of Specialization of the Subspecies of DIPODOMYS ORDII
Body Pedal Cranial Bullar Average richardsoni 4 3 3 4 3.5 oklahomae 5 8 5 2 5.0 compactus 1 19 1 3 6.0 sennetti 2 2 10 10 6.0 evexus 15 8 2 1 6.5 medius 12 1 15 11 9.75 obscurus 12 7 10.0 terrosus 25 6 4 6 10.25 fremonti 11 5 12 14 10.5 uintensis 20 7 7 9 10.75 monoensis 6 12 14 13 11.25 ordii 17 23 9 5 13.5 luteolus 27 15 8 8 14.5 extractus 8 4 25 22 14,75 chapmani 26 11 11 12 15.0 montanus 13 10 23 30 19.0 cinderensis 7 18 32 19 19.0 fetosus 9 26 17 24 19.0 utahensis 14 14 28 21 19.25 columbianus 18 22 21 17 19.5 idoneus 31 13 20 15 19.75 priscus 28 30 6 16 20.0 celeripes 3 25 30 23 20.25 cineraceus 24 28 16 18 21.5 marshalli 10 20 33 28 22.75 inaquosus 19 24 22 27 23.0 attenuatus 29 21 18 25 23.25 fuscus 16 27 24 26 23.25 longipes 23 16 26 29 23.5 pallidus 22 32 27 20 25.25 nexilis 21 31 29 33 26.0 cupidineus 30 29 19 31 27.25 palmeri 32 17 31 32 28.0
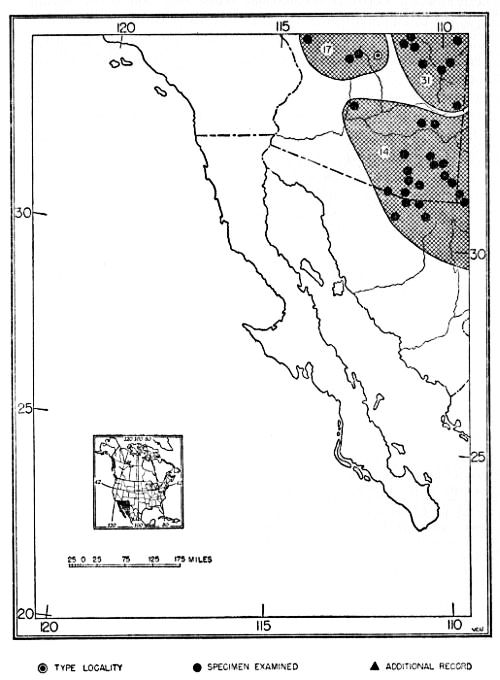
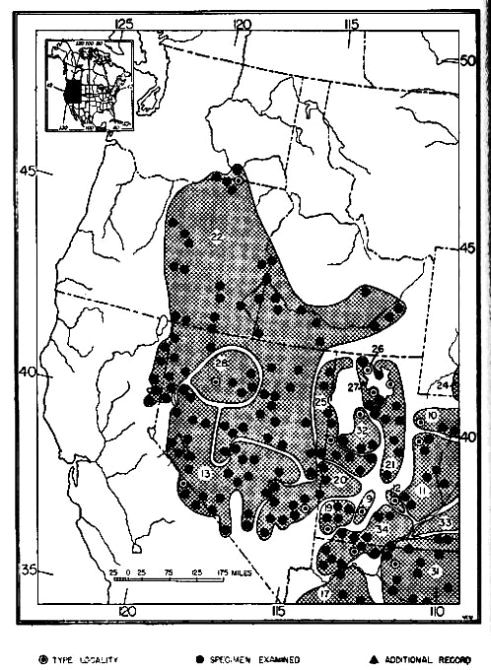
Fig. 23. Distribution of subspecies of Dipodomys ordii. 1. D. o. richardsoni 13. D. o. monoensis 25. D. o. celeripes 2. D. o. oklahomae 14. D. o. ordii 26. D. o. cineraceus 3. D. o. compactus 15. D. o. luteolus 27. D. o. marshalli 4. D. o. sennetti 16. D. o. extractus 28. D. o. inaquosus 5. D. o. evexus 17. D. o. chapmani 29. D. o. attenuatus 6. D. o. medius 18. D. o. montanus 30. D. o. fuscus 7. D. o. obscurus 19. D. o. cinderensis 31. D. o. longipes 8. D. o. terrosus 20. D. o. fetosus 32. D. o. pallidus 9. D. o. panguitchensis 21. D. o. utahensis 33. D. o. nexilis 10. D. o. uintensis 22. D. o. columbianus 34. D. o. cupidineus 11. D. o. sanrafaeli 23. D. o. idoneus 35. D. o. palmeri 12. D. o. fremonti 24. D. o. priscus
Ord Kangaroo Rat
Dipodomys ordii is a medium sized, relatively short-tailed, five-toed species of a color about average for the genus. As in other members of the genus, the hind legs and feet are disproportionately long as an adaptation to the saltatorial mode of progression. The upperparts are buffy, reddish or blackish, depending on the subspecies, but the entire ventral surface, dorsal surfaces of the hind feet, supraorbital and postauricular spots, forelimbs, hip stripes, lateral stripes of the tail and the tail at the base are pure white. The skull has a relatively short rostrum, moderate to large auditory bullae, relatively wide interparietal, relatively wide maxillary arches and grooved upper incisors.
The only other five-toed kangaroo rats with which Dipodomys ordii, at places, shares its geographic range, are Dipodomys panamintinus and Dipodomys microps. Dipodomys ordii can be distinguished from Dipodomys panamintinus by smaller size (for instance the hind foot is shorter instead of longer than, 44 mm.) and narrower expanse of maxillary arches in relation to breadth across the auditory bullae, and from Dipodomys microps by the awl-shaped, instead of chisel-shaped, lower incisors.
The species D. ordii is divisible into 35 subspecies, accounts of which follow:
Dipodops richardsoni Allen, Bull. Amer. Mus. Nat. Hist., 3:277, June 30, 1891.
Dipodomys phillippi, Knox, Trans. Kansas Acad. Sci., 4:22, 1875, (part—the part from Osborne, Kansas).
Dipodomys phillipsi ordi, Coues and Allen, Monogr. North American Rodentia, p. 542, 1877 (part—the part from Ft. Cobb, Oklahoma).
Perodipus richardsoni, Allen, Bull. Amer. Mus. Nat. Hist., 7:260, August 21, 1895 (part—the part from Pendennis, Kansas).
Cricetodipus richardsoni, Trouessart, Catalogus Mammalium, 1:581, 1897.
Perodipus montanus richardsoni, (Bailey, N. Amer. Fauna, 25:144, October 1905 (part—the part from Canadian, Texas).
Perodipus ordii richardsoni, Goldman, Proc. Biol. Soc. Washington, 30:113, May 23, 1917.
Dipodomys ordii richardsoni, Grinnell, Journ. Mamm., 2:96, May 2, 1921.
Type.—Male, no. 3025/2345, Amer. Mus. Nat. Hist.; on one of the sources of the Beaver River, Beaver County, Oklahoma; obtained on October 26, 1887, by Jenness Richardson and John Rowley, Jr. (After Allen, original description, type not seen.)
Range.—Southwestern Nebraska, eastern Colorado, northeastern New Mexico, Panhandle of Texas, and western parts of Oklahoma and Kansas; marginal localities are: in Nebraska: Bladen, Haigler; in Colorado: Olney; in New Mexico: Clayton; in Texas: 6 mi. S and 1 mi. W Quitaque, Vernon; in Oklahoma: 3 mi. S Cleo Springs; in Kansas: Medora.
Diagnosis.—Size large (see measurements). Color dark; entire dorsal surface Cinnamon-Buff, purest on sides and flanks, upper parts suffused with black; arietiform markings, pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, blackish; in some specimens the ventral stripe of tail does not extend to tip of pencil. Skull large; rostrum short and wide; nasals long; zygomata relatively heavy; auditory bullae well inflated and wide; thus with short rostrum giving appearance of nearly equilateral triangle; upper incisors long and robust.
Comparisons.—From Dipodomys ordii luteolus, D. o. richardsoni differs as follows: Size smaller in external measurements except length of body which is longer; color darker, except on plantar surfaces of hind feet and dorsal and ventral stripes of the tail which are lighter; ventral stripe of tail, in most specimens, continuous to end of pencil, whereas in D. o. luteolus ventral stripe is present on only proximal two-thirds; skull larger in all measurements taken; zygomatic arch heavier; auditory bullae relatively as well as actually more inflated; external auditory meatus egg-shaped as contrasted to nearly round in D. o. luteolus; pterygoid fossae rounded as compared to ovate in D. o. luteolus.
From Dipodomys ordii oklahomae, D. o. richardsoni differs as follows: Size larger in all measurements taken; color darker in all pigmented areas; skull larger in all respects; auditory bullae larger and more inflated ventrally; jugal straight or nearly so instead of bowed laterad; pterygoid fossae smaller; nasals straight instead of inflated as a "bulb" distally.
For comparisons with D. o. montanus and D. o. evexus, see accounts of those subspecies.
Remarks.—This race of Dipodomys ordii is readily distinguished from Dipodomys ordii evexus, from the valley of the upper Arkansas River, by larger size, larger skull and lighter color. Intergradation with Dipodomys ordii luteolus occurs rather freely in northeastern Colorado, as indicated by specimens from 3 miles northeast of Fitzsimmons, 6 miles east and 1 mile north of Denver and Barr Lake. These specimens resemble D. o. richardsoni in light color, greater inflation of the auditory bullae and the shape of the pterygoid fossae but resemble D. o. luteolus, to which they are here referred, in the length of the nasals, the least interorbital width and in the external measurements. In the southern part of the range of D. o. richardsoni intergradation occurs with Dipodomys ordii medius, as at 6 miles southwest of Muleshoe, Texas. Specimens from there have the long, wide rostrum and narrow skull of D. o. richardsoni but in the sum total of their characters more closely resemble D. o. medius. At [Pg 513] Texline, Texas, the animals show intergradation in the length and shape of the nasals and degree of convexity of the cranium but are referable to D. o. richardsoni.
In fine, intergradation occurs at all points where the range of D. o. richardsoni touches that of any other geographic race. No one series of it is as uniform as are most series of specimens of other known races. Dipodomys ordii richardsoni shows a mixture of characters. Nevertheless, each of the populations studied has characters in most of the animals that make this form recognizable as a taxonomic unit—a unit that seems, as yet, not to have become stabilized even in the central parts of its range.
Coues and Allen (1877:542) list specimens from Fort Cobb, Arkansas. It is known that the Post Office Department, for administrative purposes, attached certain towns and military installations in Indian Territory (now Oklahoma) to the State of Arkansas. Thus it is apparent that Fort Cobb, Arkansas, as recorded by Coues and Allen (loc. cit.) is Fort Cobb, Oklahoma. Specimens from Fort Cobb would be expected to be D. o. richardsoni.
Specimens examined.—Total, 351, distributed as follows:
Nebraska: Adams County: Bladen, 10 (AMNH). Dundy County: Haigler, 1 (USBS).
Colorado: Crowley County: Olney, 1 (USBS). Kiowa County: Chivington, 3 (USBS). Otero County: 18 mi. S La Junta, 4 (AMNH); Higbee, 1 (USBS). Bent County: 4 mi. SE Las Animas, 4100 ft., 3 (MVZ). Prowers County: Lamar, 9 (LACM); 1 mi. S Lamar, 4000 ft., 11 (KU). Baca County: Gaumes Ranch, 4600 ft., NW Corner, 1 (USBS).
Kansas: Cheyenne County: 23 mi. NW St. Francis, 5 (KU). Rawlins County: 2 mi. NE Ludell, 2 (KU); 1-1/2 mi. W Ludell, 1 (KU). Wallace County: Lacey Ranch, 4-1/2 mi. E and 9 mi. S Wallace, 1 (KU); unspecified, 2 (KU). Logan County: 5 mi. W Elkader, 2 (KU); unspecified, 1 (UM). Gove County: unspecified, 1 (KU). Trego County: Banner, 8 (USNM); Parrington Ranch, 12 mi. S Collyer, 2 (KU); unspecified, 8 (USNM). Ellis County: Ellis, 1 (USBS). Lane County: Pendennis, 10 (USBS). Hamilton County: Coolidge, 2 (CNHM); 1 mi. E Coolidge, 5 (KU). Pawnee County: 1 mi. S Larned, 4 (KU); 2 mi. S and 1/4 mi. W Larned, 2 (KU); 3 mi. S and 1-1/2 mi. W Larned, 10 (KU). Edwards County: Kinsley, 3 (USBS); 3-1/2 mi. E Kinsley, 5 (KU); S side Arkansas River, 2 mi. S Kinsley, 1 (KU); 1 mi. W and 3-1/2 mi. S Kinsley, 9 (KU). Stafford County: Little Salt Marsh, 15 mi. N and 3 mi. E Stafford, 2 (KU). Reno County: Medora, 1 (UM); 2 mi. W and 1/2 mi. S Medora, 4 (KU). Kiowa County: 5 mi. N Belvidere, 1(KU). Pratt County: Cairo, 2 (USBS). Sedgwick County: Wichita, 6 (AMNH). Morton County: 10 mi. N and 3 mi. E Elkhart, 34-1/2 (KU). Seward County: 1 mi. E Arkalon, 7 (KU); Liberal, 1 (KU); unspecified, 1 (KU). Meade County: Meade, 1 (USNM); 13 mi. SW Meade, 13 (6 AMNH; 7 KU); 17 mi. SW Meade, 2 (KU). Clark County: 12 mi. S Ashland, 1 (UM); unspecified, 3 (KU). Barber County: Medicine Lodge, 4 (USBS); 1 mi. W Aetna, 3 (KU); 1/2 mi. W Aetna, 2 (KU); Aetna, 3 (KU); 1 mi. SW Aetna, 1 (KU); 1-1/2 mi. SW Aetna, 1 (KU); 1 mi. S Aetna, 5 (KU); unspecified, 2 (KU). Harper County: 4-1/2 mi. NE Danville, 12 (KU); 2 mi. NE Runnymede, 3 (KU).
New Mexico: Union County: Clayton, Apache Canyon, 1 (USBS). Quay County: Glenrio, 10 (LACM).
Oklahoma: Cimmaron County: Kenton, 1 (CM). Beaver County: 1-1/2 mi. N Beaver, 7 (KU); Beaver River, 8 (7 AMNH; 1 CNHM). Harper County: 3 mi. S of Englewood, Kansas, 2 (MVZ); 4-1/2 mi. N Laverne, 1 (UM). Woods County: 2 mi. W Edith, 1 (USBS); Alva, 12 (UM); Waynoka, 18 (UM); 3 mi. SW Waynoka, 1 (USBS). Alfalfa County: 4 mi. SE Cherokee, 1 (USBS). Ellis County: Shattuck, 1 (USBS). Woodward County: Woodward, 9 (USBS). Major County: 3 mi. S Cleo Springs, 1 (USBS).
Texas: Dallam County: Texline, 8 (USBS). Lipscomb County: Lipscomb, 8 (USBS). Hemphill County: 17 mi. NE Canadian, 1 (MVZ); 1 mi. W Canadian, along Red Deer River, 12 (MVZ); 1/2 mi. W Canadian, along Red Deer River, 7 (MVZ); Canadian, 5 (USBS). Oldham County: Tascosa, 6 (USBS). Wheeler County: 1 mi. W Mobeetie, 2 (MVZ); Mobeetie, 8 (USBS); Wallace Ranch, SW Wheeler County, 1 (TCWC). Hall County: Newlin, 1 (USBS). Wilbarger County: Vernon, 5 (USBS). Floyd County: 6 mi. S and 1 mi. W Quitaque, 1 (UM).
Dipodomys oklahomae Trowbridge and Whitaker, Journ. Mamm., 21:343, August 14, 1940.
Dipodomys ordii oklahomae, Davis, Journ. Mamm., 23:332, August 14, 1942.
Type.—Female, young adult, no. 265454, U. S. Nat. Mus., Biol. Surv. Coll. (formerly Univ. of Oklahoma, Mus. Zool., no. 14517); north bank of South Canadian River, 2-1/4 mi. S Norman, Cleveland County, Oklahoma; obtained on March 16, 1934, by H. L. Whitaker, original no., X-catalog no. 29312 of U. S. Nat. Mus.
Range.—Known only from the South Canadian River Valley west of Minco, Canadian County; and east to Lexington, Cleveland County, Oklahoma.
Diagnosis.—Size medium (see measurements). Color light, entire dorsal surface near (c) Vinaceous-Buff, paler on sides with great suffusion of white; arietiform markings, pinnae of ears, plantar surfaces of hind feet, proximal ventral portion of tail and dorsal stripe on tail, brownish. Skull of medium size; rostrum wide; nasals short, projecting but slightly anteriorly to incisors; zygomatic processes of maxillae heavy; bullae not greatly inflated.
Comparisons.—Dipodomys ordii oklahomae differs from D. o. richardsoni as follows: Size smaller; color lighter in all pigmented areas; ventral stripe of tail extending only one-fourth the length rather than three fourths or to end of tail; skull smaller in all measurements taken; rostrum heavier; auditory bullae less inflated; pterygoid fossae larger; braincase slightly more inflated; nasals more expanded distally; interparietal region wider.
From Dipodomys ordii ordii, D. o. oklahomae differs in: Size larger in all measurements taken; color lighter in all pigmented areas; ventral stripe of tail extending one fourth length of tail rather than to end; skull larger in all respects; rostrum heavier; zygomatic arches heavier; bullae more inflated ventrally; cutting edge of upper incisors wider; pterygoid fossae larger; braincase more vaulted; nasals more expanded distally; orbital region larger; interparietal region wider.
Remarks.—Trowbridge and Whitaker named this kangaroo rat as a full species. The diagnostic characters were the length and breadth of the rostrum and the relatively great inflation of the auditory bullae. Also, Dipodomys oklahomae was not known to intergrade with any other named kinds. Davis (1942:332) treated D. oklahomae as a subspecies of the earlier named species Dipodomys ordii. Certain characters in specimens from the type series of both D. o. richardsoni and D. oklahomae, such as the shape and configuration of the nasals, the over-all proportion of the skull, tooth pattern and body proportions through individual variation overlap and [Pg 515] indicate that these two groups of animals belong to the same species, even though animals from intermediate geographic areas are not available to show actual intergradation. My findings corroborate Davis' conclusion that D. oklahomae should stand as Dipodomys ordii oklahomae. In spite of the great similarities shown by the two groups of animals there are still sufficient diagnostic characters between the two groups to enable them to be segregated easily as valid subspecies.
Dipodomys ordii oklahomae is, for some unknown reason, restricted to a limited geographic range. Specimens examined from the upper reaches of the South Canadian River, farther westward, are all referable to D. o. richardsoni rather than, as would be expected, to D. o. oklahomae since the habitat for these animals is continuous from the type locality of D. o. oklahomae to the upper reaches of the South Canadian River. In length and shape of the nasals, degree of inflation of the auditory bullae and width of the interorbital region these specimens from the upper reaches of the South Canadian River are intergrades between D. o. richardsoni and D. o. medius. The range of D. o. medius lies to the south of that of D. o. richardsoni and to the southwest of that of D. o. oklahomae.
The present range of D. o. oklahomae, as now understood, is the most eastern part of the range of the species Dipodomys ordii and of the genus Dipodomys. The existence of D. o. oklahomae in this area is a precarious one since its habitat is limited in extent and is periodically flooded.
Although no specimens are known from the area where intergradation between D. o. oklahomae and D. o. richardsoni would be expected to occur, it would seem that when animals from this region become available, intergradation will be shown to occur.
Specimens examined.—Total, 8, all from Oklahoma, distributed as follows: Grady County: 4 mi. N Minco, 1 (USBS). Cleveland County: 2-1/4 mi. S Norman, 7 (6 OU; 1 USBS).
Dipodomys compactus True, Proc. U. S. Nat. Mus., 11:160, January 5, 1889.
Cricetodipus compactus, Trouessart, Catalogus Mammalium, 1:581, 1897.
Perodipus compactus, Elliot, Field Columbian Museum, Zool. Ser., 2:240, 1901.
Dipodomys ordii compactus, Davis, Journ. Mamm., 23:332, August 14, 1942.
Type.—None designated but Poole and Schantz (1942:406) assumed it to be a female, no. 19665/35227, only the skin found, from Padre Island, Cameron County, Texas. April 3, 1888. Purchased from C. K. Worthen.
Range.—Padre and Mustang islands, Cameron County, Texas.
Diagnosis.—Size medium (see measurements); tail short. Color light; entire dorsal surface Light Ochraceous-Buff, purest on sides and flanks, upper parts but lightly suffused with black. A lighter color phase has entire dorsal surface Cartridge Buff, purest on sides and flanks, upper parts but lightly washed with black. In both phases, cheeks white; pinnae of ears, plantar surfaces of hind feet, dorsal stripe of tail, ventral stripe of tail (in most specimens) present on proximal third of tail only, brownish. Skull small; rostrum narrow and long; nasals long; auditory bullae inflated, but greatest breadth across bullae only slightly more than breadth across zygomatic processes of maxillae; interparietal region wide.
Comparisons.—From Dipodomys ordii sennetti, D. o. compactus differs in: Size slightly less; color lighter in all pigmented areas; skull smaller; auditory bullae slightly less inflated; interorbital width less; interparietal region wider; nasals longer.
From Dipodomys ordii attenuatus, D. o. compactus differs in: Body larger; tail shorter; normal color phase darker, and lighter color phase lighter; skull larger; rostrum wider and longer; nasals longer; interorbital region wider; auditory bullae relatively as well as actually less inflated; interparietal region wider; pterygoid fossae large and round as opposed to small and ovoid.
Compared with Dipodomys ordii medius and Dipodomys ordii ordii, D. o. compactus is smaller, lighter in color, and has less inflated auditory bullae and a smaller skull.
Remarks.—This subspecies of Dipodomys ordii was originally described as Dipodomys compactus by True in 1889 and stood as a full species until Davis (1942:332) relegated it to subspecific status under Dipodomys ordii. Davis (op. cit.) observed close resemblances in external proportions, size of mastoid bullae, width of supraoccipital, and size and shape of the interparietal, between Dipodomys ordii and Dipodomys sennetti and therefore concluded that they were only subspecies of one species. He observed that the difference between Dipodomys compactus and Dipodomys sennetti was of approximately the same degree as that between Dipodomys sennetti and Dipodomys ordii. From this he concluded that all three were subspecies of the one species Dipodomys ordii.
In any sizeable sample of Dipodomys sennetti there are crania closely resembling those of Dipodomys ordii ordii and others closely resembling those of Dipodomys compactus. The external proportions of both D. sennetti and D. compactus are duplicated in D. ordii from El Paso and conversely, specimens with the proportions of typical D. o. ordii occur in populations of D. sennetti and D. compactus. Thus, it appears that Davis' usage of the name Dipodomys ordii compactus should stand although there may be a hiatus in geographic occurrence between D. ordii and D. sennetti, as of course there is between D. sennetti and D. compactus.
In D. o. compactus there is a complete enamel ring around the occlusal surface of each molariform tooth; in D. o. ordii this ring is incomplete lingually on each of the molariform teeth and labially on the first three, and in some individuals of D. o. sennetti the enamel ring is complete and in others it is incomplete labially and lingually as in D. o. ordii.
Specimens examined.—Total, 44, all from Texas, distributed as follows: Nueces County: 19 mi. S Port Aransas, Mustang Island, 27 (17 TCWC; 10 MVZ); Mustang Island, 17 (LACM).
Dipodops sennetti Allen, Bull. Amer. Mus. Nat. Hist., 3:226, April 29, 1891.
Cricetodipus sennetti, Trouessart, Catalogus Mammalium, 1:581, 1897.
Perodipus sennetti, Elliott, Field Columbian Museum, Zool. Ser., 2:239, 1901.
Dipodomys ordii sennetti, Davis, Journ. Mamm., 23:332, August 14, 1942.
Type.—Male, no. 3478/2733. Amer. Mus. Nat. Hist.; near Brownsville, Cameron County, Texas; obtained on March 9, 1888, by J. M. Priour. (After Allen, original description, type not seen.) Type locality recorded by Bailey (1905:145) as "Santa Rosa, 85 mi. SW Corpus Christi."
Range.—Southern Texas, south of Corpus Christi; marginal localities, all in Texas are: Somerset, 8 mi. NE Los Angeles, 8 mi. E Encinal, Santa Rosa, 28 mi. E Raymondville, 2 mi. S Riviera.
Diagnosis.—Size small (see measurements). Color dark, entire dorsal surface (c) between Pinkish Buff and Cinnamon-Buff, purest on sides and flanks, upper parts mixed with black; arietiform markings, pinnae of ears, dorsal and ventral stripes of tail, plantar surfaces of hindfeet, brownish-black. Skull small; auditory bullae but slightly inflated in relation to size of skull; nasals slightly flaring distally; premaxillae extending but slightly posterior to nasals; interorbital width relatively great; external auditory meatus small; rostrum relatively long and wide; zygomatic arches relatively heavy.
Comparisons.—From Dipodomys ordii ordii, D. o. sennetti differs in: Size smaller, tail shorter; color darker; skull smaller; nasals longer; rostrum wider; auditory bullae less inflated; external auditory meatus smaller; pterygoid fossae more rounded; zygomatic arches heavier.
From Dipodomys ordii medius, D. o. sennetti differs as follows: Size smaller; color darker, but with less red in pelage; skull markedly smaller in all respects.
From Dipodomys ordii compactus, D. o. sennetti differs in: Size somewhat less; color darker; skull with total length greater; orbit smaller; least interorbital width greater; braincase more inflated; width across auditory bullae more; interparietal region wider; external auditory meatus larger; medial part of audital portion (see Howell, 1932) of auditory bullae larger.
