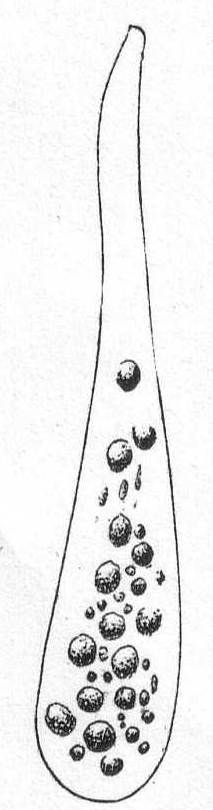
Fig. 1.—
Amœba guttala.
ENLARGE
This eBook is for the use of anyone anywhere at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org
Title: Marine Protozoa from Woods Hole
Bulletin of the United States Fish Commission 21:415-468, 1901
Author: Gary N. Calkins
Release Date: May 5, 2006 [eBook #18320]
Language: English
Character set encoding: UTF-8
***START OF THE PROJECT GUTENBERG EBOOK MARINE PROTOZOA FROM WOODS HOLE***
Comparatively little has been done in this country upon marine Protozoa. A few observations have been made by Kellicott, Stokes, and Peck, but these have not been at all complete. With the exception of Miss Stevens's excellent description of species of Lichnophora I am aware of no single papers on individual forms. Peck ('93 and '95) clearly stated the economic position of marine Protozoa as sources of food, and I need not add to his arguments. It is of interest to know the actual species of various groups in any locality and to compare them with European forms. The present contribution is only the beginning of a series upon the marine Protozoa at Woods Hole, and the species here enumerated are those which were found with the algæ along the edge of the floating wharf in front of the Fish Commission building and within a space of about 20 feet. Many of them were observed in the water and algæ taken fresh from the sea; others were found only after the water had been allowed to stand for a few days in the laboratory. The tow-net was not used, the free surface Protozoa were not studied, nor was the dredge called into play. Both of these means of collecting promise excellent results, and at some future time I hope to take advantage of them.
My observations cover a period of two months, from the 1st of July to the 1st of September. During that time I was able to study and describe 72 species representing 55 genera, all from the limited space mentioned above. In addition to these there are a few genera and species upon which I have insufficient notes, and these I shall reserve until opportunity comes to study them further.
I take this opportunity to express my thanks to Dr. Hugh M. Smith for many favors shown me while at Woods Hole.
In dealing with these marine forms from the systematic standpoint, two courses are open to the investigator. He may make numerous new species based upon minor differences in structure, or he may extend previous descriptions until they are elastic enough to cover the variations. The great majority of marine protozoa have been described from European waters, and the descriptions are usually not elastic enough to embrace the forms found at Woods Hole. I have chosen, however, to hold to the conservative plan of systematic work, and to make as few new species as possible, extending the older descriptions to include the new forms.
The different classes of Protozoa, and orders within the classes, are distributed more or less in zones. Thus the Infusoria, including the Ciliata and the Suctoria, are usually littoral in their habitat, living upon the shore-dwelling, or attached, water plants and upon the animals frequenting them. It is to be expected, therefore, that in forms here considered there should be a preponderance of Infusoria. Flagellated forms are also found in similar localities, but on the Surface of the sea as well; hence the number described in these pages is probably only a small proportion of the total number of Mastigophora in this region. The Sarcodina, including the Foraminifera and the Radiolaria, are typically deep-sea forms and would not be represented by many types in the restricted locality examined at Woods Hole. Two species, Gromia lagenoides and Truncatulina lobatula, alone represent the great order of Foraminifera, while the still larger group of Radiolaria is not represented at all.
The Protozoa described are distributed among the different orders as follows:*
The pseudopodia are lobose, sometimes absent, the body then progressing by a flowing movement; the body consists of ectoplasm and endoplasm, the latter being granular and internal, the former hyaline and external. There is always one nucleus and one vacuole, but both may be more numerous. Reproduction takes place by division or by spore-formation. Fresh-water and marine.
|
Fig. 1.— Amœba guttala. ENLARGE |
Amœba guttala Duj. Fig. 1.
A minute form without pseudopodial processes, extremely hyaline in appearance, and characterized by rapid flowing in one direction. The body is club-shape and moves with the swollen end in advance. A comparatively small number of large granules are found in the swollen portion, while the smaller posterior end is quite hyaline. Contractile vacuole absent, and a nucleus was not seen. Frequent in decomposing vegetable matter. Length 37µ. Traverses a distance of 160µ in one minute.
The fresh-water form of A. guttula has a vacuole, otherwise Dujardin's description agrees perfectly with the Woods Hole forms.
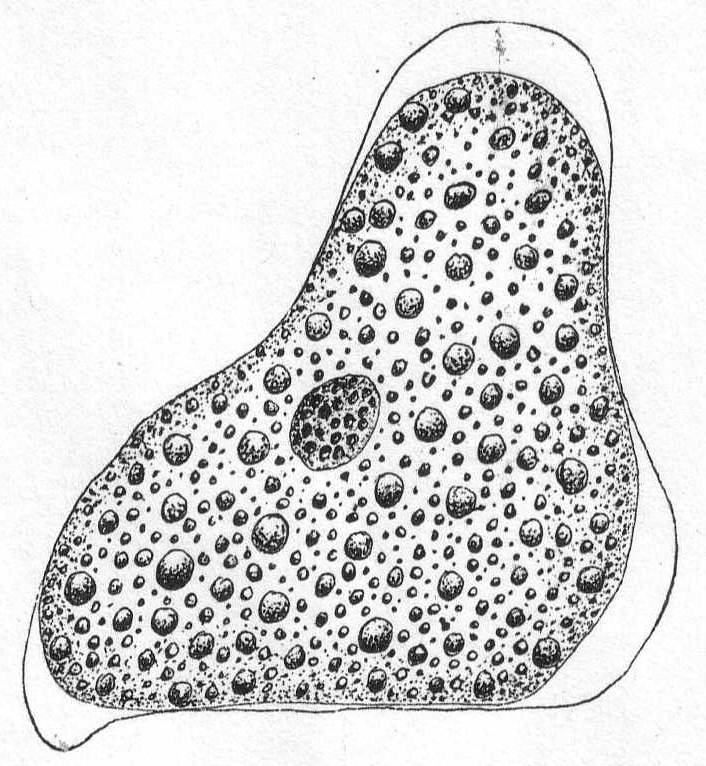
Fig. 2.—Amœba sp. ENLARGE |
Amœba ? Fig. 2.
A more sluggish form than the preceding, distinguished by its larger size, its dense granulation, and by short, rounded pseudopodia, which, as in Amœba proteus, may come from any part of the body. A delicate layer of ectoplasm surrounds the granular endoplasm, and pseudopodia formation is eruptive, beginning with the accumulation of ectoplasm. Movement rapid, usually in one direction, but may be backwards or sideways, etc. Contractile vacuole absent; the nucleus is spherical and contains many large chromatin granules. Length 80µ; diameter 56µ.
Synonym: Pachymyxa hystrix Gruber.
Marine rhizopods, globular or irregular in form, and slow to change shape. Dimorphic. Both forms multinucleate during vegetative life. Pseudopodia are long, thin, and thread-form, with rounded ends. Their function is neither food-getting nor locomotion, but probably tasting. The plasm of both forms is inclosed in a soft gelatinous membrane. In one form the jelly is impregnated with needles of magnesium carbonate (Schaudinn), but these are absent in the other form. The membrane is perforated by clearly defined and permanent holes for the exit of the pseudopodia. Reproduction occurs by division, by budding or by fragmentation, but the parts are invariably multinucleate. At the end of vegetative life the needle-bearing form fragments into numerous mononucleate parts; these develop into adults similar to the parent, but without the spines. At the end of its vegetative life this new individual fragments into biflagellated swarm-spores which may conjugate, reproducing the form with needles. Size up to 2 mm.
Trichosphærium sieboldi Schneider. Fig. 3.
With the characters of the genus. A form which I have taken to be a young stage of this interesting rhizopod is described as follows:
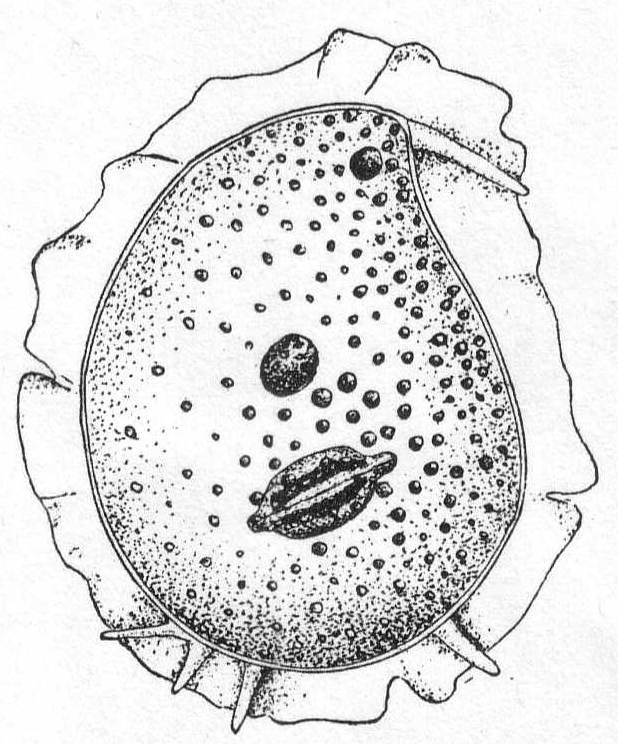
Fig. 3.— Trichosphærium sieboldi. ENLARGE |
A minute, almost quiescent, form which changes its contour very slowly. The membrane is cap-like and extends over the dome-shaped body, fitting the latter closely. The endoplasm is granular and contains foreign food-bodies. Nucleus single, spherical, and centrally located. Pseudopodia short and finger-form, emerging from the edge of the mantle-opening and swaying slowly from side to side or quiescent. The most characteristic feature is the presence of a broad, creeping sole, membranous in nature and hyaline in appearance. This membrane is the only evidence of ectoplasm, and it frequently shows folds and wrinkles, while its contour slowly changes with movements of body. The pseudopodia emerge from the body between this membrane and the shell margin. Contractile vacuole absent. Length 42µ, width 35µ. In decomposing seaweeds, etc.
Only one specimen of this interesting form was seen, and I hesitate somewhat in placing it on such a meager basis. It is so peculiar, however, that attention should be called to it in the hope of getting further light upon its structure and mode of life. Its membranous disk recalls the genus Plakopus; its mononucleate condition, its membranous disk, and the short, sometimes branched, pseudopodia make it difficult to identify with any phase in the life-history of Trichosphærium. I shall leave it here provisionally, with the hope that it may be found more abundantly another time.
The form is ovoid or globular, and the body is covered by a tightly fitting, plastic, chitin shell, which, in turn, is covered by a fine layer of protoplasm. The flexibility of the shell makes the form variable as in the amœboid types. The thickness of the shell is quite variable. The pseudopodial opening is single and terminal. The pseudopodia are very fine, reticulate, granular, and sharply pointed, and form a loose network outside of the shell opening. Nucleus single or multiple. Contractile vacuole is usually absent. Fresh and salt water.
Gromia lagenoides Gruber '84. Fig. 4.
This species is not uncommon about Woods Hole, where it is found upon the branches of various types of algæ. The body is pyriform, with the shell opening at the larger end. The chitinous shell is hyaline and plastic to a slight extent, so that the body is capable of some change in shape. The shell is thin and turned inwards at the mouth-opening, forming a tube (seen in optical section in fig. 4) through which the protoplasm passes to the outside. The walls of this tube are thicker than the rest of the shell, and in optical section the effect is that of two hyaline bars extending into the body protoplasm. A thin layer of protoplasm surrounds the shell and fine, branching, pseudopodia are given off in every direction. The protoplasm becomes massed outside of the mouth-opening and from here a dense network of pseudopodia forms a trap for diatoms and smaller Protozoa. The nucleus is spherical and contains one or two large karyosomes. The protoplasm is densely and evenly granular, without regional differentiation. I have never observed an external layer of foreign particles, such as Gruber described in the original species.
Length of shell 245µ; largest diameter 125µ.
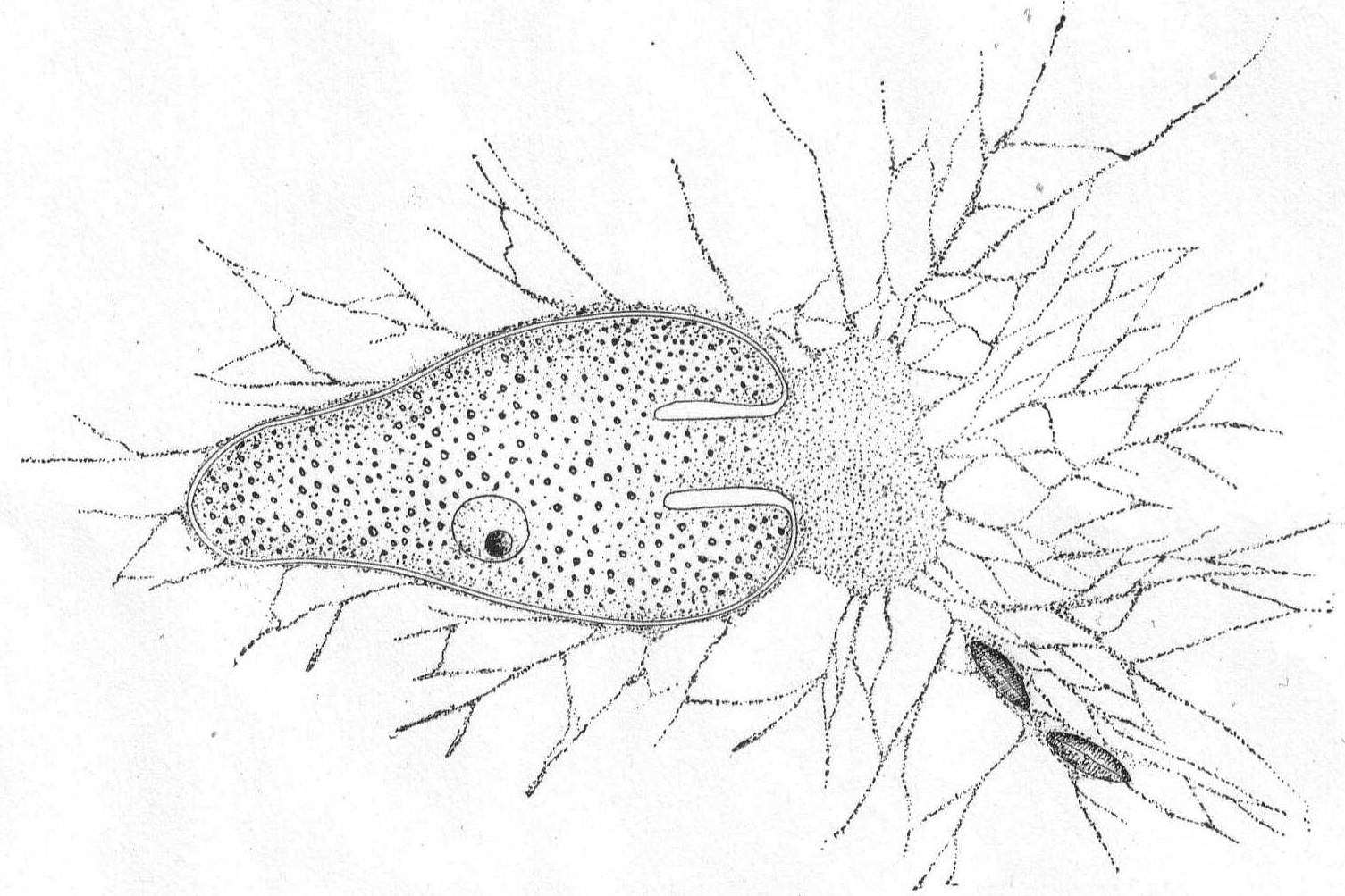
Fig. 4.—Gromia lagenoides. ENLARGE |
A group of extremely variable foraminifera in which the shell is rotaline; i. e., involute on the lower side and revolute on the upper (Brady). The shell is calcareous and coarsely porous in older forms. The characters are very inconstant, and Brady gives up the attempt to distinguish the group by precise and constant characters.
Truncatulina lobatula Walker & Jacob.
Synonyms: See Brady '84 for a long list.
"It is impossible to define by any precise characters the morphological range of the present species. Its variations are infinite." (Brady, p. 660.)
This very common form, which occurs in all latitudes, was found frequently among the algæ at Woods Hole. Its characters are so difficult to define that for the present I shall limit my record to this brief notice. Size of shell 230µ by 270µ.
The body is spherical and differentiated into granular endoplasm and vacuolated ectoplasm, but the zones are not definitely separated. There is one central nucleus and usually one contractile vacuole. The pseudopodia have axial filaments that can be traced to the periphery of the nucleus. Fresh and salt water.
Actinophrys sol Ehr., variety. Fig. 5.
Synonyms: See Schaudinn '95.
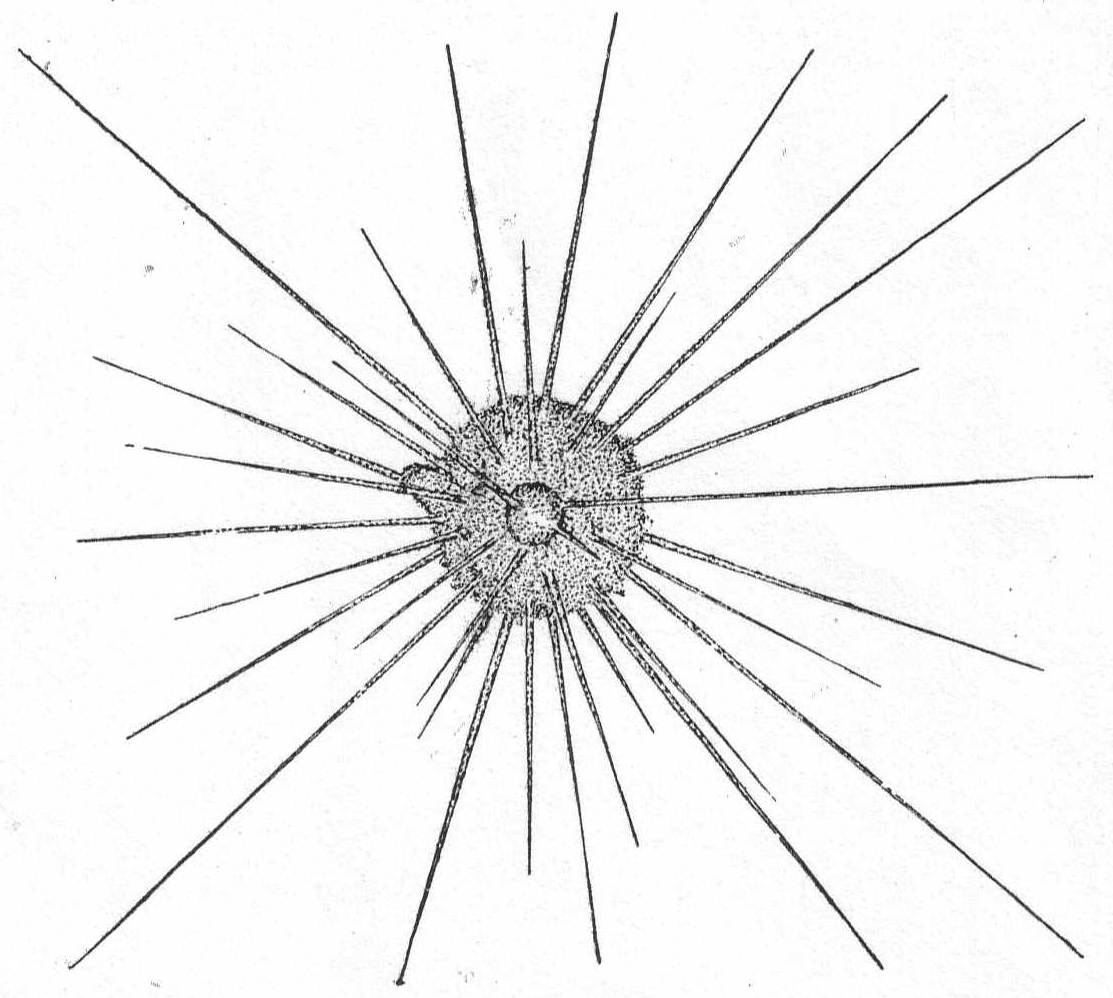
Fig. 5.—Actinophrys sol. ENLARGE |
The diameter is about 50µ; the vacuolated ectoplasm passes gradually into the granular endoplasm. This is the characterization given A. sol by Schaudinn, and it applies perfectly to the freshwater forms. If I am correct, however, in placing an Actinophrys-like form found at Woods Hole in this species, the description will have to be somewhat modified. In this form (fig. 5) there is no distinction between ectoplasm and endoplasm, and there is an entire absence of vacuoles. The nucleus is central, and axial filaments were not seen. The single specimen that I found looked much like a Suctorian of the genus Sphærophrya, but the absence of a firm cuticle and the presence of food-taking pseudopodia with granule-streaming makes it a very questionable Suctorian, and 1 place it here until further study throws more light upon it.
Diameter of body 40µ; length of pseudopodia 120 to 140µ.
The body is globular with but slight differentiation into ectoplasm and endoplasm; one nucleus in the latter; contractile vacuoles one or many; pseudopodia on all sides, thin, and with peripheral granule-streaming; surrounded by a globular, rather thick coat of jelly, which is hyaline inside and granular on the periphery. Fresh and salt water.
Heterophrys myriapoda Archer. Fig. 6.
Synonym: H. marina Hert. & Less. '74.
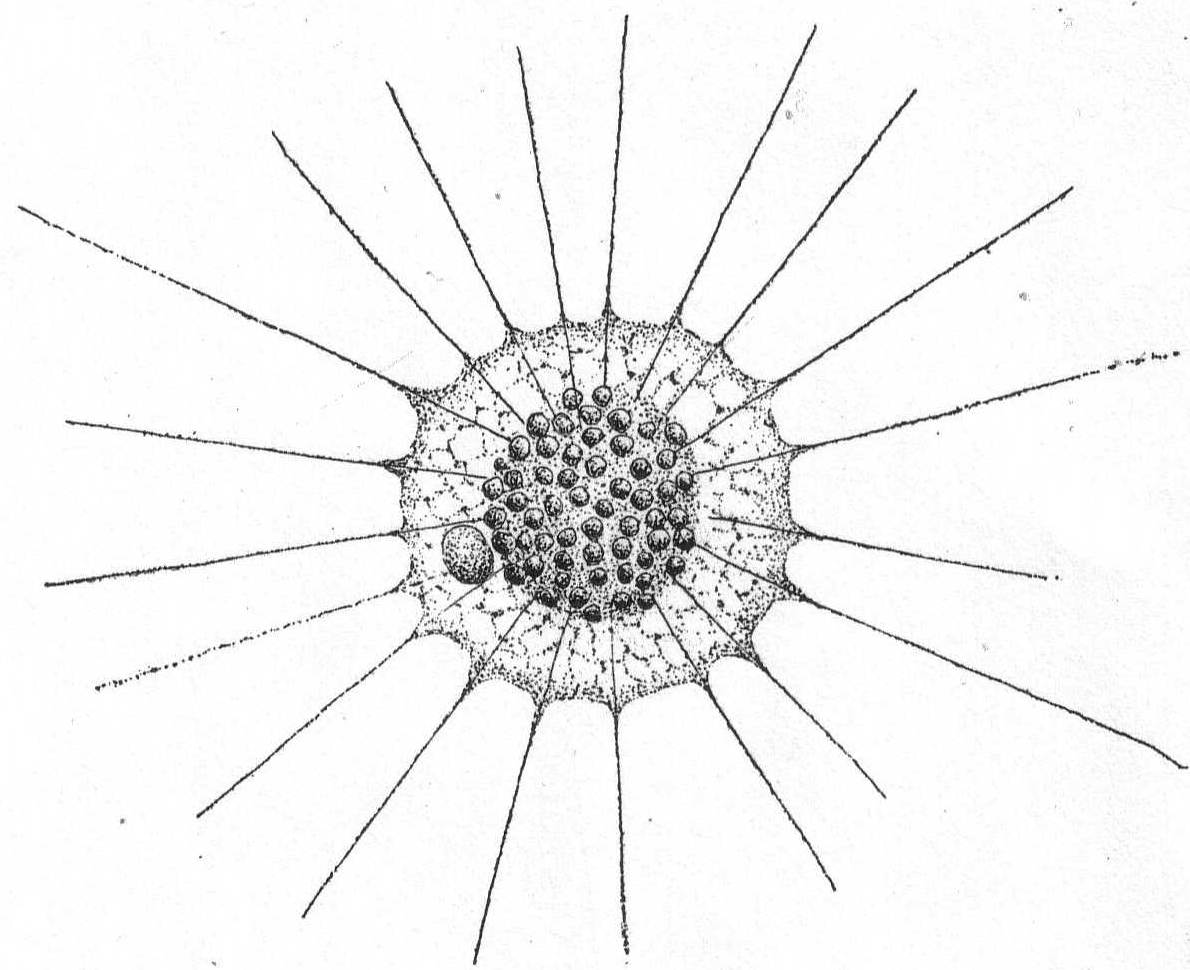
Fig. 6.—Heterophrys myriapoda. ENLARGE |
Diameter 25 to 80µ; pseudopodia twice as long as the body diameter; the plasm often contains chlorophyll bodies (Zoochlorella). The granular part of the gelatinous layer is thick (up to 10µ). The spine-like processes are very thin and short. (Schaudinn '95.) The marine form found at Woods Hole probably belongs to this species, as described by Schaudinn. The short pseudopodia which give to the periphery a fringed appearance are quite regularly placed in connection with the pseudopodia. The latter are not so long as twice the body diameter, the longest being not more than equal to the diameter of the sphere. The body inside of the gelatinous covering is thickly coated with bright yellow cells similar to those on Radiolaria. The animal moves slowly along with a rolling motion similar to that described by Pènard '90, in the case of Acanthocystis. Diameter of entire globe 35µ; of the body without the jelly 18µ. The extremely fine granular pseudopodia are 8 to 35µ long. Common among algæ.
This form was probably meant by Peck '95, when be figured "a heliozoön."
| Key to orders of Flagellidia. | |
| Small, body usually amœboid; 1 or more flagella; no mouth | Order Monadida. |
| Small; plasmic collar around the flagellum | Order Choanoflagellida. |
| With 2 or more flagella; one trails behind | Order Heteromastigida. |
| With 3 or more flagella, none of which trails | Order Polymastigida. |
| Large; firm body wall; 1 or 2 flagella; mouth or pharynx, or both | Order Euglenida. |
| Medium size; with chlorophyll, no mouth, usually colonial | Order Phytoflagellida. |
| Small; silicious skeleton; parasitic on Radiolaria or free |
Order Silicoflagellida. (One genus, Distephanus Stöhr) |
| Key to the families of Monadida. | |
| No mouth; 1 or 2 flagella: amœboid with lobose or ray-like pseudopodia | Family Rhizomastigidæ |
| Mouth at base of single flagellum; plastic; no pseudopodia | Family Cercomonadidæ |
| One flagellum; inclosed in gelatinous or membranous cups | Family Codonœcidæ |
| One flagellum; tentacle like process at base of flagellum; inclosed in cup | Family Bikœcidæ |
| One main flagellum and 1 or 2 accessory flagella | Family Heteromonadidæ |
| Key to marine genera of Monadida.* | ||
| Family Rhizomastigidæ: | ||
| 1. | Flagellum repeatedly thrown off and reassumed | Genus *Mastigamœba in part |
| 2. | Flagellum never thrown off | 3 |
| 3. | a. Pseudopodia lobose | Genus Mastigamœba |
| b. Pseudopodia ray-like | Genus Mastigophrys | |
| Family Codonœcidæ: | ||
| 1. | Goblet-shaped cups adherent by stalk | Genus *Codonœca |
| Family Heteromonadidæ: | ||
| 1. | The long flagellum vibratory | Genus *Monas |
| 2. | The long flagellum rigid; shorter one vibrates | Genus Sterromonas |
| * Presence at Woods Hole indicated by asterisk. | ||
| Key to marine genera of Choanoflagellida. | |||||
| 1. | Without gelatinous or membranous test | 3 | |||
| 2. | With gelatinous or membranous test | 4 | |||
| 3. | a. | Attached forms: | |||
| 1. | Without a stalk, or with a very short one | Genus *Monosiga | |||
| 2. | With a long, simple stalk | Genus *Codonosiga | |||
| 3. | With a long, branched, stalk | Genus Codonocladium | |||
| b. | Free-swimming | Genus Desmarella | |||
| 4. | Colonial, and with a gelatinous covering | Genus Proterospongia | |||
| * Presence at Woods Hole indicated by asterisk. | |||||
| Key to families and marine genera of Heteromastigida. | ||
| 1. | Two flagella nearly equal in size | Family Bodonidæ |
| One main and 2 accessory flagella | Family Trimastigidæ | |
| Family Bodonidæ: | ||
| 1. | Body very plastic, almost amœboid | Genus *Bodo |
| Body not plastic; with large anterior cavity, holding flagella | Genus *Oxyrrhis | |
| Family Trimastigidæ: | ||
| 1. | With an undulatory membrane between accessory flagella | Genus Trimastix |
| Without such membrane; flagella contained in a ventral groove while at rest | Genus Costia | |
| * Presence at Woods Hole indicated by asterisk. | ||
| Key to marine genera of Polymastigida. | ||
| 1. | Body flattened; ends rounded; sides hollowed; often with wing-like processes; cross section S-shaped | Genus Trepomonas |
| 2. | Body pyriform; one large asymmetrical groove; 4 flagella | Genus Tetramitus |
| 3. | Body spherical; many flagella equally distributed | Genus Multicilia |
| Key to families and marine genera of Euglenida. | ||
| 1. | With deeply-insunk pharynx; no mouth | 2 |
| With pharynx and distinct mouth | Family Peranemidæ | |
| 2 | Body plastic; usually with chromatophores and eye-spot | Family Euglenidæ |
| Body plastic; no chromatophores; no eye-spot | Family Astastidæ | |
| Family Euglenidæ: | ||
| Body Euglena-like, inclosed in shell with round opening for exit of flagellum | Genus Trachelomonas | |
| Family Astastidæ: | ||
| Body with one flagellum | Genus *Astasia | |
| Family Peranemidæ: | ||
| 1. | Body striped; plastic; two diverse flagella | Genus Heteronema |
| 2. | Body striped; not plastic; posterior flagellum longer than the other | Genus *Anisonema |
| 3. | Body striped; not plastic; with rod-like organ in pharynx | Genus Entosiphon |
| * Presence at Woods Hole indicated by asterisk. | ||
In general the form is oval and either regular in outline or irregular through the presence of many pseudopodia. One flagellum usually quite large and distinct. Differentiation of ectoplasm and endoplasm distinct or wanting. One to several contractile vacuoles. The pseudopodia are occasionally withdrawn, and the flagellum is the sole means of locomotion. In some cases the flagellum turns into a pseudopodium, and, conversely, the pseudopodium at one end may become a flagellum (see below). In some rare cases the ectoplasm secretes a gelatinous mantle. Reproduction not observed.
Fresh and salt water.
Mastigamœba simplex, n. sp. Fig. 7.
A very small form, first seen in the flagellated stage, aroused my interest by reason of the fact that its flagellum lost its regular outline and became amœboid, turning to a pseudopodium, while at the same time other pseudopodia were protruded from different parts of the periphery. In this condition ectoplasm and endoplasm could be made out with the clearest definition. After the pseudopodia were well formed, the body became flat and closely attached to the glass slide. In a short time one of the pseudopodia became longer than the rest; the body became more swollen; the pseudopodia were gradually drawn in, with the exception of the more elongate one; this became active in movement and finer in diameter, until ultimately it formed a single flagellum at the anterior of a small monadiform flagellate. The process was repeated two or three times under my observation, so that I am convinced that it was not a developmental form of some rhizopod. Several of them were seen at different times during the summer, and they were always of the same size and form in the flagellated or amœboid condition. I did not make out their reproduction, and I shall not be satisfied that this is a good species until their life history is known.
In decaying algæ. Length 10µ.
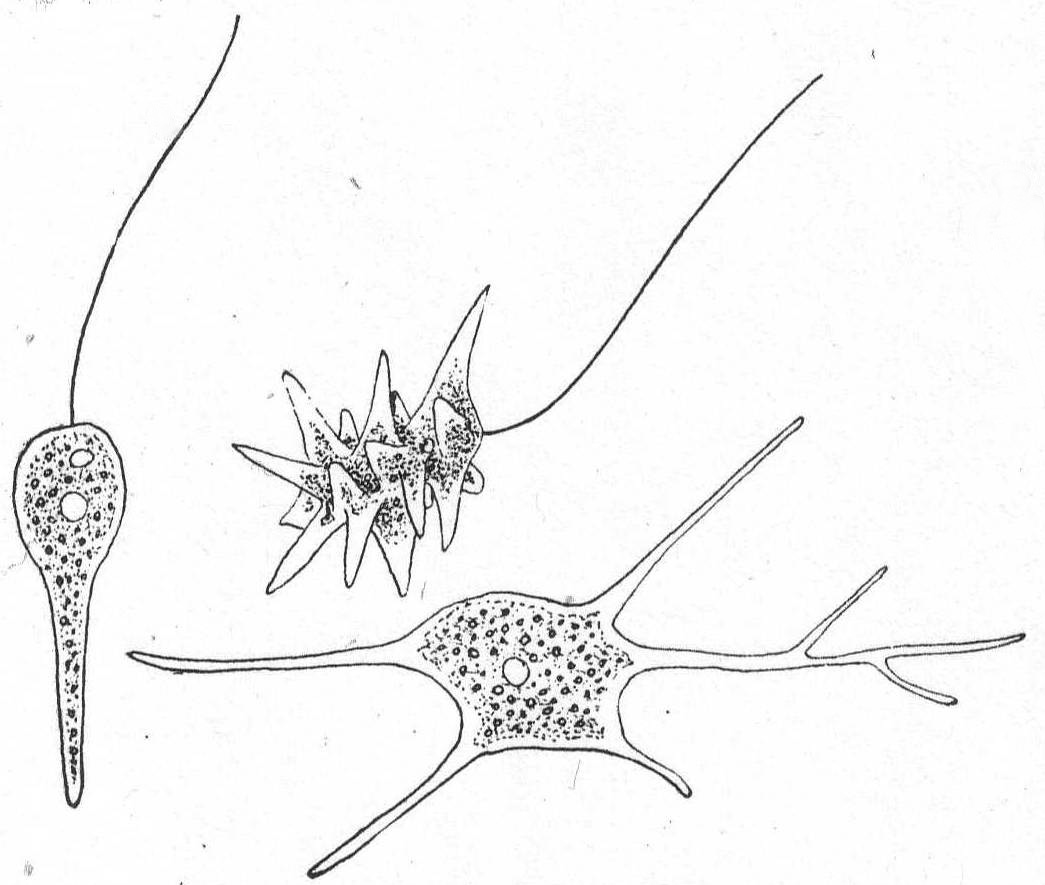
Fig. 7.—Mastigamœba simplex. ENLARGE |
(Kent '81.)
Small forms inclosed in cup or "house" of ovoid or goblet shape, colorless and probably gelatinous (chitin?) in texture, and borne upon a stalk. The monad does not completely fill the test. Contractile vacuole single, posterior.
