Significant Achievements in Space Bioscience
NASA SP-92
 Scientific and Technical Information Division 1966
Scientific and Technical Information Division 1966Foreword
This volume is one of a series which summarize the progress made during the period 1958 through 1964 in discipline areas covered by the Space Science and Applications Program of the United States. In this way, the contribution made by the National Aeronautics and Space Administration is highlighted against the background of overall progress in each discipline. Succeeding issues will document the results from later years.
The initial issue of this series appears in 10 volumes (NASA Special Publications 91 to 100) which describe the achievements in the following areas: Astronomy, Bioscience, Communications and Navigation, Geodesy, Ionospheres and Radio Physics, Meteorology, Particles and Fields, Planetary Atmospheres, Planetology, and Solar Physics.
Although we do not here attempt to name those who have contributed to our program during these first 6 years, both in the experimental and theoretical research and in the analysis, compilation, and reporting of results, nevertheless we wish to acknowledge all the contributions to a very fruitful program in which this country may take justifiable pride.
Preface
This summary of certain aspects of the space biology program of the National Aeronautics and Space Administration brings together some results of NASA research and NASA-sponsored research under grants and contracts from 1958 through 1964. Closely related research even though not sponsored by NASA is also included.
The space biology program has had a late start in comparison with the space physics program, and only a token program existed before 1962. Much of the present research involves preparation of space-flight experiments and obtainment of adequate baseline information. Perhaps half the research results reported are derived from the NASA program. Additional information is included from many other sources, especially the U.S. Air Force with its long history of work in aviation and aerospace medicine.
Relatively few biological space-flight experiments have been undertaken. These have been to test life-support systems and to demonstrate, before manned space flight, an animal's capability to survive. Few critical biological experiments have been placed in orbit by NASA, but a biosatellite program will soon make a detailed study of the fundamental biological effects of weightlessness, biorhythms, and radiation.
The search for extraterrestrial life has been limited to ground-based research and planning for planetary and lunar landings. Life-detection experiments have been developed and tested, and an important and exciting program is being planned to detect and study extraterrestrial life, if it exists.
Interest in space biology has been slow in developing, and there has been some caution and controversy in the scientific community. However, increased interest is starting to push forward the frontier of this new and important scientific field, and future outlook appears to be optimistic.
This summary was written and compiled by the members of the Bioscience Programs Division of the Office of Space Science and Applications. The report was edited and chapters 1, 3, 6, and 7 were written by Dale W. Jenkins, Chief, Environmental Biology; chapter 2, by Gregg Mamikunian, Staff Scientist, Exobiology; chapters 4 and 8, by Richard E. Belleville, Chief, Behavioral Biology; and chapter 5, by George J. Jacobs, Chief, Physical Biology.
Contents
chapter 1
The biological program of the National Aeronautics and Space Administration had a late start. A small life sciences group, organized in 1958, was concerned with life support and use of primates for system and vehicle testing for the Mercury program. Three small suborbital flights of biological materials were flown in space.
The Bioscience Program Office of the Office of Space Science and Applications was organized in 1962. The goals of the Bioscience Program are: (1) to determine if extraterrestrial life exists anywhere in the solar system and to study its origin, nature, and level of development, if it is present; (2) to determine the effects of space and planetary environments on Earth organisms, including man; (3) to conduct biological research to develop life support and protective measures for extended manned space flight; and (4) to develop fundamental theories in biology relative to origin, development, and relationship to environment. Research and development has been carried out to design life-detection experiments and instruments for future flights to Mars and to develop experiments to study the effects of the space environment on living organisms. A biosatellite program, started in 1963, has the first of six flights scheduled for 1966.
Space exploration has demanded a rigorous development, especially in the biosciences area. Investigation of the solar system for exotic life forms, the environmental extremes to which Earth organisms (including man) are being exposed, the possibilities for modification of planetary environments by biological techniques yet to be developed, and the problems of communication in biosystems are areas which have required refinement of the theoretical framework of biology before progress could be made rapidly enough to keep pace with technological advances in transportation.
Of all the sciences, biology alone has not yet benefited from comparisons with the universe beyond Earth. It is reasonable to suppose that breakthroughs might be made in biology on the basis of comparisons with life from other worlds. Organisms elsewhere may have found alternatives to processes we think of as basic characteristics of life.
In contrast, physical science has advanced sufficiently to provide a great body of laws which may be expressed in mathematical terms, and by which phenomena may be predicted with complete accuracy. A well-known characteristic of biological phenomena is variability. The Darwinian concept of evolution is perhaps the only pervading generalization in biology. This concept has been supported by evidence of a hereditary mechanism in the discovery of genes and gene mutations.
Space bioscience represents the convergence of main disciplines with a single orientation, whose direction is determined by the problems of manned space travel which have, in turn, created a host of bioengineering problems concerned with supporting man in space.
Foremost among these questions is the possibility of the existence of extraterrestrial life. The field which is concerned with the search for extraterrestrial life has come to be called "exobiology." In addition to the challenge of great technological problems which must be solved, exobiology is so closely related to the central scientific questions in biological science that it is considered by some to be the most significant pursuit in all of science.
One of the major opportunities already presented by the advances in propulsion systems is the ability to escape from the influence of the Earth, which has made possible the study of organism-environment relationships, particularly the role that environmental stimuli play in the establishment and maintenance of normal organization in living systems.
Transcending even these formidable objectives of space bioscience is an objective shared by all life sciences, the discovery of nature's scheme for coding the messages contained in biological molecules. Extraterrestrial biology seeks to find not only evidence of life now present, but the vestigial chemicals of its previous existence. The ways and means have already been made available to study molecules on whose long, recorded messages is written the autobiography of evolution—the history of living organisms extending back to the beginnings of life. On this same basis, it is now within the realm of science to foresee the means of predicting the development of life from primordial, nonliving chemical systems. Closely allied to the search for extraterrestrial life is research which seeks to identify the materials and the conditions which are the prerequisites of life.
Space bioscience research is now extending human knowledge of fundamental biological phenomena, both in space and on Earth, just as the physical sciences explore other aspects of the universe. The accomplishment of bioscience objectives is totally dependent upon advances in the technology of space flight. A highly developed launch-vehicle capability is essential to accomplish the long-duration missions required in the search for extraterrestrial life.
Life on other planets in the solar system (with emphasis on Mars) will be investigated by full exploitation of space technology which will allow both remote (orbiter) and direct (lander) observations of the planetary atmosphere, surface, and subsurface. Certain characteristics of terrestrial life, such as growth and reproduction, provide a basis for relatively simple experiments which may be used on early missions to detect the existence of life on Mars. Later missions will provide extensive automatic laboratory capabilities for analyzing many samples taken from various depths and locations. Because of the hypothetical nature of current experiment designs, it is likely that visual observations of the planet will be required. Many technical problems are involved in storing and transmitting the large amounts of data over planetary distances. Such visual observations might very well be crucial in interpreting results from other experiments. Critical to all exploration of the Moon and planets are the requirements to: (1) prevent contamination of the environment with Earth organisms and preserve the existing conditions of the planet for biological exploration; (2) provide strict quarantine for anything returned to Earth from the Moon and planets.
The biological exploration of Mars is a scientific undertaking of the greatest significance. Its realization will be a major milestone in the history of human achievement. The characterization of life, if present, and study of the evolutionary processes involved and their relationship to the evolution of terrestrial life would have a great scientific and philosophical impact. What is at stake is nothing less than knowledge of our place in nature.
Extended Earth orbital flights with subhuman specimens will be used to determine the effects on Earth organisms of prolonged weightlessness, radiation, and removal from the influence of the Earth's rotation. Such flights of biosatellites and other suitable spacecraft are expected to: (1) establish biological specifications for extending the duration of manned space flight; (2) provide a flexible means of testing unforeseen contingencies, thus providing an effective biological backup for manned missions; (3) yield experimental data more rapidly by virtue of the greater number and expendability of subjects; (4) anticipate possible delayed effects appearing in later life or in subsequent generations, through use of animal subjects with more rapid development and aging; (5) develop and test new physiological instrumentation techniques, surgical preparations, prophylactic techniques, and therapeutic procedures which are not possible on human subjects; and (6) provide a broad background of experience and data which will permit more accurate interpretations of observed effects of space flight on living organisms, including man.
chapter 2
The possibility of discovering an independent life form on a planet other than Earth presents an unequaled challenge in the history of scientific search. Therefore, the detection of life within the solar system is a major objective of space research in the foreseeable future.
The scientific data presently available concerning the possible existence of a Martian life form and the chemical constitution of the surface of Mars are disappointingly few. In fact, it is impossible to make a statement about any of the many surface features, other than the polar caps, with any degree of certainty. The observational results have been accounted for by many conflicting hypotheses which can only be resolved by the accumulation of new evidence.
The arguments supporting the existence of Martian life ([ref.1]) are based on the following observations:
The various colors, including green, exhibited by the dark areas
The seasonal changes in the visual albedo and polarization of the dark areas
The ability of the dark areas to regenerate after an extensive "duststorm"
The presence of absorption bands at 3.3µ-3.7µ, attributed to organic molecules
Conflicting interpretations of the above observations have been advanced. The argument based on the colors is inconclusive, and several workers have suggested that the color is a contrast effect with the bright-reddish continents. The meager quantitative data have been discussed by Öpik ([ref.2]) who has reduced Kozyrev's photometric observations of the very dark area of Syrtis Major to intrinsic reflectivities by allowing for the estimated atmospheric attenuation and reflectivity. Kuiper ([ref.3]) similarly demonstrated the absence of the near-infrared reflection maximum, which is characteristic of most green plants, indicating that chlorophyll was not responsible for the color.
The second and third arguments remain the most cogent. However, serious limitations are imposed on the second if the severity of the Martian climate is considered. Föcas ([ref.4]) has photometrically measured the seasonal changes in the fine structure of the dark areas of Mars and concludes that—
The dark areas of Mars show periodic variation of intensity following the cycle of the darkening element
The average intensity of the dark area, not including the action of the darkening waves, increases from the poles toward the equator
The action of each of the darkening waves decreases from the poles toward the equator. This decrease is balanced in the equatorial zone by the combined action of the two darkening waves alternately originating at the two poles. The mechanism of the darkness-generating element seems to be constant for all latitudes during the Martian year.
The variation in intensity has been explained recently by nonlife mechanisms for Depressio Hellespontica (an area showing one of the greatest seasonal changes) ([ref.2]). Similar nonlife mechanisms may be applicable to the other dark regions, and, thus, the "darkening" can be used only as circumstantial evidence in support of a Martian life form.
If inorganic interpretations of the seasonal albedo variation are accepted, then an inorganic interpretation must also be advanced for the polarization variation. Two possibilities can be suggested:
A change in surface texture, caused by varying absorption of atmospheric constituents, causing both the albedo and polarization to change in the manner observed
A change in surface texture, in which the surface material becomes rougher, which also explains the observed polarization data ([ref.5])
The third argument against the regenerative feature of the dark areas being a life process has been advanced by Kuiper ([ref.6]). It is based on atmospheric circulation causing dust, presumably lava, to be blown on the dark areas of Mars during the late summer, autumn, and winter, and then removed during the spring. Mamikunian and Moore have recently advanced the similar explanation that carbonaceous chondrites or asteroidal matter may induce the observed phenomenon if they are abundant on the planet's surface. The pulverized chondritic material will exhibit a high degree of opacity due to localization and, hence, a change in polarization characteristics and a decrease in polarization following mixing of the chondritic material with indigenous surface minerals.
The fourth observational argument, the Sinton bands ([ref.7]), has been shown to be at least doubtful. Rea, Belsky, and Calvin ([ref.8]) recorded infrared reflection spectra for a large number of inorganic and organic samples, including minerals and biological specimens, for the purpose of interpreting the 3µ-to-1µ spectrum of Mars. These authors state that a previous suggestion that the Martian "bands" be attributed solely to carbohydrates is not a required conclusion. At the same time they fail to present a satisfactory alternate explanation, and the problem remains unsolved. More recently, Rea et al. ([ref.9]) noted the similarity between the 3.58µ and 3.69µ minima in the Martian infrared spectra and those of D2O-HDO-H2O mixtures and, particularly, of HDO.
With all this marked disagreement in interpreting the observational data concerning Mars, it becomes clearly evident that an experimental approach to the detection of life on Mars should provide the maximum positive information possible. Some life-detection experiments developed with NASA support have been summarized by Quimby ([ref.10]).
The schema of the biological exploration of a planet is to conduct a series of complementary experiments proceeding from general to specific. The general experiments will examine gross characteristics of the planet's environment and surface for determining the probability of an active biota (life). Data from the general experiments will be significant in—
Defining the nature of specific experiments in which life detection is the major objective; and
Providing a high degree of confidence in undertaking specific experiments, since indications from the gross characterization of the planet in question will influence the choice and design of the specific experiments.
The biological exploration of planets is then to be defined as the search for those parameters relevant to the origin, development, sustenance, and degradation of life in a planetary environment. This definition will give rise to a critical question for each progressively specific and complex experiment to determine—
The existence of life on the planet
The degree of similarity or dissimilarity (structure and function) with respect to terrestrial life
The origin of this planetary life
The immediate objective of the biological explorations of the planet is to define the state of the planetary surface, which may exhibit the following properties:
A prebiota (defined as the absence of life)
An active biota (defined as the presence of life)
An extinct biota (defined as evidence of former life)
The identification and the detailed characterization of each of the above stages of planetary development constitute the subject matter of the biological exploration of the planets and, specifically, Mars.
THE EXPERIMENTAL INVESTIGATION OF CHEMICAL EVOLUTION
Attempts have been made to simulate and approximate models of primitive Earth conditions for abiogenic synthesis, and successful synthesis of essential biochemical constituents necessary for maintaining life has been partly accomplished.
Urey ([ref.11]) has clearly pointed out the possible role of a reducing atmosphere in the synthesis of prebiological organic molecules. Miller ([ref.12]) synthesized a variety of amino acids in a reducing atmosphere by means of an electrical discharge. A variety of organic compounds have been synthesized by the action of various energy sources upon reducing atmospheres, and several investigators have extended the Urey-Miller-type reactions to synthesize nucleic acid components ([ref.13]), adenosine triphosphate ([ref.14]), and a host of biologically essential organic compounds.
It is likely that in the synthesis of organic moieties, simple and specific molecules were first produced when the planets had a reducing atmosphere. Further complexity or degradation of the organic compounds produced varied, depending on the geochemical changes of the planet's surface, the atmospheric constituents, the degree of interaction between surface and atmosphere, and the rate of the organic synthesis. Oparin ([ref.15]) presented the most detailed mechanisms for the spontaneous generation of the first living organism arising in a sea of organic compounds synthesized in a reducing atmosphere on Earth.
It is generally accepted that, under favorable conditions, life can arise by spontaneous generation. A primary requirement for this initiation is that there be abundant organic compounds concentrated in one or more specific zones. These simple organic molecules would undergo modification to develop a greater structural complexity and specificity, finally giving rise to a "living" organism. Therefore, because of the ease with which organic compounds can be synthesized under reducing conditions, planetary surfaces may contain an abundant source of similar organic matter. However, difficulties arise in postulating steps for further organization or modification of the above synthesized organic matter into a living state. Most of the original organic matter produced in the primary reducing atmospheres of the various planets may have been quite similar. However, major variations between planets, in chemical evolution beyond the prebiotic stage, must have been the rule rather than the exception.
The primary interest in this area of research has been the realization of the possible existence of organic molecules on planetary surfaces and, particularly, Mars. Pertinent synthesis may be either biological or abiological. Research conducted in the simulation of cosmochemical synthesis has used most of the available solar spectrum. Simulation experiments devised to study the effects of these energies on the assumed early atmosphere of the Earth have yielded products that play a dominant role in molecular and biochemical organization of the cell.
Calvin ([ref.16]) irradiated water and carbon dioxide in a cyclotron, obtaining formaldehyde and formic acid. Miller ([ref.17]) found that when methane, ammonia, water, and hydrogen were subjected to a high-frequency electrical discharge, several amino acids were produced along with a variety of other organic compounds.
Corroborating experiments established that the synthesis of amino acids occurred readily. The apparent mechanism for the production of amino acids is as follows: aldehydes and hydrogen cyanide are synthesized in the gas phase by the electrical discharge. These substances react together and also together with ammonia in the water phase of the system to give hydroxy and amino nitriles, which are then hydrolyzed to hydroxy and amino acids. Among the major constituents were aspartic acid, glutamic acid, glycine, α-alanine, and β-alanine.
The "Miller-Urey" reaction mixture has been extended and several modifications introduced. Oró ([ref.18]) introduced hydrogen cyanide into the system as the primary gas component. Adenine was obtained when Oró heated a concentrated solution of hydrogen cyanide in aqueous ammonia for several days at temperatures up to 100° C. Adenine is an essential component of nucleic acids and of several important coenzymes. Guanine and urea were the two other products identified in the hydrogen cyanide reaction. Oró further obtained guanine and uracil as products of nonenzymatic reactions by using certain purine intermediates as starting materials.
Ponnamperuma ([ref.19]) also obtained adenine upon irradiation of methane, ammonia, hydrogen, and water, using a high-energy electron beam as the source of energy of irradiation. These results indicate that adenine is very readily synthesized under abiotic conditions. Adenine, among the biologically important purines and pyrimidines, has the greatest resonance energy, thus making its synthesis more likely and imparting greater radiation stability to the molecule.
The formation of adenine and guanine, the purines in RNA and DNA, by a relatively simple abiological process lends further support to the hypothesis that essential biochemical constituents of life may have originated on Earth by a gradual chemical evolution and selection. In this respect, the examination of planetary surfaces—specifically Mars—presents practical implications for current research on the problem of chemical evolution.
When Ponnamperuma et al. ([ref.14]) exposed adenine and ribose to ultraviolet light in the presence of phosphate, adenosine was produced. When the adenine and ribose were similarly exposed in the presence of the ethyl ester of polyphosphoric acid, adenosine diphosphate (ADP) and adenosine triphosphate (ATP) were produced. The abiological formation of ATP was a major stride along the path of chemical evolution, since ATP is the principal free energy source of living organisms.
Oparin ([ref.15]) postulated that α-amino acids could have been formed nonbiologically from hydrocarbons, ammonia, and hydrogen cyanide at a time when the Earth's atmosphere contained these substances in high concentrations. Oparin's hypothesis has received strong experimental support, as evidenced by the work of Miller ([ref.12]). Bernal ([ref.20]) has emphasized the role played by ultraviolet light in the formation of organic compounds at a certain stage of the Earth's evolution.
Generally it has been believed that the first proteins or foreprotein were nonbiologically formed by the polycondensation of preformed free amino acids ([ref.21]). Akabori ([ref.22]) proposed a hypothesis for the origin of the foreprotein and speculated that it must have been produced through reactions consisting of the following three steps.
The first step is the formation of aminoacetonitrile from formaldehyde, ammonia, and hydrogen cyanide.
CH2O + NH3 + HCN ————> H2N—CH2—CN + H2O
The second is the polymerization of aminoacetonitrile on a solid surface, probably absorbed on clay, followed by the hydrolysis of the polymer to polyglycine and ammonia.
x H2N—CH2—CN ————> (—NH—CH2—C—)x||NH|| + x H2OV(—NH—CH2—CO—)x + x NH3
The third step is the introduction of side chains into polyglycine by the reaction with aldehydes or with unsaturated hydrocarbons. Akabori has demonstrated experimentally the formation of cystinyl and cysteinyl residue in his above-postulated mechanism.
Fox's theory of thermal copolymerization ([ref.23]) suggests that proteins or like molecular units could have been formed in the Earth's crust, under geothermal conditions. The accumulated amino acids were heat polymerized and transported into the primary oceans for further modifications. Fox has obtained polymers consisting of all 18 amino acids usually present in proteins. The polymerization is generally done at 160° C to 200° C, although in the presence of polyphosphoric acid it can be accomplished at temperatures below 100° C. Molecular weights increased from 3600 in a proteinoid made at 160° C to 8600 in one made at 190° C.
Fox showed that when hot saturated solutions of thermal copolymers containing the 18 common amino acids were allowed to cool, large numbers of uniform, relatively firm, and elastic spherules separate. These range from 0.2µ to 60µ in diameter and are quite uniform within each preparation. Various chemical observations suggest the presence of peptide bonds in the structural organization of these proteinoids. Continuing observations of these microspheres have established further characteristics that point to the possibility of their interpretation as a kind of primitive protein macromolecule with self-organizing properties, such that a primitive form of cell, with boundary and other properties, might form.
In laboratory experiments the behavior of gram-negative and gram-positive microspheres in dilute alkali parallels that of gram-negative and gram-positive bacteria ([ref.23]). Furthermore, time-lapse studies indicate that the proteinoid microspheres undergo a septate kind of fission, mimicking cell division as shown in figure 1. Cytochemical studies show that the microsphere's boundary is membranelike in having a primitive selectivity. Electron micrographs of sections of stained microspheres also indicate the presence of a boundary.
Oparin ([ref.15]) states that the type of organization peculiar to life could only result from the evolution of a multimolecular organic system separated from its environment by a distinct boundary but constantly interacting with this environment. In his concept of coacervates as precell models, Oparin ([ref.24]) indicates that present-day protoplasm possesses a number of features similar to coacervate structure. These coacervates could represent the starting point for evolution leading to the origin of life. Moreover, in the course of their evolution the initial systems may gradually become more complex. Oparin also showed ([ref.15]) that mixing solutions of different proteins and other substances of high molecular weight produced these coacervate droplets. These droplets are characterized by the formation of a surface layer with altered structure and mechanical properties, thus providing a somewhat selective barrier in which to house a molecular system capable of replication. However, these coacervates are unstable structurally.
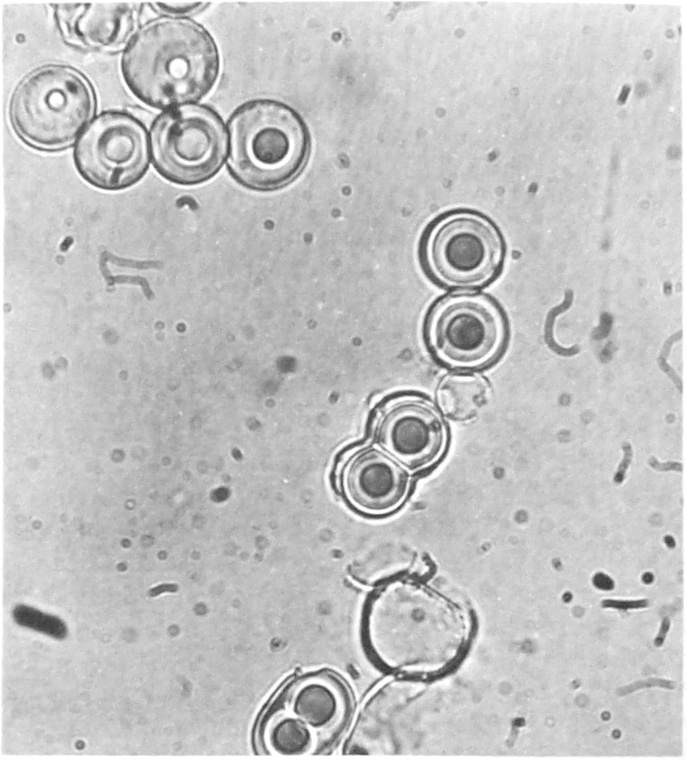
The NASA program has further provided considerable impetus for continuing research with respect to the chemical evolution of life, since its life-detection experiments may encounter prebiological molecules in their search for extraterrestrial life on other planetary surfaces.
In the area of exobiological research, the significant accomplishments to date have been—
The reconstruction of some of the pathways which may have led to the origin of life, by means of laboratory simulation of processes yielding prebiological organic molecules
The developments in experimental and theoretical biology; specifically, the role of nucleic acid-protein interactions in storage and transmission of information both within living cells and from generation to generation of cells
The suspected role of DNA in information storage and the development of new concepts of the coding mechanism in DNA that may lead to a universal biological theory embracing evolutionary, as well as homeostatic, adaptation to environment and learned behavioral systems
With the essential biochemical constituents of life and the mechanism of replication beginning to be understood, the challenge for the synthesis of living matter by abiogenic experimental techniques has become to many scientists the ultimate goal of the scientific era.
NASA has established an exobiology laboratory at Ames Research Center in addition to the sizable support of research at various academic centers of excellence for the continuation of abiogenic synthesis.
Although research on organochemical evolution is in its infancy, the data from relatively few experiments have already created an immense enthusiasm for knowledge of the biochemical pathways of evolution. This kind of research will ultimately elucidate the terrestrial evolution of life and, perhaps, the nature of life on other planetary bodies and the distribution of life in our galaxy.
This program, with its vast demands on the scientific community at large, is coordinated with related endeavors of a number of Federal agencies. It is allied with certain biochemical studies at the National Institutes of Health for the eventual elucidation of the dynamic pathways in cosmochemical synthesis of life's essential biochemical constituents.
METEORITES AND ORGANIC GEOCHEMISTRY
Meteorites
A significant area of exobiological research is the investigation of a special class of stony meteorites known as "carbonaceous chondrites." It is increasingly apparent that almost all life-detection concepts rely on the eventual analysis of the solid materials that may be available on Mars and other planetary surfaces. Cosmic dust and meteorites are two classes of material bodies that reach the Earth from outer space. The carbonaceous chondrites are the only extraterrestrial materials known to contain organic carbon.
The study of meteorites has generated an astonishing diversity of hypotheses. There is agreement at only one point: that meteorites are preserved chunks of very ancient, perhaps primordial, planetary matter and that when we are able to understand the curious structures and chemical and isotopic variations in the meteorites, we will also know a great deal about early planetary (and perhaps preplanetary) history.
Meteorites provide a more representative sample of average planetary matter than the highly differentiated crust of the Earth. Although it is known that the meteorite parent bodies ceased to be geochemically active shortly after their formation, some 4½ billion years ago, there is no consensus on the nature of the meteorite parent bodies, not even on such basic properties as size, location, and multiplicity. This is not surprising because the meteorite samples commonly available for study represent only about 10-23 to 10-26 of the parent body.
Carbonaceous Meteorites
Analysis and characterization of the chemical constituents (organic) of carbonaceous chondrites, including the possible mechanism of their formation, may be expected to improve methods of analyzing samples from the Moon and planets and of interpreting remote automated biological analyses on the planets' surfaces.
Carbon has been detected in all meteorites analyzed; however, both the amount and forms present vary considerably. Among the forms of meteorite carbon are diamond, graphite, cohenite (Fe,Ni,Co)3C, moissanite SiC, calcite CaCO3, dolomite (Ca,Mg)CO3, bruennerite (Mg,Fe)CO3. A summary of the results of carbon analyses on large numbers of meteorites is given in table I ([ref.26]).
Meteorite group |
Number analyzed |
Mean carbon content, percent by weight |
|---|---|---|
Pallasites |
10 |
0.08 |
Ureilites |
2 |
.69 |
Bronzite chondrites |
12 |
.05 |
Hypersthene chondrites |
8 |
.04 |
Enstatite chondrites |
8 |
.29 |
Carbonaceous chondrites |
16 |
2.04 |
Most meteorites possess only traces of carbon, and studies of this carbon indicate that it is composed largely of graphite, cohenite, and moissanite, with some diamond. However, studies of the carbon in the carbonaceous chondrites have failed to detect any of these forms. Some carbonates are present in a minority of the carbonaceous group, but account for only a small percentage of the total carbon (perhaps about 10 percent of the total C in type I only).
The carbonaceous chondrites contain organic carbon. The word "organic" is not used in a biological sense, merely as a chemical term to describe compounds of carbon other than carbonates, bicarbonates, and carbides. No evidence has been found of any form of carbon other than organic, except for traces of carbonates.
Various studies have demonstrated possible methods of estimating the total amount of organic matter present in meteorites. Wiik ([ref.27]) has suggested that organics can be estimated by measuring the loss of weight on ignition. Unfortunately, this method has several disadvantages and gives very low values. Corrections must be made for weight gains due to oxidation of reduced constituents, such as FeO, Fe, Ni, and Co, and for weight losses due to H2O, S, etc. The water loss is exceedingly difficult to estimate, as part comes from the combustion of organic hydrogen and part comes from the loss of mineral-bound water. The carbon also proves difficult to combust completely, and high temperatures (over 1000° C) are required for efficient conversion to CO2.
In one study the major fraction of organic matter removed proved to have a carbon content of about 47 percent ([ref.28]). Thus, if all the meteorite carbon is present as organic matter of approximately this composition, total organics must be approximately double the carbon content; that is, 2 percent by weight carbon indicates 4 percent by weight organic matter. This estimate may be too low, for Mueller ([ref.29]) has extracted a major organic fraction containing only 24 percent carbon; however, this work has not been confirmed for other meteorites.
Briggs and Mamikunian ([ref.26]) have pointed out that only 25 percent of the organic matter has been extracted, and only about 5 percent of this has been chemically characterized. Most of this 5 percent is a complex mixture of hydroxylated aromatic acids together with hydrocarbons of the aliphatic, napalicyclic, and aromatic series. Small amounts of amino acids, sugars, and fatty acids are also present.
Thus far, these chemical analyses point to an abiogenic origin for the organic matter, and no conclusive evidence exists of biological activity on the meteorite parent body. Microbiological investigations of samples of the carbonaceous chondrites have yielded only inconclusive evidence on the problem of "organized elements."
Several of these microstructures from different carbonaceous chondrites are illustrated in a paper by Mamikunian and Briggs ([ref.30]). It has been difficult to identify the organized structures, and most do not have morphologies identical to known terrestrial micro-organisms. However, they may prove to be a variety of mineral grains, droplets of organic matter and sulfur, as well as a small amount of contaminating terrestrial debris.
A comparison between the photographs of the organized elements observed in the Orgueil and Ivuna meteorites and the synthetic proteinoid microspheres observed by Fox ([ref.25]) point to similarities between the two. One inference from this finding is that the organized elements in carbonaceous chondrites were never alive but, rather, should be considered as natural experiments in molecular evolution. Also, these similarities strengthen the belief that the laboratory experiments are similar to the natural experiments in space.
In cooperation with the Smithsonian Astrophysical Observatory, NASA has a network to track meteors in the Midwest (South Dakota, Nebraska, Kansas, Oklahoma, Iowa, Missouri, and Illinois). Photographs of meteor trails are used for scientific study, and attempts are made to track and recover meteorites for examination for traces of organic material of extraterrestrial origin.
Fundamental research in terrestrial organic geochemistry has shown that ancient sediments and drill core samples subjected to organic analysis contain certain stable biochemical components of past life. This preserved record is significant not only in studies of early-life chemical pathways but also in studies of the interaction of organic matter with the geological factors. Since life on any planetary body will interact with the soil, or surface material, it is of interest to understand the relationship.
CONCEPTS FOR DETECTION OF EXTRATERRESTRIAL LIFE
It is not possible to present completely convincing evidence for the existence of extraterrestrial life. The problem often reduces to probabilities and to estimates of observational reliability. In almost all cases the evidence is optimistically considered strongly suggestive of—or, at the worst, not inconsistent with—the existence of extraterrestrial life. Alternatively, there is a pessimistic view that the evidence advanced for extraterrestrial life is unconvincing, irrelevant, or has another, nonbiological explanation.
In studies of the laboratory synthesis of life-related compounds and its significance concerning the origin of life, several results seem to suggest that organochemical synthesis is a general process, occurring perhaps on all planets which retain a reducing atmosphere. The temperature ranges must be such that precursors and reaction products are not thermally dissociated. The reaction rates for the synthesis of more complex organic molecules diminish to a negligible value when the temperature range is below 100° C.
Besides the planetary parameter of temperature, an even more fundamental necessity for a living state exists—a liquid solvent system. For terrestrial life forms, water serves this purpose. Water has this and other properties of biological significance because of hydrogen bonding between adjacent molecules in the liquid state.
Ultraviolet radiation could serve as an extraterrestrial energy source for organic synthesis. Research shows that, while an atmosphere is important, living systems can survive a wide range of ambient pressures and are little affected by a wide range of magnetic field strengths.
Oxygen is not a prerequisite for all living systems. While it is sometimes concluded that free oxygen is needed for all but the simplest organisms, less efficient metabolic processes coupled with higher food collection efficiency—or a more sluggish metabolism—would seem to do just as well. Earth is the only planet in the solar system on which molecular oxygen is known to be present in large amounts. Since plant photosynthesis is the primary source of atmospheric oxygen, it seems safe to infer that no other planet has large-scale plant photosynthesis accompanied by the production of oxygen.
The possibility of the existence of extraterrestrial life raises the important question of man's being able to detect it. Research on extraterrestrial life detection is predicated on the ability to develop ways to detect it even when the living systems are based on principles entirely different from those on Earth.
The substitution of various molecules for those of known biological significance to living organisms as we know them has been investigated; the substitution of NH2 for OH in ammonia-rich environments leads to a diverse, and biologically very promising, chemistry. The hypothesis that silicon may replace carbon does not support the construction of extraterrestrial genetics based on silicon compounds. (Silicon compounds participate in redistribution reactions which tend to maximize the randomness of silicon bonding, and the stable retention of genetic information over long time periods is thus very improbable.)
