The Project Gutenberg EBook of Evolution and Classification of the Pocket Gophers of the Subfamily Geomyinae, by Robert J. Russell This eBook is for the use of anyone anywhere at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org Title: Evolution and Classification of the Pocket Gophers of the Subfamily Geomyinae Author: Robert J. Russell Release Date: July 20, 2012 [EBook #40282] Language: English Character set encoding: ISO-8859-1 *** START OF THIS PROJECT GUTENBERG EBOOK POCKET GOPHERS--SUBFAMILY GEOMYINAE *** Produced by Chris Curnow, Joseph Cooper, Tom Cosmas and the Online Distributed Proofreading Team at http://www.pgdp.net
Throughout, an asterisk before the name (i.e., *Entoptychinae) represents an extinct family/genus/species.
University of Kansas Publications
Museum of Natural History
Vol. 16, No. 6, pp. 473-579, 9 figures in text
![]() August 5, 1968
August 5, 1968
![]()
Evolution and Classification
of the Pocket Gophers of the
Subfamily Geomyinae
BY
ROBERT J. RUSSELL
University of Kansas
Lawrence
1968
University of Kansas Publications, Museum of Natural History
Editors: E. Raymond Hall, Chairman, Henry S. Fitch,
Frank B. Cross, J. Knox Jones, Jr.
Volume 16, No. 6, pp. 473-579, 9 figs.
Published August 5, 1968
University of Kansas
Lawrence, Kansas
PRINTED BY
ROBERT R. (BOB) SANDERS, STATE PRINTER
TOPEKA, KANSAS
1968
![]()
31-4628
Evolution and Classification
of the Pocket Gophers of the
Subfamily Geomyinae
BY
ROBERT J. RUSSELL
When C. Hart Merriam wrote his monograph of the subfamily Geomyinae in 1895, he had no opportunity to examine fossil specimens. No doubt his phylogenetic conclusions and classification would have been greatly influenced had he enjoyed that opportunity because study of fossil geomyids reveals the historic sequence of phyletic development, and this sequence provides a firm basis for distinguishing specialized from primitive characters. The history of the Geomyinae has been characterized by the evolution of specializations. These evolutionary trends begin, as we presently know them, with a generalized ancestral stock in the early Miocene. The direction, degree, and rate of change, beginning with the primitive morphotype of the subfamily, has not been the same in the various lineages. The classification within the subfamily is based upon the phyletic interpretations of available data and the relationships they disclose. In turn, a new, and I hope more realistic, phylogeny and classification is offered.
Recent specimens were studied of all the known genera, subgenera and 29 of the 36 living species. Most of the species not studied are monotypic and have restricted geographic ranges. They are: Geomys colonus, G. fontanelus, and G. cumberlandius, Orthogeomys cuniculus and O. pygacanthus of the subgenus Orthogeomys, and O. dariensis and O. matagalpae of the subgenus Macrogeomys. Examination of these modern species would not radically change the estimation of the degree of phyletic development of the genera and subgenera involved. All of the major polytypic and widespread species were studied.
Specimens of the extinct genera Dikkomys, Pliosaccomys, Pliogeomys, Nerterogeomys, and Parageomys also were studied, as were examples of the extinct species Geomys quinni, Geomys tobinensis, and Orthogeomys onerosus. Considerable fossil material of living species, especially of the genera Geomys and Pappogeomys, was used.
Inasmuch as the present account concerns mainly structural changes in the subfamily Geomyinae at the level of subgenera and above, and the temporal sequence of those changes, no attempt is made in the present account to revise taxonomy below the level of subgenera. Considerable modification of the classification below that level (for species and subspecies) is to be expected in Orthogeomys and Pleistocene taxa of Geomys when available specimens are studied.
I thank Prof. Robert W. Wilson for his assistance in securing fossil geomyids for study, and those in charge of the paleontological collections at the California Institute of Technology, Prof. Bryan Patterson, formerly of the Field Museum of Natural History, and Prof. Claude W. Hibbard of the University of Michigan, Museum of Zoology. For their kindness in lending Recent species, I thank Mr. Hobart M. Van Duesen of the American Museum of Natural History, Dr. David H. Johnson of the U. S. National Museum, and Dr. Oliver P. Pearson of the California Museum of Vertebrate Zoology, the late Colin C. Sanborn of [478] the Field Museum of Natural History, and Profs. Emmet T. Hooper and William H. Burt of the University of Michigan Museum of Zoology.
I am especially grateful to Prof. E. Raymond Hall for his guidance and helpful criticisms with the manuscript. For assistance with paleontological problems, I thank Drs. Robert W. Wilson and William A. Clemens. Several persons have offered helpful suggestions and encouragement in the course of my study. For assistance of various sorts I especially thank Drs. J. Knox Jones, Jr., Rollin H. Baker, A. Byron Leonard, Sydney Anderson, James S. Findley, Robert L. Packard, and Robert G. Anderson. Advice concerning the drawings of the dentitions was generously given by Mr. Victor Hogg, and the drawings were done by Mrs. Lorna Cordonnier under his direction and by Mr. Thomas H. Swearingen. For assistance with secretarial tasks I thank Valerie Stallings, Violet Gourd, Ann Machin, Toni Ward, Sheila Miller, and my wife, Danna Russell.
Morphological features of the fossils and their stratigraphic provenience provide the information upon which phylogenetic interpretations are based. Although the most critical sequences of the fossil record are lacking, and although the existing fossils are mostly fragmentary and therefore seldom furnish ideally suitable data for the interpretations that have been made, phylogenetic conclusions drawn from fossil materials are superior to those drawn on other bases. The especially relevant characters are those disclosing primary trends in the evolution of the modern assemblages. The higher systematic categories recognized in the following account are based primarily upon such characters.
The most important characters found are in the teeth, although several structural changes in the lower jaw, especially those associated with the insertion of cranial musculature, are almost as important.
In primitive geomyines the molar consisted of two columns united at their mid-points and forming a figure 8 or H-pattern (see Fig. 4B). Both labial and lingual re-entrant folds were formed between the two columns. The primitive pattern is retained in the premolars of all known Geomyinae. Therefore, in the earliest (Miocene) members of the subfamily, the pattern of the molars was essentially like that of the premolars.
In Pliocene Geomyinae the two columns of the molars tend to merge into one. This is evident on the worn occlusal surface of the teeth; the lateral re-entrant folds are shallow vertically and progressively recede laterally until only a slight inflection remains. In the final stages of attrition, the inflection disappears and the tooth [479] is a simple elliptical column. In the Pleistocene the monoprismatic pattern appears at earlier stages of wear owing to the decrease in depth of the re-entrant folds, and in Geomyinae of Recent time the initial stages of wear on the enamel cap of infants erase the last vestiges of two columns in the molar teeth.
The general trend in evolution, therefore, has been from a bicolumnar to a monocolumnar pattern. The particular patterns of wear characterizing each genus are described in detail beyond.
The third upper molar has evolved less rapidly than the first and second and in one of the modern lineages (tribe Geomyini) tends to retain at least a vestige of the primitive bicolumnar pattern in the final stage of wear. Therefore, the loss of any trace of the bicolumnar pattern in M3 is considered to be a much specialized condition. Unfortunately, the fossil record of the third upper molar is less complete than that for the first molar and second molar; the tooth drops out of its alveolus more often than does any one of the other molariform teeth and is seldom recovered.
In the primitive genera the enamel pattern is bilophate and the enamel loop (see p. 4B) is continuous on the occlusal surface of a worn molar. Concomitant with the union of the double columns, the bilophodont pattern is reduced to a single loph, but the enamel still completely encircles the dentine.
In the molars of modern geomyines, the enamel loop is not continuous but is interrupted on the sides of the crown by vertical tracts of dentine that are exposed at the occlusal surface of the tooth during early stages of wear. Therefore, a continuous enamel band is to be found only in a juvenal individual whose teeth have been subjected to only slight attrition on the enamel cap. In molars lacking enamel on the labial and lingual sides, anterior and posterior enamel plates, or blades, are found on each molar. The premolar also has an enamel plate on the anterior surface and another on the posterior surface, and in addition both re-entrant angles are protected by a V-shaped investment of enamel. One or the other of the various plates can be reduced or lost accounting for the several distinctive tooth-patterns of the modern geomyines. If loss occurs, it usually is the anterior plate in the lower dentition and the posterior plate in the upper dentition, including the upper premolar. When reduction of the posterior plate of the upper cheek teeth occurs, enamel is first lost from the labial side of the tooth, thus leaving only a short vestigial plate on the lingual end of the crown.
The incisors are smooth with no trace of a groove in the ancestral lineage. In the specialized assemblage (tribe Geomyini) pronounced grooves are always developed on the anterior face of the upper incisor. The pattern of grooving is constant in each species and thus provides characters of taxonomic worth for grouping species into genera. The only inconstancy noted was an incisor of Geomys from the Tobin local fauna of the middle Pleistocene which has three grooves rather than the normal two (No. 6718 KU). The extra groove is an obvious abnormality, and the tooth was associated with others of the same species from the same quarry that were normally grooved.
Grooves on the lower incisors are unknown. The functional significance of grooving has been debated on numerous occasions in the literature. Grooves appear in a number of only distantly related rodents and in lagomorphs. The grooving occurs always in small herbivorous mammals, and in some way may be related to feeding habits.
The grooves provide a serrated cutting edge on the occlusal edge of the upper incisor. In the genus Geomys, for example, the two incisors, including the slight space between them, present a total of five serrations, which may facilitate cutting and piercing tuberous and fibrous roots upon which Geomys feeds. Also the sulci would perform the same function as the longitudinal groove on the side of a bayonet, and would aid the animal in extracting its upper incisors from coarse, fibrous material. In gathering food, the gopher sinks its upper incisors into a root, and then, with the upper incisors firmly anchored, slices off small chunks by means of the lower incisors. Therefore, in pocket gophers, grooving may be an adaptation for feeding on fibrous or woody material. Finally, grooves increase the enamel surface of the incisor without additional broadening of the tooth itself. There could be a selective advantage for sulcation if the extra enamel and the serrate pattern strengthen the incisors, which are under heavy stress while penetrating or prying off pieces of coarse material. Few broken incisors of pocket gophers are found.
This ridge and fossa are on the lateral surface of the ramus. The crest on the ridge begins at the base of the angular process and terminates slightly anterior to the plane of the lower premolar. The masseteric fossa receives the insertion of the rostral or superficial [481] division of the masseter muscle. The mental foramen lies immediately anterior, or anteroventral, to the fossa.
In the ancestral lineage, the ridge is distinct but relatively low; the masseteric fossa is shallow and is a poorly developed area for attachment of the superficial masseter muscle. In modern Geomyinae the ridge is massive and forms a high crest, especially anteriorly, and the masseteric fossa is a deep, prominent cup along the dorsal side of the crest. The elaboration of the crest and fossa evidently is associated with an increase in size of the superficial masseter muscle, which enlarges and provides increased power for the propalinal type of mastication. A high crest has evolved independently in both modern lineages, Thomomyini and Geomyini.
The name basitemporal fossa is suggested here to denote the deep pit that lies between the lingual base of the coronoid process and the third lower molar. The basitemporal fossa receives the insertion of the temporal muscle. The fossa, which until now has not been named, is a unique feature in advanced Geomyinae, being unknown in either primitive Geomyinae or in other rodents.
The temporal is one of several muscles holding the occlusal surface of the lower molariform dentition firmly against the upper cheek teeth during mastication. In primitive geomyines that masticate food by a planing action, the temporal muscle also moves the mandible posteriorly and food is ground between the enamel plates when the lower jaw is retracted as well as when it is moved forward.
The basitemporal fossa appears in late Pliocene geomyines and increases the attachment surface of the temporal muscles that powers the planing action important in utilizing woody and fibrous foods. The basitemporal fossa developed in only one of the modern lineages (tribe Geomyini), the same lineage in which grooved incisors evolved. Both features probably are adaptations for feeding on coarse food. The fossa is not greatly developed in either the ancestral tribe Dikkomyini or the modern tribe Thomomyini, although in some specimens a slight depression marks the site of the basitemporal fossa.
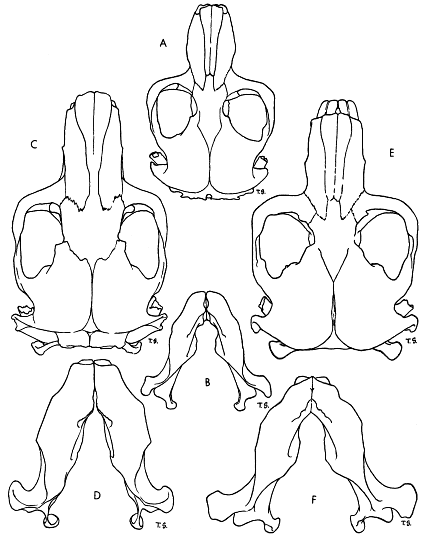
Fig. 1. Types of skulls in the subfamily Geomyinae. × 1.
A. and B. Generalized type of skull. Geomys bursarius lutescens, adult, male, No. 77955 KU, 10 mi. N Springview, Keya Paha Co., Nebraska.
A. Dorsal view of skull.
B. Ventral view of lower jaw.
C. and D. Dolichocephalic type of skull. Orthogeomys (Orthogeomys) grandis guerrerensis, adult, female, No. 39807 KU, 1/2 mi. E La Mira, 300 ft., Michoacán, México.
C. Dorsal view of skull.
D. Ventral view of lower jaw.
E. and F. Platycephalic type of skull. Pappogeomys (Cratogeomys) gymnurus tellus, adult, female, No. 33454 KU, 3 mi. W Tala, 4300 ft., Jalisco, México.
E. Dorsal view of skull.
F. Ventral view of lower jaw.
The skull in most geomyines is generalized, being neither extremely long and narrow nor short, broad and flat as in specialized skulls (see p. 1). In Pleistocene lineages of the modern tribe Geomyini, long skulls and broad skulls evolved and have been termed dolichocephalic and platycephalic specializations, respectively by Merriam (1895:88-101). He correlated them with two diametrically different mechanical methods of mastication.
In animals with dolichocephalic skulls the principal movements of the mandible in the masticatory process are anteroposterior. The resulting propalinal action of enamel plates in opposition to each other characterizes also animals with a generalized skull, and evidently is the method of mastication in the primitive geomyines, but in animals with a dolichocephalic skull the method is developed to a high degree by elongation of the cranium, mandible, and teeth. Both the mandibular and maxillary tooth-rows are relatively longer than in the generalized skull, providing a longer block for the planing action of the lower molariform teeth. All teeth, especially P4 and M3, are longer. In M3 the heel (posterior loph) in particular is elongated. Both the anterior and posterior enamel plates usually are retained in M1 and M2.
The superficial (or rostral) masseter muscle, originates on the side of the rostrum and inserts in the masseteric fossa and on the masseteric ridge. The deep masseter, especially the zygomatic part having its origin along the zygomatic arch, inserts on the angular process of the lower jaw. These two divisions of the masseter muscle have a longer pull (forward) in the dolichocephalic skull than in a non-dolichocephalic skull. The temporal and diagastric muscles retract the lower jaws.
Other, secondary, modifications of the dolichocephalic skull are shortening of the angular process of the mandible, broadening of the rostrum, and narrowing of the cranium and zygomata. Depth of the posterior part of the skull is unchanged. The skull appears to be deep and of nearly equal breadth from nasals to occiput. A good example of a dolichocephalic skull is that of Orthogeomys (see p. 1, C and D).
In the platycephalic skull, the principal masticatory movement of the mandible is anterooblique, to one side and then to the other. The oblique passage of the enamel blades of the lower teeth across those of the upper teeth produces a shearing rather than planing action (Fig. 1E, F). The anterooblique movement of the lower jaw is possible because of major architectural changes in the cranium and mandible. These changes include: (1) Broadening of the postrostral part of the skull, especially the occiput (mastoidal breadth equals or exceeds zygomatic breadth in skulls of some taxa); (2) flattening of the skull; (3) anteroposterior compression [484] of the molariform teeth, especially the molars. Therefore, the entire maxillary tooth-row is relatively shorter than in the dolichocephalic skull. Only a vestige of the heel ordinarily remains on M3. The loss of the posterior enamel blades of P4, M1, and M2 eliminates unnecessary friction, and each of these teeth is wider than long. The distance between the posterior ends of the lower jaws is increased approximately in proportion to the extent that the occiput is widened. As a result of the flattening of the skull the angular processes of the lower jaws are lateral to the zygomatic arches, and approximately on the same vertical level with them. Consequently the insertions of masticatory muscles are shifted laterally. This is especially true of the zygomatic division of the deep masseter, which inserts on the angular process. Contraction of that muscle division of one side of the skull moves the lower jaws obliquely forward. The diagastric and temporal muscles of course retract the lower jaws.
The platycephalic skull is the most specialized skull in the Geomyinae and is a result of the new (for the Geomyinae) method of mastication. The subgenus Cratogeomys (see Fig. 1, E and F) has a platycephalic skull. The trend toward platycephalic specialization has been the major feature of evolution in Cratogeomys.
The fossil record of the subfamily Geomyinae begins in the early Miocene of western North America. No geomyids have been recovered from beds of the late Miocene age. Beginning with the early Pliocene the fossil record becomes progressively more complete, and geomyines are relatively abundant in deposits of late Pliocene and Pleistocene age. Although pocket gophers of the subfamily Geomyinae are rare in lower Miocene deposits, members of the subfamily Entoptychinae are relatively common and highly diversified. Four genera and a number of species have been described (see Wood, 1936:4-25), and the subfamily ranged widely in western North America. I interpret this to mean that the geomyines were indeed uncommon in the early Miocene and their distribution restricted since so few of their remains have been recovered in comparison with entoptychines and the known records are only from the northern part of the Great Plains. On the other hand, entoptychines enjoyed a widespread distribution in western North America (see discussion beyond). Probably the geographic range of the geomyines was largely allopatric to that of the more specialized entoptychines. The zone of fossoral adaptation for herbivorous [485] rodents is ecologically narrow, and as a result competition is severe. As a rule, the outcome of episodes of intergroup competition is geographic exclusion. If these rodents were fossorial in the early Miocene—their morphology suggests they were at least semi-fossorial—mutually exclusive patterns of distribution are to be expected.
Dikkomys is the only genus of the Geomyinae known from the early and middle Miocene. Dikkomys matthewi was described by Wood (1936) on the basis of isolated teeth from lower Harrison deposits (Arikareean in age) near Agate, Sioux County, Nebraska. Later, Galbreath (1948:316-317) described the features of an almost complete mandible recovered from the younger upper Rosebud deposits, now considered by MacDonald (1963:149-150) to be middle Miocene, near Wounded Knee, Shannon County, South Dakota. More recently Black (1961:13) has described a new species, Dikkomys woodi, from the Deep River Formation, Meagher County, Montana. The Deep River Formation is late Hemingfordian (middle Miocene) in age. No remains of Dikkomys have been identified in the extensive rodent fauna of the John Day beds of the lower Miocene of Oregon, although entoptychines are abundant in these deposits.
In the present account, Dikkomys is regarded as the ancestor from which the Pliocene and modern geomyines were derived. These probably did not evolve from the subfamily Entoptychinae because the dentition of entoptychines, especially the premolars and third molars, was already highly specialized by Miocene time.
The numerous records of Thomomys and especially Geomys reported from supposed Miocene or Pliocene deposits are without foundation (see Matthew, 1899:66; 1909:114, 116, 119; 1910:67, 72; 1923a:369; 1924:66; Matthew and Cook, 1909:382; Cook and Cook, 1933:49; and Simpson, 1945:80). Most of the records of Geomys date back to the description of Geomys bisculcatus Marsh (1871:121) from the Loup Fork beds of Nebraska (near Camp Thomas on the Middle Loup River). At first Marsh and other investigators thought these beds were of the late Miocene age. Subsequently the Loup Fork fauna was determined by Matthew (1923b) to be mostly early Pliocene (Clarendonian), but with a later Pleistocene element. Recently, Schultz and Stout (1948:560) have shown that the various Loup River faunas and also those from along the Niobrara River (Hay Springs, Rushville, Gordon local faunas) are of middle Pleistocene age, the fossil-bearing beds occurring just below the Pearlette Ash. These beds are those termed the Loup Fork or North Prong of Middle Loup by the earlier workers who supposed them to be of Miocene or Pliocene age. Both Geomys and Thomomys have been recovered from most of these deposits, but they are no older than middle Pleistocene. This is not surprising in view of the primitive [486] structure of the geomyids known from Miocene and Pliocene beds, but the supposed early appearance of Geomys and Thomomys led to much confusion concerning geomyid evolution in the late Tertiary.
The dearth of geomyines in the Miocene is counterbalanced by the relatively abundant and highly differentiated gophers of the subfamily Entoptychinae. They reached the zenith of their development in this period. Four genera and a number of species are known from the western part of the United States, mostly from beds along the Pacific Coast and in the northern part of the Great Plains. The great diversification of the group in a relatively short period suggests prior movement into a new adaptive zone and subsequent specialization in different subzones and therefore an episode of radial adaptation. The radiation of the entoptychines is discussed elsewhere in the account of geomyid phylogeny, but it should be noted here that both the Geomyinae and the Entoptychinae appear in the fossil record at about the same time in the early Miocene. The principal distinguishing features of each of the two lineages were well developed at the time of their first occurrence, and the entoptychines were the more successful in early Miocene. The Entoptychinae are known only from the early and middle Miocene, unless the earlier deposits of the John Day Formation of Oregon from which mammals have been recovered are considered to be latest Whitneyian (latest Oligocene); for correlations, see Wilson (1949:75). Both lineages likely had an earlier history extending back to their divergence in the Oligocene.
The oldest and most primitive Pliocene geomyine is Pliosaccomys dubius Wilson (1936:20) from the Smith Valley local fauna of middle Pliocene (Hemphillian) age in Nevada. According to Wilson (op. cit.:15) the beds probably were deposited near the middle of Hemphillian time. Shotwell (1956:730) recorded Pliosaccomys dubius from the McKay Reservoir and from the Otis Basin (1963:73) local faunas of the middle Pliocene (Hemphillian) of Oregon, and Green (1956:155) has recovered remains of Pliosaccomys (cf. dubius) from the Wolf Creek local fauna, uppermost part of the lower Pliocene (late Clarendonian in age), of Shannon County, South Dakota. Recently, James (1963:101) has described a second species, Pliosaccomys wilsoni, of this primitive genus. The new species was found in early Pliocene deposits (late Clarendonian) from the Nettle Spring local fauna (Apache Canyon), in the Cuyama Valley, Ventura County, California. Pliosaccomys wilsoni does not differ greatly from P. dubius; however, the few differences [487] in dental characters seem to warrant specific recognition. The reduction of cusps on the metalophid of p4 from three (dubius) to two (wilsoni) and the lack of accessory cuspules on the protolophid of p4 in wilsoni are probably specializations, suggesting that P. dubius even though the more recent in age is the less advanced of the two. P. wilsoni is known only from a lower jaw of a young individual that had dp4 in place, along with m1 and m2. The permanent premolar was in the process of erupting, and the deciduous tooth was removed so that the unworn surface of p4 could be examined.
Pliosaccomys occurred geographically in the area that the Entoptychinae had occupied in the early Miocene. The Smith Valley material includes dentitions in almost all stages of wear and the chronological sequences in the development of the patterns of wear can be reconstructed. An understanding of the dental patterns of the primitive geomyines is based mostly on the interpretation of the stages of wear in Pliosaccomys.
No other pocket gopher is known from the area in which Pliosaccomys occurred, and it is unknown after middle Hemphillian age. Pliosaccomys has closer affinities with Dikkomys of the early Miocene than with any geomyid of the modern assemblage and gives no clue to the origin of the lineage culminating in the modern pocket gophers of the tribe Geomyini.
Pliogeomys buisi Hibbard (1954:353) was found in the Buis Ranch local fauna, of latest middle Pliocene, on the west side of Buckshot Arroyo, Beaver County, Oklahoma. The original material included a right ramus bearing the premolar and first two molars (the holotype) and five isolated premolars and molars. One of the molars is slightly worn and from an immature individual. One premolar is a deciduous tooth. Hibbard (op. cit.:342) identified the beds from which he obtained the Buis Ranch local fauna as from the lowermost part of the Upper Pliocene. Moreover, he judged the Buis Ranch local fauna to be only slightly older than the Saw Rock Canyon local fauna of Seward County in southwestern Kansas. Previously (Hibbard, 1953:408-410), the Saw Rock Canyon local fauna had been assessed as older than the Rexroad local faunas (latest late Pliocene) and, therefore, representative of the early part of the late Pliocene. More recently, Hibbard (1956:164) identified the Buis Ranch beds as part of the Ogallala Formation, which here occurs unconformably just beneath the Rexroad Formation (composed of strata nearly all of late Pliocene age). Therefore, he regarded the Buis Ranch beds as latest middle Pliocene in age. [488] Hibbard (1954:356) suggested that pocket gopher remains from the Saw Rock Canyon local fauna were referable to Pliogeomys buisi, and, in effect, tentatively assigned them to Pliogeomys (in his description of the genus Hibbard remarked that the upper incisor is bisulcate as in Geomys, and the only upper incisor that he mentions was one of the Saw Rock Canyon fossils and not part of the Buis Ranch material). Pliogeomys has closer affinities with modern pocket gophers of the tribe Geomyini than it does with the middle Pliocene genus Pliosaccomys.
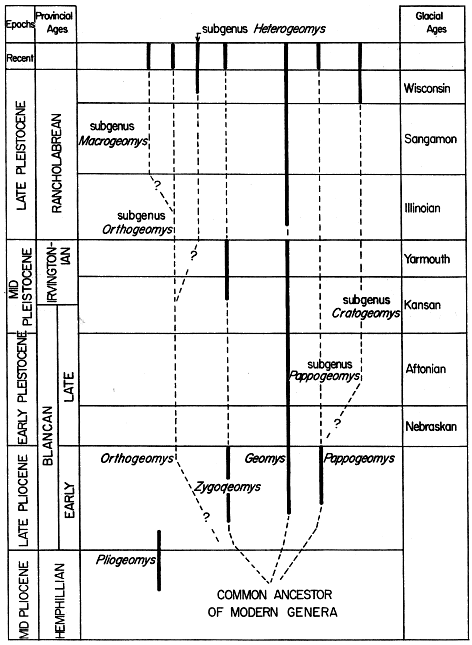
The pocket gopher fauna known from the late Pliocene was more varied than the faunas known from any earlier time. In addition to the extinct Pliogeomys, which occurs in early late Pliocene (see discussion above), the living genera Zygogeomys, Geomys, Pappogeomys (in the sense used on p. 534), and Thomomys first appear in the late Pliocene. The only other living genus, Orthogeomys, makes its first appearance in the late Pleistocene.
The earliest record of the genus Thomomys is based on a fragment of a left mandibular ramus bearing p4 and m1, Thomomys gidleyi Wilson (1933b:122), from the Hagerman local fauna of Twin Falls County, Idaho. Wilson (loc. cit.) was uncertain as to age (late Pliocene or early Pleistocene) but subsequently (1937:38 and 67-70) settled on the middle part of the late Pliocene. Hibbard (1958:11) later considered the age as early Pleistocene (suggesting that the deposits accumulated in the Aftonian interglacial interval) but subsequently (Hibbard et al., 1965:512), on the basis of potassium argon age determinations, also settled on late Pliocene.
Remains of Nerterogeomys [=Zygogeomys] have been found in the Benson local fauna, Cochise County, Arizona, and the Rexroad local fauna of Kansas. This early Blancan gopher first was described as Geomys minor by Gidley (1922:123), and was later referred by Gazin (1942:487) to his new genus Nerterogeomys. Hibbard (1950:138) identified specimens from the Fox Canyon locality, one of the localities of Meade County, Kansas, where the Rexroad local fauna is preserved, as Nerterogeomys, and tentatively referred them to the species N. minor. Nerterogeomys cf. minor has been recovered also from Locality 3 of the Rexroad local fauna (Hibbard, 1950:171) of Meade County, Kansas. Apparently these are also the small gophers about which Franzen (1947:58) wrote. She assigned them to the genus Geomys, and they may actually be a primitive form of Geomys that represents an intermediate stage in the development of the enamel pattern from the uninterrupted loops of the ancestor [489] to the discontinuous pattern of modern Geomys. I favor this interpretation; the evidence, however, is inconclusive, and I have, therefore, reluctantly allocated them, along with the other specimens of Nerterogeomys, to the genus Zygogeomys. In an early paper, Hibbard (1938:244) erroneously referred the same specimens, two upper premolars of a young individual, to the genus Thomomys, and the same material was identified with the genus Geomys, also without specific assignment, in a later paper (Hibbard, 1941b:278). Thomomys is unknown from the late Pliocene of the Great Plains. The specimens previously referred to Nerterogeomys are assigned to the genus Zygogeomys for the first time in this report; for a discussion of the systematic arrangement see the accounts beyond. The type and paratype of Nerterogeomys from the Benson local fauna of Arizona have no indication of enamel reduction.
Specimens of the genus Geomys from the late Pliocene were referred to the large Geomys quinni McGrew, first by Franzen (1947:55) and later by Hibbard and Riggs (1949:835) and Hibbard (1950:171). Geomys quinni has been obtained from the Fox Canyon locality and Locality 3 of the Rexroad local fauna. At Locality 3, both Zygogeomys (cf. minor) and Geomys quinni have been found together, but Geomys quinni can be distinguished by its much larger size and the advanced enamel pattern of the cheek teeth (see systematic accounts beyond). All age classes are represented among the specimens of Geomys quinni; therefore, it seems unlikely that the smaller gophers referred to Zygogeomys are actually the young of Geomys quinni. Hibbard (personal communication, May, 1966) informed me that specimens of Geomys from the late Pliocene (Fox Canyon and Rexroad Locality 3) are erroneously referred to G. quinni. According to Hibbard, this material represents instead two distinct undescribed species, descriptions of which have been submitted by him for publication. Allocation of late Pliocene specimens of Geomys quinni to other species will restrict quinni to the early Pleistocene.
Cratogeomys bensoni Gidley (1922:123) was of medium size. The name was based on an upper incisor bearing a single median sulcus and an associated lower jaw containing all of the cheek teeth from the Benson local fauna, Cochise County, Arizona. Additional lower jaws carrying various teeth also were recovered. The specimens might just as well have been assigned to the genus Pappogeomys since the lower dentitions of all the genera of the tribe Geomyini have the same enamel pattern, and the subgenera Pappogeomys [490] and Cratogeomys have upper incisors with median grooves. The specimens are too fragmentary to warrant more than generic identification. Mainly because of their late Pliocene age and primitive traits the specimens are here regarded as early representatives of the subgenus Pappogeomys. Discovery of the upper molariform dentition would make a more precise assignment possible.