Codonœca gracilis, n. sp. Fig. 8.
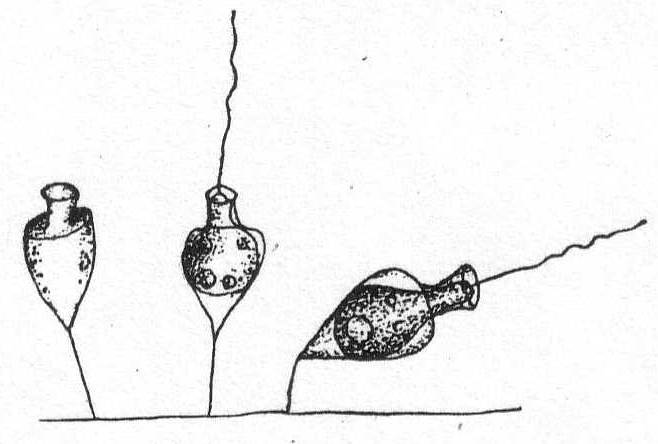
Fig. 8.— Codonœcea gracilis. ENLARGE |
The cup is urn-shaped with a well-defined neck or collar borne upon a shoulder-like end of the body. It is hyaline, colorless, and carried upon a stalk equal in length to the cup or shorter than this. The animal does not fill the cup, nor is it attached by a filament to the latter. There is a single flagellum. The nucleus is minute and lateral in position; the contractile vacuole is in the posterior end of the body. Total length of cup and stalk 21µ; of cup alone 12µ. This minute form looked so much like a choanoflagellate that I supposed it to be one until I discovered an empty case (Fig. 8).
The body is small, globular or oval and either free-swimming or fastened by one of the two flagella. The body is sometimes a little amœboid, with short pseudopodial processes. In addition to the main flagellum, there are usually one or two small flagella at the basis of the larger one. The nucleus is usually anterior, and one or two contractile vacuoles are present.
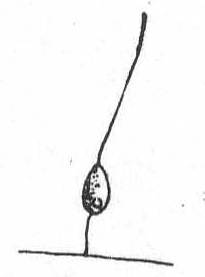 Fig. 9.—
Fig. 9.—Monas sp. ENLARGE |
Monas sp. Fig. 9.
An extremely small form (3µ) attached by a thread of protoplasm—perhaps a flagellum, to algæ. The body is ovoid and the main flagellum is about four times the length of the body. The contractile vacuole is posterior. Only one specimen was seen and upon this I shall not attempt to name the species.
Small colorless forms of Choanoflagellida, always naked and solitary. The posterior end is attached directly to the substratum, or there is a short stalk not exceeding the body in length. Kent '81 distinguished nine species, but Bütschli questioned the accuracy of many of these, and in this he was followed by Francé '97, who recognized three species—Monosiga ovata, M. fusiformis, and M. augustata. Fresh and salt water.
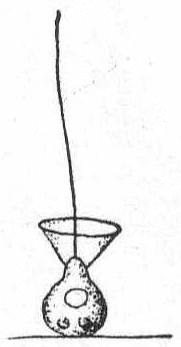 Fig. 10.—
Fig. 10.—Monosiga ovata. ENLARGE |
Monosiga ovata S. Kent '81. Fig. 10.
Synonyms: M. brevipes S. K.; M. consociata S. K.; M. limnobia Stokes.
The individuals are unstalked or provided with a very short stalk less than the body in length. The form is spherical or ovate, broadest at the base and tapering to the extremity. The collar is somewhat variable in size. In the Woods Hole forms it was about the length of the body. Oil particles present. Contractile vacuole posterior, nucleus anterior.
Fresh and salt water. Length of body without the collar 5µ.
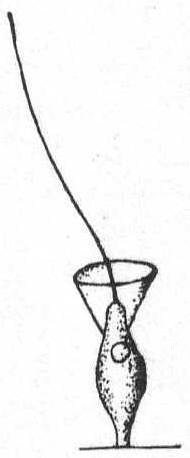
Fig. 11.— Monosiga fusiformis. ENLARGE |
Monosiga fusiformis S. K. Fig. 11.
Synonyms: M. steinii S. K.; M. longicollis S. K.
The individuals are unstalked, minute, and of a general flask-shape. The body is swollen centrally and tapers slightly at each end. There is no stalk, the body being fixed by the attenuate posterior end. There are two contractile vacuoles and one nucleus, which is situated a little above the body center. Fresh and salt water. Length without collar 9µ; length of collar 3µ.
This genus, as modified by Francé, is distinguished from the preceding by the possession of an unbranched stalk much longer than the body length. The body is naked and of various shapes, and the individuals are solitary or colonial upon a single stalk. Kent '81 enumerates no less than 10 species, which were cut down by Bütschli to 1. Francé admits 4—C. botrytis Jas. Clark; C. grossularia; C. pyriformis, and C. furcata, all S. Kent—but regards the second and third as merely form varieties of the first.
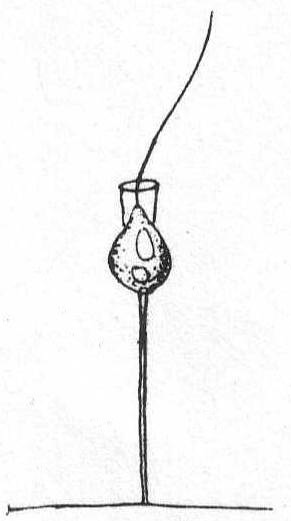 Fig. 12.—
Fig. 12.—Codonosiga botrytis. ENLARGE |
Codonosiga botrytis (Ehr. sp.) Jas. Clark '67. Fig. 12.
Francé gives the following synonyms: Epistylia botrytis Ehr.; E. digitalis Stein, Zoothamnium parasitica Stein; Anthophysa solitaria Fresenius; Codonosiga pulcherrima Jas. Clark; Monosiga gracilis S. Kent; M. globulosa S. Kent; Codonosiga pyriformis Kent; C. grossularia Kent; (Francé).
The individuals are small and provided with a long unbranched, or terminal, simply split stalk. The individuals are single or colonial. The Woods Hole form measured 22µ over all; the body was 5µ, the collar 3µ, and the stalk 14µ. No colonies were seen, and only a few individuals upon red algæ.
The body is naked, usually amœboid in its changes, and provided with two flagella, one of which is usually trailed along under and behind the animal. The anterior end is usually pointed, with the flagella arising from a minute depression; the posterior end is rounded. Specific characters very difficult to analyze. Fresh and salt water.

|
Bodo globosus. ENLARGE |
Bodo globosus Stein. Fig. 13.
The body during movement is globular or ovoid, without any anterior process. The trailing flagellum is invariably much longer than the vibratory one. The contractile vacuole lies in the anterior half of the body. Solid food particles are taken in near base of flagella.
Length of body 9 to 12µ; diameter 8 to 11µ. Common.
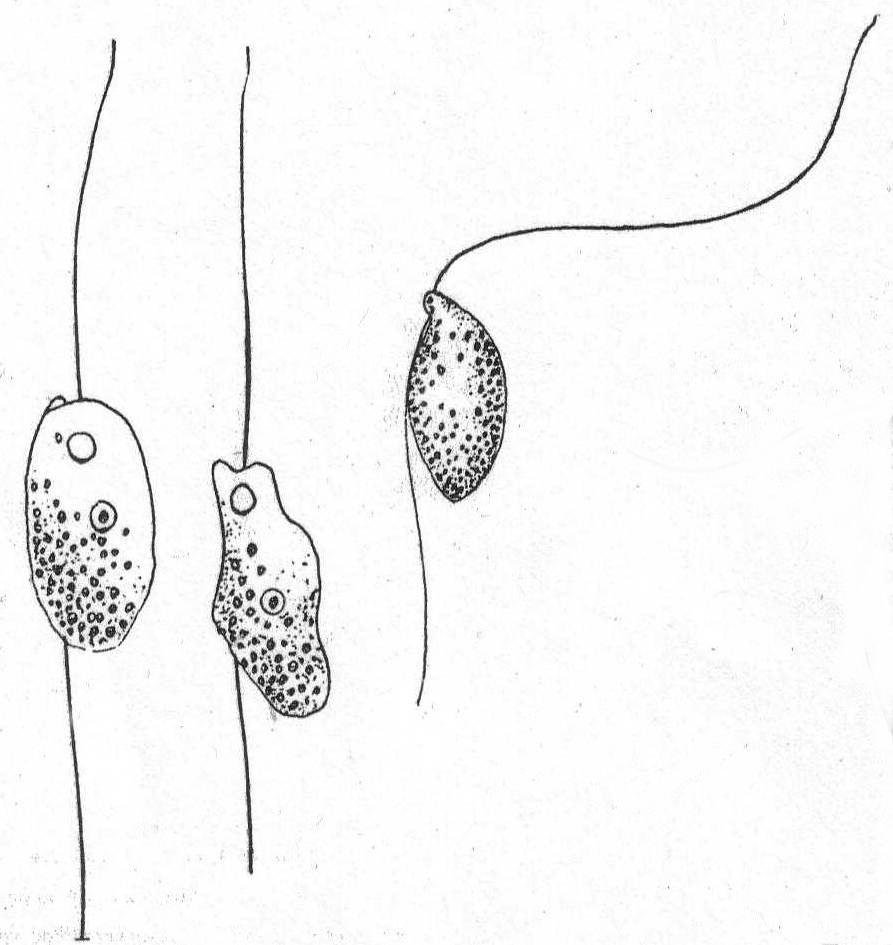
Bodo caudatus (Duj.) Stein. Fig. 14.
Synonyms: Amphimonas caudatus Duj.; Diptomastix caudata Kent.
The body is variable in shape, but usually flattened and pointed posteriorly. An anterior process is almost always present, and below this the flagella are inserted in a minute depression. The contractile vacuole is close to the base of the flagella. The flagella are about the same size, the anterior one usually somewhat longer. Common. Length 12 to 18µ.
This species was seen by Peck '95 and described as a small flagellate.
|
Fig. 14.—Bodo caudatus.
ENLARGE |
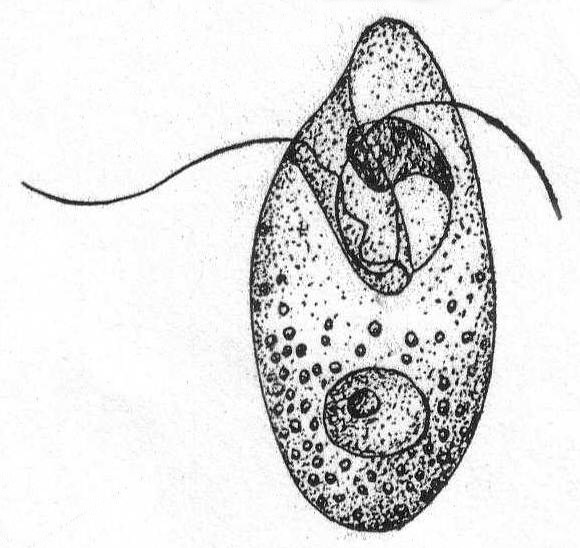
Medium-sized forms, somewhat oval in shape, with a rounded posterior end. The anterior end is continued dorsally in a somewhat attenuate pointed process. At the base of this process is a large cavity or funnel, on the dorsal wall of which, or on a projection from this wall, are two equal-size flagella. When at rest, the flagella are directed backwards. The nucleus is central. In moving, the posterior end is invariably in advance. This genus is exceptional among Mastigophora in that division is transverse instead of longitudinal.
Oxyrrhis marina Duj. Fig. 15.
With the characters of the genus. Contractile vacuole not seen. Length 28 to 40µ.
|
Oxyrrhis marina. ENLARGE |
Flagellates with one flagellum, a spindle-form body and a high degree of plasticity, the contour constantly changing. A distinct, usually striped cuticle is invariably present. "Eye-spots" are absent. Fresh and salt water.
Astasia contorta Duj. Fig. 16.
Astasia inflata Duj. '41.
The body is colorless, transparent, and flexible. It is largest in the center, thence tapering at the two extremities. The surface of the cuticle is obliquely striated, giving to the animal a distinctly twisted appearance. The contractile vacuole is in the anterior neck-like portion of the body. The flagellum is inserted in a distinct œsophageal tube, into which the contractile vacuole empties. This tube is continued into a deeper pharyngeal apparatus of unknown function.
Common in decaying algæ. Length 60µ; greatest diameter 30µ.
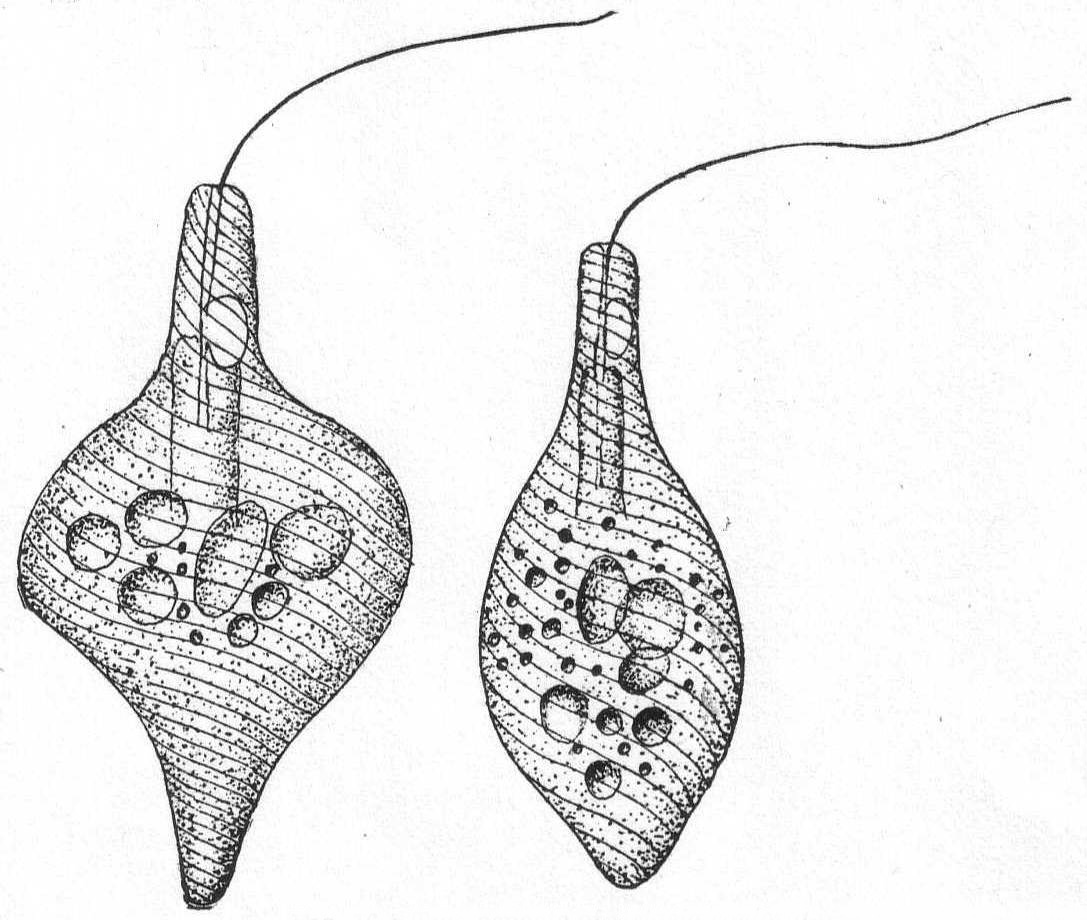
Fig. 16.—Two aspects of Astasia contorta. ENLARGE |
Flagellates with two flagella, of which one is directed forwards and is concerned with the locomotion of the animal, while the other is directed backwards and drags after the animal when in motion. Body slightly compressed dorso-ventrally (fig. 17, section). An oral furrow is present on the ventral side and the two flagella originate in it (fig. 17, at left). The vacuole is on the left side. Food vacuoles are present in the posterior part. The nucleus is central. Movement creeping.
Fresh and salt water.
Anisonema vitrea (Duj.) Fig. 17.
Synonyms: Tropidoscyphus octocostatus Stein '83; Sphenomonas Kent '81; Plœotia vitrea Senn 1900.
With the characteristics of the genus. It differs from freshwater forms in having eight furrowed surfaces running somewhat spirally from the posterior to the oral end. Length 50µ; width 23µ. This attractive flagellate was quite common in decaying algæ at Woods Hole; its shaking movement, its peculiar furrowed surfaces, and, above all, its perfectly transparent, vitreous appearance, were well described by Dujardin. Stein's Tropidoscyphus octocostatus is a fresh-water form which may possibly be a distinct species, especially as it is described with both flagella directed forwards.
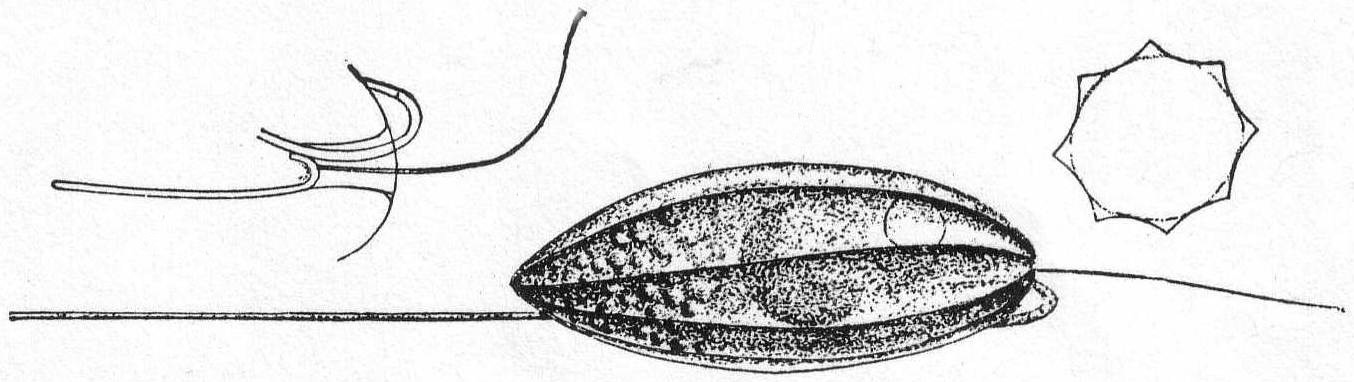
Fig. 17.—Anisonema vitrea. ENLARGE |
An aberrant flagellate bearing a single flagellum and a silicious skeleton resembling those of the Radiolaria. The skeleton consists of two rings of different diameter parallel with one another and connected by silicious bars. From the wider ring half a dozen bars radiate outwards and a similar number of short thorn-like bars point inwards obliquely. The color is yellow, and except for the flagellum the form might easily be mistaken for a Radiolarian, as has been the case repeatedly.
Distephanus speculum Stöhr.
Dictyocha speculum Stöhr; Dictyocha Auc.
With the characters of the genus.
A single specimen only of this very interesting form was seen at Woods Hole. It occurred in a collection of tow made near the end of the wharf during the evening.
| Key to families of Dinoflagellidia. | ||
| 1. | No crossfurrow; two free flagella | Family Prorocentridæ |
| 2. | One or more cross-furrows | 3 |
| 3. | Cross-furrow nearly central (cf. Oxytoxum) | Family Peridinidæ |
| Cross-furrow close to anterior end | Family Dinophysidæ | |
| Several cross-furrows and flagella |
Family Polydinidæ (One genus, Polykrikos.) |
|
| Key to marine genera of Prorocentridæ. | ||
| Diagnostic characters: The transverse furrow is absent and the two flagella arise from the anterior end of the body. The shell may be bivalved. | ||
| 1. | No tooth-like process dorsal to the flagellum | Genus *Exuviælla |
| 2. | With tooth-like process dorsal to the flagellum | Genus Prorocentrum |
| * Presence at Woods Hole indicated by asterisk. | ||
| Key to marine genera of Peridinidæ. | ||
| Diagnostic characters: The cross-furrow is nearly central (see, however, Oxytoxum); the body may or may not have a shell; the shell may or may not be composed of distinct plates; the plates are distinguished as equatorial (i.e., bordering the cross-furrow), apical, and antapical, while still another, the "rhombic plate", may be present, extending from the cross-furrow to the apex. | ||
| 1. | Without distinct shell | Genus *Gymnodinium |
| With a distinct shell | 2 | |
| 2. | Shell not composed of definite plates | 3 |
| Shell composed of definite plates | 4. | |
| 3. | Cross-furrow replaced by thin-skinned band | Genus Ptychodiscus |
| Cross-furrow well defined; reticulate markings raised on shell-surface | Genus Protoceratium | |
| Cross-furrow well defined; no markings | Genus *Glenodinium | |
| 4. | Two parts of shell equal or nearly equal | 5 |
| Two parts of shell very unequal | 11 | |
| 5. | With transverse flagellum in a distinct furrow | 6 |
| Transverse flagellum not in a furrow | 10 | |
| 6. | With horns, or with wing-like processes | 7 |
| Without processes of any kind kind | 9 | |
| 7. | Processes small, wing-like, around flagellum-fissure | Genus Diplopsalis |
| Processes horn-like | 8 | |
| 8. | Anterior part with 7 equatorial and 1 rhombic plates | Genus *Peridinium |
| Anterior part with 5 equatorial and no rhombic plates | Genus Gonyaulax | |
| Anterior part with 3 equatorial and no rhombic plates | Genus *Ceratium | |
| 9. | Anterior part with 14 equatorial and 1 rhombic plates | Genus Pyrophacus |
| Anterior part with 7 equatorial plates | Genus Goniodoma | |
| Anterior part with 4 equatorial plates | Genus Amphidoma | |
| 10. | Apical extremity drawn out into a tube | Genus Podolampas |
| pical extremity not drawn out into a tube | Genus Blepharocysta | |
| 11. | Cross-furrow deep, with great ledge-like walls | Genus Ceratocorys |
| Cross-furrow wide, no ledge-like walls | Genus Oxytoxum | |
| * Presence at Woods Hole indicated by asterisk. | ||
| Key to marine genera of Dinophysidæ. | ||
| Diagnostic characters: The cross-furrow is above the center of the body, and its edges, as well as the left edge of the longitudinal furrow, are usually produced into characteristic ledges; those of the cross-furrow usually form great funnel-like anterior processes, while those of the longitudinal furrow usually form great, lateral, wing-like processes ornamented by ribs and other markings. | ||
| 1. | Without shell; longitudinal furrow may open & close | Genus *Amphidinium |
| 2. | With shell; longitudinal furrow unchangeable | 3 |
| 3. | With distinct apical funnel | 4 |
| No apical funnel | Genus Phalacroma | |
| 4. | With great wing-like ledge | 5 |
| Ledges very small; body long, needle-like | Genus Amphisolenia | |
| 5. | Ledge of longitudinal furrow extends to posterior end | 6 |
|
Ledge of longitudinal furrow does not extend to posterior
end (Recorded by Peck ('93-'95) as very abundant at Woods Hole and in Buzzards Bay.) |
Genus Dinophysis | |
| 6. | Ledge is continued dorsally to the cross-furrow | Genus Ornithocercus |
| Ledge is not continued dorsally | 7 | |
| 7. | With deep dorsal cavity; secondary funnel not notched | Genus Citharistes |
| No dorsal cavity; secondary funnel deeply notched | Genus Histioneis | |
| * Presence at Woods Hole indicated by asterisk. | ||
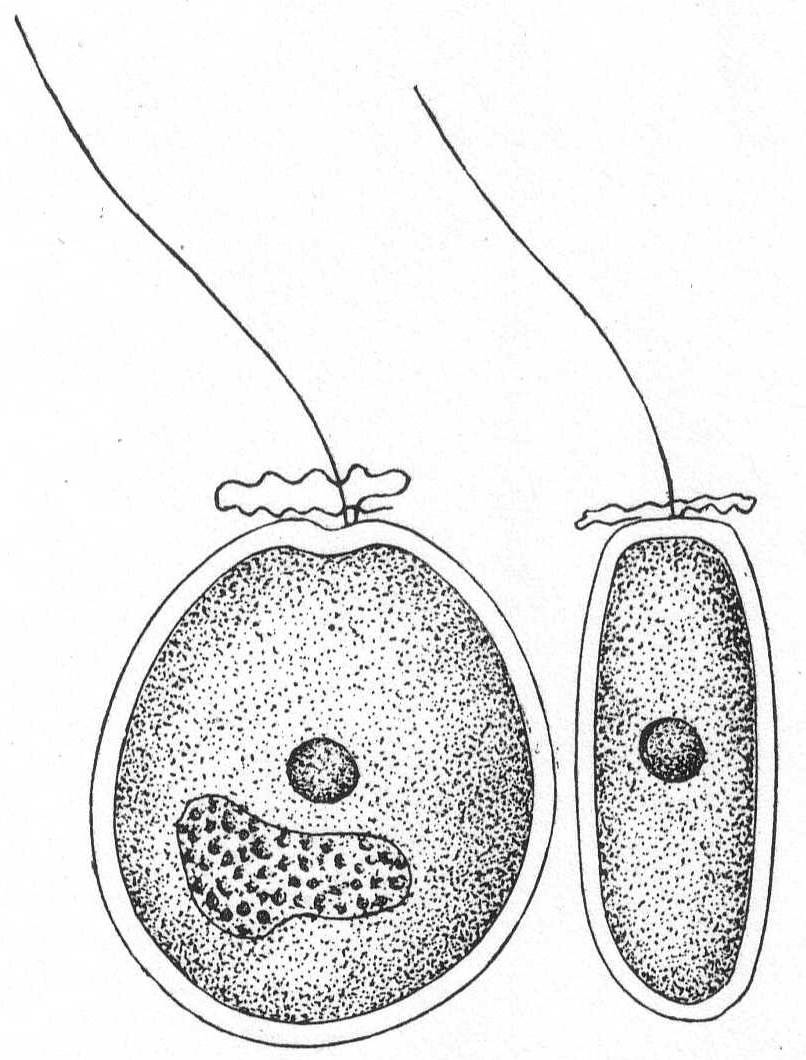
The form varies from globular to ovoid, with occasionally a sharp posterior end. Shells are usually somewhat compressed, and consist of two valves, which frequently slide one over the other in such a manner as to show the structure with great clearness. The right shell may have a distinct indentation in the anterior edge. There are two lateral, discoid, brown chromatophores, each of which possesses a central amylum granule. The nucleus is posterior. Salt water.
Exuviælla lima Ehr. Fig. 18.
Synonyms Pyxidicula Ehr.; Cryptomonas Ehr.; Prorocentrum lima Kent; Amphidinium Pouchet.
The shell is ovate, rounded and swollen posteriorly. The anterior border of both shells is slightly indented. The shell is quite thick. The animal moves through the water very slowly. Dark brown in color. Length 48µ; width 44µ.
|
Fig. 18.— Exuviælla lima. ENLARGE |
Exuviælla marina Cienkowsky. Fig. 19.
A smaller form than the preceding, more elliptical in outline, with a thinner shell and with large granules throughout the endoplasm. The nucleus is spherical and subcentral in position and possesses a distinct central granule. This may be a small variety of E. lima.

|
Fig. 19.— Exuviælla marina. ENLARGE |
The general structure of these forms is similar to that of Glenodinium; the most striking and positive difference is the absence of a shell. The animals are, as a rule, spherical, yet they may be pointed at the two ends or at one of them. They are also frequently flattened dorso-ventrally. The transverse furrow may be either circular and straight around the body or may describe a spiral course, passing even twice around the body. The flagella arise near cross-furrow or, in some cases, in longitudinal furrow. Chromatophores may or may not be present and food-taking is holozoic, in many cases at least. In some cases ectoplasm and endoplasm can be distinguished. Fresh and salt water.
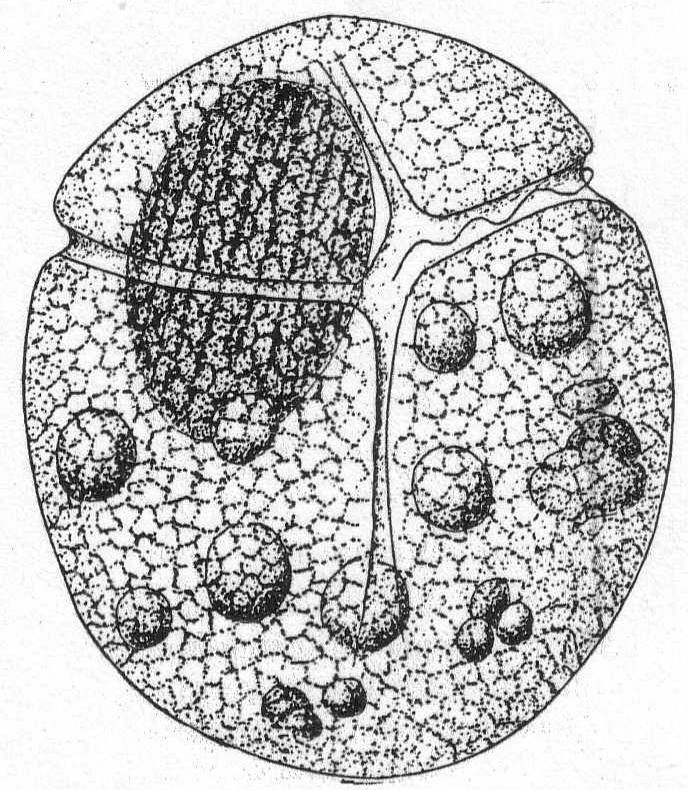
Gymnodinium gracile Bergh '82, var. sphærica, n. Fig. 20.
The body is divided by the transverse furrow into a shorter anterior and a longer posterior part. The longitudinal furrow is broader at the posterior extremity than at the cross-furrow. The structural feature upon which this new variety is made is the unvarying plumpness of the body, making it almost spherical, except for a slight flattening dorso-ventrally. The nucleus is large and ellipsoidal, with characteristic longitudinal markings of chromatin. The endoplasm is evenly granular, with a number of large ingested food bodies. The color is brown, not rose-red as in Bergh's species, nor is the Woods Hole form as large as the latter. Length of body 68µ; width 55µ. Common.
|
Fig. 20.— Gymnodinium gracile, var. sphærica. ENLARGE |
Small globular forms with two distinct furrows, one transverse around the body, the other longitudinal upon the face only. The shell is soft and structureless with a distinct aperture near the meeting point of the two furrows. The endoplasm usually, but not always, contains a bright red eye-spot.
Fresh and salt water.
Glenodinium compressa, n. sp. Fig. 21, a, b, c.
This species resembles G. acuminata of Ehrenberg except that it is strongly compressed laterally. The longitudinal furrow extends nearly to the extremity of the animal. It begins as a narrow slit and widens as it progresses upon the left side; it also becomes much deeper on this side and at the bottom of the depression the longitudinal flagellum is inserted. The transverse furrow runs evenly around the body near the upper pole, giving to the shell almost the aspect of an Amphidinium. Brown chromatophores may or may not be arranged radially about a central amylum granule. One striking characteristic is the depth of the two furrows. The nucleus is elongate and somewhat curved; it lies against the posterior wall of the rather thick shell. Not uncommon.
Length 40µ; breadth 32µ; width 18µ.
The posterior end of the animal is often somewhat pointed and this point frequently becomes attached, so that the animal whirls around upon it as upon a pivot.
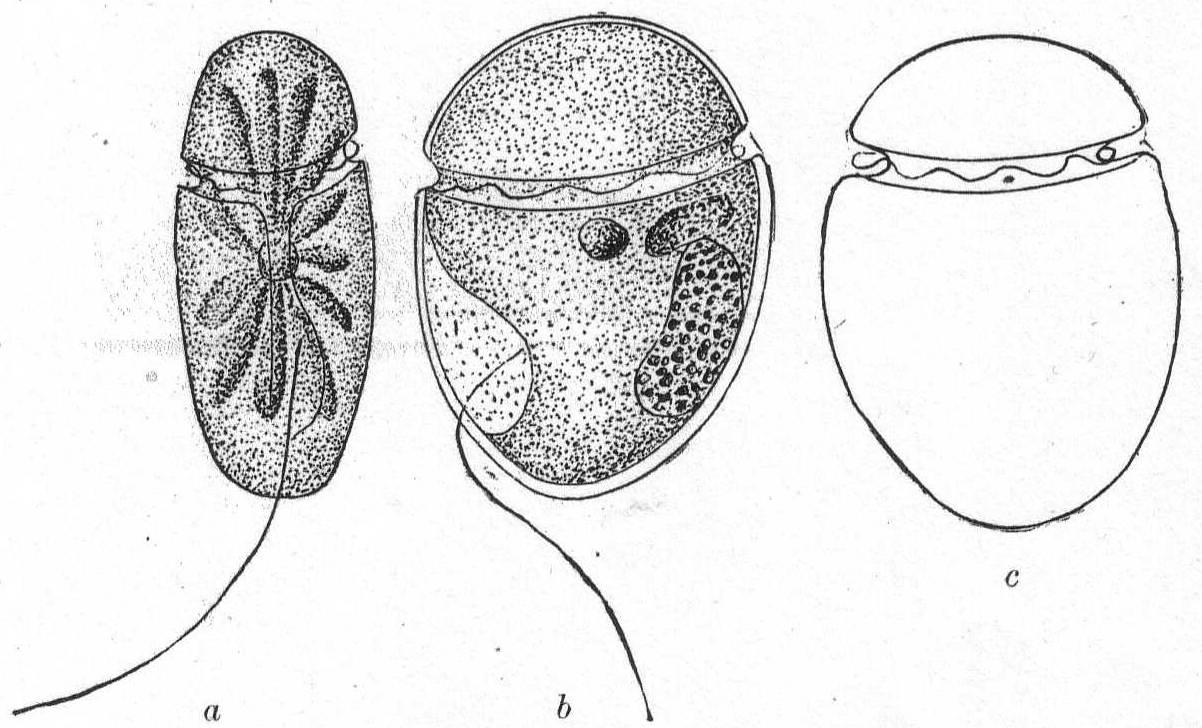 Fig. 21 a, b, c.—Glenodinium compressa, n. sp. ENLARGE |
Glenodinium cinctum Ehr. Fig. 22.
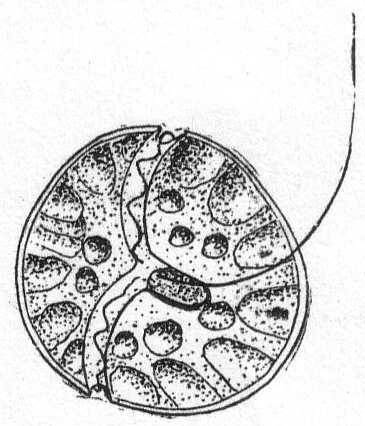
Fig. 22.— Glenodinium cinctum. ENLARGE |
The body is globular, smooth, and homogeneous. Brown chromatophores arranged radially, each in the form of a cone, the base of which rests against the shell while the points turn inward. A bright-red eye-spot may or may not be present; when present it is placed near the junction of the two furrows. The longitudinal furrow is small. Fresh water and salt.
Length and diameter the same, 21µ.
This species was observed by Peck '93.
[Illustration: Fig. 22.—Glenodinium cinctum.]
The form is globular, ovoid or elongate, the apex frequently drawn out into a long tube. The transverse and longitudinal furrows are quite distinct, the former having often a spiral course about the body. The two halves of the body are similar, the posterior being somewhat shorter; the anterior half has seven equatorial plates, an oral plate, two lateral apical plates, and one or two dorsal plates. The two antapical plates frequently have a tooth-like process. The bodies are colorless, green or brown.