Evidence relevant to life on Mars has been summarized by Sagan (ch. 1 of [ref.10]):
The Origin of LifeIn the past decade, considerable advances have been made in our knowledge of the probable processes leading to the origin of life on Earth. A succession of laboratory experiments has shown that essentially all the organic building blocks of contemporary terrestrial organisms can be synthesized by supplying energy to a mixture of the hydrogen-rich gases of the primitive terrestrial atmosphere. It now seems likely that the laboratory synthesis of a self-replicating molecular system is only a short time away from realization. The syntheses of similar systems in the primitive terrestrial oceans must have occurred—collections of molecules which were so constructed that, by the laws of physics and chemistry, they forced the production of identical copies of themselves out of the building blocks in the surrounding medium. Such a system satisfies many of the criteria for Darwinian natural selection, and the long evolutionary path from molecule to advanced organism can then be understood. Since nothing except very general primitive atmospheric conditions and energy sources are required for such syntheses, it is possible that similar events occurred in the early history of Mars and that life may have come into being on that planet several billions of years ago. Its subsequent evolution, in response to the changing Martian environment, would have produced organisms quite different from those which now inhabit Earth.
Simulation ExperimentsExperiments have been performed in which terrestrial micro-organisms have been introduced into simulated Martian environments, with atmospheres composed of nitrogen and carbon dioxide, no oxygen, very little water, a daily temperature variation from +20° to -60° C, and high ultraviolet fluxes. It was found that in every sample of terrestrial soil used there were a few varieties of micro-organisms which easily survived on "Mars." When the local abundance of water was increased, terrestrial micro-organisms were able to grow. Indigenous Martian organisms may be even more efficient in coping with the apparent rigors of their environment. These findings underscore the necessity for sterilizing Mars entry vehicles so as not to perform accidental biological contamination of that planet and obscure the subsequent search for extraterrestrial life.
Direct Searches for Life on MarsThe early evidence for life on Mars—namely, reports of vivid green coloration and the so-called "canals"—are now known to be largely illusory. There are three major areas of contemporary investigation: visual, polarimetric, and spectrographic.
As the Martian polar ice cap recedes each spring, a wave of darkening propagates through the Martian dark areas, sharpening their outlines and increasing their contrast with the surrounding deserts. These changes occur during periods of relatively high humidity and relatively high daytime temperatures. A related dark collar, not due to simple dampening of the soil, follows the edge of the polar cap in its regression. Occasional nonseasonal changes in the form of the Martian dark regions have been observed and sometimes cover vast areas of surface.
Observations of the polarization of sunlight reflected from the Martian dark areas indicate that the small particles covering the dark areas change their size distribution in the spring, while the particles covering the bright areas do not show any analogous changes.
Finally, infrared spectroscopic observations of the Martian dark areas show three spectral features which, to date, seem to be interpretable only in terms of organic matter, the particular molecules giving rise to the absorptions being hydrocarbons and aldehydes. [However, see p. 7 and Rea et al. ([ref.9]).]
Taken together, these observations suggest, but do not conclusively prove, that the Martian dark areas are covered with small organisms composed of familiar types of organic matter, which change their size and darkness in response to the moisture and heat of the Martian spring. We have no evidence either for or against the existence of more advanced life forms. There is much more information which can be garnered from the ground, balloons, Earth satellites, Mars flybys, and Mars orbiters, but the critical tests for life on Mars can only be made from landing vehicles equipped with experimental packages....
Results of Kaplan et al. ([ref.31]) indicate that Mars has no detectable oxygen, but does contain small amounts of water vapor, more abundant carbon dioxide, possibly a large surface flux of solar ultraviolet radiation, and estimated daily temperature variations of 100° C at many latitudes. Studies have shown that terrestrial micro-organisms can survive these extremely harsh environments. Furthermore, a variety of physiological and ecological adaptations might enable the biota to survive the low nighttime temperatures and intracellular ice crystallization.
Less evidence is available to support the possibility of extraterrestrial life on other planets. The Moon has no atmosphere, and extremes of temperature characterize its surface. However, the Moon could have a layer of subsurface permafrost beneath which liquid water might be trapped. The temperatures of these strata might be biologically moderate.
Studies by Davis and Libby ([ref.32]) on the atmosphere of Jupiter support the possibility of the production of organic matter in its atmosphere in a manner analogous to the processes which may have led to the synthesis of organic molecules in the Earth's early history. It is difficult to assess the possibility that life has evolved on Jupiter during the 4- or 5-billion-year period in which the planet has retained a reducing atmosphere.
The question of extraterrestrial life and of the origin of life is interwoven. Discovery of the first and analysis of its nature may very well elucidate the second.
The oldest form of fossil known today is that of a microscopic plant similar in form to common algae found in ponds and lakes. Scientists know that similar organisms flourished in the ancient seas over 2 billion years ago. However, since algae are a relatively complex form of life, life in some simpler form could have originated much earlier. Organic material similar to that found in modern organisms can be detected in these ancient deposits as well as in much older Precambrian rocks.
Although the planets now have differing atmospheres, in their early stages the atmospheres of all the planets may have been essentially the same. The most widely held theory of the origin of the solar system states that the planets were formed from vast clouds of material containing the elements in their cosmic distribution.
It is believed that the synthesis of organic compounds preceding the origin of life on Earth occurred before its atmosphere was transformed from hydrogen and hydrides to oxygen and nitrogen. This theory is supported by laboratory experiments of Calvin ([ref.16]), Miller ([ref.33]), and Oró ([ref.34]).
The Earth's present atmosphere consists of nitrogen and oxygen in addition to relatively small amounts of other gases; most of the oxygen is of biological origin. Some of the atmospheric gases, in spite of their low amounts, are crucial for life. The ultraviolet-absorbing ozone in the upper atmosphere and carbon dioxide are examples of such gases.
Significant in the search for extraterrestrial life are the data (e.g., planet's temperature) transmitted by Mariner II, which was launched from Cape Canaveral on August 27, 1962, and flew past Venus on December 14, 1962. Mariner II's measurements showed temperatures on the surface of Venus of the order of 800° F, too hot for life as known on Earth.
The question "Is life limited to this planet?" can be considered on a statistical basis. Although the size of the sample (one planet) is small, the statistical argument for life elsewhere is believed by many to be very strong. While Mars is generally considered the only other likely habitat of life in our solar system, Shapley ([ref.35]) has calculated that more than 100 million stars have planets sufficiently similar in composition and environment to Earth to support life. Of course, yet unknown factors may significantly reduce or even eliminate this probability.
SPACECRAFT STERILIZATION
The search for extraterrestrial life with unmanned space probes requires the total sterilization of the landing capsule and its contents. Scientists agree that terrestrial organisms released on other planets would interfere with exobiological explorations (refs. [ref.36]-[ref.43]). Any flight that infects a planet with terrestrial life will compromise a scientific opportunity of almost unequaled proportions. Studies on microbiological survival in simulated deep-space conditions (low temperature, high ultraviolet flux, and low dose levels of ionizing radiation) indicate that these conditions will not sterilize contaminated spacecraft (refs. [ref.44]-[ref.48]). Furthermore, many terrestrial sporeformers and some vegetative bacteria, especially those with anaerobic growth capabilities, readily survive in simulated Martian environments (refs. [ref.49]-[ref.54]). It has been estimated that a single micro-organism with a replication time of 30 days could, in 8 years of such replication, equal in number the bacterial population of the Earth. This potential could result not only in competition with any Martian life, but in drastic changes in the geochemical and atmospheric characteristics of the planet. To avoid such a disaster, certainly the first, and probably many succeeding landers on Mars, must be sterile—devoid of terrestrial life ([ref.55]). Since the space environment will not in itself kill all life aboard, the lander must leave the Earth in a sterile condition.
The sterility of an object implies the complete absence of life. The presence of life or the lack of sterility may be proven; but the absence of life or sterility cannot be proven, for the one viable organism that negates sterility may remain undetected. Many industrial products which must be guaranteed as sterile cannot be tested for sterility in a nondestructive manner. A similar situation exists in determining the sterility of a spacecraft. Certification of sterility—based on experience with the sterilizing process used, knowledge of the kinetics of the death of micro-organisms, and computation of the probability of a survivor from assays for sterility—is the only accurate approach to defining the sterility of such treated items.
Macroscopic life can be readily detected and kept from or removed from the spacecraft, but the detection and removal of microscopic and submicroscopic life is an extremely difficult task. The destruction of micro-organisms can be achieved by various chemical and physical procedures. Sterilizing agents have been evaluated not only for their ability to kill microbial life on surfaces and sealed inside components, but also for the agents' effects on spacecraft reliability as well (refs. [ref.56]-[ref.59]). Of the available agents, only heat and radiation will penetrate solid materials. Radiation is expensive, hazardous, difficult to control, and apparently damages more materials than does heat. Heat, therefore, has been selected as the primary method of spacecraft sterilization and will be used, except in specific instances where radiation may prove to be less detrimental to the reliability of critical parts ([ref.60]).
The sterilization of spacecraft is a difficult problem if flight reliability is not to be impaired. The development of heat-resistant parts will enable the design and manufacture of a heat-sterilizable spacecraft. Without careful microbiological monitoring of manufacture and assembly procedures, many bacteria could be trapped in parts and subassemblies. To permit sterilization at the lowest temperature-time regimen that will insure kill of all organisms, the microbiological load inside all parts and subassemblies must be held to a minimum.
The role of industrial clean rooms in reducing the biological load on spacecraft is currently being defined. NASA-supported studies indicate that biological contamination in industrial clean rooms for extended time periods is about 1 logarithm less (tenfold reduction), compared with conditions in a well-operated microbiological laboratory ([ref.61]). With the use of clean-room techniques and periodic decontamination by low heat cycles or ethylene oxide treatment, it should be possible to bring a spacecraft to the point of sterilization with about 106 organisms on board ([ref.60]).
The sterilization goal established for Mars landers is a probability of less than 1 in 10 000 (10-4) that a single viable organism will be present on the spacecraft. Laboratory studies of the kinetics of dry-heat kill of resistant organisms show that at 135° C the number of bacterial spores can be reduced 1 logarithm (90 percent) for every 2 hours of exposure (refs. [ref.58] and [ref.62]). The reduction in microbial count needed is the logarithm of the maximum number on the spacecraft (106) plus the logarithm of the reciprocal of the probability of a survivor (104), or a total of 10 logarithms of reduction in microbial count. Thus, with an additional 2 logarithms added as a safety factor, a total of 12 logarithms of reduction in count has been accepted as a safe value which can be achieved by a dry-heat treatment of 135° C for 24 hours. This is the heat cycle that is currently under study and being developed for use in spacecraft sterilization ([ref.60]). However, other heat treatments at temperatures as low as 105° C for periods of 300 hours or longer are under study ([ref.63]).
Based on results to date, it is reasonable to believe that a full complement of heat-sterilizable hardware will be available when needed for planetary exploration. Every effort is being made to improve the state of the art to a point where spacecraft can not only withstand sterilization temperatures, but will be even more reliable than the present state-of-the-art hardware that is not heated.
chapter 3
BIOLOGICAL EFFECTS OF WEIGHTLESSNESS AND ZERO GRAVITY
High priority has been given to studies of weightlessness. Gravity is one of the most fundamental forces that acts on living organisms, and all life on Earth except the smallest appears to be oriented with respect to gravity, although certain organisms are more responsive to it than others. The gravity force on Earth is 1 g, but this force may be experimentally varied from zero g, or weightlessness, to many thousands of g's.
Zero gravity or decreased gravity occurs during freefall, in parabolic trajectory, or during orbit around the Earth. Gravitational force decreases by the square of the distance away from the Earth's center. It is reduced about 5 percent at about 200 nautical miles' altitude. Gravitational force greater than 1 g can be obtained by acceleration, deceleration, or impact. It also can be increased by using a centrifuge which adds a radial acceleration vector to the 1 g of Earth.
On the ground, the biological effects of gravity have been studied at 1 g, and experimentally, forces of many g have been produced. In addition, modifications of the effects of the 1-g force have been induced by suspension of the organism in water or by horizontal immobilization of an erect animal such as man. The biological effects of such modification have been of significant value in understanding some of the possible consequences of human exposure to the zero-g environment of space.
Weightlessness in an Earth-orbiting satellite occurs when the continuous acceleration of Earth's gravity is exactly counterbalanced by the continuous radial acceleration of the satellite. In such a weightless state, organisms are liberated from their natural and continuous exertion against 1 g, but this liberation may carry with it certain serious physical penalties.
Some of the physical processes which probably have the greatest biological effects are (1) convective flow of fluid, e.g., protoplasmic streaming, transport of nutrient materials, oxygen, waste products, and CO2 from the immediate environment of the cell, and (2) sedimentation occurring within cells; substances of higher density sediment in a gravitational field, and those of lighter density rise. A separation of particles of different densities probably occurs. The removal of gravity would change a distribution of particles like mitochondria by 10 percent ([ref.64]).
Gravity has effects on the physical processes involved in mitosis and meiosis. Study under weightlessness might contribute to our understanding of the general cellular information-relay process.
A gravitational effect is known in the embryonic development of the frog Rana sylvatica. After fertilization, the eggs rotate in the gravitational field so that the black animal hemisphere is uppermost. Development becomes abnormal if this position is disturbed. If the egg is inverted following the first cleavage and held in this position, two abnormal animals result, united like Siamese twins. This phenomenon appears to be related to the gravitational separation of low- and high-density components of the egg. The size of the egg is about 1 to 2 mm and is suspended in water of about the same density. This system is very sensitive to gravity; and, under weightlessness, the separation of different density components might be irregular, leading to aberrant development. When certain aquatic insect eggs are inverted, subsequent development results in shortened abnormal larvae.
The directional growth of plant shoots and plant roots is probably due to this sedimentation phenomenon, particularly the effect on movement of auxins ([ref.65]).
Free convection flow is a major transport process, and under its influence the mixing of substances is much more effective than when diffusion operates alone. Free convection flow is a macroscopic phenomenon which increases not only with g, but varies also approximately with the five-fourths power of the bulk concentration involved. Whether or not convection is important at the microscopic level remains an experimentally unsolved question. The Grashoff number limits free convection to the macroscopic domain. It would appear in weightlessness that the contribution of free convective flow would be small and that only diffusion should occur. This phenomenon would cause equilibration to occur much more slowly than that occurring with free convection and diffusion. The absence of convective transfer raises a problem as to how nutrients may be obtained and waste products removed in living cells during weightlessness. In a liquid substrate, nutrients and oxygen would be depleted, and waste products would accumulate around the cell.
Absence of gravity may have far-reaching consequences in the homeostatic aspects of cell physiology. The outstanding characteristics of living cells which are most likely to be influenced by the absence of gravity are the ability of the cell to maintain its cytoplasmic membrane in a functional state, the capacity of the cell to perform its normal functions during the mitotic cycle, and the capacity of the cytoplasm to maintain the constant reversibility of its sol-gel system ([ref.66]).
Two-phase systems, e.g., air-in-water and air-in-oil, possess entirely different characteristics at zero g than at 1 g. These physical differences in phase interaction could well be suspected of interfering with the orientation and flow pattern of cell constituents, thus hindering the cellular processes involved in the movement, metabolism, and storage of nutrients and waste.
On the basis of theoretical calculations, weightlessness can be expected to have some effect even on one individual cell if its size exceeds 10 microns in diameter ([ref.64]). Cell colonies might be affected. In larger cells there may be a redistribution of enzyme-forming systems which give rise to polarization. The low surface tension of the cell membrane lends itself to hydrostatic stress distortion, implying an alteration in permeability and thus an almost certain alteration of cell properties under low gravity conditions.
Another aspect of gravity that affects the growth and development of living organisms is the directionality of the gravitational field. In fact, some plants are so sensitive that they are able to direct their growth with as little stimulus as a 1×10-6 gravitational field. Investigations of plant growth in altered gravitational fields are underway at Argonne National Laboratory and Dartmouth College.
The Argonne Laboratory has designed and developed a 4-pi, or omnidirectional, clinostat. By rotating a plant so that the force of gravity is distributed evenly over all possible directions, the directional effects of gravity are eliminated, simulating some aspects of the zero-g state. It was shown that certain plants grew more slowly and had fewer and smaller leaves, while others had about 25 percent greater replication of fronds and had greater elongation of certain plant parts. It will be extremely interesting to compare these effects under zero-g conditions in orbiting spacecraft.
The effect of gravity in transporting growth hormones in plants has been demonstrated at Dartmouth College using radiocarbon-labeled growth hormones. Plant geotropisms and growth movements have been studied and biosatellite experiments developed.
Anatomy is considered a derivative adaptation to gravity ([ref.67]). A large background of plant research exists on the effect of orientation on plant responses. Information from clinostat experiments is considered susceptible of extrapolation to low gravity conditions because the threshold period for gravitational triggering is relatively long.
Once over critical minimum dimensions, the major effects of low gravity would be assumed to occur in those heterocellular organisms that develop in more or less fixed orientation with respect to terrestrial gravity and which respond to changes in orientation with relatively long induction periods; these are the higher plant orders. On the other extreme are the complex primates which respond rapidly, but whose multiplicity of organs and correlative mechanisms are susceptible to malfunction and disorganization. It may be suggested that the heterocellular lower plants and invertebrates will be less affected. Perturbations of the environment to which the experimental organism is exposed must be limited or controlled to reduce uncertainties in interpretation of the results. At the same time, the introduction of known perturbations may assist in isolating the effects due solely to gravity. Study of de novo differentiation and other phenomena immediately after syngamy may be of particular importance. Study of anatomical changes after exposure of the organism to low gravity is important.
BIOLOGICAL EFFECTS OF SPACE RADIATION1
Radiation sources in space are of three types: galactic cosmic radiation, Van Allen belts, and solar flares with an intense proton flux. Cosmic radiation has higher energy levels than radiation produced by manmade accelerators.
The Panel on Radiation Biology, while recognizing the need for radiobiological studies of an applied nature with reference to manned flight programs, stated that it would be shortsighted for the United States to confine its efforts to the solution of immediate problems since, in the long run, successful exploration of space will be aided by the contributions of basic research. Both the immediate biological research program and the continuing program for basic studies should be built upon the large body of existing knowledge of radiation effects. The attitude that all radiobiological experiments need be repeated in the space environment should be resolutely rejected. Since fundamental radiobiology cannot be performed easily in space, it has been recommended that, wherever possible, these investigations be carried out in ground laboratories in preference to flying laboratories.
Space environment does vary from the terrestrial environment, but the variations are not so great as to lead to the expectation of strikingly different biological effects of radiation in space. However, it is conceivable that radiations whose effects are well known under terrestrial conditions may have some unsuspected biological effects when combined with unusual features of the space environment: e.g., zero g. Previous space radiobiological studies have depended solely on very low and inaccurately measured doses of ambient space radiation. It has been difficult to distinguish between the observed response levels and the random noise; thus, experiments have been inconclusive.
Biological Effects of Heavy Ions and Mesons
The biological effects of heavy ions (especially Z>2) and mesons are of specific interest to space radiobiology.
Controlled Radiobiological Experiments in Space
There is the remote possibility that the radiobiological response may be modified by factors as yet unknown and perhaps not susceptible to terrestrial study. Experiments have been designed to settle this matter including the exposure of biological materials during space flight which meet the following criteria of reliability: (1) the use of well-known biological systems, e.g., mutation induction or chromosome breakage; (2) the use of a sufficient number of individuals in the experiment to guarantee statistical precision on the results; (3) the exposure of the system to known quantities and qualities of radiation; (4) the use of adequate controls.
High-altitude balloon ascents of the 1930's initiated study of the biological effects of cosmic rays. They were limited to the exploration of secondary cosmic radiation effects. After World War II, the research extended to the use of V-2 rockets fired from the White Sands Proving Ground. Interest returned to balloons and a significant program was underway by 1950, first using mice and then hamsters, fruit flies, cats, and dogs. These flights gave no evidence of radiation damage. However, it was realized that the flights were too far south to obtain a significant exposure, and more northerly flights began in 1953. Mice and guinea pigs were flown on these later flights. Chase ([ref.68]) showed the most unequivocal results to that time, a statistically significant increase in light hairs on black animals and the streaks of white hair up to 10 times wider than expected. Brain lesions were detected in the guinea pigs flown on Man High in 1957. Many other types of biological material were sent aloft in an effort to further corroborate existing information and to investigate genetic and developmental effects of cosmic radiation.
From the earlier V-2 rocket flights to the Jupiter missile launchings of the monkeys Able and Baker, cosmic-ray research was continued, but the short flight durations of these vehicles did not provide substantial information. The USAF Discoverer satellite program has given impetus to cosmic-ray research and provided for longer "staytimes."
It has been difficult to separate radiation effects from other space-flight factors: therefore, some of the alterations observed are still subject to debate. Vibration, acceleration, and weightlessness appear to be the three most important additional parameters. Measurements of radiation dosage have been made by chemical and photographic dosimetry, ion chambers, and biological dosimetry. All evidence to date indicates that radiation exposure levels are not hazardous to man at present orbital altitudes up to 200 nautical miles. Most biological materials flown so far have been for the express purpose of investigating space-radiation levels and effects. The biological materials have ranged from tissue cultures to entire organisms and from phage and bacterial cells to man. The studies have required much of the space and weight resources allotted biology by the U.S.S.R. and the United States. They have been accompanied by ground-based controls.
The Vostok series provided the following data:
A small, but statistically significant, increase was observed in the percentage of chromosome aberrations in the rootlet cells of air-dried wheat and pea seeds after germination. In this case only, the increase did not depend on flight duration.
Lysogenic bacteria exhibited an increase of genetic alterations and increased phage production. Length of flight was associated with increased bacteriophage production by the lysogenic bacteria. There was an increase of recessive lethals coupled with nonconvergence of chromosomes (sex linked) in the fruit fly. A stimulation of cell division in wheat and pea seeds was observed. Cultures of human cells exposed to space-flight factors did not differ significantly from terrestrial controls with respect to such indicators as proliferation rate, percentage of mortality and morphological, antigenic, and cultural properties. Repeated flights of the identical HeLa cells revealed that there was a longer latent period for restoration of growth capacity than in cells carried into space once or not flown at all.
The most definite radiation effects observed were only revealed in genetic tests. No harmful influence on those characteristics affecting the viability of the organism has been discovered.
The Air Force Discoverer series launched from the west coast had a few successful flights incorporating organisms. With severe environmental stress and long recovery times, data on radiation exposure were equivocal up to Discoverer XVII and XVIII when cultures of human tissue were flown, recovered, and assessed for radiation exposure effects. Comparison with ground-based controls revealed no measurable differences.
Radiation dosimetry from the Mercury series established that minimal exposures were encountered at those orbital altitudes. A typical example is the MA-8 flight of W. M. Schirra, Jr., during which the body surface dosage was less than 30 millirads.
NASA has supported fundamental radiation studies at the Oak Ridge National Laboratory and the Lawrence Radiation Laboratory. Emphasis has been placed on the biological effects of high-energy proton radiation and particulate radiation from accelerators.
At the NASA Ames Research Center extensive fundamental studies are being carried out on the effects of radiation, especially in the nervous system. It has been demonstrated that deposits accumulate in the brain following exposure to large doses of ionizing particle radiation as well as after X-irradiation. These deposits, referred to as a "chemical lesion," result from an accumulation of glycogen. The formation of these deposits during exposure to large doses of X-irradiation was not increased in environments of 99.5 percent oxygen and increased atmospheric pressure.
SIMULATION OF PLANETARY (MARTIAN) ENVIRONMENTS
Attempts have been made to simulate to some degree the various parameters of the Martian environment, such as atmospheric composition, pressure, radiation flux, temperatures, and the day-night as well as seasonal cycles. Certain factors for Mars cannot yet be simulated, such as soil composition, gravitational field, magnetic field, and electrical field.
Caution is required in interpreting all simulation experiments. How Earth organisms respond to simulated Martian environments probably has nothing to do with life on Mars, but these experiments may show whether or not anything in the environment of Mars makes life as we know it impossible. We must expect that on Mars, life will have evolved and have adapted over long periods of time under conditions which are quite different from conditions on Earth. The simulation experiments also provide some information about the possibility of contaminating the planet Mars, or any planet, with organisms from Earth. In addition, they give us some clues about the possibilities of adaptation and evolution of life under these conditions.
From an evolutionary point of view, if life has developed on Mars, we expect it to have evolved at least to a microbial stage. On Earth, micro-organisms are the most ubiquitous and numerous forms of life. This fact should be considered in studying extraterrestrial bodies.
Micro-organisms have been selected as the best test organisms, and bacteria and fungi have been used because they are durable and easy to grow. Also, because of their rapid growth, many generations can be studied in a relatively short period of time. The organisms include chemoautotrophic bacteria, which are able to synthesize their cell constituents from carbon dioxide by energy derived from inorganic reactions; anaerobic bacteria, which grow only in the absence of molecular oxygen; photoautotrophic plants such as algae, lichens, and more complex seed plants; and small terrestrial animals.
Organisms have been collected from tundra, desert, hot springs, alpine, and saline habitats to obtain species with specialized capabilities to conserve water, balance osmotic discrepancies, store gases, accommodate to temperature extremes, and otherwise meet stresses. An attempt is made in these simulation experiments to extend these processes across the possible overlapping microenvironments which Earth and Mars may share.
Scientists have developed various special environmental simulators, including "Mars jars" and "Marsariums." These have made possible controlled temperatures, atmospheres, pressures, water activities, and soil conditions for duplicating assumed Martian surface. A complex simulator, developed by Young et al. ([ref.52]), reproduces the formation of a permafrost layer with some water tied up in the form of ice beneath the soil surface. This simulator serves as a model to study the wave of darkening, thus supporting the hypothesis that the pole-to-equator wave of darkening is correlated with the availability of subsurface water. The simulator is a heavily insulated 2-cu-ft capacity chamber with an internal pressure of 0.1 atm. The chamber contains a soil mixture of limonite and sand and an atmosphere of carbon dioxide and nitrogen. With the use of a liquid nitrogen heat exchanger at one end and an external battery of infrared lamps at the other end, the temperature simulates that of Mars from pole to equator. Thermocouples throughout the soil monitor the temperatures in the chamber.
Zhukova and Kondratyev ([ref.69]) designed a structure measuring 100×150×180 cm. Micro-organisms were placed at the surface of a copper bar made in a special groove separated by glass cloth. Copper was selected as one of the best heat-conduction materials permitting a rapid change of temperature. The lower end of the bar was immersed into a mixture of dry ice and ethyl alcohol, which made it possible to create a temperature of -60° C. Heating was performed by an incandescent spiral.
As the knowledge concerning the Martian environment becomes more refined, scientists can more accurately simulate this environment under controlled conditions in the laboratory. Determination of the effects of the Martian environment on Earth organisms will permit better theorization on the forms of life we might find on Mars and will permit us to estimate the potential survival of Earth contaminants on Mars.
However, until the environmental conditions of Mars are defined more accurately, the experiments must be changed continually to fit newly determined conditions. Therefore, existing simulation data are made less valid for comparison. The data resulting from the simulation experiments for Mars have been compiled in table II, and the experiments are summarized below.
The earliest simulation studies were carried out by the Air Force, and the studies during the past 6 years have been supported by NASA. Recently, these studies have received less support or have been terminated in favor of critical studies on the effects of biologically important environmental extreme factors on Earth organisms. These critical studies permit establishing the extreme environmental factor parameters in which Earth life can grow or survive. These data will have valuable application to the consideration of life on any planet, to the design of life-detection instruments, to the sterilization of space vehicles, and to the problem of contamination of planets.
Some exploratory experimental studies are in progress to study the capabilities of organisms to grow under the assumed conditions on Jupiter. These include studies at high pressure with liquid ammonia, methane, and other reducing compounds.
Early experiments simulating Martian conditions using soil bacteria were carried out by Davis and Fulton ([ref.70]) at the Air Force School of Aviation Medicine, San Antonio, Tex. Mixed populations of soil bacteria were put in "Mars jars" with the following conditions: 65-mm Hg pressure, 1 percent water or less, nitrogen atmosphere, sandstone-lava soil, and a temperature day-night cycle of +25° to -25° C. The moisture was controlled by desiccating the soil and adding a given amount of water. Experiments, conducted up to 10 months, demonstrated that obligate aerobes died quickly. The anaerobes and sporeformers survived. Although a small increase in the total number of organisms indicated growth, the increases in the number of bacteria may have been due to breaking up clumps of dirt.
Roberts and Irvine ([ref.71]) reported that, in a simulated Martian environment, colony counts of a sporeforming bacterium, Bacillus cereus, increased when 8 percent moisture was added. Moisture was considered more important than temperature or atmospheric gases inasmuch as a simulated Martian microenvironment containing 8 percent moisture permitted germination and growth of endospores of Clostridium sporogenes. Increases in colony counts of Bacillus cereus appeared to be influenced by temperature cycling ([ref.72]).
Species |
Survival, months |
Moisture |
Temperature, °C |
Atmospheric pressure, mm Hg |
N2, percent |
CO2, percent |
Substrate |
|---|---|---|---|---|---|---|---|
Conditions on Mars: |
14µ±7µ |
-70 to +30 |
85, 25±15, 11 |
3 to 30 |
|||
|
6 |
Low, (CaSO4) |
-60 to +20 |
76 |
95 |
5 |
Air-dried soil |
|
6 |
Low, (CaSO4) |
-60 to +20 |
76 |
95 |
5 |
Air-dried soil |
|
Growth |
Very wet |
-75 to +25 |
760 |
100 |
(?) |
Difco infusion broth |
|
10 |
1 percent or less |
-25 to +25 |
65 |
100 |
(?) |
Soil |
Bacillus cereus |
2 |
0.5 percent soil |
-25 to +25 |
65 |
94 |
2.21 |
Sandstone soil |
Clostridium sporogenes |
1 (growth) |
8.4 percent |
-25 to +25 |
65 |
94 |
2 |
Enriched soil |
Clostridium botulinum |
10 |
Lyophilized |
-25 to +25 |
65 |
95 |
0 to 0.5 |
Lava soil |
Klebsiella pneumoniae |
6 |
Lyophilized |
-25 to +25 |
65 |
95 |
0 to 0.5 |
Lava soil |
Bacillus subtilis var. globigii |
4 |
2 percent |
-25 to +25 |
85 |
95 |
0.3 |
Media |
Sarcina aurantiaca |
4 |
0.5 percent |
-25 to +25 |
85 |
95 |
0.3 |
Desert soil |
Clostridium tetani |
2 or less |
1 percent |
-60 to +25 |
85 |
95 |
0.3 |
Soil |
Aspergillus niger |
Over 6 hr |
Very dry |
-60 to +25 |
76 |
95.5 |
0.25 |
Glass cloth on copper bar |
Aspergillus oryzae |
Over 6 hr |
Very dry |
-60 to +25 |
76 |
95.5 |
0.25 |
Do. |
Mucor plumbeus |
Over 6 hr |
Very dry |
-60 to +25 |
76 |
95.5 |
0.25 |
Do. |
Rhodotorula rubra |
Over 6 hr |
Very dry |
-60 to +25 |
76 |
95.5 |
0.25 |
Do. |
Pea, bean, tomato, rye, sorghum, rice. |
0.3 |
Moist |
+25 |
75 |
100 |
0 |
Filter paper |
Winter rye |
0.6 |
Moist |
-10 to +23 |
76 |
98 |
0.24 |
Soil |
Studies of the effects of simulated Martian environments on sporeforming anaerobic bacteria were carried out by Hawrylewicz et al. ([ref.49]). They showed that the encapsulated facultative anaerobe, Klebsiella pneumoniae, survived under simulated Martian atmosphere for 6 to 8 months, but were less virulent than the freshly isolated organisms. Spores of the anaerobe Clostridium botulinum survived 10 months in the simulator. Hagen et al. ([ref.53]) found that the addition of moisture to dry-simulated Martian soil did not improve the survival of Bacillus subtilis or Pseudomonas aeruginosa. Bacillus cereus spores survived, with added organic medium plus moisture, but no germination of the spores resulted.
Hawrylewicz et al. ([ref.49]) put rocks from Antarctica bearing various lichens in simulated Martian conditions in a large desiccator. They found that the algal portion of a lichen, Trebouxia erici, showed only slight resistance to the Martian environment. They also pointed out the effect moisture had on the physical condition of lichens. The undersurface of a lichen has great water-absorbing capability, and the slightest amount of moisture on a rock surface is absorbed by the lichen which can turn green in 15 minutes.
Scher et al. ([ref.51]) exposed desert soils to simulated environmental conditions and diurnal cycles of Mars. The atmosphere consisted of 95 percent nitrogen and 5 percent carbon dioxide (no oxygen) and was dried, using calcium sulfate as a desiccant. The total atmospheric pressure was 0.1 atm. The temperature ranged from -60° to +20° C in 24-hour cycles. One hour was spent at the maximum and at the minimum temperatures. The chambers were irradiated with ultraviolet, 2537 Å, with a dose of 109 ergs/cm2, which is comparable to a daily dose found on Mars, and easily exceeds the mean lethal dose for unprotected bacteria. Soil aliquots were removed weekly and incubated at 30° C. The scoring was done both aerobically and anaerobically. Sporeforming obligate and facultative anaerobes, including Clostridium, Bacillus, and Planosarcina, and nonsporeforming facultative anaerobes, including Pseudomonas and Rhodopseudomonas, were found. The experimental chambers were frozen and thawed cyclically up to 6 months. Organisms that were able to survive the first freeze-thaw cycle were able to survive the entire experiment. The ultraviolet irradiation did not kill subsurface organisms, and a thin layer of soil served as an ultraviolet shield. All of the samples showed survivors.
Young et al. ([ref.52]) assumed that water is present on Mars, at least in microenvironments, and that nutrients would be available. The primary objective of their experiments was to determine the likelihood of contaminating Mars with Earth organisms should a space probe from Earth encounter an optimum microenvironment in terms of water and nutrients. The experiments used bacteria in liquid nutrient media. The environment consisted of a carbon dioxide-nitrogen atmosphere, and the temperature cycling was -70° to +25° C, with a maximum time above freezing of 4½ hours. Aerobacter aerogenes and Pseudomonas sp. grew in nutrient medium under Martian freezing and thawing cycles. Atmospheric pressure was not a significant factor in the growth of bacteria under these conditions.
