The University of Kansas
Lawrence, Kansas
| NUMBER 1 | APRIL 29, 1971 |
GENUS OSTEOCEPHALUS
When we initiated a study of the herpetofauna at Santa Cecilia in Amazonian Ecuador in 1966, we were immediately confronted with many kinds of animals that we could not identify with the existing literature. Comparisons of our specimens with those preserved in other museums resolved some of the problems, but many identifications could be made only after study of type specimens; even then some determinations remained questionable. We now find that in order to prepare a meaningful account of the herpetofauna of Santa Cecilia, we must complete several taxonomic studies, the limits of which extend far beyond eastern Ecuador. Because of our interests in hylids we have begun our studies on these frogs.
One of us (Trueb, 1970a) studied the cranial osteology of casque-headed hylid frogs and redefined the genus Osteocephalus but did not determine the number of species in the genus. Our work in Amazonian Ecuador resulted in the discovery of the sympatric occurrence of three species at each of two localities; one of these species was found with a fourth species at another locality. Study of museum specimens confirmed the recognition of these four species in the Amazon Basin and lower Amazonian slopes of the Andes. A fifth species from Bolivia and Perú also is included in the genus. Examination of museum specimens has provided data on the geographic [Pg 2] variation in, and distribution of, the five species. However, our conclusions pertaining to some populations need substantiation, because we have been hampered by inadequate material from areas beyond Ecuador. More than half of the 905 specimens of Osteocephalus are from Ecuador, a relatively small part of the total range of the genus.
In this paper we are presenting a taxonomic review of the genus Osteocephalus; of necessity our study has been at the alpha level. We have utilized all of the usual external characters, as well as osteological features in our definitions of the species. Tadpoles and mating calls are available for only one species, O. verrucigerus (Trueb and Duellman, 1970); these and other important systematic characters, such as karyotypes, are not available for the group at this time. Our tendency has been to take a conservative view of species; thus it is doubtful that any subsequent worker will recognize fewer species in the genus. Our observations on these frogs in Amazonian Ecuador are presented in a final section of this paper.
For the loan of specimens or for the provision of working space in their respective institutions, we are indebted to James E. Böhlke, Werner C. A. Bokermann, F. W. Braestrup, Nelly Carrillo de Espinoza, Osvaldo R. da Cunha, Josef Eiselt, M. J. Fouquette, Jr., Alice G. C. Grandison, Jean Guibé, Birgitta Hansson, Walter Hellmich, M. J. Hoogmoed, Robert F. Inger, Konrad Klemmer, Jean Lescure, Alan E. Leviton, Clarence J. McCoy, Robert H. Mount, Charles W. Myers, Umberto Parenti, Günther Peters, James A. Peters, William F. Pyburn, Juan A. Rivero, Dorothy M. Smith, Paulo E. Vanzolini, Greta Vestergren, David B. Wake, Charles F. Walker, Ernest E. Williams, and Richard G. Zweifel.
Study of specimens in European museums was made possible by a grant (No. 5063) from the Penrose Fund of the American Philosophical Society. Field work in Ecuador was partially supported by grants from the Watkins Fund of the Museum of Natural History, University of Kansas. At our base camp at Santa Cecilia, Ecuador, we enjoyed the hospitality of Ing. Ildefonso Muñoz B. Transportation in Ecuador was generously provided by the Texaco Petroleum Company. During the course of our field work Stephen R. Edwards and Thomas H. Fritts contributed directly to our study of Osteocephalus. Michael J. Tyler of the South Australian Museum provided information on the vocal sac structure. We extend our [Pg 3] sincere thanks to all of these persons for their contributions to our endeavors.
We have examined 893 preserved frogs, including the type specimens of all included nominal taxa, 8 skeletons, 1 lot of eggs, and 3 lots of tadpoles that we refer to the genus Osteocephalus; in addition skulls were removed from five preserved specimens, and radiographs were made of 12 other preserved specimens. We have been fortunate in seeing living individuals of all species, except O. pearsoni, but we have colored photographs of a living specimen of that species. Figures 1 and 2 were drawn from projected colored transparencies of living frogs. Terminology follows that of Duellman (1970b). On the distribution maps solid symbols indicate localities from which we have examined specimens; open symbols represent additional locality records based on the literature. Throughout the text specimens are listed by their catalogue numbers preceded by the appropriate museum abbreviation, as follows:
| AMNH | American Museum of Natural History |
| ANSP | Academy of Natural Sciences of Philadelphia |
| ASU | Arizona State University |
| AUM | Auburn University Museum |
| BMNH | British Museum (Natural History) |
| CAS | California Academy of Sciences |
| CAS-SU | Stanford University Collection (In California Academy of Sciences) |
| CM | Carnegie Museum |
| FMNH | Field Museum of Natural History |
| KU | University of Kansas Museum of Natural History |
| MCZ | Museum of Comparative Zoology, Harvard University |
| MIZS | Museo ed Istituto di Zoologi Sistematico, Università di Torino |
| MJP | Museo Javier Prado, Lima |
| MNHN | Muséum National d'Histoire Naturelle, Paris |
| MPEG | Museu Paraense Emiliano Goeldi, Belém |
| MVZ | Museum of Vertebrate Zoology, University of California, Berkeley |
| MZUSP | Museu de Zoología, Universidade da São Paulo |
| NHMG | Naturhistoriska Museet Göteborg |
| NHMW | Naturhistorisches Museum, Wien |
| NHRM | Naturhistoriska Riksmuseet, Stockholm |
| RMNH | Rijksmuseum van Natuurlijke Histoire, Leiden |
| SMF | Senckenbergische Museum, Frankfurt |
| UIMNH | University of Illinois, Museum of Natural History |
| UMMZ | University of Michigan Museum of Zoology |
| UP | Université de Paris |
| UPR-M | University of Puerto Rico, Mayagüez |
| UTA | University of Texas, Arlington |
| USNM | United States National Museum |
| UZM | Universitets Zoologiske Museum, Copenhagen |
| WCAB | Werner C. A. Bokermann, São Paulo, Brasil |
| ZMB | Zoologisches Museum Berlin |
| ZSM | Zoologisches Sammlung München |
Because of the taxonomic confusion that has surrounded the generic name Osteocephalus and two of the species (and their synonyms), we present a brief resumé of the taxonomic history of the group.
Among the amphibians sent to the Muséum National d'Histoire Naturelle in Paris by a Monsieur Leprieur in French Guiana was a single female specimen of a moderately large hylid having a well-ossified skull and smooth dorsal skin. This specimen escaped from the covetous eyes of Johann Tschudi, who prematurely named several species on the basis of specimens in Paris, and survived without an epithet until Duméril and Bibron (1841) proposed for it the name Hyla leprieurii. The description of the species is fairly detailed, but the specimen was not illustrated. This is the earliest trivial name now associated with Osteocephalus.
Fitzinger (1843) in his generic synopsis of amphibians and reptiles proposed the generic name Osteocephalus but did not associate a specific name with the genus. Consequently, Osteocephalus Fitzinger, 1843, is a nomen nudum. Franz Steindachner followed Leopoldo Fitzinger at the Naturhistorisches Museum in Vienna, where he had access to Fitzinger's notes and, of course, the important collections housed in that museum. Steindachner (1862) named two species of Osteocephalus on the basis of Brasilian specimens collected by Johann Natterer. Both species were named in the same publication; O. taurinus appeared on page 77, and O. favolineatus, on p. 80. This is the earliest association of the generic name Osteocephalus with a specific name and a description, both of which satisfy the Code of Zoological Nomenclature for generic availability. Therefore, Steindachner is the authority for the generic name Osteocephalus, which has O. taurinus as the type species by original designation. It is not possible to determine whether or not Steindachner's usage of Osteocephalus was the same as that intended by Fitzinger 19 years earlier.
Steindachner (1862) gave reasonably good descriptions of his two new species and provided excellent illustrations of the two specimens, both large females. Apparently impressed by the similarities between Trachycephalus nigromaculatus Tschudi, 1838, and [Pg 5] Osteocephalus taurinus, Steindachner (1867) used the combination Trachycephalus (Osteocephalus) taurinus. This ambiguous usage for the 1860's precludes our determining if Steindachner was in effect synonymizing Osteocephalus with Trachycephalus or whether he was placing Osteocephalus in a subgeneric status. Steindachner (1867) did not mention O. flavolineatus; perhaps by that time he had concluded that flavolineatus was only a color morph of taurinus.
Cope (1867) placed Hyla leprieurii in the genus Hypsiboas Wagler, 1830. Cope (1874) named Osteocephalus planiceps from Nauta, Perú. The single specimen was among the collections made by the Orton Expedition to the upper Amazon Basin and was deposited in the Academy of Natural Sciences in Philadelphia.
Boulenger (1882) placed both Osteocephalus and Trachycephalus in the synonymy of Hyla; he recognized Hyla taurina (with O. flavolineatus as a synonym), H. leprieurii, and H. planiceps. In the same publication Boulenger named Hyla buckleyi on the basis of 10 specimens in the British Museum from Ecuador; in the description he stated that buckleyi was like leprieurii and taurinus in having paired lateral vocal sacs. Boulenger held a lasting influence on taxonomic herpetology, and his generic synonymy of Osteocephalus was unchallenged until only a decade ago.
Goin (1961) presented a generic synopsis of the genera of hylid frogs, in which he recognized Osteocephalus and stated: "There are perhaps eight or ten species of this genus in South America. Certainly taurinus, britti, leprieuri, buckleyi and pearsoni belong here. O. planiceps is surely a synonym of leprieuri and I believe that garbei is as well. The status of such forms as macrotis, riopastazae, and depressa has not yet been settled." Goin defined Osteocephalus as follows: "Males with paired vocal pouches, one at each angle of the jaw; derm of head not co-ossified with skull but roof of skull exostosed." Trueb (1970a) elaborated on Goin's definition and assuredly included only O. taurinus and O. leprieurii in the genus.
Goin's inclusion of buckleyi, britti, and pearsoni in Osteocephalus was the first association of any of these names with that genus. Duellman (1970a) demonstrated that Garbeana garbei Miranda-Ribeiro, 1926, was a member of the Hyla rubra group. Hyla macrotis Andersson, 1945, is a Phrynohyas. Trueb and Duellman (1970) determined that Hyla verrucigera Werner, 1901, is the earliest name for an Osteocephalus displaying striking sexual dimorphism in coloration and texture of the dorsal skin; Hyla riopastazae Andersson, 1945 (female holotype), and Hyla orcesi [Pg 6] Funkhouser, 1956 (male holotype), were placed in the synonymy of Osteocephalus verrucigerus.
Hyla pearsoni Gaige, 1929, is a small species of Osteocephalus. Our findings substantiate Goin's suggestions relative to two other taxa. Hyla leprieurii britti Melin, 1941, from the Rio Uaupés, Brasil, and Hyla depressa Andersson, 1945, from the Río Pastaza watershed, Ecuador, are members of the genus Osteocephalus, but both are synonyms of earlier names—leprieurii and taurinus, respectively. Another name proposed by Melin (1941), Hyla (Trachycephalus) vilarsi from Taracuá, Brasil, also is placed in the synonymy of O. taurinus.
Cochran and Goin (1970) were unaware of the identities of Hyla verrucigera and riopastazae; they used the later name Osteocephalus orcesi for Colombian frogs that are correctly referred to O. verrucigerus. Although Goin (1961) placed Hyla buckleyi and H. pearsoni in Osteocephalus, Cochran and Goin (1970) recognized a "buckleyi group" in Hyla that included these two species plus a new species, Hyla cabrerai from Amazonian Colombia and Brasil (total of three specimens). Also, these authors named Hyla carri from a single Colombian specimen. Study of the types of Hyla cabrerai, H. carri, and H. festae Peracca, 1904, from Ecuador, reveal that all of these names are synonyms of Osteocephalus buckleyi.
Much of the taxonomic confusion and multiplicity of trivial names is due to the great amount of color variation in taurinus and to the sexual dimorphism in the texture of the dorsal skin in all of the species. The details of variation in these and other characters and our justifications for the synonymies are given in the accounts of the species. All of the trivial names that apply to species herein recognized as members of the genus Osteocephalus are listed in table 1.