Numerous specimens of geomyids from many localities and horizons are available from the Pleistocene of North America. Specimens of the genera Geomys and Thomomys are especially common. Few specimens are known of the genera Orthogeomys and Pappogeomys, especially from the early and middle Pleistocene, owing, probably, to slight knowledge of the early Pleistocene of México where these two genera are thought to have evolved (see map, Figure 2). This lack of knowledge about early Pleistocene deposits in México is a handicap in the present instance since the center of differentiation for several of the modern genera is judged to have been in México, probably on, and at the edge of, the Central Plateau. The relative abundance of the remains of Geomys and Thomomys from Pleistocene deposits farther north, and the marked absence of other genera, may mean that Orthogeomys and Pappogeomys did not range northward from southern and central México in most of the Pleistocene. One species of Pappogeomys eventually ranged into the southwestern United States in the late Pleistocene (toward the end of the Wisconsin) and it occurs there today, but the genus is essentially Mexican.
The fossil record of Zygogeomys, as the genus is here understood, evidently continued in the United States will into the Middle Pleistocene, depending upon the stratigraphic interpretation of the age of the Curtis Ranch local fauna from southeastern Arizona. Hibbard (1958:25) regarded the Curtis Ranch local fauna as Irvingtonian in age, a local fauna that lived either in the late Kansan glacial or the Yarmouthian interglacial, and his correlation is tentatively followed here. In deposits laid down later than those of Irvingtonian age no remains of Zygogeomys have been found. Today a single species exists as a relic in the mountains of central México and Zygogeomys may have retreated southward to its present refugium in the late Pleistocene. Perhaps, Zygogeomys occurred in northern México and the southwestern United States in the early and middle Pleistocene (see p. 2), occupying the area between the ranges of Pappogeomys to the south and Geomys to the [491] north. Competition with Pappogeomys, and especially Geomys, during Irvingtonian time may have extirpated Zygogeomys over most of this area, and by late Pleistocene (Sangamon) much of the former range of Zygogeomys came to be occupied by one or the other of its competitors. The occurrence of Geomys garbanii in southern California (see White and Downs, 1961) and the unidentified species of Geomys in Aguascalientes (Mooser, 1959; for [492] faunal correlation, see Hibbard and Mooser, 1963), both from deposits of Irvingtonian age, supports this suggestion.
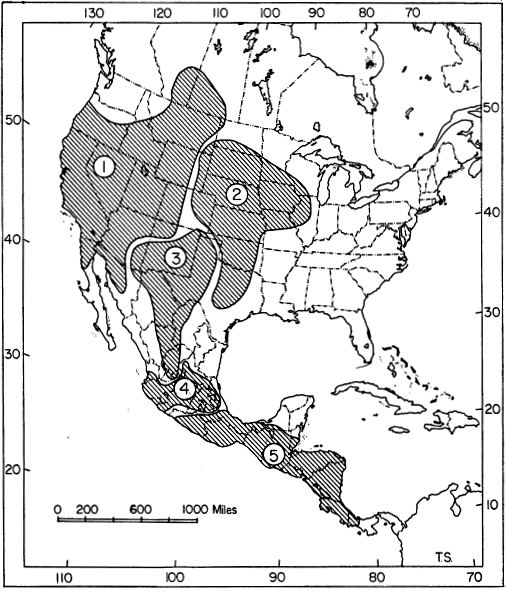
Fig. 2. Probable distribution of the Subfamily Geomyinae in the early Pleistocene (late Blancan), depicting major areas of differentiation of the modern genera.
1. Thomomys
2. Geomys
3. Zygogeomys
4. Pappogeomys
5. Orthogeomys
The earliest Pleistocene records of Thomomys are mostly isolated teeth. Although they can be identified as genus Thomomys, most of the materials are too fragmentary to be identified to species. In Thomomys two distinct patterns of occlusal surfaces of the molars can be recognized: the generalized elliptical pattern in the subgenus Pleisothomomys, not unlike the pattern in other geomyids, and the pear-shaped pattern in the subgenus Thomomys, which results from constriction of the upper molars on the labial side and constriction of the lower molars on the lingual side. Some fossils assigned to Thomomys were not examined with this distinction in mind by the persons who made the assignments. Consequently some of the identifications now in the literature may be subject to change.
Three occurrences of Thomomys are from the early and middle Pleistocene, with a possible fourth (depending upon the age of the Hay Springs local fauna of Nebraska). The earliest Pleistocene record is from the Broadwater-Lisco beds along the North Platte River in Morrill County, western Nebraska. Possibly the specimen from there was misidentified. Those beds are Lower Pleistocene, and are regarded by Schultz and Stout (1948:560-561, 573) and by Hibbard (1958:11), as having been deposited mostly during the Aftonian interglacial. There is also some indication that some of the strata were deposited late in the Nebraskan glaciation. There are no other early Pleistocene records of Thomomys. Savage (1951:228) reported the genus from the Irvington local fauna, Alameda County, California. The specimens were not identified to species, although they were described as indistinguishable from Thomomys bottae. Paulson (1961:137) recorded specimens from the Cudahy local fauna, Meade County, Kansas. These fragmentary specimens are referable to the subgenus Thomomys, owing to the strong constriction of the molars, but have not been identified to species. The Cudahy is an Irvingtonian local fauna, and is considered to have been deposited during the late Kansan glaciation. The stratum containing the Cudahy local fauna immediately underlies the Pearlette Ash. The Cudahy material includes five isolated molars and a fragmentary ramus bearing only the premolar. The genus Thomomys has been recovered also from the Hay Springs local fauna in Sheridan County, northwestern Nebraska, by Shultz and Tanner (1957:71). The Hay Springs local fauna is considered to [493] have been deposited in late Kansan glaciation or in early Yarmouth interglacial by Shultz and Tanner (op. cit.:69), or of Irvingtonian age; however, Hibbard (1958:25) regarded the beds containing this fauna as Illinoian (thus post-Irvingtonian in age), and equivalent in age to the Berends local fauna of Oklahoma and the Butler Springs and Mt. Scott local faunas of Kansas. The Thomomys from Hay Springs local fauna has not been referred to species.
The relative abundance of Geomys, and rarity of Thomomys, in Great Plains fossil beds of early and middle Pleistocene is probably due to allopatric distributions of the two genera. The Great Plains area was evidently the center of distribution and differentiation of Geomys. Perhaps Thomomys evolved earlier to the west, in the Great Basin and Pacific Coastal regions, and not on the Great Plains.
Upper Pleistocene records of Thomomys are more common. The genus was widespread in beds identified with the Illinoian and Sangamon and extended its range eastward to the Atlantic Coast. Stephens (1960:1961) reported Thomomys from the Doby Springs local fauna, Harper County, northwestern Oklahoma. The material (34 isolated teeth) was too fragmentary to permit assignment to species. The molars are constricted on one side, indicative of the subgenus Thomomys, like the Cudahy specimens reported by Paulson (see discussion above). Stephens erroneously mentioned that the enamel plate on the posterior face of the upper premolar is unique in Thomomys; this plate occurs also in Zygogeomys. The Doby Springs local fauna was recovered from beds that have been identified as Illinoian deposits, and it is correlated with the Berends local fauna in Beaver County, Oklahoma, and the Butler Springs local fauna in Meade County, Kansas (see Stephens, op. cit.: 1700).
Local faunas in Maryland and Florida of Rancholabrean age include Thomomys, in every instance referable to the subgenus Pleisothomomys on the basis of unconstricted molars. Thomomys potomacensis (Gidley and Gazin, 1933), from Cumberland Cave local fauna, Allegany County in western Maryland, is the type of the genus Pleisothomomys Gidley and Gazin (1933:354). Pleisothomomys is here regarded as a subgenus. The material used in the original description included four lower jaws, one with a complete dentition. Hibbard (1958:25) pointed out that the Cumberland Cave assemblage is a composite fauna including both glacial and interglacial forms. He placed the stratigraphic position of the fauna as definitely Upper Pleistocene, probably deposited in both Illinoian glaciation and during the Sangamon interglacial. T. potomacensis is significantly larger than T. orientalis Simpson (1928:6), [494] from the Saber-tooth Cave local fauna, Citrus County, Florida. Simpson's material included a rostral fragment with an incisor, premolar, and first molar. The Saber-tooth Cave local fauna is regarded by Kurten (1965:219) as having been recovered from Sangamon deposits. Thomomys is unknown from Wisconsin deposits in the eastern United States, and today the genus does not occur east of the Great Plains.
Thomomys of Rancholabrean provincial age from the western United States and México is known only from Wisconsin beds.
Three extinct species of Thomomys, all referable to the subgenus Thomomys, have been described. Thomomys microdon Sinclair (1905:146), based on the rostral portion of a skull without a mandible, is from the Potter Creek Cave local fauna, Shasta County, California, and has been recovered also from Samwel Cave, Shasta County, California. T. microdon closely resembles Thomomys monticola that lives in the area today. Thomomys scudderi Hay (1921:614) is from the Fossil Lake (or Christmas Lake) local fauna in central Oregon. Elftman (1931:10-11) referred these specimens to Thomomys townsendii, and he considered T. scudderi to be a synonym of T. townsendii. Davis (1937:156-158) disagreed with Elftman concerning the taxonomic status of T. scudderi, which he regarded as a valid species. According to Davis, T. scudderi is more closely allied to Thomomys bottae than to T. townsendii. Cope (1878:389; 1889:160-165) had referred the same specimens to Thomomys clusius (now Thomomys talpoides clusius). Cope considered the beds to be Pliocene in age. In all accounts of the Fossil Lake local fauna up to Hay (1921), the specimens of Thomomys were referred to the species clusius, talpoides, or bulbivorus (see Elftman, loc. cit.). The Fossil Lake local fauna is currently considered as being of Rancholabrean provincial age, probably dating from the Wisconsin glacial maximum when the lake reached its greatest size. The third extinct species described from the Wisconsin is Thomomys vetus Davis (1937:156), also from the Fossil Lake local fauna in Lake County, Oregon. Davis pointed out that T. vetus differs from T. scudderi Hay, of the same fauna, in larger size and other cranial details, and that it is closely allied to the living species Thomomys townsendii, and not to Thomomys talpoides, which is the only species of Thomomys living in the area today.
Thomomys townsendii was recovered by Gazin (1935:299) from the American Falls beds (probably Wisconsin deposits) in Idaho.
Thomomys talpoides is reported from the Howard Ranch local [495] fauna in Hardeman County, western Texas, by Dalquest (1965:69-70), who referred the isolated teeth to T. talpoides on geographic grounds, apparently on the erroneous assumption that T. talpoides was the species of Thomomys nearest geographically to Hardeman County. Hay (1927:259) reported Thomomys fuscus [= Thomomys talpoides] from late Pleistocene beds near Wenatchee, Chelan County, Washington. Hibbard (1951:229) recorded Thomomys talpoides from late Pleistocene deposits in Greeley County, Kansas, and Walters (1957:540) reported the same species from late Pleistocene deposits in Clark County, Kansas. According to Hibbard (1958:14) other remains reported as T. talpoides have been recovered from numerous areas of Wisconsin glacial drift in western North America.
Thomomys bottae has been identified from Wisconsin age deposits in western North America, as follows: Burnet Cave, Gaudalupe Mt., New Mexico (Schultz and Howard, 1935:280); Carpinteria Asphalt, California (Wilson, 1933a:70); McKittrick Asphalt, Kern County, California (J. R. Schultz, 1938:206); Rancho La Brea, Los Angeles County, California (Dice, 1925:125—specimens described as a new subspecies, T. b. occipitalis); Papago Springs Cave, Santa Cruz County, Arizona (Skinner, 1942:150 and 158—probably bottae, but possibly umbrinus on the assumption that the two are specifically instead of subspecifically distinct); Isleta Cave, Bernalillo County, New Mexico (Harris and Findley, 1964:115—some of these fossils may be post-Wisconsin in age); Potter Creek Cave and Samwel Cave, Shasta County, California (Sinclair, 1905:146—identified as T. leucodon, now a subspecies of T. bottae; also see Hay, 1927:214-215).
Thomomys umbrinus has been reported from San Josecito Cave, Nuevo León, México (Russell, 1960:542); Upper Bercerra, México (Hibbard, 1955a:51—identified only as Thomomys sp., but undoubtedly referable to T. umbrinus). Post-Wisconsin remains of Thomomys umbrinus are reported by Alvarez (1964:6) from capa II and capa III of the Cueva La Nopalera, southwestern Hidalgo. Hay (1927:222-223) reported specimens of the genus Thomomys from Wisconsin deposits in Hawver Cave, Eldorado County, California, but did not assign them to species. Gilmore (1947:158) found the remains of Thomomys umbrinus in cave deposits near Quatro Ciénegas in central Coahuila. These cave deposits may have been laid down during the Wisconsin, but more likely accumulated in the post-Wisconsin.
Remains found in the Curtis Ranch local fauna, Cochise County, in southeastern Arizona are regarded as of middle Pleistocene age. See Gazin (1942:481-484), Wilson (1937:39-40), Hibbard (1958:25), and Hibbard et al. (1965:510-511). Although some question as to the exact age of the Curtis Ranch local fauna still seems to exist, most authorities on the Pleistocene agree that the age is not Pliocene and that it is older than Rancholabrean. Gidley (1922:122) described the pocket gopher found in the Curtis Ranch beds as Geomys parvidens, which is preoccupied by Geomys parvidens Brown (1908:194), a name proposed for the pocket gopher from the Conard Fissure of Arkansas; therefore, Hay (1927:136) proposed the name Geomys persimilis for the Curtis Ranch species to replace Geomys parvidens Gidley. Geomys persimilis Hay became the type species of Gazin's genus Nerterogeomys (1942:507). In this paper, Nerterogeomys is considered to be a junior synonym of Zygogeomys.
Zygogeomys persimilis is represented by a rostral fragment bearing all the cheek teeth on the left side and the upper incisors. In addition, two lower jaws, one with the first three cheek teeth, are referred to the species (see Gazin, 1942:507). The fossils identified as Geomys from the Arroyo San Francisco, Cedazo fauna, in Aguascalientes, México, by Mooser (1959:413) may be referable instead to Zygogeomys. I have not seen the specimens and no figures are available; Mooser states that a cranium was recovered. If either the upper premolar or third molar is in place, generic identification could be made with reasonable certainty. No other fossils of Zygogeomys have been uncovered in late Pleistocene deposits and the significance of the absence of Zygogeomys has been discussed in an earlier paragraph of this section. Geomys has not been found so far south as Aguascalientes, but Zygogeomys occurs farther south now and presumably had a more extensive range on the plateau to the north in the Pleistocene.
Geomys is common in Pleistocene deposits, especially on the Great Plains. Certainly the center of differentiation for Geomys was in this region, although at times, probably when conditions were favorable, Geomys expanded its range into adjacent areas, reaching the Pacific Coast in Irvingtonian times and the Atlantic Coast at the time of the Illinoian glaciation. The earliest Pleistocene records of the genus are from the Great Plains. McGrew (1944:49) [497] described Geomys quinni from the Sand Draw local fauna, Brown County, Nebraska, considered by Hibbard (1958:11) to be Nebraskan in age. As mentioned in the account of Pliocene geomyids, Geomys quinni occurs also in the late Pliocene deposits of southwestern Kansas. Also, Geomys quinni occurs in the Broadwater-Lisco local fauna of Morrill and Garden counties, western Nebraska (Barbour and Schultz, 1937:3; Schultz and Stout, 1948:560-563; Schultz et al., 1951: table 1). The Broadwater-Lisco is currently regarded as Aftonian deposits (Schultz and Stout, loc. cit.; Hibbard, 1958:11). Hibbard (1956:174) identified Geomys quinni from the Deer Park local fauna, probably deposited during the early Aftonian interglacial, of Meade County, Kansas. Strain (1966:36) described Geomys paenebursarius on the basis of fossils obtained from early Pleistocene deposits of the Hudspeth local fauna from western Hudspeth County in the Trans-Pecos of Texas. The Hudspeth fossils were probably deposited during the Aftonian interglacial. From Kingman County, Kansas, Hibbard (op. cit.: 164) recovered isolated teeth of Geomys from the Dixon local fauna, regarded by him (op. cit.:153-154) as deposited during the latest Nebraskan glaciation, and correlated by him with the Sand Draw local fauna of Nebraska. Hibbard (1958:11) later regarded the Dixon as a transitional fauna between Nebraskan and Aftonian. The remains of Geomys from the Dixon are known only from isolated teeth. The teeth are small, and suggest that a smaller species of Geomys may have occurred along with the more common and larger G. quinni during the early Pleistocene (see discussion beyond of the Saunders Geomys). Geomys quinni was widespread and common throughout the central Great Plains from the late Pliocene (Rexroad fauna) through the early Pleistocene (Nebraskan and Aftonian deposits).
Hibbard (1956:179) referred the pocket gopher remains taken from the Saunders local fauna in Meade County, Kansas, to Geomys tobinensis, a small species having continuous enamel bands around the lower premolar in younger specimens. The Saunders local fauna was deposited in the late Aftonian and is younger than the Deer Park local fauna discussed above. Paulson (1961:138) later pointed out that the Saunders Geomys is distinct from Geomys tobinensis; hence, the small pocket gopher from the Saunders local fauna is probably an unnamed species, perhaps more closely allied to paenebursarius than to quinni. The small Geomys reported from the Aftonian Broadwater-Lisco local fauna of Nebraska (Schultz and Stout, 1948:563) may also be the same as the Saunders pocket [498] gopher, but the smaller adult specimens occurring in the same bed with larger specimens probably are females and the larger specimens males. In all living Geomyini females have smaller skulls than males.
The Irvingtonian provincial age is currently regarded as Middle Pleistocene and includes the late Kansan glaciation (that part occurring after the glacial maximum) and the Yarmouthian interglacial (see Hibbard et al., 1965:512-514). The Irvingtonian provincial age, therefore, follows the late Blancan provincial age of the early Pleistocene and is succeeded by the Rancholabrean provincial age of the late Pleistocene. No specimen of an Irvingtonian Geomys is referable to any living species. Two Irvingtonian species have been described. Hibbard (1944:735) named Parageomys tobinensis [= Geomys tobinensis] from the Tobin local fauna of Russell County, Kansas. This species since has been reported from the Cudahy local fauna of Meade County, Kansas (Paulson, 1961:137). Hibbard (1956:183) also identified as Geomys tobinensis the pocket gopher recovered from the Saunders local fauna, a late Aftonian deposit of Meade County, Kansas, and reduced the technical name Parageomys from generic to subgeneric rank. Paulson (op. cit.:138) pointed out that the Saunders specimens differ from G. tobinensis, and he, therefore, restricted the name to the small Geomys of the Cudahy and Tobin local faunas of Irvingtonian provincial age. G. tobinensis is markedly smaller than the Blancan G. quinni. The Cudahy and Tobin local faunas are of approximately the same age, and presently both are included in one unit, the Cudahy fauna. The Cudahy fauna is considered to have been deposited in late Kansan as it occurs in strata immediately below the Pearlette ash.
Recently, White and Downs (1961:8) described a new Irvingtonian species, Geomys garbanii, from the middle Pleistocene Vallecito Creek local fauna of San Diego County, California. Many well preserved fossils of the new species were recovered. Geomys garbanii is of medium size (approximately the size of one of the larger subspecies of G. bursarius), and significantly larger than the Irvingtonian Geomys tobinensis of the Great Plains. The Vallecito Creek occurrence of Geomys is the first authenticated record from the Pacific Coast region. Matthew (1902:320) erroneously referred remains of Thomomys to the genus Geomys in his revised list of Cope's earlier report on the Fossil Lake (or Silver Lake) fauna (see discussion of Thomomys above).
A number of Irvingtonian fossil remains of Geomys have not been identified with particular species. Hibbard (1941a:206) found Geomys in the Borchers local fauna (deposited in the time of the Yarmouthian interglacial) of Meade County, Kansas. Also, Geomys has been reported from several sites in Nebraska. Schultz and Tanner (1957:67) reported Geomys from the Angus fossil quarry in Nuckolls County, south-central Nebraska. The Angus fossils were found in sediments of the Sappa Formation considered by Schultz and Tanner to be a Yarmouthian deposit. Fossil quarries (Hay Springs, Rushville, and Gordon) along the south side of the Niobrara River Valley in Sheridan County, Nebraska, have also provided records of geomyids. Both a large and small species of Geomys have been reported from the more recently excavated Rushville and Gordon sites (Schultz and Stout, 1948:562-567, and table 3). In view of the great disparity in size owing to sex, these may actually be males and females of the same species, as mentioned above. The name Hay Springs has been used in reference to all three sites. The ages of the Hay Springs sites are approximately the same, but their correlation is presently under debate. Schultz and Tanner (1957:68-71) maintain that the fossils are distinctly middle Pleistocene, and that they were deposited during late Kansan glaciation, or perhaps from early Yarmouthian into early Illinoian, with the largest concentration coming from the Sappa sands of pre-Illinoian (Yarmouth) age. Hibbard (1958:25), basing his opinion on the presence of Microtus pennsylvanicus, and the stage of evolution of other species in the assemblage, regards the Hay Springs sites as probably Illinoian deposits, but certainly no older than that.
Mooser (1959:413) identified as Geomys the pocket gopher from Irvingtonian deposits in Arroyo San Francisco (loc. no. 5) near the city of Aguascalientes, México. As suggested elsewhere in this account, these fossils may be referable to Zygogeomys rather than Geomys. The Irvingtonian provincial age of this fauna was established by Hibbard and Mooser (1963:245-250). Other alleged occurrences have recently been compiled by Alvarez (1965:19-20). Maldonado-Koerdell (1948:20) noted four fossil occurrences of the genus Geomys in México. Two of these from San Josecito Cave in Nuevo León have since been identified with the genera Orthogeomys and Pappogeomys (Russell, 1960:543-548); the third listed by Maldonado-Koerdell from "near Ameca, Jalisco," was based on Brown's (1912:167) mention of some bones supposedly of the family "Geomyidae," and the fourth refers to pocket gopher remains [500] from the "Hochtals von Mexiko" listed as Geomys by Freudenberg (1921:139). His generic identification is doubtful and the specimens should be compared with Mexican genera of the Geomyinae.
Upper Pleistocene records of Geomys also are common. Upper Pleistocene is here understood to include late Illinoian, Sangamon and Wisconsin deposits; all are considered to be of Rancholabrean provincial age (see Hibbard et al., 1965:512-515) and post-Irvingtonian. The presence of remains of Bison and/or Microtus pennsylvanicus are currently considered mammalian index fossils of Rancholabrean faunas. In the Illinoian, Geomys extended its range to the Atlantic Coast in the southeastern United States. The eastern and western species-groups evidently were isolated throughout much of the late Pleistocene, and, therefore, evolved separately. Of the two, the eastern, or pinetis, species-group seems to have remained somewhat more generalized, and the western, or bursarius, species-group has become more specialized. The Rancholabrean Geomys from deposits in the southeastern United States are referable (see Ray, 1963:325) to Geomys pinetis.
Marsh (1871:121) described Geomys bisulcatus from the North Prong of the Loup River (near Camp Thomas), Nebraska. These beds are also termed the Loup Fork or Loup River fossil beds (see discussion on p. 485), and they lie along the upper reaches of the Middle Loup River in Thomas County (near Senea), Hooker County (near Mullen), and southeastern Cherry County (probably the North Prong beds northwest of Mullen). These beds were at first thought to be of Miocene age, but later were regarded as early Pliocene (see Schultz and Stout, 1948:562-566 for a historical account of expeditions to these fossil sites). Schultz and Tanner (1957:71-72) pointed out that the principal fossiliferous beds in the Middle Loup region are of middle to late Pleistocene age, with most of the fossils coming from the Crete sand and silt beds which are probably early Illinoian deposits, and, therefore, younger than the Hay Springs faunas. Some fossils may have come from the Sappa deposits dated by Schultz and Tanner (loc. cit.) as mostly Yarmouthian deposits. Geomys bisulcatus, judging from the original description and Hibbard's discussion of the cotypes (1954:357), does not differ significantly from Geomys bursarius. However, Geomys bisulcatus is tentatively retained as a valid species. Based on the evidence cited above it seems unlikely that Geomys bisulcatus occurred in pre-Irvingtonian times as often suggested in the literature.
The genus Geomys has been identified in several faunas of [501] Illinoian age, all from the Great Plains. Stephens (1960:1961) reported the genus from the Doby Springs local fauna in Harper County, Oklahoma, and Starrett (1956:1188) reported it from the Berends local fauna in Beaver County, Oklahoma. Schultz (1965:249) assigned 21 isolated teeth, including six incisors, from Butler Springs local fauna (considered by him to be late Illinoian, following the glacial maximum) to Geomys cf. bursarius. Hibbard and Taylor (1960:167) reported a baculum tentatively identified as that of Geomys from the early Illinoian Butler Springs local fauna (including the Adams fauna) of Meade County, Kansas. Hibbard (1963:206) recorded the genus Geomys from the Mt. Scott local fauna (late Illinoian deposits) of Meade County, Kansas; the specimens probably are referable to the living species bursarius. From McPherson County, Kansas, Hibbard (1952:7) reported the genus Geomys from the Kentuck Assemblage, which he (1958:25) regarded as a composite of Illinoian and Sangamon species. Specific identification of the Illinoian pocket gophers is uncertain, primarily due to the fragmentary nature of the material. On the basis of dental characters alone most specimens could be referred to G. bursarius; however the taxonomic status of G. bisulcatus is in doubt, and more complete material may indicate that the Illinoian gophers are specifically distinct from the living species. Consequently, most authors, including myself, have made no attempt to refer these specimens to species. Nevertheless, the Illinoian Geomys from the Great Plains is more closely allied to the living species of Geomys than it is to the earlier Irvingtonian species.
Geomys bursarius has been collected from a number of Sangamon fossil sites on the Great Plains. Although specific identification of specimens of Geomys from Illinoian faunas is uncertain, the Great Plains Geomys from Sangamon and later deposits probably is referable to the living species as Hibbard and Taylor (1960:165) pointed out. They found no difference between Geomys recovered from the Cragin Quarry local fauna (early Sangamon) of Meade County, Kansas, and the living species Geomys bursarius. Isolated teeth of the same species were collected from the Jinglebob local fauna of Meade County, Kansas (Hibbard, 1955b:206), a fauna of the late Sangamon. Hibbard (1943:240) also recorded the genus Geomys (referable to G. bursarius) from the Rezabek local fauna of Lincoln County, Kansas. According to Schultz et al. (1951:6 and table 1) the genus Geomys occurs in buried or "fossil" soils of Sangamon age, lying just above the Loveland Loess, in Nebraska. No specific localities were given by them, nor were any particular [502] specimens mentioned. Dalquest reported Geomys bursarius from two Sangamon faunas in northern Texas. The species is represented in the Ward Quarry local fauna of Cooke County, Texas (1962a:42), and the Good Creek local fauna of Foard County, Texas (1962b:575).
Geomys bursarius has been reported from Wisconsin fossil deposits of the Great Plains and adjacent areas as follows: Jones local fauna, Meade County, Kansas (Hibbard and Taylor, 1960:64-66); Two Creeks Forest beds of the third interstadial soils formed between Cary and Mankato glaciations, late Wisconsin (Schultz et al., 1951:8 and table 1); Cita Canyon local fauna in the northern part of the Panhandle of Texas (Johnson and Savage, 1955:39); Howard Ranch local fauna of Hardeman County in northwestern Texas (Dalquest, 1965:70); Quitaque local fauna of Motley County, Texas (Dalquest, 1964:501); Clear Creek local fauna of Denton County in north-central Texas (Slaughter and Ritchie, 1963:120); Ben Franklin local fauna, of late Wisconsin beds along the North Sulphur River in Delta County, NE Texas (Slaughter and Hoover, 1963:137); Bulverde Cave (Hay, 1920:140; 1924:247) and Friesenhahn Cave (Tamsitt, 1957:321), both in Bexar County, south-central Texas; Alton, Illinois (Hay, 1923:338-339); Wisconsin drift of Illinois, without mention of specific locality (Bader and Techter, 1959:172); Wisconsin drift of southwestern Wisconsin and northeastern Iowa (Hay, op. cit.:343); Wisconsin drift near Galena, Illinois, and mouth of Platte River in eastern Nebraska (Leidy, 1869:406).
Brown (1908:194) described Geomys parvidens from the Conard Fissure, in northern Arkansas. Hibbard (1958:25) concluded that the Conard Fissure fauna represents a glacial stage, probably the Illinoian, and Hibbard et al. (1965:510-511) regarded the fauna as a composite including both Irvingtonian and Rancholabrean elements. White and Downs (1961:21) considered G. parvidens to be a subspecies of Geomys bursarius.
The first Pleistocene occurrence of Geomys in the southeastern United States is from the Reddick I deposits reported by Gut and Ray (1963:325), who found the remains of Geomys pinetis among the fossils comprising the "rodent beds" of Marion County, Florida. Gut and Ray tentatively identified the beds as Illinoian, but Kurten (1965:219) regarded the Reddick I fauna as early Sangamon. Simpson (1928:2) reported Geomys floridanus [= pinetis] from Saber-tooth Cave deposits of Citrus County, Florida. The Saber-tooth Cave (or Lecanto Cave) local fauna is considered by Kurten (op. cit.:219) also to be a Sangamon deposit. Geomys floridanus [503] [= pinetis] was reported from the Seminole Field deposits by Simpson (1929:563); both Simpson and Kurten (op. cit.:221) agreed that the Seminole Field fauna is mainly late Wisconsin, although sub-Recent fossils occur at the tops of the beds. Ray (1958:430) collected remains of Geomys pinetis from the Melbourne Bone Bed of Brevard County, Florida. The Melbourne local fauna is considered to be from Wisconsin deposits by Kurten (op. cit.:220). The eastern species of Geomys were probably derived from Great Plains stock that reached the southeastern Coastal Plains in early Rancholabrean (Illinoian) time. Presently there is no contact between the eastern and western populations of the genus, and it is assumed that disjunction occurred as a result of Wisconsin glaciation. It is interesting to note that the genus Thomomys occurred in this region at approximately the same time; both genera occur in Saber-tooth Cave deposits.
The genus Pappogeomys is not known from Pleistocene deposits older than the Wisconsin glaciation, but a pre-Pleistocene occurrence in the Benson beds of Arizona (see discussion of the Pliocene above) shows that Pappogeomys had been differentiated by late Pliocene time. The absence of Pappogeomys, beginning in the early Pleistocene and continuing well into the late Pleistocene, is attributed to the southern distribution of the genus, where its range probably was centered on the Central Plateau of México. The paucity of early and middle Pleistocene deposits from this critical region prevents any definite statements about phyletic development within the genus. All of the late Pleistocene records pertain to the subgenus Cratogeomys (long in use as a generic name but in the present paper reduced to subgeneric rank in the genus Pappogeomys). Schultz and Howard (1935:280) found Cratogeomys [= Pappogeomys] castanops in Burnett Cave in the Guadalupe Mountains of south-central New Mexico. The Burnett deposits are probably late Wisconsin (see Schultz and Tanner, 1957:75, for discussion of the age of these deposits based on carbon-14 tests). These writers (loc. cit.) also referred the mandible of a small pocket gopher to the genus Pappogeomys [= subgenus Pappogeomys]. However, neither genera nor subgenera of the tribe Geomyini can be distinguished on the basis of their inferior dentitions. Judging from the distribution of the modern geomyines, it seems unlikely that the subgenus Pappogeomys has occurred beyond its present range in the late Pleistocene; therefore the small mandible is most likely that of a young individual of Pappogeomys castanops. Russell [504] (1960:543) referred specimens collected at San Josecito Cave in Nuevo León, México, to the group of small subspecies Cratogeomys [= Pappogeomys] castanops. Also, Russell (loc. cit.) identified a rostral fragment as of the genus Cratogeomys [= subgenus Cratogeomys] although the fragment had a combination of features different than in any named species of the genus; he did not name the fragment as a new species, preferring to wait for additional material that could clarify its taxonomic relationships.
