This eBook is for the use of anyone anywhere at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org
Title: Outlines of Lessons in Botany, Part I; From Seed to Leaf
Author: Jane H. Newell
Release Date: January 16, 2004 [eBook #10726]
Language: English
Character set encoding: ISO-8859-1
***START OF THE PROJECT GUTENBERG EBOOK OUTLINES OF LESSONS IN BOTANY, PART I; FROM SEED TO LEAF***
In this study, as in all scientific teaching, the teacher's aim should be to foster in his pupils the power of careful observation and clear expression. The actual amount of knowledge gained at school must needs be small, and often quickly forgotten, but the habit of right study is an invaluable possession.
The former method of teaching Botany was confined almost wholly to dry, technical classification. The pupil learned to find the name and order of a plant, but its structure, its habits, its life in short, were untouched by him. We know now that Nature is the best text-book. The pupil should first ask his questions of her and try to interpret her answers; then he may learn with profit what those who better understand her speech have to tell him.
This method of teaching, however, requires much, very much, of the teacher. He must be himself intelligent, well trained, and able to give time to the preparation of his lessons. It seems to us, who are but amateurs, as if it were impossible to teach thus without a thorough comprehension of the whole field. Our own ignorance oppresses us so much that we feel tempted to say that we cannot attempt it. But if the work of leading children to observe the wonders about them is to be done at all, it must be done by us, who are not masters of our subject, and we must find out for ourselves how we can best accomplish this result, since we have so little to guide us.
It is with the hope that the experience of one who has tried to do this with some fair amount of success may be of use to other puzzled experimenters, that I venture to write out some outlines of lessons in Botany for beginners.
The method of beginning with the simpler forms of life is one that appeals to the scientific tendencies of the day. It seems logical to begin with lower forms and work up to the higher. But this method is only suitable for mature minds. We do not teach a child English by showing him the sources of the language; he learns it by daily use. So also the beginning of the study of any Natural Science by the young should be the observation of the most obvious things about them, the things which they can see, and handle, and experiment upon naturally, without artificial aids. Therefore this book concerns itself only with the Flowering Plants.
The author believes that the simplest botanical study should afford the means of identifying plants, as a large part of the student's pleasure in the science will be the recognition of the things about him. The present volume affords the basis for future classification, which Part II, on flowers, will develop. It is, doubtless, as good a way, perhaps the best, to begin with a single plant, and study root, stem, leaves, and flowers as belonging to a whole, but the problem is complicated by practical difficulties. In our climate there are but two months of the school year when flowers are easily obtained. On the other hand, the material for these lessons can be got throughout the winter, and the class, well trained in methodical work, will begin the study of flowers at the season when every day brings some fresh wonder of beauty.
The author will receive gladly any criticisms or suggestions.
JANE H. NEWELL.
175 Brattle St., Cambridge.
INTRODUCTION.
The lessons here outlined are suitable for children of twelve years of age, and upwards. For younger pupils they would require much adaptation, and even then they would not be so good as some simpler method, such as following the growth of one plant, and comparing it with others at every step. The little ones profit most by describing the very simple things that they see, without much reference to theories.
The outlines follow the plan of Dr. Gray's First Lessons and How Plants Grow, and are intended to be used in connection with either of those books. The necessary references will be found at the end of every section. The book contains also references to a course of interesting reading in connection with the subjects of the lessons.
The lessons may begin, like the text-books, with the subject of Germination, if the seeds are planted before they are required for use, but it is generally preferable to use the first recitation with the class for planting the seeds, in order to have them under the direct care of the pupils. Some general talks about plants are therefore put at the beginning to occupy the time until the seedlings are ready for study.
Some Nasturtiums (Tropæolum majus) and Morning-Glories should be planted from the first in boxes of earth and allowed to grow over the window, as they are often used for illustrations.
What is Botany? The pupils are very apt to say at first that it is learning about flowers. The teacher can draw their attention to the fact that flowers are only a part of the plant, and that Botany is also the study of the leaves, the stem, and the root. Botany is the science of plants. Ask them what the Geranium is. Tell them to name some other plants. The teacher should keep a few growing plants in the schoolroom for purposes of illustration.
Ask them what else there is in the world besides plants. By this question the three kingdoms, animal, vegetable, and mineral, are brought up. It will give occasion for a discussion of the earth and what it contains, the mountains, formed of rocks and soil, the plants growing on the earth, and the animals that inhabit it, including man. Let them name the three kingdoms with some example of each. Which of these kingdoms contain living things? The words organic and inorganic can be brought in here. An organ (Εργον, meaning work) is any part that does a special work, as the leaves, the stem of a plant, and the eye, the ear of animals. An organism is a living being made up of such organs. The inorganic world contains the mineral kingdom; the organic world includes the vegetable and animal kingdoms.
One's aim in these lessons should always be to tell the pupils as little as possible. Try to lead them to think out these things for themselves.
Ask them how plants differ from animals. They will say that plants are fixed to one place, while animals can move about; that plants have no will or consciousness, and that animals have. These answers are true when we compare the higher animals with plants, but the differences become lost as we descend in the scale and approach the border land where botanist and zoologist meet on a common ground. Sea-anemones are fixed to the rock on which they grow, while some of the lower plants are able to move from place to place, and it is hardly safe to affirm that a jelly-fish is more conscious of its actions than is a Sensitive Plant, the leaves of which close when the stem is touched.
There is no real division between animals and plants. We try to classify the objects about us into groups, according to the closeness of their relationships, but we must always remember that these hard lines are ours, not Nature's. We attempt, for purposes of our own convenience, to divide a whole, which is so bound together that it cannot be separated into parts that we can confidently place on different sides of a dividing line.
1. Plants as Food-Producers.—The chief distinguishing characteristic of plants is one that the pupils may be led to think out for themselves by asking them what animals feed upon. To help them with this, ask them what they had for breakfast. Oatmeal is mentioned, perhaps. This is made from oats, which is a plant. Coffee and tea, bread made from wheat, potatoes, etc., all come from plants.[1] Beef, butter and milk come from the cow, but the cow lives upon grass. The plant, on the other hand, is nourished upon mineral or inorganic matter. It can make its own food from the soil and the air, while animals can only live upon that which is made for them by plants. These are thus the link between the mineral and animal kingdoms. Ask the scholars if they can think of anything to eat or drink that does not come from a plant. With a little help they will think of salt and water. These could not support life. So we see that animals receive all their food through the vegetable kingdom. One great use of plants is that they are food-producers.
This lesson may be followed by a talk on food and the various plants used for food.[2]
2. Clothing.—Plants are used for clothing. Of the four great clothing materials, cotton, linen, silk, and woollen, the first two are of vegetable, the last two of animal origin. Cotton is made from the hairs of the seed of the cotton plant.[1] Linen is made of the inner fibre of the bark of the flax plant. It has been cultivated from the earliest historical times.
3. Purification of the Air.—The following questions and experiments are intended to show the pupils, first, that we live in an atmosphere, the presence of which is necessary to support life and combustion (1) and (2); secondly, that this atmosphere is deprived of its power to support life and combustion by the actions of combustion (2), and of respiration (3); thirdly, that this power is restored to the air by the action of plants (4).
We have the air about us everywhere. A so-called empty vessel is one where the contents are invisible. The following experiment is a good illustration of this.
(1) Wrap the throat of a glass funnel with moistened cloth or paper so that it will fit tightly into the neck of a bottle, and fill the funnel with water. If the space between the funnel and the bottle is air-tight, the water will not flow into the bottle.
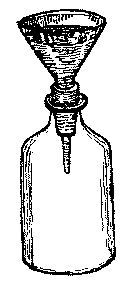
[Illustration: FIG. 1.]
Do not explain this in advance to the pupils. Ask them what prevents the water from flowing into the bottle. If they are puzzled, loosen the funnel, and show them that the water will now flow in. In the first case, as the air could not escape, the water could not flow in; in the second, the air was displaced by the heavier water.
Ask the pupils why the air in a crowded room becomes so difficult to breathe. Could a person live if he were shut up in an air-tight room for a long time? Fresh air is necessary to life. The teacher may explain that it is the oxygen in the air that supports life. Air is composed one-fifth of this gas and four-fifths of nitrogen. The gases are mixed and the nitrogen simply dilutes the oxygen, as it were.
Fresh air is necessary to support combustion as well as life. Ask them why we put out a fire by throwing a blanket or a rug over it. The following experiment illustrates this.
(2) Take a small, wide-mouthed bottle, covered with a card or cork. To this cover fasten a piece of bent wire with a taper on the end. Light the taper and lower it into the jar. It will burn a few seconds and then go out. Raise and light it again, and it will be extinguished as soon as it is plunged into the bottle. This shows that the oxygen of the air is used up by burning substances, as it is by breathing animals.
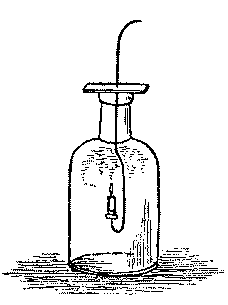
[Illustration: FIG. 2.]
The following experiment shows that fire will not burn in an atmosphere of gas from our lungs.
(3) Fill a bottle with gas by breathing into it through a bit of glass tubing, passed through a card or cork, and reaching to the bottom of the bottle. The bottle will be dimmed with moisture, showing the presence of aqueous vapor. A lighted match plunged into the bottle will be immediately extinguished. A better way, which, however, takes some skill in manipulation, is to fill the bottle with water, cover it with a flat piece of glass, and invert the bottle in a dish of water, taking care that no air bubbles enter. Then, through a bit of glass tubing, blow into the bottle till the water is expelled. Cover the mouth with the glass under water, and holding it tightly down, invert the bottle quickly. Set it down, light a match, take away the glass, and at the same instant plunge in the match. If no air has been allowed to enter, the match will go out at once. No animal could live in an atmosphere which could not support combustion.
From these experiments the pupils have seen that the life-sustaining quality of the air is used up by combustion and respiration. To bring in the subject of purification by plants, ask them why all the oxygen in the world is not exhausted by the people and the fires in it. After the subject has been explained, the following experiment can be prepared and put aside till the next lesson.
(4) Fill two bottles with air from the lungs, as in (3) having previously introduced a cutting from a plant into one of the bottles. Allow them to stand in the sun for a day or two. Then test both bottles with a burning match. If properly done, the result will be very striking. The end of the cutting should be in the water of the dish. This experiment will not succeed excepting with bottles such as are used for chemicals, which have their mouths carefully ground. Common bottles allow the air to enter between the bottle and the glass.[1]

[Illustration: FIG. 3.]
4. Fuel.—Light a match and allow it to burn until half charred. Blow it out gently, so as to leave a glowing spark. When this spark goes out it will leave behind a light, gray ash. We have to consider the flame, the charred substance, and the ash.
Flame is burning gas. In all ordinary fuels, carbon and hydrogen, in various combinations and free, make the principal part. The first effect of the heat is to set free the volatile compounds of carbon and hydrogen. The hydrogen then begins to unite with the oxygen of the air, forming water, setting free the carbon, which also unites with oxygen, forming carbonic acid gas. The burning gases cause the flame. The following experiment will illustrate this.
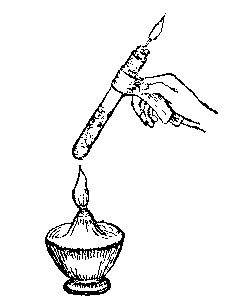
[Illustration: Fig. 4.]
(5) Fit a test-tube with a tight cork, through which a bit of glass tubing, drawn out into a jet, is passed, the tubing within being even with the cork. Place some bits of shaving in the tube, cork it, and make the cork perfectly air-tight by coating it with bees wax or paraffine. Heat the test-tube gently over an alcohol lamp. The wood turns black, and vapor issues from the jet, which may be lighted (Fig. 4). Care should be taken to expel all the air before lighting.
(6) That the burning hydrogen forms water by uniting with the oxygen of the air, may be shown by holding a cold glass tumbler over the jet, or over any flame. The glass will be dimmed by drops of moisture.
The charred part of the wood is charcoal, which is one form of carbon. Our ordinary charcoal is made by driving off all the gases from wood, by burning it under cover where only a little air can reach it. The volatile gases burn more readily than the carbon, and are the first substances to be driven off, so that the carbon is left behind nearly pure. In the same way we have driven off all the gases from the half-burned match and left the carbon. The teacher should have a piece of charcoal to show the pupils. It still retains all the markings of the wood.
If the combustion is continued, the carbon also unites with the oxygen of the air, till it is all converted into carbonic acid gas. This was the case with the match where we left the glowing spark. The gray ash that was left behind is the mineral matter contained in the wood.
(7) We can show that this gas is formed by pouring lime water into a bottle in which a candle has been burned as in (2). The water becomes milky from a fine white powder formed by the union of the carbonic acid gas with the lime, forming carbonate of lime. This is a chemical test.
The wood of the match is plainly of vegetable origin; so also is the charcoal, which is nearly pure carbon. Coal is also carbon, the remains of ancient forests, from which the gases have been slowly driven off by heat and pressure. All the common fuels are composed principally of carbon and hydrogen. When these elements unite with oxygen, carbonic acid gas and water are formed.[1]
(8) The same products are formed by respiration. We breathe out carbonic acid gas and water from our lungs. Breathe on a cold glass. It is bedewed exactly as it is by the candle flame. Breathe through a bit of glass tubing into a bottle of lime water. It becomes milky, showing the presence of carbonic acid gas. Why is this?
Every act or thought is accompanied by a consumption of material in the body, which thus becomes unfit for further use. These waste substances, composed chiefly of carbon and hydrogen, unite with oxygen breathed in from the air, forming carbonic acid gas and water, which are breathed out of the system. The action is a process of slow combustion, and it is principally by the heat thus evolved that the body is kept warm. As we are thus constantly taking oxygen from the air, a close room becomes unfit to live in and a supply of fresh air is indispensable. The cycle of changes is completed by the action of plants, which take in carbonic acid gas, use the carbon, and return most of the oxygen to the atmosphere.
APPARATUS FOR EXPERIMENTS.[1]
Two small wide-mouthed bottles. A narrow-necked bottle. A glass funnel. A bit of bent glass-tubing. A bit of straight glass-tubing. A flat piece of glass. A test-tube, with jet. An alcohol lamp. A bent wire with taper. A card. A slip of a plant. A dish and pitcher of water. Beeswax or paraffine. Shavings. Lime water. Matches.
Gray's First Lessons. Revised edition. Sect. XVI, 445-7, 437.
How Plants Grow. Chap. III, 279-288.
1. Directions for raising in the Schoolroom.—The seeds should be planted in boxes tilled with clean sand. Plates or shallow crockery pans are also used, but the sand is apt to become caked, and the pupils are likely to keep the seeds too wet if they are planted in vessels that will not drain. The boxes should be covered with panes of glass till the seedlings are well started, and should be kept at a temperature of from 65° to 70° Fahr. It is very important to keep them covered while the seeds are germinating, otherwise the sand will be certain to become too dry if kept in a sufficiently warm place. Light is not necessary, and in winter time the neighborhood of the furnace is often a very convenient place to keep them safe from frost. They should not be in the sun while germinating. When the first sprouts appear above the ground let another set be planted, and so on, till a series is obtained ranging from plants several inches high to those just starting from the seed. The seeds themselves should be soaked for a day and the series is then ready for study. The time required for their growth varies according to the temperature, moisture, etc. Dr. Goodale says they should be ready in ten days.[1]
I have never been able to raise them so quickly in the schoolroom, nor have the pupils to whom I have given them to plant done so at home. Generally, it is three weeks, at least, before the first specimens are as large as is desirable.
Germinating seeds need warmth, moisture and air. The necessary conditions are supplied in the very best way by growing them on sponge, but it would be difficult to raise enough for a large class in this manner. Place a piece of moist sponge in a jelly-glass, or any glass that is larger at the top, so that the sponge may not sink to the bottom, and pour some water into the glass, but not so much as to touch the sponge. The whole should be covered with a larger inverted glass, which must not be so close as to prevent a circulation of air. The plants can thus be watched at every stage and some should always be grown in this way. The water in the tumbler will keep the sponge damp, and the roots, after emerging from the sponge, will grow well in the moist air. Seeds can also be grown on blotting paper. Put the seeds on several thicknesses of moist blotting paper on a plate, cover them with more moist paper, and invert another plate over them, taking care to allow the free entrance of air.