Diagnostic Definition.—1) Skull broader than long; 2) dermal roofing bones of skull well ossified, exostosed, and/or co-ossified in some species; 3) prenasal and internasal bones absent; 4) parasphenoid alae posterolaterally oriented; 5) dentigerous processes of prevomers angular (/— —\); 6) vocal sacs paired, posterior, and when inflated protruding posteroventral or posterolateral to angles of jaws; 7) submentalis muscle moderate in size and araphic; 8) intermandibularis muscle undifferentiated and bearing an elongate [Pg 7] median aponeurosis; 9) parotoid glands absent or poorly developed, skin not producing viscous secretion characteristic of Phrynohyas; 10) skin on dorsum tuberculate in males, smooth in females; 11) tympanum large, 60 percent or more of diameter of eye; 12) fingers about one-third, toes more than three-fourths webbed; 13) discs large, round; 14) nuptial excrescences present in breeding males; 15) inner metatarsal tubercle not modified for digging; 16) outer metatarsal tubercle absent; 17) tarsal fold weak or absent; 18) pupil horizontal; 19) palpebrum clear; 20) known tadpoles having two upper and five lower rows of teeth.
| Trivial Name, Original Generic Name, Author, Date | Current Name |
| britti (Hyla leprieurii) Melin, 1941 | O. leprieurii |
| buckleyi (Hyla) Boulenger, 1882 | O. buckleyi |
| cabrerai (Hyla) Cochran and Goin, 1970 | O. buckleyi |
| carri (Hyla) Cochran and Goin, 1970 | O. buckleyi |
| depressa (Hyla) Andersson, 1945 | O. taurinus |
| festae (Hyla) Peracca, 1904 | O. buckleyi |
| flavolineatus (Osteocephalus) Steindachner, 1862 | O. taurinus |
| leprieurii (Hyla) Duméril and Bibron, 1841 | O. leprieurii |
| orcesi (Hyla) Funkhouser, 1956 | O. verrucigerus |
| pearsoni (Hyla) Gaige, 1929 | O. pearsoni |
| planiceps (Osteocephalus) Cope, 1874 | O. taurinus |
| riopastazae (Hyla) Andersson, 1945 | O. verrucigerus |
| taurinus (Osteocephalus) Steindachner, 1862 | O. taurinus |
| verrucigera (Hyla) Werner, 1901 | O. verrucigerus |
| vilarsi (Hyla) Melin, 1941 | O. taurinus |
Content.—As defined here, the genus contains five known species: O. buckleyi (Boulenger), O. leprieurii (Duméril and Bibron), O. pearsoni (Gaige), O. taurinus Steindachner, and O. verrucigerus (Werner).
Distribution.—The Guianas and Amazon Basin; also in the upper Orinoco and Magdalena drainages. Most localities are at elevations below 500 m, but the genus ascends the Amazonian slopes of the Andes to elevations of about 1800 m.
Size and Proportions.—Frogs of the genus Osteocephalus are moderate to large hylids. The largest species is taurinus, attaining a snout-vent length of 103.1 mm; the smallest is pearsoni, which attains a length of 54.7 mm. Considerable intraspecific geographic variation occurs in adult size, especially in taurinus. Females of all species attain a noticeably larger size than males, but no significant differences are apparent in proportions (Table 2).
(Means are given in parentheses below observed ranges)
| Species | N | Snout-vent Length |
Tibia Length/ S-V L |
Foot Length/ S-V L |
Head Length/ S-V L |
Head Width/ S-V L |
Tympanum/ Eye |
|
| O. buckleyi | ♂ | 30 | 37.9-48.1 | 0.478-0.580 | 0.375-0.444 | 0.319-0.357 | 0.329-0.368 | 0.608-0.820 |
| (43.3) | (0.520) | (0.408) | (0.343) | (0.351) | (0.711) | |||
| ♀ | 31 | 48.6-75.1 | 0.476-0.599 | 0.363-0.469 | 0.310-0.358 | 0.318-0.367 | 0.574-0.905 | |
| (61.7) | (0.553) | (0.428) | (0.333) | (0.348) | (0.734) | |||
| O. leprieurii | ♂ | 21 | 41.2-48.4 | 0.514-0.571 | 0.383-0.430 | 0.308-0.357 | 0.326-0.368 | 0.652-0.884 |
| (44.7) | (0.538) | (0.408) | (0.335) | (0.348) | (0.777) | |||
| ♀ | 21 | 46.6-61.5 | 0.516-0.592 | 0.382-0.453 | 0.314-0.343 | 0.328-0.363 | 0.698-0.909 | |
| (57.1) | (0.539) | (0.404) | (0.329) | (0.349) | (0.785) | |||
| O. pearsoni | ♂ | 2 | 45.3-46.2 | 0.481-0.504 | 0.404-0.437 | 0.322-0.335 | 0.327-0.342 | 0.660-0.673 |
| (45.8) | (0.493) | (0.421) | (0.329) | (0.335) | (0.666) | |||
| ♀ | 1 | 54.7 | 0.521 | 0.405 | 0.318 | 0.346 | 0.862 | |
| O. taurinus | ♂ | 59 | 40.3-84.6 | 0.512-0.576 | 0.387-0.445 | 0.296-0.345 | 0.301-0.355 | 0.638-0.896 |
| (66.3) | (0.541) | (0.416) | (0.318) | (0.324) | (0.752) | |||
| ♀ | 45 | 45.1-103.1 | 0.520-0.577 | 0.391-0.448 | 0.306-0.334 | 0.308-0.347 | 0.640-0.817 | |
| (75.8) | (0.542) | (0.420) | (0.321) | (0.327) | (0.758) | |||
| O. verrucigerus | ♂ | 11 | 50.4-54.3 | 0.494-0.552 | 0.409-0.442 | 0.322-0.346 | 0.328-0.344 | 0.623-0.804 |
| (53.0) | (0.519) | (0.427) | (0.333) | (0.337) | (0.730) | |||
| ♀ | 3 | 63.1-65.8 | 0.532-0.561 | 0.435-0.463 | 0.345-0.347 | 0.348-0.379 | 0.692-0.808 | |
| (64.5) | (0.545) | (0.448) | (0.346) | (0.358) | (0.731) | |||
Coloration.—All Osteocephalus are predominantly brown frogs usually with some darker dorsal markings (Figs. 1 and 2). Osteocephalus verrucigerus has a nearly uniform dark brown dorsum and no distinct transverse bars on the limbs, whereas all of the other species have distinct bars on the limbs. The dorsal markings on the body consist of irregular blotches in buckleyi, pearsoni, and taurinus but are narrow transverse marks in leprieurii. A narrow middorsal cream or yellow stripe is present in some individuals of buckleyi and taurinus but absent in all individuals of the other species. The flanks are uniform pale tan in leprieurii and uniform reddish brown in verrucigerus; in the other species the flanks are cream to brown with dark brown or black spots (also dark diagonal marks in some buckleyi). A creamy white anal stripe is present in some specimens of leprieurii but absent in all individuals of other species.
The postocular region, encompassing the tympanum, is dark brown in most specimens. In adults of pearsoni and taurinus the upper lips are dark brown. A pale cream or tan suborbital spot is present in pearsoni and in some taurinus; in some specimens of taurinus the suborbital spot is expanded posteriorly forming a labial stripe on the posterior part of the lip. The labial markings of verrucigerus are similar to the latter pattern, except that in females a distinct, light labial stripe extends the length of the lip. Osteocephalus leprieurii has a distinct, broad, pale labial stripe. The lips are barred cream and dark brown in buckleyi.
The venter is uniform creamy white or pale tan in leprieurii, uniform white in some buckleyi (most males), and uniform tan in some taurinus. The other species and some individuals of taurinus and buckleyi (most females) have dark ventral markings. These markings are most distinctive in verrucigerus, in which the venter is white with bold black mottling and spots (Fig. 3c). Those individuals of taurinus having ventral markings usually have indistinct, diffuse brown spots on the throat and chest (Fig. 3b). Osteocephalus pearsoni is characterized by a fine brown reticulation on the venter and on the anterior and posterior surfaces of the thighs in adults (Fig. 3a). Individuals of buckleyi that have ventral markings vary between the patterns illustrated for pearsoni and taurinus (Figs. 3b and c).
Ontogenetic change in coloration is slight or non-existent in buckleyi, pearsoni, and taurinus, except that juveniles lack ventral markings. A dark blotch on the back and distinct transverse bars on the limbs are evident in juveniles of verrucigerus; these markings are obscured in the adults. Juveniles of leprieurii are olive-brown with yellow dorsolateral stripes; the transverse dark marks, characteristic of the adults, appear before the stripes are lost.
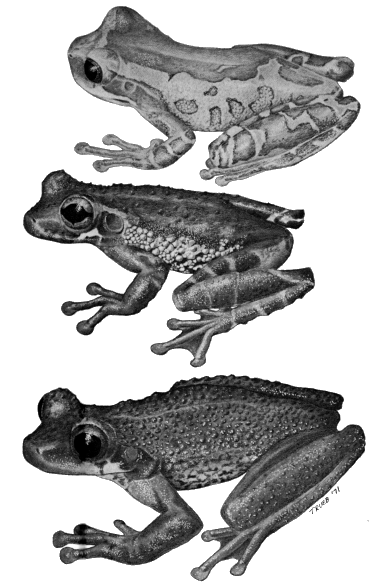
| Fig. 1. | Species of Osteocephalus: |
| Top. O. pearsoni, KU 136312, ♂; Middle. O. buckleyi, KU 123172, ♂; Bottom. O. verrucigerus, KU 123177, ♂. ×1.5. |
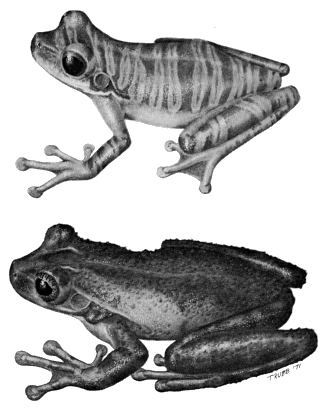
| Fig. 2. | Species of Osteocephalus: |
| Top. O. leprieurii, KU 126611, ♀; Bottom. O. taurinus, KU 126648, ♂. ×10. |
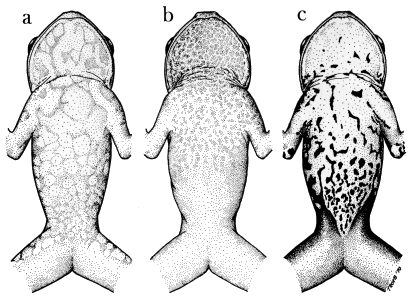
| Fig. 3. | Diagrammatic views of ventral color patterns in Osteocephalus: |
| a. O. pearsoni, UMMZ 57533, ♂; b. O. taurinus, USNM 166037, ♂; c. O. verrucigerus, KU 123185, ♀. |
Skin.—The dorsal skin of all males of Osteocephalus is tuberculate to varying degrees, whereas the dorsal skin of females is smooth, or nearly so (Fig. 4). Osteocephalus verrucigerus differs from other members of the genus by the presence of numerous, large tubercles bearing keratinized tips. The tubercles of leprieurii are numerous and spinous but much smaller than those of verrucigerus; those of taurinus are spinous but less numerous than in leprieurii. Osteocephalus buckleyi has a mixture of large and small, non-spinous tubercles, and pearsoni has only a few, small, scattered, non-spinous tubercles. Fleshy tubercles occur on the eyelids and supratympanic fold in females of buckleyi; a few small tubercles are present on the back of females of pearsoni, whereas the dorsal skin in females of the other species is smooth. The skin on the flanks of both sexes of buckleyi is weakly areolate; in the other species the flanks are smooth. The skin on the top of the head in taurinus is rugose as a consequence of co-ossification. In all species the anal opening is directed posteriorly at the upper level of the thighs.
Hands and Feet.—The feet of Osteocephalus are fully webbed or [Pg 13] nearly so. Webbing between fingers one and two is basal in all species. Webbing between fingers two, three, and four is most extensive in taurinus, in which the three fingers are about one-half webbed (Fig. 5). Osteocephalus buckleyi, pearsoni, and verrucigerus have reduced webbing between fingers two and three, and leprieurii has reduced webbing between fingers two, three, and four. All members of the genus have well-developed subconical subarticular tubercles on the fingers and toes; there is a tendency for the distal tubercle on the fourth finger to be weakly bifid. Palmar and plantar supernumerary tubercles are well developed in taurinus, moderately developed in buckleyi, leprieurii, and pearsoni, and barely evident in verrucigerus. All of the species have a noticeable fold on the wrist and enlarged prepollices, bearing horny nuptial excrescences in breeding males. The prepollex is least enlarged in buckleyi. Outer metatarsal tubercles are absent. The inner metatarsal tubercle is moderately well developed and ovoid in leprieurii and pearsoni; it is elliptical and flat in the other species. Tarsal folds are absent in all species except verrucigerus, in which the folds are barely evident.