Silverman et al. ([ref.47]) studied bacteria and a fungus under extreme—but not "Martian"—conditions. Spores of five test organisms (B. subtilis var. niger, B. megaterium, B. stearothermophilus, Clostridium sporogenes, and Aspergillus niger) and soils were exposed while under ultrahigh vacuum to temperatures of from -190° to +170° C for 4 to 5 days. Up to 25° C there was no loss in viability; at higher temperatures, differences in resistivity were observed. At 88° C, only B. subtilis and A. niger survived in appreciable numbers; at 107° C, only A. niger spores survived; none were recoverable after exposure to 120° C. B. subtilis survived at atmospheric pressure and 90° C for 5 days, but none of the other spores were viable alter 2 days. Four groups of soil organisms (mesophilic, aerobic, and anaerobic bacteria, molds, and actinomycetes) were similarly tested in the vacuum chamber. From one sample only actinomycetes survived 120° C, while one other soil sample yielded viable bacteria after exposure to 170° C. Several organisms resisted 120° C in ultrahigh vacuum for 4 to 5 days. When irradiated with gamma rays from a cobalt 60 source, differences were observed between vacuum-dried spores irradiated while under vacuum and those exposed to air immediately before irradiation. A reduction of from one-third to one-ninth of the viability of spores irradiated in vacuum occurred with vacuum-treated spores irradiated in air.
Siegel et al. ([ref.73]), in approximate simulations of Martian environments, studied tolerances of certain seed plants, such as cucumbers, corn, and winter rye, to low temperatures and lowered oxygen tensions. Lowered oxygen tensions enhanced the resistance of seedlings, particularly cucumber and rye to freezing, and lowered the minimum temperature required for germination. Germination of seeds in the absence of liquid water has also been studied. In this case, seeds of xerophytes have been suspended in air at 75-mm Hg pressure above water. The air was thus saturated. Germination was slow but did occur.
Siegel et al. (refs. [ref.73] and [ref.74]) found that the growth rate of several higher plants was enhanced by certain gases usually thought to be toxic, such as N2O. This finding is significant inasmuch as the presence of nitrogen oxides in the Martian atmosphere has been cited as evidence for the nonexistence of plants on that planet by Kiess et al. ([ref.75]). Exploratory survival tests showed that various mature plants, as well as the larvae, pupae, and adult specimens of a coleopteran insect, were undamaged when exposed to at least 40 hours of an atmosphere containing 96.5 percent N2O, 0.7 percent O2, and 2.8 percent N2.
Lichens are of interest because of their ability to survive and thrive under extreme environmental conditions on Earth. Biological activity of slow-growing lichens was detected by metabolic gas exchange, CO2 detection being especially convenient. Siegel points out that this method is sensitive and nondestructive, to be preferred to staining techniques, which at present are limited because they are only semiquantitative, subjective, and destructive of the lichen.
A Russian study of simulated planetary environments has been performed with good simulation but for periods of only 2 to 6 hours. Comments on simulation experiments made by Zhukova and Kondratyev ([ref.69]) are presented as follows:
On the basis of modern conceptions on Martian conditions it is difficult to imagine that higher forms of animals or plants exist on the planet. A Martian change of seasons similar to that of our planet empowers us to think that there is a circulation of an organic substance on Mars, which cannot exist without participation of microbic forms of life. Microorganisms are the most probable inhabitants of Mars although the possibility is not excluded that their physiological features will be very specific. That is why the solution of the problem concerning the character of life on Mars is of exceptional interest. But still the answer to this question can be verified only by simulating Martian conditions, taking into account the information obtained from astrophysicists.
Experiments aimed at creating artificial Martian climatic conditions have been started quite recently; their number is not large since they cannot be combined with the results of numerous experiments investigating the effect of extreme factors on microorganisms. The result of the effect of such physicochemical parameters of the medium as pressure, sharp temperature changes, the absence of oxygen and insolation, depends on their combination and simultaneity. These examples convincingly show that while simulating Martian conditions one should strive to the most comprehensive complex of simultaneously acting factors. The creation of individual climatic parameters acting successively leads to absolutely different, often opposite results. It should be mentioned also that refusal to imitate insolation and the performance of experiments with specimens of soil which itself has protective effect on cells of microorganisms, but not with pure culture of bacteria, are usual shortcomings in the bulk of studies on this problem.
It appears that organisms from Earth might survive in large numbers when introduced to Martian environment. Whether these organisms will be capable of growth and explosive contamination of the planet in a biological sense or not is highly questionable. The likelihood of an organism from Earth finding ideal conditions for growth on Mars seems extremely low. However, the likelihood of an organism from Earth serving as a contaminant for any life-detection device flown to Mars for the purpose of searching out carbon-based life is considerably higher. The chance that life has originated and evolved on Mars is a completely separate question and much more difficult to answer.
It would be interesting to attempt to determine possible evolutionary trends which might occur on a planet by means of selection of organisms in a simulated planetary environment. Rapid genetic selection combined with radiation and chemicals to speed up mutation rate under these conditions should reveal possible evolutionary trends under the planetary environmental conditions. This could be attempted after the planetary environments are more accurately defined.
EXTREME AND LIMITING ENVIRONMENTAL PARAMETERS OF LIFE
The question of the existence of extraterrestrial life is one of the most important and interesting biological questions facing mankind and has been the subject of much controversial discussion and conjecture. Many of the quantitative, and even qualitative, environmental constituents of the planets also are still subjects of controversy and speculation. Best guesses about a relatively unknown planetary environment, combined with lack of information about the capabilities of Earth life to grow in extreme environments, do not provide the basis for making informed scientific estimates.
Life on Earth is usually considered to be relatively limited in its ability to grow, reproduce, or survive in extreme environmental conditions. While many common plants and animals (including man) are quite sensitive to, or incapable of, surviving severe chemical and physical changes or extremes of environment, a large number of micro-organisms are highly adapted and flourish in environments usually considered lethal. Certain chemoautotrophic bacteria require high concentrations of ammonia, methane, or other chemicals to grow. Anaerobic bacteria grow only in the absence of oxygen.
Besides adapting to the extremes of environments on Earth, life is also capable of growing and reproducing under extreme environmental conditions not normally encountered: e.g., from a few rad of radiation in normal habitats to 106 or more rad from artificial sources, from 0.5 gauss of Earth magnetism to 167 000 gauss in manmade magnetic fields, and from 1-g force of gravity to 110 000 g. The extreme ranges of physical and chemical environmental factors for growth, reproduction, and survival for Earth micro-organisms are phenomenally large.
Life is ubiquitous on Earth and is found in almost every possible environment, including the most severe habitats, from the bottom of the ocean to the highest mountain tops and from cold Arctic habitats to hot springs, as well as in volcanic craters, deep wells, salt flats, and mountain snowfields. Earth life has become adapted to, and has invaded, nearly every habitat, no matter how severe. The physiological and morphological adaptations of life are exceedingly diverse and complex.
Surprisingly, the extreme parameters or ranges of the physical and chemical environmental factors permitting growth, reproduction, and other physiological processes of Earth organisms have not been critically compiled. A partial compilation of certain selected environmental factors has been made by Vallentyne ([ref.76]). A compilation of available published data on certain environmental extremes, particularly from recent NASA-supported research (compiled by Dale W. Jenkins, in press), is presented in tables III to VI. These data can serve as a starting point for a more intensive literature review by specialists, critical evaluation, standardization of end points, and especially to point out areas where critical experimentation is urgently needed.
This critical compilation involves a review of a very broad and complex range of subjects involved in many different disciplines with widely scattered literature. Since the effects of many of the specific environmental factors are harmful, it is difficult to select a point on a scale from no effect to death and use some criteria to say that normal or even minimal growth and reproduction are occurring. The effects of environmental factors are dependent on (1) the specific factor, times, (2) the concentration or energy, times, (3) the time of exposure or application of the factor. Many reports, especially older ones, do not give all of the necessary data to permit proper evaluation. A complicating factor is that the effect of each factor depends on the other factors before, during, and after its application. The condition of the organism itself is a great variable. Proper evaluation requires the critical review by a variety of biological specialists, physicists, and chemists.
To determine the potential of Earth organisms to survive or grow under other planetary environmental conditions, a number of experiments have been carried out attempting to simulate planetary environments, especially of Mars, as reviewed previously. While the results are of real interest, they do not provide much basic information. Further, as the Martian environment is more accurately defined, the experimental conditions are changed. In addition, some experimenters have altered certain factors, such as water content, to allow for potential microhabitats or for areas which might contain more water at certain times.
Physical factors |
Minimum |
Organism |
|---|---|---|
Temperature |
-30° C |
Algae (photosynthesis), pink yeast (growth) |
Magnetism |
0-50 gamma (=×10-5 gauss) |
Human |
Gravity |
0 g |
Human, plants, animals |
Pressure |
10-9 mm Hg (5 days) |
Mycobacterium smegmatis |
Microwave |
0 W/cm2 |
|
Visible |
0 ft-c |
Animals, fungi, bacteria |
Ultraviolet |
0 erg/cm2 |
|
X-ray |
0 rad |
|
Gamma ray |
0 rad |
|
Acoustic |
0 dyne/cm2 |
Physical factors |
Maximum |
Organism |
Activity |
|---|---|---|---|
Temperature |
104° C (1000 atm) |
Desulfovibrio desulfuricans |
Grows and reduces sulfate |
Magnetism |
167 000 gauss |
Neurospora
Arbacia
Drosophila
|
1 hr—no effect, Arbacia development delayed |
Gravity |
400 000 g |
Ascaris eggs |
1 hr—eggs hatch, 40 days' growth |
110 000 g |
Escherichia coli |
||
Pressure |
1400 atm |
Marine organisms |
Growth |
Microwave |
2450 Mc/sec 0.3 to 1 W/cm2 |
Drosophila |
68 hr, growth not affected |
Visible |
50 000 ft-c |
Chlorella, higher plants |
Seconds, recurrently continuous |
17 000 ft-c |
|||
Ultraviolet |
108 erg/cm2, 2537 Å |
Bean embryos |
Suppressed growth |
X-ray |
2×106 rad |
Bacteria |
Growth |
Gamma ray |
2.45×106 rad |
Microcoleus
Phormidium
Synechococcus
|
Continued growth |
Acoustic |
140 db or 6500 dyne/cm2 at 0.02 to 4.8 kcs/sec |
Man |
Threshold of pain |
Minimum temperature, °C |
Organism |
Activity or condition |
|---|---|---|
-11 |
Bacteria |
Growth (on fish) |
-12 |
Bacteria |
Growth |
-12 |
Molds |
Growth |
-15 |
Pyramidomonas |
Swimming |
-15 |
Dunaliella salina |
Swimming |
-18 |
Mold |
Growth |
-18 |
Yeast |
Growth |
-18 |
Aspergillus glaucus |
Growth (in glycerol) |
-18 to -20 |
Mold |
Growth (in fruit juice) |
-18 to -20 |
Pseudomonads |
Growth (in fruit juice) |
-20 |
Bacteria |
Growth |
-20 |
Bacteria |
Growth |
-20 |
Bacteria |
Luminescence development accelerated |
-20 to -24 |
Insect eggs (diapause) |
|
-30 |
Algae |
Photosynthesis |
-30 |
Pink yeast |
Growth (on oysters) |
-30 |
Lichens |
Photosynthesis |
-20 to -40 |
Lichens and conifers |
Photosynthesis |
-44 |
Mold spores |
Sporulation and germination |
Maximum temperature, °C |
Organism |
Activity or condition |
|---|---|---|
73 |
Thermophilic organisms |
Growth (P32 metabolism) |
73 |
Phormidium (alga) |
Acclimatized |
70 to 73 |
Bacillus calidus |
Growth and spore germination |
70 to 74 |
Bacillus cylindricus |
Growth and spore germination |
70 to 75 |
Bacillus tostatus |
Growth and spore germination |
80 |
Bacillus stearothermophilus |
Cultured in laboratory |
83 |
Sulfate-reducing bacteria |
Found in a well |
89 |
Sulfate-reducing a bacteria |
Found in oil waters |
65 to 85 |
Sulfate-reducing a bacteria |
Cultured in laboratory |
89 |
Micro-organisms |
Found in hot springs |
95 |
Bacillus coagulans |
In 80 min. sporulation activation |
110 |
Bacillus coagulans |
In 6 min, sporulation activation |
104 |
Desulfovibrio desulfuricans |
Grow and reduce sulfate at 1000 atm |
Minimum temperature °C |
Organism |
|---|---|
-190 |
Yeast bacteria, 10 species |
-197 |
Trebouxia erici from lichens |
-197 |
Protozoa, Anguillula |
-252 |
Yeasts, molds, bacteria, 10 species |
-253 |
Black currant, birch |
-273 |
Bacteria, many species |
-273 |
Bacteria, many species |
-272 |
Desiccated rotifers |
-269 |
Human spermatozoa |
Maximum temperature °C |
Organism |
Time of exposure |
|---|---|---|
140 |
Bacterial spores |
5-hr immersion |
170-200 |
Desiccated rotifers |
5 min |
151 |
Desiccated rotifers |
35 min |
150 |
Clostridium tetani |
180 min |
170 |
Aerobic bacteria, molds. actinomycetes |
5 days at 6×10-9mm Hg |
127 (dry) |
Bacteria (in activated charcoal) |
60 min |
110 (wet) |
Bacillus subtilis var. niger |
400 min |
120 |
Bacillus subtilis var. niger |
400 min |
141 |
Bacillus subtilis var. niger |
70 min |
160 |
Bacillus subtilis var. niger |
15 min |
180 |
Bacillus subtilis var. niger |
2 min |
188 |
Bacillus subtilis var. niger |
1 min |
120 (wet) |
Bacillus stearothermophilus |
25 min |
120 (dry) |
Bacillus stearothermophilus |
100 min |
141 |
Bacillus stearothermophilus |
12 min |
160 |
Bacillus stearothermophilus |
2 min |
166 |
Bacillus stearothermophilus |
1 min |
Chemical factor |
Minimum |
Organism |
|---|---|---|
O2 |
0% |
HeLa cells, Cephalobus, anaerobic bacteria |
O3 (ozone) |
0% |
|
H2 |
0% |
|
H2O |
Aw 0.48 |
Pleurococcus vulgaris |
Aw 0.5 |
Xenopsylla cheopis (prepupae) |
|
H2O2 |
0% |
|
He |
0% |
|
CO |
0% |
|
CO2 |
0% |
|
CH4 |
0% |
|
CH2O |
0% |
|
CH3OH |
0% |
|
N2 |
0% |
|
NO |
0% |
|
NO2 |
0% |
|
N2O |
0% |
|
Ar |
0% |
|
NaCl, Na2SO4, NaHCO3 |
||
H2S |
0% |
|
H2SO4 |
0% |
|
Cu++ |
||
Zn++ |
||
pH |
0 |
Acontium velatum
Thiobacillus
thioodixans
|
Eh |
-450 mV at pH 9.5 |
Sulfate-reducing bacteria |
Chemical factor |
Maximum |
Pressure, atm |
Time, days |
Organism |
Activity |
|---|---|---|---|---|---|
O2 |
100% |
1 |
Plants, animals |
Growth |
|
O3 (ozone) |
100 ppm |
5 |
Armillaria mellea |
Growth |
|
500 ppm |
5 |
Light emission |
|||
H2 |
100% |
Various plants |
Germination |
||
H2O |
Aw 1.0 |
1 |
Various aquatic organisms |
Growth |
|
H2O2 |
0.34% |
Rye |
Germination enhanced |
||
He |
100% |
Wheat, rye, rice |
Germination |
||
CO |
100% |
Rye |
Germination |
||
80% |
1.1 |
4 |
Hydrogenomonas |
Growth |
|
CO2 |
100% |
1.1 |
4 |
Rye |
Growth and germination |
CH4 |
100% |
1.1 |
4 |
Rye |
Germination |
CH2O |
50% |
Rye |
Germination |
||
CH3OH |
50% |
Rye |
Germination |
||
N2 |
100% |
.1 |
10 |
Various plants |
Germination and root growth |
NO |
18% |
.018 |
10 |
Sorghum, rice |
Germination and root growth |
NO2 |
18% |
.018 |
10 |
Rye, rice |
Germination and root growth |
N2O |
100% |
1.2 |
4 |
Rye |
Germination |
96.5% |
1.7 |
Rye |
Germination |
||
Tenebrio molitor |
Survival |
||||
Ar |
100% |
1.2 |
2 |
Rye |
Germination |
NaCl, Na2SO4, NaHCO3 |
67% |
Photosynthetic bacteria |
Growth |
||
H2S |
0.96 g/liter |
Desulfovibrio desulfuricans |
Growth |
||
H2SO4 |
7% |
Acontium velatum |
Growth |
||
Thiobacilli |
Growth, reproduction |
||||
Cu++ |
12 g/liter |
Thiobacillus ferrooxidans |
Growth |
||
Zn++ |
17 g/liter |
Thiobacillus ferrooxidans |
Growth |
||
pH |
13 |
Plectonema nostocorum |
Growth |
||
Nitrobacter |
Growth |
||||
Nitrosomonas |
Growth |
||||
Eh |
850 mV at pH 3 |
Iron bacteria |
Growth |
chapter 4
EFFECTS OF THE SPACE ENVIRONMENT ON BEHAVIOR
NASA was established in 1958, shortly after the Russian launching of the second Earth satellite Sputnik II, the first vehicle to carry life into orbit around the Earth. This accomplishment was preceded by the pioneering work of Henry et al. ([ref.77]), in which animals were exposed briefly to low-gravity states in Aerobee rockets. A motion-picture camera photographed the behavior of two white mice in rotating drums during this series of flights, which marked the first time that simple psychological tests were made on animals in the weightless condition. While this behavioral experiment was relatively simple, it provided the basic concepts for recent studies which involved rotation of animals during the weightless state. Subsequent flights such as Project MIA (Mouse-in-Able) reflected a preoccupation with physiologic measures (refs. [ref.78] and [ref.79]), although the flights of Baker and Able included preflight and postflight performance studies ([ref.80]). Able's behavior was recorded in detail on in-flight film, but none of the behavior was programed or under experimental control.
The first flights in which behavior or performance was explicitly programed were those of Sam and Miss Sam in flights of the Little Joe rocket with the Mercury capsule, launched from Wallops Island in 1959 and 1960 ([ref.81]). The first major space achievement in the behavioral sciences was the successful in-flight measurement of the behavior of the chimpanzee Ham in early 1961, in which the pretrained animal performed throughout the flight. The second achievement along these lines was in 1962 when the chimpanzee Enos made several orbits around Earth and performed continuously on a complex behavioral task. The tasks which the animals performed during these flights have been described in detail by Belleville et al. ([ref.82]), and the results of the in-flight performance have been presented by Henry and Mosely ([ref.83]). These early flights provided much of the technological framework on which current biological experiments on organisms during flights of extended duration are based. Due largely to the efforts of Grunzke (refs. [ref.84] and [ref.85]), the apparatus needed to sustain animals during space flight, such as zero-g watering and feeding devices, are now commonplace ([ref.86]). Advanced systems of programing stimulus presentations and recording responses, developed for Project Mercury, may now be seen in many basic research laboratories throughout the country.
Several other noteworthy advances have been made as an outgrowth of the Mercury animal flights. Immediately before the orbital flight MA-5, in which the chimpanzee Enos was employed, it was unexpectedly found that this 5-year-old animal was hypertensive. Subsequent centrifuge studies showed that its vascular responses exceeded those of a control group. Consideration of the animal's preflight experience led to speculation concerning the origin of this hypertension. An explanation of the high-blood-pressure responses detected in Enos has been pursued by Meehan et al. ([ref.87]). Persistent hypertension has been produced in other laboratory chimpanzees restrained in the same manner as those participating in space flight and exposed to demanding performance tasks, a demonstration which has important implications for prolonged manned space flight and for cardiovascular medicine in general.
Studies more directly concerned with behavior and performance have been extended from those of Project Mercury. These extensions have been in the following directions: (1) the establishment and maintenance of complex behavioral repertoires under conditions of full environmental control, (2) the refinement of behavioral techniques for assessing sensory and motor processes, and (3) the maintenance of sustained performance under conditions of long-term isolation and confinement and preliminary extension of such experimental analysis to man.
Numerous studies with primate subjects, including several at Ames Research Center, have been devoted to developing methods for maintaining optimum performance in environments with limited sources of stimulation. Monkeys, baboons, and chimpanzees, for example, have been isolated for periods of longer than 2 years with no decrement in performance on complicated behavioral tasks ([ref.88]). The behavioral techniques used in these studies are closely related to those employed on human subjects under NASA sponsorship at the University of Maryland ([ref.89]). The essence of these techniques is in the proper programing of environmental stimuli ([ref.90]). It is not sufficient to provide the subject with his physiological requirements for survival, but he must be given the psychological motivation for using these provisions. This statement, of course, is an oversimplification of the problem, but it serves to illustrate the essence of these experimental programs.
Gravity has long been known as one of the major factors influencing various life processes and the orientation of both plants and animals. One of the most challenging problems of space research has been to define this influence more precisely. Related to the effect of gravity on living processes is the problem of the effects of weightlessness. Of particular interest to psychologists are the possible modifications an altered gravitational environment might produce in behavioral patterns basic to the animal's maintenance and survival, such as eating, sensory and discriminative processes, development and maturation, and learning capacity ([ref.91]).
One prominent method of studying gravitational effects is to simulate an increase in gravity by centrifugation. Smith et al. ([ref.92]) and Winget et al. ([ref.93]) have investigated the effects of long-term acceleration on birds, primarily chickens, while Wunder (refs. [ref.94] and [ref.95]) and his coworkers (refs. [ref.96]-[ref.99]) have used fruit flies, mice, rats, hamsters, and turtles. The general findings are that, when animals are subjected to a prolonged period of acceleration of moderate intensity, they exhibit decreased growth, delayed maturation, and an increase in the size of certain muscles and organs, dependent on the species. With regard to the decreased growth effect, the data of these investigators show some exceptions. When the gravitational increase is kept below a certain limit, growth was greater than that of controls in the fruit fly, turtle, mouse, and chicken. The limit below which enhancement of growth was observed varied with the species studied.
The data on food intake do not present a consistent picture. Wunder ([ref.94]) found that food intake in accelerated mice was markedly reduced from that of nonaccelerated control animals. Smith, however, found that in chickens, food intake increased up to 36 percent over controls and has derived an exponential relation between food intake and acceleration. After six generations of selective breeding, Smith has produced a strain of chickens better adapted to prolonged exposure to high g.
A very relevant finding of their research with birds was that exposure to chronic acceleration in some way appears to interfere with habituation to rotatory stimulation. Chickens who were being subjected to chronic acceleration were given repeated rotatory stimulation tests to estimate their labyrinthine sensitivity. This study revealed that centrifuged animals showed a marked reduction in labyrinthine sensitivity. This result appeared to persist after the acceleration was terminated. In animals who developed gait or postural difficulties as a result of acceleration, there was no evidence of a postnystagmus in response to the rotatory stimulation test, which the investigators point out may be evidence of a lesion in the labyrinth or its neural pathways.
Smith has implicated social factors as interfering with acceleration effects. His subjects were typically accelerated four or six to a cage. When groups were mixed midway through the experiment, they exhibited a higher mortality rate and incidence of acceleration symptoms than did groups whose constituency remained unchanged.
At the U.S. Naval School of Aerospace Medicine, numerous studies have been conducted on the effects of slow rotation on the behavior and physiology of humans and animals ([ref.100]). Rotation initially produces decrements in performance, but adaptation to a rotating environment ensues quite rapidly (refs. [ref.101]-[ref.103]). Perceptual distortion, nystagmus, nausea, and other signs of discomfort are common responses to slow rotation. These symptoms are generally reduced with continued exposure (adaptation). Interestingly, however, adaptation is delayed when the subjects are exposed to a fixed reference outside their rotating environment.
At NASA-Ames, rodents have been used in experiments by Weissman and Seldeen to delimit the stimulus effects of rotation. In these experiments the subjects must discriminate between different speeds of rotation in order to obtain food reinforcement. The results thus far provide evidence that these animals are capable of discriminating between the different speeds at which they are being rotated. The range of speeds studied was 0-25 rpm, with tests of discrimination being made at intervals of less than 5 rpm. Experiments such as these will lead to the development of techniques for measuring rotational sensitivity in many species, including man.
The optimum configuration of manned spacecraft will depend, in part, upon biomedical considerations. A voluminous literature now exists on the possible hazards to man of prolonged exposure to zero-g conditions. Should prolonged weightlessness prove to be a serious detriment to health, consideration must be given to design concepts which provide artificial gravity.
No data exist on the minimum gravity requirements necessary to sustain basic biological functions for extended periods. A limit of 0.2 g has been given as the lower level at which man can walk unaided ([ref.104]). It has also been recommended that angular velocity be maintained at the lowest possible level in order to minimize the occurrence of vestibular disturbances. These recommendations are based on human-factor requirements, rather than upon biological considerations, which may significantly modify these values. In recent studies, a technique has been devised which promises to provide reliable criteria for biological acceptability, since it is based on fundamental biological and behavioral principles.
As animals progress up the evolutionary stale, their survival depends less and less upon stereotyped physiological reactions which occur in reflex fashion, in response to environmental stimulation. In higher organisms, survival depends more upon the capacity of organisms to modify their behavior. At the highest levels of functional efficiency, the ultimate form of adaptation is seen—the manipulation of the environment by the organism. Developments in behavioral science now permit us to utilize the adaptive behavior of animals to investigate many problems of biological interest. Recent studies on the self-selection of gravity levels represent a further attempt to exploit the adaptive capacities of animals, in order to provide information relevant to problems of space exploration.
One such project allows animals to select their own gravity environment in an apparatus designed to create g-forces through centrifugal action by rotation at 60 rpm ([ref.105]). The surface of this centrifuge is parabolic, so that the resultant of the centrifugal g and the Earth's gravity is always normal to the surface. When the animal moves away from the center, increasing the radius of rotation, it is exposed to increasing gravity. Motion toward the center reduces the gravity level. By this means, an animal is free to select its own gravity environment.
When the animal moves toward or away from the center, he is moving from one tangential velocity to another. He is therefore acted upon by a third force—due to Coriolis acceleration. The effects of Coriolis forces are a major problem difficult to eliminate in studies such as these, but they must be taken into account in the design of spacecraft which produce artificial gravity by rotation. Motion of the head in any direction not parallel to the centrifugal force vector would result in bizarre stimulation of the semicircular canals and consequent motion sickness. This effect is likely to become even more pronounced if the sensitivity of these organs is increased by prolonged exposure to reduced gravity. Methods such as these are currently being developed for conducting a refined psychophysical analysis of gravity, including studies by Lange and Broderson on the perception of angular, linear, and Coriolis acceleration.
The results of animal studies such as these will be of great value in arriving at a decisive judgment concerning the need for artificial gravity in a manned orbiting space station, or other vehicles designed for long-term occupancy.
To aid in the interpretation of in-flight data, other studies are underway to determine the functions of the vestibular system, as a principal brain center related to orientation in space and to the physiology of posture and movement, as well as with the influences of acceleration, rotation, and weightlessness. Experiments are presently being conducted on monkeys and cats in order to trace these complex neurological connections and to determine their functional organization.
BIOLOGICAL INFORMATION SYSTEMS
The nature of memory has been the subject of considerable speculation in the past. It has long been felt intuitively that retention of information in the central nervous system involves either an alteration of preexisting material or structure, or, alternatively, synthesis of materials not present previously. The cellular site of operational alteration was unknown but, again intuitively, was felt to be closely associated with the synapses. The problems faced by early investigators were great; but nevertheless much information relevant to the question of biological information storage was obtained. With the relatively recent advent of more refined tools and methodologies, there has been rapid progress.
A significant amount of the work which has been conducted in the area of biological information and communication systems is easily classified as "basic research" (refs. [ref.106]-[ref.109]). This discussion will be limited to those aspects closely related to the fields of molecular biology and experimental psychology, which seem to have universal application to all known animal life forms. Studies involving the basic principles of acquisition, processing, storage, and retrieval of information in living systems are emphasized.
Early Work
Early speculations on the operational nature of memory have been based upon relatively little experimental evidence. Charles Darwin observed that domestic rabbits had smaller brains than their wild counterparts, and attributed this to lack of exercise of their intellect, senses, and voluntary movements. Unfortunately, subsequent studies of the brains of men with greatly differing intellectual capability did not substantiate the hypothesis. Idiots sometimes had larger brains than geniuses. Later, an idea proposed by Ramon y Cajal came into favor. Since brain cells did not increase in number after birth, he proposed that memory involved the establishment of new and more extended intercortical connections. Unfortunately, methods were not available to test this hypothesis adequately and it has remained until quite recently in the realm of conjecture.
Another major hypothesis was that there were two or more stages in the information storage process. The final form the information took in the brain was called a brain engram, or memory trace. However, prior to the formation of the engram, a transitory process denoted as "reverberational memory" was postulated to exist for a relatively short time (minutes to hours) (refs. [ref.106] and [ref.107]). This hypothesis was used by Pauling to explain why an elderly chairman of a board could brilliantly summarize a complex 8-hour meeting and yet, after its conclusion and his return to his office, not even remember having attended the meeting. Thus, this individual's reverberational memory functioned well, but advanced years had seriously impaired his brain's ability to form a permanent engram. Similar, although less dramatic, observations in other situations are not uncommon. A wide variety of experiments have been conducted to study this aspect of memory and to relate it to the process whereby the information is transformed to a more stable form (refs. [ref.110]-[ref.112]).
More recently, the concept of a specific biochemical activity during the process of long-term storage of information has gained considerable favor. Initially, neither the site nor the nature of the change was well defined. Quite recent studies by Krech et al. (refs. [ref.113] and [ref.114]), Bennett et al. ([ref.115]), Rosenzweig et al. (refs. [ref.116] and [ref.117]) support the view that alteration of the levels of acetylcholinesterase at cortical synapses play an important role in information storage. These studies will be discussed in a later section. However, these authors do not claim that the changes observed are unambiguously related to the storage of memory. It may well be that the alterations observed are in some way related to this process but are still secondary to some other, more basic, process.
An alternative hypothesis is that the information resides in its ultimate form in some more central structure of the neurone than the synapse. (It has even been postulated that the basic information is stored in nonneuronocortical material.) Perhaps Halstead was the first to postulate the involvement of nucleoprotein in this process ([ref.107]). From the biochemist's point of view, this is an extremely attractive hypothesis. Both proteins and nucleic acids possess sufficient possible permutations of structure to permit storage of a lifetime's accumulation of information in an organ the size of the brain. From the previously known ability of the nucleic acids to code genetic information, they are the prime suspects. However, from the known regulatory ability of nucleic acids in specific protein synthesis, it is possible that the final repository is protein.
Recent Biochemical Studies
Among the foremost investigators of the chemistry and biochemistry of the central nervous system is Holger Hyden at the University of Göteborg, Sweden. He and others (refs. [ref.118]-[ref.120]) have for many years performed elegant microanalytical studies of single nerve cells. The evidence which Hyden has obtained is consistent with the hypothesis that the initial electrical reverberations in the brain induce a change in the molecular structure of the ribonucleic acid (RNA) of the neurones which, in turn, leads to a subsequent deposition of specific proteins. It is well known from other investigations that a major role of RNA in any type of cell is to specify and mediate synthesis of the protein enzymes of the cells. Thus, in this hypothesis, it is only necessary to postulate the modification of brain RNA by the activities associated with reverberational memory. Particularly pertinent to this hypothesis are observations that—
Large nerve cells have a very high rate of metabolism of RNA and proteins, and, of the somatic cells, are the largest producers of RNA.
Vestibular stimulation by passive means leads to an increase in the RNA content of the Deiters nerve cells of rabbits ([ref.121]). The protein content of these cells is also increased.
Changes in the RNA composition of neurones and glia of the brainstem occur during a learning situation. Animals were trained over a period of 4 to 5 days to climb a steeply inclined wire to obtain food. The big nerve cells and the glia of their lateral vestibular apparatus were analyzed, since the Deiters neurones present in this structure are directly connected to the middle ear. The amount of RNA was found to be increased in the nerve cells; and, more significantly, the adenine-to-uracil ratio of both the nuclear RNA of nerve cells and glia cells became significantly increased ([ref.119]). A variety of control experiments were conducted. Although there was an increase in RNA content of these cells in animals exposed to passive stimulation, there was no change in the ratio of adenine to uracil. Nerve cells from the reticular formation, another portion of the brain, had only an increased content of RNA with no base-ratio change. Animals subjected to a stress experiment involving the vestibular nucleus showed only an increase in content of RNA. Littermates living in cages on the same diet as learning animals showed no change in content of RNA. Thus, it would appear that the change in the base ratio of the RNA synthesized is not due to increased neurone function per se, but is more directly related to the learning process. The fact that this was nuclear RNA implies that it was immediately related to chromosomal DNA.
Neuronal RNA with changed cytosine-guanine ratios synthesized during a short period of induced protein synthesis could be blocked by actinomycin D. It was concluded, therefore, that the RNA was immediately DNA dependent and directly related to the genetic apparatus.
Rats which were normally right handed were forced to modify their handedness in order to obtain food. The RNA of nerve cells in that part of the cortex, whose destruction destroys the ability to transfer handedness, was analyzed. A significant increase in RNA of nerve cells of the fifth to sixth cortical layers on the right side of the brain was observed. The corresponding nerve cells on the opposite side of the same brain served as controls. There was an increase in RNA and a significant increase in the purine bases relative to the pyrimidine bases in the learning side of the cortex. When the animals were not forced to learn a new procedure, only an increase of RNA was observed, with no change in base ratio.