Hibbard (1955a:52-53) identified Cratogeomys [= Pappogeomys] tylorhinus from the Becerra Superior deposits in the valley of Tequixquic in the northern part of the state of México. The Wisconsin age of these beds suggests an earlier Pleistocene derivation of the gymnurus-group of species.
Several specimens of the subgenus Cratogeomys have been reported from beds of latest Wisconsin (certainly after the glacial maximum) or post-Wisconsin age. Gilmore (1947:158) found fossil remains of Cratogeomys [= Pappogeomys] castanops commonly in Quaternary cave deposits on the mountain slopes in the vicinity of Cuatro Ciénegas, in central Coahuila. These deposits actually may be of post-Wisconsin origin (see discussion above). Alvarez (1964:8) obtained fragments of Cratogeomys [= Pappogeomys] tylorhinus from sub-Recent deposits of Capa III in the Cueva La Nopalera in southwestern Hidalgo, México. Pappogeomys merriami lives in the area today. Mayer-Oakes (1959:373) reported remains of Cratogeomys [= Pappogeomys] merriami from levels eight and eleven of the excavations at El Risco II, in the northern part of Mexico City. The ages of these deposits are unknown to me, but they probably are no older than late Wisconsin with most of the beds dating from the post-Wisconsin.
This genus is not known from the Pleistocene, except for its occurrence in the San Josecito cave deposits of southwestern Nuevo León, México (Russell, 1960:544). Although Orthogeomys does not occur in the immediate vicinity of the cave at the present time, the northern limits of its range is nearby in southern Tamaulipas. The Orthogeomys from San Josecito Cave differs from living species, and has been named Heterogeomys [= Orthogeomys] onerosus Russell (loc. cit.), and is evidently referable to the subgenus Heterogeomys. As mentioned before, the San Josecito Cave local fauna represents deposits of Wisconsin glaciation.
The account of the Tucan or Indian mole by Hernandez (sometimes listed as Fernandez) in 1651 probably is the earliest published one of a geomyid (see Merriam, 1895:201; Coues, 1877:607-608). Linnaeus in 1758 did not mention geomyids. In 1772, Kerr described Hernandez's Tucan under the name Sorex mexicana on the basis of Hernandez's account without having seen any specimens. Lichtenstein in 1827 applied the technical name Ascomys mexicana to three specimens collected by Deppe from unknown localities on the tableland of México. Merriam (loc. cit.) pointed out that the name mexicanus of Lichtenstein in 1827 is a nomen nudum, and that it is preoccupied by mexicanus used by Kerr in 1792. The latter can not be technically identified with any particular species of geomyid.
Bartram in 1791 wrote of the pocket gopher of Florida, without formally describing it. The first available technical name is Mus bursarius of Shaw in 1800. Rafinesque in 1817 proposed the first generic names for the geomyids when he described Geomys and Diplostoma. In 1839, Waterhouse referred the genus Geomys to his family Arvicolidae, considered by him to be a subgroup of muroids. In 1841, he suggested that Geomys was related to Bathyergus and Spalax. Waterhouse in 1848 (p. 8) treated the pocket gophers as a subgroup of rodents under the group name Saccomyina, in which he included the genera Heteromys, Saccomys, Perognathus, and Dipodomys. Hence, Waterhouse was the first to recognize the relationship between the heteromyids and geomyids. In the next year Gervais erected the family Pseudostomidae for a group of specialized squirrels to include Geomys and Thomomys and the same genera (at least in part) of heteromyids that Waterhouse classified in the "family" Saccomyina.
In 1839 the name Thomomys was proposed by Maximilian (Wied-Neuwied). All of the generic names previously proposed for pocket gophers were considered by subsequent authors to be synonyms of Geomys.
A third family name, Sciurospalacoides, was proposed by Brandt (1855:188) who referred Geomys and Thomomys to that family. He placed his new family phylogenetically between the family Sciuridae and the family Spalacoides (a group in which Brandt included the genera Spalax, Sipheus, and Ellobius). Brandt took exception to the classification of Waterhouse (1848), who united the geomyids and heteromyids in one family. Brandt placed the [506] heteromyid genera in other groups: Perognathus in the Muridae, and Macrocolus [= Dipodomys] in the Macrolini, a subfamily of the family Dipodoides.
Modern classification of the pocket gophers begins with Baird in 1858. The important classifications are summarized in Table 1; a few that do not depart essentially from those listed have been omitted owing to limited space for the tabular arrangement, but are discussed in the following account.
Baird probably was strongly influenced by the arrangement proposed by Waterhouse in 1848, but was opposed to separating geomyids from heteromyids as was done by Brandt. Baird was convinced of the close relationship of the geomyids and heteromyids, and referred both groups to one family, the Saccomyidae, as Waterhouse had done earlier. In order to recognize the morphological specializations he used two subfamilies, Geomyinae and the Saccomyinae. In the 20 years that followed, some authors followed Brandt and others followed Baird.
Gill, in 1872 (p. 71), proposed a classification essentially like Baird's of 1858, but Gill raised Baird's subfamilies to the rank of family (see Table 1). In referring all pocket gophers to the Geomyidae, Gill used that name as a family term for the first time. Also he established the superfamily Saccomyoidea to include his two families, Geomyidae and Saccomyidae; therefore, the Saccomyoidea was equivalent to the group Saccomyina of Waterhouse (1848) and the Saccomyidae of Baird (1858). Coues (1877), in his classic monograph of the Geomyidae followed the arrangement proposed by Gill in treating the pocket gophers as a family. Alston in 1876 proposed another classification based on Baird (1858), with two subfamilies, the Geomyinae and the Heteromyinae, united together in the family Geomyidae; thus, he recognized that the genus Saccomys Frédéric Cuvier, 1823, was a synonym of Heteromys Desmarest, 1817, as had been pointed out by Gray (1868:201) and Peters (1874:356). Coues (1877:487-490) acknowledged the invalidity of the genus Saccomys, but refused to give up the name in supergeneric classification. Winge, first in 1887 and subsequently in 1924, classified the geomyids and heteromyids together in the family Saccomyidae as did Baird in 1858, and like Coues, Winge too ignored the synonymy of Saccomys with Heteromys and insisted on retaining the technical terms Saccomyidae and Saccomyini.
Up to the time of Merriam's classic revision of the Recent Geomyidae in 1895 all the known species of living pocket gophers were referred to two genera, Geomys and Thomomys. Merriam described [507] much new material, especially from México and Central America, and proposed seven new genera (see Table 1). His complete and detailed study of the dentitions and osteology of the skull remains today as the definitive work on this subject, and is the point where most studies of the Geomyidae must begin. His treatment of the Recent genera survived for 52 years without change until Hooper (1946:397) arranged Platygeomys as a synonym of Cratogeomys. However, Merriam's genera have been recognized in all subsequent classifications except for the current review (see Table 1).
Cope described the first known fossil geomyids in 1878, and published an excellent review of the two genera, Pleurolicus and Entoptycus, in 1884 (pp. 855-870, pl. 64, figs. 1-9). Both genera were recovered from the John Day Miocene deposits of Oregon. Cope did not propose a new systematic arrangement of these geomyids, but referred them to the family Saccomyidae and mentioned that the Saccomyidae was equivalent to the family Geomyidae of Alston. Winge, in 1887, followed Cope in referring Pleurolicus and Entoptycus to the Saccomyidae along with the living genera Thomomys and Geomys. Miller and Gidley (1918), in their synopsis of the supergeneric groups of rodents, proposed a new subfamily, Entoptychinae, to include the divergent Miocene pocket gophers. Miller and Gidley also revived the old subfamily Geomyinae of Baird (1858), but restricted its application to the modern pocket gophers and their immediate ancestors. In 1936, A. E. Wood revised the taxa of the subfamily Entoptychinae, and described the first Miocene genus, Dikkomys, of the Geomyinae. He followed the supergeneric classification of Miller and Gidley (1918).
The recent classifications of Simpson (1945) and Wood (1955) have combined the classifications of Merriam (1895) and Wood (1936). Wood (1955) brought up to date the list of genera, including those that were described after the publication of Simpson's classification (1945). In Table 1, the list of genera is principally from Simpson (1945) but generic names used by Wood (1955) are included. This is the currently accepted classification.
The new classification proposed in this paper (see Table 1) includes three tribes proposed as vertical units; they are intended to stress the phyletic trends in the known evolutionary sequences by placing immediate ancestors together with their descendants.
Pliogeomys is placed in the same tribe (Geomyini) as Zygogeomys, Geomys, Orthogeomys, and Pappogeomys. That tribe includes the most specialized Geomyinae. Zygogeomys, Geomys, Orthogeomys, and Pappogeomys are lineages resulting from a Pleistocene radiation in which all the lineages diverged from a common Pliocene ancestor. The radiation of the Geomyini was well under way by the close of the late Pliocene. Although Pliogeomys may not be the actual ancestor, it closely resembles the primitive morphotype.
Table 1.—History of the classification of the Superfamily Geomyoidea
| Baird 1858 | Gill 1872 Coues 1877 |
Winge 1887 and 1924 | Merriam 1895 Ellerman 1940 |
Wood 1935 Wood 1936 |
Simpson 1945 Wood 1955 |
Names used in present paper | |
| Family Saccomyidae |
Family Geomyidae |
Family Saccomyidae |
Family Geomyidae |
Family Geomyidae |
Family Geomyidae |
Family Geomyidae |
|
| Subfamily Geomyinae |
"Group" Geomyini |
Subfamily Geomyinae |
Subfamily Geomyinae |
Subfamily Geomyinae |
|||
| Tribe Dikkomyini |
|||||||
| *Dikkomys | *Dikkomys | *Dikkomys | |||||
| *Pliosaccomys | *Pliosaccomys | ||||||
| Tribe Thomomyini |
|||||||
| *Pleisothomomys | *Pleisothomomys |
|
Thomomys | ||||
| Thomomys | Thomomys | Thomomys | Thomomys | Thomomys | Thomomys | ||
| Tribe Geomyini |
|||||||
| *Pliogeomys | *Pliogeomys | ||||||
| Zygogeomys | Zygogeomys | Zygogeomys |
|
Zygogeomys | |||
| *Nerterogeomys | |||||||
| Geomys | Geomys | Geomys | Geomys | Geomys | Geomys |
|
Geomys |
| *Parageomys | |||||||
| [509] | Orthogeomys | Orthogeomys | Orthogeomys |
|
Orthogeomys | ||
| Heterogeomys | Heterogeomys | Heterogeomys | |||||
| Macrogeomys | Macrogeomys | Macrogeomys | |||||
| Pappogeomys | Pappogeomys | Pappogeomys |
|
Pappogeomys | |||
| Cratogeomys | Cratogeomys | Cratogeomys | |||||
| Platygeomys | Platygeomys | Platygeomys | |||||
| Subfamily Entoptychinae |
Subfamily Entoptychinae |
Subfamily Entoptychinae |
|||||
| *Pleurolicus | *Pleurolicus | *Pleurolicus | *Pleurolicus | ||||
| *Gregorymys | *Gregorymys | *Gregorymys | |||||
| *Grangerimus | *Grangerimus | *Grangerimus | |||||
| *Entoptychus | *Entoptychus | *Entoptychus | *Entoptychus | ||||
| Geomyidae incertae sedis |
Geomyidae incertae sedis |
||||||
| "Group" Gymnoptychine** |
*Gidleumys | *Diplolophus | *Diplolophus | ||||
| Gymnoptychus | *Griphomys | *Griphomys | |||||
| Subfamily Saccomyinae |
Family Saccomyidae |
"Group" Saccomyini |
Family Heteromyidae |
Family Heteromyidae |
Family Heteromyidae |
||
* Denotes extinct genera.
** Winge included in his family Saccomyidae the "group" Gymnoptychine and the contained genus Gymnoptychus Cope, 1873, which genus currently is placed in the family Eomyidae. The type of Gymnoptychus Cope, 1873, is synonymous with Ischyromys Leidy, 1856, and the valid name for the genus is Adjidaumo Hay, 1899.
Pliosaccomys, on the other hand, represents the terminal stages of a long trend that began with the Dikkomys-like Geomyinae of the early Miocene. In this lineage, the rate of evolution in the dentition and the skull was slow; therefore, the differences between early Miocene (Dikkomys) and middle Pliocene (Pliosaccomys) are not great and the two are united into the tribe Dikkomyini. The Dikkomyini is the ancestral geomyinen trunk from which the modern groups have diverged.
The Pliocene ancestor of Thomomys is unknown but probably resembled Pliosaccomys, with which it may have been a contemporary. Thomomys is the least specialized of the modern Geomyinae, and, consequently, shows the most resemblance to the ancestral tribe. The specializations of Thomomys, however, clearly preclude its reference to the tribe Dikkomyini; therefore, it is set apart in the monotypic tribe Thomomyini. That tribe has not undergone an adaptive radiation comparable to that of the tribe Geomyini or that of the Entoptychinae in the early Miocene. Here, for the first time, Thomomys is set apart in classification from the other living pocket gophers.
Merriam's genera Orthogeomys, Heterogeomys, and Macrogeomys are closely related. Each of these taxa is retained as a subgenus of a single genus, Orthogeomys. Some species of Macrogeomys seem to be more closely allied to the subgenus Orthogeomys and others to the subgenus Heterogeomys. A revision of the genus is needed; it might show that the currently recognized subgenera are artificial, and that a different arrangement of the species would more clearly express their evolutionary relationships. The subgenus Heterogeomys seems to be the most nearly uniform of the subgenera, and it is the least specialized. Radiation within the genus may have begun relatively recently, but the many special adaptations for tropical environments suggest that the genus has been in the Neotropical Zone a long time. Therefore, discovery of an early dichotomy from the common ancestral stock of the tribe would come as no surprise.
Nerterogeomys Gazin here is arranged as a junior synonym of Zygogeomys. Both are less specialized than any of the other Geomyini, except Pliogeomys. The single living species (Zygogeomys tricopus) is obviously a relic. Its range is small. The two subspecies [511] differ only in minor features. The living species does have a few unique characteristics, only to be expected in the surviving species of a long phyletic lineage. Some of these are specializations. Otherwise, Zygogeomys and Nerterogeomys are closely related and the latter is best placed as a synonym of the former. Both are admittedly closely related to Geomys. Zygogeomys and Geomys share several characters, particularly primitive ones; there is considerable parallelism, especially marked in Irvingtonian species of Geomys. Nevertheless, Geomys is more specialized, particularly in the dentition, and it has developed some Pappogeomys-like specializations. Zygogeomys has retained more of the primitive characters of the tribe. A strong case could be made for recognizing only one genus, Geomys, containing Zygogeomys as one of two subgenera. Nevertheless, the characters separating Zygogeomys and Geomys are of considerable importance and I consider the two kinds to be distinct genera.
The species of Geomys, both living and extinct, form a distinct and well-marked group. The genus is less primitive in most respects than Zygogeomys and Orthogeomys and it is less specialized than Pappogeomys, excluding the ancestral stock (subgenus Pappogeomys). Some specimens of species of Irvingtonian age (Geomys tobinensis and Geomys garbanii, especially the former) retain primitive enamel plates as does Zygogeomys; but this is true of only a small percentage of the individuals. Also the adult dental pattern developed somewhat later in ontogeny in these middle Pleistocene species of Geomys than in either Recent or late Pliocene and early Pleistocene representatives (Geomys paenebursarius, Geomys quinni) of the genus. Whether these features represent a stage in the evolution of the late Pleistocene and Recent species or a terminal stage in members of a sterile and primitive branch of the main line of evolution of Geomys is uncertain. At present I favor the latter explanation, and view G. paenebursarius and G. quinni as early progressive species that evolved dental specializations that were maintained in the main line of phylogeny.
Hibbard proposed the generic name Parageomys (1944:55), but later regarded it as a subgenus of Geomys (1956:182) that includes those species retaining continuous enamel bands until relatively late in ontogeny; no other differences have been noted. When the early phylogeny of Geomys is better understood, Parageomys may serve as a subgeneric taxon in which the primitive species of Geomys can be grouped, but as of now Parageomys is arranged as a synonym of Geomys.
Pappogeomys and Cratogeomys also form a natural group. Their close relationship is best reflected in formal taxonomy by including them in the same genus. Their dissimilarities are of the sort that separate a primitive ancestral lineage from a divergent and progressively more specialized assemblage. The fossil record is inadequate, and I can only speculate that Cratogeomys diverged from primitive Pappogeomys-stock in the earlier Pleistocene, at least before the end of the Irvingtonian. Cratogeomys probably originated on the Mexican Plateau and probably underwent its subsequent evolution there. The living species of the subgenus Pappogeomys are evidently relics of the ancestral stock of the genus. Hooper (1946:397), I think correctly, considered Platygeomys as congeneric with Cratogeomys, although the highest degree of specialization of the genus is attained in those species formerly classed in the genus Platygeomys. Even so, in my opinion, the differences are insufficient to warrant even subgeneric recognition.
Rodents of the superfamily Geomyoidea specialized for completely fossorial life (early Pliocene to Recent); specialized earlier (late? Oligocene and early Miocene) for semi-fossorial habits; body thickset, fusiform without apparent neck (in modern geomyids); legs short; forelegs especially stout; eyes and ears small (pinna reduced to inconspicuous crest concealed beneath pelage); tail tactile, shorter than head and body; lips closing behind incisors; cheek pouches external, fur-lined; baculum rodlike, arched, having expanded quadriform platelike base; pelage long, soft without underfur, covering body in thick coat (in some species of Orthogeomys scant, harsh or scattered bristles); color varying from pale tints of buffy (almost white) to metallic black.
Skull thick-walled, massive, angular, relatively broad, and flattened; distinctly murine form, but having zygomasseteric structure of advanced sciuromorphs, including small infraorbital canal (that transmits no part of masseter muscle) and well-developed, broad zygomatic plate; zygomata massive and widely flaring, jugals stout; rostrum robust, relatively broad and deep, and without evidence of transverse canal (as in Heteromyidae); anterior projection of nasals only slightly exceeding that of upper incisors; interorbital region usually constricted, narrower than rostrum; anterior opening of infraorbital canal far forward on side of rostrum, about half way between zygomatic plate and upper incisor and just behind premaxillary-maxillary suture, its opening countersunk in oblique sulcus (for protection from muscle contraction); postorbital process lacking, except for rudimentary knoblike projection in subgenus Macrogeomys; palate relatively narrow, its deeply sculptured surface sloping steeply downward posteriorly causing region supporting maxillary tooth-row to be markedly depressed; palatine bone reduced, forming, on two abruptly different levels, posterior margin of hard palate behind tooth-rows; parietals compressed and narrow, and most of cerebral cavity roofed by squamosals (in some species squamosals overlap lateral parts of parietals); tympanic bullae completely inferior in position and fully ossified, external meatus being developed laterally as elongated tube; mastoid not inflated, but broadly exposed at posterolateral margin of the skull; occiput large, its surface usually rugose, and paroccipital processes large and flangelike, at least in advanced groups (early Pliocene to Recent); ramus relatively short and stout, having distinct [513] crest and ridges for muscle attachments; coronoid process well developed, erect; articular condyle prominent; angular process prominent, reflected laterally, and in modern groups lateral extension protruding from posterior border of ramus nearly at right angle; capsule for root of lower incisor, prominent between angular process and articular condyle.
Anterior surface of incisors broad and flat, always smooth on lower teeth, but either smooth or grooved on upper teeth depending on taxon; cheek teeth hypsodont, becoming progressively higher crowned in modern groups, rooted in primitive groups (late? Oligocene to middle Pliocene), rootless and ever-growing in modern groups (late Pliocene to Recent); upper and lower premolars persistently bicolumnar; upper and lower molars bicolumnar only in primitive groups (late? Oligocene and early Miocene), becoming progressively monocolumnar in advanced groups (early Pliocene to Recent), primitive bicolumnar pattern being retained on occlusal surface only in early stages of ontogeny and in third molar throughout life; enamel pattern of occlusal surface of cheek teeth based on sextituberculate prototype (see Wood and Wilson, 1936:388-391), having cusps arranged in two transverse rows of three cusps each, excepting three anterior cusps of premolars that are arranged in trefoil, especially on p4 (sometimes only one or two, rather than three, cusps develop in a particular set, especially in p4), conules absent; protostyle and endostyle in upper teeth and protostylid and hypostylid in lower teeth formed from cingulum; cusps of each row uniting with wear into transverse enamel lophs (or lophids), each tooth having two lophs, one on anterior column, protoloph and protolophid, and one on posterior column, hypoloph and hypolophid, that unite with additional wear forming continuous enamel band; enamel lacking on sides of each column in advanced lineages, thereby restricting enamel to anterior and posterior walls; with extreme reduction, posterior plates of upper teeth and, more commonly, anterior plates of lower molars, missing. Dental formula: 1/1, 0/0, 1/1, 3/3.
Key to the Subfamilies of Geomyidae
Subfamily Entoptychinae Miller and Gidley, 1918
Anterior face of upper incisor usually smooth, sometimes bearing faint groove in center or near medial margin of tooth, at least in Gregorymys; cheek teeth hypsodont, medium to high crowned, and rooted in all but Entoptychus (has rootless, ever-growing teeth); cheek teeth identical in form, premolars resembling molars and lower cheek teeth mirror images of upper teeth; crowns biprismatic, having two columns joined at edge of protomeres (for description of term, see discussion of primitive morphotype on p. 537) and with persistent lateral fissure between them; lateral re-entrant fold deep, penetrating at least half width of crown, from external side in upper teeth and internal side in lower teeth (in specialized genus Entoptychus lophs, upon additional wear, join also at edge of parameres, thus uniting columns at both ends and thereby enclosing interior part of lateral fissure as a transverse fossette in center of tooth); enamel investment of prisms usually complete, including inflection bordering re-entrant folds, occlusal pattern becoming interrupted with wear only in Entoptychus, where enamel disappears first from sides of crowns (following [514] union of anterior and posterior columns at both sides) and later, in final stages of attrition, from anterior wall of lower molars and posterior wall of upper molars.
Maxillary bone without pronounced vertical depth in part supporting cheek teeth, its inferior border only slightly lower than inferior border of premaxillary and alveolar lips of molariform teeth consequently approximately level with, or slightly below, alveolar lip of upper incisor; squamosal without lateral expansion, therefore, meatal tube of auditory bulla separated from zygomatic process of squamosal by deep, well-developed postglenoid notch; angular part of mandible below alveolar level of mandibular cheek teeth; angular process only slightly reflected laterally; coronoid process low, tip only slightly above condyle.
For information concerning the structure and relationships of the known genera, and for accounts of species, see Wood (1936). A list of the named genera in order of specialization is as follows:
*Pleurolicus Cope, 1878. Proc. Amer. Phil. Soc., 18:66.
*Gregorymys Wood, 1936. Amer. Mus. Novit., 866:9.
*Grangerimus Wood, 1936. Amer. Mus. Novit., 866:13.
*Entoptychus Cope, 1878. Proc. Amer. Phil. Soc., 18:64.
Five new species have been described since Wood's (1936) revision. They are: Pleurolicus clasoni MacDonald (1963:180); Gregorymys kayi Wood (1950:335); Gregorymys montanensis Hibbard and Keenmon (1950:198); Grangerimus dakotensis MacDonald (1963:182); Grangerimus sellardsi Hibbard and Wilson (1950:623).
Subfamily Geomyinae Baird, 1858
Anterior face of upper incisor primitively smooth, grooves consistently developed only in one modern lineage (Geomyini); cheek teeth hypsodont, primitively rooted and having crown of medium height (late Oligocene to middle Pliocene), being higher crowned, rootless and ever-growing in modern lineages (late Pliocene to Recent); primitively crowns of cheek teeth biprismatic, having two columns joined at mid-points by narrow isthmus and entire crown sheathed in continuous band of enamel; premolars retaining primitive biprismatic form, anterior and posterior columns never uniting at edge of protomeres or parameres, and with both lateral re-entrant folds persistent throughout life; primitive biprismatic pattern becoming decidedly modified in molars (except in M3), having two prisms progressively uniting into one column by reduction and loss of lateral inflections, primitive biprismatic patterns being retained only in early stages of ontogeny; third upper molars retaining, at least partially, primitive bicolumnar pattern (except in Thomomyini), with relatively broad isthmus and horizontally shallow re-entrant folds, lingual fold sometimes wanting; enamel pattern becoming discontinuous (late Pliocene to Recent) owing to loss of enamel from sides of each column; remaining enamel restricted to anterior and posterior plates, or cutting blades, and enamel bordering lateral inflections in premolars (considering both sides together, these plates constitute essentially two transverse cutting blades); enamel pattern of M3 varying, depending on taxon; with specialization, anterior plates of lower molars and posterior plates of upper premolar and molars may be reduced or lost; except in primitive species (early Miocene), no enamel fossettes retained in adult dentitions.
Maxillary bone having pronounced vertical depth in part supporting cheek teeth, inferior border arching downward well below inferior border of premaxillary; consequently, alveolar lips of molariform teeth decidedly below level of alveolar lip of upper incisor; squamosal with marked lateral expansion at expense of postglenoid notch; notch compressed and reduced between meatal tube of auditory bulla and zygomatic process of squamosal; angular part of mandible mostly above alveolar level of mandibular cheek teeth; angular process reflected laterally at right angles to axis of ramus and developed into heavy knoblike projection; coronoid process well developed, tip decidedly higher than [515] condyle; fossorial specializations remarkably well developed in advanced lineages, degree of specialization of primitive Miocene species unknown but probably only semi-fossorial as in Entoptychinae.
Key to the Tribes of the Geomyinae
Genotype.—Dikkomys Wood, 1936.
Chronologic and geographic range.—Early to Middle Pliocene (early Arikareean to mid-Hemphillian) in western United States. Known from Miocene fossil sites in Montana, South Dakota, and Nebraska and Pliocene sites in South Dakota, Oregon, Nevada, and southern California. For precise localities see accounts of Dikkomys and Pliosaccomys beyond.
Diagnosis.—Small Geomyinae; lacking specializations of more advanced tribes; upper incisors smooth, at least in Pliosaccomys; molariform teeth always rooted and having crowns of medium height; enamel investment of cheek teeth complete and uninterrupted in all stages of wear; crowns of molars primitively biprismatic, having two columns united at mid-points, thus forming narrow isthmus separating lateral re-entrant folds as in premolars, and, with wear, also uniting secondarily at protomeres (with exception of third lower molars), consequently, isolating remnant of that inflection as shallow fossette (columns uniting first at protomeres in Pliosaccomys); anterior and posterior columns of first and second molars, both above and below, becoming progressively united into one column in advanced Dikkomyini (early and middle Pliocene), but m3 (M3 unknown) retaining primitive biprismatic pattern, with columns joined at centers but never at protomeres (for details of dentition see generic accounts); [516] mandible stout, its angle mostly above mandibular tooth-row; masseteric ridge low; basitemporal fossa barely discernable in some fragments of Pliosaccomys; postcranial skeleton unknown.
Key to the Genera of the Tribe Dikkomyini
1936. Dikkomys Wood, Amer. Mus. Novit., 866:26, July 2.
Type.—Dikkomys matthewi Wood, 1936, from Lower Harrison deposits near Agate, Sioux County, Nebraska.
Chronologic range.—Early Miocene, from early Arikareean (Lower Harrison local fauna of Nebraska) to middle Miocene, late Hemingfordian (Upper Rosebud local fauna, South Dakota, and the Deep River Formation, Montana). According to MacDonald (1963:149-150), the Upper Rosebud is middle Miocene rather than early Miocene.
Description.—Size small, about as in small kinds of Thomomys; known only from fragmentary mandible, including molariform dentition in place, and isolated cheek teeth, including M1 (see Wood, 1936:26-28 and fig. 32; Galbreath, 1948:316-317 and fig. 1; and Black, 1961:13-14 and fig. 58); upper incisors unknown; cheek teeth hyposodont, persistently rooted, and having crowns of medium height compared with Recent geomyids; enamel investment complete and uninterrupted in all molariform teeth in all stages of wear; P4 unknown, but probably formed like p4; p4 persistently biprismatic, two crowns joined at mid-points by relatively narrow isthmus separating lateral re-entrant folds; anterior lophid of p4 having distinct anteroexternal inflection; molars also biprismatic throughout life; two lophids of lower molars first uniting at mid-points as in p4, and, with additional wear, m1 and m2 secondarily uniting at edge of protomeres and forming isolated enamel fossette between point of connection (detailed description of stages of wear discussed in account of phylogeny of subfamily); m3 permanently joined at mid-point only, without lateral union at edge of protomeres; upper molars, judging by M1 (M2 and M3 unknown), having same pattern as lower molars, but first union of lophs decidedly on lingual side of center, consequently, lingual re-entrant fold small; M1 probably developing U-pattern in advanced stages of wear by union of protomeres, with minute lingual fossette developing in transition as lophs secondarily become united at lingual edge of columns; mandible stout and geomyidlike; masseteric ridge weakly developed; basitemporal fossa absent.
Evidently, Dikkomys matthewi is more primitive than Dikkomys woodi. The modified H-pattern in m1 and m2, with the metalophid and hypolophid joined at both their mid-points and also at their protomeres (by union of the protostylid and hypostylid in the lower dentition), is persistent throughout life. Therefore, the enclosed enamel fossette is not eradicated with wear. In m1 and m2 of Dikkomys woodi, the fossette is shallower, and, at least in advanced stages of wear, it would disappear, therefore, forming a U-pattern on the occlusal surface, as in M1 and M2, but lateral inflection horizontally shallow rather than deep as in entoptychines.
Specimen (No. P 26284 FMNH) reported as Dikkomys matthewi by Galbreath (1948:316) is referable to the recently described species Dikkomys woodi Black, 1961.
Specimens examined.—One, no. P 26284, Field Mus. Nat. Hist., from upper Rosebud, Shannon Co., South Dakota.
Referred species.—two:
Dikkomys matthewi Wood, 1936. Amer. Mus. Novit., 866:26, July. Type from early Arikareean Lower Harrison deposits (early Miocene) near Agate, Sioux County, Nebraska.