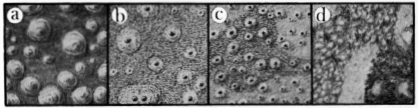
| Fig. 4. | Segments of dorsal skin of males of Osteocephalus showing size and arrangement of tubercles: |
| a. O. verrucigerus, KU 123183; b. O. taurinus, USNM 166033;
c. O. leprieurii, KU 126616; d. O. buckleyi, USNM 165999. Each square = 1 sq. cm. |
Cranium.—As a genus, the cranial structure is remarkably uniform and quite generalized when viewed in the context of the family Hylidae. The skulls are broad and relatively flat, each being only slightly more broad than long and about one-third as high as long. In dorsal aspect the snouts are broadly rounded; the snout of buckleyi is somewhat less rounded and appears to be slightly longer than the snouts of other species. This subtle difference relates to the relative narrowness of the premaxillaries in buckleyi.
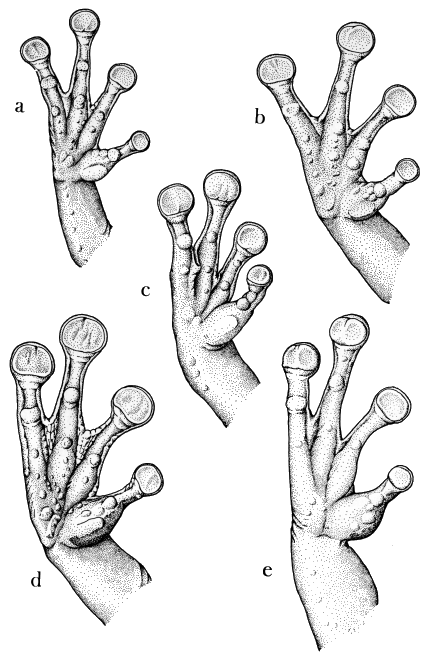
| Fig. 5. | Palmar views of hands of males of Osteocephalus: |
| a. O. buckleyi, KU 109506; b. O. leprieurii, KU 126627; c. O. pearsoni, MCZ 15565; d. O. taurinus, KU 126653; e. O. verrucigerus, KU 123177. ×4. |
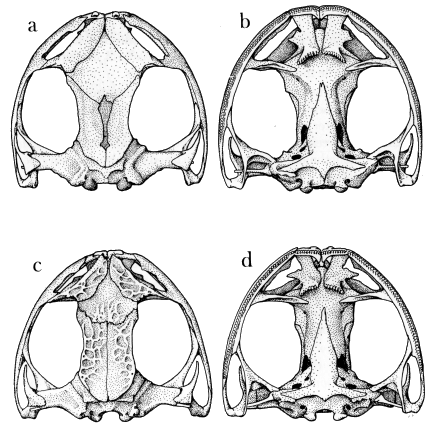
| Fig. 6. | Skulls of two species of Osteocephalus: |
| a and b. O. leprieurii, KU 125961; c and d. O. pearsoni, UMMZ 67465. ×3. |
The genus is characterized by well-developed dermal roofing bones and an unusually large exposure of the sphenethmoid dorsally (Fig. 6). The conformation of the sphenethmoid exposed dorsally is determined by the marginal positions of the adjacent, overlapping elements—the nasals and frontoparietals. Medially the nasals overlap the lateral margins of the sphenethmoid. Anteromedially, the nasals converge in leprieurii and taurinus, are narrowly separated in buckleyi and pearsoni, or are more widely separated in verrucigerus. In all species the nasals terminate at the anterodorsal corner of the orbit. The frontoparietals of buckleyi, leprieurii, and taurinus have an anterolateral extension, which marginally overlaps the dorsolateral edge of the sphenethmoid and articulates with the posterodorsal corner of the nasal in buckleyi and taurinus; the bones are narrowly separated in leprieurii. The frontoparietals of pearsoni and verrucigerus have more extensive median ossification and less extensive anterolateral ossification. Thus, in those species the posteromedian portion of the sphenethmoid is obscured, and the [Pg 16] lateral margins are partly exposed. The frontoparietal fontanelle is completely covered in all species, except buckleyi and leprieurii, in which an irregular, median separation of the frontoparietals exposes a small portion of the fontanelle. The posterolateral margins of the frontoparietals lie medial to the epiotic eminences.
Dermal ornamentation, involving the nasals, frontoparietals, and sphenethmoid, occurs in taurinus and, to a limited extent, in pearsoni. In the latter species marginal portions of the frontoparietals, the dorsal surfaces of the nasals, and the posteromedial part of the exposed sphenethmoid are slightly exostosed, resulting in an open, reticulate pattern of dermal sculpturing of exceedingly low relief (Fig. 6c). Osteocephalus taurinus is characterized by casquing, co-ossification, and a rather intricate pattern of dermal sculpturing, which was described in detail and illustrated by Trueb (1970a).
The squamosals of all species are moderately large structures having otic plates that overlie the lateral portion of the cristae paroticae. The posterior rami are short; the zygomatic rami of all species, except taurinus, extend slightly more than one-half of the distance to the maxillary. In taurinus the zygomatic ramus extends nearly to, or articulates with, the maxillary.
The maxillary arches are complete and relatively robust. The alary processes of the premaxillaries are vertically oriented in leprieurii, pearsoni, and taurinus and very slightly inclined posteriorly in buckleyi and verrucigerus. The maxillaries are uniformly characterized by the absence of postorbital processes and by the presence of preorbital processes that articulate with the maxillary processes of the nasals. The partes facialae of the maxillaries are moderately developed in all species, except taurinus, in which the pars fascialis tends to articulate with the lateral margin of the nasal in well-ossified individuals. The partes palatinae are poorly developed in all species, except buckleyi, in which the pars palatina of the premaxillary is moderately robust.
The pterygoids are uniformly tri-radiate structures. The anterior rami terminate at about the mid-level of the orbit, and the medial rami articulate firmly with the anterolateral corner of the otic capsule. The palatines are well-developed elements bearing ventral ridges; the ridges are somewhat irregular in buckleyi, taurinus, and verrucigerus but smooth in leprieurii and pearsoni. The parasphenoids are large elements characterized by acuminate cultriform processes and posterolaterally inclined alary processes. The basal areas of the cultriform processes bear small odontoid protuberances in buckleyi, taurinus, and verrucigerus, whereas they are smooth in [Pg 17] leprieurii and pearsoni. The prevomers are remarkably uniform, moderately well-developed structures. In each species the anterior ramus lies adjacent to the premaxillary, and the lateral wings form the anterior, medial, and posteromedial margins of the internal nares. The dentigerous processes are characteristically large and angular and bear numerous teeth. The sphenethmoid and otoccipitals are well ossified; a dermal sphenethmoid is present only in taurinus.
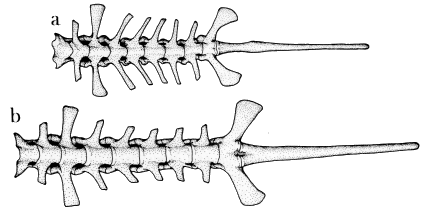
| Fig. 7. | Dorsal views of vertebral columns of two species of Osteocephalus: |
| a. O. leprieurii, KU 125962, ♀; b. O. buckleyi, USNM 165997, ♀. ×2. |
Vertebral Column.—The cervical cotyles are uniformly widely displaced. The neural arches are low and non-imbricate. The transverse processes of the third presacral vertebrae are approximately equal in width to the sacral diapophyses in all species, except buckleyi, in which the processes of the third presacral are slightly narrower than the diapophyses. Osteocephalus buckleyi is further distinguished by the presence of narrow transverse processes on presacrals five through eight (Fig. 7b); males have narrower processes than do females. The processes are moderately wide but subequal in width in pearsoni, taurinus, and verrucigerus, whereas they are nearly equivalent in width to one another and to the sacral diapophyses in leprieurii (Fig. 7a). The sacral diapophyses of all species are moderately dilated and posterolaterally inclined. The coccyx bears a distinct dorsal ridge and has a bicondylar articulation with the sacrum.
Pectoral Girdle.—The pectoral girdles are fully arciferal and bear small, cartilaginous omosterna and moderately large cartilaginous sterna. The coracoids are robust, and the clavicles are strongly arched. Procoracoid cartilage seems to be absent. The [Pg 18] scapulae are large, longer than the clavicles, and bicapitate proximally. The suprascapulae are moderately large and well ossified in leprieurii and taurinus. The suprascapula of verrucigerus is poorly ossified, and that of buckleyi is not ossified.
Pelvic Girdle.—The ilia of buckleyi, taurinus, and verrucigerus lack any indication of a crest on the shaft, whereas leprieurii has a low crest. The dorsal acetabular expansion of the ilia is moderately low in taurinus and verrucigerus, but distinctly lower in buckleyi and leprieurii. The ilia of all species bear low dorsal protuberances. The ischia of leprieurii, taurinus, and verrucigerus are moderately expanded; that of buckleyi is somewhat less expanded dorsally. The pubis of leprieurii, taurinus, and verrucigerus are calcified, whereas that of buckleyi remains cartilaginous.
Throat Musculature and Vocal Sac Structure.—Tyler (1971) described the throat myology of Osteocephalus. The genus is characterized by a moderate-sized araphic submentalis muscle and an undifferentiated intermandibularis having an elongate median aponeurosis. The intermandibularis and submentalis are completely independent in buckleyi, whereas in the other species there is a small attachment between these muscles.
According to Tyler (pers. com.), Osteocephalus has three distinctive types of vocal sac structure which result from differences in the development of the interhyoideus muscle and the overlying skin. In leprieurii and verrucigerus the supramandibular portions of the interhyoideus form a simple tubular, posterolateral extension; there is no modification of the associated skin. Osteocephalus buckleyi and pearsoni have more extensive development of the supramandibular portions of the interhyoideus; furthermore, the associated skin forms a broad, loose fold extending from the ventromedial surface of the throat dorsolaterally to the base of the supratympanic fold. Like buckleyi and pearsoni, the supramandibular portion of the interhyoideus is much expanded in taurinus. The vocal sac structure of taurinus differs from that of other members of the genus in that the skin of taurinus forms an everted pouch, which dangles loosely beneath the supratympanic fold.
| 1. | Inner edge of third finger webbed to mid-length of antepenultimate phalange; dorsum brown with dark brown spots or median blotch; skull in adults casqued and co-ossified with prominent supraorbital flanges |
| O. taurinus | |
| [Pg 19] | Inner edge of third finger webbed to base of antepenultimate phalange; dorsum plain or marked with dark blotches or transverse bars; skull in adults smooth or slightly exostosed, lacking supraorbital flanges |
| 2 | |
| 2. | Skin on flanks areolate; dorsum in males bearing a mixture of large and small non-spinous tubercles; lips distinctly barred |
| O. buckleyi | |
| Skin on flanks smooth; dorsum in males bearing tubercles of uniform size; lips not barred | |
| 3 | |
| 3. | Dorsal pattern consisting of narrow transverse dark bars; dorsum in males bearing numerous small spinous tubercles |
| O. leprieurii | |
| Dorsal pattern not consisting of transverse bars; dorsal tubercles large or few in number | |
| 4 | |
| 4. | Dorsum uniformly dark brown; venter heavily mottled with black, especially in females; dorsum in males bearing large, keratinized tubercles |
| O. verrucigerus | |
| Dorsum tan with irregular dark brown blotches; venter cream with fine brown reticulations; dorsum in males bearing few, small non-spinous tubercles | |
| O. pearsoni | |
Justification of Synonymy.—Boulenger (1882:362) listed 11 specimens in his description of Hyla buckleyi. We have examined all of these and conclude that one (BMNH 1947.2.13.42) is O. leprieurii. Cochran and Goin (1970:213) restricted the type locality to Canelos, Provincia Pastaza, Ecuador; we here select BMNH 1947.2.13.44 from that locality as the lectotype. This specimen is a male having a snout-vent length of 37.9 mm; the diameter of the tympanum [Pg 20] is 3.5 mm, 81.4 percent of the diameter of the eye. The type series, exclusive of BMNH 1947.2.13.42 (= O. leprieurii) consists of six males having snout-vent lengths of 37.9-44.6 (mean 40.4) mm, and four females having snout-vent lengths of 50.0-53.9 (mean 51.5) mm. The dorsum in the males bears a mixture of large and small tubercles, whereas the dorsum in females is nearly smooth. The skin on the flanks, especially the axilla, is areolate. The coloration consists of a creamy tan ground color with irregular reddish brown markings on the back and broad transverse bars on the limbs. The dorsal markings are narrowly bordered by creamy white; those on the back consist of an interorbital bar and a pair of longitudinal marks beginning in the scapular region and usually diverging posteriorly in the sacral region or converging into a broad median blotch. One specimen has a middorsal creamy white stripe from the tip of the snout to the vent. In all of the types large dark brown spots are present on the flanks and posterior surfaces of the thighs. The ventral surfaces are pale creamy tan with or without diffuse brown spots on the throat and chest.