Frank Morrell, head of the Neurology Department at Stanford Medical School, has also been active in this field during the past 6 years. He has found that if a primary epileptic lesion is induced on one side of the cortex, a secondary mirror lesion eventually develops in the contralateral homologous cortex. This secondary lesion, which showed self-sustaining epileptiform discharge, could be isolated, whereupon the epileptiform discharge disappeared. This was interpreted as learned behavior of the secondary lesion. From changes in the staining properties of the secondary lesion, Morrell concluded that changes in RNA had occurred in the cell. Changes in the composition of the RNA could not be shown by these techniques.
At the University of California at Berkeley, Drs. Rosenzweig, Bennett, and Krech have conducted extensive studies related to this topic. These investigators have directed their efforts toward demonstrating alterations in the cerebral cortex of animals exposed to continuing learning situations or continuously deprived of sensory stimulation. In a recent publication ([ref.116]), which also summarizes a considerable amount of previous work, they report studies which demonstrate the following:
Rats given enriched experience develop, in comparison with their restricted littermates, greater weight and thickness of cortical tissue and an associated proportional increase in total acetylcholinesterase activity of the cortex.
The gain in weight of cortical tissue is relatively larger than the increase in enzymatic activity. Acetylcholinesterase activity increases in other portions of the brain even though tissue weight decreases.
The changes appear in a variety of lines of rats, although differing in amount between strains.
The changes are observed in both the young and adult animals.
The previous studies were comparisons between experience-enriched animals and animals maintained in isolation. Animals which were housed in colonies, but given no special treatment, showed intermediate effects in those situations studied.
The Berkeley group emphasized that the finding of changes in the brain subsequent to experience does not prove that the changes have anything to do with memory storage, but do establish the fact that the brain can respond to environmental pressure. However, the results are compatible with the hypothesis that long-term memory storage involves the formation of new somatic connections among neurones. Calculations of the amount of additional material required to permit this to exist are compatible with the increases observed.
A number of investigators have studied the effects of antimetabolites and drugs on the learning process. Since their specific metabolic effects are known in other tissues, the rationale is that if these materials do interfere with memory, then specific types of metabolic activities may be implicated in the deposition of the engram.
One of the initial studies of this type was conducted by Dingman and Sporn ([ref.122]), presently at the National Institute of Mental Health. They showed that 8-azaguanine, a purine antagonist, injected intra-cisternally was incorporated into the RNA of the brains of rats. Associated with this incorporation was an impairment of the maze-learning ability of the animals. These findings have been confirmed.
Flexner and his associates injected puromycin, an inhibitor of protein synthesis, into the brains of mice, which were then trained to perform in a maze. Losses of short-term or long-term memory were obtained, depending upon the site of the injection. The results indicate that the hippocampal region is the site of recent memory.
The hippocampal region is of interest in connection with memory processes for a number of other reasons. Adey et al. ([ref.123]) and his group observed a transient fall in electrical impedance in this region when cats learned to perform in a T-maze in response to a visual cue. It was supposed that the electrodes were situated within glial cells of the dendritic zone of the hippocampal pyramidal cell layer. Extinction of the learned habit abolished the briefly evoked impedance changes, which subsequently reappeared with retraining.
A number of other studies more or less indirectly implicate RNA in the learning processes. For instance, in retinal cells of rabbits raised in darkness, there was virtually no ribonucleoprotein as compared with normal amounts in the cells of animals raised in light ([ref.124]). Further, maintenance of normal electrical activity of isolated perfused cat brains is highly dependent upon the presence of the ribonucleic acid precursors, uridine and cytidine, in the perfusate ([ref.125]), and severe derangements occur if any of a variety of pyrimidine antagonists are added ([ref.126]). Brief electrical stimulation of cat cortical tissue causes an increase in nucleic acid cytidine and adenine, thus indicating a synthesis of altered polynucleotides. Finally, injections of RNA in animals have shown interesting effects. When given at a dose of 116 mg/kg daily for 1 month, rats showed an enhanced response and greater resistance to extinction in a shock-motivated behavioral response. It has been shown by another group that injections of RNA enhance the ability of young animals to learn various tasks.
Planaria have been used in a variety of studies which seem to bear on the problem of memory. Quite recent evidence by Bennett, Calvin, and their associates has cast somewhat of a pall over the studies; nevertheless, the work may have some validity. Interest in the use of flatworms, particularly planaria, for study of memory began with a demonstration by McConnell that these simple animals could undergo conditioning ([ref.127]). Subsequently, it was found that some conditioning was retained when the animal was transected and allowed to regenerate. The retention of training was found in both new animals, although the very simple brain, really only two ganglia, was in the head section ([ref.128]).
Apparently, some diffusely distributed component of the animal was responsible for retention of learning. Evidence has accumulated to indicate that this material is RNA. Among this evidence is the following:
The two halves of a trained planaria were allowed to regenerate in a solution containing RNA-destroying enzymes. Whereas the head ends retained some training, no retention was observed in the animals derived from the tail end ([ref.129]).
When pieces of trained planaria were fed to untrained animals, the untrained cannibal required a shorter time to become trained to a criterion. It would appear that the digestive system of planaria is so simple that the material responsible for the transfer of the information was not broken down.
When RNA, obtained from trained planaria, is injected into the digestive tract of untrained animals, there is a transfer of information.
NEUROPHYSIOLOGY2
Neurophysiological studies concern the functions of the nervous system—in particular the central nervous system (CNS)—under normal, simulated, and actual flight conditions. Of paramount importance is the maintenance of equilibrium and orientation in three-dimensional space. The ability of man and his close relatives among the vertebrates to maintain these functions depends on an integrated sensory input from the vestibular organ; the eyes; the interoceptors of the muscles, tendons, joints, and viscera; and the exteroceptors of the skin.
Certain parameters of the environmental and space-flight conditions drastically affect man's ability to maintain equilibrium and spatial orientation. Centrifugal forces modify or reverse the directional vector of gravity. Linear acceleration may increase enormously, as may angular stimulation. The sensory organs listed above are unreliable under such conditions. The very organ which is designed specifically to furnish information on spatial orientation may malfunction in man while he is in flight. Thus, with respect to sensory orientation, these labyrinthine organs are by no means precision instruments.
The use of classical histological methods and the observation of equilibrium disturbances resulting from operative interference with the internal ear have in the past been the two principal sources of knowledge concerning the structure and function of the labyrinth, but the answers given to various questions vary considerably in their value. The development of electrophysiological techniques and the refinement in recent years of the ultrastructural analysis by means of the electron microscope may allow more precise experimental studies of the correlation of function and structure.
Before considering vestibular impulses in their bulbar and descending spinal pathways, a recent study concerning the generation of impulses in the labyrinth must be mentioned. Von Bekesy's finding ([ref.131]) of the direct current potentials in the cochlea aroused speculation about the existence of similar labyrinthine potentials. Such dc potentials were also detected in the semicircular canal of the guinea pig by Trincker ([ref.132]), who measured the potential changes in the endolymph, surface of the cupula, or side of the crista during cupular deflection. It seems likely, however, that the effects do not represent the physicochemical changes in the cupula but the electrical potentials in the nerve and nerve endings of the crista. Attempts at differentiating these effects have failed so far. Great expectations are brought by the advances of microchemistry, microphysiology, and physical chemistry with regard to the excitatory processes, the generation of the nerve impulse. Quite apart from a need to understand vestibular nerve discharges and patterns more adequately in such terms, the analysis of the vestibular system has in the past revealed general biological principles which were not readily discernible through the examination of other tissues ([ref.133]).
The neural connections of the vestibular organ consist of numerous chains of neurons, reciprocally linked in many ways and having their synapses in various anatomical nuclei. All the chains work in intimate collaboration, and the final pattern of reflex responses is attributable largely to the highly complex integrating activity of the center. The labyrinthine function is automatic, carried out in a reflex fashion: in other words, mostly below the level of consciousness. The brain centers through which the labyrinth elicits the various appropriate muscular reactions of the head, body, limbs, and eyes—the righting, the postural, and the ocular reflexes—represent an intricate mechanism. Before we can hope for a satisfactory understanding of their functional organization, we will have to know their anatomy in more detail. Thus, we are confronted with a fruitful field for the exploration of basic mechanisms of neuronal activity. Major advances dining the last years have provided us with new information about the neuroanatomy of the vestibular system (refs. [ref.134]-[ref.137]).
Vestibular impulses entering the brainstem ascend and descend the neuroaxis and cross the midline. It was previously believed that the vestibular apparatus had only subcortical projections. Recently, however, it has been established by means of electrophysiological methods that the organ is represented by a projection area in the cerebral cortex of some animals (refs. [ref.138]-[ref.141]). The use of brief electrical stimulation of the vestibular nerve in order to elicit a cortical response has been of great value for the mapping of these areas.
Among a great variety of sensory receptors, the vestibular ones are capable of evoking the most widespread somatovisceral effects throughout the body. Moreover, vestibular effects seem to be imperious and less dependent upon the state of readiness of the nervous system. As a consequence of the extensive distribution of vestibular effects, there are many opportunities for central integration. Proprioceptive and vestibular systems are both known to be active in posture and locomotion; streams of impulses arising from the receptors in each of these systems must converge to influence the activity of the final common path. The state of the motor centers of the spinal cord, as affected by vestibular stimulation, has been tested by dorsal root and other sensory input interventions. These experiments have provided us with insight into the mechanisms concerned with the vestibular control of spinal reflexes (refs. [ref.142]-[ref.146]).
It has long been known that the vestibular apparatus is essential for the development of motion sickness. Commonplace subjective experience of nausea relates to visceral changes mediated through autonomic efferent pathways and may ultimately involve rhythmic somatic nerve discharges to skeletal muscles responsible for retching and vomiting. However, very little is known about the central nervous mechanisms responsible for elaboration of the whole syndrome. Since the maintenance of vestibular bombardment for some length of time seems essential for the development of motion sickness, one would presume this to be an instance of slow temporal summation. Experimental findings demonstrate a powerful effect of temporal summation upon somatic motor outflow during vestibular stimulation ([ref.147]), and not upon parasympathetic outflow.
The practical implication of these studies is closely related to physiological effects of weightlessness. Based on experimental evidence from short weightless periods obtained in aircraft, it was concluded that "when the exposure becomes longer, there may develop minor physiologic disturbances which, if cumulative or irritating, may cause or enhance psychiatric symptoms" ([ref.148]). Although the zero-g condition, per se, does not cause spatial disorientation if visual cues are provided, the astronauts reported a temporary loss of orientation during the orbital flight while they were engaged in activities which diverted their attention. However, no disturbing sensory inputs were observed during the weightless period. Violent head maneuvers within the limited mobility of the helmet were performed in every direction without illusions or vertigo. The subjective sensations of "tumbling forward" after sustainer engine cutoff reported by the Mercury astronauts, and Titov's motion sickness attacks, which were particularly dismaying during head movements, were well within the entire range of psychosomatic experiences already obtained during aerodynamic trajectories ([ref.149]). Interestingly enough it now appears that the otolithic output in mammals including man is the differential of linear acceleration, and therefore unaffected by zero g.
Of interest in this connection are the problems which may be encountered during and following long-term exposure to weightlessness. Although there is no evidence of adverse effects on operative behavior, the possibility of biological disturbances on a cellular or subcellular level, which may cause a deterioration of the somatic basis, has been repeatedly stressed. Whether effects of this sort will occur or whether the organism will be able to adapt is still an open question. Since motion sensitivity based on vestibular stimulation differs widely among individuals, the selection of astronauts may solve the problem of zero-g vestibular disturbance. Reports from the MA-8 (Sigma 7) and Vostok III and IV flights seem to support this assumption. Moreover, experiments are being made in the slow rotation room at the Naval School of Aviation Medicine to study the Coriolis effects which arise when "artificial gravity" is produced by angular acceleration. Since man can adapt to wave motion on shipboard within a few days, a similar process may be expected to occur in the case of long-term weightlessness ([ref.150]).
chapter 5
To support biological investigations in space and to accumulate baseline data needed for manned space flight, NASA has conducted a program in laboratory research and theory. A multidisciplinary approach has included such fields as ecology, physiology, organic and biological chemistry, engineering, electronics, and optics. Emphasis in this program has been placed on qualitative and theoretical rather than purely descriptive research, and the investigation of fundamental biological phenomena at all levels, from the molecular to the total life form.
MOLECULAR BIOLOGY
Research in molecular biology has included chemical, physical, biological, and theoretical investigations of prebiological conditions on Earth and, possibly, on other planets; studies of cellular inclusions; genetic material (DNA and RNA) and coding; as well as energy transfer in biological systems.
The understanding of prebiological conditions on Earth, and possible conditions on other planets, depends upon the nature of the complex chemical species which might be encountered. Scientists have shown that biologically important compounds, such as amino acids, can be generated by applying an electrical discharge, ultraviolet radiation, or heat to a gaseous mixture. Biologically interesting compounds can be removed from such a system by condensation or absorption; however, in the limited time and space available in such experiments, many compounds are not produced in sufficient quantity to be measured.
The National Biomedical Research Foundation (NBRF) and the National Bureau of Standards (NBS) are conducting an investigation on equilibria in multielement systems. The distribution of molecular species at equilibrium is independent of the way equilibrium was reached and is dependent only on pressure, temperature, and elemental composition. Many of the conditions which might have arisen naturally can be approximated by thermodynamic equilibrium. Compounds which can be formed at equilibrium need no special mechanism to explain their presence. However, special mechanisms have to be sought for those compounds which could not be so produced and which would have been required for the structure and nutrition of the first living organisms.
In the absence of precise knowledge of the composition of the primitive planetary atmospheres, equilibrium concentrations with a wide range of temperatures, pressures, and elemental compositions are being investigated by NBRF and NBS. These investigators have postulated that the maximum atmospheric pressure may have approached 100 atm if the primitive Earth was sufficiently hot and if an appreciable portion of the water on Earth's surface today was present on primitive Earth. (If the present oceans were to evaporate, the surface pressure would be approximately 300 atm.) Low pressures of 10-6 atm and temperatures between 500° and 1000° K are being used.
A large range of N, O, C, and H compositions are being investigated for interesting and plausible combinations of factors. In these calculations an IBM 7090 computer is being used to obtain data on a very large number of combinations of chemicals. Other chemical species will be added as the research continues. Some results of this study give an insight into the variety of biologically significant chemicals which might have existed during Earth's primitive prebiological condition or may now exist on the surfaces and in the atmospheres of other planets (refs. [ref.151]-[ref.153]). The general method described by White et al. ([ref.152]), minimizing the free energy of the system, was used. The solution was approached by an iterative process, starting with an initial guess of concentrations of the compounds. At each step, M+1 linear equations are solved where M is the number of elements in the system.
In addition to listing of the concentrations of all compounds included in each problem, the results of three-element problems have been expressed on a triangular composition diagram for convenience. A coarse grid of 60 points is used to survey all elemental compositions, with finer grids being used in regions of particular interest. The calculated concentrations of the compounds at each composition are stored, and finally a series of triangular diagrams is printed out, each showing the concentrations of as many as four compounds at the grid points.
Figure 2 shows the results obtained in the C, H, and O systems. Organic compounds in concentrations greater than 10-20 mole fraction are found everywhere except where free O2, is present. Solid carbon theoretically becomes stable along the lower dashed line at 500° K. However, reactions producing it are very slow. The supersaturated region beyond the line of potential carbon formation was also investigated. A threshold was found where polynuclear aromatic compounds are sufficiently concentrated to form a liquid phase. These conditions may have been involved in the primordial formation of asphaltic petroleum.
Jukes and associates ([ref.154]) at the University of California at Berkeley have been investigating the code for amino acids in protein synthesis, the key for translating the sequence of bases in DNA into the sequence of amino acids in proteins. The amino acid code was solely a matter of theory until Nirenberg and Matthaei ([ref.155]) at the National Institutes of Health carried out a crucial experiment. This experiment bridged the last remaining gap separating theoretical genetics and test-tube biochemistry. It now became experimentally possible to search for codes for all 20 amino acids concerned in the synthesis of proteins.
The amino acid bases of DNA are: A, adenine; C, cytosine; G, guanine; T, thymine; and U, uracil, which replaces thymine in RNA. There are only 16 ways of arranging A, C, G, and T in pairs. For this and other reasons it is thought that a triplet of three consecutive bases is needed to code for each amino acid. The sequences of bases in a strand of DNA are known to be unrestricted with respect to the order in which they occur; apparently any one of the four bases can be next to any of the other four, although, of course, each base must be paired with the corresponding complementary base in the adjacent strand. Since the same freedom is true of the amino acid sequences in the polypeptide chains of proteins, any one of the 20 amino acids can occur next to any other. Moreover, the sequences in DNA are subject to mutational changes in which one base replaces another, or bases are added to or deleted from the DNA. Such rearrangements plus the possibility of lengthening of DNA molecules are numerous enough to account for all the genetics of living forms since the first appearance of life on Earth.
Most of our knowledge is based on experiments with synthetic RNA carried out with extracts of E. coli. The majority of the work has been at Nirenberg's laboratory at the National Institutes of Health and at Ochoa's laboratory at New York University ([ref.155]). Various combinations of A, C, G, and U were used in preparing the synthetic RNA molecules that are used in experiments to explore the code. These molecules are made by incubating a mixture of ribonucleoside diphosphates with a specific enzyme, polynucleotide phosphorylase. An important property of this enzyme is that it condenses the nucleoside diphosphates into polynucleotide strands containing random sequences depending on the proportion of each base. For example, if the enzyme were furnished with a mixture of 5 parts of A and 1 part of C, it would make strands containing, on the average, 25 sequences of AAA, 5 of AAC, 5 of ACA, 5 of CAA, and 1 each of ACC, CAC, and CCA. The proportion of triplets within the strands of a polynucleotide is reflected in the proportion of amino acids in polypeptides that are obtained in the cell-free system. Most of the present knowledge of the amino acid code is based on this concept. All the proposed codes have been discovered by this experimental approach where synthetic RNA molecules are used as "artificial" messenger RNA.
Representative of another class of activities in molecular biology is the examination of passive ion flux across axon membranes. This work is being done by Goldman at the National Naval Medical Center. The question of stimulus transmission by nerve tissue is far from simple, and the ion concentrations associated with nerve membranes is a significant part of the answer. Because the space environment may very well produce alterations in these ion potentials, an investigation of their natures and significance becomes extremely important. A working theory is now being developed as a result of this study.
Vital cell processes, chemical transformations, and mechanisms that provide energy for cell maintenance and activity have been studied by Kiesow (refs. [ref.157] and [ref.158]) at the Naval Medical Research Institute. The common objective of all phases of this project is the elucidation of reaction steps in which energy and matter are transformed in living systems. Compared with photosynthetic organisms, chemosynthetic bacteria offer distinct advantages for the study of energy assimilation. These studies have led to the following experimental findings.
With the energy from oxidation of nitrite, NO2— to nitrate, NO3— as an inorganic source, and with added organic chemical energy from the hydrolysis of adenosinetriphosphate (ATP) to adenosinediphosphate (ADP) and inorganic phosphate, chemosynthetic bacteria are capable of reducing diphosphopyridinenucleotide (DPN+) to DPNH, in a coupled oxidoreduction-dephosphorylation. Thus, in the crucial step of chemosynthesis, ATP is consumed, not produced. However, in simultaneously proceeding cell respiration, the energy donor, DPNH, is oxidized and generates more ATP than is required for DPN+ reduction. This "breeder cycle" for DPNH—with different ratios of cell respiration and biosynthesis—results in a net production of either DPNH, or ATP, or both. Production of DPNH in the cycle leads immediately to the assimilation of C14 from HC14O3—. These observations explain the bacteria's energy source without the classical hypotheses of either direct phosphorylation or direct CO2 reduction by inorganic chemical or electromagnetic energy. The cycle transforms the free energy of nitrite oxidation into the free energy of the organic compounds. Cell respiration and elementary biosynthesis proceed through structure-bound enzyme systems in the same fraction of subcellular particles. Three components, two cytochromes and one flavoprotein, have been identified. A thermodynamic analysis of the DPNH "breeder cycle" appears to be attainable by measurements of redox potentials and calorimetric determinations of heats of reaction.
Studies are also being conducted by Pollard and associates at Pennsylvania State University in an attempt to formulate a theoretical basis for the description of the processes of synthesis, growth, division, and differentiation of the living cell. Such a theory would be basic to an understanding of very primitive life forms or prebiological material which might be found elsewhere in the universe. For these purposes, studies are being undertaken in macromolecular reproduction which differ from the studies involving cellular genetic material. Theories concerning the problem of replication of cellular structures and information storage in two-dimensional systems are being developed. Theories are also being developed about the mechanisms which control and regulate receptor and enzymatic activities within the cell.
One study involved the rate of mutation in cells and disposed of the suggestion that the process of mutation consists of a "tunneling" of proton from one base to another in DNA. Such a suggestion can no longer be advanced as a major explanation of mutations.
Work is also being conducted on the centrifugation of cells of E. coli. It has been shown that cells exposed to as little as 100 g have a modification in their function. This has been looked at from the point of view of thymine uptake, which would be concerned with the formation of DNA, and also from the point of view of the induction of an enzyme, which would correspond to the transcription of the DNA. Preliminary experiments in the latter case indicate considerable centrifugation effect. The thymine uptake is affected, but not nearly as much as formerly thought. Further work is in progress in this area.
Important work has been completed on the cells of E. coli grown on maltose, which can be induced to produce betagalactosidase by the addition of thiomethyl galactoside. If cells are irradiated shortly after induction, the transcription of the DNA ceases and the enzyme produced by the messenger RNA is observed to reach a maximum. This enables the calculation of the half-life of unstable messenger RNA. The half-life for this decay is readily measurable, and values are given over a temperature range of 17° C (5.2 minimum) to 45° C (0.56 minimum). These agree very well with half-lives measured by others by inducing for short times and measuring the course of enzyme formation. The rate of transcription is involved in the kinetics of cessation of enzyme induction, and the rate of transcription can be measured. Arrhenius plots for this rate and the rate of decay are given, and the activation energies measured are about 16 000 cal/mole. The cessation of transcription is linked to the degradation, possibly of only one strand, of DNA.
Pollard has suggested that one important action of ionizing radiation is concerned with the transcription of the genetic message into RNA. Clayton and Adler ([ref.159]) showed that induced catalase synthesis in Rhodopseudomonas spheroides is inhibited by low doses of X-rays, giving experimental support to the idea. Pollard and Vogler ([ref.160]), using cells in which the process of induction involved permease, showed that there is some sensitivity to gamma radiation. Novelli et al. ([ref.161]) found a reduced sensitivity as compared with colony formation, but it is still a considerable sensitivity.
The process of induction of an enzyme indicates that the transcription of the genetic message is repressed by something which can be acted on by a small molecule, the inducer, to remove repression and permit the formation of messenger RNA, which then acts to make the enzyme. The messenger RNA undergoes decay through a process which is still not clear. Very elegant measurements by Kepes ([ref.162]) show that for the messenger RNA for betagalactosidase, the half-life is 1.02 min at 37° C and 2.05 min at 25° C. The time of onset of enzyme formation after induction was found to be about 3 minutes.
If the process of transcription is indeed sensitive to ionizing radiation, then the irradiation of cells which have just been induced should show formation of the enzyme to the extent of formation of new messenger RNA within a few minutes, plus the formation of the enzyme while the messenger RNA is decaying. This pattern was found by Clayton and Adler. The experiments conducted by Pollard and associates amplify and extend their work and also agree with the work of Kepes ([ref.162]).
BIOINSTRUMENTATION
Fernandez-Moran (refs. [ref.163]-[ref.165]), at the University of Chicago, has devised a new multielectrode electrostatic lens which he has incorporated into an electron microscope. This necessitated the development of a novel high-voltage power source and voltage regulator of extreme stability and accuracy. Some promising work has now been done on superconducting lenses. In a series of experiments with a simple electron microscope without pole pieces, using high-field superconducting niobium-zirconium solenoid lenses in an open air core, liquid helium Dewar, electron microscopic images of test specimens have been recorded while operating at 32 200 gauss in a persistent current mode, with regulated accelerating potentials of 4 to 8 kilovolts. These preliminary experiments have demonstrated the exceptional stability of the images (both short term and long term) over a period of 4 to 8 hours and the relatively high quality of the images.
Progress has been made on the viscosimeter for high intrinsic viscosities. This is now working, and the viscosity of DNA preparations has been measured. It is hoped to use the viscosimeter to study the variation in DNA viscosity as a function of the cell cycle.
An instrument is under development by Wald at the University of Pittsburgh to automatically analyze cytogenetic material and, thus, extend cytogenetic methodology both for research and as a biological monitoring procedure, using automatic electronic scanning and computer analysis of chromosomes. Chromosomal aberrations can thus be monitored under unusual and abnormal conditions such as weightlessness and radiation, since chromosomes are very sensitive to stress situations. In this device a sample will be prepared and automatically inserted under a microscope lens. The device will then scan, identify, and photograph on 35-mm film a predetermined number of mitotic cells and process the film. The data will be recorded under the direct control of a digital computer. The computer will perform a detailed quantitative analysis of the pictorial data.
Significant effort has been expended in the development of instrumentation for measuring and recording electrophysiological information. One such instrument, developed by the Franklin Institute, Philadelphia, Pa., is a temperature-sensing microprobe. This microprobe is an implantable and remote broadcasting instrument. These developments are associated, in part, with training programs so that competent individuals may be trained not only in electronics but also in the biological uses of the devices they construct.
A project of interest, conducted at the Stanford Research Institute, is the investigation of the uses of an extremely sensitive method for measuring magnetic susceptibility having the possibility of detecting macroscopic quantum effects in macromolecules of biological interest. Good progress has been made in the first 15 months of a project devoted to the development and initial use of equipment specifically designed for this purpose. A new superconducting circuit, together with superconducting magnetic shields, has been constructed. This apparatus can measure the magnetic susceptibility of small organic samples at temperatures between 1° and 300° K in fields up to 40 000 gauss. It can detect flux changes of 107 gauss-cm2, which is equivalent to detecting a change in specific susceptibility of 1 in 109 in a 100-mg sample under an applied field of 10 000 gauss.
Several hundred preliminary measurements were made on samples of coronene. The most reliable of these were in agreement with published values of the magnetic susceptibility of coronene. Experience during these measurements led to changes which have resulted in an apparatus well suited to the measurements on macromolecules. An improved version of the superconducting circuit now available shows promise of a further improvement in sensitivity by a factor of more than a thousand ([ref.166]).
Living organisms possess many unique processes and systems which are complex and poorly understood. The new theoretical approaches, combined with laboratory studies, are expected to result in advances which will expand both our scientific and technological horizons.
chapter 6
BALLOONS
Biological and medical experiments carried out on balloon flights, both manned and unmanned, antedate the establishment of NASA. Aside from the early use of balloons in flights that could be called simply flight-survival studies, balloons have made important contributions to our present knowledge of the effects of cosmic radiation and to various aspects of space travel.
The achievements of the Strato-Lab and Man High series by the U.S. Navy and Air Force include a wealth of information on balloon travel and on the survival of man at altitudes close to and above 100 000 feet. Generally, balloon launches of animals, which reached a maximum in 1953 when 23 balloons were released, have established the feasibility of a program of extended manned balloon flights to high altitudes.
Atmospheric life studies outside the area of cosmic radiation effects have been comparatively few. Results from two manned flights, Strato-Lab I and II, indicate that the flights did produce pronounced changes in white blood cell count; however, the data suggest that these changes were due to psychological rather than physical stress. Exposure to altitudes above 90 000 feet for a total of 62 hours did not produce any general behavioral change in two Java monkeys, according to other balloon flights. Many of these flights were effective in testing equipment, telemetering devices, and in pointing the way for other flights.
Stratoscope I and II, originally undertaken by the Office of Naval Research (ONR), are projects involving various astronomical observations with the aid of a balloon-borne telescope and television and camera systems. NASA cooperated with ONR on Stratoscope II (36-inch telescope compared with Stratoscope I's 12-inch telescope) which has already resulted in significant discoveries about the nature of the planets and stars. Water vapor has been identified in the atmosphere of cool red stars and an analysis of the Martian spectra showed a greater abundance of carbon dioxide than had previously been believed. Since the balloon-borne telescope was carried beyond Earth's obscuring atmosphere, the Stratoscope projects have yielded valuable photographs of the Sun, stars, and various planets.
ROCKETS AND SATELLITES
Historically, biological experiments aboard rockets and satellites have been limited to a "piggyback" and "noninterference" basis on military rockets. For the past few years, however, as the effort toward manned space flight leading to lunar and Martian landings increased, more attention was devoted to experiments designed to show the effects of the space environment on living systems. As in the balloon flight programs, the U.S. Army, Navy, and Air Force played an important role, reaching what might be considered a high point with the successful launch and recovery of a ballistic rocket experiment with monkeys Able and Baker. Aerobee rockets as well as Thor IRBM's carried biological payloads consisting of mice and monkeys on six launches, contributing to our knowledge of the effects of weightlessness and radiation on higher animals.
Van der Wal and Young ([ref.78]) used Thor-Able combinations to serve as boosters for lifting a 20-pound biocapsule to a peak altitude of 1400 miles and over a distance of about 5300 miles from Cape Canaveral to the west coast of Africa. Weightlessness was attained for a period of almost 40 minutes. During reentry into the atmosphere, a peak deceleration of about 60 g was reached. Each of the three capsules flown carried one mouse (Mouse-in-Able); two of the mice were instrumented for heart-rate telemetry. Although all three mice were lost, the two experiments with Laska and Benji yielded physiological results.
The experimenters designed effective instrumentation for registering the electrical activity of the mouse's heart through a single commutated telemetry channel. Records were obtained for both animals during various portions of the flight. The results indicate that both animals were alive when the nose cones hit the water.
Two South American squirrel monkeys (Gordo and Baker) and a rhesus monkey (Able) were launched into space from Cape Canaveral in 1958 and 1959 by U.S. Army Jupiter missiles. The vehicles reached speeds of approximately 10 000 mph and altitudes of 300 miles on flights which lasted about 15 min.
Time courses of cardiac and respiratory rates ([ref.80]) of the two squirrel monkeys showed that the noise of the engine at liftoff immediately produced an increase in their heart rates. Respiration also increased temporarily, but slowed later with increasing acceleration. Heart rates fluctuated considerably during launch acceleration, which reached about 15 g at cutoff.
The period of free flight and weightlessness was characterized by pronounced fluctuations of heart activity in the postacceleration phase. Thereafter, the heart rate of Baker remained relatively constant, whereas the cardiac activity of Gordo fluctuated markedly and decreased slowly almost to the end of his flight. Slight changes, which were transient and not pathological in nature, were also noted in the electrocardiogram. Gordo's respiration was very shallow during maximum launch acceleration, when Baker's reached its highest value, only to be approximated again during reentry when forces of about 35 g were encountered.
Able's cardiac and respiratory rates indicated that, after an initial startle reaction, the heart rate dropped transiently and then increased steeply, reaching a maximum of 259 during the 10-second interval at peak acceleration. Respiration increased only slightly throughout the launching phase. There was a period of tachycardia during postacceleration weightlessness, after which the heart rate declined steadily and was disturbed only by several startling missile events. At the end of the subgravity phase, Able's cardiac rate was slightly below normal.
Although the periods of high g force and free flight were short, the extremes were considerable, and the changes from one state to the next were rapid. In spite of this, the cardiovascular, hemodynamic, and electrocardiographic phenomena were remarkably well maintained. Apparently the animals were not in serious plight at any time. That psychological factors entered into the observed phenomena is clearly evident from the increase in cardiac rate associated with the noise of the engine prior to liftoff and also from the cinematographic record of facial expressions. Nevertheless, the integrated responses indicated that the animals' physiological states remained sufficiently normal to insure a safe flight.
LITTLE JOE FLIGHTS
The first step in an attempt at animal verification of the adequacy of the Mercury flight program was the development of two tests by NASA in collaboration with the U.S. Air Force School of Aviation Medicine in which there would be a biomedical evaluation of the accelerations experienced during the abort of a Mercury flight at and shortly after liftoff. These flights were launched at the NASA Wallops Station with a Little Joe solid-fuel launch vehicle.
Two Little Joe launches were made with activation of the escape rockets during the boost phase to secure maximum acceleration; only a brief period of weightlessness was attained. The first launch was on December 4, 1959, and the other on January 21, 1960. A 36 by 18-inch sealed, 125-pound, cylindrical capsule containing the subject, an 8-pound Macaca mulatta, the necessary life-support system, and associated instrumentation was flown in a "boilerplate" model of the Mercury spacecraft. The rhesus monkeys were named "Sam" and "Miss Sam."
The flight profile included maximum accelerations of about 10 to 12 g and periods of about 3 minutes at 0±0.02 g. The peak altitude obtained in the last ballistic flight was about 280 000 feet. The experimental capsule was pressurized at 1 atmosphere with 100 percent oxygen at the start of the experiment and fell to just below a half atmosphere of oxygen due to breathing during flight. The capsule temperature was kept between 10° and 20° C in both flights.
The measurements taken from the rhesus monkeys were the electrocardiogram, respiration, body temperature, eye movements, and bar pressing, but only partial results were obtained in the first flight. Oxygen tension, total pressure, capsule temperature, and relative humidity were recorded. Both animals were recovered alive and did not show pathologic alterations in their physiologic and psychological reactions.