Fresh and salt water.
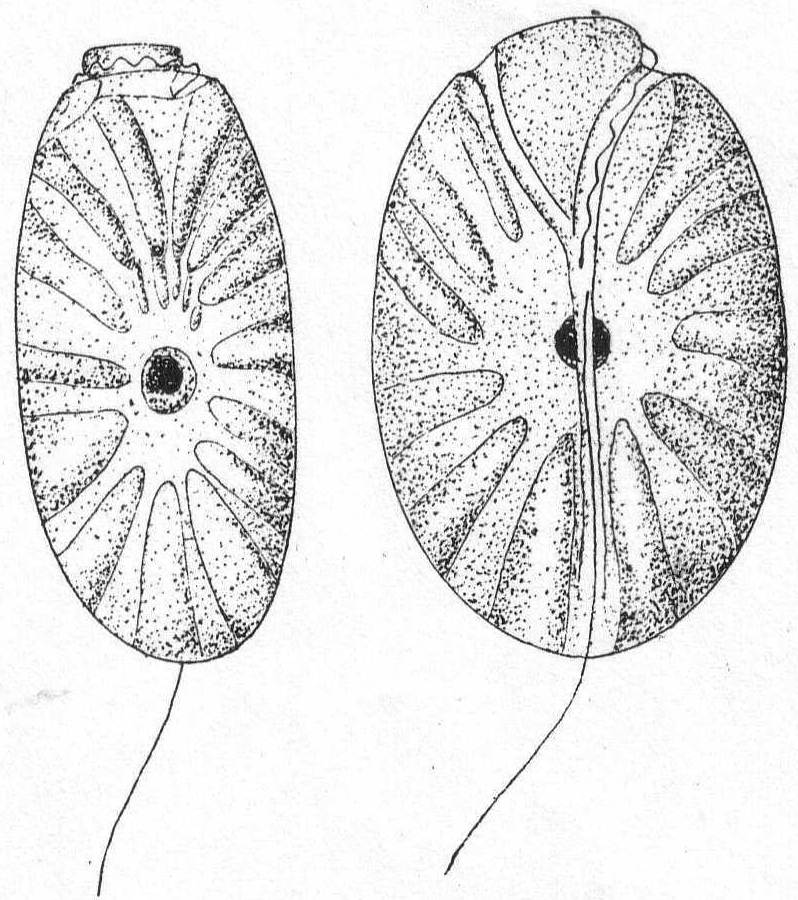
Peridinium digitale Pouchet. Fig. 23.
Synonyms: Protoperidinium digitale Pouchet; Protoperidinium Bergh p. p.; P. divergens Peck.
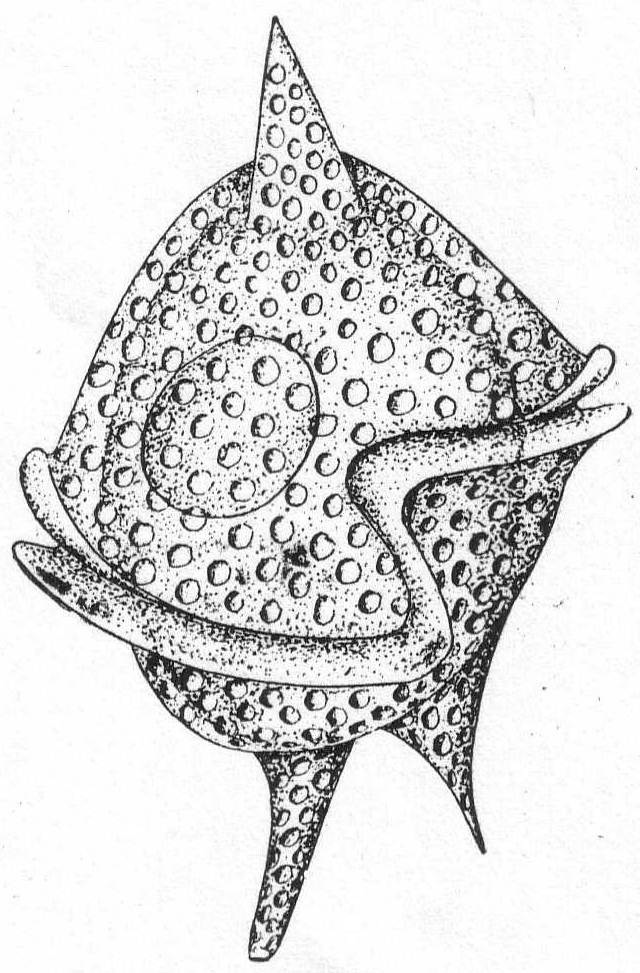
Fig. 23.— Peridinium digitale. ENLARGE |
The shell is covered with pits of large size. The posterior part is hemispherical and surmounted by a single horn or spine. The transverse furrow is very oblique, and its two extremities are united by a sigmoid longitudinal furrow. The anterior half bears two spines or horns of different size, and variable. The nucleus is spherical or ellipsoidal and placed in the posterior half of the shell.
Length 68µ; diameter 54µ. Common.
Although the description of Pouchet's P. digitale differs in some respects from a careful description of the Woods Hole form, I think the species are the same. The chief difference is in the single horn of the posterior half; in Pouchet's form this is furrowed by a narrow groove which runs to the S-shaped longitudinal furrow. In the Woods Hole form I was unable to make out such a furrow. The flagella, also, were not seen. This same form was pictured by Peck '95 as P. divergens.
Peridinium divergens Ehr. Fig. 24.
Synonym: Ceratium divergens Kent.
The shell is spheroidal, widest centrally, attenuate and pointed posteriorly; the anterior portion is armed with two short, pointed horns, each of them having a toothed process at the basal portion of the inner margin. They are frequently colorless and beautifully transparent, the body being free from large opaque granules; again they are colored brown or yellow. The nucleus is large and elongate and finely granular. 75µ long and 68µ in diameter. Common.
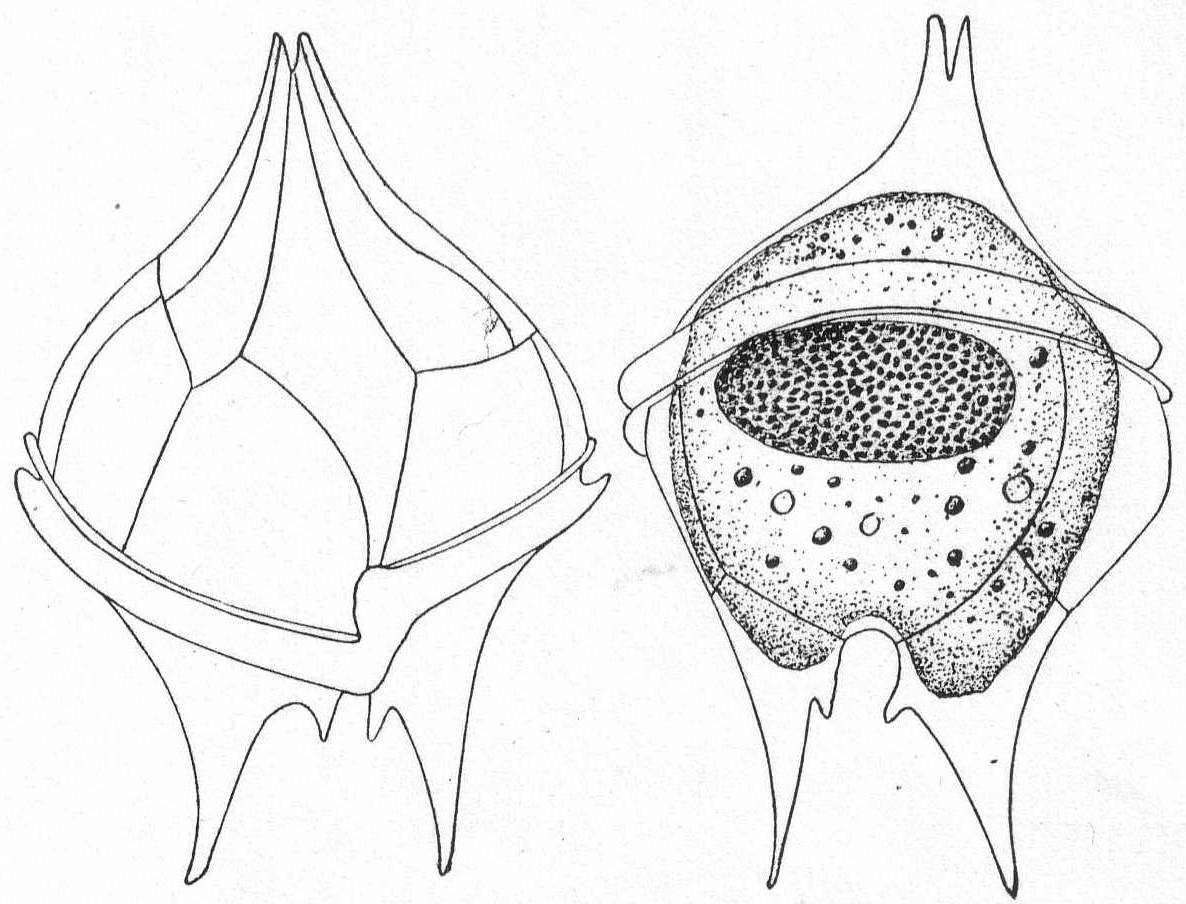
Fig. 24.—Ventral and dorsal aspects of Peridinium divergens. ENLARGE |
The general shape is a flattened sphere with three long processes or horns. The cross-furrow is either spiral or circular; the longitudinal furrow is usually wide and occupies the greater part of the anterior half of the shell. The shell is thick, reticulate or striped, and sometimes provided with short spines; often distinctly porous. The anterior half is composed of 3 equatorial and 3 apical plates, the latter being continued into the horn-like process. The posterior half is composed of 3 equatorial and one apical plate continued into the posterior horn. The right posterior plate is continued into a similar horn which may remain rudimentary or be continued into a considerable process. Similarly the left posterior horn is usually developed, but remains small. There may be from 2 to 3, 4, and 5 horns. Chromatophores usually present, green to yellow brown.
Fresh and salt water.
Ceratium tripos Ehr. Fig. 25.
The body is somewhat triangular and bears three horns, two of which are shorter than the other one and slightly curved upward.
Length, including the horns, 290µ.
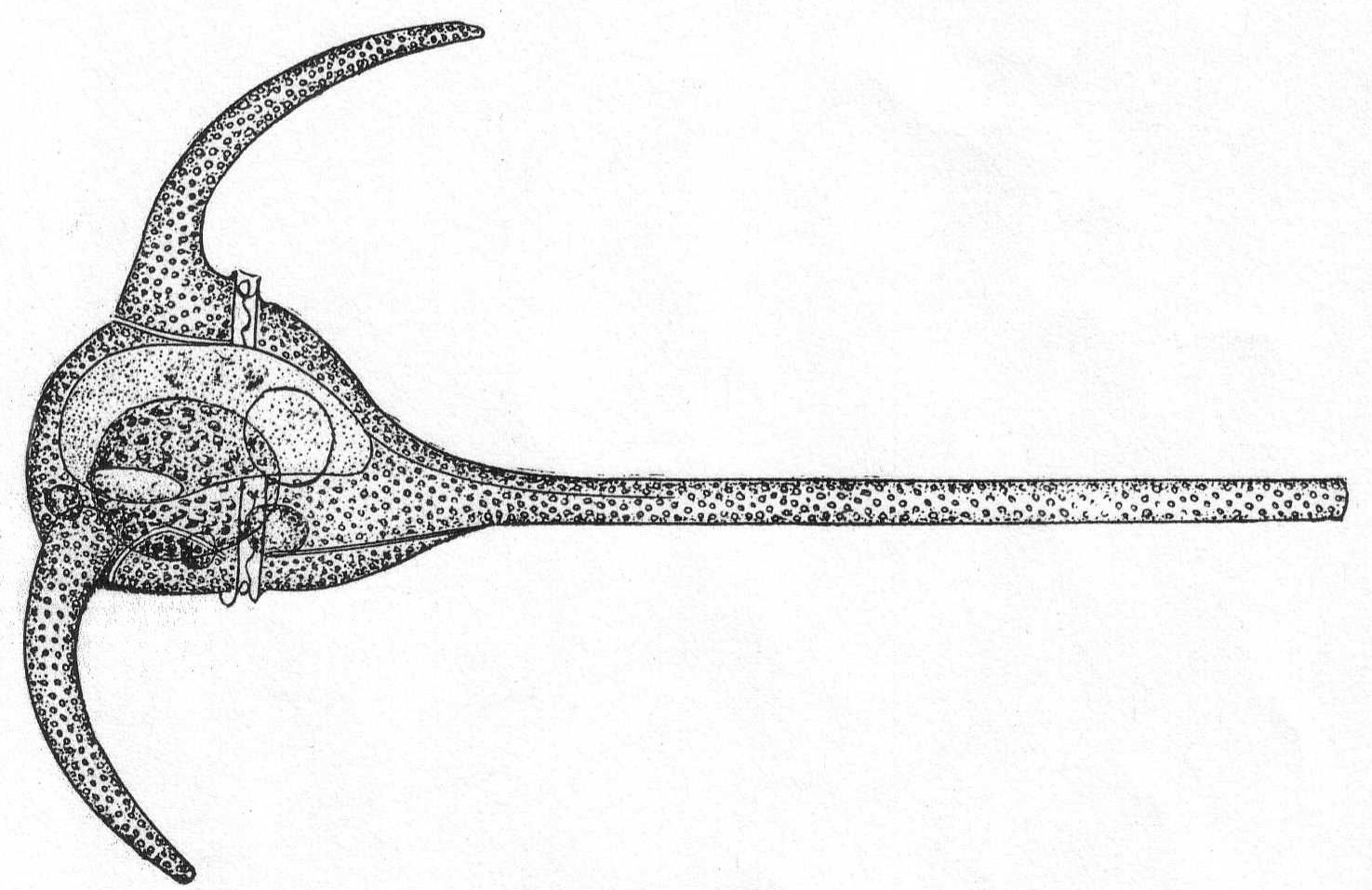
Fig. 25.—Ceratium tripos. ENLARGE |
Ceratium fusus Ehr. Fig. 26.
Synonym: Peridinium fusus Ehr.
The animal is very elongate, due to the presence of two long horns at the extremities of the body. Color, yellow with chromatophores. Length 285µ; width 23µ.
Both of these species are common in the tow and in the algæ at the edge of the wharf. Both of them are mentioned by Peck in '93 and '95.
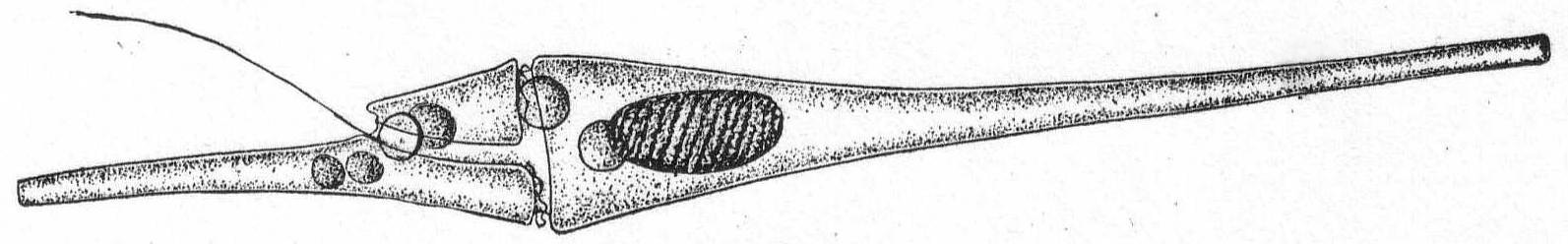
Fig. 26.—Ceratium fusus. ENLARGE |
The body is ovoid to globular and usually much flattened dorso-ventrally. The anterior portion is very much reduced and is somewhat head-like or cap-like. The longitudinal furrow extends through the entire posterior body length and is apparently capable of widening and narrowing. It is probably naked (see here Klebs, Pouchet, Bütschli), although Stein maintained that there is a delicate cuticle-like shell. Chromatophores of brown or green colors present and usually grouped radially about a central amylum granule. The nucleus is posterior.
Fresh and salt water.
Amphidinium operculatum Clap. & Lach. Fig. 27.
The body is oval and flattened. The transverse furrow is at the extremity (posterior) of the body and the small portion, which is thus apparently cut off, is the cap-like or operculum-like structure which gives the name to the species. Klebs maintains that the two furrows are not connected, but in this he is certainly mistaken, provided we have the same species under consideration. Very common about Woods Hole.
Length from 40 to 50µ; width 30µ; thickness 15µ.
|
Fig. 27.— Amphidinium operculatum. ENLARGE |
| Key to Infusoria. | |||
| 1. | With cilia | Subclass Ciliata. 3 | |
| 2. | Without cilia (in adult state) tentacles | Subclass Suctoria | |
| 3. | a. | Without a specialized fringe of large cilia (ad. zone) | Order Holotrichida |
| b. | With general covering of cilia + adoral zone | Order Heterotrichida | |
| c. | With cilia on ventral side + adoral zone | Order Hypotrichida | |
| d. | With cilia in region of adoral zone, and about mouth only | Order Peritrichida | |
| Key to families of the Holotrichida. | ||||||
| A. | Mouth closed except during food ingestion; no undulating membrane | 1 | ||||
| Mouth always open; with undulating membrane | 2 | |||||
| 1. | Gymnostomina. | |||||
| a. | Mouth terminal or subterminal. Food is swallowed and not introduced by currents | Family Enchelinidæ | ||||
| b. | Mouth terminal or subterminal; body frequently drawn out into long process; mouth may have specialized framework | Family Trachelinidæ | ||||
| c. | Mouth central or posterior; pharynx with supporting framework | Family Chlamydodontidæ | ||||
| 2. | Trichostomina. | |||||
| a. | Mouth anterior or central; pharynx short or absent; peristomial depression faint or absent | Family Chiliferidæ | ||||
| b. | Mouth central; pharynx long, tubular; cilia in two broad zones | Family Urocentridæ | ||||
| c. | Mouth posterior; form asymmetrical; cilia dispersed or limited to oral region | Family Microthoracidæ | ||||
| d. | Mouth anterior or central. Peristomial depression clearly marked. |
Paramœcidæ (One genus, Paramœcium) |
||||
| e. | Mouth at end of long peristome running along ventral side; body dorso-ventrally or laterally compressed; left edge of peristome with great, sail-like undulating membrane | Family Pleuronemidæ | ||||
| f. | Mouth and pharynx distinct, posterior; cilia uniform. Parasites in ruminants. | Family Isotrichidæ | ||||
| g. | Mouth absent; body vermiform, cilia uniform. Usually parasites. | Family Opalinidæ | ||||
| Key to marine genera of Enchelinidæ | |||
| Diagnostic characters: Form ellipsoid or ovoid; the mouth is invariably terminal and is usually round—more rarely slit-formed; it is closed except when food is taken. An œsophagus when present is a short, invariably non-ciliated tube which is usually surrounded by a more or less clearly defined buccal armature. The anus is usually terminal. Large food particles are swallowed, never introduced by currents. | |||
| 1. | Body naked | 3 | |
| 2. | Body inclosed in a shell or coat | 7 | |
| 3. | a. | Cilia uniform about the entire body; body symmetrical | 4 |
| b. | Cilia in the mouth region longer than the others; body symmetrical | 5 | |
| c. | Bristles, or tentacles, in addition to cilia | 6 | |
| 4. | Mouth terminal; body ellipsoidal to ovoid | Genus Holophrya | |
| 5. | a. | Mouth terminal; body elongate, flexible, and elastic | Genus Chænia |
| b. | Mouth terminal; "neck" highly elastic; entire body flexible; conical "head" | Genus *Lacrymaria | |
| c. | Mouth terminal; "neck" highly elastic; entire body flexible; "head" square | Genus *Trachelocerca | |
| d. | Mouth terminal; "neck" highly elastic; no separate mouth-bearing portion | Genus Lagynus | |
| 6. | a. | Body asymmetrical; bristles in addition to cilia | Genus Stephanopogon |
| b. | Body symmetrical; 4 small tentacles from mouth; cilia and cirri in girdles | Genus *Mesodinium | |
| 7. | Shell composed of small sculptured pieces; cilia long, uniform | Genus *Tiarina | |
| * Presence at Woods Hole indicated by asterisk. | |||
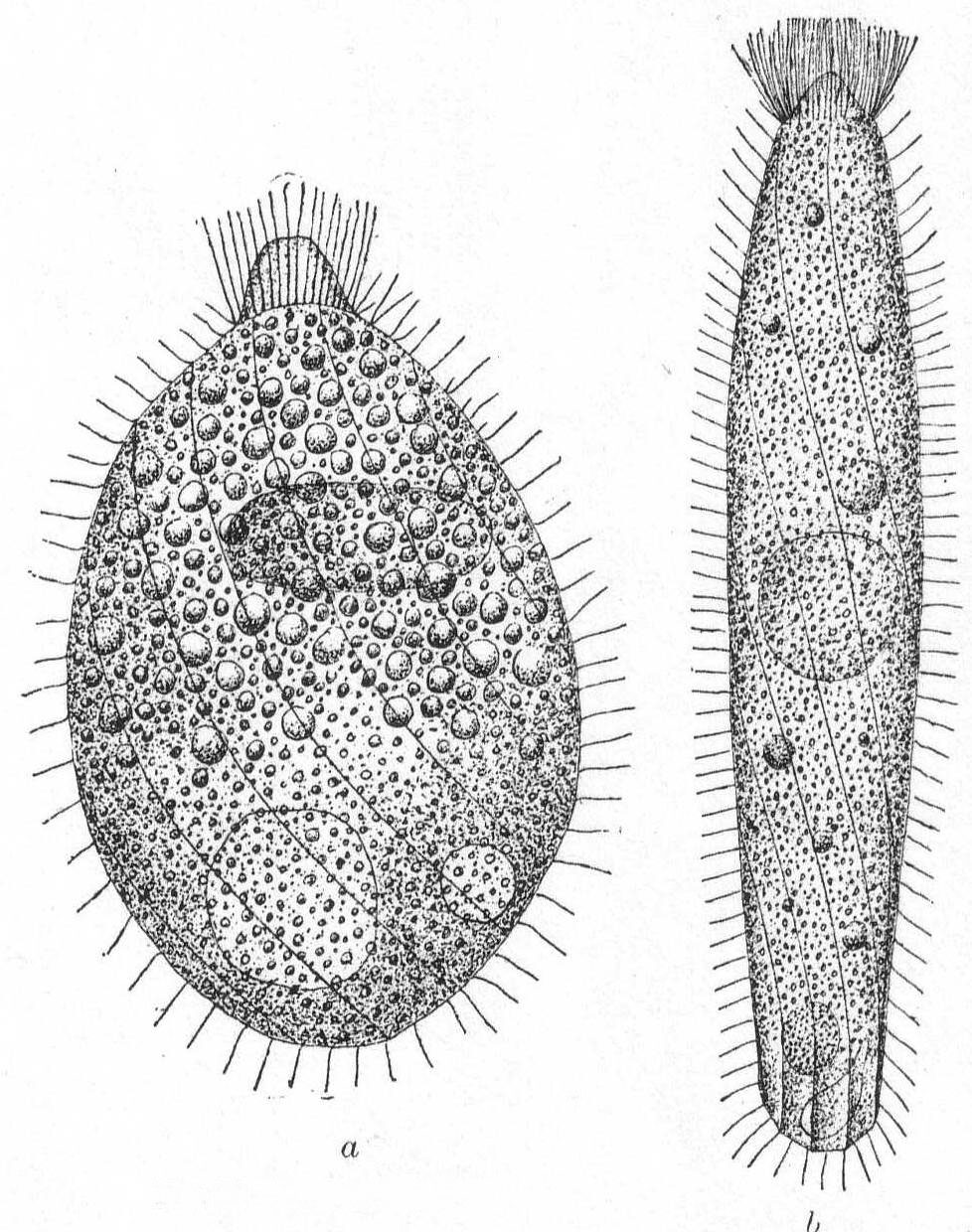
Body short to very long flask-shape; for the most part contractile, especially in the neck region. The posterior end is rounded or pointed. The main character is the mouth-bearing apex, which "sets like a cork in the neck of the flask." One or more circles of long cilia at the base of the mouth portion or upon it. The body is spirally striped. Contractile vacuole terminal, with sometimes one or two further forward. Macronucleus central, globular to elongate, sometimes double. Food mainly bacteria. Fresh and salt water.
Lacrymaria lagenula Clap. & Lach. Fig. 28, a, b.
Synonym: L. tenuicula Fromentel '74.
Body more or less flask-shape, two or three times as long as broad, with conical apex, which is slightly elastic and protrusible; surface obliquely striate, with well-defined lines, 14 to 16 in number; cilia uniform on the body, with a crown of longer ones at the base of the conical proboscis. The body cilia are not thickly placed except around the proboscis. The endoplasm is thickly packed with large granules (food particles) in the anterior half and with finely granular particles in the posterior half. The elongate macronucleus lies a little above the center among the larger granules; the contractile vacuole is double, one on each side of the median line and at the posterior end of the body among the finer granules. The anus is posterior. Length 90µ to 160µ; greatest width assumed 65µ. When fully expanded the posterior end assumes a curious polyhedral form. (Fig. 28 b.)
This form differs slightly from others of the same species as described by different observers, the most striking difference being the presence of two contractile vacuoles in place of the usual one. These are very slow to fill and grow to a large size before diastole. The membrane is very tough and retains its form easily under pressure of the cover glass. Another characteristic feature is the flattening of the surfaces between the striæ. Decaying algæ.
|
Fig. 28.— Lacrymaria lagenula. ENLARGE |
Lacrymaria coronata Cl. & Lach. '58. Fig. 29.
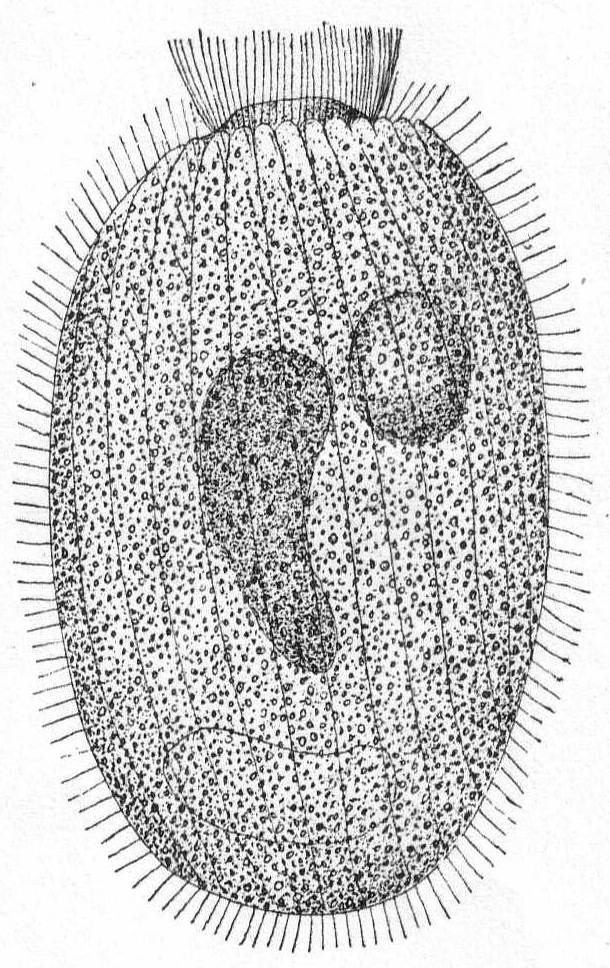
Fig. 29.— Lacrymaria coronata. ENLARGE |
Synonyms: L. lagenula Cohn '66; Möbius '88; L. cohnii ? Kent '81; L. versatilis Quen. '67.
Form flask-like and similar to L. lagenula, contractile but tough. The contractile vacuole is terminal, the proboscis is short, slightly raised and separated from the body by a deep cleft; the buccal cilia are inserted part way up on the proboscis. Form changeable, from short, sac-like to elongate and vermiform. Length 85µ.
This species is not very different from L. lagenula, but I noted that in addition to the elongate nucleus, the body striæ are much more apparent here and seem to sink into the cuticle, giving the periphery, especially at the collar region, a curious crenulated effect. The endoplasm is very densely granular and colored a blue-green, probably from food particles. The number of striæ is much larger than in the preceding species. The membrane is very tough and retains the shape of the body, even with the full pressure of the cover glass. Micronucleus and trichocysts were not observed.
The only well-known representative is very elongate, large (up to 3 mm. Van Beneden), and very contractile. The main feature of importance in distinguishing it is the 4-part structure of the mouth region, which, however, may not be obvious. Pharynx faint and smooth. Contractile vacuole terminal. Macronucleus in one central body or in numerous pieces scattered throughout the cell. Salt water.
Trachelocerca phœnicopterus Cohn '66. Fig. 30.
Synonyms: T. sagitta Ehr. '40, Stein '59; T. tenuicollis Quennerstedt '67, Kent '81; T. minor Gruber '87, Shevyakov '96.
The body is extremely elongate and ribbon-like, and this, combined with its wonderful power of extension and retraction, makes it one of the most curious and interesting of microscopic forms. The anterior end is square or cylindrical; the type species has a four-sided mouth, but many specimens may be found which have a plain cylindrical mouth region. One reason for this may be the fact that the extremity gets broken off. In one instance I noticed a very large form with the anterior end under some debris, which evidently held it tight, for the body of the ciliate was thrashing back and forth and twisting itself into knots, etc., like a nematode worm. Finally, the anterior end broke off with about one-tenth of the body; the remainder, in an hour, had regenerated a new anterior end with long cilia, but with no indication of four sides. The small anterior piece was also very lively, moving about and eating like the normal animal; its history, however; was not followed. This species appears to be variable in other ways as well; thus, in some cases the posterior end is rounded (cf. Entz '84); in others it is pointed (cf. Kent '81, Cohn '66, et al.).
Again, the macronucleus may be a single round body (Entz '84, Bütschli '88) or in two parts (Kent '81), or in many parts scattered about the body (Gruber). In the Woods Hole forms the tail is distinctly pointed and turned back sharply, forming an angle at the extremity. The cilia on this angular part are distinctly longer than the rest. The function of this posterior part is apparently to anchor the animal while it darts here and there upon the tail as a pivot, contracting and expanding the while. The body is finely striated with longitudinal markings; when contracted there are no transverse markings nor annulations. The nucleus is in the form of many fragments scattered throughout. Length of large specimen 1.7 mm.
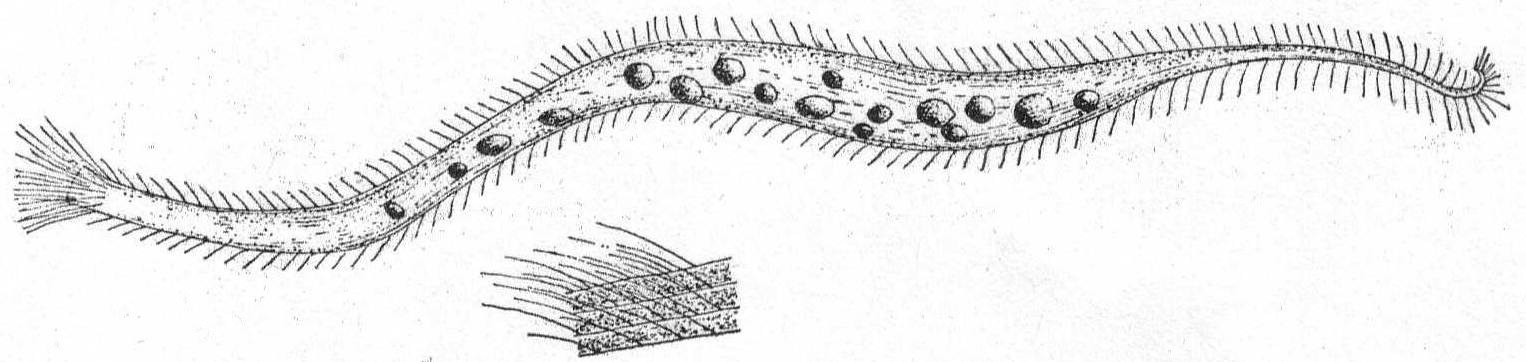
Fig. 30.—Trachelocerca phœnicopterus. ENLARGE |
The main part of the body is globular or conical, with a short, platform-like oral region, and a deep annular groove about the middle of the body. The œsophagus is rather long, and smooth or longitudinally striped. One or more rings of cirri rise in the groove. If more than one ring of cirri are present, the anterior set usually point forward and lie close to the anterior part of the body. The posterior set, on the other hand, cling close to the posterior region of the body and give to it a peculiar encapsuled appearance. The most characteristic feature is the presence of four short tentacle-like processes which can be protracted and retracted from the oral region. (Mereschowsky says that the entire anterior half is more or less contractile.) The macronucleus is horseshoe-shaped or ovoid and is situated in the posterior half of the body. The contractile vacuole is also posterior.
Movement consists in rapid swimming, with rotation on its axis, or in creeping by means of its anterior cirri, or in sudden jumping, by which it apparently clears a distance of 20 times its diameter in one bound. Mouth parts may also be used for attachment to foreign bodies. The moving periods alternate with quiescent periods, during which the organisms with their outstretched and radiating cirri resemble the heliozoön Actinophrys.
Mesodinium cinctum, n. sp. Fig. 31.
Body spherical to pyriform, constricted near the middle, the constriction dividing the body into dissimilar parts. The anterior part is broadly pyriform, somewhat plastic and hyaline, with an oral extremity which is sometimes hollow, sometimes evaginated and convex. Upon this flexible anterior part there are four short but distensible tentacles. The posterior part is granular and usually filled with food particles; it is well rounded and holds the nucleus and contractile vacuole. The entire body is surrounded by a fine cuticle. The nucleus is elongate and extends through the greater part of the posterior half. The contractile vacuole lies on one side, near the girdle. The mouth is on the anterior pole in the tentacle region. The motile organs are cirri and cilia, all inserted in the constriction. There are two sets of cirri and one of cilia; the latter stand out radially from the girdle and are usually in motion. The cirri of one set, the anterior, extend forward about twice the length of the anterior half; those of the posterior set closely engirdle the lower half, reaching not quite to the posterior extremity. These are somewhat hyaline and are closely approximated, giving the impression of a tight-fitting crenulate casing about the lower half. The cirri are sharply pointed, much broader at the base, and the two sets are so placed that, looked at from above, they have the appearance of a twisted cord. (Fig. 31 b.) Movement erratic; sometimes the animal swims steadily forward with mouth in front; again it shoots across the field of the microscope, either backward or forward or sideways, through the action of its powerful cirri. It is often quiet, usually mouth downward, and is held in place by adhesion of the tentacles. In this position it looks strikingly like a heliozoön.
Length 35µ; greatest width 30µ. Not uncommon.
The chief features by which this species is distinguished from the frequently described M. pulex of Europe are the number of anterior cirri and the ring of true cilia in place of the central girdle of cirri. The European form is described with four anterior bristles; the present form has from 28 to 32. The radial cilia differ decidedly from the more powerful cirri and they are not in one plane, so that counting is difficult; they are not closely set. The presence of tentacles makes these forms of especial theoretical interest, especially in the light of the origin of Suctoria.