The holotype of Hyla festae is a female having a snout-vent length of 75.0 mm; the diameter of the tympanum is 3.9 mm, 57.4 percent of the diameter of the eye. The skin is smooth on the dorsum and areolate on the anterior part of the flanks. The dorsum is pale brown with a large median longitudinal dark brown blotch on the back and broad transverse bars, narrowly outlined by cream, on the limbs. Dark brown spots are present on the flanks; the posterior surfaces of the thighs are dark brown. The throat and belly are grayish white with irregular dark brown spots.
The holotype of Hyla carri is a female having a snout-vent length of 66.1 mm; the diameter of the tympanum is 4.7 mm, 81.0 percent of the diameter of the eye. The skin on the dorsum is smooth with scattered small tubercles and areolate on the anterior part of the flanks. The dorsum is tan with irregular dark brown blotches on the back and transverse bars on the limbs; all dorsal markings are narrowly outlined by creamy white. Dark brown spots are present on the flanks; the venter and posterior surfaces of the thighs are tan without dark spots.
The holotype of Hyla cabrerai is a female having a snout-vent length of 52.7 mm; the diameter of the tympanum is 4.0 mm, 76.9 percent of the diameter of the eye. The skin on the dorsum is weakly tuberculate and that on the anterior part of the flanks is areolate. The dorsum is creamy tan with dark brown markings (interorbital bar, reticulations on occiput, three longitudinal streaks on back, [Pg 21] and broad transverse bars on limbs). Irregular dark brown spots are present on the flanks. The venter is pinkish tan with small reddish brown spots on the throat and darker brown spots on the chest and belly.
In their description of Hyla cabrerai, Cochran and Goin (1970:217) stated: "This species, together with buckleyi and pearsoni certainly make a closely knit groupߪ. Both buckleyi and cabrerai have long hind legs, with the extended heel reaching to the tip of the snout, while in pearsoni the extended heel reaches only to the eye. H. buckleyi has the belly dusky, while it is heavily spotted in cabrerai and is reticulated in pearsoni. H. cabrerai seems to have the heaviest hands with the most webbing between the fingers; the other two species have the webbing reduced between the fingers." The description of Hyla cabrerai was based on three specimens. We have examined the holotype and one paratype (WCAB 13284 from Territorio do Amapá, Brasil). Another paratype in the private collection of C. J. Goin from Caño Tuí, between Mitú and Raudal de Yurupari, Comisaria de Vaupés, Colombia, was not examined.
Cochran and Goin (1970:211) based their description of Hyla carri on one gravid female and stated: "A large Hyla with the vomerine teeth in two ʌʌ shaped patches between the somewhat squarish choanae; reduced webs between the fingers; and a pattern of dorsal dark blotches bordered by light margins. The species is not similar to any other species known in Colombia. It is perhaps most closely related to Hyla claresignata of Brazil, from which it can be differentiated by its more heavily spotted dorsum, larger tympanum, and lack of dark anal spots."
Except for the inclusion of the name in checklists, Hyla festae has not been mentioned in the literature since the original description.
The wholesale synonymization of names, which, on the bases of their published diagnoses, seem to apply to distinctly different species, is possible with the application of uniform criteria to the types and series of other specimens. In measurements and proportions the type specimens of the nominal taxa all fall within the range of variation exhibited by a series of 18 males and 15 females from Provincia Pastaza, Ecuador, except the ratio of the diameter of the tympanum to that of the eye in the female holotype of Hyla festae. In that specimen the ratio is 0.574, whereas the ratio in the 15 females from Provincia Pastaza is 0.587-0.905 (mean 0.736).
Ventral coloration is the most variable character among the types. The venter in the type of Hyla festae is boldly spotted; it is [Pg 22] distinctly spotted in cabrerai, uniform tan in carri, and tan, flecked, or spotted in the type series of buckleyi. The ventral coloration in series of specimens from Amazonian Ecuador encompasses that observed in all of the types, except that of festae, which has more ventral spotting than any other individual.
The webbing on the hand usually excludes the penultimate phalanges of the fingers, but in some specimens from Amazonian Ecuador the webbing encompasses the proximal parts of the penultimate phalanges of the fingers. In a few of these specimens, the holotype of festae, and one paratype of cabrerai the webbing extends to the middle of the penultimate phalanges of the third and fourth fingers. In the holotype of cabrerai the webbing extends to the middle of the penultimate phalanges of the third and fourth fingers and to the base of the disc of the second finger.
The types of the nominal taxa and series of specimens from Guyana and Amazonian Ecuador display noticeable variation in dorsal coloration. The variety of dorsal patterns of all of the types is included in the variation displayed by the other specimens. All specimens have some amount of dark spotting on the flanks; all have vertically barred lips, on which a pale subocular spot usually is evident. Probably the most unifying physical characteristic of all of the specimens is the nature of the skin on the anterior part of the flank. The skin is elevated amidst an irregular network of depressions. This areolate dermal condition is present in all specimens and does not occur in other species of Osteocephalus. The degree of tubercularity of the skin on the dorsum is variable and sexually dimorphic. All males are tubercular, whereas small females are smooth or have only a few scattered tubercles. Large females usually have pronounced tubercles on the eyelids and supratympanic fold.
In their description of Hyla carri, Cochran and Goin (1970:211) misrepresented the nature of the dentigerous processes of the prevomers, which are angular, not ʌ-shaped. Their suggestion that the Colombian Hyla carri is related to Hyla claresignata in southeastern Brasil is unfounded. The latter species is smaller (40 mm), has a yellow dorsum and venter, dark brown spots dorsolaterally, oblique dentigerous processes of the prevomers, small tympanum, and smooth skin dorsally.
The ventral coloration of the type of Hyla festae resembles that of Osteocephalus verrucigerus, but the type differs from verrucigerus by having areolate skin on the flanks and distinct dark markings on the dorsum. In verrucigerus the skin on the flanks is smooth, [Pg 23] and the dorsum is uniform dark brown, except for a tan snout in females.
Comparisons of the types of the nominal species with series of specimens from Guyana, Colombia, Ecuador, and Perú suggest strongly that the types are representative of one taxon, the oldest name for which is Hyla buckleyi Boulenger, 1882. Consequently, we place Hyla festae Peracca, 1904, Hyla carri Cochran and Goin, 1970, and Hyla cabrerai Cochran and Goin, 1970, as junior synonyms of Hyla buckleyi Boulenger, 1882.
Diagnosis.—1) Size moderate, sexual dimorphism extreme; maximum observed snout-vent length in males 48.1 mm, in females 75.1 mm; 2) skin on dorsum in males bearing a mixture of large and small non-spinous tubercles; 3) skin on flanks, especially anteriorly, areolate; 4) web usually extending only to base of antepenultimate phalange on inner edge of third finger; 5) dorsum pale tan or green with irregular, longitudinal, dark brown blotches, usually narrowly outlined with cream; 6) venter cream or tan, suffused with brown or marked with brown spots in some specimens; 7) lips marked with vertical brown and cream bars; 8) flanks creamy tan with irregular brown spots and/or diagonal marks; 9) dermal roofing bones of skull lacking exostosis; 10) dermal sphenethmoid absent; 11) nasals widely separated medially; 12) anteromedial margin of frontoparietal at mid-level of orbit; 13) frontoparietal fontanelle partially exposed; 14) palatine serrate; 15) parasphenoid bearing odontoids; 16) zygomatic ramus of squamosal extending approximately one-half of distance to maxillary arch; 17) transverse processes of third presacral vertebra narrower than sacral diapophyses; transverse processes of presacral vertebrae 3-8 subequal in width and narrower in males than in females; 18) intermandibularis and submentalis muscles independent; 19) supramandibular portion of interhyoideus extensively developed; associated skin forming broad loose fold.
Osteocephalus buckleyi can be distinguished readily from all other species in the genus by the presence of areolate skin anteriorly on the flanks and by the rather boldly contrasting dorsal pattern. Furthermore, females are distinctive in having tubercles on the eyelids and supratympanic folds.
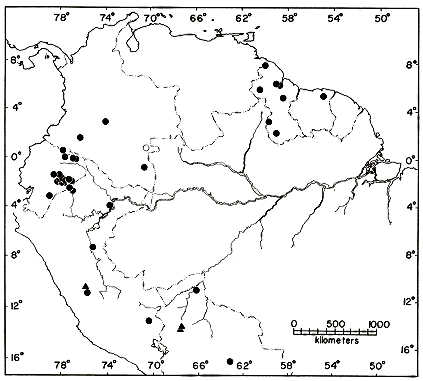
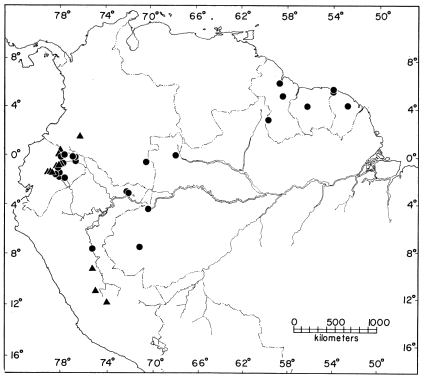
Distribution.—The periphery of the Amazon Basin, in the Guianas and Territorio do Amapá in northeastern Brasil; the upper Amazon Basin from southern Colombia to east-central Bolivia; one locality (Acevedo) in upper Río Magdalena drainage in Colombia (Fig. 8). All localities are at elevations of less than 700 m. Records [Pg 24] for Pallatanga and Santiago in Provincia Chimborazo, Ecuador (high on the Pacific slopes of the Andes), are considered to be erroneous. 78 specimens from 40 localities.
Remarks.—In life the dorsum is green with dark markings. A male (KU 123171) from Santa Cecilia, Ecuador, was: "Dorsum green with dark brown blotches. Anterior and posterior surfaces of thighs dull blue. Venter brown, flecked with white. Iris greenish bronze with brown horizontal triangles and ventromedian brown line." (W. E. Duellman, field notes, 16 June 1968.) A female (KU 126646) from Lago Agrio, Ecuador, was: "Dorsum pale green with darker green blotches and creamy yellow middorsal stripe. Lateral blotches bronze-tan. Flanks tan with black blotches. Anterior surfaces of thighs dark brown. Dorsal and posterior surfaces of thighs and shanks tan with dark brown blotches. Webbing brown. Suborbital spot green. Postorbital bar black. Belly grayish brown in appearance—tips of granules white; intergranular spaces brown. Iris golden bronze with black flecks peripherally and median, horizontal, [Pg 25] reddish brown streak." (W. E. Duellman, field notes, 12 May 1969.)
No ontogenetic change in coloration has been noted.
Justification of Synonymy.—The holotype of Hyla leprieurii is a female having a snout-vent length of 46.6 mm. The diameter of the tympanum is 3.7 mm, 69.8 percent of the diameter of the eye. The dorsal roofing bones are smooth, and the skin on the dorsum is smooth. The penultimate phalanges of the fingers are not included in the webbing. When we examined the specimen on 2 July 1969, it was slightly soft and somewhat faded to a peculiar grayish green color with faint darker transverse bars on the limbs. Duméril and Bibron (1841:554) described the coloration, as follows: "The loreal region is black. A stripe of the same color extends from the posterior border of the orbit to the corner of the mouth, passing through the tympanum. All of the dorsal parts are grayish white with large transverse brown bands, which are more expanded and less regularly outlined on the back than on the limbs. There is one of these on the occiput that is in a triangular shape. All of the venter is white." (Free translation from French.)
The holotype of Hyla leprieurii britti is a male having a snout-vent length of 48.1 mm. The diameter of the tympanum is 3.6 mm, 65.5 percent of the diameter of the eye. The skin on the dorsum is tubercular; the tubercles are small on head and on the dorsal surfaces of the limbs and slightly larger on the back. The penultimate phalanges of the fingers are not included in the webbing. Melin (1941:43) stated: "Above blackish brown with a very indistinct band between the eyes; iris with mottle of metallic lustre; hinder parts of upper jaw whitish; sides of body mottled with blackish brown; hind limbs (especially tibiae and tarsi) with narrow, diffuse cross bars; beneath whitish with slight brown mottle along jaw." We examined the type on 17 February 1969; at that time it was dull brown above with faint, narrow, dark brown, transverse bars on [Pg 26] the back and dorsal surfaces of the limbs. A cream subocular spot was evident, and the venter was creamy white.