If possible, it is by far the best way to have the seeds growing in the schoolroom, and make it a regular custom for the pupils to observe them every morning and take notes of their growth.
These lessons on seeds are suitable for pupils of every age, from adults to the youngest children who go to school. The difference should be only in the mode of treatment; but the same principles should be brought out, whatever the age and power of comprehension of the pupil.
For these lessons the following seeds should be planted, according to the above directions:
Morning-Glory, Sunflower or Squash, Bean, Pea, Red Clover, Flax, Corn, Wheat, and Oats.[1] If they can be procured plant also acorns, Pine-seeds, Maple-seeds, and horsechestnuts.
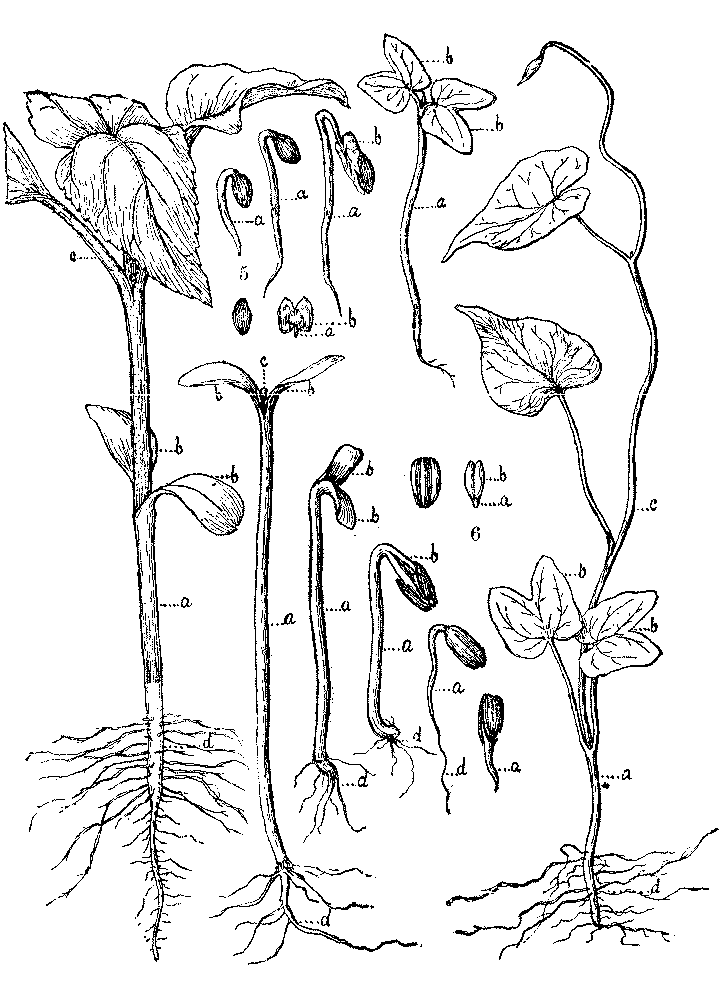
2. Study of Morning-Glory, Sunflower, Bean, and Pea.—For reasons hereafter given, I consider the Morning-Glory the best seedling to begin upon. Having a series, as above described, before them, the pupils should draw the seedlings. When the drawings are made, let them letter alike the corresponding parts, beginning with the plantlet in the seed, and using new letters when a new part is developed. The seed coats need not be lettered, as they do not belong to the plantlet.
[Illustration: FIG. 5.—Germination of Morning Glory, a, caulicle; b, cotyledons; c, plumule; d, roots.]
[Illustration: FIG. 6.—Germination of Sunflower.]
After drawing the Morning-Glory series, let them draw the Sunflower or Squash in the same way, then the Bean, and finally the Pea. Let them write answers to the following questions:
MORNING-GLORY.[1]
Tell the parts of the Morning-Glory seed.
What part grows first?
What becomes of the seed-covering?
What appears between the first pair of leaves?
Was this to be seen in the seed?
How many leaves are there at each joint of stem after the first pair?
How do they differ from the first pair?
SUNFLOWER OR SQUASH.
What are the parts of the seed?
What is there in the Morning-Glory seed that this has not?
How do the first leaves change as the seedling grows?
BEAN.
What are the parts of the seed?
How does this differ from the Morning-Glory seed?
How from the Sunflower seed?
How do the first pair of leaves of the Bean change as they grow?
How many leaves are there at each joint of stem?[1]
How do they differ from the first pair?
PEA.
What are the parts of the seed? Compare it with the Morning-Glory, Sunflower, and Bean.
How does it differ in its growth from the Bean?
What have all these four seeds in common?
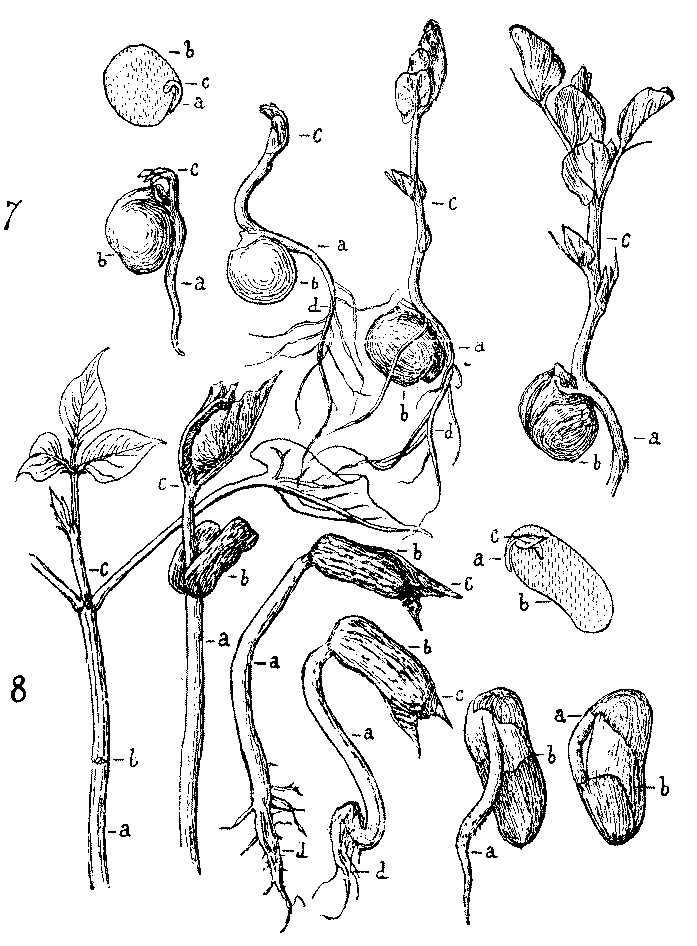
[Illustration: FIG. 7.—Germination of Pea. a, caulicle; b, cotyledons; c, plumule; d, roots.]
[Illustration: FIG. 8.—Germination of Bean.]
What has the Morning-Glory seed that the others have not?
What have the Bean and Pea that the Morning-Glory has not?
How does the Pea differ from all the others in its growth?
What part grows first in all these seeds?
From which part do the roots grow?
What peculiarity do you notice in the way they come up out of the ground?[1]
The teacher must remember that, unless the pupils have had some previous training, they will first have to learn to use their eyes, and for this they will need much judicious help. They should be assisted to see what is before them, not told what is there. It is absolutely necessary that these questions should be thoroughly understood and correctly answered before any conclusions are drawn from them. For this purpose abundant material is indispensable. It is better not to attempt these lessons on seeds at all, unless there is material enough for personal observation by all the pupils.
After this preliminary work has been done, the names of the parts can be given to the pupils. They may be written under each drawing thus,—A=Caulicle;[1] B=Cotyledons; C=Roots; D=Plumule. The whole plantlet in the seed is the embryo or germ, whence the sprouting of seeds is called germination.
I consider this the best order to study the seeds because in the Morning-Glory the cotyledons are plainly leaves in the seed; and in the Squash or Sunflower[1] the whole process is plainly to be seen whereby a thick body, most unlike a leaf, becomes an ordinary green leaf with veins.[2] In the Sunflower the true leaves are nearly the same shape as the cotyledons, so that this is an especially good illustration for the purpose. Thus, without any hint from me, my pupils often write of the Bean, "it has two thick leaves and two thin leaves." In this way the Bean and Pea present no difficulty. The cotyledons in the first make apparently an unsuccessful effort to become leaves, which the second give up altogether.
The teacher's object now is to make the pupils understand the meaning of the answers they have given to these questions. In the first place, they should go over their answers and substitute the botanical terms they have just learned for the ones they have used.
COMPARISON OF THE PARTS OF THE SOAKED SEEDS.
Morning-Glory. A seed covering. Some albumen. Two cotyledons. A caulicle.
Sunflower. An outer covering.[1] An inner covering. Two cotyledons. A caulicle.[2]
Bean. A seed covering. Two cotyledons. A caulicle. A plumule.
Pea. The same as the Bean.
They have also learned how the first leaves in the last three differ from those of the Morning-Glory, being considerably thicker in the Sunflower, and very much thicker in the Bean and Pea. Why should the Morning-Glory have this jelly that the others have not? Why do the first leaves of the Sunflower change so much as the seedling grows? What becomes of their substance? Why do those of the Bean shrivel and finally drop off? By this time some bright pupil will have discovered that the baby-plant needs food and that this is stored around it in the Morning-Glory, and in the leaves themselves in the others. It is nourished upon this prepared food, until it has roots and leaves and can make its own living. The food of the Morning-Glory is called albumen; it does not differ from the others in kind, but only in its manner of storage.[1]
Also the questions have brought out the fact that the Bean and Pea have the plumule ready formed in the seed, while the Morning-Glory and Sunflower have not. Why should this be? It is because there is so much food stored in the first two that the plumule can develop before a root is formed, while in the others there is only nourishment sufficient to enable the plantlet to form its roots. These must make the second leaves by their own labor.
3. Comparison with other Dicotyledons.—The pupils should now have other seeds to compare with these four. Let them arrange Flax, Four o-clock, Horsechestnut, Almond, Nasturtium, Maple-seeds, etc., under two heads.
| Seeds with the Food stored outside the plantlet (Albuminous) | Seeds with the Food stored in the embryo itself (Exalbuminous) |
| Flax. Four-o'clock. Morning-Glory. | Acorn. Horsechestnut. Almond. Maple. Sunflower. Squash. Bean. Pea. Nasturtium. |
They may also be divided into those with and without the plumule.
| Without Plumule | With Plumule |
| Flax. Maple. Sunflower. Four-o'clock. Morning-Glory. | Acorn. Horsechestnut. Almond. Bean. Pea. Squash. Nasturtium. |
Those with plumules will be seen to have the most abundant nourishment. In many cases this is made use of by man.
These last can be again divided into those in which the cotyledons come up into the air and those where they remain in the ground.
| In the Air. | In the Ground. |
| Bean. Almond. Squash. | Acorn. Horsechestnut. Pea. Nasturtium. |
In the latter the cotyledons are so heavily gorged with nourishment that they never become of any use as leaves. As Darwin points out, they have a better chance of escaping destruction by animals by remaining in the ground.
The cotyledons are very good illustrations of the different uses to which a single organ may be put, and the thorough understanding of it will prepare the pupils' minds for other metamorphoses, and for the theory that all the various parts of a plant are modified forms of a very few members.
4. Nature of the Caulicle.—Probably some of the pupils will have called the caulicle the root. It is, however, of the nature of stem. The root grows only at the end, from a point just behind the tip; the stem elongates throughout its whole length. This can be shown by marking the stem and roots of a young seedling with ink. India ink must be used, as common ink injures the plants. Dip a needle in the ink and prick a row of spots at equal distances on a young root. Corn is very good for this purpose, but Morning-Glory or Bean is better for experiments on the stem. The plants should then be carefully watched and the changes in the relative distance of the spots noted. The experiment is very easily conducted with the seedlings growing on sponge, with their roots in the moist air of the tumbler, as before described.
Dr. Goodale says of this experiment,—"Let a young seedling of corn be grown on damp paper in the manner described in No. 1,[1] and when the longest root is a few centimetres long let it be marked very carefully by means of India ink, or purple ink, put on with a delicate camel's-hair pencil just one centimetre apart. Plants thus marked are to be kept under favorable conditions with respect to moisture and warmth, so that growth will be as rapid as possible. The marks on the older part of the root will not change their relative distance, but the mark at the tip will be carried away from the one next it, showing that the growth has taken place only at this point. Such experiments as the one described are perfectly practicable for all classes of pupils except the very youngest. How far the details of these experiments should be suggested to the pupils, or rather how far they should be left to work out the problem for themselves, is a question to be settled by the teacher in each case. The better plan generally is to bring the problem in a very clear form before the whole class, or before the whole school, and ask whether anybody can think of a way in which it can be solved; for instance, in this case how can it be found out whether roots grow only at their tip or throughout their whole length. If the way is thought out by even a single pupil the rest will be interested in seeing whether the plan will work successfully."
I have been more successful in pricking the roots than in marking them with a brush.
The caulicle can be proved by the manner of its growth to be of the nature of stem, not root. The main root grows from its naked end. Roots can also grow from the sides of the caulicle, as in Indian Corn. In this, it acts precisely as does the stem of a cutting. It can be prettily shown with the seedlings by breaking off a bean at the ground and putting the slip in water. It will throw out roots and the pupil will readily understand that the caulicle does the same thing.
Darwin has made very interesting experiments on the movements of seedlings. If the teacher wishes to repeat some of the experiments he will find the details very fully given in "The Power of Movement of Plants."[1] The pupils can observe in their growing seedlings some of the points mentioned and have already noticed a few in their answers. They have said that the caulicle was the part to grow first, and have spoken of the arched form of the young stem. Their attention should also be drawn to the root-hairs, which are well seen in Corn, Wheat, and Oats. They absorb the liquid food of the plants. A secondary office is to hold the seed firmly, so that the caulicle can enter the ground. This is shown in Red Clover, which may be sown on the surface of the ground. It puts out root-hairs, which attach themselves to the particles of sand and hold the seed. These hairs are treated more fully in the lessons on roots.
5. Leaves of Seedlings.—Coming now to the question as to the number of leaves at each joint of the stem, the Morning-Glory, Sunflower, and Bean will present no difficulty, but probably all the pupils will be puzzled by the Pea. The stipules, so large and leaf-like, look like two leaves, with a stem between, bearing other opposite leaves, and terminating in a tendril, while in the upper part it could not be told by a beginner which was the continuation of the main stem. For these reasons I left this out in the questions on the Pea, but it should be taken up in the class. How are we to tell what constitutes a single leaf? The answer to this question is that buds come in the axils of single leaves; that is, in the inner angle which the leaf makes with the stem. If no bud can be seen in the Pea, the experiment may be tried of cutting off the top of the seedling plant. Buds will be developed in the axils of the nearest leaves, and it will be shown that each is a compound leaf with two appendages at its base, called stipules, and with a tendril at its apex. Buds can be forced in the same way to grow from the axils of the lower scales, and even from those of the cotyledons, and the lesson may be again impressed that organs are capable of undergoing great modifications. The teacher may use his own judgment as to whether he will tell them that the tendril is a modified leaflet.
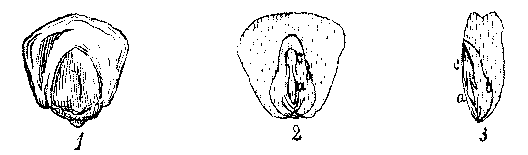
[Illustration: FIG. 9. 1. Grain of Indian Corn. 2. Vertical section, dividing the embryo, a, caulicle: b, cotyledon; c, plumule. 3. Vertical section, at right angles to the last.]