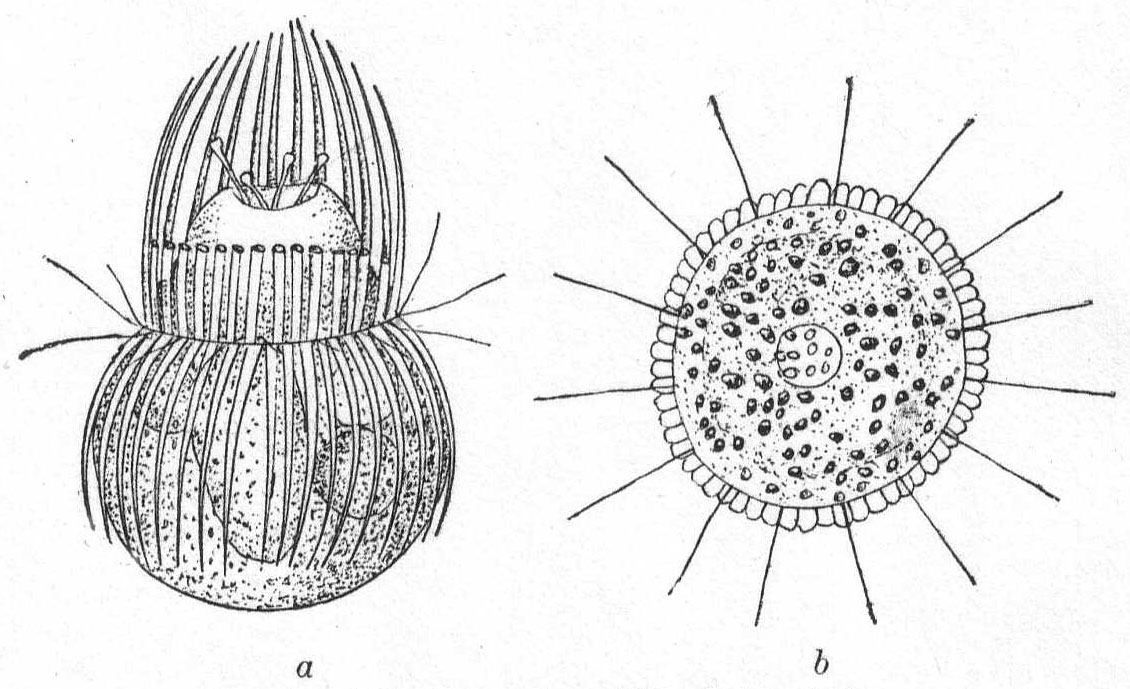
Fig. 31.—Side and top views of Mesodinium cinctum. ENLARGE |
Body subcylindrical, pointed posteriorly, two and one-half times as long as broad; encased in covering composed of separate pieces arranged in five girdles. The pieces bear processes which rest against neighboring pieces of the girdle. Mouth large, anus terminal near contractile vacuole. The macronucleus is simple and round. Salt water.
Tiarina fusus (Cl. & Lach.) emend R. S. Bergh.
Synonyms: Coleps fusus Cl. & Lach. '58; Daday '86; Möbius '88, Lauterborn '94; Shevyakov '86.
This form, which resembles Coleps rather closely, was placed as a separate genus by R. S. Bergh. The skeletal parts consist of five zones of needles composed of an organized substance and embedded in the cortical plasm, the last zone coming to a point at the posterior end. The needles have lateral processes, which give a latticed appearance to the casing. The cilia are long, with a specialized crown of still longer ones at the oral end; they arise outside of the skeletal elements and do not pass between them, as in Coleps.
| Key to marine genera of Trachelinidae. | |||
| Diagnostic characters: Body bilateral, or asymmetrical by local prolongations; usually compressed or flattened laterally, the left side more convex than the right. The essential feature is the position and character of the mouth. This is either a long slit extending from the anterior end well down the ventral surface, or the posterior part only of a ventral furrow remains open as a round or elongate mouth some distance from the anterior end. The entire mouth region of the body is usually drawn out into an elongate tapering proboscis which is generally curved dorsally at the extremity. An œsophagus is short or absent altogether; when present it is supported by a stiff buccal armature. Cilia are uniform about entire body or limited to the flat right side. Food is swallowed. | |||
| 1. | a. | Proboscis easily distinguished from the main body | 2 |
| b. | Proboscis not marked off from main body; body flat; both surfaces striated | Genus *Loxophyllum | |
| 2. | a. | Mouth runs the entire length of proboscis; entire body uniformly ciliated | Genus Amphileptus |
| b. | Mouth runs the entire length of proboscis; body flat; right side only is ciliated | Genus *Lionotus | |
| c. | Proboscis much drawn out, flexible; mouth at its base | Genus Dileptus | |
| * Presence at Woods Hole indicated by asterisk. | |||
The body is flat and somewhat leaf-shape, flexible, and elastic. The anterior end is somewhat proboscis-like and flexible, but is not sharply demarcated as in Lionotus. The central portion of the body is developed into a more or less arched dorsal mass, which usually contains the nuclei and contractile vacuoles. As a result of this local thickening, the body is surrounded by a thin hyaline margin. This, however, may be absent on the right side in some species. The mouth reaches from the anterior extremity to a short distance from the end, and usually approaches the left edge. An anus is present near the posterior end of the dorsal swelling. Trichocysts are numerous on the ventral surface, and often on the dorsal surface, where they are inclosed in minute papilla-like swellings. Cilia-distribution controverted. Maupas and Bütschli hold that ventral surface alone is ciliated; others (Kent and Dujardin) that cilia are uniformly distributed. The entire body, dorsal and ventral surfaces alike, are uniformly striated. The contractile vacuole lies posteriorly, on the right side and in the dorsal swelling. In the fresh-water form L. meleagris, it is connected with a long canal whose swellings are frequently taken for additional contractile vesicles (Bütschli); in the marine form described below the canal is not developed and a series of vacuoles takes its place; these are all contractile. The macronucleus may be single, double, quadruple, band-formed, or rosette-formed. Movement is steadily progressive and peculiarly gliding. Fresh and salt water.
Loxophyllum setigerum Quenn. '67.
Synonyms: Litosolenus armatus Stokes '93; Litosolenus verrucosa Stokes '93.
The body is flattened, irregular in outline, obtusely pointed anteriorly, the point being turned to the right; rounded posteriorly. The left edge is nearly straight, the right considerably arched with a few setæ on the posterior half. Contractile vacuoles are numerous, dorsal in position and on the right side. The macronueleus is beaded, the several spheres connected.
Variety armatum (Cl. & Lach.) Fig. 32.
Under the name Litosolenus armatus, Stokes described a form from brackish water near New York, which should unquestionably be referred to the genus Loxophyllum, and I believe to Quennerstedt's species setigerum. While the latter possesses only a few setæ, the former has a number of them, and Stokes described his species as having a variable number. For this reason I include the Woods Hole form under the tentative name armatum, as a variety of Quennerstedt's L. setigerum. The flat margins are distinctly striated longitudinally, and faintly marked radially, on the dorsal surface. Longitudinal elevated striæ also run the length of the dorsal hump and upon the entire ventral surface. The ventral surface is alone ciliated. Upon the edges of the flat border are sharp-pointed, colorless, spine-like processes, situated at equal distances around the entire periphery except at the anterior end. Each spine is thick at the base and tapers to a full point which is curved upward—i. e., dorsally (fig. 32, a, b). The entire body is plastic and contractile, turning its leaf-like edge readily over objects upon which it creeps. The cilia are fine and uniform, with a tendency to lengthen in the oral region.
Length 100µ; greatest width assumed on contraction 85µ; when normal about 50µ.
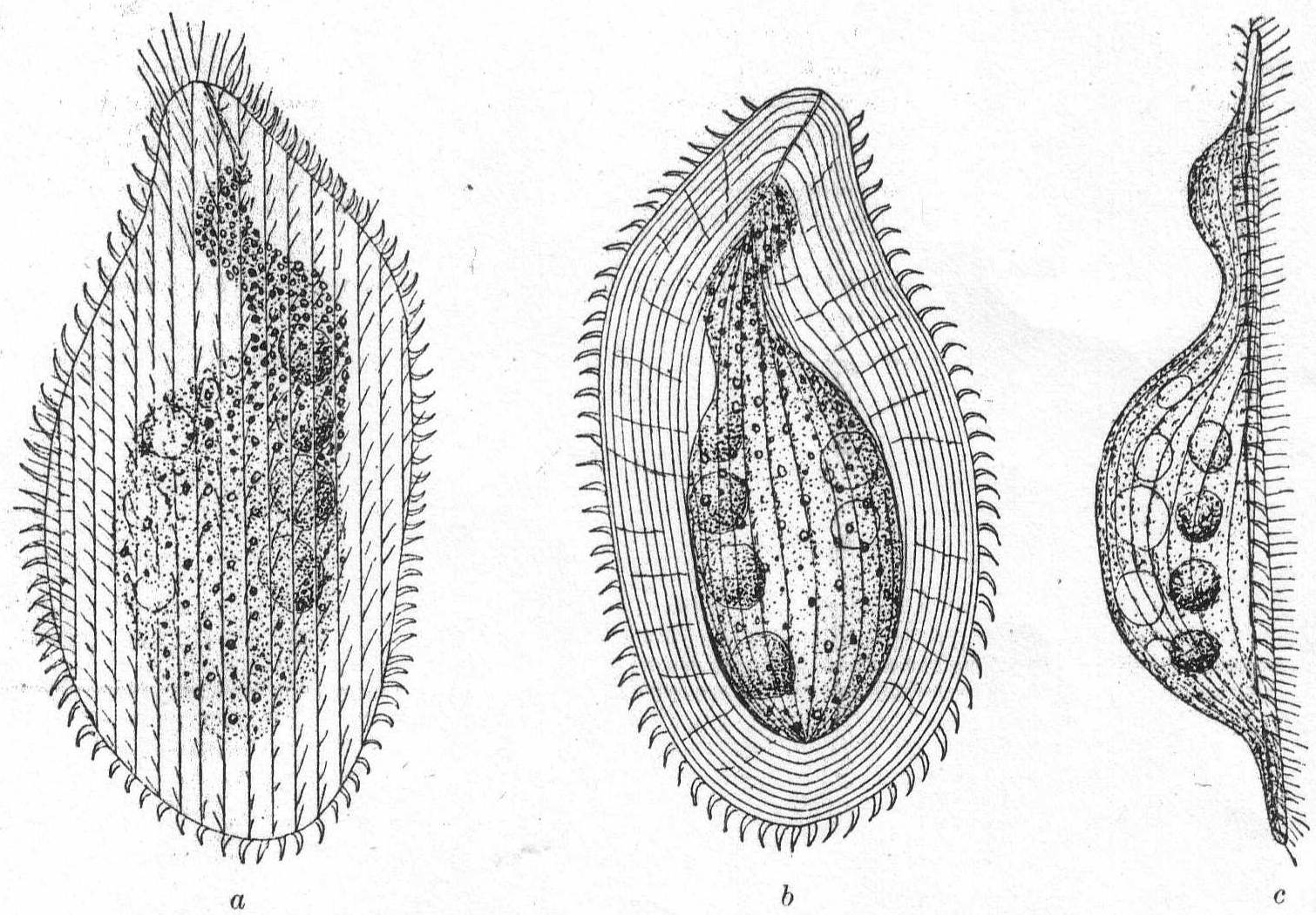
Fig. 32.—Loxophyllum setigerum, var. armatum. a, b, c, ventral, dorsal, and lateral aspects. ENLARGE |
The body is elongate and somewhat lance-shaped, widest at the central part and tapering to a point at the anterior end. The posterior end may be similarly tapered or rounded. The anterior end frequently proboscis-like, flat, and flexible, while the entire body is more or less elastic and contractile. The right side is flattened and alone provided with cilia, while the left side of the body proper is arched; on the left side of the proboscis is a row of coarse cilia resembling an adoral zone, and a row of trichocysts. A long peristome stretches down the thin, ventral side of the proboscis, and the mouth proper is situated at the junction of the proboscis and body; the mouth, as a rule, is invisible. The ciliated right side alone is striated in the majority of species. The contractile vacuole may be single or multiple, usually in the posterior region of the body and dorsal in position. The macronucleus is usually double, rarely single or quadruple, but may occasionally break into numerous smaller pieces. Movement, free-swimming or gliding, with especial tendency to get under clumps of foreign matter.
Fresh and salt water.
Lionotus fasciola Ehr. Fig. 33.
Synonyms. Amphileptus fasciola Ehr. '38;
Dujardin '41; Lachmann '56; Cohn '66, Diesing '65.
Loxophyllum fasciola Claparède & Lachmann '58;
Balbiani '61.
Loxophyllum duplostriatum Maupas '83. Shevyakov '96.
Body frequently brown or brilliant yellow in color, somewhat sigmoid in form with tapering anterior end, the extremity of which is turned dorsally. The proboscis is about half the entire length and is not sharply marked from the rest of the body but tapers gradually, its base being equal to the diameter of the body at its middle point. The body is slightly contractile and the posterior end is carried to a rounded point, but not into a distinct tail. Unlike the fresh-water variety, this one has no hyaline margin nor hyaline caudal region, and the contractile vacuole is double or multiple on the dorsal side near the posterior end. Cilia are present only on the under (right) side, with, however, a row of large cilia marking the course of the elongate mouth, upon its left side. The right side is striated, the left arched and without markings. The endoplasm is finely granular with, however, larger food particles in the process of digestion, while specimens are occasionally seen with the natural form completely lost through distortion caused by over-large captures (Cf. also Wrzesniowski '70, p. XXIII, fig. 32). Movement continuous, slow, and gliding; very little tendency to jerking movements. Macronucleus double, both parts spherical, and placed in about the center of the larger part of the body; closely approximated but not, as Schewiakoff described, connected. In conjugation, a large form unites with a smaller one, the mouth parts being connected. Details of conjugation and macronuclei not made out. Length 200µ to 600µ.
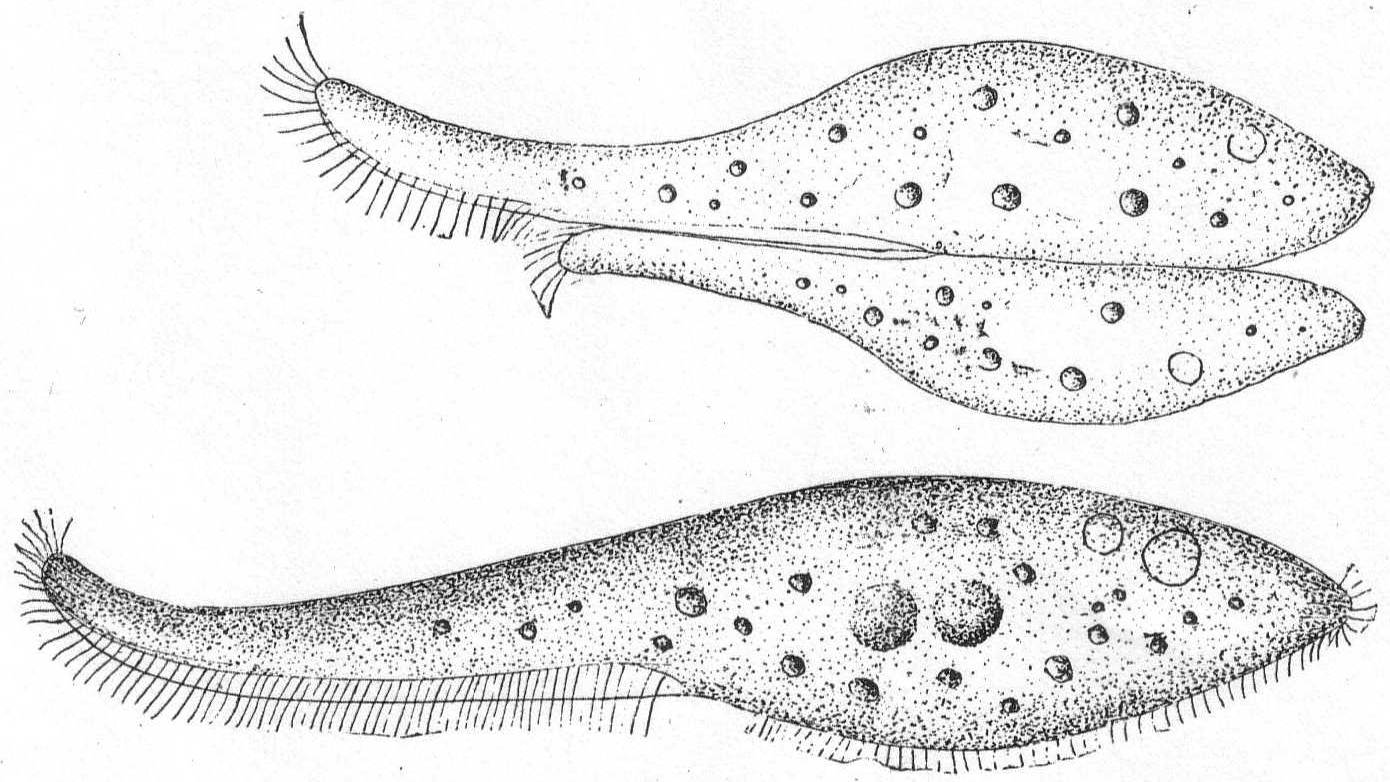
Fig. 33.—Lionotus fasciola. ENLARGE |
| Key to marine genera of Chlamydodontidæ. | |||
| Diagnostic characters: Form usually ellipsoid, never very elongate. Transverse section of body circular or elliptical. The mouth is usually some distance from the anterior end and may be in the posterior part. Sometimes it is in the center of the ventral surface, again on the right side. The œsophagus invariably has a well-developed buccal armature, or a smooth peculiarly built œsophageal tube. Food particles of large size. | |||
| 1. | Body cylindrical. Cilia about entire body | Genus *Nassula | |
| Body flat | 2 | ||
| 2. | a. | Without a caudal process | 3 |
| b. | With a caudal process | 5 | |
| 3. | a. | Anterior end angular | 4 |
| b. | Anterior end rounded | Genus Chlamydodon | |
| 4. | a. | Dorsal striæ and cilia present, ventral cilia longer | Genus Orthodon |
| b. | Dorsal striæ and cilia absent; posterior end not pointed | Genus *Chilodon | |
| c. | Dorsal striæ and cilia absent; posterior end pointed | Genus Scaphidiodon | |
| 5. | a. | Caudal spine with posterior bristle-like cilia | 6 |
| b. | Caudal spine without posterior bristle-like cilia; ventral cilia reduced | Genus Trochilia | |
| 6. | a. | With pigment spot on anterior angle | Genus Ægyria |
| b. | Without such pigment spot | Genus Onychodactylus | |
| c. | Cilia on right edge only of greatly reduced ventral surface | Genus *Dysteria | |
| * Presence at Woods Hole indicated by asterisk. | |||
The body is ovoid or cylindrical, with well-rounded ends, and in some cases slightly flattened. The mouth is ventral and placed some distance from anterior end (1/4 to 1/3 total length). A slight depression on the ventral surface marks the mouth region, which is further indicated by larger and more powerful cilia. The rest of the body is uniformly ciliated. The entire body is marked by clearly defined spiral stripes. The mouth is circular and the œsophagus is supported by a considerable armature, which usually extends dorsally and to the left, rarely to the right. In some cases the structure of this armature is indistinct; again it can be clearly seen to consist of definite rods (Stäbchen). The anus is probably always terminal. Contractile vacuoles are variable in different species. In some cases there is but one, which is placed at the posterior end or centrally on the ventral side; in others there may be four—two dorsal and two ventral. In many cases trichocysts are uniformly distributed. Sometimes the body is colorless; again, and more often, it is brightly colored with red, blue, brown, or black pigment. The macronucleus is globular and central, occasionally band-form and with numerous attached micronuclei. Food substance varied, usually vegetable matter, see, however, below. Cysts are globular. Movement is a steady progression, combined with rolling.
Nassula microstoma Cohn '66. Fig. 34.
Synonyms: Paramœcium microstomum Cl. et Lach. '58, Gourret et Roeser '88; Isotricha microstomum Kent '81.
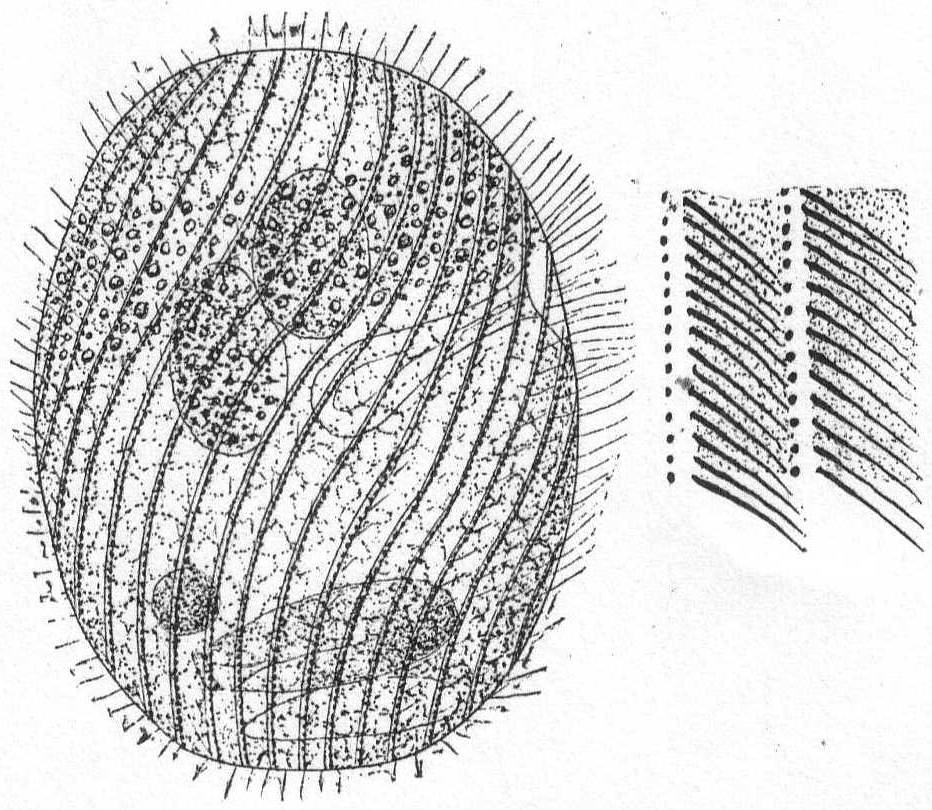
Fig. 34.—Nassula microstoma. ENLARGE |
Body subcylindrical, rounded at each extremity, not quite twice as long as broad. A slight depression on one surface marks the position of the mouth, this depression being indicated by a row of longer cilia. The mouth is extremely small and is surrounded by a curious buccal armature. This is not made up of bars or rods, as in most species of Nassula, but appears perfectly smooth and uniform except for the considerable swelling at the inner end. The cuticle is firm and unyielding and marked by longitudinal and somewhat spiral rows of cilia and trichocysts. Under the microscope this is one of the most pleasing forms found at Woods Hole. Its color is yellowish brown from the presence of brilliant particles of coloring matter held in the cortical plasm, and, as it slowly rolls along, these particles and the black trichocysts give to the organism a peculiar sparkling effect. The macronucleus is almost central; the contractile vacuole posterior. The endoplasm appears well filled with food bodies, some of which could be distinguished as Amphidinium and Glenodinium.
Length 55µ; greatest diameter 30µ.
Small forms, greatly flattened dorso-ventrally and almost egg-form in outline. The anterior end is bent distinctly to the left and forms a characteristic process, which, together with the entire margin of the body, is soft and flexible. The posterior end is, as a rule, broadly rounded. The ventral surface is finely striate, and this surface alone is ciliated. The lines of cilia converge at the mouth, and at this region the cilia are somewhat larger and more distinct, thus forming a functional adoral zone. The mouth is median and is situated in the anterior half of the body. It is surrounded by a well-defined armature, composed usually of from 10 to 16 rods. The contractile vacuoles are quite varied and from one to many in number, the number increasing with the size of the individual. The macronucleus is usually single, elliptical in form, and centrally placed; one micronucleus. Reddish granular pigment and trichocysts are occasionally present.
Chilodon cucullulus Müll., sp. Fig. 35.
Synonyms; Colpoda cucullus O. F. Müller; Loxodes cucullulus; Chilodon uncinatus Ehr. '58, Perty '52, Dujardin '41; L. dentatus Duj., etc.
This extremely variable form has received so many different names that it hardly pays to enumerate them. It is one of the commonest and most widely spread ciliates known, although at Woods Hole I was surprised to see it so rarely. It is the type species of the genus and needs no further description. The specimens observed at Woods Hole had numerous contractile vacuoles and were 42 to 45µ long and from 28 to 32µ wide.
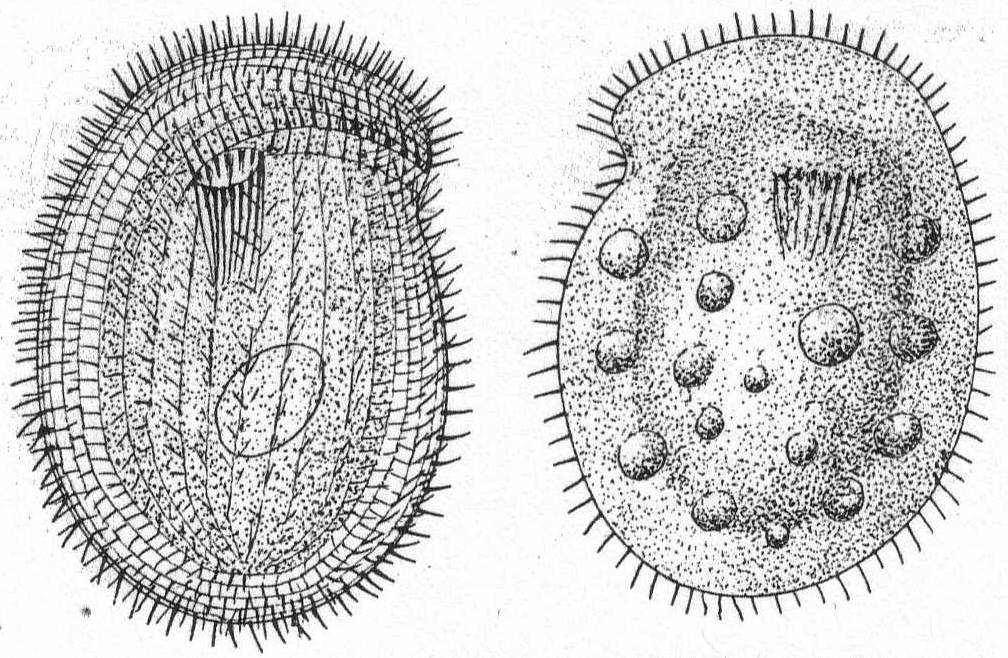
|
Fig. 35.— Chilodon cucullulus. ENLARGE |
Small forms, firm in outline, and colorless or slightly colored. The body is somewhat clam-shaped, flattened, slightly curved or straight on the right side, the other more convex. The true ventral side is only a narrow strip along the right and anterior edge of the body, the apparent ventral side being a fold of the very large dorsal surface which comes around ventrally, forming a valved structure somewhat analogous to a clam shell. Cilia are limited to the outer edge of the small ventral surface, which also bears a peculiar spine at the posterior end. Behind this spine are larger cilia. The mouth opening lies in the anterior widened portion of the ventral surface and is connected with a smooth tubular pharynx. The right half of the dorsal side, i.e., the apparent dorsal side, is arched and bears longitudinal ridges. Two to four contractile vacuoles are placed on the ventral side. The macronucleus is usually dorsal, elliptical, and cleft, with one micronucleus attached. Fresh and salt water.
Dysteria lanceolata Cl. et Lach. Fig. 36.
Synonym: Cypridium lanceolatum Kent '81.
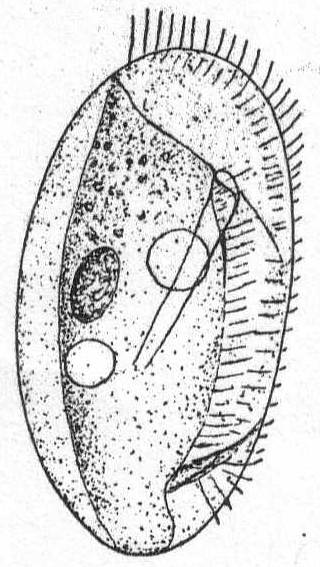
Fig. 36.— Dysteria lanceolata. ENLARGE |
Outline of the flattened body ovoid; body consists of two valve-like portions; the edge of the right valve is nearly straight, that of the left valve more or less sinuous; anteriorly it is cut away, obliquely and posteriorly it has a deep indentation in which the seizing spine rests. The cilia are confined to the ventral surface, here reaching, however, from the anterior dorsal extremity to below the posterior indentation. Posteriorly the cilia become larger, corresponding to the larger cirri of D. armata, which are posterior to the spine. The mouth lies between the two valves and is surrounded by a long and smooth buccal armature which passes downward and backward to the left a distance equal to about half the entire body length. The macronucleus is situated in the dorsal region in the central part of the body. There are two contractile vacuoles, one behind the center of the buccal armature, the other near the inner end of this organ. Movement is in circles, the animal moving around quite rapidly when not attached by its posterior process. It is colorless and measures 45µ in length by 27µ in width. Claparède & Lachmann and Shevyakov describe it as 70µ long.
| Key to marine genera of Chiliferidæ. | ||
| Diagnostic characters: Mouth never lies behind the middle of the body; the œsophagus is but slightly developed. The undulating membranes are placed either on the edge of the mouth or in the œsophagus. A peristomial depression leading to the mouth is absent or very slightly indicated. | ||
| 1. | Mouth in the anterior half, undulating membrane on left edge only; right edge continued in a long ventral furrow | Genus *Frontonia |
| 2. | Two undulating membranes; mouth central; no caudal bristles | Genus *Colpidium |
| 3. | Two undulating membranes; caudal bristle | Genus *Uronema |
| * Presence at Woods Hole indicated by asterisk. | ||
Form elongate and cylindrical, or often flattened dorso-ventrally, with round or pointed ends. It is usually plastic and contractile. Cilia are evenly distributed about the body and are similar in length. The large, open mouth lies on the anterior half of the ventral surface, and is elongate and oval in outline. On its left edge is a well-defined membrane which stretches across to the right side of the mouth. On the right edge is a small, longitudinally striped tract which is free from trichocysts and smooth in appearance. This tract is continued posteriorly in a long furrow, which in some cases reaches the posterior end of the animal. A few rows of cilia in this furrow vibrate differently from the others and give the effect of a membrane (Bütschli). The œsophagus is extremely short and hard to make out. The body is usually covered uniformly with trichocysts, often of considerable size. There are 1 or 2 vacuoles with long canals radiating throughout the endoplasm. The macronucleus is oval and centrally placed. Micronuclei vary from one to many. An anal opening is placed at the end of the long ventral furrow. The plasm is colorless or green by the presence of Zoochlorella, or colored brown or black by pigments. In these cases there is a considerable pigment mass on the anterior end. Movement is regular, forward, and combined with rotation. Food consists of foreign objects, diatoms, other protozoa and the like. Fresh and salt water.
Frontonia leucas Ehr. Fig. 37.
Synonyms: Frontonia vernalis Ehr. '38; Bursaria leucas Allman '55, Carter '56; Panophrys leucas Duj. '41, Stein '67; Panophrys vernalis Dujardin '41, Stein '67; P. chrysalis Duj. '41, Fromentel '74; Cyrtostomum leucas Stein '67, Kent '81.
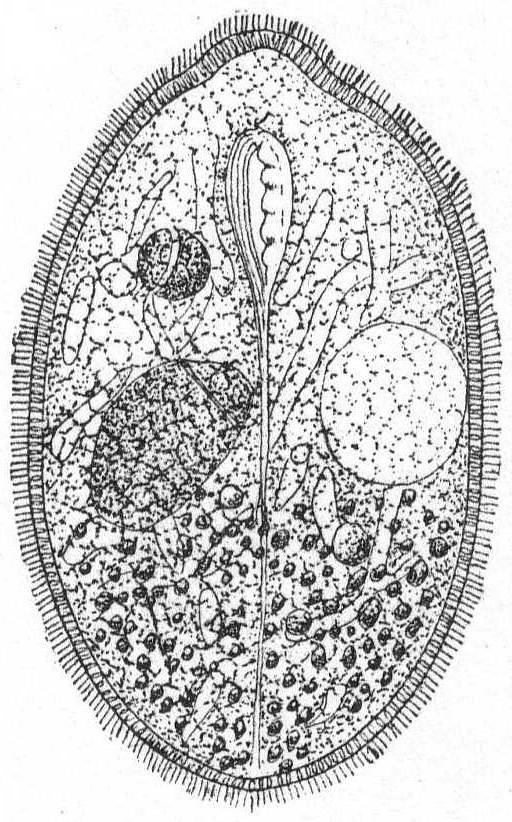
Fig. 37.— Frontonia leucas. ENLARGE |
Form ovoid, elongate, occasionally a little flattened dorso-ventrally. Mouth in the anterior third of the body. The left edge of the mouth carries a distinct undulating membrane; the right edge is plain, longitudinally striated and bears cilia. It is slightly depressed and the depression is carried posteriorly in the form of a shallow furrow which reaches to the posterior end. The contractile vacuole is on the left side, the spheroidal nucleus on the right side of the furrow. The body is uniformly covered with fine cilia, and the periphery is uniformly studded with large trichocysts, except along the furrow. Food consists of dinoflagellates and other small forms. Color dark brown to black.
Length 330µ; width 200µ.
This form differs considerably from the fresh-water Frontonia leucas as described by Schewiakoff '89, especially in the extreme length of the peristomial furrow, in the position of the nucleus and contractile vacuole, and in the nature of the water canals. These in the Woods Hole form are very irregular in size and very much branched, not uniform as in Lieberkühn's (see Bütschli) figure of Frontonia leucas, nor radiating as in Schewiakoff's description. This may be the same species as Frontonia marina, of Fabre-Domergue '91, whose description and figure I have not seen.
The general form is oval, slightly compressed laterally with the dorsal side strongly arched. The ventral side is slightly incurved. The anterior end is somewhat smaller than the posterior end, which is broadly rounded. The mouth is placed some distance from the anterior end in an oral depression and opens into a tubular œsophagus. There are usually two undulating membranes which do not extend beyond the mouth borders. The right undulating membrane extends down into the œsophagus and appears to be attached to the walls of the latter. The body stripes in front of the mouth are twisted to the left. The anus is terminal and the contractile vacuole may be terminal or situated forwards in the dorsal region. The macronucleus is spherical and has one micronucleus attached. Food consists mainly of bacteria. Movement rapid, but interrupted.
Fresh and salt water, common in infusions.
Colpidium colpoda Ehr., sp. Fig. 38.
Synonyms: Colpidium cucullus Kent '81; C. striatus Stokes '85; Kolpoda cucullus Duj. '41; Paramœcium colpoda Ehr. '38, Quennerstedt '67; Plagyiopyla nasula Kent '81, G. & R. '86; Glaucoma pyriformis G. & R. '86; Tillina campyla Stokes '85, '88.