Dikkomys woodi Black, 1961. Postilla, Yale Peabody Museum, 48:13, January 16. Type from Deep River Formation, late Hemingfordian (middle Miocene), Meagher County, Montana; also known from Upper Rosebud deposits (middle Miocene) near Wounded Knee, Shannon County, South Dakota.
1936. Pliosaccomys Wilson, Carnegie Inst. Washington Publ., 473:20, May 21.
Type.—Pliosaccomys dubius Wilson, 1936, from Smiths Valley local fauna in Lyon County, Nevada.
Chronologic range.—Early Pliocene, late Clarendonian (Wolf Creek local fauna, South Dakota, and Nettle Springs local fauna, California) to Middle Pliocene, middle part of Hemphillian (Smiths Valley local fauna, Nevada, and McKay Reservoir and Otis Basin local faunas, Oregon).
Description.—Size small (alveolar length of mandibular tooth-row measuring 6.0 in holotype), about as in Thomomys monticola; upper incisor relatively broad and flat, having anterior face smooth, without trace of grooving; crowns of cheek teeth of medium height and rooted; enamel investment continuous and uninterrupted in all stages of wear; premolars permanently, biprismatic; P4 having anterior prism subtriangular and decidedly smaller that sub-crescentic posterior prism, and joined near centers by narrow, obliquely oriented isthmus; p4 having anterior prism subovate, posterior prism strongly compressed anteroposteriorly, and joined at mid-points by relatively broad and straight isthmus; first and second molars, both above and below, monoprismatic in final (?adult) stage of wear, derived ontogenetically from primitive bilophate pattern by coalescence of two columns into one; M1 and M2 mirror images of m1 and m2 in pre-final stages of wear, two columns first uniting at edge of protomeres forming U-pattern, and primitive H-pattern never developing in either series (for detailed description of stages of wear, see account of phylogeny, p. 546); m3 (M3 unknown, but probably with same form as in Geomyini, see p. 552) persistently biprismatic, two columns joined by relatively broad isthmus at centers, consequently, forming H-pattern of primitive ancestors; rostrum heavy and broad as in modern geomyids; palate narrow and strongly ribbed; mandible stout; masseteric ridge and fossa well developed; basitemporal fossa absent.
Specimens examined.—Six, nos. 1796 (holotype)—1799, 1804 and 1806 (CIT) now in the Los Angeles County Museum, all from Smiths Valley local fauna, Middle Pliocene, Nevada.
Referred species.—two:
*Pliosaccomys dubius Wilson, 1936. Carnegie Inst. Washington Publ., 743:20, May 21. Known from early and middle Pliocene faunas including Wolf Creek local fauna (late Clarendonian), Shannon County, South Dakota; McKay Reservoir local fauna and Otis Basin local fauna (Hemphillian), Oregon; type from Smiths Valley local fauna (probably middle Hemphillian), Lyon County, Nevada.
*Pliosaccomys wilsoni James, 1963. Univ. California Publ. Geol. Sci., 45:101, June 26. Type from Nettle Springs local fauna of late Clarendonian (early Pliocene), Ventura County, California.
Type.—Thomomys Wied-Neuwied, 1839.
Chronologic and geographic range.—Known from late Pliocene (early Blancan) to Recent. Known primarily from western North America from southern Canada south to Central México in Pliocene, Pleistocene and Recent and in middle and late Pleistocene of Maryland and Florida.
Diagnosis.—Size small to medium (basilar length exclusive of T. bulbivorus, measuring from approximately 24 to 45, including both males and females); upper incisors without grooving, excepting fine, indistinct sulcus rarely near inner margin (grooving more common in T. monticola than in other Recent species); crowns of cheek teeth high, rooted and ever-growing; all molars, including M3, monoprismatic and anteroposteriorly compressed, sometimes (especially in subadults) having slight inflection on labial side in upper teeth and lingual side in lower teeth; molars bicolumnar in pre-final stages of wear (seen in juvenal teeth only), patterns of wear in both upper and lower molars resembling those of Pliosaccomys, except that crowns of m3 and M3 unite into single column in final stages of wear; enamel pattern interrupted in all cheek teeth, loss occurring only at sides of each column; transverse enamel blade completely covering posterior face of both P4 and p4; all upper and lower molars with two transverse enamel blades, one on anterior surface and one on posterior surface, of each tooth, including M3; small third plate sometimes persistent on broad side of tooth, labial side in upper molars and lingual side in lower molars (T. bulbivorus); skull generalized, neither unusually narrow and deep or broad and flat; usually without marked cresting or rugosity; masseteric ridge well developed and massive; basitemporal fossa absent, sometimes shallow depression forming in T. townsendii; pelage soft, never harsh or hispid, covering body with thick coat of hair; forefoot exceptionally small for fossorial mammal, claws not especially long; body form remarkably fossorial.
The tribe Thomomyini is monotypic, including only the genus Thomomys.
1839. Thomomys Wied-Neuwied, Nova Acta Phys. Med. Acad. Caesar. Leop.-Carol., 19(1):377.
1836. Oryctomys Eydoux and Gervais (in part), Mag. de Zool., 6:20, pl. 21. Type: Oryctomys (Saccophorus) bottae, from coast of California, probably near Monterey.
1903. Megascapheus Elliot, Field Columb. Mus., Publ. 76, Zool. Ser., 3(11):190, July 25. Type: Diplostoma bulbivorum Richardson, from Columbia River, probably near Portland, Ore.
1933. Pleisothomomys Gidley and Gazin, Jour. Mamm. 14:354. Type: Pleisothomomys potomacensis Gidley and Gazin, from Pleistocene, Cumberland Cave local fauna, Allegany County, Maryland.
Chronologic range.—Known from late Pliocene to Recent.
Description.—Same as that given for the tribe Thomomyini above.
Discussion.—Features characterizing Thomomys and the tribe Thomomyini are more advanced than those characterizing the tribe Dikkomyini. Also, the [519] Thomomyini retain more of the primitive features of the Geomyinae than do the more specialized tribe Geomyini.
Specializations are few, but include the third molar being a single column both above and below, enamel plates, and a masseteric ridge.
Key to the Subgenera of Thomomys
Subgenus Pleisothomomys Gidley and Gazin
1933. Pleisothomomys Gidley and Gazin, Jour. Mamm., 14:354, November 13.
Type.—Pleisothomomys potomacensis Gidley and Gazin, 1933.
Chronologic range.—Late Pliocene (Hagerman local fauna, Idaho) to late Pleistocene. The latest records are from the fauna of Saber-tooth Cave, Florida, a late Pleistocene assemblage that probably was deposited in the Sangamon. The middle and late Pleistocene records are from the eastern United States, suggesting that the subgenus Pleisothomomys was restricted to that region while the subgenus Thomomys occupied the western United States and parts of Canada and México as it does today.
Description and Comparison.—Separated from subgenus Thomomys only on basis of sub-crescentic shaped molars (only jaw fragments and isolated teeth known), seemingly a primitive feature of the genus. This dental structure continued into the late Pleistocene; none of the Recent species expresses this feature of the molars, although the molars of Thomomys vetus of the late Pleistocene (Wisconsin deposits), referred to the subgenus Thomomys on the basis of its alleged relationship to Thomomys townsendii (see Davis, 1937:156-158), are less distinctly pear-shaped, and are more sub-crescentic, than in any other known species of the subgenus Thomomys. Pleisothomomys Gidley and Gazin (loc. cit.) was proposed as a genus but is here considered as of no more than subgeneric worth, and is recognized because of the apparent constancy of the sub-crescentic molars in the earlier members of the genus and in those populations of Thomomys occurring in Pleistocene times in the eastern United States.
Referred species.—Three (all extinct):
*Thomomys gidleyi Wilson, 1933. Carnegie Inst. Washington Publ. 440:122, December. Type from Hagerman beds, late Pliocene, Idaho.
*Thomomys potomacensis Gidley and Gazin, 1933. Jour. Mamm., 14:354, November 13. Type from Cumberland Cave, middle and late Pleistocene, Maryland.
*Thomomys orientalis Simpson, 1928. Amer. Mus. Novit., 328:6, October 26. Type from Saber-tooth Cave, late Pleistocene, Florida.
Subgenus Thomomys Wied-Neuwied
1839. Thomomys Wied-Neuwied, Nova Acta Phys.-Med. Acad. Caesar. Leop. Carol., 19(1):377.
1903. Megascapheus Elliot, Field Columb. Mus., Publ. 76, Zool. Ser., 3 (11):190, July 25. Type: Diplostoma bulbivorum Richardson, from Columbia River, probably near Portland, Oregon.
Type.—Thomomys rufescens Wied-Neuwied, 1839.
Chronologic range.—Early Pleistocene (Broadwater-Lisco local fauna, Nebraska) to Recent. Numerous records, mostly isolated teeth, from nearly all stratigraphic levels of the Pleistocene (for details, see account of fossil record).
Description.—Molars pear-shaped in cross-section, becoming abruptly narrow at one end of the tooth. The teeth of the late Pleistocene species Thomomys vetus are less distinctly pear-shaped than other referred species (see remarks in the description of the subgenus Pleisothomomys).
Essentially on the basis of its significantly larger size and details of the skull, Elliott (1903:190) proposed subgeneric recognition of Thomomys bulbivorus and described the subgenus Megascapheus to include it. Also the molars of Thomomys bulbivorus usually have a small enamel plate, both above and below, bordering the persistent inflection on the protomere end of the tooth; each lateral plate is isolated from the transverse plates on the anterior and posterior walls of the tooth. In my opinion these features do not warrant subgeneric recognition; however, these characters do distinctly separate Thomomys bulbivorus from other groups of species, and the character of the molars suggests retention of a primitive trait. Therefore, I propose that the unique structure of this species be recognized by setting it apart in the bulbivorus species-group.
Referred species.—Ten species, three extinct, placed in three species-groups (the numerous subspecies of this genus are listed in Miller and Kellogg, 1955:276-332, and Hall and Kelson, 1959:412-447).
bulbivorus species-group
Thomomys bulbivorus (Richardson, 1829). Fauna Boreali-Americana, 1:206. Type from Columbia River, probably near Portland, Oregon.
umbrinus species-group
*Thomomys scudderi Hay, 1921. Proc. U. S. Nat. Mus., 49:614. Type from Fossil Lake beds, late Pleistocene, Oregon.
Thomomys umbrinus (Richardson, 1829). Fauna Boreali-Americana, 1:202. Type from southern México, probably near Boca de Monte, Veracruz.
Thomomys bottae (Eydoux and Gervais, 1836). Mag. de Zool., Paris, 6:23. Type from coast of California, probably near Monterey.
*Thomomys vetus Davis, 1937. Jour. Mamm., 18:156, May 12. Type from Fossil Lake beds, late Pleistocene, Oregon.
Thomomys townsendii (Bachman, 1839). Jour. Acad. Nat. Sci. Philadelphia, 8:105. Type probably from near Nampa, Canyon Co., Idaho (erroneously given as "Columbia River").
talpoides species-group
*Thomomys microdon Sinclair, 1905. Bull. Dept. Geol. Univ. California, 4:145-161. Type from Potter Creek Cave, late Pleistocene, California.
Thomomys monticola J. A. Allen, 1893. Bull. Amer. Mus. Nat. Hist., 5:48, April 28. Type from Mt. Tallac, 7500 ft., El Dorado Co., California.
Thomomys talpoides (Richardson, 1828). Zool. Jour., 3:518. Type locality fixed at near Fort Carlton (Carlton House), Saskatchewan River, Saskatchewan, Canada.
Thomomys mazama Merriam, 1897. Proc. Biol. Soc. Washington, 11:214, July 15. Type from Anna Creek, 6000 ft., near Crater Lake, Mt. Mazama, Klamath Co., Washington.
Genotype.—Geomys Rafinesque, 1817.
Chronologic and geographic range.—Known from late middle Pliocene deposits to Recent. The range of living members extends from extreme southern Manitoba and the southeastern United States south to southern Panamá, and probably northern Colombia, South America.
Diagnosis.—Size small to large (condylobasal length of skull 33.0 to 73.0 in adults, including both sexes); sexual dimorphism marked, sometimes strongly, females being smaller than males, especially in cranial dimensions; upper incisors invariably grooved, number and position of grooves varying according to genus; cheek teeth high-crowned and ever-growing, except in one primitive genus (Pliogeomys); all three lower molars and M1 and M2 monoprismatic, and elliptical in cross-section in final stages of wear (teeth of young, subadult, and adult animals); primitive biprismatic patterns (as known from Recent specimens) occurring only in pre-final stages of wear (teeth of juveniles only); biprismatic patterns of lower molars as in Dikkomys, and upper molars as in Pliosaccomys (for detailed description of these patterns, see account beyond of the phylogeny of the Geomyinae); m3 becoming monoprismatic, anteroposteriorly compressed and elliptical in cross-section like m1 and m2, but M3 remaining, with rare exceptions (see accounts of Geomys and Pappogeomys beyond), at least partially biprismatic throughout life, having one or both lateral inflections usually persisting (with exceptions) and developing various occlusal shapes (subtriangular, elongate, obcordate, suborbiculate, or quadriform) but never elliptical.
Enamel of cheek teeth reduced to interrupted plates, with exception of p4 in Pliogeomys; plate on posterior wall of P4 variable, occurring completely across posterior surface in primitive members, but progressively reduced to lingual side only or completely lost in modern genera (see generic accounts beyond for detailed description); both anterior and posterior plates usually retained in M1 and M2, posterior plate sometimes reduced to lingual side or completely lost (as in Pappogeomys) but anterior plate always completely retained; M3 usually having three plates, one anterior and two lateral; posterior plate wanting (sometimes lingual plate moved to posterior position); plates retained completely across posterior walls of all lower cheek teeth with no reduction, but anterior plates of m1-3 always lacking, except in primitive genus Pliogeomys (only Geomyini having both anterior and posterior enamel plates on lower molars).
Skull primitively generalized, but becoming specialized towards either dolichocephaly (Orthogeomys) or platycephaly (Pappogeomys) in two modern genera; skull highly specialized for fossorial life; mandible stout and deep, angular process being high and diverging laterally at right angles to ramus; masseteric ridge and fossa weakly developed in primitive members, becoming well developed and massive in modern genera; basitemporal fossa absent in primitive forms (Pliogeomys and early members of Zygogeomys); pelage usually soft, but harsh and hispid in some genera; forefeet broad and massive, claws long and stout for digging; body form remarkably fossorial.
The tribe Geomyini includes the most highly specialized members of the subfamily Geomyinae.
Key to the Genera of the Tribe Geomyini
1954. Pliogeomys Hibbard, Michigan Acad. Sci., Arts and Letters, 39:353.
Genotype.—Pliogeomys buisi Hibbard, 1954, from Buis Ranch local fauna (middle Pliocene), Beaver County, Oklahoma.
Chronologic range.—Latest Middle Pliocene, known only from the highest part of the Hemphillian mammalian fauna (Buis Ranch local fauna, Oklahoma). Professor Hibbard informs me (personal communication) that he found the type, a right ramus, lying on the surface near the base of the fossil beds. The isolated teeth of small geomyids from the Saw Rock Canyon local fauna (see Hibbard, 1953:392) may also be referable to this genus. The Saw Rock Canyon local fauna may also be middle Pliocene in age but is considered to be from the later part of the late Pliocene, and, therefore, somewhat younger than the Buis Ranch local fauna (Hibbard, op. cit.:342).
Description and discussion.—The size of members of this small genus of the Geomyinae is about the same as in smaller adults of Geomys bursarius. According to Hibbard (op. cit.:353), the holotype is smaller than specimens from the Rexroad local fauna referred to Geomys quinni and larger than specimens referred to Zygogeomys cf. minor. The cheek teeth are rooted, and the crowns are as high as those of living geomyids. The upper incisor is bisulcate, and the inner groove is fine and indistinct in places.
Of the molariform dentition only the lower premolar and first two lower molars are known. The enamel investment of p4 is complete, and would not be subject to interruption at any stage of wear; the two prisms are joined at their mid-points, and the isthmus of dentine is relatively broad (as in Pliosaccomys) when compared with modern pocket gophers of this tribe. Also, the re-entrant folds, rather than having parallel sides, diverge broadly to the sides. The divergence is especially noticeable in the labial fold. The lower deciduous premolar would have formed essentially the same enamel pattern with wear as observed in Nerterogeomys [= Zygogeomys] cf. minor (see Hibbard, 1954:fig. 5, A and B) and Pliosaccomys dubius (see Wilson, 1936; pl. 1, fig. 1). Each molar is a single column in the final stages of wear; pre-final stages are unknown. Anterior and posterior enamel plates are present on m1 and m2 (m3 has not been recovered). The dentine tracts of m1 are exposed over a relatively wide surface; therefore, the enamel plates are distinctly separated. The tracts of dentine of m2 are much narrower than in m1 and the enamel plates are barely separated at the anterolateral margin of the tooth. Possibly the enamel band of m2 was continuous in an earlier stage of wear.
The mandible is stout and its general construction not unlike that in modern geomyines. The capsule at the base of the angular process that receives the terminal end of the lower incisor is well developed. The base of the angular processes is preserved, and suggests that the process was short and decidedly smaller than in living examples of the tribe. The masseteric ridge is distinct but weakly developed, and not at all massive as in living pocket gophers. The mental foramen is immediately anterior, and slightly ventral, to the anterior extension of the crest. The basitemporal fossa is absent as such, but its position is marked by a slight depression.
Specimens examined.—Two rami; nos. 29147 (holotype) and 33446; several isolated teeth 30194 and 30195, including an upper incisor and a dp4 (deciduous lower premolar), all from Univ. Michigan Mus. Paleo.
Referred species.—One.
*Pliogeomys buisi Hibbard, 1954. Papers Michigan Acad. Sci., Arts, and Letters, 39:353. Type from Buis local fauna, latest middle Pliocene, Beaver County, Oklahoma.
1895. Zygogeomys Merriam, N. Amer. Fauna, 8:195, January 31.
1942. Nerterogeomys Gazin, Proc. U. S. Nat. Mus., 92:507 (type, Geomys persimilis Hay, 1927).
Type.—Zygogeomys trichopus Merriam, 1895, from Nahuatzen, Michoacán.
Chronologic range.—Late Pliocene (Benson and Curtis Ranch local faunas, Arizona, and ?Rexroad Formation, Kansas) to Recent.
Description and discussion.—The size is small to medium for the subfamily Geomyinae. This genus is distinguished principally by the retention of primitive features. In the living species, the skull is generalized, rather than specialized toward either extreme dolichocephaly or platycephaly. The angular process is short, barely exceeding the lateral extensions of the mastoid process of the squamosal. The rostrum is remarkably narrow in relation to its length. The jugal is reduced and displaced ventrally, causing the maxillary arm of the zygomata to articulate with the squamosal arm of the zygomata along the dorsal border of the zygomatic arch (a feature observed also in Orthogeomys cherriei costaricensis).
The upper incisor, recovered in material from the late Pliocene and middle Pleistocene, is bisulcate as in the genus Geomys and the primitive genus Pliogeomys. The enamel plate across the posterior wall of P4 is either complete (late Pliocene to late Pleistocene) or restricted to the lingual half of the tooth (always restricted in living species). The Pliocene specimens of the Rexroad local fauna referred to Nerterogeomys cf. minor by Hibbard (1950:138-139) are exceptional. In these specimens the length and position of the posterior enamel plate is variable; however, all but one specimen had persistant enamel. Evidently, in approximately 43 per cent of the specimens, a complete enamel blade was present (see Paulson, 1961:139), and in the others (except the one without any enamel) the plate was restricted to a small area of the ventral surface, usually on the lingual side of the loph. Hibbard suggested that the decrease in size of the plate, and its restriction to the lingual side, may be a function of age. Hence, most adults would be characterized by the reduced posterior plate on the upper premolar. Although age may be the important factor, intragroup variation cannot be ruled out. It is of interest to note that in all specimens from the Benson (type series of P. minor) and Curtis Ranch local faunas, the former of late Pliocene age and the latter of middle Pleistocene age, the enamel plates are complete on the posterior face of the upper premolar. As mentioned before, the specimens from Kansas may actually represent the transitional stages of the early evolution of Geomys in which the posterior plate of P4 is entirely lost. The enamel pattern of p4 is like that in other members of the tribe (excepting the genus Pliogeomys). The re-entrant angles of P4 and p4 are widely open (obtuse) in the examples recovered from late Pliocene and middle Pleistocene deposits, representing retention of a trait that is primitive in the Geomyini (see account of phylogeny).
M1 and M2 are elliptical in cross-section and each has an enamel plate on both the anterior and posterior surface. In the living species (Z. trichopus), the posterior enamel plate fails to reach the labial margin of the tooth and is restricted to the lingual two-thirds of the posterior surface; however, the enamel plates are complete in the late Pliocene species (Z. minor) and the middle Pleistocene species (Z. persimilis), being only slightly separated from the anterior plate by narrow tracts of dentine on the ends of the tooth. M3 is partly biprismatic in the living species, the two incompletely divided lophs being separated by a distinct outer sulcus. The posterior loph is elongated and forms a conspicuous heel paralleling the evolution of this trait in the genus Orthogeomys; therefore, the crown is longer than wide. The posterior part of the tooth is protected by two lateral enamel plates; of the two, the lingual plate is especially long and extends to the end of the heel. M3 has not been recovered in the Pliocene species, but in the middle Pleistocene species (Z. persimilis) M3 is subtriangular, no longer than wide, and the lateral inflections are weakly developed. The trend towards elongation of M3 evidently occurred in late Pleistocene evolution of the genus. All three of the inferior molars are elliptical, and only the posterior enamel plate is present (as in all other genera of the tribe except Pliogeomys).
The masseteric ridge of the mandible is well developed. In the late Pliocene species Z. persimilis and Z. minor the mental foramen is directly beneath the anterior extension of the masseteric ridge, but in the living species, Z. trichopus, the foramen lies well anterior to the ridge. The basitemporal fossa in the living [525] species is well developed and deep; in the Pliocene species it is usually distinct but shallow (late Pliocene specimens of Z. minor).
Referred species.—Three (two extinct and one living; the last has two subspecies):
*Zygogeomys minor (Gidley), 1922. U. S. Geol. Surv. Prof. Paper, 131:123, December 26. Type from Benson local fauna (late Pliocene), Cochise County, Arizona; also known from the Rexroad local fauna, Meade County, Kansas.
*Zygogeomys persimilis Hay, 1927. Carnegie Inst. Washington Publ., 136. Originally described by Gidley, 1922 (U. S. Geol. Surv. Prof. Papers, 131:123, December 26) as Geomys parvidens which was preoccupied by G. parvidens Brown, 1908. Type from Curtis Ranch local fauna (middle Pleistocene), Cochise County, Arizona.
Zygogeomys trichopus trichopus Merriam, 1895. N. Amer. Fauna, 8:196, January 31. Type from Nahuatzen, Michoacán.
Zygogeomys trichopus tarascensis Goldman, 1938. Proc. Biol. Soc. Washington, 51:211, December 23. Type from 6 mi. SE Pátzcuaro, 8,000 ft., Michoacán.
1817. Geomys Rafinesque, Amer. Monthly Mag., 2(1):45, November.
1817. Diplostoma Rafinesque, Amer. Monthly Mag., 2(1):44-45, November. Included species: Diplostoma fusca Rafinesque [= Mus bursarius Shaw] and Diplostoma alba Rafinesque [= Mus bursarius Shaw] from the Missouri River region.
1820. Saccophorus Kuhl, Beitr. Zool. und Vergl. Anat., pp. 65, 66. Type: Mus bursarius Shaw, from upper Mississippi Valley.
1823. Pseudostoma Say, Long's Expd. Rocky Mts., I, pp. 406. Type: Pseudostoma bursaria [= Mus bursarius Shaw], from upper Mississippi Valley.
1825. Ascomys Lichtenstein, Abh. K. Akad. Wiss. Berlin (1822), p. 20., fig. 2. Type: Ascomys canadensis Lichtenstein [= Mus bursarius Say], probably from upper Mississippi Valley.
1944. Parageomys Hibbard, Bull. Geol. Soc. Amer., 55:735, June. Type: Parageomys tobinensis Hibbard, from Pleistocene, Cudahy (Tobin) local fauna, Russell Co., Kansas.
Type.—Geomys pinetis Rafinesque, 1817, restricted to Screven County, Georgia, in region of the pines.
Chronologic range.—Late Pliocene faunas of Blancan age (Rexroad, Kansas, and Sand Draw, Nebraska, local faunas) to Recent. Reported from numerous Pleistocene deposits of all stratigraphic levels, especially from the Great Plains, where common today.
Description and discussion.—Pocket gophers of this genus are medium-sized geomyids; none is so small as the average-sized Thomomys. The skull is generalized and lacks the dolichocephalic and platycephalic specializations seen in the genera Orthogeomys and Pappogeomys, respectively. Geomys closely resembles Zygogeomys, but retains fewer of the primitive characters of the ancestral stock. At the same time, Geomys has several specializations. Even so, a considerable amount of parallelism is evident in the phyletic trends of the two genera.
The upper incisor of Geomys is bisulcate as in Pliogeomys and Zygogeomys; the deeper grove is medial and the shallower grove lies near the inner border [526] of the tooth. The premolar, above and below, is bicolumnar; and two columns are joined at their mid-points (deep re-entrant angles separate the columns at the sides). A permanent enamel plate protects the anterior face of the anterior loph, and enamel bands outline each of the re-entrant folds. In p4 a complete enamel plate covers the posterior surface of the posterior loph. All of the enamel bands are interrupted by tracts of dentine, except in the initial stages of wear of the occlusal surface of the newly erupted tooth. For a short time in living Geomys, the enamel bands are continuous as observed in juveniles of Geomys bursarius major (KU 5628, 8531, and 41540). But, the enamel cap is thin and the dentine tracts, which are high on the sides of the tooth, are soon revealed by a minimum of wear on the crown. Therefore, the adult, or final, pattern characterized by interrupted enamel plates emerges early in life and remains throughout the life of the individual. Evidence from fossil Geomys, especially from specimens from early and late Pleistocene deposits, suggests that the final adult pattern appears later, ontogenetically, than in Recent specimens. Some of the fossil premolars in initial stages of wear have continuous and uninterrupted bands of enamel. Geomys quinni of the late Pliocene and early Pleistocene has the interrupted pattern seen in late Pleistocene and Recent Geomys. Also, in late Pliocene and early Pleistocene species, the re-entrant folds diverge laterally and form "open" angles. In later taxa (middle Pleistocene to Recent) the folds are compressed and parallel-sided, and the "open" folds are found only in the early stages of wear.
The posterior enamel plate of P4 disappears in the final stages of wear as the interrupted enamel pattern is formed. In the late Pleistocene and Recent Geomys, the loss of the posterior plate occurs early in life, usually in the first phases of wear on the occlusal surface of the newly erupted tooth, but in fossils of Geomys of corresponding ontogenetic age from the early and middle Pleistocene, the posterior plate is retained in some individuals until a later phase of wear, thereby delaying the appearance of the final pattern. Indeed, in five or fewer per cent of the individuals (see Paulson, 1961:138-139; and White and Downs, 1961:18) a vestige of enamel is retained throughout life or at least until late in adulthood. In Geomys tobinensis, for example, a thin, but transversely complete, plate of enamel occurs all the way down to the base of the loph (Paulson, loc. cit.) and would persist throughout life. In Geomys garbanii, a vestige on the lingual side of the posterior surface of a fully adult specimen was noted by White and Downs (loc. cit.). Vestiges of the posterior plate occur less frequently in living geomyids. Paulson (loc. cit.) found a posterior plate in one of 75 specimens of Geomys bursarius dutcheri. A young (suture present between exoccipitals and supraoccipital) female of Geomys pinetis austrinus (KU 23358) has a vestige of the posterior plate on the lingual side of the tooth as White and Downs (loc. cit.) observed in a specimen of Geomys garbanii. The enamel, I suspect, tends to be thicker on the lingual than on the labial side of the loph and extends farther down the lingual surface in some individuals; therefore, wear on the occlusal surface erodes it down to the dentine more rapidly on the labial than on the lingual side. The tendency of enamel to be retained is a primitive feature.
A lower molar of Geomys is a single elliptical column, and enamel is restricted to the posterior surface as in Zygogeomys, Orthogeomys, and Pappogeomys. Paulson (loc. cit.) found a thin enamel plate on the anterior surfaces of the lower molars in about five per cent of the individuals of Geomys tobinensis [527] from the Cudahy local fauna (middle Pleistocene, deposits of the late Kansan glaciation). An anterior plate is unknown in other members of the tribe Geomyini, except in the primitive genus Pliogeomys of the middle Pliocene. Occurrence of the plate in Geomys tobinensis is an atavistic trait. Primitive dental patterns occur occasionally in geomyids, as pointed out above, but the frequency of occurrence in G. tobinensis is higher than would be expected.
M1 and M2, like the lower molars, are elliptical in cross-section. Complete enamel plates on the anterior and posterior surfaces are separated by tracts of dentine on the sides of each tooth. M3 is usually suborbicular (sometimes subtriangular) in cross-section. The tooth is not especially elongated posteriorly and usually has no definite heel; therefore, it is not significantly longer than wide. Living species of Geomys rarely have a well defined outer re-entrant fold on M3; less than 10 per cent of the individuals (and usually only one side in each individual in which it occurs) have it, although a shallow inconspicuous groove occurs more frequently. The biprismatic molar characteristic of the ancestral morphotype is less often found in Geomys than in any other living member of the tribe Geomyini. The outer re-entrant fold and biprismatic pattern are more often present in the extinct species Geomys garbanii of the Middle Pleistocene than in other species. Less than 24 per cent of the third upper molars in Geomys garbanii lack a tract of the re-entrant fold and more than 38 per cent have a well developed outer fold (see White and Downs, 1961:13, 18). The bicolumnar pattern, although incomplete, would be clearly evident in those teeth having a well marked re-entrant fold; the pattern occurs less frequently in those teeth with no fold or only a slight one. M3 of geomyids is not usually recovered and, therefore, the occlusal pattern of M3 is unknown in most extinct kinds of Geomys. In Recent Geomys the fold is more common in the eastern pinetis species-group than in the western bursarius species-group.
The masseteric ridge on the outer side of the mandible is well developed in all species of the genus. The position of the mental foramen relative to the anterior part of the ridge varies with individuals and according to species. The basitemporal fossa is always present, but is shallower in the late Pliocene and Pleistocene species than in Recent species. The angular process is short.
Referred species.—The twelve species, five of which are extinct, are as follows:
quinni species-group
*Geomys quinni McGrew, 1944. Geol. Ser., Field Mus. Nat. Hist., 9 (546):49, January 20. Type from Sand Draw local fauna (late Pliocene), Brown County, Nebraska; also known from Broadwater-Lisco local faunas (early Pleistocene), Morrill and Garden counties, Nebraska, Deer Park local fauna (early Pleistocene), Meade County, Kansas.