Remarks.—Dipodomys sennetti, along with Dipodomys compactus, was regarded by Davis as conspecific with Dipodomys ordii. Reasons for placing these two kinds of kangaroo rats as subspecies of D. ordii are given in the account of Dipodomys ordii compactus.
This subspecies is known only from north of the Rio Grande which may serve as a barrier to the spread of the animal into northern Tamaulipas.
Specimens examined.—Total, 20, all from Texas, distributed as follows: Atascosa County: Somerset, 2 (TCWC). LaSalle County: 8 mi. NE Los Angeles, 1 (TCWC); 8 mi. E Encinal, 1 (TCWC). Kleberg County: 2 mi. S Riviera, 9 (TCWC). Jim Hogg County: Hebronville, 8 (LACM). Brooks County: Falfurrias, 2 (LACM). Willacy County: 28 mi. E Raymondville, 2 (TCWC).
Dipodomys ordii evexus Goldman, Journ. Washington Acad. Sci., 23:468, October 15, 1933.
Perodipus montanus richardsoni Warren, Mammals of Colorado, p. 76, 1910 (part—the part from Salida, Colorado).
Type.—Male, adult, no. 150990, U. S. Nat. Mus. Biol. Surv. Coll.; Salida, Chaffee County, Colorado (altitude 7000 ft.); obtained on November 10, 1907, by Merritt Cary, original no. 1245.
Range.—Upper Arkansas River Valley of south-central Colorado, from Salida to Pueblo.
Diagnosis.—Size large (see measurements). Color dark, entire dorsal surface between (16") Pinkish Cinnamon and Cinnamon-Buff, purest on sides and flanks, upper parts strongly suffused with black; arietiform markings, pinnae of ears, plantar surfaces of hind feet and dorsal and ventral stripes of tail, blackish. Skull of medium size; rostrum short and wide; nasals short; auditory bullae but slightly inflated; braincase but slightly vaulted.
Comparisons.—From Dipodomys ordii richardsoni, D. o. evexus differs as follows: Size smaller in all measurements taken; color darker; ears darker, dorsal and ventral stripes on tail darker, arietiform markings darker and more distinct, plantar surfaces of hind feet darker; skull smaller in all measurements; length, as expressed in percentage of width of skull, greater in D. o. evexus (66 per cent in D. o. evexus, 62 per cent in D. o. richardsoni which gives the appearance of a long, narrow skull as contrasted with a rather short, wide skull); auditory bullae less expanded laterally, posteriorly and ventrally; interparietal region relatively wider in proportion to greatest width across auditory bullae; cutting edge of upper incisors narrower; pterygoid fossae smaller and more nearly circular.
Compared with Dipodomys ordii luteolus, D. o. evexus differs as follows: Size somewhat smaller in external measurements; color darker in all pigmented areas; skull smaller in two of the seven measurements taken but in the other five measurements somewhat larger; auditory bullae less inflated; cutting edge of upper incisors narrower; zygomatic arch heavier; pterygoid fossae smaller and more nearly circular; external auditory meatus ovoid as contrasted to nearly circular; paroccipital processes smaller.
From Dipodomys ordii nexilis, D. o. evexus differs in: Color darker; rostrum wider and shorter; interorbital region wider; breadth across maxillary arches greater; auditory bullae less inflated; interparietal region larger; zygomatic arches heavy and bowed laterad; molariform teeth smaller; cutting edge of upper incisors narrower.
For comparison with Dipodomys ordii montanus see account of that subspecies.
Remarks.—This race of kangaroo rat, described from the upper Arkansas River Valley, closely resembles Dipodomys ordii luteolus but differs in darker color, slightly smaller body and larger skull.
No evidence of intergradation with any other race was noted. To the south the range of D. o. evexus is separated from that of D. o. montanus by a high, transverse ridge of the Rocky Mountains which is inhospitable to these animals. Much territory inhospitable to Dipodomys intervenes also between the ranges of D. o. evexus and 3883D. o. luteolus, but there are areas connecting the northern part of the range of D. o. evexus and the southwestern part of the known range of D. o. luteolus, in which Dipodomys may occur. If kangaroo rats occur in these areas it is to be expected that they will show intergradation between the two subspecies concerned.
Specimens examined.—Total, 24, all from Colorado, distributed as follows: Chaffee County: Salida, 10 (3 USBS; 7 AMNH). Fremont County: Canyon City, 13 (USBS). Pueblo County: Pueblo, 1 (USBS).
Perodipus montanus richardsoni, Bailey, N. Amer. Fauna, 25:144, October, 1905 (part—the part from Santa Rosa, New Mexico).
Type.—Male, no. 118526, U. S. Nat. Mus. Biol. Surv. Coll.; Santa Rosa, Guadalupe County, New Mexico; obtained on October 5, 1902, by Jas. H. Grant, original no. 565.
Range.—From north-central New Mexico, southeastward to west-central Texas; marginal localities are, in New Mexico: 15 mi. N Ojo Caliente, Gallina Mts., Deer Creek, San Pedro; in Texas: 20 mi. N Monahans, Colorado, 7 mi. E Post, 6 mi. SW Muleshoe.
Diagnosis.—Size medium (see measurements). Color dark; entire dorsal surface (14") between Orange-Cinnamon and Cinnamon, purest on sides and flanks, dorsal surface lightly washed with black; arietiform markings, pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, brownish-black. Skull of medium size; nasals long; medial mastoid portion of auditory bullae well inflated caudad; braincase vaulted; external auditory meatus small; rostrum short and truncate; medial auditory portion of auditory bullae relatively little inflated; pterygoid fossae ovate; zygomatic arches slender and relatively straight; junction of jugal and zygomatic process of maxilla heavy.
Comparisons.—From Dipodomys ordii richardsoni, D. o. medius differs as follows: Tail longer; hind foot shorter; color darker; arietiform markings more distinct; white lateral stripes of tail narrower; ventral stripe of tail in most specimens complete to end of pencil; postauricular spots less pronounced; hip stripe narrower and in some specimens almost obliterated; skull smaller in all measurements taken; angle of dorsal extension of premaxilla with zygomatic process of maxilla more nearly 90°; braincase more vaulted; medial [Pg 520] mastoid portion of auditory bullae more inflated, and coming to more of a point; medial auditory portion of auditory bullae more inflated ventrally; rostrum shorter and narrower; external auditory meatus smaller.
From Dipodomys ordii montanus, D. o. medius differs in: Color lighter in all pigmented areas; skull larger in all respects; rostrum shorter and heavier; bullae more inflated; zygomata, while nearly straight, are bowed slightly laterally; pterygoid fossae more ovate; foramen magnum larger; pterygoid foramina smaller.
Compared with Dipodomys ordii ordii and Dipodomys ordii sennetti, D. o. medius is larger and darker. The skull is also larger in all measurements taken.
Compared with Dipodomys ordii longipes, D. o. medius is darker and smaller.
Remarks.—This hitherto undescribed race of Dipodomys ordii can readily be distinguished from any of its near neighbors by the characters set forth under diagnosis and comparisons.
Intergradation is noted with D. o. ordii, D. o. longipes, D. o. montanus and D. o. richardsoni. Among named races D. o. medius shows closest affinities with D. o. richardsoni but the two are easily separable. The northwestern extremity of the range of D. o. medius is an area of intergradation in which no specimens are clearly of one subspecies or the other. In specimens from 5 miles east of Abiquiu, New Mexico, three-way intergradation occurs. These animals are like D. o. medius in size, D. o. longipes in color and their cranial proportions are as in D. o. montanus. At Deer Creek, New Mexico, and at Monahans, Texas, the animals show intergradation in size of body and agree with D. o. ordii in cranial proportions. In specimens from 6 miles southwest of Muleshoe, Texas, intergradation with D. o. richardsoni in the shape of the skull and width of the rostrum is noted. In the sum total of characters studied, however, these specimens are referable to D. o. medius.
Specimens examined.—Total, 129, distributed as follows:
New Mexico: Rio Arriba County: 2 mi. SE El Rito, 2 (KU); Rio Alamosa, 15 mi. N Ojo Caliente, 1 (USBS); 6 mi. E Abiquiu, 4 (USBS); Rinconada, 5 (USBS); Espanola, 6 (USBS). Sandoval County: 12 mi. NW Alameda, 5500 ft., 3 (MVZ). Santa Fe County: Seton's Ranch, near Santa Fe, 1 (USBS); 8 mi. SW Santa Fe, 8 (KU); San Pedro, 3 (USBS). San Miguel County: Pecos, 2 (USBS); 3 mi. S Pecos, 2 (USBS); Rowe, 6 (LACM). Bernallilo County: Bear Canyon, Sandia Mountains, 7 (USBS); Pajarito, 3 (MVZ). Guadalupe County: Santa Rosa, 10 (USBS). Lincoln County: 44 mi. NW Roswell, 5 (MVZ). De Baca County: 8 mi. N Fort Sumner, 9 (USBS). Roosevelt County: Kenna, 4 (LACM). Curry County: 4 mi. W and 2-3/4 mi. N Clovis, 1 (MVZ). Chaves County: 40 mi. N Roswell, 2 (USBS); 35 mi. N Roswell, 2 (USBS); 15 mi. NE Roswell, 8 (LACM); Stinking Springs Lake, 3 (USBS).
Texas: Bailey County: 6 mi. SW Muleshoe, 5 (UM); 9 mi. SW Muleshoe, 2 (UM). Garza County: 7 mi. E Post, 5 (UM). Martin County: Stanton, 4 (USBS). Howard County: 6 mi. NE Coahoma, 7 (UM); 1 mi. S Coahoma, 1 (UM); 5 mi. W Big Springs, 2400 ft., 1 (MVZ). Mitchell County: Colorado, 5 (USBS). Winkler County: 20 mi. N Monahans, 1 (USBS). Ward County: Monahans, 1 (USBS).
Perodipus obscurus Allen, Bull. Amer. Mus. Nat. Hist., 19:603, November 12, 1903.
Dipodomys ordii obscurus, Grinnell, Journ. Mamm., 2:96, May 2, 1921.
Type.—Male, adult, no. 20957, Amer. Mus. Nat. Hist.; Rio Sestin, northwestern Durango, Mexico; obtained on April 13, 1903, by J. H. Batty. (Type not seen.)
Range.—Northwestern and northern Durango, Mexico; marginal localities are: Rosario, Rio Sestin, Mt. San Gabriel, Rio del Bocas, Villa Ocampo.
Diagnosis.—Size small (see measurements). Color dark, entire dorsal surface (16") between Pinkish Cinnamon and Cinnamon-Buff, purest on sides, flanks and cheeks, upper parts strongly suffused with black; arietiform markings, plantar surfaces of hind feet, pinnae of ears, dorsal and ventral stripes of tail, brownish. Skull of medium size, nasals long and flared distally; rostrum long and narrow; interorbital region relatively narrow; auditory bullae less inflated than in Dipodomys ordii palmeri; interparietal region narrow; zygomatic arches heavy and bowed laterad; pterygoid fossae ovoid; braincase but slightly vaulted.
Comparisons.—From Dipodomys ordii palmeri, D. o. obscurus differs in: Size larger; color lighter; nasals shorter and more flared distally; interorbital width less; lacrimal processes larger; auditory bullae less inflated; pterygoid fossae ovoid as opposed to subcircular; zygomatic arches heavier; rostrum shorter and wider.
From Dipodomys ordii ordii, D. o. obscurus differs as follows: Size smaller; color darker; skull smaller; nasals longer; rostrum narrower and shorter; interorbital width greater; interparietal region narrower; narrower across auditory bullae; zygomatic arches heavier and more bowed laterally; pterygoid fossae more ovoid; breadth across maxillary arches greater; external auditory meatus smaller.
With Dipodomys ordii attenuatus and Dipodomys ordii sennetti, D. o. obscurus needs no comparison since it is larger and darker than either of those subspecies and can readily be told from the latter by the greater expansion of the auditory bullae.
For comparison with Dipodomys ordii fuscus see account of that subspecies.
Remarks.—D. o. obscurus seemingly is not a far-ranging subspecies. The only examples referable to it come from a relatively restricted area of Durango, Mexico. One specimen from Rio del Bocas, Durango, is not typical and shows the characters described for the animals from Chihuahua City and from Casas Grandes. I have considered the possibility that this specimen is an intergrade between D. o. obscurus and an unnamed subspecies ranging to the northeastward. The other specimens in the series from Rio del Bocas are typical of D. o. obscurus.
Specimens examined.—Total, 69, all from Durango, distributed as follows: Rosario, 20 (AMNH); Villa Ocampo, 5 (AMNH); Rio Sestin, 30 (28 AMNH; 2 CNHM); Mt. San Gabriel, 2 (AMNH); Rio del Bocas, 11 (AMNH); Rancho Santuario, 1 (AMNH).
Fig. 24. Known occurrences and probable geographic range of the subspecies of Dipodomys ordii in the southeastern fourth of the range of the species. 1. D. o. richardsoni 7. D. o. obscurus 30. D. o. fuscus 2. D. o. oklahomae 14. D. o. ordii 31. D. o. longipes 3. D. o. compactus 16. D. o. extractus 35. D. o. palmeri 4. D. o. sennetti 23. D. o. idoneus 6. D. o. medius 29. D. o. attenuatus
Dipodomys ordii terrosus Hoffmeister, Proc. Biol. Soc. Washington, 55:165, December 31, 1942.
Dipodomys phillipsi ordi, Coues and Allen, Monogr. North American Rodentia, p. 541, August, 1877 (part—the part from Yellowstone River, Montana).
Perodipus montanus richardsoni, Cary, N. Amer. Fauna, 49:124, December, 1926 (part—the part from Glendive, Montana).
Type.—Male, no. 93477, Mus. Vert. Zool., Univ. California; Yellowstone River, 5 mi. W Forsyth, 2,750 ft., Rosebud County, Montana; obtained on June 2, 1940, by J. R. Alcorn, original no. 1528.
Range.—Extreme southwestern Saskatchewan and southeastern Alberta, eastern half of Montana, northern Wyoming and probably extreme western North Dakota; marginal localities are: 50 mi. W Swift Current, Saskatchewan; "near Medicine Hat," Alberta; in Wyoming, Sheep Creek, and 23 mi. SW Newcastle; in Montana, Medicine Rocks (14 mi. NE Ekalaka), and Glendive.
Diagnosis.—Size large (see measurements). Color dark, entire dorsal surface near (c) Ochraceous-Buff, purest on sides and flanks; upper parts mixed with black; arietiform markings, pinnae of ears, dorsal and ventral stripes of tail and plantar surfaces of hind feet brownish-black. Skull large; rostrum short, wide and deep; braincase slightly vaulted; auditory bullae markedly inflated ventrally; zygomatic arches heavy and bowed laterad; upper incisors long and robust.
Comparisons.—From Dipodomys ordii priscus, D. o. terrosus differs as follows: Size larger in all measurements taken, except for length of hind feet, which is less; color darker in all pigmented areas; skull larger in all parts measured except width of interparietal, which is less; auditory bullae more inflated ventrally; zygomatic processes of maxillae wider; rostrum deeper and shorter.
From Dipodomys ordii richardsoni, D. o. terrosus differs as follows: Size larger; color darker in all pigmented areas; ventral stripe of tail extending farther distally; skull larger except in width across auditory bullae, which is the same.
For comparison with Dipodomys ordii luteolus see account of that subspecies.
Remarks.—As noted in the comparisons, D. o. terrosus is larger and darker than D. o. priscus, D. o. luteolus or D. o. richardsoni, its closest geographic neighbors, and does not resemble any of them, but rather resembles D. o. longipes and D. o. evexus in size and appearance, both of which are distantly removed geographically.
Like other subspecies of the species D. ordii, D. o. terrosus prefers sandy soils to those of any other type. Two miles east and 1 mile south of Forsyth, Montana, animals were trapped on lenses of sandy soil. These lenses alternated with areas of black loam of similar size. It was noteworthy that burrows were found only in the areas of sandy soil, although paths used by the rats when foraging [Pg 524] did extend onto and several crossed the lenses of black loam. We were not permitted to excavate any of these burrows, but conversation with farmers of the immediate vicinity indicated that the burrows were not deep. An eight-inch disc would frequently plow out nests and food caches. It was said that each of several caches contained as much as a peck of wheat.
Intergradation was noted in animals from 23 miles southwest of Newcastle and Arvada, Wyoming. In animals from both localities the pterygoid fossae are more as in D. o. luteolus but referable to D. o. terrosus. The specimens from Arvada, although immature, possessed cranial characters which were intermediate between those of D. o. terrosus and D. o. luteolus but the specimens are referable to the former.
Specimens examined.—Total, 74, distributed as follows:
Montana: Petroleum County: 24 mi. N Roundup, 8 mi. SW Flatwillow, 2 (UM). Garfield County: Jordan, 10 (1 UM; 2 MVZ; 7 AMNH). Dawson County: Glendive, 9 (USNM). Musselshell County: Harvey Ranch, Melstone, 3 (MVZ). Rosebud County: Yellowstone River, 5 mi. W Forsyth, 2750 ft., 7 (MVZ); 2 mi. E and 1 mi. S Forsyth, 2600 ft., 8 (KU). Custer County: Miles City, 1 (USBS). Yellowstone County: Billings, 2 (1 USBS; 1 MVZ). Big Horn County: Fort Custer, 1 (USBS); Crow Agency, 1 (USBS). Powder River County: Powderville, 4 (USBS). Carter County: Medicine Rocks, 15 mi. N Ekalaka, 2 (USBS); Medicine Rocks, 14 mi. N Ekalaka, 2 (USBS); Clark's Fork, 1 (USBS).
Wyoming: Big Horn County: Dry Creek, 10 mi. W Germania, 1 (USBS); 3 mi. E Germania, 1 (USBS); Greybull, 2 (USBS); Bighorn Basin, 1 (USBS). Sheridan County: Arvada, 8 (USBS). Campbell County: Little Powder River, 1 (USBS). Weston County: Newcastle, 2 (USBS); 23 mi. SW Newcastle, 4 (USBS). Fremont County: Wilson's Ranch, Sheep Creek, S base Owl Creek Mountains, 1 (USBS).
Additional records.—Canada (Anderson, 1946:131): Alberta: near Medicine Hat, 1; Saskatchewan: near Shackleton, 45-50 mi. NW Swift Current, 1; near Tompkins, 50 mi. W Swift Current, 1.
Dipodomys ordii fremonti Durrant and Setzer, Bull. Univ. Utah, 35 (no. 26):21, June 30, 1945.
Type.—Female, no. 15661, Carnegie Museum, Pittsburgh, Pennsylvania; Torrey, 7000 ft., Wayne County, Utah; obtained on July 19, 1938, by W. F. and F. H. Wood, original no. 1562.
Range.—Known only from the type locality.
Diagnosis.—Size small (see measurements). Color dark, entire dorsal surface Cinnamon-Buff, purest on sides, flanks and cheeks; upper parts strongly suffused with black; arietiform markings, pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, brownish. Skull small; upper incisors long; rostrum deep; jugal bowed laterally; diastema long; upper molariform tooth-row long.
Comparisons.—From Dipodomys ordii panguitchensis, D. o. fremonti differs in: Color lighter in all pigmented areas, particularly ears which are light brown in D. o. fremonti and black in D. o. panguitchensis; skull larger in all measurements taken; upper incisors longer; rostrum deeper; auditory bullae deeper; jugal bowed laterally rather than straight; diastema longer.
From Dipodomys ordii cupidineus, longipes, nexilis, uintensis and sanrafaeli, D. o. fremonti can readily be distinguished by its smaller size and generally darker color.
Remarks.—This subspecies of Dipodomys ordii inhabits the upper reaches of the Fremont River in west-central Wayne County, Utah. D. o. fremonti appears to be isolated and is known only from the type locality. D. o. fremonti is so remarkably different from any other subspecies of Dipodomys ordii that a long period of isolation from the ancestral stock (which probably gave rise also to Dipodomys ordii utahensis and Dipodomys ordii panguitchensis) is indicated. Although intergradation is not known to occur with other kinds, differentiation has not progressed far enough for these animals to be recognized as a distinct species.
The subspecies closest, geographically, to D. o. fremonti is D. o. cupidineus from which D. o. fremonti differs more than from any of the other named forms.
Specimens examined.—Total, 9, from Utah, as follows: Wayne County: Torrey, 7000 ft., 9 (CM).
Dipodomys ordii uintensis Durrant and Setzer, Bull. Univ. Utah, 35 (no. 26):27, June 30, 1945.
Perodipus longipes, Allen, Bull. Amer. Mus. Nat. Hist., 8:246, November 1896 (part—the part from Uncompahgre Indian Reservation, Utah).
Dipodomys ordii luteolus, Moore, Journ. Mamm., 11:88, February, 1930 (part—the part from Vernal, Utah).
Type.—Male, adult, no. 11634, Carnegie Museum, Pittsburgh, Pennsylvania; Red Creek, 6,700 ft., 2 mi. N Fruitland, Duchesne County, Utah; obtained on August 15, 1936, by J. K. and M. T. Doutt, original no. 3433.
Range.—Uinta basin of the White, Green and Duchesne river drainage in northeastern Utah; marginal occurrences are: 2 mi. N Fruitland, 10 mi. S Ouray, Vernal.
Diagnosis.—Size large (see measurements); hind foot short. Color dark; entire dorsal surface, near (c) Cinnamon-Buff, purest on sides and flanks, with moderate suffusion of black on upper parts; cheeks white; arietiform markings, pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, brownish. Skull large; frontomaxillary suture convex mediad; lacrimal process large; styloid process projects, on ventral surface of tympanic bulla, beyond middle of external auditory meatus; nasals flared distally.
Comparisons.—From Dipodomys ordii priscus, D. o. uintensis differs in: Hind foot shorter; color darker; styloid process projects on ventral part of tympanic bulla well anterior to middle of external auditory meatus as opposed to projecting to middle; depth of foramen magnum, expressed in percentage of width across posterior margin of occipital condyles, greater (86 per cent in D. o. uintensis and 81 per cent in D. o. priscus); frontomaxillary suture convex mediad as opposed to nearly straight; lacrimal processes larger; nasals more flared distally.
From Dipodomys ordii nexilis, D. o. uintensis differs as follows: Size smaller; color lighter; interorbital breadth greater; frontomaxillary suture convex mediad as opposed to concave; lacrimal processes larger; nasals more flared distally; narrower across auditory bullae; basal length greater; zygomatic arches bowed laterad as opposed to relatively straight.
From Dipodomys ordii longipes, D. o. uintensis differs as follows: Size smaller; color darker; auditory bullae wider, longer and deeper; frontomaxillary suture convex mediad as opposed to nearly straight; greatest breadth across auditory bullae less.
For comparison with Dipodomys ordii sanrafaeli see account of that subspecies.
Remarks.—This large, rather dark race inhabits the desert valleys of the White, Green and Duchesne rivers in northeastern Utah. The race nearest geographically, as well as morphologically, is Dipodomys ordii priscus. Intergradation occurs with the latter subspecies at Vernal, Uintah County, in cranial measurements and in color. On the basis of color alone D. o. uintensis can be distinguished from D. o. sanrafaeli, the geographic race to the south. Specimens from Jensen are intermediate in color and cranial measurements between Dipodomys ordii nexilis and D. o. uintensis but are referable to the latter.
Specimens examined.—Total, 40, all from Utah, distributed as follows: Duchesne County: Red Creek, 6700 ft., 2 mi. N Fruitland, 4 (CM); 10 mi. S Myton, 1 (UU); 20 mi. S Myton, 1 (RH). Uintah County: Vernal, 1 (BYU); 20 mi. E Ouray, 5 (CM); Junction Green and White rivers, 4800 ft., 2 mi. S Ouray, 5 (CM); Pariette Bench, 5000 ft., 8 mi. S Ouray, 8 (CM); Desert Springs, 10 mi. S Ouray, 4 (CM); Pariette Bench, 12 mi. S Ouray, 2 (CM); Jensen, 5 (BYU); E side Green River, 3 mi. S Jensen, 4 (CM).
Dipodomys ordii sanrafaeli Durrant and Setzer, Bull. Univ. Utah, 35 (no. 26):26, June 30, 1945.
Dipodomys ordii longipes, Stanford, Journ. Mamm., 12:360, November, 1931 (part—the part from King's Ranch, Utah).
Type.—Female, adult, no. 4612, Museum of Zoology, University of Utah; 1-1/2 mi. N Price, 5567 ft., Carbon County, Utah; obtained on June 5, 1940, by Ross Hardy and H. Higgins, original no. 1901.
Range.—East-central Utah, east into west-central Colorado. Marginal occurrences are: in Utah, 12 mi. E Price, 1-1/2 mi. N Price, Notom, King's Ranch, 12 mi. SW Green River, 16 mi. NW Moab; in Colorado, State Line and Grand Junction.
Diagnosis.—Size large (see measurements). Color dark, entire dorsal surface Cinnamon-Buff, purest on sides and flanks with but slight suffusion of black on upper parts; cheeks white; arietiform markings, pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, brownish-black. Skull large; pterygoid fossae ovoid; lacrimal processes small; width across maxillary arches relatively great; auditory bullae well inflated; diastema short.
Comparisons.—From Dipodomys ordii longipes, D. o. sanrafaeli differs as follows: Size smaller; color lighter, more cinnamon, pinnae of ears lighter; [Pg 527] skull smaller; auditory bullae smaller; pterygoid fossae ovoid rather than round; wider across occipital condyles; narrower across zygomatic processes of maxillae.
From Dipodomys ordii cupidineus, D. o. sanrafaeli can be recognized by its larger size, lighter color and larger skull.
For comparisons with Dipodomys ordii nexilis, Dipodomys ordii priscus and Dipodomys ordii uintensis see accounts of those subspecies.