6. Monocotyledons.—These are more difficult. Perhaps it is not worth while to attempt to make the pupils see the embryo in Wheat and Oats. But the embryo of Indian Corn is larger and can be easily examined after long soaking. Removing the seed-covering, we find the greater part of the seed to be albumen. Closely applied to one side of this, so closely that it is difficult to separate it perfectly, is the single cotyledon. This completely surrounds the plumule and furnishes it with food from the albumen. There is a line down the middle, and, if we carefully bend back the edges of the cotyledon, it splits along this line, showing the plumule and caulicle within. The plumule consists of successive layers of rudimentary leaves, the outer enclosing the rest (Fig. 10, 1, c). The latter is the first leaf and remains undeveloped as a scaly sheath (Fig. 10, 2, c). In Wheat and Oats the cotyledon can be easily seen in the largest seedlings by pulling off the dry husk of the grain. The food will he seen to have been used up.
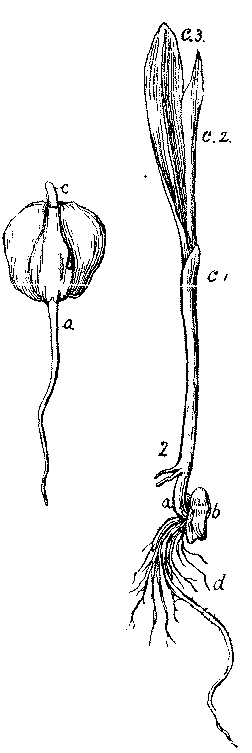
[Illustration: FIG. 10. 1. Germination of Indian corn. 2. Same more advanced. a, caulicle; c1, first leaf of the plumule, sheathing the rest; c2, second leaf; c3, third leaf of the plumule; d, roots.]
The series of Corn seedlings, at least, should be drawn as before and the parts marked, this time with their technical terms. The following questions should then be prepared.
CORN.
What are the parts of the seed?
Compare these parts with the Morning-Glory, Sunflower, Bean, and Pea.
Where is the food stored?
How many cotyledons have Corn, Wheat, and Oats?
How many have Bean, Pea, Morning-Glory, and Sunflower?
Compare the veins of the leaves of each class and see what difference you can find.
This will bring up the terms dicotyledon and monocotyledon. Di means two, mono means one. This difference in the veins, netted in the first class, parallel in the second, is characteristic of the classes. Pupils should have specimens of leaves to classify under these two heads. Flowering plants are divided first into these two classes, the Dicotyledons and the Monocotyledons.
If Pine-seeds can be planted, the polycotyledonous embryo can also be studied.
7. Food of seedlings.—The food of the Wheat seedling may be shown in fine flour. [1]"The flour is to be moistened in the hand and kneaded until it becomes a homogeneous mass. Upon this mass pour some pure water and wash out all the white powder until nothing is left except a viscid lump of gluten. This is the part of the crushed wheat-grains which very closely resembles in its composition the flesh of animals. The white powder washed away is nearly pure wheat-starch. Of course the other ingredients, such as the mineral matter and the like, might be referred to, but the starch at least should be shown. When the seed is placed in proper soil, or upon a support where it can receive moisture, and can get at the air and still be warm enough, a part of the starch changes into a sort of gum, like that on postage stamps, and finally becomes a kind of sugar. Upon this sirup the young seedling feeds until it has some good green leaves for work, and as we have seen in the case of some plants it has these very early."
The presence of starch can be shown by testing with a solution of iodine. Starch is turned blue by iodine and may thus be detected in flour, in seeds, in potatoes, etc.
After all this careful experimental work the subject may be studied in the text-book and recited, the recitation constituting a thorough review of the whole.
A charming description of the germination of a seed will be found in the Reader. V. The Birth of Picciola.
Gray's Lessons. Sect. II, 8-14. III. How Plants Grow. Sect. I, 22, 23. II.
This subject can be treated more conveniently while the young seedlings are still growing, because their roots are very suitable for study. It seems best, therefore, to take it up before examining the buds.
1. Study of the Roots of Seedlings.—One or two of the seedlings should be broken off and the slips put into a glass of water. They will be studied later. Bean and Sunflower are the best for the purpose.
Begin by telling the pupils to prepare for their first lesson a description of the roots of their seedlings. Those grown on sponge or paper will show the development of the root-hairs, while those grown on sand are better for studying the form of the root. Give them also some fleshy root to describe, as a carrot, or a radish; and a spray of English Ivy, as an example of aërial roots.
Throughout these lessons, the method is pursued of giving pupils specimens to observe and describe before teaching them botanical terms. It is better for them to name the things they see than to find examples for terms already learned. In the first case, they feel the difficulty of expressing themselves and are glad to have the want of exact terms supplied. This method is discouraging at first, especially to the younger ones; but, with time and patience, they will gradually become accustomed to describe whatever they can see. They have, at any rate, used their eyes; and, though they may not understand the real meaning of anything they have seen, they are prepared to discuss the subject intelligently when they come together in the class. If they will first write out their unassisted impressions and, subsequently, an account of the same thing after they have had a recitation upon it, they will be sure to gain something in the power of observation and clear expression. It cannot be too strongly urged that the number of facts that the children may learn is not of the slightest consequence, but that the teacher should aim to cultivate the quick eye, the ready hand, and the clear reason.
The root of the Morning-Glory is primary; it is a direct downward growth from the tip of the caulicle. It is about as thick as the stem, tapers towards the end, and has short and fibrous branches. In some plants the root keeps on growing and makes a tap-root; in the Bean, it soon becomes lost in the branches. These are all simple, that is, there is but one primary root. Sometimes there are several or many, and the root is then said to be multiple. The Pumpkin is an example of this. The root of the Pea is described in the older editions of Gray's Lessons as being multiple, but it is generally simple. Indian Corn, also, usually starts with a single root, but this does not make a tap-root, and is soon followed by many others from any part of the caulicle, or even from the stem above, giving it the appearance of having a multiple root.
The root of the Radish is different from any of these; it is fleshy. Often, it tapers suddenly at the bottom into a root like that of the Morning-Glory with some fibres upon it. It is, in fact, as the Morning-Glory would be if the main root were to be thickened up by food being stored in it. It is a primary tap-root. The radish is spindle-shaped, tapering at top and bottom, the carrot is conical, the turnip is called napiform; some radishes are shaped like the turnip.
The aërial roots of the English Ivy answer another purpose than that of giving nourishment to the plant. They are used to support it in climbing. These are an example of secondary roots, which are roots springing laterally from any part of the stem. The Sweet Potato has both fleshy and fibrous roots and forms secondary roots of both kinds every year.[1] Some of the seedlings will probably show the root-hairs to the naked eye. These will be noticed hereafter.
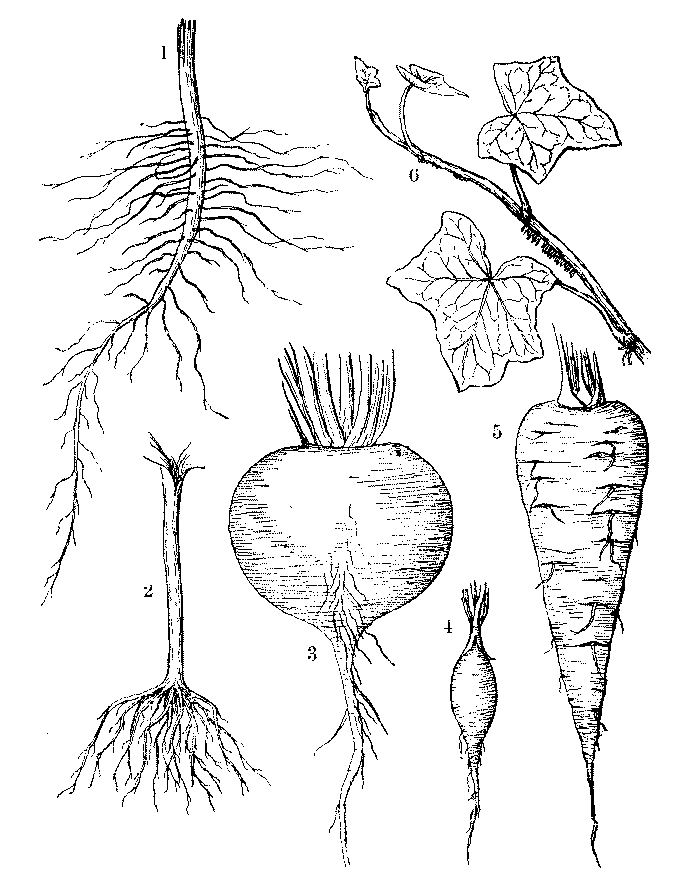
[Illustration: FIG. 11.—1. Tap-root. 2. Multiple root of Pumpkin. 3. Napiform root of Turnip. 4. Spindle-shaped root of Radish. 5. Conical root of Carrot. 6. Aërial roots of Ivy.]
It is my experience that pupils always like classifying things under different heads, and it is a good exercise. The following table may be made of the roots they have studied, adding other examples. Dr. Gray says that ordinary roots may be roughly classed into fibrous and fleshy.[1] Thomé classes them as woody and fleshy.[2]
| ROOTS. | |||
| Primary. | Secondary. | ||
| Fibrous. | Fleshy. | Roots of cuttings | |
| Aërial roots. | |||
| Sweet potatoes.[1] | |||
| Simple. | Multiple. | Simple. | |
| Morning Glory. | Pumpkin | Carrot. | |
| Sunflower. | Radish. | ||
| Pea. | Turnip. | ||
| Bean. | Beet. | ||
| Corn. | Corn. | ||
2. Fleshy Roots.—The scholars are already familiar with the storing of food for the seedling in or around the cotyledons, and will readily understand that these roots are storehouses of food for the plant. The Turnip, Carrot, and Beet are biennials; that is, their growth is continued through two seasons. In the first year, they make a vigorous growth of leaves alone, and the surplus food is carried to the root in the form of a syrup, and there stored, having been changed into starch, or something very similar. At the end of the first season, the root is filled with food, prepared for the next year, so that the plant can live on its reserve fund and devote its whole attention to flowering. These roots are often good food for animals. There are some plants that store their surplus food in their roots year after year, using up in each season the store of the former one, and forming new roots continually. The Sweet Potato is an example of this class. These are perennials. The food in perennials, however, is usually stored in stems, rather than in roots, as in trees. Annuals are generally fibrous-rooted, and the plant dies after its first year. The following experiment will serve as an illustration of the way in which the food stored in fleshy roots is utilized for growth.
Cut off the tapering end of a carrot and scoop out the inside of the larger half in the form of a vase, leaving about half of the flesh behind. Put strings through the upper rim, fill the carrot cup with water, and hang it up in a sunny window. Keep it constantly full of water. The leaf-buds below will put forth, and grow into leafy shoots, which, turning upwards, soon hide the vase in a green circle. This is because the dry, starchy food stored in the carrot becomes soft and soluble, and the supply of proper food and the warmth of the room make the leaf-buds able to grow. It is also a pretty illustration of the way in which stems always grow upward, even though there is enough light and air for them to grow straight downwards. Why this is so, we do not know.
3. Differences between the Stem and the Root.—Ask the pupils to tell what differences they have found.
| Stems. | Roots. |
| Ascend into the air. | Descend into the ground. |
| Grow by a succession of similar parts, each part when young elongating throughout. | Grow only from a point just behind the tip. |
| Bear organs. | Bear no organs. |
There are certain exceptions to the statement that roots descend into the ground; such as aërial roots and parasitic roots. The aërial roots of the Ivy have been mentioned. Other examples of roots used for climbing are the Trumpet Creeper (Tecoma radicans), and the Poison Ivy (Rhus Toxicodendron). Parasitic roots take their food ready-made from the plants into which they strike. The roots of air-plants, such as certain orchids, draw their nourishment from the air.
The experiment of marking roots and stem has been already tried, but it should be repeated. Repetition of experiments is always desirable, as it fixes his conclusions in the pupil's mind. The stem grows by a succession of similar parts, phytomera, each part, or phyton, consisting of node, internode, and leaf. Thus it follows that stems must bear leaves. The marked stems of seedlings show greater growth towards the top of the growing phyton. It is only young stems that elongate throughout. The older parts of a phyton grow little, and when the internode has attained a certain length, variable for different stems and different conditions, it does not elongate at all.
The root, on the contrary, grows only from a point just behind the tip. The extreme tip consists of a sort of cap of hard tissue, called the root-cap. Through a simple lens, or sometimes with the naked eye, it can be distinguished in most of the roots of the seedlings, looking like a transparent tip. "The root, whatever its origin in any case may be, grows in length only in one way; namely, at a point just behind its very tip. This growing point is usually protected by a peculiar cap, which insinuates its way through the crevices of the soil. If roots should grow as stems escaping from the bud-state do,—that is, throughout their whole length—they would speedily become distorted. But, since they grow at the protected tips, they can make their way through the interstices of soil, which from its compactness would otherwise forbid their progress."[1]
The third difference is that, while the stem bears leaves, and has buds normally developed in their axils, roots bear no organs. The stem, however, especially when wounded, may produce buds anywhere from the surface of the bark, and these buds are called adventitious buds. In the same manner, roots occasionally produce buds, which grow up into leafy shoots, as in the Apple and Poplar.[1]
It should be made perfectly clear that the stem is the axis of the plant, that is, it bears all the other organs. Roots grow from stems, not steins from roots, except in certain cases, like that of the Poplar mentioned above. This was seen in the study of the seedling. The embryo consisted of stem and leaves, and the roots were produced from the stem as the seedling grew.
For illustration of this point, the careful watching of the cuttings placed in water will be very instructive. After a few days, small, hard lumps begin to appear under the skin of the stem of the broken seedling Bean. These gradually increase in size until, finally, they rupture the skin and appear as rootlets. Roots are always thus formed under the outer tissues of the stem from which they spring, or the root from which they branch. In the Bean, the roots are in four long rows, quartering the stem. This is because they are formed in front of the woody bundles of the stem, which in the seedling Bean are four. In the Sunflower the roots divide the circumference into six parts. In some of my cuttings of Beans, the stem cracked in four long lines before the roots had really formed, showing the parenchyma in small hillocks, so to speak. In these the gradual formation of the root-cap could be watched throughout, with merely a small lens. I do not know a better way to impress the nature of the root on the pupil's mind. These forming roots might also be marked very early, and so be shown to carry onward their root-cap on the growing-point.
4. Root-hairs. These are outgrowths of the epidermis, or skin of the root, and increase its absorbing power. In most plants they cannot be seen without the aid of a microscope. Indian Corn and Oats, however, show them very beautifully, and the scholars have already noticed them in their seedlings. They are best seen in the seedlings grown on damp sponge. In those grown in sand, they become so firmly united to the particles of soil, that they cannot be separated, without tearing the hairs away from the plant. This will suggest the reason why plants suffer so much from careless transplanting.
The root-hairs have the power of dissolving mineral matters in the soil by the action of an acid which they give out. They then absorb these solutions for the nourishment of the plant. The acid given out was first thought to be carbonic acid, but now it is supposed by some experimenters to be acetic acid, by others to vary according to the plant and the time. The action can be shown by the following experiment, suggested by Sachs.

[Illustration: Fig. 12. I. Seedling of Sinapis alba showing root-hairs. II. Same, showing how fine particles of sand cling to the root-hairs. (Sachs.)]
Cover a piece of polished marble with moist sawdust, and plant some seeds upon it. When the seedlings are somewhat grown, remove the sawdust, and the rootlets will be found to have left their autographs behind. Wherever the roots, with their root-hairs have crept, they have eaten into the marble and left it corroded. The marks will become more distinct if the marble is rubbed with a little vermilion.
In order that the processes of solution and absorption may take place, it is necessary that free oxygen should be present. All living things must have oxygen to breathe, and this gas is as needful for the germination of seeds, and the action of roots and leaves, as it is for our maintenance of life. It is hurtful for plants to be kept with too much water about their roots, because this keeps out the air. This is the reason why house-plants are injured if they are kept too wet.
A secondary office of root-hairs is to aid the roots of seedlings to enter the ground, as we have before noticed.
The root-hairs are found only on the young parts of roots. As a root grows older the root-hairs die, and it becomes of no further use for absorption. But it is needed now for another purpose, as the support of the growing plant. In trees, the old roots grow from year to year like stems, and become large and strong. The extent of the roots corresponds in a general way to that of the branches, and, as the absorbing parts are the young rootlets, the rain that drops from the leafy roof falls just where it is needed by the delicate fibrils in the earth below.[1]
5. Comparison of a Carrot, an Onion, and a Potato.—It is a good exercise for a class to take a potato, an onion, and a carrot or radish to compare, writing out the result of their observations.