Melin (1941:42) stated that the holotype of Hyla leprieurii britti "ߪ resembles a good deal H. leprieurii Dum. & Bibr. As, however, it differs from the latter species by its very concave loreal region, small tympanum, and almost uniformly brownish colour, it may at least form a subspecies of leprieuriiߪ." The pattern of narrow transverse bars on the backs of the holotypes of H. leprieurii and H. britti is a condition shared only by these two nominal taxa that are placed in Osteocephalus. Melin noted that britti differed from leprieurii in the depth of the loreal concavity and in the size of the tympanum. Neither of these differences is noteworthy in comparison with series of specimens. The depth of the loreal concavity is a highly subjective character, and we note no differences between the types. The ratio of the diameter of the tympanum to the diameter of the eye is relatively smaller in both holotypes (0.698 in leprieurii—♀; 0.655 in britti—♂) than in series of fresh specimens from Lago Agrio, Ecuador (0.652-0.884, mean 0.785 in 17 males; 0.700-0.909, mean 0.790 in 20 females). The smaller proportions in the types may be due to geographic variation or to shrinkage as a result of many years in preservative (130+ years for leprieurii; 45 for britti).
Comparisons of the holotypes with series of specimens from Ecuador, Guyana, and Surinam indicate that one morphological species occurs throughout the upper Amazon Basin and the Guianas and that both type specimens are representatives of one species. Consequently, we consider Hyla leprieurii Duméril and Bibron, 1841, to be a monotypic species with Hyla leprieurii britti Melin, 1941, as a junior synonym.
In their account of Osteocephalus leprieurii, Cochran and Goin (1970:323) stated: "The specimen described and illustrated (MCZ 28042) has been directly compared with the types of leprieurii, planiceps, and vilarsi by the junior author and there seems to be no doubt that all are conspecific. Another specimen (CNHM 69716) has been directly compared with the types of planiceps and vilarsi and these, likewise, are considered conspecific." With this justification Cochran and Goin (1970:322) included Osteocephalus planiceps Cope, 1874, and Hyla vilarsi Melin, 1941, in the synonymy of Osteocephalus leprieurii.
We do not concur with Cochran and Goin's synonymy and contend that planiceps and vilarsi are synonyms of Osteocephalus taurinus; we give our reasons in the account of that species. We have examined the specimens listed as O. leprieurii by Cochran and [Pg 27] Goin; several of them, including CNHM (= FMNH) 69716, are taurinus. Thus, due to Cochran and Goin's confusion of two taxa, their comparisons of certain specimens with types has little meaning.
Cochran and Goin did not include Hyla leprieurii britti in their synonymy of Osteocephalus leprieurii but did discuss the name in their account of Osteocephalus orcesi (= O. verrucigerus), as follows (1970:319): "When we first examined one of the specimens we felt sure that we had Melin's Hyla britti at hand, but on direct comparison with the type of britti the two proved to be different. After studying the type of orcesi (SUNHM 13150) we have no doubt that the specimens at hand are orcesi and that britti is a different, probably valid species."
Diagnosis.—1) Size moderate, sexual dimorphism evident; maximum observed snout-vent length in males 48.4 mm, in females, 61.5 mm; 2) skin on dorsum in males bearing numerous, minute, spinous tubercles; 3) skin on flanks smooth; 4) web extending to base of antepenultimate phalange on inner edge of third finger; 5) dorsum tan or olive-brown with transverse brown or olive bars; 6) venter creamy white or pale tan without markings; 7) lips marked with creamy tan labial stripe and suborbital spot; 8) flanks pale tan with no markings; 9) dermal roofing bones of skull lacking exostosis; 10) dermal sphenethmoid absent; 11) nasals juxtaposed medially; 12) anteromedial margin of frontoparietal between mid- and anterior levels of orbit; 13) frontoparietal fontanelle partially exposed; 14) palatine not serrate; 15) parasphenoid lacking odontoids; 16) zygomatic ramus of squamosal extending about one-half of distance to maxillary arch; 17) transverse processes of presacral vertebrae 3-8 about equal in width to one another and to sacral diapophyses; 18) intermandibularis and submentalis muscles connected; 19) supramandibular portion of interhyoideus forming simple tubular posterolateral extension; associated skin unmodified.
Osteocephalus leprieurii differs from all other members of the genus by having transverse dark bars on the back. Two other hylids (Hyla lanciformis and multifasciata) in the Amazon Basin have transverse dark marks on the dorsum. Both of these differ from leprieurii by having pointed snouts, much longer hind limbs, and smooth skin dorsally.
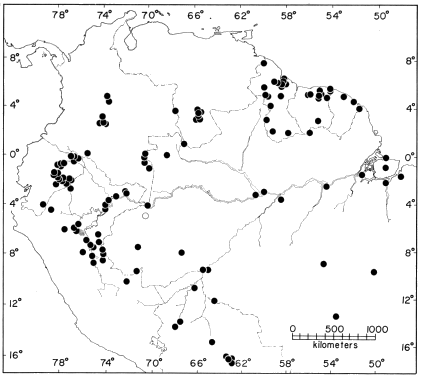
Distribution.—The periphery of the Amazon Basin, in the Guianas and the upper part of the basin in southern Colombia, Ecuador, Perú, and extreme western Brasil (Fig. 9). Most localities are at elevations of less than 500 m, but the species ascends the lower [Pg 28] Andean slopes to elevations of 1100 m. 265 specimens from 31 localities.
Remarks.—Most adults of leprieurii have distinct transverse markings on the back; these are variable in width, extent, and arrangement. In some specimens, such as USNM 166557, some of the transverse bars are fragmented into spots; in a few specimens the dorsal pattern consists solely of small dark spots arranged in transverse rows. Such specimens have a dorsal pattern resembling that of some taurinus. The transverse nature of the dorsal markings is further modified in some specimens, such as USNM 166555, in which the dark bars are fragmented and oblique.
Extreme ontogenetic change in color pattern is exhibited by this species (Fig. 10). Juveniles having snout-vent lengths of less than 28 mm have an olive-brown dorsum with a pale cream stripe across the head and broad, cream, dorsolateral stripes; transverse dark bars are absent on the body and limbs. Individuals having snout-vent lengths of 30-35 mm have dark brown transverse bars on [Pg 29] the back and limbs but still retain the light dorsolateral stripes, whereas the stripes are lost in larger individuals.

| Fig. 10. | Ontogenetic change in color pattern in Osteocephalus leprieurii: |
| a. KU 126644; b. KU 126640; c. KU 126625. ×2. |
Coloration in life of specimens from Lago Agrio, Ecuador: "In males the dorsal ground color varies from dark brown to ochre-tan; dorsal markings uniformly dark brown. Most specimens have dark brown and cream anal stripes; labial area cream-colored. Flanks vary from tan to white. Ventral coloration varies from salmon to tan to white. The iris is bronze with a greenish cast and black reticulations. In females the dorsal coloration is the same as in males, except that dark marks tend to be outlined with cream; venter tannish salmon." (W. E. Duellman, field notes, 12 May 1969).
Justification of Synonymy.—Goin (1961:13) suggested that Hyla pearsoni Gaige was an Osteocephalus, but Cochran and Goin (1970:217) [Pg 30] considered pearsoni to be a Hyla. The presence of exostosed dermal roofing bones, angulate prevomerine dentigerous processes, and the structure of the vocal sacs are characters which place the species in Osteocephalus.
Diagnosis.—1) Size moderate, sexual dimorphism evident; maximum observed snout-vent length in males 46.2 mm, in females 54.7 mm; 2) skin on dorsum in males bearing a few, small, scattered non-spinous tubercles; 3) skin on flanks smooth; 4) web extending to base of antepenultimate phalange on inner edge of third finger; 5) dorsum tan with irregular brown blotches; 6) venter cream with fine brown reticulations; 7) lips dark with pale vertical bar below eye; 8) flanks pale tan with round, brown spots; 9) dermal roofing bones of skull slightly exostosed; 10) dermal sphenethmoid absent; 11) nasals narrowly separated medially; 12) anteromedial margin of frontoparietal between mid- and anterior levels of orbit; 13) frontoparietal fontanelle covered; 14) palatine not serrate; 15) parasphenoid lacking odontoids; 16) zygomatic ramus of squamosal extending about one-half distance to maxillary arch; 17) transverse processes of third presacral vertebra approximately equal in width to sacral diapophyses; transverse processes of presacral vertebrae 3-8 subequal in width; 18) intermandibularis and submentalis muscles connected; 19) supramandibular portion of interhyoideus extensively developed; associated skin forming broad loose fold.
Osteocephalus pearsoni can be distinguished most readily from other members of the genus by the brown reticulate pattern on the venter, round brown spots on the flanks, and smooth skin on the flanks. Also, it is the least tuberculate species in the genus.
Distribution.—Upper Amazon Basin and Amazonian slopes of the Andes in central Perú (1620 m in Río Ucayali drainage) and northern Bolivia (less than 500 m in Río Beni drainage) (Fig. 8). 6 specimens from 3 localities.
Remarks.—The specimen from Yaupi, Perú (KU 136312) is a subadult female having a snout-vent length of 39.8 mm. In life the coloration was: "Dorsum light pinkish brown with large rich chocolate brown blotch from eyes to anterior tips of ilia; numerous small chocolate blotches on flanks; dorsal surfaces of thighs and shanks, canthus, and supraorbital region to insertion of forearm chocolate brown; supralabial border and short bar from eye to lip bronze-white; venter bronze-white with numerous tiny chocolate brown flecks [tending to form reticulations on throat and chest]; anterior and posterior surfaces of thighs light olive-brown; iris largely black with gold flecks." (Thomas H. Fritts, field notes, 23 [Pg 31] March 1970.) On the basis of this one subadult, it seems likely that reticulations on the venter develop with age.
Justification of Synonymy.—The holotype of Osteocephalus taurinus is a female having a snout-vent length of 103.9 mm. The diameter of the tympanum is 6.8 mm, 77.3 percent of the diameter of the eye. The skull is strongly exostosed, and the lateral edges of the frontoparietals are elevated so as to form distinct ridges. The skin on the dorsum is smooth. When we examined the type on 5 August 1969, the specimen was soft and badly faded to a pale creamy tan with pale brown transverse bars on the hind limbs and spots on the flanks. Steindachner (1862:79) described the coloration of the type: "In the preserved specimen the dorsum of the entire body, including fore and hind limbs, is a light yellow-brown color, which becomes lighter towards the venter. The belly is whitish, as are the undersides of the arms and legs. The throat is indistinctly marbled with brown. Roundish dark brown flecks are randomly distributed in a considerable number along the side of the body up to the eye; the tympanum is more or less fully surrounded by brown. A few discrete spots, always more or less drawn out in length, on the sides of the body, are also found on the posterior part of the back. The dorsal surfaces of the fore and hind feet are marked with somewhat obliquely arranged brown transverse bands, which are more intensively colored near the margin than in the middle of the band." (free translation from German.)
The holotype of Osteocephalus flavolineatus is a female having [Pg 32] a snout-vent length of 81.8 mm. The diameter of the tympanum is 6.0 mm, 71.4 percent of the diameter of the eye. The skull is strongly exostosed, and the lateral edges of the frontoparietals are elevated so as to form a ridge on each side. The skin on the dorsum is very weakly tuberculate. We examined the type on 9 August 1969 and found it to be in excellent condition. The color pattern is unchanged from that described by Steindachner (1862:81). The dorsum is tan with irregular brown blotches on the back, spots on the flanks, and transverse bars on the limbs. A narrow creamy white, middorsal stripe extends from the snout to the vent. The subocular area is creamy tan, and the venter is tan. Boulenger (1882:363) questionably synonymized flavolineatus with taurinus. We have observed that a middorsal cream stripe occurs in about 10 percent of the specimens of taurinus and in some specimens of buckleyi. This is a common color morph in many species of Eleutherodactylus. In the absence of distinguishing morphological characteristics we can only conclude that the middorsal stripe is a pattern variant and that Boulenger was correct in synonymizing flavolineatus with taurinus.
The holotype of Osteocephalus planiceps is a male having a snout-vent length of 58.5 mm. The diameter of the tympanum is 4.9 mm, 77.8 percent of the diameter of the eye. The skull is moderately exostosed, and the lateral edges of the frontoparietals are distinctly elevated. The skin on the dorsum is tuberculate. Cope (1874:122) described the coloration of the type as follows: "Color above uniform dark brown, concealed surfaces on the limbs similar and without any markings. Sides a little varied with the white of the belt. A light border to the upper lip, and lighter line from the orbit to the angle of the mouth; dermal scapular fold pale edged. Femur and tibia with dark crossbands on the exposed surfaces." We examined the holotype on 25 September 1969, and found it to be soft and rubbed. The coloration remains much the same as described by Cope, who provided no means of distinguishing planiceps from taurinus. The coloration and morphometric and structural characters of the type of planiceps all fall within the range of variation displayed by series of O. taurinus from the upper Amazon Basin.