Remarks.—Intergradation between Dipodomys ordii cupidineus and D. o. sanrafaeli is noted in the intermediate size of body in a single specimen from Notom. Intergradation in color and cranial characters occurs between Dipodomys ordii nexilis and D. o. sanrafaeli in specimens from 16 miles northwest of Moab. All these specimens, however, are referable to D. o. sanrafaeli.
Animals from that part of the range of D. o. sanrafaeli west of the Green River are typical while those to the east of the river are all intergrades. Animals from 16 miles northwest of Moab, Utah, and from three localities in Colorado, even though intergrades with D. o. nexilis, are all referable to D. o. sanrafaeli. It appears that the Green River does not act as a complete barrier in this area since in the winter it occasionally freezes over, thus allowing the animals to cross. It is thought that kangaroo rats do not hibernate but remain more or less active throughout the winter. Man-made conveniences, such as bridges, might also serve as means of dispersal, permitting these animals to cross otherwise prohibitive barriers. Where there are no bridges across the Green River, farther to the south, the rats apparently do not cross the river; steep, rocky canyon-walls and the lack of ice on the water in winter lessen the chances of small mammals crossing from one side to the other.
Specimens examined.—Total, 30, distributed as follows:
Utah: Carbon County: 12 mi. NE Price, 2 (CM); 3 mi. NE Price, 1 (RH); 1-1/2 mi. N Price, 2 (1 RH; 1 UU); Wellington, 1 (RH). Emery County: "San Rafael, 21 mi. out," 1 (USAC); 12 mi. SW Green River, 2 (CM). Grand County: 1 mi. E Green River, 1 (MVZ); 16 mi. NW Moab, 2 (CM). Wayne County: Notom, 1 (BYU). Garfield County: King's Ranch, 4800 ft., 3 (2 UU; 1 USAC).
Colorado: Mesa County: State Line, 11 (MVZ); Fruita, 1 (USBS); Grand Junction, 2 (USBS).
Dipodomys ordii panguitchensis Hardy, Proc. Biol. Soc. Washington, 55:90, June 25, 1942.
Type.—Male, adult, no. 4375, Museum of Zoology, University of Utah; one mile south of Panguitch, 6666 ft., Garfield County, Utah; obtained on August 31, 1940, by Ross Hardy, original no. 2151.
Range.—Known only from the type locality.
Diagnosis.—Size small (see measurements). Color dark, entire dorsal surface near Olive-Brown, purest on sides and flanks, upper parts strongly suffused with black; cheeks white; arietiform markings, pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail which are wider than white lateral stripes, blackish. Skull small; rostrum relatively short and wide; interorbital region wide; interparietal region wide; foramen magnum elongate dorsoventrally; pterygoid fossae ovoid.
Comparisons.—From Dipodomys ordii utahensis, which it closely resembles, D. o. panguitchensis differs in: Size larger; color darker; interparietal region wider; foramen magnum elongate dorsoventrally as opposed to nearly round; pterygoid fossae ovoid as opposed to nearly round.
This subspecies can be distinguished from Dipodomys ordii fetosus, Dipodomys ordii celeripes and Dipodomys ordii cupidineus by its darker color and generally larger size.
For comparisons with Dipodomys ordii cinderensis and Dipodomys ordii fremonti see accounts of those subspecies.
Remarks.—This geographic race inhabits the upper reaches of the Sevier River Valley in the vicinity of Panguitch, Utah. Natural barriers to kangaroo rats, such as the Cedar Mountains to the west, high plateau country to the south, the Paunsaugunt Plateau to the east and the narrow canyons of the Sevier River to the north prevent these animals from extending their range or from coming into physical contact with any adjacent geographic races. This isolation has resulted in a fairly stable population. Some animals, however, show characters, such as the width of the rostrum, and the shape and length of the nasals which are intermediate between those of topotypes of D. o. utahensis and the type series of D. o. panguitchensis.
Specimens examined.—Total, 3, all from Utah, distributed as follows: Garfield County: 1 mi. S Panguitch, 6666 ft., 3 (2 RH; 1 UU).
Perodipus monoensis Grinnell, Univ. California Publ. Zool., 21:46, March 29, 1919.
Dipodomys ordii monoensis, Grinnell, Journ. Mamm., 2:96, May 2, 1921.
Type.—Female, adult, no. 27002, Museum of Vertebrate Zoology, University of California; Pellisier Ranch, 5 mi. N Benton Station, 5600 ft., Mono County, California; obtained on September 21, 1917, by J. Dixon, original no. 6384.
Range.—Northeastern Inyo and Mono counties, California, north to southern Pershing County and east to eastern Nye County, Nevada; marginal occurrences are: in California, 5 mi. N Benton Station and Deep Spring Valley; in Nevada, Arlemont, 2 mi. NW Palmetto, 1 mi. N Beatty, 5 mi. W White Rock Spring, Big Creek at Quinn Canyon Mts., 2-1/2 mi. S Lock's Ranch, 2 mi. S Millett P. O., 13-1/2 mi. NW Goldfield, Fingerrock Wash, Eastgate, 1/2 mi. NE Toulon, 21 mi. W and 2 mi. N Lovelock, 1/2 mi. S Pyramid Lake, West Walker River in Smith's Valley, and 10 mi. S Yerington.
Diagnosis.—Size medium (see measurements). Color pale, entire dorsal surface (c) between Pinkish Buff and Cinnamon-Buff, purest on sides, flanks and cheeks, with but slight suffusion of black in upper parts; pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, brownish. Skull medium in size; rostrum relatively long and narrow; nasals relatively short; interorbital region narrow; interparietal region relatively wide; lacrimal processes small; auditory bullae relatively small; pterygoid fossae circular; zygomatic arches robust and relatively straight; foramen magnum nearly circular.
Comparisons.—From Dipodomys ordii columbianus, D. o. monoensis differs as follows: Size larger; color lighter; skull larger; rostrum longer and narrower; interorbital region narrower; breadth across auditory bullae less; lacrimal processes larger; braincase less vaulted; auditory bullae more inflated ventrally; pterygoid fossae smaller; zygomatic arches more robust; cutting edge of upper incisors wider.
From Dipodomys ordii fetosus, D. o. monoensis differs in: Hind foot shorter; color lighter; skull smaller; rostrum shorter and narrower; interorbital width less; interparietal region larger; lacrimal processes smaller; auditory bullae less inflated.
For comparison with Dipodomys ordii inaquosus see account of that subspecies.
Remarks.—This subspecies retains all of its diagnostic characters throughout nearly all parts of its geographic range. Intergradation occurs in animals from the southern end of Pyramid Lake, Big Smoky Valley and near Toquima Peak, Nevada; these animals, although typical of D. o. monoensis in coloration, resemble D. o. columbianus cranially. Three-way intergradation between D. o. columbianus, D. o. fetosus and D. o. monoensis is noted in animals from east-central Nye County, Nevada. These animals resemble D. o. monoensis in size, D. o. fetosus in color and resemble D. o. columbianus in certain cranial features. These animals are referred to D. o. monoensis. Animals from Toulon, Nevada, in the inflation of the auditory bullae, the vault of the braincase, the color and the total length show intergradation with D. o. inaquosus but are referable to D. o. monoensis.
Specimens examined.—Total, 264, distributed as follows:
California: Mono County: Pellisier Ranch, 5 mi. N Benton Station, 17 (7 DRD; 10 MVZ); Benton, 5639 ft., 2 (1 LACM; 1 MVZ); Taylor Ranch, 2 mi. S Benton Station, 5300 ft., 2 (MVZ). Inyo County: Deep Springs Valley, 1 (LACM).
Nevada: Washoe County: 1/2 mi. S Pyramid Lake, 3950 ft., 1 (MVZ); 1-1/2 mi. N Wadsworth, 4100 ft., 2 (MVZ). Pershing County: 21 mi. W and 2 mi. N Lovelock, 4000 ft., 2 (MVZ); 3-1/4 mi. NNE Toulon, 3900 ft., 1 (MVZ); 3 mi. NNE Toulon, 3900 ft., 6 (MVZ); 1/2 mi. NE Toulon, 1 (MVZ); Toulon, 3930 ft., 5 (MVZ). Churchill County: Truckee Canal, 2 mi. SW Hazen, 4000 ft., 1 (MVZ); 1 mi. NW Soda Lake, 4000 ft., 2 (MVZ); 1 mi. S Soda Lake, 4000 ft., 1 (MVZ); 5 mi. W Fallon, 1 (MVZ); 4 mi. W Fallon, 4000 ft., 3 (MVZ); 1 mi. W Mountain Well, 5350 ft., 3 (MVZ); Eastgate, 4400 ft., 13 (MVZ). Lyon County: 6 mi. N Fernley, 1 (MVZ); 1 mi. SE Wadsworth, 4200 ft., 7 (MVZ); 3/4 mi. N Fernley Underpass, Fernley, 1 (MVZ); 1/2 mi. N Fernley Underpass, Fernley, [Pg 530] 1 (MVZ); Wilson Canyon, 8 mi. NE Wellington, 4700 ft., 1 (MVZ); West Walker River, Smith's Valley, 4700 ft., 4 (MVZ); 10 mi. S Yerington, Mason Valley, 4500 ft., 6 (MVZ). Mineral County: 8 mi. SE Schurz, 4100 ft., 18 (MVZ); Fingerrock Wash, Stewart Valley, 5400 ft., 4 (MVZ); Cat Creek, 4 mi. W Hawthorne, 4500 ft., 1 (MVZ); Huntoon Valley, 5700 ft., 1 (MVZ). Nye County: 2 mi. S Millett P. O., 5500 ft., 1 (MVZ); 4 mi. SE Millett P. O., 5500 ft., 11 (MVZ); 5 mi. SE Millett P. O., 5500 ft., 5 (MVZ); 4 mi. S Millett P. O., 5500 ft., 2 (MVZ); Millman Ranch, Moore Creek, 6400 ft., 19 mi. SE Millett P. O., 9 (MVZ); Meadow Creek Ranger Station, Toquima Mts., 2 (MVZ); Monitor Valley, 9 mi. E Toquima Mts., 7000 ft., 19 (MVZ); Fish Spring Valley, 1/2 mi. N Fish Lake, 6500 ft., 2 (MVZ); Railroad Valley, 2-1/2 mi. S Lock's Ranch, 5000 ft., 5 (MVZ); Hot Creek Valley 3-1/2 mi. E Hot Creek, 5650 ft., 1 (MVZ); Hot Creek Valley, 4/5 mi. S Hot Creek, 5900 ft., 1 (MVZ); 5-1/2 mi. NE San Antonio, 5700 ft., 1 (MVZ); San Antonio, 5400 ft., 2 (MVZ); 9 mi. W and 3 mi. S Tybo, 6200 ft., 2 (MVZ); Ralston Valley, 15-1/2 mi. NE Tonopah, 5800 ft., 2 (MVZ); Railroad Valley, 2-1/2 mi. S Lock's Ranch, 5000 ft., 5 (MVZ); Hot Creek Valley Creek, 5800 ft., 1 (MVZ); Ralston Valley, 34 mi. E and 1 mi. N Tonopah, 5650 ft., 2 (MVZ); Old Mill, N end Reveille Valley, 6200 ft., 6 (MVZ); 1-1/2 mi. S Silverbow, Kawich Mountains, 1 (MVZ); 5 7/10 mi. SE Kawich, 2 (MVZ); 5 mi. W White Rock Spring, 6950 ft., Belted Range, 2 (MVZ); 1 mi. N Beatty, 1 (DRD). Esmeralda County: 13-1/2 mi. NW Goldfield, 4850 ft., 3 (MVZ); 7 mi. N Arlemont, 5500 ft., 6 (MVZ); Arlemont, 11 (MVZ); Mouth Palmetto Wash, 7 (DRD); 2 mi. NW Palmetto, 7 (DRD); 1 mi. NW Palmetto, 1 (DRD); Palmetto, 7 (DRD); 1 mi. SE Palmetto, 7 (DRD); Pigeon Spring, 6400 ft., 1 (MVZ); Indian Spring, Mt. Magruder, 20 (DRD).
D(ipodomys) ordii Woodhouse, Proc. Acad. Nat. Sci. Philadelphia, 6:224, 1853.
Dipodomys phillipsi ordi, Coues and Allen, Monogr. North American Rodentia, p. 541, 1877 (part—the part from El Paso, Texas).
Dipodops ordii, Merriam, N. Amer. Fauna, 4:42, October, 1890 (part—the part from El Paso, Texas).
Cricetodipus ordii, Trouessart, Catalogus Mammalium, 1:581, 1897.
Perodipus ordi, Elliot, Field Columbian Museum, Zool. Ser., 2:238, 1901.
Perodipus montanus richardsoni, Bailey, N. Amer. Fauna, 25:144, October, 1905 (part—the part from Carlsbad, New Mexico).
Perodipus ordii, Goldman, Proc. Biol. Soc. Washington, 30:113, May 23, 1917.
Type.—None designated. Species characterized from specimens obtained by Dr. Woodhouse at El Paso, Texas.
Range.—Southeastern Arizona, southern New Mexico, western Texas and north-central Mexico; marginal occurrences are: in Arizona, 20 mi. NE Calva, Oracle and Calabasas; in Sonora, Nogales; in Chihuahua, Casas Grandes, Corralitos and Santa Rosalia; in Texas, 16 mi. E Van Horn and 30 mi. N Van Horn; in New Mexico, 40 mi. N Roswell, 40 mi. SE Corona and Mangos Valley.
Diagnosis.—Size small (see measurements). Color dark, entire dorsal surface (16") between Pinkish Cinnamon and Cinnamon-Buff, purest on sides and flanks, upper parts suffused with black; arietiform markings, pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, brownish-black. Skull of medium size; rostrum narrow and relatively long; braincase flattened; auditory bullae but slightly inflated; zygomatic arches slender and straight; upper incisors short and narrow.
Comparisons.—From Dipodomys ordii richardsoni, D. o. ordii differs as follows: Size smaller; color somewhat lighter; dorsal and ventral stripes of tail brownish instead of blackish; skull smaller in all measurements taken.
From Dipodomys ordii montanus, D. o. ordii differs in: Size smaller; color lighter; skull narrower across bullae and rostrum; wider interorbitally; all other measurements taken the same, thus imparting to the skull, in dorsal view, a longer, narrower appearance.
From Dipodomys ordii chapmani, D. o. ordii differs in: Size smaller; color lighter; skull smaller; zygomatic arches narrower at suture of jugal and zygomatic process of maxillary; rostrum narrower; upper incisors smaller; nasals shorter; least interorbital width less.
For comparisons with Dipodomys ordii sennetti and Dipodomys ordii compactus see accounts of those subspecies.
Remarks.—Intergradation occurs with Dipodomys ordii chapmani, Dipodomys ordii longipes, Dipodomys ordii medius and Dipodomys ordii attenuatus. Only along the Rio Grande near El Paso, Texas, are individuals of a population uniform. On the periphery of the range, specimens from a given locality may resemble D. o. ordii or D. o. longipes or may be intermediate between these two subspecies.
Animals from the Organ Mountains near Globe Springs, 20 miles north of Cliff, New Mexico, and those labeled with reference to Van Horn, Texas, on the average are darker, have a wider expanse across the auditory bullae, a wider interorbital region and, in most specimens, more distally flared nasals than the norm of other populations. This deviation from the normal is interpreted not as intergradation with any other subspecies but rather as individual variation in a given population.
Specimens from 40 miles southeast of Corona and 10 miles northeast of Socorro, New Mexico, show intergradation between D. o. ordii and D. o. montanus in size of body, configuration of nasals and cranial size. For a discussion of intergradation with D. o. longipes, D. o. medius and D. o. chapmani see the remarks in the accounts of those subspecies.
Specimens from Cananea and Santa Cruz, Sonora, Casas Grandes Viejo, Chihuahua City and Colonia Diaz, Chihuahua, are not typical of D. o. ordii but rather show the influence of some race probably to the southeast that is as yet unknown. In a series of nine specimens from near Casas Grandes Viejo, Chihuahua, four individuals are typically D. o. ordii, four resemble an apparently undescribed form and one specimen is intermediate between D. o. ordii and the unnamed subspecies. This unnamed race differs from D. o. ordii in having darker pelage, darker dorsal stripe on tail, larger body, wider interorbital region, longer skull, greater breadth across the bullae, less vaulted braincase, more robust zygomatic arches and the foramen magnum more deeply notched on both the dorsal and [Pg 532] ventral rims. It seems that these animals mentioned above should be referred to D. o. ordii at least until such time as material becomes available from eastern Chihuahua, northern Coahuila and northeastern Durango.
Specimens examined.—Total, 451, distributed as follows:
Arizona: Gila County: Rice, 2 (USBS). Maricopa County: Marinette, 8 (CAS). Pinal County: Oracle, 4 (USBS). Graham County: 20 mi. NE Calva, 5 (USBS); Fort Grant, 11 (7 USBS; 4 MVZ). Pima County: Ft. Lowell, 3 (DRD); 11-1/2 mi. S Tucson, 1 (MVZ); Continental, 20 (8 USBS; 12 DRD); La Osa, 11 (USNM); Babiquivari Mountains 10 mi. N International Boundary, 1 (DRD). Santa Cruz County: 2 mi. S Tumacacori Mission, 1 (DRD); 7 mi. N Patagonia, 4500 ft., 12 (CAS); 3 mi. N Patagonia, 1 (CAS); Calabasas, 7 (6 USBS; 1 USNM). Cochise County: 8 mi. W Bowie, 1 (LACM); Wilcox, 4163 ft., 25 (12 USBS; 5 UM; 4 MVZ; 4 USNM); 12 mi. SE Dos Cabezos, 2 (UM); 1 mi. WSW Chiricahua National Monument, 5000 ft., 1 (MVZ); Portal, 4500 ft., 2 (USBS); Fairbank, 8 (3 AMNH; 5 CNHM); Mouth Pinery Canyon, 4 (USBS).
New Mexico: Torrance County: Gran Quivira, Mesa Jumanes, 6 (USBS). Catron County: Mangos Valley, 3 (USBS); Alma, 3 (USBS); Pleasanton, 7 (USBS). Socorro County: Gallina Mountains, 1 (USBS); 10 mi. NE Socorro, 2 (USBS); 3 mi. N Socorro, 3 (MVZ); Socerro, 1 (USBS); Range, 2 mi. SW Socerro, 4700 ft., 2 (MVZ); Lava Mesa; S Clyde, 4300 ft., 1 (MVZ); Dry Creek, 3 (USBS); San Augustine Plain, 12 mi. N Monica Springs, 2 (USBS). Lincoln County: Guyo Canyon, 40 mi. SE Corona, 1 (USBS); 4 mi. W Carrizozo, 2 (UM). Grant County: Gila, 5 (USBS); Cactus Flat, 20 mi. N Cliff, 3 (USBS); Cliff, Gila River, 4470 ft., 1 (USBS); Silver City, 2 (USBS); Redrock, 2 (USBS); 9 mi. N Faywood, 2 (USBS); Hachita, 1 (USBS); Dog Spring, 11 (USNM); Deer Creek, Culberson Ranch, 2 (USBS). Sierra County: Fairview, 6500 ft., 1 (USBS); Cuchillo, 4700 ft., 3 (USBS); Lake Valley, 5000 ft., 3 (USBS). Otero County: Tularosa, 5 (USBS); 10 mi. SW Tularosa, 2 (CNHM); Quartz Sands, SW Tularosa, 2 (MVZ); White Sands, 12 mi. W Alamogordo, 1 (MVZ); 2 mi. S Alamogordo, 2 (UM); 3 mi. S Alamogordo, 1 (UM); 5 mi. S Alamogordo, 1 (UM); 9 mi. SW Alamogordo, 1 (UM); 12 mi. SW Alamogordo, 2 (UM); 15 mi. SW Alamogordo, 1 (LACM); White Sands, 18 mi. SW Alamogordo, 7 (MVZ); 19 mi. SW Alamogordo, 3 (UM); White Sands National Monument, 24 (13 CNHM; 11 LACM); 1/2 mi. SW Escondido, 4000 ft., 2 (MVZ); Jarilla, 1 (USBS). Hidalgo County: 10 mi. NW Lordsburg, 6 (LACM); 4 mi. NW San Luis Pass, 5200 ft., Animas Valley, 5 (MVZ). Luna County: Deming, 11 (USBS). Dona Ana County: Garfield, 4 (USBS); 6 to 8 mi. NE Las Cruces, 1 (CAS); 15 mi. W Las Cruces, 1 (LACM); 11 mi. W Las Cruces, 2 (CAS); Las Cruces, 3 (USBS); Organ Mountains, near Globe Springs, 2 (USBS); Coe's Ranch, El Paso Road, 35 mi. N El Paso, Texas, 2 (USBS); 1/4 mi. N Strauss, 2 (CAS); 1 mi. E Strauss, 4100 ft., 7 (MVZ); 35 mi. W El Paso, Texas, 2 (USNM); 20 mi. W El Paso, Texas, 1 (USNM); 10 mi. W El Paso, Texas, 1 (USNM); Mexican Boundary, near monument 5, Lat. 31° 47'; Long. 30° 15', 13 (USNM). Eddy County: 3 mi. NW Carlsbad, 7 (MVZ); 2 mi. E Carlsbad, 1 (KU); Eddy, 10 (USBS).
Texas: El Paso County: 3 mi. NE City Limits, El Paso, 3764 ft., 15 (12 MVZ; 3 TCWC); El Paso, 3 (USBS); near El Paso, 7 (USNM). Hudspeth County: 1 mi. NW old Fort Hancock, 3900 ft., 3 (MVZ); Fort Hancock, 4 (USNM); 4 mi. NW Sierra Blanca, 1 (LACM). Culberson County: 35 mi. N Van Horn, 5 (TCWC); 30 mi. N Van Horn, 1 (TCWC); 16 mi. E Van Horn, 3 (TCWC); 16 mi. SE Van Horn, 5 (TCWC); Kent, 1 (USBS). Reeves County: 5 mi. E Toyahvale, 1 (USBS). Jeff Davis County: 14-1/2 mi. S Fort Davis, 2 (UM). Presidio County: 10 mi. NE Marfa, 1 (UM).
Sonora: Nogales, 2 (USNM); Santa Cruz, 4 (USNM); 5 mi. N Cananea, 4750 ft., 4 (MVZ); Alamo Wash, 35 mi. NW Magdalena, 5 (DRD); Sonora, 2 (AMNH).
Chihuahua: 4.3 mi. W Casas Grandes Viejo, 5000 ft., 8 (MVZ); 1.5 mi. W Casas Grandes Viejo, 1 (MVZ); Casas Grandes, 2 (USBS); Gallego, 1 (USBS); Colonia Diaz, 6 (USBS); Las Trincheras, 9 mi. S by road Boquillos de Condios, 4 (MVZ); Santa Rosalia, 6 (USBS); Chihuahua, 7 (USBS); 5 mi. SE Chihuahua, 5250 ft., 4 (MVZ); Corallitos, 4 (1 USBS; 3 MVZ).
Perodipus ordii luteolus Goldman, Proc. Biol. Soc. Washington, 30:112, May 23, 1917.
Dipodomys phillipsi ordi, Coues and Allen, Monogr. of N. Amer. Rodentia, p. 541, August, 1877 (part—the part from Niobrara River, Nebraska).
Perodipus montanus richardsoni, Cary, N. Amer. Fauna, 33:141, August 17, 1911 (part—the part from Sterling, Colorado).
Dipodomys ordii luteolus, Grinnell, Journ. Mamm., 2:96, May 2, 1921.
Type.—Male, adult, no. 160408, U. S. Nat. Mus. Biol. Surv. Coll.; Casper, Natrona County, Wyoming; obtained on September 2, 1909, by Merritt Cary, original no. 1690.
Range.—Southeastern Wyoming, northeastern Colorado, northwestern half of Nebraska and southwestern South Dakota; marginal occurrences are: in Wyoming, Casper, Sun and Ft. Steele; in Colorado, Loveland, Hugo and Akron; in Nebraska, Birdwood Creek, Neligh and Valentine; in South Dakota, Batesland and Buffalo Gap.
Diagnosis.—Size medium (see measurements). Color light, entire dorsal surface between Light Ochraceous-Buff and Ochraceous-Buff, purest on sides and flanks; upper parts but lightly washed with black; arietiform markings, plantar surfaces of hind feet, pinnae of ears and dorsal and ventral stripes of tail, blackish. Skull medium in size; jugal weak; braincase slightly inflated; nasals slightly flared distally.
Comparisons.—From Dipodomys ordii terrosus, D. o. luteolus differs as follows: Size smaller, except that tail and ear are longer; color lighter in all pigmented areas; skull smaller in every measurement taken; auditory bullae less inflated; zygomatic processes of maxillae smaller; cutting edge of upper incisors narrower; zygomatic arch weaker.
From Dipodomys ordii priscus, D. o. luteolus differs in: Size larger except hind foot which is shorter; dorsal and ventral stripes of tail, plantar surfaces of hind feet, arietiform markings and pinnae of ears, in most specimens, darker; auditory bullae less inflated; nasals shorter; rostrum wider; total length of skull shorter; zygomatic arch weaker; foramen magnum more ovate.
From Dipodomys ordii evexus, D. o. luteolus differs as follows: Hind foot longer; color lighter in all pigmented areas; auditory bullae more inflated; pterygoid fossae more expanded laterally; width across maxillary arches less; interorbital region narrower; zygomatic arch weaker; external auditory meatus almost round as opposed to ovoid.
Comparison with Dipodomys ordii richardsoni is made in account of that subspecies.
Remarks.—Dipodomys ordii luteolus resembles D. o. priscus in size and color but can readily be told from it and D. o. richardsoni when specimens from the central portions of the ranges of the subspecies are compared. At and near the periphery of the range, especially in that part which adjoins the range of D. o. richardsoni, intergradation occurs. Specimens from Kennedy, Perch and Neligh, Nebraska, approach D. o. richardsoni in the shape of the pterygoid [Pg 534] fossae and nasal bones, but in all other characters they resemble D. o. luteolus to which subspecies they are here referred. Specimens from Loveland and 20 miles east of Avalo, Colorado, show intergradation with D. o. richardsoni in the width of the rostrum and size of the zygomatic arch but are referable to D. o. luteolus.