MERCURY ANIMAL TEST FLIGHTS
In the Mercury animal test program a Redstone missile carried the chimpanzee Ham on a ballistic flight to a height of 155 miles to provide animal verification of the success with which the Mercury system could be applied to manned flight. The male chimpanzee was trained to perform a two-phased reaction task during the 16 minutes of flight. The chimpanzee Enos was put into orbit for 3 hours and 20 minutes. Results of the two flights gave the following information:
Pulse and respiration rates during both the ballistic (MR-2) and the orbital (MA-5) flights remained within normal limits throughout the weightless state. Effectiveness of heart action, as evaluated from the electrocardiograms and pressure records, was also unaffected by the flights.
Blood pressures, both arterial and venous, were not significantly changed from preflight values during 3 hours of the weightless state.
The performance of a series of tasks involving continuous and discrete avoidance, fixed ratio responses for food reward, delayed response for a fluid reward, and solution of a simple oddity problem was unaffected by the weightless state.
Animals trained in the laboratory to perform during simulated acceleration, noise, and vibration of launch and reentry were able to maintain performance throughout an actual flight.
From the results of the MR-2 and MA-5 flights, the following conclusions were drawn:
The numerous objectives of the Mercury animal test program were met. The MR-2 and MA-5 tests preceded the first ballistic and orbital manned flights, respectively, and provided valuable training in countdown procedures and range monitoring and recovery techniques. The bioinstrumentation was effectively tested and the adequacy of the environmental control system was demonstrated.
A 7-minute (MR-2) and a 3-hour (MA-5) exposure to the weightless state were experienced by the subjects in an experimental design which left visual and tactile references unimpaired. There was no significant change in the physiological state or performance of the animals as measured during a series of tasks of graded motivation and difficulty.
Questions were answered concerning the physical and mental demands that the astronauts would encounter during space flight, and it was shown that these demands would not be excessive.
It was also demonstrated that the young chimpanzee can be trained to be a highly reliable subject for space-flight studies.
The suborbital ballistic flight of Ham on January 31, 1961, was the prelude to Alan R. Shepard's suborbital space flight, while the orbital flight of Enos on November 29, 1961, preceded the orbital flight of John H. Glenn.
The fact that we now categorize these events as belonging to the rather distant past, although they occurred only about 4 years ago, serves to emphasize the pace of development in the exploration of space. While the chimpanzee program may pale in the light of subsequent successes, its scientific and technological contribution should not be overlooked.
The significance of this project can be fully appreciated, and its contribution judged, only by considering the lack of knowledge existing at the time of its conception. In addition to its essential training function, this project verified the feasibility of manned space flight through operational tests of the Mercury life-support system. It demonstrated that complex behavioral processes and basic physiological functions remained essentially unperturbed during brief exposures to space flight. The Mercury chimpanzee program marked the first time that physiological and behavioral assessment techniques were combined for evaluating the functional efficiency of the total organism in space.
Perhaps the ultimate contribution of this program was in providing the framework of knowledge upon which future scientific experiments on biological organisms, exposed to flights of extended durations, must be based. Biosatellite experiments designed to seek more subtle and elusive effects of prolonged space flight on biological functioning will require even more refined and difficult techniques, but will depend heavily on the groundwork laid in these early steps of Project Mercury.
A summary of the more important animal suborbital and orbital flights during the period 1957 to 1964 is presented in table VII.
In another NASA-supported flight, NERV 1, various experiments were carried in a suborbital flight of 20 minutes. Neurospora molds showed a surprisingly high level of mutation, but the control molds also had high rates.
The Discoverer XVII and XVIII flights, to which the Air Force contributed, resulted in many interesting findings relative to the responses of living systems to space flight. On the Discoverer XVII flight, samples of human gamma globulin and rabbit antiserum specific for human gamma globulin showed an increase in reactivity, and samples of synovial and conjunctival cells showed no changes in their cytological characteristics.
Discoverer XVIII was launched during a massive solar flare which lasted for the first 13 hours of the 48-orbit, 3-day flight. Neurospora conidia, nerve tissue, algae, human bone marrow, eyelid tissue, gamma globulin, and cancer cells were put in orbit. The results indicated that biological specimens may be able to withstand radiation from solar flares with a minimum of shielding and that aluminum shielding may be better than lead.
In 1949, the U.S.S.R. began a systematic, uninterrupted research program in biological space experimentation. They have studied the effects of physical stress, immune reactions, psychobiology and behavior, genetics, and responses to environmental factors such as spacecraft dynamics and ambient radiation. The organisms and biological materials included tobacco mosaic and influenza viruses; T2 and T4 bacteriophage; Bacillus aerogenes; lysogenic bacteria; Clostridium butyricum; Escherichia coli; actinomycetes; yeasts; Chlorella pyrenoidosa; seeds of fir, pine, onion, corn, lettuce, wheat, cabbage, carrot, buckwheat, cucumber, beet, Euonymus, fennel, mustard, pea, broad bean, tomato, and nutmeg; Tradescantia paludosa; Ascaris eggs; snail spawn; Drosophila melanogaster; loach roe; frog eggs and sperm; guinea pigs; mice; rats; hamsters; rabbits; dogs; monkeys; human and rabbit skin; HeLa tissue cultures and other tissues (refs. [ref.167] and [ref.168]).
Year |
Animal subject |
Flight profile |
|---|---|---|
United States |
||
1958 |
Mice Wickie, Laska, and Benji |
1400 miles. None of the three flights were recovered. |
1958 |
Squirrel monkey Old Reliable |
300-mile maximum altitude over a 1300-mile distance via a Jupiter rocket. Not recovered. |
1959 |
Rhesus monkeys Able and Baker |
300-mile maximum altitude over a 1500-mile distance via a Jupiter rocket. Recovered. |
1959 |
Black mice |
500 seconds of weightlessness in Discoverer III via a Thor-Able rocket. The Discoverer vehicle did not go into orbit and the animals were lost. |
1959 |
Rhesus monkey Sam |
53-mile altitude in Little Joe. Recovered. |
1960 |
Rhesus monkey Miss Sam |
9-mile altitude in Little Joe. Recovered. |
1960 |
C-57 black mice |
650-mile altitude over a 5000-mile distance via Atlas RVX-2A. Recovered. |
1961 |
Chimpanzee Ham |
156-mile altitude over a 414-mile distance via a Redstone booster, Mercury capsule. Recovered. |
1961 |
Chimpanzee Enos |
2 Earth orbits. 183 minutes of weightlessnessat an apogee of 146 miles anda perigee of 99 miles. Atlas booster, Mercury capsule. Recovered. |
Soviet Union |
||
1958 |
Dogs Belyanka and Pestraya |
280-mile altitude in hermetically sealed cabin. Recovered. |
1959 |
Dog Otyazhnaya and a rabbit |
Over 100-mile altitude. Recovered. |
1960 |
Dogs Belka and Strelka, 21 black and 21 white mice |
16 Earth orbits (24 hours) via Sputnik V. First successful recovery of living creature from orbital flight. |
1960 |
Dogs Pchelka and Mushka |
16 Earth orbits (24 hours). Spacecraft destroyed during reentry. |
1961 |
1 dog, mice, guinea pigs, and frogs |
1 Earth orbit at an apogee of 155 miles and a perigee of 114 miles. Recovered. |
1961 |
Dog Laetzpochka |
1 Earth orbit. Recovered. |
France |
||
1961 |
Rat Hector |
95-mile attitude in a capsule boosted by a Veronique rocket. Recovered. |
1963 |
Cat Felicette |
95-mile altitude in a capsule boosted by a Veronique rocket. Over 5 min of weightlessness. Recovered. |
THE NASA BIOSATELLITE PROGRAM3
The space environment offers a unique opportunity to study the basic properties of living Earth organisms with new tools and opens up new areas of research for which biological theory fails to provide adequate predictions. Unique components of the space environment of biological importance are weightlessness or greatly decreased gravity, the imposition of an environment disconnected from Earth's 24-hour rotation (particularly its effect on biorhythms), and cosmic radiation with energies and particle sizes unmatched by anything produced artificially on Earth ([ref.169]).
As progress is made in the manned exploration of space, the biological effects of its unique environmental factors become of greater importance. It is essential to determine the effects of space environment on man's ability to perform physical and mental tasks. In addition, it is necessary to develop those systems required for his survival and for his physiological and psychological well-being, both in space and in his subsequent resumption of normal life patterns. Despite nearly a century of research and development in environmental physiology, a number of phenomena will be encountered in long-term space flight with which we have had neither the experience that would enable us to predict the effects nor to develop the necessary protective or remedial measures ([ref.170]). Many of the experimental programs in bioscience are being carried out or planned so that the deleterious effects of these phenomena may be determined, predicted, or avoided before they are encountered in manned flight.
Biological experimentation has been carried out in orbiting spacecraft by Soviet and American scientists preparatory to manned space flight. These first-generation exploratory experiments had the following objectives:
To discover whether complex organisms could survive space conditions and to test life-support systems
To determine whether complex organisms (dogs and primates) could survive launch, orbital space flight, reentry, and recovery
To determine the effects of space radiation and any obvious effects of weightlessness on biological organisms
These biological studies indicate that manned space flight was practicable, and the various cosmonaut and astronaut flights have proven the validity of the results.
The National Academy of Sciences' Space Science Board summer study ([ref.171]) recommended that—
NASA should exploit special features of the space environment as unique situations for the general analysis of the organism-environment relationships including, especially, the role environmental inputs play in the establishment and maintenance of normal organization in the living system. NASA should support studies in ground-based and in orbiting laboratories [biosatellites] on the biological effects of gravity fields both above and below normal. This should be considered a major responsibility of NASA in the area of environmental opportunities. NASA should support studies of biological rhythms in plants and animals including man as part of its effort in environmental biology. Investigate by observation of rhythms in organisms in space in (a) polar and equatorial low orbits; (b) orbits less than, equal to and greater than 22,000 miles. Properly designed experiments should be conducted to explore the effects of different environmental factors when these impinge simultaneously on test organisms.
The Panel on Gravity of the Space Science Board ([ref.67]) stated that the major effects of low gravity would be expected in heterocellular organisms that develop in more or less fixed orientation with respect to terrestrial gravity and which respond to changes in orientation with relatively long induction periods, including the higher plants. On the other extreme are the complex primates which respond rapidly, but whose multiplicity of organs and correlative mechanisms make the occurrence of malfunction and disorganization probable, but not certain. The Panel recommended emphasis on early embryogenesis and histogenesis, particularly of plants during exposure to low gravity, and anatomical studies after low gravity. They stated that perturbations of the environment to which the experimental organism is exposed must be limited or controlled to reduce uncertainties in interpretation of results. At the same time, the introduction of known perturbations may assist in isolating the effects due solely to gravity. Ground-based clinostats and centrifuges should be used in conjunction with the experiments, and an attempt should be made to extrapolate effects of low gravity with the clinostat.
The study of the effects of unique or unknown space environmental factors will probably yield unexpected results which may drastically modify future technical approaches. The results from these biosatellite studies will have broad application to longer term, manned space flight, including manned space stations and lunar and planetary exploration.
The biosatellite program is a second-generation series of carefully planned and selected experiments, including some highly sophisticated experiments which have required several years of baseline study and equipment development. These orbiting recoverable biosatellites will provide opportunities for critical testing of major biological hypotheses in the areas of genetics, evolution, and physiology.
The scientific community showed great interest in the biosatellite program, and scientists from universities, industry, and Government have submitted 185 flight experiments involving primates and other mammals, vertebrate and invertebrate animals, micro-organisms, and plants.
The selected biosatellite experiments include studies at the cellular, tissue, and organism levels, including embryological development and growth experiments at the tissue level and physiological, behavioral, reproductive, and genetic studies at the organism level. The experiments are divided into six categories:
Primates
Mammals (nonprimate)
Animal, cellular, and egg
Plant morphogenesis, photosynthesis, and growth
Biorhythm
Radiation
Twenty experiments have been selected for flight to study the effects of weightlessness and decreased gravity during 3- to 30-day orbital periods. The experiments include a wide variety of plants and animals from single-celled organisms to higher plants and animals. The effects of weightlessness on the primate will be studied, especially the central nervous, the cardiovascular, and the skeletal systems during 30-day orbits.
Experiments have been selected to study the genetic and somatic effects of weightlessness combined with a known source of radiation (Sr85) to determine if there are any antagonistic or synergistic effects ([ref.172]). Experiments are also included for studying the effects of the unique environment of the Earth-orbiting satellite and removal from the Earth's rotation in relation to biological rhythms of plants and animals.
Six biosatellites are included in the presently approved program, with the first flight in 1966. They will be launched from Cape Kennedy by the improved two-stage, thrust-augmented Thor-Delta into a nearly equatorial circular orbit at an altitude of 180-200 miles for periods up to 30 days. Recovery will be by Air Force airplane during capsule/parachute descent. The spacecraft weigh 1000-1200 pounds, have a 280-pound recoverable capsule and, while in orbit, will not experience greater than 1/10 000 g of acceleration. The life-support system will provide an environment at sea-level pressure of 80 percent nitrogen, 20 percent oxygen, and no more than 0.5 percent carbon dioxide with a temperature of 75° F ±5° F.
All experiments are in various stages of development or testing and flight test hardware has been and is being constructed. The experiments and hardware are being subjected to preflight tests simulating launch and recovery stresses. Rhesus, pigtail, and squirrel monkeys have been subjected to the dynamic forces of the simulated flight under conditions of complete, partial, and no restraint. Three types of centrifuges have been used to simulate the flight profile. Primates were fully instrumented with deep brain electrode implants, implanted catheters, and other implanted sensors. During centrifugation, motion pictures were taken. These primates were semirestrained in form-fitted couches which allowed movement of the body while facing the accelerative force in a ventrodorsal position (eyeballs in). In this series of tests, all primates were normal following the tests and exhibited no unusual behavior or effects. X-rays showed that implanted catheters and electrodes remained in place, and there were no movements causing tissue damage. However, when the primates were placed with their backs toward the accelerative force, dorsoventral (eyeballs out), the animals suffered visible damage. At 6 g there was no visible stress, but at 8 g swelling of the lower eyelids was noticeable. At 11 g both eyelids were swollen shut. In the biosatellite program, primates will be placed in the semirestraint couches in a position facing accelerative forces, ventrodorsal (eyeballs in), to prevent these effects.
chapter 7
BIOREGENERATIVE LIFE-SUPPORT SYSTEMS
Placing a man in space requires a complete life-support system capable of supplying sufficient oxygen, food, and water and removing excess carbon dioxide, water vapor, and human body wastes. In addition, the oxygen, carbon dioxide, and pressure must be maintained at a suitable level. Any accumulated toxic products and noxious odors must be removed.
In the spacecraft the human is confined in a restricted environment in which it is necessary to establish a balanced microcosm or closed ecological system. This is an enormous biological and bioengineering problem. Weight, size, simplicity of operation, and reliability particularly are important factors.
For relatively short missions involving one or several astronauts, food, oxygen, and water can be stored and made available as required, and the various waste products can be stored. On longer missions, particularly those involving more than one astronaut, efficient chemical or biological regenerative systems will be required. Any regenerative system introduces a fixed cost in weight of processing equipment and energy requirements.
Chemical, or partially regenerative, methods for providing breathing oxygen by the regeneration of metabolic products such as water vapor and carbon dioxide include the thermal decomposition of water and CO2, photolysis and radiolysis of water, electrolysis of fused carbonates and aqueous solutions, and the chemical reduction of CO2 with H2, followed by electrolysis of the water formed. Chemical regenerative systems have been developed to remove excess carbon dioxide and water vapor from the atmosphere. Nonbiological regenerative systems are time limited by the amount of food, water, and oxygen that can be carried or recovered. These physical-chemical processes show great potential, but they also present many difficulties, including requirements for extremely high temperatures and considerable amounts of power, the formation of highly toxic materials, and high susceptibility to inactivation. None of the presently studied nonbiological processes can function as completely as a bioregenerative system. All these nonbiological systems have unrealistic supply requirements and produce unusable wastes. Consequently, for long planetary missions the bioregenerative systems, though also beset with problems, are potentially far superior to their physical and chemical counterparts.
Table VIII shows average daily metabolic data for a 70-kg astronaut. A man breathes about 10 cubic feet of air per minute, or 400 000 liters, daily. The expired air contains about 4 percent carbon dioxide. Man normally breathes air containing 0.03 percent CO2, but can withstand comfortably about 1.5 percent CO2. Anything in excess of 1.5 percent will produce labored breathing, headaches, and, if greatly exceeded, death. A man exhales about 1.1 pounds of water per day and this, in addition to water from perspiration and other sources, must be removed from the air.
O2 input, kg |
0.862 |
CO2 output, kg |
1.056 |
Drinking water, liters |
2.5 |
Food rehydrating water, liters |
1 |
Caloric value of food, kcal |
3000 |
Water output: |
|
Urine, liters |
1.6 |
Respiration and perspiration, liters |
2.13 |
Feces, kg |
0.09 |
Total heal output, Btu |
11 100 |
Two types of biological regenerative systems have been proposed. The photosynthetic closed ecological system was proposed as early as 1951. This involves the use of single-celled algae or higher plants, including floating aquatic and terrestrial plants, and requires the interaction of light energy with CO2 and H2O to produce O2 and plant cells. Another system, proposed in 1961, involves electrolysis of water into oxygen and hydrogen, and the concurrent use of Hydrogenomonas bacteria which take up hydrogen, some oxygen, carbon dioxide, and urine yielding water and bacterial cells.
System |
Requirements/1 man4 |
Requirements/3 men (270 man-day mission)5 |
||
|---|---|---|---|---|
Weight, kg |
Power, kW |
Weight, kg |
Power, kW |
|
Partial chemoregenerative |
7 332 |
1.75 |
||
LiOH |
125 |
1.40 |
||
NaOH |
155 |
7.68 |
||
CO2-H2 |
34 |
.36 |
||
Full bioregenerative—algae: |
||||
Artificial illumination |
116 |
6 10.40 |
591 |
25.00 |
Solar illumination |
103 |
1.70 |
356 |
.60 |
Electrolysis-hydrogenomonas |
55 |
.25 |
129 |
2.60 |
The values given in table IX indicate relative weights and powers required by various systems to provide the gaseous environment for manned space cabins. If one considers operating temperatures and hazards, other systems may offer advantages which offset the weight and power advantages of the hydrogen reduction of LiOH systems.
Research is being conducted by NASA on life-support-system technology applicable to missions planned for 20 years in the future. Life-support systems include the requirements for supplying breathing gases, control of contaminants in the cabin atmosphere, water reclamation, food supply, and personal hygiene. The disciplines involved in such systems include biology and microbiology, cryogenic fluid handling at zero g, heat transfer, and thermal integration with other systems, such as power. The physiological, psychological, and sociological problems of the crew are also being considered.
Photosynthetic System
Green plants contain chlorophyll which captures light energy thermodynamically required to convert carbon dioxide and water into carbohydrate which can subsequently be transformed into other foods such as protein and fat. During this process, carbon dioxide is consumed, and an approximately equal amount of oxygen gas is liberated. As a first approximation, photosynthesis is the reverse of the oxidative metabolism of animal life:
OxidationC6H12O6 + 6O2 ———————> 6CO2 + 6H2O + heatPhotosynthesis6CO2 + 6H2O + light ———————> C6H12O6 + 6O2
The photosynthetic process in plants and respiration during photosynthesis have been studied intensively, and several metabolic pathways have been elucidated. Mechanisms are being studied to explain the inhibitory effect of strong visible light on this process. This program may lead to the use of chloroplasts or chlorophyll without cells in future photosynthetic bioregenerative systems for long-term space travel.
One of the prime considerations of a closed ecological system is that the environmental gases shall remain physiologically tolerable to all of the ecologic components. Ideally, a photosynthetic gas exchange organism should possess a high ratio of gas exchange to total mass (considering all equipment and material incidental to growth, harvesting, processing, and utilization); and a controllable assimilation rate to maintain steady-state gas composition. It should also be (1) amenable to confining quarters which may be imposed by inflexibility of rocket or space station design; (2) genetically and physiologically stable and highly resistant to anticipated stresses; (3) edible and capable of supplying most or all human nutritional requirements; (4) capable of utilizing raw or appropriately treated organic wastes; and (5) amenable to water recycling as demanded by other components of the ecosystem.
Higher Plants
Efforts to utilize multicellular plants as photosynthetic gas exchangers have been somewhat neglected, since it has been assumed by many that algae would be more efficient. The family Lemnaceae (duckweeds) are small primitive aquatic plants with a minimum of tissue differentiation. Practically all of the cells of the plant contain chlorophyll and are capable of photosynthetic activity. They reproduce principally by asexual budding of parent leaflike fronds. They can be grown readily on moist surfaces ([ref.177]) on almost any medium suitable for the growth of autotrophic plants. With duckweeds the problems of gaseous exchange and harvesting are simplified and the volume of medium can be greatly decreased as compared with algae.
Ney ([ref.177]) obtained a very high gas exchange rate with duckweeds. Using small cultures under controlled optimal conditions of temperature, light (600-1000 ft-c), and CO2, concentration, he estimated that 2.3 m2 of frondal surface of duckweed, at a gas exchange rate of 10.8 liters m2/hr would provide sufficient gas exchange for one man. This would produce about 25 grams of dry plant material per hour.
A few nutritional studies have been carried out with duckweeds. Nakamura ([ref.178]) considered Wolffia as a possible source of food for space travel and found that it contained carbohydrate 25-60 percent, protein 8-10 percent, fat 18-20 percent, minerals 6-8 percent (all dry weights), and vitamins B2, B6, and C, with C the most abundant.
One of the desirable features of a duckweed system is that the gas exchange is direct between the atmosphere and the plant and does not require dissolving the respiratory gases in a bulky fluid system which introduces special engineering difficulties in zero- or low-gravity conditions.
In the design of equipment for photosynthetic studies, careful consideration should be given to the material used in the construction of the unit. Most plastic materials are subject to photo-oxidative degradation, with CO as one of the products. When air is recirculated through plastic tubing and transparent rigid plastics in the presence of light, considerable quantities of CO are given off. With high-intensity illumination such as sunlight, a CO buildup of several hundred parts per million is not uncommon. Also, plant pigments such as the carotenoids and chlorophylls will react similarly when exposed to light of high intensity. If the plants die, then CO is released quite rapidly.
At Colorado State University the responses of plants to high-intensity radiation (ultraviolet to infrared) are being studied. Plants from high mountaintops that are exposed to greater ultraviolet light are being studied for specialized adaptations. The effect of temperature on photosynthesis is being explored. Various plants are also being studied under germ-free conditions.
Screening of higher plants for possible use in bioregenerative systems at Connecticut Agriculture Experiment Station resulted in the selection of corn, sugarcane, and sunflower. Under optimal conditions it has been shown that 100 to 130 ft2 of leaf surface are required to support an astronaut.
Plants considered as possible food sources include soybeans, peanuts, rice, and tomatoes, which can be combined with algae to give a well-balanced and reasonably varied diet. Hydroponic systems use large quantities of water, but progress is being made in reducing this.
The possibility of using animals in the closed ecological system is open to question, particularly in the absence of gravity, and much work remains to be done on using plant materials as animal food and on the disposal of wastes. Animals which have been considered are crustaceans, fish, chickens, rabbits, and goats.
Algae
Algae have the fastest growth rate and are among the most efficient plants for oxygen and food production. It has been amply demonstrated by Myers ([ref.179]) and other workers that Chlorella can be used in a closed ecological system to maintain animals such as mice and a monkey. The use of algae for supplying O2 and food, and for removing CO2 and odors has been considered by many authors for use in spacecraft, space platforms, and for establishing bases on the Moon or Mars.
Estimates of total efficiency are based on extrapolated laboratory data and vary widely, since many different types of data have been used as a basis for these estimates.
The respired air containing about 4-5 percent CO2 is bubbled into the Chlorella culture, at either atmospheric or increased pressure. Air containing a high percentage of oxygen and saturated with moisture is released from the algal system.
The use of algae for several purposes might require from one to three separate algal systems. For food production, Chlorella produces 50 percent protein and 50 percent lipids in high-nitrogen media. In low-nitrogen media, it produces 85 percent lipids. Proper choice of Chlorella strains and media will produce not only the necessary calories but also the necessary specific nutrients required. Certain strains are more effective in O2 production, and others in the use of urine and other wastes.
Some of the early estimates, using Chlorella grown at 25° C, for supplying these requirements for a single man in space include the following: 168 kg of algal suspension ([ref.179]), 200 kg of algal suspension and 50 kg of equipment including pumps (refs. [ref.180] and [ref.181]), and 100 kg of algal suspension and 50 cubic feet for equipment and gas exchange ([ref.182]). Using the blue-green alga Synechocystis, 600 kg of algal suspension would be required, according to Gafford and Craft. These estimates are based on preliminary studies, are quite high, and are not of real practical value.
Other studies have indicated an extremely efficient algal system which offers a real potential for a practical and effective gas exchanger ([ref.183]). A thermophilic strain of Chlorella with an optimum growth temperature of 39° C and an optimum temperature for photosynthesis of about 40° C can increase its cell mass 10 000-fold per day. When operating at one-half maximum efficiency, this alga produces 100 times its cell volume of oxygen per hour. Burk et al. ([ref.183]) state: "Future engineering development should lead to a space requirement, per adult person, of no more than 3 to 5 cubic feet of algal culture, equipment, and instrumentation for adequate purification of air." The requirements of this system would require additional energy in the form of light and of small amounts of nitrogenous and mineral material for the algae. The light source used by Burk et al. ([ref.183]) is a tungsten filament quartz lamp the size of a pencil, which has a long life, produces a luminous flux 5-10 times greater than sunlight on Earth, and operates at a 10-12 percent light efficiency.
Research is being carried out on algal regenerative systems by about 40 or 50 laboratories in the United States. NASA is supporting several basic studies on photosynthesis, the physiology of algae, and engineering pilot-plant development. Much of the research on algae is being supported by the Air Force.
Most algal studies have been carried out in small units and the data obtained have been used as a basis for extrapolating logistic values for the use of these organisms in manned space vehicles. Myers ([ref.179]) has shown that the quantity of algae necessary to support a man (with an assumed O2 requirement of 625 liters per day) would yield about 600-700 grams dry weight of new cells per day. If algal growth in mass cultures could be maintained in a steady-state concentration of 2.5 gram dry weight per liter with such a growth rate as to yield 10 grams weight per liter per day, the volume of algal culture would be 60-70 liters and the total mass of the system would approximate 200-250 pounds.
Using an 8-liter system, Ward et al. ([ref.176]) have produced algal concentrations of 5-7 grams of dry algae per liter with a high-temperature algal strain. The maximum growth rate observed with the culture was 0.375 gram dry weight per liter per hour, or 9 grams dry weight per liter per day. This was accomplished by using 1-centimeter layers of culture and a light intensity of 8000 foot-candles. The culture system consisted of a rectangular plastic chamber having an area of 0.5 square meter and illuminated on each side to an intensity of 4000 foot-candles (cool-white). To produce 25 liters of oxygen per hour, an area of 8.3 square meters (85 square feet) would be required.
The major problem in large-scale production of algae is that of illumination. Conversion of electricity to light has an efficiency of only 10 to 20 percent. In addition, the maximum efficiency of light utilization by Chlorella algae lies in the range of 18-22 percent. This results in a maximum efficiency of only 4 percent for photosynthetic systems. Another problem involved in conversion of electricity to light is the production of heat which has to be removed even with thermophilic algae. With a human demand of 600 liters of oxygen per day, the minimum electrical requirement becomes 4 kW. No large-scale culture has yet been managed at anything close to this minimum figure.
Another problem is the poor penetration of light into concentrated cultures of algae. This necessitates construction of large tanks of only about ¼-inch thickness. This results frequently in fouling of the surfaces of the tank by algae and makes the removal of the excess algae difficult. Production of 1 liter of oxygen results in the production of 1 gram dry weight of algae. Although a small amount of CO is produced by some algae, it can probably be removed by catalytic oxidation. Other problems include mutation and genetic drift of the algae and the necessity for maintaining bacteria-free cultures. There are also difficulties in maintaining a sterile culture if urine is to be used as a nitrogen source. While there is a potential for using algae as food, more research is required before it can be determined what quantity and methods of processing can be used. Research and development on algae is much greater than on both the higher plants and the electrolysis-Hydrogenomonas systems together.
The difference between the photosynthetic and electrolysis-chemosynthetic systems is the way electrical energy is made available to the organisms. In the photosynthetic system, electrical energy is converted to light which the algae or plants transform into chemical energy. In the chemosynthetic process, electrical energy is transformed into the chemical energy of hydrogen gas which is used by the bacteria. Both organisms use the chemical energy available to them to synthesize cell material with similar degrees of efficiency. The problem is to make the conversion of electricity to available chemical energy as efficient as possible.
In photosynthetic systems much energy is lost in the conversion of electricity to light, a process only 10-20 percent efficient at best. When this is combined with the loss from the inefficient use of light by plants, an overall efficiency of about 4 percent is obtained. In the electrolysis-Hydrogenomonas system, the two steps are very efficient. Electrolysis cells can operate at up to 85 percent efficiency and the overall efficiency can be up to seven times that of a photosynthetic system.
ELECTROLYSIS-HYDROGENOMONAS SYSTEM
Electrolysis is carried out in a closed unit containing an electrolyte (KOH solution) with an anode and a cathode. These cells produce a maximum yield (60-80 percent or more) in gas production per unit of power consumption. According to Dole and Tamplin ([ref.184]), a unit capable of producing enough oxygen to sustain one man would be highly reliable, weigh approximately 18 kg, and require a power input of 0.25 kW.
One approach to zero-gravity operation is to rotate the electrolysis cell as described by Clifford and McCallum ([ref.185]) and Clifford and Faust ([ref.186]). The smallest known electrolysis cell under development uses this artificial gravity to separate oxygen from the anode and electrolyte, while the dry hydrogen gas permeates through the foil cathode, fabricated from palladium-silver alloy. This electrolysis cell, which would provide breathing oxygen for three men, has a volume of 1.4 liters, weighs 4.5 kg, and requires 0.67 kW, excluding auxiliary equipment, and has an efficiency of 84 percent.
The chemosynthetic conversion is carried out by the hydrogen bacteria. By the oxidation of molecular hydrogen, supplied from the electrolysis of water, energy is made available for biosynthesis. The generation of this "biological energy" is mediated by the stable enzyme hydrogenase which is present in the bacteria. On the average, the oxidation of 4 moles of H2 is required for the conversion of 1 mole of CO2 (the hourly production of a man). The removal of this amount of CO2 would thus require the cleavage of 4 moles of water. In addition, to supply oxygen for human respiration (at a rate of 1 mole of O2 per hour) the cleavage of two additional moles of water is required. Therefore, the chemosynthetic regeneration and human respiration together would require, on the average, the splitting of 6 moles of water per hour.
The material balance for electrolysis, biosynthesis, and human metabolism, with gram molecular weights in parentheses, are shown in equations (1) to (3), respectively:
6H2O ———————> 3O2 +6H2(108) ———————> (96) + (12) (1)
The bacterial synthesis requires 6 moles of H2, 2 moles of O2, and 1 mole of CO2 (from the astronaut), as shown in equation 2:
6H2 + 2O2 + CO2 ———————> CH2O + 5H2O(12) + (64) + (44) ———————> (30) + (90) (2)
The respiration of the astronaut requires 1 "food" mole (CH2O) representing about 120 kcal, and 1 mole of O2, as shown in equation 3:
CH2O + O2 ———————> CO2 + H2O(30) + (32) ———————> (44) + (18) (3)
The metabolic data in table VIII show that the CO2 of the astronaut and the bacteria must balance at about 1.056 kg per day.
The water relations are not completely balanced, but are fairly close. About 2.6 liters per day of water are split by electrolysis. The astronaut has an intake of 3.5 liters of water per day, 2.5 liters for drinking and 1 liter for preparing dehydrated food. The output is about 1.6 liters of urine and 2.1 liters of water of respiration and perspiration per day, or a total output of 3.7 liters, with the 0.2-liter excess due mainly to water of metabolism. The bacteria-produced water, amounting to 2.2 liters per day, and the excess from the astronaut would supply 2.4 liters toward balancing the 2.6 liters of water electrolyzed.
Bacterial Culture
Hydrogen bacteria are characterized by their ability to metabolize and multiply in a strictly inorganic medium, when supplied with H2, CO2 and O2 in required amounts. They can be grown in batch culture or in continuous culture using different methods of supplying entire medium or components on a demand feed system.
A medium was developed for batch culture of Hydrogenomonas eutropha by Repaske ([ref.187]) with quantitation of a number of components including trace minerals. Experiments by Bongers ([ref.188]) showed that a simplified medium, using laboratory-grade chemicals, could be used. A definite requirement was found for magnesium and ferrous iron (Fe++). The optimal growth requirements observed for Hydrogenomonas eutropha are shown in table X.
Culture parameter |
Optimum value |
|---|---|
Cell density, g (dry weight)/liter |
10 |
Temperature, °C |
35 |
Pressure, atm |
1 |
pH (phosphate buffer) |
6.8 (6.4-8.0) |
H2, percent |
75 |
O2, percent |
15 |
CO2, percent |
10 |
Urea CO(NH2)2, g/liter |
1 |
MgSO4·7H2O, g/liter |
0.1 |
Fe(NH4)2(SO4)2, g/liter |
0.008 |
The effects of temperatures ranging from 20° to 42.5° C on the growth rates of Hydrogenomonas eutropha were studied by Bongers ([ref.189]), and the optimal temperature was found to be about 35° C. Experiments at 25° and 35° C indicated that the efficiency of energy conversion was essentially identical at both temperatures. Hydrogenomonas requires, as part of its substrate, a mixture of three gases: hydrogen, oxygen, and carbon dioxide. Experiments were performed by Bongers ([ref.189]) to determine the toleration limits of the three gases. Growth rates were found to be identical when hydrogen varied from 5 to 80 percent. Nearly identical growth was obtained when CO2 partial pressures were 5 to 60 percent, being slightly lower at higher partial pressures. The organism was highly sensitive to oxygen concentration. Dissolved oxygen concentrations above 0.13 mM were found to inhibit cell division; energy utilization was also affected by oxygen concentration. At 0.2 mM oxygen concentration, the efficiency of energy conversion was approximately half the value observed with 0.05 mM.