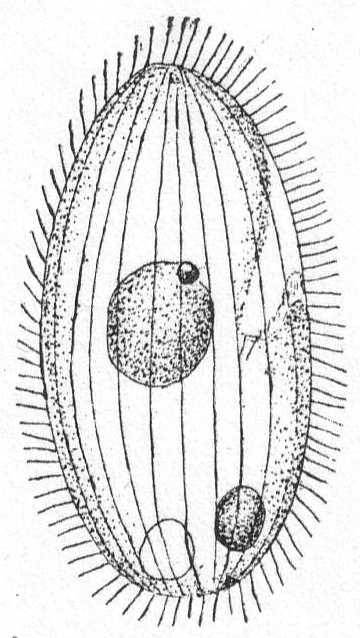
Fig. 38.— Colpidium colpoda. ENLARGE |
The body is oval, somewhat larger posteriorly, and a little compressed dorso-ventrally. The anterior end is twisted a little from the right to the left (more evident in fresh-water forms), and leans somewhat toward the ventral side. Under this portion, on the ventral side, lies the mouth in a large depression just above the middle of the body. The entire body is covered with uniform and delicate cilia, which are placed in longitudinal rows. These rows are almost straight on the dorsal side, but bend on the ventral surface, following the contour of the twisted anterior portion. The endoplasm is finely granular; the œsophagus leading into it is very distinct. Schewiakoff ('89) describes two membranes, an inner and an outer; Maupas ('83) describes them as right and left. In the present species I was able to make out only one. The macronucleus is central, spherical in form, and bears a single minute micronucleus. The contractile vacuole is posterior and dorsal to the long axis of the body. The anus is ventral to this axis and also posterior. Length 45µ, width 20µ. Common.
This marine variety is much smaller than the fresh-water form and the form differs in a number of respects, viz, in the anterior torsion and in the structure of the mouth. These may be, however, only individual variations of a widely spread species, and I believe it is perfectly safe to describe this as Colpidium colpoda.
Minute forms; colorless and constant in body form. The form is oval, slightly compressed on the ventral side, while the dorsal side is distinctly arched. The membrane is distinctly marked by rather widely separated striæ. These occasionally have a spiral course about the body; in all cases they can be easily counted. The mouth is large and placed near the center of the ventral surface. It is sometimes approached by a very shallow depression or peristome from the anterior end, and marked by two rows of cilia. An undulating membrane extends down the mouth. Oesophagus absent. A long, stiff bristle extends outwards from the posterior end. The contractile vacuole is terminal or subterminal and near the anal opening. The macronucleus is spherical, centrally placed, and with one micronucleus closely applied. Movement is rapid and usual forwards in a straight line, often found resting, however, with outstretched cilia in contact with some foreign body. Food mainly bacteria. Fresh and saltwater, usually in decomposing vegetable substances.
Uronema marina Duj. Fig. 39.
Synonyms: Enchelys triquetra Dujardin; E. corrugata Duj.; Cryptochilum griseolum Maupas '83; Philaster digitiformis Fabre-Domergue '85.
Small animals with ellipsoidal form and about twice as long as broad. The mouth lies in the upper half of the body and bears a well-developed undulating membrane upon its left edge. The membrane is longitudinally striped and covered with long and vibratile cilia. The right edge of the mouth bears cilia which are about the same in size as the body cilia, but are more closely inserted (Schewiakoff). The most characteristic feature is the long caudal bristle, which is extremely delicate and about two-thirds the length of the body. Schewiakoff thinks this bristle has a sensory function. I could not make this out, for although other protozoa ran against this bristle, often bending it well over to one side, the animal showed no sign of irritability but lay quiescent. A spherical macronucleus with attached micronucleus lies in the center of the body. The contractile vacuole is posterior in front of the bristle. The macronucleus was found to be double, as though just divided, in a large percentage of cases. This may be a precocious division of the nucleus long before signs of the body division are evident. Such a phenomenon, however, is rare, the macronucleus usually dividing at a late stage of cell division. Length 30 to 50µ; width 15 to 20µ. Common in decomposing algæ.
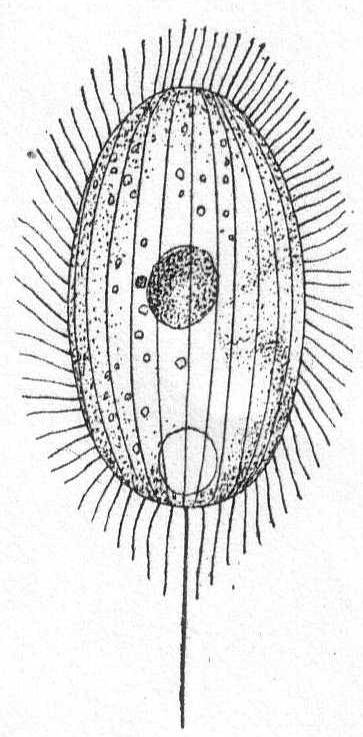
|
Fig. 39.— Uronema marina. ENLARGE |
| Key to marine genera of Pleuronemidæ. | |
| Diagnostic characters: The mouth is at the end of a long peristome running along the ventral side; the body is dorso-ventrally or laterally compressed. The entire left edge of the peristome is provided with an undulating membrane which occasionally runs around the posterior end of the peristome to form a "pocket" leading to the mouth. The right edge of the peristome is provided with a less-developed membrane. There may or may not be a well-developed pharynx. | |
| Body small; not produced into neck-like elongation | Genus *Pleuronema |
| Body medium-sized; anterior end produced into neck-like elongation | Genus *Lembus |
| * Presence at Woods Hole indicated by asterisk. | |
Small to medium-sized ciliates, with an unchanging form. They are somewhat lens-shape and laterally compressed, the two surfaces about equally arched. The ventral surface is nearly straight or but slightly arched; the dorsal is quite convex. The anterior and posterior extremities are equally rounded. The peristome begins as a small depression, but becomes larger until it takes in nearly all of the ventral surface. The depression becomes much deeper at about the center of the body, and is especially marked on the left side of the peristome. In this deeper portion is the mouth, with an almost imperceptible œsophagus. Upon the left edge of the peristome is a high, undulating membrane, sail-like in appearance when extended. This may stretch around the posterior edge of the peristome and upon the right aide, thus forming a pocket by means of which the food particles are directed into the mouth. The rest of the right edge of the peristome is occupied by closely approximated powerful cilia (Bütschli) or a second undulating membrane (Stein). The body cilia are relatively long. Trichocysts and caudal bristles may be present. The contractile vacuole is subterminal and dorsal; it is questionable whether there are canals leading to it. A round macronucleus with one micronucleus is in the anterior half of the body. The anus is terminal. Food is chiefly bacteria. Movement combines springing with swimming and rotation. Fresh and salt water.
Pleuronema chrysalis Ehr., sp. Fig. 40.
Synonyms: Pleuronema crassa Dujardin '41; P. marina Duj. '41; Fabre-Domergue '85; P. coronata Kent '81; Paramœcium chrysalis Ehr. '38; Lembadion orale G. & R. '88; Histiobalantium agile Stokes '85, '88.
The body is ovoid, slightly flattened, rounded at both ends, the anterior end sharper than the posterior. The ventral surface is almost entirely taken up by a peristome which extends from the anterior end posteriorly three-quarters of the body length. The posterior end of the peristome is straight, the left curved, following the depressed portion. The body is covered with fine cilia in longitudinal lines, except on the peristome. The mouth is small and situated in the hollow of the peristome near the left border. On the left peristome edge is a large undulating membrane. It begins near the anterior end of the body and increases in height posteriorly following the peristome edge around on the right side. This posterior bend of the membrane causes the appearance of a full sail, so often seen. It can be entirely withdrawn and folded together in the peristome. On the right edge of the peristome are large, powerful cilia. The contractile vacuole is central and dorsal; the macronucleus is in the anterior half of the body, with one attached micronucleus. Food consists of bacteria. Not very common. Fresh and salt water. It often remains quiet, with membrane and cilia outstretched, as though dead, but suddenly gives a spring and is gone.
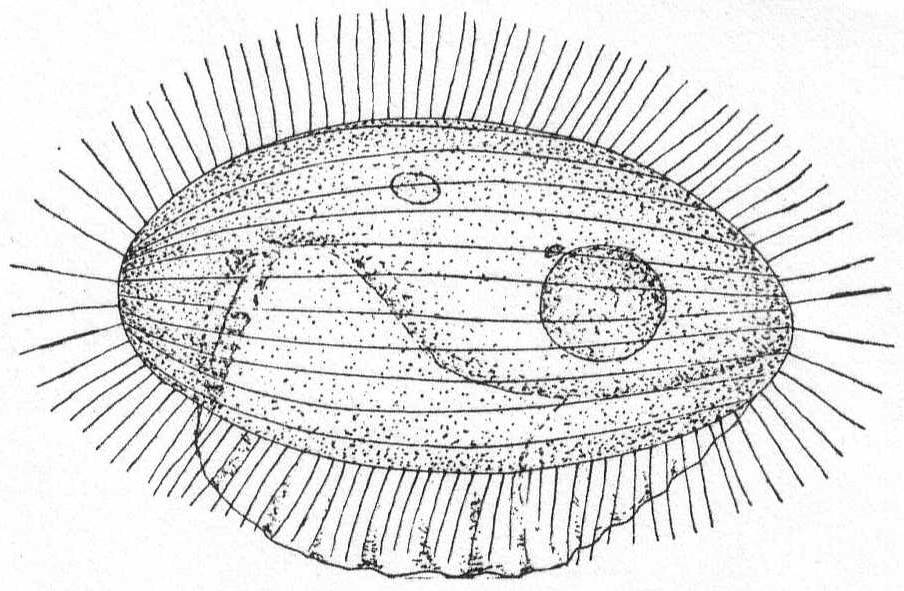
|
Fig. 40.— Pleuronema chrysalis. ENLARGE |
Pleuronema setigera, n. sp. Fig. 41.
Body colorless, elongate, and with the general form of a cucumber, the posterior end being somewhat pointed. The mouth and relatively small peristome are situated in the lower third of the body. The peristome begins as a shallow furrow at the center of the ventral surface and dips sharply into the buccal depression, which is deep and turned toward the posterior end. The left edge of the peristome bears a high undulating membrane, which extends anteriorly only as far as the center of the body; posteriorly it passes around to the right edge of the peristome, thus forming the characteristic membranous pocket. Inside the oral depression is a second undulating membrane, running down to the mouth. This is small and without an œsophagus. The body is clothed with long setose cilia which are frequently fully outstretched when the animal is resting, a slight tremor of the large membrane alone indicating vitality. Posteriorly these appendages are drawn out into long filiform setæ, the number varying in different individuals from three to nine or ten. These are extremely fine and difficult to see without a high power (e.g. 1/12 oil) and careful focussing of the substage condensor. Like P. chrysalis, the resting periods are terminated by sudden springs, otherwise the movements are steady and forward. The macronucleus is central, and the contractile vacuole posterior and terminal. Length 45µ to 50µ; greatest diameter 17µ. In decaying algæ.
It was this form, I believe, that Peck '95 described as a "ciliate."
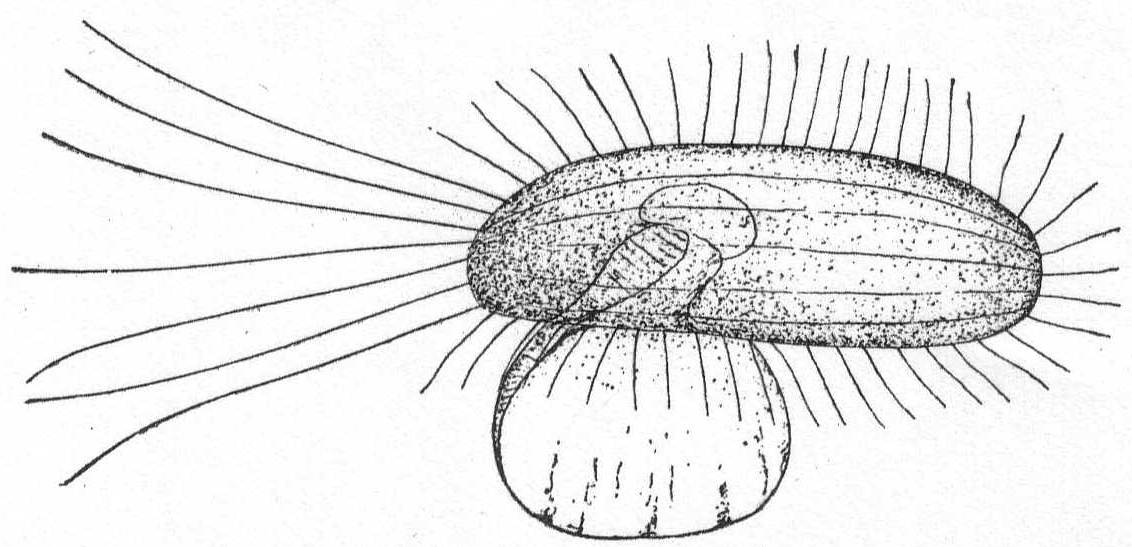
|
Fig. 41.— Pleuronema setigera. ENLARGE |
Free-swimming animals of elongate form, more or less elastic, and flexible, bending readily to avoid obstacles, etc. The anterior half is usually drawn out into a slightly curved neck-like portion. The peristome is a small groove leading from the anterior end to the mouth about midway down the ventral side of the body. Bütschli, following Quennerstedt, describes an undulating membrane on each side of the peristome groove. Other observers, however, usually describe but one, the left, which is clearly defined and stretches out some distance from the body, while the right border is described as having smaller but very active cilia. The general body surface is clothed with fine, uniform cilia, and body striæ are usually absent. One or more caudal bristles may be present. The contractile vacuole is posterior and terminal, and may be multiple. The macronucleus is spherical and perhaps double (Kent). Food is chiefly bacteria, and the animals are frequently found with the anterior end embedded in zooglœa masses. Salt water, usually in infusions.
Lembus infusionum, n. sp. Fig. 42.
The body is elongate, lancet-shaped, with a tapering anterior extremity. The dorsal outline is concave through the bending of the anterior end, while the ventral outline presents an even, convex curve. The mouth lies slightly above the center of the body and marks the posterior limit of the ventral peristomial groove, which curves slightly from the anterior extremity. Each side of this groove bears an undulating membrane, the left being much larger and conspicuously striated. The general form of this left membrane is triangular, the widest part is anterior, the narrowest at the mouth. The right membrane is similar in form, but smaller and more active. The endoplasm is colorless and finely granular, not regionally differentiated. The ectoplasm consists of a relatively thick cortical plasm specially noticeable in the posterior half of the body and a delicate cuticle which bears almost imperceptible longitudinal markings—the insertion points of the fine cilia. The body is covered with uniform cilia except at the anterior extremity. Here they are much larger and bristle-like. I was unable to find any cilia in the peristome. One long caudal bristle, one-quarter of the length of the body, trails out behind. The macronucleus is spheroidal and placed near the center of the body; a conspicuous micronucleus lies near it. A row of contractile vacuoles extends from the posterior end. I have seen as many as six of nearly equal size and one or two smaller ones. The intervals of contraction are quite long. Length 70 to 75µ; greatest diameter 10 to 12µ.
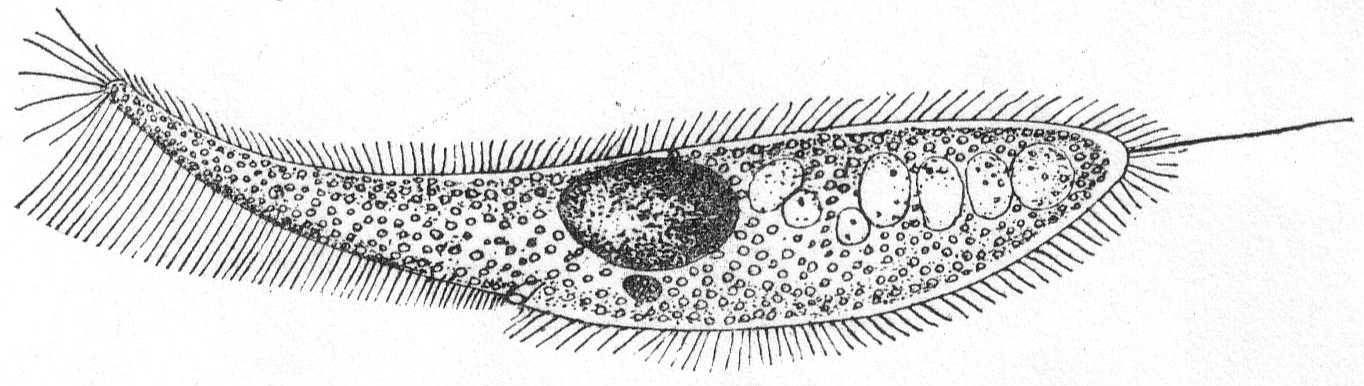
Fig. 42.—Lembus infusionum. ENLARGE |
L. infusionum resembles L. elongatus in its general form and in its mode of life, for it excavates a retreat in zooglœa masses and lies there for considerable periods perfectly quiet. It differs from L. elongatus and from L. velifer (probably the same as L. elongatus of Claparède & Lachmann) in the presence of the caudal bristle, in the absence of annular markings, number of contractile vacuoles, and in the slightly smaller size. It resembles Lembus verminus (Müller) as described by Kent (Proboscella vermina), and L. intermedius as described by Gourret & Roeser (Lembus verminus syn.)in the absence of annular markings and in the presence of a caudal bristle. It differs from the former, however, in the absence of a tentacle-like process, and from both in the absence of a double nucleus and in the presence of many vacuoles. These features are so characteristic of all the specimens examined that I have concluded, somewhat reluctantly, to give it a specific name. It is common in old infusions of algæ, especially after decomposition is well advanced. Its food consists of bacteria.
Lembus pusillus Quennerstedt 1869. Fig. 43.
Synonym: L. subulatus Kent 81.
This species is much smaller than the preceding, and might easily be mistaken for Uronema marina. It is subcylindrical in form, the anterior end bluntly pointed, the posterior end rounded. The oral apparatus is quite different from Uronema. The mouth, as in the preceding species, is at the end of a long peristomial groove extending from the anterior end to the middle of the body. The edges of the peristome bear undulating membranes as in L. infusionum. Like the latter, there is one caudal bristle, but unlike it there is only one posterior contractile vacuole, while the endoplasm is filled with large granules or food balls. The cuticle is distinctly striated with longitudinal markings, and the cilia are uniform in length.
Habitat similar to that of L. infusionum, in zooglœa masses. Length 26 to 30µ; diameter 7 to 8µ.
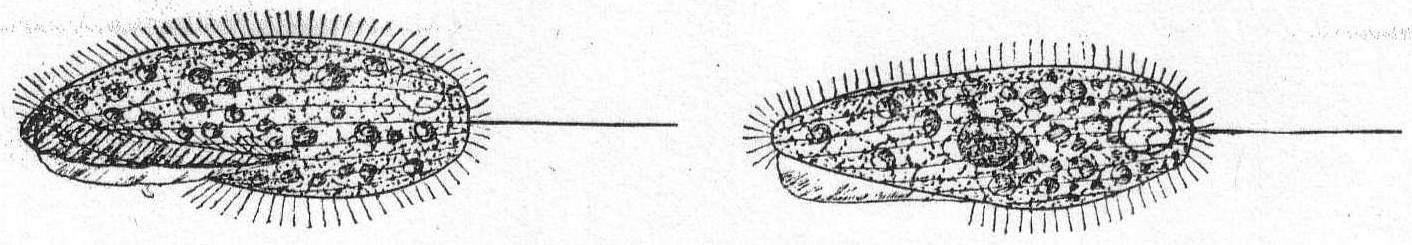
Fig. 43.—Lembus pusillus. ENLARGE |
Although Quennerstedt's description of L. pusillus makes no mention of a caudal bristle, the size and other characters are so closely similar that I hesitate to make a new species. The bristle is extremely delicate, scarcely thicker than a cilium, and easily overlooked, yet with proper focussing of the condenser I found it on every specimen examined.
| Key to marine genera of Opalinidæ. | |
| Diagnostic characters: The form is oval, and the body may be short or drawn out to resemble a worm. They are characterized mainly by the absence of mouth and pharynx. | |
| Anterior end not pointed; body cylindrical; tapering | Genus *Anoplophrya |
| Anterior end pointed; body elongate; cylindrical; tapering | Genus Opalinopsis |
| * Presence at Woods Hole indicated by asterisk. | |
The general form is elongate, cylindrical or slightly flattened, with rounded ends, the posterior end tapering. The body is striated with clearly defined, often depressed lines, which run longitudinally and sometimes spirally. The contractile vacuoles are usually placed in rows upon the edges. The macronucleus is almost always long and band-formed, rarely oval, and generally extending through the entire length of the body. Micronuclei have been made out in one case. Reproduction is effected by simple cross division or by budding at the posterior end, and is frequently combined with chain formation. The main characteristic is the entire absence of mouth and œsophagus, the animals being parasitic in the digestive tract of various annelids. Parasites, salt-water forms.
Anoplophrya branchiarum. Stein '52. Fig. 44.
A. circulans Balbiani.
The body is cylindrical to pyriform, in the latter case broadened anteriorly. Cuticle distinctly marked by longitudinal striations which take the form of depressions and give to the body a characteristic melon shape. The endoplasm contains a number of large refringent granules—probably body products. The nucleus is elongate, somewhat curved, and coarsely granular. A micronucleus lies in the concavity. The cilia are long, inserted rather widely apart along the longitudinal markings. The contractile vacuole is single and is located at the pointed end, which is directed backwards during locomotion. One specimen found free swimming among some algæ.
Length 104µ; greatest diameter 36µ.
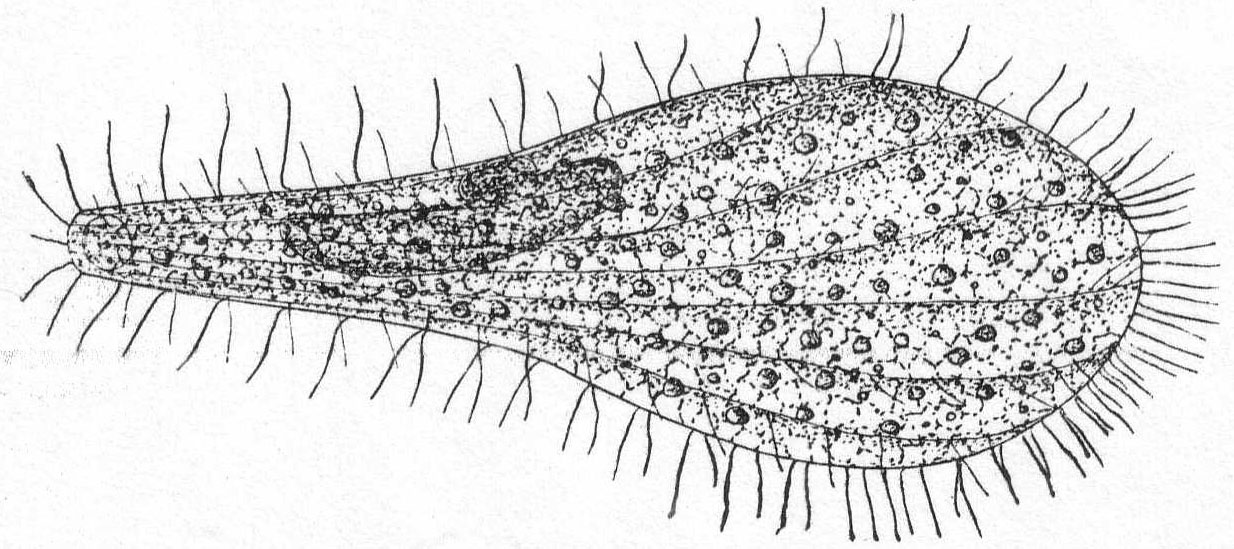
Fig. 44.—Anoplophrya branchiarum. ENLARGE |
I was much surprised to find this form swimming about freely in the water; its mouthless condition showed it to belong to the family of parasites, the Opalinidæ. As the name indicates, however, this species is an ectoparasite upon the gills, and Stein gave the name branchiarum to a fresh-water form parasitic upon Gammarus pulex. The Woods Hole form is so strikingly similar to the figure of G. branchiarum that, although the name was given to a fresh-water form, it obviously applies to this marine variety. One important difference is the presence of only one contractile vacuole in the marine form.
| Key to families of Heterotrichida. | |||
| Cilia cover the body | 1 | ||
| Cilia reduced to certain localized areas | 2 | ||
| 1. | Polytrichina. | ||
| a. | The mouth terminates a long peristomial furrow having an adoral zone along the entire left edge | Family Plagiotomidæ | |
| b. | Peristomial area a broad triangular area ending in mouth | Family Bursaridæ | |
| c. | Peristomial depression short; limited to the anterior end; its plane at right angles to the long axis of body; surface of peristome striated and ciliated; no undulating membranes | Family Stentoridæ | |
| 2. | Oligotrichina. | ||
| a. | Peristome without cilia; cilia limited to one or more girdles about body | Family Halteriidæ | |
| One marine genus | *Strombidium | ||
| b. | Thecate forms; the body is attached by a stalk to the cup; within the adoral zone is a ring of cilia. | Family Tintinnidæ | |
| c. | The peristomial depression is deep and funnel-like; cuticle thick, with posterior spine-like processes. | Family Ophryoscolecidæ | |
| * Presence at Woods Hole indicated by asterisk. | |||
| Key to the marine genera of Plagiotomidæ. | ||
| Diagnostic characters: The peristome is a narrow furrow which begins, as a rule, close to the anterior end and runs backward along the ventral side, to the mouth, which is usually placed between the middle of the body and the posterior end. A well-developed adoral zone stretches along the left side of the peristome, and is usually straight. | ||
| 1. | Body cylindrical; size medium; peristome long and turns sharply to the left at the extremity | Genus Metopus |
| 2. | No torsion in the peristome; undulating membrane is confined to the posterior part of peristome | Genus Blepharisma |
| 3. | No peristomial torsion; body highly contractile; no undulating membrane | Genus Spirostomum |
| Key to the marine genera of Bursaridæ. | ||
| Diagnostic characters: The body is usually short and pocket-like, but may be elongate. The chief characteristic is the peristome, which is not a furrow, but a broad triangular area deeply insunk and ending in a point at the mouth. The adoral zone is usually confined to the left peristome edge, or it may cross over to the right anterior edge. | ||
| 1. | The anterior half of the body tapers to nearly a point in front; the peristome is narrowest at the apex; the mouth is the entire peristome base. | Genus Balantidium |
| 2. | The anterior end does not taper; the peristome is widest at the end of the body; the mouth is clearly defined. | Genus *Condylostoma |
| * Presence at Woods Hole indicated by asterisk. | ||
Colorless and more or less flexible animals of medium size. The general form is elongate and cylindrical or somewhat smaller anteriorly. The posterior end is broadly rounded, the anterior end somewhat truncate and oblique. The peristome is broad and triangular, the base of the triangle being the entire anterior end of the body. The entire length of the peristome is one-fourth or less of the body length. The mouth is large and placed at the apex of the peristomial triangle and opens into a comparatively small œsophagus. The right edge of the peristome is lamellate and bears a clearly defined undulating membrane. The adoral zone is well developed upon the left edge of the peristome, from which it passes around anteriorly to the right edge. The surface of the peristome is free from cilia, but the rest of the body is uniformly coated with small active cilia. Contractile vacuoles are not safely determined. Bütschli thinks there is probably one terminal vacuole, but some observers deny this (e.g. Maupas). Others describe them on the dorsal side of the posterior end (Quennerstedt). The macronucleus is long and beaded and placed upon the right side. Micronuclei are numerous and scattered along the macronucleus. The anus is terminal and dorsal. Food consists of large and small particles. Movement rapid, free swimming, alternating with resting periods; in some cases an undulating or wriggling movement is seen, showing clearly the flexibility of the body. Fresh and salt water.
Condylostoma patens Müller. Fig. 45.
The body is elongate, somewhat sac-like, five or six times as long as broad, plastic, and frequently contains brightly colored food granules. The triangular peristome takes up the greater part of the anterior end, and the mouth is situated at the sharper angle of the triangle, about one-fourth of the total length from the anterior end. The cuticle is longitudinally striated, the lines having a slightly spiral course. They are not closely set, and fine cilia are thickly inserted along their edges. The endoplasm is granular and viscous. The motile organs consist of an adoral zone of membranelles, which stretch along the left edge of the peristome and the front edge of the body. The right edge of the peristome supports an undulating membrane. The nucleus is moniliform and extends the full length of the left side; a number of micronuclei are distributed along its course (Maupas).
Length 400µ; diameter at widest part 105µ. Maupas gives the length from 305µ to 495µ; and Stein 376µ to 564µ. Very common.
For a more extended account of the structures, see the excellent description by Maupas '83.
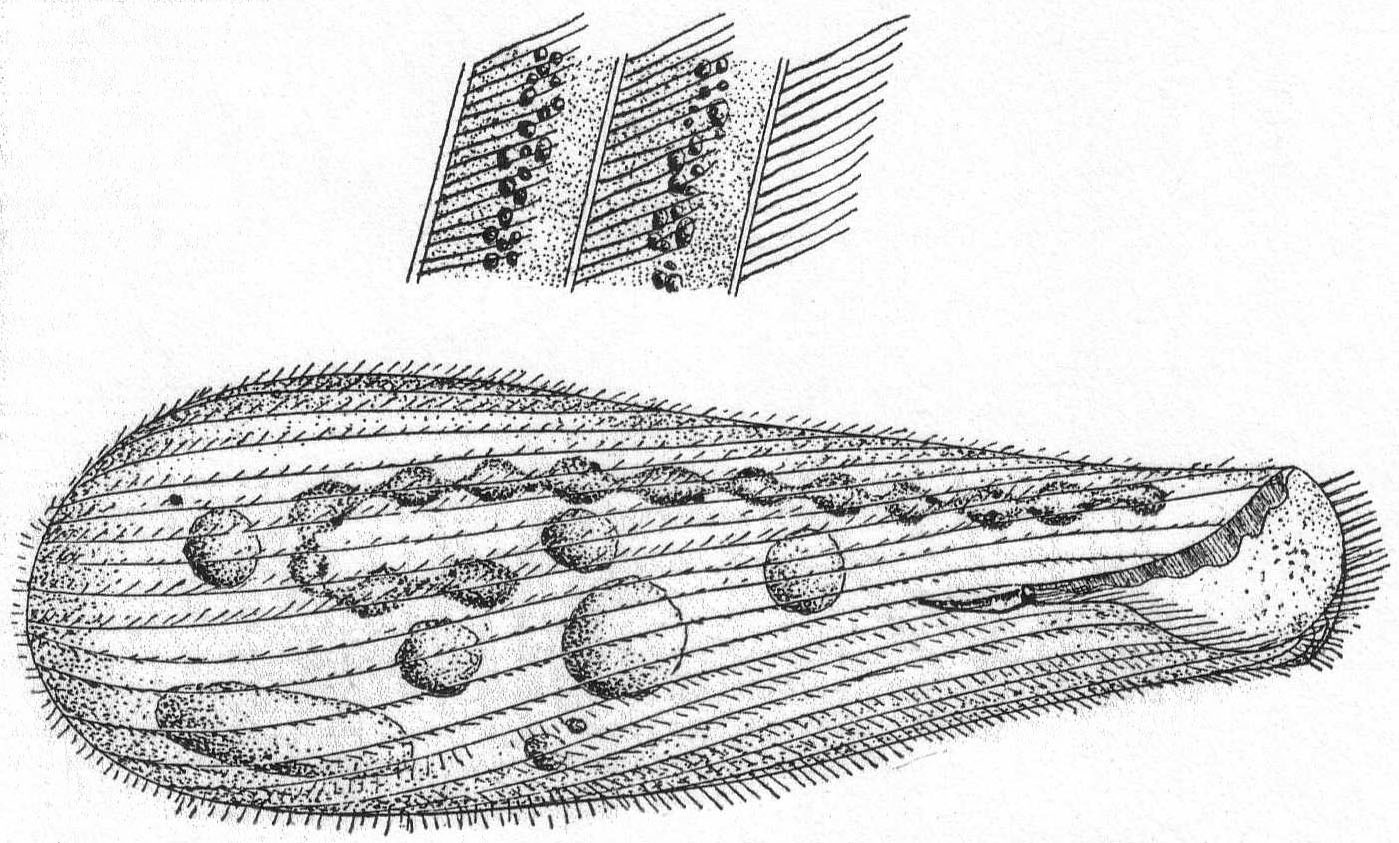
Fig. 45.—Condylostoma patens. ENLARGE |
| Key to the marine genera of Stentoridæ. | ||
| Diagnostic characters: The peristome is relatively short and limited to the front end of the animal, so that its plane is nearly at right angles to that of the longitudinal axis of the body. The adoral zone of cilia either passes entirely around the peristome edge or ends at the right-hand edge. The surface of the peristome is spirally striated and provided with cilia. Undulating membranes are absent. | ||
| 1. | Peristome circular in outline; limited to the anterior end | Genus Stentor |
| 2. | The peristome is drawn out into two wing-like processes; tube-dwelling | Genus Folliculina |
Small, colorless (except for ingested food) animals with characteristic springing movements. The form is usually constant, but in some cases may be plastic like Astasia; it is usually globular or conical, the posterior end being more or less pointed, the anterior end broadest. The latter is surrounded by a complete circle of the adoral zone, the oral end of which passes into a peristomial depression which extends deep into the middle of the body. The mouth, with a very small œsophagus, lies at the bottom of the inturned peristome. The region surrounded by the adoral zone is frequently drawn out into an anterior process, occasionally bearing a pigment mass. The ventral surface in some cases bears cilia, which may be distributed or restricted to a row of large cilia. Trichocysts are usually present and may be widely spread, limited to the posterior region, or arranged in a girdle about the middle. The contractile vacuole is simple, and posterior in position. The macronucleus is spherical and usually central in position. Movement is rapid swimming, combined with resting and floating periods, the latter usually terminated by a sudden leap.
Fresh and salt water; more common in the latter.
Strombidium caudatum Fromentel '74. Fig. 46, a, b, c.
Fromentel described a fresh-water form of this genus with a caudal appendage. The body is pyriform, broadly truncate on the anterior end, in the middle of which rises a papilliform process (Schnabel). On this process is a heap of pigment granules, which, however, are not constant. A ring of long cirri surround the anterior end and pass into the peristome, and from the left edge of this line of cirri a large adoral zone continues down to the mouth. The peristome is elongate and sac-form, and the mouth lies at the posterior extremity. With the exception of a caudal filament there are no other motile organs; this is about half as long as the body, structureless, hyaline, and sharply pointed. It splits up into a bundle of fine fibers upon treatment with caustic potash (c). The cirri emerge from minute hollows in the edge of the anterior border. The cortical plasm contains peculiar rod-like bodies, which look more like lines or markings than like rods or trichocysts. The nucleus is large, spherical, and placed in the center of the body. The contractile vacuole is posterior.
Length without appendage is about 35µ; greatest diameter 15 to 18µ. In decaying vegetable matter. Common.