*Geomys paenebursarius Strain, 1966. Bull. Texas Memorial Mus., 10:36. Type from Hudspeth local fauna (early Pleistocene), Hudspeth County, Texas.
*Geomys tobinensis Hibbard, 1944. Bull. Geol. Soc. Amer., 55:736. Type from Tobin local fauna (middle Pleistocene), Russell County, Kansas; also known from Cudahy local fauna (middle Pleistocene), Meade County, Kansas.
*Geomys garbanii White and Downs, 1961. Contrib. Sci., Los Angeles Co. Mus., 42:1-34, June 30. Type from Vallecito Creek local fauna (middle Pleistocene), San Diego County, California.
*Geomys bisulcatus Marsh, 1871. Amer. Jour. Sci., 3:121. Type from Loup River fossil beds, near Camp Thomas, Nebraska (probably late Pleistocene).
bursarius species-group
*Geomys parvidens Brown, 1908. Mem. Amer. Mus. Nat. Hist., 9:194. (An extinct subspecies of Geomys bursarius according to White and Downs, 1961:6). Type from Conard Fissure local fauna (late Pleistocene), northern Arkansas.
Geomys bursarius (Shaw, 1800). Trans. Linn. Soc. London, 5:227. Type from somewhere in Upper Mississippi Valley, North America.
Geomys arenarius Merriam, 1895. N. Amer. Fauna, 8:139, January 31. Type from El Paso, El Paso County, Texas.
Geomys personatus True, 1889. Proc. U. S. Nat. Mus., 11:159, January 5. Type from Padre Island, Cameron County, Texas.
pinetis species-group
Geomys pinetis Rafinesque, 1806. Amer. Monthly Mag., 2 (1):45, November. Type locality restricted to Screven County, Georgia.
Geomys colonus Bangs, 1898. Proc. Boston Soc. Nat. Hist., 28:178, March. Type from Arnot Plantation, about 4 mi. W St. Marys, Camden County, Georgia.
Geomys cumberlandius Bangs, 1898. Proc. Boston Soc. Nat. Hist., 28:180, March. Type from Stafford Place, Cumberland Island, Camden County, Georgia.
Geomys fontanelus Sherman, 1940. Jour. Mamm., 21:341, August 13. Type from 7 mi. NW Savannah, Chatham County, Georgia.
1895. Orthogeomys Merriam, N. Amer. Fauna 8:172, January 31.
1895. Heterogeomys Merriam, N. Amer. Fauna 8:179, January 31 (type, Geomys hispidus Le Conte, 1862).
1895. Macrogeomys Merriam, N. Amer. Fauna 8:185, January 31 (type, Geomys heterodus Peters, 1865).
Type.—Geomys scalops Thomas, 1894, from Tehuantepec, Oaxaca, México.
Chronologic range.—Late Pleistocene Wisconsin deposits (San Josecito Cave local fauna, Nuevo León, México) to Recent.
Description and discussion.—Species of this genus are of medium to large size. The skull is strongly dolichocephalic in most species; the posterior part of the skull is especially narrow. The angular processes are remarkably short, especially in relation to the length of the mandible. The nasals and rostrum are relatively broad and heavy. The pelage is coarse, and often hispid. In some species the hairs are so sparsely distributed that the body appears almost naked, and none has so dense a covering of hair as do other genera. The genus occurs entirely within the tropical life-zones, and most of the external features seem to be associated with adaptation to tropical conditions.
The upper incisor is unisulcate; the sulcus is usually near the inner border of the tooth, but in some species (subgenus Orthogeomys) it is more medial, and in a few individuals with an extremely wide groove the outer lip of the sulcus may actually reach the middle of the tooth. The groove is compressed or open. The premolar is a double column united at the mid-point. The two prisms are of approximately equal size, and the lateral re-entrant folds are so compressed that their sides are parallel. Enamel plates cover the anterior surface and border the re-entrant angles in both upper and lower premolars. As in other members of the tribe, the lower premolar has a fourth enamel plate on the posterior surface of the posterior lophid. In the upper premolar, the enamel plate is reduced to a narrow blade on the lingual side of the loph [529] as in the living species of the genus Zygogeomys. In the subgenus Orthogeomys the posterior plate is usually absent, and otherwise is narrow and near the lingual border of the tooth.
Each lower molar, in the final stage of wear, consists of a single elliptical column having an enamel plate only on the posterior surface. The first and second upper molars are single elliptical columns having one enamel plate on the anterior surface and another on the posterior surface. The plates are separated by a tract of dentine on each side of the tooth. The third upper molar is partly bilophodont, and the two lophs are separated by a deep outer re-entrant fold. In many of the species an inner re-entrant fold also is retained, but in the adult tooth it is less distinct than the outer. In all of the species the posterior loph is long and forms a conspicuous heel; consequently the crown is significantly longer then wide. Moreover, the posterior loph has an enamel plate on each side. The labial plate always borders the outer re-entrant fold, and in the subgenus Orthogeomys is infrequently separated into two small plates.
The mandible is relatively long. Its masseteric ridge is well developed and massive. The basitemporal fossa is usually deep and well defined; it tends to be shallow in the subgenus Orthogeomys, and in young individuals is hardly more than a slight depression.
Key to the Subgenera of Orthogeomys
1895. Orthogeomys Merriam, N. Amer. Fauna, 8:172, January 31.
Type.—Geomys scalops Thomas, 1894, from Tehuantepec, Oaxaca, México.
Chronologic range.—Known only from the Recent.
Description.—Skull elongated and narrow (many skulls of nearly uniform breadth throughout), being extreme in dolichocephalic specializations; mandibles long and narrow, rami not spreading laterally, being more nearly parallel-sided than in other subgenera; angular processes short; breadth across zygomata not significantly exceeding breadth across mastoid processes (in many skulls considerably less); interorbital area remarkably broad, lacking deep constriction; frontals between orbits greatly inflated laterally, postorbital prominence inconspicuous; mesopterygoid fossa extending to level of posterior margin of M3; I having sulcus broader than in other subgenera, mostly on inner half of anterior surface but sometimes overlapping mid-line; enamel plate lacking from [530] posterior wall of P4, rarely retaining narrow vestige near lingual border of posterior loph; M3 having distinct heel, bicolumnar pattern with inner re-entrant fold usually minute, occlusal length less than in other subgenera, length less than combined lengths of M1-2; hair generally coarse, sometimes hispid, sparse, in lowland forms, so sparse as to impart appearance of nakedness.
Referred species and subspecies.—Fourteen taxa:
Orthogeomys grandis alleni Nelson and Goldman, 1930. Jour. Mamm., 11:156, May 9. Type from near Acapulco, 2000 ft., Guerrero.
Orthogeomys grandis annexus Nelson and Goldman, 1933. Proc. Biol. Soc. Washington, 46:195, October 26. Type from Tuxtla Gutierrez, 2600 ft., Chiapas.
Orthogeomys grandis carbo Goodwin, 1956. Amer. Mus. Novit., 1757:5, March 8. Type from Excurano, 2500 ft., Cerro de San Pedro, 20 km. W Mixtequilla, Oaxaca.
Orthogeomys grandis felipensis Nelson and Goldman, 1930. Jour. Mamm., 11:157, May 9. Type from Cerro San Felipe, 10 mi. N Oaxaca, Oaxaca.
Orthogeomys grandis huixtlae Villa, 1944. Anal. Inst. Biol. Univ. Nac. México, 15:319. Type from Finca Lubeca, 12 km. NE Huixtla, 850 m., Chiapas.
Orthogeomys grandis grandis (Thomas, 1893). Ann. Mag. Nat. Hist., ser. 6, 12:270, October. Type from Dueñas, Guatemala.
Orthogeomys grandis latifrons Merriam, 1895. N. Amer. Fauna, 8:178, January 31. Type from Guatemala, exact locality unknown.
Orthogeomys grandis nelsoni Merriam, 1895. N. Amer. Fauna, 8:176, January 31. Type from Mt. Zempoaltepec, 8000 ft., Oaxaca.
Orthogeomys grandis pluto Lawrence, 1933. Proc. New England Zool. Club, 13:66, May 8. Type from Cerro Cantoral, north of Tegucigalpa, Honduras.
Orthogeomys grandis scalops (Thomas, 1894). Ann. Mag. Nat. Hist., ser. 6, 13:437, May. Type from Tehuantepec, Oaxaca.
Orthogeomys grandis soconuscensis Villa, 1949. Anal. Inst. Biol. Univ. Nac. México, 19:267, April 8. Type from Finca Experanza, 710 m., 45 km. (by road) NW Huixtla, Chiapas.
Orthogeomys grandis guerrerensis Nelson and Goldman, 1930. Jour. Mamm., 11:158, May 9. Type from El Limón, in valley of Río de las Balsas approximately 20 mi. NW La Unión, Guerrero.
Orthogeomys cuniculus Elliot, 1905. Proc. Biol. Soc. Washington, 18:234, December 9. Type from Zanatepec, Oaxaca.
Orthogeomys pygacanthus Dickey, 1928. Proc. Biol. Soc. Washington, 41:9, February 1. Type from Cacaguatique, 3500 ft., Dept. San Miguel, El Salvador.
1895. Heterogeomys Merriam, N. Amer. Fauna, 8:179, January 21.
Type.—Geomys hispidus Le Conte, 1852, from near Jalapa, Veracruz.
Chronologic range.—Late Pleistocene, Wisconsin deposits (San Josecito Cave local fauna, Nuevo León) to the Recent.
Description.—Skull dolichocephalic (less so than in the other subgenera); zygomata more widely spreading than in Orthogeomys; ramus and angular process short; interorbital area noticeably constricted; frontals between orbits neither exceptionally broad or inflated; mesopterygoid fossa extending to level of posterior margin of M3; I having sulcus on inner third of anterior surface usually narrower than in subgenus Orthogeomys; enamel plate on posterior wall of P4 restricted to lingual half of loph; M3 distinctly biprismatic, posterior loph usually circumscribed by shallow inner re-entrant fold and outer [531] deep fold well developed in all members of genus; posterior loph forming conspicuous heel longer than in subgenus Orthogeomys; occlusal length equal to or slightly less than combined lengths of M1-2; hair coarse and hispid but never so sparse as to impart appearance of nakedness.
Referred species and subspecies.—Eleven taxa:
*Orthogeomys onerosus (Russell, 1960). Univ. Kansas Publ., Mus. Nat. Hist., 9 (21):544, January 14. Type from San Josecito Cave local fauna, Upper Pleistocene, Nuevo León.
Orthogeomys hispidus cayoensis (Burt, 1937). Occ. Papers Mus. Zool., Univ. Michigan, 365:1, December 16. Type from Mountain Pine Ridge, 12 mi. S El Cayo, British Honduras.
Orthogeomys hispidus chiapensis (Nelson and Goldman, 1929). Proc. Bio. Soc. Washington, 42:151, March 30. Type from Tenejapa, 16 mi. NE San Cristobal, Chiapas.
Orthogeomys hispidus concavas (Nelson and Goldman, 1929). Proc. Biol. Soc. Washington, 42:148, March 30. Type from Pinal de Amoles, Querétaro.
Orthogeomys hispidus hispidus (Le Conte, 1852). Proc. Acad. Nat. Sci. Philadelphia, 6:158. Type from near Jalapa, Veracruz.
Orthogeomys hispidus latirostris (Hall and Alvarez, 1961). Anal. Escuela Nac. Ciencias Biol., 10:121, December 20. Type from Hacienda Tamiahua, Cabo Rojo, Veracruz.
Orthogeomys hispidus negatus (Goodwin, 1953). Amer. Mus. Novit., 1620:1, May 4. Type from Gomez Ferias, 1300 ft., about 45 mi. S Ciudad Victoria, 10 km. W Pan American Highway, Tamaulipas.
Orthogeomys hispidus tehuantepecus (Goldman, 1939). Jour. Washington Acad. Sci., 29:174, April 15. Type from mountains 12 mi. NW Santo Domingo and about 60 mi. N Tehuantepec, 1600 ft., Oaxaca.
Orthogeomys hispidus torridas (Merriam, 1895). N. Amer. Fauna, 8:183, January 31. Type from Chichicaxtle, Veracruz.
Orthogeomys hispidus yucatanensis (Nelson and Goldman, 1929). Proc. Biol. Soc. Washington, 42:150, March 30. Type from Campeche, Campeche.
Orthogeomys lanius (Elliot, 1905). Proc. Biol. Soc. Washington, 18:235, December 9. Type from Xuchil, Veracruz.
1895. Macrogeomys Merriam, N. Amer. Fauna, 8:185, January 31.
Type.—Geomys heterodus Peters, 1865, from Costa Rica, exact locality unknown.
Chronologic range.—Known only from the Recent.
Description.—Skull dolichocephalic in varying degree (overlapping subgenera Orthogeomys and Heterogeomys in this respect); mandibles elongated, not spreading far laterally; angular processes decidedly short; breadth across zygomata in no instance significantly exceeding mastoid breadth; interorbital area strongly constricted; frontals between orbits slightly inflated laterally (especially in forms having more strongly dolichocephalic skulls); postorbital prominence conspicuous; anterior margin of mesopterygoid fossa terminating well behind M3; I having narrow and deep sulcus entirely on inner third of anterior surface; enamel plate on posterior wall of P4 restricted to inner half of loph; M3 bilophodont (outer and inner re-entrant folds each circumscribing a loph), posterior loph remarkably elongated and forming pronounced heel, length of crown more than combined lengths of M1-2; hair wooly in some individuals, harsh in others but seldom hispid, never so sparse as in subgenus Orthogeomys; some species having white markings, especially on lumbar region and head.
Referred species and subspecies.—Eleven taxa:
Orthogeomys heterodus cartagoensis (Goodwin, 1943). Amer. Mus. Novit., 1227:2, April 22. Type from Paso Ancho, Province Cartago, Costa Rica.
Orthogeomys heterodus dolichocephalus (Merriam, 1895). N. Amer. Fauna, 8:189, January 31. Type from San José, Costa Rica.
Orthogeomys heterodus heterodus (Peters, 1865). Monatsb. preuss. Acad. Wiss., Berlin, 1865:177. Type from Costa Rica, exact locality unknown.
Orthogeomys cavator nigrescens (Goodwin, 1943). Amer. Mus. Novit., 1227:3, April 22. Type from El Muneco (Río Navarro), 10 mi. S Cartago, 4000 ft., Province Cartago, Costa Rica.
Orthogeomys cavator pansa (Bangs, 1902). Bull. Mus. Comp. Zool., 39:44, April. Type from Bogava (= Bugaba), 600 ft., Chiriquí, Panamá.
Orthogeomys dariensis (Goldman, 1912). Smithsonian Misc. Coll., 60(2):8, September 20. Type from Cana, 2000 ft., mountains of eastern Panamá.
Orthogeomys underwoodi (Osgood, 1931). Field Mus. Nat. Hist., Publ. 295, Zool. Ser., 185:143, Aug. 3. Type from Alto de Jabillo Pirris, between San Geronimo and Pozo Azul, western Costa Rica.
Orthogeomys cherriei carlosensis (Goodwin, 1943). Amer. Mus. Novit., 1227:3, April 22. Type from Cataratos, San Carlos, Alajuela, Costa Rica.
Orthogeomys cherriei cherriei (J. A. Allen, 1893). Bull. Amer. Mus. Nat. Hist., 5:337, December 16. Type from Santa Clara, Costa Rica.
Orthogeomys cherriei costaricensis (Merriam, 1895). N. Amer. Fauna, 8:192, January 31. Type from Pacuare, Costa Rica.
Orthogeomys matagalpae (J. A. Allen, 1910). Bull. Amer. Mus. Nat. Hist., 28:97, April 30. Type from Peña Blanca, Matagalpa, Nicaragua.
1895. Pappogeomys Merriam, N. Amer. Fauna, 8:145, January 31.
1895. Cratogeomys Merriam, N. Amer. Fauna, 8:150, January 31. Type: Geomys merriami Thomas.
1895. Platygeomys Merriam, N. Amer. Fauna, 8:162, January 31. Type: Geomys gymnurus Merriam; Hooper, Jour. Mamm., 27:397, November 25, 1946.
Type.—Geomys bulleri Thomas, 1892, from near Talpa, west slope Sierra de Mascota, 8500 ft. (actually about 5000 ft.), Jalisco.
Chronologic range.—Late Pliocene, from deposits of early Blancan age (Benson local fauna, Arizona) to the Recent. However in the Pleistocene, only late Pleistocene records are known, and Pappogeomys has not been found in early (late Blancan) or middle (Irvingtonian) Pleistocene local faunas. Presumably the genus was restricted to México during the Pleistocene until post-Wisconsin time.
Description and discussion.—The size ranges from as little as in the smaller kinds of Thomomys to the maximum attained in the subfamily and matched elsewhere perhaps in only a few of the larger subspecies of Orthogeomys grandis. Depending on the species and subgenus, the form of the skull varies from generalized to specialized. The generalized skulls are short and not especially narrow; the zygomatic arches are spread laterally so far that the breadth across them exceeds the breadth across the mastoid processes. The most specialized skulls are platycephalic and the breadth across the [533] mastoid processes equals or exceeds the breadth across the zygomatic arches (even so, the zygomatic arches are still relatively widespread). In correlation with the great breadth of the posterior part of the cranium, the rami of the mandibles diverge widely posteriolaterally and the angular processes are remarkably elongated. The rostrum is moderately broad in most species, but not nearly so broad and heavy as in Orthogeomys.
The single deep, median sulcus on the outer surface of the upper incisor is slightly displaced to the inner side of the tooth. The posterior surface of P4 lacks enamel (small vestige found on lingual end of posterior wall in only two adult individuals—UA 3260 and KU 100442, of the subgenus Pappogeomys); the other three plates are fully developed as usual. The p4 is provided with four fully developed enamel plates, in the pattern characteristic of the tribe Geomyini. In the p4 of the late Pliocene species (P. bensoni) the re-entrant angles are open (obtuse), a trait that is evidently primitive in the Geomyini.
All three lower molars are single, compressed, elliptical columns with enamel on only the posterior surfaces. M1 and M2 are also elliptical in cross-section and decidedly anteroposteriorly compressed, like the lower molars. Nevertheless, the enamel pattern is variable; enamel plates may be retained completely across both the anterior and posterior walls of M1 and M2 or only the anterior plate may be retained without reduction and the posterior plate may be reduced so that only a vestige is retained on the lingual fourth of the tooth or the posterior plate may be completely lost.
M3 tends to remain at least incompletely bilophodont by reason of retaining a permanent labial re-entrant fold in most species (with exceptions in Pappogeomys bulleri and some old adults of P. castanops). Primitively the occlusal surface of M3 is subtriangular (subgenus Pappogeomys), but in the castanops species-group of the advanced subgenus Cratogeomys, the posterior loph usually is reduced and the occlusal surface is quadriform or obcordate. Curiously, the trend towards reduction of the posterior loph is reversed in one subspecies (P. merriami fulvescens) and, the loph has elongated into a pronounced heel in some specimens, resembling the condition in Orthogeomys. The entire range of variation occurs in P. m. fulvescens. The subtriangular pattern is retained in the most specialized species of Cratogeomys where that pattern is associated with extreme platycephaly in the gymnurus species-group. In most species the posterior loph supports two lateral plates, the outer one always bordering the labial re-entrant fold. In Pappogeomys bulleri and in the castanops species-group, the outer re-entrant fold of M3 tends to be obsolete, and the tooth becomes quadriform or suborbiculate in some individuals and loses the bilophodont pattern that characterizes other species. The lingual enamel plate is displaced to the posterior surface of the tooth, and one or both plates may disappear with advancing age. Consequently, only the anterior enamel plate remains in some adults, and constitutes the maximum degree of reduction of enamel on M3 in the Geomyinae. In many adults of Pappogeomys bulleri, the enamel investment of the posterior loph is complete and the two lateral plates are connected, without interruption around the posterior apex of the tooth, evidently representing the retention of a primitive character of the ancestral lineage.
The m3 of P. bensoni from the late Pliocene is distinguished by minute lateral inflections suggesting the primitive biprismatic pattern. Also the posterior enamel plates of m1 and m2 are remarkably long, extending around the [534] ends of the tooth. The associated upper incisor was unisulcate as in the modern species, and the basitemporal fossa of the mandible is well developed and deep.
The lower jaw is stout and relatively short. The masseteric ridge is well developed and has an especially thick crest. The basitemporal fossa is deep. In most living species, the pelage is soft and dense, but in one species, Pappogeomys fumosus, the hairs are coarse and hispid somewhat as in Orthogeomys.
Key to the Subgenera of Pappogeomys
1895. Pappogeomys Merriam, N. Amer. Fauna, 8:145, January 31.
Type.—Geomys bulleri Thomas, 1892, from near Talpa, west slope Sierra de Mascota, 8500 ft. (actually about 5000 ft.), Jalisco.
Chronologic range.—Late Pliocene (Benson local fauna, Arizona) to Recent, but no specimens known from Pleistocene.
Description.—Small, approximately same size as small subspecies of Thomomys umbrinus but forefeet larger and claws longer; skull of generalized shape, broad, relatively short, smoothly rounded, not especially compressed dorso-ventrally; zygomatic breadth great but not exceeding mastoid breadth; zygomata relatively slender for geomyid and lacking platelike expansions at lateral angles; rostrum relatively narrow; sagittal crest lacking, owing to impressions of temporal muscles remaining separated; angular process of mandible not especially elongated; enamel plates extending completely across posterior wall of M1 and M2, except in one species, P. alcorni, where posterior plate of M1 remains only on lingual fourth of posterior wall (remainder of plate lacking); with wear, plates sometimes exceptionally thin completely across posterior face of M2 and especially M1 in a few individuals of P. bulleri much as Paulson (1961:138-139) describes in extinct Geomys tobinensis; one or both plates rarely disappear in final stages of attrition in old individuals resulting in same dental pattern found in Cratogeomys; M1 and M2 retaining enamel plate on anterior wall throughout life; M3 usually subtriangular in cross-section but sometimes suborbiculate or ovoid, crown slightly bilophodont owing to shallowness of labial re-entrant angle in modern species; posterior loph of M3 not especially elongated and crown not significantly longer than wide; both lateral enamel plates of M3 usually well developed and approximately equal in length, occasionally plates reduced in length and rarely one or both plates are lost with wear in old individuals; patch of whitish or buffy hairs surrounding nose of most individuals.
The primitive character of the lower dentition, as described in the species account above, suggest that Cratogeomys [= Pappogeomys] bensoni Gidley should be referred to the subgenus Pappogeomys rather than Cratogeomys. Only the upper dentition would make positive identification possible; however, reference to the subgenus Pappogeomys seems to be the best arrangement at this time.
Referred species.—Three (one extinct):
*Pappogeomys bensoni (Gidley), 1922. U. S. Geol. Surv. Prof. Papers, 131:123. Type from Benson local fauna (late Pliocene), Cochise County, Arizona.
Pappogeomys alcorni Russell, 1957. Univ. Kansas Publ. Mus. Nat. Hist., 9(11):359. Type from 4 mi. W Mazamitla, Jalisco.
Pappogeomys bulleri Thomas, 1892. Ann. Mag. Nat. Hist., Ser. 6, vol. 10:196, August. Type from "near Talpa," west slope of Sierra Madre de Mascota, Jalisco.
1895. Cratogeomys Merriam, N. Amer. Fauna, 8:150, January 31.
1895. Platygeomys Merriam, N. Amer. Fauna, 8:162, January 31. Type: Geomys gymnurus Merriam, 1892.
Type.—Geomys merriami Thomas, 1893, from "Southern México," probably in Valley of México.
Chronologic range.—Late Pleistocene, from Wisconsin deposits (San Josecito Cave, Nuevo León, Upper Bercerra, México, and Burnet Cave, New Mexico, local faunas) to the Recent.
Description.—Size medium to large; skull becoming angular and rugose with age, and tending towards platycephaly and dorso-ventral compression; zygomata stout, each bearing platelike expansion at anterolateral angle into which anterior end of jugal becomes morticed; breadth across zygomata great relative to length of skull; rostrum relatively broad; squamosals expanding medially with age eventually growing over lateral parts of parietals, and sometimes also expanding laterally displacing postglenoid notch; sagittal crest well developed in adults of both sexes, but especially high and bladelike in males; lambdoidal crest prominent in all but young animals, having dorsal outline broadly convex posteriorly in most species but strongly sinuous in gymnurus-group; enamel plate on posterior wall of P4 absent; enamel plates present only on anterior walls of M1 and M2; M3 variform in occlusal shape (as described in species account), either subtriangular (gymnurus-group), quadriform or obcordate (castanops-group, with exceptions as noted before); lateral plates of M3 usually present in all species, labial plate approximately as long as lingual plate in gymnurus-group (like that in subgenus Pappogeomys) or distinctly shorter in castanops-group (labial plate scarcely extending beyond border of labial re-entrant fold); one or both lateral plates tending to disappear with wear in castanops-group, with lingual plate usually disappearing first; breadth across angular processes clearly more than breadth across zygomatic processes, especially in gymnurus-group.
Remarks.—In the species of the castanops-group the skulls can be spoken of as generalized and the least platycephalic of the subgenus. Indeed, the species of the castanops-group are hardly more specialized in this respect than is the subgenus Pappogeomys. In these skulls the breadth across the squamosal processes is less than that across the zygomatic arches, although the two dimensions are almost equal in some examples of P. merriami of the castanops-group (where squamosal breadth varies from 85 to 98% of zygomatic breadth). In the species having marked platycephalic skulls (gymnurus species-group) the breadth across the squamosal processes equals or exceeds the breadth across the zygomatic arches (squamosal breadth rarely 97 to 99% of zygomatic breadth), except in P. zinseri and P. tylorhinus zodius.
The variable character of the third upper molar as between species suggests that this tooth is presently undergoing active evolution. The structure of this tooth, although differing between taxa, is remarkably stable in other kinds of Geomyini. The most remarkable modification of M3 in Cratogeomys [536] is the obcordate pattern developed in P. merriami of the castanops-group. The posterior loph and entire tooth is shortened somewhat resembling in shape that of Thomomys. Moreover, the posterior loph is twisted labially; consequently, its posterior surface now forms the labial border of the weakly defined posterior loph. Owing to the torsion, the lingual enamel plate has been rotated to the posterior surface of the tooth. Therefore, the tooth is provided with two transverse enamel plates, including the plate on the anterior wall of the tooth. The labial plate is greatly reduced, its total surface being restricted to the small labial inflection. The highly specialized obcordate M3 is not found in the most specialized platycephalic skulls characteristic of the gymnurus species-group. Instead the gymnurus-group retains the primitive subtriangular pattern without significant modification.
Referred species.—Seven:
castanops species-group
Pappogeomys castanops (Baird, 1852). Report Stanbury's Exp'd. to Great Salt Lake, p. 313, June. Type from "Prairie road to Bent's Fort," near present town of Las Animas, Colorado.
Pappogeomys merriami (Thomas, 1893). Ann. Mag. Nat. Hist., ser. 6, 12:271, October. Type from "southern Mexico," probably Valley of México (see Merriam, 1895:152).
gymnurus species-group
Pappogeomys fumosus (Merriam, 1892). Proc. Biol. Soc. Washington, 7:165, September 29. Type from 3 mi. W Colima, Colima.
Pappogeomys gymnurus (Merriam, 1892). Proc. Biol. Soc. Washington, 7:166, September 29. Type from Zapotlan (Ciudad Guzman), Jalisco.
Pappogeomys neglectus (Merriam, 1902). Proc. Biol. Soc. Washington, 15:68, March 22. Type from Cerro de la Calentura, about 8 mi. NW Pinal de Amoles, Querétaro.
Pappogeomys tylorhinus (Merriam, 1895). N. Amer. Fauna, 8:167, January 31. Type from Tula, Hidalgo.
Pappogeomys zinseri (Goldman, 1939). Jour. Mamm., 20:91, February 15. Type from Lagos, Jalisco.
The fossil record of the Geomyidae provides a sequence of morphotypes, each representing a stage in the phyletic development of the family. Most of the preserved specimens probably represent the stufenreihe rather than the ahnenreihe, as Simpson (1953:219-220) points out. Even so, the stufenreihe closely approximates the general trend of evolution, and the level of structural organization in the different stages of phyletic development may be ascertained. The actual ancestral series of most lineages probably will remain unknown, but hopefully some of the existing gaps will be filled by future discoveries. From the established record, several clearly defined lineages can be distinguished; in fact the sequence of origin, pattern of evolution, and specializations, of the principal lineages are reasonably well expressed.
In the earliest known geomyids from the Upper Oligocene and Lower Miocene, the premolars and molars are biprismatic and bilophodont. In rodents, this is itself a specialized pattern, and is thought to have evolved from a more primitive sextituberculate prototype by the union of individual cusps, and probably also cuspules, forming the two transverse enamel lophs. The primitive, common ancestor of the Geomyidae and Heteromyidae with sextituberculate teeth in the early Tertiary is unknown.
As soon as geomyids attained the early bilophodont stage of evolution, the basic morphological structure of the family was established. The family probably first became clearly distinguished from other Geomyoidea at this stage. In the early bilophodont stages of evolution, owing to the relatively deep valley between them, the two columns probably failed to unite in the normal cycle of wear, as they do in all later geomyids. Griphomys described by Wilson (1940:93) from the late Eocene of California, has a bilophate pattern in which the anterior and posterior lophs are separated by a persistent transverse valley. The occlusal pattern of Griphomys closely resembles a stage through which the ancestors of the early Miocene geomyids must have passed in their pre-Miocene evolution, as Wilson suggests (1949:115-116). Although he (1940:95; 1949:110-118) tentatively referred Griphomys to the superfamily Geomyoidea and Simpson (1945:80) went so far as to refer it to the family Geomyidae, with a notation of incertae sedis, its exact relationship to the pocket gophers is uncertain. However, the structure of the molariform dentition of Griphomys does not exclude it from the phyletic ancestry of the Geomyidae. In subsequent stages of evolution the anterior and posterior columns become united. Thereby part of the valley floor between the transverse prisms was progressively elevated, to the stage where attrition on the occlusal surface would unite the two columns. On the unworn enamel cap of living geomyids the two transverse enamel folds are separated by a shallow but well defined valley, briefly reflecting the ancient ancestral pattern.
Union of the lophs may have been either at the mid-points of the two columns or at the edge of their protomeres. [A protomere is the half of a tooth containing the protocone or protoconid—lingual side of upper tooth and labial side of lower tooth. The paramere is the opposite half of a given tooth—labial side of upper tooth and [538] lingual side in lower tooth. See Miller and Gidley, 1918:434.] Union of the columns at the mid-points would have produced the figure-8 occlusal pattern (or H-pattern), which is characteristic of the early Miocene Geomyinae (Dikkomys). Union of the two columns at the protomeres would have produced the U-shaped pattern of the Entoptychinae, which also occurred in the early Miocene and were contemporary with the earliest Geomyinae. Since pre-Miocene geomyids are unknown, the actual phyletic development of the dentition is a matter of speculation. Probably the development of the two divergent lineages, one leading to the Entoptychinae and the other to the subfamily Geomyinae, occurred in the Oligocene (as depicted in Fig. 3). Of the two lineages, the subfamily Geomyinae, in my view, is the more primitive and less specialized. Support for this view is furnished by a reconstruction of the pattern of occlusal wear in Dikkomys and Pliosaccomys, especially on the first and second molars.