Another parameter of importance is the total volume of suspension which would be required to balance the metabolic needs of one man. The volume of suspension is determined by the conversion capacity of a unit volume. This capacity is a function of the cell concentration; hence, the more cells that can be packed in a unit volume of suspension (and adequately provided with H2, O2, and CO2), the less the volume of suspension required.
Results of experiments by Bongers (refs. [ref.190] and [ref.191]) on conversion capacity-density relationships show that the rate of CO2 conversion obtained with suspensions up to approximately 10 grams (dry weight) per liter is linear with relation to density. This indicates that the supply of H2, O2, and CO2 is adequate. Upon a further increase in cell concentration, the conversion rate still increases but not linearly. The highest amount of CO2 taken up per liter of suspension was approximately 2 liters per hour. At these very high cell concentrations, the relationship between rate of conversion and density is no longer linear. This is demonstrated when the conversion rate is calculated per unit cell weight instead of per unit suspension volume. The rate per gram dry weight per liter decreases from 146 to 68 ml of CO2 per hour. With a suspension at a density of approximately 10 grams, the conversion of 1.1 liters of CO2 per liter per hour is obtained. At a CO2 output of 22 liters per man per hour, 20 liters of suspension would be sufficient to balance the gas exchange needs of one man.
At higher cell concentrations, less volume of suspension would suffice if gas equilibration could be maintained at the higher consumption rates to avoid anaerobic conditions which could lead to a shift in metabolism. In the final analysis, the technical problem of gas transfer from the gas to the liquid phase determines the optimal cell concentration and, therefore, the required suspension volume.
From data presently available, it can be concluded that, using the slow-growing H. facilis, the volume of suspension required to support one man is about 500 liters. Using H. eutropha, Schlegel ([ref.192]) calculated a suspension volume of 66 liters with 1 gram dry weight of bacteria per liter.
In recent NASA-supported research, the amount of culture medium has been estimated using improved cultivation methods and conditions. For batch culture, the data show that from 10 to 66 liters would be required per man, with a best practical estimate of 20 liters at 9 to 10 grams dry weight of bacteria per liter ([ref.191]). For continuous culture using the turbidostat, the present data indicate a demand for some 30 liters of suspension, and a volume of 20 liters (at approximately 10 grams dry weight of bacteria per liter) as a realistic goal.
In the foregoing section, the material balance for gases and water was discussed. It was shown that a close match could be obtained with these components of the closed environment.
Less abundant, though no less important, are the nonwater components of urine and feces. The urine is important for the content of fixed nitrogen and other products of man's metabolism and serves as a very effective substrate for cultivation of hydrogen bacteria. Maximum closure of the system necessitates utilization of the urea in urine as a nitrogen source.
The average man produces 1.2 to 1.6 liters of urine per 24-hour period. This contains about 0.00005 gram per liter of iron, 0.113 gram per liter of magnesium, and 24.5 grams per liter of urea ([ref.193]). As shown in table X, each liter of bacterial medium requires 0.008 gram per liter of Fe(NH4)2(SO4)2, about 0.1 gram of MgSO4·7H2O, and 1.0 gram per liter of urea. In comparing the daily urine output with the estimated required ingredients of a bacterial medium, a relatively close balance is observed, with the exception of iron.
For the fixation of 24 moles of CO2 (288 grams of C) produced per man per day, the production of about 640 grams dry bacterial mass is required. At an average N-content of 12 percent, the nitrogen requirement would be some 100 grams. A comparison of daily output (urine) and daily requirement by the bacterial suspension reveals that only 10 to 33 percent of this amount could be recovered from average urine. To obtain a material balance, either the man must be fed a protein-rich diet or the bacterial suspension must be grown under conditions which lead to the production of a cell mass relatively low in protein content. Experiments have indicated that nitrogen starvation of the bacterial culture might be a promising solution. Culture "staging" (cultivation under nitrogen-rich conditions, followed by cultivation in the absence of substrate nitrogen and subsequent harvesting for food processing) will probably be the most promising means of nitrogen economy in the closed environment. As discussed in a following section, a biomass of relatively high lipid content can be obtained under conditions of nitrogen starvation.
Continuous Culture of Hydrogenomonas Bacteria
Growth of hydrogen bacteria in a batch culture, after an initial period of adjustment, becomes steady and rapid during the exponential growth phase. This steady state of growth is temporary and ceases when nutrient substrate or gas concentrations drop to limiting values. For long periods a continual supply of nutrients must be provided. Growth then occurs under steady-state conditions for prolonged periods, and such factors as pH, concentration of nutrient, oxygen, and metabolic products (which change during batch culture) are all maintained constant in continuous culture.
Two methods can be used for control of continuous cultures: the turbidostat and the chemostat. In the turbidostat, regulation of medium input and cell concentration is controlled by optically sensing the turbidity of the culture.
The dilution rate varies with the population density of the culture and maintains the density within a narrow range. Organisms grow at the maximum rate characteristic of the organism and the conditions. The growth rate can be changed by modifying the nutrient medium, gas concentration, or incubation temperature. A disadvantage of the turbidostat is that all nutrient concentrations in the culture chamber are necessarily higher than the minimum, resulting in inefficient utilization of nutrients.
The turbidostat system for continuous culture of Hydrogenomonas bacteria, developed by Battelle Memorial Institute ([ref.194]), includes electrolysis of water in a separate unit. Hydrogen and oxygen are fed separately up to the point of injection into the culture vessel, and the mixed volume is kept very small to minimize am possibility of explosion. However, the two gases may be injected simultaneously if there is a demand for both.
In the chemostat, growth of the organisms is limited by maintaining one essential nutrient concentration below optimum. A constant feed of medium, with one nutrient in limiting concentration and with constant removal of culture at the same rate, is used to achieve the steady state. The dilution rate is set at an arbitrary value, and the microbial population is allowed to find its own level. By appropriate setting of the dilution rate, the growth rate may be held at any desired value from slightly below the maximum possible to nearly zero. This constitutes a self-regulating system and allows selection of a desired growth rate.
A combined electrolysis-chemostat method, developed by Magna Corp., maintained the hydrogen-producing electrode of an electrolysis cell in the bacterial culture. Resting cells of Hydrogenomonas eutropha consumed hydrogen produced at the cathode of an electrolysis cell built into a specially constructed Warburg flask. Attempts to immobilize Hydrogenomonas cells on a porous conductor were partially successful. This system could lower the volume requirements compared with those for the isolated subsystems. Disadvantages of this integrated system include electrolysis of the bacterial medium, possibly resulting in toxic breakdown products, and the possible effects of electric power and the KOH electrolyte on the bacteria. The main disadvantage of an integrated system would be the disparity between optimal conditions for efficient electrolysis and efficient bacterial conversion, particularly temperature and pH, with the combination possibly resulting in considerably higher power and weight demands.
Both continuous-culture approaches are being studied with NASA support. The turbidostat offers the greatest potential efficiency in weight and volume, but uses nutrient materials less efficiently and is more complex. The chemostat is less efficient in weight and volume, but has greater simplicity and reliability.
Hydrogenomonas eutropha has been grown in 15-liter batch cultures and in 2.1-liter continuous cultures. A 20-liter continuous culture, sufficient to balance the requirements of a man, is under development.
The potential problem areas in large-scale continuous production of the bacteria include assuring genetic stability, preventing or controlling bacteriophage and foreign bacterial contamination, and preventing heterotrophic growth caused by exposure to organic material from the urine. Genetics of hydrogen bacteria and phage infection have been studied by DeCicco. Research on these problems indicates that they are not of major importance, but cause significant effects and must be eliminated or controlled.
Bacterial Composition and Nutrition
Hydrogenomonas bacteria can be used for at least part of the astronauts' diet. The washed bacteria have a mild taste and are being studied for their total energy content, protein and lipid digestibility, and vitamin content. Carbon and nitrogen balances, and respiratory quotient are to be determined in animals fed the bacteria as their sole food source. No toxic constituents have been discovered. Sonicated and cooked bacteria, when fed to white rats as 12 percent of the solids of a nutritionally balanced diet, were eaten readily and produced no ill effects. Net utilization of the protein appears to be somewhat lower than casein and about the same as legume proteins.
The composition of Hydrogenomonas eutropha is shown in table XI. The composition of the bacteria varies with the age and growth phase of the cells and with the medium and gas available. It is possible to modify the growth conditions to grow the type of bacteria desired for nutritive purposes.
Hydrogenomonas cells contain about 75 percent water. Of the dry weight, about 74 percent is protein, calculated as 6.25 times the nitrogen content. Table XI shows the amino acid composition to be comparable with other bacterial proteins, except for higher tryptophan and methionine values.
Constituent |
Percent by weight |
|
|---|---|---|
Moisture |
74.55 |
|
Fat |
.44 |
|
Ash |
1.73 |
|
Nitrogen |
3.02 |
(wet) |
11.87 |
(dry) |
|
Protein (N × 6.25) |
18.90 |
(wet) |
74.26 |
(dry) |
|
Amino acids (dry weight)8 |
||
Alanine |
4.47 |
|
Arginine |
3.41 |
|
Aspartic acid |
4.32 |
|
Cystine |
.08 |
|
Glutamic acid |
7.67 |
|
Glycine |
2.76 |
|
Histidine |
.95 |
|
Isoleucine |
2.17 |
|
Leucine |
4.04 |
|
Lysine |
2.65 |
|
Methionine |
1.14 |
|
Phenylalanine |
2.20 |
|
Proline |
2.06 |
|
Serine |
1.80 |
|
Threonine |
2.15 |
|
Tryptophan |
.78 |
|
Tyrosine |
1.79 |
|
Valine |
3.03 |
|
The lipid content of rapidly growing cells is normally quite low (0.45 to 2.3 percent crude ether extractable lipids). The most important lipid is poly-beta-hydroxybutyric acid, which is stored under the growing conditions of insufficient nitrogen or oxygen supply (refs. [ref.187] and [ref.191]). Under these conditions, this unusual polymer constitutes up to 80 percent of the dry weight. While the monomer itself, beta-hydroxybutyric acid, is rapidly and efficiently used in cell metabolism, the nutritive value of the polymer is yet to be determined. The fatty acids found include lauric, myristic, palmitic, palmitoleic, heptadecaenoic, C17 saturated(?), stearic, linoleic, and linolenic(?) ([ref.195]).
Application to Spacecraft System
A bioregenerative life-support system will be required in long manned space flight, especially with several astronauts such as would be required for a manned mission to Mars in the 1980 time period. While almost 15 years is a long leadtime, the biological research and engineering problems are formidable, and a system would have to be developed at least 5 years before the mission.
The power and weight requirements for both chemical and biological regenerative life-support systems were presented in table VIII. These should be considered tentative best estimates based on present data.
The use of bioregenerative systems in spacecraft systems has been studied by Bongers and Kok ([ref.175]) who put the electrolysis-Hydrogenomonas system in proper perspective with the following statement:
The bioregenerative systems are more or less in a transitory phase between research and development. The power data can be considered fairly accurate, at least within ±20 percent. The postulated weight data, however, represent approximations, particularly with respect to auxiliary equipment and construction materials. Also omitted are the weight penalties most probably involved in the processing of the solid output of the exchangers, elegantly defined as potential food. Further research is required in this area to evaluate the regenerative systems, especially the bacteria, with respect to this potential. Furthermore, as yet there is no experimental proof that the growth rates of the heavy bacterial suspensions can be realized in a large design, determined on a relatively small scale with fairly precise control of physiological conditions and gas exchange. This aspect may affect considerably the weight involved in a chemosynthetic balanced system. Nevertheless, at present, this approach still seems most promising.
CABIN ATMOSPHERES9
In the first U.S. manned space flight program, Project Mercury, and in the face of very severe weight limitations, a cabin atmosphere of pure oxygen at one-third atmospheric pressure was adopted. This choice probably represented the greatest simplification which could be achieved readily and, at the same time, provide protection against some of the risks of rapid decompression. Although breathing pure oxygen at higher pressures was known to be attended by some undesirable physiological effects, the short duration of the flights to be undertaken, and the low pressure employed, suggested that no harmful results would result in this case. That these expectations were generally borne out is now history. Preparations for space flights of longer duration—many weeks or months—present similar problems and require special attention to phenomena which may be either undetectable or of trivial significance on a time scale of a few days.
Physiological Criteria in the Choice of Cabin Atmosphere
If maintenance of normal respiratory function were the only consideration, a cabin atmosphere of about sea-level composition and pressure might be an ideal and straightforward choice for manned spacecraft. In fact, this atmosphere has been used in the manned space flights conducted by the U.S.S.R. No other atmosphere has been shown to be more satisfactory from the physiological point of view, and the tedious respiratory studies which should accompany the use of other atmospheres can be avoided. Nevertheless, the formidable problems of spacecraft design and the necessary precautions for safeguarding the crew from accident require that other atmospheric compositions and pressures be considered. For example, if a cabin at 1-atm pressure were decompressed to space suit pressure (0.3 atm), the occupants would develop decompression sickness; i.e., "bends."
Several engineering considerations argue for low cabin pressures and pure oxygen composition. Among these are structural design, weight of atmospheric gas storage and control equipment, and the difficulty of contriving pressure suits which allow operation at pressures near one atmosphere. Such departures from the normal human gaseous environment, however, require the demonstration of an acceptable level of safety and physiological performance.
The limits of the composition and pressure of acceptable cabin atmospheres are then set by—
A pure oxygen atmosphere at a pressure which will provide an alveolar oxygen partial pressure equal to that provided by air at sea level
A mixed gas (oxygen and inert gas) atmosphere having a pressure and composition that will allow decompression to the highest acceptable suit pressure without the risk of bends
A numerical value for the lower limit (1) is approximately 0.2 atm of pure oxygen. The upper limit (2) is determined by the operating pressure and composition of the space-suit atmosphere and may be of the order of 0.5 atm for a cabin atmosphere of 50 percent oxygen. It is necessary to determine the astronaut's ability to survive and perform his duties in any atmosphere selected.
Atelectasis and Pulmonary Edema
Localized or diffuse collapse of alveoli in the lungs may, if the condition persists, lead to arterial hypoxia which may be extremely undesirable under the stresses of space flight. The alveoli are probably unstable when pure oxygen is breathed; they tend to collapse if there is blockage of the airways, especially at low pressures. This collapse occurs because each of the gases present in the alveoli (oxygen, water vapor, and carbon dioxide) is subject to prompt and complete absorption from the alveoli by the blood.
The alveoli are normally stabilized against collapse by the presence of inert and relatively insoluble gas (nitrogen) and an internal coating of lipoprotein substances with low surface tension.
Theoretical and experimental results strongly suggest the desirability of using oxygen-inert gas atmospheres for long missions to avoid atelectasis and other gas absorption phenomena, such as retraction of the eardrum. However, further experimental evidence is required both to confirm this point and to establish its upper limit of suitability of pure oxygen atmospheres.
At Ohio State University in 1962, scientists studied the effect on young rats exposed for 27 days to 100 percent oxygen (with no nitrogen), at a reduced barometric pressure equivalent to 33 000 feet altitude. The rats showed no difference in growth rate, oxygen consumption, food and water intake, or behavior from control rats in air at 1 atm.
Oxygen Toxicity
It has long been known that breathing pure oxygen at normal atmospheric pressure often produces pulmonary irritation and other toxic effects both in man and animals. This knowledge has occasioned concern over the use of pure oxygen atmospheres in spacecraft.
The effect of 100 percent oxygen at a simulated altitude of 26 000 feet for 6 weeks was studied using white rats at Oklahoma City University under a NASA grant. Radioactive carbon techniques revealed a 15-percent reduction of metabolism in the 100-percent oxygen-exposed rats, compared with rats in air at 1 atmosphere. There was a 20-percent decrease in lipid metabolism in the liver compared with controls, but no decrease in heart metabolism. There was no gross change in body weight.
The White Leghorn chick between 2 and 7 weeks old is markedly resistant to the toxic effects of 1 atm of O2. Continuous exposure (Ohio State University) for as long as 4 weeks did not cause deaths, obvious morbidity, or any signs of pulmonary damage on gross autopsy. Nevertheless, the hyperoxia had some adverse effects, primarily reducing the growth rate to between three-fourths to one-fourth of normal; reducing feed intake per unit body weight to three-fourths of normal; slowing respiratory rate by 30 percent; decreasing erythrocytes, hemoglobin, and hematocrit by 9 to 12 percent; and causing reversible histological changes in the lungs. Arterial O2 tensions were elevated over 300-mm Hg, but arterial pCO2 and blood pH were unaffected. No residual effects were noted upon return to air breathing. It is possible that the anatomical peculiarities of the avian lung play some role in the chicks' resistance to hyperoxia, but it is also possible that this resistance is a function of age, similar to the tolerance shown by the young rat but not the adult.
Carbon Dioxide Tolerance
Studies of CO2 tolerance in submarine crews indicate that no loss of performance is involved if the concentration in air at normal pressure does not exceed 1.5 percent with exposures of 30 to 40 days. However, biochemical adaptive changes were observed at this concentration.
Inert-Gas Components
If other investigations establish the need for an inert gas in manned spacecraft atmospheres, gases other than nitrogen may be considered. Compared with nitrogen, the physical properties or helium and neon offer advantages with respect to solubility in body fluids, storage weight, and thermal properties.
Studies at Ohio State University in 1964, under a NASA grant, showed that helium substituted for nitrogen in a closed container causes humans to feel "cold" at a normally comfortable temperature. Studies with animals have shown that in a helium atmosphere there is greater heat loss due to the increased conducting capacity and probably greater evaporative capacity. In 6 days at 21 percent oxygen and 79 percent helium at 1-atmosphere pressure, young rats grew at the same rate as controls, but drank more water, excreted more urine, and had a higher rate of food and oxygen consumption than controls in air at 1 atmosphere. Men are being tested on a bicycle ergometer in saturated and low relative humidity helium atmospheres to study heat balance.
Mice were exposed to 80 percent argon and 20 percent oxygen continuously at 1-atmosphere pressure for 35 days at Oklahoma City University. Carbon 14 studies of metabolism showed a slight slowing and a twofold to threefold increase in fat deposition.
Bends
Decompression, whether accidental (due to damage of the spacecraft) or intentional (as in the use of the pressure suit outside the capsule), carries the risk of bends if the inert gases dissolved in the tissues and body fluids come out of solution. The magnitude of this risk is determined to a very considerable extent by—
Individual susceptibility
The extent to which the nitrogen (or other inert gas) concentrations of tissues and body fluids have been reduced
The magnitude and rate of the inert-gas, partial pressure change on decompression
The probability of getting bends is reduced by—
Selection of bends-resistant individuals
Thorough denitrogenation before flight
Limitation of decompressive pressure changes by appropriate choice of cabin atmosphere pressure and composition
Space-suit pressure setting
In some cases, further improvements might be obtained by using, in the cabin atmosphere, an inert-gas component which has a lower solubility in tissue and body fluids or less tendency than nitrogen to form bubbles.
Fire Hazard
Experience indicates that fires in pure oxygen atmospheres, even at low pressures (e.g., 1/3 atm), are extremely difficult to extinguish. While this phenomenon has nothing to do with respiratory physiology, the risk on flights of long duration may be so serious as to demand special measures. Unless effective countermeasures can be devised, this risk may argue very strongly against the use of such atmospheres in the future. Further experimental investigation is required.
Acceleration Effects on the Lungs and Pulmonary Circulation
Forces produced by high acceleration overdistend one part and compress another part of the lungs. Blood flow diminishes in some parts of the lungs and increases in others. Fluid leaks from the blood into the tissues and into the air sacs in parts of the lungs. These effects cause difficulty in breathing, low arterial oxygen saturation, and impaired consciousness during high sustained acceleration and, to a lesser extent, after its cessation. They must be considered when selecting the best gas to be breathed, since a high partial pressure of oxygen is favorable for consciousness, but a low inert-gas concentration during acceleration is unfavorable for rapid lung recovery afterward.
PHYSIOLOGICAL PROBLEMS
A study of the manned space flights and laboratory observations to date suggests that during long periods of weightlessness, some physiological difficulties may arise which may produce serious effects on human performance. Although recent experience gives no grounds for expecting insuperable difficulties, neither the quantity nor quality of the available observations permits the conclusion that long-term exposure to weightlessness will not have serious consequences. The critical role to be played by the astronaut demands that every effort be made to identify in advance those phenomena which may affect performance, and to study their qualitative and quantitative relationships so that proper precautions can be taken.
Lawton ([ref.197]), in reviewing the literature on prolonged weightlessness, found few instances in which physiological function was truly gravity dependent. He stated that the physiological systems likely to be most affected by weightlessness were the musculoskeletal system, the cardiovascular system, and the equilibrium senses. Subsequent experience proved this to be the case. McCally and Lawton ([ref.198]) analyzed the data from experiments since 1961 and concluded that much more basic laboratory work is necessary. Studies using immobilization, immersion, and cabin-confinement techniques were recommended approaches toward simulating weightlessness.
Much of the difficulty in obtaining precise information of anticipated problems arises from a lack of knowledge of normal mammalian physiology. Many of these deficiencies can be remedied in the laboratory. In space-flight development, however, two distinct investigational approaches can be adopted. The first of these may be characterized as empirical and incremental; that is, the capabilities of the astronaut are explored in successive flights involving relatively modest increases in difficulty or severity of the environmental conditions. In this way it is hoped to ascertain the human limitations without running too great a risk. The second approach can be described as fundamental: determining by a series of controlled experiments the effects of exposure to space-flight conditions upon comparative mammalian physiology, with emphasis on man. A fundamental understanding of the observed effects would be sought so that predictions for new situations and possible ways to control them could be made with confidence.
It is not possible now to predict for flights of 30 days or more—
The effects of sudden reimposition of reentry accelerations and terrestrial gravity
Changes in body fluid distribution and composition
The effects of violent physical effort on respiratory and cardiovascular systems in prolonged weightlessness
Central nervous system functions, especially coordination, skilled motor performance, judgment, and sleep-wakefulness cycles
NASA has emphasized that planning for manned space programs involves a systematic extension from physiological observations in animals to man, and finally the establishment of man as part of the man-vehicle system design. Moreover, these studies require the evaluation of central nervous, cardiovascular, respiratory, gastrointestinal, and other systems as a matrix in mutual interdependence. There is particular interest in the effects of weightlessness on flights exceeding 30 days.
Mammalian flights of about 30 days also merit attention, including the development of the life-support systems which must precede such a program. Development of facilities for biological experiments may well be an important requirement for studies in anticipation of manned flights of longer duration than Apollo. Unless the biological satellite programs of the type mentioned above are successful in providing the necessary data, a manned orbiting laboratory may also be important in studies of shorter range.
General Studies of Biological Rhythmicity
The effects of weightlessness on the organism as a whole may be manifested by important changes in certain integrated behavioral patterns having an inherently rhythmic character. Modifications in basic behavioral patterns and performance may occur as disruptions of rhythmic physiological phenomena, which are themselves the end product of interrelated functional activity in a number of physiological systems, such as the neuroendocrine, cardiovascular, and central nervous systems.
Measurements of interdependent components of biological rhythmicity are beginning to be analyzed by methods well established in physics—including correlation and spectral analyses, and phase modulation and variance in rhythmic processes. A wide variety of physiological functions can be treated as periodic variables in the analysis, including rhythmicities in cardiac output and blood pressure, respiration, brain waves, and the slower tides of appetite, and sleep-wakefulness. The importance of such investigations argues for their inclusion in forthcoming flight programs. Their experimental simplicity is an additional advantage. Biorhythms have been discussed in more detail in the section on "Environmental Biology."
Effects of Weightlessness on the Cardiovascular System
Earlobe oximetry, indirect measurements of blood flow and of blood pressure by finger plethysmography or impedance plethysmography, and ballistocardiographic techniques have potential application to manned space flight.
Adaptation to prolonged exposure to weightlessness or to lunar gravity may cause difficulties when the astronaut is exposed again to reentry forces and terrestrial gravity. It is possible that these adaptive changes may thus produce unacceptable effects on performance or cause risk to life. It is important to obtain experimental evidence on this subject.
It is common knowledge that following a stay in bed, dizziness, faintness, and weakness characterize arising, and that a feeling of general weakness may persist for several days. The phenomenon has been investigated in a number of laboratories. One approach has been to put healthy young subjects to bed, and even in extensive casts for periods of 2 or 3 weeks or more. Two major findings have emerged from these studies. First, a substantial adjustment in the blood circulatory system occurs, which is termed the "hypodynamic state." Second, there is a large decrease in the skeletal and muscle mass of the body.
There are two kinds of evidence for the hypodynamic state: measurement of parameters of circulatory function, and measurement of the response of the individuals to a quantitatively imposed mild gravitational load. After 3 weeks in bed, otherwise healthy persons exhibit an increase of more than 20 percent in heart rate; a reduction of 10 to 20 percent in total blood volume, primarily as a result of reduction of plasma volume; and a decrease in heart size of about 8 percent. Coupled with these cardiovascular changes is a reduction of 10 percent in the basal metabolic rate. It appears as though the circulation and metabolism are reset to a lower functional level commensurate with the reduced demands placed on the whole organism.
After 3 weeks of bed rest, all of the subjects tested showed pronounced orthostatic hypotension. After tilting, the average heart rate increased by 37 beats per minute, the systolic blood pressure fell some 12-mm Hg, and some of the subjects fainted. The measurements were continued for 16 days after the bed-rest period, and it was round that recovery was not quite complete when the experiment was terminated.
There is little question that in prolonged exposures to the weightless state, there is a fair probability of extensive circulatory adjustments, the seriousness of which cannot yet be foretold. While it is likely that the astronauts will adapt successfully to long periods of weightlessness at some new circulatory functional level, the remote possibility exists that the circulatory changes may be progressive to the point of ultimate failure.
Metabolic Effects of Weightlessness
Without metabolic information, accurate planning of environmental systems for long flights is difficult. Importance is also attached to early evaluation of weightlessness effects on body-fluid equilibria. The results of Earth orbital flights and of terrestrial water-immersion experiments suggest the occurrence of undesirable changes, although no effects leading to operational incapacity have yet arisen.
In both recumbency and immersion, a similar redistribution of body fluids occurs. It has been suggested that recumbency may affect an extracellular fluid-volume receptor mechanism which by decreasing aldosterone secretion by the adrenal gland, would decrease sodium reabsorption by the renal tubules. Aldosterone excretion decreases during recumbency and during standing in water, but increases while standing in air. There is also evidence for cardiac atrial volume receptor mechanisms which respond to increased filling of the left atrium with reflex inhibition of release of pituitary antidiuretic hormone (ADH), resulting in diuresis (Henry-Gauer reflex).
Altered fluid equilibrium in buoyant states is accompanied by shifts in intracellular and extracellular electrolyte distribution, especially sodium and potassium. Evidence from recumbency studies indicates a strong correlation between loss of erect posture or weight bearing and excretion of calcium stores in bone.
A bone X-ray densitometry method has been developed by Mack, at Texas Woman's University, for accurately determining the loss of bone mineral (±2 percent accuracy) in humans and animals. The heel bone and spine are X-rayed using a calibrated aluminum wedge as a standard. This technique will be used for preflight and postflight analysis of the primate being flown in the 30-day biosatellite. Comparative appraisal of bone mineral behavior in astronauts participating in the Gemini and Apollo programs will be invaluable for future flight missions.
Bed rest and immobilization studies by Mack have shown loss of skeletal mineral and increased calcium in the urine and excreta. Four bed-rest studies, each extending for 2 weeks, compared different levels of calcium intake. Four men were used in each study and served as their own controls during extended ambulatory periods. During 2-week periods, up to 10 percent of calcium mineral was lost from the heel bone. Calcium was also determined in the urine and feces. In other studies, isometric exercises reduced loss of bone mineral during bed rest.
Excretion of calcium in the urine is accompanied by risk of its deposition as calculi or "kidney stones" in the urinary tract. Currently, changes in calcium metabolism resulting from weightlessness over periods up to 2 weeks is not considered a hazard requiring precautionary measures.
Flights in excess of 2 weeks, however, constitute a problem serious enough to warrant study on the 11-day orbital flights and the 30-day biosatellite primate mission. Therapeutic immobilization, post-poliomyelitis immobility, and experimental restraint in normal subjects lead to a negative calcium balance, with hypercalciuria.
Central Nervous System Functions in Weightlessness
The wide range of individual tolerances to the disturbing effects of vestibular stimulation has emphasized the importance of this factor in astronaut selection. At the same time, vestibular functions must be considered jointly with visual task performance, since both have special significance for such maneuvers as vehicle docking. Vestibular function in the weightless state remains almost completely unknown. Limited evidence from animal and manned space flights suggests that head turning, resulting from vestibular stimulation, may seriously interfere with visuomotor performance, but that susceptibility to these disturbances is significantly different between individuals and that partial adaptation occurs relatively quickly.
NASA is currently collecting extensive baseline electroencephalogram data under controlled conditions in a form suitable for mathematical analysis. Data are being taken from about 200 subjects in major national and overseas centers. It is intended that this study will assist in astronaut selection and monitoring in space.
Studies on many effects of weightlessness on nervous functions require monitoring of the autonomic nervous system, including such autonomic effects as gastrointestinal activity, secretion, lacrimation, salivation, sweating, and the central control of respiration. Urinary estimations of catecholamines and 5-hydroxyindoleacetic acid would provide important data on autonomic system activity if collected in flight and compared with preflight and postflight controls.
Major areas have been outlined in which prolonged weightlessness may be expected to interfere with performance, judgment, and, ultimately, chances of survival. These include cardiovascular, metabolic, central nervous, psychophysiological, and biorhythmic effects. They have been dealt with separately and in sequence, but have not been intended to be viewed as hierarchic. The relative scarcity of data necessarily precludes such an evaluation.
Soviet experience with zero gravity and weightlessness has increased their emphasis on this space-flight factor and was an important topic at the May 1964 COSPAR meeting. Discussion of the postflight medical status of Bykovsky (5-day flight) and Tereshkova (3-day flight) revealed a concern for the significance of prolonged weightlessness and the presence of postflight physical debility and fatigue following Vostok flights 3 through 6. These changes persisted for several days. Among the physiological conditions singled out for mention were—
Body fluids— Cosmonauts have shown a postflight weight loss of 1.9 to 2.4 kg apparently resulting from a redistribution of body fluid in response to elimination of the hydrostatic pressure gradients caused by Earth gravity. There is the suggestion that this redistribution is complete within the first 24 hours of flight. Titov is reported to have been dehydrated alter his flight with early hemoconcentration. These findings directly support predictions made from ground-based research.
Cardiovascular— Postflight orthostatic tachycardia is reported for Titov as long as 23 hours after landing; at 48 hours there was significant residual intolerance to the upright posture. Cosmonauts have demonstrated a 20- to 35-percent increase in oxygen consumption during the standard postflight exercise test.
In both of these areas there was a return to normal within the postflight period of study. The Soviets have continued their biological experiments in space with the Vostok/Voshkod series. Fixing of histologic specimens in flight by Bykovsky demonstrated a critical role for man and made possible an expanded experimental program. Biopackages have become more complex with each succeeding flight.
With the exception of postflight orthostatic intolerance after the third and fourth Mercury flights, changes as a result of exposure to a zero-gravity environment have not been noted by U.S. investigations in space. Ground-based research proceeds here at an advanced pace and is supported in large measure by both the USAF and NASA. A study of the relationships among renal and systemic hemodynamics, neurohumoral cardiovascular regulation, and renal excretory function in differently positioned subjects is underway, as are studies of acceleration tolerance.
DEPRESSED METABOLISM
In anticipation of prolonged manned space flights, NASA has sponsored research related to metabolism depression. The daily food requirements, for example, of astronauts during a voyage of several months can constitute a major portion of the weight and storage capacity of the spacecraft. A somewhat promising and fundamental approach to this problem is the reduction of the astronauts' daily metabolic requirements. It has been suggested that astronauts on prolonged space missions be put in a state of suspended animation until their destination is reached. Though this sounds fantastic, 10 years ago no cell had been frozen to cryogenic temperatures and survived. Today it is commonplace for tissues to be frozen, stored at low temperatures, and thawed and then to maintain their viability and function.
Animal metabolism may be depressed by reducing body temperature, as in hibernation and hypothermia. Other means by which metabolism can be lowered include drugs and electronarcosis. Hibernation is a nonstressful state and results in a great decrease in metabolism. However, human beings are not hibernators, and much research is needed before the mechanism of hibernation is understood, and the possibility of inducing it in humans evaluated. Hypothermia is the direct cooling of the body to temperatures where metabolism is substantially depressed. Extracorporeal circulation systems combined with cooling are in routine use in most medical centers throughout the world. Hypothermia is not an ideal solution, however, since general body hypothermia is a stressful condition. Pharmacologic induction of hypothermia can be accomplished by such drugs as chlorpromazine and harbamil. Other drugs can be used to depress metabolism, but all have some disadvantage.
In recent years there has been a growing interest in electronarcosis, the induction of sleep by an electric current. Although potentially valuable, this method is far from routine application.