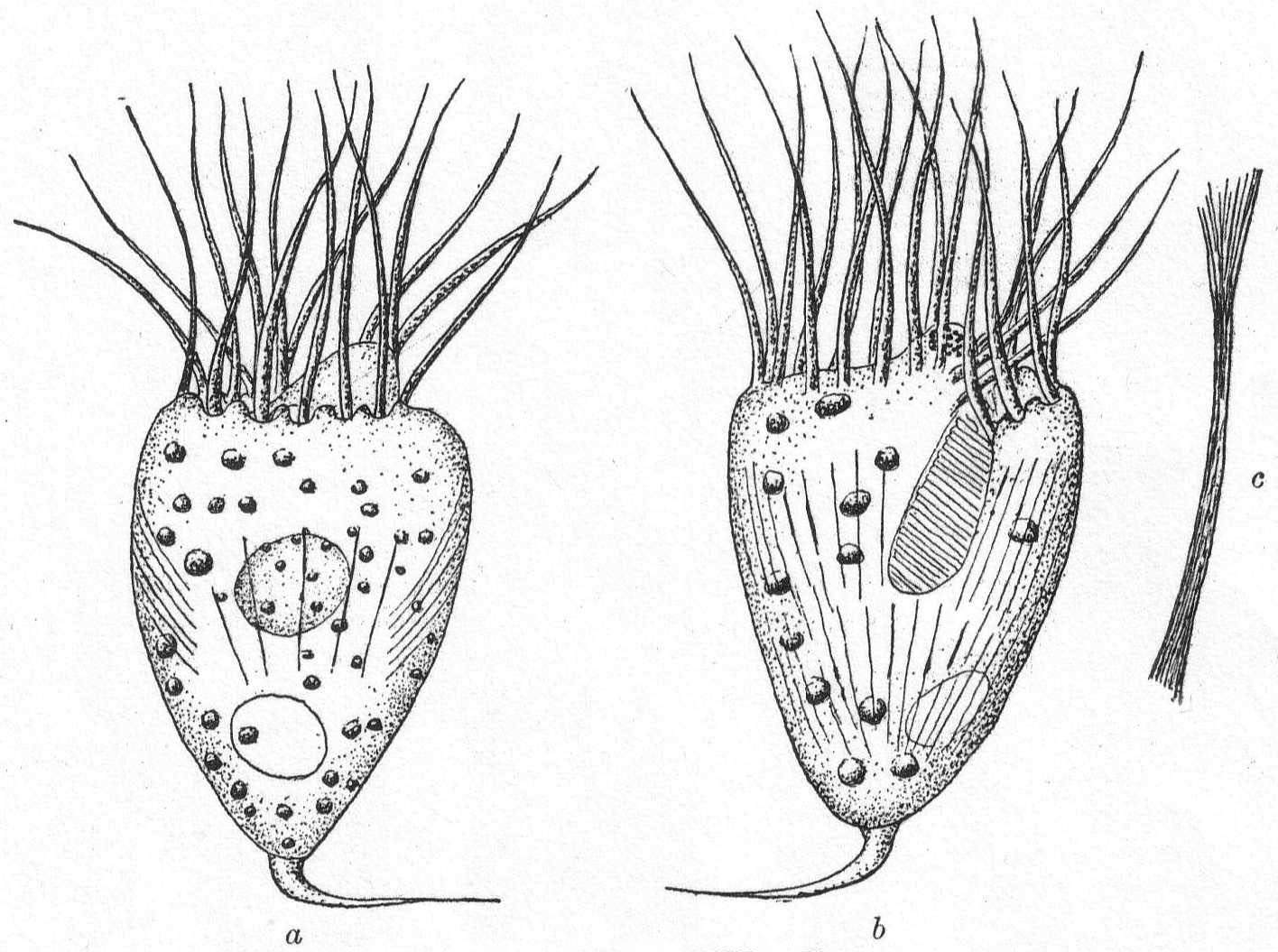
Fig. 46.—Strombidium caudatum. ENLARGE |
Although Fromentel's species is incompletely described, it is very evident that the organism corresponds fairly well with the Woods Hole variety. His was a fresh-water type; this is marine, but the caudal filament and the contractile vacuole are similar. Certainly in this case the organism can not be regarded as a Vorticella broken off its stalk, as Kent '81 suspected. The anterior process with its pigment spot; the cirri, the spherical nucleus, the position of the vacuole, etc., are all opposed to such an interpretation which Kent applied to the original species. Neither can it be a Tintinnoid. I place it provisionally as S. caudatum.
| Key to the marine genera of Tintinnidæ. | ||
| Diagnostic characters: Body attached by a stalk to a cup. Inside the zone of membranelles is a ring of cilia (par-oral). | ||
| 1. | The test is gelatinous and more or less covered by foreign particles | Genus Tintinnidium |
| 2. | The test is chitinous and clear. No foreign particles. | Genus Tintinnus |
| 3. | The test is chitinous; covered by foreign particles, growth rings frequent | Genus *Tintinnopsis |
| 4. | The test is chitinous, often covered by foreign particles. The test is marked by discoid, circular, or hexagonal spots. | Genus Codonella |
| 5. | The test is perforated by pores of circular or hexagonal form. | Genus Dictyocysta |
| * Presence at Woods Hole indicated by asterisk. | ||
Medium-sized ciliates, inclosed in a chitinous lorica with embedded sand crystals. The form of the house, or lorica, varies greatly. In some cases the mouth opening is wide, giving the lorica a bell form; it may be long and tubular, short and spherical, or variously indented. The animal is attached, as in the closely allied genus Tintinnus, by a peduncle to the bottom of the lorica. The anterior end of the animal is inclosed by two complete circles of cilia; one, the outer, forming the adoral zone, is composed of thick tentacle-like membranelles, the other consists of shorter cilia within the adoral zone. The mouth leads into a curved œsophagus containing rows of downward-directed cilia (Daday). The entire body is covered with cilia, but as the lorica is always opaque these can be made out only when the animal is induced to leave the house. The only difference between this genus and Tintinnus is the covering of foreign bodies—usually sand crystals. Movement is rapid and restless, and peculiarly vibratory, owing to the apparent awkwardness in moving the house. Salt water.
Tintinnopsis beroidea Stein, var. plagiostoma Daday. Fig. 47.
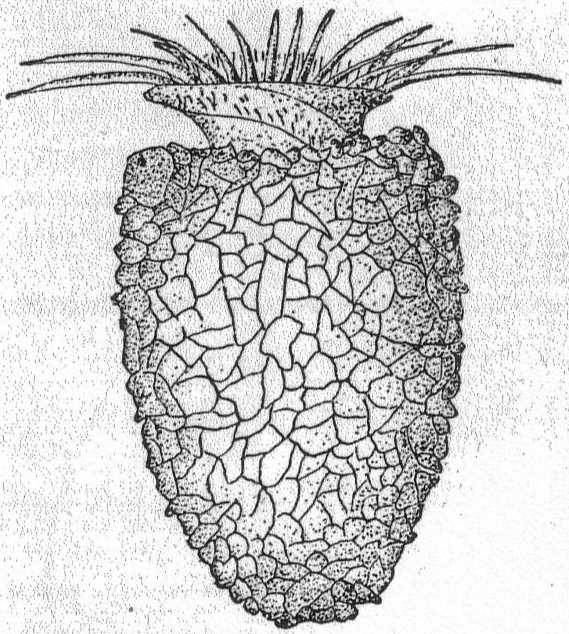
Fig. 47.— Tintinnopsis beroidea. ENLARGE |
Synonym: Codonella beroidea Entz '84.
The shell is colorless, thimble-shaped, with a broadly rounded posterior end. The body is cylindrical. The internal organs were not observed. Membranelles 24 in number. Length 50µ; greatest diameter 40µ.
Var. compressa Daday '87.
The posterior end of the shell is pointed, the lower third of the shell is swollen, the upper third is uniform in diameter and without oral inflation or depression. Nucleus not seen.
Length 70µ; greatest diameter 48µ.
Tintinnopsis davidoffi Daday. Fig. 48.
The shell is large, elongated, and provided with a considerable spine. The chitin of the shell is covered with silicious particles of diverse size. The internal structures were not observed.
Length of shell and spine 230µ; diameter of the oral aperture 54µ.
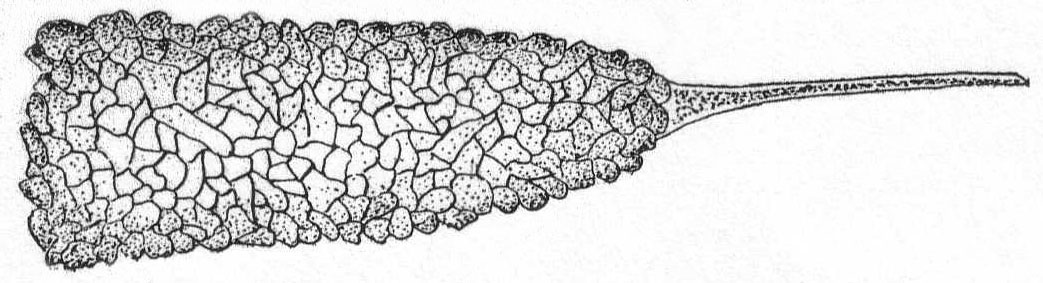
Fig. 48.—Tintinnopsis davidoffi. ENLARGE |
The variations of these species are considerable, and as the internal structures, such as the nucleus, are essential in fixing their systematic position, I place them as above, provisionally, and until further observations can be made.
| Key to families of Hypotrichida. | ||
| a. | Peristome indistinct; cilia on ventral surface uniform and not differentiated into cirri |
Family Peritromidæ One genus, *Peritromus |
| b. | Peristome more or less indistinct; cilia reduced to a few rows on the ventral surface; anal and frontal cirri present | Family Oxytrichidæ |
| c. | Cilia entirely reduced; frontal and anal cirri present or reduced; macronucleus band-formed or spherical | Family Euplotidæ |
| d. | Peristome reduced to left edge and does not reach over the anterior margin |
Family Aspidiscidæ One genus, *Aspidisca |
| * Presence at Woods Hole indicated by asterisk. | ||
The body is flat, colorless or tinged with yellow, and contractile. It is elliptical in outline, with broadly rounded ends; in some cases the left edge is slightly incurved, the right edge convex. The ventral surface is flat, the dorsal surface is arched in the middle region of the body. The edges being flat are somewhat more transparent than the remainder of the body. The ventral surface is striated by longitudinal straight or slightly curved lines, the dorsal surface is smooth and without cilia. (Maupas describes bristles on the back, but this is not corroborated.) The adoral zone is fairly well developed, but not distinctly marked off from the remaining ventral surface. It begins on the right side and extends entirely around the frontal margin and down the left side below the middle of the body, where it turns suddenly to the right, entering the slightly insunk peristome. The mouth leads into a short, indistinct œsophagus. One contractile vacuole is situated in the dorsal swelling at the posterior end of the animal. Macronucleus double, one in each side of the dorsal swelling. Movement is slow and creeping, with a peculiar method of contracting the more hyaline edge, which may turn upward or around a foreign object.
Fresh (?) and salt water.
Peritromus emmæ Stein. Fig. 49.
With the characters of the genus.
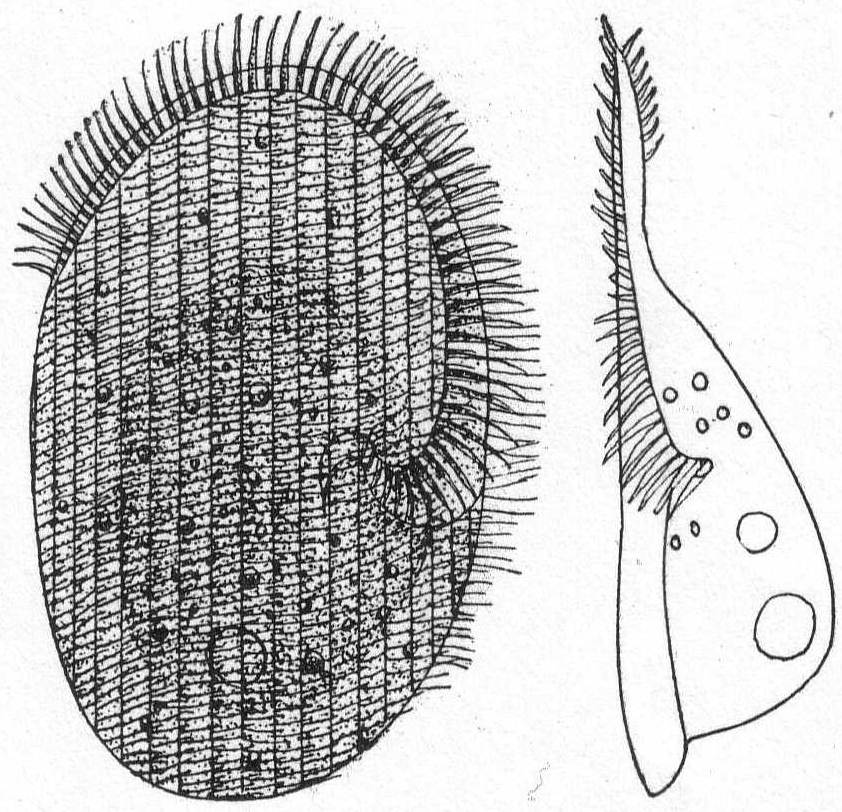
|
Fig. 49.— Peritromus emmæ, ventral and lateral aspects. ENLARGE |
| Key to the marine genera of Oxytrichidæ. | ||
| Diagnostic characters: The peristome is not always marked off from the frontal area. In the most primitive forms the cilia on the ventral surface are similar to those of the preceding family (Peritromidæ). Usually some of the anterior and some of the posterior cilia are fused into cirri, distinguished as the frontal and anal cirri, respectively. In the majority of forms all of the cilia are thus differentiated; strong marginal cirri are formed in perfect rows, and ventral cirri in imperfect rows. In addition to the adoral zone there is an undulating membrane on the right side of the peristome, and in some cases a row of cilia between the membrane and the adoral zone. These are the par-oral cilia and they form the par-oral zone. | ||
| 1. | The posterior end is pointed or tail-like | 2 |
| The posterior end is rounded; not tail-like | 5 | |
| 2. | The front end is pointed | 3 |
| The front end is rounded | 4 | |
| 3. | Frontal and anal cirri absent; often tube-forming | Genus Stichotricha |
| Eight frontal and 3 caudal cirri; not tubiculous | Genus Gonostomum | |
| 4. | Anal cirri present; with or without short lateral bristles | Genus *Epiclintes |
| Anal cirri absent; no bristles | Genus Uroleptus | |
| 5. | With frontal cirri | 6 |
| No frontal cirri; 2 to 3 rows of ventral cirri; anal cirri small | Genus Holosticha | |
| 6. | Right margin of peristome straight as far as the anterior end; 5 rows ventral cirri; 5 anal cirri | Genus Oxytricha |
| Right margin of peristome curved | 7 | |
| 7. | Five rows or less of ventral cirri | 8 |
| More than 5 rows of ventral cirri | Genus Urostyla | |
| 8. | Membranelles normal; 5-10 anal cirri; no caudal cirri | Genus *Amphisia |
| Membranelles normal; 5 to 10 anal cirri; 3 caudal cirri | Genus Stylonychia | |
| Membranelles very large and powerful; adoral zone not continued to mouth; 5 anal cirri | Genus Actinotricha | |
| * Presence at Woods Hole indicated by asterisk. | ||
Very active, contractile, colorless forms of rather small size. In the fully expanded condition the body is oval and long, with its greatest width in the center or at the front half of the body. The posterior end is always drawn out into a relatively long tail, which is extremely elastic. The peristome is short and stretches around the front end of the animal. In the frontal region are from one to three rows of cirri. The ventral surface is covered with longitudinal rows of cilia, the number of rows being in dispute (6 to 7 according to Stein; 9 according to Mereschowsky and Rees) Some of these cilia project from the lateral edges and from the posterior end, where they are slightly elongated. The anus is dorsal and placed at the beginning of the posterior process. Macronucleus probably double. Movement is rapid and restless, the tail process contracting to jerk the body backward. Salt water.
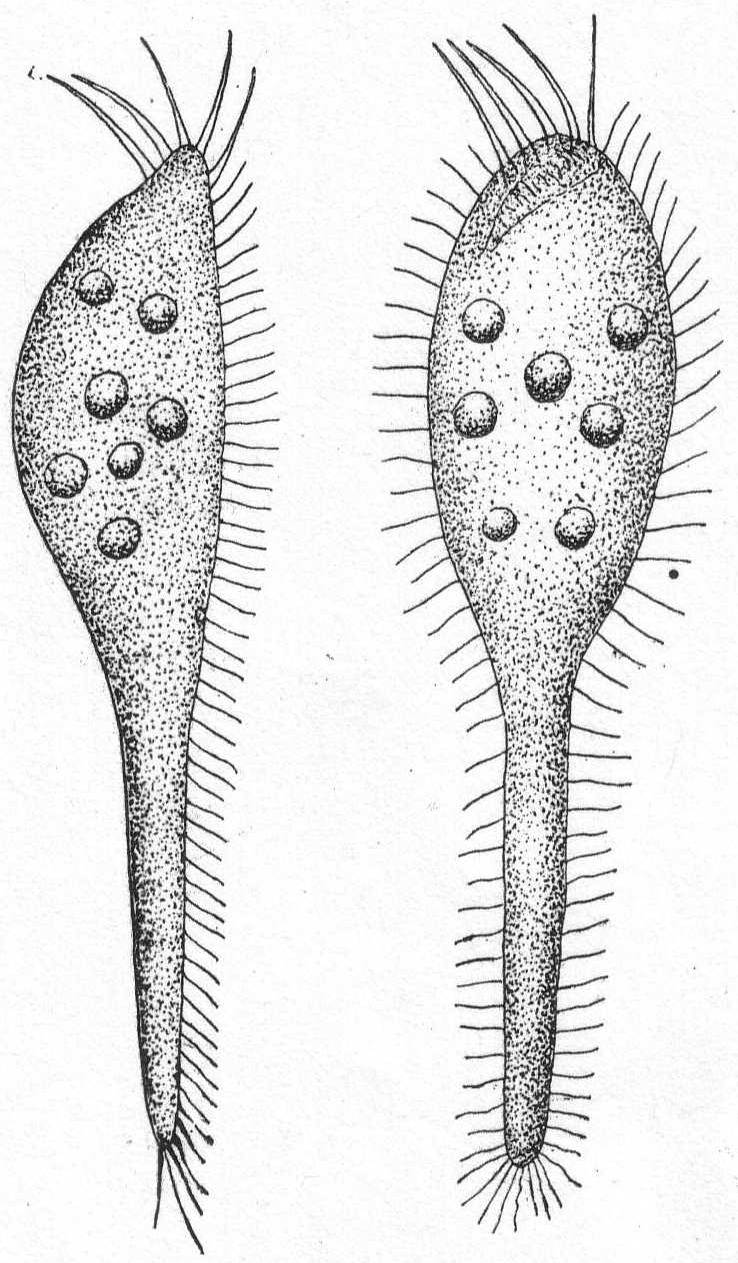 Fig. 50.— Epiclintes radiosa. ENLARGE |
Epiclintes radiosa Quenn. Fig. 50.
Synonym: Metra radiosa Quenn.
The body is elongate, slightly narrowed anteriorly, and drawn out posteriorly into a long, retractile, tail-like portion. Five large cirri extend outward from the anterior extremity. The caudal portion may be extended to a distance equal to twice the length of the body or contracted to half the length. The peculiar nervousness of this form made it extremely difficult to study, and the oral region was imperfectly made out. The anterior cirri appear to line the upper left border of the peristome, which is marked by a row of large cilia. The peristome begins upon the right side of the anterior end and passes backward and to the left, narrowing at this point. The mouth is very small and difficult to see. It is apt to stay in one locality under zooglœa, switching back and forth with great vivacity, or hanging on by the posterior cilia while the anterior end stretches out in the surrounding medium. Nucleus and contractile vacuole were not observed. Length 45µ.
The body is plastic and soft, colorless or slightly tinged with yellow or red. In form it is oval or elongate, the posterior end is rounded and slightly reduced in diameter, but does not form a distinct tail. The anterior end is also rounded and similarly reduced in width. There are two rows of marginal cirri (Randcirren), which may be placed some distance from the edge, and two or three rows of ventral cirri between them. There are from 3 to 5 frontal cirri of larger size than those of the ventral rows, and from 5 to 10 anal cirri. (The genus Holosticha is similar in all respects save the presence of frontal cirri.) The macronucleus is double; the contractile vacuole is central and on the left side. The peristome is long and rather narrow and carries an undulating membrane on its right margin. Fresh and salt water.
Amphisia kessleri Wrzes. '77. Fig. 51.
Synonyms: Trichoda gibba Müller; Oxytricha gibba Stein '59; O. velox? Quen. '69; O. kessleri Wrzes. '77.
Body elongate, slightly sigmoid and swollen in the center, about 3-1/2 times as long as broad; the rounded anterior end is turned to the left, the similarly rounded posterior end to the right; both ends taper slightly. The peristome is long and narrow, with a distinct adoral zone which appears broken in its course. To the right of this adoral zone is a single line of preoral cilia. On the right border of the peristome is an undulating membrane. The three frontal cirri form a triangle and the five smaller anal cirri form a continuous line with the broken row of ventral cirri. There are two and one-half rows of ventral cirri and the marginal cirri are drawn in until they are ventral in position.
Length 135µ; greatest width 40µ.
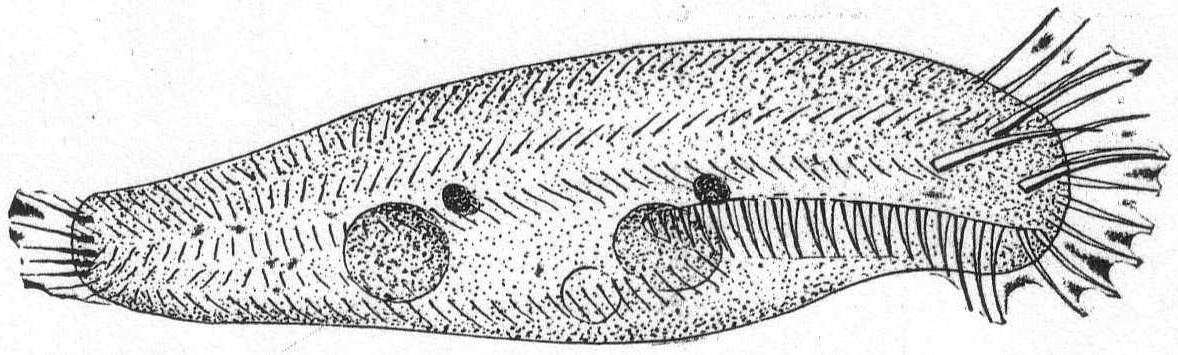
Fig. 51.—Amphisia kessleri. ENLARGE |
This variety differs from O. kessleri as described by Wrzesniowski in having three frontal cirri instead of four. Another difference is in the structure of the nuclei and in their position. These differences are too minute to warrant a specific name. O. velox of Quennerstedt is probably the same as 0. kessleri, but differs in having three complete rows of ventral cirri. O. velox has three frontal cirri in a line, thus differing from the Woods Hole form.
| Key to the marine genera of Euplotidæ. | ||
| Diagnostic characters: Cilia, as well as the frontal, marginal, and ventral cirri, very much reduced; the anal cirri, on the other hand, are always present. The macronucleus is band-form. | ||
| 1. | Frontal cirri more than 8 | 2 |
| Frontal cirri less than 8 | 3 | |
| 2. | Eleven marginal cirri on the left side; 11 frontal cirri | Genus Certesia |
| Four marginal cirri, 2 on each side; 9 to 10 frontal cirri | Genus *Euplotes | |
| 3. | Seven frontal, 5 anal, 3 right marginal, and 2 left marginal cirri | Genus *Diophrys |
| No frontal, 5 anal, 3 right, and 2 left marginal cirri | Genus *Uronychia | |
| * Presence at Woods Hole indicated by asterisk. | ||
Small to medium-sized forms. Rigid in form, colorless, or green by chlorophyl. They are quite flat on the ventral surface but decidedly arched dorsally, and the contour is usually oval. The anterior end is broadly rounded to truncate; the posterior end is similarly rounded, or may be somewhat pointed. The mouth is placed centrally or near the left margin, and from it the right edge of the peristome forms a curved line to the left, which bends forward, thus making the greater part of the left edge the peristomial area. In front the peristome bends sharply to the right and extends as far as the right end of the adoral zone. Upon the frontal and median ventral surface are 9 to 10 great cirri (Bauchwimpern of Stein). Posteriorly five great anal cirri stretch out beyond the posterior body margin. In addition to these there are two smaller marginal cirri upon the left body edge, and two similar ones on the hinder part of the body. The dorsal surface is rarely smooth, but usually is marked by longitudinal ridges, and rows of dorsal bristles have been described. The single contractile vacuole lies on the right side in the region of the anal cirri, sometimes just above them, sometimes below. The anus is posterior and on the right side. The characteristic macronucleus is long and band-form, its main portion being usually on the left side with an anterior and a posterior arm toward the right. Movement is rapid swimming, which, however, is frequently broken by creeping periods, during which the animals appear to be examining the foreign body on which they creep.
Fresh and salt water.
Euplotes charon Ehr. Fig. 52.
Synonyms: Trichoda charon Müller; Plœsconia charon; P. affinis, subrotunda, radiosa, longiremis, Dujardin '41.
The body is oval, small, and somewhat variable in length. The carapace is strongly marked upon the dorsal side by deep longitudinal grooves, 6 to 8 in number; the grooves may be absent, however. The adoral zone extends to the posterior third of the body, the mouth and œsophagus are directed anteriorly. There are 10 ventral cirri, 7 of which are on or near the frontal border and 3 near the right edge. There are 5 posterior cirri and 4 anal cirri, of much smaller size. The cirri may or may not be fimbriated, the latter condition indicating the approaching disintegration of the body and is abnormal. The macronucleus is long and band-formed or horseshoe shape. The contractile vacuole lies on the right side dorsal to the posterior cirri.
Fresh and salt water. Length 45µ; diameter 25µ.
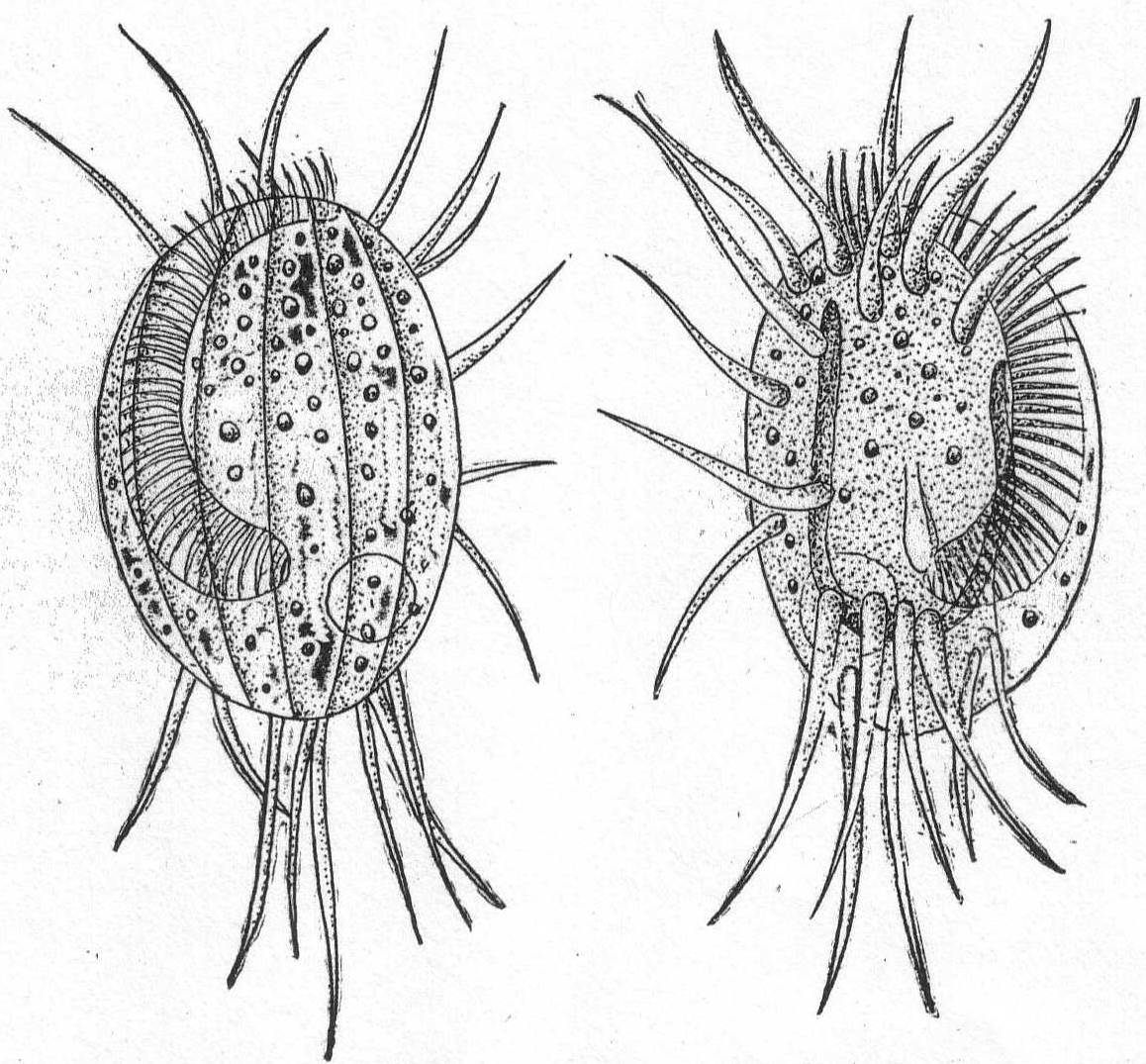
|
Fig. 52.— Euplotes charon, dorsal and ventral aspects. ENLARGE |
Euplotes harpa Stein. Fig. 53.
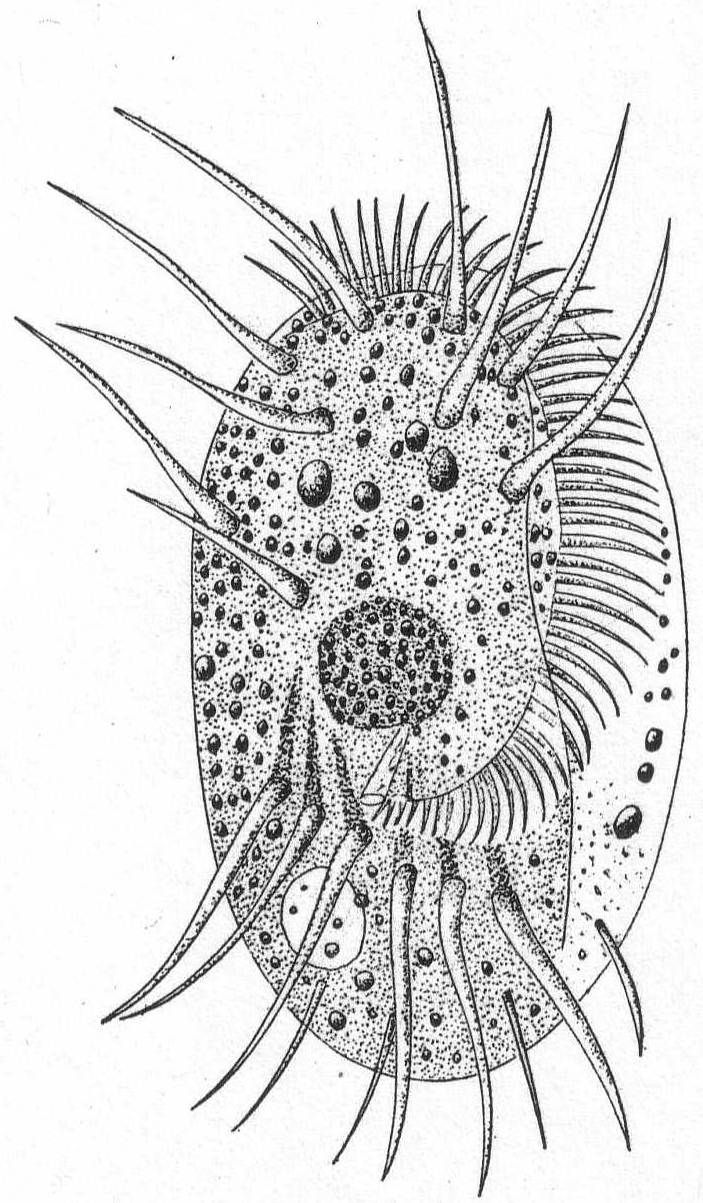
|
Fig. 53.— Euplotes harpa. ENLARGE |
The body is elongate, oval, somewhat widened anteriorly, and has rounded ends. The frontal margin is three-toothed. Ten ventral cirri. Dorsal surface provided with 8 longitudinal markings. The peristome is long and broad, with considerable variation. The adoral zone consists of powerful membranelles arranged in a continuous curve from the mouth to the extreme right frontal margin. Seven of the 10 ventral cirri are situated at the anterior extremity; the remainder are arranged in a triangle on the right edge. The anal cirri, 5 in number, are long and stiff; the marginal cirri smaller and finer. The nucleus and contractile vacuole are similar to those of the preceding species.
Length 95µ; width 54µ.
Medium size, colorless to yellow, rigid in form. The body contour is oval, the anterior end being rounded or slightly reduced, the posterior end usually cut in on the right side. The peristome is broad but less extensive than in Euplotes, and may extend beyond the middle of the body. Its right edge is convex toward the right side, extends forward and does not turn again to the right. The anterior ventral surface has 7 to 8 scattered cirri and just behind the mouth is a transverse row of large anal cirri. In the sharp in-cut of the posterior end are three great angular cirri. Two lateral cirri are placed on the left of the median line between the mouth and the anal cirri, and usually in a slight hollow. The contractile vacuole is on the right side in the vicinity of the anal cirri. The macronucleus is in two parts, each band-form, one anterior, the other posterior in position. Movement is rapid and steady. Salt water.
Diophrys (Styloplotes) appendiculatus Stein '59. Fig. 54.
Synonyms: Styloplotes appendiculatus Stein '59; Kent '81; Quennerstedt '67, etc.