In Dikkomys, the anterior and posterior column first unite near their mid-points in the first stages of wear thus producing a figure-8 shaped (H-shaped) occlusal pattern in the premolar and all three molars. Evidently in the first two upper molars, the columns unite closer to their lingual margins than their mid-points, but at any rate both outer and inner re-entrant folds are evident at this stage of wear. With continued attrition on m1 and m2 of Dikkomys, the anterior and posterior columns secondarily unite at the edge of their labial margins thus enclosing a fossette of enamel in the labial half of the tooth. The lateral coalescence at the ends of the protomeres occurs because of the shallow vertical depth of the labial re-entrant fold, and the fossette itself does not reach the base of the crown and with continued wear it too would disappear, but not until the last stages of wear, at least in Dikkomys matthewi. The lingual re-entrant fold is deep, and therefore, persistent through all stages of wear. Although the amount of wear required for its effacement would be great, the occlusal configuration of the first and second lower molars in Dikkomys could be eventually ground down to a U-pattern as in the entoptychids. Only one upper molar of Dikkomys, the first, has been recovered (see Wood, 1936:23, fig. 32B). Although the tooth is in an early stage of wear, the lingual valley is minute. Less attrition than required in m1 and m2 would progressively reduce the lingual fold until it too would essentially form a U-pattern, perhaps retaining a slight lingual inflection. Hence, the first upper molar becomes a mirror image of the first lower molar, and the second upper molar probably had the same [539] pattern as the first (at least it does so in Pliosaccomys). Both of the lateral re-entrant folds of the premolar are deep vertically, and consequently would not disappear with occlusal wear. Therefore, the H-pattern of the premolars is retained throughout life.
The m3 (M3 unknown for Dikkomys or Pliosaccomys) also has deep lateral folds; hence, it too retains the H-pattern in all stages of attrition, although the isthmus between the two prisms may become wider in the final phases of wear (as it does in Pliosaccomys).
In Pliosaccomys, the stages of wear are essentially the same as those described for Dikkomys, except that the anterior and posterior loph of the first and second molars tend to unite closer to one side of the tooth, lingual side in upper molars and labial in lower. Only a slight inflection of the re-entrant fold is evident on the side of union, and the inflection disappears in the first phases of wear as the columns unite. Concomitant with the lateral shift in the initial point of coalescence of the transverse lophs, the occlusal penetration of the re-entrant fold from the opposite side increases in horizontal depth, and the fold extends medially more than half way across the occlusal surface, thus forming a pattern essentially like that of the entoptychids. The U-pattern in Pliosaccomys appears in the initial stages of wear without going through an earlier H-pattern as is the case in its Miocene ancestors of the genus Dikkomys, unless the minute inflection is considered as indicative of that stage. The two columns of the premolar and m3 are joined near their mid-points as in Dikkomys; therefore, they retain their primitive H-pattern, a feature unique to the Geomyinae.
The evolutionary trend toward an ontogenetically earlier U-pattern in the first two molars in the primitive lineage of the Geomyinae suggests that the U-pattern characteristic of the Entoptychinae was simply an earlier tendency toward the same specialization that occurred later in the subfamily Geomyinae. If so, early entoptychines would have been characterized by an H-pattern in the first stages of attrition, like Dikkomys, and later developed union at the edge of the protomeres. However, in the entoptychines, all the molariform dentition, and not merely the first and second molar, became specialized; consequently the U-pattern was produced on the occlusal surfaces of each of the cheek teeth. As in Pliosaccomys, the transitional phase, in which the two columns were united at their mid-points, was eventually eliminated from the pattern of wear and only the U-pattern, that now appeared in the initial stages of wear, was retained. In the entoptychines of the early Miocene there is no suggestion of the H-pattern that characterizes [540] the Geomyinae, except in the position of the cusps before wear in the lower molars of Pleurolicus sulcifrons, which, according to Wood (1936:6), suggests the H-pattern. In earlier unknown Oligocene stages of evolution, the prisms possibly united first at their mid-points, and the columns may have joined at the side of the tooth only in the terminal stages of wear. The U-pattern of pre-Miocene entoptychines, therefore, may have become the dominant occlusal pattern only in the later stages of phyletic development.
According to the recently expressed views of several paleontologists, the Entoptychinae constitute the primitive lineage of the family and the early Geomyinae constitute a specialized offshoot of the entoptychine ancestral assemblage. The structure of the Entoptychinae, especially of the less advanced genera, closely approximates that of the hypothetical primitive morphotype. But, according to my view, the subfamily Geomyinae constitutes the ancestral assemblage and its structure is essentially that of the primitive morphotype of the family. At any rate the structure of the early geomyines more closely approximates the structure of the ancestral stock than the more divergent entoptychines. Therefore, the genus Dikkomys of the early Miocene, the first known geomyine, is considered to be a generalized geomyid, and, although it is a contemporary of the more specialized entoptychid assemblage, is considered to be more closely allied to the ancestral stock.
The entoptychines were the dominant and most highly differentiated geomyids of the early and middle Miocene. Nevertheless, they became extinct in the middle Miocene, and the geomyines of that time survived and later gave rise to the modern pocket gophers. Therefore, the early history of the family Geomyidae is characterized by an early radiation and trend toward specialization, followed by survival of the less specialized Geomyinae and extinction of the more specialized Entoptychinae.
The most abundant geomyids of the early and middle Miocene, the Entoptychinae, consisted of at least 24 species (see Wood, 1936:4-25) classified in four genera: Pleurolicus, Gregorymys, Grangerimus, and Entoptychus. The genera were essentially contemporaneous (see Figure 3). Even so, the subfamily was morphologically varied, pointing to an earlier origin in the Oligocene (actually a part of the John Day Fauna, including Pleurolicus may be correlated with late Oligocene Whitneyian age) followed by a relatively rapid radiation including all four genera in the early [541] Miocene. Two genera, Pleurolicus and Gregorymys, continued into the Middle Miocene (Hemingfordian). This divergence, specialization, and subsequent radiation suggest that the entoptychines evolved into a new major adaptive zone, in the sense described by Simpson (1945:199-206).
The radiation is correlated geographically and temporally with the southward retreat of the Neotropical flora of the Tertiary from the western United States and southward movement of the Arctic flora of the Tertiary (see Axlerod, 1950; Berry, 1937:31-46; Chaney, 1947:139-148; and Kendeigh, 1961:280-283). In the early Tertiary the Neotropical-tertiary geoflora occurred northward to at least 49° latitude in western North America, and the boreal Arctic-tertiary flora was restricted to a circumpolar zone. The southward and eastward shift of the Neotropical-tertiary flora, associated with the drying and chilling of the continent, began in the middle or late Oligocene and was concurrent with the divergence and radiation of the Entoptychinae. Beginning in late Oligocene and continuing at least into middle Miocene, most of the region in which the entoptychines occurred was occupied by the Arcto-tertiary geoflora of which the temperate forest division contributed the dominate plant associations. The maples, chestnuts, dogwoods, beeches, walnuts, oaks, elms, birches, and sycamores of that flora were the forerunners of today's eastern deciduous forest. It is my view that the entoptychines became adapted to the conditions of this paleoecological environment and radiated rapidly in the Arikareean when the major change occurred in climax vegetation. The ancestral stock of the Geomyinae was not so successful in the Arcto-tertiary climax, and most of it probably was displaced southward along with the tropical flora.
The skeleton in the entoptychines is not so strongly fossorial as in the modern geomyids (Wilson, 1949:117), and these early geomyids probably were semi-fossorial with somewhat the same burrowing habits as those of the living mountain beaver (Aplodontia). Inasmuch as the morphology and taxonomy of the entoptychines were discussed in detail by Cope (1884) and reviewed later by Wood (loc. cit.), there is no need to recount the details here. According to Wood (op. cit., 27-28), Pleurolicus occupied a central position in the entoptychid radiation and perhaps appeared slightly earlier than the other genera. Wilson (1949) suggested that the lower part of the John Day may actually be Upper Oligocene rather than Lower Miocene, and this arrangement is followed here. Also, Pleurolicus is less specialized than the [542] other genera and occurs in deposits of both the Great Plains and the Pacific Coast. Gregorymys, also little specialized, occurred only on the Great Plains. The more specialized genera, Grangerimus and Entoptycus, evidently appeared somewhat later than Pleurolicus and evolved from it. Except for a record from southern Texas reported recently by Hibbard and Wilson (1950:621-623) and the new species described by MacDonald (1963:182) from the Sharps Formation of South Dakota (early Arikareean), Grangerimus is known only from the Pacific coast. Entoptycus was restricted to the Pacific Coast (John Day fauna).
Entoptycus is the most specialized of the known genera; it has pronounced fossorial adaptations, especially in the skull. Its molariform teeth are rootless and ever-growing as in the modern geomyines. Moreover, the continuous enamel bands on only moderately worn teeth become separated in the final stages of wear into anterior and posterior enamel plates by tracts of dentine that extend toward the crown on the sides of each tooth. This extension was made possible by the union of the two columns at both the lingual and labial margins of the tooth forming an O-pattern, and the crown is essentially monoprismatic save for the isolated enamel fossette in the center of the tooth. The fossette is all that remains of the lateral re-entrant fold that characterized the preceding U-pattern of the earlier stages of wear. Late in the sequence of wear, the anterior enamel plate is lost in the lower molars and the posterior plate in the upper molars. The U-pattern characterizes the final stages of attrition in the other genera of the Entoptychinae; none developed the dental specializations seen in Entoptycus. Rootless, ever-growing cheek teeth, discontinuous enamel patterns, and monoprismatic molars were not evolved in the subfamily Geomyinae until the late Pliocene.
Phyletic Trends in Subfamily Geomyinae
The subfamily Geomyinae is made up of three groups, recognized taxonomically for the first time in this account as tribes—Dikkomyini, Thomomyini, and Geomyini (for full discussion of classification, see previous account). The phylogeny proposed by me is illustrated in Figure 3. The tribe Dikkomyini is characterized by generalized and primitive features that together form the basic structural foundation of the subfamily. Evolution within the Dikkomyini resulted in the acquisition and perfection of fossorial adaptations. The Thomomyini and Geomyini are considerably more specialized than the ancestral Dikkomyini from which they evolved. [543] The Geomyini are clearly more specialized than the Thomomyini, suggesting closer affinity between the Thomomyini and the Dikkomyini than between the Geomyini and the Dikkomyini. The specializations in the dentition and the associated changes in the skull of the Thomomyini and Geomyini permit more efficient mastication of fibrous vegetation. Along with these specializations, fossorial adaptations inherited from the Dikkomyini are retained without noteworthy modification.
Dikkomys, the earliest known genus of the tribe Dikkomyini, can be taken as a starting point of evolution for the subfamily Geomyinae. The Pliocene genus Pliosaccomys is the only other known geomyine having primitive features closely resembling those of Dikkomys. The relatively close but previously unrecognized relationship between Dikkomys and Pliosaccomys can be understood when patterns of wear on the occlusal surfaces of the cheek teeth are taken into account. It appears that Pliosaccomys descended from Dikkomys-like stock, if not Dikkomys itself. Although Dikkomys is towards the beginning of this phyletic sequence and Pliosaccomys towards the end of the sequence, the primitive features shared by the two provide a generalized morphotype for the subfamily Geomyinae.
In the molariform dentition, an almost complete series of stages of wear in Pliosaccomys has been preserved, and those of Dikkomys can be reconstructed with reasonable accuracy from those that are known (see Fig. 4):
(1) In the initial stage of wear in Dikkomys the anterior and posterior columns are separated by an intervening valley (Fig. 4A), and the occlusal surface of each column bears a loph of dentine surrounded by a ring of enamel: protoloph on the anterior column and metaloph on the posterior column of the upper teeth (protolophid and hypolophid in corresponding positions in the lower teeth). Actually this stage is not preserved in the known material of Dikkomys, but does occur in both geomyines and entoptychines in all stages of evolution, and it must have also occurred in Dikkomys in order for the next two stages, which are preserved, to have developed.
(2) The occlusal surfaces are ground down to a level where the enamel loops of the two columns join at their mid-points, thus forming an H-shaped pattern (Fig. 4B), or more exactly a pattern resembling a figure 8. Probably this was the primitive pattern in the final stage of wear in the geomyid ancestor of the Oligocene.
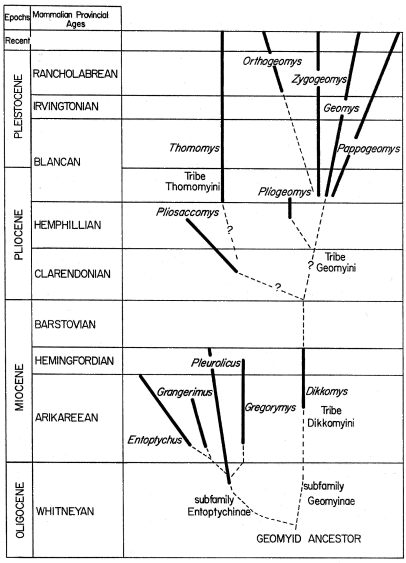
Fig. 3. Diagram depicting geologic range and probable phyletic relationships of the family Geomyidae. Dashed lines represent parts of lineages that are not represented by fossil records, and solid lines represent parts of lineages verified by actual specimens. Question marks indicate uncertainty of suggested ancestry of known taxa. The relationships within the subfamily Entoptychinae are modified after Wood (1936), and the temporal range of the Miocene geomyids have been adjusted to agree with current stratigraphic correlations. Hence, Pleurolicus, Gregorymys and Dikkomys are illustrated as ranging into the Hemingfordian, rather than being confined to the Arikareean (see MacDonald, 1963, and Black, 1961).
(3) In the pre-final stage of wear, the anterior and posterior lophs of the first and second molars unite secondarily at the edge of their protomeres (labial side in the lower and lingual in the upper), thus enclosing an isolated enamel fossette (Fig. 4C). Lateral union occurs in the lower teeth because the vertical depth of the labial re-entrant angle is less than the depth of the lingual re-entrant fold. In the upper teeth the reverse is true. The re-entrant angle on one side of the premolar is as deep vertically as the angle on the other side of that tooth, and both reach the base of the crown; therefore, they do not disappear at any stage of attrition. The same pertains in the third lower molar.
(4) In the final stage of wear (Fig. 4D), the enamel fossette disappears as a result of continued attrition on the occlusal surface in the upper series. The fossette may vary somewhat in vertical depth in m1 and m2, but the amount of wear required for its effacement would be greater than in the upper teeth. Therefore, upon wear, the U-pattern would become characteristic of the final stage in M1 (and probably also M2), but the modified H-pattern described in Fig. 4C would prevail in m1 and m2. Perhaps, in extremely worn teeth, the labial fossette of m1 and m2 would disappear. If this advanced stage of effacement is obtained, then the two columns would be united across the entire surface of their protomeres from the center of the crown to its labial edge, and the occlusal pattern would be in the shape of a U.
The occlusal pattern, at least in M1 and M2, in the final stages of wear in Dikkomys resembles that in the subfamily Entoptychinae, but the U-pattern develops on only the first and probably the second molar in Dikkomys and not on all of the cheek teeth as it does in the entoptychines. Judging from the material that has been described, the U-pattern did not develop in the lower teeth of Dikkomys until the Hemingfordian (D. woodi), upper Rosebud, and specimens of D. matthewi from the earlier Arikareean, lower Harrison, suggest that the modified H-pattern, with secondary coalescence at the edge of the protomeres, persisted throughout life, without developing the U-pattern in the final stages of wear.
Essentially the same patterns of wear characterize the genus Pliosaccomys, except that the earlier stages were telescoped and the second stage was omitted while another (final) stage was added. The stages are reconstructed in sequence in figure 4, and all are based on preserved dentitions, as follows:
(1) The first phases of wear produced the pattern (p. 4E and I) described for Dikkomys in the previous account (Fig 4A).
(2) A small additional amount of wear produced the 2nd stage (Fig. 4F and J) characterized by a U-pattern, formed by union of the anterior and posterior columns at the edge of the protomeres of the first and second molars, both above and below, without first forming an H-shaped pattern. Union at the mid-points thus was omitted from the sequence of wear in these two teeth. In the premolars and third molars the primitive H-pattern did form, as in Dikkomys. The pattern of wear in the first two molars is the same as in the entoptychines of the early Miocene. The trend of evolution through which the Pliosaccomys lineage passed must have featured a progressively earlier union at the edge of the tooth until the lateral coalescence occurred simultaneously with the median union. At that stage, emphasis was shifted to the union at the edge of the tooth, and eventually the teeth failed to unite at their mid-points and the U-pattern developed directly. Therefore, the horizontally deep re-entrant fold that separates the two lophs of the U-pattern is equivalent to one fold plus the apex of the opposite fold.
(3) The horizontal re-entrant fold of the U-pattern was remarkably shallow vertically and disappeared with little additional wear. Thus the two parts of M1, and also of M2, are united into a single column except for a slight inflection on the labial side and this is true also of m1 and m2 except for a slight inflection on the lingual side (Fig. 4G and K). The inflection appears to have persisted in the upper teeth (Fig. 4H), but evidently with slight wear, disappeared in the lower teeth (Fig. 4L). The final monocolumnar pattern was attained early ontogenetically, evidently before the permanent premolar had fully erupted; hence, the earlier stages occurred only in transition, persisted for only a brief interval in the teeth of juveniles, and the final stage developed in the young animal and lasted throughout the rest of its life in Pliosaccomys. In Dikkomys the two columns never united into a single column, and a bilophodont occlusal pattern persisted throughout life.
The early phyletic development of the subfamily Geomyinae took place in the tribe Dikkomyini from the early Miocene into the early Pliocene. Compared with the rapid evolution of the specializations that distinguish the Entoptychinae, the structural changes in the early Geomyinae occurred at a remarkably slow rate. In fact the lineage changed but little from Dikkomys to Pliosaccomys, in parts of the animal that can be compared, as illustrated by the low-crowned and rooted cheek teeth, the continuous enamel bands, the lack of grooving of the upper incisor, the retention [547] of the primitive H-pattern, both above and below, in the premolar and third lower molar, and the ridges and fossae of the mandible to which the muscles of mastication attach. The only major changes detected in the known fragments are in the pattern of wear and the final configuration of the first and second molars, as described above. The unification of the two lophs in each of these two teeth into a single column was a significant step in the evolution of the Geomyinae, and is a stage between the primitive bilophodont pattern of the early and middle Miocene geomyines having continuously bicolumnar teeth and the monolophodont pattern in the modern pocket gophers of both lineages in which these teeth consist of a single column in all but the initial stages of wear. The monocolumnar structure of the first and second molars in the final stages of wear, therefore, is closer to that in the lineage of Thomomys than it is to that of Dikkomys. Other specializations in the dentition of Pliosaccomys, especially in m1 and m2 where the H-pattern has been completely eliminated from the sequence of wear, are too far advanced for Pliosaccomys to have given rise to the tribe Geomyini. The teeth in the immediate ancestor of the Geomyini must have been less specialized in m1 and m2, perhaps about as in Dikkomys. In the m1 and m2 of the tribe Geomyini, the H-pattern is formed in the initial stages of wear; therefore, in the early Pliocene ancestor, presently unknown in the fossil record, the H-pattern probably was present. Even so, the ancestor of the Geomyini and that of Pliosaccomys probably were closely allied otherwise, and both probably had attained the highly specialized fossorial adaptations characterizing all modern pocket gophers, before the divergence of Pliosaccomys and the Geomyini took place.
The evidence points to a major divergence of the geomyines that lived in the latest Miocene or the early Pliocene (probably the latter) and that gave rise to the two modern lineages, Thomomyini and Geomyini (see Fig. 3). One, the most primitive of the two, gave rise to the Thomomyini lineage that eventually evolved into Thomomys. Pliosaccomys is closely allied to the ancestry of this lineage, although it is probably not the actual ancestor, as mentioned previously. Aside from the aforementioned specializations of the first and second molars, the features of the Thomomyini are less advanced than in the other specialized lineage (tribe Geomyini). Primitive traits retained in the tribe Thomomyini (and also characteristic of the ancestral tribe Dikkomyini) are: (1) Small size, in general no larger than the ancestral morphotype; (2) lack of grooving on the upper incisor (although a slight [548] rudimentary groove is developed rarely in some living species); (3) retention of anterior and posterior enamel plates in lower and upper cheek teeth; (4) premolars having widely open re-entrant folds; (5) smooth and generalized skull lacking marked angularity, regosity or cresting (neither the sagittal nor the lambdoidal crest are ordinarily well developed except in Thomomys bulbivorus); (6) forefoot small, less modified for digging than in the Geomyini.
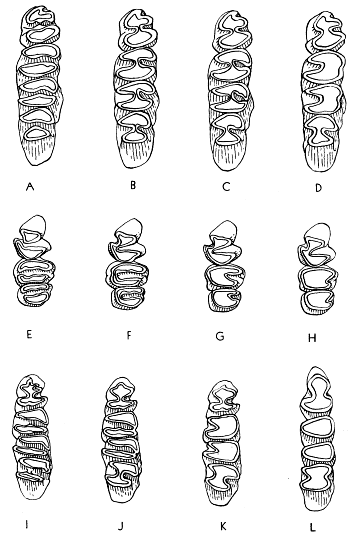
Fig. 4. Drawings of the molariform dentitions of Dikkomys and Pliosaccomys (Tribe Dikkomyini) depicting the patterns of wear on the occlusal surfaces. Ontogenetically, the stages of wear are arranged from left to right in each row. Stages not represented by actual specimens have been carefully reconstructed from information provided by known stages in the sequence of wear and the dentitions of other geomyines. × 5.
A-D. Dikkomys woodi, right lower tooth-row, including p4-m3. Patterns based on No. P26284 (FMNH) from Upper Rosebud (Middle Miocene), Shannon Co., South Dakota (B above).
E-H. Pliosaccomys dubius, left upper tooth-row, including P4-M2 (M3 unknown). Patterns based on Nos. 1798 and 1799 (LAM) from Smiths Valley (Middle Pliocene), Lyon Co., Nevada.
I-L. Pliosaccomys dubius, right lower tooth-row, including p4-m3. Patterns based on Nos. 1796 (holotype), 1804, and 1806 (LAM) from Smiths Valley (Middle Pliocene), Lyon Co., Nevada.
The lineage of the Thomomyini is essentially rectilinear and without the major branching seen in the tribe Geomyini. The one genus, Thomomys, appears first in the Upper Pliocene (early Blancan time), and the specializations characterizing the lineage had already developed by that time. Evidently, the early stages of divergence from the ancestral stock resulted in the development of rootless, ever-growing, more hypsodont cheek teeth, simplification of M3, and enlargement of the masseteric ridge on the mandible. The enamel investment on the sides of the molariform teeth is interrupted owing to intrusion of tracts of dentine on the sides of each column. Even so, complete anterior and posterior plates are retained on all of the cheek teeth (Fig. 5, K and L) and there is no trend toward additional loss of enamel as in the Geomyini. The enamel on the sides of the column has little functional value, and its elimination probably reduces friction during the anteroposterior movements of the lower jaw, thereby increasing the efficiency of the cutting blades on the anterior and posterior wall of the tooth. The simplification of M3 was achieved by union of the two columns of the primitive pattern into a single column and obliteration of both the labial and lingual re-entrant folds in the first stages of wear. The adult tooth (see Fig. 5L) is without trace of the bilophate pattern and is not elongated; therefore, its structure is essentially the same as that of the first and second upper molars.
In the Thomomyini, the two lophs of the unworn molars unite entirely across the width of their surfaces with the first traces of wear (see Fig. 5, I and J), owing to the shallow and uniform depth of the transverse valley. In the molars, the final pattern is acquired, therefore, before the deciduous premolar has been replaced by the permanent tooth. A relatively shallow re-entrant inflection between the ends of the parameres sometimes is retained, although it also will disappear with slight additional wear. Therefore, both lophs tend to unite completely with the first stages of wear in the Thomomyini, thus omitting both U and H patterns from the sequence of wear. This is the highest degree of specialization attained in the Geomyidae in regard to the patterns of wear, since a sequence of [550] bilophodont patterns appear in both the Dikkomyini and Geomyini before the monoprismatic pattern is developed.
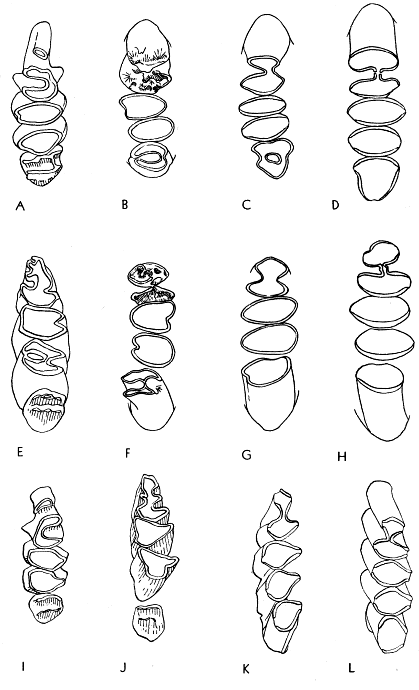
Fig. 5. Drawings of molariform dentitions representative of the tribes Geomyini and Thomomyini depicting patterns of wear on the occlusal surface. A-D represent, in ontogenetic sequence from left to right, upper tooth-rows of the tribe Geomyini. E-H represent, in the same sequence of stages, lower tooth-rows of the tribe Geomyini. I-L represents both upper and lower tooth-rows of both pre-final and final stages of wear in the tribe Thomomyini. All × 5.
A and E. Geomys bursarius majusculus, No. 2948 (KU), Douglas Co., Kansas. Right upper (A) including DP4-M3; lower left (E) including dp4-m3.
B and F. Pappogeomys bulleri burti, No. 100444 (KU), 10 mi. NNW Barra de Navidad, Jalisco. Right upper (B) including P4-M3; right lower (F) including p4-m3 (both P4 and p4 with unworn enamel caps).
C and G. Pappogeomys bulleri albinasus, No. 31044 (KU), 10 mi. S and 8 mi. W Guadalajara, Jalisco. Right upper (C) including P4-M3; right lower (G) including p4-m3.
D and H. Pappogeomys bulleri albinasus, No. 31002 (KU), W side La Venta, 13 mi. W and 4 mi. N Guadalajara, Jalisco. Right upper (D) including P4-M3; right lower (H) including p4-m3.
I and J. Thomomys talpoides bridgeri, No. 6865 (KU), 2 mi. up Mink Creek, Pocatella, Bannock Co., Idaho. Left upper (I), DP4-M3; left lower (J), dp4-m3.
K and L. Thomomys talpoides fossor, No. 13205 (KU), Wasson Ranch, 3 mi. E Creede, Mineral Co., Colorado. Right lower (K), p4-m3; left upper (L), P4-M3.
Relationship of the Geomyini with the ancestral Dikkomyini is most clearly demonstrated in the sequence of wear on the occlusal surfaces of the molars. As in all geomyids, the upper part of the [551] crown is biprismatic in the newly erupted tooth, and the two columns are separated by an intervening valley. With slight attrition on the unworn enamel cap, the weakly developed cusps merge and form a transverse enamel loop on each of the two columns (see third molar in Fig. 5, A and E), each loop enclosing a core of dentine that had become exposed. The valley between the two columns is shallow, and upon further wear of the tooth, the two loops unite. The two columns become joined at different points in the upper and lower molars depending on the varying depth of the valley in different teeth. Therefore, upper and lower molars develop distinctly different occlusal configurations.
In the lower molars, the pattern characteristic of Dikkomys (Fig. 4C) is preserved without significant modification, as illustrated in an immature specimen of Geomys (see Fig. 5E). The H-pattern and modified H-pattern are developed in the same sequence of wear in the Geomyini. A juvenal female (not illustrated), KU 2931, provides an example of the intermediate H-pattern. In this specimen, the protolophid and hypolophid of the left m2 are united only at their mid-points, indicating that the pattern of wear occurs in the same sequence in the Geomyini as it did in the Miocene genus Dikkomys. After the two columns have become united at their mid-points, a secondary union is formed at the edge of their [552] protomeres, thus enclosing the enamel fossette as illustrated in Figure 5E (this is the modified H-pattern mentioned above). However, the fossette itself is shallow and soon disappears with slight wear. At this stage, the occlusal configuration would be in a U-pattern (m1 in Fig. 5E). The lingual re-entrant fold is also shallow in vertical depth; therefore, it is obliterated by wear following the eradication of the labial fossette. Consequently, the two columns are united into one. In m3 (see Figs. 5E, F, and G), the two columns merge by progressive lateral expansion of the medial isthmus.
In the first and second upper molars, the two columns unite across the entire surface of their protomeres from near the lingual edge of the crown to near its center. A minute inner inflection may be temporarily retained in some teeth. At this stage (see Fig. 5B), the parameres are still separated by the labial fissure, and the occlusal pattern is in the shape of a U, resembling, but not exactly duplicating, the pre-final pattern of Ml and M2 in the genus Pliosaccomys (see Fig. 4H). The labial fissure is shallow, and, with further wear, the inflection is worn away and the parameres also unite, thereby forming a monoprimatic crown in the final stage. In M3, the two lophs first become united near the edge of their protomeres (see Fig. 5B), therefore forming a U-pattern similar to that developed in Ml and M2 of Pliosaccomys. The connection of the two lophs is not directly at the end of the protomere; consequently a shallow lingual inflection remains. The lingual edge of the valley is also shallow, and, with continued wear a second union of the two lophs takes place near the ends of their parameres, and the deeper, interior part of the valley remains as an isolated enamel fossette (see Fig. 5C). The two primary lophs of the tooth are now joined near both sides, having shallow lingual and labial re-entrant angles on the sides and the enamel island in the center. With continued effacement of the occlusal surface, the fossette will be eradicated, and the pattern of the occlusal surface will become the partially biprismatic pattern of the final stages (adult) of wear (see Fig. 5D). M3's of Dikkomys and Pliosaccomys are not known; however, it seems reasonable to assume that the pattern of wear in the M3 of Dikkomyini was not essentially different from that of the Geomyini, except that it is likely that the U-pattern of the second stage of wear in the Geomyini was probably the final stage in the genus Dikkomys.