No specimens here referred to D. o. luteolus were found to intergrade with D. o. priscus.
Specimens examined.—Total, 250, distributed as follows:
South Dakota: Perkins County: 9 mi. N Bison, 8 (MVZ). Meade County: Smithsville, 2 (USBS). Jackson County: 20 mi. SSE Phillip, in Haakon County, 1 (MVZ). Custer County: Elk Mountain, 1 (MHS); Buffalo Gap, 2 (USNM). Bennett County: Big Spring Canyon, Batesland, 7 (CNHM); Rosebud Indian Agency, 1 (USBS).
Wyoming: Fremont County: 2-1/2 mi. W Shoshoni, 80 (KU); Granite Mountain, 1 (UM). Natrona County: 1 mi. NE Casper, 19 (KU); Casper, 7 (USBS); Sun, 8 (USBS). Converse County: Douglas, 2 (USBS). Niobrara County: Van Tassel Creek, 1 (CM). Carbon County: Fort Steele, 1 (USBS). Goshen County: Rawhide Butte, 1 (USBS).
Nebraska: Sioux County: Warbonnet Canyon, 6 (MHS); Glen, 1 (MHS); Agate, 3 (1 UM; 2 CNHM); Jim Creek, 1 (MHS). Box Butte County: 16 mi. NE Alliance, 4 (UM). Sheridan County: 12 mi. S Gordon, 3 (MVZ); 5 mi. N Antioch, 4 (UM); 4 mi. N Antioch, 3 (UM). Cherry County: Valentine, 9 (6 USBS; 2 USNM; 1 NU); 15 mi. E Gordon, 1 (CNHM); Niobrara River, 3 (USBS); 18 mi. NW Kennedy, 3 (2 USBS; 1 MHS); Valentine Lake Refuge, 1 (NU); Kennedy, 10 (3 MVZ; 5 USBS; 2 USNM); 30 mi. S Wood Lake, 4 (CNHM); Cherry, 7 (NU); near Clark's Canyon, 7 (USBS). Rock County: Perch, 12 (10 AMNH; 2 CNHM). Antelope County: Neligh, 8 (1 USBS; 7 MHS). Scotts Bluff County: Mitchell, 1 (MHS). Hooker County: Kelso, 5 (UM). Thomas County: Halsey, 1 (MHS); Dismal River, 2 (USBS). Custer County: 1 mi. NW Gavin, 6 (2 MVZ; 4 UM); Gavin, 3 (UM); 8 mi. NW Anselmo, 1 (UM); 7 mi. NW Anselmo, 1 (UM). Lincoln County: Birdwood Creek, 2 (USBS); Brady, 4 (NU); Hackberry Lake, 13 (UM).
Colorado: Larimer County: Loveland, 8 (USBS). Weld County: 20 mi. E Avalo, 3 (USBS); 8 mi. E Pawnee Buttes, 1 (USBS); Greeley, 1 (USBS). Morgan County: 30 mi. S Fort Morgan, 1 (UM). Logan County: Sterling, 8 (USBS). Adams County: Barr Lake, 4 (CMNH); 6 mi. E and 1 mi. N Denver, 1 (CMNH); 3 mi. NE Fitzsimons, 1 (CMNH). Washington County: Akron, 2 (UM). Lincoln County: Limon, 1 (USBS); Hugo, 1 (USBS). El Paso County: Colorado Springs, 2 (MVZ).
Type.—Male, adult, no. 76562, Museum of Vertebrate Zoology, University of California; 1 mi. E Samalayuca, 4500 ft., Chihuahua, Mexico; obtained on May 15, 1937, by William B. Richardson, original no. 2148.
Range.—Known only from the type locality.
Diagnosis.—Size medium (see measurements). Color pale, entire dorsal surface Pinkish Buff, purest on sides and flanks, dorsal surface but slightly suffused with black; pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail light-brownish. Skull medium in size; nasals long; rostrum relatively as well as actually wide; least interorbital breadth wide; auditory bullae relatively little inflated; braincase but slightly vaulted; external auditory meatus ovoid; zygomatic arches relatively robust and but slightly bowed laterad; pterygoid fossae subcircular; lacrimal processes small; foramen magnum deeply notched on dorsal border.
Comparisons.—From Dipodomys ordii ordii, D. o. extractus differs as follows: Size larger; color lighter; arietiform marks absent in D. o. extractus but pronounced in D. o. ordii; skull larger; rostrum wider; nasals longer; maxillary arches wider; auditory bullae more inflated laterally; braincase less vaulted; pterygoid fossae subcircular as opposed to circular; foramen magnum more deeply evaginated dorsally and ventrally.
Fig. 25. Known occurrences and probable geographic range of the subspecies of Dipodomys ordii in the northeastern fourth of the range of the species. 1. D. o. richardsoni 8. D. o. terrosus 18. D. o. montanus 2. D. o. oklahomae 10. D. o. uintensis 24. D. o. priscus 5. D. o. evexus 11. D. o. sanrafaeli 31. D. o. longipes 6. D. o. medius 15. D. o. luteolus 33. D. o. nexilis
From Dipodomys ordii obscurus, D. o. extractus differs as follows: Size larger; color lighter; skull larger in all respects, notably in least interorbital width, greatest length of skull and breadth across auditory bullae; zygomatic arches less robust and straight as opposed to robust and bowed laterally; pterygoid fossae subcircular as opposed to circular; auditory bullae more inflated ventrally.
From Dipodomys ordii idoneus, D. o. extractus differs in: Size larger; color lighter; skull larger in all measurements taken except least interorbital width and breadth across maxillary arches which are smaller; nasals longer and narrower; braincase less vaulted; zygomatic arches less robust and more nearly straight; auditory bullae less inflated ventrally.
Remarks.—This subspecies of Dipodomys ordii is the palest of any of the known races from Mexico. It inhabits an area of light-colored sandy soil at the type locality.
The only subspecies with which D. o. extractus really needs comparison is Dipodomys ordii ordii from which it differs as described above. The series which is herein described contains two individuals that resemble D. o. ordii cranially and three that resemble it in color although they are somewhat paler. One specimen, MVZ no. 76560, is the palest of any of the series, has a short tail and in these respects resembles the pale form of Dipodomys ordii compactus. It is apparent from the skull of this latter individual that it is not D. o. compactus but D. o. extractus.
Dipodomys ordii ordii inhabits, in most of its range, dark-colored soils, whereas D. o. extractus inhabits light-colored soils. Possibly the differentiation that has occurred is a result of an ecological separation much as occurs with Perognathus on the white sands and black lavas of the Tularosa Basin of New Mexico (see Benson, 1933).
Specimens examined.—Total, 14, all from Chihuahua, as follows: 1 mi. E Samalayuca, 4500 ft., 14 (MVZ).
Dipodomys chapmani Mearns, Bull. Amer. Mus. Nat. Hist., 2:291, February 21, 1890.
Cricetodipus chapmani, Trouessart, Catalogus Mammalium, 1:581, 1897.
Perodipus chapmani, Elliot, Field Columbian Museum, Zool. Ser., 2:237, 1901.
Perodipus ordii chapmani, Goldman, Proc. Biol. Soc. Washington, 30:113, May 23, 1917.
Dipodomys ordii chapmani, Grinnell, Journ. Mamm., 2:96, May 2, 1921.
Cotypes.—Male, no. 2400, and female, no. 2398, Amer. Mus. Nat. Hist.; Fort Verde, Yavapai County, Arizona; obtained on January 26, 1887, and 49October 1, 1885, respectively, by Edgar A. Mearns. (Type not seen.)
Range.—Central Arizona from the Grand Canyon of the Colorado River, southeastward to, and probably beyond, Fort Verde; marginal occurrences are: lower end Prespect Valley, Kirkland, Prescott, Camp Verde, Bill Williams Mountain.
Diagnosis.—Size medium (see measurements). Color dark; entire dorsal surface between (10") Pinkish Cinnamon and Cinnamon-Buff, purest on sides and flanks, upper parts strongly suffused with black; arietiform markings, pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, blackish. Skull medium in size; rostrum long and narrow; nasals long; auditory bullae slightly distended; braincase vaulted.
Comparisons.—For comparisons with Dipodomys ordii cupidineus, Dipodomys ordii longipes and Dipodomys ordii ordii see accounts of those subspecies.
Remarks.—This subspecies of Dipodomys ordii is relatively isolated from other subspecies of the species. The Colorado River with its deep canyons is interposed between the ranges of D. o. chapmani and D. o. cupidineus. A high range of mountains separates D. o. chapmani from the range of D. o. longipes. The only race with which D. o. chapmani probably comes into contact is D. o. ordii to the southeast. No specimens are available from the southeastern part of the range of D. o. chapmani. Animals from the northwestern part of the range of D. o. ordii, without exception, have dark-colored pelage which is characteristic of D. o. chapmani. In the shape of the nasals, vault of the braincase, interorbital width and auditory bullae, animals from Rice, Arizona, resemble D. o. chapmani, but in other characters resemble D. o. ordii to which subspecies they are referred. Animals from 20 miles northeast of Calva, Arizona, are dark-colored and are intermediate between the dark D. o. chapmani and the pale D. o. ordii in size of the cranium and body but are referable to D. o. ordii. In the northwestern part of the range of D. o. chapmani the animals are not typical in that they have a shorter, wider rostrum and a heavier zygomatic arch than topotypes.
Specimens examined.—Total, 90, all from Arizona, distributed as follows: Mohave County: lower end Prospect Valley, 4500 ft., Grand Canyon, 7 (USBS); Kingman, 9 (LACM). Coconino County: Aubrey Valley, 10 mi. S Pine Spring, 4 (USBS); Bill Williams Mountain, 1 (USNM). Yavapai County: 3 mi. N Fort Whipple, 11 (USBS); Fort Whipple, 4 (2 USBS; 2 USNM); Willow Creek, 4 mi. N Prescott, 4 (MVZ); 1 mi. W Camp Verde, 1 (MVZ); Camp Verde, 33 (15 USBS; 3 MVZ; 3 AMNH; 9 LACM; 3 DRD); 1/2 mi. S Camp Verde, 4 (MVZ); Kirkland, 9 (USBS); Turkey Creek, 3 (USBS).
Dipodomys montanus Baird, Proc. Acad. Nat. Sci. Philadelphia, 7:334, April, 1855.
Perodipus montanus, Merriam, Proc. Biol. Soc. Washington, 17:140, July 14, 1904.
Perodipus ordii montanus, Goldman, Proc. Biol. Soc. Washington, 30:113, May 23, 1917.
Dipodomys ordii montanus, Grinnell, Journ. Mamm., 2:96, May 2, 1921.
Type.—Male, no. 490/1631, U. S. Nat. Mus.; near Fort Massachusetts, Costilla County, Colorado; obtained in 1853 by F. Kreutzfeldt (Pacific Railroad Survey).
Range.—The San Luis Valley of south-central Colorado and north-central New Mexico; marginal occurrence are: in Colorado, 22 mi. E Mosca, Saguache, Alamosa and Antonito; in New Mexico, 4 mi. SW Cimmaron.
Diagnosis.—Size small (see measurements). Color dark, entire dorsal surface between (c) Vinaceous-Buff and Avellaneous, purest on sides and flanks, upper parts strongly suffused with black; arietiform markings, pinnae of ears, plantar surfaces of hind feet and dorsal and ventral stripes of tail, blackish. Skull small; rostrum slender; interorbital width narrow; auditory bullae strongly inflated; braincase but slightly vaulted; interparietal region wide; zygomatic arch bowed laterally.
Comparisons.—From Dipodomys ordii richardsoni, D. o. montanus differs as follows: Size smaller; color darker in all pigmented areas; ventral stripe of tail continuous to end of pencil in all specimens, whereas in D. o. richardsoni the stripe is incomplete in most specimens. In all specimens the dorsal and ventral stripes of the tail are darker in D. o. montanus; skull smaller in all measurements taken; jugal straight or nearly so, as contrasted to bowed laterally; pterygoid fossae shallower and more ovoid, that is to say, wings of pterygoid flare laterally; ventral projection of auditory bullae more pronounced; rostrum, immediately in front of zygomatic process of maxilla, nearly parallel to median plane along long axis of skull, while in D. o. richardsoni this union forms a decided obtuse angle; paroccipital processes smaller; foramen magnum smaller and more ovoid as opposed to larger and more circular.
From Dipodomys ordii evexus, D. o. montanus differs as follows: Size smaller in all measurements taken except width across auditory bullae which is greater; color darker, ventral stripe of tail continuous to end of pencil; least width of supraoccipital less (2.5 mm. in D. o. montanus and 3.5 mm. in D. o. evexus); extension of premaxillae posterior to nasals greater; braincase more inflated dorsally; bullae relatively as well as actually larger and more inflated ventrally; pterygoid fossae narrower anteroposteriorly and wider laterally than in D. o. evexus; jugal straight or nearly so as contrasted to bowed laterally; lacrimal processes smaller.
For comparison with Dipodomys ordii medius see account of that subspecies.
Fig. 26. Known occurrences and probable geographic range of the subspecies of Dipodomys ordii in the southwestern fourth of the range of the species. 14. D. o. ordii 17. D. o. chapmani 31. D. o. longipes
Remarks.—This subspecies of Dipodomys ordii appears to be restricted to the San Luis Valley of Colorado and New Mexico. Intergradation, between Dipodomys ordii ordii and D. o. montanus, is noted in animals from Gran Quivira, Mesa Jumanes, New Mexico. These animals have the frontomaxillary suture and extension of the premaxilla as in D. o. ordii and the nasals and inflation of the auditory bullae as in D. o. montanus. In the majority of characters they are, however, referable to the latter. Intergradation is noted also in animals from 4 miles southwest of Cimmaron, New Mexico, in that they resemble D. o. richardsoni in the shape of the nasals and the width and length of the rostrum but in the remainder of the characters studied resemble D. o. montanus to which they are referred.
Baird, in the original description, gives "near Fort Massachusetts" as the type locality for this subspecies. Miller (1923) lists the type locality as "Fort Massachusetts (now Fort Garland)." Fort Massachusetts and Fort Garland are two different ex-army posts. Old Fort Massachusetts was situated on what is now the Trinchera Ranch which is north of the town of Fort Garland. Old Fort Garland is approximately one-half mile south of the town of Fort Garland. The two abandoned army posts were no more than twelve miles apart. In 1946, at neither abandoned post could I find any habitat suitable for kangaroo rats.
Specimens examined.—Total, 148, distributed as follows:
Colorado: Saguache County: Saguache, 1 (USBS). Alamosa County: 1.6 mi. NE Medano Springs Ranch Headquarters, 8 (MVZ); Great Sand Dunes National Monument, 10 (MVZ); 8 mi. S Great Sand Dunes National Monument, 4 (MVZ); Medano Ranch, 15 mi. NE Mosca, 11 (2 USBS; 9 MVZ); 22 mi. E Mosca, 5 (AMNH); Alamosa, 1 (UM). Conejos County: Antonito, 1 (USBS); 7 mi. E Antonito, 3 (USBS); 12 mi. E Antonito, 2 (USBS). Costilla County: 4-1/2 mi. N Blanca, 8100 ft., 3 (MVZ); 1 mi. NE Blanca, 7950 ft., 1 (MVZ); 3 mi. NW Fort Garland, 6 (CMNH); 1 mi. NW Fort Garland, 3 (MVZ); 15 mi. W Fort Garland, 7800 ft., 4 (KU); Fort Garland, 64 (31 USBS; 28 LACM; 5 MVZ); 5 mi. SSE Fort Garland, 4 (AMNH); 5 mi. S Fort Garland, 2 (AMNH).
New Mexico: Colfax County: 4 mi. SW Cimmaron, 5 (AMNH); 3 mi. SE Cimmaron, 8 (AMNH); 1-1/2 mi. W Philmont Ranch Headquarters, 2 (AMNH).
Dipodomys ordii cinderensis Hardy, Proc. Biol. Soc. Washington, 57:53, October 31, 1944.
Type.—Male, no. 4611, Mus. Zool., Univ. Utah; at approximately 4,000 ft. on sandy soil, immediately north of the northern of two large cinder cones in Diamond Valley, 10 mi. N St. George, Washington County, Utah; obtained on February 13, 1944, by Ross Hardy, original no. 2690.
Range.—Diamond Valley, Washington County, Utah, north through Mountain Meadows, east as far as Cedar City, Iron County, Utah, north through the Escalante Desert to Lund, Iron County, and west almost to the Utah-Nevada boundary; marginal occurrences, all in Utah are: 11 mi. SE Lund; N end Mountain Meadows; Diamond Valley; 10 mi. W Cedar City; 4-1/2 mi. NW Summit and 6 mi. W Parowan.
Diagnosis.—Size small (see measurements). Color dark, upper parts near Buffy Brown, strongly suffused with black, purest on sides and flanks; pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, arietiform markings, black. Skull small; rostrum narrow; auditory bullae relatively as well as actually well inflated; external auditory meatus small; interparietal region narrow.
Comparisons.—From Dipodomys ordii fetosus, D. o. cinderensis differs in: Size smaller; color darker; skull smaller in all measurements taken.
From Dipodomys ordii panguitchensis, D. o. cinderensis differs as follows: Size smaller; color lighter; skull smaller except for auditory bullae which are larger; external auditory meatus smaller; maxillary tooth-row longer.
Compared with Dipodomys ordii longipes and Dipodomys ordii cupidineus, D. o. cinderensis is smaller, darker and can easily be distinguished.
For comparisons with Dipodomys ordii celeripes and Dipodomys ordii utahensis see accounts of those subspecies.
Remarks.—This race confined to Utah, and inhabiting northern Washington County and most of Iron County, is restricted almost exclusively to areas of loose shifting sand. Intergradation with Dipodomys ordii fetosus occurs near Lund. The animals from this locality intergrade in size of body and in color, but in the majority of cranial characters resemble D. o. cinderensis. In all characters studied D. o. cinderensis appears to be more closely related to Dipodomys ordii utahensis than to any other subspecies with which it comes into actual contact.
Animals from near Parowan have both cranial and external characters by which they could be referred to either D. o. panguitchensis or D. o. cinderensis. Owing to the existence of the physical barrier (Cedar Mountains) separating the two populations, they are referred to D. o. cinderensis. Dipodomys ordii cinderensis is not a well differentiated race but appears to possess characters sufficiently diagnostic to distinguish it from contiguous forms.
Specimens examined.—Total, 74, all from Utah, distributed as follows: Iron County: 11 mi. SE Lund, 50 (46 RH; 4 MVZ); 4-1/2 mi. NW Summit and 6 mi. W Parowan, 9 (RH); 10 mi. W Cedar City, 1 (USAC); 5 mi. W Cedar City, 1 (USAC); Cedar City, 2 (BYU). Washington County: Diamond Valley, 9 (RH); N end Mountain Meadows, 2 (RH).
Type.—Female, adult, no. 48451, Museum of Vertebrate Zoology, University of California; 2 mi. N Panaca, 4,800 ft., Lincoln County, Nevada; obtained on June 24, 1931, by Ward C. Russell, original no. 1658.
Range.—Southeastern Nevada and western Beaver and Millard counties, Utah; marginal occurrences in Nevada, 2 mi. SE Pioche, 15 mi. WSW Sunnyside, 16-1/2 mi. WSW Sunnyside, 14 mi. NNE Sharp, 8-1/2 mi. NE Sharp, 15 mi. S Groom Baldy, 10 mi. E Crystal Spring, Panaca; in Utah, Pine Valley, 50 mi. W Milford and 5 mi. S Garrison.
Diagnosis.—Size medium (see measurements). Color dark, entire dorsal surface (16") between Pinkish Cinnamon and Cinnamon-Buff, purest on sides and flanks with strong admixture of black in upper parts; cheeks white; pinnae of ears, arietiform markings, plantar surfaces of hind feet, dorsal and ventral stripes of tail, blackish. Skull medium in size; rostrum wide and short; interorbital region narrow; braincase but slightly vaulted; lacrimal processes large; zygomatic arches weak and bowed laterally; pterygoid fossae large and subcircular.
Comparisons.—From Dipodomys ordii utahensis, D. o. fetosus differs as follows: Hind foot longer; color lighter; skull larger; rostrum, relatively as well as actually, shorter and wider; nasals longer and wider; interorbital region narrower; braincase less vaulted; interparietal region narrower; auditory bullae more inflated laterally, posteriorly and ventrally; foramen magnum smaller.
From Dipodomys ordii celeripes, D. o. fetosus differs as follows: Larger; color darker; arietiform markings present; skull larger; rostrum longer and wider; nasals longer and less inflated distally; interorbital region wider; auditory bullae somewhat more inflated; zygomatic arches more robust.
For comparisons with Dipodomys ordii monoensis, D. o. columbianus, D. o. pallidus, D. o. panguitchensis and D. o. cinderensis see accounts of those subspecies.
Remarks.—Dipodomys ordii fetosus has a small geographic range. Hall (1946:416) points out that the interorbital breadth in topotypes is less than in those from any other part of the range. This feature, therefore, is not useful in characterizing this subspecies. Intergradation with Dipodomys ordii columbianus is noted in the greater total length and the somewhat darker color in animals from Coal Valley and Garden Valley, Nevada. In the part of the range which lies in Utah, none of the animals is typical and all are intergrades in color and cranial characters with Dipodomys ordii celeripes.
Specimens examined.—Total, 136, distributed as follows:
Nevada: Nye County: White River Valley, 15 mi. WSW Sunnyside, 5500 ft., 26 (MVZ); White River Valley, 16-1/2 mi. WSW Sunnyside, 5500 ft., 6 (MVZ); 14 mi. NNE Sharp, 1 (MVZ); Garden Valley, 15 mi. NE Sharp, 1 (MVZ); Garden Valley, 14-1/2 mi. NE Sharp, 3 (MVZ); 8-1/2 mi. NE Sharp, 17 (MVZ). Lincoln County: E side Coal Valley, 14 mi. N Seeman Pass, 4850 ft., 4 (MVZ); 2 mi. SE Pioche, 1 (MVZ); E side Coal Valley, 10 mi. N Seeman Pass, 1 (MVZ); Desert Valley, 20 mi. SW Pioche, 1 (MVZ); 2 mi. N Panaca, 4800 ft., 17 (16 MVZ; 1 CAS); Desert Valley, 21 mi. W Panaca, 10 (MVZ); Crystal Spring, Pahranagat Valley, 4000 ft., 2 (MVZ); 10 mi. E Crystal Springs, 5000 ft., 1 (MVZ); 15 mi. S Groom Baldy, 1 (MVZ).
Utah: Millard County: 5 mi. S Garrison, 5400 ft., 12 (MVZ); Pine Valley, Sec. 33, T. 25 S, R. 17 W, 5000 ft., 16 (UU); Warm Cove, Sec. 34, T. 25 S, R. 18 W, 5500 ft., 2 (UU); Desert Range Experiment Station, 50 mi. W Milford, 5252 ft., 10 (9 UU; 1 BYU).
Perodipus montanus utahensis Merriam, Proc. Biol. Soc. Washington, 17:143, July 14, 1904.
Perodipus ordii utahensis, Goldman, Proc. Biol. Soc. Washington, 30:113, May 23, 1917.
Dipodomys ordii columbianus, Grinnell, Journ. Mamm., 2:96, May 2, 1921.
Dipodomys ordii columbianus, Hall, Univ. California Publ. Zool., 37:5, April 10, 1931 (part—the part from Ogden, Utah.)
Type.—Male, adult, no. 55115, U. S. Nat. Mus. Biol. Surv. Coll.; Ogden, Weber County, Utah; obtained on July 15, 1893, by Vernon Bailey, original no. 4085.
Range.—Between Great Salt Lake and the Wasatch Mountains from extreme northern Utah, south to northern Sevier County, Utah; marginal occurrences, all in Utah, are: Promontory Point, Ogden, St. John, Cedar Valley, Nephi, Aurora, Spring City, Provo and 4 mi. N Draper.
Diagnosis.—Size small (see measurements). Color dark, entire dorsal surface (a) between Cinnamon-Buff and Clay Color, purest on sides, flanks and cheeks, with heavy admixture of black in upper parts; arietiform markings, pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail blackish. Skull small; rostrum short and narrow; interorbital region moderately wide; interparietal large, spatulate anteriorly; lacrimal processes relatively large; braincase vaulted; zygomatic arches robust and straight; auditory bullae relatively, as well as actually, greatly inflated.
Comparisons.—From Dipodomys ordii marshalli, D. o. utahensis differs as follows: Size smaller; color darker; skull smaller; rostrum longer and narrower; interorbital region wider; lacrimal processes larger; interparietal region larger; auditory bullae less inflated; palate longer and narrower.
From Dipodomys ordii pallidus, D. o. utahensis differs in: Size smaller; color darker; skull smaller; rostrum shorter and narrower; interorbital region narrower; auditory bullae less inflated laterally, posteriorly and ventrally; interparietal region larger; lacrimal processes smaller; foramen magnum with ventral notch as opposed to rounded ventrally; pterygoid fossae circular as opposed to subcircular; palate longer and narrower.
From Dipodomys ordii cinderensis, D. o. utahensis differs as follows: Size larger, except hind foot which is shorter; color somewhat lighter, more reddish; skull larger; rostrum longer; nasals longer; braincase more vaulted; pterygoid fossae circular as opposed to subcircular; auditory bullae less inflated laterally but more inflated ventrally.
For comparisons with Dipodomys ordii columbianus, D. o. fetosus, D. o. marshalli, D. o. panguitchensis and D. o. cineraceus see accounts of those subspecies.