The carrot is a fleshy root, as we have already seen. The onion consists of the fleshy bases of last year's leaves, sheathed by the dried remains of the leaves of former years, from which all nourishment has been drawn. The parallel veining of the leaves is distinctly marked. The stem is a plate at the base, to which these fleshy scales are attached. In the centre, or in the axils of the scales, the newly-forming bulbs can be seen, in onions that are sprouting. If possible, compare other bulbs, as those of Tulip, Hyacinth, or Snowdrop, and the bulb of a Crocus, in which the fleshy part consists of the thickened base of the stem, and the leaves are merely dry scales. This is called a corm.
The potato is a thickened stem. It shows itself to be a stem, because it bears organs. The leaves are reduced to little scales (eyelids), in the axils of which come the buds (eyes). The following delightful experiment has been recommended to me.
In a growing potato plant, direct upwards one of the low shoots and surround it with a little cylinder of stiff carpet paper, stuffed with sphagnum and loam. Cut away the other tuber-disposed shoots as they appear. The enclosed shoot develops into a tuber which stands more or less vertical, and the scales become pretty little leaves. Removing the paper, the tuber and leaves become green, and the latter enlarge a little. A better illustration of the way in which organs adapt themselves to their conditions, and of the meaning of morphology, could hardly be found.
Gray's First Lessons. Sect. v, 65-88. How Plants Grow. Chap. I, 83-90.
1. There is an astonishing amount to be learned from naked branches, and, if pursued in the right way, the study will be found exceedingly interesting. Professor Beal, in his pamphlet on the New Botany,[1] says:—
"Before the first lesson, each pupil is furnished or told where to procure some specimen for study. If it is winter, and flowers or growing plants cannot be had, give each a branch of a tree or shrub; this branch may be two feet long. The examination of these is made during the usual time for preparing lessons, and not while the class is before the teacher. For the first recitation each is to tell what he has discovered. The specimens are not in sight during the recitation. In learning the lesson, books are not used; for, if they are used, no books will contain a quarter of what the pupil may see for himself. If there is time, each member of the class is allowed a chance to mention anything not named by any of the rest. The teacher may suggest a few other points for study. The pupils are not told what they can see for themselves. An effort is made to keep them working after something which they have not yet discovered. If two members disagree on any point, on the next day, after further study, they are requested to bring in all the proofs they can to sustain their different conclusions. For a second lesson, the students review the first lesson, and report on a branch of a tree of another species which they have studied as before. Now they notice any point of difference or of similarity. In like manner new branches are studied and new comparisons made. For this purpose, naked branches of our species of elms, maples, ashes, oaks, basswood, beech, poplars, willows, walnut, butternut, hawthorns, cherries, and in fact any of our native or exotic trees or shrubs are suitable. A comparison of the branches of any of the evergreens is interesting and profitable. Discoveries, very unexpected, are almost sure to reward a patient study of these objects. The teacher must not think time is wasted. No real progress can be made, till the pupils begin to learn to see; and to learn to see they must keep trying to form the habit from the very first; and to form the habit they should make the study of specimens the main feature in the course of training."
HORSECHESTNUT (Æsculus Hippocastanum).
We will begin with the study of a branch of Horsechestnut.[1] The pupils should examine and describe their specimens before discussing them in the class-room. They will need some directions and hints, however, to enable them to work to any advantage. Tell them to open both large and small buds. It is not advisable to study the Horsechestnut bud by cutting sections, as the wool is so dense that the arrangement cannot be seen in this way. The scales should be removed with a knife, one by one, and the number, texture, etc., noted. The leaves and flower-cluster will remain uncovered and will be easy to examine. The gum may be first removed by pressing the bud in a bit of paper. The scholars should study carefully the markings on the stem, in order to explain, if possible, what has caused them. The best way to make clear the meaning of the scars is to show them the relation of the bud to the branch. They must define a bud. Ask them what the bud would have become the next season, if it had been allowed to develop. It would have been a branch, or a part of one. A bud, then, is an undeveloped branch. They can always work out this definition for themselves. Conversely, a branch is a developed bud, or series of buds, and every mark on the branch must correspond to something in the bud. Let them examine the specimens with this idea clearly before their minds. The lesson to prepare should be to write out all they can observe and to make careful drawings of their specimens. Ask them to find a way, if possible, to tell the age of the branch.
At the recitation, the papers can be read and the points mentioned thoroughly discussed. This will take two lesson-hours, probably, and the drawing may be left, if desired, as the exercise to prepare for the second recitation.
[1]The buds of Horsechestnut contain the plan of the whole growth of the next season. They are scaly and covered, especially towards the apex, with a sticky varnish. The scales are opposite, like the leaves. The outer pairs are wholly brown and leathery, the succeeding ones tipped with brown, wherever exposed, so that the whole bud is covered with a thick coat. The inner scales are green and delicate, and somewhat woolly, especially along the lapping edges. There are about seven pairs of scales. The larger terminal buds have a flower-cluster in the centre, and generally two pairs of leaves; the small buds contain leaves alone, two or three pairs of them. The leaves are densely covered with white wool, to protect them from the sudden changes of winter. The use of the gum is to ward off moisture. The flower-cluster is woolly also.
The scars on the stem are of three kinds, leaf, bud-scale, and flower-cluster scars. The pupils should notice that the buds are always just above the large triangular scars. If they are still in doubt as to the cause of these marks, show them some house-plant with well-developed buds in the axils of the leaves, and ask them to compare the position of these buds with their branches. The buds that spring from the inner angle of the leaf with the stem are axillary buds; those that crown the stems are terminal. Since a bud is an undeveloped branch, terminal buds carry, on the axis which they crown, axillary buds give rise to side-shoots. The leaf-scars show the leaf-arrangement and the number of leaves each year. The leaves are opposite and each pair stands over the intervals of the pair below. The same is observed to be true of the scales and leaves of the bud.[1] All these points should be brought out by the actual observation of the specimens by the pupils, with only such hints from the teacher as may be needed to direct their attention aright. The dots on the leaf-scar are the ends of woody bundles (fibro-vascular bundles) which, in autumn, separated from the leaf. By counting these we can tell how many leaflets there were in the leaf, three, five, seven, nine, or occasionally six or eight.
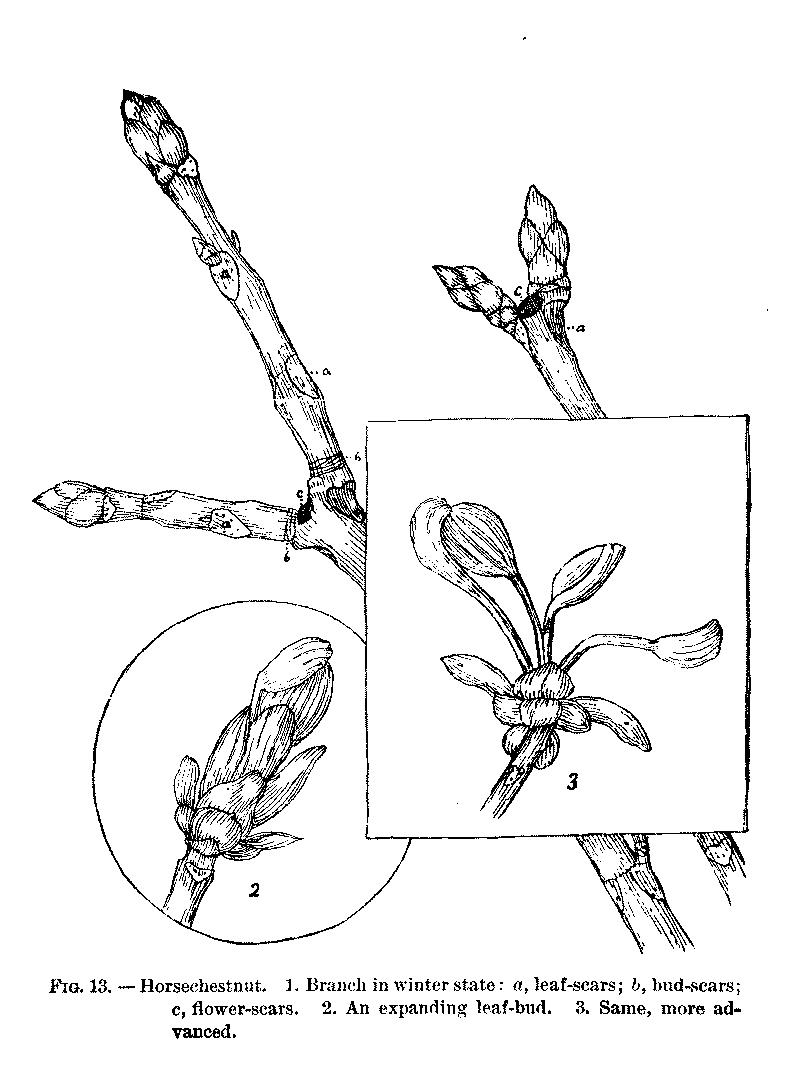
[Illustration: FIG. 13.—Horsechestnut. I. Branch in winter state: a, leaf-scars; b, bud-scars; c, flower-scars. 2. An expanding leaf-bud. 3. Same, more advanced.]
The Bud Scale-Scars. These are rings left by the scales of the bud and may be seen in many branches. They are well seen in Horsechestnut. If the pupils have failed to observe that these rings show the position of former buds and mark the growth of successive years, this point must be brought out by skilful questioning. There is a difference in the color of the more recent shoots, and a pupil, when asked how much of his branch grew the preceding season, will be able to answer by observing the change in color. Make him see that this change corresponds with the rings, and he will understand how to tell every year's growth. Then ask what would make the rings in a branch produced from one of his buds, and he can hardly fail to see that the scales would make them. When the scholars understand that the rings mark the year's growth, they can count them and ascertain the age of each branch. The same should be done with each side-shoot. Usually the numbers will be found to agree; that is, all the buds will have the same number of rings between them and the cut end of the branch, but occasionally a bud will remain latent for one or several seasons and then begin its growth, in which case the numbers will not agree; the difference will be the number of years it remained latent. There are always many buds that are not developed. "The undeveloped buds do not necessarily perish, but are ready to be called into action in case the others are checked. When the stronger buds are destroyed, some that would else remain dormant develop in their stead, incited by the abundance of nourishment which the former would have monopolized. In this manner our trees are soon reclothed with verdure, after their tender foliage and branches have been killed by a late vernal frost, or consumed by insects. And buds which have remained latent for several years occasionally shoot forth into branches from the sides of old stems, especially in certain trees."[1]
The pupils can measure the distance between each set of rings on the main stem, to see on what years it grew best.
The Flower-Cluster Scars. These are the round, somewhat concave, scars, found terminating the stem where forking occurs, or seemingly in the axils of branches, on account of one of the forking branches growing more rapidly and stoutly than the other and thus taking the place of the main stem, so that this is apparently continued without interruption. If the pupils have not understood the cause of the flower-cluster scars, show them their position in shoots where they are plainly on the summit of the stem, and tell them to compare this with the arrangement of a large bud. The flower-cluster terminates the axis in the bud, and this scar terminates a branch. When the terminal bud is thus prevented from continuing its growth, the nearest axillary buds are developed.[1] One shoot usually gets the start, and becomes so much stronger that it throws the other to one side. The tendency of the Horsechestnut to have its growth carried on by the terminal buds is so strong that I almost feel inclined to say that vigorous branches are never formed from axillary buds, in old trees, except where the terminal bud has been prevented from continuing the branch. This tendency gives to the tree its characteristic size of trunk and branches, and lack of delicate spray. On looking closely at the branches also, they will be seen to be quite irregular, wherever there has been a flower-cluster swerving to one side or the other.
There is one thing more the pupils may have noticed. The small round dots all over the young stem, which become long rifts in the older parts, are breaks in the epidermis, or skin of the stem, through which the inner layers of bark protrude. They are called lenticels. They provide a passage for gases in and out of the stem. In some trees, as the Birch, they are very noticeable.
After discussing the subject thoroughly in the class-room, the pupils should rewrite their papers, and finally answer the following questions, as a species of review. I have thus spent three recitations on the Horsechestnut. The work is all so new, and, if properly presented, so interesting, that a good deal of time is required to exhaust its possibilities of instruction. If the teacher finds his scholars wearying, however, he can leave as many of the details as he pleases to be treated in connection with other branches.
QUESTIONS ON THE HORSECHESTNUT.
How many scales are there in the buds you have examined?
How are they arranged?
How many leaves are there in the buds?
How are they arranged?
Where does the flower-cluster come in the bud?
Do all the buds contain flower-clusters?
What is the use of the wool and the gum?
Where do the buds come on the stem?
Which are the strongest?
How are the leaves arranged on the stem?
Do the pairs stand directly over each other?
What are the dots on the leaf-scars?
How old is your branch?
How old is each twig?
Which years were the best for growth?
Where were the former flower-clusters?
What happens when a branch is stopped in its growth by flowering?
What effect does this have on the appearance of the tree?
In some parts of the country the Horsechestnut is not so commonly planted as in New England. In the southern states the Magnolia may be used in its stead, but it is not nearly so simple an example of the main points to be observed.[1]
MAGNOLIA UMBRELLA.
The bud may be examined by removing the scales with a knife, as in Horsechestnut, and also by cutting sections. The outer scales enfold the whole bud, and each succeeding pair cover all within. They are joined, and it is frequently difficult to tell where the suture is, though it can generally be traced at the apex of the bud. On the back is a thick stalk, which is the base of the leaf-stalk. Remove the scales by cutting carefully through a single pair, opposite the leaf-stalk, and peeling them off. The scales are modified stipules, instead of leaf-stalks, as in Horsechestnut. The outer pair are brown and thick, the inner green, and becoming more delicate and crumpled as we proceed toward the centre of the bud. The leaves begin with the second or third pair of scales. The first one or two are imperfect, being small, brown, and dry. The leaves grow larger towards the centre of the bud. They are covered with short, silky hairs, and are folded lengthwise, with the inner surface within (conduplicate). In the specimens I have examined I do not see much difference in size between the buds with flowers and those without. In every bud examined which contained a flower, there was an axillary bud in the axil of the last, or next to the last, leaf. This bud is to continue the interrupted branch in the same way as in Horsechestnut.
There are from six to ten good leaves, in the buds that I have seen. Those without flowers contain more leaves, as in Horsechestnut. In the centre of these buds the leaves are small and undeveloped. The flower is very easy to examine, the floral envelopes, stamens and pistils, being plainly discernible. The bud may also be studied in cross-section. This shows the whole arrangement. The plan is not so simple as in Horsechestnut, where the leaves are opposite. The subject of leaf-arrangement should be passed over until phyllotaxy is taken up.
The scars on the stem differ from Horsechestnut in having no distinct bands of rings. The scales, being stipules, leave a line on each side of the leaf-scar, and these are separated by the growth of the internodes. In the Beech, the scales are also stipules; but, whereas in the Magnolia there are only one or two abortive leaves, in the Beech there are eight or nine pairs of stipules without any leaves at all. The rings thus become separated in Magnolia, while in the Beech the first internodes are not developed, leaving a distinct band of rings, to mark the season's growth. The Magnolia is therefore less desirable to begin upon. The branches are swollen at the beginning of a new growth, and have a number of leaf-scars crowded closely together. The leaf-scars are roundish, the lower line more curved. They have many dots on them. From each leaf-scar runs an irregular line around the stem. This has been left by the stipules.
The flower-scar is on the summit of the axis, and often apparently in the axil of a branch, as in Horsechestnut. Sometimes the nearest axillary bud is developed; sometimes there are two, when the branch forks. The axillary buds seldom grow unless the terminal bud is interrupted. The tree therefore has no fine spray.
LILAC (Syringa vulgaris).
Ask the scholars to write a description of their branches and to compare them with Horsechestnut. These papers should be prepared before coming into the class, as before.