Judging from the pre-final stages of wear, the dentition of the [553] Geomyini provides a curious combination of patterns that resemble in part the Miocene genus Dikkomys and in part the early and middle Pliocene genus Pliosaccomys. There is no significant variation in the premolars or third molars (at least in the lower teeth) of the Geomyinae from the early Miocene to late Pliocene; therefore, deviations of major significance are in the character of the first and second molars. In the Geomyini, the patterns of wear of m1 and m2 are the same as those of Dikkomys, and are distinctly different from those of Pliosaccomys where the two columns first unite at the edge of their protomeres to form a U-pattern, rather than at their mid-points to form an H-pattern. Even though the intermediate stages of ontogeny in m1 and m2 of Pliosaccomys and the Geomyini are entirely different, the bicolumnar crowns of both eventually unite, upon wear, into a single column. On the other hand, the patterns of M1 and M2 in the Geomyini most closely resemble those of Pliosaccomys, rather than Dikkomys. In this regard it should be pointed out that the upper molars of Dikkomys are presently represented by only one tooth, an M1 in an early stage of wear. As described already, the patterns of M1-2 evidently would be mirror images of m1-2 in corresponding stages of wear. However, the initial union of the two columns, in the M1 that is known, is somewhat to the lingual side of center and the relatively small lingual valley does not reach the base of the crown, indicating, that eventually with wear, the two columns of Dikkomys might have become united across the entire surface of their protomeres as in Pliosaccomys. Even so, the two columns of M1 do initially join closer to their mid-points than they do in Pliosaccomys, and, if they did actually unite across their protomeres, the union would have occurred with subsequent wear. That is, the first occlusal pattern would be H-shaped (but with the connection closer to the lingual than the labial side), as in m1 and m2, and it would become U-shaped only after additional wear. This sequence of patterns of M1 and M2, as already pointed out, does not pertain in Pliosaccomys or the Geomyini, since the U-pattern is formed with the first union of the two columns at the edge of their protomeres, and the primitive H-pattern is never developed, unless one counts the slight lingual inflection, that occasionally is formed just after the two columns unite, as being indicative of the primitive pattern. As in the lower teeth, the bicolumnar crowns of early ontogeny in both Pliosaccomys and the Geomyini become eventually united, with wear, into a single column.
Based upon the foregoing evidence, it would seem likely that the Geomyini evolved from an early Pliocene (perhaps late Miocene) Dikkomyini ancestor that had evolved the specializations of M1 and M2 that characterize its relative, Pliosaccomys, but had not also evolved the specializations of m1 and m2 that distinguish Pliosaccomys. Therefore, the ancestor of the Geomyini differed from the Pliosaccomys-Thomomyini lineage in its retention, unmodified, of the primitive patterns in m1 and m2 that characterized the earliest known Geomyines (Dikkomys). The same patterns are preserved in m1 and m2 of its modern descendents, the living Geomyini. In the Pliosaccomys-Thomomyini lineage the pattern of m1 and m2 are entirely different, as described above.
The earliest record of the Geomyini is the extinct genus Pliogeomys (see Fig. 6) in the latest Hemphillian (middle Pliocene) and earliest Blancan (late Pliocene). Pliogeomys is more primitive than any modern genus of the Geomyini, seems to have been a late survivor of the primitive stock, but was itself probably a collateral lineage and not on the direct line of descent. The cheek teeth in Pliogeomys are rooted and less hypsodont than in the late Pliocene examples of the modern genera, and the anterior enamel plate of the lower molars shows no indication of reduction, as would be expected if Pliogeomys were in the direct line of evolution. Separation of Pliogeomys from the main stem of the Geomyini probably occurred after several specializations had already been achieved by the Geomyini. Two inheritances might have been grooving on the upper incisors and some reduction in amount of enamel on the sides of the cheek teeth. The dentine tracts on the sides of the cheek teeth of Pliogeomys are narrow (see Fig. 7A) and barely separate the enamel blades and there is no discernible reduction in the anterior enamel blades on its lower molars. Those blades evidently were lost in the main lineage before the Pleistocene radiation of the living genera took place. Pliogeomys is in an intermediate stage in evolution, and was not so advanced as was the main lineage at the time Pliogeomys died out. Its structure does provide clues as to phyletic development that took place in the main lineage.
Specialized trends in the early phylogeny of the Geomyini included: development of rootless, ever-growing cheek teeth and an increase in hypsodonty; loss of the bicolumnar structure of the first and second molars, and, consequently, the formation of a single elliptical column in the final stage of wear; interruption of the enamel investment of the molariform teeth and formation of anterior [555] and posterior enamel plates; and enlargement of the masseteric ridge and fossa. Each of these trends occurred independently in the Thomomyini, and each is an example of parallelism in the phyletic evolution of the two lineages. Three additional specializations lacking in the Thomomyini are the grooving on upper incisors, loss of anterior enamel plate in lower molars, and development of a basitemporal fossa on the mandible. Evidently, two grooves evolved in the ancestral incisors in the same bisculcate pattern preserved in Pliogeomys, Zygogeomys and Geomys. The innermost groove is weakly developed in Pliogeomys, suggesting that this character was in an intermediate stage of evolution in the ancestral lineage at the time that Pliogeomys split off. Numerous other specializations in the Geomyini appeared later, but evolved in the different genera that diverged from the ancestral lineage and are discussed separately in the next account. Only two of the major features characterizing the Dikkomyini are retained in the Geomyini: the H-pattern on the occlusal surface of the m1 and m2 developed during the initial stages of wear, and the bicolumnar pattern of M3. Adaptive radiation produced the living genera of the Geomyini in the late Pliocene and early Pleistocene (see Fig. 6) and subsequent specialization of the ancestral morphology followed.
Parallelism in the molars of later geomyines and the Entoptychinae is illustrated by the lateral interruption of the enamel investment and loss of enamel plates and by the omission of the H-pattern stage in the first and second molars (in Pliosaccomys). Resemblance of dentitions in certain stages of wear in Pliosaccomys and in entoptychines led some investigators, for instance, Hibbard (1953:357), to suggest that Pliosaccomys descended from one of the less specialized entoptychines, possibly Grangerimus but probably Gregorymys. Actually, the highly specialized upper and lower premolars and third molars of the entoptychines rule them out as ancestors of the later geomyines. The evolution of entoptychine-like features in Pliosaccomys is regarded as an example of iteration, a pattern of parallelism (see Simpson, 1953:248-253) where an allochronic and independent lineage undergoes the same evolutionary trend that phyletically characterized an earlier lineage, usually after the latter has become extinct. In this case, the lineage giving rise to Pliosaccomys passed through the same phyletic stages in its evolution in the early Pliocene (and possibly the late Miocene) as did the entoptychines in the late Oligocene and early Miocene.
Another parallelism by iteration, occurring in the middle and late Pliocene in both the Thomomyini and Geomyini, is the loss of enamel from the lateral surfaces of the cheek teeth, and, in the Geomyini only, the eventual loss of the anterior plate in the lower teeth and the posterior plate in the upper teeth. Both features were evolved more than an epoch earlier in the specialized entoptychid genus Entoptychus of the lower Miocene. In Entoptychus, only the posterior plate of the lower molars and the anterior plate of the upper molars remained in the final stages of attrition, although a central enamel fossette, a remnant of the re-entrant fold, remained throughout life. Iteration is also expressed in the subfamily Geomyinae by the development of grooving on the upper incisor and the formation of the basitemporal fossa. A shallow but distinct basitemporal fossa occurs between the coronoid process and the third lower molar in the genus Entoptychus and a sulcated upper incisor, a single shallow groove usually near the median border of the tooth, is found in the genus Gregorymys of the subfamily Entoptychinae. Both features are regarded as advanced specializations in the tribe Geomyini, even though each was evolved in the entoptychines of the Lower Miocene.
The postcranial skeleton of living genera of pocket gophers, as befits animals that spend most of their life within underground burrows, are highly specialized for a fossorial life. Elements of the postcranial skeleton recovered from Lower Miocene deposits indicate that the entoptychines were only semi-fossorial (see Cope, 1884:857; Wood, 1936:4-5; Wilson, 1949:117-118). One of the basic trends of the entoptychines was towards greater fossorial adaptation; the skeleton of Entoptychus shows a greater degree of fossorial adaptation than earlier genera of the subfamily. There is no reason to suppose that the geomyine genus Dikkomys, which lived at the same times as the entoptychines, had acquired any more advanced fossorial adaptations than had the entoptychines.
The most pronounced fossorial adaptations seem to have evolved only in the ancestral lineage of the modern geomyines, probably in the latter part of the Miocene and in the early Pliocene, before the modern Thomomyini and Geomyini diverged. Extreme fossorial adaptations in herbivorous rodents, such as those characteristic of the modern pocket gophers and their immediate ancestors, are thought to have evolved only in response to pronounced arid conditions. The Entoptychinae and evidently the early geomyines lived in environments that were either tropical or temperate, and under conditions more mesic than I would consider necessary to bring [557] about selection pressure resulting in fossorial specializations. In late Oligocene and early Miocene, according to Axelroad (1958:433-509), arid conditions did not exist in the United States, and the only xerophytic environments in North America occurred on the Central Plateau of México. Moreover (Axelroad, loc. cit.), arid conditions did not develop in the western United States until the early Pliocene. Geomyids evidently became extinct in this region at the close of the Middle Miocene, and none appear in fossil deposits in the western United States until the latest Lower Pliocene (Clarendonian). The reappearance of geomyids, Pliosaccomys, in the western United States coincides with a trend toward aridity and the northward movement of the Madro-tertiary geoflora into the Great Basin and Great Plains from its place of origin on the Central Plateau of México (Axelroad, loc. cit.). Later, in the middle and later Pliocene, the Madro-tertiary geoflora gave rise to the modern xerophytic plants that now characterize the desert vegetation of North America.
The Madro-tertiary climax does not appear as a major flora until the Miocene, but probably originated earlier. According to Axelroad (loc. cit.), this xerophytic flora evolved from elements of the Neotropical-tertiary geoflora that became adapted to arid conditions that developed in the rain shadow of the high mountains flanking the Central Plateau of México. Originally, the Madro-tertiary flora consisted of small trees, shrubs, and grasses. Although some elements of this flora moved northward in the late Miocene, the major part of it remained in México until the early Pliocene. In the western United States, mountain formation increased in intensity in the Pliocene and continued on into the early Pleistocene. As the mountains became more elevated, especially the Sierra Nevada and Cascade ranges, they blocked the prevailing winds from the Pacific Ocean and extensive aridity developed on their leeward side. As xeric conditions became widespread, the Madro-tertiary flora successfully occupied the drier regions of southern California, the Great Basin, and the western parts of the Great Plains.
While the Entoptychinae probably evolved in response to the Arcto-tertiary flora, the late Tertiary geomyines probably evolved in response to the Madro-tertiary geoflora on the Central Plateau of México. Some of these early geomyines, especially ancestors of the modern lineages, probably were pushed southward by competition with the more specialized entoptychines. Most geomyines were pushed out of the northern area of distribution, except for Dikkomys that survived in association with the entoptychids throughout the [558] early and middle Miocene. During this time, and probably continuing on into the late Miocene, the geomyines occurring to the south in México became adapted to the arid environments of the Madro-tertiary geoflora.
Of course, information is lacking about climates in several parts of the late Miocene and early Pliocene. When such information becomes available it conceivably could modify the hypothesis outlined immediately above.
The principal trend of evolution in these semi-fossorial rodents was toward more complete fossorial adaptation, and the pronounced fossorial features characteristic of the modern pocket gophers were perfected. This trend continued in response to the intense selection pressures in this arid environment. The principal structural characters effected were in the postcranial anatomy, especially in the skeletal and muscular systems. Consequently, it is not surprising that in skull and dentition, Pliosaccomys differs but little from Dikkomys. Therefore, most of the basic structural specializations so far developed for subterranean existence probably had evolved by the time geomyines moved back north in the early Pliocene. Both modern lineages, the tribes Thomomyini and Geomyini, have essentially the same fossorial features, and it seems unlikely that these features were acquired independently in the relatively short period of time available to them after their divergence; probably they were inherited from a common ancestor. These probabilities indicate that the evolution of the fossorial specialization was in the later phyletic development of the tribe Dikkomyini.
Plio-Pleistocene radiation of Geomyini
Unlike the lineage of the Thomomyini that remained essentially rectilinear through out its history, the Geomyini in the late Pliocene and the early Pleistocene underwent adaptive radiation in a degree comparable to the earlier radiation of the Entoptychinae, and all of the later history of the tribe is dominated by the radiation—the resulting structural diversity. At least four lineages were produced by the Plio-Pleistocene radiation (see Fig. 6); each originated at essentially the same time (late Pliocene) presumably from the same ancestral stock. Each of these lineages within the Geomyini has given rise to one of the four modern genera: Zygogeomys, Geomys, Orthogeomys, and Pappogeomys.
The immediate, unknown, ancestor probably lived on the Central Plateau of México. After the radiation began the ancestors of Geomys and Zygogeomys extended their ranges northward.
Features of the hypothetical morphotype, that would permit derivation of the modern genera would include the following: (1) Skull generalized, neither excessively long and narrow or short and broad; (2) skull smoothly rounded, without pronounced angularity, rugosity or cresting (sagittal crest probably lacking, even in old individuals); (3) zygomata slender, without lateral platelike expansions; (4) rostrum moderately broad; (5) upper incisors bisulcate, two grooves in pattern found in Pliogeomys, Zygogeomys and Geomys; (6) lateral re-entrant angles of premolars obtuse; (7) p4 having four enamel plates (one on anterior wall, one on posterior wall, and two lateral plates) and lower molars having one enamel plate on the posterior wall of tooth (anterior plate is lacking); (8) P4 having four enamel plates, in same pattern as described for p4, M1 having two enamel plates (one anterior and one posterior), M2 same as M1, M3 having three plates (one anterior, two lateral on sides of posterior loph, none posterior); (9) M3 subtriangular in cross-section, distinctly bicolumnar, two columns marked by shallow re-entrant folds and connected by broad isthmus; (10) masseteric ridge large, forming high crest bordering masseteric fossa; (11) basitemporal fossa shallow; (12) angular process of mandible short, its lateral projection barely exceeding that of zygomatic arch.
In relation to the primitive morphotype, increase in size, simplification of dentition, and changes in shape of skull are regarded as specializations. Considerable parallelism between the four lineages is seen. But each lineage is distinguished by a combination of specialized features, and three by a few unique specializations.
Among trends resulting in simplification of the dentition, reduction of enamel on the posterior wall of the upper cheek teeth has occurred in various degrees in all lineages of the Geomyini even to loss of all enamel on the posterior wall of the premolars and molars in two genera. Loss of some enamel is more common on P4 than on M1-2, and has occurred in all genera (see Figs. 7 and 9.)
In evolutionary sequence loss of enamel from M1 and M2 usually occurs after, but never preceding, the reduction of enamel on P4. Loss of enamel plates from the posterior face of M1 and M2 is associated with the evolution of an efficient anterotransverse shearing action of the teeth.
On the anterior wall of those teeth no reduction of the cutting blade has been observed; a complete anterior plate is retained in all living Geomyini.
Presence of both the posterior and anterior plates decreases the efficiency of transverse shearing, by providing two upper plates (anterior plate of one tooth and posterior plate of the preceding tooth) over which the lower cutting blade simultaneously must pass with each movement. The advantages of shearing over the more common mechanics of planing are largely lost unless the posterior plates are eliminated. Also, none of the living Geomyini have retained a definitive posterior enamel plate on M3, the last upper molar; but two well-developed lateral plates, that extend almost all of the way back to the posterior apex of M3, have been retained, and, together function as a posterior plate. Loss of either or both of the lateral plates of M3 is rare, and occurs only in old individuals. Their loss in the final stages of wear may represent the beginning of a new trend in those species where it occurs (the castanops-group of the subgenus Cratogeomys). In any case, reduction of enamel takes place by transverse shortening of the plate through the complete loss of enamel on one end, the diminution beginning first on the labial end and proceeding by progressive atrophy to the lingual end of the plate. Evidently, when enamel has been eliminated from the labial end of a plate, the rate of loss decreases markedly, and the last stages of evolution, terminating in complete loss of an enamel plate, occurs more slowly. Evolution may be arrested before complete loss has occurred, and that part of the enamel that remains forms a short, vestigial plate restricted to the lingual one-fourth or one-third of the wall. The enamel pattern of the lower dentition is the same in all of the diverging lineages, with no evidence of additional loss of enamel from that which had already occurred in their common ancestor (see Figs. 7 and 9). Reduction and loss of enamel plates began and was terminated in the lower dentition before reduction began in the upper dentition.
Other dental specializations have occurred in the shape of the third upper molar and in the pattern of grooving in the upper incisor. Unlike M3 of the Thomomyini, that of the Geomyini differs in shape from M2, and its enamel investment differs from that of M2. Primitively, M3 was probably subtriangular in cross-section, and the posterior loph evidently projected posteriorly as a short, rudimentary heel that formed the apex of the triangle. Other shapes [562] of M3 are considered to be specializations that have been derived from the primitive form. In addition to the primitive subtriangular pattern, the M3 of living Geomyini may be suborbicular, quadriform, elongate, or obcordate in shape. Usually each lineage is [563] characterized by only one pattern, but in one genus (Pappogeomys) all patterns occur. Of the different forms, the elongate and obcordate seem to be the most highly specialized deviations from the triangular-shaped tooth. The bicolumnar pattern is accentuated in the elongate type (Fig. 7D, F, H) by deep lateral re-entrant folds, on both the lingual and labial sides, and by the elongation of the posterior loph into a pronounced heel. Teeth having this pattern have been illustrated by Merriam (1895:76-82) in Figures 27 (6 and 7), 28 (c and d), 34 (7 through 15), and 35 (8).
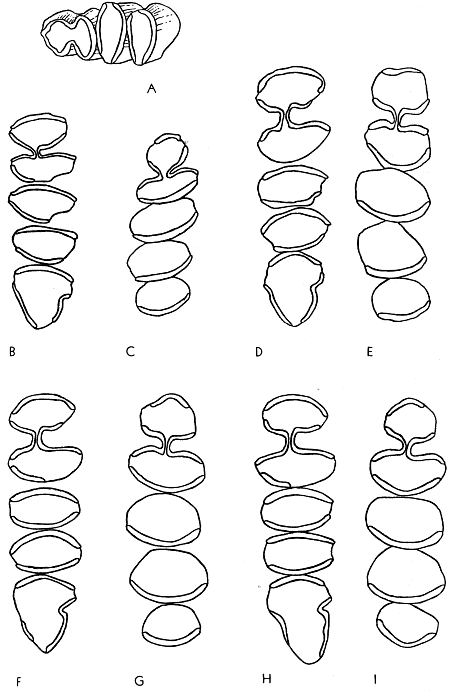
Fig. 7. Molariform dentitions of the Tribe Geomyini. Drawings illustrating enamel patterns characteristic of Pliogeomys, Zygogeomys, and the subgenera of Orthogeomys (Orthogeomys, Heterogeomys and Macrogeomys). × 5.
A. Pliogeomys buisi, No. 29157 (UMMP), holotype, Buis Ranch (Upper Middle Pliocene), Beaver Co., Oklahoma. Right lower, p4-m2 (m3 unknown).
B and C. Zygogeomys trichopus trichopus, adult female, No. 51971 (FMNH), Mt. Tancítaro, 10,500 ft., Michoacán. Left upper (B), P4-M3; right lower (C), p4-m3.
D and E. Subgenus Orthogeomys. Orthogeomys grandis guerrerensis, adult female, No. 39807 (KU), 1/2 mi. E La Mira, 300 ft., Michoacán. Left upper (D), P4-M3; right lower (E), p4-m3.
F and G. Subgenus Heterogeomys. Orthogeomys hispidus hispidus, adult female, No. 23975 (KU), 4 km. W Tlapacoyan, 700 ft., Veracruz. Left upper (F), P4-M3; right lower (G), p4-m3.
H and I. Subgenus Macrogeomys. Orthogeomys heterodus cartagoensis, adult female, No. 60664 (KU), Rancho Redando, Volcán Lrozá, Prov. San José, Costa Rica. Left upper (H), P4-M3; right lower (I), p4-m3.
The subcordate form is characterized by pronounced anteroposterior compression, and retention of a distinct labial re-entrant fold. The posterior loph apparently has been rotated in such a way that what was previously its posterior border now lies on the outer margin of the tooth; therefore, the axis of the posterior loph is strongly oblique in relation to the anteroposterior bearing of the maxillary tooth-row, and the median enamel plate also has been rotated and so lies transversely across the posterior wall of the tooth. Owing to the rotation of the posterior loph, the apex of the obcordate tooth is at its lingual side. The subcordate type is illustrated by Merriam (loc. cit.) in Figures 27 (3 and 4), 28 (a and b), 34 (3 and 4), and 35 (5, 6, and 7). The suborbicular and quadriform types are less specialized than the two described above. Both are characterized by reduction, often obliteration, of the bicolumnar pattern of the subtriangular ancestral form, especially marked by the decrease in depth of the lateral re-entrant folds and the decrease in length of the posterior projection of the posterior loph. With these [564] changes, the tooth becomes essentially monocolumnar, its occlusal surface oval in outline in one and squarish in shape in the other. Occlusal views of the suborbicular form are presented by Merriam (loc. cit.) in Figure 33 (1, 5, 6, 7, 11, and 12) and the quadriform tooth is depicted in Figure 29. Grooved upper incisors are characteristic of the living Geomyini, but variation occurs in the number of grooves, and, if only one groove is present, its position on the anterior face of the tooth varies. Except for the previously mentioned (p. 480) abnormal tooth having three grooves, incisors with no more than two grooves are found in these pocket gophers, and this number of grooves is taken to be primitive. Loss of one or the other of the two grooves of the bisulcate pattern, therefore, is regarded as specialization. However, complete loss of both grooves never occurs in the Geomyini. Each of the four major lineages is characterized by one of the three patterns of grooving, and the particular groove-pattern is remarkably stable in each group.
Shape of skull varies from dolichocephalic to platycephalic. The morphology of each has been described in foregoing accounts. The dolichocephalic skull is highly specialized for planing, a grinding action of the teeth; whereas, the platycephalic skull is highly specialized for shearing, a slicing action of the teeth. Of course, concomitant specializations of the dentition, as described above, are closely associated with both specialized trends in the skull. Most kinds of living Geomyini have generalized skulls that show no tendency toward either of the specialized conditions.
Increase in size of body and skull is seen in most Pleistocene lineages of the Geomyini. Judging from the smallness of the skull in late Pliocene species, representing the base of three of these lineages, the ancestral species of the living assemblage were no larger than the living species of the subgenus Pappogeomys or the smaller subspecies of Geomys bursarius. The recorded range of variation in condylobasal length is 36.1 to 45.5 in Pappogeomys bulleri, including both adult males and females. Probably the skulls of the ancestral species were not significantly larger. Maximum dimensions of males in living species are 74.5 (subgenus Cratogeomys) and 75.0 (subgenus Orthogeomys). These are more than twice the minima observed in Pappogeomys bulleri.
This is the least specialized and most primitive of the four lineages, has a generalized type of skull, two grooves on the anterior face of each upper incisor, an enamel plate on the posterior wall of [565] P4, open or divergent lateral re-entrant angles on the premolars, and a bicolumnar and elongated M3. All of these features are primitive and essentially as in the ancestral morphotype. No other modern genus retains so much of the primitive structure. Phyletic trends in Zygogeomys are not well documented in the fossil record; and only a few fossils are known and they are fragmentary as discussed before. The genus is represented in the late Pliocene (Z. minor), middle Pleistocene (Z. persimilis), and Recent (Z. trichopus). The living species is a relict population in the mountains of Central México. Judging from the known material, the phyletic trends in the genus have been increase in size, reduction of enamel on the posterior face of P4 (occurring only in the living species) where a short enamel plate is retained on the lingual side of the tooth (see Fig. 7B), loss of the outer fourth of the enamel blade on the posterior wall of M1 and M2 (also occurring only in the living species), development of a more pronounced heel on the M3 by progressive elongation of the posterior loph, reduction in size of the jugal and its displacement ventrally, which allows the maxillary and squamosal bones to meet along the dorsal border of the zygomatic arch. The last specialization is seen in at least one taxon of Orthogeomys (Orthogeomys cherriei costaricensis). In my opinion, too much weight has been given to this feature in past classifications. Reduction of enamel in the upper dentition evidently occurred in the late Pleistocene, since the posterior plates on the upper cheek teeth were complete in specimens from the middle Pleistocene (Z. persimilis).
Geomys, slightly more specialized than Zygogeomys, must also be regarded as one of the most primitive of the living genera. Primitive features that have been retained are the generalized type of skull, the bisulcate pattern of grooves on the upper incisor, and the retention of enamel plates on both the anterior and posterior walls of M1 and M2 (see Fig. 9A). All of these primitive features are shared with Zygogeomys. In addition, three other trends, or specializations, in evolution characterize the phyletic development of Geomys. One major trend is toward loss of the enamel plate from the posterior wall of P4. No trace of enamel remains on the posterior wall of this tooth in late Pleistocene or Recent species of Geomys, and at least one of the earlier species (quinni) was also characterized by loss of this enamel plate. Secondly, M3 retains only a vestige of the primitive bicolumnar pattern after the initial stages of wear. In most Recent specimens, especially of the species [566] G. bursarius, the lateral re-entrant fold and the heel of M3 are small, and the re-entrant inflection is hardly evident. The lateral fold is more frequently well-developed in Irvingtonian species than in living species (White and Downs, 1961:13), illustrating progressive loss of the bicolumnar pattern in Pleistocene evolution. A third trend involves the modification of the lateral folds of the premolars. Primitively the angles of these folds are broadly open or divergently V-shaped, and some of the earliest species of Geomys, for example G. quinni, have retained this feature throughout life. Nevertheless, the main trend is toward progressive compression of the folds resulting in their walls being more nearly perpendicular, and parallel, to the long axis of the tooth. Obtuse re-entrant angles persist in premolars of young individuals of Irvingtonian species, but the adults are characterized by well-compressed folds, as in Recent species.
Remains of Geomys are abundant, especially from Pleistocene deposits of the Great Plains, but in most instances specific assignment is difficult or impossible since only isolated teeth or fragments of skulls have been preserved. Estimates of phyletic relationships of the known species of Geomys are depicted in Figure 8; those estimates are useful in discussing the phyletic development of the genus. One of the earliest known species, Geomys quinni, ranges from Upper Pliocene to the later stages of the Lower Pleistocene (Aftonian interglacial deposits). The dentition of G. quinni is essentially the same as in the living species except that open lateral re-entrant angles are retained in the premolars. Geomys paenebursarius, also of the early Pleistocene, is a smaller species and seems to be more directly in the line of evolution of the modern species. As yet unnamed smaller species of Geomys from the Rexroad fauna (late Pliocene) and Saunders fauna (latest Aftonian) may also be on the main line of evolution. Surprisingly, Geomys tobinensis and Geomys garbanii of later Irvingtonian provincial age are less specialized than either Geomys quinni or Geomys paenebursarius. It is likely that G. tobinensis and the unnamed species from the Dixon are closer to the main line of descent than G. paenebursarius suggesting that the direct ancestral lineage of the living species of Geomys was more conservative and less specialized than Geomys paenebursarius of the Lower Pleistocene. Geomys quinni and G. paenebursarius seem to have acquired specialized dental features in the early Pleistocene. Geomys quinni was successful on the Great Plains, and persisted into the late Blancan. The main line may be represented in the early Pleistocene by [567] Geomys paenebursarius from the Hancock formation of the Texas Trans-Pecos. The structure of G. paenebursarius indicates that it is in or close to the main line of descent, and probably evolved from one of the more primitive late Pliocene species of Geomys from the Rexroad fauna.
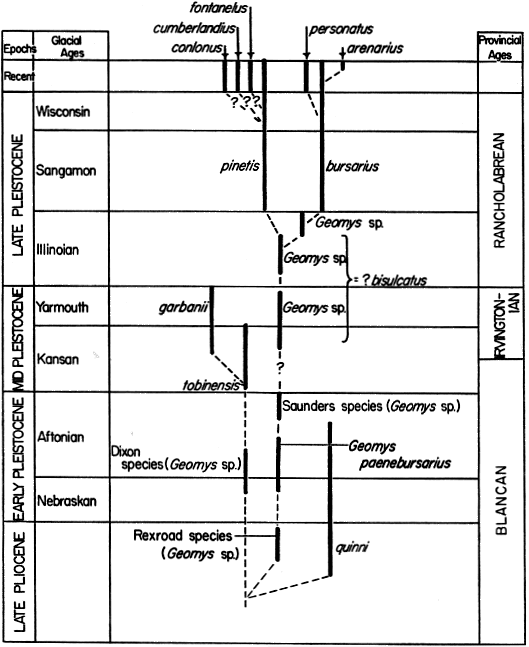
Fig. 8. Tentative arrangement of species of the genus Geomys, depicting phylogenetic trends and probable relationships within the genus.
Isolated teeth, to which the name Geomys bisulcatus probably applies, from Illinoian deposits on the Great Plains, show that the dentition characteristic of the living Geomys had been developed by that time. Actually, the Illinoian material is too fragmentary to [568] show clearly its taxonomic or phyletic affinities with the species of the later Pleistocene. Even so, the two main stocks of living Geomys, G. bursarius and G. pinetis, had certainly been differentiated by Sangamon time. The other living species evidently evolved from one or the other of these two stocks in a period of isolation from the main population, probably in either the Wisconsin or post-Wisconsin. For example, Geomys arenarius clearly differentiated from populations of Geomys bursarius that were isolated by the eastward retreat of the main population from the southwestern United States as that region became more arid in the post-Wisconsin.
In review, it seems that the Recent species, represented basically by bursarius and pinetis, evolved from Illinoian species (Geomys bisulcatus?), which descended in turn from the more primitive species of the early Pleistocene, possibly Geomys paenebursarius or possibly from descendants of the Saunders species. Actually the Saunders species may prove to be Geomys paenebursarius. At any rate, three trends that took place during the Pleistocene stage of evolution, in the direction of the modern species, were an increase in size, progressive loss of the posterior enamel plate on P4, and a decrease in the vertical depth of the enamel cap as a result of which the dentine is reached in the initial phases of attrition on the tooth of a juvenile. Geomys garbanii, occurring at the periphery of the range of the genus, is regarded as a sterile offshoot of the primitive tobinensis-line of evolution.
This is one of the more specialized genera of the Geomyini. Save for one record in the late Pleistocene (Orthogeomys onerosus), there is no fossil history of the genus upon which to reconstruct its phylogeny; therefore, its phyletic development must be estimated by comparing it and the primitive morphotype of the tribe. Results of that comparison suggest that Orthogeomys has closer affinities with Zygogeomys than with any of the other genera, and that Orthogeomys may have originated in an early dichotomy of primitive Zygogeomys stock instead of descending from the ancestral stock of the tribe. Except for the unisulcate incisors and the longer posterior loph on the third upper molars, the teeth of the two genera do not differ significantly. As in Zygogeomys, the enamel blade on the posterior wall of P4 has been reduced to a short plate restricted to the lingual third of the tooth (see Fig. 7F and H). In Orthogeomys, the trend in reduction of enamel is carried to its extreme only in the subgenus Orthogeomys, where this plate has been completely [569] lost in most taxa (see Fig. 7D). The most significant trends in Orthogeomys, and the principal basis for recognizing the genus, are the dolichocephalic specializations of the skull, as described elsewhere, and the adaptive traits that have equipped the genus for living in tropical environments. The dolichocephalic features are more sharply defined in the subgenera Orthogeomys and Macrogeomys, and are less developed in the subgenus Heterogeomys. Aside from the general dolichocephalic specializations, trends in Orthogeomys include: Increase in size; loss of the median one of the two grooves on the anterior face of the upper incisor in the ancestral stock; increase in the anteroposterior length of each of the cheek teeth, as well as the aforementioned elongation of the posterior loph of M3; compression of the lateral angles of the premolars; and the remarkable increase in the size of the rostrum.