The type of Hyla vilarsi is a gravid female having a snout-vent length of 62.7 mm. The diameter of the tympanum is 4.8 mm, 73.8 percent of the diameter of the eye. The dorsal roofing bones of the skull are moderately exostosed, and the lateral edges of the frontoparietals are distinctly elevated. The skin on the dorsum is smooth. [Pg 33] Melin (1941:42) described the coloration of the holotype as follows: "Above uniform reddish brown; upper eyelids and sides of head darkish brown; below the rostral edge a narrow dark band, continuing as a broader light-edged one through the eye and tympanum towards the base of the forelimb and then farther on continuing along the sides as a line of black spots; sides of upper jaw whitish with traces of dark cross bars (one distinct under the eye); sides of body darkish with black spots and marble, often on a whitish ground; thighs, tibiae, and tarsi each with two broad light-edged, dark cross bars on a brownish ground (less distinct on thighs); sides of thighs finely mottled with brown; beneath whitish with small, sparse spots along jaw, on the chest and sides." We examined the type on 17 February 1969, at which time the specimen was somewhat desiccated, especially the hands and feet. The coloration remains much the same as described by Melin, except that he failed to note the presence of four elongate spots on the back.
The status of the names Osteocephalus planiceps Cope and Hyla vilarsi Melin was confused by Cochran and Goin (1970:322), who assigned these names to the synonymy of O. leprieurii. Bokermann (1966:64) placed Hyla vilarsi in the synonymy of Osteocephalus taurinus without justification. The type specimens of both planiceps and vilarsi have moderately exostosed dermal roofing bones and distinct cranial ridges. The type of planiceps has moderately large tubercles on the dorsum, and the type of vilarsi has spots on the throat, chest, and flanks and longitudinal markings on the back. All of these features are characteristic of taurinus and not of leprieurii, which lacks exostosis and cranial ridges and has transverse markings on the back, no spots on the throat, chest, and flanks, and in males has small dorsal tubercles.
The type of Hyla depressa is a male having a snout-vent length of 69.8 mm. The diameter of the tympanum is 5.2 mm, 77.6 percent of the diameter of the eye. The dorsal roofing bones of the skull are moderately exostosed, and the lateral edges of the frontoparietals are elevated. The skin on the dorsum is tuberculate. The dorsum is dull brown with a broad darker brown longitudinal mark having indistinct lateral edges from the snout to the post-sacral area. A narrow cream middorsal line extends from the snout to the vent. The side of the head is dark brown, palest posteroventral to the orbit. The posterior surfaces of the thighs are dull brown; the flanks are pale brown, and the ventral surfaces are pale creamy tan. Dark brown transverse bars are present on the limbs. When we examined the type on 3 January 1969, it was in excellent condition. [Pg 34] Andersson (1945:75) contrasted the type of Hyla depressa with leprieurii and buckleyi, but he did not compare his specimen with taurinus, from which it exhibits no distinguishing features.
Osteocephalus taurinus is a widespread and variable species, and it has received several specific names. We are convinced that Osteocephalus taurinus Steindachner, 1862, is the oldest available name for this large Amazonian species. The following names are junior synonyms: Osteocephalus flavolineatus Steindachner, 1862; Osteocephalus planiceps Cope, 1874; Hyla (Trachycephalus) vilarsi Melin, 1941; Hyla depressa Andersson, 1945.
Diagnosis.—1) Size large; sexual dimorphism evident; maximum observed snout-vent length in males 84.6 mm, in females 104 mm; 2) skin on dorsum in males bearing many moderately large, spinous tubercles; 3) skin on flanks smooth; 4) web extending to middle of antepenultimate phalange on inner edge of third finger; 5) dorsum brown usually with a large medial dark brown blotch or, less frequently, several dark spots; narrow middorsal yellow line present in some; 6) venter cream or tan with or without small, irregular brown flecks; 7) lips brown with vertical cream bar below eye in some, expanded into pale labial stripe posteriorly in some females; 8) flanks tan or cream with or without small, irregular brown spots; 9) dermal roofing bones of skull exostosed, casqued, and co-ossified (in large adults); 10) dermal sphenethmoid present; 11) nasals juxtaposed medially; 12) anteromedial margin of frontoparietals at mid-level of orbit; 13) frontoparietal fontanelle covered; 14) palatine serrate; 15) parasphenoid bearing odontoids; 16) zygomatic ramus of squamosal usually articulating with maxillary arch; 17) transverse processes of third presacral vertebra approximately equal in width to sacral diapophyses; transverse processes of presacral vertebrae 3-8 subequal in width; 18) intermandibularis and submentalis muscles connected; 19) supramandibular portion of interhyoideus extensively developed; associated skin forming everted pouch.
The moderately rugose dorsum (in males), large size, extensive webbing on the hand, and frontoparietal flanges in adults serve to distinguish taurinus from other members of the genus.
Distribution.—The Amazon Basin, the upper Orinoco Basin, and the Guianas. Most localities are below 500 m, but the species ascends the lower Amazonian slopes of the Andes to elevations of about 1000 m (Fig. 11). A record from Caracas, Venezuela, and those from Provincia Carchi and Provincia Esmeraldas, Ecuador, are considered to be erroneous. The latter specimens were included [Pg 35] in a collection sold to the University of Illinois; contained in the collection are many common Amazonian species unknown from the Pacific lowlands. 516 specimens from 151 localities.
Remarks.—This widespread species is highly variable in size and coloration. Striking differences in snout-vent length are evident in series from various parts of the range. The smallest calling males (CAS-SU 12351-6 from Rio Tapirapé, Brasil) have snout-vent lengths of 46.5-60.3 (mean 53.3) mm, whereas the largest (FMNH 140254, KU 92243-6, WCAB 9997, 10001, 10003-4 from Igarapé Marmelo, Brasil) have snout-vent lengths of 71.5-84.6 (mean 77.6) mm. Mean values of snout-vent lengths of males from other localities are: Río Pastaza drainage, Ecuador 73.8 mm, Surinam 67.7 mm, Río Ucayali drainage, Perú 57.6 mm, and Guyana 55.5 mm. Although the difference between the smallest and largest adults is highly significant, populations bridging the gap do exist. Furthermore, the geographic arrangement of small versus large frogs is a confusing mosaic. We have entertained the thought that we have included more than one species in taurinus, but on the basis of preserved [Pg 36] specimens we are unable to detect consistent differences distinguishing two or more taxa.
The coloration and pattern of taurinus are so variable that no one series of statements can describe samples drawn from the entire range of species. We have been unable to determine geographic trends in color pattern; instead the variation within a given sample can encompass the variety known in most other samples. Two minor exceptions do exist. A narrow middorsal light stripe is present in some individuals from throughout the range, but striped specimens are most common in the upper Amazon Basin. The absence of dorsal markings is uncommon in the entire species, but it is most frequent in individuals from the Guianas. A few individuals, such as KU 105230, have scattered white spots on the dorsum.
The coloration of four males in life from Lago Agrio, Ecuador (KU 126652-5) was: "Dorsal ground color tan to dark brown with darker brown markings. Flanks creamy tan to yellow with brown or black flecks or mottling. Venter uniform creamy yellow or yellow with brown spots or reticulations. Iris greenish yellow with radiating black streaks and a median, horizontal reddish brown streak." (W. E. Duellman, field notes, 12 May 1969.) A female from Santa Cecilia, Ecuador (KU 123173), was: "Dorsum mottled olive-green and tan. Flanks tan with brown spots. Belly and throat creamy white, becoming tan posteriorly. Edge of upper jaw olive-green." (W. E. Duellman, field notes, 16 June 1968.) Another female from Santa Cecilia (KU 123175), was: "Brown dorsally with cream-colored mottling. Transverse bars on legs darker brown with cream-colored edges. Margin of upper lip creamy yellow. Anterior and posterior surfaces of thighs tan. Flanks white with brown spots. Venter creamy white. Iris greenish bronze with heavy radiating reticulations of black." (W. E. Duellman, field notes, 22 July 1968.)
The tendency for females to have a labial stripe posteriorly and the absence of dorsal tubercles in females has resulted in the identification of many such specimens as O. leprieurii.
Ontogenetic change in coloration is slight in taurinus. Most juveniles (less than 40 mm in snout-vent length) can be identified readily. There is a tendency for the dorsal markings of juveniles to consist of several small spots. Apparently with growth the spots usually coalesce, forming a large median blotch, but some adults retain the juvenile pattern. Cochran and Goin (1970:251) erroneously identified several juveniles from Colombia as Hyla palpebrogranulata Andersson.
Justification of Synonymy.—Trueb and Duellman (1970:605) discussed the assignment of the names in the synonymy of O. verrucigerus; only a brief resumé is given here.
The extant type of Hyla verrucigera is a juvenile male having a snout-vent length of 32.0 mm. The dorsum is smooth except for tubercles on the eyelids; the skin is loose, and the body is soft. The specimen is faded to a pale brown; indistinct dark spots are present on the back, and transverse bars are evident on the limbs.
The holotype of Hyla riopastazae is a gravid female having a snout-vent length of 64.7 mm. The dorsum is smooth. The dorsal ground color is pale brown with indistinct brown transverse bars on the limbs. The throat, chest, and belly are cream with brown spots and mottling.
The holotype of Hyla orcesi is an adult male having a snout-vent length of 52.6 mm. The dorsum is heavily tuberculate. The dorsum is dark brown with faint transverse bars on the forearms and feet; the ventral surfaces are creamy brown.
Trueb and Duellman (1970) provided conclusive evidence that the types of H. verrucigera, riopastazae, and orcesi are a juvenile, adult female, and adult male, respectively, of one species, the earliest available name for which is Hyla verrucigera Werner, 1901.
Diagnosis.—1) Size moderate, sexual dimorphism evident; maximum observed snout-vent length in males 54.3 mm, in females 65.8 mm; 2) skin on dorsum in males bearing large, keratinized tubercles; 3) skin on flanks smooth; 4) web extending to base of antepenultimate phalange on inner edge of third finger; 5) dorsum uniformly dark brown or black, with tan snout in females; 6) venter creamy white, heavily mottled with black or dark brown, especially in females; 7) lips marked with pale tan labial stripe and suborbital bar; 8) flanks dull reddish brown; 9) dermal roofing bones of skull lacking exostosis; 10) dermal sphenethmoid absent; 11) nasals [Pg 38] widely separated medially; 12) anteromedial margin of frontoparietals at anterior border of orbit; 13) frontoparietal fontanelle covered; 14) palatine serrate; 15) parasphenoid bearing odontoids; 16) zygomatic ramus of squamosal extending approximately one-half of distance to maxillary arch; 17) transverse processes of third presacral vertebra approximately equal in width to sacral diapophyses; transverse processes of presacral vertebrae 3-8 subequal in width; 18) intermandibularis and submentalis muscles connected; 19) supramandibular portion of interhyoideus forming simple, tubular, posterolateral extension; associated skin unmodified.
Osteocephalus verrucigerus can be distinguished from other members of the genus by its uniformly dark dorsum, heavily mottled venter, and large, spinous tubercles on the dorsum in males.
Distribution.—Lower Amazonian slopes (500-1840 m) of the Andes and on the western fringe of the Amazon Basin in Ecuador and Perú; one locality (Acevedo) in upper Río Magdalena drainage in Colombia (Fig. 9). 40 specimens from 13 localities.
Remarks.—In life the dorsum in males is dull olive-green; the groin, anterior and posterior surfaces of the thighs, inner surfaces of limbs, and upper arms are dark brown. The ventral surfaces of the limbs are pinkish tan; the other ventral surfaces are pale creamy tan with reddish brown spots. The suborbital spot is pale greenish tan, and the iris is deep reddish brown. In females the dorsum is dull olive-brown; the anterior part of the head is tan, and the suborbital spot is yellowish tan. The groin and hidden surfaces of the limbs are dark reddish brown. The ventral surfaces of the limbs are brown; the throat and chest are creamy white, and the belly is reddish tan, both with dark brown mottling.
Considerable ontogenetic change occurs in coloration. Juveniles are pale above with a dark median dorsal blotch and dark transverse bars on the limbs. The venter is white. The change consists principally of an increase in dark pigment and subsequent obliteration of the juvenile pattern.
Tadpoles of this species have moderately long tails with low fins, robust bodies, two rows of labial papillae with median part of the upper lip bare, and two upper and five lower rows of teeth. Trueb and Duellman (1970) described the eggs, tadpoles, mating call, and variation in the adults.
Among the 33 genera currently recognized in the family Hylidae, there are two basic types of vocal sac structure (Duellman, 1970b), [Pg 39] namely the subgular type and the lateral type. Only four hylid genera, all Neotropical lowland groups, are known to possess paired lateral vocal sacs; these are Osteocephalus, Argenteohyla, Phrynohyas, and Trachycephalus. The geographical distributions and morphological characteristics of these four genera suggest that they are more closely related to one another than with any other hylid genera.