Outstanding advances have been made in metabolism suppression. Recent progress in the biochemistry and physiology of hibernation and hypothermia have shown that the oxygen requirements of individual mammals, organs, and tissues can be reduced. When the chemical composition of the blood and the cardiac output are sufficient to meet cellular requirements, regulatory mechanisms remain effective and animal survival is assured. In contrast, when oxygen transport is interrupted, a reduction in cellular activity occurs and regulation is impaired. In induced hypothermia, the low temperature slows the rates of all processes and modifies the action of metabolites and other substances. This in itself is not harmful, as shown by the true hibernating animal (e.g., ground squirrel), but will become disastrous as soon as anoxia and chemical imbalance begin to develop.
The phenomenon of natural hibernation is being investigated in the laboratory in the hope that the unusual tolerance of hibernating animals to reduced metabolism and low body temperature may some day be produced artificially in ordinary laboratory animals and man. Experiments with the ground squirrel, a typical hibernator, show that the artificially cooled ground squirrel does not tolerate such long periods of low body temperature as does a naturally hibernating animal.
Other studies of the brown adipose tissue (fat), which is present in most hibernating mammals, show it to be essential to hibernation. Indications that brown fat has a thermogenic role in rats exposed to low temperatures suggest that this may be the case in true hibernators ([ref.199]). Arousal of the hibernating animal by cold is triggered by sympathetically activated thermogenesis in areas of brown fat so located, relative to the vasculature, that the heat is transferred to areas of the body concerned with normal metabolic and nervous activity.
Soviet work comparing various depressed metabolic states and resistances to acceleration shows deep winter hibernation to be most effective, followed by deep hypothermia, and drug narcosis as the least effective.
Experimental evidence is being accumulated to show that hibernation and hypothermia somewhat protect animals against radiation. Clinical studies on irradiation of cancer patients indicate that lowering the body temperature reduces cellular metabolism and thus decreases tissue sensitivity to gamma radiation ([ref.200]).
The use of prolonged hypothermia, hibernation, drugs, and electronarcosis appears to hold some potential for reducing astronauts' metabolic requirements. If one or mote of these methods become practical, human requirements for food and oxygen could be drastically reduced. Simultaneously, these methods may afford radiation protection and acceleration tolerance.
NUTRITION IN SPACE10
The human body can use food stores so that the nutritional requirements can be reduced for a short time. This will vary widely among individuals and each individual may exhibit characteristic patterns of nutritional behavior. During reduced food intake, muscular efficiency may not change significantly over a period of 4 to 6 days; unfortunately, however, mental activity begins to decline after 24 hours. Feeding requirements can be divided into two categories: short term (for missions of less than 21 days) and long term. Since dehydration can occur in a matter of hours under adverse conditions, water requirements must be considered as a special case.
Water Requirements
Water requirements are extremely critical and the amount supplied should not under any circumstances be kept to a minimum. Rather, a large margin of safety should be allowed.
Present data on water requirements show a very strong dependence upon suit inlet temperatures. In the absence of an accurately controlled suit temperature, water requirements can easily double. If this should occur, the mission would probably have to be aborted, since it is doubtful if electrolyte balance would be maintained at such high rates of water loss. Normal or even extreme conditions of the terrestrial environment usually include diurnal variation in temperature which may modify water needs. These conditions will not be obtained in the spacecraft.
In addition to ground-based experiments, measurements of water intake should be made under actual flight conditions. Data from short-term flights should be used for extrapolation to longer missions.
Formula Diets
The tacit assumption which now prevails, "Astronauts even on short-term missions require a diet of great variety," is apparently not well supported. In many parts of the world, people live on a monotonous diet consisting of only a few types of food with no apparent ill effects, provided their nutritional requirements are satisfied. Experimental evidence from many sources (e.g., the Army Medical Research and Nutrition Laboratory) shows that individuals can be kept on a single disagreeable formula diet for as long as 60 to 90 days without harm. Since highly motivated individuals are chosen for space flights, it is unlikely that they would object to the monotony of a formula diet and would probably prefer its simplicity. Also, there are definite possibilities of developing a much more acceptable formula than present types. There is no reason to anticipate adverse effects from the use of formula diets in short-term flights.
Formula diets would be extremely desirable for short-term flights. A formula diet (a rehydrated liquid formula could be used) would considerably reduce the number of manipulations and the time required for in-flight preparation, compared to a varied diet. These two improvements could contribute materially to the safety of a flight, since the astronauts would not be preoccupied with food preparation for so long a period, and the food could be dispensed without removing suit components, such as gloves. Storage requirements could be simplified with this type of diet. Weight, however, would not be lowered without the development of more refined formulas than those now available. Formula diets could readily be adapted to the determined metabolic requirements of the individual astronaut. Packaging problems will be simplified by using formula diets, which can easily be given a variety of flavors and colors.
Waste
The problem of waste production is intimately related to nutrition and can be solved or simplified by dietary changes. Any diet should be adjusted for the minimum production of feces, before and during even short flights. Water will be sequestered by accumulation in the feces, and the net loss, under normal conditions, would be approximately 40 to 60 grams per man per day. Flatus can be a serious problem, since considerable concentrations of toxic gases may accumulate. The purification system for the recirculated atmosphere must be able to remove these, although the diet should be planned to minimize the problem. The collection of urine and its storage is of importance, particularly on short-term flights, and individual packaging and labeling of urine specimens will be necessary for the analyses.
Metabolism
An accurately measured intake of nutrients, calories, and water is necessary for determining metabolic demands imposed in any space flight. There is insufficient knowledge to predict total metabolic requirements under the numerous stresses which can be anticipated. Simulator studies are of great importance even for short-duration flights.
The two most important variables to be considered in establishing the minimal diet are protein and energy requirements. NASA is supporting research at the University of California (Berkeley) to determine these requirements and to estimate individual variation in healthy young men. The possibility of minimizing need through biological adaptation is being explored.
It is difficult to estimate the minimum protein requirement of an adult man. The generally accepted criterion of minimum adequate protein nutrition in the adult is the maintenance of nitrogen balance at minimum intake. The minimum protein requirements depend on endogenous nitrogen loss. Analysis of the little data available indicates a best estimate of 2 mg of nitrogen per kilocalorie of basal energy expenditure. However, this figure is higher than that noted in experiments in some human subjects.
After minimum nitrogen requirements and minimum amino acid requirements have been established, studies will be directed toward investigating caloric restriction and adaptation to restriction of calories. It has been suggested that caloric restriction in animals and man results in apparent decreased energy need for the same activity. This apparent paradox has never been explained. It has been shown that there is adaptation to repeated episodes of caloric restriction both in animals and man, so that subsequent periods of caloric restriction result in decreased rate of weight loss, nitrogen loss, and longer survival.
Additional experiments are urgently required to determine the metabolic demands for minerals—in particular, the metabolic balance of calcium, potassium, sodium, and phosphorus. Under conditions of high water consumption, large mineral losses are to be expected. Failure to replace these can cause an imbalance which could impair the efficiency of the individual to the extent of endangering the flight.
Analysis of samples taken in flight, both of urine and feces, should be made. Respiratory quotients can be determined in flight, blood samples should be taken before and immediately after flight for analyzing selected components (in simulator studies these could be taken periodically), and nutritional intakes (which would be facilitated by formula diets) must be measured and analyzed.
Short-Range Technology
There are many practical difficulties in providing for food storage and accessibility in spacecraft. The packaging of food materials, both dehydrated and liquid, has proceeded satisfactorily under the supervision of the Food and Container Institute. If packaging materials are to be made to withstand very high relative humidities and large variations in temperature, additional investigations are required, since such containers are not yet available. In packaging, serious consideration must be given to the ease with which the food may be reached and eaten.
If dehydrated formula foods are to be fed on short-term missions, additional work is required on the rehydration of such formulas. Present methods of water measurement under weightless conditions are not satisfactory, and better methods will have to be contrived.
Long-Term Nutritional Problems
There is a dearth of metabolic information, even for short-duration flights, without which changes in metabolic patterns to longer flights cannot be extrapolated. However, using scattered information, certain changes which may be encountered can be hypothesized. Decalcification of bone and changes in water-holding capacity of the body may be anticipated. It is also possible that changes in proportion of fat to lean body mass could be experienced and should be considered in nutritional planning. Nutritional requirements depend on size, particularly lean body mass, sex, physiological state, and individual metabolic rates. Therefore, individuals for space flight should be screened with these factors in mind if it is desirable to minimize food intake in long flights. The factors which influence the total nutritional requirements of the individual also influence his mental and physical responses to stress.
Synthetic Foods
The development of food materials other than those derived directly from animal or vegetable origin is of interest. Advantages of such diets may be low residue, ease of storage, rehydration, and manipulation. Experiments with chemically defined synthetic diet for humans have been carried out by Medical Sciences Research Foundation, San Mateo, Calif. The complete liquid diet is composed of required amino acids, fat, carbohydrate, vitamins, and minerals. A cubic foot of the diet (50 percent solids in H2O) supplies 2500 calories per day for 1 month, and has been given a variety of artificial flavors.
This synthetic diet has been fed to human volunteers for 6 months in a pilot study at the California Medical Facility, Vacaville, Calif., and the results are being reviewed. Schwarz Bioresearch, Inc., is studying the storage, stability, and packaging of chemically defined synthetic diets for human and animal flights.
Food Production in Space
Long-term feeding in space depends upon a payload of stored food unless food is produced during flight. If sufficient propulsive energy is available, the duration of missions using stored food may be quite long. However, in emergencies in which a mission lasts longer than planned, survival may depend on the ability to produce food extraterrestrially. Eventually it will be desirable or necessary to produce food beyond the confines of Earth.
The nutritional requirements of the crew will be influenced by such factors as activity, physical and psychological stress, individual size of the members, and individual metabolic rates. The food intake will have to be adjusted to meet these requirements. It is necessary to know the nutritional requirements of each astronaut and the way in which these are altered by the conditions of space flight in order to estimate needs on long missions. Without this information, the food supplies for the longer flights may be too much, too little, or improperly balanced. Where dependence would not be on stored food alone, but on food produced en route, more exact information on requirements is needed to determine the capacity of food production units.
In the discussion of bioregenerative systems, it was suggested that food materials could be produced by photosynthetic organisms (e.g., algae, duckweed, and other higher plants) or by nonphotosynthetic organisms (e.g., Hydrogenomonas). In contrast to the use of living organisms, reprocessing waste materials by chemical treatment or the actual synthesis of high-energy compounds has been suggested. No chemical system has yet been demonstrated as workable for the economical production of food in space, and the systems considered produce materials which may be converted to food, but are not food as such.
Algal cultures have had the most extensive investigation as food in space, but the technical problems of using this material as a food source have not yet been solved. It is apparent from the investigations to date that algae will require treatment before they can be used as food. In limited trials, difficulties have been experienced with amino acid deficiencies, digestibility, high residues, and gastric distress. Processing methods which would be applicable in space travel and the possibility of secondary conversion by other animals or plants should be systematically investigated.
chapter 8
SIGNIFICANCE TO SCIENCE
One of the most critical research areas of the space program is bioscience. Of both practical and philosophical significance in exploring the origins of life and the possibilities of life on other planets, bioscience also promises much in medical aspects. Space offers biologists completely new environmental factors, such as the effects of zero gravity and of removal from Earth's rotation. These effects have been studied in attempts to advance understanding of basic mechanisms of physiology and biological rhythms. These studies can be of great value in dealing with problems of disease and metabolic disorders.
Biological research is fundamental to the problem of successfully protecting and sustaining man in the peculiar and hostile space environment. Understanding human requirements and variations in their response to various environmental factors offers value in medical research for human survival and comfort. The many technological discoveries and advancements in electronic and engineering equipment greatly enhance medical diagnosis, treatment of disease, and the extension of human life.
The life sciences, biology and medicine, are fundamental to the success of manned exploration of space, which marks a unique and significant development in the long history of man's conquest of new frontiers. Those who pioneered other frontiers on land and sea and in the air were not forced to await biological and medical research. Even the pioneers of aerial flight began their efforts without first seeking biomedical data. The search for such data followed flight experience and, indeed, was made only after problems arose.
Project Mercury, NASA's first program for manned space flight, stimulated immediate and extensive studies in the life sciences to sustain man in space. Before a vehicle could be designed to carry an astronaut into space, anticipated biomedical problems associated with space flight were studied. Life-support systems were designed to offer adequate protection from environmental stresses peculiar to space, such as zero gravity, removal from Earth's rotation, and high-energy cosmic radiation. These life-support systems used knowledge already gained from research for manned space flight by the U.S. Air Force.
Our entry into space has put us at the threshold of fundamental and far-reaching discoveries in the biological realm which have profound implications for other areas of human thought and endeavor. As man goes farther into space, the hazards increase; but past accomplishments indicate that the road ahead holds more promise than peril and that the vistas of knowledge that may be foreseen are as vast as space itself.
Almost everything which now can be said about the effects of extraterrestrial environments and about life on the Moon or the planets lies in the realm of pure speculation. There is one prediction, however, that can be made with considerable certainty by reason of historical precedent—the opportunity to investigate a totally new area, such as is offered by space exploration, is certain to produce a burst of scientific interest as soon as the path is charted by a few pioneers. Over the next few decades a progressively larger proportion of biological interest will turn to space. We may well expect that the discoveries made here will revolutionize some of our concepts of biology.
It should be fully realized that the accumulation and dissemination of biological and other scientific information is not only of great value to science and humanity but is of tremendous import to the prestige of the Nation.
SIGNIFICANCE FOR PRACTICAL APPLICATIONS
It can be predicted as confidently for space biology as for other space sciences that the economic costs will be amply repaid in the long run by applications of space-oriented biotechnology to other fields of biology and medicine. There are inevitable substantial, though indirect, contributions of NASA's continuing efforts in space biology.
NASA-supported biological research has many practical applications and "spinoffs" which contribute to the fields of health and medicine, food and agriculture, and industry and manufacturing. Some of these are presented to show the range and value of applications which have resulted from basic and applied biological research. In addition to those listed are many others from the biosatellite program, particularly in the fields of bioengineering and miniaturization.
Health and Medicine
Solar cells, which have powered space systems, are now being used as a power source in studies on brain function. A miniaturized solar cell developed by General Electric provides enough power, under ordinary house lights, to stimulate an animal's brain and to telemeter respiratory, cardiovascular, and brain-wave data while the animal is allowed to move about freely. Such a system is now used by the National Institute of Mental Health Laboratory at Rethesda, Md.
Scientists at the Ames Research Center have devised a new technique for studying organic compounds, whether synthesized in the laboratory or produced by a living system. This technique is based on a property of matter called optical activity. Previous methods of measuring optical activity have been plagued by low sensitivity. The new method is many tunes more sensitive and represents a real contribution to modern analytical instrumentation.
Studies on calcium metabolism and bed rest simulating weightlessness are adding knowledge on the prevention of demineralization of the skeleton; treatment of Paget's disease and osteoporosis prevention of muscular atrophy; the cause and treatment of renal calculi (kidney stones); optimal calcium for the human diet; and the factors influencing calcium absorption, metabolism, and excretion. The results will have great importance in bone healing and repair, care and treatment of fracture cases, treatment of paraplegics, and treatment of polio patients and similar cases. These grant studies at Texas Woman's University have also proven that the X-ray bone densitometry method can accurately detect changes in the skeleton.
A primary objective of the planetary exploration program is the detection of possible extraterrestrial life. The study of the fundamental properties of living things on Earth is restricted to the type of life which has evolved and survived here. Life which has been exposed to totally different environmental conditions may have markedly different physiological characteristics. The impact of the new information obtainable from the study of extraterrestrial life upon the sciences of medicine and biology will unquestionably be of fundamental and far-reaching importance. Advancement in the treatment of disease and the problems of aging are among the many possible consequences.
New developments in such techniques as ultraviolet spectrophotometry, polarimetry, and gas chromatography will find use in the detection of biochemicals and other compounds in hospitals and in toxicology and pathology laboratories. They will also be useful in studies of atmospheric pollutants such as smog.
Studies of the chemistry of living systems, molecular biology, and biophysics of cellular processes will create a better understanding of the basic mechanisms of life, leading to an understanding of both inherited and acquired disease, especially neoplastic conditions and chemical disturbances incident to mental disease.
The University of Pittsburgh is conducting a study to increase the availability of cytological technique in research and as a monitoring procedure by developing an automatic electronic scanning device using computer analysis for recording, counting, and sorting chromosomes. Structural changes in blood cell chromosomes can indicate the degree of radiation damage as well as damage resulting from various environmental stresses. Accordingly, this instrument, when developed, can be used as a radiation dosimeter in civil defense by swiftly detecting the degree and type of chromosomal aberrations in blood cells. Thus, casualties in nuclear attack could be quickly detected and treated. This system would also be useful for nuclear industrial plants and for military maneuvers. In medicine, various disease trends could be monitored. (Chromosomes exhibit anomalies in leukemia and mental retardation as well as in other states.) In space exploration and experimentation, the device can spot monitor radiation dose levels as well as changes resulting from any of the environmental stresses experienced in space. This apparatus can be modified for use as an extraterrestrial-life-detecting instrument by scanning the growth of cells (or cellular inclusions), computing rates, and telemetering changes to the researcher.
Investigations of rhythmic phenomena of various physiological systems can result in knowledge of the utmost importance to medicine. Rhythmic phenomena are found in the cardiovascular system of normal humans. Changes in these rhythms have the potential of foretelling abnormalities (heart disease, arteriosclerosis) before outward signs are manifested, allowing for earlier diagnosis, treatment, and control or cure.
The spacecraft sterilization program requires the use of rooms having the lowest attainable level of bacterial contamination. The rate of dissemination of bacteria from the humans in the room is basic to the problem. Data on this matter are being obtained through support of the Communicable Disease Center of the U.S. Public Health Service. The findings are affecting the measures used in surgical practice to lower infection rates.
Studies on the physiology of hibernation in mammals are important to understand temperature regulation and the mechanism of survival at low body temperatures. The purpose of this type of research is to understand and use reduced metabolic activity in astronauts on future extended space flight. Other applications involve studies of the mechanisms of injury and freezing biological organisms, for improving techniques in hypothermic surgery, pathology, and preservation of tissue for human grafting.
Food and Agriculture
Gathering agricultural information by remote sensing of Earth's surface from aircraft, balloons, and satellites has a potential application in research and development. Current needs for data gathered in this way include crop and livestock surveys for marketing planning; soil mapping; crop disease, insect, and weed surveys; soil conservation management and research; and crop acreage control programs. As population and world trade increase, the needs will become even more intense for regularly scheduled synoptic surveys of the world's agricultural lands for crop plantings and harvests; determining the condition of crops as affected by drought, disease, or insect outbreaks; and studies of the lands suitable for agricultural development in underdeveloped countries. The only way that worldwide synoptic surveys can be made is by using orbiting platforms.
The NASA nutrition program for developing diets for prolonged manned and animal space flight lends itself to civil defense purposes; military maneuvers where space and weight are prime considerations; polar and desert exploration; reducing hunger in underdeveloped countries; and detecting metabolic diseases as well as diseases of infancy and old age. For space research such a diet can be used on prolonged manned space flights, animal experiments in space, manned orbiting laboratories, and space and planetary stations. Studies on the packaging and stability of foods under various conditions of humidity, temperature, and radiation will lead to better processing and storage.
Learning how microbial spores are transported by air is important to biology, agriculture, and medicine. Besides spreading crop destruction, microbial spores produce allergic responses in some human beings. To obtain the facts, not only the biology of micro-organisms but also the weather factors that induce the flight of mature spores must be known. Thus, both biological and meteorological problems are involved. Data obtained under a NASA contract with the General Mills Electronic Division (now part of Litton Industries, Inc.) indicate that spores of fungi are present in low numbers in the stratosphere. A reservoir of spores exists which cannot be brought down by the normal scrubbing mechanisms of rainfall and other meteorological disturbances in the troposphere. This finding has important implications for reducing the spread of agricultural crop diseases and for protecting persons suffering from allergies. This project has indicated the necessity for designing novel biological samplers for use in the stratosphere. Such samplers will aid in determining various pollutants of the atmosphere.
The NASA program for developing sterile spacecraft for the biological exploration of Mars will contribute improved methods of sterilization that can be applied to the canning industry. Studies on sterilization at low temperatures for long periods of time are being supported by NASA at the Massachusetts Institute of Technology and the Communicable Disease Center and the Sanitary Engineering Center of the Public Health Service. The developing capability is making possible the heat sterilization of products that never before could be thoroughly sterilized.
In preparing for missions to search for extraterrestrial life, research on the psychrophilic or cold bacteria, on halophytic or salt bacteria, and on specialized bacteria and other organisms growing in extreme environments is defining the extremes under which life can exist. Increased knowledge about organisms that can grow in or on refrigerated, dried, or salted foods and other materials should have practical applications for food storage and preservation. Research on psychrophilic bacteria is being conducted by Whirlpool Corp. and the NASA Ames Research Center.
Theoretical studies of Martian life involve investigations of plant and bacterial spores. Many of these forms are spoilage organisms and some produce lethal toxins. This work has potential importance for food processing and for obtaining more precise knowledge of how wounds become infected. The program for investigating possible forms of life on Mars includes a thorough study of anaerobic micro-organisms. This research has led to the discovery of new types of nitrogen-fixing bacteria other than the familiar types found in the root nodules of leguminous plants. Thus, it may be possible to use these microorganisms, or the principles involved, in the incorporation of vital atmospheric nitrogen into terrestrial soils which are now unproductive.
Industry and Manufacturing
Batteries that have been developed in the space program to endure high sterilization temperatures for extended times will have greatly increased shelf life at normal storage temperatures and will be serviceable after many hours of baking at high temperatures.
Currently, the highest quality tape recorders are subject to imperfect reproduction because the tapes are heat labile; i.e., they soften and stretch when warm. The development of high-quality magnetic tapes for space-data recorders is an outgrowth of the materials developed to meet spacecraft sterilization requirements. These improved tapes will be useful for all types of recording—industry, automation controls, home, and studio.
OUTLOOK FOR BIOSCIENCE—MAJOR PROBLEMS
The problems undertaken are among the most challenging, if not the most challenging, man faces on the space frontier. These include the quest for the origin of life, the explanation of life and life processes, the elucidation of the environment's role in establishing and maintaining normal organization in living organisms, the possibility of extraterrestrial life on other planets—the concern of exobiology. The greatest promise for their solution lies in advances in biological theory rather than other avenues of research; therefore, it is fortunate that the need to solve them has come at a time when developments in experimental biology are at a high level. In addition, technological developments in electronics and engineering are providing new and wonderful instruments for this great exploration into the sources of life. Many of these have had practical application that has made possible important advances in medical diagnosis and treatment.
The broad national space goals initially charted by NASA have gone beyond space flight in near-Earth orbit to lunar and interplanetary exploration by man and machine. For such missions, more intensive and comprehensive research in the life sciences is needed. Before manned voyages for extended periods into deep space will be possible, solutions must be found for problems such as the development of bioregenerative life-support systems, communication with nonhuman species, and the development of new methods for transferring knowledge to the human brain.
The problems are all of the type that could perhaps be solved by truly great advances in biological theory, and probably not by any other avenue.
Notes
| [1] | This section includes part of the Summary of the Panel on Radiation Biology of the Environmental Biology Committee Space Science Board, NAS/NRC (1963), and results of research by the Bioscience Programs, NASA. |
| [2] | Excerpt from [ref.130]. |
| [3] | From [ref.169]. |
| [4] | From [ref.174]. |
| [5] | From [ref.175]. |
| [6] | From [ref.176]. |
| [7] | Includes instrumentation and food storage. |
| [8] | Trace amounts of the following were also found: methionine sulfoxide, citrulline, alpha-amino-n-butyric acid, homocitrulline, glucosamine, galactosamine, methionine sulfoximine, ethionine, and ethanolamine. |
| [9] | Includes part of [ref.196]. |
| [10] |
References
| [ref.1] | Rea, D. G.: The Evidence for Life on Mars. Nature, vol. 200, 1963, p. 114. |
| [ref.2] | (1, 2) Öpik, E. J.: Spectroscopic Evidence of Vegetation on Mars. Irish Astron. J., vol. 5, 1958, pp. 12-13. |
| [ref.3] | Kuiper, G. P.: Visual Observations of Mars, 1956. Astrophys. J., vol. 125, 1957, p. 307. |
| [ref.4] | Föcas, J. H.: Seasonal Evolution of the Fine Structure of the Dark Areas of Mars. Planetary Space Sci., vol. 9, 1962, p. 371. |
| [ref.5] | Dollfus, A.: Interpretation of the Polarization of Light Reflected by the Different Regions of the Surface of Mars. Compt. Rend., vol. 233, 1951, p. 467. |
| [ref.6] | Kuiper, G. P.: On the Martian Surface Features. Publ. Astron. Soc. Pacific, vol. 67, 1955, p. 271. |