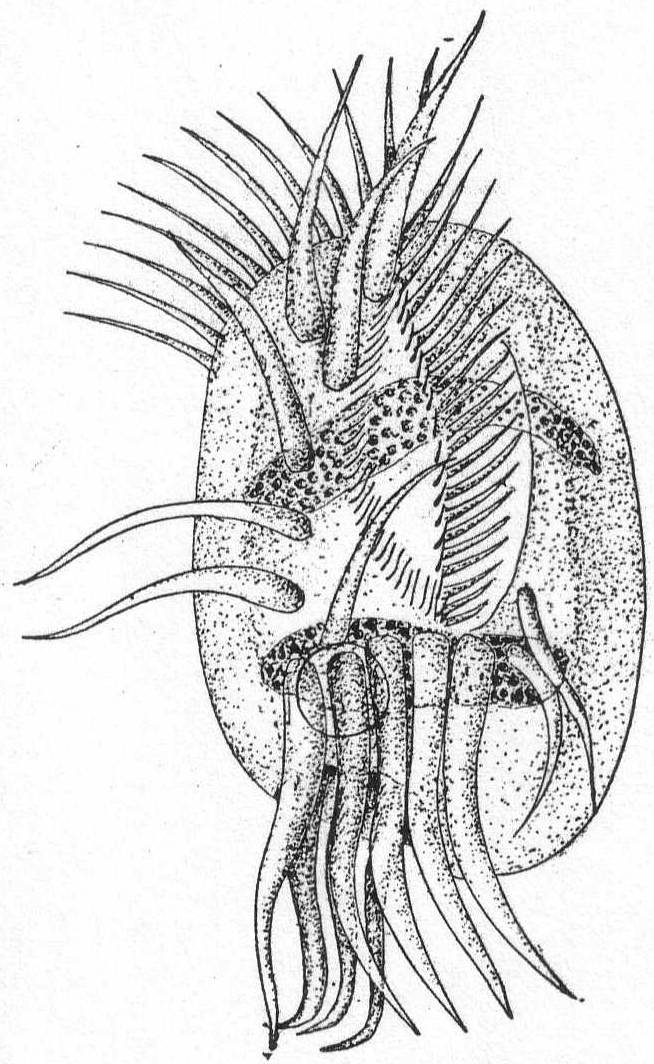
Fig. 54.— Diophrys appendiculatus. ENLARGE |
The general form resembles Euplotes. Its outline is oval and regular except at the posterior end on the right side, where there is a considerable indentation. The frontal margin is characterized by a row of powerful membranelles, which become smaller at the peristome and at the mouth they are of characteristically small size. The ventral cirri are 7 in number. Five of them are in one row from the anterior end down the right side nearly to the anal cirri; 1 is on the frontal border between the first two; 1 lies just anterior to the second anal cirrus from the right side. The 5 anal cirri are large and powerful and extend some distance beyond the posterior end of the body. In all specimens observed these cirri curve to the left. Dorsal to the anal cirri and placed deep into the dorsal pit are 3 large, sharply curved cirri, which in most cases are fimbriated, but when the specimens are normal these are pointed and curve abruptly to the right. Two smaller cirri lie to the left of the group of anal cirri. The peristome is well-marked by the adoral zone, and upon its right border there is a row of cilia, and a similar row of cilia runs along the base of the oral membranelle. The macronucleus is double and consists of two elongate cylindrical masses lying parallel with one another. One of these is in the anterior region; the other is posterior. The contractile vacuole lies dorsal to the anal cirri and anterior to the three dorsal cirri. The movement and general activities resemble those of Euplotes.
Length 50µ; diameter 25µ.
Medium-sized colorless ciliates of usually constant body form. The body is somewhat short and oval in outline. The anterior end is broadly truncate, the posterior end rounded or slightly pointed. The ventral and dorsal surfaces are considerably arched and the latter usually has a number of rows of longitudinal stripes. The open peristome is broad and reaches back to the middle of the ventral surface and beyond. According to Stein, the two edges can approach each other, thus opening and closing the peristomial area. Its right edge forms a greater angle with the front edge than in the genus Euplotes, and the left edge forms a greater angle with the front edge than in that genus. The left edge also appears to cover over the adoral zone slightly. There are no ventral cirri in front, but on the posterior ventral surface are 7 great springing cirri. Five of these are inserted on the right aide in a deep in-sinking, and the other 2 in a similar depression on the left ventral surface. Above the 5 right-side cirri, i.e., dorsal to them, but in the same depression, are 3 angular cirri. A few edge cirri are found to the left of them and another to the right of the 5 cirri. The contractile vacuole is on the left side between the main groups of cirri. The macronucleus is band-form or spherical, and is situated in the middle region of the body. Movement consists in forward swimming with sudden springs.
Salt water.
Uronychia setigera, n. sp. Fig. 55.
This species is very common in the Woods Hole waters. It is small, colorless, and very active. The most characteristic feature is the posterior end with its relatively enormous cirri, which are apparently large enough for an animal four times its size. The form is ovoid, widened posteriorly.
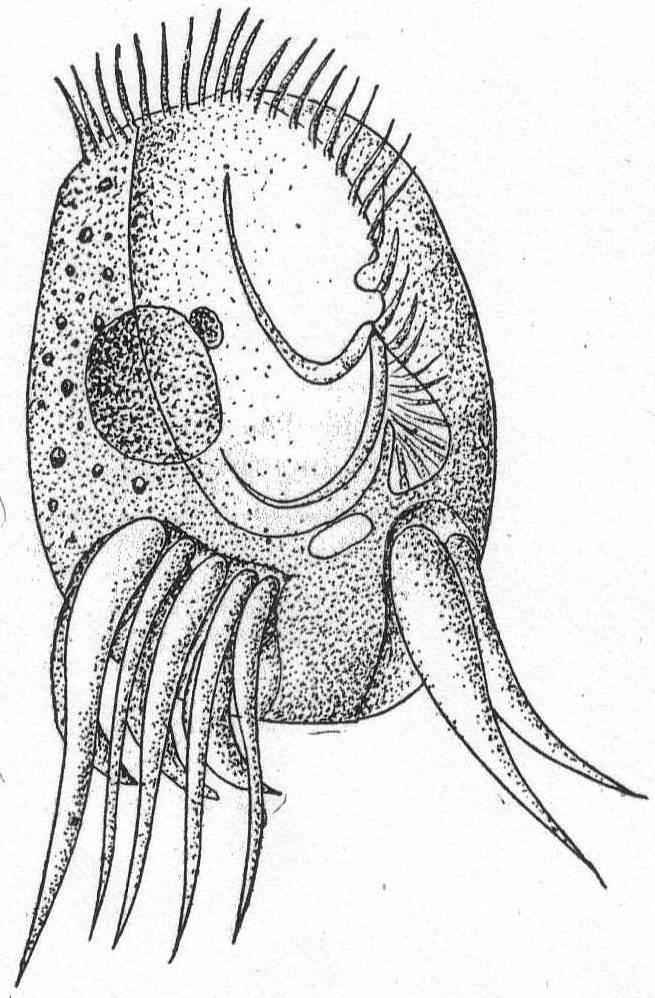
Fig. 55.— Uronychia setigera. ENLARGE |
The ventral surface is flat and has two excavations in the posterior end. The right hollow is larger and contains 5 great cirri of unequal size, the extreme right one being the largest. The left hollow contains 2 cirri, also of dissimilar size. Dorsal to the 5 right cirri are 3 sickle-formed cirri, which are usually fimbriated. These are pointed and curve regularly to the left. The peristome is wide and open, and a small pocket-like hollow on its left border indicates the region of the mouth. The adoral zone runs into this pocket and the mouth is located in its lower right-hand corner. In U. transfuga the right border is generally described as having a membrane of extreme delicacy. I was unable to see such a membrane in this form, but in its place there are 2 flagella-like cirri extending from the margin of the mouth-opening into the peristome, and these vibrate slowly. I do not believe these could be the moving edge of an undulating membrane, for they are quite distinct. The macronucleus is spherical instead of band-form, and a single micronucleus is closely attached. This is unlike the European species U. transfuga, in which the nucleus is elongate. The contractile vacuole lies between the two sets of posterior cirri. There are no marginal folds like those of the European species.
Length 40µ; width 25µ. Common.
Small, colorless, and rigid forms, with nearly circular to oval contour. The left side is usually straight, or at least but slightly convex. The right side is much more convex, and the right margin is considerably thickened. The ventral side is flat, the dorsal surface convex, with from one to several longitudinal ridges which run more or less parallel with the right edge. The peristome is limited to the left edge, where it forms a small depression which may or may not reach the anterior border, but which in no case runs around the anterior margin. The left peristome margin in some cases grows over the peristome depression toward the right, thus making a sort of cover for the peristome. In the posterior region is a deep depression, from which 5 to 12 cirri take their origin. Seven or 8 cirri are placed in the anterior half of the ventral surface and are arranged more or less in rows. The anus is on the right side in the region of the anal cirri (Stein). The contractile vacuole is generally on the right side and similarly located. The macronucleus is a horseshoe-shaped body. Movement rapid, somewhat in circles, and rather uniform.
Fresh and salt water.
Aspidisca hexeris Quennerstedt '67. Fig. 56.
The carapace is elliptical, about 1-1/2 times as long as broad, rounded at the extremities. The left border of the carapace bears a spur-like projection. The ventral cirri are short and thick, and are very characteristic of the species. When moving slowly they look much like nicely-pointed paint brushes, but when the animal is compressed they quickly become fibrillated, and then look like extremely old and worn brushes. These cirri are placed in depressions in the ventral surface and each one appears to come from a specific shoulder. At the posterior end an oblique hollow bears 6 unequal cirri placed side by side. The extreme right cirrus is the largest, and they become progressively smaller to the opposite end. Dorsal to these lies the contractile vacuole. The peristome is in the posterior half of the body and an undulating membrane extends from it into the œsophagus. The dorsal surface is longitudinally striated by 5 or 6 lines, which are usually curved. The nucleus is horseshoe-shaped and lies in the posterior half of the body. Length 68µ; diameter 48µ.
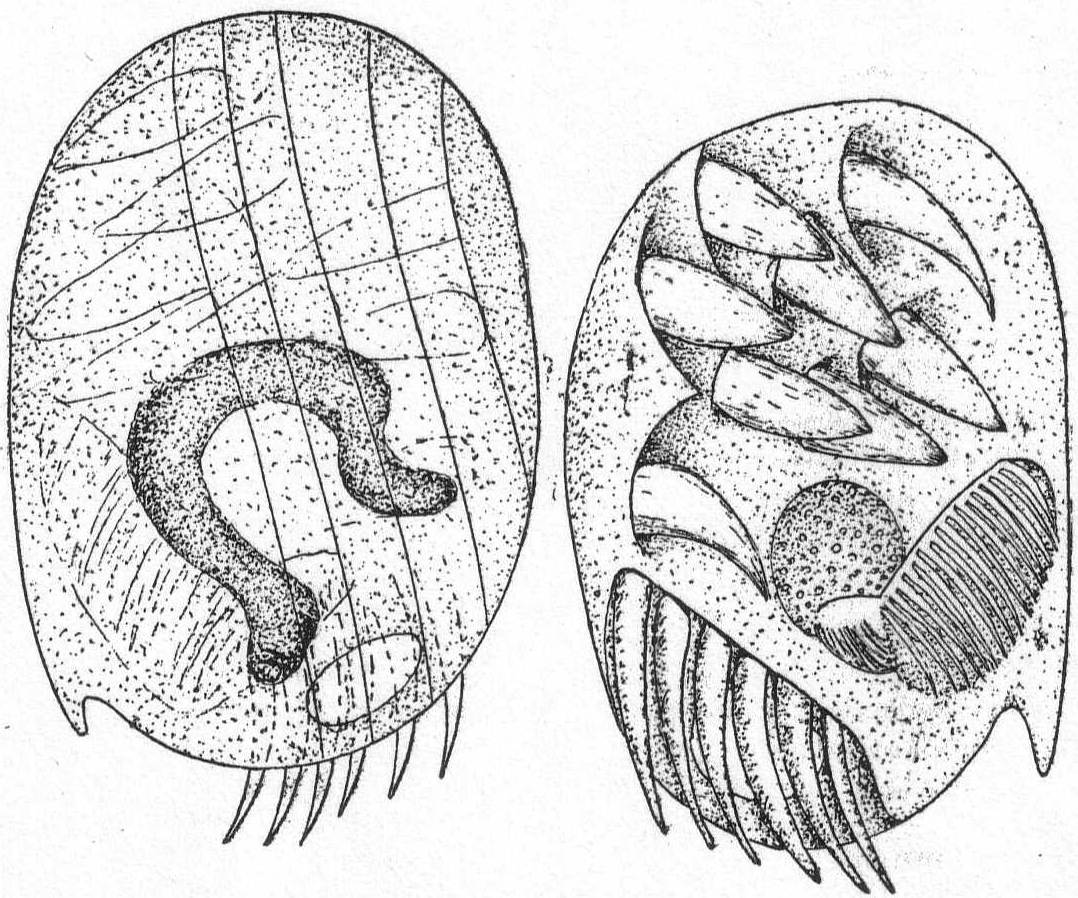
|
Fig. 56.— Aspidisca hexeris. ENLARGE |
This form was incorrectly mentioned as Mesodinium sp. by Peck '95:
In the figure given by Quennerstedt there are only 7 ventral cirri. In the Woods Hole form there are 8, 7 of which are anterior, 6 of them about one central one. The eighth cirrus is by itself, near the base of the largest posterior cirrus. These cirri, in spite of their size, are easily overlooked and more easily confused, but by using methylene blue they can be seen and counted.
Aspidisca polystyla Stein. Fig. 57.
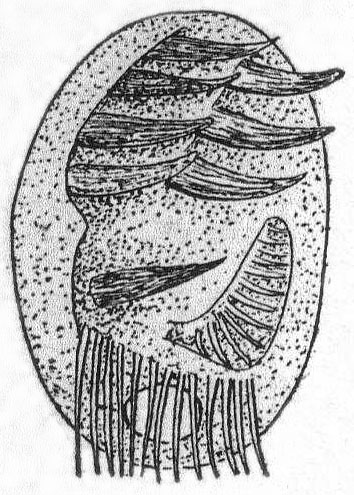
Fig. 57.— Aspidisca polystyla. ENLARGE |
This species is similar to A. hexeris, but is smaller, very transparent, and without the spur-like process on the left edge of the carapace. The chief difference, however, lies in the number of anal cirri. These are 10 in number and they are arranged obliquely as in the preceding species, with the largest one on the right and the smallest on the left. The ventral cirri are 8 in number, and are arranged in two rows, one of which, the right, has 4 cirri closely arranged, the other having 3 cirri close together and one at some distance, near the largest anal cirrus. The peristome, contractile vacuole, and nucleus are similar to the preceding. Length 36µ; width 22µ.
Stein assigns only 7 ventral cirri to this species, but he also describes 2 very fine bristle like cilia (p. 125) and pictures them in figs. 18, 19, 20, and 21 of his Taf. III in the same relative position as my eighth cirrus. I am positive that cilia do not occur on the ventral face of this form, and that the characteristic cirri are the sole locomotor organs.
| Key to families of Peritrichida. | ||
| a. | Peristome drawn out into funnel-like process; parasitic | Family Spirochonidæ |
| b. | Adoral zone and circlet of cilia at opposite end. Adoral zone left-wound. Parasitic. |
Family Lichnophoridæ (one genus, *Lichnophora) |
| c. | Adoral zone a left-wound spiral. Attached or unattached forms. | Family Vorticellidæ |
| * Presence at Woods Hole indicated by asterisk. | ||
Small or medium-sized colorless animals, extremely elastic and flexible. The anterior part, bearing the adoral zone, is round or oval in ventral view, and has a flat ventral and a highly arched dorsal surface. The posterior end of the animal is reduced to a stalk-like structure which is broadened at the extremity to form a sucking disk. The surface of this disk and the surface of the peristome may be brought into the same plane by the characteristic bending of the stalk portion. A ciliated girdle is placed at the edge of the sucking disk. A well-developed adoral zone incloses the peristome; it begins at the mouth on the left side and includes nearly all of the peristome in its left-wound spiral, the extremity approaching closely the end near the mouth. The macronucleus is a long-beaded structure, or it may be in several parts connected by strands (Gruber). The contractile vacuole is on the left side in the region of the mouth. Salt water.
Lichnophora macfarlandi Stevens. Fig. 58.
The body is elongate; oral disk variable in form, attachment disk clearly defined and constant. The stalk is very contractile and elastic, constantly changing in shape. When detached from the host the animal moves with a very irregular and indefinite motion. When attached it moves freely over the surface on its pedal disk. The latter is bordered by four membranes composed of cilia. A distinct axial fiber extends from the pedal disc to the peristome and gives off a number of branches. This fiber is analogous to the myonemes in Vorticella. An indistinct longitudinal furrow can be made out occasionally. The nucleus is in 5 or 6 separate pieces, of which 1 is found in the pedal disk and 1 or 2 in the neck.
On the egg capsules of Crepidula plana; also reported upon annelids at Woods Hole.
Length 60µ from disk to extremity of the peristomial disk.
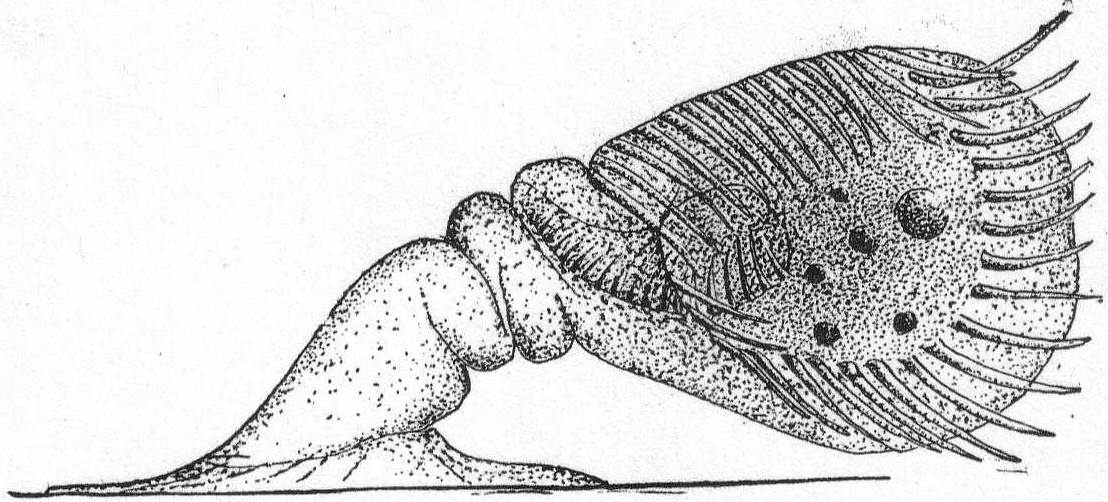
|
Fig. 58.— Lichnophora macfarlandi. ENLARGE |
This form does not agree in all respects with Stevens's species, but the agreement is so close in other respects that I believe it can be safely identified as L. macfarlandi. The mode of life is different, and the macronucleus is different, there being from 25 to 30 fragments in Stevens's form and only 5 or 6 in the present one. There is, however, the same evidence of chain formation in both of them. The length of the oral cilia in Stevens's form is 18µ in fixed and 30µ in living forms. In the Woods Hole form the cilia are not more than half that length.
| Key to the marine genera of Vorticellidæ. | |||
| Diagnostic characters: Attached or unattached forma of peritrichous ciliates in which the adoral zone seen from above forms a right-wound spiral. A secondary circlet of cilia around the posterior end may be present either permanently or periodically. | |||
| 1. | Posterior ciliated girdle permanent around an attaching disk | 3 | |
| 2. | Posterior ciliated girdle, temporary during motile stage | 4 | |
| 3. | Body cylindrical: | ||
| (a) | With ring of stiff bristles above the ciliated girdle | Genus Cyclochæta | |
| (b) | Without accessory ring of bristles; with velum | Genus Trichodina | |
| Body conical; general surface ciliated | Genus Trichodinopsis | ||
| 4. | No test and no stalk | Genus Scyphidia | |
| 5. | No test; with stalk containing contractile thread | 8 | |
| 6. | No test; with stalk but without contractile thread | Genus Epistylis | |
| 7. | With a test; with or without a stalk | Genus *Cothurnia | |
| 8. | Individuals solitary | Genus *Vorticella | |
| Individuals colonial; entire colony contractile | Genus *Zoothamnium | ||
| Individuals colonial; parts only of the colony contractile | Genus Carchesium | ||
| * Presence at Woods Hole indicated by asterisk. | |||
Medium-sized ciliates of general bell-like form. They may be colorless, or yellow and green through the presence of Zoochlorella. When not contracted, the peristome end is widespread, rarely narrowed. The adoral zone and peristome agree with the details given in the family characteristics. The chief character is the attachment of the posterior end by means of a single, longer or shorter, stalk, which contains a highly contractile thread easily distinguished in the living animal. Another character is the absence of colony formation. Contractile vacuole, single or double, usually connected with a sac-like reservoir. The macronucleus is invariably long and band-formed, with attached micronucleus. Fresh and salt water.
So many species of Vorticella have been described that the task of collecting data and of arranging the synonyms is extremely irksome and difficult. Stokes enumerates 66 species, inhabiting fresh and salt water, and several other new species have been added since his work. I am impressed with the fact that new species have been created without proper regard for the manifold variations which nearly all of the Ciliata show, and I believe the 66 species might be safely reduced to 12 or 15.
Vorticella patellina Müller. Fig. 59.
Body campanulate, widest at anterior border, from which it tapers directly to the pedicle. The diameter of the peristome is a little larger than the length of the body. The ciliary disk is but little elevated. The cuticle is not striated and the body plasm is quite transparent. Length 52µ.
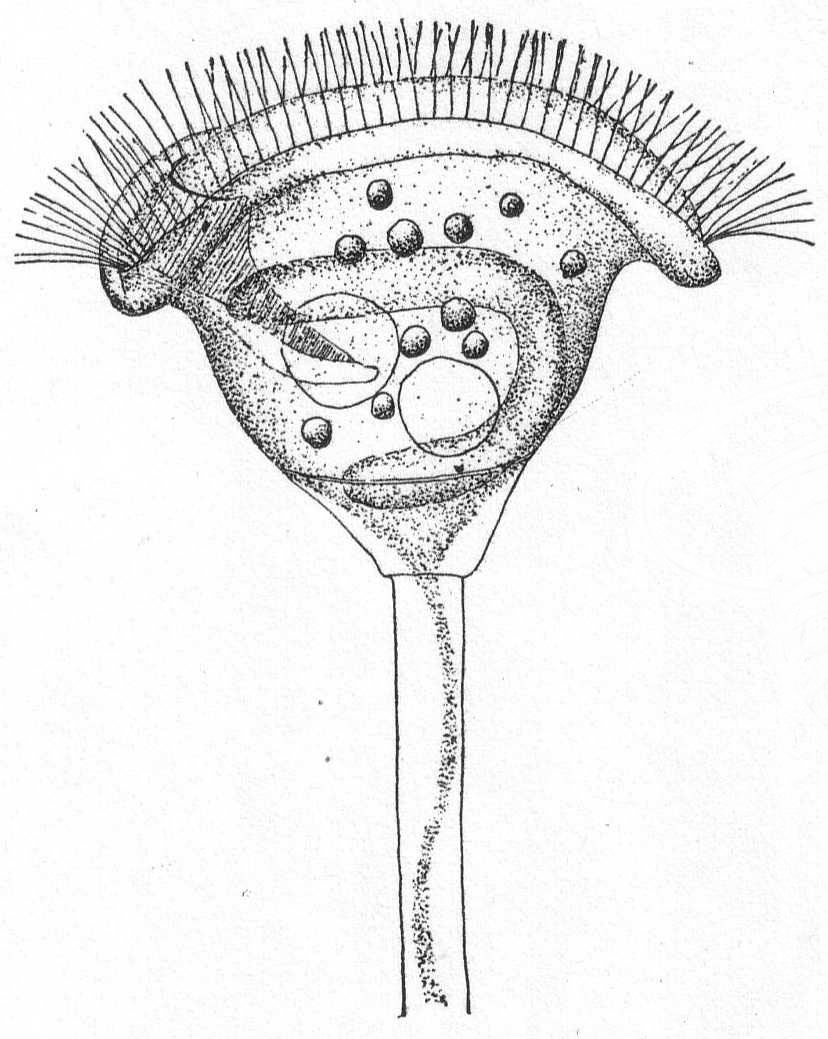
|
Fig. 59.— Vorticella patellina. ENLARGE |
Vorticella marina Greeff. Fig. 60.
The body is conical but variable, and may he short or elongate, so that relative length and breadth offer no chance of identification. In general the body is campanulate. The distinguishing feature is the transverse annulation of the bell.
Small, but common, and grows in small social groups. Length 35µ.
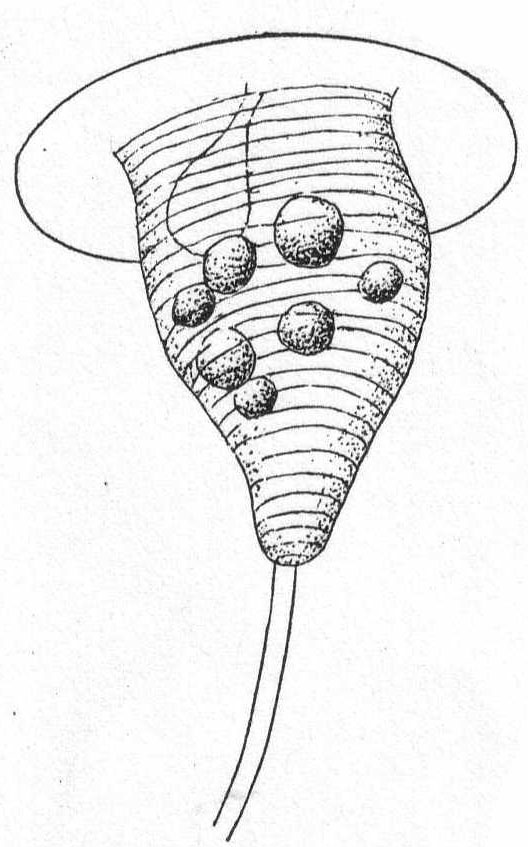
|
Fig. 60.— Vorticella marina. ENLARGE |
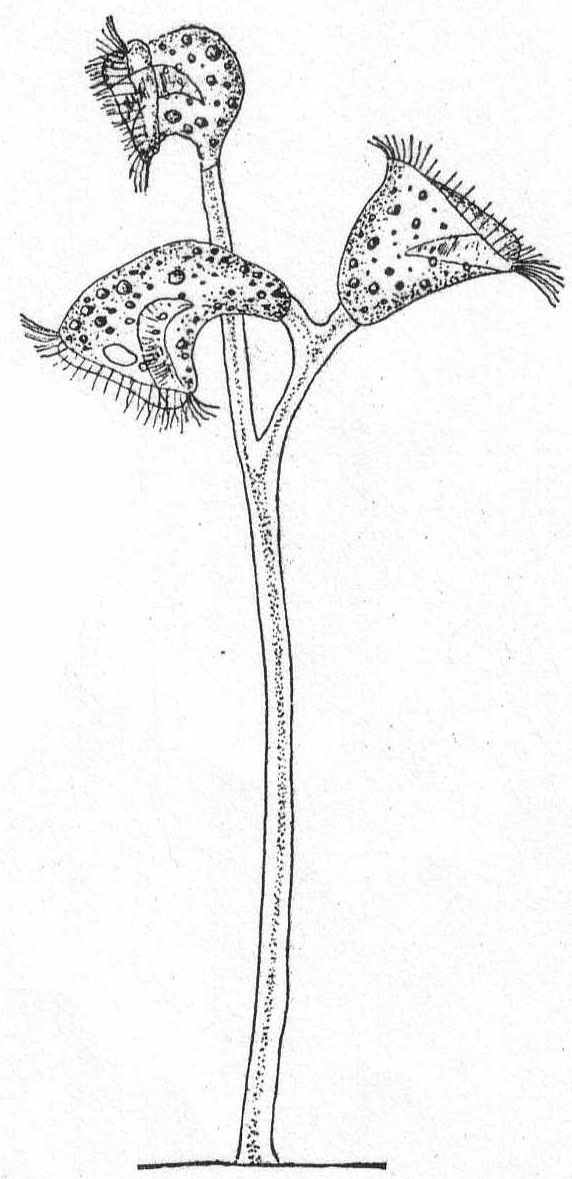
Fig. 61.— Zoothamnium elegans. ENLARGE |
Colorless and highly contractile forms growing in small or large colonies. The form and structure of the individuals is not different from Vorticella. The colonies are usually richly branched upon the dichotomous plan and the entire colony is contractile. The main character is that with each division of the individual the stalk also divides, each daughter cell getting one-half of the parent stem. The stems therefore remain in communication, so that a simultaneous contraction results, and the colony as a whole is withdrawn. In some species so-called macrogonidia, or larger sexual individuals, are developed alongside the usual ones. Fresh and salt water.
Zoothamnium elegans D'Udekem '64? Fig. 61.
The bodies are variable—peristomial border widely dilated, tapering and attenuate posteriorly. The pedicle is slender, smooth, and transparent, and branches sparsely at its distal extremity. There are but few zooids (3 to 4). The ciliary disk projects conspicuously beyond the peristomial border. The pharyngeal cleft is very distinct and extends beyond center of body. Length of body 80µ.
Colorless forms of medium size-in some cases they may be green by Zoochlorella. The general structure is similar to that of Vorticella, but the individuals are elongate and occupy houses. The macronucleus is invariably long and band-form. The distinguishing character is the colorless or brownish lorica of quite variable form but always attached. These houses may be finger-formed, with widened center, or widened mouth, or constricted mouth, and the like. Ring-formed swellings are frequently developed. Sometimes the mouth becomes twisted and the lorica is therefore bilateral. The houses are attached either directly to some foreign object or by means of a short stalk. The animals are similarly fastened to the lorica, sometimes directly, sometimes by means of a short stalk. When they contract they draw back to the bottom of the lorica; when expanded they usually stretch out of the mouth opening. In some forms there is an operculum, by means of which the opening of the shell can be closed when the animal is retracted. Fresh and salt water.
The number of species of Cothurnia has become so great that the difficulty in placing forms is almost sufficient to discourage the systematist; as Bütschli well remarks, the variations in the theca have been made the basis of new species so many times that the genus is almost as confused as Difflugia among the rhizopods or Campanularia among the hydroids. The length of cup, of stalk, the presence of annulations on stalk or cup, etc., have given rise to many specific names, the majority of which I believe can be discarded. According to such differentials the same branch of an alga holding a hundred specimens of Cothurnia crystallina yield 10 or 12 species, whereas they are merely growth stages of one and the same form.
Cothurnia crystallina Ehr. Fig. 62.
Synonyms: Vaginicolla crystallina Ehr., Perty, Eichwald; V. grandis Perty; V. pedunculata Eichwald; Cothurnia crystallina Claparède & Lachmann, D'Udek.; C. gigantea D'Udek; C. maritima, C. crystallina Cohn; C. grandis Meresch.
The form of the cup shows the greatest differences; sometimes it is cylindrical, sometimes elongate thimble-shape, sometimes pouch-shape, corrugated or smooth on the sides, and wavy or smooth on border. Frequently the basal part becomes stalk-like, but this is very short. When present, the stalk may or may not have a knob-like swelling. The animal within the cup may or may not be borne on a stalk, and this stalk may or may not be knobbed. The cups are colorless or brown. The animal is very contractile and may stretch half its length out of the cup or retract well into it. There is no operculum. The length of the cup varies from 70µ to 200µ (C. gigantea; Vag. grandis, etc.). From Entz.
There is nothing to add to Entz's characterization of this species, which is found both in fresh and salt water. The variability of the cup and stalk is quite noticeable in the Woods Hole forms.
 Fig. 62.—Cothurnia crystallina. ENLARGE |
Cothurnia imberbis Ehrenberg, var. curvula Entz. Fig. 63.
Synonyms: C. imberbis Kent et al.; C. curvula Entz; C. socialis Gruber?
The lorica is swollen posteriorly, narrowest at the oral margin, bent on its axis and is supported on a short stalk. It is perfectly smooth and without annulations. The animal itself has no definite stalk. When fully expanded the animal emerges but slightly from the margin of the cup. Fresh and salt water. On red algæ. Dimensions of Woods Hole form: Cup 50 to 55µ long; greatest diameter 22µ; length of stalk 4 to 5µ.
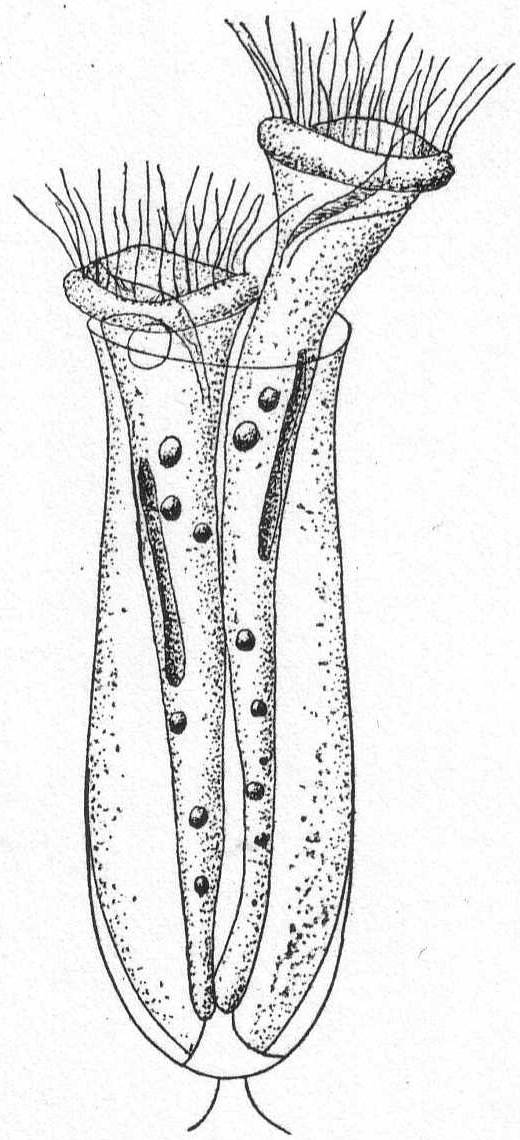
|
Fig. 63.—Cothurnia imberbis.
ENLARGE |
Cothurnia nodosa Claparède & Lachmann. Fig. 64.
| A. | Smooth cup.—Cothurnia maritima Ehr., Eichwald, Stein, Kent. |
| B. | Cross-ringed cup.—C. pupa Eichwald, Stein, Cohn; C. nodosa Cl & L.; V. crystallina Entz '78; C. pontica Meresch., Kent; C. cohnii and pupa Kent; C. longipes Kellicott '94. |
The cup is elongated, swollen centrally, tapering at oral end and conical at base or rounded. Oral opening either circular or elliptical. Cross rings may or may not be present, and the cup is either smooth or annulate. Length of cup 70µ to 80µ. The stalk which supports the cup is extremely variable in length. The animal is borne upon a stalk of variable length within the cup.
Entz states that the many variations which this species exhibits run into each other so gradually that he does not believe it wise to separate them. The Woods Hole forms which I found on algæ of various kinds were nearly of a size, and did not vary much from the one figured. Kellicott '94 described a Cothurnia from Woods Hole under the name of C. longipes, which I believe is only a long-stemmed variety of C. nodosa. My form has the following dimensions: Cup 75µ; cup stalk 38µ; animal stalk 14µ.
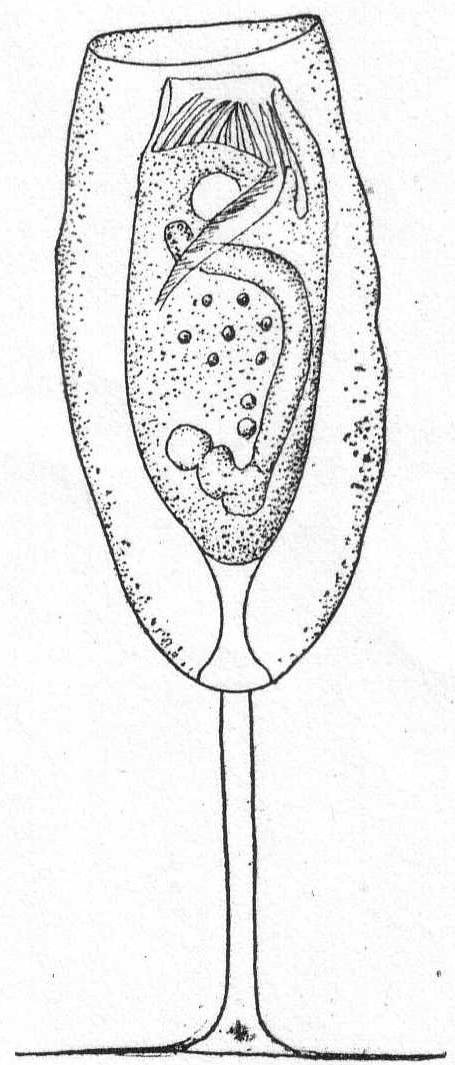
|
Fig. 64.—Cothurnia nodosa.