Remarks.—The systematic status of D. o. utahensis has long been in doubt. Several workers have, at various times, considered it to be indistinguishable from Dipodomys ordii columbianus. Durrant and Setzer (1945:29) recognized it as a valid subspecies and gave diagnostic characters by which it could be distinguished from D. o. columbianus.
Intergradation has been noted in specimens from the eastern and southeastern shores of Great Salt Lake. In color and the width of the auditory bullae these specimens are intermediate between D. o. utahensis and Dipodomys ordii marshalli but in the sum total of characters are referable to the latter. Specimens from 20 miles southwest of Nephi and from U. B. (Yuba) Dam are darker and intermediate in rostral width and breadth of the auditory bullae but are referable to Dipodomys ordii celeripes.
Specimens examined.—Total, 91, all from Utah, distributed as follows: Box Elder County: Promontory Point, 1 (USNM). Weber County: Ogden, 4293 ft., 20 (7 BYU; 7 MVZ; 4 USNM); Little Mountain, 1 (USNM). Davis County: Antelope Island, Great Salt Lake, 4250 ft., 5 (USNM). Tooele County: Bauer, 4500 ft., 6 (UU); St. John, 4300 ft., 4 (UU); Little Valley, Sheeprock Mountains, 5500 ft., 1 (UU); Clover Creek, Onaqui Mountains, 5500 ft., 1 (UU). Salt Lake County: Plain, 4 mi. N Draper, 4500 ft., 1 (UU). Utah County: Fairfield, Cedar Valley, 4800 ft., 15 (9 BYU; 6 UU); W Lake Mountains, 9 (BYU); Provo, 4510 ft., 10 (MVZ); Sand Dunes W Curtis Station, 4 (BYU). Juab County: Nephi, 2 (1 USNM; 1 MVZ); 4 mi. W Nephi, 1 (RH). San Pete County: Spring City, 4 (LACM). Sevier County: 1 mi. W Aurora, 5190 ft., 6 (5 UU; 1 USNM).
Perodipus ordi columbianus Merriam, Proc. Biol. Soc. Washington, 9:115, June 21, 1894.
Dipodomys phillipii, Baird, U. S. P. R. R. Expl. and Surveys, Gen. Rept., p. 412, 1857 (part—the part from Ft. Walla Walla, O. T.).
Cricetodipus ordii columbianus, Trouessart, Catalogus Mammalium, 1:581, 1897.
Perodipus ordii columbianus, Goldman, Proc. Biol. Soc. Washington, 30:113, May 23, 1917.
Dipodomys ordii columbianus, Grinnell, Journ. Mamm., 2:96, May 2, 1921.
Type.—Female, adult, no. 24181/31594, U. S. Nat. Mus., Dept. of Agriculture, Coll.; Umatilla, Plains of Columbia, Umatilla County, Oregon; obtained on October 18, 1890, by Clark P. Streator, original no. 386.
Range.—Southeastern Washington, eastern Oregon, southwestern Idaho, northwestern, northern and most of northeastern Nevada; marginal occurrences are: in Washington, 2 mi. SW Paterson, 4 mi. E Burbank and Wallula; in Oregon, 7 mi. E Madras, 2 mi. E Prineville and Guano Valley; in California, Eagleville, Red Rock, Observation Peak, 5 mi. E Litchfield, Honey Lake, Vinton; in Nevada, 3-1/2 mi. E Flanigan, 6 mi. S Pahrum Peak, 2-1/2 mi. E and 11 mi. N Gerlach, 2 mi. SW Quinn River Crossing, 1 mi. SE Tuscarora, 5 mi. N Beowawe, 1/2 mi. S Beowawe, 2-1/2 mi. NE Smiths Creek Ranch, Bells Ranch, 5 mi. W Austin, 8 mi. W Eureka, 4 mi. S Shoshone, 5 mi. SE Greens Ranch, and 22 mi. N Deeth; in Idaho, 8 mi. W Rodgerson, 6 mi. SW American Falls, 4 mi. NE American Falls, 5 mi. NW Michaud, Arco, Hammett and Payette; in Oregon, Umatilla.
Diagnosis.—Size medium (see measurements). Color dark, entire dorsal surface (16") between Pinkish Cinnamon and Cinnamon-Buff, purest on sides and flanks with strong admixture of black on upper parts; cheeks white; arietiform markings, pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, blackish. Skull medium in size; rostrum relatively [Pg 545] long and narrow; nasals long and expanded distally; interorbital region narrow; lacrimal processes small; braincase but slightly vaulted; auditory bullae relatively greatly inflated; pterygoid fossae circular; zygomatic arches robust.
Comparisons.—From Dipodomys ordii utahensis, D. o. columbianus differs as follows: Color lighter, dorsal and ventral stripes of tail brownish as opposed to black; ventral stripe of tail, in most specimens, incomplete to end of pencil as opposed to complete; lacrimal processes smaller; extension of premaxillae dorsad wider; interparietal generally narrower, smaller, quadrangular and truncate anteriorly, as opposed to spatulate and rounded anteriorly; braincase less vaulted; zygomatic arches bowed laterally as opposed to straight; styloid processes larger; cutting edge of upper incisors narrower.
From Dipodomys ordii fetosus, D. o. columbianus differs in: Hind foot shorter; color generally darker; skull larger; rostrum shorter and wider; nasals somewhat shorter; braincase more vaulted; auditory bullae less inflated; interparietal region wider; ventral border of foramen magnum more rounded (less indented); zygomatic arches straight as opposed to bowed laterally.
For comparisons with Dipodomys ordii inaquosus, Dipodomys ordii monoensis and Dipodomys ordii celeripes see accounts of those subspecies.
Remarks.—In most subspecies of Dipodomys ordii having large geographic ranges, there is a certain amount of variation, but D. o. columbianus remains relatively stable throughout its large range. Where extreme variation is noted in D. o. columbianus it is obviously the result of intergradation. This intergradation is noted between Dipodomys ordii monoensis and D. o. columbianus in Lyon County, Nevada, where the animals are referable to the former race cranially but to the latter on the basis of color. Animals from Kelton, Utah, are in color as Dipodomys ordii marshalli and cranially combine characters of D. o. columbianus, D. o. utahensis and D. o. marshalli. The sum total of their characters places them with D. o. marshalli. Durrant and Setzer (1945:29), and the present writer in this account, do not record any animals from Utah as belonging to D. o. columbianus. Nevertheless, the influence of D. o. columbianus is seen in the animals from Kelton, Utah.
Specimens examined.—Total, 588, distributed as follows:
Washington: Benton County: Blalock Island, 2 mi. SW Paterson, 2 (MVZ). Walla Walla County: 4 mi. E Burbank, 4 (MVZ); 2 mi. SSE Burbank, 27 (MVZ); Wallula, 7 (4 MVZ; 3 DRD).
Oregon: Gilliam County: Arlington, 1 (MVZ). Morrow County: 2-1/2 mi. SW Irigon, 6 (MVZ); Cecil, 1 (MVZ). Umatilla County: Umatilla, 10 (MVZ). Jefferson County: 7 mi. E Madras, 2 (DRD). Crook County: 2 mi. NE Prineville, 1 (MVZ); 7 mi. W Prineville, 8 (MVZ); 4 mi. SW Prineville, 23 (MVZ); Crooked River, at mouth of Bear Creek, 8 (MVZ). Lake County: Fort Rock, 1 (DRD); NE edge Alkali Lake, 8 (MVZ); 9 mi. S Adel, mouth 20 Mile Creek, 1 (MVZ); Guano Ranch, Guano Valley, 2 (DRD). Harney County: Malheur Lake, 9 (LACM); Narrows, 1 (DRD); 1 mi. S Narrows, 3 (MVZ); 5 mi. SW Narrows, 19 (MVZ); Smith Creek, 10 mi. SE Diamond, 1 (MVZ); Lake Alvord, 1 (MVZ); 1-1/2 mi. E Denio, 4 (MVZ). Malheur County: 3 mi. N Vale, 2 (MVZ); 8 mi. N Jordan Valley, 1 (MVZ); 1/2 mi. S Rome, 5 (MVZ); 1 mi. S Rome, 11 (MVZ).
Idaho: Payette County: Payette, 1 (DRD). Butte County: Arco, 1 (MVZ). Elmore County: Hammett, 1 (MVZ). Bingham County: 5 mi. E Shelley, 1 (MVZ); Blackfoot, 5 (MVZ); 3 mi. S Blackfoot, 6 (MVZ); Aberdeen, 1 (MVZ). Owyhee County: S bank Snake River, Homedale, 13 (MVZ); Indian Cove, 4 (MVZ); 5 mi. SE Murphy, 3 (MVZ); Castle Creek, 8 mi. S Oreana, 1 (MVZ); S Fork Owyhee River, 12 mi. N Nevada line, 1 (MVZ). Minidoka County: Acequia, 8 (MVZ); 4 mi. N Rupert, 2 (MVZ). Twin Falls County: Salmon Creek, 8 mi. W Rodgerson, 7 (MVZ). Power County: 5 mi. NW Michaud, 4 (MVZ); 4 mi. NE American Falls, 4 (MVZ); 6 mi. SW American Falls, 10 (MVZ).
California: Modoc County: Eagleville, 12 (CAS); 2 mi. E Eagleville, 7 (CAS); 2-1/2 mi. E Eagleville, 24 (CAS); 3 mi. E Eagleville, 6 (CAS); 5 mi. E Eagleville, 3 (CAS). Lassen County: 2 mi. W Red Rock P. O., 1 (MVZ); 1 mi. W Red Rock P. O., 4 (MVZ); Dransfield, 6 mi. E Ravendale, 5300 ft., 4 (CAS); 7 mi. E Ravendale, 1 (MVZ); 7 mi. N Observation Peak, 21 (MVZ); 6 mi. N Observation Peak, 7 (MVZ); Observation Peak, 31 (MVZ); Pete's Valley, 4 (MVZ); 5 mi. E Litchfield, 12 (MVZ); 6 mi. W Wendell, 3975 ft., 3 (CAS); 8 mi. SW Wendell, 5 (CAS); Honey Lake, 2 (LACM); 4-1/2 mi. WNW Stacy, 5 (MVZ); 3 mi. NW Warm Springs, 1 (MVZ). Plumas County: Beckwith, 1 (DRD); 2 mi. WNW Vinton, 2 (MVZ); Vinton, 18 (14 LACM; 4 MVZ); 1 mi. E Vinton, 7 (MVZ); near Vinton, 21 (DRD).
Nevada: Washoe County: 3 mi. N Vya, 3 (MVZ); 4-1/2 mi. NE Painted Point, 4 (MVZ); 8-1/2 mi. E Vya, 1 (MVZ); 3 mi. E Painted Point, 1 (MVZ); Long Valley Ranch, 3 mi. S Vya, 1 (MVZ); 10-1/2 mi. S Vya, 1 (MVZ); 11 mi. S Vya, 2 (MVZ); 13 mi. S Vya, 4 (MVZ); Hausen, 4800 ft., 8 (MVZ); 10 mi. SE Hausen, 4675 ft., 7 (MVZ); 2-1/2 mi. E and 11 mi. N Gerlach, 4050 ft., 4 (MVZ); Smoke Creek, 9 mi. E California line, 3900 ft., 5 (MVZ); 40° 28' N Lat., 6 mi. E California line, 4000 ft., 3 (MVZ); Horse Canyon, 3 mi. NW Pahrum Peak, 5000 ft., 1 (MVZ); Fox Canyon, 6 mi. S Pahrum Peak, 4800 ft., 4 (MVZ); N side Sand Pass, 3950 ft., 2 (MVZ); 4 mi. NW Flanigan, 4200 ft., 1 (MVZ); 3-1/2 mi. NW Flanigan, 4200 ft., 1 (MVZ); 3-1/2 mi. E Flanigan, 2 (MVZ); 2-3/4 mi. SW Pyramid, 4300 ft., 2 (MVZ). Humboldt County: 1 mi. S Denio, Oregon, 4200 ft., 6 (MVZ); Quinn River Crossing, 4100 ft., 1 (MVZ). Elko County: Mary's River, 22 mi. N Deeth, 3 (MVZ); 1 mi. SE Tuscarora, 5900 ft., 2 (MVZ); 5 mi. W Halleck, 5200 ft., 7 (MVZ); 3 mi. W Halleck, 5300 ft., 2 (MVZ). Lander County: 1 mi. E Battle Mountain, 1 (MVZ); Reese River Valley, 6 mi. N Austin, 2 (MVZ); Malloy Ranch, 5 mi. W Austin, 3 (MVZ); 2-1/2 mi. NE Smiths Creek Ranch, 5800 ft., 1 (MVZ); Campbell Creek, 6900 ft., 3 (MVZ); Campbell Creek Ranch, 5500 ft., 8 (MVZ). Eureka County: 5 mi. N Beowawe, 7 (MVZ); 1/2 mi. S Beowawe, 1 (MVZ); Pine Creek, 2 mi. E Palisades, 7 (MVZ); Evans, 4 (MVZ); Winzell, 3 (MVZ); 4 mi. SE Romano, Diamond Valley, 1 (MVZ); 8 mi. W Eureka, 12 (MVZ). White Pine County: 5 mi. SE Greens Ranch, Steptoe Valley, 1 (MVZ); Cherry Creek, 6600 ft., 2 (MVZ); 1 mi. E Illipah, 6100 ft., 3 (MVZ); 5-1/2 mi. SE Ely, 6500 ft., 5 (MVZ); Spring Valley, 5900 ft., 4 mi. S Shoshone, 6 (MVZ). Nye County: Bells Ranch, Reese River, 6890 ft., 1 (MVZ).
Type.—Male, adult, no. 90029, Museum of Zoology, University of Michigan; San Juan, 12 mi. W Lerdo, 3,800 ft., Durango, Mexico; obtained on March 1, 1946, by F. and F. Hammerstrom.
Range.—Known only from the type locality.
Diagnosis.—Size small (see measurements). Color pale, entire dorsal surface (c) between Light Ochraceous-Buff and Ochraceous-Buff, purest on sides, flanks and cheeks; upper parts but slightly washed with black; arietiform markings, lateral parts of pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, brownish; medial part of pinnae of ears flesh-colored. Skull medium in size; rostrum wide and relatively long; braincase but slightly inflated; least interorbital width great; interparietal region relatively narrow; upper incisors strongly recurved; zygomatic arches relatively heavy and slightly bowed laterally; auditory bullae greatly inflated ventrally, laterally and posteriorly.
Comparisons.—From Dipodomys ordii ordii, D. o. idoneus differs in: Size [Pg 547] smaller; color lighter; skull larger; nasals longer and somewhat more inflated distally as opposed to straight; interorbital width greater; breadth across maxillary arches greater; interparietal region narrower; auditory bullae more inflated posteriorly, laterally and ventrally and projecting anteriorly into orbit; extension of nasals anterior to upper incisors less; zygomatic arches heavier and more bowed laterally; paroccipital processes smaller; foramen magnum elongate as opposed to nearly circular.
From Dipodomys ordii obscurus, D. o. idoneus differs as follows: Size smaller; color lighter; skull larger, rostrum wider; nasals shorter and more expanded distally; interorbital region wider; width across maxillary arches greater; interparietal region narrower; auditory bullae more inflated laterally, posteriorly and ventrally; zygomatic arches shorter, heavier and less bowed laterally.
From Dipodomys ordii palmeri, D.o. idoneus differs in: Size smaller; color lighter; skull larger; rostrum shorter and wider; nasals shorter and more expanded distally; interorbital region wider; auditory bullae more inflated laterally, posteriorly and ventrally; zygomatic arches heavier.
From Dipodomys ordii fuscus, D. o. idoneus differs as follows: Size smaller; color lighter; skull smaller; rostrum narrower and shorter; interorbital width greater; braincase more vaulted; nasals longer; interparietal region generally wider; auditory bullae less inflated; breadth across maxillary arches greater; zygomatic arches heavier and more bowed laterally; pterygoid fossae ovoid as opposed to subcircular; occipital condyles more anterior to dorsal evagination when viewed ventrally.
Dipodomys ordii idoneus differs from Dipodomys ordii attenuatus in larger size, generally darker color and larger skull.
Remarks.—This subspecies of Dipodomys ordii is paler than either of the subspecies with adjoining geographic ranges. Measurements of the skull, in proportion to external measurements, are larger than in any other subspecies of Dipodomys ordii. Cranially, this new subspecies is more closely allied to Dipodomys ordii fuscus but in color appears to be much closer to Dipodomys ordii attenuatus from which it differs markedly in cranial features.
Specimens examined.—Total, 3, all from Durango, distributed as follows: San Juan, 12 mi. W Lerdo, 3 (UM).
Dipodomys ordii priscus Hoffmeister, Proc. Biol. Soc. Washington, 55:167, December 31, 1942.
Perodipus longipes, Allen, Bull. Amer. Mus. Nat, Hist., 8:246, November, 1896 (part—the part from Kinney Ranch, Wyoming).
Perodipus montanus richardsoni, Cary, N. Amer. Fauna, 33:141, August 17, 1911 (part—the part from Lay, Colorado).
Periodipus [Dipodomys] ordii luteolus, Svihla, Journ. Mamm.; 12:262, August, 1931 (part—the part from Linwood, Utah).
Type.—Male, no. 89119, Mus. Vert. Zool., Univ. California; Kinney Ranch, 21 mi. S Bitter Creek, 7100 ft., Sweetwater County, Wyoming; obtained on September 16, 1939, by Donald T. Tappe, original no. 766.
Range.—Southwestern Wyoming, extreme northeastern Utah and northwestern Colorado; marginal occurrences are: in Wyoming, 7 mi. N Ft. Washakie and 10 mi. SW Granger; in Colorado, Lay and Snake River south of Sunny Peak.
Diagnosis.—Size medium (see measurements). Color light, entire dorsal surface between (c) Light Ochraceous-Buff and Ochraceous-Buff, purest on sides, flanks and cheeks; upper parts slightly suffused with black; arietiform markings, pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, grayish to blackish in different individuals. Skull medium in size; rostrum long and narrow; nasals long; auditory bullae relatively narrow and skull long, giving appearance of much narrower skull than is actually the case; zygomata weak; upper incisors moderately wide at cutting edge.
Comparisons.—From Dipodomys ordii uintensis, D. o. priscus differs as follows: Hind foot longer; color lighter in all pigmented areas, and skull smaller in all measurements taken.
From Dipodomys ordii sanrafaeli, D. o. priscus differs in: Color decidedly less red in upper parts; ventral stripe of tail not continuous to end of pencil; nasals longer; auditory bullae less inflated.
For comparisons with Dipodomys ordii luteolus and Dipodomys ordii terrosus see accounts of those subspecies.
Remarks.—The characters of this subspecies are stable throughout most of its geographic range. It is not known to intergrade with adjacent forms. One specimen, a male, available from the Snake River, south of Sunny Peak, Colorado, however, is not typical of D. o. priscus, in that it has a wider and deeper rostrum than any other specimen from the entire range. Between D. o. priscus and D. o. luteolus, the skull, but not the color, provides diagnostic differences.
The Red Desert of Wyoming is mostly sandy but there are large exposures of rock and gravel. At the type locality of D. o. priscus rockier soil predominates but is interspersed with stabilized sand dunes where the kangaroo rats are abundant. Traps set on the rocky soils, in a continuous line between the dunes, yielded no kangaroo rats.
Kangaroo rats in the area of the Kinney Ranch, Wyoming, apparently form a good source of food for owls, since many bones (jaws and limb elements) were found in owl pellets at the bases of cliffs approximately 5 miles northeast of the ranch buildings.
Specimens examined.—Total, 72, distributed as follows:
Wyoming: Fremont County: 7 mi. N Fort Washakie, 1 (USBS); Fort Washakie, 1 (USBS); Wind River, 1 (USBS). Sweetwater County: Eden, 1 (USBS); 5 mi. E Rock Springs, 1 (UM); 10 mi. SW Granger, 13 (UM); Bitter Creek, 9 (6 AMNH; 3 CNHM); Kinney Ranch, 21 mi. S Bitter Creek, 18 (MVZ); Shell Creek, 25 mi. S Bitter Creek, 3 (CM); 30 mi. S Bitter Creek, 3 (KU); 33 mi. S Bitter Creek, 8 (KU). Carbon County: 20 mi. W Baggs, 2 (USBS).
Utah: Daggett County: E bank Green River, 4 mi. E Linwood, 4 (CM).
Colorado: Moffat County: Snake River, south of Sunny Peak, 1 (USBS); 20 mi. NW Sunbeam, 4 (CM); Bear River, Lay, 1 (USBS). Rio Blanco County: W side White River, 1 mi. N Rangely, 1 (CM).
Dipodomys ordii columbianus, Hall, Univ. California Publ. Zool., 37:5, April 10, 1931.
Type.—Male, adult, no. 1956, Museum of Zoology, University of Utah; Trout Creek, 4600 ft., Juab County, Utah; obtained on May 5, 1937, by Stephen D. Durrant, original no. 1168.
Range.—Eastern Nevada, western and west-central Utah, east to eastern Sevier County, Utah; marginal occurrences are: in Nevada, Tecoma, Cobre and 8 mi. SE Mt. Moriah; in Utah, E side Clear Lake, U. B. (Yuba) Dam, 10 mi. SW Nephi, and Trout Creek.
Diagnosis.—Size small (see measurements). Color pale, entire dorsal surface Pinkish Buff, purest on sides, flanks and cheeks, with but slight suffusion of black in upper parts; pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, brownish. Skull small; rostrum relatively short and wide; nasals relatively short; lacrimal processes small; braincase but slightly vaulted; auditory bullae but little inflated; interorbital region narrow; styloid processes project on ventral surface of auditory bullae beyond middle of external auditory meatus; zygomatic arches robust and bowed laterally; pterygoid fossae ovoid.
Comparisons.—From Dipodomys ordii marshalli, D. o. celeripes differs as follows: Size smaller, hind foot longer, tail shorter; color lighter; skull smaller; rostrum wider and shorter; interorbital region narrower; auditory bullae less inflated; lacrimal processes smaller; palate shorter; pterygoid fossae larger.
From Dipodomys ordii cinderensis, D. o. celeripes differs as follows: Size smaller, tail shorter; color lighter; rostrum wider and shorter; nasals shorter; interorbital width less; auditory bullae less inflated; foramen magnum wider and shorter; pterygoid fossae more circular.
From Dipodomys ordii columbianus, D. o. celeripes differs in: Size smaller; color lighter; rostrum wider and shorter; interorbital width less; interparietal region wider; auditory bullae, generally, less inflated; upper incisors longer and more robust; pterygoid fossae larger.
For comparisons with Dipodomys ordii fetosus, Dipodomys ordii pallidus and Dipodomys ordii panguitchensis see accounts of those subspecies.
Remarks.—It appears that wherever the range of D. o. celeripes meets that of an adjoining race, intergradation occurs freely. Practically all of the Nevadan specimens referable to this subspecies are intergrades with Dipodomys ordii columbianus in size and color. Specimens from west of Delta, Utah, are intermediate in size between Dipodomys ordii pallidus and D. o. celeripes but their cranial characters are more as in D. o. celeripes. Intergrades with Dipodomys ordii utahensis are discussed under the account of that subspecies.
The characters differentiating D. o. celeripes from any contiguous subspecies are not present in every specimen even in the type series.
Specimens examined.-Total, 96, distributed as follows:
Nevada: Elko County: 13 mi. N Montello, 5000 ft., 2 (MVZ); 1-1/2 mi. N Tecoma, 4900 ft., 6 (MVZ); Tecoma, 4900 ft., 8 (MVZ); Cobre, 6100 ft., 3 (MVZ); Salt Springs, 4200 ft., 1 (MVZ). White Pine County: 2 mi. W Smith Creek Cave, 6300 ft., Mt. Moriah, 2 (MVZ); Hendry Creek, 8 mi. SE Mt. Moriah, 6200 ft., 1 (MVZ).
Utah: Tooele County: Clifton Flat, 7 mi. SW Gold Hill, 6149 ft., 4 (UU); Parrish Ranch, 5 mi. N Ibapah, 5175 ft., 1 (UU); Ibapah, 5000 ft., 23 (UU). Juab County: Trout Creek, 4600 ft., 21 (19 UU; 1 USAC; 1 MVZ); 20 mi. SW Nephi, 2 (UU); U. B. (Yuba) Dam, Sevier River, 5000 ft., 10 (UU). Millard County: 4 mi. S Gandy, 5000 ft., 1 (MVZ); Smith Creek, 6 mi. S Gandy, 5400 ft., 2 (MVZ); Hendry Creek, 5000 ft., 17 mi. S Gandy, 4 (MVZ); Oak City, 5000 ft., 1 (UU); White Valley, 60 mi. W Delta, 1 (UU); 35 mi. W Delta, 1 (UU); East side Clear Lake, 4600 ft., 4 (3 UU; 1 USAC); 2 mi. E Clear Lake, 4600 ft., 2 (UU).
Dipodomys ordii cineraceus Goldman, Journ. Mamm., 20:352, August 14, 1939.
Type.—Male, no. 263890, U. S. Nat. Mus. Biol. Surv. Coll.; Dolphin Island, Great Salt Lake, 4250 ft., Box Elder County, Utah; obtained on June 4, 1938, by William H. Marshall, original no. 57.
Range.—Dolphin Island, Great Salt Lake, Utah.
Diagnosis.—Size medium (see measurements). Color pale, upper parts near Pale Pinkish Buff, but lightly washed with black, purest on sides and flanks; arietiform markings, pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, brownish. Skull medium in size; rostrum short; nasals relatively short; interparietal region relatively wide; auditory bullae but slightly inflated.
Comparisons.—From Dipodomys ordii utahensis, D. o. cineraceus differs in: Size smaller; color lighter; nasals longer; auditory bullae larger; total length of skull greater.