| [ref.7] | Sinton, W. M.: Spectroscopic Evidence for Vegetation on Mars. Astrophys. J., vol. 126, 1957, p. 231. |
| [ref.8] | Rea, D. G.; Belsky, T.; and Calvin, M.: Interpretation of the 3- to 4-Micron Infrared Spectrum of Mars. Science, vol. 141, 1963, p. 923. |
| [ref.9] | (1, 2) Rea, D. G.; O'Leary, B. T.; and Sinton, W.: Mars and the Origin of the 3.58- and 3.69-Micron Minima in the Infrared Spectra. Science, vol. 147, 1965, p. 1286. |
| [ref.10] | (1, 2) Quimby, F. H.: Concepts for Detection of Extraterrestrial Life. NASA SP-56, 1964. |
| [ref.11] | Urey, H. C.: The Planets. Their Origin and Development. Yale Univ. Press (New Haven), 1952. |
| [ref.12] | (1, 2) Miller, S. L.: Mechanism of Synthesis of Amino Acids by Electric Discharge. Biochim. Biophys. Acta, vol. 23, 1957, p. 480. |
| [ref.13] | Oró, J.: Synthesis of Adenine From Ammonium Cyanide. Biochem. Biophys. Res. Commun., vol. 2, 1960, p. 407. |
| [ref.14] | (1, 2) Ponnamperuma, C.; Sagan, C.; and Mariner, R.: Synthesis of Adenosine Triphosphate Under Possible Primitive Earth Conditions. Nature, vol. 199, 1963, p. 222. |
| [ref.15] | (1, 2, 3, 4) Oparin, A. I.: The Origin of Life. Macmillan Co. (New York), 1938. |
| [ref.16] | (1, 2) Calvin, M.: Reduction of Carbon Dioxide in Aqueous Solutions by Ionizing Radiation. Science, vol. 114, 1951, p. 416. |
| [ref.17] | Miller, S. L.: A Production of Amino Acids Under Possible Primitive Earth Conditions. Science, vol. 117, 1953, pp. 528-529. |
| [ref.18] | Oró, J.: Studies in Experimental Cosmochemistry. Ann. N. Y. Acad. Sci., vol. 108, 1963, pp. 464-481. |
| [ref.19] | Ponnamperuma, C.: Chemical Evolution and the Origin of Life. Nature, vol. 201, 1964, pp. 337-340. |
| [ref.20] | Bernal, J. D.: The Physical Basis of Life. Routledge & Kegan-Paul (London), 1951. |
| [ref.21] | Fox, S.: Anhydrocopolymerization of Amino Acids Under the Influence of Hypothetically Primitive Terrestrial Conditions. Federation Proc., vol. 13, 1954, p. 211. |
| [ref.22] | Akabori, S.: On the Origin of the Fore-Protein. In The Origin of Life on the Earth. A. I. Oparin et al., eds., Pergamon Press (New York), 1955, p. 189. |
| [ref.23] | (1, 2) Harada, K.; and Fox, S. W.: The Thermal Synthesis of Amino Acids From a Hypothetically Primitive Terrestrial Atmosphere. In The Origin of Prebiological Systems and Their Molecular Matrices. Academic Press (New York and London), 1965. |
| [ref.24] | Oparin, A. I.: Biochemical Processes in the Simplest Structures. In The Origin of Life on the Earth. Pergamon Press (New York), 1959. |
| [ref.25] | (1, 2) Fox, S. W.: Simulated Natural Experiments in Spontaneous Organization of Morphological Units for Proteinoid. In The Origin of Prebiological Systems and of their Molecular Matrices. Academic Press (New York), 1965. |
| [ref.26] | (1, 2) Briggs, M. H.; and Mamikunian, G.: Organic Constituents of the Carbonaceous Chondrites. Space Sci. Rev., vol. 1, 1963, p. 647. |
| [ref.27] | Wilk, H. B.: Composition of Some Stony Meteorites. Geochim. Cosmochim. Acta, vol. 9, 1956, p. 279. |
| [ref.28] | Briggs, M. H.: Organic Extracts of Some Carbonaceous Meteorites. Life Sciences, vol. 2, 1963, p. 1. |
| [ref.29] | Mueller, G.: Interpretation of the Microstructures of Carbonaceous Chondrites. Nature, vol. 196, 1962, p. 929. |
| [ref.30] | Mamikunian, G.; and Briggs, M. H.: Some Microstructures of Complex Morphology Observed in Preparations of Carbonaceous Choridrites Made Under Sterile Conditions. Nature, vol. 197, 1963, p. 1245. |
| [ref.31] | Kaplan, L. D.; Munch, G.; and Spinrad, H.: An Analysis of the Spectrum of Mars. Astrophys. J., vol. 139, 1964, p. 1. |
| [ref.32] | Davis. D. R.; and Libby, W. F.: Positive Ion Chemistry. Science, vol. 144, 1961, p. 991. |
| [ref.33] | Miller, S. L.: Production of Some Organic Compounds Under Possible Primitive Earth Conditions. J. Am. Chem. Soc., vol. 77, 1955, p. 235. |
| [ref.34] | Oró, J.: Experimental Cosmochemistry. In Current Aspects of Exobiology. G. Mamikunian and M. H. Briggs, eds., Pergamon Press (New York), 1965. |
| [ref.35] | Shapley, H.: Of Stars and Men. Beacon Press (Boston), 1958, pp. 11 and 157. |
| [ref.36] | Lederberg, J.; and Cowie, D. B.: Moondust. Science, vol. 127, 1958, pp. 1473-1475. |
| [ref.37] | CETEX: Development of International Efforts To Avoid Contamination by Extraterrestrial Exploration. Science, vol. 128, 1958, pp. 887-889. |
| [ref.38] | CETEX: Contamination by Extraterrestrial Exploration. Nature, vol. 183, 1959, pp. 925-928. |
| [ref.39] | Sagan, C.: Biological Contamination of the Moon. Proc. Natl. Acad. Sci. U.S., vol. 46, 1960, pp. 393-401. |
| [ref.40] | Davies, R. W.; and Communizis, M. G.: Sterilization of Space Vehicles To Prevent Extraterrestrial Biological Contamination. Proc. 10th Intern. Astro. Congr., Springer-Verlag (Vienna), 1960, pp. 495-504. |
| [ref.41] | Lederberg, J.: Exobiology: Approaches to Life Beyond the Earth. Science, vol. 132, 1960, pp. 393-400. |
| [ref.42] | Brown, A. H.: Report of the Working Subgroup on Space Probe Sterilization. A Review of Space Research. Natl. Acad. Sci.-Natl. Res. Council, Publ. 1079, 1962, ch. 10. |
| [ref.43] | Imshenetskii, A. A.: Prospects of the Development of Exobiology. In Space Research III, W. Priester, ed., North-Holland Publ. Co. (Amsterdam), 1963. |
| [ref.44] | Portner, D. M.; Spiner, D. R.; Hoffman, R. K.; and Phillips, C. R.: Effect of Ultrahigh Vacuum on Viability of Microorganisms. Science, vol. 134, 1961, p. 2047. |
| [ref.45] | Morelli, F. A.; Fehlner, F. P.; and Stembridge, C. H.: Effects of Ultra-High Vacuum on Bacillus subtilis var. niger. Nature, vol. 196, 1962, pp. 106-107. |
| [ref.46] | Davis, N. S.; Silverman, G. J.; and Keller, W. H.: Combined Effects of Ultrahigh Vacuum on the Viability of Some Spores and Soil Organisms. Appl. Microbiol., vol. II, 1963, pp. 202-211. |
| [ref.47] | Silverman, G. J.; Davis, N. S.; and Keller, W. H.: Exposure of Microorganisms to Simulated Extraterrestrial Space Ecology. In Life Sciences and Space Research II, M. Florkin and A. Dollfus, eds., North-Holland Publ. Co. (Amsterdam), 1964, pp. 372-384. |
| [ref.48] | Imshenetskii, A. A.; Bogrov, N.; and Lysenko, S.: Resistance of Microorganisms to High Vacuum. Doklady Akad. Nauk SSSR, vol. 154, 1964, pp. 1188-1190. |
| [ref.49] | (1, 2, 3) Hawrylewicz, E. J.; Gowdy, B.; and Ehrlich, R.: Microorganisms Under a Simulated Martian Environment. Nature, vol. 193, 1962, p. 497. |
| [ref.50] | Packer, E.; Scher, S.; and Sagan, C.: Biological Contamination of Mars. Part II. Cold and Aridity as Constraints on the Survival of Terrestrial Microorganisms in Simulated Martian Environments. Icarus, vol. 2, 1963, pp. 292-316. |
| [ref.51] | Scher, S.; Packer, E.; and Sagan, C.: Biological Contamination of Mars: I. Survival of Terrestrial Microorganisms in Simulated Martian Environments. In Space Research IV. P. Muller, ed., North-Holland Publ. Co. (Amsterdam), 1964. |
| [ref.52] | (1, 2) Young, R. S.: Deal, P. H.; Bell, J.; and Allen, J. L.: Bacteria Under Simulated Martian Conditions. In Space Research IV, P. Muller, ed., North-Holland Publ. Co. (Amsterdam), 1963. |
| [ref.53] | Hagen, C. A.; Hawrylewicz, E. J.; and Ehrlich, R.: Survival of Microorganisms in a Simulated Martian Environment. Appl. Microbiol., vol. 12, 1964, pp. 215-218. |
| [ref.54] | Hawrylewicz, E. J.; Hagen, C. A.; and Ehrlich, R.: Response of Microorganisms to a Simulated Martian Environment. In Life Sciences and Space Research III, M. Florkin, ed., North-Holland Publ. Co. (Amsterdam), 1965. |
| [ref.55] | Nicks, O. W.; and Reynolds, O. E.: Decontamination and Sterilization of Lunar and Planetary Spacecraft. Science, vol. 142, 1963, pp. 539-540. |
| [ref.56] | Phillips, C. R.; and Hoffman, R. K.: Sterilization of Interplanetary Vehicles. Science, vol. 132, 1960, pp. 991-995. |
| [ref.57] | Hobby, G.: Review of NASA-JPL Spacecraft Sterilization Program. A Review of Space Research. Natl. Acad. Sci.-Nat. Res. Council, Publ. 1079, 1962, ch. 10. App. III. |
| [ref.58] | Bruch, C. W.; Koesterer, M. G.; and Bruch, M. K.: Dry-Heat Sterilization: Its Development and Application to Components of Exobiological Space Probes. Develop. Ind. Microbiol., vol. 4, 1963, pp. 334-342. |
| [ref.59] | Koesterer, M. G.: Thermal Death Studies on Microbial Spores and Some Considerations for the Sterilization of Spacecraft Components. Develop. Ind. Microbiol., vol. 6, 1964, pp. 268-276. |
| [ref.60] | (1, 2, 3) Hall, L. B.; and Bruch, C. W.: Procedures Necessary for the Prevention of Planetary Contamination. In Life Sciences and Space Research III, M. Florkin, ed., North-Holland Publ. Co. (Amsterdam), 1965. |
| [ref.61] | Portner, D. M.; Hoffman, R. K.; Decker, H. M.; and Phillips, C. R.: The Level of Microbial Contamination in a Clean Room During a One-Year Period. Rept. 11-65 on NASA Interagency Agreement R-35, 1964. |
| [ref.62] | Bruch, C. W.: Some Biological and Physical Factors in Dry-Heat Sterilization: A General Review. In Life Sciences and Space Research II. M. Florkin and A. Dollfus, eds., North-Holland Publ. Co. (Amsterdam), 1964. |
| [ref.63] | Bruch, C. W.: Dry-Heat Sterilization for Planetary-Impacting Spacecraft. Tech. Rept. on Spacecraft Sterilization. Jet Propulsion Lab., 1965. |
| [ref.64] | (1, 2) Pollard, E.: Pilot Theoretical Study of the Effect of Weightlessness and Densely Ionizing Radiation on Single Cells. Progr. Rept. on Contract NsG-182-62. NASA, 1962. |
| [ref.65] | Lyon, C. J.: Auxin Transport in Leaf Epinasty. Plant Physiol., vol. 38, 1963, pp. 567-574. |
| [ref.66] | McKinney, R.; Montgomery, P. O'B.; and Gell, C. F.: A Study of the Effects of Zero Gravity on Cell Physiology. In Physical and Biological Phenomena in a Weightless State. Second AAS Symp. Phys. Biol. Phenomena Under Zero Gravity Conditions. E. T. Benedikt and R. W. Halliburton, eds., Advan. Astronaut. Sci., vol. 14, 1963, pp. 291-306. |
| [ref.67] | (1, 2) Committee on Environmental Biology: Report of Panel on Gravity. Space Science Board, Natl. Acad. Sci.-Natl. Res. Council, 1964. |
| [ref.68] | Chase, H. B.: Cutaneous Effects of Primary Cosmic Radiation. J. Aviation Med., vol. 25, 1954, p. 388. |
| [ref.69] | (1, 2) Zhukova, A. I.; and Kondratyev, I. I.: On Artificial Martian Conditions Reproduced for Microbiological Investigations. In Life Sciences and Space Research III, M. Florkin, ed., North-Holland Publ. Co. (Amsterdam), 1965, pp. 120-126. |
| [ref.70] | Davis, I.; and Fulton, J. D.: Microbiological Studies on Ecological Considerations of the Martian Environment. USAF SAM Review, vol. 2, 1959, p. 60. |
| [ref.71] | Roberts, T. S.; and Irvine, L. A.: Studies With a Simulated Martian Environment. Germination and Growth of Bacterial Spores. Tech. Doc. Rept. 63-75. USAF School Aerospace Med., 1963. |
| [ref.72] | Roberts, T. L.; and Wynn, E. S.: Studies With a Simulated Martian Environment. Bacterial Survival and Soil Moisture Content. Tech. Doc. Rept. 62-121. USAF School Aerospace Med., 1962. |
| [ref.73] | (1, 2) Siegel, S. M.; Giumarro, C.; and Latterall, R.: Behavior of Plants Under Extraterrestrial Conditions: Seed Germination in Atmospheres Containing Nitrogen Oxides. Proc. Natl. Acad. Sci. U.S., vol. 52, 1961, pp. 11-13. |
| [ref.74] | Siegel, S. M.; Halpern, L. A.; Giumarro, C.; Renwick, G.; and Davis, G.: Martian Biology: The Experimentalist's Approach. Nature, vol. 193, 1963, pp. 329-331. |
| [ref.75] | Kiess, C. C.; Corliss, C. H.; and Kiess, H.: Evidence for Oxides of Nitrogen in the Atmosphere of Mars. Science, vol. 131, 1960, p. 1319. |
| [ref.76] | Vallentyne, J. R.: Environmental Biophysics and Microbial Ubiquity. Ann. N.Y. Acad. Sci., vol. 108, 1963, pp. 342-352. |
| [ref.77] | Henry, J. P.; Ballinger, E. R.; Maher, P. J.; and Simons, D. G.: Animal Studies of the Subgravity State During Rocket Flight. J. Aviation Med., vol. 23, 1952, p. 421. |
| [ref.78] | (1, 2) Van der Wal., F. L.; and Young, W. D.: A Preliminary Experiment With Recoverable Biological Payloads in Ballistic Rockets. Project MIA. Rept. 715-58, Am. Rocket Soc., 1958. |
| [ref.79] | Van der Wal, F. L.; and Young, W. D.: Project MIA (Mouse-in-Able) Experiment on Physiological Response to Space Flight. ARS J., vol. 29, 1954, p. 716. |
| [ref.80] | (1, 2) Graybiel, A.; Holmes, R. A.; Beischer, D. E.; Champlin, G. E.; Pedigo, G. P.; Hixson, W. C.; Davis, T. R. A.; Barr, N. L.; Kistler, W. G.; Niven, J. T.; Wilbarger, E.; Stullken, D. E,; Augerson, W. S.; Clark, R.; and Berrian, J. H.: An Account of Experiments in Which Two Monkeys Were Recovered Unharmed After Ballistic Space Flight. Aerospace Med., vol. 30, 1959, p. 871. |
| [ref.81] | Green, C. D.; Welch, B. E.; Brown, W. L.; Lamb, L. E.; Tang, P. C.; Gisler, D. B.; and Blodgett, H. C.: Studies of Escape From Ballistic Space Vehicles. Rept. 61-29, School Aviation Med., sec. 1, pp. 1-24; sec. 2, pp. 1-16, 1961. |
| [ref.82] | Belleville, R. E.; Rohles, F. H.; Grunzke, M. E.; and Clark, F. C.: Development of a Complex Multiple Schedule in the Chimpanzee. J. Exp. Anal. Behav., vol. 6, 1963, pp. 549-556. |
| [ref.83] | Henry, J. P.; and Mosely, J. D.: Results of the Project Mercury Ballistic and Orbital Chimpanzee Flights. NASA SP-39, 1963. |
| [ref.84] | Grunzke, M. E.: A Liquid Dispenser for Primates. J. Exp. Anal. Behav., vol. 4, 1961, p. 326. |
| [ref.85] | Grunzke, M. E.: Feeding Devices for Use With Primates in Space Flight. Tech. Doc. Rept. 61-35, USAF Missile Develop. Center, 1961. |
| [ref.86] | Gilbert, G. A.: A Zero Gravity Pellet Dispenser for Use With Primates in Long-Term Space Flights. ARL Tech. Rept. 64-15, 1964. |
| [ref.87] | Meehan, J. P.; Fineg, J.; and Mosely, J. D.: The Effect of Restraint and Training on the Arterial Pressure of the Immature Chimpanzee. Federation Proc., vol. 23, 1964. |
| [ref.88] | Findley, J.; and Weissman, N.: "Counting" in a Baboon. Paper presented at Psychonomic Soc. Meeting (New York), 1961. |
| [ref.89] | Findley, J.; and Brady, J. V.: Exposure to Total and Continuous Environmental Control With a Single Human Organism. Paper presented at Symp. at 7th Annual Meeting, Human Factors Soc. (Palo Alto, Calif.), 1963. |
| [ref.90] | Findley, J.: An Experimental Outline for Building and Exploring Multioperant Behavior Repertories. J. Exp. Anal. Behav., vol. 5, 1962, pp. 113-116. |
| [ref.91] | Beasley, J.; and Seldeen, B.: The Effect of Prolonged Acceleration on Eating and Performance. NASA TM X-54:077, 1964. |
| [ref.92] | Smith, A. H.; Winget, C. M.; and Kelly, C. F.: Growth and Survival of Birds Under Chronic Acceleration. Growth, vol. 23, 1959, p. 97. |
| [ref.93] | Winget, C. M.; Smith, A. H.; and Kelly, C. F.: Effects of Chronic Acceleration on Induced Nystagmus in the Fowl. J. Appl. Physiol., vol. 17, 1962, p. 709. |
| [ref.94] | (1, 2) Wunder, C. C.: Food Consumption of Mice During Continual Centrifugation. Proc. Iowa Acad. Sci., vol. 68, 1961, pp. 616-624. |
| [ref.95] | Wunder, C. C.: Survival of Mice During Chronic Centrifugation. Aerospace Med., vol. 33, 1962, pp. 866-870. |
| [ref.96] | Wunder, C. C.; Lutherer, L. C.; and Dodge, C. H.: Survival and Growth of Organisms During Life-Long Exposure to High Gravity. Aerospace Med., vol. 34, 1963, pp. 5-11. |
| [ref.97] | Dodge, C. H.; and Wunder, C. C.: Growth of Turtles During Continual Centrifugation. Proc. Iowa Acad. Sci., vol. 69, 1962, pp. 594-599. |
| [ref.98] | Oyama, J.; and Platt, W.: Effects of Deceleration on Rats Exposed to Prolonged Centrifugation. Nature, vol. 203, 1964, pp. 766-767. |
| [ref.99] | Matthews, B. H. C.: Adaptation to Centrifugal Acceleration. J. Physiol., vol. 122, 1953, p. 31. |
| [ref.100] | Riccio, D.: The Effects of Vestibular Stimulation on Performance Under Several Schedules of Reinforcement. Paper presented at Eastern Psych. Assoc. Meeting (Atlantic City, N.J.), 1965. |
| [ref.101] | Guedry, F. E.; and Biberman, N.: Apparent Adaptation Effects in Vestibular Reactions. Rept. 293, USA Med. Res. Lab., 1957. |
| [ref.102] | Guedry, F. E.; Cramer, R. L.; and Koella, W. P.: Experiments on the Rate of Development and Rate of Recovery of Apparent Adaptation Effects in the Vestibular System. Rept. 338. USA Med. Res. Lab., 1958. |
| [ref.103] | Guedry, F. E.; and Graybiel, A.: The Appearance of Compensatory Nystagmus in Human Subjects as a Conditional Response During Adaptation to a Continuously Rotating Environment. Rept. 531, USA Med. Res. Lab., 1962. |
| [ref.104] | Loret, B. J.: Optimization of Manned Orbital Satellite Vehicle Design With Respect to Artificial Gravity. ASD Tech. Rept. 61-688, Wright-Patterson AFB, Ohio, 1961. |
| [ref.105] | Belleville, R. E.; Clark. F. C.; and Lange, K. O.: The Behavior of Small Animals Under the Accelerative Conditions Found in Space Travel. NASA TT F-9080, 1964. |
| [ref.106] | (1, 2) Hebb, D. O.: The Organization of Behavior. John Wiley & Sons, Inc., 1949. |
| [ref.107] | (1, 2) Jeffress, L. A.: Cerebral Mechanisms in Behavior. John Wiley & Sons, Inc., 1951. |
| [ref.108] | Konorsky, J.: Mechanisms in Animal Behavior. Symp. Soc. Exp. Biol., vol. 4, Academic Press, 1950. |
| [ref.109] | Eccles, J. C.: The Neurophysiological Basis of Mind. Oxford Univ. Press (London), 1953. |
| [ref.110] | Thorpe, W. H.: Learning and Instinct in Animals. Methuen (London), 1956. |
| [ref.111] | Sperry, R. W.: On the Neural Basis of the Conditioned Response. Brit. J. Anim. Behav., vol. 3, 1955, p. 41. |
| [ref.112] | Burns, B. D.: The Mammalian Cerebral Cortex. Monographs Physiol. Soc. (London), vol. 5, 1958. |
| [ref.113] | Krech, D.; Rosenzweig, M. R.; and Bennett, E. L.: Dimensions of Discrimination and Level of Cholinesterase Activity in Cerebral Cortex of the Rat. J. Comp. Physiol. Psychol., vol. 49, 1956, p. 261. |
| [ref.114] | Krech, D.; Rosenzweig, M. R.; and Bennett, E. L.: Correlation Between Brain Cholinesterase and Brain Weight Within Two Strains of Rats. Am. J. Physiol., vol. 196, 1959, pp. 31-32. |
| [ref.115] | Bennett, E. L.; Rosenzweig, M. R.; Krech, D.; Karlsson, H.; Dye, N.; and Ohlander, A.: Individual, Strain, and Age Differences in Cholinesterase Activity of the Rat Brain. J. Neurochem., vol. 3, 1958, p. 153. |
| [ref.116] | (1, 2) Rosenzweig, M. R.; Krech, D.; and Bennett, E. L.: Effect of Pentobarbital Sodium on Adaptive Behavior in the Rat. Science, vol. 123, 1956, pp. 371-372. |
| [ref.117] | Rosenzweig, M. R.; Krech, D.; and Bennett, E. L.: A Search for Relations Between Brain Chemistry and Behavior. Psychol. Bull., vol. 57, 1960, p. 476. |
| [ref.118] | Hyden, H.: Biochemistry of the Central Nervous System. Pergamon Press (New York), 1959. |
| [ref.119] | Hyden, H.; and Egyhazi, E.: Nuclear RNA Changes of Nerve Cells During a Learning Experiment in Rats. Proc. Natl. Acad. Sci. U.S., vol. 48, 1962, p. 1366. |
| [ref.120] | Brachet, J.; and Mirsky, A. E.: The Cell: Biochemistry, Physiology, Morphology. Vol. IV, ch. 5. Academic Press (New York), 1960. |
| [ref.121] | Tower, D. B.; and Schade, J. P.: Structure and Function of the Cerebral Cortex. Elsevier (New York), 1960. |
| [ref.122] | Dingman, W.; and Sporn, M. B.: The Incorporation of 8-Azaguanine Into Rat Brain RNA and Its Effect on Maze Learning by the Rat. An Inquiry Into the Biochemical Basis of Memory. J. Psychiatric Res., vol. 1, 1961, pp. 1-11. |
| [ref.123] | Adey, W. R.; Kado, R. T.: Didio, J.; and Schindler, W. J.: Impedance Changes in Cerebral Tissue Accompanying a Learned Discriminative Performance in the Cat. Exp. Neurol., vol. 7, 1963, pp. 282-293. |
| [ref.124] | Brattgard, S. O.: Acta Radiol. Suppl., vol. 96, 1952, p. 1. |
| [ref.125] | Geiger, A.: Correlation of Brain Metabolism and Function by the Use of a Brain Perfusion Method In Situ. Physiol. Rev., vol. 38, 1958, pp. 1-20. |
| [ref.126] | Geiger, A.: Metabolism of the Nervous System. D. Richter, ed., Pergamon Press (New York), 1957. |
| [ref.127] | Thompson, R.; and McConnell, J.: Classical Conditioning in the Planarian Dugesia dorotocephala. J. Comp. Physiol. Psychol., vol. 48, 1955, p. 65. |
| [ref.128] | McConnell, J. V.; Jacobson, A. L.; and Kimble, D. P.: Effects of Regeneration Upon Retention of a Conditioned Response in the Planarian. J. Comp. Physiol. Psychol., vol. 52, 1959, p. 1. |
| [ref.129] | Corning, W. C.; and John, E. E.: Effects of Ribonuclease on Retention of a Conditioned Response in Regenerating Planarians. Science, vol. 134, 1961, p. 1363. |
| [ref.130] | Gerathewohl, S. J.; and Gernandt, B. E.: Physiological and Behavioral Sciences, Vol. 1, NASA SP-11, 1962, pp. 399-413. |
| [ref.131] | Von Bekesy, G. J.: DC Potentials and Energy Balance of the Cochlear Partition. Acoust. Soc. Am., vol. 23, 1951, p. 576. |
| [ref.132] | Trincker, D.: Bestandspotentiale in Bogengangssystem des Meerschweinchens und ihre Anderungen bei Experimentellen Cupula-Ablenkunges. Arch. Ges. Physiol., Bd. 264, 1957, p. 351. |
| [ref.133] | Livingston, R. B.: Neural Mechanisms of the Auditory and Vestibular Systems. Ch. 25, G. L. Rasmussen and W. F. Windle, eds., Charles C Thomas (Springfield, Ill.), 1960. |
| [ref.134] | Brodal, A.: Neural Mechanisms of the Auditory and Vestibular Systems. G. L. Rasmussen and W. F. Windle, eds., Charles C Thomas (Springfield, Ill.), 1960. |
| [ref.135] | Brodal, A.; Pompeiano, O.; and Walberg, F.: The Vestibular Nuclei and Their Connections, Anatomy, and Functional Correlations. Charles C Thomas (Springfield, Ill.), 1960. |
| [ref.136] | Carpenter, M. B.: Neural Mechanisms of the Auditory and Vestibular Systems. Ch. 22, G. L. Rasmussen and W. F. Windle, eds., Charles C Thomas (Springfield, Ill.), 1960. |
| [ref.137] | Dow, R. S.; and Moruzzi, G.: The Physiology and Pathology of the Cerebellum. Univ. Minn. Press (Minneapolis), 1958. |
| [ref.138] | Kempinsky, W. H.: Cortical Projection of Vestibular and Facial Nerves in the Cat. J. Neurophysiol., vol. 14, 1951, p. 203. |
| [ref.139] | Mickle, W. A.; and Ades, H. W.: A Composite Sensory Projection Area in the Cerebral Cortex of the Cat. Am. J. Physiol., vol. 170, 1952, p. 682. |
| [ref.140] | Walzl, E. M.; and Mountcastle, V.: Projection of Vestibular Nerve to Cerebral Cortex of the Cat. Am. J. Physiol., vol. 159, 1949, p. 595. |
| [ref.141] | Anderson, S.; and Gernandt, B. E.: Acta Oto-Laryngol. Suppl., vol. 116, 1954, p. 10. |
| [ref.142] | Gernandt, B. E.; and Thulin, C. A.: Vestibular Mechanisms of Facilitation and Inhibition of Cord Reflexes. Am. J. Physiol., vol. 172, 1953, pp. 653-660. |
| [ref.143] | Gernandt, B. E.; and Terzuolo, C. A.: Effect of Vestibular Stimulation on Strychnine-Induced Activity of the Spinal Cord. Am. J. Physiol., vol. 183, 1955, pp. 1-8. |
| [ref.144] | Gernandt, B. E.; Katsuki, Y.; and Livingston, R. B.: Functional Organization of Descending Vestibular Influences. J. Neurophysiol., vol. 20, 1957, pp. 453-469. |
| [ref.145] | Gernandt, B. E.; and Gilman, S.: Vestibular and Propriospinal Interactions and Protracted Spinal Inhibition by Brain Stem Activation. J. Neurophysiol., vol. 23, 1960, pp. 269-287. |
| [ref.146] | Gernandt, B. E.; and Gilman, S.: Interactions Between Vestibular, Pyramidal and Cortically Evoked Extrapyramidal Activities. J. Neurophysiol., vol. 23, 1960, pp. 516-533. |
| [ref.147] | Akert, K.; and Gernandt, B. E.: Electroencephalog. Clin. Neurophysiol., vol. 14, 1962, p. 383. |
| [ref.148] | Gerathewohl, S. J.; and Ward, J. E.: The Physics and Medicine of the Upper Atmosphere and Space. O. O. Benson and H. Strughold, eds., John Wiley & Sons, Inc., 1960, ch. 26. |
| [ref.149] | Gerathewohl, S. J.: Personal Experiences During Short Periods of Weightlessness Reported by Sixteen Subjects. Astronaut. Acta, vol. 2, 1956, pp. 2O5-217. |
| [ref.150] | Henry, J. P.; Augerson, W. S.; Belleville, R. E.; Douglas, W. K.; Grunzke, M. K.; Johnston, R. S.; Laughlin, P. C.; Mosely, J. D.; Rohles, F. H.; Voas, R. B.; and White, S. C.: Effects of Weightlessness in Ballistic and Orbital Flight. Aerospace Med., vol. 33, 1962, pp. 1056-1068. |
| [ref.151] | Dayhoff, M. O.; Lippincott, E. R.; and Eck, R. V.: Thermodynamic Equilibria in Prebiological Atmospheres. Science, vol. 146, 1964, p. 1461. |
| [ref.152] | Armstrong, G. T.; Furukawa, G. T.; and Hilsenrath, J.: A Survey of Thermodynamic Properties of the Compounds of the Elements CHNOPS. NBS Rept. 8521, U.S. Dept. Com., 1964. |
| [ref.153] | White, W. B.; Johnson, S. M.; and Dantzig, G. B.: J. Chem. Phys., vol. 28, 1958, p. 751. |
| [ref.154] | Jukes, T. H.: Present Status of the Amino Acid Code. J. Am. Dietet. Assoc., vol. 45, 1964, p. 517. |
| [ref.155] | (1, 2) Nirenberg, M. W.; and Matthaei, J. H.: The Dependence of Cell-Free Protein Synthesis in E. coli upon Naturally Occurring or Synthetic Polyribonucleotides. Proc. Natl. Acad. Sci. U.S., vol. 47, 1961, p. 1588. |
| [ref.156] | Ochoa, S.: Chemical Basis of Heredity, the Genetic Code. Experientia, vol. 20, 1964, p. 57. |
| [ref.157] | Kiesow, L.: The Energy-Transforming Step in Nitrobacter-Chemosynthesis. Biochem. Z., Bd. 338, 1963, p. 400. |
| [ref.158] | Kiesow, L.: On the Assimilation of Energy From Inorganic Sources in Autotrophic Forms of Life. Proc. Natl. Acad. Sci. U.S., vol. 52, 1964, p. 980. |
| [ref.159] | Clayton, R. K.; and Adler, H. I.: Protein Synthesis and Viability in X-Irradiated Rhodopseudonomonas spheroides. Biochim. Biophys. Acta, vol. 56, 1962, p. 257. |
| [ref.160] | Pollard, E.; and Vogler, C.: Radiation Action on Some Metabolic Processes in E. coli. Radiation Res., vol. 15, 1961, p. 109. |
| [ref.161] | Novelli, G. D.; Kameyama, T.; and Eisenstadt, J. M.: Cold Spring Harbor Symp. Quant. Biol., vol. 26, 1961, p. 133. |
| [ref.162] | (1, 2) Kepes, A.: Kinetics of Induced Enzyme Synthesis. Determination of the Mean Life of a Galactosidase-Specific Messenger RNA. Biochim. Biophys. Acta, vol. 76, 1963, p. 293. |
| [ref.163] | Fernandez-Moran, H.: New Approaches in Correlative Studies of Biological Ultrastructure by High-Resolution Electron Microscopy. J. Roy. Microscop. Soc., vol. 83, 1964, p. 183. |
| [ref.164] | Fernandez Moran, H.: Electron Microscope-Medicines Research: Tool of Unfulfilled Promise. J. Am. Med. Assoc. 189, 1964, p.31. |
| [ref.165] | Fernandez Moran, H.: Electron Microscope With High Field Superconducting Solenoid Lenses. Proc. Natl. Acad. Sci. U.S., vol.53, 1965, p. 445. |
| [ref.166] | Deaver, B. S. Jr.; Swedlund, J. B.; and Bradley, H. Jr.: Magnetic Properties of Some Macromolecules of Biological Interest. Final Rept. PHU 4644. Stanford Res. Inst., 1964. |
| [ref.167] | Sisakyan, N. M.: Problems of Space Biology. Vol. I, USSR Acad. Sci. Publ. House (Moscow) 1962. |
| [ref.168] | Sisakyan, N. M. and Yazdovskiy V. I.,: Problems of Space Biology. Vol. III. USSR Acad. Sci. Publ. House (Moscow), 1964. |
| [ref.169] | (1, 2) Jenkins D. W.,: The NASA Biosatellite Program. In Life Sciences and Space Research III, M. Florkin, ed,. North-Holland Publ. Co. (Amsterdam), 1965, pp. 230-240. |
| [ref.170] | Pack N.: The Effects of Weightlessness on Mammals. Space Biology: Proc. 24th Annual Biol. Colloq., Oregon St. Univ., 1963, pp. 65-74. |
| [ref.171] | Anon.: A Review of Space Research. Ch. 9, Natl. Acad. Sci.-Natl. Res. Council. Publ. 1079, 1962. |
| [ref.172] | Committee on Environmental Biology: Report of Panel on Radiation Biology. Space Science Board, Natl. Acad. Sci.-Natl. Res. Council, 1963. |
| [ref.173] | Taleot, J. M.: Life Support in Space Operations. Air Univ. Rev., vol. 16, 1965, pp. 42-52 |
| [ref.174] | Del Duca, M. G.; Konecci, E. B.; and Ingelfinger A. L.: Life Support-the Next Generation. Space Aeronautics, vol. 41, 1964, pp. 84-91. |
| [ref.175] | (1, 2) Bongers. L.; and Kok B.: Life Support for Space Missions. Develop. Ind. Micro-biol., vol. 5, 1964, pp. 183-195. |
| [ref.176] | (1, 2) Ward, C. H.; Wilks. S. S.; and Craft, H. L.; Use of Algae and Other Plants in the Development of Life Support Systems. Am. Biol. Teacher, vol. 25, 1963, pp. 512-521. |
| [ref.177] | (1, 2) Ney L. F.: Gas Exchange by the Duckweed Family. ONR Contract No. Nonr-2887(00), Stanford Res. Inst., 1960. |
| [ref.178] | Nakamura, H.: A Study of Wolffia as a New Food. Rept. Microalgal Res. Inst. Japan., vol. 1, 1960, pp. 7-13. |
| [ref.179] | (1, 2, 3) Myers, J.: Basic Remarks on the Use of Plants as Biological Gas Exchangers in a Closed System. In Epitome of Space Medicine. J. Aviation Med., vol. 25, 1954, pp. 407-411. |
| [ref.180] | Bowman, N. J.: The Food and Atmosphere Control Problems on Space Vessels. J. Brit. Interplanet. Soc, vol. 12, 1953, pp. 118-123. |
| [ref.181] | Bowman, N. J.: The Food and Atmosphere Control Problems on Space Vessels. Part II. The Use of Algae for Food and Atmosphere Control. J. Brit. Interplanet. Soc. Vol. 12, 1953, pp. 159-166. |
| [ref.182] | Bassham, J. A.: Experiments With Photosynthetic Gas Exchangers. Proc. World Symp. Appl. Solar Energy, Phoenix, Ariz., 1955, pp. 35-50. |
| [ref.183] | (1, 2, 3) Burk D.; Hobby, G.; and Gaucher, T.: Closed-Cycle Air Purification With Algae. Proc. First Intern. Symp. Submarine Space Med. Macmillan Co. (New York), 1958. |
| [ref.184] | Dole, S. H.; and Tamplin, A. R.: The Sabatier Reaction for Inorganic Recovery of Oxygen in Manned Space Capsules. In Closed Circuit Respiratory Systems Symp. Tech. Rept. 60-574, Wright Air Develop. Div., 1960, pp. 239-287. |
| [ref.185] | Clifford, J. E.; and McCallum, J.: Research on the Electrolysis of Water Under Weightless Conditions. AMRL-Tech. Doc. Rept. 62-44, 1962. |
| [ref.186] | Clifford, J.; and Faust, C.: Research on Electrolysis of Water With a Hydrogen Diffusion Cathode To Be Used in a Rotating Cell. AMRL-Tech. Doc. Rept. 62-94, 1962. |
| [ref.187] | (1, 2) Rupaske, R.: Nutritional Requirements for Hydrogenomonas eutropha. J. Bacteriol, vol. 83, 1962, pp. 418-422 |
| [ref.188] | Bongers, L.: Chemosynthetic Gas Exchanger. Quarterly Progress Rept. No. 4 on NASA Contract NASw-971, 1965, 23 pp. |
| [ref.189] | (1, 2) Bongers, L.: Chemosynthetic Gas Exchanger. Ann. Rept. on NASA Contract NASw-713, 1964, 44 pp. |
| [ref.190] | Bongers, L.: Chemosynthetic Gas Exchanger. Rept. RM 153-1963. Martin-Marietta Space Systems Div. (Baltimore, Md.), 1963 |
| [ref.191] | (1, 2, 3) Bongers, L.: Sustaining Life in Space—A New Approach. Aerospace Med., vol. 35, 1964, pp. 139-144. |
| [ref.192] | Schlegel, H.G.: Die Verwendung von H2-oxydierenden Bakterien zur Regenerierung der Atemluft. Raumfahrtforschung, Bd. 8. Nr. 2, 1964, pp. 65-67. |
| [ref.193] | Webb. P., ed.: Bioastronautics Data Book. NASA SP-3006, 1964. |
| [ref.194] | (1, 2) Foster. J. F.; and Litchfield, J. H.: A Continuous Culture Apparatus for the Microbial Utilization of Hydrogen Produced by Electrolysis of Water in Closed-Cycle Space Systems. Biotech. Bioeng., vol. 6, 1964, pp. 441-456. |
| [ref.195] | Callaway. D.; and Margen, S.: Nutritional Properties of Hydrogenomonas eutropha. Semiann. Progr. Rept., NASA Grant NGR-05-003-(089), 1965. |
| [ref.196] | Working Group on Gaseous Environment for Manned Spacecraft: Summary Report. Space Science Board. NAS/NRC, 1963. |
| [ref.197] | Lawton, R. W.: Physiological Considerations Relevant to the Problem of Prolonged Weightlessness. Astronaut. Set. Rev., vol. 4, 1962, pp. 1-16. |
| [ref.198] | McCally, M.; and Lawton. R. W.: The Pathophysiology of Disuse and the Problem of Prolonged Weightlessness: A Review. AMRL-Tech. Doc. Rept. 63-3, Aerospace Med. Div. (Wright-Patterson AFB, Ohio), 1963. |
| [ref.199] | Smith, R. E.; and Hock, R. J.: Brown Fat: Thermogenic Effector of Arousal in Hibernators. Science, vol. 140, 1963, p. 199. |
| [ref.200] | Storer. J. B.; and Hempelman L. H.: Hypothermia and Increased Survival Rate of Infant Mice Irradiated With X-rays. Am. J. Physiol., vol. 171, 1952, pp. 341-348. |
| [ref.201] | Working Group on Nutrition and Feeding Problems: Summary Report. Space Science Board. Man-in-Space Comm., NAS/NRC, 1963. |
| [ref.202] | Anon.: Conference on Nutrition in Space and Related Waste Problems. NASA SP-70, 1964. |
* U.S. GOVERNMENT PRINTING OFFICE 1966 - O - 798 520
Transcription note
The following typographic errors have been corrected:
p. 2, l. 16: to be called '"exobiology." ——> to be called "exobiology."
Table III.- Ultraviolet/Maximum: 108 erg/cm2, 2537° Å ——> 108 erg/cm2, 2537 Å
p. 58, l. 14-15: approximately 300 atm) ——> approximately 300 atm.)
p. 80, l. -2: (600-1000 ft=c) ——> (600-1000 ft-c)
p. 87, l. 1: 0.2-mM ——> 0.2 mM
p. 99, l. 1-2: faintsess ——> faintness
p. 104, l. 2: hiberation ——> hibernation
p. 116, l. 19: processsing ——> processing
p. 121, l. 1: Hoffman, R. K., ——> Hoffman, R. K.;
p. 124, l. -10: Rosenszweig ——> Rosenzweig
p. 128, l. 29: AMRL Tech. Doc. Rept. ——> AMRL-Tech. Doc. Rept.
Variant spelling: both forms microorganism and micro-organism have been retained, as quoted from different sources or bibliographic reference titles.
Tables: Where necessary, the widths of columns have been adjusted, and some tables have been split to avoid them being excessively wide. In the original text, Table VI was split over two pages but has been rejoined in this version.
References: For ease of searching, references in the text, as well as those in the list of references, have been enclosed in square brackets, e.g. [ref.3].
UTF-8 characters: This version contains UTF-8 characters to represent Greek letters, subscripts, superscripts and some punctuation. If these characters do not appear correctly, you may need to select a different font.