Of the four genera, Osteocephalus is the most generalized in morphology, and, like Phrynohyas, has no specialized habits. Osteocephalus and Argenteohyla are similarly distinguished from Phrynohyas and Trachycephalus on the basis of vocal sac structure. The vocal sacs of Osteocephalus and Argenteohyla are posterior and protrude posterolateral to the angles of the jaws when they are inflated, whereas those of Phrynohyas and Trachycephalus are more lateral and protrude posterior to the angles of the jaws when inflated.
Although Osteocephalus and Argenteohyla have similar vocal sac structure, they are obviously distinct. The monotypic Argenteohyla is a rather specialized, semifossorial frog (Trueb, 1970b), characterized by smooth skin, moderate-sized digital discs, and a large inner metatarsal tubercle. The general architecture of the skull is not unlike that of Osteocephalus; the skulls of both are well roofed, broader than long, and characterized by posterolaterally oriented parasphenoid alae. Argenteohyla bears small, slightly curved prevomerine dentigerous processes in contrast to the large, angular processes of Osteocephalus. The skull of Argenteohyla shows specializations, apparently adaptations to its semifossorial mode of existence, which further distinguish the genus from Osteocephalus. In comparison with Osteocephalus, the cranium of Argenteohyla is slightly depressed anteriorly, the roofing bones extensively casqued, and the palatines robust.
Osteologically, Osteocephalus more closely resembles Phrynohyas than either of the other two genera, but Osteocephalus and Phrynohyas are clearly distinct on the basis of their respective vocal sac structure. Like Osteocephalus, skulls of the members of the genus Phrynohyas are broader than long, have extensive dermal roofing bones, and have posterolaterally oriented parasphenoid alae. In contrast to Osteocephalus, the dentigerous processes of the prevomers are curved, rather than angular in Phrynohyas. Furthermore, the latter genus is singularly distinguished from Osteocephalus, Argenteohyla, and Trachycephalus by having extensively developed [Pg 40] parotoid glands that produce a viscous, milky volatile secretion.
Trachycephalus is the most readily identifiable of the four genera under discussion. Members of this genus are large frogs with heavily casqued and co-ossified skulls (Trueb, 1970a). The dermal roofing bones bear ornate and characteristic patterns of sculpturing. The medial ramus of the pterygoid does not articulate with the otic capsule, and the parasphenoid alae are laterally, rather than posterolaterally, oriented. A dermal sphenethmoid is present, and the parasphenoid bears odontoids. The basic structure of the skull has many characters in common with both Osteocephalus and Phrynohyas. The obvious modifications of dermal roofing bones and of palatal and suspensory elements seem to be specializations adapting members of the genus Trachycephalus to their peculiar phragmotic habits. The vocal sac structure of Trachycephalus is like that of Phrynohyas and therefore further distinguishes it from Osteocephalus.
Morphologically, Osteocephalus seems to be sufficiently diverse and generalized so as to represent a modern derivative of an ancestral type which might have given rise to Phrynohyas, Trachycephalus, and Argenteohyla. The specialized vocal sac structure in Phrynohyas and Trachycephalus suggests that these two genera may be rather closely allied and represent a single phyletic line from an ancestral stock similar to Osteocephalus. Argenteohyla is quite distinct from Phrynohyas and Trachycephalus and apparently represents a distinct phyletic line from the ancestral stock.
All of our observations on members of this genus have been made at four localities: 1) Santa Cecilia at an elevation of 340 meters on the Río Aguarico, a tributary of the Río Napo, 2) Lago Agrio, 330 meters, about 14 kilometers east of Santa Cecilia, 3) Puerto Libre, 570 meters, on the Río Aguarico just east of its formation by the confluence of the Río Cofanes and Río Chingua, and 4) south slope of the Cordillera del Dué, above the Río Coca, 1150 meters. Osteocephalus leprieurii was found at all four localities, and buckleyi was found at all but the last; taurinus was found at Santa Cecilia and Lago Agrio, and verrucigerus was found only in the Cordillera del Dué. Our data are based on collections of 113 frogs and three lots of tadpoles, as well as observations on calling sites and young. The observations are summarized by species, as follows:
[Pg 41] Osteocephalus buckleyi.—No breeding activity was observed. Males were found only at night in March, June, and July. One was perched on a Heliconia leaf in a swamp at Puerto Libre, and two were on bushes in the forest at Santa Cecilia. A gravid female was found on a recently felled tree at Lago Agrio on the night of 12 May 1969.
Osteocephalus leprieurii.—Males were heard calling sporadically at Puerto Libre in July 1968, and at Santa Cecilia in May 1969. A small chorus was found on the night of 12 May 1969 at Lago Agrio, where the frogs were perched on branches of fallen trees over a temporary pool. The call is a soft rattling chuckle. In late April and May many gravid females and males with well-developed nuptial excrescences were obtained from trees as they were felled at Lago Agrio. The reproductive condition of the frogs indicates that they probably breed in May. One individual called nearly every night from a large tree at Puerto Libre between 4-17 July 1968. The tree was felled on the latter date, but no frog was found. Two nights later apparently the same individual called from a bromeliad at a height of about 10 m on a large bamboo adjacent to the felled tree; the frog was collected when the bamboo was cut down.
Throughout the rainy months that we have worked in Ecuador (April-August) we have found occasional individuals perched on bushes or low trees at night. Large numbers of adults were observed only during a clearing operation which resulted in the felling of many large trees. Thus, it seems likely that leprieurii is a tree-top inhabitant. A partially digested adult male was removed from the stomach of a Hemiphractus proboscideus.
At Santa Cecilia many recently metamorphosed young and juveniles were found in June and July 1968. Most of these were on low bushes or herbs in swamp forest at night; some were found in unfolded Heliconia leaves by day, and one was observed on the forest floor by day. Snout-vent lengths of 18 specimens are 12.3-17.0 (mean 15.1) mm. The smaller frogs were recently metamorphosed as evidenced by the melanophore deposits above the vent. The coloration of the young is strikingly different from that of the adults (see account of O. leprieurii), so the association of the young and adults was not made until individuals with intermediate patterns were obtained at Lago Agrio in May 1969. Probably juveniles obtained in June and July are the offspring of an April or May breeding. We have been unable to associate tadpoles with this species.
Osteocephalus taurinus.—A small chorus occurred at Lago Agrio [Pg 42] on 12 May 1969. Males were calling from the ground adjacent to a small pool amidst recently felled trees. The males were very wary and, when approached, jumped onto limbs and ran up branches; this behavior was noted by Bokermann (1964). The call consists of a series of low-pitched, short notes—like a slow trill—four to six notes per call group. Call groups are repeated two, three, or four times followed by a lapse of several minutes. Although no amplectant pairs were found, several gravid females were collected at Lago Agrio in May, so it can be safely assumed that the species breeds in May. From April through July occasional individuals were observed on bushes and trees at night. During clearing operations at Lago Agrio several individuals were obtained from the tops of trees as they were felled.
Osteocephalus verrucigerus.—Observations were made in a broad, shallow ravine, in which there was a small stream. On 2-4 August 1968, males were observed calling from low bushes and rocks at the edge of a quiet pool in the stream. The call consists of a series of well-pulsed, low-pitched, guttural notes produced at the rate of 5-10 per minute. One amplectant pair was found at the base of a bush adjacent to the pool on 3 August. Another female was found on a branch of a tree 2 m above the ground and 10 m from the stream. Tadpoles of this species were found in the quiet silt-bottomed pool.
The localities for each of the specimens examined are given in the following paragraphs. The arrangement of the data is as follows: alphabetically by country, state (department or province), and locality; alphabetically by the first letter in the abbreviations for the museums, and numerically after each museum abbreviation. Specimens lacking precise locality data are listed first in the most restricted political unit possible; localities which have not been found on maps or the positions of which are not known to us are given in quotation marks. Where more than one specimen is included under one museum number, the number of specimens is given in parentheses after the museum number. Unless noted otherwise, all specimens are alcoholics.
BOLIVIA: El Beni: Ivón, BMNH 1967.2070-1. Santa Cruz: Buenavista, CM 4333, 4339, UMMZ 66563-5.
BRASIL: Amapá: No specific locality, WCAB 13284.
COLOMBIA: Amazonas: Río Guacaya, USNM 152759. Huila: Acevedo, [Pg 43] Río Suaza, FMNH 69702. Nariño: Rumiyacu, FMNH 54756. Meta: Río Guejar, Campamento La Macarena, USNM 152199.
ECUADOR: No specific locality, NHMW 6209, WCAB 35499. Chimborazo: Pallatanga, BMNH 1947.2.13.46; Santiago, FMNH 42529. Morona-Santiago: "Río Santiago" (= Río Zamora), MIZS 2950. Napo: Lago Agrio, KU 126646; Puerto Libre, Río Aguarico, KU 123172; Santa Cecilia, AUM 8138, KU 105208-9, 109506, 123171. Pastaza: Alpayacu, BMNH 1912.11.1.64; Canelos, BMNH 1947.2.13.40-1, 1947.2.13.43-5; Colonia Mena, Río Conambo, ZSM 33/1962; Don Tomás, USNM 166014; Guaché, Río Pastaza, AMNH 79986; Río Bobonaza, USNM 166005; Río Capahuari, USNM 166554; Río Conambo at Río Shiona-yacu, USNM 166018; Río Copataza, upper Río Pastaza, USNM 166007-13; Río Pastaza, NHRM 1946; Río Pucyacu, USNM 165997 (skeleton), 165998-6001; Río Rutuno, USNM 166006; Río Villano, USNM 166002-4; Sarayacu, BMNH 1947.2.13.36-9, MCZ 26090, ZMB 10166.
GUYANA: Mazaruni-Potaro: Kartabo, AMNH 70971; Membaru River, upper Mazaruni River, UMMZ 85168; Oko Mountains, FMNH 26722-3. North West: Amakura River, Haulover, UMMZ 83558-9. Rupununi: Marudi River, AMNH 46233; Shudi-kar-wau, AMNH 49252. West Demerara: Dunoon, UMMZ 52449, 52508.
PERÚ: Junín: Chanchamayo, BMNH 1911.12.13.79-80. Loreto: Andoas, AMNH 79984-5; Cashiboya, AMNH 43454; San Antonio, Río Itaya, AMNH 43218. Puno: Yahuaramayo, BMNH 1913.2.25.7.
SURINAM: Suriname: Powakka, CM 44217.
SOUTH AMERICA: No specific locality, NHMW 6208.
BRASIL: Acre: Tarauacá, FMNH 83247. Amazonas: Rio Javarí, Benjamin Constant, CAS-SU 12620; Río Uaupés, north of Rio Japú, NHMG 489.
COLOMBIA: Amazonas: Gino-goje, lower Río Apoporis, MCZ 28038, 28040-2, 28044, USNM 152136-8.
ECUADOR: No specific locality, WCAB 35452-3; "Napo-Pastaza," USNM 166571. Napo: Avila, UMMZ 92093; south slope Cordillera del Dué, KU 123170; Lago Agrio, KU 125961-2 (skeletons), 126611-44, UMMZ 129326 (2); Limón Cocha, Río Napo, KU 99210-6, UIMNH 63087-9, 63098, 63106-9, 63118-9, 64802-4, 64858, 87998-9, 88001-30, 88437-8, 88580, 88604-5, 89852-97, 89999-90000; Loreto, CAS-SU 11439, WCAB 36526; Puerto Libre, Río Aguarico, KU 123190-1; Puerto Napo, UIMNH 55818-20; Río Cotapino, UMMZ 92094; Río Napo, UMMZ 92078; Santa Cecilia, AUM 8099, 8102, 8113-5, 8127-9, 8131, 8137, 8139-46, 8148, KU 105210-20, 109509-11, 111971, 122964-87, 123169, 126645. Pastaza: Canelos, BMNH 1947.2.13.42, KU 120915; Río Alpayacu, UMMZ 92079; Río Arajuno, USNM 166560-2, WCAB 40176; Río Oglán, USNM 16655203, 166558; Río Rutuno, USNM 166559; Río Shilcayacu, below Puyo, USNM 166557; Río Villano, USNM 166551.
FRENCH GUIANA: No specific locality, MNHN 4629. Inini: Lunier River, MNHN 98/217.
GUYANA: Mazaruni-Potaro: Kartabo, AMNH 70967-8, 70972, 70976. Rupununi: Shudi-kar-wau, AMNH 49255. West Demerara: Demerara Falls, BMNH 72.10.16.23, 72.10.16.37-8.