The buds are four-sided. The scales and leaves are opposite, as in Horsechestnut. The outer pair sometimes have buds in their axils. Remove the scales one by one with a knife, or better, with a stout needle. The scales gradually become thinner as we proceed, and pass into leaves, so that we cannot tell where the scales end and leaves begin. After about six pairs are removed, we come, in the larger buds, to leaves with axillary flower-clusters. The leaves grow smaller and the flower-clusters larger till we come to the centre, where the axis is terminated by a flower-cluster. There is a great difference in the buds on different bushes and on shoots of the same bush, some being large, green, and easy to examine, others small, hard, and dark-colored. It is better, of course, to select as soft and large buds as possible for examination.
[Illustration: FIG. 14.—Lilac. I. Branch in winter state: a, leaf-scar; b, bud-scar (reduced). 2. Same, less reduced. 3. Branch, with leaf-buds expanded. 4. Series in a single bud, showing the gradual transition from scales to leaves.]
That the scales are modified leaves is plainly shown by the gradual transition they undergo, and also by the fact that buds are developed in their axils. If any of these can be shown to the pupils, remind them of the experiment where the top of a seedling Pea was cut off and buds forced to develop in the axils of the lower scales.[1] The transition from scales to leaves can be well studied by bringing branches into the house, where they will develop in water, and towards spring may even be made to blossom. Cherry, Apple, Forsythia, and other blossoming trees and shrubs can be thus forced to bloom. Place the branches in hot water, and cut off a little of their ends under water. If the water is changed every day, and the glass kept near the register or stove, they will blossom out very quickly. These expanded shoots may be compared with the buds. The number of leaves in the bud varies.
The leaf-scars of Lilac are horseshoe-shaped and somewhat swollen. It can often be plainly seen that the outer tissue of the stem runs up into the scar. It looks as if there were a layer of bark, ending with the scar, fastened over each side of the stem. These apparent layers alternate as well as the scars. The epidermis, or skin of the leaves, is in fact always continuous with that of the stem. There are no dots on the leaf-scars.
The rings are not nearly so noticeable as in Horsechestnut, but they can be counted for some years back.
The flower-cluster can often be traced by a dried bit of stem remaining on the branch.
The terminal bud in the Lilac does not usually develop, and the two uppermost axillary buds take its place, giving to the shrub the forked character of its branching. In all these bud studies, the pupil should finish by showing how the arrangement of the buds determines the growth of the branches.
QUESTIONS ON THE LILAC.
How do the scales differ from those of Horsechestnut?
How many scales and leaves are there?
How are they arranged?
Where does the flower-cluster come in the bud?
Do all the buds contain flower-clusters?
How does the arrangement of leaves and flower-clusters differ from that of Horsechestnut?
How old is your branch?
Which buds develop most frequently?
How does this affect the appearance of the shrub?
COPPER BEECH (Fagus sylvatica, var. purpurea).
The buds are long and tapering, the scales thin and scarious, the outer naked, the inner with long, silky hairs. Remove the scales one by one, as in Lilac. The outer four or six pairs are so minute that the arrangement is not very clear, but as we proceed we perceive that the scales are in alternate pairs, as in Horsechestnut; that is, that two scales are exactly on the same plane. But we have learned in the Lilac that the scales are modified leaves, and follow the leaf-arrangement of the species. The Beech is alternate-leaved, and we should therefore expect the scales to alternate. The explanation is found as we go on removing the scales. At the eighth or ninth pair we come upon a tiny, silky leaf, directly between the pair of scales, and, removing these, another larger leaf, opposite the first but higher up on the rudimentary stem, and so on, with the rest of the bud. There are five or more leaves, each placed between a pair of scales. Our knowledge of the parts of a leaf shows us at once that the scales must be modified stipules, and that therefore they must be in pairs.[1] Other examples of scales homologous with stipules are the American Elm, Tulip-tree, Poplar and Magnolia. The leaves are plaited on the veins and covered with long, silky hairs. The venation is very distinct. The outer leaves are smaller and, on examining the branch, it will be seen that their internodes do not make so large a growth as the leaves in the centre of the bud.

[Illustration: FIG. 15.—Copper Beech. 1. Branch in winter state: a, leaf-scar; b, bud-scar. 2. Branch, with leaf-buds expanding, showing the plicate folding of the leaves.]
The leaf-scars are small, soon becoming merely ridges running half round the stem.
The bud-rings are very plain and easily counted. For this reason, and because it branches freely, it is a good tree for measurements of growth, as is seen in the following tables. Nos. 1, 2, 3 and 4: were made by a class of girls, from fourteen to sixteen, from a tree on my lawn. No. 5 was made by a pupil, whom I taught by correspondence, from a tree of the same species in another town. No. 6 was made by myself from my own tree. The measurements of the first four tables were somewhat revised by me, as they were not perfectly accurate. The pupils should always be cautioned to measure from the beginning of one set of rings to the beginning of the next.[1]
NO. 1.
| YEARS. | GROWTH OF. MAIN AXIS. (in.) | 1ST BRANCH. | 2nd BRANCH. | 3RD BRANCH. | 4TH BRANCH. |
| '79 | 8-1/2 | — | — | — | — |
| '80 | 4-1/2 | 2 | 1-7/8 | — | — |
| '81 | 3-1/2 | 1-1/8 | 2-5/8 | — | — |
| '82 | 6 | 5/8 | 4-1/4 | 5-7/8 | — |
| '83 | 7-3/8 | 3-3/8 | 5-1/4 | 4 | 5-3/4 |
| '84 | 2 | 1/2 | 3/4 | 3/8 | 5-3/8 |
| '85 | 5/8 | 1/4 | 3/8 | 1/2 | 1 |
| '86 | 5-5/8 | 7/8 | 4-3/8 | 3-1/8 | 5 |
NO. 2.
| YEARS. | GROWTH of MAIN AXIS. (in.) | 1ST BRANCH | 2nd BRANCH | 3RD BRANCH | 4TH BRANCH | 5TH BRANCH | — | — |
| '79 | 8 | — | — | — | — | — | — | — |
| '80 | 3-1/2 | 5-1/4 | 5-1/2 | 5-5/8 | — | — | — | — |
| '81 | 4-3/4 | 3/4 | 1/2 | 2-1/2 | 2 | — | — | — |
| '82 | 5-3/4 | 7/8 | 2 | 3/4 | 3/8 | 1/2 | — | — |
| '83 | 5-1/4 | 4-3/4 | 5-1/2 | 4 | 3-1/4 | 2-3/8 | 1-3/4 | — |
| '84 | 1/2 | 1 | 3/4 | 3/8 | 1 | 3/4 | 1 | 3/8 |
| '85 | 2-3/4 | 1-3/4 | 4-3/8 | 3/4 | 3/4 | 2-1/8 | 3-1/4 | 1-1/4 |
| '86 | 7-1/2 | 5-1/2 | 6-3/4 | 3 | 3 | 4-1/2 | 3-1/8 | 5 |
NO. 3.
| YEARS. | GROWTH of MAIN AXIS. (in.) | 1ST BRANCH | 2ND BRANCH | 3RD BRANCH | 4TH BRANCH | 5TH BRANCH |
| '80 | 8-1/4 | — | — | — | — | — |
| '81 | 4-1/2 | 3-1/2 | 3-3/4 | — | — | — |
| '82 | 5-1/2 | 3/4 | 1-1/2 | 1 | — | — |
| '83 | 3-1/4 | 3-3/4 | 4-1/2 | 3/4 | 2 | 1-1/4 |
| '84 | 5-1/2 | 1/2 | 3/4 | 1 | 1/2 | 3 |
| '85 | 1/2 | 1-3/4 | 1/2 | 3/8 | 1 | 1/2 |
| '86 | 4-1/4 | 3-3/8 | 2-3/8 | 1-1/4 | 2-1/4 | 1-1/2 |
NO. 4.
| YEARS | GROWTH of MAIN AXIS (in.) | 1ST BRANCH | 2nd BRANCH | 3RD BRANCH | 4TH BRANCH |
| '81 | 7-3/4 | — | — | — | — |
| '82 | 8-3/4 | 6 | 6 | — | — |
| '83 | 6-3/4 | 5-1/4 | 4 | 4-3/4 | 5-1/2 |
| '84 | 4-1/2 | 5/8 | 1-5/8 | 2-1/4 | 3-1/4 |
| '85 | 2 | 5/8 | 3/16 | 2 | 3/4 |
| '86 | 10-3/4 | 1-3/4 | 1/4 | 7-1/4 | 3-1/2 |
NO. 4. (cont.)
| YEARS | 5TH BRANCH | 6TH BRANCH | 7TH BRANCH | 8TH BRANCH | 9TH BRANCH |
| '81 | — | — | — | — | — |
| '82 | — | — | — | — | — |
| '83 | — | — | — | — | — |
| '84 | 3/4 | 2-1/2 | — | — | — |
| '85 | 7/8 | 5/8 | 1/4 | 3/4 | — |
| '86 | 4-3/4 | 6-3/8 | 1 | 2-1/4 | 6-1/2 |
NO. 5.
| YEARS | GROWTH of MAIN AXIS (in.) | 1ST BRANCH | 2nd BRANCH | 3RD BRANCH | 4TH BRANCH | 5TH BRANCH | 6TH BRANCH |
| '82 | 6-7/8 | — | — | — | — | — | — |
| '83 | 6-1/2 | 4-3/4 | 4-1/4 | — | — | — | — |
| '84 | 4-3/4 | 1/4 | 1-3/4 | 3-1/2 | — | — | — |
| '85 | 4-1/2 | 3/4 | 1 | 2-3/4 | 2-3/4 | — | — |
| '86 | 6-1/4 | 2-1/4 | 4-3/4 | 6-3/4 | 2-3/4 | 5-3/4 | — |
| '87 | 6-3/4 | 1-1/8 | 3-1/4 | 4 | 2-1/4 | 3 | 5-1/2 |
NO. 6.
| YEARS | MAIN AXIS (in.) | 1ST BRANCH | 2ND BRANCH | 3RD BRANCH | 4TH BRANCH | ||
| 1st side shoot. | 2nd side shoot. | ||||||
| '80 | 6-1/4 | — | — | — | — | — | — |
| '81 | 8-3/4 | 6-3/4 | — | — | — | — | — |
| '82 | 8-1/2 | 6-1/4 | 6-7/8 | — | — | — | . |
| '83 | 4-3/4 | 1-1/2 | 2-3/8 | — | — | 4 | . |
| '84 | 3-1/2 | 3-1/8 | 5-1/8 | — | — | 1-3/4 | 7/8 |
| '85 | 4-1/2 | 3/8 | 4-3/4 | 2-1/4 | — | 6 | 1 |
| '86 | 6+ | 6-3/4 | 12-1/8 | 5-1/2 | 10-1/2 | 8-7/8 | 5-1/8 |
| '87 | bough broken. | 2-1/2 | 8-3/4 | 4-1/4 | 4-1/4 | 4-6/8 | 3-3/4 |
One question brought up by these measurements is whether there is any correspondence in growth between the main axis and its branches. It appears in these tables that there is a general correspondence, in this tree at least. In the recitation of the class, whose tables are given above (Nos. 1, 2, 3 and 4), we took all the measurements of these four branches for the year 1885 and added them. We did the same for 1886, and compared the results. The total growth for 1885 was 31-15/16 inches; for 1886, leaving out the measurement of the twig whose entire growth was in that year, 109-3/4 inches or nearly 3-1/2 times as much. The proportion held in a general way throughout, there being only a single case of a branch where the growth was greater in the first year.[1] But there is a point that must not be overlooked in this connection. The branches of the Beech seem to grow about equally well in the first, second, third, or any succeeding year. In some trees, as the Ash, the axillary buds make a large growth, and the succeeding terminal buds carry on the branch much more slowly; in other trees, as the Cherry, a branch grows very slowly in the first few years and then suddenly takes a start. These facts would appear in tables of growth, made from branches of these trees, but the addition of results for any particular year would have no significance.
In table No. 5, the addition of the measurements for 1885 and 1886 shows the growth in the latter year to be about twice that of the former. This branch came from a tree in another town. We have tried also to discover whether the number of leaves each year has any relation to growth. I cannot see that it has, but it requires many experiments to determine these points. To study this, make tables of the number of leaves on the branch each year. I think teachers would find it interesting to keep all data of this kind of work done by their classes, with a view to tabulation and comparison. The scholars themselves are exceedingly interested in anything that partakes of the nature of an original investigation.[1]
The leaf-arrangement of the Beech is alternate, on the one-half plan. The small twigs turn upwards, so that all the spray is on the upper side, giving a flat appearance to the branch.[1] This gives the leaves a better exposure to the light. Both the terminal and axillary buds grow freely, thus forming long, straight limbs, with many branches and much fine spray.
The bark of the Beech is beautifully smooth. The extreme straightness of the trunk and limbs is very striking, and may be compared to the crooked limbs of the Horsechestnut, where the branch is continually interrupted by the flower-cluster. In the Beech the flowers are axillary.
QUESTIONS ON THE BEECH.
How are the scales of the Beech bud arranged?
How many leaves are there in the bud?
How does the arrangement of the scales and leaves in the bud differ from that of the Horsechestnut?
How are the leaves folded in the bud?
What is the arrangement of the leaves on the stem?
How does this differ from Horsechestnut and Lilac?
How old is your branch?
How old is each twig?
What years were the best for growth?
How does the growth of the branches differ from that of Horsechestnut? From Lilac?
Explain these differences with reference to the growth and arrangement of the buds?
In what direction do the twigs grow?
How does this affect the appearance of the tree?
Compare the amount of spray of the Beech and Horsechestnut and explain the reason of the difference.
These questions are only intended for review, they are never to be used for the first study of the specimen.
AMERICAN ELM (Ulmus Americana).
The buds are covered with brown scales, which are hairy on the edges. The flower-buds are larger than the leaf-buds and are in the axils of the lower leaves of the preceding year. Each leaf in the bud is enclosed by a pair of scales. They are so small that the pupils, unused to delicate work, will hardly discover them. Under a glass they can be seen to be ovate, folded on the midrib with the inner face within (conduplicate), and with an ovate scale joined to the base of the leaf on either side. The scales thus show themselves to be modified stipules. The venation of the leaves is very plain. The scales are much larger than the leaves. The flower-buds contain a cluster of flowers, on slender green pedicels. The calyx is bell-shaped, unequal, and lobed. The stamens and pistil can be seen. The flower-clusters do not seem to leave any mark which is distinguishable from the leaf-scar.
[Illustration: FIG. 16.—American Elm. 1. Branch in winter state: a, leaf-scars; b, bud-scars; d, leaf-buds; e, flower-buds. 2. Branch, with staminate flower-buds expanding. 3. Same, more advanced. 4. Branch, with pistillate flowers, the leaf-bud also expanding. ]
The leaf-scars are small and extend about half around the stem. The arrangement is alternate on the one-half plan. There are three dots on the scar.
The rings are quite plain. The tree can be used to make tables of growth, like those of the Beech.
The buds will probably be too small for examination by the pupils, at present, but their position and development can be studied, and are very instructive. As the leaf-buds are all on the ends of the branchlets, the twigs and branches will be just below the bud-rings, and then there will be a space where no twigs nor branches will be found, till the next set of rings is reached. This gives the branches more room to develop symmetrically. The terminal buds do not develop in the Elm, in old trees, the bud axillary to the last leaf of the season taking its place, and most of the other axillary buds growing also. This makes the tree break out into very fine spray. A tree like the Elm, where the trunk becomes lost in the branches, is called deliquescent; when the trunk is continued to the top of the tree, as in the Spruce, it is excurrent.
The small, feathery twigs and branches that are often seen on the trunks and great limbs of the elm grow from buds which are produced anywhere on the surface of the wood. Such buds are called adventitious buds. They often spring from a tree when it is wounded.
"The American elm is, in most parts of the state, the most magnificent tree to be seen. From a root, which, in old trees, spreads much above the surface of the ground, the trunk rises to a considerable height in a single stem. Here it usually divides into two or three principal branches, which go off by a gradual and easy curve. Theses stretch upwards and outwards with an airy sweep, become horizontal, the extreme half of the limb, pendent, forming a light and regular arch. This graceful curvature, and absence of all abruptness, in the primary limbs and forks, and all the subsequent divisions, are entirely characteristic of the tree, and enable an observer to distinguish it in the winter and even by night, when standing in relief against the sky, as far as it can be distinctly seen."[1]
QUESTIONS ON THE AMERICAN ELM.