From Dipodomys ordii marshalli, D. o. cineraceus differs in: Size smaller; color darker; skull larger in all measurements taken.
Remarks.—The race D. o. cineraceus along with Dipodomys microps russeolus is found on Dolphin Island, Great Salt Lake, Utah. Dolphin Island is connected with the mainland by a bar that could allow animals from the island to disperse onto the mainland. There are no animals from the mainland, however, that are referable to D. o. cineraceus or to D. m. russeolus. The animals available from the mainland, at Kelton, although geographically near Dolphin Island, are referable to D. o. marshalli and to D. m. bonnevillei. The paucity of material from the mainland, adjacent to Dolphin Island, may be significant. Future trapping in this area is needed before we can be certain that D. o. cineraceus is isolated.
Specimens examined.—Total, 2, both from Utah, as follows: Box Elder County: Dolphin Island, Great Salt Lake, 4250 ft., 2 (USNM).
Dipodomys ordii marshalli Goldman, Proc. Biol. Soc. Washington, 50:223, December 28, 1937.
Type.—Female, adult, no. 262655, U. S. Nat. Mus. Biol. Surv. Coll.; Bird Island, Great Salt Lake, 4,300 ft., Tooele County, Utah; obtained on June 22, 1937, by W. H. Marshall, X-catalog no. 27969.
Range.—Bird, Carrington, Badger and Stansbury islands, Great Salt Lake; around western edge of Great Salt Lake north to Kelton, Box Elder County, Utah; around southern and southeastern edge of the lake to mouth of the Jordan River; marginal occurrences are: all in Utah, Kelton, 2 mi. W Grantsville, 14 mi. W Salt Lake City and Bird Island, Great Salt Lake.
Diagnosis.—Size medium (see measurements). Color pale, entire dorsal surface near Pinkish Buff, purest on sides, flanks and cheeks with but slight suffusion of black on upper parts; arietiform markings, pinnae of ears, plantar surfaces of hind feet, and dorsal and ventral stripes of tail, brownish. Skull medium in size; rostrum narrow and long; pterygoid fossae ovoid; cutting edge of upper incisors narrow; external auditory meatus round and small; jugal straight or nearly so.
Comparisons.—From Dipodomys ordii pallidus, D. o. marshalli differs in: Hind foot smaller; color darker, arietiform markings more distinct; dorsal and ventral stripes of tail more pronounced; skull smaller; palate shorter and wider; jugals lighter; external auditory meatus smaller; cutting edge of upper incisors narrower; nasals shorter and less flared distally.
From Dipodomys ordii utahensis, D. o. marshalli differs in: Size smaller; color lighter; skull smaller, except least interorbital width and depth of auditory bullae which are greater; cutting edge of upper incisors narrower; nasals less flared distally.
For comparison with Dipodomys ordii celeripes see account of that subspecies.
Remarks.—This subspecies was described from Bird Island, Great Salt Lake, and was thought by Marshall (1940:153), who collected the specimens, to be restricted to that island. However, specimens taken over a period of years on the nearby mainland, by students from the University of Utah, are referable to this subspecies. Although these animals from the mainland are referable to D. o. marshalli, they have some characteristics of D. o. utahensis from the eastern and southern mainland. Four specimens from Kelton, also on the mainland, are not typical of D. o. marshalli; nevertheless, more than half of their characters of taxonomic worth are as in D. o. marshalli to which the animals are referred.
Great Salt Lake has not been so effective in isolating the animals living on the islands as heretofore has been thought. After these supposedly isolated kinds of animals from Great Salt Lake were named, some other workers have shown several of the kinds to have [Pg 552] extensive ranges on the mainland. Some of the kinds ranging also on the mainland are: Dipodomys ordii marshalli, Dipodomys microps subtenuis and Peromyscus crinitus pergracilis. Of the animals named from the islands, I suppose that those which require the semiarid habitat found in this area will all be found to have large ranges on the adjacent mainland and that each of the kinds of animals which do not require the above habitat, and which lived in this region during the Pleistocene, will be found to be restricted to the island from which it was named.
Specimens examined.—Total, 47, all from Utah, distributed as follows: Box Elder County: Kelton, 4300 ft., 7 (5 MVZ; 2 UU). Tooele County: 2 mi. W Grantsville, 1 (CAS); Bird Island, Great Salt Lake, 4300 ft., 1 (USNM); Carrington Island, Great Salt Lake, 4300 ft., 1 (USNM); Stansbury Island, Great Salt Lake, 4300 ft., 10 (6 UU; 4 USNM). Salt Lake County: 18 mi. W Salt Lake City, 4260 ft., 16 (UU); 17 mi. W Salt Lake City, 4320 ft., 7 (UU); 16 mi. W Salt Lake City, 4300 ft., 3 (UU); 14 mi. W Salt Lake City, 4300 ft., 1 (UU).
Dipodomys ordii inaquosus Hall, Proc. Biol. Soc. Washington, 54:58, May 20, 1941.
Type.—Male, adult, no. 73580, Museum of Vertebrate Zoology, University of California; 11 mi. E and 1 mi. N Jungo, 4,200 ft., Humboldt County, Nevada; obtained on July 26, 1936, by Ward C. Russell, original no. 5026.
Range.—Southeastern Humboldt and northern Lander counties, Nevada; marginal occurrences, all in Nevada, are: 7 mi. N Winnemucca, 8 mi. E and 1 mi. N Jungo, 15 mi. SW Winnemucca, 23 mi. NW Battle Mountain, Izenhood, and 18 mi. NE Iron Point.
Diagnosis.—Size medium (see measurements). Color pale, entire dorsal surface Pinkish Buff, purest on sides and flanks with but slight suffusion of black on the upper parts; cheeks white; arietiform marks indistinct, pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, brownish. Skull medium in size; rostrum short and wide; nasals relatively long and expanded distally; braincase but slightly vaulted; interorbital width narrow; extension of premaxillae posterior to nasals relatively great; interparietal relatively large; pterygoid fossae subcircular; foramen magnum ovoid; auditory bullae relatively greatly expanded ventrally.
Comparisons.—From Dipodomys ordii columbianus, D. o. inaquosus differs as follows: Size larger; color lighter; skull larger; nasals longer; rostrum longer and wider; interorbital region narrower; braincase more vaulted; interparietal larger; auditory bullae more inflated ventrally; pterygoid fossae subcircular as opposed to nearly circular; zygomatic arches more robust.
From Dipodomys ordii monoensis, D. o. inaquosus differs in: Size larger; color lighter; skull larger; nasals longer; rostrum wider; interorbital width greater; braincase more vaulted; auditory bullae more inflated laterally, ventrally and posteriorly; interparietal region narrower; pterygoid fossae subcircular as opposed to circular; lacrimal processes larger; foramen magnum ovoid as opposed to circular.
Remarks.—In pallor D. o. inaquosus closely approaches Dipodomys ordii celeripes from eastern Nevada and western Utah. The two populations are separated, however, by a large area inhabited by the darker Dipodomys ordii columbianus. Specimens from 18 mi. NE Iron Point, Nevada, which Hall (1946:413) refers to D. o. columbianus, are intergrades in color, size and cranial characters and here are referred to D. o. inaquosus instead of to D. o. columbianus. Animals from Toulon are intergrades between D. o. inaquosus and Dipodomys ordii monoensis but are referable to the latter.
Specimens examined.—Total, 47, all from Nevada, distributed as follows: Humboldt County: 18 mi. NE Iron Point, 4600 ft., 1 (MVZ); 7 mi. N Winnemucca, 4400 ft., 4 (MVZ); 1 mi. N Winnemucca, 4600 ft., 8 (MVZ); 5 mi. NE Golconda, 7 (MVZ); 3 mi. SW Winnemucca, 4500 ft., 1 (MVZ); 4 mi. SW Winnemucca, 4500 ft., 1 (MVZ); 5 mi. SW Winnemucca, 4600 ft., 2 (MVZ); 8 mi. E and 1 mi. N Jungo, 4200 ft., 6 (MVZ); 11 mi. E and 1 mi. N Jungo, 4200 ft., 7 (MVZ); 10 mi. SW Winnemucca, 4500 ft., 1 (MVZ); 23 mi. NW Battle Mountain, 4 (MVZ). Pershing County: 15 mi. SW Winnemucca, 1 (MVZ). Lander County: Izenhood, 2 (MVZ); 3 mi. S Izenhood, 2 (MVZ).
Dipodomys ordii attenuatus Bryant, Occas. Papers, Museum of Zool., Louisiana State Univ., no. 5:65, November 10, 1939.
Type.—Male, no. 80429, Museum of Vertebrate Zoology, University of California; Mouth of Santa Helena Canyon, 2146 ft., Big Bend of the Rio Grande, Brewster County, Texas; obtained on November 19, 1938, by Adrey E. Borell, original no. 5581.
Range.—Big Bend region of Brewster County, Texas.
Diagnosis.—Size small (see measurements). Color light, entire dorsal surface between Pinkish Buff and Pinkish Cinnamon, purest on sides and flanks, upper parts but lightly mixed with black; cheeks and narrow outer margin of ear, pure white; arietiform markings absent; pinnae of ears, dorsal and ventral stripes of tail, brownish; plantar surfaces of hind feet, blackish. Skull small; rostrum long and narrow; nasals short; interparietal longer than wide; auditory bullae longer than deep.
Comparisons.—Compared with Dipodomys ordii ordii, D. o. attenuatus differs in: Size smaller; color lighter; skull smaller; nasals thinner; rostrum narrower; auditory bullae less inflated.
From Dipodomys ordii richardsoni, D. o. attenuatus differs in smaller size and lighter color. Skull smaller in all respects.
For comparison with Dipodomys ordii sennetti see account of that subspecies.
Fig. 27. Known occurrences and probable geographic range of the subspecies of Dipodomys ordii in the northwestern fourth of the range of the species. 9. D. o. panguitchensis 20. D. o. fetosus 28. D. o. inaquosus 10. D. o. uintensis 21. D. o. utahensis 31. D. o. longipes 11. D. o. sanrafaeli 22. D. o. columbianus 32. D. o. pallidus 12. D. o. fremonti 24. D. o. priscus 33. D. o. nexilis 13. D. o. monoensis 25. D. o. celeripes 34. D. o. cupidineus 17. D. o. chapmani 26. D. o. cineraceus 19. D. o. cinderensis 27. D. o. marshalli
Remarks.—This small race of Dipodomys ordii was known formerly only from the type locality. This study has shown it to range northward and slightly westward. At four places, 6, 20, and 47 miles south of Marathon and 10 miles west of San Vicente, Texas, this race can be said to be typical, in that specimens from these places agree with those from the type locality. At two other localities, 5 miles east of Toyahvale and Kent, Texas, animals show an intergrading tendency toward Dipodomys ordii ordii. The animals from these latter places are darker colored and are intermediate in size and cranial characters. Since these animals show more resemblance to D. o. ordii in the majority of characters they are referred to that race, rather than to D. o. attenuatus.
Specimens examined.—Total, 14, all from Texas, distributed as follows: Brewster County: 6 mi. S Marathon, 1 (USBS); 20 mi. S Marathon, 1 (USBS); Cooper's Well, 47 mi. S Marathon, 4 (MVZ); 10 mi. W San Vicente, 1 (UM); Johnson's Ranch, Big Bend Rio Grande, 4 (2 TCWC; 2 UM); mouth Santa Helena Canyon, 3 (1 TCWC; 2 MVZ).
Type.—Male, adult, no. 93886, U. S. Nat. Mus., Biol. Surv. Coll.; Juamave, Tamaulipas, Mexico; obtained on June 3, 1898, by E. W. Nelson and E. A. Goldman, original no. 12437.
Range.—Nuevo León, extreme northern Zacatecas, Tamaulipas and northern San Luis Potosí, Mexico; marginal occurrences are: Coahuila, La Ventura; Tamaulipas, Tula, Juamave and Nuevo Laredo.
Diagnosis.—Size medium (see measurements). Color dark, entire dorsal surface (a) between Cinnamon-Buff and Clay Color, purest on sides, flanks and cheeks, upper parts strongly suffused with black; posterior surfaces and fold of pinnae of ears, white; arietiform markings, plantar surfaces of hind feet, inside of pinnae of ears, dorsal and ventral stripes of tail, blackish. Skull of medium size; nasals relatively short; interorbital width narrow; auditory bullae relatively as well as actually greatly inflated posteriorly, laterally and ventrally; braincase vaulted; zygomatic arches light and straight; pterygoid fossae ovoid; rostrum short and wide.
Comparisons.—From Dipodomys ordii palmeri, D. o. fuscus differs in: Size larger; color lighter; skull larger; rostrum shorter and wider; interorbital width less; nasals shorter; auditory bullae more expanded laterally, posteriorly and ventrally; interparietal region narrower; pterygoid fossae more circular; upper incisors wider at cutting edge; zygomatic arches straight rather than bowed laterally.
From Dipodomys ordii obscurus, D. o. fuscus differs as follows: Size larger; color lighter; skull larger; rostrum wider and shorter; interorbital region wider; width across maxillary arches greater; auditory bullae more expanded laterally, posteriorly and ventrally; zygomatic arches light and straight instead of heavy and bowed laterally; pterygoid fossae more nearly circular; braincase more vaulted; foramen magnum smaller.
From Dipodomys ordii ordii, D. o. fuscus differs in: Size larger; color darker; skull larger; rostrum shorter and wider; nasals short and more flared distally; interorbital width greater; auditory bullae more inflated posteriorly, laterally and ventrally; interparietal region narrower; width across maxillary arches greater; pterygoid fossae larger and more nearly circular; braincase more vaulted.
From Dipodomys ordii sennetti, D. o. fuscus can be distinguished by larger [Pg 556] size, longer tail, darker color, and cranially by larger auditory bullae, longer nasals, wider interorbital region and greater width across maxillary arches.
Remarks.—This dark subspecies of Dipodomys ordii is most closely allied to Dipodomys ordii palmeri from, which it can be distinguished by the characters set forth under comparisons.
It seems that the Rio Grande serves as a barrier to the dispersal southward of kangaroo rats from the north side and vice versa. D. o. fuscus ranges from the type locality to Nuevo Laredo and has not been found on the north side of the river, being replaced there, up-river, by Dipodomys ordii medius and down-river by Dipodomys ordii sennetti.
Specimens examined.—Total, 53, distributed as follows:
Coahuila: San Juan Neponuceno, 5 mi. N La Ventura, 4 (MVZ); La Ventura, 7 (USBS).
Tamaulipas: Nuevo Laredo, 2 (USBS); Miquihuana, 14 (4 USBS; 9 MVZ; 1 AMNH); Juamave, 8 (USBS); Tula, 5 (UM); 3 mi. N Lulú, 5 (MVZ): Lulú, 8 (MVZ).
Dipodops longipes Merriam, N. Amer. Fauna, 3:72, September 11, 1890.
Perodipus ordii, Allen, Bull. Amer. Mus. Nat. Hist., 5:71, April 28, 1893 (part—the part from Bluff, Utah).
Cricetodipus longipes, Trouessart, Catalogus Mammalium, 1:581, 1897.
Perodipus longipes, Elliot, Field Columbian Museum, Zool. Ser., 2:239, 1901.
Perodipus ordii longipes, Goldman, Proc. Biol. Soc. Washington, 30:113, May 23, 1917.
Dipodomys ordii longipes, Grinnell, Journ. Mamm., 2:96, May 2, 1921.
Dipodomys ordii cleomophila Goldman, Journ. Washington Acad. Sci., 23:469, October 15, 1933, type from 5 miles northeast of Winona, Coconino County, Arizona.
Type.—Male, no. 17703/24639, U. S. Nat. Mus. Dept. Agric. Coll.; foot of Echo Cliffs, Painted Desert, Arizona; obtained on September 22, 1889, by C. Hart Merriam, original no. 512.
Range.—Southeastern Utah, northeastern Arizona to immediately south of the Little Colorado River, northwestern New Mexico and extreme southwestern Colorado; marginal occurrences are: in Arizona, 20 mi. NE Lees Ferry, Foot Echo Cliffs in Painted Desert, 5 mi. NE Winona, Winslow and Holbrook; in New Mexico, 10 mi. SW Quemado, Riley, Laguna, Chama Canyon and Shiprock.
Diagnosis.—Size large (see measurements). Color dark, entire dorsal surface between (16") Pinkish Cinnamon and Cinnamon-Buff, purest on sides and flanks with but slight suffusion of black on upper parts; cheeks white; arietiform markings, pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, brownish. Skull large; rostrum long and narrow; nasals long and thin; auditory bullae greatly inflated; styloid processes project on ventral surface of auditory bullae beyond middle of external auditory meatus.
Comparisons.—From Dipodomys ordii chapmani, D. o. longipes differs as follows: Size larger; color lighter; skull larger in all measurements taken; auditory bullae much more inflated.
From Dipodomys ordii cupidineus, D. o. longipes differs in: Size larger; color lighter; rostrum longer and narrower; nasals longer; interorbital width greater; breadth across zygomatic processes of maxillae greater; auditory bullae more inflated; zygomatic arches heavier and more bowed laterally.
For comparisons with Dipodomys ordii medius and Dipodomys ordii nexilis see accounts of those subspecies.
Remarks.—The large reddish Dipodomys ordii longipes is notably distinct from those subspecies which have adjoining ranges. The subspecies closest, geographically and morphologically, is Dipodomys ordii nexilis with which D. o. longipes intergrades, in size of body and color of pelage, north of the San Juan River, Utah. This intergradation is probably the result of a migration of D. o. longipes around the head waters of, or above the place in, the river where there is constantly flowing water. It is probably impossible for these animals to cross a constantly flowing stream as wide as the San Juan is in its lower reaches. The San Juan River, however, in 1896, and again in 1934, was so low that it ceased to flow below Aneth (Gregory, 1938:6, 19). Consequently, at these times, Dipodomys could have crossed the river from one side to the other, at least above the Goosenecks. Intergradation is noted also with Dipodomys ordii medius at Chama Canyon, New Mexico, where the color is like that of D. o. medius, although the cranial characters are as in D. o. longipes; specimens from Chama Canyon are referred to the latter. Ten miles southwest of Quemado and at Riley, New Mexico, intergradation with Dipodomys ordii ordii occurs in the shape of the nasals and the intermediate size of the skull. In color and external measurements these animals more closely resemble D. o. longipes.
Goldman (1933:469) described Dipodomys ordii cleomophila from 5 miles northeast of Winona, Coconino County, Arizona, and gave as distinctive characters "upper parts distinctly darker, ... black facial markings more distinct; skull slightly different." These comparative statements were made in reference to Dipodomys ordii longipes. While it is true that the animals which Goldman examined are darker and show some slight cranial differences, these animals, in my opinion, are not sufficiently distinct to warrant their separation from D. o. longipes. In the degree of expansion of the auditory bullae, the width of the rostrum, the length of the nasals and interorbital width, animals from 5 miles northeast of Winona (2 specimens) are within the range of individual variation shown by D. o. longipes. The color is darker, being more as in Dipodomys ordii chapmani. Specimens examined [Pg 558] from 10 miles north of Angell, Springerville, Tanner Tank and Cedar Ranch Wash, Arizona (listed by Goldman as referable to D. o. cleomophila), and those from Wupatki Ruins, Arizona, agree with D. o. longipes except that the width across the maxillary arches is greater. This greater width across the maxillary arches and the darker color of animals from Winona and Wupatki Ruins, in my opinion, afford insufficient basis for the recognition of the subspecies Dipodomys ordii cleomophila Goldman which, therefore, is placed as a synonym of Dipodomys ordii longipes (Merriam).
Specimens examined.—Total, 244, distributed as follows:
Utah: San Juan County: 1 mi. N Bluff, 3500 ft., 1 (UU); 1/2 mi. N Bluff, 3300 ft., 7 (UU); Bluff, 2 (UU); Johns Canyon, San Juan River, 5150 ft., 2 (UU).
Arizona: Coconino County: 20 mi. NE Lees Ferry, 1 (USNM); 2 mi. S Endische, 2 (MVZ); 2 mi. SE Endische Spring, Navajo Mountain, 1 (MVZ); 1/2 mi. W Navajo Spring, Echo Cliffs, 4 (MVZ); 5 mi. S Navajo Mountain, 2 (MVZ); 7 mi. SE Navajo Mountain, 1 (MVZ); Moa Vae, 10 (MVZ); 2 mi. S Grand Canyon Bridge, 1 (USBS); Tuba, 4 (1 USBS; 3 AMNH); 12 mi. above mouth of Moencopi Wash, 2 (USBS); 14 mi. NW Cedar Ridge, 1 (USBS); 5 mi. N Cameron, 1 (MVZ); Painted Desert, 6 mi. NE Cameron, 1 (MVZ); 5 mi. E Little Colorado River on Tuba road, 2 (MVZ); Cameron, 4200 ft., 17 (8 LACM; 9 MVZ); Wupatki Ruins, 4 (MVZ); Deadmans Flat, 6400 ft., NE San Francisco Mt., 3 (MVZ); 3 mi. NW Winona, 4 (USBS); 5 mi. NE Winona, 2 (USBS); 10 mi. N Angell, Walnut Tank, 8 (USBS); Cedar Ranch Wash, 3 mi. above mouth, 5 (USBS); Tanner Tank, 2 (USBS); Tanners Crossing, Painted Desert, 1 (USBS). Navajo County: Right fork, Segi-ot-Sosie Canyon, 11 mi. NW Kayenta, 2 (MVZ); Bat Woman Canyon, 13 mi. W Kayenta, 1 (MVZ); Kayenta, 4 (USBS); Segi Canyon, 12 mi. WSW Kayenta, 1 (MVZ); Segi Canyon, 13 mi. WSW Kayenta, 1 (MVZ); Oraibi, 6000 ft., 9 (4 USBS; 5 MVZ); Walpi, 2 (USBS); Keam Canyon, 12 (6 USBS; 3 USNM; 3 MVZ); Winslow, 6 (USBS); Zuni Well, 7-1/2 mi. N Adamana, 9 (MVZ); Holbrook, 10 (USBS); Long Canyon, 3 (AMNH); Bubbling Springs, 2 (AMNH). Apache County: Dinnehotso, 5000 ft., 3 (MVZ); Lukachukai, 1 (USBS); Chin Lee, 6 (USBS); 9 mi. SW Chin Lee, 1 (USBS); 7 mi. from mouth, Canyon de Chelly, 1 (USBS); Fort Defiance, 2 (USNM); Ganado, 1 (USBS); Springerville, 2 (USBS); 3 mi. SE Springerville, 5 (USBS); unspecified, 4 (2 USBS; 2 USNM); Trash Tank, Grand Canyon, 2 (USBS).
New Mexico: San Juan County: Ship Rock, 4 (USBS); Fruitland, 9 (USBS); Blanco, 1 (USBS); Chaco Canyon National Monument, 4 (MVZ). Rio Arriba County: Chama Canyon, 1 (USBS). McKinley County: 2 mi. W Mariano Lake, 3 (MVZ); Gallup, 6 (USBS); Wingate, 6 (USBS); Thoreau, 2 (USBS); Zuni River, 1 (USBS). Sandoval County: Juan Tafoya, 1 (USBS). Valencia County: 8 mi. SE Grants, 7 (UM); 9 mi. SSE Grants, 5 (UM); 2-1/2 mi. E El Morro P. O., Zuni Mts., 1 (LACM); Laguna, 1 (USBS); Laguna Indian Reservation, 7 (LACM). Catron County: Cougar Ranch, NE Adams Diggings, 1 (CAS); 10 mi. SW Quemado, 1 (USBS). Socorro County: 1 mi. S Bernardo, 1 (MVZ); Riley, 1 (USBS).
Dipodomys ordii pallidus Durrant and Setzer, Bull. Univ. Utah, 35 (no. 26):24, June 30, 1945.
Type.—Male, adult, no. 3526, Museum of Zoology, University of Utah; Old Lincoln Highway, 18 mi. SW Orr's Ranch in Skull Valley, 4,400 ft., Tooele County, Utah; obtained on June 6, 1940, by S. D. Durrant, original no. 1905.
Range.—Low valleys of west-central Utah in Tooele, Juab and Millard counties; marginal occurrences are: all in Utah, 18 mi. SW Orr's Ranch, 7 mi. S Fish Springs, Hinckley and Lynndyl.
Diagnosis.—Size medium (see measurements). Color light, entire dorsal surface Light Pinkish Cinnamon, purest on sides and flanks, with but slight suffusion of black on upper parts; cheeks white; arietiform markings, pinnae of ears, plantar surfaces of hind feet, dorsal and ventral stripes of tail, brownish. Skull large; auditory bullae long, wide and well inflated; external auditory meatus elongate with a notch on the dorsal border; nasals long and flaring distally.
Comparisons.—From Dipodomys ordii celeripes, D. o. pallidus differs in: Size larger; color generally darker, although some specimens are lighter; skull larger; nasals longer and more flared distally; interorbital width greater; auditory bullae larger; external auditory meatus larger.
From Dipodomys ordii fetosus, D. o. pallidus differs in: Size larger; color lighter; ventral stripe of tail indistinct as opposed to pronounced; skull larger; nasals longer and more flared distally; auditory bullae larger; external auditory meatus larger; palate shorter and broader.
For comparisons with Dipodomys ordii marshalli and Dipodomys ordii utahensis see accounts of those subspecies.
Remarks.—Among named subspecies of Dipodomys ordii, D. o. pallidus most closely resembles, morphologically, D. a. marshalli, its nearest geographic neighbor to the north and east. Intergradation with Dipodomys ordii utahensis is noted in color and intermediacy of body size of specimens from Clover Creek. The majority of cranial characters, however, show these animals to be referable to D. o. utahensis rather than to D. o. pallidus. Specimens taken at Lynndyl and Hinckley show intergradation in size of body, length and configuration of the nasals and the degree of inflation of the auditory bullae between Dipodomys ordii celeripes and D. o. pallidus. The majority of characters studied show these latter animals to be referable to D. o. pallidus. Specimens from 35 and 60 miles west of Delta, in size of body and shape and inflation of the auditory bullae, are intermediate between D. o. pallidus and D. o. celeripes. These specimens are here referred to D. o. celeripes.