PERÚ: Loreto: Estirón, Río Ampiyacu, MZUSP 31033-4; Pebas, CAS-SU 3158, 3160; Roaboya, AMNH 43064.
[Pg 44] SURINAM: No specific locality, MCZ 2036, RMNH 11468. Marowijne: Camp 3, RMNH 13045-6; Wane Creek North, RMNH 11469-70. Saramacca: Right Coppename River, RMNH 11467.
BOLIVIA: El Beni: upper Río Beni, below mouth of Río Mapiri, MCZ 15565, UMMZ 57548, 67464-5; Rurrenbaque, UMMZ 57533.
PERÚ: Pasco: Yaupi, KU 136312.
BOLIVIA: El Beni: Ivón, BMNH 1967.2040; Reyes, UMMZ 57532. La Paz: San Ernesto, Mapiri District, BMNH 1901.8.2.54. Santa Cruz: Buenavista, AMNH 33951-2, 33958, BMNH 1927.8.1.19, 1927.8.1.118, FMNH 27091, UMMZ 63319-21, 63959(2), 63961(2), 66566(2), 66567, 66568(2), 66569 (2), 66570, 66571(2), 66575-6, 68196; Río Mamore, 2 km N Boca Chaparé, AMNH 79324; Sara, CM 3840-1; Surutu, CM 3814-5.
BRASIL: No specific locality: "Interior," BMNH 74.7.16.8-9. Acre: Plácido de Castro, MZUSP 6518; Tarauacá, WCAB 2496. Amazonas: Cucuí, NHMW 16495; Manacapurú, ZMB 28492, ZSM 278/1925; Manáus, MCZ 56281, NHMW 16492; Maués, AMNH 69623, 76177; Taracuá, NHMG 488, WCAB 18463-4. Mato Grosso: Mabuca, MZUSP 4272; Posto Coluene, Rio Xingú, WCAB 812; "Puerto Cabello," AMNH 3154; Tapirapé, AMNH 73647-62, CAS-SU 12351-6, MNHN 46/324. Pará: No specific locality, MPEG 623-6; Belém, KU 129866; Cachimbo, FMNH 175876, UIMNH 42149, WCAB 813; Cametá, NHMW 15892; Gurupá, BMNH 96.6.29.13; Ilha de Marajó, BMNH 1923.11.9.20-4; Ilha Mexicana, ZSM 111/1911, 112/1912; "Ponto Dois Indios," BMNH 1939.1.5.5; Santarém, BMNH 75.10.22.1-4, MCZ 354. Rondonia: Abuná, CAS 49773-4, FMNH 64239; Forte Principe da Beira, WCAB 10230; Igarapé Marmelo, FMNH 140254, KU 84725 (skeleton), 92243-6, 92247-8 (skeletons), WCAB 9997, 10001, 10003-4; Porto Velho, MZUSP 16343.
COLOMBIA: Amazonas: Gino-goje, lower Río Apoporis, USNM 152139; Leticia, USNM 152010-1; Raudal de la Playa, lower Río Apoporis, MCZ 28050; Río Apoporis, MCZ 28060. Boyacá: Sutatenza, USNM 152054-6. Cundinamarca: Medina, MCZ 16269-71, USNM 152089-90, 152092-7, 152757. Meta: El Mico, Río Guejar, USNM 152203; Río Duda, Sierra de Macarena, AMNH 79914; Río Guapaya, Sierra de Macarena, FMNH 81332; Río Guaviari, Casa de Piedra, UTA No number. Putumayo: Río Mecaya, FMNH 69711-4, 69716. Vaupés: Gomogoje, lower Río Apoporis, MCZ 28048.
ECUADOR: No specific locality, WCAB 35451, 35785; "Oriente," UMMZ 90418. Carchí: below Salinas, USNM 166059. Esmeraldas: Carondelet, UIMNH 53560-9; Lagartera, Río Caoni, UIMNH 53441, 53458-79. Morona-Santiago: Macuma, UIMNH 63142-3, 63145, 63147, 63151, 63154, 63157, USNM 166060. Napo: Avila, UMMZ 92077; Cuyabeno, UIMNH 63158, 90111; Lago Agrio, KU 126647-55; Limón Cocha, Río Napo, AUM 8132-4, KU 99207-8, 99421-3, 99424 (skeleton), 99425, UIMNH 64801, 87798, 87800, 88032-5, 88576, 90066, 90082, 90102, 90104, 90314, 90984; Loreto, WCAB 35352; Río Cotapino, UMMZ 92080; Río Napo, UMMZ 84120; San José Abajo, AMNH 1295, 1449, 22180, 79990; Santa Cecilia, AUM 8117, 8150, KU 105230-3; south slope Volcán Sumaco, USNM 166570. Pastaza: No specific locality, ZSM 31/1956; Arajuno, USNM 165995; Bufeo, lower Río Bobonaza, USNM 166046-8; Canelos, BMNH 80.12.5.179, 1947.2.13.48, UMMZ 89066; [Pg 45] Don Tomás, Río Bobonaza, USNM 166049-50; Montalvo, CAS-SU 10320, USNM 165987-9, 166058, 166566; 2.5 km SE Puyo, USNM 166051; Río Arajuno, USNM 166043-5; Río Arajuno (headwaters), USNM 166053; Río Bobonaza, WCAB 3613-4, 35504; Río Capahuari, USNM 165990, 166555-6; Río Capahuari (headwaters), USNM 166057; Río Conambo, USNM 166569, ZSM 28/1962, 35/1962; Río Conambo at Río Ollaguanga, USNM 166568; Río Conambo at Río Shiona-yacu, USNM 166019, 166563-5; Río Corrientes, USNM 195994, 166020-38, WCAB 3841-2; Río Huiyo-yacu, Pico de Conambo, USNM 166052; Río Pastaza, MCZ 19697; Río Pastaza (drainage), NHRM 1966, USNM 165996; Río Pindo, USNM 166039-41; Río Pindo at Río Tigre (village), USNM 165992-3, 166042; Río Pucayacu, USNM 166054, 166056; Río Rutuno, USNM 166055; Río Solis, upper Río Bobonaza, WCAB 39914; Río Villano, USNM 165991, 166567; Sarayacu, BMNH 80.12.5.213, 80.12.5.239-40, MZUSP 323; Shell Mera, KU 99420. Zamora-Chinchipe: "Yani-Inzari," AMNH 43259, 43394; Zamora, AMNH 78928.
FRENCH GUIANA: Cayenne: Crique Grégoire, UP 40; Maripa, Oyapok River, UP 72; Oyapok River, UZM 1473. Inini: Crique Gabrielle, UP 118-20.
GUYANA: No specific locality: RMNH 1873(3), ZMB 3102(2). East Demerara: Atkinson Field, ASU 11622. Mazaruni-Potaro: Chinapora River, upper Potaro River, BMNH 1905.11.1.20-1; Kamakusa, AMNH 21416, 21418-9, 21422; Kartabo, AMNH 11689, 11691, 11697-9, 11703, 11706-8, 23107, 39730, 70966, 70969-70, 70973-5, USNM 118057; Moraballi Creek, Essequibo River, BMNH 1930.10.10.47-51; Oko Mountains, FMNH 26692-705; upper Potaro River, Tung District, BMNH 1905.11.1.40; Rockstone, FMNH 26591. North West: Amakura River, Haulover, UMMZ 83735. Rupununi: north of Acaray River, west of New River, KU 69747-8; Kuyuwini Landing, AMNH 46283; Pakaraima Mountains, BMNH 1933.6.19.49; Shudi-kar-wau, AMNH 10665, 39637, 49256(2). West Demerara: Demerara, CAS 54773-4; Demerara Falls, BMNH 72.10.16.16-22, 72.10.16.25-32; Dunoon, MCZ 4834, UMMZ 46736, 52493-4, 52502, 52504-5, 57271; Vryheid, BMNH 78.12.13.18.
PERÚ: Amazonas: Río Cenepa, AMNH 43400. Huanuco: Monte Alegre, Río Pachitea, AMNH 43014, 43019. Loreto: Achinamisa, Río Huallaga, AMNH 42178, 42502; Andoas, Río Pastaza, AMNH 79991; Cashiboya, AMNH 43388, 43453; Estirón, Río Ampiyacu, CAS 93264-74, 93276, 93278-9, 93281, 93283-6, 93289, 93311, 93327; Igarapé Champuia, upper Río Curiuja, MZUSP 10339; Iquitos, AMNH 42204, 42442, 43468, NHMW 6118; Lago de Miraño, mouth of Río Napo, AMNH 42712, 43186; Nauta, ANSP 11399; Ollanta, AMNH 42865; Pampa Hermosa, Río Cushabatay, AMNH 43124, 43146; Pebas, CAS-SU 6375; Pucallpa, MJP 101(2), 140(3); Punga, Río Tapiche, AMNH 43194; "Rancho de Indiana, Iquitos District," MVZ 16890; upper Río Abujao, AMNH 42908; Río Itaya, AMNH 42755; upper Río Pisqui, AMNH 43536; Río Tapiche at Río Contaya, AMNH 42983; Río Utoquinia at Brasilian frontier, AMNH 43137; Sobral, Río Tamaya, AMNH 43242; Yurimaguas, BMNH 84.2.18.50. San Martín: Cainarachi, AMNH 42763; Moyobamba, ZSM 19/1914.
SURINAM: No specific locality, BMNH 70.3.10.67, NHMW 18433.3. Brokopondo: Afobaka, RMNH 16536; Brownsweg, RMNH 16537; Railway km. 121, RMNH 16534. Marowijne: Djai Creek, RMNH 16513-4; Maroni River, ZMB 8240, 8531; Nassaugebergte, RMNH 16517-33; Paloemeu, USNM 159025; Swamp Camp, RMNH 16515. Nickerie: Sipaliwini, RMNH 16538. Saramacca: Left Coppename River, RMNH 16535; Tibiti, RMNH 16516. [Pg 46] Suriname: Berlijn, RMNH 15064; Powakka, CM 44226; Zanderij, CM 50568.
VENEZUELA: Amazonas: Cerro Duida, UPR-M 2875; Cerro Marahuaca, UPR-M 114-5; Esmeralda, AMNH 23174; Iniridi, SMF 2640; La Culebra, MCZ 28572, UPR-M 117; Laguna, between Tama Tama and Esmeralda, UPR-M 2760; Río Pescado, AMNH 23177; Tapara, UPR-M 113. Distrito Federal: Caracas, BMNH 51.7.17.182.
COLOMBIA: Huila: Acevedo, Río Suaza, FMNH 69709-10.
ECUADOR: No specific locality, ZMB 16589. Napo: Avila, UMMZ 90413; south slope Cordillera del Dué, KU 123176-88, 123189 (skeleton), 124208 (eggs), 124209-11 (tadpoles); L'Alegria, USNM 167472-3; Río Pacayacu, tributary of Río Cotapino, CAS-SU 13150; southeast slope Volcán Sumaco, CAS-SU 11442. Pastaza: Abitagua, CAS-SU 5067, FMNH 25791, 27619, UMMZ 90414, 92092; Alpayaca, Río Pastaza, BMNH 1912.11.1.64; Mera, UMMZ 90412(4). Tungurahua: Baños, NHRM 1960.
PERÚ: Ayacucho: La Mar, Sivia, Río Apurimac, FMNH 39853. Huanuco: Río Pachitea, midway between Puerto Victoria and Puerto Inca, CAS-SU 17745. Junín: Satipo, MJP 38.
Amazon by Prof. Orton. Proc. Acad. Nat. Sci. Philadelphia, 25:120-137.
The University of Kansas Publications, Museum of Natural History, beginning with volume 1 in 1946, was discontinued with volume 20 in 1971. Shorter research papers formerly published in the above series are now published as Occasional Papers, Museum of Natural History. The Miscellaneous Publications, Museum of Natural History, began with number 1 in 1946. Longer research papers are published in that series. Monographs of the Museum of Natural History were initiated in 1970.
Institutional libraries interested in exchanging publications may obtain the Occasional Papers and Miscellaneous Publications by addressing the Exchange Librarian, University of Kansas Library, Lawrence, Kansas 66044. Individuals may purchase separate numbers of all series. Prices may be obtained upon request addressed to Publications Secretary, Museum of Natural History, University of Kansas, Lawrence, Kansas 66044.
Except for the list of corrections below and minor corrections not listed, the text presented here is that of the original printed version.
| Page | Correction | |
| 5 | is ⇒ in | |
| 5 | buckley ⇒ buckleyi | |
| 17 | scaral ⇒ sacral | |
| 19 | Provicia ⇒ Provincia | |
| 25 | in ⇒ is | |
| 25 | metalic ⇒ metallic | |
| 41 | bromeiad ⇒ bromeliad |