How do the flower-buds differ from the leaf-buds in position and appearance?
What is the arrangement of the leaves?
What other tree that you have studied has this arrangement?
How old is your branch?
Where would you look to see if the flower-cluster had left any mark?
Why is it that several twigs grow near each other, and that then comes a space without any branches?
What buds develop most frequently?
How does this affect the appearance of the tree?
What is a tree called when the trunk is lost in the branches?
BALM OF GILEAD (Populus balsamifera, var. candicans).
The buds are pointed: the terminal slightly angled, the axillary flattened against the stem.[1] Some of the axillary buds contain leaves and some flowers; the appearance of the leaf-buds and flower-buds being the same. The scales of the bud are modified stipules. The terminal buds have about three pairs of the outer scales brown and leathery. The inner scales, as well as the leaves, are coated with resinous matter, which has a strong odor and a nauseous taste. The smaller outer scales have no corresponding leaf, and apparently are modified stipules of the leaves of the preceding year, but the larger ones have a leaf to each pair of scales. The outer and inner leaves are small, the middle ones larger. Comparing the branch, it will be seen that these leaves make the largest growth of internode. The leaves are rolled towards the midrib on the upper face (involute). There are about ten which are easily seen and counted, the inner ones being very small, with minute scales. The axillary buds have a short thick scale on the outer part of the bud, then about three pairs of large scales, each succeeding one enwrapping those within, the outer one brown and leathery. The scales of the flower-buds are somewhat gummy, but not nearly so much so as those of the leaf-buds. Within is the catkin. Each pistil, or stamen (they are on separate trees, dioecious) is in a little cup and covered by a scale, which is cut and fringed.
The leaf-scars are somewhat three-lobed on the young parts, with three dots, indicating the fibro-vascular bundles, which ran up into the leaf. The scars are swollen, making the young branches exceedingly rough. In the older parts the scars become less noticeable. Strong young shoots, especially those which come up from the root, are strongly angled, with three ridges running up into each leaf-scar, making them almost club-shaped. There are often from twenty to thirty leaves in one year's growth, in such shoots, and all the leaves are not rudimentary in the bud. The growth in this case is said to be indefinite. Usually in trees with scaly buds the plan of the whole year's growth is laid down in the bud, and the term definite is applied. Branches, like the Rose, that go on growing all summer grow indefinitely.
The bud-scale scar is quite different from the other trees which we have examined. It is not composed of definite rings, but of leaf-scars with long ridges running from each side of them, showing the scales to be modified stipules. The leaf-scars have become somewhat separated by the growth of the internodes. In the Beech, there are eight, or more, pairs of scales with no leaves, so that the internodes do not develop, and a ring is left on the branch.
The flower-cluster leaves a concave, semicircular scar, in the leaf-axil.

[Illustration: FIG. 17.—Balm-of-Gilead. 1. Branch in winter state: a, leaf-scar; b, bud-scar. 2. Branch, with leaf-buds expanded. 3. Branch, with catkin appearing from the bud.]
The terminal buds are the strongest and not very many axillary buds develop, so that the tree has not fine spray.
The leaf-arrangement is alternate, on the 2/5 plan. Phyllotaxy is not yet to be taken up, but the pupils should be shown the different angles of the branching of the twigs, and told to compare them with Beech and Elm.
QUESTIONS ON THE BALM OF GILEAD.
In which buds are the flower-clusters?
Are there flowers and leaves in the same buds?
What are the scales of the bud?
How are the leaves folded in the bud?
How do the axillary and terminal buds differ?
What are the dots on the leaf-scars?
Why is there no distinct band of rings as in Beech?
How old is your branch?
Where do you look for flower-cluster scars?
Which buds are the strongest?
How does this affect the appearance of the tree?
What makes the ends of the branches so rough?
Compare the arrangement of the twigs and branches with Beech and Elm, with Horsechestnut and Lilac.
TULIP-TREE (Liriodendron Tulipifera).
The buds are small, flat, and rounded at the apex. They are sheathed by scales, each leaf being covered by a pair, whose edges cohere. The outer pair are brown and are the stipules of the last leaf of the preceding year. The leaves are conduplicate, as in Magnolia, and have the blade bent inwards on the petiole (inflexed). Their shape is very clearly to be seen, and no bud is more interesting in the closeness of its packing. Axillary buds are often found within. The flowers grow high upon the trees and towards the ends of the branches.
The leaf-scars are round with many dots. The scar of the stipules is a continuous line around the stem, as in Magnolia.
CHERRY (Prunus Cerasus).
The leaf-buds are terminal, or in the axils of the upper leaves of the preceding year; the flower buds are axillary. There is but one bud in each axil, and usually two or three flowers in each bud, but the leaves on the twigs are crowded and the flowers therefore appear in clusters. The blossom-buds are larger and more rounded than the leaf-buds.
The buds of the tree develop very easily in the house, and as they are so small they can be better studied in watching them come out, than by attempting to dissect them, unless the scholars are sufficiently advanced to use the microscope easily. It is always bad for a pupil to attempt to describe what he sees but imperfectly. He will be sure to jump at any conclusions which he thinks ought to be correct.
The leaf-scars are semicircular, small and swollen.
The bud-rings are plain. The twigs make a very small growth in a season, so that the leaf-scars and rings make them exceedingly rough.
The flower-cluster scars are small circles, with a dot in the centre, in the leaf-axils. The flowers come before the leaves.
The leaf-arrangement is alternate on the 2/5 plan. The pupils may compare the branching with that of their other specimens.
RED MAPLE (Acer rubrum).
This is a good specimen for the study of accessory buds. There is usually a bud in the axil of each lower scale of the axillary buds, making three side by side. We have already noticed this as occurring sometimes in Lilac. It is habitually the case with the Red Maple. The middle bud, which is smaller and develops later, is a leaf-bud. The others are flower-buds.
The leaf-scars are small, with three dots on each scar. The rings are very plain. The flower-cluster leaves a round scar in the leaf-axil, as in Cherry.
The leaves are opposite and the tree branches freely. The twigs seem to be found just below the bud-rings, as the upper leaf-buds usually develop best and the lower buds are single, containing flowers only.
NORWAY SPRUCE (Picea excelsa).
The buds are terminal, and axillary, from the axils of the leaves of the preceding year, usually from those at the ends of the branchlets. They are covered with brown scales and contain many leaves.
[Illustration: FIG. 18.—Branch of Cherry in winter state: a, leaf-scar; b, bud-scar; c, flower-scar.]
[Illustration: FIG. 19.—Branch of Red Maple in winter state (reduced). 2. Flower-buds]
The leaves are needle-shaped and short.[1] They are arranged densely on the branches, alternately on the 8/21 plan (see section on phyllotaxy). When they drop off they leave a hard, blunt projection which makes the stem very rough. As the terminal bud always develops unless injured, the tree is excurrent, forming a straight trunk, throwing out branches on every side. The axillary buds develop near the ends of the branchlets, forming apparent whorls of branches around the trunk. In the smaller branches, as the tree grows older, the tendency is for only two buds to develop nearly opposite each other, forming a symmetrical branch.
The bud-scales are persistent on the branches and the growth from year to year can be traced a long way back.
The cones hang on the ends of the upper branches. They are much larger than in our native species of Black and White Spruce.
The Evergreens are a very interesting study and an excellent exercise in morphology for the older scholars.
2. Vernation. This term signifies the disposition of leaves in the bud, either in respect to the way in which each leaf is folded, or to the manner in which the leaves are arranged with reference to each other. The pupils have described the folding of the leaves in some of their specimens.
In the Beech, the leaf is plicate, or plaited on the veins. In the Elm, Magnolia, and Tulip-tree, it is conduplicate, that is, folded on the midrib with the inner face within. In the Tulip-tree, it is also inflexed, the blade bent forwards on the petiole. In the Balm of Gilead, the leaf is involute, rolled towards the midrib on the upper face.
Other kinds of vernation are revolute, the opposite of involute, where the leaf is rolled backwards towards the midrib; circinate, rolled from the apex downwards, as we see in ferns; and corrugate, when the leaf is crumpled in the bud.
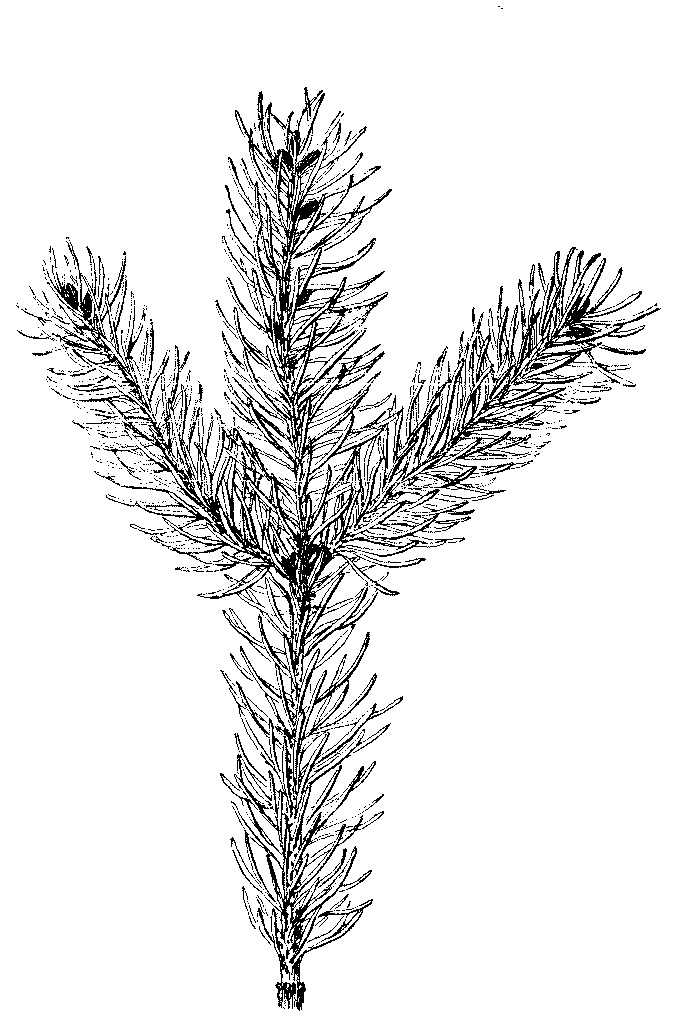
[Illustration: FIG. 20.—Branch of Norway Spruce.]
In all the trees we have studied, the leaves simply succeed each other, each leaf, or pair of leaves, overlapping the next in order. The names of the overlapping of the leaves among themselves, imbricated, convolute, etc., will not be treated here, as they are not needed. They will come under æstivation, the term used to describe the overlapping of the modified leaves, which make up the flower.[1]
3. Phyllotaxy. The subject of leaf-arrangement is an extremely difficult one, and it is best, even with the older pupils, to touch it lightly. The point to be especially brought out is the disposition of the leaves so that each can get the benefit of the light. This can be seen in any plant and there are many ways in which the desired result is brought about. The chief way is the distribution of the leaves about the stem, and this is well studied from the leaf-scars.
The scholars should keep the branches they have studied. It is well to have them marked with the respective names, that the teacher may examine and return them without fear of mistakes.
In the various branches that the pupils have studied, they have seen that the arrangement of the leaves differs greatly. The arrangement of leaves is usually classed under three modes: the alternate, the opposite, and the whorled; but the opposite is the simplest form of the whorled arrangement, the leaves being in circles of two. In this arrangement, the leaves of each whorl stand over the spaces of the whorl just below. The pupils have observed and noted this in Horsechestnut and Lilac. In these there are four vertical rows or ranks of leaves. In whorls of three leaves there would be six ranks, in whorls of four, eight, and so on.
When the leaves are alternate, or single at each node of the stem, they are arranged in many different ways. Ask the pupils to look at all the branches with alternate leaves that they have studied, and determine in each case what leaves stand directly over each other. That is, beginning with any leaf, count the number of leaves passed on the stem, till one is reached that stands directly over the first.[1] In the Beech and the Elm the leaves are on opposite sides of the stem, so that the third stands directly over the first. This makes two vertical ranks, or rows, of leaves, dividing the circle into halves. It is, therefore, called the 1/2 arrangement. Another way of expressing it is to say that the angular divergence between the leaves is 180°, or one-half the circumference.
The 1/3 arrangement, with the leaves in three vertical ranks, is not very common. It may be seen in Sedges, in the Orange-tree, and in Black Alder (Ilex verticillata). In this arrangement, there are three ranks of leaves, and each leaf diverges from the next at an angle of 120°, or one-third of the circumference.
By far the commonest arrangement is with the leaves in five vertical ranks. The Cherry, the Poplar, the Larch, the Oak, and many other trees exhibit this. In this arrangement there are five leaves necessary to complete the circle. We might expect, then, that each leaf would occupy one-fifth of the circle. This would be the case were it not for the fact that we have to pass twice around the stem in counting them, so that each leaf has twice as much room, or two-fifths of the circle, to itself. This is, therefore, the 2/5 arrangement. This can be shown by winding a thread around the stem, passing it over each leaf-scar. In the Beech we make one turn of the stem before reaching the third leaf which stands over the first. In the Apple the thread will wind twice about the stem, before coming to the sixth leaf, which is over the first.
Another arrangement, not very common, is found in the Magnolia, the Holly, and the radical leaves of the common Plantain and Tobacco. The thread makes three turns of the stem before reaching the eighth leaf which stands over the first. This is the 3/8 arrangement. It is well seen in the Marguerite, a greenhouse plant which is very easily grown in the house.
Look now at these fractions, 1/2, 1/3, 2/5, and 3/8. The numerator of the third is the sum of the numerators of the first and second, its denominator, the sum of the two denominators. The same is true of the fourth fraction and the two immediately preceding it. Continuing the series, we get the fractions 5/13, 8/21, 13/34. These arrangements can be found in nature in cones, the scales of which are modified leaves and follow the laws of leaf-arrangement.[1]
[1]"It is to be noted that the distichous or 1/2 variety gives the maximum divergence, namely 180°, and that the tristichous, or 1/3, gives the least, or 120°; that the pentastichous, or 2/5, is nearly the mean between the first two; that of the 3/8, nearly the mean between the two preceding, etc. The disadvantage of the two-ranked arrangement is that the leaves are soon superposed and so overshadow each other. This is commonly obviated by the length of the internodes, which is apt to be much greater in this than in the more complex arrangements, therefore placing them vertically further apart; or else, as in Elms, Beeches, and the like, the branchlets take a horizontal position and the petioles a quarter twist, which gives full exposure of the upper face of all the leaves to the light. The 1/3 and 2/5, with diminished divergence, increase the number of ranks; the 3/8 and all beyond, with mean divergence of successive leaves, effect a more thorough distribution, but with less and less angular distance between the vertical ranks."
For directions for finding the arrangement of cones, see Gray's Structural Botany, Chap. IV, Sect. 1.
The subject appears easy when stated in a text-book, but, practically, it is often exceedingly difficult to determine the arrangement. Stems often twist so as to alter entirely the apparent disposition of the leaves. The general principle, however, that the leaves are disposed so as to get the best exposure to air and light is clear. This cannot be shown by the study of the naked branches merely, because these do not show the beautiful result of the distribution.[1] Many house plants can be found, which will afford excellent illustrations (Fig. 21). The Marguerite and Tobacco, both easily grown in the house, are on the 3/8 plan. The latter shows the eight ranks most plainly in the rosette of its lower leaves. The distribution is often brought about by differences in the lengths of the petioles, as in a Horsechestnut branch (Fig. 22) where the lower, larger leaves stand out further from the branch than the upper ones; or by a twist in the petioles, so that the upper faces of the leaves are turned up to the light, as in Beech (Fig. 23). If it is springtime when the lessons are given, endless adaptations can be found.
[Illustration: FIG. 21. Branch of Geranium, viewed from above.]