ENLARGE |
| Key to families of Suctoria. | ||
| a. | Unattached forms; ventral cilia present; one suctorial tentacle | Hypocomidæ |
| b. | Attached forms; thecate and athecate tentacles simple, one or two in number | Urnulidæ |
| c. | Thecate; posterior end of cup drawn out into stalk; walls perforated for exit of tentacles | Metacinetidæ |
| d. | Stalked or unstalked; globular; tentacles of different kinds, some knobbed, others pointed |
Podophryidæ (2 genera *Ephelota, *Podophrya) |
| e. | Naked or thecate; stalked or not; tentacles numerous, usually knobbed and all alike | Acinetidæ |
| f. | Naked; athecate; tentacles numerous, all alike, knobbed and grouped in tufts. They may be simple or branched. | Dendrosomidæ |
| g. | Sessile forms resting on basal surface or on a portion raised like a stalk; tentacles many; short and knobbed; distributed on apical surface or localized on branched arms | Dendrocometidæ |
| h. | Stalked or sessile; tentacles long, rarely knobbed, supported on proboscis-like processes | Ophryodendridæ |
| * Presence at Woods Hole indicated by asterisk. | ||

Fig. 65.— Podophrya gracilis. ENLARGE |
The body is globular, with tentacles radiating in all directions. The tentacles may be very short or very long. The stalk also is either short or long, and some species form stalks but rarely (P. libera). The macronucleus is centrally placed and globular to ovoid in form. The contractile vacuole is usually single. Reproduction takes place by division; the distal half developing cilia and becoming a swarm-spore. Fresh and salt water.
Podophrya gracilis, n. sp. Fig. 65.
Of all the Podophrya that have been described not one approaches this minute form in the relative length of the stalk. The body is spherical and is covered with short capitate tentacles. The stalk is extremely slender, bent, and without obvious structure. There are one or two contractile vacuoles in the distal half of the body. The nucleus is small and is situated near the insertion-point of the stalk. Reproduction not observed. Diameter of body 8µ; length of stalk 40µ. Only one specimen seen.
Small to medium-sized and large forms; colorless to brown. The body is globular or oval or wedge-shape, sometimes quadrangular. The stalk is variable, sometimes 1 mm. in length. The diameter of the stalk increases from the point of attachment to the body of the animal; it is usually striated either longitudinally or transversely, or both. The tentacles are of two kinds and are usually confined to the anterior half of the body. Some are long and sharp-pointed and adapted for piercing; others are short, cylindrical, usually retracted and capitate, adapted for sucking. Contractile vacuoles vary from one to many. The macronucleus is nearly central in position and usually of horseshoe shape, but is frequently branched and irregular. Reproduction is accomplished by external multiple budding, usually from the anterior half of the body. Salt water.
Ephelota coronata Str. Wright. Fig. 66.
Synonyms: Hemiophrya gemmipara S. K.; Podophrya gemmipara Hertwig.
The body is spheroidal, ovate, or pyriform, with numerous sharp-pointed tentacles and a few straight, uniform tentacles. The stalk is about three times the length of the body and tapers from its widest part at the insertion in the body to the narrowest part at the point of attachment. It may or may not be longitudinally striated. This is one of the commonest of the Suctoria found at Woods Hole. It is usually present on Campanularian hydroids, but may be found on algæ and Bryozoa.
Length of body 90µ to 200µ.
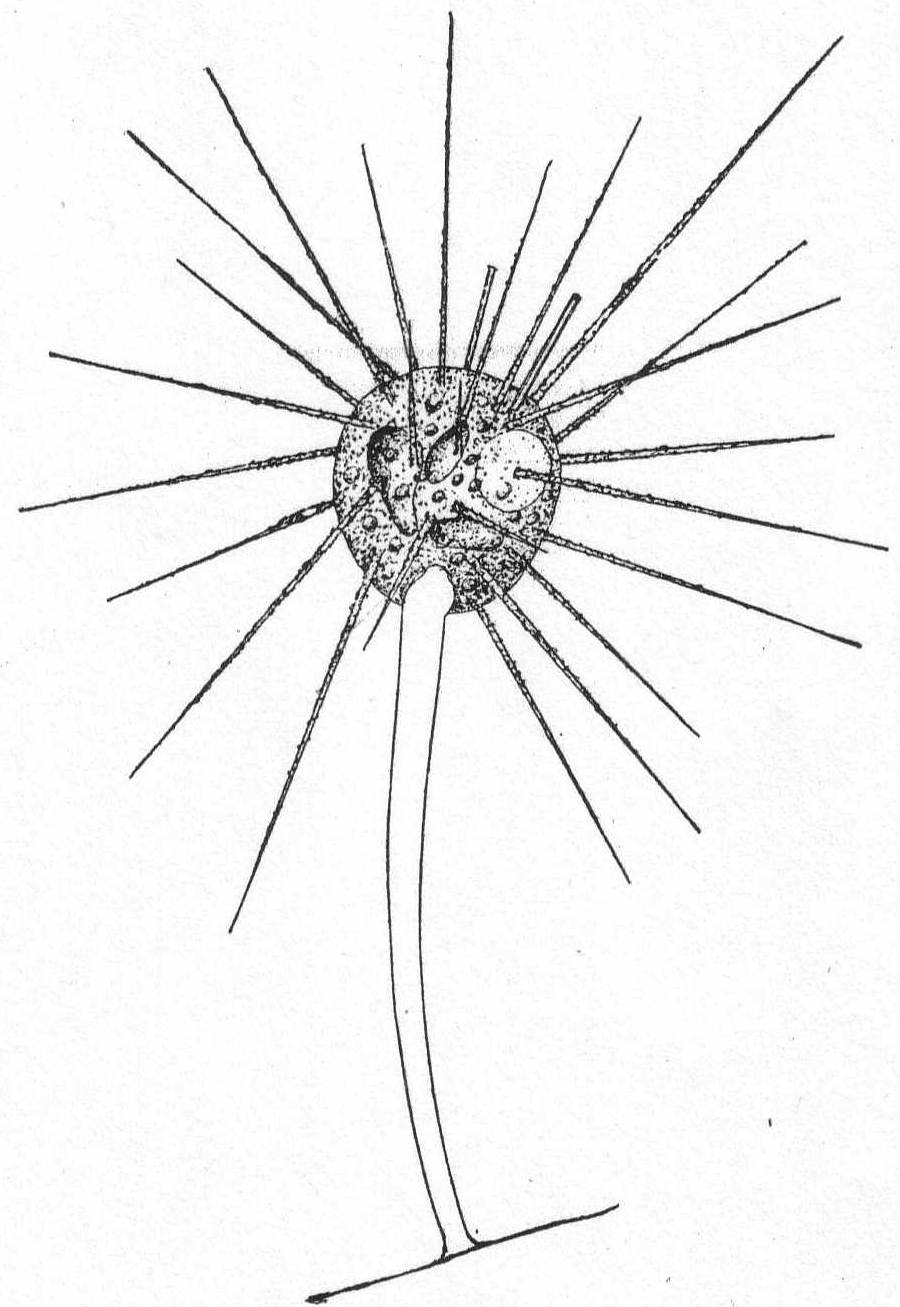
|
Fig. 66.— Ephelota coronata. ENLARGE |
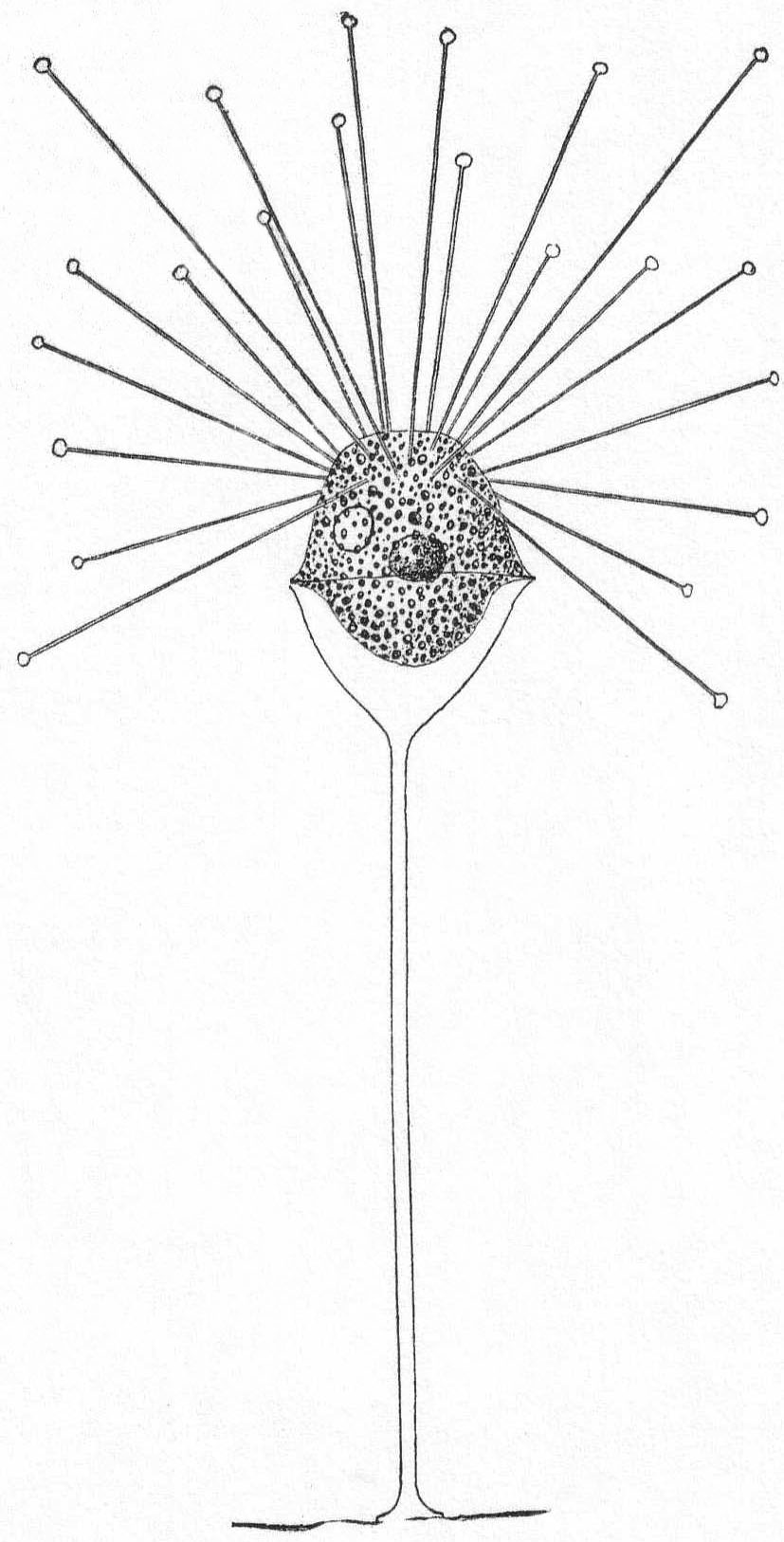 Fig. 67.—Acineta divisa. ENLARGE |
Small to medium-sized forms. The distinguishing feature is that the stalk is swollen at the distal extremity to form a cup or basin in which the animal rests. The cup may be developed until the body is nearly inclosed. The macronucleus is spherical or band form. The contractile vacuole is usually single. Budding, so far as known, is endogenous. Fresh and salt water.
Acineta divisa Fraipont '79. Fig. 67.
This extremely graceful form is common on Bryozoa at Woods Hole. The cup is shaped like a wine glass and is specifically characterized by a cup-formed membrane upon which the animal rests. The animal thus has the appearance of being suspended on the edge of the cup. The stalk is slender and about 4 times the length of the body. The tentacles are all capitate and distributed, and about 2-1/2 times the body length. They sway back and forth very slowly. The nucleus is spherical and central in position. The contractile vacuole lies near the periphery.
Length of body 27µ; of stalk 100µ; of extended tentacle 65µ.
Acineta tuberosa Ehr. Fig. 68.
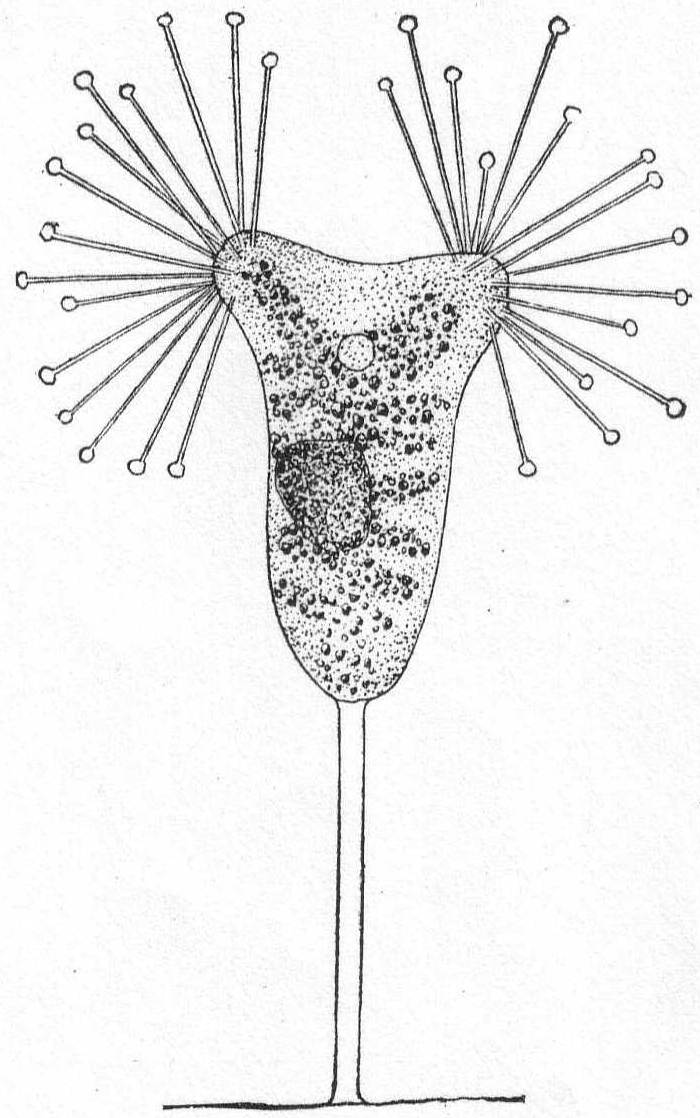
|
Fig. 68.— Acineta tuberosa. ENLARGE |
Large forms of Suctoria with tentacles arranged in fascicles. The stalk is variable in length and the cup is frequently so delicate that it can barely be made out. A specific characteristic is the break in continuity of the cup at different points, and through these places the tentacles emerge in bundles. The tentacles are capitate and in the Woods Hole form, 15 in number in each of the two bundles. The endoplasm is granular and yellowish in color. The coloring matter is frequently arranged in patterns. The nucleus is spheroidal. The contractile vacuole is in the anterior third of the body about midway between the bundles of tentacles. Reproduction not observed.
Length of body 330µ.
Small forms to medium size; no cups or stalks. The body is spherical to elongate, usually, however, more or less irregularly lobed and changeable. The tentacles are in fascicles which are usually borne upon lobed or swollen places. The body is always more or less spread out. Contractile vacuoles variable. The macronucleus is spherical, elongate, band-formed or horseshoe-shaped. Reproduction takes place by endogenous budding, and the swarm spores are flat or lenticular with a distinct ciliary girdle. They are frequently parasitic. Fresh and salt water.
Trichophrya salparum Entz '84. Fig. 69.
Bütschli '88; Schewiakoff '93; Trichophrya ascidiarum Lachmann '59; René Sand 1901.
The body is somewhat cup-form, with a large, flat base. The anterior border is rounded, each of the ends being somewhat truncate and carrying a bundle of tentacles all capitate and similar. These may be continued internally as far as the nucleus (Sand). The cytoplasm is uncolored, but may contain some brilliant granules. The nucleus is granular, and spherical, band or horseshoe formed.
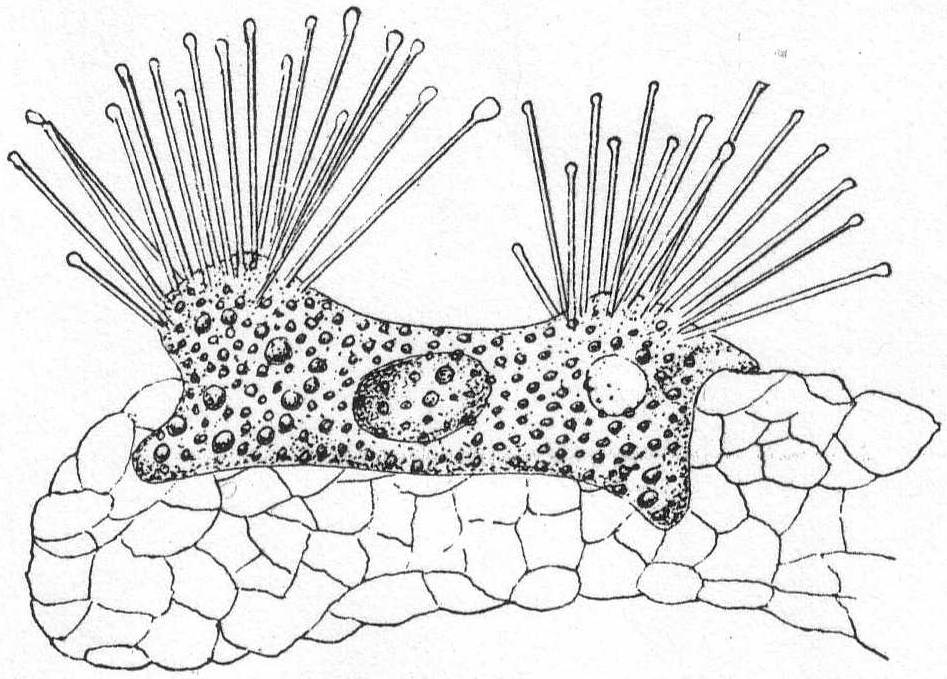
|
Fig. 69.— Trichophrya salparum. ENLARGE |
This species was found by Dr. G. Hunter on the branchial bars of the Ascidian Molgula manhattensis, where great numbers of them are often parasitic.
AUERBACH, L. '54. Ueber Encystierung von Oxytricha pellionella. Zeit. wiss. Zool., V, 1854.
–––– '55. Ueber die Einzelligkeit der Amoeben. Zeit. wiss. Zool., VII, 1855.
BALBIANI E. G. '61. Recherches sur les phénomènes sexuelles des Infusoires. Jour. de la physiol:, IV, 1861.
–––– '85. Sur un Infusoire parasite du sang de l'Aselle aquatique. Rec. zool. Suisse, II, 1885.
BERGH, R. S. '79. Tiarina fusus Cl. & Lach. Vid. Med. f. d. Nat. Foren. Kjobenhavn.
–––– '82. Der Organismus der Cilioflagellaten. Morph. Jahr., VII, p. 177.
BRADY, H. B. '79. Report on the Foraminifera dredged by H. M. S. Challenger during the years 1873-1876. Challenger Reports, Zoology, IX.
BRANDT, K. '96. Die Tintinnodeen. Bibliot. Zoolog., XX, 1896.
BÜTSCHLI, O. '76. Studien über die ersten Entwickelungsvorg. der Eizelle, die Zelltheilung, und die Conjugation der Infusorien. Abhand. der Senck. naturf. Ges. Freiburg, X, 1876.
–––– '83-'88. Die Protozoen. Bronn's Klassen u. Ord. des Tierreichs.
CARPENTER, W. B. '56. Researches on the Foraminifera. Phil. Trans. (2), p. 547.
CARTER, H. J. '56. Notes on the fresh-water Infusoria of the island of Bombay. Ann. Mag. Nat. Hist. (2), XVIII.
–––– '63. On the coloring matter of the Red Sea. Id. (3), XI.
–––– '63a. On Amœba princeps and its reproductive cells. Id. (3), XII.
–––– '64. On fresh-water Rhizopods of England and India. Id. (3), XIII, p. 13.
–––– '65. On the fresh- and salt-water Rhizopods of England and India. Id. (3), XV, p. 277.
CIENKOWSKY, L. '55. Bemerkungen über Stein's Acinetenlehre. Bull. Phys. Math. Acad., St. Petersburg, XIII, p. 297 (also in Ann. Mag. Nat. Hist. (3), XIII).
–––– '55. Ueber Cystenbildung bei Infusorien. Zeit. wiss. Zool., VI, p. 301.
–––– '61. Beiträge zur Kenntniss der Monaden. Arch. f. mik. Anat., I, 1865, p. 203.
CLAPARÈDE, E. '54. On Actinophrys sol. Ann. Mag. Nat. Hist. (2), XV, 1854.
–––– '60. Recherches sur les annélides, etc., observés dans les Hébrides. Mém. Soc. phys. d'hist. nat., Genève, XVI, 1860.
–––– '67. Miscellanées zoologiques. Ann. d. sc. nat. zool. (5), VIII, 1867.
CLAPARÈDE ET LACHMANN '58-'60. Etudes sur les Infusoires et les Rhizopodes. Mém. Inst. genèvoise, V, VI, VII.
COHN, F. '54. Beiträge z. Entwicklungsges. der Infusorien. Zeit. wiss. Zool. III, 1851; IV, 1853.
–––– '57. Ueber Fortpflanzung von Nassula elegans Ehr. Zeit. wiss. Zool., IX, 1857, p. 143.
COSTE, M. '64. Développement des Infusoires ciliés dans une macération de foin. Ann. d. ac. nat. zool.(5), II.
DADAY, E. v. '86. Ein kleiner Beitrag der Infusorien-Fauna des Golfes v. Neapel. Mitt. Zool. St. Neap., VI, p. 481.
–––– '88. Monographie der Familie der Tintinnodeen. Mitt. Zool. St. Neap., VIII, p. 473.
DAVIS, J. '79. On a new species of Cothurnia. Jour. Roy. Mic. Soc., II, 1879.
DUJARDIN, F. '35. Recherches sur les organismes inférieures. Ann. d. sc. nat. zool. (2), IV, p. 343.
–––– '41. Histoire naturelle des Zoophytes infusoires. Paris, 1841.
EHRENBERG, C. G. '31. Ueber die Entwickelung und Lebensdauer d. Infusionsthiere. Abhand. d. Berlin Ak., 1831, p. 1.
–––– '38. Die Infusionsthierchen als vollkommne Organismen. Leipzig, 1838.
ENGELMANN, Th. W. '78. Zur Physiologie der Contractilen Vacuolen der Infusionsthiere. Zool. Anz., I, p. 121.
ENTZ, G. '84. Ueber Infusorien d. Golfes v. Neapel. Mitt. Zool. St. Neap., V, p. 289.
–––– '85. Zur naheren Kenntniss der Tintinnodeen. Mitt. Zool. St. Neap., VI, p. 185.
FABRE-DOMERGUE, P. '85. Note sur les Infusoires ciliés de la baie de Concarneau. Jour. d. l'anat. et de la phys., XXI, p. 554.
–––– '91. Matériaux pour servir a l'histoire des Infusoires ciliés. Ann. d. mic., III, 1890-91, p. 49.
FOL, H. '83. Further contribution to the knowledge of the Tintinnodea. Ann. Mag. Nat. Hist. (5), XII, p.73.
FRANCÉ, R. H. '97. Der organismus der Craspedomonaden. Budapest.
FROMENTEL. E. DE. '74. Etudes sur les microzoaires, etc. Paris, 1874.
GOURRET ET ROESER. '88. Contributions à l'étude des Protozoaires de la Corse. Arch. d. biol., VIII p. 139.
GREEFF, R. '66. Ueber einige in der Erde lebende Amoeben und andere Rhizopoden. Arch. f. mik. Anat., II, p. 299.
–––– '70. Untersuchungen ueber den Bau u. d. Naturgeschichte d. Vorticellen. Arch. f. Naturges., XXXVI, XXXVII.
–––– '71. Ueber die Actinophryens oder Sonnenthierchen des Süssenwassers als echte Radiolarien. Sitz. Ber. d. Niederrh. Ges. i. Bonn, XXVIII, p. 4.
–––– '75. Ueber Radiolarien u. Radiolarienartige Rhizopoden des Süssenwassers 2. Arch. f. mik. Anat., XI.
–––– '88. Studien über Protozoen. Sitz. Ber. Ges. z. Bef. d. ges. Nat., Marburg, 1888, p. 91.
GRENACHER, H. '68. Ueber Actinophrys sol Ehr. Verh. phys.-med. Ges. Freiburg (n. F.), I.
GRUBER, A. '83. Beobachtungen an Chilodon curvidentis n. sp. Festschr. 56. Vers. Deutsch. Naturf. gewid. v. d. Naturf. Ges. Freiburg, II, p. 1.
–––– '84. Die Protozoen des Hafens v. Genua. Nova Act. d. K. Leop.-Car. Deutsch. Akad. d. Naturf., XLVI, p. 475.
–––– '87. Ueber der Bedeutung der Conjugation bei den Infusorien. Ber. d. Naturf. Ges. Freiburg, II, p. 31.
HÆCKEL, E. '73. Zur Morphologie der Infusorien. Jena Zeit., VII, p. 516.
HERTWIG, R. '76. Ueber Podophrya gemmipara, nebst Bemerkungen zum Bau u. d. systemat. Stellung d. Acineten. Morph. Jahr., IV, p. 20.
HERTWIG U. LESSER. Ueber Rhizopoden u. denselben nahe stehende Organismen. Arch. F. mik. Anat., X, Suppl., p. 35.
HUXLEY, T. H. '57. On Dysteria, a new genus of Infusoria. Jour. Mic. Sci., V, p. 78.
ISHIKAWA, C. Ueber eine in Misaki vorkommende Art v. Ephelota, etc. Journ. Coll. Sci. Imp. Univ. Tokyo, Japan, X, pt. 2.
JAMES-CLARK, H. '66. On the Spongiæ ciliatæ as Infusoria flagellata, etc. Mem. Bost. Soc. Nat. Hist. (3), I, p. 1.
KENT, W. SAVILLE. '81. Manual of the Infusoria. London, 1881.
KLEBS, G. '84. Ein kleiner Beitrag z. Kenntniss d. Peridineen. Bot. Zeit., XLII, p. 721.
–––– '92. Flagellaten Studien 1. Zeit. wiss. Zool., LV.
LABBÉ, A. '95. Sur les Protozoaires marins de Roscoff. Arch. d. zool. expér. (3), N. et R., p. XIV.
LAUTERBORN, R. '94. Beiträge z. Süsswasserfauna der Insel Helgoland. Wiss. Meeresunt. Komm. wiss. Unt. d. Meere Kiel (2), I, p. 215.
LEIDY, J. '77. Remarks upon Rhizopods and notice of a new form. Proc. Ac. Sci. Phila., 1877, p. 293.
–––– '79. Fresh water Rhizopods of North America. Washington, 1879.
LIEBERKÜHN, N. '56. Ueber Protozoen. Notes from a letter to C. Th. v. Siebold. Zeit. wiss. Zool., VIII, p. 307.
MAUPAS, E. '81. Contributions à l'étude des Acinétiens. Arch. d. zool. expér. (1), IX, p. 299.
–––– '83. Contributions à l'étude morphologique et anatomique des Infusoires ciliés. Id. (2), I.
–––– '83a. Les sucto-ciliés de M. Mereschowsky. Comp. Ren., XCV, p. 1381.
–––– '88. Recherches expérimentales sur la multiplication des Infusoires ciliés. Arch. d. zool. exper. (2), VI, p. 165.
MERESKOWSKY, C. '78. Studien über Protozoen des nordlichen Russland. Arch. f. mik. Anat., XVI, p. 163.
–––– '81. On some new or little-known Infusoria. Ann. Mag. Nat. Hist. (5), VII, 1881.
MOEBIUS, K. '88. Bruchstücke einer Infusorienfauna der Kieler Bucht. Arch. f. Naturg., 1888.
PECK, J. I. '93. On the food of the menhaden. Bull. U. S. Fish Com., 1893, p. 113.
–––– '95. The sources of marine food. Bull. U. S. Fish Com., 1895, p. 351.
PERTY, M. '49. Mikrosk. Organ. der Alpen u. d. Italien. Schweiz. Mitt. d. Naturf. Ges. in Bern, 164-165.
–––– '52. Zur Kenntniss kleinster Lebensformen, etc., Bern.
POUCHET, G. '83; '85. Contributions à l'histoire des Péridiniens marins. Jour. de l'anat. et de la phys., XIX, XXI.
QUENNERSTEDT, A. '65; '67; '69. Bidrag till Sveriges Infusorie-fauna. Lunds Univ. Ärsskrift, II, IV, VI.
SAND, R. 1901. Etude monographique sur le groupe des Infusoires tentaculifères. Ann. d. la Soc. belge de microscopie, XXIV, XXV, XXVI.
SCHAUDINN, F. '95. Die Heliozoen. Das Tierreich, 1895.
SCHEWIAKOFF, W. '89. Beiträge z. Kenntniss der Holotrichen Ciliaten. Bib. Zool., V, p. 1.
–––– '93. Ueber einige ecto- u. ento-parasitische Protozoen der Cyclopiden. Bull. Soc. nat. Moscou, 1893. I.
SCHUETT, F. '95. Die Peridineen d. Plankton-Expedition, 1. Kiel u. Leipzig.
SCHULTZE, F. E. '74; 75. Rhizopodenstudien. Arch. f. mik. Anat., X, XI, XIII.
SCHULTZE, M. '62. Ueber d. Organismus d. Polythalamien. Leipzig, 1862.
SHEVYAKOV ?. '96. Monograph on Holotrichous ciliates. (In Russian.) Mem. of the St. Petersburg Acad., VII.
STEIN, F. '59; '78; '83. Der Organismus der Infusionsthiere. I. Infusoria, '59; II. Infusoria, '78; III. Flagellata, '83.
–––– '49. Untersuchung über die Entwicklungs d. Infusorien. Arch. f. Natur., I, p. 92.
–––– '54. Die Infusionsthiere auf ihre Entwicklungsgeschichte untersucht. Leipzig, 1854.
–––– '60. Ueber Leucophrys patula u. über 2 neue Infusoriengattungen Gyrocoris u. Lophomonas. Sitz. Ber. d. K.-böhm. Ges. d. Wiss. d. Prag, 1860, p. 4.
–––– '64. Ueber die neue Gattung Epiclintes. Id., 1864, I.
STERKI, V. '78. Beiträge z. Morphologie der Oxytrichinen. Zeit. wiss. Zool., XXXI, p. 29.
–––– '98. On the classification of ciliate Infusoria. Amer. Natur., XXXII, p. 425.
STEVENS, N. M. 1901. Studies on ciliate Infusoria. Proc. Calif. Acad. Sciences, III, 1.
STOKES, A. C. '84. Notices of some new parasitic Infusoria. Amer. Nat., XVIII, p. 1081.
–––– '85. Some apparently undescribed Infusoria from fresh water. Id., XIX, p. 18.
–––– '87. Some new hypotrichous Infusoria from American fresh waters. Ann. Mag. Nat. Hist. (5), 20.
TATEM, T. G. '67. New species of microscopic animals. Q. J. M. S. (n.s.), VII, p. 251.
VEJDOWSKY, F. '79. Monographie der Enchytraeiden. Prag, 1869.
WALLENGREN, H. '94. Studier ofver ciliata Infusorier, 1. Slagtet Lichnophora. Lund, 1894.
WALLICH, G. C. '63. Observations on an undescribed indigenous Amœba. Ann. Mag. Nat. Hist. (3), XI, XII.
WESTON, J. '56. On the Actinophrys sol. Q. J. M. S., IV, p. 116.
WRZESNIOWSKI A. '61. Observations sur quelques Infusoires. Ann. d. sc. nat. zool. (6), XVI.
–––– '69. Ein Beitrag zur Anatomie der Infusorien. Arch. f. mik. Anat., V, p. 25.
–––– '70. Beobachtungen über Infusorien a. d. Umgebung v. Warschau. Zeit. wiss. Zool., XX, p. 467.
***END OF THE PROJECT GUTENBERG EBOOK MARINE PROTOZOA FROM WOODS HOLE***
******* This file should be named 18320-h.txt or 18320-h.zip *******
This and all associated files of various formats will be found in:
http://www.gutenberg.org/1/8/3/2/18320
Updated editions will replace the previous one--the old editions will be renamed.
Creating the works from public domain print editions means that no one owns a United States copyright in these works, so the Foundation (and you!) can copy and distribute it in the United States without permission and without paying copyright royalties. Special rules, set forth in the General Terms of Use part of this license, apply to copying and distributing Project Gutenberg-tm electronic works to protect the PROJECT GUTENBERG-tm concept and trademark. Project Gutenberg is a registered trademark, and may not be used if you charge for the eBooks, unless you receive specific permission. If you do not charge anything for copies of this eBook, complying with the rules is very easy. You may use this eBook for nearly any purpose such as creation of derivative works, reports, performances and research. They may be modified and printed and given away--you may do practically ANYTHING with public domain eBooks. Redistribution is subject to the trademark license, especially commercial redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License (available with this file or online at
http://www.gutenberg.org/license).
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH F3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS,' WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTIBILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need, is critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation web page at http://www.gutenberg.org/fundraising/pglaf.
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at
809 North 1500 West, Salt Lake City, UT 84116, (801) 596-1887, email
[email protected]. Email contact links and up to date contact
information can be found at the Foundation's web site and official
page at http://www.gutenberg.org/about/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
[email protected]
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit http://www.gutenberg.org/fundraising/pglaf
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations.
To donate, please visit: http://www.gutenberg.org/fundraising/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart is the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For thirty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Each eBook is in a subdirectory of the same number as the eBook's
eBook number, often in several formats including plain vanilla ASCII,
compressed (zipped), HTML and others.
Corrected EDITIONS of our eBooks replace the old file and take over
the old filename and etext number. The replaced older file is renamed.
VERSIONS based on separate sources are treated as new eBooks receiving
new filenames and etext numbers.
Most people start at our Web site which has the main PG search facility:
http://www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.
EBooks posted prior to November 2003, with eBook numbers BELOW #10000,
are filed in directories based on their release date. If you want to
download any of these eBooks directly, rather than using the regular
search system you may utilize the following addresses and just
download by the etext year.
http://www.gutenberg.org/dirs/etext06/
(Or /etext 05, 04, 03, 02, 01, 00, 99,
98, 97, 96, 95, 94, 93, 92, 92, 91 or 90)
EBooks posted since November 2003, with etext numbers OVER #10000, are
filed in a different way. The year of a release date is no longer part
of the directory path. The path is based on the etext number (which is
identical to the filename). The path to the file is made up of single
digits corresponding to all but the last digit in the filename. For
example an eBook of filename 10234 would be found at:
http://www.gutenberg.org/dirs/1/0/2/3/10234
or filename 24689 would be found at:
http://www.gutenberg.org/dirs/2/4/6/8/24689
An alternative method of locating eBooks:
http://www.gutenberg.org/dirs/GUTINDEX.ALL
*** END: FULL LICENSE ***