Specimens examined.—Total, 34, all from Utah, distributed as follows: Tooele County: Old Lincoln Highway, 18 mi. SW Orr's Ranch in Skull Valley, 4400 ft., 9 (UU). Juab County: Fish Springs, 4400 ft., 4 (UU); 7 mi. S Fish Springs, 4400 ft., 4 (UU). Millard County: 1 mi. N Lynndyl, 4768 ft., 5 (UU); Lynndyl, 4768 ft., 1 (UU); Hinckley, 4600 ft., 11 (UU).
Dipodomys ordii nexilis Goldman, Journ. Washington, Acad. Sci., 23:470, October 5, 1933.
Perodipus longipes, Warren, Mammals of Colorado, p. 77, 1910 (part—the part from Coventry, Colorado).
Dipodomys ordii longipes, Moore, Journ. Mamm., 10:260, August, 1929 (part—the part from Monticello, Utah).
Type.—Male, adult, no. 149938, U. S. Nat. Mus. Biol. Surv. Coll.; 5 mi. W Naturita, Montrose County, Colorado; obtained on July 20, 1907, by Merritt Cary, original no. 1068.
Range.—West-central Colorado, southwest into San Juan County, Utah, north of the San Juan River; northwest into Grand County, Utah, to the [Pg 560] Colorado River; and westward probably as far as the Colorado River; marginal occurrences are: in Utah, Cisco, 18 mi. NE Moab, and Blanding; in Colorado, 5 mi. W Naturita, and Coventry.
Diagnosis.—Size medium (see measurements). Color dark, entire dorsal surface between (a) Cinnamon-Buff and Clay Color, purest on sides and flanks, upper parts strongly suffused with black; arietiform markings, pinnae of ears, plantar surfaces of hind feet, dorsal and ventral tail-stripes blackish. Lateral white stripes of tail less than one fourth of diameter of tail. Skull large; rostrum long and narrow; nasals long; auditory bullae strongly inflated; braincase not inflated; pterygoid fossae ovoid.
Comparisons.—From Dipodomys ordii sanrafaeli, D. o. nexilis differs in: Color darker; skull smaller in all measurements taken; auditory bullae noticeably smaller; width across zygomatic processes of maxillae less.
From Dipodomys ordii longipes, D. o. nexilis differs in: Size smaller; color darker in all respects; rostrum wider; breadth across auditory bullae less; interorbital width greater; breadth across zygomatic processes of maxillae less; cutting edge of upper incisors wider; zygomatic arch lighter and more nearly straight.
Comparison with Dipodomys ordii evexus is made in account of that subspecies.
Remarks.—This animal is apparently not abundant at any place in its range. Two different attempts, by me, to obtain topotypes were unsuccessful. A single specimen was obtained and that was only a skull saved from a mutilated animal that was taken away from a rattlesnake. The habitat at the type locality is such that a person would not expect it to be inhabited by kangaroo rats. The soil is a heavy clay with a generous admixture of stones but in isolated spots there are light sandy soils which should be suitable for kangaroo rats. Even so, 500 traps set in the area of the type locality over a period of two nights yielded no Dipodomys.
This subspecies of Dipodomys ordii, inhabiting west-central Colorado and southeastern Utah, is darker than any other subspecies with which its range comes in contact. The races to both the north and south are larger, with the exception of the hind foot which is longer in D. o. nexilis than in either D. o. sanrafaeli or D. o. longipes. The affinities of D. o. nexilis are with D. o. longipes rather than with any of the other known subspecies of Dipodomys ordii.
Intergradation between D. o. nexilis and D. o. longipes is shown by animals from Bluff, San Juan County, Utah, which, however, are referable to the latter. In animals from sixteen miles northwest of Moab, Grand County, Utah, there is intergradation in size of skull and in color between D. o. nexilis and D. o. sanrafaeli. The specimens are referred to D. o. sanrafaeli.
D. o. nexilis is apparently prevented from extending its range to the northward by the presence there of D. o. sanrafaeli. To the south D. o. nexilis encounters D. o. longipes and the San Juan River. To the east it is limited by the Rocky Mountains and to the west by the deep canyons of the Colorado River.
Warren (1942:183) did not apply the name D. o. nexilis to Ord kangaroo rats in Colorado. He listed specimens from Montrose, Montezuma and Mesa counties as being referable to D. o. longipes. The specimens from Montezuma County, Colorado, probably are D. o. longipes and those from Montrose and Mesa counties (for which Warren gives no precise localities) are D. o. nexilis.
Specimens examined.—Total, 35, distributed as follows:
Utah: Grand County: Cisco, 4 (CM); 18 mi. NE Moab, 6000 ft., 1 (UU). San Juan County: 15 mi. N Monticello, 4 (MVZ); Blanding, 1 (UU).
Colorado: Delta County: Hotchkiss, 1 (USBS). Montrose County: Paradox, 1 (DRD); Bedrock, 4 (AMNH); 5 mi. W Naturita, 2 (USBS); Coventry, 3 (1 USBS; 2 AMNH).
Dipodomys ordii cupidineus Goldman, Journ. Washington Acad. Sci., 14:372, September 19, 1924.
Type.—Male, no. 243093, U. S. Nat. Mus. Biol. Surv. Coll.; Kanab Wash, at southern boundary of Kaibab Indian Reservation, Arizona; obtained on October 12, 1922, by E. A. Goldman, original no. 23384.
Range.—Northeastern Arizona and south-central Utah; marginal occurrences are: in Utah, Escalante, near Paria and S of Virgin; in Arizona, 6 mi. N Wolf Hole, 20 mi. S Wolf Hole, 5 mi. S Trumbull Spring and 10 mi. S Jacobs Pools in Houserock Valley.
Diagnosis.—Size large (see measurements). Color relatively dark, entire dorsal surface between (16") Pinkish Cinnamon and Cinnamon-Buff, purest on sides and flanks, upper parts washed with black; arietiform markings, pinnae of ears, dorsal and ventral stripes of tail, plantar surfaces of hind feet, blackish. Skull large; auditory bullae well inflated; interorbital width relatively narrow; rostrum long and narrow; nasals long and slightly flared distally; pterygoid fossae ovoid; external auditory meatus small and ovoid; jugal light and relatively straight.
Comparisons.—From Dipodomys ordii chapmani, D. o. cupidineus differs in: Size larger; color lighter in all pigmented areas; skull larger; rostrum shorter and wider; nasals shorter; auditory bullae more inflated, particularly median tympanic portion; least interorbital width less.
From Dipodomys ordii nexilis, D. o. cupidineus differs as follows: Size smaller; color lighter in all pigmented areas; skull smaller; rostrum relatively longer and narrower; least interorbital width less; auditory bullae less inflated; breadth across maxillary arches less; foramen magnum circular as opposed to ovoid; cutting edge of upper incisors narrower; zygomatic arches lighter.
For comparisons with Dipodomys ordii cinderensis, Dipodomys ordii panguitchensis and Dipodomys ordii longipes see accounts of those subspecies.
Remarks.-This subspecies is prevented from intergrading, to the east, with D. o. longipes by the deep canyons of the Colorado River, with D. o. cinderensis by the Virgin River and with D. o. panguitchensis by a series of high mountain ranges. Intergradation does, however, occur with D. o. sanrafaeli in animals from Escalante, Garfield County, Utah, but they are referable to D. o. cupidineus. The Colorado River, farther to the north, again serves as a barrier in preventing intergradation between D. o. nexilis and D. o. cupidineus.
Specimens examined.—Total, 122, distributed as follows:
Utah: Garfield County: Mouth of Calf Creek, Escalante River, 3 (BYU); Ten Mile Spring, 3 (BYU); Escalante, 3 (BYU). Washington County: near Short Creek road, south of town of Virgin, 18 (RH). Kane County: near Paria, 1 (UU); Willow Tank Springs, 7 (BYU); 8 mi. NW Kanab, 4800 ft., 1 (UU); near Sand Dunes, 7 (RH); Kanab, 15 (12 MVZ; 2 BYU; 1 UU); 1 mi. S Kanab, 4400 ft., 2 (UU).
Arizona: Mohave County: near S Boundary Kaibab Indian Reservation, 4 (MVZ); 6 mi. N Wolf Hole, 8 (MVZ); 4 mi. N Wolf Hole; 3 mi. NW Diamond Butte, 7 (USBS); 20 mi. S Wolf Hole, 1 (USBS); 1 mi. W Diamond Butte, 1 (USBS); 6 mi. N Mt. Trumbull, 1 (USBS); Nixon Springs, 6250 ft., W Base Mt. Trumbull, 6 (3 USBS; 3 MVZ); 5 mi. S Trumbull Spring, 1 (USBS). Coconino County: Fredonia, 3 (USBS); 2 mi. W Lees Ferry, 3 (MVZ); Jacobs Pools, 4000 ft., Houserock Valley, 15 (MVZ); 10 mi. S Jacobs Pools, Houserock Valley, 2 (MVZ); Houserock Valley, 10 (USBS).
Dipodops ordii palmeri Allen, Bull. Mus. Comp. Zool., 8:187, 1881.
Cricetodipus ordii palmeri, Trouessart, Catalogus Mammalium, 1:581, 1897.
Perodipus ordii palmeri, Goldman, Proc. Biol. Soc. Washington, 30:113, May 23, 1917.
Dipodomys ordii palmeri, Grinnell, Journ. Mamm., 2:96, May 2, 1921.
Cotypes.—Two males, adults, nos. 5886 and 5887, Mus. Comp. Zool.; San Luis Potosí, Mexico; obtained on May 1, 1878, and September 1, 1878, respectively, by Dr. Edward Palmer. (Types not seen.)
Range.—Eastern Zacatecas, Aguascaliente, northern Jalisco, San Luis Potosí, Hidalgo, and probably Querétaro; marginal occurrences are: Zacatecas, Canitas and Berriozobal; Aguascaliente, 1 mi. N Chicalote; Jalisco, 9 mi. N Encarnación; Guanajuato, Celaya; Hidalgo, Irolo.
Diagnosis.—Size small (see measurements). Color dark, entire dorsal surface (h) between Cinnamon and Sayal Brown, purest on sides and flanks, upper parts strongly suffused with black; posterior surfaces and fold of pinnae of ears, white; arietiform markings, plantar surfaces of hind feet, inside of pinnae of ears, dorsal and ventral stripes of tail, blackish. Skull small; nasals long; rostrum long and narrow; interorbital region relatively wide; degree of inflation of auditory bullae about average for species; zygomatic arches light and bowed laterally; pterygoid fossae subcircular; braincase but slightly vaulted.
Comparisons.—From Dipodomys ordii ordii, D. o. palmeri differs as follows: Size somewhat larger; color darker; skull larger; nasals longer and flaring distally; interorbital width greater; narrower across auditory bullae; interparietal region narrower; breadth across maxillary arches greater; zygomatic arches [Pg 563] bowed laterally as opposed to relatively straight; pterygoid fossae more nearly circular; rostrum longer and narrower.
From Dipodomys ordii sennetti, D. o. palmeri differs in larger size, darker color, longer tail and longer skull.
For comparisons with Dipodomys ordii obscurus and Dipodomys ordii fuscus see accounts of those subspecies.
Remarks.—Specimens from Berriozobal, Zacatecas, in the width of the rostrum and the configuration of the nasals, are intermediate between Dipodomys ordii fuscus and D. o. palmeri. In color these animals are more nearly like typical representatives of D. o. palmeri. The animals from Jalisco and Guanajuato are not typical but the characters differentiating them from D. o. palmeri are merely modifications of such slight degree that they all have been placed with that subspecies.
Specimens examined.—Total, 56, distributed as follows:
Zacatecas: Canitas, 3 (USBS); Berriozobal, 10 (USBS).
San Luis Potosí: Potrero Santa Ana, 7.6 mi. S Matehuala, 2 (MVZ); Jesus Maria, 12 (USBS); 2 mi. NW San Luis Potosí, 8 (MVZ).
Aguascalientes: 1 mi. N Chicalote, 1900 m., 4 (MVZ).
Jalisco: 9 mi. N Encarnación, 1900 m., 1 (MVZ).
Guanajuato: Celaya, 8 (USBS).
Hidalgo: Ixmiquilpan, 1 (USBS); Irolo, 12 (USBS).
1. There are thirty-five recognizable subspecies of the species Dipodomys ordii of which four are herein, for the first time, recognized by name. Three subspecies, D. o. oklahomae, D. o. sennetti and D. o. compactus, previously were regarded by most authors as full species.
2. The species Dipodomys ordii is divisible into six complexes, or groups, of subspecies on both geographic and morphological bases.
3. Dipodomys ordii is the most generalized Recent species of the genus.
4. The extremes of geographic variation in Dipodomys ordii are greater than in any other species of the genus.
5. Color, at least in Dipodomys ordii, does not seem to be correlated with amount of moisture but rather with color of soil.
6. Clinal variation, from north to south, is shown in the decreasing length of the nasals. This decrease in length of nasals and resultant decrease in size of the nasal chamber may be correlated with the decrease of humidity of the environment.
7. Subspeciation has been enhanced by the late Quaternary mountain building which was prevalent over the western United States.
8. In general, the most primitive kinds of Dipodomys ordii occur at the periphery of the range of the species.
9. Natural selection plus geographical and ecological isolation has undoubtedly been operative in speciation and in subspeciation.
10. Dipodomys deserti is found to be the most specialized species in the genus.
11. Six, rather than nine, groups of species are recognized, on the basis of morphology, as comprising the genus Dipodomys.
12. The center of dispersal for the genus Dipodomys appears to have been in the southwestern United States and the adjoining part of Mexico. A secondary center of differentiation is apparent in the low, hot valleys of central California.
13. Parallel development of species is noted between Dipodomys of the parental center and Dipodomys of the isolated valleys of central California; Dipodomys in the California center is the less specialized.
14. The trend of the dipodomyines, as indicated by the fossil record, has been toward a saltatorial specialization with consequent morphological changes.
15. The morphological change in the direction of saltatorial specialization is clearly evident in the compacting and aligning of the viscera as well as in the lengthening of the distal segments of the hind legs and the tail, tufting of the tail, enlargement of the auditory bullae, shortening of the neck and fusion of the cervical vertebrae for stability and other modifications of the skeleton.
Measurements (in Millimeters) of Dipodomys ordii
| Total length | Length of tail | Length of hind foot | Greatest length of skull | Greatest breadth across bullae | Breadth across maxillary arches | Width of rostrum | Length of nasals | Least interorbital width | Basilar length | |
D. o. richardsoni 1-1/2 mi. N Beaver, Oklahoma (KU) |
||||||||||
♂ Mean (4) |
251.2 |
131.5 |
40.5 |
41.0 |
24.7 |
22.0 |
4.3 |
14.9 |
13.0 |
26.3 |
Maximum |
256 |
136 |
41 |
41.8 |
25.1 |
22.3 |
4.5 |
15.4 |
13.2 |
27.3 |
Minimum |
243 |
133 |
40 |
40.4 |
24.4 |
21.5 |
4.2 |
14.6 |
12.7 |
26.0 |
♀ KU 17962 |
253 |
135 |
42 |
40.6 |
25.4 |
22.6 |
4.6 |
15.1 |
14.2 |
25.7 |
KU 17963 |
246 |
133 |
40 |
39.4 |
24.0 |
... |
4.3 |
14.1 |
12.5 |
25.6 |
KU 17963 |
255 |
136 |
41 |
41.2 |
25.5 |
22.1 |
4.5 |
15.1 |
13.8 |
26.2 |
D. o. oklahomae 2-1/4 mi. S Norman, Oklahoma (USBS) |
||||||||||
♂ Mean (4) |
239.7 |
130.0 |
39.5 |
39.0 |
23.9 |
21.5 |
4.3 |
13.7 |
13.3 |
24.9 |
Maximum |
254 |
140 |
42 |
40.3 |
24.6 |
22.4 |
4.4 |
14.3 |
13.6 |
25.8 |
Minimum |
227 |
125 |
37 |
38.0 |
23.0 |
20.9 |
4.2 |
13.4 |
12.8 |
23.9 |
♀ USBS 265456 |
245 |
133 |
40 |
39.1 |
24.5 |
22.0 |
4.4 |
14.4 |
13.5 |
25.3 |
OU 20140 |
234 |
120 |
39 |
38.7 |
23.8 |
21.7 |
4.4 |
14.3 |
13.5 |
24.8 |
D. o. compactus 19 mi. S Port Aransas, Mustang Island, Texas (TCWC) |
||||||||||
♂ Mean (10) |
234.6 |
119.5 |
39.3 |
37.0 |
21.8 |
19.4 |
4.1 |
13.8 |
11.9 |
24.2 |
Maximum |
251 |
134 |
41 |
37.9 |
22.5 |
20.8 |
4.2 |
14.6 |
12.3 |
25.3 |
Minimum |
228 |
112 |
38 |
35.6 |
21.3 |
18.2 |
3.9 |
12.9 |
11.6 |
22.8 |
♀ Mean (7) |
231.0 |
116.0 |
39.0 |
37.1 |
22.4 |
19.7 |
4.0 |
13.9 |
... |
24.2 |
Maximum |
235 |
120 |
41 |
38.2 |
22.9 |
20.5 |
4.1 |
14.7 |
... |
24.8 |
Minimum |
224 |
110 |
38 |
36.0 |
21.7 |
19.1 |
3.8 |
13.2 |
... |
23.8 |
D. o. sennetti 2 mi. S Riviera, Texas (TCWC) |
||||||||||
♂ Mean (5) |
217.6 |
112.2 |
35.8 |
37.2 |
23.4 |
20.1 |
4.0 |
13.6 |
13.1 |
24.2 |
Maximum |
222 |
115 |
38 |
38.2 |
24.1 |
20.7 |
4.3 |
14.4 |
13.2 |
24.6 |
Minimum |
208 |
104 |
34 |
36.3 |
23.0 |
19.4 |
3.8 |
13.0 |
12.6 |
23.8 |
♀ Mean (4) |
218.3 |
112.0 |
36.0 |
37.2 |
23.3 |
20.1 |
4.0 |
13.9 |
13.1 |
24.1 |
Maximum |
226 |
115 |
37 |
37.7 |
23.9 |
20.8 |
4.1 |
14.4 |
13.4 |
24.4 |
Minimum |
209 |
108 |
35 |
36.7 |
22.8 |
19.0 |
3.9 |
13.3 |
12.6 |
23.5 |
D. o. evexus Salida, Colorado (AMNH) |
||||||||||
♂ AMNH 28802 |
267.0 |
140.0 |
42.0 |
... |
23.0 |
... |
4.2 |
... |
... |
25.0 |
AMNH 28804 |
271 |
156 |
43 |
39.3 |
23.5 |
22.1 |
4.4 |
14.6 |
14.0 |
25.8 |
AMNH 28805 |
252 |
137 |
39 |
... |
... |
21.5 |
4.2 |
... |
... |
24.5 |
♀ Mean (4) |
261.0 |
147.0 |
42.2 |
38.0 |
23.5 |
21.6 |
4.3 |
14.1 |
13.4 |
24.6 |
Maximum |
264 |
149 |
43 |
38.1 |
24.0 |
22.1 |
4.4 |
14.2 |
13.8 |
25.0 |
Minimum |
258 |
144 |
42 |
37.9 |
22.7 |
20.8 |
4.2 |
14.0 |
13.0 |
24.3 |
D. o. medius Santa Rosa, New Mexico (USBS) |
||||||||||
♂ Mean (6) |
258.5 |
143.3 |
38.0 |
39.5 |
25.2 |
21.4 |
4.1 |
14.2 |
12.9 |
24.8 |
Maximum |
266 |
151 |
40 |
40.6 |
25.9 |
22.5 |
4.2 |
15.2 |
13.1 |
25.6 |
Minimum |
251 |
1321 |
37 |
38.9 |
24.8 |
20.6 |
4.0 |
13.5 |
12.8 |
24.1 |
♀ USBS 118527 |
261.0 |
145.0 |
40.0 |
40.4 |
25.4 |
22.2 |
4.5 |
15.7 |
13.5 |
25.7 |
USBS 127310 |
... |
... |
... |
38.1 |
24.3 |
20.8 |
4.1 |
13.5 |
13.5 |
... |
| [Pg 566]
D. o. obscurus Rio Sestin, Durango (AMNH) |
||||||||||
♂ Mean (6) |
... |
... |
... |
36.3 |
22.9 |
19.7 |
3.7 |
13.1 |
12.6 |
22.8 |
Maximum |
... |
... |
... |
38.0 |
23.4 |
21.0 |
3.9 |
13.5 |
13.0 |
23.9 |
Minimum |
... |
... |
... |
35.2 |
22.6 |
19.1 |
3.6 |
11.9 |
12.4 |
21.7 |
♀ AMNH 20945 |
... |
... |
... |
36.5 |
22.8 |
19.9 |
3.8 |
13.1 |
12.9 |
22.3 |
AMNH 20951 |
... |
... |
... |
36.0 |
22.3 |
19.6 |
3.5 |
13.0 |
... |
22.5 |
AMNH 20958 |
... |
... |
... |
35.9 |
22.7 |
19.4 |
3.4 |
12.6 |
12.3 |
22.7 |
D. o. terrosus Jordan, Montana |
||||||||||
♂ AMNH 41442 |
280.0 |
155.0 |
44.0 |
42.7 |
26.5 |
23.6 |
4.0 |
15.2 |
14.5 |
27.0 |
AMNH 41443 |
267.0 |
155.0 |
40.0 |
40.5 |
24.8 |
21.1 |
4.0 |
14.5 |
13.1 |
25.1 |
AMNH 41444 |
279.0 |
162.0 |
41.0 |
40.8 |
25.7 |
21.6 |
4.4 |
14.6 |
13.4 |
25.9 |
♀ AMNH 41441 |
265.0 |
149.0 |
40.5 |
41.4 |
25.4 |
22.2 |
4.3 |
14.7 |
13.0 |
26.8 |
MVZ 25658 |
273.0 |
154.0 |
41.0 |
41.3 |
25.1 |
22.4 |
4.0 |
14.9 |
13.8 |
26.0 |
D. o. fremonti Torrey, Utah |
||||||||||
♂ CM 15663 |
250.0 |
141.0 |
40.0 |
37.0 |
23.3 |
19.3 |
3.6 |
13.5 |
12.0 |
23.2 |
CM 15670 |
248.0 |
136.0 |
38.0 |
37.3 |
23.8 |
19.8 |
3.8 |
13.7 |
11.6 |
23.5 |
♀ CM 15666 |
258.0 |
141.0 |
39.0 |
37.5 |
23.5 |
19.9 |
3.7 |
13.8 |
11.9 |
23.5 |
CM 15667 |
252.0 |
140.0 |
39.0 |
36.8 |
23.5 |
20.5 |
3.7 |
13.5 |