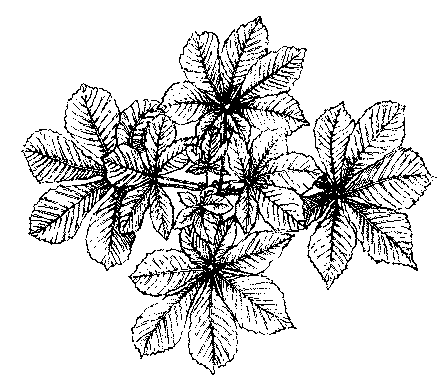
[Illustration: FIG. 22.]
[Illustration: FIG. 23.]
Gray's First Lessons. Sect. IV. VII, §4. How Plants Grow. Chap. I, 51-62; I, 153.
The stem, as the scholars have already learned, is the axis of the plant. The leaves are produced at certain definite points called nodes, and the portions of stem between these points are internodes. The internode, node, and leaf make a single plant-part, and the plant is made up of a succession of such parts.
The stem, as well as the root and leaves, may bear plant-hairs. The accepted theory of plant structure assumes that these four parts, root, stem, leaves, and plant-hairs, are the only members of a flowering plant, and that all other forms, as flowers, tendrils, etc., are modified from these. While this idea is at the foundation of all our teaching, causing us to lead the pupil to recognize as modified leaves the cotyledons of a seedling and the scales of a bud, it is difficult to state it directly so as to be understood, except by mature minds. I have been frequently surprised at the failure of even bright and advanced pupils to grasp this idea, and believe it is better to let them first imbibe it unconsciously in their study. Whenever their minds are ready for it, it will be readily understood. The chief difficulty is that they imagine that there is a direct metamorphosis of a leaf to a petal or a stamen.
Briefly, the theory is this: the beginnings of leaf, petal, tendril, etc., are the same. At an early stage of their growth it is impossible to tell what they are to become. They develop into the organ needed for the particular work required of them to do. The organ, that under other circumstances might develop into a leaf, is capable of developing into a petal, a stamen, or a pistil, according to the requirements of the plant, but no actual metamorphosis takes place. Sometimes, instead of developing into the form we should normally find, the organ develops into another form, as when a petal stands in the place of a stamen, or the pistil reverts to a leafy branch. This will be more fully treated under flowers. The study of the different forms in which an organ may appear is the study of morphology.
1. Forms of Stems.—Stems may grow in many ways. Let the pupils compare the habits of growth of the seedlings they have studied. The Sunflower and Corn are erect. This is the most usual habit, as with our common trees. The Morning Glory is twining, the stem itself twists about a support. The Bean, Pea and Nasturtium are climbing. The stems are weak, and are held up, in the first two by tendrils, in the last by the twining leaf-stalks. The English Ivy, as we have seen, is also climbing, by means of its aërial roots. The Red Clover is ascending, the branches rising obliquely from the base. Some kinds of Clover, as the White Clover, are creeping, that is, with prostrate branches rooting at the nodes and forming new plants. Such rooting branches are called stolons, or when the stem runs underground, suckers. The gardener imitates them in the process called layering, that is, bending down an erect branch and covering it with soil, causing it to strike root. When the connecting stem is cut, a new plant is formed. Long and leafless stolons, like those of the Strawberry are called runners. Stems creep below the ground as well as above. Probably the pupil will think of some examples. The pretty little Gold Thread is so named from the yellow running stems, which grow beneath the ground and send up shoots, or suckers, which make new plants. Many grasses propagate themselves in this way. Such stems are called rootstocks. "That these are really stems, and not roots, is evident from the way in which they grow; from their consisting of a succession of joints; and from the leaves which they bear on each node, in the form of small scales, just like the lowest ones on the upright stem next the ground. They also produce buds in the axils of these scales, showing the scales to be leaves; whereas real roots bear neither leaves nor axillary buds."[1] Rootstocks are often stored with nourishment. We have already taken up this subject in the potato, but it is well to repeat the distinction between stems and roots. A thick, short rootstock provided with buds, like the potato, is called a tuber. Compare again the corm of Crocus and the bulb of Onion to find the stem in each. In the former, it makes the bulk of the whole; in the latter, it is a mere plate holding the fleshy bases of the leaves.
2. Movements of Stems.—Let a glass thread, no larger than a coarse hair, be affixed by means of some quickly drying varnish to the tip of the laterally inclined stem of one of the young Morning-Glory plants in the schoolroom. Stand a piece of cardboard beside the pot, at right angles to the stem, so that the end of the glass will be near the surface of the card. Make a dot upon the card opposite the tip of the filament, taking care not to disturb the position of either. In a few minutes observe that the filament is no longer opposite the dot. Mark its position anew, and continue thus until a circle is completed on the cardboard. This is a rough way of conducting the experiment. Darwin's method will be found in the footnote.[1]
The use of the glass filament is simply to increase the size of the circle described, and thus make visible the movements of the stem. All young parts of stems are continually moving in circles or ellipses. "To learn how the sweeps are made, one has only to mark a line of dots along the upper side of the outstretched revolving end of such a stem, and to note that when it has moved round a quarter of a circle, these dots will be on one side; when half round, the dots occupy the lower side; and when the revolution is completed, they are again on the upper side. That is, the stem revolves by bowing itself over to one side,—is either pulled over or pushed over, or both, by some internal force, which acts in turn all round the stem in the direction in which it sweeps; and so the stem makes its circuits without twisting."[1]
The nature of the movement is thus a successive nodding to all the points of the compass, whence it is called by Darwin circumnutation. The movement belongs to all young growing parts of plants. The great sweeps of a twining stem, like that of the Morning-Glory, are only an increase in the size of the circle or ellipse described.[1]
When a young stem of a Morning-Glory, thus revolving, comes in contact with a support, it will twist around it, unless the surface is too smooth to present any resistance to the movement of the plant. Try to make it twine up a glass rod. It will slip up the rod and fall off. The Morning-Glory and most twiners move around from left to right like the hands of a clock, but a few turn from right to left.
While this subject is under consideration, the tendrils of the Pea and Bean and the twining petioles of the Nasturtium will be interesting for comparison. The movements can be made visible by the same method as was used for the stem of the Morning-Glory. Tendrils and leaf petioles are often sensitive to the touch. If a young leaf stalk of Clematis be rubbed for a few moments, especially on the under side, it will be found in a day or two to be turned inward, and the tendrils of the Cucumber vine will coil in a few minutes after being thus irritated.[1] The movements of tendrils are charmingly described in the chapter entitled "How Plants Climb," in the little treatise by Dr. Gray, already mentioned.
The so-called "sleep of plants" is another similar movement. The Oxalis is a good example. The leaves droop and close together at night, protecting them from being chilled by too great radiation.
The cause of these movements is believed to lie in changes of tension preceding growth in the tissues of the stem.[1] Every stem is in a state of constant tension. Naudin has thus expressed it, "the interior of every stem is too large for its Jacket."[2] If a leaf-stalk of Nasturtium be slit vertically for an inch or two, the two halves will spring back abruptly. This is because the outer tissues of the stem are stretched, and spring back like india-rubber when released. If two stalks twining in opposite directions be slit as above described, the side of the stem towards which each stalk is bent will spring back more than the other, showing the tension to be greater on that side. A familiar illustration of this tension will be found in the Dandelion curls of our childhood.
The movements of the Sensitive Plant are always very interesting to pupils, and it is said not to be difficult to raise the plants in the schoolroom. The whole subject, indeed, is one of the most fascinating that can be found, and its literature is available, both for students and teachers. Darwin's essay on "Climbing Plants," and his later work on the "Power of Movement in Plants," Dr. Gray's "How Plants Behave," and the chapter on "Movements" in the "Physiological Botany," will offer a wide field for study and experiment.
3. Structure of Stems.—Let the pupils collect a series of branches of some common tree or shrub, from the youngest twig up to as large a branch as they can cut, and describe them. Poplar, Elm, Oak, Lilac, etc., will be found excellent for the purpose.
While discussing these descriptions, a brief explanation of plant-structure may be given. In treating this subject, the teacher must govern himself by the needs of his class, and the means at his command. Explanations requiring the use of a compound microscope do not enter necessarily into these lessons. The object aimed at is to teach the pupils about the things which they can see and handle for themselves. Looking at sections that others have prepared is like looking at pictures; and, while useful in opening their eyes and minds to the wonders hidden from our unassisted sight, fails to give the real benefit of scientific training. Plants are built up of cells. The delicate-walled spherical, or polygonal, cells which make up the bulk of an herbaceous stem, constitute cellular tissue (parenchyma). This was well seen in the stem of the cutting of Bean in which the roots had begun to form.[1] The strengthening fabric in almost all flowering plants is made up of woody bundles, or woody tissue.[2] The wood-cells are cells which are elongated and with thickened walls. There are many kinds of them. Those where the walls are very thick and the cavity within extremely small are fibres. A kind of cell, not strictly woody, is where many cells form long vessels by the breaking away of the connecting walls. These are ducts. These two kinds of cells are generally associated together in woody bundles, called therefore fibro-vascular bundles. We have already spoken of them as making the dots on the leaf-scars, and forming the strengthening fabric of the leaves.[3]
We will now examine our series of branches. The youngest twigs, in spring or early summer, are covered with a delicate, nearly colorless skin. Beneath this is a layer of bark, usually green, which gives the color to the stem, an inner layer of bark, the wood and the pith. The pith is soft, spongy and somewhat sappy. There is also sap between the bark and the wood. An older twig has changed its color. There is a layer of brown bark, which has replaced the colorless skin. In a twig a year old the wood is thicker and the pith is dryer. Comparing sections of older branches with these twigs, we find that the pith has shrunk and become quite dry, and that the wood is in rings. It is not practicable for the pupils to compare the number of these rings with the bud-rings, and so find out for themselves that the age of the branch can be determined from the wood, for in young stems the successive layers are not generally distinct. But, in all the specimens, the sap is found just between the wood and the bark, and here, where the supply of food is, is where the growth is taking place. Each year new wood and new bark are formed in this cambium-layer, as it is called, new wood on its inner, new bark on its outer face. Trees which thus form a new ring of wood every year are called exogenous, or outside-growing.
Ask the pupils to separate the bark into its three layers and to try the strength of each. The two outer will easily break, but the inner is generally tough and flexible. It is this inner bark, which makes the Poplar and Willow branches so hard to break. These strong, woody fibres of the inner bark give us many of our textile fabrics. Flax and Hemp come from the inner bark of their respective plants (Linum usitatissimum and Cannabis sativa), and Russia matting is made from the bark of the Linden (Tilia Americana).
We have found, in comparing the bark of specimens of branches of various ages, that, in the youngest stems, the whole is covered with a skin, or epidermis, which is soon replaced by a brown outer layer of bark, called the corky layer; the latter gives the distinctive color to the tree. While this grows, it increases by a living layer of cork-cambium on its inner face, but it usually dies after a few years. In some trees it goes on growing for many years. It forms the layers of bark in the Paper Birch and the cork of commerce is taken from the Cork Oak of Spain. The green bark is of cellular tissue, with some green coloring matter like that of the leaves; it is at first the outer layer, but soon becomes covered with cork. It does not usually grow after the first year. Scraping the bark of an old tree, we find the bark homogeneous. The outer layers have perished and been cast off. As the tree grows from within, the bark is stretched and, if not replaced, cracks and falls away piecemeal. So, in most old trees, the bark consists of successive layers of the inner woody bark.
Stems can be well studied from pieces of wood from the woodpile. The ends of the log will show the concentric rings. These can be traced as long, wavy lines in vertical sections of the log, especially if the surface is smooth. If the pupils can whittle off different planes for themselves, they will form a good idea of the formation of the wood. In many of the specimens there will be knots, and the nature of these will be an interesting subject for questions. If the knot is near the centre of the log, lead back their thoughts to the time when the tree was as small as the annular ring on which the centre of the knot lies. Draw a line on this ring to represent the tree at this period of its growth. What could the knot have been? It has concentric circles like the tree itself. It was a branch which decayed, or was cut off. Year after year, new rings of wood formed themselves round this broken branch, till it was covered from sight, and every year left it more deeply buried in the trunk.
Extremely interesting material for the study of wood will be found in thin sections prepared for veneers. Packages of such sections will be of great use to the teacher.[1] They show well the reason of the formation of a dividing line between the wood of successive seasons. In a cross section of Oak or Chestnut the wood is first very open and porous and then close. This is owing to the presence of ducts in the wood formed in the spring. In other woods there are no ducts, or they are evenly distributed, but the transition from the close autumn wood, consisting of smaller and more closely packed cells, to the wood of looser texture, formed in the following spring, makes a line that marks the season's growth.
Let each of the scholars take one of the sections of Oak and write a description of its markings. The age is easily determined; the pith rays, or medullary rays, are also plain. These form what is called the silver grain of the wood. The ducts, also, are clear in the Oak and Chestnut. There is a difference in color between the outer and inner wood, the older wood becomes darker and is called the heart-wood, the outer is the sap-wood. In Birds-eye Maple, and some other woods, the abortive buds are seen. They are buried in the wood, and make the disturbance which produces the ornamental grain. In sections of Pine or Spruce, no ducts can be found. The wood consists entirely of elongated, thickened cells or fibres. In some of the trees the pith rays cannot be seen with the naked eye.
Let the pupils compare the branches which they have described, with a stalk of Asparagus, Rattan, or Lily. A cross section of one of these shows dots among the soft tissue. These are ends of the fibro-vascular bundles, which in these plants are scattered through the cellular tissue instead of being brought together in a cylinder outside of the pith. In a vertical section they appear as lines. There are no annular rings.
If possible, let the pupils compare the leaves belonging to these different types of stems. The parallel-veined leaves of monocotyledons have stems without distinction of wood, bark and pith; the netted-veined leaves of dicotyledons have exogenous stems.
Dicotyledons have bark, wood, and pith, and grow by producing a new ring of wood outside the old. They also increase by the growth of the woody bundles of the leaves, which mingle with those of the stem.[1] Twist off the leaf-stalk of any leaf, and trace the bundles into the stem.
Monocotyledons have no layer which has the power of producing new wood, and their growth takes place entirely from the intercalation of new bundles, which originate at the bases of the leaves. The lower part of a stem of a Palm, for instance, does not increase in size after it has lost its crown of leaves. This is carried up gradually. The upper part of the stem is a cone, having fronds, and below this cone the stem does not increase in diameter. The word endogenous, inside-growing, is not, therefore, a correct one to describe the growth of most monocotyledons, for the growth takes place where the leaves originate, near the exterior of the stem.
Gray's First Lessons. Sect. VI. Sect, XVI, §1, 401-13. §3. §6, 465-74.
How Plants Grow. Chap. 1, 82, 90-118.
We have studied leaves as cotyledons, bud-scales, etc., but when we speak of leaves, we do not think of these adapted forms, but of the green foliage of the plant.
1. Forms and Structure.—Provide the pupils with a number of green leaves, illustrating simple and compound, pinnate and palmate, sessile and petioled leaves. They must first decide the question, What are the parts of a leaf? All the specimens have a green blade which, in ordinary speech, we call the leaf. Some have a stalk, or petiole, others are joined directly to the stem. In some of them, as a rose-leaf, for instance, there are two appendages at the base of the petiole, called stipules. These three parts are all that any leaf has, and a leaf that has them all is complete.
Let us examine the blade. Those leaves which have the blade in one piece are called simple; those with the blade in separate pieces are compound. We have already answered the question, What constitutes a single leaf?[1] Let the pupils repeat the experiment of cutting off the top of a seedling Pea, if it is not already clear in their minds, and find buds in the leaf-axils of other plants.[2]
An excellent way to show the nature of compound leaves is to mount a series showing every gradation of cutting, from a simple, serrate leaf to a compound one (Figs. 24 and 25). A teacher, who would prepare in summer such illustrations as these, would find them of great use in his winter lessons. The actual objects make an impression that the cuts in the book cannot give.
[Illustration: FIG. 24.—Series of palmately-veined leaves.]
[Illustration: FIG. 25.—Series of pinnately-veined leaves.]
Let the pupils compare the distribution of the veins in their specimens. They have already distinguished parallel-veined from netted-veined leaves, and learned that this difference is a secondary distinction between monocotyledons and dicotyledons.[1] The veins in netted-veined leaves are arranged in two ways. The veins start from either side of a single midrib (feather-veined or pinnately-veined), or they branch from a number of ribs which all start from the top of the petiole, like the fingers from the palm of the hand (palmately-veined). The compound leaves correspond to these modes of venation; they are either pinnately or palmately compound.