The genus Pappogeomys, as it is conceived of in this study, is comprised of two subgenera; one, Pappogeomys, is generalized and primitive, and the other, Cratogeomys, is specialized, and includes the most highly specialized of the modern pocket gophers. The subgenus Pappogeomys is regarded as the ancestral lineage, and the subgenus Cratogeomys is regarded as an early offshoot, probably in the early Pleistocene, that became progressively more specialized in the course of its subsequent evolution. In the same period of time, the subgenus Pappogeomys changed little. It is known only from late Pliocene fragments and from the living species. The ancestral morphotype is preserved in Pappogeomys. Primitive characters are: (1) Small size; (2) skull generalized and smoothly rounded; (3) temporal ridges separate (not uniting into a sagittal crest); (4) enamel plates retained on both anterior and posterior walls of M1 and M2; (5) M3 bilophate, its posterior loph short. Basic specializations are few and include loss of the inner groove from the anterior face of the upper incisor; anteroposterior compression of the lateral re-entrant folds of the premolars; and loss of enamel from the posterior wall of P4. All three features have been perpetuated in the advanced subgenus Cratogeomys, suggesting that they were already developed in the early evolution of the subgenus Pappogeomys before Cratogeomys diverged. Agreement with Geomys is demonstrated by the lack of enamel on the posterior wall of P4 (see Fig. 9) and by retention of the posterior enamel plate on M1 and M2. In Pappogeomys (Pappogeomys) alcorni the enamel from the posterior face of M1 has been lost from all but the [570] lingual fourth or so of the posterior wall (Fig. 9E). Reduction of enamel in M1 provides an example of parallelism with the more advanced subgenus Cratogeomys, discussed below.
There is no record as yet of the early evolution of the subgenus Cratogeomys. The features that characterize the subgenus were [571] already well developed in the first known fossils which are from Wisconsin deposits of the late Pleistocene. Cratogeomys is not a homogenous assemblage; instead it is composed of two groups of living species, the generalized castanops group and the specialized gymnurus group. The castanops group may be survivors of the ancestral lineage that diverged in two different stages in the phyletic development of the main line. Even so, the castanops group has acquired its peculiar specializations. Indeed, P. merriami of the castanops group differs from the hypothetical stem more than does P. castanops. Judging from the structure of the living species of the subgenus Cratogeomys and from the primitive subgenus Pappogeomys, the subgenus Cratogeomys featured five major trends: (1) Increase in size; (2) formation of sagittal crest by union of the temporal impressions; (3) increase in rugosity and angularity of the skull; (4) progressive development of platycephalic specializations, including the elongation of the angular process of the mandible; (5) complete loss of enamel plates from the posterior wall of M1 and M2. Each trend is thought to be adaptive.
Fig. 9. Molariform dentitions of the Tribe Geomyini. Drawings illustrating enamel patterns characteristic of Geomys and Pappogeomys (including the subgenera Pappogeomys and Cratogeomys). × 5.
A and B. Geomys bursarius bursarius, adult female, No. 46275 (KU), Elk River, Sherborne Co., Minnesota. Left upper (A), P4-M3; right lower (B), p4-m3.
C and D. Subgenus Pappogeomys. Pappogeomys bulleri albinasus, adult female, No. 31002 (KU), W side La Venta, 13 mi. W and 4 mi. N Guadalajara, Jalisco. Left upper (C), P4-M3; right lower (D), p4-m3.
E and F. Subgenus Pappogeomys. Pappogeomys alcorni, adult female, No. 31051 (KU), holotype, 4 mi. W Mazamitla, 6600 ft., Jalisco. Left upper (E), P4-M3; right lower (F), p4-m3.
G and H. Subgenus Cratogeomys. Pappogeomys gymnurus tellus, adult female, No. 31051 (KU), 1 mi. NE Tala, 4400 ft., Jalisco. Left upper (G), P4-M3; right lower (H), p4-m3.
Loss of enamel is a trend common to all living genera of the tribe Geomyini, but the greatest loss has occurred in Cratogeomys. It has lost the plates on the posterior walls of M1 and M2 (Fig. 9G). If the lateral plates of M3 are considered as one functional plate and the lateral plates on either side of P4 together as two transverse plates, then, the transverse cutting blades in Cratogeomys number seven in the upper and seven in the lower cheek teeth compared with 10 in the upper and seven in the lower in the primitive morphotype. Indeed, in some species of the subgenus, one or both of the lateral plates on M3 is also lost, usually in old age, resulting in even greater reduction of enamel. Loss of enamel from the posterior [572] walls of the upper molars may be associated with changes in the mechanics of mastication from anteroposterior planing to anterotransverse shearing, as discussed elsewhere. Merriam (1895:95-96) argues convincingly that the posterior cutting blades of the upper molars would hinder efficient shearing action of the teeth; hence, selection would favor their reduction and eventual loss. Changes in the shape of the skull also seem to be correlated with the shift from a planing to a shearing type of mastication. More efficient shearing action, which depends upon lateral movement of the jaw, can be developed if the functional muscles insert farther laterally than is possible in the generalized type of skull. Therefore, platycephalic specializations involved lateral expansion of the braincase and mandible. Pronounced lateral expansion has been developed only in the gymnurus group of species, suggesting that the dental specializations evolved earlier in the evolution of the subgenus than did the platycephalic specializations of the skull, and that the castanops group separated from the gymnurus group before the common ancestor had developed the more extreme trends in platycephaly. It is interesting to note that the subtriangular M3 (Fig. 9G) postulated for the ancestral morphotype and that characterizes the subgenus Pappogeomys is retained also in the gymnurus group.
Alston, E. R.
1876. On the classification of the order Glires. Proc. Zool. Soc. London, 1876:61-98, 1 pl., June.
Alvarez, T.
1964. Nota sobre restos oseos de mamiferos del Reciente, encontrados cerca de Tepeapulco, Hidalgo, Mexico. Publ. Inst. Nac. Antro, e Hist., 15:1-15.
1965. Catálago Paleomastozoológico Mexicano. Publ. Inst. Nac. Antro, e Hist., 17:1-70.
Axlerod, D. I.
1950. Studies in late Tertiary paleobotany. Carnegie Inst. Washington Publ., 590:1-322.
1958. Evolution of the Madro-tertiary geoflora. Bot. Rev., 24:433-509.
Bader, R. S., and Techter, D.
1959. A list and bibliography of the fossil mammals of Illinois. Nat. Hist. Misc., Chicago Acad. Sci., 172:1-8, October 30.
Baird, S. F.
1858. Mammals. Part I. General report upon the zoology of the several Pacific railroad routes. Repts., explorations and surveys for a railroad route from the Mississippi River to the Pacific Ocean. Washington, D. C., vol. 8, pp. xlviii + 757, 35 figs., 43 pls., July 14.
Barbour, H., and Schultz, C. B.
1937. An early Pleistocene fauna from Nebraska. Amer. Mus. Nat. Hist. Novit., 942:1-10, September 10.
Berry, E. W.
1937. Tertiary floras of North America. Bot. Rev., 3:31-46.
Black, C. C.
1961. Rodents and lagomorphs from the Miocene Fort Logan and Deep River Formations of Montana. Postilla, Yale Peabody Museum, 48:1-20, 6 figs., January 16.
Brandt, J. F.
1855. Beiträge zur mähern Kenntiss der Säugethiere Russlands. Acad. Imp. des Sciences St. Petersburg, 7:79-336, 11 pls.
Brown, B.
1908. The Conard fissure, a Pleistocene bone deposit in northern Arkansas with description of two genera & 20 new species of mammals. Mem. Amer. Mus. Nat. Hist., 9:157-208, 2 pls., 3 figs.
1912. Brachyostracon, a new genus of glyptodont from Mexico. Bull. Amer. Mus. Nat. Hist., 31:167-177, August 2.
Chaney, R. W.
1947. Tertiary centers and migration routes. Ecol. Monog., 17:139-148.
Cook, H. J., and Cook, M. C.
1933. Faunal lists of the Tertiary Vertebrata of Nebraska and adjacent areas. Nebraska Geol. Surv., Paper No. 5:49.
Cope, E. D.
1878. Description of new extinct Vertebrata from the upper Tertiary and Dakota formations. Bull. U. S. Geol. and Geog. Surv., 4:379-396.
1884. The White River and John Day faunae, pp. 759-1002, pl. 64, figs. 1-9. In The Vertebrata of the Tertiary formations of the west. Book I. Part second. Rept. U. S. Geol. Surv. Terr. (F. V. Hayden), vol. 3.
1889. The vertebrate fauna of the Equus beds. Amer. Nat., 23:160-165.
Coues, E.
1877. Monographs of North American Rodentia. No. 8, Saccomyinae, pp. 483-542, and No. 10, Geomyinae, pp. 601-629, pl. 7, August.
Dalquest, W. W.
1962a. A record of the giant bison (Bison latifrons) from Cooke County, Texas. Texas Jour. Sci., 13:41-44, March.
1962b. The Good Creek Formation of Texas, and its fauna. Jour. Paleont., 36:568-582.
1964. A new Pleistocene local fauna from Motley County, Texas. Trans. Kansas Acad. Sci., 67:499-505, 4 figs., December 11.
1965. New Pleistocene Formation and local fauna from Hardeman County, Texas. Jour. Paleont., 39:63-72, 2 figs., January.
Dice, L. R.
1925. Rodents and lagomorphs of the Rancho La Brea Deposits. Carnegie Inst. Washington Publ., 349:119-130.
Davis, W. B.
1937. Variations in Townsend pocket gophers. Jour. Mamm., 18:145-158, May 12.
Ellerman, J. R.
1940. The families and genera of living rodents. Vol. I. Rodents other than Muridae. British Museum (Nat. Hist.), London, pp. xxvi + 689, 189 figs., June 8.
Elliott, D. G.
1903. A list of mammals obtained by Edmund Heller from the coast region of northern California and Oregon. Field Columb. Mus. Publ., 76, zool. ser. vol. 3(11):175-197, July 25.
Elftman, H. O.
1931. Pleistocene mammals of Fossil Lake, Oregon, Amer. Mus. Novit., 481:1-21, 10 figs., July 14.
Franzen, D. S.
1947. The pocket gopher, Geomys quinni McGrew, in the Rexroad fauna, Blancan age, of southwestern Kansas. Trans. Kansas Acad. Sci., 50:55-59.
Freudenberg, W.
1921. Geologie von Mexiko. Berlin, pp. viii + 232.
Galbreath, E. C.
1848. An additional specimen of the rodent Dikkomys from the Miocene of Nebraska. Trans. Kansas Acad. Sci., 51:316-317.
Gazin, C. L.
1935. Annotated list of Pleistocene Mammalia from American Falls, Idaho, Jour. Washington Acad. Sci., 25:297-302.
1942. The late Cenozoic vertebrate fauna from the San Pedro Valley, Arizona. Proc. U. S. Nat. Mus., 92(3155):475:518, 2 pls., 9 figs.
Gervais, P.
1849. Rongeurs. In Dictionnaire universel d'historie naturelle, Dirigé par M Ch. d'Orbigny, Paris, M. M. Renard, Martinet et Cie, vol. 11:198-204.
Gidley, J. W.
1922. Preliminary report on fossil vertebrates of the San Pedro Valley, Arizona, with descriptions of new species of Rodentia and Lagomorpha. U. S. Geol. Surv. Prof. Papers, 131:119-130, pls. 34 and 35.
Gidley, J. W., and Gazin, C. L.
1933. New Mammalia in the Pleistocene fauna from Cumberland Cave. Jour. Mamm., 14:343-357, 9 figs.
Gill, T. H.
1872. Arrangement of the families of mammals with analytical tables. Smithsonian Misc. Coll., 11, art. 1, pp. vi + 98.
Gilmore, R. M.
1947. Report on a collection of mammal bones from archeologic cave-sites in Coahuila, Mexico. Jour. Mamm., 38:147-165, June 1.
Gray, J. E.
1868. Synopsis of the species Saccomyinae or pouched mice in the collection of the British Museum. Proc. Zool. Soc. London, 1868:199-206, May.
Green, M.
1956. The lower Pliocene Ogallala-Wolf Creek vertebrate fauna, South Dakota. Jour. Paleo., 30(1):146-169, 12 figs., January.
Gut, H. J., and Ray, C. E.
1963. The Pleistocene vertebrate fauna of Reddick, Florida. Quart. Jour. Florida Acad. Sci., 26:315-328.
Hall, E. R., and Kelson, K. R.
1959. The mammals of North America. 2 vols., xxx + 1083 pp., 553 figs., 500 maps, March 31.
Harris, A. H., and Findley, J. S.
1964. Pleistocene—Recent fauna of the Isleta Caves, Bernalillo County, New Mexico. Amer. Jour. Sci., 262:114-120, January.
Hay, O. P.
1920. Description of some Pleistocene vertebrates found in the United States. Proc. U. S. Nat. Mus., 57:83-146, 8 pls., 4 figs.
1921. Description of species of Pleistocene vertebrate types of specimens of most of which are preserved in the U. S. National Museum. Proc. U. S. Nat. Mus., 59:599-642.
1923. The Pleistocene of North America and its vertebrated animals from the states east of the Mississippi River and from the Canadian provinces east of longitude 95°. Carnegie Inst. Washington, Publ. 322: 1-499, February 24.
1924. The Pleistocene of the middle region of North America and its vertebrated animals. Carnegie Inst. Washington, Publ. 322A:1-346.
1927. The Pleistocene of the western region of North America and its vertebrate animals. Carnegie Inst. Wash., Publ. 322B, pp. v + 346, 12 pls., 21 maps, 10 text figs.
Hibbard, C. W.
1938. An Upper Pliocene fauna from Meade County, Kansas. Trans. Kansas Acad. Sci., 40:239-265, 5 pls., 2 figs.
1941a. The Borchers fauna, a new Pleistocene interglacial fauna from Meade County, Kansas. Bull. State Geol. Surv. Kansas, 38:197-220, 2 pls., July 14.
1941b. Mammals of the Rexroad Fauna from the Upper Pliocene of southwestern Kansas. Trans. Kansas Acad. Sci., 44:265-313, 4 pls.
1943. The Rezabek fauna, a new Pleistocene fauna from Lincoln County, Kansas. Univ. Kansas Sci. Bull., 29:235-247, October 15.
1944. Stratigraphy and vertebrate paleontology of Pleistocene deposits of southwestern Kansas. Geol. Soc. Amer. Bull., 55:707-754, 3 pls., 20 figs.
1950. Mammals of the Rexroad Formation from Fox Canyon, Meade County, Kansas. Univ. Michigan Paleo. Contrib., 8(6):113-192, 5 pls., 23 figs.
1951. Thomomys talpoides (Richardson) from a late Pleistocene deposit in Kansas. Jour. Mamm., 32:229-230.
1952. Vertebrate fossils from late Cenozoic deposits of central Kansas. Univ. Kans. Paleont. Contrib., Vertebrata, 2:1-14, March 20.
1953. The Saw Rock Canyon fauna and its stratigraphic significance. Papers Michigan Acad. Sci., Arts, and Letters, 38:387-411.
1954. A new Pliocene vertebrate fauna from Oklahoma. Papers Michigan Acad. Sci., Arts, Letters, 39:339-359.
1955a. Pleistocene vertebrates from the upper Bercerra (Bercerra Superior) Formation, valley of Tecquixquiac, Mexico, with notes on other Pleistocene forms. Contribution from Mus. Paleo., Univ. Michigan, 12:47-96, 9 pls., 5 figs., February 18.
1955b. The Jinglebob interglacial (Sangamon?) fauna from Kansas and its climatic significance. Contrib. Mus. Paleo., Univ. Michigan, 12: 179-228, 2 pls., 8 figs., September 1.
1956. Vertebrate fossils from the Meade Formation of southwestern Kansas. Papers Michigan Acad. Sci., Arts, and Letters, 41:145-200.
1958. Summary of North American Pliestocene mammalian local faunas. Papers of Michigan Acad. Sci., Arts, and Letters, 43:1-32.
1963. A late Illinoian fauna from southwestern Kansas and its climatic significance. Papers Michigan Acad. Sci., Arts and Letters, 48:187-221, 8 figs.
Hibbard, C. W., and Keenmon, K. A.
1950. New evidence of the Lower Miocene age of the Blacktail Deer Creek Formation in Montana. Contrib. Univ. Michigan Mus. Paleo., 8:193-204, 3 figs., 1 map, July 28.
Hibbard, C. W., and Mooser, O.
1963. A porcupine from the Pleistocene of Aguascalientes, Mexico. Contrib. Mus. Paleo. Univ. Michigan, 18:245-250, November 22.
Hibbard, C. W., Ray, D. E., Savage, D. E., Taylor, D. W., Guilday, G. E.
1965. Quaternary mammals of North America. In Quaternary of the United States. Eds. H. E. Wright, Jr., and D. G. Frey. Princeton Univ. Press, Princeton, pp. x + 922.
Hibbard, C. W., and Riggs, E.
1949. Upper Pliocene vertebrates from Keef Canyon, Meade County, Kansas. Geol. Soc. Amer. Bull., 60(5):829-860, 11 figs., 5 pls.
Hibbard, C. W., and Taylor, D. W.
1960. Two late Pleistocene faunas from southwestern Kansas. Contrib. Univ. Michigan Mus. Paleo., 16(1):1-223, 16 pls., 18 figs.
Hibbard, C. W., and Wilson, J. A.
1950. A new rodent from subsurface stratum in Bee County, Texas. Jour. Paleo., 24:621-623, September.
Hooper, E. T.
1946. Two genera of pocket gophers should be congeneric. Jour. Mamm., 27:397-399, November, 1965.
James, G. T.
1963. Paleontology and nonmarine stratigraphy of the Cuyama Valley Badlands, California. Part I. Geology, faunal interpretations, and systematic descriptions of Chiroptera, Insectivora, and Rodentia. Univ. California Publ. Geol. Sci., 45:iv + 170, 8 pls., 53 figs., June 26.
Johnson, C. S., and Savage, D. E.
1955. A survey of various Late Cenozoic vertebrate faunas of the Panhandle of Texas, Part I. Univ. California Publ. Geol. Sci., 31:27-50.
Kendeigh, C. S.
1961. Animal Ecology. Prentice Hall, Inc., Englewood Clifts, New Jersey, pp. x + 468.
Kurten, B.
1965. The Pleistocene Felidae of Florida. Bull. Florida State Mus., 9(6):215-273, June 8.
Leidy, J.
1869. The extinct mammalian fauna of Dakota and Nebraska—together with a synopsis of the mammalian remains of North America. Philadelphia, 472 pp., 30 pls.
MacDonald, J. R.
1963. The Miocene faunas from the Wounded Knee area of western North Dakota. Bull. Amer. Mus. Nat. Hist., 125:141-328, 30 figs., August 26.
Maldonado-Koerdell, M.
1948. Los vertebrados fosiles del Cuaternario en Mexico. Rev. Soc. Mex. Hist. Nat., 9:1-35, June.
Marsh, O. C.
1871. Notice of some new fossil mammals and birds from the Tertiary formations of the west. Amer. Jour. Sci. and Art, 3:120-127.
Matthew, W. D.
1899. A provisional classification of the fresh-water Tertiary of the West. Bull. Amer. Mus. Nat. Hist., 12:19-75, March 31.
1902. List of the Pleistocene fauna from Hay Springs, Nebraska. Bull. Amer. Mus. Nat. Hist., 16:317-322, September 25.
1909. Faunal lists of the Tertiary mammalia of the west. Bull. U. S. Geol. Surv., 361:91-138.
1910. Notes on the osteology and relationships of Paramys, and the affinities of the Ischyromyidae. Bull. Amer. Mus. Nat. Hist., 28: 43-72, 19 figs., March 22.
1923a. Fossil bones in the rock. The fossil quarry near Agate, Sioux County, Nebraska. Nat. Hist., 23:358-369.
1923b. The occurrence of the Hesperopithecus tooth. Amer. Mus. Novit., 53:11-13, January 6.
1924. Third Contribution to the Snake Creek Fauna. Bull. Amer. Mus. Nat. Hist., 50:59-210, July 3.
Matthew, W. D., and Cook, H. J.
1909. A Pliocene fauna from western Nebraska. Bull. Amer. Mus. Nat. Hist., 26:361-414.
Mayer-Oakes, W. J.
1959. A stratigraphic excavation at El Risco, Mexico. Proc. Amer. Phil. Soc., 103:332-373, June 15, 1959.
McGrew, P. O.
1944. An early Pleistocene (Blancan) fauna from Nebraska. Field Mus. Nat. Hist., Geol. Ser., 9(2):33-66, January 20.
Merriam, C. H.
1895. Monographic revision of the pocket gophers, family Geomyidae (exclusive of the species of Thomomys). N. Amer. Fauna, 8:1-258, 19 pls., 4 maps, 81 figs., January 31.
Miller, G. S., Jr., and Gidley, J. W.
1918. Synopsis of the supergeneric groups of rodents. Jour. Washington Acad. Sci., 8:431-448, July 19.
Miller, G. S., Jr., and Kellogg, R.
1955. List of North American Recent mammals. Bull. U. S. Nat. Mus., 205:xii + 954, March 3.
Mooser, O.
1959. La fauna "Cedazo" del Pleistoceno en Aguascalientes. An. Inst. Biol. Mex., 29(1 and 2):409-452, 29 figs.
Paulson, G. R.
1961. The mammals of the Cudahy fauna. Papers Michigan Acad. Sci., Arts, and Letters, 46:127-153.
Peters, W.
1874. Uber die Taschenmause und eine neue Art derselben, Heteromys adspersus, aus Panama. Monatsber. K. Acad. Wissensch. Berlin, pp. 354-359, 1 pl.
Ray, C. E.
1958. Additions to the Pleistocene mammalian fauna from Melbourne, Florida. Bull. Mus. Comp. Zool., 119:421-449, 5 figs., November.
Russell, R. J.
1960. Pleistocene pocket gophers from San Josecito Cave, Nuevo León, México. Univ. Kansas Publ., Mus. Nat. Hist., 9:539-548, January 14.
Savage, D. E.
1951. Late Cenozoic vertebrates of the San Francisco Bay Region. Univ. California Publ. Geol. Sci., 28:215-314, March 23.
Schultz, C. B., and Howard, E. B.
1935. The fauna of Burnet Cave, Guadalupe Mountains, New Mexico. Proc. Acad. Nat. Sci. Philadelphia, 87:273-298, 37 figs., 6 pls.
Schultz, C. B., Lueninghoener, G. C., and Frankforter, W. D.
1951. A graphic résumé of the Pleistocene of Nebraska. Bull. Univ. Nebraska State Mus., 3(6):1-41, July.
Schultz, C. B., and Stout, T. M.
1948. Pleistocene mammals and terraces in the Great Plains. Bull. Geol. Soc. Amer., 59:553-589, June.
Schultz, C. B., and Tanner, L. G.
1957. Medial Pleistocene fossil vertebrate localities in Nebraska. Univ. Nebraska State Mus., 4(4):59-81, September.
Schultz, G. E.
1965. Pleistocene vertebrates from the Butler Springs local fauna, Meade County, Kansas. Papers Michigan Acad. Sci., Arts, and Letters, 50:235-265.
Schultz, J. R.
1938. A late Quaternary mammal fauna from the tar seeps of McKittrick, California. Carnegie Inst. Washington Publ., 487:111-215, July 6.
Shotwell, J. A.
1956. Hemphillian mammalian assemblage from northeastern Oregon. Bull. Geol. Soc. Amer., 67:717-738, 7 figs., June.
1963. The Juntura Basin: studies in earth history and paleoecology. Mammalian fauna of the Drewsey Formation, Bartlett Mountains, Drinkwater and Otis Basin local faunas. Trans. Amer. Phil. Soc., n.s., 53(1):1-77, April.
Simpson, G. G.
1928. Pleistocene mammals from a cave in Citrus County, Florida. Amer. Mus. Novit., 328:1-4, October 26.
1929. Pleistocene Mammalian fauna of the Seminole Field, Pinellas County, Florida. Bull. Amer. Mus. Nat. Hist., 56:561-599.
1945. The principles of classification and a classification of mammals. Bull. Amer. Mus. Nat. Hist., 85: pp. xvi + 350, October 5.
1953. The major features of evolution. Columbia Univ. Press, New York, pp. xx + 434, 52 figs.
Sinclair, W. J.
1905. New Mammalia from the Quaternary caves of California. Bull. Dept Geol. Univ. California, 4:145-161, pls. 19 and 23.
Skinner, M. F.
1942. The fauna of Papago Springs Cave, Arizona. Bull. Amer. Mus. Nat. Hist., 80:143-220, November 6.
Slaughter, B. H., and Hoover, B. R.
1963. Sulphur River Formation and the Pleistocene mammals of the Ben Franklin local fauna. Jour. Grad. Resc. Center, 31(3):132-148, June.
Slaughter, B. H., and Ritchie, F.
1963. Pleistocene mammals of the Clear Creek local fauna, Denton County, Texas. Jour. Grad. Resc. Center, 31(3):117-131, June.
Starrett, A.
1956. Pleistocene mammals of the Berends fauna of Oklahoma. Jour. Paleo., 30:1187-1192.
Stephens, J. J.
1960. Stratigraphy and paleontology of a late Pleistocene Basin, Harper Co., Oklahoma. Bull. Geol. Soc. Amer., 71:1675-1702, November.
Strain, W. S.
1966. Blancan mammalian fauna and Pleistocene formations, Hudspeth County, Texas. Bull Texas Memorial Mus., 10:1-55, 8 figs., 13 pls., February.
Tamsitt, J. R.
1957. Peromyscus from the Late Pleistocene of Texas. The Texas Jour. Sci., 9(3):355-363, September.
Waterhouse, G. R.
1839. Observations on the Rodentia, with a view to point out the groups, as indicated by the structure of the crania, in this order of mammals. Mag. Nat. Hist., new series, 3:90-96, 184-188, 274-279, 593-600, 1 pl.
1841. Observations on the Rodentia. Ann. Mag. Nat. Hist., 8:81-84, pl. 2.
1848. A natural history of the mammalia. Vol. 2, containing the order Rodentia, or gnawing mammalia. Hippolyte Bailliere, London, 500 pp., 21 pls.
Waters, J. H.
1957. Second find of Thomomys talpoides from late Pleistocene in Kansas. Jour. Mamm., 37:540-542.
White, J. A., and Downs, T.
1961. A new Geomys from the Vallecito Creek Pleistocene of California, with notes on variation in Recent and fossil species. Contr. in Science, Los Angeles Co. Mus., 42:1-34, June 30.
Wilson, R. W.
1933a. Pleistocene mammalian fauna from the Carpinteria asphalt. Carnegie Inst. Washington Publ., 440:59-76, November.
1933b. A rodent fauna from the later Cenozoic beds of southwestern Idaho. Carnegie Inst. Washington Publ. 440:117-135, 8 figs., 2 pls., December.
1936. A Pliocene rodent fauna from Smith's Valley, Nevada. Carnegie Inst. Washington Publ., 473:15-34.
1937. Pliocene rodents of western North America. Carnegie Inst. Washington Publ. 487:21-73, July 23.
1940. Two new Eocene rodents from California. Carnegie Inst. Washington Publ., 514:85-95, June 27.
1949. Early Tertiary rodents of North America. Carnegie Inst. Washington Publ., 584:67-164, June 22.
Winge, H.
1924. Pattedyr-Slaegter. Vol. 2, Rodentia, Carnivora, Primates. H. Hagerups Forlag, Copenhagen, 321 pp.
Wood, A. E.
1935. Evolution and relationship of the heteromyid rodents. Ann. Carnegie Mus., 24:73-262, May 13.
1936. Geomyid rodents from the Middle Tertiary. Amer. Mus. Novit., 866:1-31, July 2.
1950. A new geomyid rodent from the Miocene of Montana. Ann. Carnegie Mus., 31:335-338, 1 fig.
1955. A Revised classification of rodents. Jour. Mamm., 36:165-187, May 26.
Wood, A. E., and Wilson, R. W.
1936. A suggested nomenclature for the cusps of the cheek teeth of rodents. Jour. Paleo., 10:388-391, 2 figs., July.
Transmitted May 29, 1967.
![]()
Transcriber's Notes
All obvious typographical errors were corrected. Minor changes were made to standardize the text to match the most prevalent form used.
Typographical Corrections
| Page | Correction | |
| 477 | cumberlandicus => cumberlandius | |
| 535 | breath => breadth |
End of the Project Gutenberg EBook of Evolution and Classification of the
Pocket Gophers of the Subfamily Geomyinae, by Robert J. Russell
*** END OF THIS PROJECT GUTENBERG EBOOK POCKET GOPHERS--SUBFAMILY GEOMYINAE ***
***** This file should be named 40282-h.htm or 40282-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/4/0/2/8/40282/
Produced by Chris Curnow, Joseph Cooper, Tom Cosmas and
the Online Distributed Proofreading Team at
http://www.pgdp.net
Updated editions will replace the previous one--the old editions
will be renamed.
Creating the works from public domain print editions means that no
one owns a United States copyright in these works, so the Foundation
(and you!) can copy and distribute it in the United States without
permission and without paying copyright royalties. Special rules,
set forth in the General Terms of Use part of this license, apply to
copying and distributing Project Gutenberg-tm electronic works to
protect the PROJECT GUTENBERG-tm concept and trademark. Project
Gutenberg is a registered trademark, and may not be used if you
charge for the eBooks, unless you receive specific permission. If you
do not charge anything for copies of this eBook, complying with the
rules is very easy. You may use this eBook for nearly any purpose
such as creation of derivative works, reports, performances and
research. They may be modified and printed and given away--you may do
practically ANYTHING with public domain eBooks. Redistribution is
subject to the trademark license, especially commercial
redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License available with this file or online at
www.gutenberg.org/license.
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS', WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTABILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation information page at www.gutenberg.org
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at 809
North 1500 West, Salt Lake City, UT 84116, (801) 596-1887. Email
contact links and up to date contact information can be found at the
Foundation's web site and official page at www.gutenberg.org/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
[email protected]
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit www.gutenberg.org/donate
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations.
To donate, please visit: www.gutenberg.org/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart was the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For forty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Most people start at our Web site which has the main PG search facility:
www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.