11.5 |
23.2 |
D. o. uintensis 2 mi. N Fruitland, Utah |
||||||||||
♂ CM 11634 |
253.0 |
140.0 |
40.0 |
37.9 |
23.3 |
20.3 |
3.9 |
13.5 |
12.5 |
23.9 |
CM 11640 |
260.0 |
150.0 |
41.0 |
38.2 |
24.2 |
20.6 |
4.0 |
13.2 |
12.5 |
24.5 |
D. o. sanrafaeli 12 mi. SW Green River, Utah |
||||||||||
♂ CM 15649 |
265.0 |
144.0 |
41.0 |
... |
... |
... |
... |
14.1 |
... |
... |
♀ CM 15647 |
253.0 |
138.0 |
42.0 |
... |
... |
... |
... |
13.5 |
... |
... |
D. o. panguitchensis 1 mi. S Panguitch, Utah |
||||||||||
♂ RH 2151 |
257.0 |
145.0 |
41.0 |
... |
... |
... |
... |
13.3 |
12.0 |
... |
RH 2152 |
252.0 |
135.0 |
40.0 |
... |
... |
... |
... |
13.5 |
11.9 |
... |
♀ RH 2153 |
240.0 |
132.0 |
38.0 |
... |
... |
... |
... |
12.7 |
11.3 |
... |
D. o. monoensis 5 mi. N Benton Station, California (MVZ) |
||||||||||
♂ MVZ 26993 |
232.0 |
125.0 |
40.0 |
36.6 |
23.7 |
20.0 |
3.7 |
13.0 |
... |
23.4 |
MVZ 26995 |
240.0 |
125.0 |
39.0 |
36.8 |
23.0 |
19.8 |
3.6 |
13.4 |
11.5 |
23.3 |
MVZ 26997 |
230.0 |
122.0 |
39.0 |
37.2 |
23.2 |
20.1 |
3.6 |
13.4 |
12.0 |
23.9 |
♀ Mean (5) |
228.0 |
125.4 |
38.4 |
36.6 |
23.1 |
19.7 |
3.5 |
13.1 |
11.8 |
23.4 |
Maximum |
240.0 |
137.0 |
39.0 |
37.2 |
24.2 |
20.0 |
3.7 |
13.2 |
12.2 |
23.6 |
Minimum |
220.0 |
120.0 |
38.0 |
36.0 |
22.6 |
19.3 |
3.4 |
13.0 |
11.5 |
23.0 |
| [Pg 567]
D. o. ordii Near El Paso, Texas (USNM) |
||||||||||
♂ Mean (5) |
... |
... |
... |
37.1 |
24.1 |
19.6 |
3.7 |
13.0 |
12.7 |
23.5 |
Maximum |
... |
... |
... |
38.1 |
24.8 |
20.5 |
3.8 |
13.2 |
13.3 |
24.3 |
Minimum |
... |
... |
... |
36.3 |
23.4 |
18.8 |
3.5 |
12.8 |
12.1 |
22.4 |
♀ Mean (6) |
235.2 |
128.0 |
37.4 |
37.3 |
24.0 |
19.6 |
3.7 |
13.3 |
12.8 |
23.5 |
Maximum |
245.0 |
136.0 |
39.0 |
38.0 |
24.1 |
20.0 |
3.8 |
13.5 |
13.4 |
24.4 |
Minimum |
222.0 |
120.0 |
35.0 |
36.3 |
24.0 |
19.1 |
3.6 |
13.0 |
12.4 |
23.0 |
D. o. luteolus 1 mi. NE Casper, Wyoming (KU) |
||||||||||
♂ Mean (12) |
265.6 |
152.2 |
42.2 |
38.9 |
24.1 |
20.8 |
4.3 |
13.9 |
13.0 |
24.6 |
Maximum |
281.0 |
163.0 |
43.0 |
39.5 |
25.0 |
22.1 |
4.4 |
14.5 |
13.7 |
25.7 |
Minimum |
254.0 |
145.0 |
42.0 |
37.5 |
23.8 |
19.9 |
4.2 |
13.0 |
12.5 |
24.0 |
♀ Mean (7) |
260.7 |
148.0 |
41.0 |
38.6 |
24.2 |
20.9 |
4.3 |
13.9 |
12.9 |
24.7 |
Maximum |
269.0 |
153.0 |
43.0 |
40.5 |
25.7 |
21.9 |
4.4 |
14.9 |
13.8 |
25.5 |
Minimum |
250.0 |
139.0 |
40.0 |
37.6 |
23.0 |
20.0 |
4.2 |
13.3 |
12.5 |
24.0 |
D. o. extractus 1 mi. E Samalayuca, Chihuahua (MVZ) |
||||||||||
♂ Mean (11) |
235.1 |
128.2 |
37.3 |
37.5 |
24.1 |
20.1 |
3.5 |
13.2 |
13.0 |
23.4 |
Maximum |
251.0 |
142.0 |
39.0 |
38.6 |
24.6 |
20.8 |
3.8 |
13.8 |
13.5 |
24.4 |
Minimum |
224.0 |
121.0 |
35.0 |
36.2 |
23.2 |
19.3 |
3.3 |
12.6 |
12.4 |
22.3 |
♀ MVZ 76568 |
251.0 |
132.0 |
39.0 |
39.6 |
25.4 |
22.2 |
3.7 |
13.8 |
14.1 |
24.8 |
MVZ 76569 |
244.0 |
140.0 |
37.0 |
37.3 |
23.6 |
20.0 |
3.6 |
13.5 |
12.8 |
24.0 |
MVZ 76570 |
230.0 |
120.0 |
37.0 |
35.7 |
23.4 |
19.5 |
3.2 |
12.4 |
12.1 |
22.5 |
D. o. chapmani Camp Verde, Arizona (USBS) |
||||||||||
♂ Mean (6) |
258.0 |
148.0 |
39.0 |
38.9 |
24.2 |
21.1 |
3.8 |
14.2 |
13.5 |
24.3 |
Maximum |
268.0 |
157.0 |
41.0 |
40.1 |
24.7 |
22.1 |
3.9 |
14.8 |
13.9 |
24.7 |
Minimum |
244.0 |
138.0 |
38.0 |
38.1 |
23.6 |
20.5 |
3.8 |
13.3 |
13.1 |
24.1 |
♀ Mean (4) |
255.4 |
151.0 |
38.2 |
37.1 |
23.9 |
20.1 |
3.8 |
13.4 |
13.5 |
23.3 |
Maximum |
262.0 |
152.0 |
39.0 |
37.7 |
24.3 |
20.1 |
3.9 |
13.7 |
14.9 |
23.5 |
Minimum |
245.0 |
144.0 |
37.0 |
36.0 |
23.7 |
20.1 |
3.6 |
13.1 |
12.6 |
23.0 |
D. o. montanus Fort Garland, Colorado (USBS) |
||||||||||
♂ Mean (11) |
252.0 |
140.5 |
41.0 |
37.7 |
24.2 |
20.1 |
4.1 |
13.6 |
12.8 |
23.8 |
Maximum |
263.0 |
150.0 |
42.0 |
39.0 |
25.0 |
21.0 |
4.5 |
14.0 |
13.2 |
24.7 |
Minimum |
233.0 |
131.0 |
39.0 |
36.6 |
23.6 |
19.0 |
3.9 |
12.7 |
12.4 |
23.0 |
♀ Mean (11) |
256.0 |
141.0 |
40.8 |
37.2 |
23.9 |
19.6 |
4.1 |
13.4 |
12.5 |
23.7 |
Maximum |
259.0 |
145.0 |
42.0 |
38.4 |
24.4 |
21.4 |
4.3 |
14.1 |
13.3 |
24.4 |
Minimum |
237.0 |
132.0 |
40.0 |
36.0 |
23.0 |
18.2 |
3.9 |
12.3 |
11.9 |
22.6 |
D. o. cinderensis 11 mi. SE Lund, Utah |
||||||||||
♂ MVZ 102059 |
244.0 |
135.0 |
41.0 |
35.7 |
23.2 |
19.2 |
3.6 |
12.2 |
11.2 |
22.7 |
MVZ 102056 |
237.0 |
130.0 |
41.0 |
36.4 |
23.7 |
20.6 |
3.5 |
13.0 |
12.1 |
23.1 |
♀ MVZ 102057 |
229.0 |
122.0 |
41.0 |
36.3 |
23.5 |
20.2 |
3.6 |
13.0 |
11.5 |
23.4 |
MVZ 102058 |
241.0 |
127.0 |
40.0 |
36.8 |
24.0 |
20.0 |
3.6 |
13.2 |
11.7 |
23.2 |
| [Pg 568]
D. o. fetosus 2 mi. N Panaca, Nevada (MVZ) |
||||||||||
♂ Mean (4) |
242.5 |
133.7 |
42.3 |
37.4 |
23.7 |
20.0 |
3.7 |
13.3 |
11.7 |
23.7 |
Maximum |
249.0 |
140.0 |
43.0 |
37.7 |
24.0 |
20.3 |
3.8 |
13.9 |
12.0 |
23.8 |
Minimum |
233.0 |
126.0 |
42.0 |
36.8 |
23.2 |
19.5 |
3.7 |
12.7 |
11.5 |
23.6 |
♀ Mean (4) |
229.0 |
125.5 |
40.6 |
35.8 |
23.3 |
19.5 |
3.6 |
12.8 |
11.3 |
22.5 |
Maximum |
235.0 |
129.0 |
41.5 |
37.0 |
24.1 |
20.0 |
3.6 |
13.3 |
11.6 |
23.2 |
Minimum |
224.0 |
122.0 |
40.0 |
34.3 |
23.0 |
19.0 |
3.6 |
11.9 |
11.0 |
21.9 |
D. o. utahensis Ogden, Utah |
||||||||||
♂ MVZ 44005 |
243.0 |
133.0 |
40.5 |
35.9 |
23.1 |
19.2 |
3.5 |
12.8 |
12.4 |
22.4 |
♀ MVZ 44006 |
238.0 |
133.0 |
38.0 |
36.1 |
... |
19.8 |
3.7 |
13.5 |
12.2 |
22.9 |
MVZ 44007 |
240.0 |
137.0 |
39.0 |
35.6 |
22.9 |
... |
3.6 |
12.8 |
11.7 |
22.4 |
MVZ 44008 |
241.0 |
134.0 |
40.0 |
36.4 |
23.5 |
19.8 |
3.6 |
13.3 |
12.3 |
... |
D. o. columbianus Umatilla, Oregon |
||||||||||
♂ MVZ 45317 |
243.0 |
140.0 |
40.0 |
37.2 |
23.6 |
20.0 |
3.8 |
13.5 |
12.2 |
23.4 |
MVZ 45318 |
236.0 |
137.0 |
40.0 |
36.8 |
23.5 |
20.1 |
3.6 |
13.2 |
... |
23.4 |
MVZ 45322 |
240.0 |
132.0 |
40.0 |
36.4 |
23.4 |
19.9 |
3.7 |
13.0 |
12.1 |
23.3 |
♀ MVZ 45314 |
239.0 |
130.0 |
39.0 |
35.8 |
22.7 |
19.8 |
3.7 |
13.2 |
11.9 |
22.7 |
MVZ 45315 |
251.0 |
140.0 |
41.0 |
36.6 |
23.8 |
19.6 |
3.6 |
13.3 |
12.0 |
23.1 |
D. o. idoneus 12 mi. W Lerdo, Durango |
||||||||||
♂ UM 90027 |
... |
... |
35.0 |
37.3 |
24.3 |
20.7 |
3.8 |
13.0 |
14.1 |
23.1 |
UM 90029 |
230.0 |
129.0 |
35.0 |
37.2 |
23.5 |
20.0 |
3.6 |
13.5 |
13.4 |
23.3 |
D. o. priscus 33 mi. S Bitter Creek, Wyoming (KU) |
||||||||||
♂ Mean (7) |
259.0 |
148.0 |
44.0 |
39.1 |
24.3 |
20.7 |
4.1 |
14.3 |
13.1 |
24.9 |
Maximum |
265.0 |
152.0 |
45.0 |
40.4 |
25.1 |
21.2 |
4.3 |
15.2 |
13.6 |
25.5 |
Minimum |
251.0 |
144.0 |
43.0 |
38.0 |
23.7 |
20.0 |
4.0 |
13.8 |
12.7 |
23.7 |
♀ Mean (4) |
257.0 |
147.0 |
43.0 |
39.4 |
24.6 |
20.8 |
4.2 |
14.3 |
13.1 |
24.7 |
Maximum |
264.0 |
152.0 |
45.0 |
40.4 |
25.2 |
21.9 |
4.3 |
14.9 |
13.3 |
25.2 |
Minimum |
249.0 |
138.0 |
40.0 |
38.1 |
23.5 |
20.1 |
4.1 |
14.0 |
12.7 |
24.0 |
D. o. priscus 33 mi. S Bitter Creek, Wyoming (KU) |
||||||||||
♂ Mean (4) |
208.7 |
109.0 |
40.2 |
35.3 |
22.9 |
19.3 |
3.6 |
12.6 |
11.2 |
... |
Maximum |
220.0 |
115.0 |
41.0 |
36.6 |
23.6 |
20.2 |
3.8 |
13.5 |
11.5 |
... |
Minimum |
203.0 |
100.0 |
39.0 |
34.0 |
22.1 |
18.4 |
3.5 |
11.7 |
11.1 |
... |
♀ UU 1957 |
219.0 |
110.0 |
39.0 |
35.3 |
23.4 |
19.1 |
3.6 |
11.9 |
11.8 |
22.4 |
UU 1961 |
223.0 |
120.0 |
40.0 |
37.0 |
23.6 |
20.4 |
3.6 |
13.1 |
12.1 |
23.9 |
D. o. cineraceus Dolphin Island, Great Salt Lake, Utah |
||||||||||
♀ USNM 263893 |
228.0 |
129.0 |
39.0 |
37.2 |
23.4 |
19.7 |
3.5 |
13.6 |
11.6 |
23.0 |
USNM 263894 |
230.0 |
132.0 |
38.0 |
37.1 |
23.7 |
20.1 |
3.6 |
13.4 |
11.9 |
23.6 |
| [Pg 569]
D. o. marshalli Stansbury Island, Great Salt Lake, Utah |
||||||||||
♂ UU 2968 |
238.0 |
128.0 |
40.0 |
36.4 |
23.5 |
... |
3.5 |
13.0 |
12.0 |
23.2 |
UU 2969 |
241.0 |
136.0 |
40.0 |
36.3 |
23.7 |
19.2 |
3.7 |
12.9 |
... |
23.0 |
♀ UU 2972 |
... |
... |
... |
35.9 |
23.5 |
19.9 |
3.6 |
12.9 |
11.9 |
23.1 |
D. o. inaquosus 11 mi. E and 1 mi. N Jungo, Nevada (MVZ) |
||||||||||
♂ Mean (4) |
243.0 |
137.0 |
40.0 |
36.6 |
23.5 |
19.8 |
3.7 |
13.5 |
11.8 |
23.1 |
Maximum |
247.0 |
140.0 |
40.5 |
36.8 |
24.0 |
20.0 |
3.8 |
13.8 |
12.1 |
23.5 |
Maximum |
238.0 |
131.0 |
39.0 |
36.4 |
23.3 |
19.5 |
3.6 |
13.1 |
11.5 |
23.0 |
♀ MVZ 73577 |
242.0 |
139.0 |
40.5 |
36.4 |
23.9 |
19.2 |
3.7 |
13.2 |
11.4 |
22.9 |
MVZ 73579 |
231.0 |
130.0 |
40.5 |
36.3 |
23.2 |
19.0 |
3.5 |
12.7 |
11.0 |
23.2 |
MVZ 73582 |
254.0 |
138.0 |
41.0 |
38.2 |
23.9 |
20.0 |
3.6 |
13.9 |
12.2 |
... |
D. o. attenuatus Johnson's Ranch, Texas |
||||||||||
♂ TCWC 3633 |
235.0 |
126.0 |
41.0 |
37.3 |
24.1 |
20.1 |
3.7 |
13.7 |
... |
23.4 |
TCWC 3634 |
237.0 |
136.0 |
37.0 |
35.5 |
22.1 |
18.8 |
3.5 |
13.2 |
12.2 |
22.3 |
UM 79121 |
245.0 |
143.0 |
37.3 |
35.7 |
22.3 |
19.1 |
3.6 |
12.4 |
12.9 |
23.3 |
♀ UM 79122 |
238.0 |
140.0 |
38.0 |
34.6 |
22.5 |
18.5 |
3.5 |
11.9 |
13.0 |
21.9 |
D. o. fuscus Juamave, Tamaulipas (USBS) |
||||||||||
♂ Mean (4) |
248.0 |
149.0 |
38.5 |
37.6 |
24.1 |
20.2 |
3.6 |
12.6 |
12.7 |
23.6 |
Maximum |
254.0 |
152.0 |
40.0 |
38.3 |
24.7 |
20.4 |
3.7 |
13.1 |
12.9 |
23.7 |
Minimum |
241.0 |
146.0 |
38.0 |
37.0 |
23.5 |
19.7 |
3.6 |
12.3 |
12.5 |
23.6 |
♀ Mean (4) |
244.7 |
146.2 |
38.5 |
37.4 |
24.1 |
19.9 |
3.4 |
12.9 |
12.7 |
23.2 |
Maximum |
254.0 |
155.0 |
40.0 |
37.7 |
24.5 |
20.5 |
3.6 |
13.0 |
12.9 |
23.4 |
Minimum |
240.0 |
138.0 |
38.0 |
37.2 |
23.7 |
19.2 |
3.3 |
12.7 |
12.4 |
23.2 |
D. o. longipes Kayenta, Arizona |
||||||||||
♂ USBS 247915 |
255.0 |
145.0 |
41.0 |
39.3 |
25.5 |
20.7 |
3.9 |
13.8 |
13.0 |
24.3 |
USBS 247916 |
242.0 |
138.0 |
40.0 |
38.4 |
25.4 |
20.6 |
3.8 |
13.3 |
12.7 |
24.6 |
♀ USBS 247552 |
255.0 |
135.0 |
40.0 |
39.9 |
25.7 |
21.2 |
4.0 |
13.8 |
... |
25.4 |
D. o. pallidus 18 mi. SW Orr's Ranch, Utah (UU) |
||||||||||
♂ Mean (4) |
236.6 |
131.3 |
42.3 |
37.6 |
24.2 |
20.4 |
3.8 |
13.4 |
11.8 |
23.6 |
Maximum |
240.0 |
134.0 |
43.0 |
37.9 |
24.6 |
20.9 |
4.0 |
13.6 |
12.1 |
24.2 |
Minimum |
230.0 |
128.0 |
41.0 |
37.0 |
23.7 |
19.8 |
3.7 |
12.9 |
11.5 |
23.4 |
♀ UU 3528 |
229.0 |
132.0 |
40.0 |
36.6 |
23.5 |
20.0 |
3.7 |
12.5 |
12.0 |
... |
D. o. nexilis 5 mi. W Naturita, Colorado |
||||||||||
♂ USBS 149940 |
261.0 |
156.0 |
44.0 |
38.7 |
25.1 |
18.9 |
4.1 |
13.8 |
11.4 |
24.6 |
♀ USBS 149941 |
265.0 |
142.0 |
45.0 |
40.5 |
26.3 |
21.5 |
4.2 |
14.4 |
13.1 |
25.3 |
| [Pg 570]
D. o. cupidineus Houserock Valley, Arizona (USBS) |
||||||||||
♂ Mean (6) |
239.0 |
138.0 |
39.8 |
36.9 |
24.1 |
19.3 |
3.8 |
12.8 |
12.0 |
23.6 |
Maximum |
244.0 |
146.0 |
41.0 |
37.5 |
24.9 |
20.1 |
4.0 |
13.0 |
12.6 |
24.3 |
Minimum |
225.0 |
130.0 |
38.5 |
36.3 |
23.5 |
18.7 |
3.6 |
12.6 |
11.7 |
23.2 |
♀ Mean (4) |
244.7 |
141.0 |
40.0 |
37.4 |
24.2 |
19.7 |
3.8 |
13.2 |
12.3 |
23.9 |
Maximum |
247.0 |
147.0 |
40.5 |
37.7 |
24.4 |
20.0 |
3.9 |
13.5 |
12.6 |
24.6 |
Minimum |
240.0 |
137.0 |
39.5 |
37.1 |
23.9 |
19.5 |
3.7 |
12.9 |
12.0 |
23.7 |
D. o. palmeri Jesus María, San Luis Potosí (USBS) |
||||||||||
♂ Mean (6) |
242.0 |
141.6 |
37.6 |
36.7 |
23.6 |
20.0 |
3.3 |
12.9 |
12.9 |
22.6 |
Maximum |
255.0 |
157.0 |
39.0 |
37.3 |
24.4 |
20.8 |
3.6 |
13.6 |
13.4 |
23.0 |
Minimum |
229.0 |
129.0 |
35.0 |
35.9 |
23.2 |
19.4 |
3.0 |
12.5 |
12.6 |
22.0 |
♀ Mean (6) |
239.5 |
139.6 |
37.1 |
36.4 |
23.8 |
19.9 |
3.4 |
12.8 |
13.1 |
22.7 |
Maximum |
252.0 |
148.0 |
38.0 |
37.3 |
23.9 |
20.0 |
3.6 |
13.3 |
13.2 |
23.5 |
Minimum |
225.0 |
128.0 |
36.0 |
35.9 |
23.7 |
19.5 |
3.3 |
12.2 |
13.0 |
22.0 |
Anderson, R. M. 1947. Catalogue of Canadian Recent mammals. National Museum of Canada, Bull. 102, Biol. Ser., 31, pp. v + 238, January 24, 1947.
Bailey, V.1905. Biological survey of Texas. N. Amer. Fauna, 25:1-222, 24 figs., Government Printing Office, October 24, 1905.
Benson, S. B.1933. Concealing coloration among some desert rodents of the southwestern United States. Univ. California Publ. Zool., 40:1-70, 2 pls., 8 figs. in text, June 13, 1933.
Burt, W. H.1936. A study of the baculum in the genera Perognathus and Dipodomys. Journ. Mamm, 17:145-156, May 14, 1936.
Coues, E.1875. A critical review of the North American Saccomyidae. Proc. Acad. Nat. Sci. Philadelphia, 1875:272-327, 3 figs. in text, August 31, 1875.
Coues, E., and Allen, J. A.1877. Monographs of North American Rodentia, pp. x + 1081, 77 figs., 1877.
Davis, W. B.1942. The systematic status of four kangaroo rats. Journ. Mamm., 23:328-333, August 13, 1942.
Durrant, S. D., and Setzer, H. W.1945. The distribution and taxonomy of kangaroo rats (genus Dipodomys) of Utah. Bull. Univ. Utah, 35(26):l-39, 2 figs., 4 maps, June 30, 1945.
Gazin, C. L.1942. The late Cenozoic vertebrate faunas from the San Pedro Valley, Arizona. Proc. U. S. Nat. Mus., 92:475-518, 2 pls., 9 figs., 1942.
Gidley, J. W.1922. Preliminary report on fossil vertebrates of the San Pedro Valley, Arizona, with descriptions of new species of Rodentia and Lagomorpha. U. S. Geol. Surv. Prof. Papers, 131-E, pp. 119-131, 2 pls., 1922.
Goldman, E. A.1933. New mammals from Arizona, New Mexico, and Colorado. Journ. Washington Acad. Sci., 23:463-473, October 15, 1933.
Gregory, H. E.1938. The San Juan Country: a geographic and geologic reconnaissance of southeastern Utah. U. S. Geol. Surv. Prof. Papers, 188, pp. v + 123, 26 pls., 4 figs., 1938. [Pg 572]
Grinnell, J.1919. Four new kangaroo rats from west-central California. Proc. Biol. Soc. Washington, 32:203-206, December 31, 1919. 1922. A geographical study of the kangaroo rats of California. Univ. California Publ. Zool., 24(1): 1-124, 7 pls., 24 figs., June, 1922.
Hall, E. R., and Dale, F. H.1939. Geographic races of the kangaroo rat Dipodomys microps. Occas. Papers, Mus. Zool., Louisiana State Univ., 4:47-63, 2 figs., November 10, 1939.
Hall, E. R.1946. Mammals of Nevada, pp. xi + 710, frontispiece, 11 pls., 473 figs., Univ. California Press, Berkeley and Los Angeles, California,. July 1, 1946.
Hibbard, C. W.1937. Additional fauna of Edson Quarry of the Middle Pliocene of Kansas. Amer. Mid. Nat., 18(3):460-464, May, 1937. 1939. Notes on additional fauna of Edson Quarry of the Middle Pliocene of Kansas. Trans. Kansas Acad. Sci., 42:457-462, 6 figs., 1939.
Howell, A. B.1932. The saltatorial rodent Dipodomys: The functional and comparative anatomy of its muscular and osseous systems. Proc. Amer. Acad. Arts and Sci., 67(10):377-536, 28 figs., December, 1932. 1944. Speed in animals, their specialization for running and leaping, pp. xi + 270, 55 figs., University of Chicago Press, Chicago, Illinois, 1944.
Howell, A. B., and Gersh, I.1935. Conservation of water by the rodent Dipodomys. Journ. Mamm., 16:1-9, February 14, 1935.
Marshall, W. H.1940. A survey of the mammals of the islands in Great Salt Lake, Utah. Journ. Mamm., 21:144-159, May 14, 1940.
Mayr, E.1942. Systematics and the origin of species, pp. x + 334, 29 figs., Columbia University Press, New York, New York, 1942.
Midgley, E. E.1938. The visceral anatomy of the kangaroo rat. Journ. Mamm., 19:304-317, 16 figs., August 18, 1938.
Miller, G. S., Jr.1924. List of North American Recent mammals. 1923, U. S. Nat. Mus. Bull., 128:xvi + 673, 1924.
Ridgway, R.1912. Color standards and color nomenclature, pp. iii + 43, 53 pls., published by the author. Washington, D. C., 1912. [Pg 573]
Schultz, J. R.1938. A late Quaternary mammal fauna from the tar seeps of McKittrick, California. Carnegie Inst., Washington, 487:113-215, 17 pls., 12 figs., July 6, 1938.
Warren, E. R.1942. The mammals of Colorado, their habits and distribution, pp. xviii + 330, 50 pls., University of Oklahoma Press, Norman, Oklahoma, 1942.
Wilson, R. W.1939. Rodents and Lagomorphs of the late Tertiary Avawatz Fauna, California. Carnegie Inst., Washington, 514:33-38, 1 pl., May 18, 1939.
Wood, A. E.1935. Evolution and relationship of the heteromyid rodents with new forms from the Tertiary of western North America. Annals Carnegie Museum, 24:73-262, 157 figs., 3 tables, May 13, 1935.
Transmitted May 15, 1948.
22-6114
End of the Project Gutenberg EBook of Speciation in the Kangaroo Rat,
Dipodomys ordii, by Henry W. Setzer
*** END OF THIS PROJECT GUTENBERG EBOOK SPECIATION IN KANGAROO RAT ***
***** This file should be named 42810-h.htm or 42810-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/4/2/8/1/42810/
Produced by Chris Curnow, Matthias Grammel, Joseph Cooper,
The Internet Archive for some images and the Online
Distributed Proofreading Team at http://www.pgdp.net
Updated editions will replace the previous one--the old editions
will be renamed.
Creating the works from public domain print editions means that no
one owns a United States copyright in these works, so the Foundation
(and you!) can copy and distribute it in the United States without
permission and without paying copyright royalties. Special rules,
set forth in the General Terms of Use part of this license, apply to
copying and distributing Project Gutenberg-tm electronic works to
protect the PROJECT GUTENBERG-tm concept and trademark. Project
Gutenberg is a registered trademark, and may not be used if you
charge for the eBooks, unless you receive specific permission. If you
do not charge anything for copies of this eBook, complying with the
rules is very easy. You may use this eBook for nearly any purpose
such as creation of derivative works, reports, performances and
research. They may be modified and printed and given away--you may do
practically ANYTHING with public domain eBooks. Redistribution is
subject to the trademark license, especially commercial
redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License available with this file or online at
www.gutenberg.org/license.
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS', WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTABILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation information page at www.gutenberg.org
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at 809
North 1500 West, Salt Lake City, UT 84116, (801) 596-1887. Email
contact links and up to date contact information can be found at the
Foundation's web site and official page at www.gutenberg.org/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
[email protected]
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit www.gutenberg.org/donate
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations.
To donate, please visit: www.gutenberg.org/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart was the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For forty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Most people start at our Web site which has the main PG search facility:
www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.