These ribs and veins are the woody framework of the leaf, supporting the soft green pulp. The woody bundles are continuous with those of the stem, and carry the crude sap, brought from the roots, into the cells of every part of the leaf, where it is brought into contact with the external air, and the process of making food (Assimilation 4) is carried on. "Physiologically, leaves are green expansions borne by the stern, outspread in the air and light, in which assimilation and the processes connected with it are carried on."[1]
The whole leaf is covered with a delicate skin, or epidermis, continuous with that of the stem.[1]
2. Descriptions.—As yet the pupils have had no practice in writing technical descriptions. This sort of work may be begun when they come to the study of leaves. In winter a collection of pressed specimens will be useful. Do not attach importance to the memorizing of terms. Let them be looked up as they are needed, and they will become fixed by practice. The pupils may fill out such schedules as the following with any leaves that are at hand.
SCHEDULE FOR LEAVES.
| 1. BLADE | Arrangement | Alternate[1] |
| Simple or compound. (arr. and no. of leaflets) | Simple | |
| Venation | Netted and feather-veined | |
| Shape | Oval | |
| Apex | Acute | |
| Base | Oblique | |
| Margin | Slightly wavy | |
| Surface | Smooth | |
| 2. PETIOLE | Short; hairy | |
| 3. STIPULES | Deciduous | |
| Remarks. Veins prominent and very straight. | ||
In describing shapes, etc., the pupils can find the terms in the book as they need them. It is desirable at first to give leaves that are easily matched with the terms, keeping those which need compound words, such as lance-ovate, etc., to come later. The pupils are more interested if they are allowed to press and keep the specimens they have described. It is not well to put the pressed leaves in their note books, as it is difficult to write in the books without spoiling the specimens. It is better to mount the specimens on white paper, keeping these sheets in brown paper covers. The pupils can make illustrations for themselves by sorting leaves according to the shapes, outlines, etc., and mounting them.
3. Transpiration.—This term is used to denote the evaporation of water from a plant. The evaporation takes place principally through breathing pores, which are scattered all over the surface of leaves and young stems. The breathing pores, or stomata, of the leaves, are small openings in the epidermis through which the air can pass into the interior of the plant. Each of these openings is called a stoma. "They are formed by a transformation of some of the cells of the epidermis; and consist usually of a pair of cells (called guardian cells), with an opening between them, which communicates with an air-chamber within, and thence with the irregular intercellular spaces which permeate the interior of the leaf. Through the stomata, when open, free interchange may take place between the external air and that within the leaf, and thus transpiration be much facilitated. When closed, this interchange will be interrupted or impeded."[1]
In these lessons, however, it is not desirable to enter upon subjects involving the use of the compound microscope. Dr. Goodale says: "Whether it is best to try to explain to the pupils the structure of these valves, or stomata, must be left to each teacher. It would seem advisable to pass by the subject untouched, unless the teacher has become reasonably familiar with it by practical microscopical study of leaves. For a teacher to endeavor to explain the complex structure of the leaf, without having seen it for himself, is open to the same objection which could be urged against the attempted explanation of complicated machinery by one who has never seen it, but has heard about it. What is here said with regard to stomata applies to all the more recondite matters connected with plant structure."[1]
There are many simple experiments which can be used to illustrate the subject.
(1) Pass the stem of a cutting through a cork, fitting tightly into the neck of a bottle of water. Make the cork perfectly air-tight by coating it with beeswax or paraffine. The level of the liquid in the bottle will be lowered by the escape of water through the stem and leaves of the cutting into the atmosphere.
(2) Cut two shoots of any plant, leave one on the table and place the other in a glass of water.[1] The first will soon wilt, while the other will remain fresh. If the latter shoot be a cutting from some plant that will root in water, such as Ivy, it will not fade at all. Also, leave one of the plants in the schoolroom unwatered for a day or two, till it begins to wilt. If the plant be now thoroughly watered, it will recover and the leaves will resume their normal appearance.
Evaporation is thus constantly taking place from the leaves, and if there is no moisture to supply the place of what is lost, the cells collapse and the leaf, as we say, wilts. When water is again supplied the cells swell and the leaf becomes fresh.
(3) Place two seedlings in water, one with its top, the other with its roots in the jar. The latter will remain fresh while the first wilts and dies.
Absorption takes place through the roots. The water absorbed is drawn up through the woody tissues of the stem (4), and the veins of the leaves (5), whence it escapes into the air (6).
(4) Plunge a cut branch immediately into a colored solution, such as aniline red, and after a time make sections in the stem above the liquid to see what tissues have been stained.[1]
(5) "That water finds its way by preference through the fibro-vascular bundles even in the more delicate parts, is shown by placing the cut peduncle of a white tulip, or other large white flower, in a harmless dye, and then again cutting off its end in order to bring a fresh surface in contact with the solution,[1] when after a short time the dye will mount through the flower-stalk and tinge the parts of the perianth according to the course of the bundles."[2]
(6) Let the leaves of a growing plant rest against the window-pane. Moisture will be condensed on the cold surface of the glass, wherever the leaf is in contact with it. This is especially well seen in Nasturtium (Tropæolum) leaves, which grow directly against a window, and leave the marks even of their veining on the glass, because the moisture is only given out from the green tissue, and where the ribs are pressed against the glass it is left dry.
Sometimes the water is drawn up into the cells of the leaves faster than it can escape into the atmosphere.[1] This is prettily shown if we place some of our Nasturtium seedlings under a ward-case. The air in the case is saturated with moisture, so that evaporation cannot take place, but the water is, nevertheless, drawn up from the roots and through the branches, and appears as little drops on the margins of the leaves. That this is owing to the absorbing power of the roots, may be shown by breaking off the seedling, and putting the slip in water. No drops now appear on the leaves, but as soon as the cutting has formed new roots, the drops again appear.
This constant escape of water from the leaves causes a current to flow from the roots through the stem into the cells of the leaves. The dilute mineral solutions absorbed by the roots[1] are thus brought where they are in contact with the external air, concentrated by the evaporation of water, and converted in these cells into food materials, such as starch. The presence of certain mineral matters, as potassium, iron, etc., are necessary to this assimilating process, but the reason of their necessity is imperfectly understood, as they do not enter in the products formed.
The amount of water exhaled is often very great. Certain plants are used for this reason for the drainage of wet and marshy places. The most important of these is the Eucalyptus tree.[1]
"The amount of water taken from the soil by the trees of a forest and passed into the air by transpiration is not so large as that accumulated in the soil by the diminished evaporation under the branches. Hence, there is an accumulation of water in the shade of forests which is released slowly by drainage.[1] But if the trees are so scattered as not materially to reduce evaporation from the ground, the effect of transpiration in diminishing the moisture of the soil is readily shown. It is noted, especially in case of large plants having a great extent of exhaling surface, such, for instance, as the common sunflower. Among the plants which have been successfully employed in the drainage of marshy soil by transpiration probably the species of Eucalyptus (notably E. globulus) are most efficient."[2]
4. Assimilation.—It is not easy to find practical experiments on assimilation. Those which follow are taken from "Physiological Botany" (p. 305).
Fill a five-inch test tube, provided with a foot, with fresh drinking water. In this place a sprig of one of the following water plants,—Elodea Canadensis, Myriophyllum spicatum, M. verticillatum, or any leafy Myriophyllum (in fact, any small-leaved water plant with rather crowded foliage). This sprig should be prepared as follows: Cut the stem squarely off, four inches or so from the tip, dry the cut surface quickly with blotting paper, then cover the end of the stein with a quickly drying varnish, for instance, asphalt-varnish, and let it dry perfectly, keeping the rest of the stem, if possible, moist by means of a wet cloth. When the varnish is dry, puncture it with a needle, and immerse the stem in the water in the test tube, keeping the varnished larger end uppermost. If the submerged plant be now exposed to the strong rays of the sun, bubbles of oxygen gas will begin to pass off at a rapid and even rate, but not too fast to be easily counted. If the simple apparatus has begun to give off a regular succession of small bubbles, the following experiments can be at once conducted:
(1) Substitute for the fresh water some which has been boiled a few minutes before, and then allowed to completely cool: by the boiling, all the carbonic acid has been expelled. If the plant is immersed in this water and exposed to the sun's rays, no bubbles will be evolved; there is no carbonic acid within reach of the plant for the assimilative process. But,
(2) If breath from the lungs be passed by means of a slender glass tube through the water, a part of the carbonic acid exhaled from the lungs will be dissolved in it, and with this supply of the gas the plant begins the work of assimilation immediately.
(3) If the light be shut off, the evolution of bubbles will presently cease, being resumed soon after light again has access to the plant.
(5) Place round the base of the test tube a few fragments of ice, in order to appreciably lower the temperature of the water. At a certain point it will be observed that no bubbles are given off, and their evolution does not begin again until the water becomes warm.
The evolution of bubbles shows that the process of making food is going on. The materials for this process are carbonic acid gas and water. The carbonic acid dissolved in the surrounding water is absorbed, the carbon unites with the elements of water in the cells of the leaves, forming starch, etc., and most of the oxygen is set free, making the stream of bubbles. When the water is boiled, the dissolved gas is driven off and assimilation cannot go on; but as soon as more carbonic acid gas is supplied, the process again begins. We have seen by these experiments that sunlight and sufficient heat are necessary to assimilation, and that carbonic acid gas and water must be present. The presence of the green coloring matter of the leaves (chlorophyll) is also essential, and some salts, such as potassium, iron, etc., are needful, though they may not enter into the compounds formed.
The food products are stored in various parts of the plant for future use, or are expended immediately in the growth and movements of the plant. In order that they shall be used for growth, free oxygen is required, and this is supplied by the respiration of the plant.
Some plants steal their food ready-made. Such a one is the Dodder, which sends its roots directly into the plant on which it feeds. This is a parasite.[1] It has no need of leaves to carry on the process of making food. Some parasites with green leaves, like the mistletoe, take the crude sap from the host-plant and assimilate it in their own green leaves. Plants that are nourished by decaying matter in the soil are called saprophytes. Indian Pipe and Beech-Drops are examples of this. They need no green leaves as do plants that are obliged to support themselves.
Some plants are so made that they can use animal matter for food. This subject of insectivorous plants is always of great interest to pupils. If some Sundew (Drosera) can be obtained and kept in the schoolroom, it will supply material for many interesting experiments.[1] That plants should possess the power of catching insects by specialized movements and afterwards should digest them by means of a gastric juice like that of animals, is one of the most interesting of the discoveries that have been worked out during the last thirty years.[2]
5. Respiration.—Try the following experiment in germination.
Place some seeds on a sponge under an air-tight glass. Will they grow? What causes them to mould?
Seeds will not germinate without free access of air. They must have free oxygen to breathe, as must every living thing. We know that an animal breathes in oxygen, that the oxygen unites with particles of carbon within the body and that the resulting carbonic acid gas is exhaled.[1] The same process goes on in plants, but it was until recently entirely unknown, because it was completely masked during the daytime by the process of assimilation, which causes carbonic acid to be inhaled and decomposed, and oxygen to be exhaled.[2] In the night time the plants are not assimilating and the process of breathing is not covered up. It has, therefore, long been known that carbonic acid gas is given off at night. The amount, however, is so small that it could not injure the air of the room, as is popularly supposed. Respiration takes place principally through the stomata of the leaves.[3] We often see plants killed by the wayside dust, and we all know that on this account it is very difficult to make a hedge grow well by a dusty road. The dust chokes up the breathing pores of the leaves, interfering with the action of the plant. It is suffocated.
The oxygen absorbed decomposes starch, or some other food product of the plant, and carbonic acid gas and water are formed. It is a process of slow combustion.[4] The energy set free is expended in growth, that is, in the formation of new cells, and the increase in size of the old ones, and in the various movements of the plant.
| ASSIMILATION PROPER. | RESPIRATION. |
| Takes place only in cells containing chlorophyll. | Takes place in all active cells. |
| Requires light. | Can proceed in darkness. |
| Carbonic acid absorbed, oxygen set free. | Oxygen absorbed, carbonic acid set free. |
| Carbohydrates formed. | Carbohydrates consumed. |
| Energy of motion becomes energy of position. | Energy of position becomes energy of motion. |
| The plant gains in dry weight. | The plant loses dry weight. |
[**Proofers Note: Two footnote marks [3] and [4] above in original text, but no footnote text is in the original text.]
This process of growth can take place only when living protoplasm is present in the cells of the plant. The substance we call protoplasm is an albuminoid, like the white of an egg, and it forms the flesh of both plants and animals. A living plant can assimilate its own protoplasm, an animal must take it ready-made from plants. But a plant can assimilate its food and grow only under the mysterious influence we call life. Life alone brings forth life, and we are as far as ever from understanding its nature. Around our little island of knowledge, built up through the centuries by the labor of countless workers, stretches the infinite ocean of the unknown.
Gray's First Lessons. Sect. VII, XVI, §2, §4, §5, §6, 476-480.
How Plants Grow. Chap. I, 119-153, Chap. III, 261-280.
***END OF THE PROJECT GUTENBERG EBOOK OUTLINES OF LESSONS IN BOTANY, PART I; FROM SEED TO LEAF***
******* This file should be named 10726-h.txt or 10726-h.zip *******
This and all associated files of various formats will be found in:
https://www.gutenberg.org/1/0/7/2/10726
Updated editions will replace the previous one--the old editions will be renamed.
Creating the works from public domain print editions means that no one owns a United States copyright in these works, so the Foundation (and you!) can copy and distribute it in the United States without permission and without paying copyright royalties. Special rules, set forth in the General Terms of Use part of this license, apply to copying and distributing Project Gutenberg-tm electronic works to protect the PROJECT GUTENBERG-tm concept and trademark. Project Gutenberg is a registered trademark, and may not be used if you charge for the eBooks, unless you receive specific permission. If you do not charge anything for copies of this eBook, complying with the rules is very easy. You may use this eBook for nearly any purpose such as creation of derivative works, reports, performances and research. They may be modified and printed and given away--you may do practically ANYTHING with public domain eBooks. Redistribution is subject to the trademark license, especially commercial redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License (available with this file or online at
https://gutenberg.org/license).
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH F3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS,' WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTIBILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need, is critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation web page at https://www.pglaf.org.
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Its 501(c)(3) letter is posted at
https://pglaf.org/fundraising. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at
809 North 1500 West, Salt Lake City, UT 84116, (801) 596-1887, email
[email protected]. Email contact links and up to date contact
information can be found at the Foundation's web site and official
page at https://pglaf.org
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
[email protected]
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit https://pglaf.org
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including including checks, online payments and credit card
donations. To donate, please visit: https://pglaf.org/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart was the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For thirty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Each eBook is in a subdirectory of the same number as the eBook's
eBook number, often in several formats including plain vanilla ASCII,
compressed (zipped), HTML and others.
Corrected EDITIONS of our eBooks replace the old file and take over
the old filename and etext number. The replaced older file is renamed.
VERSIONS based on separate sources are treated as new eBooks receiving
new filenames and etext numbers.
Most people start at our Web site which has the main PG search facility:
https://www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.
EBooks posted prior to November 2003, with eBook numbers BELOW #10000,
are filed in directories based on their release date. If you want to
download any of these eBooks directly, rather than using the regular
search system you may utilize the following addresses and just
download by the etext year.
http://www.ibiblio.org/gutenberg/etext06
(Or /etext 05, 04, 03, 02, 01, 00, 99,
98, 97, 96, 95, 94, 93, 92, 92, 91 or 90)
EBooks posted since November 2003, with etext numbers OVER #10000, are
filed in a different way. The year of a release date is no longer part
of the directory path. The path is based on the etext number (which is
identical to the filename). The path to the file is made up of single
digits corresponding to all but the last digit in the filename. For
example an eBook of filename 10234 would be found at:
https://www.gutenberg.org/1/0/2/3/10234
or filename 24689 would be found at:
https://www.gutenberg.org/2/4/6/8/24689
An alternative method of locating eBooks:
https://www.gutenberg.org/GUTINDEX.ALL
*** END: FULL LICENSE ***